RESEARCH ARTICLE
Effects of Structure and Meaning on Cortical
Tracking of Linguistic Units in Naturalistic Speech
a n o p e n a c c e s s
j o u r n a l
Cas W. Coopmans1,2
, Helen de Hoop2
, Peter Hagoort1,3
, and Andrea E. Martin1,3
1Max Planck Institute for Psycholinguistics, Nijmegen, The Netherlands
2Centre for Language Studies, Radboud University, Nijmegen, The Netherlands
3Donders Institute for Brain, Cognition and Behaviour, Radboud University, Nijmegen, The Netherlands
Keywords: EEG, mutual information, compositionality, idioms, jabberwocky
ABSTRACT
Recent research has established that cortical activity “tracks” the presentation rate of syntactic
phrases in continuous speech, even though phrases are abstract units that do not have direct
correlates in the acoustic signal. We investigated whether cortical tracking of phrase structures
is modulated by the extent to which these structures compositionally determine meaning. To
this end, we recorded electroencephalography (EEG) of 38 native speakers who listened to
naturally spoken Dutch stimuli in different conditions, which parametrically modulated the
degree to which syntactic structure and lexical semantics determine sentence meaning.
Tracking was quantified through mutual information between the EEG data and either the
speech envelopes or abstract annotations of syntax, all of which were filtered in the frequency
band corresponding to the presentation rate of phrases (1.1–2.1 Hz). Overall, these mutual
information analyses showed stronger tracking of phrases in regular sentences than in stimuli
whose lexical-syntactic content is reduced, but no consistent differences in tracking between
sentences and stimuli that contain a combination of syntactic structure and lexical content.
While there were no effects of compositional meaning on the degree of phrase-structure
tracking, analyses of event-related potentials elicited by sentence-final words did reveal
meaning-induced differences between conditions. Our findings suggest that cortical tracking
of structure in sentences indexes the internal generation of this structure, a process that
is modulated by the properties of its input, but not by the compositional interpretation of
its output.
INTRODUCTION
How the brain parses a continuous speech stream into discrete, hierarchically organized units
of linguistic representation remains an important question in the neurobiology of language
(Giraud & Poeppel, 2012; Martin, 2016, 2020; Meyer et al., 2020). A possible mechanism that
the brain might use to extract linguistic information relies on phase alignment between neural
activity and quasi-regular properties of the speech signal. This process, called cortical tracking,
results from the tendency of neural systems to adjust to the timing of (quasi-)regular aspects of
external stimuli, and has been argued to facilitate segmentation and parsing of continuous
speech (for reviews, see Ding & Simon, 2014; Giraud & Poeppel, 2012; Kösem & van
Wassenhove, 2017; Obleser & Kayser, 2019; Peelle & Davis, 2012; Rimmele et al.,
2018; Schroeder & Lakatos, 2009; Zoefel & VanRullen, 2015).
Citation: Coopmans, C. W., de Hoop, H.,
Hagoort, P., & Martin, A. E. (2022).
Effects of structure and meaning on
cortical tracking of linguistic units in
naturalistic speech. Neurobiology of
Language, 3(3), 386–412. https://doi.org
/10.1162/nol_a_00070
DOI:
https://doi.org/10.1162/nol_a_00070
Supporting Information:
https://doi.org/10.1162/nol_a_00070
Received: 14 October 2021
Accepted: 2 March 2022
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Author:
Cas W. Coopmans
cas.coopmans@mpi.nl
Handling Editor:
Alec Marantz
Copyright: © 2022
Massachusetts Institute of Technology
Published under a Creative Commons
Attribution 4.0 International
(CC BY 4.0) license
The MIT Press
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
3
3
8
6
2
0
3
1
3
9
0
n
o
_
a
_
0
0
0
7
0
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cortical tracking of linguistic units
Cortical speech tracking:
The brain response related to slow
fluctuations in the speech stimulus.
Speech envelope:
Changes in the amplitude of a speech
stimulus over time.
Jabberwocky sentence:
A structurally coherent and
prosodically natural sentence in
which all content words are replaced
by pseudowords.
Compositional meaning:
The meaning of an expression that is
a function of the meanings of its parts
and their mode of combination.
Idiom:
A more or less fixed phrase or
expression with a figurative,
non-literal meaning.
Cortical tracking is well established for low-level aspects of the linguistic signal, which
have clear correlates in the physical instantiation of speech (e.g., the speech envelope).
Strikingly, recent work has shown that words and phrases, which are not clearly discernable
in the speech signal and have to be internally constructed, are also cortically tracked (Ding
et al., 2016; Ding, Melloni, et al., 2017). Moreover, these high-level linguistic properties
influence lower-level speech processing, as shown by the fact that cortical tracking of the
speech envelope is modulated by the listener’s knowledge of the language (Broderick et al.,
2019; Di Liberto et al., 2018; Kaufeld et al., 2020).
These studies indicate that the inferred content of a signal affects the extent to which the
brain tracks that signal (see also Keitel et al., 2018; Martin, 2020; ten Oever & Martin, 2021).
What it is still elusive, however, is which aspects of content determine cortical speech track-
ing. In a recent paper, Kaufeld et al. (2020) showed that the neural signal aligns more strongly
with periodically occurring linguistic units, such as syntactic phrases, when these contain
meaningful information and are therefore relevant for linguistic processing. Specifically, cor-
tical tracking of phrase structure was stronger for regular sentences than for control stimuli that
were matched in terms of either lexical semantics (word lists) or both prosody and syntactic
structure (jabberwocky sentences), suggesting that this neural response is driven by the com-
positional meaning of sentence structures. However, the difference between sentences and
these control conditions can be described not only in terms of the output of compositional
processing (i.e., the fact that sentence structures have a meaningful compositional interpreta-
tion), but also in terms of the factors that go into structural composition. To investigate which of
these aspects of linguistic content affect cortical tracking of linguistic structure, the current
electroencephalogram (EEG) study investigates cortical tracking of linguistic units (phrases,
words, syllables) when these are embedded in stimuli that are parametrically varied in terms
of the amount of linguistic information. These stimuli ranged from regular compositional sen-
tences to structure-meaning divergent forms (idioms, syntactic prose), structures with reduced
lexical-syntactic content ( jabberwocky) and unstructured word lists. We thus test how the rela-
tionship between structure and meaning in spoken language affects cortical tracking of linguis-
tic information.
Cortical Tracking of Linguistic Structure
Low-frequency cortical activity closely tracks the amplitude envelope of the speech signal
(Ahissar et al., 2001; Doelling et al., 2014; Gross et al., 2013; Kayser et al., 2015; Keitel
et al., 2017, 2018; Luo & Poeppel, 2007). Because the low-frequency periodicity of the speech
envelope correlates with the syllable rate (i.e., in the theta band), it has been argued that cor-
tical activity in this frequency range tracks syllable-sized linguistic units (Giraud & Poeppel,
2012; Luo & Poeppel, 2007; Peelle & Davis, 2012; Poeppel & Assaneo, 2020). However,
speech contains temporal regularities at multiple timescales; high-level linguistic units, such
as syntactic phrases, also exhibit quasi-regular temporal structure, yet only a small number of
studies have investigated cortical tracking of phrase structure.
A main method to study tracking of abstract structure has relied on careful control of the
presentation rate of linguistic information, whereby this information is frequency tagged. The
idea behind this approach is that when information is presented repeatedly at a specific fre-
quency, the neural response to that type of information synchronizes with its presentation rate.
In a series of MEG/EEG (magnetoencephalography/electroencephalography) studies, Ding and
colleagues have shown that neural activity becomes phase-locked to the presentation rate of
phrases and sentences, even though these abstract units are not physically discernable in the
Neurobiology of Language
387
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
3
3
8
6
2
0
3
1
3
9
0
n
o
_
a
_
0
0
0
7
0
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cortical tracking of linguistic units
Mutual information:
A measure of the statistical
dependence between two signals.
auditory signal itself (Blanco-Elorrieta et al., 2020; Ding et al., 2016; Ding, Melloni, et al.,
2017; Getz et al., 2018; Makov et al., 2017; Sheng et al., 2019). Such phase-locked responses
are found only if the input can be grouped into phrases, showing that they are based on lin-
guistic knowledge, not acoustic information (Ding et al., 2016; Martin & Doumas, 2017). And
while it has been disputed that what is tracked is really abstract structure rather than lexical
semantics (Frank & Yang, 2018), recent studies have shown that lexical accounts cannot fully
explain the data (Burroughs et al., 2021; Jin et al., 2020).
These frequency-tagging studies are very artificial because they rely on synthesized speech
that is isochronously presented, but similar effects are reported in studies with more naturalistic
materials. In one such study by Keitel et al. (2018), participants listened to naturally spoken sen-
tences that were embedded in noise, after which they had to perform a comprehension task. All
sentences were annotated for the occurrence of phrases, words, and syllables, yielding linguis-
tically relevant frequency bands that were specific for their stimulus materials. Within each fre-
quency band, speech tracking was quantified through mutual information between the speech
envelope and neural activity. At the timescale of words and phrases, tracking was stronger for
correctly comprehended than for incorrectly comprehended sentences, showing that speech
tracking in these frequency bands is related to successful language comprehension.
Using a similar approach, Kaufeld et al. (2020) presented participants with naturally spoken
stimuli in three conditions: regular sentences, jabberwocky sentences (i.e., same prosody and
structure, but different lexical content), and word lists (i.e., same lexical content, but different
structure and prosody). Backward versions of all stimuli were used to control for acoustic dif-
ferences. At the phrasal timescale, speech tracking was stronger for regular sentences than for
both jabberwocky sentences and word lists, while these differences were absent in the acous-
tic control conditions. These findings thus show that the brain is more attuned to phrases when
they contain meaningful information and are therefore relevant for language comprehension
(Kaufeld et al., 2020). In particular, the fact that phrase-level speech tracking is stronger for
sentences than for jabberwocky suggests that this response is modulated by the semantic con-
tent of phrases (see also Brennan & Martin, 2020; Martin, 2020; Martin & Doumas, 2017).
It is still an open question, however, whether semantic content should be interpreted as
lexical-semantic content—the fact that sentences are structured sequences composed of real
words—or rather, compositional-semantic content—the fact that these real words in sentences
compose into meaningful constituents. The most prominent difference between regular and jab-
berwocky sentences is that the former contain real content words, which are replaced by pseu-
dowords in jabberwocky sentences. Real words and pseudowords differ in both semantic and
lexical-syntactic content, with the latter strongly affecting linguistic structure building (e.g.,
Hagoort, 2005, 2017; Matchin & Hickok, 2020). It is thus possible to interpret the difference
in phrase-level speech tracking between sentences and jabberwocky in two ways: Either it
reflects the fact that words in sentences can be composed into meaningful constituents (i.e.,
reflecting the outcome of structure building; Kaufeld et al., 2020), or it reflects the fact that
the lexical-syntactic information carried by content words allows words in sentences to be easily
composed in the first place (i.e., reflecting the input to structure building). In the latter case, these
findings reflect the brain’s attempt to build a structural representation of the linguistic input,
regardless of its interpretation. The present study aims to tease apart these two possibilities.
Background of the Present Study
We contrast regular sentences, whose meaning is compositionally derived from their structure
and lexical components, with stimuli in which the mapping between structure and meaning is
Neurobiology of Language
388
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
3
3
8
6
2
0
3
1
3
9
0
n
o
_
a
_
0
0
0
7
0
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cortical tracking of linguistic units
less transparent. If it is indeed the case that phrase-level speech tracking is driven by the
structure-meaning correspondence of sentences, the tracking response should be stronger
for sentences than for controls that are divergent in their structure-meaning relationship. As
examples of the latter, we used one naturally occurring stimulus (idioms) and one artificial
stimulus (syntactic prose), both of which contain the same structural and lexical-semantic
information as regular (compositional) sentences, but are putatively less compositional in
the sense that their meaning does not derive fully from a combination of their structure and
lexical components. Parametrically reducing the amount of linguistic information, we also
included jabberwocky sentences and unstructured word lists.
We note that compositional processing is not an all-or-none phenomenon (Baggio, 2021;
Titone & Connine, 1999), and idioms and syntactic prose are not processed entirely non-
compositionally. However, a compositional analysis of the sentences in these conditions either
does not yield a sensible interpretation (syntactic prose) or does not yield the intended interpreta-
tion (idioms). We therefore assume that compositional processes will be overall less engaged in the
comprehension of idioms and syntactic prose than in the comprehension of regular sentences.
Idioms are conventionalized co-occurrence restrictions whose figurative meaning must be
learned (Cacciari, 2014; Cacciari & Glucksberg, 1991; Jackendoff, 1995, 2017). They adhere
to basic grammatical rules but are semantically idiosyncratic: The figurative meaning of idioms
is not fully derived from a semantic composition of their component parts (Cacciari & Glucksberg,
1991; Jackendoff, 1995, 2017; Sprenger et al., 2006). As an example, consider the Dutch idiom
een vinger aan de pols houden (literally, “to keep a finger on the wrist”), whose figurative mean-
ing is “to check whether everything goes right.” Clearly, this figurative meaning is non-
compositional and conventionalized, but in terms of structure the idiom is not an unanalyzed
whole. The idiom is a verb phrase whose verb inflects in the past tense in the same way it does in
regular sentences (i.e., as in English, houden “to keep” is irregular, inflecting to hield “kept” in
the past tense), and it has the regular argument structure of the verb houden “to keep,” which is
used ditransitively and can be modified by adverbs in the usual way.
The idea that idioms contain regular syntactic structure is supported by evidence from lan-
guage processing, which shows that the structure of idioms is accessed in both comprehension
and production (Cutting & Bock, 1997; Konopka & Bock, 2009; Peterson et al., 2001; Sprenger
et al., 2006). This structure is linked to the idiom’s meaning in a highly idiosyncratic way, but
language users who process the idiom in real time cannot know this beforehand and will
therefore initially attempt to derive its interpretation compositionally. Behavioral experiments
show that while effects of compositionality can be found in the early stages of idiom compre-
hension, literal processing can to some extent be terminated after the phrase or sentence is
recognized as being an idiom, at which point its idiomatic meaning is retrieved from semantic
memory (Cacciari, 2014; Cacciari & Corradini, 2015; Cacciari & Tabossi, 1988; Holsinger &
Kaiser, 2013; Libben & Titone, 2008; Peterson et al., 2001; though see Smolka et al., 2007).
Evidence from electrophysiological brain recordings also suggests that compositional pro-
cesses can be interrupted in the comprehension of idioms (Canal et al., 2017; Rommers
et al., 2013; Vespignani et al., 2010). We therefore consider idioms suited to serve as exper-
imental sentences whose meaning is not fully derived from their component parts. These
effects of compositionality might not be apparent immediately (i.e., before the idiom recogni-
tion point), but we suspect that compositional processes will be overall less engaged for idioms
than for regular sentences.
