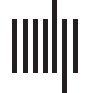RESEARCH ARTICLE
A Weak Shadow of Early Life Language
Processing Persists in the Right Hemisphere of the
Mature Brain
a n o p e n a c c e s s
j o u r n a l
Kelly C. Martin1
William D. Gaillard1,3
, Anna Seydell-Greenwald1,2
, Peter E. Turkeltaub1,2
, Madison M. Berl1,3
,
, and Elissa L. Newport1,2
1Center for Brain Plasticity and Recovery, Georgetown University Medical Center, Washington, DC
2MedStar National Rehabilitation Hospital, Washington, DC
3Children’s National Hospital, Washington, DC
Keywords: language, development, pediatric stroke, fMRI, lateralization, developmental plasticity
ABSTRACT
Studies of language organization show a striking change in cerebral dominance for
language over development: We begin life with a left hemisphere (LH) bias for language
processing, which is weaker than that in adults and which can be overcome if there is a LH
injury. Over development this LH bias becomes stronger and can no longer be reversed.
Prior work has shown that this change results from a significant reduction in the magnitude
of language activation in right hemisphere (RH) regions in adults compared to children.
Here we investigate whether the spatial distribution of language activation, albeit weaker in
magnitude, still persists in homotopic RH regions of the mature brain. Children aged 4–13
(n = 39) and young adults (n = 14) completed an auditory sentence comprehension fMRI
(functional magnetic resonance imaging) task. To equate neural activity across the
hemispheres, we applied fixed cutoffs for the number of active voxels that would be
included in each hemisphere for each participant. To evaluate homotopicity, we generated
left-right flipped versions of each activation map, calculated spatial overlap between the LH
and RH activity in frontal and temporal regions, and tested for mean differences in the
spatial overlap values between the age groups. We found that, in children as well as in
adults, there was indeed a spatially intact shadow of language activity in the right frontal
and temporal regions homotopic to the LH language regions. After a LH stroke in
adulthood, recovering early-life activation in these regions might assist in enhancing
recovery of language abilities.
INTRODUCTION
In the mature human brain, the cerebral hemispheres exhibit relative biases for certain cogni-
tive abilities. The classic example is the left hemisphere bias for language processing, which is
observed in the majority of adults regardless of handedness (Broca, 1861; regarding handed-
ness: Knecht, Deppe, et al., 2000; Knecht, Dräger, et al., 2000). However, the developmental
course of this bias is not fully understood. Studies of infants who suffered a unilateral brain
injury provide compelling evidence that the two cerebral hemispheres are at first equivalent
Citation: Martin, K. C., Seydell-
Greenwald, A., Berl, M. M., Gaillard,
W. D., Turkeltaub, P. E., & Newport,
E. L. (2022). A weak shadow of early life
language processing persists in the
right hemisphere of the mature brain.
Neurobiology of Language, 3(3),
364–385. https://doi.org/10.1162/nol_a
_00069
DOI:
https://doi.org/10.1162/nol_a_00069
Supporting Information:
https://doi.org/10.1162/nol_a_00069
Received: 26 August 2021
Accepted: 10 February 2022
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Author:
Kelly C. Martin
kcm81@georgetown.edu
Handling Editor:
Denise Klein
Copyright: © 2022
Massachusetts Institute of Technology
Published under a Creative Commons
Attribution 4.0 International
(CC BY 4.0) license
The MIT Press
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
3
3
6
4
2
0
2
3
4
0
3
n
o
_
a
_
0
0
0
6
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Weak shadow of language processing in the non-dominant hemisphere
Language equipotentiality:
The theory that the left and right
hemispheres have equal capability
to support language processing early
in life.
Cerebral lateralization:
A bias for one cerebral hemisphere
to dominate a certain type of
information processing.
Task fMRI activation magnitude:
The size of the resulting t value at
each voxel when an experimental
condition is contrasted with a control
condition.
(equipotential) in the sense that either can support language when critical regions are irrevers-
ibly damaged early in development (Lenneberg, 1967, 1969). Indeed, language can develop
successfully in the right hemisphere after a left hemisphere stroke around the time of birth (Fair
et al., 2010; Guzzetta et al., 2008; Ilves et al., 2014; Newport et al., 2017; Staudt et al., 2001,
2002; Stiles et al., 2012; Tillema et al., 2008) or following hemispherectomy for refractory
epilepsy in the first years of life (Boatman et al., 1999; Bulteau et al., 2015; Liégeois, Connelly,
et al., 2008; Liégeois, Cross, et al., 2008). In the healthy brain, however, the left and right
hemispheres do not engage equally during language processing: The left hemisphere exhibits
language dominance that has been measured as early as the first months of life (Peña et al.,
2003). This lateralization strengthens throughout childhood until it stabilizes around the age of
10 (Berl, Mayo, et al., 2014). Taken together, it appears that we begin life with a left hemi-
sphere bias for language processing which can be overcome if there is a left hemisphere injury,
but over the course of typical cognitive and brain maturation, we develop a strongly left-
lateralized language system.
In the early years of typical development, in addition to the left hemisphere bias cited
above, the right hemisphere also participates in the language functions that will later
become strongly left-lateralized (for example, sentence processing; Berl, Mayo, et al.,
2014; Gaillard et al., 2000; Holland et al., 2007). A recent study by Olulade et al. (2020)
used functional magnetic resonance imaging (fMRI) to measure language activation during
sentence processing in the left and right hemispheres of typically developing 4–6-year-old
children, as well as of older children and adults. All participants performed an auditory
sentence comprehension task, in which a button-pushing task to spoken sentences was
contrasted with a button-pushing task to the same sentences played backwards (therefore
unintelligible but acoustically matched to the spoken sentences). In this study, sentence
comprehension robustly activated left hemisphere frontal and temporal regions at all ages.
Crucially, however, Olulade et al. also found significant clusters of activity in right hemi-
sphere frontal and temporal regions in the individual activation maps for most of the youn-
gest children. When they compared 4–6-year-olds, 7–9-year-olds, 10–13-year-olds, and
young adults, they found a continuous pattern of decline in the number of participants
showing significant clusters of language activity in right frontal and temporal regions. The
right frontal cortex also exhibited an age-related decline in activation magnitude. The
authors suggested that the right hemisphere frontal and temporal regions that support
language after early life injury are the ones also responsive during language processing in
the typically developing brain. Through an unknown mechanism of brain and cognitive
maturation, the magnitude of this response declines and renders language activity scarcely
detectable in the right hemisphere of adults.
The consequence of this maturational change is that when an adult suffers a stroke to left
hemisphere language centers, unlike what is observed after early-life stroke, language is often
permanently impaired. Even when right hemisphere areas are activated during language pro-
cessing in these patients, the response tends to be weak and is at best associated with limited
language recovery (Martin et al., 2022; Skipper-Kallal et al., 2017a, b). How then do right
hemisphere frontal and temporal regions transition from being equipotential for language pro-
cessing early in life to being unavailable for language processing in the mature brain?
The answer to this question is not only critical to understanding how language may be
recovered in the right hemisphere after a stroke in adulthood, but may also illuminate the prin-
ciples that govern how cognitive abilities change during typical development. One possibility
is that the right hemisphere regions that are equipotential for language early in life may con-
tinue to respond during language processing in adults, but only weakly. This “weak shadow”
Neurobiology of Language
365
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
3
3
6
4
2
0
2
3
4
0
3
n
o
_
a
_
0
0
0
6
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Weak shadow of language processing in the non-dominant hemisphere
Homotopic:
Brain regions that are parallel to one
another in opposite hemispheres.
of language activity in right hemisphere regions may reflect populations of language-
responsive neurons that are either fewer in number than in similar regions of the left hemi-
sphere, or are less specifically tuned for linguistic input. An alternative possibility is that these
right hemisphere regions are recruited for other cognitive functions in the mature brain, and
there is no longer a spatially intact trace of the language-responsive neuronal populations that
existed early in life.
An fMRI study of healthy adults by Just et al. (1996) pointed to the existence of a “weak
echo” of language activation in the right hemisphere counterparts of Broca’s and Wernicke’s
areas. They observed that as sentence complexity increased, the amount of activation in
Broca’s and Wernicke’s areas also increased—and in homotopic right hemisphere regions,
the amount of activation (which was much lower than in the left hemisphere regions)
increased as well. To our knowledge, no study since Just et al.’s (1996) has intentionally
investigated whether a weak language response persists in homotopic right hemisphere
regions of the healthy adult brain. This investigation is important to our understanding of
language system organization, and whether homotopic right hemisphere regions may be
viable under certain conditions to recover language processing after stroke in adulthood.
The primary reason why homotopic language activity in the adult brain is underreported
in fMRI studies is because of the way we define activity in our analyses. Most studies using
fMRI to localize the neural correlates of sentence processing report only those areas with
sufficiently strong activity to survive conventional statistical thresholding (e.g., voxelwise
threshold of p < 0.001 and cluster-defining threshold of p < 0.05 on supra-threshold voxels).
Using these cutoffs, right hemisphere areas are rarely above threshold during sentence pro-
cessing in adults, or sometimes emerge but are rarely the focal point of discussion. For
example, a recent fMRI study by Quillen et al. (2021) investigated the dissociation of brain
region recruitment for linguistic and nonlinguistic tasks in a sample of healthy adults.
Although they were not specifically examining the response in homotopic regions, they
found the same pattern of activity modulation in mirroring left and right inferior frontal gyrus,
pars triangularis, and posterior superior temporal sulcus for easy and difficult semantic judg-
ments, and the same pattern of deactivation to perceptual judgments, but with a much
weaker percentage signal change in the right hemisphere regions. Quillen et al. were posi-
tioned to observe this similarity because in their region of interest (ROI) selection using lan-
guage activation data from an orthogonal task, they opted to include right hemisphere
regions that “did not reach statistical significance but were clearly homotopic to left hemi-
sphere language areas” (Quillen et al., 2021, p. 7). These findings accord with the observa-
tion of Just et al. (1996) that right hemisphere regions may respond similarly to core left
hemisphere language regions when processing linguistic information. However, customary
analysis decisions regarding what levels of activity magnitude are reported can mean that
there is a relative absence of these findings in the published literature. It is possible that a
weak shadow of the language engagement in right frontal and temporal regions that was
observed early in life may indeed still persist in the adult right hemisphere, but we have
largely been ignoring it.
One approach to defining activity that would capture the localization of both weak and
strong neuronal responses is to examine a particular number of active voxels without setting
a threshold for the individual t values. Fedorenko and colleagues routinely employ a similar
method for questions about the spatial localization of language activity (Mollica et al., 2020;
Shain et al., 2020). This type of top voxel approach would make it possible to equalize the
amount of activity being assessed to determine, for example, whether the weak language
response in right hemisphere regions is localized homotopic to the strong language response
Neurobiology of Language
366
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
3
3
6
4
2
0
2
3
4
0
3
n
o
_
a
_
0
0
0
6
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Weak shadow of language processing in the non-dominant hemisphere
in left hemisphere regions. As noted above, Olulade et al. (2020) examined which brain
regions were active in child and adult participants during an auditory sentence comprehension
task. Their results suggest that language recruits right frontal and temporal regions with high
consistency in young children but this recruitment systematically declines over age. However,
a weak response to language stimuli in right frontal and temporal cortex would not survive the
conventional statistical cutoff used, even at the individual level. In order to measure whether
the spatial distribution of activity is similar on the two sides of the brain, the quantity of activity
in the left and right hemispheres needs to be equalized.
