RESEARCH ARTICLE
Language Experience Impacts Brain Activation for
Spoken and Signed Language in Infancy: Insights
From Unimodal and Bimodal Bilinguals
Evelyne Mercure1,2,3
, Samuel Evans2,4
, Laura Pirazzoli3,5, Laura Goldberg2,
Harriet Bowden-Howl2,6, Kimberley Coulson-Thaker2,7, Indie Beedie2,
, and Mairéad MacSweeney2
, Mark H. Johnson3,8
Sarah Lloyd-Fox3,8
1Goldsmiths, University of London, London, UK
2University College London, London, UK
3Birkbeck – University of London, London, UK
4University of Westminster, London, UK
5Boston Children’s Hospital, Boston, Massachusetts, US
6University of Plymouth, Plymouth, Devon, UK
7University of Hertfordshire, Hatfield, Hertforshire, UK
8University of Cambridge, Cambridge, Cambridgeshire, UK
Keywords: deaf, fNIRS, infants, sign language, speech, infant-directed language
ABSTRACT
Recent neuroimaging studies suggest that monolingual infants activate a left-lateralized
frontotemporal brain network in response to spoken language, which is similar to the network
involved in processing spoken and signed language in adulthood. However, it is unclear how brain
activation to language is influenced by early experience in infancy. To address this question, we
present functional near-infrared spectroscopy (fNIRS) data from 60 hearing infants (4 to 8 months
of age): 19 monolingual infants exposed to English, 20 unimodal bilingual infants exposed to
two spoken languages, and 21 bimodal bilingual infants exposed to English and British Sign
Language (BSL). Across all infants, spoken language elicited activation in a bilateral brain
network including the inferior frontal and posterior temporal areas, whereas sign language elicited
activation in the right temporoparietal area. A significant difference in brain lateralization was
observed between groups. Activation in the posterior temporal region was not lateralized in
monolinguals and bimodal bilinguals, but right lateralized in response to both language modalities
in unimodal bilinguals. This suggests that the experience of two spoken languages influences brain
activation for sign language when experienced for the first time. Multivariate pattern analyses
(MVPAs) could classify distributed patterns of activation within the left hemisphere for spoken and
signed language in monolinguals (proportion correct = 0.68; p = 0.039) but not in unimodal or
bimodal bilinguals. These results suggest that bilingual experience in infancy influences brain
activation for language and that unimodal bilingual experience has greater impact on early brain
lateralization than bimodal bilingual experience.
INTRODUCTION
Areas of the frontal and temporal cortex are crucial to language processing in adulthood.
These regions are already activated in response to spoken language in the first few days or
weeks of life (Altvater-Mackensen & Grossmann, 2016; Dehaene-Lambertz, Dehaene, &
Hertz-Pannier, 2002; Dehaene-Lambertz et al., 2006, 2010; May, Gervain, Carreiras, &
a n o p e n a c c e s s
j o u r n a l
Citation: Mercure, E., Evans, S.,
Pirazzoli, L., Goldberg, L., Bowden-
Howl, H., Coulson-Thaker, K., …
MacSweeney, M. (2020). Language
experience impacts brain activation for
spoken and signed language in
infancy: Insights from unimodal and
bimodal bilinguals. Neurobiology of
Language, 1(1), 9–32. https://doi.org/
10.1162/nol_a_00001
DOI:
https://doi.org/10.1162/nol_a_00001
Supporting Information:
https://doi.org/10.1162/nol_a_00001
Received: 4 April 2019
Accepted: 9 September 2019
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Author:
Evelyne Mercure
e.mercure@gold.ac.uk
Handling Editor:
Kate Watkins
Copyright: © 2019
Massachusetts Institute of Technology
Published under a Creative Commons
Attribution 4.0 International
(CC BY 4.0) license.
The MIT Press
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
1
1
9
1
8
6
7
9
0
9
n
o
_
a
_
0
0
0
0
1
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Experience impacts brain activation for language in infancy
Werker, 2018; Minagawa-Kawai et al., 2010; Pena et al., 2003; Perani et al., 2011; Sato et al.,
2012; Shultz, Vouloumanos, Bennett, & Pelphrey, 2014; Vannasing et al., 2016). As in adults,
brain responses to speech in infants are often found to be greater in amplitude in the left than
in the right hemisphere (Altvater-Mackensen & Grossmann, 2016; Dehaene-Lambertz et al.,
2002, 2010; Minagawa-Kawai et al., 2010; Pena et al., 2003; Shultz et al., 2014; Vannasing
et al., 2016), but this left lateralization is not always observed in infants (Dehaene-Lambertz
et al., 2006; May, Byers-Heinlein, Gervain, & Werker, 2011; May et al., 2018; Perani et al.,
2011).
The similarities between the adult and infant language networks suggest an early neural
specialization for language processing in human development. However, the presence of this
pattern from birth does not necessarily imply that it is established in the absence of experience.
A fetus can hear from the 24th to 25th gestational week (Birnholz & Benacerraf, 1983). The
preference of neonates for their mother’s voice and the language heard in utero indicate that
fetuses use the sounds and vibrations of their mother’s voice to begin learning the foundations
of voice and language processing prenatally (DeCasper & Fifer, 1980; Moon, Cooper, & Fifer,
1993). Even in preterm newborns (born at 28 to 32 weeks of gestation), who have very limited
prenatal experience of language, some aspects of the neural circuits for language appear to
be already in place. Indeed the response to syllables in the posterior temporal cortex of
these preterm infants is faster and more sustained in the left than the right hemisphere
(Mahmoudzadeh et al., 2013).
Despite this early neural specialization for language, there is also a clear role of experience
in shaping the neural substrate of language. In newborns and in older infants, brain activation
for the familiar language has been shown to be larger in amplitude (Fava, Hull, & Bortfeld,
2014; May et al., 2011; Minagawa-Kawai et al., 2010; Sato et al., 2012) and more left later-
alized (Sato et al., 2012; Vannasing et al., 2016) than for an unfamiliar language. Moreover, in
the left temporal and temporoparietal cortex of newborns, forward speech elicits more activa-
tion than backward speech for a familiar language, whereas this difference is not observed for
an unfamiliar language (May et al., 2018; Sato et al., 2012; but see also May et al., 2011).
Another way of assessing how experience shapes the neural substrate of language is by
comparing infants with different language experience, such as monolinguals and bilinguals.
When infants are exposed to two spoken languages from birth, they acquire two linguistic
codes (two sets of sounds, two lexicons, two sets of grammatical rules) and learn to keep them
apart, even though they experience a reduced amount of each of these codes compared to
monolinguals (Costa & Sebastián-Gallés, 2014; Werker, 2012). Although this process is ex-
tremely complex, bilinguals usually follow the same milestones of early language develop-
ment that monolinguals follow (Costa & Sebastián-Gallés, 2014; Werker, 2012). It has been
suggested that native and non-native phonetic contrasts are differently represented in the
brains of bilingual compared to monolingual infants (Ferjan Ramírez, Ramírez, Clarke,
Taulu, & Kuhl, 2017; Garcia-Sierra et al., 2011; Petitto et al., 2012). To our knowledge, there
are no published studies comparing the neural responses to familiar and unfamiliar languages
in bilingual and monolingual babies above the phonetic level.
Hearing infants with deaf mothers who use a sign language also grow up to be bilingual.
They offer a unique window into experience-dependent plasticity. If a deaf mother uses a sign
language (such as British Sign Language [BSL]), as her preferred mode of communication, the
speech and language experience of her hearing infant is likely to be very different from that of
hearing infants of hearing mothers. Prenatally, if the mother is using mainly sign language in
her daily interactions, the fetus is likely to have a reduction in language experience, given the
Neurobiology of Language
10
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
1
1
9
1
8
6
7
9
0
9
n
o
_
a
_
0
0
0
0
1
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Experience impacts brain activation for language in infancy
reduced exposure to maternal voice in utero. After birth, hearing infants with deaf mothers
experience a sign language, such as BSL, and a spoken language, such as English, which
can be used by the mother, as well as by hearing relatives and the rest of the hearing commu-
nity. For this reason, these infants can be referred to as “bimodal bilinguals,” as opposed to
“unimodal bilinguals,” who are exposed to two spoken languages. The language produced by
a deaf mother to and around her infant is likely to include less audiovisual spoken language
than that of a hearing mother. Many deaf signers may use speech to communicate with hear-
ing people, but the extent to which they actually “voice” their speech and produce sound, as
opposed to silently mouth, is extremely variable (Bishop & Hicks, 2005). When addressing her
infant, a deaf mother may use signed and/or spoken language, but spoken utterances by deaf
mothers tend to be reduced in length and frequency compared to that of hearing mothers
(Woll & Kyle, 1989).
Studies that have addressed spoken language development in bimodal bilingual children
are usually based on a single child or a few children, and they often report inconsistent results.
Early vocabulary development in bimodal bilinguals has been reported to be similar to that of
monolinguals (Brackenbury, Ryan, & Messenheimer, 2006; Capirci, Iverson, Montanari, &
Voltera, 2002; Griffith, 1985), better than monolinguals (Daniels, 1993), or poorer than mono-
linguals (Murphy & Slorach, 1983; Schiff-Meyers, 1993). Poorer performance in bimodal bi-
lingual children compared to monolingual norms has also been reported on assessments of
phonology, comprehension, and/or grammar (Hofmann & Chilla, 2015; Johnson, Watkins,
& Rice, 1992; Murphy & Slorach, 1983; Schiff & Ventry, 1976; Schiff-Meyers, 1993).
However, because bimodal bilinguals grow up learning two languages, a more appropriate
contrast is with unimodal bilinguals than with monolinguals. Such comparison suggests that
bimodal bilinguals may achieve the early linguistic milestones in spoken and in signed lan-
guage at the same time as children learning two spoken languages (Hofmann & Chilla, 2015;
Petitto et al., 2001).