In syntactic prose, real words are used to construct syntactically correct but nonsensical
sentences (e.g., Bastiaansen & Hagoort, 2015; Kaan & Swaab, 2002; Marslen-Wilson & Tyler,
Neurobiology of Language
389
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
3
3
8
6
2
0
3
1
3
9
0
n
o
_
a
_
0
0
0
7
0
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cortical tracking of linguistic units
1980; Mazoyer et al., 1993). As an example, consider the Dutch sentence een prestatie zal het
concept naar de mouwen leiden, which translates as “an achievement will lead the concept to
the sleeves.” This sentence adheres to the rules of Dutch syntax, including constraints on word
order and argument structure, but a compositional analysis of the sentence does not yield an
interpretation that makes sense. Not many studies have investigated the brain processes
involved in comprehending syntactic prose, but one relevant study found increased EEG
gamma-band power for regular sentences compared to syntactic prose (Bastiaansen &
Hagoort, 2015). Notably, two other EEG studies reported similar effects in the gamma band
when comparing regular sentences and idioms (Canal et al., 2017; Rommers et al., 2013),
tentatively suggesting that the contrast between sentences and both idioms and syntactic prose
affects similar neurocognitive processes.
In addition to these two conditions, we also used jabberwocky sentences and word lists (see
also Kaufeld et al., 2020). With these five conditions in total (see examples in Table 1), our
design parametrically varies the amount of linguistic information present in the stimuli. All
conditions except jabberwocky sentences contained real content words, and all conditions
except word lists had the same syntactic structure. Moreover, for all syntactically structured
Table 1. Dutch example stimuli of all five conditions
Condition
Sentence
Stimulus
De jongen gaat zijn zusje met haar huiswerk helpen.
Lexical
semantics
X
Syntactic
structure
X
Meaningful compositional
interpretation
X
the boy will his sister with her homework help
“The boy will help his sister with her homework.”
Idiom
De directie zal een vinger aan de pols houden.
X
the directorate will a finger on the wrist keep
Literal: “The directorate will keep a finger on the
wrist.”
Figurative: “The directorate will check whether
everything goes right.”
Syntactic
prose
Een prestatie zal het concept naar de mouwen
X
leiden.
an achievement will the concept to the sleeves lead
“An achievement will lead the concept to the
sleeves.”
Jabberwocky De jormen gaat zijn lumse met haar luisberk
malpen.
the jormen will his lumse with her luisberk malp
“The jormen will malp his lumse with her luisberk.”
Word list
De gaat jongen zusje huiswerk zijn haar helpen met
X
the will boy sister homework his her help with
X
X
X
Note. English translations are provided above. Only the underlined words in the idiom stimulus are part of the conventionalized idiom.
Neurobiology of Language
390
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
3
3
8
6
2
0
3
1
3
9
0
n
o
_
a
_
0
0
0
7
0
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cortical tracking of linguistic units
conditions, a compositional interpretation can be derived. However, a compositional combi-
nation of the words in idioms does not yield their figurative meaning, a compositional com-
bination of the words in syntactic prose does not yield a coherent semantic interpretation,
and a compositional combination of the (pseudo)words in jabberwocky sentences is under-
specified. In other words, regular sentences differ from the other syntactically structured con-
ditions not in whether they allow compositional processing in principle, but in whether a
compositional combination of the structure and the lexical components yields a straightfor-
ward meaningful interpretation.
The Present Study
Participants listened to spoken stimuli in these conditions while their EEG was recorded.
We quantified cortical tracking between the speech envelopes and the EEG data by means
of mutual information (MI), which is an information-theoretic measure that quantifies the
statistical dependence between two random variables (Cogan & Poeppel, 2011; Gross
et al., 2013; Ince et al., 2017; Kayser et al., 2015; Keitel et al., 2017). MI was computed
in three frequency bands, corresponding to the occurrence of phrases (1.1–2.1 Hz), words
(2.3–4.7 Hz), and syllables (3.4–4.9 Hz) in our stimuli (Kaufeld et al., 2020; Keitel et al.,
2018). Following previous research, we controlled for spectral differences between sen-
tences and word lists by including backward versions of both stimuli. These backward ver-
sions preserve many of the spectral properties of their forward version (especially rhythmic
components) but are unintelligible (Gross et al., 2013; Kaufeld et al., 2020; Keitel et al.,
2017; Park et al., 2015).
We were particularly interested in the coherence between speech and EEG in the phrase
frequency band. For this measure of phrase-level speech tracking we consider two possibili-
ties. If it is affected by the extent to which a compositional analysis of the input yields a mean-
ingful structural representation, we expect higher MI for regular sentences than for all other
conditions. Instead, if phrase-level speech tracking reflects the construction of a structural rep-
resentation regardless of its compositional interpretation, we do not expect MI for regular sen-
tences to differ from MI for idioms and syntactic prose. Yet, we do predict MI to be higher for
regular sentences than for jabberwocky and word lists, because the latter two contain less
information based on which a structural representation can be constructed (i.e., cues from
argument structure, word order).
MATERIALS AND METHODS
Participants
We recruited 40 participants (30 female, average age = 24.6 years, age range = 19–31 years)
from the participant pool of the Max Planck Institute for Psycholinguistics. All participants
were right-handed native speakers of Dutch, who reported normal hearing and did not have
a history of language impairment. After receiving information about the experimental proce-
dures, participants gave written informed consent to take part in the experiment, which was
approved by the Ethics Committee of the Faculty of Social Sciences at Radboud University
Nijmegen. They were reimbursed for their participation. After preprocessing, we excluded
two participants due to low numbers of artifact-free trials. The analyses reported are based
on a sample of 38 participants.
Neurobiology of Language
391
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
3
3
8
6
2
0
3
1
3
9
0
n
o
_
a
_
0
0
0
7
0
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cortical tracking of linguistic units
Syntactic prose sentence:
A sentence that is syntactically
correct but semantically incoherent.
Materials
Experimental items
An example of one stimulus item for each condition is given in Table 1. The Sentence condi-
tion contained sentences with compositional meaning, which is derived from a combination of
the word meanings and their structural combination. To give an example of a translated stim-
ulus item, the meaning of The boy will help his sister with her homework is a function of the
meaning of the individual words and the syntactic structure of the sentence. The structure was
the same for all syntactically structured conditions (i.e., Sentence, Idiom, Syntactic prose, and
Jabberwocky), which start with a noun phrase (NP) and an auxiliary verb (e.g., The boy will …),
followed by a verb phrase consisting of an NP, a prepositional phrase (PP), and a non-finite
lexical verb ( V), which is phrase-final in Dutch (e.g., … help his sister with her homework).
For the Idiom condition we selected a set of commonly used and well-known Dutch idioms
that had the same NP-PP-V structure. The majority of these idioms were selected from
stimulus lists shared by Hubers et al. (2018) and Rommers et al. (2013). The idioms were
embedded in carrier sentences by the addition of a sentence-initial NP and an auxiliary verb,
which are not part of the conventionalized structure. We only analyzed those idioms that were
known to the participants. Idiom knowledge was established for each participant by means of
a post-experiment questionnaire (see the section Idiom Knowledge Test). Syntactic prose sen-
tences are grammatically well-formed and contain real words, but these are difficult to com-
pose into a coherent semantic representation. The stimulus sets in both the sentence condition
and the syntactic prose condition were matched with the idioms on the total number of syl-
lables and on the lexical frequency of the content words (frequencies extracted from the
SUBTLEX-NL database of Dutch word frequencies; Keuleers et al., 2010). Jabberwocky sen-
tences were generated with the Wuggy pseudoword generator (Keuleers & Brysbaert, 2010),
which generates pseudowords that obey the phonotactic constraints of Dutch. We created
jabberwocky versions of all items in the sentence condition by substituting each content word
with a pseudoword that was matched in number of syllables, subsyllabic structure, and sylla-
ble transition frequency. The function words (auxiliaries, determiners, prepositions, pronouns)
were kept the same, allowing for the construction of the same syntactic structure with a com-
positional interpretation. Items in the Word list condition contained the same words as those in
the corresponding sentence item, but were scrambled in such a way that no syntactic com-
binations could be formed.
We created 85 stimuli for all conditions, of which the first five served as practice trials,
which were not analyzed. Only the idiom condition had 90 items, which allowed us to pre-
serve roughly the same number of trials as in the other conditions after excluding unknown
idioms.
Audio recordings
The stimuli were recorded in a sound-attenuated booth by a female native speaker of Dutch
(sampling rate = 44.1 kHz (mono), bit depth = 16). After recording, the intensity of all stimuli
was scaled to 70 dB in Praat ( Version 6.1.02; Boersma & Weenink, 2019). Backward stimuli
for the Sentence and Word list conditions were created by reversing each stimulus recording
in Praat.
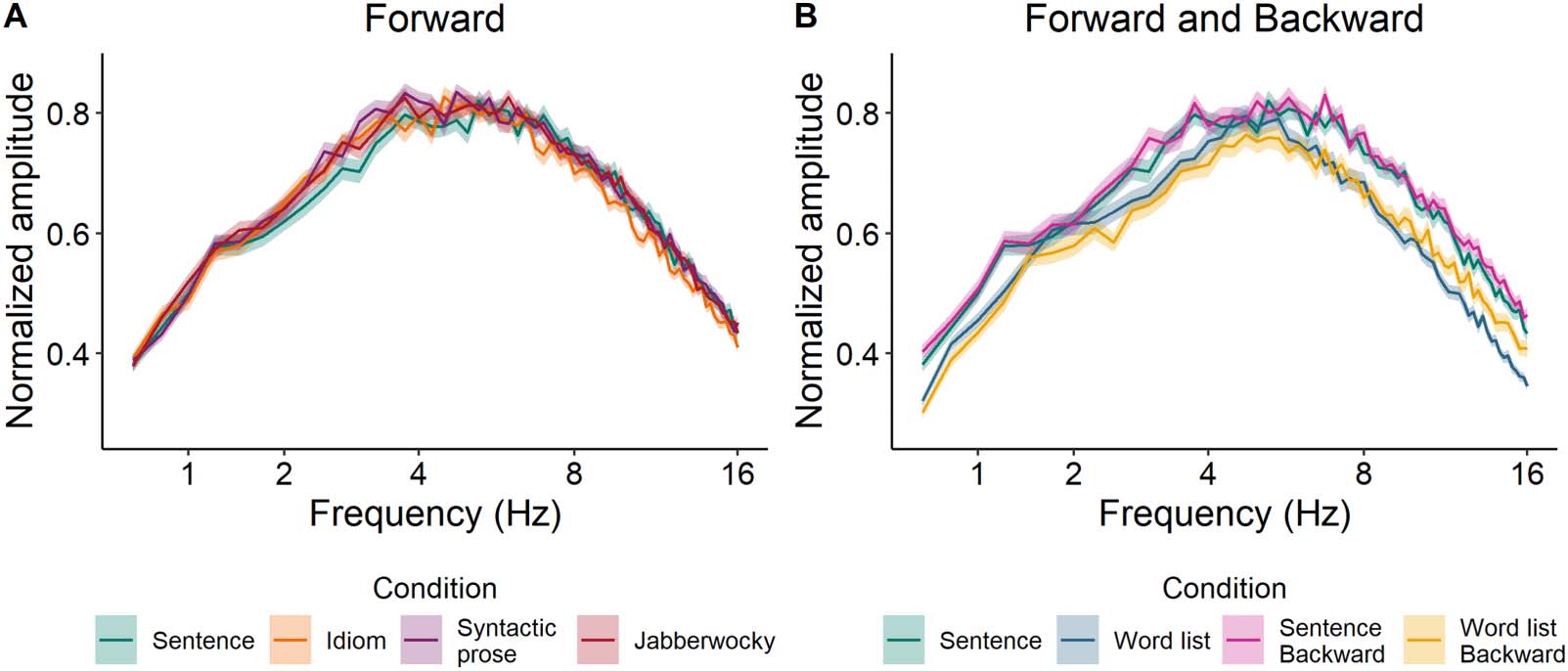
Figure 1 shows the modulation spectra of all Forward conditions as well as the Backward
version of sentences and word lists. These figures indicate that all Forward conditions are pro-
sodically very similar (Figure 1A), except for the word list condition (Figure 1B), which deviates
from the sentence condition at several frequencies. (See Supplementary Information S1.
Neurobiology of Language
392
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
3
3
8
6
2
0
3
1
3
9
0
n
o
_
a
_
0
0
0
7
0
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cortical tracking of linguistic units
Figure 1. Modulation spectra of the forward versions of all conditions, computed following the procedure described in Ding, Patel, et al.
(2017). Backward versions of sentences and word lists were included because of the differences between the forward versions of these
two conditions.
Supporting Information can be found at https://doi.org/10.1162/nol_a_00070.) While not
ideal, we believe that this prosodic difference between stimuli with regular syntactic structure
and those without structure is inherent in the contrast between these conditions. Because our
main interest is the comparison between the syntactically structured conditions (Sentence vs.
Idiom, Syntactic prose, Jabberwocky), it is important that these conditions do not systemati-
cally differ in acoustic properties.
Annotations
We manually annotated the forward recordings in Praat (Boersma & Weenink, 2019) with
respect to the presence of phrases, words, and syllables. Specifically, for each stimulus we
annotated the position in the recording where a linguistic unit ends (Figure 2A). For both
Figure 2. Three different annotations of linguistic structure for the Dutch translation of the sentence the boy will help his sister with her
homework. (A) Schematic illustration of the three different timescales of the linguistic units of information (phrases, words, and syllables)
contained in the sentence. From the annotation of these timescales, we derived frequency bands for each linguistic unit. (B) Phrase-level anno-
tation, where words that integrate a phrase are coded as 1 for their entire duration, while all other words are coded as 0 (bracket presence). (C)
Phrase-level annotation, where the value assigned to each word corresponds to the number of phrases that the word integrates (bracket count).
Neurobiology of Language
393
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
3
3
8
6
2
0
3
1
3
9
0
n
o
_
a
_
0
0
0
7
0
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cortical tracking of linguistic units
words and syllables, this corresponds to the boundary between successive units. For
phrases this corresponds to the position of closing phrase boundaries. For example, in
[de jongen] [gaat [zijn zusje] [met haar huiswerk] helpen] the closing bracket denotes
the offset of a phrase whose onset is denoted by the corresponding opening bracket. As
word lists by definition do not contain phrases, we marked “phrases” in these stimuli by
annotating the offsets of the words that are at positions of closing phrase boundaries in the
In the example sentence above, phrases are closed after
corresponding sentence item.
the second, fifth, eighth, and ninth word, leading to the following phrase annotation for the
corresponding word list: [de gaat] [jongen [zusje huiswerk] [zijn haar helpen] met]. Con-
verting the onsets and offsets of these annotations to frequencies resulted in the following
frequency bands: 1.1–2.1 Hz for phrases, 2.3–4.7 Hz for words, and 3.4–4.9 Hz for
syllables.