In the current work, we wanted to determine whether language activation in homotopic
right hemisphere areas persists into adulthood. We know from Olulade et al.’s (2020) findings
that activation magnitude in the right hemisphere during language processing changes with
age in the right hemisphere, but we do not know whether the spatial distribution of activation,
albeit weaker in magnitude, still persists in homotopic right hemisphere regions of the mature
brain. Here we investigate whether there is a weak shadow of language activity in the right
frontal and temporal cortex in adults by examining whether the localization of activity is
homotopic in the two hemispheres irrespective of the magnitude of this activity using a top
voxel approach. We hypothesized that in adults there would indeed be a spatially intact
shadow of language activation in the homotopic right frontal and temporal regions, and that
the homotopicity between the left and right hemispheres in adults would be similar to the
homotopicity we measured in 4–6-, 7–9-, and 10–13-year-old children. These results would
indicate that in the mature brain, tissue that once supported language processing early in life
may still be available to some degree in right hemisphere regions. This finding is potentially
important for language recovery after a left hemisphere stroke in adulthood: Perhaps recover-
ing some or all of the early-life activation in these regions might be relevant to enhancing
recovery of language processing abilities in adults after stroke.
METHODS
To investigate whether a weak shadow of language activation persists in homotopic right
hemisphere regions of the mature brain, we re-analyzed the language activation data from
children and adults reported in Olulade et al. (2020). Crucially, we equated the number of
active voxels in the left and right hemisphere using the top voxel approach described below,
and then calculated how overlapping the right hemisphere language activation was when
transposed onto the activity in left hemisphere regions. This analysis decision made it possible
to compare relatively weak activity in the right hemisphere to relatively strong activity in the
left, to determine whether the localization of language processing is precisely homotopic at
different ages. See Olulade et al. (2020) for a more comprehensive description of participants’
task and IQ (intelligence quotient) performance, MRI (magnetic resonance imaging) acquisi-
tion, and fMRI preprocessing and first-level statistical maps.
Participants
This sample included neurologically healthy children (4–13 years old; who also served as
healthy controls in Berl, Mayo, et al., 2014; Berl, Zimmaro, et al., 2014; Gaillard et al.,
2007) and young adults (18–29 years old; Table 1). All included participants were right-
handed, had IQs in the average or above-average range (measured using age-appropriate
behavioral tests; see Olulade et al., 2020), and were native speakers of English with no signif-
icant exposure to another language before age 4.
Neurobiology of Language
367
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
3
3
6
4
2
0
2
3
4
0
3
n
o
_
a
_
0
0
0
6
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Weak shadow of language processing in the non-dominant hemisphere
Table 1.
Age distribution
Age
n (females)
4–6-year-olds
10 (6)
7–9-year-olds
14 (8)
10–13-year-olds
15 (7)
Adults
14 (7)
Age mean (SD)
5.5 (0.77)
8.5 (0.86)
11.4 (0.92)
21.4 (3.09)
Age range
4.5–6.7
7.4–9.8
10.0–13.0
18.4–29.1
Note. Adapted from Olulade et al. (2020).
Magnetic Resonance Imaging
Auditory description decision fMRI task
Participants were scanned while they performed an Auditory Description Decision Task
(ADDT) developed by Gaillard and colleagues (Berl, Mayo, et al., 2014; Berl, Zimmaro,
et al., 2014; Gaillard et al., 2007). Participants heard short auditory descriptions of different
nouns (Forward Speech condition, e.g., “a big gray animal is an elephant”) and pressed a but-
ton when they judged it to be accurate (70% of sentences were accurate descriptions), and
they also heard unintelligible sound sequences (Reverse Speech condition, the same sentences
from the forward condition played backwards) and pressed a button following a beep at the
end (70% of sequences were followed by a beep). The 5-min task run was divided into ten
30-s blocks, each containing 10 stimuli presented every 3 s with 1 s of response time following
each stimulus. The five blocks of forward and five blocks of reverse speech were interleaved,
always starting with reverse speech. The frequencies of the target words in the sentences were
adjusted by age group to equalize difficulty and to keep all age groups at high accuracy. Word
frequencies were taken from a corpus of children’s reading materials, with the same materials
used for the oldest children (10–12-year-olds) and adults (see Berl, Mayo, et al., 2014, for
details). Prior to their scan, participants practiced the task on a parallel set of materials in a
mock scanner to acclimate them to the environment and familiarize them with the task. All
participants achieved at least 85% accuracy on the in-scanner task.
Scanner and auditory equipment
MRI data were collected on a 3 Tesla Siemens MAGNETOM Trio scanner with a 12-channel
headcoil at Georgetown University’s Center for Functional and Molecular Imaging. Auditory
stimuli for the fMRI task were presented through Sensimetrics Model S14 insert headphones,
and participants wore additional Bilsom ear defenders to reduce the interference of scanner noise.
Researchers confirmed that participants could clearly hear the stimuli over the scanner noise.
Scan sequences
A high-resolution anatomical image was collected: Siemens MPRAGE, 176 sagittal slices, TR =
1.9 s, TE = 2.52 ms, flip angle = 9 deg, 1 × 1 × 1 mm voxels, whole-brain coverage. Functional
images were collected during the ADDT: echo-planar images, 50 horizontal slices, descending
order, TR = 3 s, TE = 30 ms, flip angle = 90 deg, 3 × 3 × 3 mm voxels, whole-brain coverage,
100 volumes for a total of 5 min. A second run was collected if the participant moved exces-
sively, but only one run was analyzed.
Functional MRI Preprocessing and First-Level Statistical Maps
The first-level statistical maps from Olulade et al. (2020) were used in the current analyses. See
Olulade et al. (2020) for a complete description of preprocessing and statistical analysis of
Neurobiology of Language
368
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
3
3
6
4
2
0
2
3
4
0
3
n
o
_
a
_
0
0
0
6
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Weak shadow of language processing in the non-dominant hemisphere
Figure 1. Analysis workflow. We (A) flipped the activation map for each participant, (B) masked activation in regions of interest (lan-
guage regions defined by a meta-analysis database), (C) applied a top voxel cutoff to equalize the quantity of activation within the ROI in
the left and flipped right maps, and (D) calculated the spatial overlap in the left and flipped right activation areas with a dice coefficient.
L: left, R: right.
these data. Briefly, functional images were slice-time corrected, realigned to the first volume of
the run, co-registered to the native-space anatomical image, spatially normalized into MNI
(Montreal Neurological Institute) standard space, and then smoothed (8 mm FWHM (full-width
at half-maximum) Gaussian kernel) using SPM-8 ( Wellcome Trust Centre for Neuroimaging at
University College London, https://www.fil.ion.ucl.ac.uk/spm/doc/) and the VBM-8 toolbox
(developed by Christian Gaser, University of Jena, dbm.neuro.uni-jena.de/vbm8/ VBM8
-Manual.pdf). Prior studies have demonstrated that adults and children in the age range we
studied can be registered to a common anatomical template without introducing a significant
age bias (Burgund et al., 2002; Ghosh et al., 2010). Volumes with extreme motion (>0.75 mm
between volumes) were deweighted by a “bad scan” regressor in the statistical model; if the
number of bad scans exceeded 25%, the run was not analyzed. The first-level general linear
model included 14 predictors: Forward and reverse speech condition time courses convolved
with a canonical hemodynamic response function, motion estimates for rotation and transla-
tion along the x, y, and z axes, the “bad scan” regressor, a global signal regressor, and a high-
pass filter of four cosine basis functions. The resulting forward and reverse speech condition
beta maps were contrasted using voxel-wise t tests to identify voxels that were more active
during language processing. These activation maps (example in Figure 1A) were used in sub-
sequent analyses.
Flipped Activation Maps
To calculate how symmetrical (homotopic) task-related activity was within each individual
activation map, we flipped each participant’s activation map across the midline (using
SPM-12’s reorient utility) and normalized it to their MNI-space-warped anatomical image
(using SPM-12’s normalization function). Thus their right hemisphere activation was warped
to their left hemisphere anatomy in MNI space (Figure 1A). We then calculated the spatial
overlap between the left hemisphere activation and the flipped right hemisphere activation
to quantify how symmetrical (homotopic) language activity was for each participant
(Figure 1D).
Neurobiology of Language
369
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
3
3
6
4
2
0
2
3
4
0
3
n
o
_
a
_
0
0
0
6
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Weak shadow of language processing in the non-dominant hemisphere
Regions of Interest
To investigate homotopicity within the language network, we used the Neurosynth open-
source online meta-analysis database (https://neurosynth.org/; Yarkoni et al., 2011) to generate
a map of activation locations for all studies that reported the keyword “language” at least once
in the abstract. Briefly, the Neurosynth platform calculates a two-way ANOVA (analysis of var-
iance) to test for the association between the keyword “language” and voxel activation, and
the resulting F values are then z-scored: Regions with positive z scores are more consistently
reported as active in studies that used the term “language” than in studies that did not. These
associations are then thresholded with a false discovery rate criterion of 0.01. We created sep-
arate binarized versions of the frontal and temporal areas from this map by assigning a 1 to all
voxels with a z-score of 1 or greater (using SPM-12’s image calculator utility). These ROI
masks (Figure 1B) were resliced to match the dimensions of the activation maps. We isolated
the frontal and temporal activations for the original and flipped versions of each participant’s
activation map by multiplying them by each of the binarized ROI masks. We chose to examine
frontal and temporal language activations separately because these regions may have slightly
different relationships with their homotopic counterparts; for example, it has been shown that
lateralization stabilizes in temporal regions earlier than frontal regions in development (Berl,
Mayo, et al., 2014).
Equalizing the Activation Areas (Top Voxel Approach)
Asking whether the activity during language processing is homotopic in individual participants
requires determining how overlapping the spatial locations of the activity in the left and right
hemisphere are. It is well known that the amount of activity in the left hemisphere is greater
than it is in the right, so we must equalize the quantity of activation in the left and right hemi-
spheres before we can compare how much they overlap. To accomplish this, we determined
four different cutoffs per ROI for the number of active voxels to be included for overlap com-
parison (Figure 1C and Table 2). These cutoffs were matched across participants and in the
original and flipped activation maps. When a participant had fewer voxels with t values that
met the statistical threshold than the average, we included the voxels with the next highest
t values for the spatial overlap comparison. We examined the results using four different cutoffs
to ensure that the spatial overlap we calculated would not be biased by the particular cutoff
selected. To determine these cutoffs, for the frontal and temporal ROIs separately, we first tal-
lied the number of voxels active in the original activation map (i.e., left hemisphere language
centers) for each participant at four statistical thresholds (p < 0.01, 0.005, 0.001, 0.0005) with
minimal cluster thresholding (k = 4). We then found the average number of voxels active
within each ROI across all participants at each threshold, and selected these average values
as the top voxel cutoffs (Figure 1C and Table 2). For each participant, we then ranked the
voxels within each ROI from highest t value to lowest, and selected the number of top voxels
equal to each cutoff. This approach ensures that the same number of voxels are examined in
each participant to preclude developmental changes in activation magnitude from contami-
nating results. We applied the same cutoffs to both the original and flipped activation maps
Table 2. Number of voxels analyzed in regions of interest
Frontal
ROI Size
12,922
Level 1 (p < 0.01)
3,311
Level 2 (p < 0.005)
2,763
Level 3 (p < 0.001)
1,794
Level 4 (p < 0.0005)
1,484
Temporal
12,685
2,588
2,188
1,498
1,278
Neurobiology of Language
370
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
3
3
6
4
2
0
2
3
4
0
3
n
o
_
a
_
0
0
0
6
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Weak shadow of language processing in the non-dominant hemisphere
within each ROI to ensure that the same number of voxels were examined in the left and right
hemispheres (Figure 1D). For the individual participant values that went into these averages,
see supplementary materials Figure S3 and Table S1 (Supporting Information can be found at
https://doi.org/10.1162/nol_a_00069).