It is unclear how bimodal bilingual experience affects brain activation for spoken language
in infancy. Moreover, the neural representation for sign language has never been studied in
infancy. Adult neuroimaging studies robustly demonstrate that sign language is processed in a
brain network similar to that of spoken language in deaf and hearing adults who are fluent in
sign language (Capek et al., 2008; Emmorey, 2001; Hickok, Bellugi, & Klima, 1996;
MacSweeney et al., 2004; MacSweeney, Capek, Campbell, & Woll, 2008; Petitto et al.,
2000). This is a strong argument for the idea that classical language areas in the left perisylvian
cortex are specialized for the processing of natural languages independent of their modality.
However, it is unclear if this activation pattern in adulthood represents an adaptation that takes
years of language experience and language learning to be established or if it can be observed
from infancy.
The present study aims to clarify how the development of the neural system’s supporting lan-
guage perception is influenced by the infant’s language experience. To address this we use func-
tional near-infrared spectroscopy (fNIRS). A technique that has been used successfully to study
brain representation for language in infants (Altvater-Mackensen & Grossmann, 2016; Fava
et al., 2014; May et al., 2011, 2018; Minagawa-Kawai et al., 2010; Pena et al., 2003; Sato
et al., 2012; Vannasing et al., 2016), fNIRS measures hemodynamic responses elicited by neu-
ronal activation. This technique offers a balance of spatial and temporal resolution, is relatively
robust to movement artifacts, and requires less infant tolerance than does fMRI or MEG (Lloyd-
Fox, Blasi, & Elwell, 2010). Using traditional univariate approaches to fNIRS data analysis (Aslin,
Shukla, & Emberson, 2015; Lloyd-Fox et al., 2010), we describe patterns of neural activation in
Neurobiology of Language
11
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
1
1
9
1
8
6
7
9
0
9
n
o
_
a
_
0
0
0
0
1
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Experience impacts brain activation for language in infancy
response to spoken and signed language in 60 hearing infants. Furthermore, we assess how these
patterns of language activation for spoken and signed language differ in three groups of infants with
different language experience: monolinguals, unimodal bilinguals, and bimodal bilinguals. Using
region-of-interest (ROI) analyses, we test the hypothesis that, in response to spoken language, bi-
modal bilinguals show reduced amplitude of activation and reduced lateralization in frontotem-
poral language areas compared with monolinguals and unimodal bilinguals, due to reduced
input of auditory spoken language from their mothers. On the other hand, we predict that bimodal
bilinguals will show increased amplitude of activation in frontotemporal language areas and in-
creased lateralization in response to sign language compared to monolinguals and unimodal bilin-
guals, who have never experienced sign language.
A second aim of the current study is to clarify how the effects of language familiarity on
brain activation are influenced by an infant’s language experience. Based on the literature re-
viewed above, it is predicted that a familiar language, here spoken English, will be associated
with increased activation compared to an unfamiliar language in all infants. Moreover, it is
predicted that these familiarity effects will be reduced in both groups of bilinguals compared
to monolinguals, as they are likely to have experienced a reduced amount of the familiar lan-
guage, since their language exposure is split between two languages. We also predict that in
bimodal bilingual infants, a familiar sign language will be associated with increased activation
compared to an unfamiliar sign language. These predictions will be assessed with univariate
and ROI analyses.
In addition to addressing these questions using univariate and ROI analyses, we use mul-
tivariate pattern analysis (MVPA) to compare distributed patterns of brain activation associated
with spoken versus signed language, and with familiar versus unfamiliar languages. The use of
multivariate analyses potentially offers greater sensitivity to experimental effects by pooling
weakly discriminative information that is distributed across measurement channels (Haynes
& Rees, 2006; Norman, Polyn, Detre, & Haxby, 2006). Support vector machines, in particular,
are robust to the inclusion of noninformative channels, hence avoiding the arbitrary selection of
channels of interest (McGettigan et al., 2012). Although fNIRS has fewer channels compared to
fMRI voxels, multivariate analysis methods have been used successfully in studies in both adults
and children (Bogler, Mehnert, Steinbrink, & Haynes, 2014; Gu et al., 2018; Heger, Mutter,
Herff, Putze, & Schultz, 2013; Hosseini et al., 2011; Ichikawa et al., 2014; Luu & Chau,
2008; Misawa, Shimokawa, & Hirobayashi, 2014). Applying MVPA to infant fNIRS data is chal-
lenging given the low trial numbers, variability, and noise in the data. Nevertheless, Emberson,
Zinszer, Raizada, and Aslin (2017) successfully used MVPA to decode visual and auditory stim-
uli in infant fNIRS data as well as two different types of audiovisual stimulation.
In the current study, we hypothesize that MVPA can be used to classify brain activation in
response to spoken and signed language in infants who have experienced only spoken lan-
guage (monolinguals and unimodal bilinguals). We also hypothesize that information in the
left hemisphere will be critical to this classification of language modalities. Moreover, we hy-
pothesize that spoken and signed language will be associated with more similar patterns of
neural activity in infants who have experience of both modalities (bimodal bilinguals) com-
pared to monolinguals and unimodal bilinguals. Therefore we predict that decoding spoken
and signed language using MVPA will be more successful in monolinguals and unimodal bi-
linguals than in bimodal bilinguals. Finally, we hypothesize that a familiar language can be
discriminated from an unfamiliar language based on distributed patterns of activation. This
classification is predicted to be more successful in monolinguals than in either of the bilingual
groups due to greater exposure to the familiar language in monolinguals.
Neurobiology of Language
12
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
1
1
9
1
8
6
7
9
0
9
n
o
_
a
_
0
0
0
0
1
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Experience impacts brain activation for language in infancy
MATERIALS AND METHODS
Participants
Data are presented from 60 infants between 4 and 8 months of age. An additional 34 infants par-
ticipated in the study but were not included in the analyses due to equipment malfunction (n = 3),
withdrawal (n = 1), channel rejection, and looking time criteria (n = 30; see Data Processing
section for details). This dropout rate is similar to that for other infant fNIRS studies (Lloyd-Fox
et al., 2010). Infants were from three groups with different language experience: 19 monolingual
infants with hearing parents (8 girls; mean age = 6.1 months, SD = 1.0), 20 unimodal bi-
lingual infants with hearing parents (6 girls; mean age = 6.0 months, SD = 0.9), and 21 bi-
modal bilingual infants with deaf parents (11 girls; mean age = 6.4 months, SD = 1.2). Age
did not differ between groups (F(2) = 0.67; p = 0.518; η2 = 0.023) and did not differ in var-
iance between groups (F(2, 57) = 1.5; p = 0.225; see Supplementary Figure 1A for distribu-
tion). Maternal education did not differ between groups (Pearson chi-square (6) = 6.6; p =
0.356; see Supplementary Figure 1B). Annual household income varied greatly between
families and there was an underrepresentation of bimodal bilingual families in the highest
income categories (Pearson chi-square (24) = 73.4; p < 0.001; see Supplementary Figure 1C).
Children came from 58 different families (one family had twins and one family returned
later to participate with a younger sibling). Most infants were born at term (37 to 42 weeks
of gestation), except for two infants who were born slightly before term (35 to 36 weeks of
gestation) for whom a corrected age was used. Infants had no severe hearing or vision prob-
lems, and no history of seizure or other serious mental or physical health issues according to
their parents.
Monolingual infants were exposed to English only and both parents were hearing monolin-
guals. Unimodal bilinguals were frequently and regularly exposed to English and one or more
additional spoken language(s) (see Supplementary Table 1 for additional languages and lan-
guage combinations). All infants in this group had a hearing bilingual/multilingual mother.
Most unimodal bilingual infants also had a bilingual/multilingual father (n = 15), whereas five
had a monolingual father. Bimodal bilinguals were frequently and regularly exposed to BSL
and English. All infants in this group had a deaf mother who used BSL as her preferred mode of
communication. Nineteen bimodal bilingual infants had a severely/profoundly deaf father,
one had a hearing father, and one had a single deaf mother. Infants exposed to French or
French Sign Language of Belgium (LSFB) were excluded from the study because these lan-
guages were used as unfamiliar languages in the present study.
The Mullen Scales for Early Learning (Mullen, 1995) was administered to all infants to as-
sess for any differences in development that could influence the interpretation of brain imaging
results. An analysis of variance (ANOVA) with three groups on Mullen t scores revealed no
group effect on visual reception, F(2) = 1.7; p = 0.188; η2 = 0.057; fine motricity, F(2) =
1.8; p = 0.183; η2 = 0.058, and gross motricity scales, F(2) = 2.2; p = 0.119; η2 = 0.072
(see Table 1). A significant effect of group was observed on the receptive language scale,
F(2) = 3.9; p = 0.025; η2 = 0.122. Post hoc t tests revealed that bimodal bilinguals out-
performed unimodal bilinguals (p = 0.013) and monolinguals (p = 0.028) on receptive language
skills, whereas there was no difference between monolinguals and unimodal bilinguals (p =
0.785). The items included in this scale for this age group were mostly communicative in
nature, such as interacting with their reflection in a mirror, or turning around when an ex-
perimenter called their name from behind their back. None of these items was rated by pa-
rental report. The expressive language scale was excluded because its administration
disadvantaged bimodal bilinguals (see Procedure for details).
Neurobiology of Language
13
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
1
1
9
1
8
6
7
9
0
9
n
o
_
a
_
0
0
0
0
1
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Experience impacts brain activation for language in infancy
Table 1. Mullen Scales of Early Learning t scores in the receptive language, visual reception, fine motricity, and gross motricity scales in each
group
Receptive language
Visual reception
Fine motricity
Gross motricity
Monolinguals (n = 19)
44.2 (12.8) [20–64]
Unimodal bilinguals (n = 20)
43.5 (11.9) [20–63]
Bimodal bilinguals (n = 21)
52.9 (10.5) [20–66]
53.5 (12.2) [26–80]
51.7 (9.4) [26–64]
56.0 (10.8) [31–77]
46.8 (11.1) [27–64]
46.6 (9.9) [27–64]
52.8 (8.1) [40–68]
51.9 (12.1) [32–76]
51.6 (10.4) [26–68]
49.5 (9.9) [30–67]
Group mean (standard deviation), [range].