To provide additional evidence that our results index cortical tracking of abstract (syntactic)
information, rather than mere acoustic differences between the conditions, we performed an
additional MI analysis in which the speech stimuli were replaced by abstract versions of these
stimuli in which we only encoded phrase-structure information (Brodbeck, Presacco, & Simon,
2018; Kaufeld et al., 2020). For each forward stimulus, we marked all time points correspond-
ing to phrase-final words with a 1 and marked all other time points with a 0 (bracket presence;
Figure 2B). Phrase-final words are those words at which syntactic and/or semantic composition
can take place. For example, the time points corresponding to the underlined words in the
sentence [de jongen] [gaat [zijn zusje] [met haar huiswerk] helpen] (i.e., boy, sister, homework,
help) were marked by a 1, because they close syntactic phrases, while all other time points
were marked by a 0. Again, annotating phrase-final words in word lists is impossible, so we
marked phrases in the same way as described above, marking time points corresponding to
words with a 1 if these words are in a position that indexes a phrase-final word in the corre-
sponding sentence.
These abstract annotations of bracket presence are actually insufficient to represent phrase
structure, because sentences are hierarchically embedded structures rather than linearly
concatenated phrases. To represent this property, we incorporated bracket count as yet
another type of abstract annotation (Brennan & Martin, 2020; Brennan et al., 2012; Brennan
et al., 2016; Nelson et al., 2017), which is correlated with bracket presence but contains
more detailed syntactic information. This variable counts the number of phrases that are
completed at a particular word (derived from bottom-up tree traversal), corresponding to the
closing brackets in [[de jongen] [gaat [[zijn zusje] [met [haar huiswerk]] helpen]]]. The value
assigned to each word for its entire duration corresponds to the number of phrases that the word
integrates (Figure 2C).
Experimental Design
Participants listened to all stimuli in all seven conditions, which were presented in a block
design. The order in which the seven blocks were presented was pseudo-randomized, with
the following constraints: The two backward conditions were never presented in adjacent
blocks, and the block with word lists and the block with idioms always preceded the block
with sentences. Regarding the word lists, this presentation order was used to reduce the pos-
sibility that participants would project (their memory of ) the phrase structure of the sentences
onto the word lists. Regarding the idioms, this order was used to reduce the possibility that
participants would try to derive a compositional analysis of their meaning. Within each block,
the order of the items was randomized.
Neurobiology of Language
394
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
3
3
8
6
2
0
3
1
3
9
0
n
o
_
a
_
0
0
0
7
0
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cortical tracking of linguistic units
Procedure
Participants were individually tested in a soundproof booth. They were instructed to attentively
listen to the audio, which was presented over loudspeakers, while looking at a fixation cross
displayed at the center of the screen. After each trial, participants had to advance to the next
trial by pressing a button. They were allowed to take short breaks between blocks. The EEG
experiment lasted approximately 60–70 min and was followed by an idiom knowledge test.
Idiom Knowledge Test
The EEG experiment was followed by a digital questionnaire in which participants were asked
to indicate whether they knew the figurative meaning of the idioms that were presented in the
experiment. For each idiom, they had to indicate this on a keyboard. If they answered “yes,”
they had to type the meaning using the keyboard. If they answered “no,” they were asked to
indicate what they thought the meaning could be. Idioms were rated as known when the par-
ticipant answered “yes” and gave a correct description of the meaning of the idiom. For each
participant, we included only idioms rated as known into subsequent analyses. On average,
participants knew 78 of the 90 idioms (86.7%, range = 64–90).
Speech Preprocessing
The speech envelope is the acoustic power of the speech signal at a given time in a given
frequency range. Here, we estimated the broadband speech envelope by averaging across
all ranges, following the procedure described in Chandrasekaran et al. (2009) and adopted
by subsequent studies (Gross et al., 2013; Kaufeld et al., 2020; Kayser et al., 2015; Keitel
et al., 2017). Using the Chimera toolbox (Smith et al., 2002), we band-pass filtered the audi-
tory signal into 8 frequency bands between 100–8000 Hz (third-order Butterworth filter, for-
ward and reverse), such that the bands spanned equal widths on the cochlear frequency map
(1.i in Figure 3). The cutoff frequencies of the bands (in Hz) were: 100, 228, 429, 743, 1233,
2000, 3198, 5071, and 8000. We computed the Hilbert transform of the signal in each of these
frequency bands and took the absolute value as an estimate of the narrowband envelope (1.ii
in Figure 3). We downsampled each narrowband speech envelope to 150 Hz, and averaged
across all 8 bands to derive the broadband speech envelope (1.iii in Figure 3).
EEG Recording and Preprocessing
The EEG was recorded using an MPI custom actiCAP 64-electrode montage (Brain Products,
Munich, Germany), of which 59 electrodes were mounted in the electrode cap (see
Supplementary Information S2 for electrode layout). Eye blinks were registered by one elec-
trode below the left eye, and eye movements were registered by two electrodes, placed on
the outer canthi of both eyes. One electrode was placed on the right mastoid, the reference
electrode was placed on the left mastoid and the ground was placed on the forehead. The
EEG signal was amplified through BrainAmp DC amplifiers and referenced online to the left
mastoid. The data were acquired at a sampling rate of 500 Hz, using a band-pass filter of
0.016–249 Hz.
Preprocessing was performed using the Fieldtrip toolbox (Oostenveld et al., 2011) in Matlab
( Version 2016a). If channels were broken or showed heavy drifts, they were replaced by a
weighted average of their neighbors. The data were then low-pass filtered at 50 Hz (36db/
oct), re-referenced to the average of all electrodes and segmented into epochs ranging from
the onset to the offset of the audio recording. We manually rejected trials that contained
Neurobiology of Language
395
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
3
3
8
6
2
0
3
1
3
9
0
n
o
_
a
_
0
0
0
7
0
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cortical tracking of linguistic units
Figure 3. Visual representation of the analysis pipeline.
(movement) artifacts and trials in which an unknown idiom was presented (on a by-idiom, by-
participant basis; based on the post-experiment questionnaire). We used independent compo-
nent analysis (ICA; using ICA weights from a version of the data which was downsampled to
300 Hz and high-pass filtered at 1 Hz) to filter artifacts resulting from eye movements and
steady muscle activity. Last, we automatically rejected epochs in which the difference
between the maximum and minimum voltage exceeded 150 μV. In total, we excluded 9.2%
of the data (range of averages across conditions = 6.4%–12.1%). Each EEG segment was
downsampled to 150 Hz to match the sampling rate of the speech envelopes. The prepro-
cessed data were then subjected to mutual information analysis.
Mutual Information Analysis
To quantify cortical speech tracking in each frequency band, we computed MI between the
band-limited Hilbert representations of the broadband speech envelope and the EEG signal
(see Figure 3). In our experiment, MI measures the average reduction in uncertainty about
the EEG signal given that the speech envelope (or annotation of syntax) is known, and can
thus be used as a measure of the relatedness of the two signals (Ince et al., 2017). We followed
the procedure described in Kaufeld et al. (2020), which involved the following steps for speech
signals and EEG trials separately: First, each signal was band-pass filtered in the frequency
bands of interest (Figure 3, 2.i), using third-order Butterworth filters (forward and reverse).
We then extracted the complex components from each filtered signal using a Hilbert transform
(Figure 3, 2.ii), whose real and imaginary parts were normalized separately using the copula
normalization method developed by Ince et al. (2017). We derived instantaneous phase and
power and concatenated the resulting signals from all trials (Figure 3, 2.iii). MI was computed
for each electrode, participant, and condition separately, in the following way:
MI(EEG;Speech) = H(EEG) + H(Speech) − H(EEG,Speech)
Here, H(EEG) is the entropy of the (Hilbert representation of the) EEG signal, H(Speech) the
entropy of the (Hilbert representation of the) broadband speech envelope, and H(EEG, Speech)
Neurobiology of Language
396
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
3
3
8
6
2
0
3
1
3
9
0
n
o
_
a
_
0
0
0
7
0
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cortical tracking of linguistic units
their joint entropy. To accommodate speech-brain lag, we computed MI at five different lags,
ranging from 60 to 140 ms, in steps of 20 ms. Statistical analysis was done on the average MI
across all five lags.
The same steps were taken for the abstract stimuli, except that the band-pass filter was
applied in the phrase frequency band only. MI was computed between the Hilbert represen-
tations of the abstract stimuli and the EEG signals corresponding to all forward conditions. For
clarification, we use the term speech tracking to refer to MI computed between EEG and the
speech envelopes, and syntax tracking to refer to MI computed between EEG and the abstract
annotations of syntax.
Statistical Analysis of MI Values
We fitted linear mixed-effects models (Baayen et al., 2008) to the log-transformed, trimmed
(2.5% at both tails of the distribution of each condition) MI values in each frequency band
and in a centroparietal cluster of electrodes (electrodes 1, 3, 4, 5, 8, 9, 10, 11, 28, 29, 30,
33, 35, 36, 37, 40, 41, 42, 43, based on Kaufeld et al., 2020; see Supplementary
Information S2 for electrode layout) using lme4 (Bates et al., 2015) in R (R Core Team,
2021). In each frequency band we ran two separate models for the MI analysis between
EEG and speech. The first model compared MI for Sentences to MI for Idioms, Syntactic prose,
and Jabberwocky. This model contained the four-level factor Construction as fixed effect,
which was treatment-coded with Sentence as the reference level. Participant was added as
a random effect, which had a random intercept and Construction as random slope. Because
we had backward versions of Sentences and Word lists, we compared Sentences to Word lists
in a two-by-two analysis. This involved a second model with Structure (Sentence vs. Word list),
Direction (Forward vs. Backward), and their interaction as fixed effects. Structure and Direc-
tion were deviation coded (−0.5, 0.5), and participant was added as random effect, with a
random intercept and the interaction between Structure and Direction as random slope. This
second model evaluates whether the MI difference between Sentences and Word lists in the
forward version is different from the same difference in the backward version.
For the MI analyses in which the speech envelope was replaced by abstract annotations, we
ran a model with the five-level factor Construction (Sentence, Idiom, Syntactic prose, Jabber-
wocky, Word list) as fixed effect. This model compared MI for Sentences to MI for Idioms,
Syntactic prose, Jabberwocky, and Word lists. Construction was again treatment-coded with
Sentence as the reference level. Participant was added as a random effect, which had a ran-
dom intercept and Construction as random slope. In all analyses we evaluated whether adding
a fixed effect increased predictive accuracy by comparing a model with that fixed effect to a
model without that fixed effect using R’s anova() function.
ERP Preprocessing and Analysis
To evaluate whether the different forward conditions were processed as intended, we com-
pared the event-related potentials (ERPs) elicited by the sentence-final lexical verb in all syn-
tactically structured conditions (i.e., Sentence, Idiom, Syntactic prose, and Jabberwocky).
Word lists were not included because the lexical verbs in word lists were not sentence-final
(see Table 1), due to which the ERP windows segmented around these verbs also contained
activity evoked by the subsequent word. We were specifically interested in the N400, a
negative-going ERP component that peaks between 300 and 500 ms after the onset of each
content word and is sensitive to predictability and semantic congruency (Baggio & Hagoort,
2011; Kutas & Federmeier, 2011).
Neurobiology of Language
397
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
3
3
8
6
2
0
3
1
3
9
0
n
o
_
a
_
0
0
0
7
0
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cortical tracking of linguistic units
Because the segments corresponding to the N400 for these sentence-final verbs lasted
beyond the offset of the audio recordings, they were not captured in the segments we used
for MI analysis. We therefore used a separate preprocessing pipeline for the ERP analysis, in
which the data were low-pass filtered at 40 Hz (36db/oct), re-referenced to the average of the
left and right mastoid, and segmented into epochs ranging from −250 to 1,500 ms relative to
the onset of the sentence-final verb in each audio recording. All other preprocessing steps
were identical to those reported in the section EEG Recording and Preprocessing. In total,
we excluded 4.8% of the data (range of averages across conditions = 4.1%–5.3%). Before sta-
tistical analysis, the EEG data were baseline-corrected using a 250 ms baseline window pre-
ceding the sentence-final verb.
For the N400 region of interest, we calculated the voltage in the centroposterior electrodes
3, 8, 9, 15, 27, 28, 35, 40, 41, 47 in a 300–500 ms time window after the onset of the
sentence-final word, for each trial and each participant (based on Coopmans & Nieuwland,
2020, see Supplementary Information S2 for electrode layout). These voltage values were
compared via a linear mixed-effects analysis (Baayen et al., 2008) in R (R Core Team,
2021). The mixed-effects model contained Construction as fixed effect, which was
treatment-coded with Sentence as the reference level to which the conditions Idiom, Syntactic
prose, and Jabberwocky were individually compared. We included participant as random
effect, which had a random intercept and Construction as random slope. The models with
and without Construction were compared with R’s anova() function.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
3
3
8
6
2
0
3
1
3
9
0
n
o
_
a
_
0
0
0
7
0
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
RESULTS
Speech Tracking
In the phrase frequency band, we ran two separate mixed-effects models. The first model eval-
uates whether MI is modulated by the type of Construction that was presented, comparing
Sentences to the other syntactically structured conditions (Idioms, Syntactic prose, and Jabber-
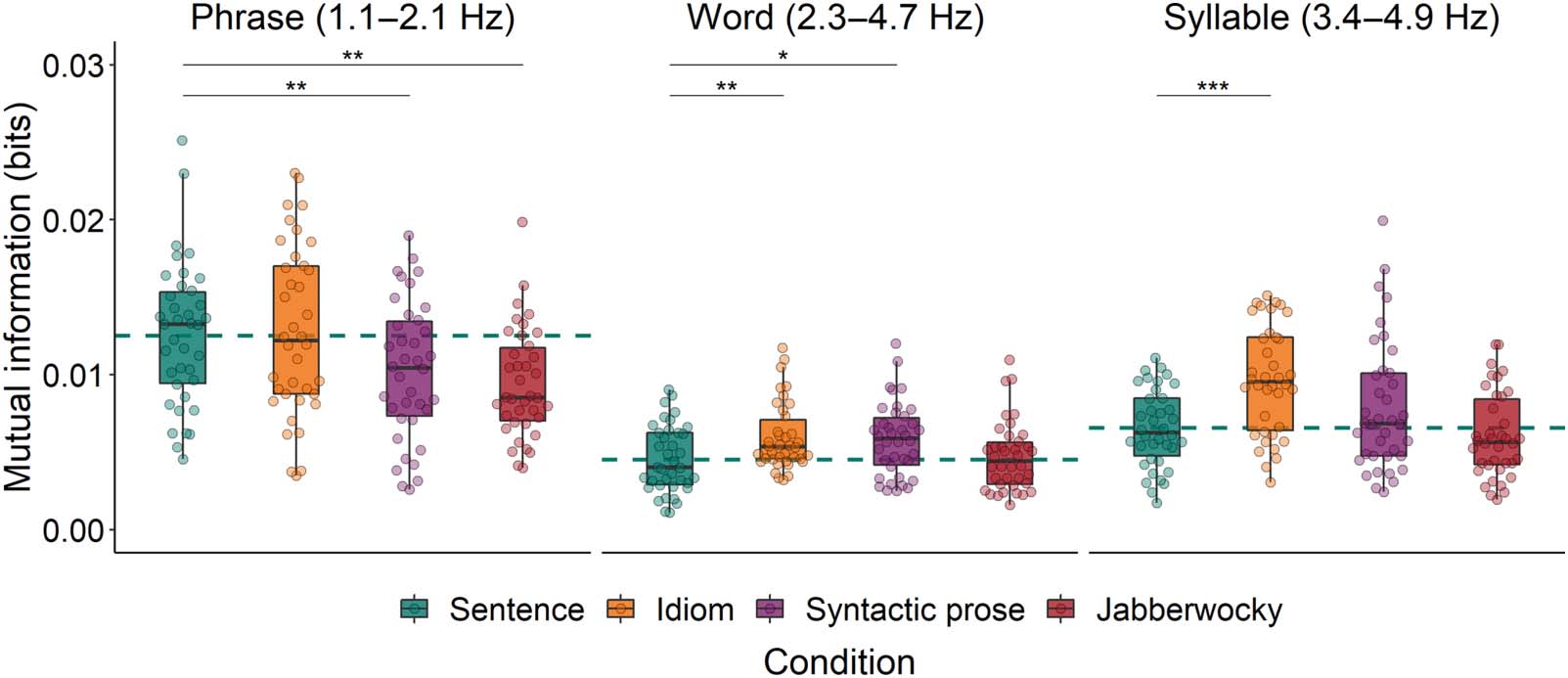
wocky). Model comparison showed that Construction predicted MI (χ2 = 15.30, p = 0.002; see
left panel of Figure 4). Specifically, MI was higher for Sentences than for both Jabberwocky and
Figure 4. Mutual information between EEG and the speech envelopes of all syntactically structured conditions in the phrase, word, and syllable
frequency bands. Drops reflect average per participant. The dashed horizontal line reflects the average of the Sentence condition. *p < 0.05,