Spatial Overlap Calculation (Dice Coefficient)
We calculated a dice coefficient (Figure 1D) to summarize the number of overlapping voxels
for each pair of maps (the left and right hemisphere language activations, i.e., the flipped and
unflipped ADDT maps; x and y):
Þ
ð
2 x ⋂ y
x þ y
For each participant, we averaged the dice coefficient calculated at each top voxel cutoff level
to obtain one summary value of spatial overlap for each region. We grouped these values by
age range to compare children to adults.
To test whether our measured dice coefficients are greater than would be measured by
chance, we shuffled binary vectors the same length as the ROIs, with the same number of
simulated active voxels to match each top voxel cutoff applied to the actual data, and calcu-
lated dice coefficients ten thousand times.
We also performed a between-subjects comparison of each person’s right hemisphere activ-
ity with the left hemisphere activations of all other participants in the same age group (labeled
R with Others’ L). This comparison captures the variability in language localization in the left
and right hemispheres between similarly aged participants and allows us to determine whether
within-participant homotopicity exceeds the overlap arising purely from the fact that language
activations are spatially similar to a degree in all subjects and all hemispheres. In this case, the
x and y in the dice equation above would be one participant’s right hemisphere activation and
another participant’s left hemisphere activation.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
3
3
6
4
2
0
2
3
4
0
3
n
o
_
a
_
0
0
0
6
9
p
d
.
/
l
Penetrance Maps
Penetrance maps highlight the localization of the top voxels for all participants in each age
group, for the most lenient cutoff (number of top voxels corresponding to an average voxelwise
p < 0.01 in the individual participant activation maps; Table 2 and Figure 1C). These maps,
rendered on the MNI-152 standard template using MRIcroGL (https://www.nitrc.org/projects
/mricrogl), visualize where language activity was the most consistent across individuals.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Statistical Comparisons
Frequentist statistical analyses were performed using R through R-Studio (https://www.R
-project.org/). Bayesian statistical analyses were performed using the JASP open-source sta-
tistical software (version 0.14.1; https://jasp-stats.org/download/). We ran separate tests for
each ROI because the size of the ROIs and number of voxels compared to calculate the
dice coefficients differed.
Homotopicity comparison (Right with Own Left)
We statistically compared the four age groups to measure whether there were age-related dif-
ferences in the dice coefficients calculated for each participant’s left and right hemisphere acti-
vations (language homotopicity). We ran a one-way ANOVA for each ROI to examine the
Neurobiology of Language
371
Weak shadow of language processing in the non-dominant hemisphere
effects of age group (4–6-, 7–9-, 10–13-year-olds, and adults) on the homotopicity dice coef-
ficient (overlap between left and flipped right activity) averaged across all four top voxel cutoff
levels. Because our primary prediction was a lack of difference between age groups on the
overlap comparison of right with own left (homotopicity), we also ran complementary one-
way Bayesian ANOVA, which quantifies the amount of evidence in favor of the null hypothesis
(no mean differences) as well as whether there is insufficient evidence to support the presence
or absence of an effect.
Between-subjects right-left comparison (Right with Others’ Left)
We also statistically compared the language homotopicity dice coefficients to a between-
subjects measure of language localization consistency in both hemispheres (a dice coefficient
calculated between each participant’s right hemisphere activity and the left hemisphere activ-
ity for all other participants in the same age group). We ran a 4 × 2 mixed effects ANOVA for
each ROI to examine the effects of age group (between-subjects factor) and overlap compar-
ison (within-subjects factor with two levels: right with own left (homotopicity) and right with
others’ left) on the dice coefficient measured across all four levels. We also ran a complemen-
tary 4 × 2 Bayesian repeated measures ANOVA to quantify the sufficiency of evidence in favor
of the null hypothesis.
RESULTS
When activation magnitude was equated through our top voxel method, there were no differ-
ences between the developing and mature brain in the degree to which left and right frontal
and temporal activations were homotopic. The average dice coefficients (Table 3) across the
four top voxel cutoffs tested (Table 2) were similar across the four age groups in the frontal and
temporal ROIs (Figure 2). One-way ANOVAs measuring the effect of age group on the homo-
topicity dice coefficient in each ROI showed no significant main effect of age group (Table 4;
Frontal: F(3, 49) = 1.018, p = 0.393; Temporal: F(3, 49) = 0.237, p = 0.87). See supplementary
materials for additional correlation results (Figures S1 and S2). Under the Bayesian one-way
ANOVA framework, we found that the null model predicted the observed data 3.661 times
better than the age group model in the frontal ROI, and 7.813 times better in the temporal
ROI, which is considered moderate evidence to support the absence of an effect of age on
homotopicity (Table 5).
Table 3. Dice coefficient descriptive statistics
Age
4–6-year-olds
7–9-year-olds
10–13-year-olds
Adults
Mean (SD)
Frontal ROI
R with Own L
0.33 (0.09)
0.33 (0.12)
0.33 (0.10)
0.27 (0.09)
R with Others’ L
Temporal ROI
0.23 (0.05)
0.24 (0.04)
0.21 (0.04)
0.22 (0.03)
R with Own L
0.29 (0.13)
0.26 (0.11)
0.28 (0.11)
0.28 (0.07)
R with Others’ L
0.21 (0.07)
0.20 (0.09)
0.26 (0.07)
0.20 (0.04)
Neurobiology of Language
372
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
3
3
6
4
2
0
2
3
4
0
3
n
o
_
a
_
0
0
0
6
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Weak shadow of language processing in the non-dominant hemisphere
Figure 2. Average homotopicity for each age group. The pairwise dice coefficient averages for individual participants were binned by age
group. There were no mean differences in the average homotopic overlap between age groups (means and standard deviations in Table 3, one-
way ANOVAs in Tables 4 and 5). The white dashed lines indicate the dice coefficient that would be measured between random arrangements
of the same number of active voxels in the same size space (0.1809 in the frontal and 0.1488 in the temporal ROI), which is substantially lower
than the group means for all age groups.
Table 4. One-way ANOVAs
SSn
SSd
dfn
dfd
F
p
Predictor
Frontal ROI
Age Group
0.031
0.496
Temporal ROI
Age Group
0.008
0.541
3
3
49
49
1.018
0.39
0.237
0.87
Note. Dice coefficient for homotopic overlap by age group. Sum of squares in the numerator (SSn) and denom-
inator (SSd); degrees of freedom in the numerator (dfn) and denominator (dfd); F statistic; p value.
Models
Frontal ROI
Null model
Age Group
Temporal ROI
Null model
Age Group
Table 5.
Bayesian one-way ANOVAs
P(M)
P(M|data)
0.500
0.500
0.500
0.500
0.785
0.215
0.887
0.113
BFM
3.661
0.273
7.813
0.128
error %
0.003
0.001
Note. Dice coefficient for homotopic overlap by age group. Prior model probability (P(M)); posterior model
probability (P(M|data)); change from prior model odds to posterior model odds (BFM; here, Bayes Factor for
the Null model relative to model containing Age Group, and Bayes Factor for the model containing Age Group
relative to the Null model).
Neurobiology of Language
373
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
3
3
6
4
2
0
2
3
4
0
3
n
o
_
a
_
0
0
0
6
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Weak shadow of language processing in the non-dominant hemisphere
The dice values measuring homotopicity in our participants were substantially greater than
what would be measured between random arrangements of the same number of active voxels
in the same size space. The actual dice coefficient averaged across top voxel cutoff levels in
the frontal ROI for adults (0.2731) was approximately 147 standard deviations above the sim-
ulated dice for the frontal ROI (average of 0.1809 and standard deviation of 0.00062775
across top voxel cutoffs); similarly, the actual temporal ROI dice for adults (0.2754) was
approximately 291 standard deviations above the simulated temporal ROI dice (average of
0.1488 and standard deviation of 0.00043544 across top voxel cutoffs).
The spatial distribution of language activity is somewhat consistent across individuals (e.g.,
robust activation of inferior frontal and superior temporal regions), but also somewhat idiosyn-
cratic (e.g., more variable recruitment and activation robustness of additional regions;
Fedorenko & Kanwisher, 2009). Thus the high dice coefficients above might reflect general
left-right symmetry of commonly activated language regions, or might additionally reflect
symmetry of more idiosyncratic individualized localization of language processors. To dis-
criminate between these two possibilities, we conducted a second reference comparison, this
time comparing individual participant overlap to the overlap between each participant’s right
hemisphere activation and the left hemisphere activations of the other participants in the same
age group. Large overlap values between participants’ right hemisphere language activations
and other participants’ left hemisphere language activations would indicate that language is
localized with great consistency in the left and right hemispheres across individuals in each
age group. If individuals’ own between-hemispheres overlap (homotopicity) exceeds the over-
lap of their right hemisphere activity with other participants’ left hemisphere activity, this
would indicate that the idiosyncratic nuances of individual language networks are homotopic
above and beyond the left-right symmetry of common language regions.
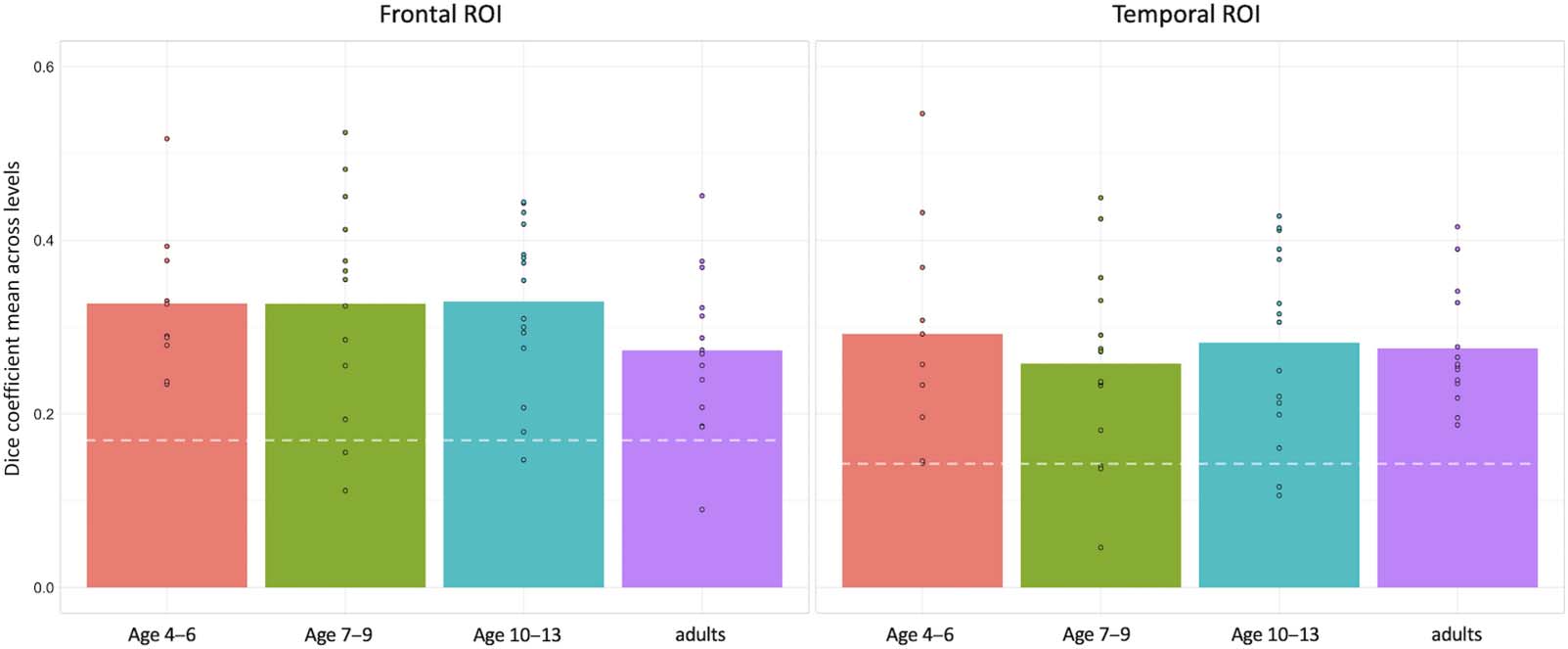
We found that within-participant homotopicity was greater than between-subjects right-left
overlap in both frontal and temporal regions (Figure 3). Two-way mixed effects ANOVAs (cal-
culated separately for the frontal and temporal ROIs) measuring the effect of age group
(between-subjects factor) and overlap comparison (within-subjects factor with two levels: right
with own left, and right with others’ lefts) on the dice coefficient revealed a significant main
effect of overlap comparison (Frontal: F(1, 49) = 37.797, p < 0.000001; Temporal: F(1, 49) =
Figure 3. Group averages for each overlap comparison. For each participant in an age group, we compared the overlap of the activity in their
right with their own left hemisphere (homotopicity) to the overlap between their right with all other participants’ left hemispheres in the age
group (colorful vs. gray bars, with lines connecting individual participants) to interpret whether within-participant homotopicity was greater
than between-subjects’ right-left overlap. See Table 3 for means and standard deviations, and Tables 6 and 7 for two-way mixed effects
ANOVAs.