Bimodal bilinguals were recruited through social media and websites specifically aimed at
the deaf community. Infants with hearing parents were contacted from the Birkbeck Babylab
database of volunteers recruited from advertisements at parent-and-baby groups, parenting
websites, and publications. Deaf families were geographically spread across Great Britain,
while infants with hearing parents came mostly from London and its surroundings. Travel ex-
penses were reimbursed, and a baby t-shirt and certificate of participation were offered to fam-
ilies. All parents provided written informed consent prior to participation, after explanations of
the study in English or BSL, depending on the parents’ preferred mode of communication. The
protocol was approved by the Birkbeck and UCL Research Ethics Committees and conforms to
the Declaration of Helsinki.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
1
1
9
1
8
6
7
9
0
9
n
o
_
a
_
0
0
0
0
1
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
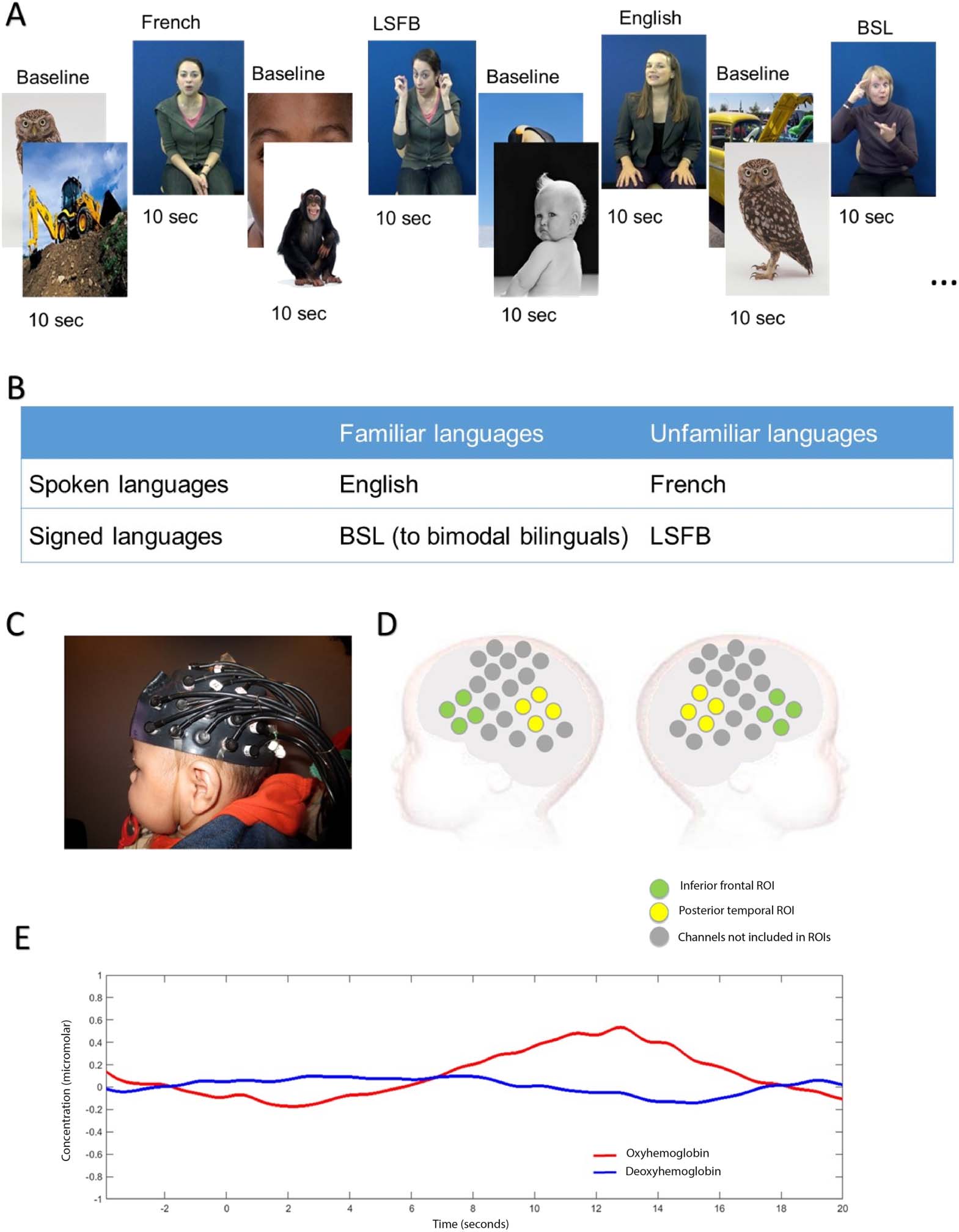
Stimuli
Experimental stimuli consisted of audiovisual videos of extracts from two children’s story-
books. All infants were presented with four different experimental conditions: infant-directed
English (spoken language, familiar to all infants), infant-directed French (spoken language, un-
familiar to all infants), infant-directed BSL (sign language, familiar to infants with deaf mothers),
and infant-directed LSFB (sign language, unfamiliar to all infants). BSL and LSFB are parts of
different families of sign languages, respectively, the British Sign Language family and the
“Langue des Signes Française” family (https://glottolog.org). Four female models who were bi-
lingual in a different combination of languages (English-French, English-BSL, French-LSFB, or
BSL-LSFB) contributed four videos each (two videos per language). Videos were 9 to 12 s long
(mean 10.5 s) and consisted of approximately three sentences from two children’s stories. These
videos were interleaved with 10-s baseline trials in which static images of animals, babies, and
modes of transportation were presented. The experiment started with the baseline condition.
Experimental conditions were presented in pseudo-random order, alternating between spoken
and signed language modalities (see Figure 1A).
Experimental Design
All infants were presented with four experimental conditions, which could be merged in two
language modalities: spoken languages (English + French) and sign languages (BSL + LSFB)
(see Figure 1B). Two levels of analysis are presented in this article. First, modality analyses
assess brain activation for spoken language (English + French), and brain activation for sign
language (BSL + LSFB). Then familiarity analyses are performed within each language modal-
ity. These compare activation for a familiar spoken language (English) with activation for an
unfamiliar spoken language (French) in each group of infants. Sign language familiarity anal-
yses compare brain activation for a familiar sign language (BSL) with brain activation for an
Neurobiology of Language
14
Experience impacts brain activation for language in infancy
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
1
1
9
1
8
6
7
9
0
9
n
o
_
a
_
0
0
0
0
1
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 1.
Stimuli and fNIRS measurement. A. Example of stimulus sequence and timings. B.
Summary of experimental conditions and experimental design. C. Picture of infant wearing the
NTS fNIRS headgear used in the current study. D. Illustration of channel location in relation to
an infant’s head. Channels depicted in green are part of the inferior frontal region of interest, while
channels depicted in yellow are part of the posterior temporal region of interest. E. Example of
HbO2 and HHb grand averaged hemodynamic responses in a left posterior temporal channel in
response to spoken language. Highlighted time windows represent periods of experimental stimulus
and analyses.
Neurobiology of Language
15
Experience impacts brain activation for language in infancy
unfamiliar sign language (LSFB) within the bimodal bilingual group. Sign language familiarity
analyses are not performed in monolinguals and unimodal bilinguals because both sign lan-
guages are unfamiliar to them.
Procedure
Infants sat on their caregiver’s lap wearing a custom-built fNIRS-Centre for Brain and Cognitive
Development (CBCD) headgear formed of 46 channels with a 2 cm source-detector separation
(16 sources and 16 detectors), covering areas of the frontal, temporal, and temporoparietal
cortex (see Figure 1C and 1D). Caregivers were instructed to prevent infants from grabbing
the headgear and to refrain from interacting with their infant unless the infant became fussy
or sought interaction. Infants were seated approximately 100 cm from a 117 cm plasma screen
in a dimly lit and sound-attenuated room. The experiment ended once infants had viewed 20
experimental trials or if the infant became fussy or uninterested. The fNIRS data were recorded
using the NTS optical topography system (Gowerlabs Ltd. L, UK) with two continuous wave-
lengths of source light: 770 and 850 nm at a sampling rate of 10 Hz. Infant’s behavior during
the study was filmed for off-line coding of looking time.
After completion of the fNIRS study, the Mullen Scales of Early Learning was administered by a
trained experimenter. During these sessions, the parent and experimenter interacted in English for
hearing parents and in BSL and/or English for deaf parents. As described in the Mullen Scales of
Early Learning manual, very little verbal instruction was given to infants of this age group, and when
they were required, parents were asked to give these instructions to their infant and/or repeat them
in the language of their choice. This method was used to avoid a potential disadvantage to bilingual
infants. All sessions were filmed and the receptive language, visual reception, fine motricity, and
gross motricity scales were scored off-line by two independent scorers. For items for which scores
differed between scorers, an experienced third coder made the final decision. The expressive lan-
guage scale was excluded because many of the items could be scored based on parental report and
several deaf mothers reported not being aware of the details of their child’s babbling. Moreover,
because sign language was used to communicate with deaf parents during these evaluation ses-
sions, bimodal bilinguals were exposed to less spoken language than monolinguals and unimodal
bilinguals, which could negatively affect their vocal productions during these sessions.
Data Processing
Infant’s looking time during stimulus presentation was coded from videos by a researcher who
was unfamiliar with the hypotheses. Only experimental trials in which the infant was looking
at the screen for at least 60% of the trial duration were included for analyses. This criterion is
similar to that for other fNIRS studies employing visual stimuli with infants (Di Lorenzo, Blasi,
et al., 2019; Lloyd-Fox et al., 2013; Lloyd-Fox, Papademetriou, et al., 2014).
The fNIRS system measured the light attenuation from each source-detector pair (channel).