**p < 0.01, ***p < 0.001.
Neurobiology of Language
398
Cortical tracking of linguistic units
Table 2.
Sentences to speech tracking for Idioms, Syntactic prose, and Jabberwocky
Fixed effects of the models that compare speech tracking (i.e., speech-brain MI) for
Estimate
SE
df
t value
p value
Phrase frequency band
Intercept
Sentence-Idiom
Sentence-Syntactic prose
Sentence-Jabberwocky
Word frequency band
Intercept
Sentence-Idiom
Sentence-Syntactic prose
Sentence-Jabberwocky
Syllable frequency band
Intercept
Sentence-Idiom
Sentence-Syntactic prose
Sentence-Jabberwocky
−4.77
−0.05
−0.30
−0.38
−5.82
0.31
0.26
0.02
−5.44
0.40
0.12
−0.11
0.07
0.10
0.10
0.11
0.09
0.10
0.10
0.13
0.08
0.09
0.10
0.08
37.5
38.0
38.0
38.0
37.6
37.9
38.0
38.0
37.5
38.3
38.0
38.0
−70.68
<0.001
−0.52
−2.88
−3.36
0.61
0.007
0.002
−65.05
<0.001
3.08
2.61
0.15
−69.83
4.58
1.15
−1.39
0.004
0.013
0.88
<0.001
<0.001
0.26
0.17
Note. The estimates are from three different models, corresponding to the phrase, word, and syllable frequency
bands. SE = standard error; df = degrees of freedom.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
3
3
8
6
2
0
3
1
3
9
0
n
o
_
a
_
0
0
0
7
0
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Syntactic prose, but not different from MI for Idioms (see Table 2 for the estimates of the fixed
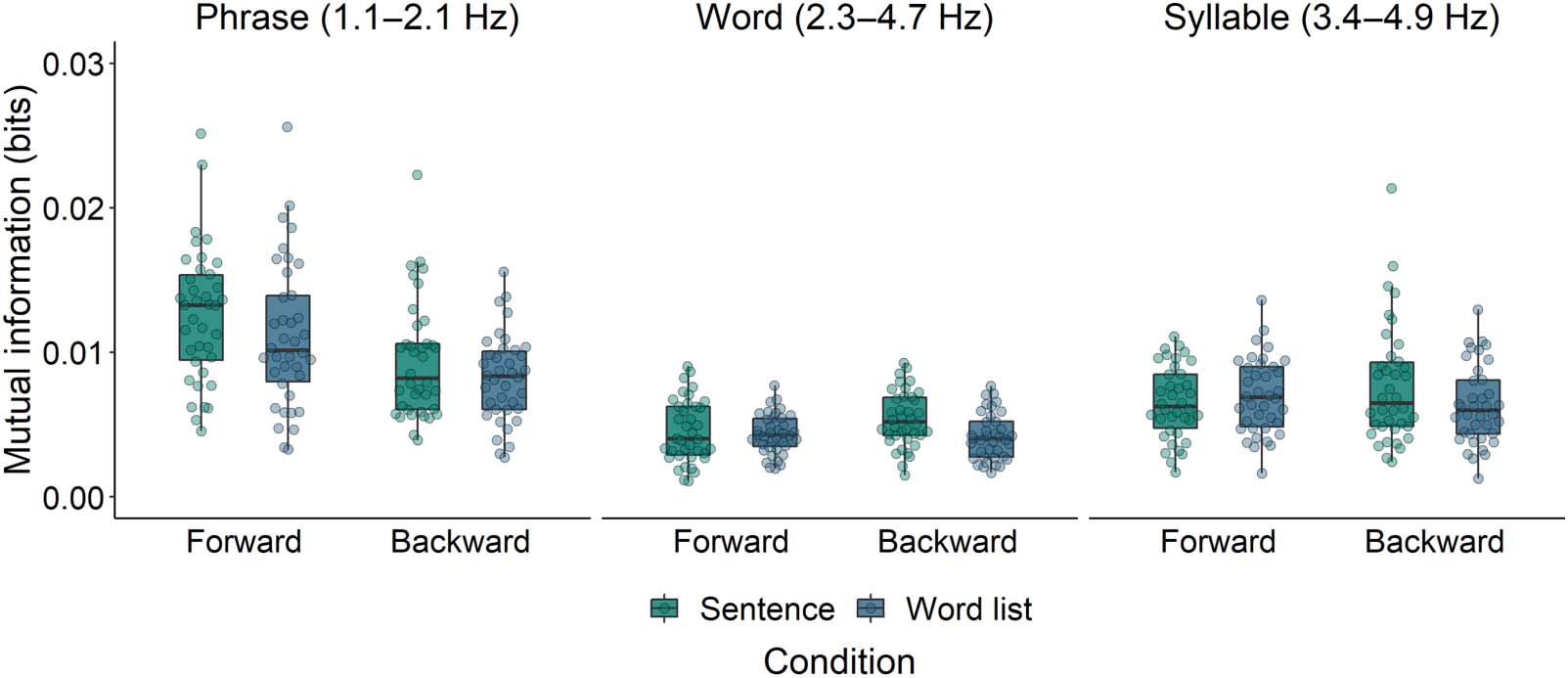
effects). The second model evaluated the interaction between Structure (Sentence vs. Word
list) and Direction (Forward vs. Backward). Model comparison revealed that Sentences elicited
higher MI than Word lists (χ2 = 4.19, p = 0.041; see left panel of Figure 5), and that Forward
stimuli elicited higher MI than Backward stimuli (χ2 = 14.37, p < 0.001). The interaction was
not significant (χ2 = 0.72, p = 0.40), which means that the difference between Sentences and
Word lists was not solely driven by the linguistic differences between their forward versions
and thus (at least partially) also reflects differences in acoustics. The estimates of the fixed
effects of this second model are presented in Table 3.
In the word frequency band, the first model showed that Construction predicted MI (χ2 =
11.71, p = 0.008; see middle panel of Figure 4), but this effect was not driven by the same
contrasts as the effect in the phrase frequency band. That is, MI was lower for Sentences than
for both Idioms and Syntactic prose, but not different from MI for Jabberwocky (Table 2). The
second model showed a marginal difference between Sentences and Word lists (χ2 = 3.70, p =
0.054; see middle panel of Figure 5), and no difference between Forward and Backward
stimuli (χ2 = 0.33, p = 0.56). The interaction between Structure and Direction was not signif-
icant (χ2 = 2.95, p = 0.086; Table 3).
In the syllable frequency band, the first model again showed that Construction predicted MI
(χ2 = 22.15, p < 0.001; see right panel of Figure 4). MI was lower for Sentences than for Idioms,
Neurobiology of Language
399
Cortical tracking of linguistic units
Figure 5. Mutual information between EEG and the speech envelopes of both forward and backward versions of Sentences and Word lists in
the phrase, word, and syllable frequency bands. Drops reflect average per participant.
Table 3.
vs. Word list) and Direction (Forward vs. Backward) on speech tracking (i.e., speech-brain MI)
Fixed effects of the interaction models, which evaluate the effects of Structure (Sentence
Estimate
SE
df
t value
p value
Phrase frequency band
Intercept
Structure
Direction
Structure*Direction
Word frequency band
Intercept
Structure
Direction
Structure*Direction
Syllable frequency band
Intercept
Structure
Direction
Structure*Direction
−5.00
−0.15
−0.28
−0.12
−5.77
−0.11
0.09
0.25
−5.43
−0.13
0.03
0.23
0.04
0.07
0.07
0.14
0.04
0.06
0.08
0.14
0.04
0.09
0.08
0.15
37.8
38.1
38.0
38.2
37.6
38.2
37.9
38.2
38.0
37.8
37.6
38.1
−123.55
−2.07
−4.10
−0.85
−161.66
−1.82
1.08
1.75
−135.37
−1.50
0.45
1.55
<0.001
0.046
<0.001
0.40
<0.001
0.077
0.29
0.088
<0.001
0.14
0.66
0.13
Note. The estimates are from three different models, corresponding to the phrase, word, and syllable frequency
bands. SE = standard error; df = degrees of freedom.
but not different from MI for Syntactic Prose and Jabberwocky (Table 2). The second model
showed no difference between Sentences and Word lists (χ2 = 0.98, p = 0.32; see right panel of
Figure 5), nor between Forward and Backward stimuli (χ2 = 0.10, p = 0.76). The interaction
between Structure and Direction was also not significant (χ2 = 2.33, p = 0.13; Table 3).
Neurobiology of Language
400
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
3
3
8
6
2
0
3
1
3
9
0
n
o
_
a
_
0
0
0
7
0
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cortical tracking of linguistic units
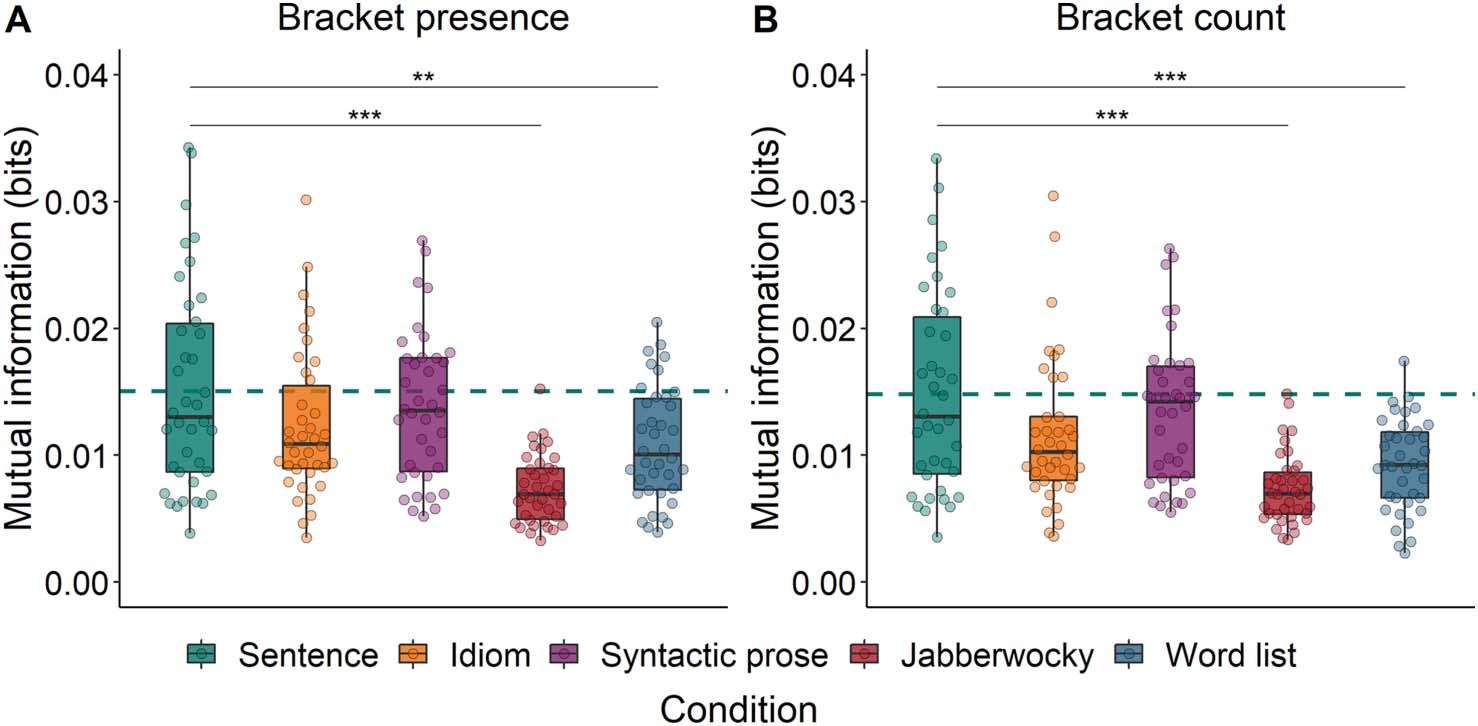
Syntax Tracking
We then evaluated whether Construction (i.e., Sentence, Idiom, Syntactic prose, Jabberwocky,
and Word list) predicted MI between the EEG signal and the abstract annotations of syntactic
structure. When these annotations reflected bracket presence, Construction indeed predicted
MI (χ2 = 35.98, p < 0.001; Figure 6A). MI was higher for Sentences than for both Jabberwocky
and Word lists, but not different from MI for Idioms or Syntactic prose (see Table 4).
Figure 6. Mutual information between EEG and abstract annotations (bracket presence (A) and
bracket count (B)) in the phrase frequency band. Drops reflect average per participant. The dashed
horizontal line reflects the average of the Sentence condition. **p < 0.01, ***p < 0.001.