Neurobiology of Language
374
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
3
3
6
4
2
0
2
3
4
0
3
n
o
_
a
_
0
0
0
6
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Weak shadow of language processing in the non-dominant hemisphere
Table 6.
Two-way mixed effects ANOVAs
SSn
SSd
dfn
dfd
F
p
Predictor
Frontal ROI
Age Group
Overlap Comparison
0.022
0.325
0.195
0.253
Age Group * Overlap Comparison
0.015
0.253
Temporal ROI
Age Group
Overlap Comparison
0.027
0.526
0.092
0.239
Age Group * Overlap Comparison
0.014
0.239
3
1
3
3
1
3
49
49
49
49
49
49
1.128
0.347
37.797
1.38e−07
*
0.973
0.413
0.844
0.477
18.903
6.93e−05
*
0.974
0.413
Note. Dice coefficient by age group (between-subjects factor) & overlap comparison (within-subjects factor).
Sum of squares in the numerator (SSn) and denominator (SSd); degrees of freedom in the numerator (dfn) and
denominator (dfd); F statistic; p value.
* denotes statistically significant effects.
18.903, p < 0.0001), but no significant main effect of age group ( p > 0.3 for both ROIs) or age
group by overlap comparison interaction ( p > 0.4 for both ROIs; Table 6). See supplementary
materials for individual participant data (Figure S5). We also calculated two-way repeated
measures ANOVAs using a Bayesian statistics framework. For both ROIs, the null model pre-
dicted the observed data approximately 5–8 times better than the model that included age
group alone, which is considered moderate evidence to support the absence of an effect of
age group on homotopicity (Table 7).
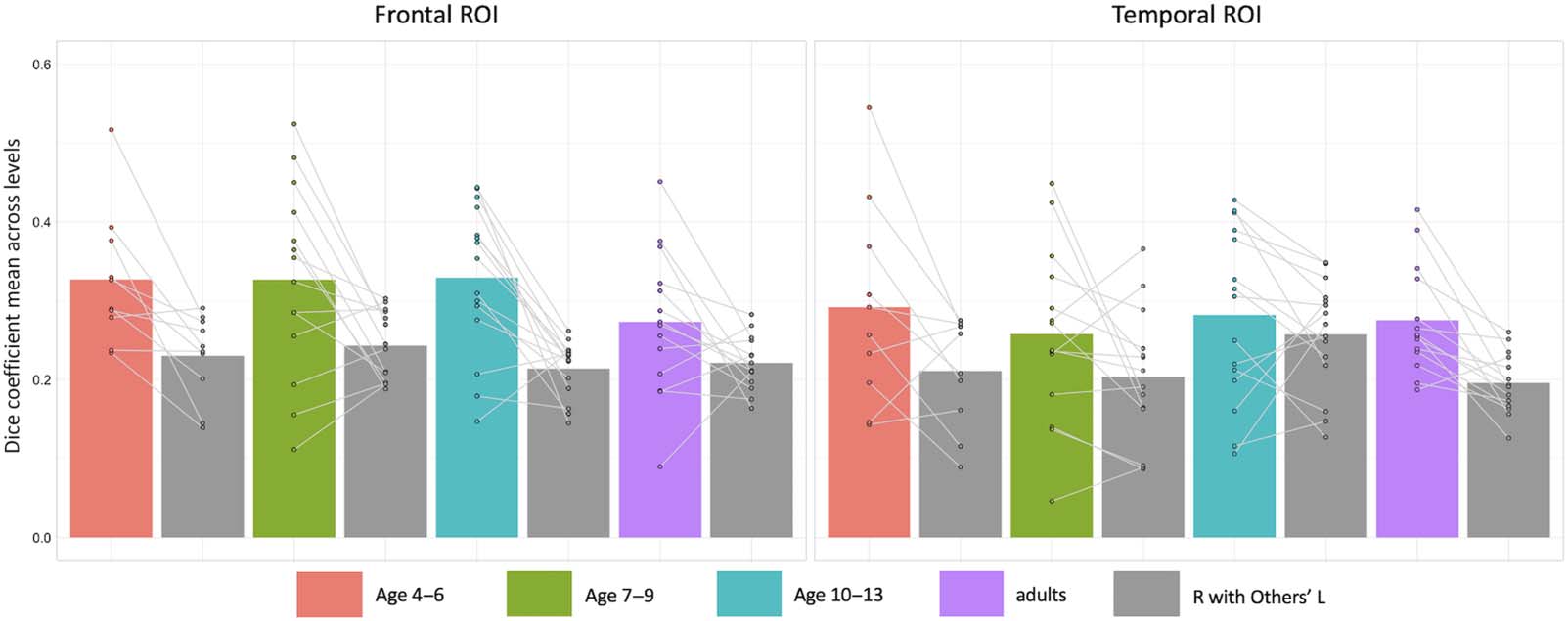
Penetrance maps showing the consistency of homotopic activity localization (Figure 4)
reveal that the inferior frontal gyrus pars triangularis and orbitalis and mid-to-posterior superior
temporal sulcus appear to be the most consistent areas of homotopic activity across age
groups. Interestingly, the ventral occipitotemporal cortex, for which the left hemisphere com-
ponent is implicated in reading word forms in literate adults, also appears to be homotopically
activated by our auditory comprehension task with some consistency in adult but not young
child participants. See supplementary materials for the penetrance maps of the activation con-
sistency for each age group in the left and right hemisphere ROIs respectively (Figure S4).
DISCUSSION
In the current work we investigated whether there is a weak shadow of language activity in the
right frontal and temporal cortex of children and adults by using a top voxel approach to
examine the degree to which the activity is similar in the two hemispheres irrespective of
the magnitude of this activity. We know from prior research (Olulade et al., 2020) that activity
magnitude in the right hemisphere during language processing declines with age, but we did
not know whether this activity, albeit weaker in magnitude, still continues to some degree in
homotopic right hemisphere regions. The current approach allowed us to investigate the loca-
tion of activation while ignoring activation magnitude by comparing the same number of
active voxels in the left and right hemispheres. To make this comparison, we examined dice
coefficients calculated between participants’ left and flipped right hemispheres in frontal and
temporal ROIs, averaged across the four top voxel cutoff levels investigated. We then grouped
these values by age range to compare children to adults. We find that in older children as well
Neurobiology of Language
375
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
3
3
6
4
2
0
2
3
4
0
3
n
o
_
a
_
0
0
0
6
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Weak shadow of language processing in the non-dominant hemisphere
Models
Frontal ROI
Table 7.
Bayesian repeated measures ANOVA
P(M)
P(M|data)
BFM
BF01
BF10
error %
Overlap Comparisons: R with Own L and R with Others’ L
Null model
(includes subject)
0.200
3.235e−7
1.294e−6
1.000
1.000
Overlap Comparison
0.200
0.775
13.790
4.174e−7
2,369,000
1.052
Age Group + Overlap
0.200
0.173
0.838
1.867e−6
5,355,301
1.010
Comparison
Age Group + Overlap
0.200
0.052
0.218
6.273e−6
159,417
1.081
Comparison +
Age Group *
Comparison
Age Group
Temporal ROI
0.200
5.364e−8
2.146e−7
6.032
0.166
0.649
Overlap Comparisons: R with Own L and R with Others’ L
Null model
0.200
0.002
0.010
Overlap Comparison
0.200
0.759
12.569
Age Group + Overlap
0.200
0.189
0.933
Comparison
1.000
0.003
0.013
1.000
304.903
0.743
75.990
1.357
Age Group + Overlap
0.200
0.049
0.208
0.050
19.838
1.577
Comparison +
Age Group *
Comparison
Age Group
0.200
5.154e−4
0.002
4.827
0.207
0.389
Note. Dice coefficient by age group (between-subjects factor) and overlap comparison (within-subjects factor).
Prior model probability (P(M)); posterior model probability (P(M|data)); change from prior model odds to pos-
terior model odds (BFM); Bayes Factor for the Null model relative to current model (BF01); Bayes Factor for
current model relative to the Null model (BF10).
Figure 4. Penetrance maps showing homotopic activation consistency for each age group. Homotopic overlap maps (flipped right hemi-
sphere activity compared to left hemisphere activity) were merged for each participant in the respective age group at the Level 1 cutoff
(the top 3,311 and 2,588 voxels in the frontal and temporal regions respectively; ROI coverage displayed on the left). Darker blue areas reflect
regions of greater consistency in the localization of homotopic language activity, and lighter blue areas reflect greater variance.
Neurobiology of Language
376
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
3
3
6
4
2
0
2
3
4
0
3
n
o
_
a
_
0
0
0
6
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Weak shadow of language processing in the non-dominant hemisphere
as in adults there is indeed a spatially intact shadow of language activity in homotopic right
frontal and temporal regions. These results indicate that in the mature brain, tissue that once
supported language processing early in life may still be available to some degree in right hemi-
sphere regions.
Twenty-five years ago, Just et al. (1996) demonstrated that as the complexity of visually
presented sentences increases, the number of active voxels increases in left hemisphere
Broca’s and Wernicke’s areas—as well as in their mirror-image counterparts in the right
hemisphere of the healthy adult brain. The amount of activation in the right hemisphere frontal
and temporal regions was much lower than in the left, but the linguistic difficulty modulated
these regions in the same way. Just et al. (1996) called this observation a “weak echo” of
language processing in the right hemisphere regions that are homotopic to canonical language
areas. In the years since, no study to our knowledge has explicitly investigated the “weak
echo” observed by Just et al. in neurotypical adults, although some have replicated it inciden-
tally, as discussed above (e.g., Quillen et al., 2021). In the current work, we intended to bring
the concept of a “weak echo,” or a “weak shadow,” to center stage. By equalizing the volume
of activation in the left and right hemispheres, we find that the degree of symmetry between a
person’s right hemisphere activation and their left hemisphere activation is stable from child-
hood through adulthood. Further, the degree of right-left symmetry is greater within individuals
than the right-left symmetry across individuals of similar age, demonstrating that even idiosyn-
cratic aspects of language organization that vary across individuals are symmetrical above and
beyond typical rough symmetry that might be observed in group maps. These findings suggest
that the language system is organized asymmetrically only in the sense that the left hemisphere
response is stronger during language processing, but ignoring response magnitude, language
activation is spatially organized in homotopic regions even in the mature brain.