These light attenuation measures were used to calculate changes in oxyhemoglobin (HbO2)
and deoxyhemoglobin (HHb) chromophore concentration (μmol) and used as hemodynamic
indicators of brain activity (Obrig & Villringer, 2003). Prior to conversion to concentration
data, the attenuation measurements within each channel for each infant were inspected using
artifact-detection thresholding algorithms (Lloyd-Fox et al., 2009, 2010). Channels with poor
signal readings, excess variability in the data measured with the coefficient of variation, or
large baseline drifts were excluded from further analyses. Channels were excluded if the co-
efficient of variation of their attenuation exceeded 15% or if their normalized power was larger
than 50% of the total power. This procedure aimed to exclude channels for which there was
Neurobiology of Language
16
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
1
1
9
1
8
6
7
9
0
9
n
o
_
a
_
0
0
0
0
1
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Experience impacts brain activation for language in infancy
not enough light from the source reaching the corresponding detector (e.g., due to hair block-
ing either optode or one of the optodes being unclipped from the array), for which the noise
characteristics per wavelength were significantly different, or for which channels contained
strong frequency components unrelated to the experiment. The specific thresholds used were
based on previous experience using the NTS Gowerlabs system (Lloyd-Fox et al., 2009), as it is
recommended in infant studies to tailor thresholds to the particular study and fNIRS system
employed (Di Lorenzo, Pirazzoli, et al., 2019). Infants with more than 15 rejected channels
were excluded from all analyses. For each infant, the near-infrared intensity signal was low-
pass filtered, using a cutoff frequency of 1.7 Hz to account for heart rate and certain types of
instrumentation noise. This cut-off frequency is in line with recent studies using this fNIRS sys-
tem (Lloyd-Fox et al., 2019). The data were then segmented into blocks of 24 s of data con-
sisting of 4 s of the baseline trial prior to the onset of the stimulus, the experimental stimulus
trial (10 s), plus the following baseline (10 s). To account for baseline drifts attributable to
potential build-up of activation from trial to trial and/or slow fluctuations that could be of
physiological origin, each block of attenuation data was de-trended with a linear fit between
the average of the first 4 s and the average of the last 4 s. The attenuation data were then
converted into changes in concentration in HbO2 and HHb using the modified Beer–
Lambert law (Delpy et al., 1988) with an assumption of an age-appropriate differential
pathlength factor of 5.13 (Duncan et al., 1995). A second level of automatic artifact detection
and rejection was then conducted on a trial-by-trial level (within each channel) to identify
excessive movement artifacts (Lloyd-Fox et al., 2010, 2019). Trials were removed if there were
concentration changes greater than ±3 μmol during the 4-s baseline prior to the onset of the ex-
perimental stimuli, or if changes exceeded ±5 μmol during the experimental trial itself. These
thresholds were set at different levels to ensure the rejection of abrupt changes in signal caused
by motion while taking into account changes in hemoglobin levels caused by experimental con-
ditions. This second step was designed to identify isolated trials with artifacts, for example, as
caused by a sudden movement by the infant, which may not have triggered identification at
the channel inspection stage of the intensity signal.
An average hemodynamic response curve was computed for each participant in each condition
(English, French, BSL, and LSFB) based on valid channels in valid trials. Experimental conditions
with fewer than three valid trials were excluded (i.e., trials passing channel rejection and looking
time criteria). If one experimental condition was excluded (e.g., English), the other condition of the
same modality was also excluded (e.g., French). Given these criteria, spoken language familiarity
analyses were based on 14 monolinguals, 18 unimodal bilinguals, and 13 bimodal bilinguals (see
Supplementary Table 2 for number of trials). Sign language familiarity analyses were based on eight
bimodal bilinguals. Infants were included in modality analyses if they had at least three valid trials
for each modality: spoken language (English + French) and sign language (BSL + LSFB). Using these
criteria, modality analyses were based on 19 monolinguals, 20 unimodal bilinguals, and 21
bimodal bilinguals (see Supplementary Table 2 for number of trials).
A grand average including all infants was computed for each channel during the presentation
of each language modality (spoken and signed language) and used to select an analysis time
window from 8 to 16 s poststimulus onset (see Figure 1E). This period was selected to include
the range of maximal concentration changes observed for HbO2 and HHb based on visual in-
spection of the current data and was informed by data analysis approaches using a similar par-
adigm in previous cohorts (Lloyd-Fox et al., 2009, 2013; Lloyd-Fox, Blasi, Mercure, Elwell, &
Johnson, 2012). The maximum amplitude variation from baseline was generally observed at
around 12 s. This is later than in most adult studies (Cutini, Moro, and Bisconti, 2012), but sim-
ilar to previous infant studies with stimuli of this length and complexity (Lloyd-Fox et al., 2012,
Neurobiology of Language
17
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
1
1
9
1
8
6
7
9
0
9
n
o
_
a
_
0
0
0
0
1
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Experience impacts brain activation for language in infancy
2019). Peak amplitude variation from baseline was calculated within this time window for each
infant in each experimental condition for HbO2 and HHb and compared statistically using two-
tailed t tests. Given the relatively large width of the time window and potential individual var-
iability in peak latency, peak amplitudes were analyzed instead of mean amplitudes. To resolve
statistical problems of multiple comparisons for these group analyses we applied the false dis-
covery rate (FDR) correction (Benjamini & Hochberg, 1995) based on 46 channels. Given the
exploratory nature of this first step in the analyses, we report our findings both before and after
correction. Both a significant increase in HbO2 and significant decrease in HHb are commonly
accepted as indicators of cortical activation in infant studies (Lloyd-Fox et al., 2010). In instances
where HbO2 and HHb increased or decreased at the same time, the signal was considered in-
consistent with a hemodynamic response (Lloyd-Fox et al., 2010; Obrig & Villringer, 2003) and
was therefore not reported as a significant result.
ROI Selection
Two a priori ROIs corresponding to classical language areas were defined: the inferior frontal
region and posterior temporal region (see Figure 1D). Both regions have been widely docu-
mented to be involved in language processing and both are activated in a wide variety of
language tasks in adults including prelexical phonemic processing, word retrieval and arti-
culation, as well as processing semantic and syntactic ambiguity in adults (Price, 2010).
Channels for each of these regions were selected based on anatomical coregistration of fNIRS
data with individual MRI scans from a group of infants in the same age range (4–7 months;
Lloyd-Fox, Richards, et al., 2014). The inferior frontal ROI included eight channels (four per
hemisphere) previously coregistered within the inferior frontal lobe in 90%–100% of 4- to
7-month-olds. The posterior temporal ROI was composed of eight channels (four per hemi-
sphere) covering the posterior temporal area and temporoparietal junction. Of these eight
channels, four channels (two per hemisphere) were coregistered in the temporal lobe in
90%–100% of 4- to 7-month-olds, with the middle and superior temporal gyrus as the iden-
tified macroanatomical structures for each channel. Two channels (one per hemisphere) were
coregistered in the parietal, frontal, or temporal area, with the superior temporal gyrus or post-
central gyrus as the most frequent identified macroanatomical structures. The last two chan-
nels (one per hemisphere) were adjacent, but not included in the co-registration with MRI of
Lloyd-Fox, Richards, et al. (2014), as they were not part of the headgear used in their study.
These channels were located over the temporoparietal junction and are most likely to overlay
the posterior part of the superior temporal gyrus or the supramarginal gyrus.
Multivariate Pattern Analyses
To compare brain activation for different experimental conditions at the network level, includ-
ing the inferior frontal and posterior temporal ROIs, MVPAs were used. To test for hemispheric
asymmetries, patterns of activation were compared across all channels, and then within each
hemisphere. Classification was conducted with a linear support vector machine packaged in
MATLAB using a soft margin with the default C value of 1 for binary classifications. The pat-
terns submitted to the analysis were the maximum amplitude in the predefined time window
from the average of all trials for each participant and each experimental condition, such that
each participant contributed a single pattern to each analysis (e.g., spoken vs. signed
language, or familiar vs. unfamiliar languages). Channels were excluded following the same
criteria described in Data Processing. Data were z-scored within each channel across all in-
fants to ensure that features were in comparable scales for classification. We used a leave-one-
participant-out approach, such that the classifier was trained on a balanced training set of
Neurobiology of Language
18
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
1
1
9
1
8
6
7
9
0
9
n
o
_
a
_
0
0
0
0
1
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Experience impacts brain activation for language in infancy
neural responses from all of the participants excluding the to-be-classified participant (see
Emberson et al., 2017 for a similar approach). The model was trained on the averaged pattern
derived from the fNIRS epoch data for each of the two experimental conditions from all
participants except one. It was then tested against the held-out pattern from the remaining
participant. As such, each participant contributed two trials, one from each condition to be
classified and could be classified as correctly guessed or otherwise. Hence the full data set of
60 participants yielded accuracy based on successful classifier guesses from 120 trials, and
within-group analyses of 20 participants derived 40 classification trials on which accuracy was
based. The classification reported is the proportion of correctly guessed trials. To ensure that
classification was not biased, permutation testing was conducted by randomly permuting
condition labels for each participant, such that the labels were either randomly maintained
or swapped prior to training and testing of the classifier. This ensured that the participant struc-
ture was preserved in the label shuffling. For binary classifications (i.e., spoken vs. signed
language or familiar vs. unfamiliar languages), 1,000 permutations were conducted and a
probability value was ascertained by generating a null distribution and identifying the number of
observed values that was greater than or equal to the accuracy derived from the nonshuffled
data (Pereira, Mitchell, & Botvinick, 2009). The observed value was included in both the
numerator and denominator for calculating the p value, such that if the classification accuracy
observed from the data was higher than all the observed permutation values, this would result in a
value of p = 1/1001 (Ruxton & Neuhäuser, 2013). Between-group and between-hemisphere
analyses were conducted by ascertaining a null difference distribution by subtracting values
derived from two null distributions. The p value of these analyses represents the fraction of the
sample that is greater than or equal to the accuracy actually observed when using the correct
labels.
RESULTS
To establish a full picture of the neural activation in response to each language modality, we
begin by presenting channel-by-channel analyses for the spoken and the signed language mo-
dalities in all infants taken as a whole, as well as in each group of infants. These are followed
by channel-by-channel analyses of the impact of language familiarity on these patterns of ac-
tivation. Next, we focus on frontotemporal ROIs to statistically compare the activation within
these language areas of the left and right hemispheres, between groups of infants, in response
to each language modality and language familiarity. Finally, we present MVPAs aiming to con-
trast language modalities and language familiarity at the network level.