Table 4.
Sentences to syntax tracking for Idioms, Syntactic prose, Jabberwocky, and Word lists
Fixed effects of the models that compare syntax tracking (i.e., annotation-brain MI) for
Estimate
SE
df
t value
p value
Bracket presence
Intercept
Sentence-Idiom
Sentence-Syntactic prose
Sentence-Jabberwocky
Sentence-Word list
Bracket count
Intercept
Sentence-Idiom
Sentence-Syntactic prose
Sentence-Jabberwocky
Sentence-Word list
−4.62
−0.19
−0.12
−0.74
−0.35
−4.64
−0.24
−0.13
−0.71
−0.48
0.10
0.13
0.10
0.13
0.12
0.10
0.13
0.11
0.13
0.11
37.6
37.5
38.3
37.8
37.9
37.6
37.6
38.1
37.8
38.1
−47.17
<0.001
−1.51
−1.23
−5.68
−3.02
0.14
0.23
<0.001
0.005
−46.85
<0.001
−1.83
−1.23
−5.30
−4.25
0.076
0.23
<0.001
<0.001
Note. The estimates are from two different models, corresponding to annotations reflecting respectively bracket
presence and bracket count. SE = standard error; df = degrees of freedom.
Neurobiology of Language
401
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
3
3
8
6
2
0
3
1
3
9
0
n
o
_
a
_
0
0
0
7
0
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cortical tracking of linguistic units
The same pattern of results was found for the analysis of MI between the EEG signal and the
annotations of bracket count, which differed across Constructions (χ2 = 34.12, p < 0.001;
Figure 6B). MI was higher for Sentences than for Jabberwocky as well as for Word lists, but
not different from MI for Idioms or Syntactic prose (see Table 4).
Overall, both analyses show that, at the frequency band corresponding to abstract phrase
structure, the brain tracks the structure of sentences more strongly than the structure of both
jabberwocky and word lists. It is interesting to note that the pattern of results is very similar for
bracket count and bracket presence, suggesting that the more detailed syntactic information
contained in the bracket count annotations does not add predictive accuracy with respect to
phrase tracking (contrary to previous work, e.g., Brennan et al., 2016).
An anonymous reviewer rightly noted that the sentences in our conditions differ in co-
occurrence frequency, with syntactic prose and jabberwocky sentences having lower transi-
tional probabilities than regular sentences and idioms. We do not think this difference can
account for our phrase-level effects, because it would predict a pattern of results that is dif-
ferent from what we found. First, it would predict no differences between syntactic prose
and jabberwocky sentences, because they have similarly low transitional probabilities.
Yet, these two conditions do elicit differences in cortical tracking of phrase structure. To test
this, we repeated our linear mixed-effects analysis in the phrase frequency band, but with
Syntactic prose as the reference level for the four-level factor Construction. When MI is com-
puted between EEG and the abstract syntactic annotations, it is higher for Syntactic prose
than for Jabberwocky, both when the annotations reflect bracket presence (β = −0.57, SE =
0.09, t = −6.32, p < 0.001; Figure 6A) and when they reflect bracket count (β = −0.62, SE =
0.09, t = −6.55, p < 0.001; Figure 6B). There were no differences between Syntactic prose
and Jabberwocky in terms of speech tracking (β = −0.08, SE = 0.13, t = −0.59, p < 0.001). Sec-
ond, it would predict differences between idioms and regular sentences, because the words in
idioms are part of a fixed expression and therefore have high transitional probabilities.
However, no such differences between sentences and idioms were found at the phrase level
in either speech tracking (see the section Speech Tracking) or syntax tracking.
ERPs to Sentence-Final Verb
The MI analyses showed no consistent differences in phrase tracking between Sentences and
Idioms, whereas the difference between Sentences and Syntactic Prose was inconclusive (i.e.,
difference in speech tracking but no difference in syntax tracking). This absence of expected
differences might indicate either that the brain does not track the syntactic structure of these
stimuli differently (i.e., the conditions are perceived as being different, but this does not affect
phrase tracking), or that the conditions were not processed as being very different). To evaluate
the latter possibility, we compared the ERPs elicited by the sentence-final lexical verb in all
syntactically structured conditions. The results show that the stimuli in the different conditions
were processed as expected.
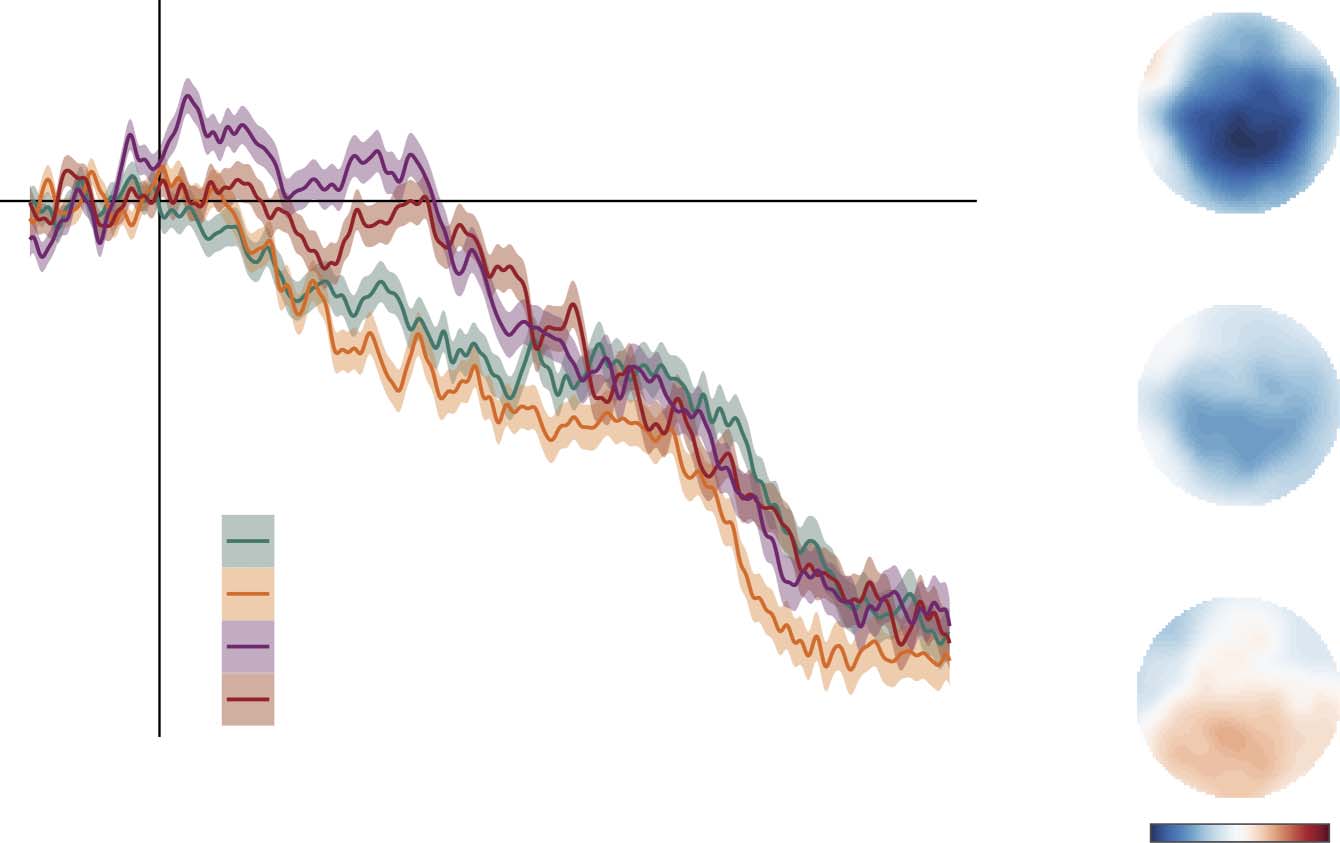
As indicated by the sentence-final ERPs in Figure 7A, the variable Construction was asso-
ciated with modulations of activity in the N400 region of interest (χ2 = 45.9, p < 0.001). The
ERP elicited by sentence-final verbs in Sentences was less negative than the ERP elicited by
sentence-final verbs in both Syntactic prose (β = −1.56, SE = 0.25, t = −6.37, p < 0.001) and
Jabberwocky (β = −0.89, SE = 0.25, t = −3.51, p < 0.001), but more negative than the ERP
elicited by sentence-final verbs in Idioms (β = 0.60, SE = 0.27, t = 2.26, p = 0.029). Note that
the effects seem to start quite early, in particular for Syntactic prose (Figure 7A). This might
have to do with the fact that the words preceding the verb in those stimuli are semantically
Neurobiology of Language
402
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
3
3
8
6
2
0
3
1
3
9
0
n
o
_
a
_
0
0
0
7
0
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cortical tracking of linguistic units
(A) Grand-average ERPs at the centroposterior cluster of electrodes, time-locked to the onset of the sentence-final verb in the four
Figure 7.
syntactically structured conditions. Negative voltage is plotted upwards, and color-shaded areas show the within-subjects standard error of the
mean per time sample. (B) Topographical plots of the voltage differences between conditions in the 300–500 ms time window of interest.
odd (and thus elicit a strong N400), and with differences between conditions in the pre-verb
parts in general. Figure 7B contains the topographical plots of the voltage differences in the
300–500 ms time window of interest.
DISCUSSION
In this EEG study with naturally spoken stimuli, we investigated whether cortical tracking of
phrase structure is modulated by the degree to which this structure is meaningful. Participants
were presented with stimuli that contained different degrees of structural meaning. We mea-
sured tracking by computing mutual information between the EEG data and either the speech
envelopes (speech tracking) or abstract annotations of syntax (syntax tracking). Both signals
were filtered in the frequency band corresponding to the occurrence of phrases. These anal-
yses showed overall stronger tracking of phrases in regular sentences than in stimuli with
reduced lexical-syntactic content ( jabberwocky) or without syntactic structure (word lists),
but no consistent differences in phrase-level tracking between sentences and divergent stimuli
that contained a combination of both structure and lexical meaning (idioms, syntactic prose).
As analyses of sentence-final ERPs showed clear differences between the conditions in terms of
their sentence-level meaning, we take these findings to suggest that cortical tracking of linguis-
tic structure reflects the internal generation of that structure, whether it transparently maps
onto semantic meaning or not.
Effects of Composition in Processing Idioms and Syntactic Prose
We contrasted regular sentences to two semi-compositional conditions: Idioms and syntactic
prose. We reasoned that compositional processes would be less engaged during the compre-
hension of idioms and syntactic prose (Canal et al., 2017; Peterson et al., 2001; Rommers
et al., 2013; Vespignani et al., 2010), though there are several factors that likely influence
the extent to which participants will try to derive a compositional interpretation from these
Neurobiology of Language
403
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
3
3
8
6
2
0
3
1
3
9
0
n
o
_
a
_
0
0
0
7
0
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cortical tracking of linguistic units
sentences. Theories of idiom comprehension assume that before the idiom is recognized as
being an idiomatic construction, standard literal processing is engaged (see Cacciari &
Tabossi, 1988; Libben & Titone, 2008; Sprenger et al., 2006; Titone & Connine, 1999). As
the idioms in our experiment were embedded in neutral carrier sentences, they could not
be predicted from context, so the idiom recognition point might occur late. Sentence-final
ERPs indicated that this was not too late to affect online comprehension: The N400 for
sentence-final verbs was less negative in idioms than in regular sentences, suggesting that
the idiom was recognized and retrieved before sentence offset. Nevertheless, any effects of
compositionality are likely restricted to late time points, reducing the overall effect of compo-
sitionality on cortical tracking on phrase structure.
In addition, there are several differences within the group of idioms that might affect com-
positional processing. Many of the idiomatic constructions have variable slots that can be
filled by compositional information (Jackendoff, 2017). The initial noun phrase in the idiom
NP door de vingers zien (“to condone” NP), for instance, must be interpreted literally and will
likely receive a compositional analysis. Relatedly, even in idioms that do not take variables,
the literal meanings of the words are sometimes part of the idiom’s figurative meaning. For
example, the idiom de regels aan je laars lappen, which literally means “to patch the rules
to your boot” (figuratively: “to ignore the rules”) is actually about rules (though not about
boots), so the NP de regels “the rules” will be processed literally. A question for future research
is whether this variation within the class of idioms affects cortical tracking. We could not inves-
tigate this possibility, as it requires individual-item analyses that were impossible given the way
MI was computed. Yet it would be interesting to see whether cortical tracking of linguistic units
is affected by idiomatic variation, such as the transparency, decomposability, and syntactic
flexibility of the idiom (see Cacciari, 2014; Cacciari & Glucksberg, 1991; Gibbs et al.,
1989; Libben & Titone, 2008).
At the phrase timescale, we did not find a consistent difference between regular sentences
and syntactic prose. While speech tracking was stronger for regular sentences than for syntac-
tic prose (left panel of Figure 4), this difference was absent in measures of syntax tracking
(Figure 6). Before we give our interpretation of these results, it is important to emphasize that
the computation of speech tracking involves the speech envelope. While we filtered the signals
in narrow frequency bands that were based on manual annotations of linguistic information in
our stimuli (see the section Annotations), and while we checked for acoustic differences via
analysis of the modulation spectra (Figure 1 and S1), we cannot rule out the possibility that
any difference between the conditions in terms of speech tracking is driven by acoustic differ-
ences between the speech recordings. Such an effect would be in line with the fact that sen-
tences and syntactic prose did not show differences in terms of syntax tracking, which is based
on abstract annotations of syntax without any acoustic differences.
A possible reason for the lack of a consistent effect is that the difference between regular
sentences and syntactic prose in terms of compositional processing is not the same at the
phrase and sentence levels. At the sentence level, these conditions were clearly differentia-
ble, as indicated by the ERP results. The N400 for sentence-final verbs was larger in syntac-
tic prose than in regular sentences, showing that participants noticed the sentence-level
semantic incongruency of syntactic prose. At the phrase-level, however, these conditions
might impose similar demands on compositional processing, in particular if the type of com-
position is mostly syntactic. In contrast to previous research (Brodbeck et al., 2018; Kaufeld
et al., 2020), much of the phrase-level compositional processing in our stimuli can be con-
ceived of as mostly syntactic (i.e., containing a combination of a determiner and a noun)
rather than semantic or conceptual. This applies even more strongly to our measure of
Neurobiology of Language
404
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
3
3
8
6
2
0
3
1
3
9
0
n
o
_
a
_
0
0
0
7
0
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cortical tracking of linguistic units
syntax tracking, which is based on abstract annotations of phrase structure. These annota-
tions do not distinguish between combinations like the boat and red boat, even though the
latter phrase involves more semantic and conceptual composition. It is not unlikely that the
presence of semantically impoverished combinations affects the overall degree of phrase-
level tracking (i.e., making syntax tracking a less sensitive measure), especially in light of
the finding that manipulations of lexical-semantic and conceptual composition affect mea-
sures of brain activity on top of the neural response to syntactic aspects of composition
(e.g., Fedorenko et al., 2016, 2020; Kaufeld et al., 2020; Schell et al., 2017; Westerlund
& Pylkkänen, 2014; Zhang & Pylkkänen, 2015).