Mechanisms of Language Lateralization
In the context of evolution and ontogeny, it makes sense for language processing to recruit
regions symmetrically rather than asymmetrically. Mammals are generally symmetrical across
the left-to-right body axis, which is thought to be more adaptive than asymmetry: We move lin-
early through our environment with the support of symmetrical limbs that are controlled by the
bilateral somatomotor cortices, and our bilateral sensory systems allow us to detect and react to
incoming information from every angle (Concha et al., 2012; Corballis, 2009). Crucially, the
brain develops along four—not three—axes: anteroposterior, dorsoventral, and then each hemi-
sphere independently develops along its own mediolateral axis (Palmer, 2004), which generally
results in symmetrical structures. The sensory and motor systems develop symmetrically
(Corballis, 2009), and the language system depends on these systems for input and output.
Theoretically, then, language recruitment should be somewhat symmetrical. Our results indicate
that this may be the case, even in adults. Of course, other apparently lateralized systems may
also be activated homotopically in the mature brain. Here we simply claim that even the
language system, which is considered highly lateralized, shows substantial symmetry in the
mature brain regarding which areas within the two hemispheres respond to language.
At the same time, we know that, in terms of function, there is an asymmetry in the degree to
which adults rely on the hemispheres for language: Injury to left hemisphere regions, but not
right hemisphere regions, in adulthood renders many central aspects of language chronically
impaired. It is not well understood what properties of the left and right hemispheres create this
functional asymmetry for language processing. Prominent modern theories propose that audi-
tory neurons in the left and right hemispheres have slight biases for better processing of tem-
poral versus spectral properties of auditory information respectively, which leads speech
Neurobiology of Language
377
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
3
3
6
4
2
0
2
3
4
0
3
n
o
_
a
_
0
0
0
6
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Weak shadow of language processing in the non-dominant hemisphere
processing to be dominated by the left hemisphere while processing music and prosody are
dominated by the right hemisphere (Albouy et al., 2020; Zatorre et al., 1992; Zatorre & Belin,
2001). Asymmetries in the size or volume of particular brain structures have also, though with
notable inconsistency, been related to language lateralization, including the planum tempor-
ale (Foundas et al., 1994), Heschl’s gyrus (Penhune et al., 1996), posterior inferior frontal gyrus
(Dorsaint-Pierre et al., 2006), superior longitudinal fasciculus (Powell et al., 2006), and arcuate
fasciculus (Barrick et al., 2007). Sign languages in congenitally deaf individuals show the same
functional asymmetries (e.g., Newman et al., 2015), so these biases must apply across modal-
ities. All these theories focus on particular features that differ between the two hemispheres to
explain not only why language lateralizes but also why it lateralizes to the left hemisphere so
consistently. However, the theory that will best describe which properties of the left and right
hemispheres create this functional asymmetry for language processing will need also to
account for the evidence that lateralization is less strong—and is even reversible in extreme
circumstances—early in life. Based on the divergence in outcomes after injury in adulthood
versus early childhood, it is evident that certain properties of the neural correlates that perform
language processing must change over the course of development.
One of the signs of this maturational change is a decline in the activity magnitude of homo-
topic right hemisphere regions during language processing. This was shown by Olulade et al.
(2020) in the same sample included in the current study. It was previously unknown whether,
in addition to the magnitude declining, the spatial organization of this activity also disassem-
bles over the course of development—for example, because the right hemisphere regions that
were active during language processing early in life had perhaps become dedicated to other
cognitive functions. However, our results show that when one compares the same quantity of
activity in the left and right hemispheres, the activation is as homotopic in adults as it is in
children. We interpret this to mean that the same right hemisphere regions that are active dur-
ing language processing early in life are still responsive in adults, but very weakly.
Because language can develop successfully in homotopic right hemisphere regions after
early life injury to left hemisphere perisylvian cortex, it seems likely that there are language-
capable processors in homotopic right hemisphere regions early in development (Newport
et al., 2017, 2022). While some studies have argued that the best language outcomes after
early life stroke are supported by recruitment of intact left hemisphere tissue (Raja Beharelle
et al., 2010), this pattern of organization is generally observed in participants with periventri-
cular strokes that spared left hemisphere perisylvian cortical regions, and even in these cases,
the language system will sometimes recruit right perisylvian cortex (Staudt et al., 2002). Many
studies, including our own (Newport et al., 2017, 2022) have found right hemisphere activa-
tion and successful language even when no left hemisphere tissue is spared. The weak activity
we observe in homotopic right hemisphere areas in adults may be a weakened response of
these early-life language-capable processors.
What kinds of maturational changes might lead to a weaker response to language input?
The number of language-responsive neuronal populations may decline in these right hemi-
sphere regions, for example, through the normal developmental process of synaptic pruning
as a response to environmental experience. Or rather than being eliminated through pruning,
it is possible that cellular inhibition of these populations may increase throughout life (e.g.,
through transcallosal connections or more local lateral inhibition). A third possibility is a func-
tional rather than structural change: Language-responsive neuronal populations in these right
hemisphere regions may undergo a change in their stimulus response properties, such as
broadened tuning that leads to reduced specificity for the characteristics of language to which
they responded earlier in life. Developmental changes in connectivity may also engender
Neurobiology of Language
378
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
3
3
6
4
2
0
2
3
4
0
3
n
o
_
a
_
0
0
0
6
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Weak shadow of language processing in the non-dominant hemisphere
cerebral dominance for language processing. One study of two-day-old infants found strong
interhemispheric connectivity that contrasted with the relatively stronger intrahemispheric
connectivity observed in adults (Perani et al., 2011). In older children (age 7 and up) and
adults, strong intrahemispheric (primarily left hemisphere) connectivity outweighs the strength
of interhemispheric connectivity (Ailion et al., 2022; Mbwana et al., 2021). The weakening of
interhemispheric connections or of intrahemispheric right hemisphere connections early in life
may drive the later reduction of right hemisphere sentence processing activity over the course
of childhood. Whatever the mechanisms that drive developmental reductions in right hemi-
sphere activation, our findings constrain the possible mechanisms by demonstrating that while
the level of right hemisphere activity decreases during development, the overall organization
of bihemispheric regions involved in sentence processing remains symmetrical.
Rather than reflecting the presence of early-life language-capable processors, the weak lan-
guage response in the right hemisphere may be a weak echo of left hemisphere language acti-
vation, that is, co-activation with the dominant left hemisphere language regions that declines
in magnitude with age. This was the interpretation Just et al. (1996) made about their findings
in healthy adults. Brain imaging in adults who have their hemispheres disconnected either
surgically as a treatment for chronic epilepsy (split-brain) or by a stroke would potentially
elucidate whether weak but homotopically organized right hemisphere language activation
persists in the right hemisphere without transcallosal input from the left hemisphere. Unlike
middle cerebral artery stroke patients, these split brain patients retain both left and right hemi-
sphere areas for potential language processing, only without transcallosal communication.
However, published fMRI studies in such patients are strikingly scarce, and studies of language
fMRI tasks in these patients appear to be absent from the literature. Evidence from noninvasive
brain stimulation work in healthy adults has shown that phonological decision-making is
temporarily impaired by inhibiting either the left or right posterior inferior frontal gyrus
(Hartwigsen, Price, et al., 2010), or the left or right supramarginal gyrus (Hartwigsen,
Baumgaertner, et al., 2010). In addition, another study found that inhibiting left posterior
superior temporal sulcus led to an increase in activity in the homotopic right temporal area,
and improved speed in native versus foreign word recognition, though it is unclear whether
the behavioral improvement was driven by the decreased activation in the left and/or the
increased activation in the right temporal areas (Andoh & Paus, 2011). These findings each
suggest that the language-evoked activity in right hemisphere regions reflects language
processing and not merely a physiological echo of activity in the left. And, under certain
conditions (e.g., suppressed left hemisphere activity), these theoretical language-capable
processors in right hemisphere regions may become re-involved in language processing.
Implications for Recovery from Aphasia
After a left hemisphere stroke in adulthood, language impairments tend to be chronic. In
patients who have spared language areas in the left hemisphere, language processing con-
tinues to recruit these regions (however, see DeMarco et al., 2021), and in some cases also
homotopic right hemisphere regions (Turkeltaub et al., 2011). Some studies have argued that
better language outcomes after adult stroke depend on reengaging the intact left hemisphere
tissue (see Anglade et al., 2014, and Turkeltaub, 2015, for a review of language recovery in the
left versus right hemisphere after adult stroke). However, recovery of language functions after a
left hemisphere stroke clearly relies on homotopic right hemisphere regions in some individ-
uals: For example, several case reports have described individuals who acquired aphasia after
a left hemisphere stroke, recovered substantially, and then suffered a right hemisphere stroke
that caused a loss of recovered language abilities (Barlow, 1877; Basso et al., 1989; Turkeltaub
Neurobiology of Language
379
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
3
3
6
4
2
0
2
3
4
0
3
n
o
_
a
_
0
0
0
6
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Weak shadow of language processing in the non-dominant hemisphere
et al., 2012). This right hemisphere recruitment may reflect the reengagement of the right
hemisphere regions that were more strongly active during language processing early in life,
a prediction that is made by Newport et al.’s Developmental Origins Hypothesis (Newport
et al., 2017; see also Berl, Mayo, et al., 2014; Mbwana et al., 2009; Rosenberger et al.,
2009). However, the chronic persistence of severe aphasia in most adults with large left hemi-
sphere strokes demonstrates that the right hemisphere is often not capable of sustaining normal
language ability in adults after stroke. The present results suggest that even pre-injury, these
right hemisphere regions continue to be weakly active in adults during language processing.
An important question raised by this finding is: If there is a weak shadow of language process-
ing still intact in the right hemisphere, why can we not make use of it when we need to? One
caveat is that in the case of stroke, the injury is so sudden and abrupt that there is no time for
right hemisphere regions to transition back to processing language. In contrast, slow growing
tumors in the left hemisphere do in fact produce right hemisphere language recruitment and
better language outcomes compared to rapidly growing tumors (Thiel et al., 2006). If the weak
shadow of early life language processing in the right hemisphere can be reanimated, it may
need more time than a stroke allows.