Univariate Analyses
Language modality
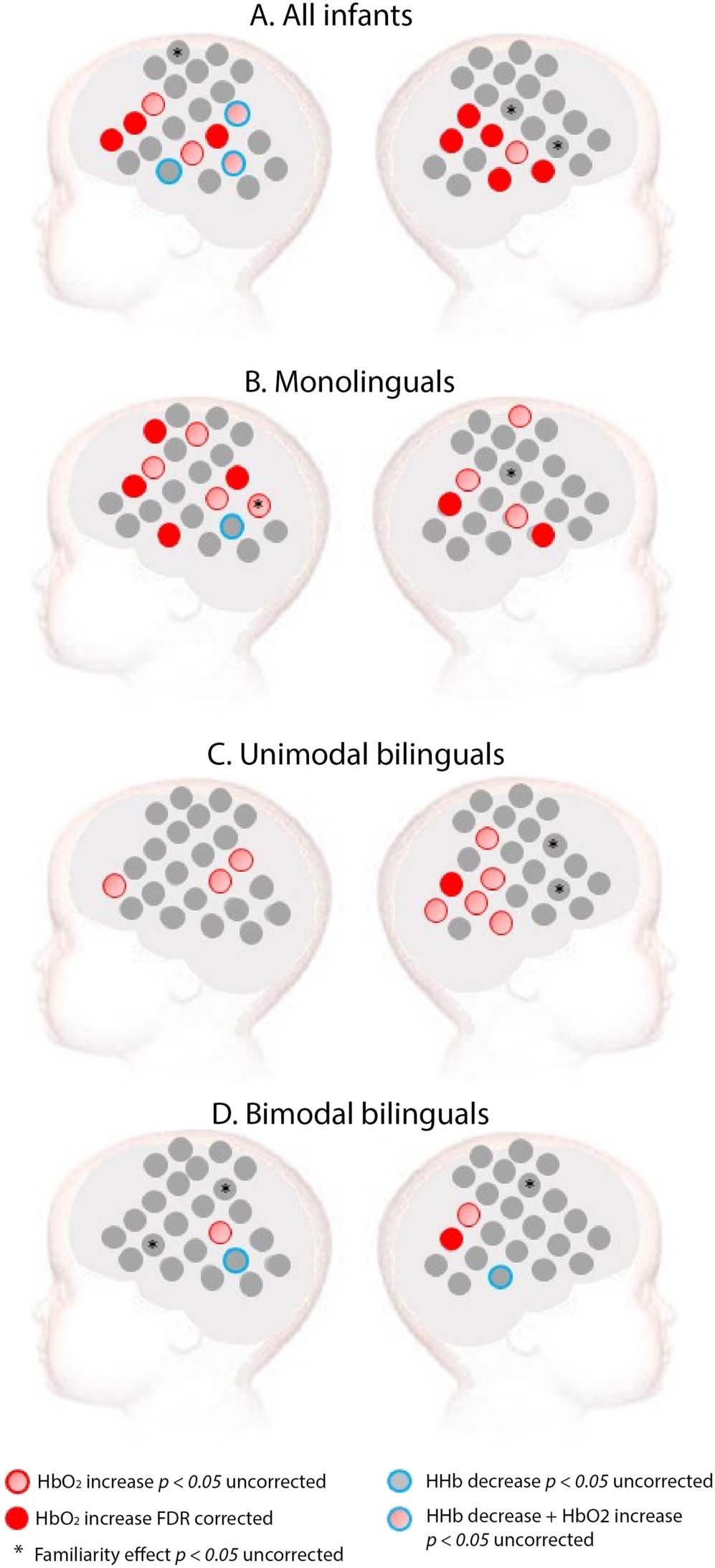
When all 60 infants were considered as a single group, widespread activation was observed in
response to spoken language versus baseline. Significant increase of HbO2 was found in a
bilateral network that included the temporal and the inferior frontal areas of the brain (see
Figure 2). When monolingual infants were considered alone, a significant increase of HbO2
was found in a bilateral network that included the temporal and the inferior frontal areas of the
brain (see Supplementary Figure 3 for grand averaged hemodynamic response in each chan-
nel, group, and experimental condition). Unimodal bilinguals showed bilateral activation with
an uncorrected p value, but the only channel surviving FDR correction was located in the right
posterior temporal area. Bimodal bilinguals had fewer active channels at an uncorrected
p value compared to the other two groups, especially in the left inferior frontal region.
Neurobiology of Language
19
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
1
1
9
1
8
6
7
9
0
9
n
o
_
a
_
0
0
0
0
1
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Experience impacts brain activation for language in infancy
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
1
1
9
1
8
6
7
9
0
9
n
o
_
a
_
0
0
0
0
1
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 2. Activation for spoken language in (A) all infants, (B) monolinguals, (C) unimodal bilin-
guals, and (D) bimodal bilinguals. Red: Channels showing a significant increase in HbO2 in
channel-by-channel analyses at the FDR-corrected level. Pink: Channels showing a significant in-
crease in HbO2 in channel-by-channel analyses at an uncorrected statistical level of p < 0.05. Blue:
Channels showing a significant decrease in HHb at an uncorrected statistical level of p < 0.05. No
channel showed a significant HHb effect at an FDR-corrected level. Stars indicate a significant dif-
ference between a familiar (English) and an unfamiliar (French) spoken language at an uncorrected
statistical level of p < 0.05.
Neurobiology of Language
20
Experience impacts brain activation for language in infancy
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
1
1
9
1
8
6
7
9
0
9
n
o
_
a
_
0
0
0
0
1
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
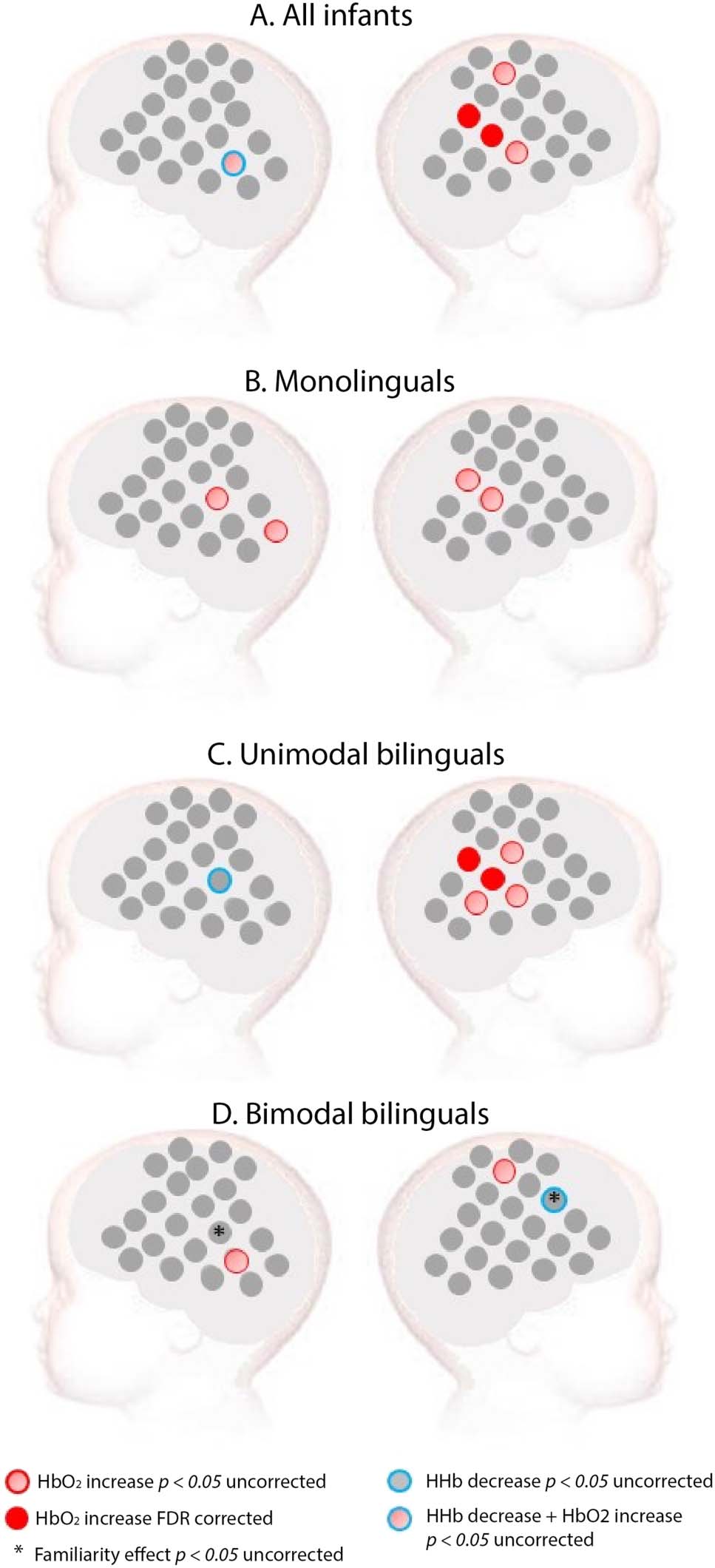
Figure 3. Activation for sign language in (A) all infants, (B) monolinguals, (C) unimodal bilinguals,
and (D) bimodal bilinguals (D). Red: Channels showing a significant increase in HbO2 in channel-
by-channel analyses at the FDR-corrected level. Pink: Channels showing a significant increase in
HbO2 in channel-by-channel analyses at an uncorrected statistical level of p < 0.05. Blue: Channels
showing a significant decrease in HHb at an uncorrected statistical level of p < 0.05. No channel
showed a significant HHb effect at an FDR-corrected level. Stars indicate a significant difference
between a familiar (BSL) and unfamiliar (LSFB) sign language at an uncorrected statistical level of
p < 0.05. These familiarity effects were assessed only in the bimodal bilingual group given that
both signed languages were unfamiliar to infants of the other groups.