Sentences vs. Word Lists: Structure and Acoustics
We observed stronger phrase-level speech tracking for sentences than for word lists, and stron-
ger tracking for forward stimuli than for backward stimuli (left panel of Figure 5). The differ-
ence between forward and backward speech has been reported before and is often related to
differences in their intelligibility (Gross et al., 2013; Kaufeld et al., 2020; Park et al., 2015).
However, other studies have failed to find such a relationship (Howard & Poeppel, 2010; Peña
& Melloni, 2012; Zoefel & VanRullen, 2016), and it has been suggested that positive correla-
tions between speech tracking and intelligibility are actually driven by the spectro-temporal
properties of the unintelligible control condition (for discussion, see Kösem & van Wassenhove,
2017; Zou et al., 2019).
Contrasting with the results reported by Kaufeld et al. (2020), the MI difference between
sentences and word lists did not differ across forward and backward versions of these condi-
tions. We therefore cannot exclude the possibility that the difference between (forward) sen-
tences and (forward) word lists reflects acoustic differences. Indeed, analysis of the modulation
spectra shows that sentences were reliably different from word lists in terms of spectral properties
(Figure 1B and Supplementary Information S1). However, the presence of acoustic differences
does not necessarily negate the effect of syntactic differences. When the speech envelopes were
replaced by abstract annotations of syntactic structure (bracket presence or bracket count), we
found stronger MI for sentences than for word lists, presumably because these annotations do not
reflect any syntactic information in word lists. This leaves open the possibility that syntactic
structure did have an effect, but that it could not be detected in measures of speech tracking
due to the masking effect of acoustic variance.
In order to find evidence for cortical tracking of phrase structure, we should find not only a
difference in the forward condition (as a function of syntax), but also no difference in the back-
ward condition. The latter might be quite difficult to obtain with our measures, because naturally
produced sentences and word lists are acoustically quite different, and acoustic properties of the
input can account for much of the variance in the neural response to speech (e.g., Brodbeck
et al., 2018; Doelling et al., 2014). Moreover, the fact that syntactic information and supraseg-
mental modulations (e.g., prosodic phrases, intonation phrases) fluctuate at similar frequencies
and both affect delta-band activity (Bourguignon et al., 2013; Ghitza, 2017; Meyer et al., 2017;
Rimmele et al., 2021) makes it plausible that any structure-driven differences between sentences
and word lists were partially masked by their acoustic differences. Supporting this possibility,
analysis of spectral power in the phrase frequency band showed a bilateral distribution for all
conditions (see Supplementary Information S3; see also e.g., Keitel et al., 2017; Molinaro &
Lizarazu, 2018). This suggests that the neural signal in this frequency band is also affected by
low-frequency modulations in suprasegmental information, which are present in both forward
and backward recordings.
Neurobiology of Language
405
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
3
3
8
6
2
0
3
1
3
9
0
n
o
_
a
_
0
0
0
7
0
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cortical tracking of linguistic units
Cortical Tracking of Lexicalized Structure
At the timescale of phrases, speech tracking was stronger for sentences than for jabberwocky
and syntactic prose, but not different from idioms (left panel of Figure 4). In partial agreement
with these results, syntax tracking was stronger for sentences than for jabberwocky and word
lists, but not different from tracking for idioms and syntactic prose (Figure 6). Overall, cortical
tracking of phrase structure seems to be enhanced for regular sentences compared to stimuli
whose lexical-syntactic content is reduced ( jabberwocky, word lists), but it is not consistently
different from stimuli that contain both lexical content and syntactic structure (idioms, syntac-
tic prose). This pattern of results is in line with the view that cortical tracking of syntactic struc-
ture reflects the generation of structure (Martin, 2020; Martin & Doumas, 2017, 2019; Meyer
et al., 2020), whether this structure transparently maps onto a semantic interpretation or not.
Most current neurobiological models of language processing assume structure building to
be a lexicalized process (Baggio, 2021; Hagoort, 2005, 2017; Martin, 2020; Matchin &
Hickok, 2020). Words are associated with structures that are stored in the mental lexicon in
the form of treelets. During language comprehension, these structures are combined to create
the hierarchical structures of phrases and sentences. In this lexical-syntactic conception of
structure building, the syntax determines which words can be combined, but the words them-
selves are the units of combination. As such, this process is affected by its input, in terms of
both structure and lexical content. Input-wise, both jabberwocky and word lists are markedly
different from sentences. Word lists contain content and function words but lack (cues to)
syntactic structure, which means that adjacent words cannot be combined into phrasal units.
Jabberwocky sentences are structured sequences that contain both function words and
inflectional morphology, but they lack content words and therefore miss the information car-
ried by their argument structure (e.g., the different relations in saw the book on the table and
put the book on the table). The lexical-syntactic difference between regular and jabberwocky
sentences thus explains why they elicit different degrees of phrase-level speech tracking:
Structure-building processes are more weakly activated by lexically impoverished input. This
idea is supported by evidence showing that lexical information affects neurocognitive measures
of structure building (Burroughs et al., 2021; Fedorenko et al., 2016, 2020; Kaufeld et al., 2020;
Matchin et al., 2019; Mollica et al., 2020).
Interestingly, in a recent study Mai and colleagues (2016) reported a different result, namely
stronger delta-band speech tracking for stimuli containing pseudowords than stimuli contain-
ing real words. However, because their design was markedly different from ours, these
findings do not necessarily contradict our interpretation. First, their pseudoword condition
contained lists rather than sentences, so their two conditions differ in both structure
(sentences vs. lists) and lexical-syntactic content (real words vs. pseudowords). Second,
Mai et al. (2016) attribute their delta-tracking effect to the sound matching task they used,
arguing that the semantic and syntactic content of real-word sentences facilitates the recog-
nition of their phonological content. This reduces demands on phonological processing, as
indexed by reduced speech tracking in the delta band. Because we instructed participants to
listen for comprehension rather than to detect a specific sound, we think that our results are
better accounted for in terms of lexical-syntactic structure building than in terms of phonol-
ogical processing.
Idioms and syntactic prose are similar to sentences in lexical-syntactic structure, but they
differ either in the extent to which their interpretation is compositionally derived from this
structure (idioms) or in the extent to which a compositional interpretation of this structure
makes sense (syntactic prose). Given the absence of consistent differences between these
Neurobiology of Language
406
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
3
3
8
6
2
0
3
1
3
9
0
n
o
_
a
_
0
0
0
7
0
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cortical tracking of linguistic units
conditions, the interpretation most strongly supported by our data is that phrase-level tracking
reflects the lexically-driven computation of syntactic structures in the service of semantic com-
position (Kaufeld et al., 2020; Martin, 2020; Martin & Doumas, 2017, 2019; Meyer et al.,
2020). The computations involved in building hierarchical structure are most strongly activated
by syntactically structured sequences of real words. Given the right input, these computa-
tions generate a compositional structure, whether the input can easily compose semantically
or not. A similar idea has been proposed for the segmentation of complex word forms into
stems and grammatical affixes (Marslen-Wilson, 2007). Behavioral and neuroimaging evi-
dence shows that this morphophonological process is triggered by both real words and pseu-
dowords, as long as they contain the diagnostic properties of inflectional affixes in English
(Post et al., 2008; Tyler et al., 2005). To explain the lexical insensitivity of this process, Marslen-
Wilson (2007, p. 180) argued that “without a decompositional analysis, the system cannot rule
out the possibility that the pseudo-regular trade is actually the morpheme tray in the past tense, or
that snade is the past tense of the potential real stem snay.” Analogously, without a decomposi-
tional analysis at the level of phrase structure, the system cannot rule out the possibility that “col-
orless green ideas sleep furiously” (Chomsky, 1957, p. 15) actually involves sleeping ideas or
that to kick the bucket actually involves buckets being kicked. The conclusion that the compo-
sitional meaning of these forms is either semantically incoherent (syntactic prose) or not identical
to their intended figurative meaning (idioms) can only be drawn after a compositional analysis
has taken place. In a sense, then, the structure-building processes know how to build structure
(i.e., adhering to syntactic rules, subcategorization restrictions), but not what is being built (i.e.,
whether a compositional analysis yields an interpretation that makes sense). Supporting this
functional distinction between generation and interpretation, both behavioral and neurobiolog-
ical evidence show a difference between sentences and both idioms and syntactic prose in terms
of compositional meaning, but not in terms of syntactic structure building (Bastiaansen &
Hagoort, 2015; Canal et al., 2017; Konopka & Bock, 2009; Peterson et al., 2001; Rommers
et al., 2013; Vespignani et al., 2010).
Effects of Composition on Word-Level Speech Tracking
At the timescale of words, speech tracking was stronger for both idioms and syntactic prose
than for sentences (middle panel of Figure 4). These word-level effects might be related to the
differential predictability of words in these conditions, as it has been shown that speech track-
ing is enhanced for unpredictable target words that are presented in low-constraining sentence
contexts (Donhauser & Baillet, 2020; Molinaro et al., 2021). In these contexts, target words
cannot be predicted by top-down mechanisms, so the brain might rely more strongly on the
bottom-up input to ensure successful comprehension (Donhauser & Baillet, 2020; Molinaro
et al., 2021). Words in syntactic prose are semantically odd and thus very unpredictable.
On this account, the brain attunes more strongly to unexpected words (in syntactic prose),
whose content can only be derived from a bottom-up analysis, than to expected words (in
sentences), whose content can be predicted by top-down mechanisms, explaining the differ-
ence between syntactic prose and regular sentences in terms of word-level speech tracking.
ERP analysis showed that sentence-final verbs elicited a less negative N400 in idioms than
in regular sentences, indexing facilitated activation or integration of the verb in idioms (e.g.,
Moreno et al., 2002; Rommers et al., 2013). This indicates that sentence-final verbs were more
predictable in idioms than in sentences, in line with the behavioral literature (Cacciari et al.,
2007; Cacciari & Tabossi, 1988). This predictability-related ERP difference is opposite to the
difference between sentences and syntactic prose, which suggests that the difference between
idioms and sentences in terms of word-level speech tracking (middle panel of Figure 4) does
Neurobiology of Language
407
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
3
3
8
6
2
0
3
1
3
9
0
n
o
_
a
_
0
0
0
7
0
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cortical tracking of linguistic units
not have the same origin as the difference between syntactic prose and sentences. It is unclear
why words would be tracked more closely in idioms, but one possibility is that participants
were relatively more attuned to words in idioms because words are the linguistic unit on
which participants have to rely to activate and retrieve the full idiom from memory (for dis-
cussion of the activation of properties of the individual words, see Cacciari, 2014; Cacciari &
Tabossi, 1988; Hubers et al., 2021; Sprenger et al., 2006).
CONCLUSION
Despite a constantly growing literature on cortical speech tracking, it is still unclear which
aspects of high-level linguistic content drive neural activity into alignment with the speech sig-
nal. In this EEG study with naturally spoken stimuli, we used an experimental design with a
parametric modulation of linguistic information, comparing compositional sentences to stimuli
that diverged in terms of their relationship between structure and meaning (idioms, syntactic
prose, jabberwocky, word lists). We found that the brain tracks syntactic phrases more closely
in regular sentences than in stimuli whose lexical-syntactic content is reduced, but we found no
consistent differences in phrase tracking between sentences and stimuli that contained a com-
bination of both syntactic structure and lexical content. These findings refine a recent account
of cortical speech tracking, which holds that it indexes the generation of linguistic structure
(Martin & Doumas, 2017, 2019; Meyer et al., 2020). Specifically, they suggest that phrase-level
speech tracking is modulated by the lexical-syntactic properties of the input to structure build-
ing, not by the compositional interpretation of its output. This is in line with neurobiological
models of language processing in which structure building is a lexicalized process.
ACKNOWLEDGMENTS
We thank Greta Kaufeld for helpful suggestions on the Mutual Information analysis and Eva
Poort for the recordings of her voice.
FUNDING INFORMATION
Andrea E. Martin, Nederlandse Organisatie voor Wetenschappelijk Onderzoek (https://dx.doi
.org/10.13039/501100003246), Award ID: 016.Vidi.188.029. Peter Hagoort, Nederlandse
Organisatie voor Wetenschappelijk Onderzoek (https://dx.doi.org/10.13039/501100003246),
Award ID: 024.001.006. Andrea E. Martin, Max-Planck-Gesellschaft (https://dx.doi.org/10
.13039/501100004189), Award ID: Lise Meitner Research Group.
AUTHOR CONTRIBUTIONS
Cas W. Coopmans: Conceptualization; Data curation; Formal analysis; Investigation; Meth-
odology; Project administration; Validation; Visualization; Writing – original draft; Writing –
review & editing. Helen de Hoop: Conceptualization; Methodology; Writing – review &
editing. Peter Hagoort: Conceptualization; Methodology; Writing – review & editing.
Andrea E. Martin: Conceptualization; Methodology; Supervision; Writing – review & editing.
REFERENCES
Ahissar, E., Nagarajan, S., Ahissar, M., Protopapas, A., Mahncke, H.,
& Merzenich, M. M. (2001). Speech comprehension is correlated
with temporal response patterns recorded from auditory cortex.
Proceedings of the National Academy of Sciences, 98(23),
13367–13372. https://doi.org/10.1073/pnas.201400998,
PubMed: 11698688
Baayen, R. H., Davidson, D. J., & Bates, D. M. (2008). Mixed-
effects modeling with crossed random effects for subjects and
Neurobiology of Language
408
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
3
3
8
6
2
0
3
1
3
9
0
n
o
_
a
_
0
0
0
7
0
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cortical tracking of linguistic units
items. Journal of Memory and Language, 59(4), 390–412. https://
doi.org/10.1016/j.jml.2007.12.005
Baggio, G. (2021). Compositionality in a parallel architecture for
language processing. Cognitive Science, 45(5), Article e12949.
https://doi.org/10.1111/cogs.12949, PubMed: 34018238
Baggio, G., & Hagoort, P. (2011). The balance between memory
and unification in semantics: A dynamic account of the N400.
Language and Cognitive Processes, 26(9), 1338–1367. https://
doi.org/10.1080/01690965.2010.542671
Bastiaansen, M. C. M., & Hagoort, P. (2015). Frequency-based
segregation of syntactic and semantic unification during online
sentence level language comprehension. Journal of Cognitive
Neuroscience, 27(11), 2095–2107. https://doi.org/10.1162/jocn
_a_00829, PubMed: 26042498
Bates, D., Mächler, M., Bolker, B., & Walker, S. (2015). Fitting linear
mixed-effects models using lme4. Journal of Statistical Software,
67(1), 1–48. https://doi.org/10.18637/jss.v067.i01
Blanco-Elorrieta, E., Ding, N., Pylkkänen, L., & Poeppel, D. (2020).