Therapies that aim to increase right hemisphere activity in left hemisphere stroke patients to
improve language outcomes have reported some success. Intention therapy, for example,
trains patients to name pictures while they simultaneously perform complex movements with
their left hand that activate the contralateral right hemisphere (Crosson et al., 2007; Raymer
et al., 2002; Richards et al., 2002). Melodic intonation therapy trains patients to produce
common words and phrases by tapping out each syllable and producing exaggerated, melodic
prosody (Albert et al., 1973; Norton et al., 2009). A number of studies with small patient sam-
ple sizes have found that this combination of rhythmic motor and verbal exercises over a series
of treatment sessions facilitates improvements in language production in non-fluent patients
(Bonakdarpour et al., 2003; Schlaug et al., 2008; Van der Meulen et al., 2014; Wilson
et al., 2006; Zumbansen et al., 2014). The benefit of these approaches may be even greater
when brain stimulation is simultaneously applied: One study found that when MIT was paired
with excitatory transcranial direct current stimulation to the right posterior inferior frontal gyrus
for three consecutive days, stroke patients with moderate to severe non-fluent aphasia showed
significant improvements in speech fluency compared to when MIT was paired with sham
stimulation for the same duration in the same individuals (Vines et al., 2011). Such rhythmic-
melodic interventions have been associated with changes in right frontal and temporal regions
in adult stroke patients with aphasia (Schevenels et al., 2020; Schlaug et al., 2008; Wan et al.,
2014) though studies have also reported an increase in activation of residual left hemisphere
regions in some patients (Belin et al., 1996). It is possible that the spatially intact weak shadow
of early life language areas in right hemisphere frontal and temporal regions may be reengaged
by treatment approaches such as these in some left hemisphere adult patients with aphasia.
Our between-subjects consistency results also highlight the importance of accounting for
individual variability in the localization of this right hemisphere weak shadow for targeted
brain stimulation treatments. Future studies should examine the presence of a weak shadow
in older adults to confirm whether it is still present in this demographic, which is more similar
to the age range of when adults commonly have strokes (e.g., Szaflarski et al., 2006, but
without strict thresholding of activity magnitude).
Limitations
The approach that we used to calculate how homotopic language activity was in our partic-
ipants is limited to capturing active voxels that are directly parallel to one another—providing
Neurobiology of Language
380
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
3
3
6
4
2
0
2
3
4
0
3
n
o
_
a
_
0
0
0
6
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Weak shadow of language processing in the non-dominant hemisphere
a very conservative estimate of homotopicity. Other techniques could be used to make the
criterion for homotopicity more lenient, such as including right hemisphere activations that
fall within some Euclidean distance from the left hemisphere language centers. Also, our par-
ticipant group sizes were small (n = 10–14); a larger sample should be investigated to confirm
the robustness of results, and with other language tasks. Finally, our interpretations about the
localization of language-responsive brain regions are constrained by the univariate contrast of
the two task conditions we created (activation during speech minus activation during reverse
speech). Other analysis approaches that examine the patterns of response across voxels (mul-
tivariate methods) or direct measurement of the neural response using electrocorticography in
special clinical populations might provide a different perspective on how the participation of
right hemisphere regions in language processing changes over development.
Conclusions
We find that a weak shadow of early life language activity persists in homotopic regions of the
adult right hemisphere. These results indicate that while the strength of activity declines with
age in right frontotemporal regions, weak activity during language processing is still spatially
organized homotopic to core language centers. This finding is potentially important for lan-
guage recovery after a left hemisphere stroke in adulthood: Perhaps recovering some or all of
the early-life activation in these regions might be relevant to enhancing recovery of language
abilities in adults after stroke.
ACKNOWLEDGMENTS
This work was supported by funds from Georgetown University and MedStar Health; from the
Solomon James Rodan Pediatric Stroke Research Fund, the Feldstein Veron Innovation Fund,
and the Bergeron Visiting Scholars fund to the Center for Brain Plasticity and Recovery; by
NIH Grants K18DC014558 to ELN, K23NS065121 to MMB, R01NS244280 to WDG,
R01DC016902 to ELN and WDG, by a T32 to KM from NINDS 5T32NS041218 to Georgetown
University Center for Neural Injury and Recovery; and by M01RR020359 and P30HD040677 to
the IDDRC U54 HD090257 at Children’s National Health System and Georgetown University.
FUNDING INFORMATION
Kelly C. Martin, Georgetown University (https://dx.doi.org/10.13039/100008064). Elissa L.
Newport, Solomon James Rodan Pediatric Stroke Research Fund. Elissa L. Newport, Feldstein
Veron Innovation Fund. Elissa L. Newport, Bergeron Visiting Scholars Fund. Elissa L. Newport,
National Institutes of Health (https://dx.doi.org/10.13039/100000002), Award ID:
K18DC014558. Madison M. Berl, National Institutes of Health (https://dx.doi.org/10.13039
/100000002), Award ID: K23NS065121. William D. Gaillard, National Institutes of Health
(https://dx.doi.org/10.13039/100000002), Award ID: R01NS244280. Elissa L. Newport,
National Institutes of Health (https://dx.doi.org/10.13039/100000002), Award ID:
R01DC016902. William D. Gaillard, National Institutes of Health (https://dx.doi.org/10.13039
/100000002), Award ID: R01DC016902. Kelly C. Martin, National Institute of Neurological
Disorders and Stroke (https://dx.doi.org/10.13039/100000065), Award ID: 5T32NS041218.
The Intellectual and Developmental Disabilities Research Center U54 HD090257 at the
Children’s National Health System (which also includes Georgetown University), National
Institutes of Health (https://dx.doi.org/10.13039/100000002), Award ID: M01RR020359. The
Intellectual and Developmental Disabilities Research Center U54 HD090257 at the Children’s
National Health System (which also includes Georgetown University), National Institutes of
Health (https://dx.doi.org/10.13039/100000002), Award ID: P30HD040677.
Neurobiology of Language
381
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
3
3
6
4
2
0
2
3
4
0
3
n
o
_
a
_
0
0
0
6
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Weak shadow of language processing in the non-dominant hemisphere
AUTHOR CONTRIBUTIONS
Kelly C. Martin: Conceptualization: Supporting; Formal analysis: Lead; Funding acquisition:
Supporting; Investigation: Lead; Methodology: Equal; Visualization: Lead; Writing – original
draft: Lead; Writing – review & editing: Lead. Anna Seydell-Greenwald: Conceptualization:
Supporting; Data curation: Lead; Investigation: Supporting; Methodology: Equal; Visualization:
Supporting; Writing – original draft: Supporting; Writing – review & editing: Supporting.
Madison M. Berl: Funding acquisition: Supporting; Resources: Supporting; Writing – review
& editing: Supporting. William D. Gaillard: Funding acquisition: Supporting; Writing – review
& editing: Supporting. Peter E. Turkeltaub: Conceptualization: Equal; Investigation: Equal;
Methodology: Equal; Supervision: Equal; Visualization: Supporting; Writing – original draft:
Supporting; Writing – review & editing: Supporting. Elissa L. Newport: Conceptualization:
Equal; Funding acquisition: Supporting; Investigation: Equal; Supervision: Equal; Visualization:
Supporting; Writing – original draft: Supporting; Writing – review & editing: Supporting.
REFERENCES
Ailion, A. S., You, X., Mbwana, J. S., Fanto, E. J., Krishnamurthy, M.,
Vaidya, C. J., Sepeta, L. N., Gaillard, W. D., & Berl, M. M. (2022).
Functional connectivity as a potential mechanism for language
plasticity. Neurology, 98(3), e249–e259. https://doi.org/10.1212
/WNL.0000000000013071, PubMed: 34795045
Albert, M. L., Sparks, R. W., & Helm, N. A. (1973). Melodic intona-
tion therapy for aphasia. Archives of Neurology, 29(2), 130–131.
https://doi.org/10.1001/archneur.1973.00490260074018,
PubMed: 4717723
Albouy, P., Benjamin, L., Morillon, B., & Zatorre, R. J. (2020).
Distinct sensitivity to spectrotemporal modulation supports
brain asymmetry for speech and melody. Science, 367(6481),
1043–1047. https://doi.org/10.1126/science.aaz3468, PubMed:
32108113
Andoh, J., & Paus, T. (2011). Combining functional neuroimaging with
off-line brain stimulation: Modulation of task-related activity in lan-
guage areas. Journal of Cognitive Neuroscience, 23(2), 349–361.
https://doi.org/10.1162/jocn.2010.21449, PubMed: 20146606
Anglade, C., Thiel, A., & Ansaldo, A. I. (2014). The complementary
role of the cerebral hemispheres in recovery from aphasia after
stroke: A critical review of literature. Brain Injury, 28(2),
138–145. https://doi.org/10.3109/02699052.2013.859734,
PubMed: 24456053
Barlow, T. (1877). On a case of double hemiplegia, with cerebral
symmetrical lesions. British Medical Journal, 2(865), 103–104.
https://doi.org/10.1136/bmj.2.865.103, PubMed: 20748595
Barrick, T. R., Lawes, I. N., Mackay, C. E., & Clark, C. A. (2007).
White matter pathway asymmetry underlies functional lateraliza-
tion. Cerebral Cortex, 17(3), 591–598. https://doi.org/10.1093
/cercor/bhk004, PubMed: 16627859
Basso, A., Gardelli, M., Grassi, M. P., & Mariotti, M. (1989). The
role of the right hemisphere in recovery from aphasia. Two case
studies. Cortex, 25(4), 555–566. https://doi.org/10.1016/S0010
-9452(89)80017-6, PubMed: 2612175
Belin, P., Zilbovicius, M., Remy, P., Francois, C., Guillaume, S.,
Chain, F., Rancurel, G., & Samson, Y. (1996). Recovery from non-
fluent aphasia after melodic intonation therapy: A PET study.
Neurology, 47(6), 1504–1511. https://doi.org/10.1212/ WNL.47
.6.1504, PubMed: 8960735
Berl, M. M., Mayo, J., Parks, E. N., Rosenberger, L. R., VanMeter, J.,
Ratner, N. B., Vaidya, C. J., & Gaillard, W. D. (2014). Regional
differences in the developmental trajectory of lateralization of the
language network. Human Brain Mapping, 35(1), 270–284.
https://doi.org/10.1002/hbm.22179, PubMed: 23033058
Berl, M. M., Zimmaro, L. A., Khan, O. I., Dustin, I., Ritzl, E., Duke,
E. S., Sepeta, L. N., Sato, S., Theodore, W. H., & Gaillard, W. D.
(2014). Characterization of atypical language activation patterns
in focal epilepsy. Annals of Neurology, 75(1), 33–42. https://doi
.org/10.1002/ana.24015, PubMed: 24038442
Boatman, D., Freeman, J., Vining, E., Pulsifer, M., Miglioretti, D.,
Minahan, R., Carson, B., Brandt, J., & McKhann, G. (1999). Lan-
guage recovery after left hemispherectomy in children with
late-onset seizures. Annals of Neurology, 46(4), 579–586.
https://doi.org/10.1002/1531-8249(199910)46:4<579::AID
-ANA5>3.0.CO;2-K, PubMed: 10514094
Bonakdarpour, B., Eftekharzadeh, A., & Ashayeri, H. (2003).
Melodic intonation therapy in Persian aphasic patients. Aphasiol-
ogy, 17(1), 75–95. https://doi.org/10.1080/729254891
Broca, P. (1861). Remarks on the seat of the faculty of articulated
language, following an observation of aphemia (loss of speech).
Bulletin de La Société Anatomique, 6, 330–357. https://garfield
.library.upenn.edu/classics1990/A1990DY08800001.pdf
Bulteau, C., Grosmaitre, C., Save-Pédebos, J., Leunen, D.,
Delalande, O., Dorfmüller, G., Dulac, O., & Jambaqué, I.