Neurobiology of Language
21
Experience impacts brain activation for language in infancy
The only channel surviving FDR correction in bimodal bilinguals was in the right posterior
temporal area. None of the channels demonstrating a significant decrease in HHb survived
FDR correction.
In contrast, sign language elicited a significant increase in HbO2 compared to baseline,
mainly in the right temporoparietal area when all infants were considered as a single group
(see Figure 3). In monolinguals, sign language elicited increased HbO2 in the temporoparietal
area of both hemispheres with an uncorrected p value, but no channel survived FDR correc-
tion. In unimodal bilinguals, a significant increase of HbO2 was observed in the right temporo-
parietal area. Surprisingly, in bimodal bilinguals, only a few scattered channels showed a
significant response to sign language at an uncorrected p value, none of which survived
FDR correction (see Figure 3).
These analyses suggest that spoken language elicited more widespread activation than
signed language, and that the lateralization of this activation differs between groups. These
observations were assessed using ROI analyses, and differences in patterns of activation be-
tween each language modality were assessed with MVPA. In accordance with previous infant
research (Cristia et al., 2013; Gervain et al., 2011; Lloyd-Fox et al., 2010) the majority of the
significant channel-by-channel effects were in HbO2 for both spoken and signed languages.
No HHb effect was significant after FDR correction, so all subsequent familiarity analyses, ROI
analyses, and MVPAs focus on HbO2.
Language familiarity
The peak amplitude of HbO2 was directly contrasted for English (familiar spoken language to
all infants) and French (unfamiliar spoken language to all infants) on a channel-by-channel
basis. Analyses of all infants together suggested that a few channels elicited a difference be-
tween familiar and unfamiliar spoken language. This was also the case when each group of
infants was analyzed separately. However, these channels did not cluster together and none of
these effects survived FDR correction (see Figure 2).
In bimodal bilinguals, we also tested for sign language familiarity effects. BSL (familiar) elic-
ited increased activity compared to LSFB (unfamiliar) in one right frontal channel, whereas the
opposite effect was found in one left temporoparietal channel (see Figure 3). However, these
effects did not survive FDR correction. Familiarity effects are not reported for the other two
groups of infants, since both sign languages were unfamiliar to them.
ROI Analyses
Language modality
To gain a better understanding of the activation within language areas, two ROI analyses were
defined corresponding to the inferior frontal and posterior temporal areas. Peak HbO2 re-
sponse was averaged for all channels within each ROI for each infant, in each hemisphere
and condition. Activation in each ROI was analyzed in a 2 × 2 × 3 ANOVA with language
modality (spoken, signed) × hemisphere (left, right) × group (monolinguals, unimodal bilin-
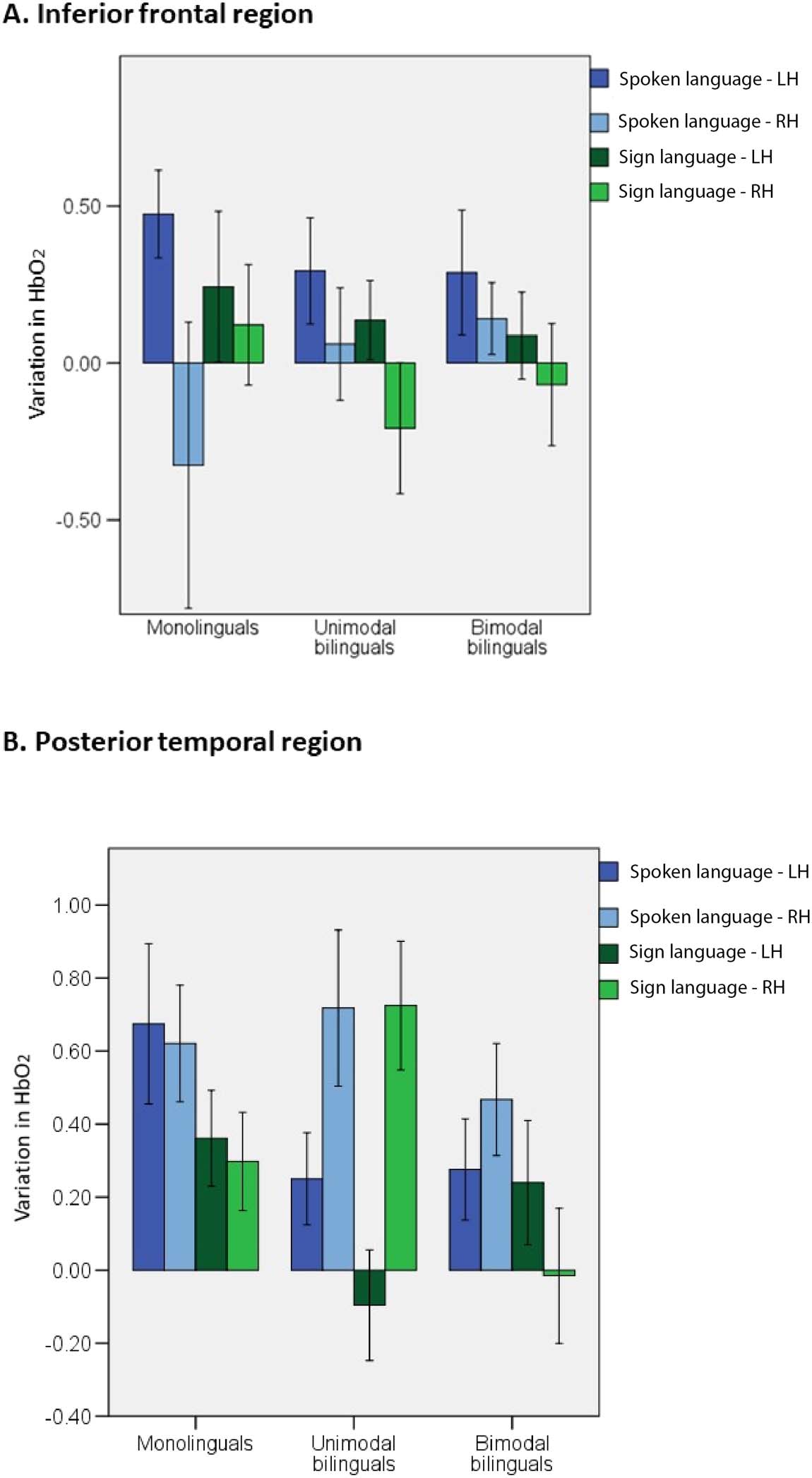
guals, bimodal bilinguals). In the inferior frontal ROIs (see Figure 1D), there was a significant
effect of hemisphere, F(1, 57) = 6.3; p = 0.015; η2 = 0.100, but no other effect or interaction
reached significance level (see Figure 4). Activation in the inferior frontal region was left-
lateralized regardless of group and language modality. This result suggests that activation
within the inferior frontal ROI is not specific to spoken language and not significantly influenced
by an infant’s experience of different language modalities. We note with interest the increased
Neurobiology of Language
22
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
1
1
9
1
8
6
7
9
0
9
n
o
_
a
_
0
0
0
0
1
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Experience impacts brain activation for language in infancy
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
1
1
9
1
8
6
7
9
0
9
n
o
_
a
_
0
0
0
0
1
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. Mean variation in HbO2 in the inferior frontal and posterior temporal ROIs for each
language modality in each group of infants. Error bars represent standard error.
variability in right hemisphere activation in the inferior frontal region to spoken language in the
monolingual group. Although left hemisphere activation was strong in most monolingual infants,
a strong right hemisphere deactivation was observed in many monolingual infants in response to
spoken language. It may be that this right hemisphere deactivation develops with functional
Neurobiology of Language
23
Experience impacts brain activation for language in infancy
specialization for language. The monolingual group may be more advanced in the process of
functional specialization for language because of reduced variability in language input. Right
hemisphere deactivation to spoken language may be most reliably observed in older infants
and as language proficiency develops. These speculative interpretations require further investi-
gation, ideally in a group of infants with a smaller age range.
In the posterior temporal region, there was a significant effect of modality, F(1, 55) = 6.8; p =
0.012; η2 = 0.109. This region activated more in response to spoken than signed language. There
was also a significant interaction between hemisphere × group, F(2, 55) = 6.8; p = 0.002; η2 = 0.199.
Separate ANOVAs in each group revealed that activation in the posterior temporal area was right
lateralized in unimodal bilinguals in response to spoken and signed language, F(1, 17) = 9.8; p =
0.006; η2 = 0.365, whereas monolinguals, F(1, 18) = 0.2; p = 0.653; η2 = 0.011, and bimodal bi-
linguals, F(1, 20) = 0.1; p = 0.778; η2 = 0.004, showed no difference in left and right activation. This
result suggests that the experience of two spoken languages in infancy influences lateralization
of activation in response to language of any modality within the posterior temporal ROI.
Language familiarity
In addition, analyses within each ROI were performed to assess for familiarity effects within each
modality. For spoken language, a 2 × 2 × 3 ANOVA with familiarity (familiar, unfamiliar) ×
hemisphere (left, right) × group (monolinguals, unimodal bilinguals, bimodal bilinguals) did
not reveal any familiarity effects in the temporal or inferior frontal ROIs (all p > 0.14; η2 <
0.001 for the inferior frontal area and η2 = 0.005 for the posterior temporal area). For sign lan-
guage, familiarity effects were assessed only within bimodal bilinguals, as both language modal-
ities were unfamiliar to infants of other groups. No familiarity effect was significant in either ROI
(all p > 0.14; η2 = 0.147 for the inferior frontal area and η2 = 0.149 for the posterior temporal
area). This analysis may have been underpowered given that only eight infants survived inclu-
sion criteria. A power analysis reveals that 12 participants would have been required to achieve
90% power.