Understanding requires tracking: Noise and knowledge interact
in bilingual comprehension. Journal of Cognitive Neuroscience,
32(10), 1975–1983. https://doi.org/10.1162/jocn_a_01610,
PubMed: 32662732
Boersma, P., & Weenink, D. (2019). Praat: Doing phonetics by
computer ( Version 6.1.02) [Computer software]. https://www
.fon.hum.uva.nl/praat/
Bourguignon, M., De Tiège, X., Op De Beeck, M., Ligot, N., Paquier,
P., Van Bogaert, P., Goldman, S., Hari, R., & Jousmäki, V. (2013).
The pace of prosodic phrasing couples the listener’s cortex to the
reader’s voice. Human Brain Mapping, 34(2), 314–326. https://doi
.org/10.1002/hbm.21442, PubMed: 22392861
Brennan, J. R., & Martin, A. E. (2020). Phase synchronization varies
systematically with linguistic structure composition. Philosoph-
ical Transactions of the Royal Society B: Biological Sciences,
375(1791), Article 20190305. https://doi.org/10.1098/rstb.2019
.0305, PubMed: 31840584
Brennan, J. [R.], Nir, Y., Hasson, U., Malach, R., Heeger, D. J., &
Pylkkänen, L. (2012). Syntactic structure building in the anterior
temporal lobe during natural story listening. Brain and Language,
120(2), 163–173. https://doi.org/10.1016/j.bandl.2010.04.002,
PubMed: 20472279
Brennan, J. R., Stabler, E. P., Van Wagenen, S. E., Luh, W.-M., &
Hale, J. T. (2016). Abstract linguistic structure correlates with
temporal activity during naturalistic comprehension. Brain and
Language, 157–158, 81–94. https://doi.org/10.1016/j.bandl
.2016.04.008, PubMed: 27208858
Brodbeck, C., Presacco, A., & Simon, J. Z. (2018). Neural source
dynamics of brain responses to continuous stimuli: Speech pro-
cessing from acoustics to comprehension. NeuroImage, 172,
162–174. https://doi.org/10.1016/j.neuroimage.2018.01.042,
PubMed: 29366698
Broderick, M. P., Anderson, A. J., & Lalor, E. C. (2019). Semantic
context enhances the early auditory encoding of natural speech.
Journal of Neuroscience, 39(38), 7564–7575. https://doi.org/10
.1523/JNEUROSCI.0584-19.2019, PubMed: 31371424
Burroughs, A., Kazanina, N., & Houghton, C. (2021). Grammatical
category and the neural processing of phrases. Scientific Reports,
11(1), Article 2446. https://doi.org/10.1038/s41598-021-81901-5,
PubMed: 33510230
Cacciari, C. (2014). Processing multiword idiomatic strings: Many
words in one? The Mental Lexicon, 9(2), 267–293. https://doi.org
/10.1075/ml.9.2.05cac
Cacciari, C., & Corradini, P. (2015). Literal analysis and idiom
retrieval in ambiguous idioms processing: A reading-time study.
Journal of Cognitive Psychology, 27(7), 797–811. https://doi.org
/10.1080/20445911.2015.1049178
Cacciari, C., & Glucksberg, S. (1991). Understanding idiomatic
expressions: The contribution of word meanings. In G. B. Simpson
(Ed.), Understanding word and sentence (pp. 217–240). Elsevier.
https://doi.org/10.1016/S0166-4115(08)61535-6
Cacciari, C., Padovani, R., & Corradini, P. (2007). Exploring the
relationship between individuals’ speed of processing and their
comprehension of spoken idioms. European Journal of Cognitive
P s y c ho l o g y, 1 9 ( 3 ) , 4 1 7 –4 4 5 . h t t p s : / / d o i . o rg / 1 0 . 1 0 8 0
/09541440600763705
Cacciari, C., & Tabossi, P. (1988). The comprehension of idioms.
Journal of Memory and Language, 27(6), 668–683. https://doi
.org/10.1016/0749-596X(88)90014-9
Canal, P., Pesciarelli, F., Vespignani, F., Molinaro, N., & Cacciari, C.
(2017). Basic composition and enriched integration in idiom pro-
cessing: An EEG study. Journal of Experimental Psychology:
Learning, Memory, and Cognition, 43(6), 928–943. https://doi
.org/10.1037/xlm0000351, PubMed: 28068127
Chandrasekaran, C., Trubanova, A., Stillittano, S., Caplier, A., &
Ghazanfar, A. A. (2009). The natural statistics of audiovisual
speech. PLOS Computational Biology, 5(7), Article e1000436.
https://doi.org/10.1371/journal.pcbi.1000436, PubMed:
19609344
Chomsky, N. (1957). Syntactic structures. Mouton. https://doi.org
/10.1515/9783112316009
Cogan, G. B., & Poeppel, D. (2011). A mutual information analysis
of neural coding of speech by low-frequency MEG phase infor-
mation. Journal of Neurophysiology, 106(2), 554–563. https://doi
.org/10.1152/jn.00075.2011, PubMed: 21562190
Coopmans, C. W., & Nieuwland, M. S. (2020). Dissociating activa-
tion and integration of discourse referents: Evidence from ERPs
and oscillations. Cortex, 126, 83–106. https://doi.org/10.1016/j
.cortex.2019.12.028, PubMed: 32065957
Cutting, J. C., & Bock, K. (1997). That’s the way the cookie
bounces: Syntactic and semantic components of experimentally
elicited idiom blends. Memory & Cognition, 25(1), 57–71.
https://doi.org/10.3758/BF03197285, PubMed: 9046870
Di Liberto, G. M., Lalor, E. C., & Millman, R. E. (2018). Causal cor-
tical dynamics of a predictive enhancement of speech intellig-
ibility. NeuroImage, 166, 247–258. https://doi.org/10.1016/j
.neuroimage.2017.10.066, PubMed: 29102808
Ding, N., Melloni, L., Yang, A., Wang, Y., Zhang, W., & Poeppel, D.
(2017). Characterizing neural entrainment to hierarchical linguis-
tic units using electroencephalography (EEG). Frontiers in Human
Neuroscience, 11, Article 481. https://doi.org/10.3389/fnhum
.2017.00481, PubMed: 29033809
Ding, N., Melloni, L., Zhang, H., Tian, X., & Poeppel, D. (2016).
Cortical tracking of hierarchical linguistic structures in connected
speech. Nature Neuroscience, 19(1), 158–164. https://doi.org/10
.1038/nn.4186, PubMed: 26642090
Ding, N., Patel, A. D., Chen, L., Butler, H., Luo, C., & Poeppel,
D. (2017). Temporal modulations in speech and music. Neu-
roscience & Biobehavioral Reviews, 81(Pt. B), 181–187.
https://doi.org/10.1016/j.neubiorev.2017.02.011, PubMed:
28212857
Ding, N., & Simon, J. Z. (2014). Cortical entrainment to continuous
speech: Functional roles and interpretations. Frontiers in Human
Neuroscience, 8, Article 311. https://doi.org/10.3389/fnhum
.2014.00311, PubMed: 24904354
Doelling, K. B., Arnal, L. H., Ghitza, O., & Poeppel, D. (2014).
Acoustic landmarks drive delta–theta oscillations to enable
speech comprehension by facilitating perceptual parsing.
Neurobiology of Language
409
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
3
3
8
6
2
0
3
1
3
9
0
n
o
_
a
_
0
0
0
7
0
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cortical tracking of linguistic units
NeuroImage, 85(Pt. 2), 761–768. https://doi.org/10.1016/j
.neuroimage.2013.06.035, PubMed: 23791839
Donhauser, P. W., & Baillet, S. (2020). Two distinct neural timescales
for predictive speech processing. Neuron, 105(2), 385–393.
https://doi.org/10.1016/j.neuron.2019.10.019, PubMed:
31806493
Fedorenko, E., Blank, I. A., Siegelman, M., & Mineroff, Z. (2020).
Lack of selectivity for syntax relative to word meanings through-
out the language network. Cognition, 203, Article 104348.
https://doi.org/10.1016/j.cognition.2020.104348, PubMed:
32569894
Fedorenko, E., Scott, T. L., Brunner, P., Coon, W. G., Pritchett, B.,
Schalk, G., & Kanwisher, N. (2016). Neural correlate of the con-
struction of sentence meaning. Proceedings of the National Acad-
emy of Sciences, 113(41), E6256–E6262. https://doi.org/10.1073
/pnas.1612132113, PubMed: 27671642
Frank, S. L., & Yang, J. (2018). Lexical representation explains corti-
cal entrainment during speech comprehension. PLOS One, 13(5),
Article e0197304. https://doi.org/10.1371/journal.pone
.0197304, PubMed: 29771964
Getz, H., Ding, N., Newport, E. L., & Poeppel, D. (2018). Cortical
tracking of constituent structure in language acquisition. Cogni-
tion, 181, 135–140. https://doi.org/10.1016/j.cognition.2018.08
.019, PubMed: 30195135
Ghitza, O. (2017). Acoustic-driven delta rhythms as prosodic markers.
Language, Cognition and Neuroscience, 32(5), 545–561. https://doi
.org/10.1080/23273798.2016.1232419
Gibbs, R. W., Nayak, N. P., & Cutting, C. (1989). How to kick the
bucket and not decompose: Analyzability and idiom processing.
Journal of Memory and Language, 28(5), 576–593. https://doi.org
/10.1016/0749-596X(89)90014-4
Giraud, A.-L., & Poeppel, D. (2012). Cortical oscillations and
speech processing: Emerging computational principles and oper-
ations. Nature Neuroscience, 15(4), 511–517. https://doi.org/10
.1038/nn.3063, PubMed: 22426255
Gross, J., Hoogenboom, N., Thut, G., Schyns, P., Panzeri, S., Belin,
P., & Garrod, S. (2013). Speech rhythms and multiplexed oscilla-
tory sensory coding in the human brain. PLOS Biology, 11(12),
Article e1001752. https://doi.org/10.1371/journal.pbio
.1001752, PubMed: 24391472
Hagoort, P. (2005). On Broca, brain, and binding: A new frame-
work. Trends in Cognitive Sciences, 9(9), 416–423. https://doi
.org/10.1016/j.tics.2005.07.004, PubMed: 16054419
Hagoort, P. (2017). The core and beyond in the language-ready brain.
Neuroscience & Biobehavioral Reviews, 81(Pt. B), 194–204. https://
doi.org/10.1016/j.neubiorev.2017.01.048, PubMed: 28193452
Holsinger, E., & Kaiser, E.
(2013). Processing (non)composi-
tional expressions: Mistakes and recovery.
Journal of Exper-
imental Psychology: Learning, Memory, and Cognition, 39(3),
866–878. https://doi.org/10.1037/a0030410, PubMed:
23088547
Howard, M. F., & Poeppel, D. (2010). Discrimination of speech
stimuli based on neuronal response phase patterns depends on
acoustics but not comprehension. Journal of Neurophysiology,
104(5), 2500–2511. https://doi.org/10.1152/jn.00251.2010,
PubMed: 20484530
Hubers, F., Cucchiarini, C., Strik, H., & Dijkstra, T. (2021). Individ-
ual word activation and word frequency effects during the pro-
cessing of opaque idiomatic expressions. Quarterly Journal of
Experimental Psychology, 1–17. https://doi.org/10.1177
/17470218211047995, PubMed: 34507505
Hubers, F. C. W., van Ginkel, W., Cucchiarini, C., Strik, H., & Dijkstra,
A. F. J. (2018). Normative data on Dutch idiomatic expressions:
Native speakers [Data set]. Data Archiving and Networked Services
(DANS). 75(6). https://doi.org/10.17026/dans-zjx-hnsk
Ince, R. A. A., Giordano, B. L., Kayser, C., Rousselet, G. A., Gross,
J., & Schyns, P. G. (2017). A statistical framework for neuroimag-
ing data analysis based on mutual information estimated via a
Gaussian copula. Human Brain Mapping, 38(3), 1541–1573.
https://doi.org/10.1002/hbm.23471, PubMed: 27860095
Jackendoff, R. (1995). The boundaries of the lexicon. In M. Everaert,
E. J. van der Linden, A. Schenk, & R. Schroeder (Eds.), Idioms:
Structural and psychological perspectives (pp. 133–165). Erlbaum.
Jackendoff, R. (2017). In defense of theory. Cognitive Science, 41(S2),
185–212. https://doi.org/10.1111/cogs.12324, PubMed:
26611772
Jin, P., Lu, Y., & Ding, N. (2020). Low-frequency neural activity
reflects rule-based chunking during speech listening. eLife, 9,
Article e55613. https://doi.org/10.7554/eLife.55613, PubMed:
32310082
Kaan, E., & Swaab, T. Y. (2002). The brain circuitry of syntactic
comprehension. Trends in Cognitive Sciences, 6(8), 350–356.
https://doi.org/10.1016/S1364-6613(02)01947-2, PubMed:
12140086
Kaufeld, G., Bosker, H. R., ten Oever, S., Alday, P. M., Meyer, A. S.,
& Martin, A. E. (2020). Linguistic structure and meaning organize
neural oscillations into a content-specific hierarchy. Journal of
Neuroscience, 40(49), 9467–9475. https://doi.org/10.1523
/JNEUROSCI.0302-20.2020, PubMed: 33097640
Kayser, S. J., Ince, R. A. A., Gross, J., & Kayser, C. (2015). Irregular
speech rate dissociates auditory cortical entrainment, evoked
responses, and frontal alpha. Journal of Neuroscience, 35(44),
14691–14701. https://doi.org/10.1523/ JNEUROSCI.2243-15
.2015, PubMed: 26538641
Keitel, A., Gross, J., & Kayser, C. (2018). Perceptually relevant
speech tracking in auditory and motor cortex reflects distinct lin-
guistic features. PLOS Biology, 16(3), Article e2004473. https://
doi.org/10.1371/journal.pbio.2004473, PubMed: 29529019
Keitel, A., Ince, R. A. A., Gross, J., & Kayser, C. (2017). Auditory
cortical delta-entrainment interacts with oscillatory power in
multiple fronto-parietal networks. NeuroImage, 147, 32–42.
https://doi.org/10.1016/j.neuroimage.2016.11.062, PubMed:
27903440
Keuleers, E., & Brysbaert, M. (2010). Wuggy: A multilingual pseu-
doword generator. Behavior Research Methods, 42(3), 627–633.
https://doi.org/10.3758/BRM.42.3.627, PubMed: 20805584
Keuleers, E., Brysbaert, M., & New, B. (2010). SUBTLEX-NL: A new
measure for Dutch word frequency based on film subtitles.