(2015). Language recovery after left hemispherotomy for Rasmus-
sen encephalitis. Epilepsy & Behavior, 53, 51–57. https://doi.org
/10.1016/j.yebeh.2015.07.044, PubMed: 26519666
Burgund, E. D., Kang, H. C., Kelly, J. E., Buckner, R. L., Snyder,
A. Z., Petersen, S. E., & Schlaggar, B. L. (2002). The feasibility
of a common stereotactic space for children and adults in fMRI
studies of development. NeuroImage, 17(1), 184–200. https://doi
.org/10.1006/nimg.2002.1174, PubMed: 12482076
Concha, M. L., Bianco, I. H., & Wilson, S. W. (2012). Encoding
asymmetry within neural circuits. Nature Reviews Neuroscience,
13(12), 832–843. https://doi.org/10.1038/nrn3371, PubMed:
23165260
Corballis, M. C. (2009). The evolution and genetics of cerebral
asymmetry. Philosophical Transactions of the Royal Society B:
Biological Sciences, 364(1519), 867–879. https://doi.org/10
.1098/rstb.2008.0232, PubMed: 19064358
Crosson, B., Fabrizio, K. S., Singletary, F., Cato, M. A., Wierenga,
C. E., Parkinson, R. B., Sherod, M. E., Moore, A. B., Ciampitti, M.,
Neurobiology of Language
382
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
3
3
6
4
2
0
2
3
4
0
3
n
o
_
a
_
0
0
0
6
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Weak shadow of language processing in the non-dominant hemisphere
Holiway, B., Leon, S., Rodriguez, A., Kendall, D. L., Levy, I. F., &
Gonzalez Rothi, L. J. (2007). Treatment of naming in nonfluent
aphasia through manipulation of intention and attention: A
phase 1 comparison of two novel treatments. Journal of the Inter-
national Neuropsychological Society, 13(4), 582–594. https://doi
.org/10.1017/S1355617707070737, PubMed: 17521480
DeMarco, A. T., van der Stelt, C. M., Paul, S., Dvorak, E., Lacey, E.,
Snider, S., & Turkeltaub, P. E. (2021). Testing the perilesional neu-
roplastic recruitment hypothesis in aphasia. BioRxiv. https://doi
.org/10.1101/2021.08.06.455431
Dorsaint-Pierre, R., Penhune, V. B., Watkins, K. E., Neelin, P., Lerch,
J. P., Bouffard, M., & Zatorre, R. J. (2006). Asymmetries of the pla-
num temporale and Heschl’s gyrus: Relationship to language lat-
eralization. Brain, 129(5), 1164–1176. https://doi.org/10.1093
/brain/awl055, PubMed: 16537567
Fair, D. A., Choi, A. H., Dosenbach, Y. B. L., Coalson, R. S., Miezin,
F. M., Petersen, S. E., & Schlaggar, B. L. (2010). The functional
organization of trial-related activity in lexical processing after
early left hemispheric brain lesions: An event-related fMRI study.
Brain and Language, 114(2), 135–146. https://doi.org/10.1016/j
.bandl.2009.09.001, PubMed: 19819000
Fedorenko, E., & Kanwisher, N. (2009). Neuroimaging of language:
Why hasn’t a clearer picture emerged? Language and Linguistics
Compass, 3(4), 839–865. https://doi.org/10.1111/j.1749-818X
.2009.00143.x
Foundas, A. L., Leonard, C. M., Gilmore, R., Fennell, E., & Heilman,
K. M. (1994). Planum temporale asymmetry and language
dominance. Neuropsychologia, 32(10), 1225–1231. https://doi
.org/10.1016/0028-3932(94)90104-X, PubMed: 7845562
Gaillard, W. D., Berl, M. M., Moore, E. N., Ritzl, E. K., Rosenberger,
L. R., Weinstein, S. L., Conry, J. A., Pearl, P. L., Ritter, F. F., &
Sato, S. (2007). Atypical language in lesional and nonlesional
complex partial epilepsy. Neurology, 69(18), 1761–1771.
https://doi.org/10.1212/01.wnl.0000289650.48830.1a,
PubMed: 17967992
Gaillard, W. D., Hertz-Pannier, L., Mott, S. H., Barnett, A. S.,
LeBihan, D., & Theodore, W. H. (2000). Functional anatomy of
cognitive development: fMRI of verbal fluency in children and
adults. Neurology, 54(1), 180–180. https://doi.org/10.1212
/ WNL.54.1.180, PubMed: 10636145
Ghosh, S. S., Kakunoori, S., Augustinack, J., Nieto-Castanon, A.,
Kovelman, I., Gaab, N., Christodoulou, J. A., Triantafyllou, C.,
Gabrieli, J. D. E., & Fischl, B. (2010). Evaluating the validity of
volume-based and surface-based brain image registration for
developmental cognitive neuroscience studies in children 4 to
11 years of age. NeuroImage, 53(1), 85–93. https://doi.org/10
.1016/j.neuroimage.2010.05.075, PubMed: 20621657
Guzzetta, A., Pecini, C., Biagi, L., Tosetti, M., Brizzolara, D.,
Chilosi, A., Cipriani, P., Petacchi, E., & Cioni, G. (2008). Language
organisation in left perinatal stroke. Neuropediatrics, 39(3),
157–163. https://doi.org/10.1055/s-0028-1085465, PubMed:
18991195
Hartwigsen, G., Baumgaertner, A., Price, C. J., Koehnke, M., Ulmer,
S., & Siebner, H. R. (2010). Phonological decisions require both
the left and right supramarginal gyri. Proceedings of the National
Academy of Sciences, 107(38), 16494–16499. https://doi.org/10
.1073/pnas.1008121107, PubMed: 20807747
Hartwigsen, G., Price, C. J., Baumgaertner, A., Geiss, G., Koehnke,
M., Ulmer, S., & Siebner, H. R. (2010). The right posterior inferior
frontal gyrus contributes to phonological word decisions in the
healthy brain: Evidence from dual-site TMS. Neuropsychologia,
48(10), 3155–3163. https://doi.org/10.1016/j.neuropsychologia
.2010.06.032, PubMed: 20600177
Holland, S. K., Vannest, J., Mecoli, M., Jacola, L. M., Tillema, J.-M.,
Karunanayaka, P. R., Schmithorst, V. J., Yuan, W., Plante, E., & Byars,
A. W. (2007). Functional MRI of language lateralization during
development in children. International Journal of Audiology,
46(9), 533–551. https://doi.org/10.1080/14992020701448994,
PubMed: 17828669
Ilves, P., Tomberg, T., Kepler, J., Laugesaar, R., Kaldoja, M.-L., Kepler,
K., & Kolk, A. (2014). Different plasticity patterns of language
function in children with perinatal and childhood stroke. Journal
of Child Neurology, 29(6), 756–764. https://doi.org/10.1177
/0883073813489350, PubMed: 23748202
Just, M. A., Carpenter, P. A., Keller, T. A., Eddy, W. F., & Thulborn,
K. R. (1996). Brain activation modulated by sentence compre-
hension. Science, 274(5284), 114–116. https://doi.org/10.1126
/science.274.5284.114, PubMed: 8810246
Knecht, S., Deppe, M., Dräger, B., Bobe, L., Lohmann, H.,
Ringelstein, E. B., & Henningsen, H. (2000). Language lateraliza-
tion in healthy right-handers. Brain, 123(1), 74–81. https://doi.org
/10.1093/brain/123.1.74, PubMed: 10611122
Knecht, S., Dräger, B., Deppe, M., Bobe, L., Lohmann, H., Flöel, A.,
Ringelstein, E.-B., & Henningsen, H. (2000). Handedness and
hemispheric language dominance in healthy humans. Brain,
123(12), 2512–2518. https://doi.org/10.1093/ brain/123.12
.2512, PubMed: 11099452
Lenneberg, E. H. (1967). The biological foundations of language.
Hospital Practice, 2(12), 59–67. https://doi.org/10.1080
/21548331.1967.11707799
Lenneberg, E. H. (1969). On Explaining Language: The development
of language in children can best be understood in the context of
developmental biology. Science, 164(3880), 635–643. https://doi
.org/10.1126/science.164.3880.635, PubMed: 5813477
Liégeois, F., Connelly, A., Baldeweg, T., & Vargha-Khadem, F.
(2008). Speaking with a single cerebral hemisphere: fMRI lan-
guage organization after hemispherectomy in childhood. Brain
and Language, 106(3), 195–203. https://doi.org/10.1016/j.bandl
.2008.01.010, PubMed: 18329093
Liégeois, F., Cross, J. H., Polkey, C., Harkness, W., & Vargha-
Khadem, F. (2008). Language after hemispherectomy in childhood:
Contributions from memory and intelligence. Neuropsychologia,
46(13), 3101–3107. https://doi.org/10.1016/j.neuropsychologia
.2008.07.001, PubMed: 18657558
Martin, K. C., Ketchabaw, W. T., & Turkeltaub, P. E. (2022). Plastic-
ity of the language system in children and adults. Handbook of
Clinical Neurology, 184, 397–414. https://doi.org/10.1016/B978
-0-12-819410-2.00021-7, PubMed: 35034751
Mbwana, J., Berl, M., Ritzl, E., Rosenberger, L., Mayo, J., Weinstein,
S., Conry, J., Pearl, P., Shamim, S., & Moore, E. (2009). Limita-
tions to plasticity of language network reorganization in localiza-
tion related epilepsy. Brain, 132(2), 347–356. https://doi.org/10
.1093/brain/awn329, PubMed: 19059978
Mbwana, J. S., You, X., Ailion, A., Fanto, E. J., Krishnamurthy, M.,
Sepeta, L. N., Newport, E. L., Vaidya, C. J., Berl, M. M., &
Gaillard, W. D. (2021). Functional connectivity hemispheric
contrast (FC-HC): A new metric for language mapping. Neuro-
Image: Clinical, 30, Article 102598. https://doi.org/10.1016/j
.nicl.2021.102598, PubMed: 33858809
Mollica, F., Siegelman, M., Diachek, E., Piantadosi, S. T., Mineroff,
Z., Futrell, R., Kean, H., Qian, P., & Fedorenko, E. (2020). Com-
position is the core driver of the language-selective network.
Neurobiology of Language, 1(1), 104–134. https://doi.org/10
.1162/nol_a_00005
Newman, A. J., Supalla, T., Fernandez, N., Newport, E. L., &
Bavelier, D. (2015). Neural systems supporting linguistic structure,
Neurobiology of Language
383
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
3
3
6
4
2
0
2
3
4
0
3
n
o
_
a
_
0
0
0
6
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Weak shadow of language processing in the non-dominant hemisphere
linguistic experience, and symbolic communication in sign lan-
guage and gesture. Proceedings of the National Academy of Sci-
ences, 112(37), 11684–11689. https://doi.org/10.1073/pnas
.1510527112, PubMed: 26283352
Newport, E. L., Landau, B., Seydell-Greenwald, A., Turkeltaub,
P. E., Chambers, C. E., Dromerick, A. W., Carpenter, J., Berl,
M. M., & Gaillard, W. D. (2017). Revisiting Lenneberg’s hypoth-
eses about early developmental plasticity: Language organiza-
tion after left-hemisphere perinatal stroke. Biolinguistics, 11,
407–422. https://www.biolinguistics.eu/index.php/biolinguistics
/article/view/597. https://doi.org/10.5964/bioling.9105,
PubMed: 30556058
Newport, E. L., Seydell-Greenwald, A., Landau, B., Turkeltaub,
P. E., Chambers, C. E., Martin, K. C., Rennert, R., Giannetti, M.,
Dromerick, A. W., Carpenter, J. L., Berl, M. M., & Gaillard, W. D.
(2022). Language and developmental plasticity after perinatal
stroke [Manuscript in preparation]. Departments of Neurology and
Rehabilitation Medicine, Georgetown University Medical Center.