Multivariate Pattern Analyses
Language modality
Patterns of activation for spoken and signed language were compared. Each participant’s aver-
age maximum amplitude of HbO2 in the predefined time window in response to spoken lan-
guage (English + French) and signed language (BSL + LSFB) was entered in MVPAs. Data from all
channels were analyzed (dimensions of feature vector = 46) and then separate analyses were
performed for each hemisphere (feature vector = 23) to test for hemispheric differences. When
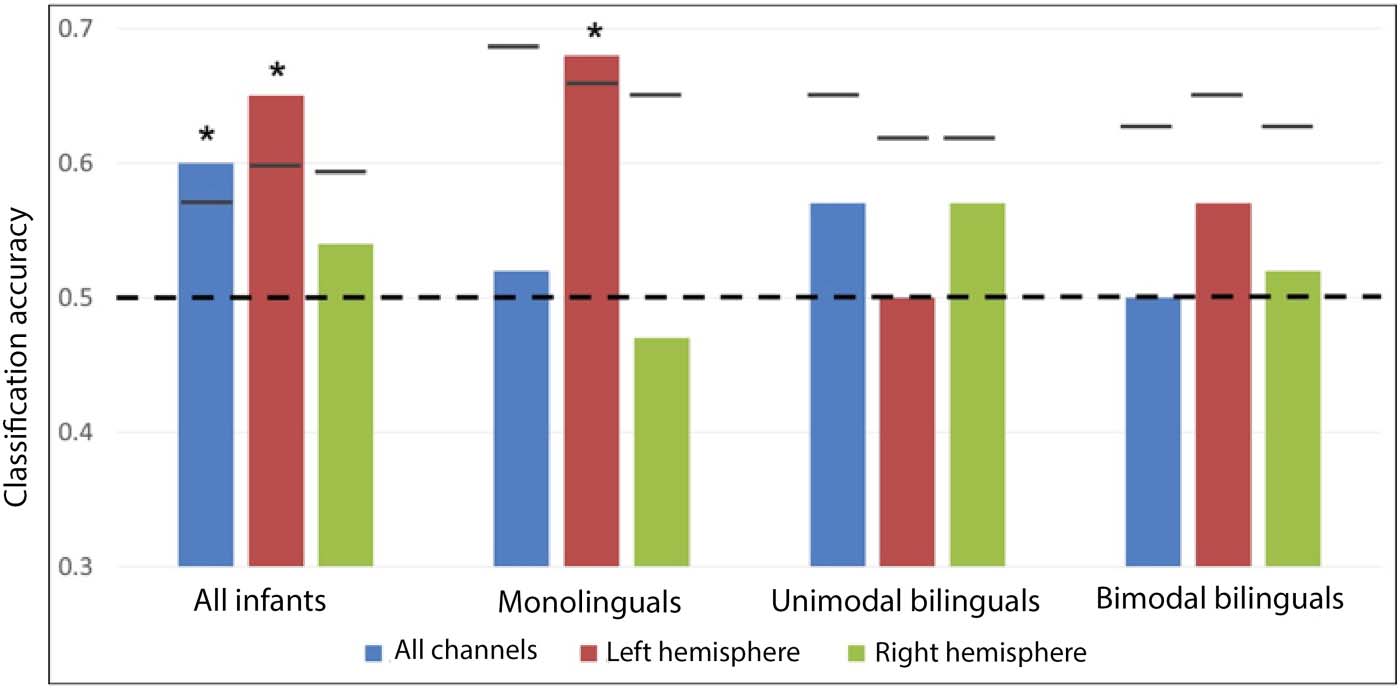
all 60 infants were taken as a single group (see Figure 5 for MVPA results and Supplementary
Figure 2 for a channel-by-channel comparison of language modalities), patterns of activation for
spoken and signed language could be classified at a level greater than chance using all 46 chan-
nels (proportion correct = 0.60; permutation p value = 0.029), or left hemisphere channels (pro-
portion correct = 0.65; permutation p value = 0.010), but not right hemisphere channels
(proportion correct = 0.54; permutation p value = 0.270). The difference in classification accu-
racy between the left and right hemispheres was not significant ( p = 0.218).
In monolinguals, patterns of activation for spoken and signed language could be classified
at a level greater than chance using left hemisphere channels (proportion correct = 0.68; p =
0.039), but not right hemisphere channels (proportion correct = 0.47; p = 0.733) or all chan-
nels (proportion correct = 0.52; p = 0.520). The difference in classification accuracy between
the left and right hemispheres was not significant (p = 0.118). It was predicted that classification
Neurobiology of Language
24
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
1
1
9
1
8
6
7
9
0
9
n
o
_
a
_
0
0
0
0
1
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Experience impacts brain activation for language in infancy
Figure 5. Classification accuracy for spoken versus signed language. Empirical proportion of cor-
rect classification for spoken versus signed language by MVPAs. Empirical proportion of correct
classification is presented for all channels, left hemisphere channels, and right hemisphere channels
in each group of infants. The dotted line represents chance level (0.5), short lines represent the
upper bound of the 95% confidence interval, and stars represent classification models that are sig-
nificantly more successful than chance.
would be less successful in bimodal bilinguals compared to monolinguals and unimodal bilinguals
given that these infants have experience in both language modalities. However, MVPAs could not
classify spoken and signed language with an accuracy greater than chance in either group of bi-
linguals using all channels (all p > 0.2), left channels (all p > 0.5), or right channels (all p > 0.2; see
Supplementary Table 3). Furthermore, although classification based on left hemisphere channels
was successful in only monolinguals, when classification accuracies in the left hemisphere were
compared between groups, this difference was not statistically significant (monolinguals vs. uni-
modal bilinguals [p = 0.123]; monolinguals vs. bimodal bilinguals [p = 0.406]).
These results suggest that spoken and signed language modalities are associated with
differentiated brain activation patterns in monolingual infants, but not in both groups of bilin-
guals. However, the strength of this conclusion is constrained by the absence of a difference in
accuracies between groups.
Language familiarity
Patterns of activation for familiar and unfamiliar spoken languages were compared at the net-
work level using MVPAs. The average of maximum amplitude of HbO2 of all trials in the pre-
defined time window for each participant in response to spoken English (familiar) and spoken
French (unfamiliar) was entered in MVPAs. Classification was not successful in any group
based on all channels, left or right hemisphere channels (see Supplementary Table 4).
Classification of familiar (BSL) and unfamiliar sign languages (LSFB) in bimodal bilinguals
was not significantly more successful than chance (see Supplementary Table 4). These group
analyses were based on small samples and may lack power.
DISCUSSION
The present study investigated the neural activation for spoken and signed language in three
groups of infants with very different language experiences. Although the three groups had
Neurobiology of Language
25
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
1
1
9
1
8
6
7
9
0
9
n
o
_
a
_
0
0
0
0
1
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Experience impacts brain activation for language in infancy
different experience of speech, language, and communication, they shared the experience of
normal hearing and the exposure to English as a familiar spoken language. When this group of
60 infants was considered as a whole, we found activation in response to audiovisual infant-
directed spoken language in a wide bilateral network, which included the posterior temporal
and the inferior frontal areas. This pattern of activation is similar to the network involved in
spoken language processing in adults (Price, 2010) and also to that previously reported in in-
fants (Altvater-Mackensen & Grossmann, 2016; Dehaene-Lambertz et al., 2002, 2006, 2010;
May et al., 2018; Minagawa-Kawai et al., 2010; Pena et al., 2003; Perani et al., 2011; Sato
et al., 2012; Shultz et al., 2014; Vannasing et al., 2016). Sign language elicited activation in a
few channels located in the right temporoparietal area. Multivariate analyses suggest that spo-
ken and signed language elicited different patterns of neural activation, which could be de-
coded with an accuracy greater than chance based on all channels and based on left
hemisphere channels, but not based on right hemisphere channels. This suggests that the left
hemisphere is more sensitive to language modality than the right hemisphere in infancy, even
though only one third of the infants had prior experience of sign language as a mode of
communication.
It is important to note that these patterns of activation emerge from differences in brain
oxygenation between experimental conditions and baseline. During these baseline intervals, in-
fants were presented with a variety of static pictures to keep their attention to the screen. These
consisted of pictures of animals, babies, and modes of transportation. Any difference in activa-
tion between experimental conditions and baseline may be triggered not only by the presence of
language (spoken or signed language), but also by biological motion and/or the consistent pres-
ence of faces in both language conditions. In that respect, we can interpret these patterns of
activation as associated with the audiovisual communicative experience of language.
Having established the pattern of activation in response to spoken and signed language
across the whole group of infants, we examined our specific aims of clarifying the role of lan-
guage experience in shaping brain activation for language. We compared brain activation for
spoken and for signed language and for familiar and unfamiliar languages in each group of
infants.
Language Modality
In monolinguals, activation for spoken language compared to baseline was found in a large
bilateral network of channels including the inferior frontal and posterior temporal regions,
whereas sign language did not elicit any activation compared to baseline, which survived cor-
rection for multiple comparisons. In unimodal bilingual infants, both spoken and signed lan-
guage elicited activation in the right temporoparietal area. Bimodal bilinguals showed
activation to spoken language in the right posterior temporal area, while sign language did
not elicit any activation that survived correction for multiple comparisons.
Our original predictions were that reduced experience of spoken language in bimodal bi-
linguals compared to monolinguals and unimodal bilinguals would be associated with re-
duced amplitude and lateralization of activation in frontotemporal language areas for
spoken language. Conversely, we predicted that the bimodal bilingual’s experience of sign
language would lead to increased amplitude of activation in frontotemporal language areas
and increased lateralization for sign language. Activation in the inferior frontal region of inter-
est was left lateralized regardless of group and language modality. Therefore, left lateralized
activation in this region in infancy is not specific to spoken language, and can also be ob-
served for sign language regardless of the infant’s experience of this language modality. It is
Neurobiology of Language
26
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
1
1
9
1
8
6
7
9
0
9
n
o
_
a
_
0
0
0
0
1
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Experience impacts brain activation for language in infancy
possible that left lateralized activation is elicited in the inferior frontal area in response to stim-
uli with a complex structure such as that of spoken or signed language, and/or by stimuli of a
social communicative nature.