Behavior Research Methods, 42(3), 643–650. https://doi.org/10
.3758/BRM.42.3.643, PubMed: 20805586
Konopka, A. E., & Bock, K. (2009). Lexical or syntactic control of
sentence formulation? Structural generalizations from idiom pro-
duction. Cognitive Psychology, 58(1), 68–101. https://doi.org/10
.1016/j.cogpsych.2008.05.002, PubMed: 18644587
Kösem, A., & van Wassenhove, V. (2017). Distinct contributions of
low- and high-frequency neural oscillations to speech comprehen-
sion. Language, Cognition and Neuroscience, 32(5), 536–544.
https://doi.org/10.1080/23273798.2016.1238495
Kutas, M., & Federmeier, K. D. (2011). Thirty years and counting:
Finding meaning in the N400 component of the event-related
brain potential (ERP). Annual Review of Psychology, 62(1),
621–647. https://doi.org/10.1146/annurev.psych.093008
.131123, PubMed: 20809790
Libben, M. R., & Titone, D. A. (2008). The multidetermined nature
of idiom processing. Memory & Cognition, 36(6), 1103–1121.
https://doi.org/10.3758/MC.36.6.1103, PubMed: 18927029
Neurobiology of Language
410
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
3
3
8
6
2
0
3
1
3
9
0
n
o
_
a
_
0
0
0
7
0
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cortical tracking of linguistic units
Luo, H., & Poeppel, D. (2007). Phase patterns of neuronal
responses reliably discriminate speech in human auditory cortex.
Neuron, 54(6), 1001–1010. https://doi.org/10.1016/j.neuron
.2007.06.004, PubMed: 17582338
Mai, G., Minett, J. W., & Wang, W. S.-Y. (2016). Delta, theta, beta,
and gamma brain oscillations index levels of auditory sentence
processing. NeuroImage, 133, 516–528. https://doi.org/10.1016/j
.neuroimage.2016.02.064, PubMed: 26931813
Makov, S., Sharon, O., Ding, N., Ben-Shachar, M., Nir, Y., &
Golumbic, E. Z. (2017). Sleep disrupts high-level speech parsing
despite significant basic auditory processing. Journal of Neurosci-
ence, 37(32), 7772–7781. https://doi.org/10.1523/JNEUROSCI
.0168-17.2017, PubMed: 28626013
Marslen-Wilson, W. D. (2007). Morphological processes in lan-
guage comprehension. In M. G. Gaskell (Ed.), Oxford hand-
book of psycholinguistics (pp. 175–186). Oxford University
Press. https://doi.org/10.1093/oxfordhb/9780198568971.013
.0011
Marslen-Wilson, W. [D.], & Tyler, L. K. (1980). The temporal struc-
ture of spoken language understanding. Cognition, 8(1), 1–71.
https://doi.org/10.1016/0010-0277(80)90015-3, PubMed:
7363578
Martin, A. E. (2016). Language processing as cue integration:
Grounding the psychology of language in perception and neuro-
physiology. Frontiers in Psychology, 7, Article 120. https://doi.org
/10.3389/fpsyg.2016.00120, PubMed: 26909051
Martin, A. E. (2020). A compositional neural architecture for
language. Journal of Cognitive Neuroscience, 32(8), 1407–1427.
https://doi.org/10.1162/jocn_a_01552, PubMed: 32108553
Martin, A. E., & Doumas, L. A. A. (2017). A mechanism for the
cortical computation of hierarchical linguistic structure. PLOS
Biology, 15(3), Article e2000663. https://doi.org/10.1371
/journal.pbio.2000663, PubMed: 28253256
Martin, A. E., & Doumas, L. A. A. (2019). Predicate learning in
neural systems: Using oscillations to discover latent structure.
Current Opinion in Behavioral Sciences, 29, 77–83. https://doi
.org/10.1016/j.cobeha.2019.04.008
Matchin, W., Brodbeck, C., Hammerly, C., & Lau, E. (2019). The
temporal dynamics of structure and content in sentence compre-
hension: Evidence from fMRI-constrained MEG. Human Brain
Mapping, 40(2), 663–678. https://doi.org/10.1002/hbm.24403,
PubMed: 30259599
Matchin, W., & Hickok, G. (2020). The cortical organization of
syntax. Cerebral Cortex, 30(3), 1481–1498. https://doi.org/10
.1093/cercor/bhz180, PubMed: 31670779
Mazoyer, B. M., Tzourio, N., Frak, V., Syrota, A., Murayama, N.,
Levrier, O., Salamon, G., Dehaene, S., Cohen, L., & Mehler, J.
(1993). The cortical representation of speech. Journal of Cogni-
tive Neuroscience, 5(4), 467–479. https://doi.org/10.1162/jocn
.1993.5.4.467, PubMed: 23964919
Meyer, L., Henry, M. J., Gaston, P., Schmuck, N., & Friederici, A. D.
(2017). Linguistic bias modulates interpretation of speech via neu-
ral delta-band oscillations. Cerebral Cortex, 27(9), 4293–4302.
https://doi.org/10.1093/cercor/bhw228, PubMed: 27566979
Meyer, L., Sun, Y., & Martin, A. E. (2020). Synchronous, but not
entrained: Exogenous and endogenous cortical rhythms of
speech and language processing. Language, Cognition and Neu-
roscience, 35(9), 1089–1099. https://doi.org/10.1080/23273798
.2019.1693050
Molinaro, N., & Lizarazu, M. (2018). Delta(but not theta)-band cor-
tical entrainment involves speech-specific processing. European
Journal of Neuroscience, 48(7), 2642–2650. https://doi.org/10
.1111/ejn.13811, PubMed: 29283465
Molinaro, N., Lizarazu, M., Baldin, V., Pérez-Navarro, J., Lallier, M.,
& Ríos-López, P. (2021). Speech-brain phase coupling is
enhanced in low contextual semantic predictability conditions.
Neuropsychologia, 156, Article 107830. https://doi.org/10.1016
/j.neuropsychologia.2021.107830, PubMed: 33771540
Mollica, F., Siegelman, M., Diachek, E., Piantadosi, S. T., Mineroff,
Z., Futrell, R., Kean, H., Qian, P., & Fedorenko, E. (2020). Com-
position is the core driver of the language-selective network.
Neurobiology of Language, 1(1), 104–134. https://doi.org/10
.1162/nol_a_00005
Moreno, E. M., Federmeier, K. D., & Kutas, M. (2002). Switching
languages, switching palabras (words): An electrophysiological
study of code switching. Brain and Language, 80(2), 188–207.
https://doi.org/10.1006/brln.2001.2588, PubMed: 11827443
Nelson, M. J., El Karoui, I., Giber, K., Yang, X., Cohen, L., Koopman,
H., Cash, S. S., Naccache, L., Hale, J. T., Pallier, C., & Dehaene, S.
(2017). Neurophysiological dynamics of phrase-structure building
during sentence processing. Proceedings of the National Academy
of Sciences, 114(18), E3669–E3678. https://doi.org/10.1073/pnas
.1701590114, PubMed: 28416691
Obleser, J., & Kayser, C. (2019). Neural entrainment and atten-
tional selection in the listening brain. Trends in Cognitive Sci-
ences, 23(11), 913–926. https://doi.org/10.1016/j.tics.2019.08
.004, PubMed: 31606386
Oostenveld, R., Fries, P., Maris, E., & Schoffelen, J.-M. (2011). Field-
Trip: Open source software for advanced analysis of MEG, EEG,
and invasive electrophysiological data. Computational Intelli-
gence and Neuroscience, 2011, Article 156869. https://doi.org
/10.1155/2011/156869, PubMed: 21253357
Park, H., Ince, R. A. A., Schyns, P. G., Thut, G., & Gross, J. (2015).
Frontal top-down signals increase coupling of auditory
low-frequency oscillations to continuous speech in human lis-
teners. Current Biology, 25(12), 1649–1653. https://doi.org/10
.1016/j.cub.2015.04.049, PubMed: 26028433
Peelle, J. E., & Davis, M. H. (2012). Neural oscillations carry speech
rhythm through to comprehension. Frontiers in Psychology, 3,
Article 320. https://doi.org/10.3389/fpsyg.2012.00320,
PubMed: 22973251
Peña, M., & Melloni, L. (2012). Brain oscillations during spoken
sentence processing. Journal of Cognitive Neuroscience, 24(5),
1149–1164. https://doi.org/10.1162/jocn_a_00144, PubMed:
21981666
Peterson, R. R., Burgess, C., Dell, G. S., & Eberhard, K. M. (2001).
Dissociation between syntactic and semantic processing during
idiom comprehension. Journal of Experimental Psychology:
Learning, Memory, and Cognition, 27(5), 1223–1237. https://
doi.org/10.1037/0278-7393.27.5.1223, PubMed: 11550750
Poeppel, D., & Assaneo, M. F. (2020). Speech rhythms and their neu-
ral foundations. Nature Reviews Neuroscience, 21(6), 322–334.
https://doi.org/10.1038/s41583-020-0304-4, PubMed: 32376899
Post, B., Marslen-Wilson, W. D., Randall, B., & Tyler, L. K. (2008).
The processing of English regular inflections: Phonological cues
to morphological structure. Cognition, 109(1), 1–17. https://doi
.org/10.1016/j.cognition.2008.06.011, PubMed: 18834584
R Core Team. (2021). R: A language and environment for statistical
computing [Computer software]. R Foundation for Statistical
Computing. https://www.r-project.org
Rimmele, J. M., Morillon, B., Poeppel, D., & Arnal, L. H. (2018).
Proactive sensing of periodic and aperiodic auditory patterns.
Trends in Cognitive Sciences, 22(10), 870–882. https://doi.org
/10.1016/j.tics.2018.08.003, PubMed: 30266147
Rimmele, J. M., Poeppel, D., & Ghitza, O. (2021). Acoustically
driven cortical δ oscillations underpin prosodic chunking.
Neurobiology of Language
411
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
3
3
8
6
2
0
3
1
3
9
0
n
o
_
a
_
0
0
0
7
0
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cortical tracking of linguistic units
Eneuro, 8(4), Article 0562-20.2021. https://doi.org/10.1523
/ENEURO.0562-20.2021, PubMed: 34083380
Rommers, J., Dijkstra, T., & Bastiaansen, M. (2013). Context-dependent
semantic processing in the human brain: Evidence from idiom com-
prehension. Journal of Cognitive Neuroscience, 25(5), 762–776.
https://doi.org/10.1162/jocn_a_00337, PubMed: 23249356
Schell, M., Zaccarella, E., & Friederici, A. D. (2017). Differential
cortical contribution of syntax and semantics: An fMRI study
on two-word phrasal processing. Cortex, 96, 105–120. https://
doi.org/10.1016/j.cortex.2017.09.002, PubMed: 29024818
Schroeder, C. E., & Lakatos, P. (2009). Low-frequency neuronal
oscillations as instruments of sensory selection. Trends in Neuro-
sciences, 32(1), 9–18. https://doi.org/10.1016/j.tins.2008.09.012,
PubMed: 19012975
Sheng, J., Zheng, L., Lyu, B., Cen, Z., Qin, L., Tan, L. H., Huang,
M.-X., Ding, N., & Gao, J.-H. (2019). The cortical maps of hier-
archical linguistic structures during speech perception. Cerebral
Cortex, 29(8), 3232–3240. https://doi.org/10.1093/cercor
/bhy191, PubMed: 30137249
Smith, Z. M., Delgutte, B., & Oxenham, A. J. (2002). Chimaeric sounds
reveal dichotomies in auditory perception. Nature, 416(6876),
87–90. https://doi.org/10.1038/416087a, PubMed: 11882898
Smolka, E., Rabanus, S., & Rösler, F. (2007). Processing verbs in
German idioms: Evidence against the configuration hypothesis.
Metaphor and Symbol, 22(3), 213–231. https://doi.org/10.1080
/10926480701357638
Sprenger, S. A., Levelt, W. J. M., & Kempen, G. (2006). Lexical
access during the production of idiomatic phrases. Journal of
Memory and Language, 54(2), 161–184. https://doi.org/10.1016
/j.jml.2005.11.001
ten Oever, S., & Martin, A. E. (2021). An oscillating computational
model can track pseudo-rhythmic speech by using linguistic pre-
dictions. eLife, 10, Article e68066. https://doi.org/10.7554/eLife
.68066, PubMed: 34338196
Titone, D. A., & Connine, C. M. (1999). On the compositional and
noncompositional nature of idiomatic expressions. Journal of
Pragmatics, 31(12), 1655–1674. https://doi.org/10.1016/S0378
-2166(99)00008-9
Tyler, L. K., Stamatakis, E. A., Post, B., Randall, B., & Marslen-Wilson,
W. (2005). Temporal and frontal systems in speech comprehen-
sion: An fMRI study of past tense processing. Neuropsychologia,
43(13), 1963–1974. https://doi.org/10.1016/j.neuropsychologia
.2005.03.008, PubMed: 16168736
Vespignani, F., Canal, P., Molinaro, N., Fonda, S., & Cacciari, C.
(2010). Predictive mechanisms in idiom comprehension. Journal
of Cognitive Neuroscience, 22(8), 1682–1700. https://doi.org/10
.1162/jocn.2009.21293, PubMed: 19580384
Westerlund, M., & Pylkkänen, L. (2014). The role of the left anterior
temporal lobe in semantic composition vs. semantic memory.
Neuropsychologia, 57, 59–70. https://doi.org/10.1016/j
.neuropsychologia.2014.03.001, PubMed: 24631260
Zhang, L., & Pylkkänen, L. (2015). The interplay of composition
and concept specificity in the left anterior temporal lobe: An
MEG study. NeuroImage, 111, 228–240. https://doi.org/10.1016
/j.neuroimage.2015.02.028, PubMed: 25703829
Zoefel, B., & VanRullen, R. (2015). The role of high-level processes
for oscillatory phase entrainment to speech sound. Frontiers in
Human Neuroscience, 9, Article 651. https://doi.org/10.3389
/fnhum.2015.00651, PubMed: 26696863
Zoefel, B., & VanRullen, R. (2016). EEG oscillations entrain their
phase to high-level features of speech sound. NeuroImage,
124, 16–23. https://doi.org/10.1016/j.neuroimage.2015.08.054,
PubMed: 26341026
Zou, J., Feng, J., Xu, T., Jin, P., Luo, C., Zhang, J., Pan, X., Chen, F.,
Zheng, J., & Ding, N. (2019). Auditory and language contribu-
tions to neural encoding of speech features in noisy environ-
ments. NeuroImage, 192, 66–75. https://doi.org/10.1016/j
.neuroimage.2019.02.047, PubMed: 30822469
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
3
3
8
6
2
0
3
1
3
9
0
n
o
_
a
_
0
0
0
7
0
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neurobiology of Language
412