Norton, A., Zipse, L., Marchina, S., & Schlaug, G. (2009). Melodic
intonation therapy: Shared insights on how it is done and why it
might help. Annals of the New York Academy of Sciences,
1169(1), 431–436. https://doi.org/10.1111/j.1749-6632.2009
.04859.x, PubMed: 19673819
Olulade, O. A., Seydell-Greenwald, A., Chambers, C. E.,
Turkeltaub, P. E., Dromerick, A. W., Berl, M. M., Gaillard,
W. D., & Newport, E. L. (2020). The neural basis of language
development: Changes in lateralization over age. Proceedings
of the National Academy of Sciences, 117(38), 23477–23483.
https://doi.org/10.1073/pnas.1905590117, PubMed: 32900940
Palmer, A. R. (2004). Symmetry breaking and the evolution of
development. Science, 306(5697), 828–833. https://doi.org/10
.1126/science.1103707, PubMed: 15514148
Peña, M., Maki, A., Kovačić, D., Dehaene-Lambertz, G., Koizumi,
H., Bouquet, F., & Mehler, J. (2003). Sounds and silence: An
optical topography study of language recognition at birth.
Proceedings of the National Academy of Sciences, 100(20),
11702–11705. https://doi.org/10.1073/pnas.1934290100,
PubMed: 14500906
Penhune, V., Zatorre, R., MacDonald, J., & Evans, A. (1996). Inter-
hemispheric anatomical differences in human primary auditory
cortex: Probabilistic mapping and volume measurement from
magnetic resonance scans. Cerebral Cortex, 6(5), 661–672.
https://doi.org/10.1093/cercor/6.5.661, PubMed: 8921202
Perani, D., Saccuman, M. C., Scifo, P., Anwander, A., Spada, D.,
Baldoli, C., Poloniato, A., Lohmann, G., & Friederici, A. D.
(2011). Neural language networks at birth. Proceedings of the
National Academy of Sciences, 108(38), 16056–16061. https://
doi.org/10.1073/pnas.1102991108, PubMed: 21896765
Powell, H. R., Parker, G. J., Alexander, D. C., Symms, M. R.,
Boulby, P. A., Wheeler-Kingshott, C. A., Barker, G. J., Noppeney,
U., Koepp, M. J., & Duncan, J. S. (2006). Hemispheric asymme-
tries in language-related pathways: A combined functional MRI
and tractography study. NeuroImage, 32(1), 388–399. https://doi
.org/10.1016/j.neuroimage.2006.03.011, PubMed: 16632380
Quillen, I. A., Yen, M., & Wilson, S. M. (2021). Distinct neural cor-
relates of linguistic and non-linguistic demand. Neurobiology of
Language, 2(2), 202–225. https://doi.org/10.1162/nol_a_00031,
PubMed: 34585141
Raja Beharelle, A., Dick, A. S., Josse, G., Solodkin, A., Huttenlocher,
P. R., Levine, S. C., & Small, S. L. (2010). Left hemisphere
regions are critical for language in the face of early left focal
brain injury. Brain, 133(6), 1707–1716. https://doi.org/10.1093
/brain/awq104, PubMed: 20466762
Raymer, A. M., Rowland, L., Haley, M., & Crosson, B. (2002). Non-
symbolic movement training to improve sentence generation in
transcortical motor aphasia: A case study. Aphasiology, 16(4–6),
493–506. https://doi.org/10.1080/02687030244000239
Richards, K., Singletary, F., Rothi, L. J. G., Koehler, S., & Crosson, B.
(2002). Activation of intentional mechanisms through utilization
of nonsymbolic movements in aphasia rehabilitation. Journal of
Rehabilitation Research and Development, 39(4), 445–454.
https://www.rehab.research.va.gov/jour/02/39/4/richards.html.
PubMed: 17638142
Rosenberger, L. R., Zeck, J., Berl, M. M., Moore, E. N., Ritzl, E. K.,
Shamim, S., Weinstein, S. L., Conry, J. A., Pearl, P. L., Sato, S.,
Vezina, L. G., Theodore, W. H., & Gaillard, W. D. (2009). Inter-
hemispheric and intrahemispheric language reorganization in
complex partial epilepsy. Neurology, 72(21), 1830–1836. https://
doi.org/10.1212/ WNL.0b013e3181a7114b, PubMed: 19470965
Schevenels, K., Price, C. J., Zink, I., De Smedt, B., & Vandermosten,
M. (2020). A review on treatment-related brain changes in apha-
sia. Neurobiology of Language, 1(4), 402–433. https://doi.org/10
.1162/nol_a_00019
Schlaug, G., Marchina, S., & Norton, A. (2008). From singing to
speaking: Why singing may lead to recovery of expressive lan-
guage function in patients with Broca’s aphasia. Music Percep-
tion, 25(4), 315–323. https://doi.org/10.1525/mp.2008.25.4
.315, PubMed: 21197418
Shain, C., Blank, I. A., van Schijndel, M., Schuler, W., & Fedorenko,
E. (2020). fMRI reveals language-specific predictive coding dur-
ing naturalistic sentence comprehension. Neuropsychologia,
138, Article 107307. https://doi.org/10.1016/j.neuropsychologia
.2019.107307, PubMed: 31874149
Skipper-Kallal, L. M., Lacey, E. H., Xing, S., & Turkeltaub, P. E.
(2017a). Right hemisphere remapping of naming functions
depends on lesion size and location in poststroke aphasia. Neu-
roplasticity of Language Networks, 2017, Article 8740353.
https://doi.org/10.1155/2017/8740353, PubMed: 28168061
Skipper-Kallal, L. M., Lacey, E. H., Xing, S., & Turkeltaub, P. E.
(2017b). Functional activation independently contributes to
naming ability and relates to lesion site in post-stroke aphasia.
Human Brain Mapping, 38(4), 2051–2066. https://doi.org/10
.1002/hbm.23504, PubMed: 28083891
Staudt, M., Grodd, W., Niemann, G., Wildgruber, D., Erb, M., &
Krägeloh–Mann, I. (2001). Early left periventricular brain lesions
induce right hemispheric organization of speech. Neurology,
57(1), 122–125. https://doi.org/10.1212/ WNL.57.1.122,
PubMed: 11445639
Staudt, M., Lidzba, K., Grodd, W., Wildgruber, D., Erb, M., &
Krägeloh-Mann, I. (2002). Right-hemispheric organization of
language following early left-sided brain lesions: Functional
MRI topography. NeuroImage, 16(4), 954–967. https://doi.org
/10.1006/nimg.2002.1108, PubMed: 12202083
Stiles, J., Reilly, J. S., Levine, S. C., Trauner, D. A., & Nass, R. (2012).
Neural plasticity and cognitive development: Insights from chil-
dren with perinatal brain injury. Oxford University Press.
https://doi.org/10.1093/acprof:osobl/9780195389944.001.0001
Szaflarski, J. P., Holland, S. K., Schmithorst, V. J., & Byars, A. W.
(2006). fMRI study of language lateralization in children and
adults. Human Brain Mapping, 27(3), 202–212. https://doi.org
/10.1002/hbm.20177, PubMed: 16035047
Thiel, A., Habedank, B., Herholz, K., Kessler, J., Winhuisen, L.,
Haupt, W. F., & Heiss, W.-D. (2006). From the left to the right:
How the brain compensates progressive loss of language func-
tion. Brain and Language, 98(1), 57–65. https://doi.org/10.1016
/j.bandl.2006.01.007, PubMed: 16519926
Neurobiology of Language
384
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
3
3
6
4
2
0
2
3
4
0
3
n
o
_
a
_
0
0
0
6
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Weak shadow of language processing in the non-dominant hemisphere
Tillema, J. M., Byars, A. W., Jacola, L. M., Schapiro, M. B.,
Schmithorst, V. J., Szaflarski, J. P., & Holland, S. K. (2008). Reprint
of “Cortical reorganization of language functioning following
perinatal left MCA stroke” [Brain and Language 105 (2008),
99–111]. Brain and Language, 106(3), 184–194. https://doi.org
/10.1016/j.bandl.2008.08.001, PubMed: 18762059
Turkeltaub, P. E. (2015). Brain stimulation and the role of the right
hemisphere in aphasia recovery. Current Neurology and Neuro-
science Reports, 15(11), Article 72. https://doi.org/10.1007
/s11910-015-0593-6, PubMed: 26396038
Turkeltaub, P. E., Coslett, H. B., Thomas, A. L., Faseyitan, O.,
Benson, J., Norise, C., & Hamilton, R. H. (2012). The right
hemisphere is not unitary in its role in aphasia recovery. Cortex,
48(9), 1179–1186. https://doi.org/10.1016/j.cortex.2011.06.010,
PubMed: 21794852
Turkeltaub, P. E., Messing, S., Norise, C., & Hamilton, R. H. (2011).
Are networks for residual language function and recovery consis-
tent across aphasic patients? Neurology, 76(20), 1726–1734.
https://doi.org/10.1212/ WNL.0b013e31821a44c1, PubMed:
21576689
Van der Meulen, I., van de Sandt-Koenderman, W. M. E.,
Heijenbrok-Kal, M. H., Visch-Brink, E. G., & Ribbers, G. M.
(2014). The efficacy and timing of melodic intonation therapy
in subacute aphasia. Neurorehabilitation and Neural Repair,
28(6), 536–544. https://doi.org/10.1177/1545968313517753,
PubMed: 24449708
Vines, B. W., Norton, A. C., & Schlaug, G. (2011). Non-invasive
brain stimulation enhances the effects of melodic intonation
therapy. Frontiers in Psychology, 2, Article 230. https://doi.org
/10.3389/fpsyg.2011.00230, PubMed: 21980313
Wan, C. Y., Zheng, X., Marchina, S., Norton, A., & Schlaug, G.
(2014). Intensive therapy induces contralateral white matter
changes in chronic stroke patients with Broca’s aphasia. Brain
and Language, 136, 1–7. https://doi.org/10.1016/j.bandl.2014
.03.011, PubMed: 25041868
Wilson, S. J., Parsons, K., & Reutens, D. C. (2006). Preserved sing-
ing in aphasia: A case study of the efficacy of melodic intonation
therapy. Music Perception, 24(1), 23–36. https://doi.org/10.1525
/mp.2006.24.1.23
Yarkoni, T., Poldrack, R. A., Nichols, T. E., Van Essen, D. C., &
Wager, T. D. (2011). Large-scale automated synthesis of human
functional neuroimaging data. Nature Methods, 8(8), 665–670.
https://doi.org/10.1038/nmeth.1635, PubMed: 21706013
Zatorre, R. J., & Belin, P. (2001). Spectral and temporal processing
in human auditory cortex. Cerebral Cortex, 11(10), 946–953.
https://doi.org/10.1093/cercor/11.10.946, PubMed: 11549617
Zatorre, R. J., Evans, A. C., Meyer, E., & Gjedde, A. (1992). Later-
alization of phonetic and pitch discrimination in speech process-
ing. Science, 256(5058), 846–849. https://doi.org/10.1126
/science.256.5058.846, PubMed: 1589767
Zumbansen, A., Peretz,
(2014). The combina-
I., & Hébert, S.
tion of rhythm and pitch can account for the beneficial effect
of melodic intonation therapy on connected speech improve-
ments in Broca’s aphasia. Frontiers in Human Neuroscience, 8,
Article 592. https://doi.org/10.3389/fnhum.2014.00592,
PubMed: 25157222
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
3
3
6
4
2
0
2
3
4
0
3
n
o
_
a
_
0
0
0
6
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neurobiology of Language
385