In contrast, there was greater activation in the posterior temporal ROI for spoken than for
signed language regardless of hemisphere. Moreover, lateralization within this region signifi-
cantly differed between groups of infants. Contrary to our prediction, this significant effect was
not driven by bimodal bilinguals, but by unimodal bilinguals who showed right lateralization
in this area in response to both spoken and signed language. Activation in the posterior tem-
poral ROI was not lateralized for spoken and signed language in both monolinguals and bi-
modal bilinguals. Previous studies with proficient unimodal bilingual adults suggest that both
languages engage similar neural regions, located primarily in the left hemisphere (Abutalebi,
Cappa, & Perani, 2001; Liu & Cao, 2016; Perani et al., 1998). Some studies comparing mono-
lingual and unimodal bilingual adults who acquired their languages early have revealed an
increased participation of the right hemisphere during language processing in bilinguals
(Hull & Vaid, 2007; Połczyn´ ska, Japardi, & Bookheimer, 2017). The present study extends
these results to show increased right lateralization in preverbal unimodal bilinguals during
the processing of infant-directed spoken language. Moreover, this pattern of increased right
hemisphere engagement extended to the processing of sign language in unimodal bilingual
infants. That is, experiencing two spoken languages from birth influences brain activation in
response to sign language in unimodal bilinguals, even when experienced for the very first
time.
It is possible that the unimodal bilingual experience in infancy contributes to developing an
increased sensitivity to the general structure of languages, which in turn would lead to similar
patterns of activation for an unfamiliar language modality in this group. Indeed, both spoken
and signed languages are formed of a hierarchical structure where phonological, semantic,
and syntactic elements are combined to produce utterances (Sandler & Lillo-Martin, 2006).
It is also possible that this similarity in structure leads to a similar pattern of activation for both
language modalities.
In addition, unimodal bilinguals may have an increased sensitivity to visual speech articu-
lation. This could explain why unimodal bilingual infants are better than monolinguals at dis-
criminating two foreign spoken languages based on silent articulation (Sebastián-Gallés,
Albareda-Castellot, Weikum, & Werker, 2012). Other studies also demonstrated increased at-
tention capture and maintenance by face stimuli in unimodal bilingual infants compared to
both monolinguals and bimodal bilinguals (Mercure, Kushnerenko, et al., 2018; Mercure,
Quiroz, et al., 2018). The stimuli used in the present study contained some English lip move-
ments often present in natural BSL. It is therefore possible that the activation for sign language
in unimodal bilingual infants is in part associated with the processing of these visual cues of
articulation. Sign language stimuli without any lip movements or with the mouth obscured
would be required to evaluate this possibility.
Perhaps surprisingly, our data suggest that compared to the bimodal bilingual experience,
the unimodal bilingual experience appears to have more impact on lateralization of activation
for language perception. Spoken languages are all produced by movements of the vocal tract
and perceived as audiovisual speech. In that respect, they are more similar than a spoken and
signed language. Learning two similar but not identical codes may require increased cognitive
resources in order to differentiate them, compared to learning a spoken and a signed language.
Spoken and signed languages are more easily differentiated and can also be produced simul-
taneously, often referred to as code-blending (Emmorey, Luk, Pyers, & Bialystok, 2008). In that
Neurobiology of Language
27
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
1
1
9
1
8
6
7
9
0
9
n
o
_
a
_
0
0
0
0
1
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Experience impacts brain activation for language in infancy
respect, a unimodal bilingual experience in infancy may have a stronger impact than a bi-
modal bilingual experience on the process of neural specialization in infancy. Unimodal bi-
lingual infants may focus on the rhythm of the speech input to support the differentiation of
spoken languages, while the differentiation of spoken and signed languages is perceptually
more salient. This increased focus on speech rhythm may explain the increased right hemi-
sphere involvement in unimodal bilinguals compared to other groups of infants (Riecker,
Wildgruber, Dogil, Grodd, & Ackermann, 2002).
Bimodal bilinguals demonstrated a smaller number of active channels in response to both
language modalities compared to monolinguals and unimodal bilinguals. They also tended to
have smaller amplitudes of activation in both ROIs (the inferior frontal and posterior temporal
regions) compared to monolinguals. However, no group effects were found on the mean am-
plitude of the response to spoken and signed language in these ROIs. Although this pattern of
results may provide some weak support for the hypothesis of reduced amplitude of activation
for spoken language in this group, the trend also applied to sign language, which was contrary
to our hypotheses. Of interest, this trend for reduced activation in response to both language
modalities was observed in the absence of a receptive language deficit in this group. To the
contrary, bimodal bilingual infants outperformed the other two groups of infants on commu-
nicative skills, as measured by the receptive language scale of the Mullen Scales of Early
Learning. This is interesting given that this scale was designed with hearing infants of hearing
parents in mind and included items that may not be as familiar to infants with deaf parents,
such as calling their names from behind their back. It is interesting to note that sighted infants
with blind parents have also been shown to outperform sighted infants of sighted parents on
visual receptive skills (Senju et al., 2013). Like sighted infants with blind parents, bimodal bi-
linguals need to adapt their communicative strategies to communication partners with differ-
ent needs. This flexibility may lead to more mature skills in some aspects of their development.
That bimodal bilinguals had better communicative skills than the other two groups, as mea-
sured by the receptive language scale of the Mullen Scales of Early Learning, yet showed very
few active channels in response to spoken and signed language, could suggest that reduced or
more focal language activation may be associated with better communicative skills. However,
future studies with larger samples are required to test this possibility. It would also be interest-
ing to see whether the same association is observed with more language-specific abilities, such
as the development of vocabulary and syntax later in childhood.
Multivariate analyses suggested that spoken and signed language elicited different patterns
of activation in monolinguals, which could be classified at an accuracy greater than chance
based on left hemisphere activity. The classification accuracy (68%) was reasonably high,
especially given that it was performed between participants. This accuracy is comparable to
trial-level decoding found in adult fMRI studies (Evans et al., 2013; Formisano, De Martino,
Bonte, & Goebel, 2008; McGettigan et al., 2012; Misaki, Kim, Bandettini, & Kriegeskorte,
2010). This is also similar to the accuracy found in fNIRS studies in between-infant decoding
(Emberson et al., 2017; data set no. 2, mean = 72%) and between-subject decoding in adults
(Zinszer, Bayet, Emberson, Raizada, & Aslin, 2017; mean = 70%). On the other hand, classi-
fication was not successful in any of the two groups of bilinguals. Both groups of bilinguals
have more diversity in their spoken language experience compared to monolinguals. Not only
is their experience characterized by more than one language, but it is also likely to include
more variable models within a language, including foreign accents, and/or less precise pho-
nological distinctions produced by deaf adults (Lane & Webster, 1991; Waldstein, 1990). This
increased variability in input could influence the process of neural specialization for language
in these bilingual infants, leading to more variable brain representation.
Neurobiology of Language
28
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
1
1
9
1
8
6
7
9
0
9
n
o
_
a
_
0
0
0
0
1
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Experience impacts brain activation for language in infancy
Language Familiarity
The second aim of this study was to assess the impact of language experience on the brain
signature for language familiarity. A spoken language familiar to all infants was presented
(English), as well as a spoken language unfamiliar to all infants (French). Based on the litera-
ture, increased activation was predicted for the familiar versus the unfamiliar language (Fava
et al., 2014; Minagawa-Kawai et al., 2010; Sato et al., 2012; Vannasing et al., 2016), espe-
cially in the left hemisphere. It was also predicted that familiarity effects would be smaller in
both bilingual groups compared to monolinguals because of their reduced exposure to the
familiar language. In each group of infants, a few channels showed differential activation
for the familiar language compared to the unfamiliar language, but none of these channels
clustered and no familiarity effects survived FDR correction. Moreover, ROI analyses revealed
no effect of familiarity and no interaction of familiarity with group or hemisphere. MVPAs
could not classify familiar and unfamiliar spoken languages based on distributed patterns of
activation in all infants or in any of the infant groups. The difference in the patterns of brain
activation for a familiar and an unfamiliar language in preverbal infants may not have suf-
ficient stability across participants to allow categorization. Moreover, infants included in this
study had a wide age range that is associated with various stages in the process of language-
specific specialization (Fava et al., 2014). This could explain the lack of significant familiarity
effects at the group level in the present study.
A familiar sign language was also expected to elicit increased activation and lateralization
in comparison with an unfamiliar sign language in bimodal bilinguals. This hypothesis was not
supported by the data, as no effects of sign language familiarity were observed. Familiar and
unfamiliar sign languages could not be classified based on MVPA in bimodal bilinguals. Only
eight infants reached the inclusion criteria to allow comparison of the two sign language con-
ditions. For this reason, this analysis may be underpowered and these null results should be
interpreted cautiously.
Summary
The results of the present study suggest that the neural substrates of language are influenced by
early experience of language. Contrary to our predictions, our primary finding was related to
the patterns observed in unimodal bilingual infants. These hearing babies exposed to two spo-
ken languages from birth, appear to engage the right hemisphere to a greater extent than the
left during perception of spoken language and also sign language, of which they had no pre-
vious experience. Unimodal bilingual experience may lead to increased sensitivity to the
structure of languages and/or to increased sensitivity to lip patterns that often accompany sign
language. Patterns of activation for spoken and signed language could be successfully classi-
fied in monolinguals, but not in any of the bilingual groups. Bimodal bilinguals demonstrated a
tendency for less activation in response to both language modalities; however, this trend was
accompanied by an increase in behaviorally measured communicative skills. Our results in-
dicate that the neural substrate of language is plastic in infancy and influenced by language
input and language modality, suggesting that there may be different neural paths to language
development.
ACKNOWLEDGMENTS
The authors would like to thank (1) all families who participated in the study; (2) the Deafness
Cognition and Language Research Centre (DCAL) for their support in the form of an Economic
Neurobiology of Language
29
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
1
1
9
1
8
6
7
9
0
9
n
o
_
a
_
0
0
0
0
1
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Experience impacts brain activation for language in infancy
and Social Research Council Research Centre Grant [RES-620-28-0002], with special thanks to
Bencie Woll and Jo Atkinson for their feedback on the experimental design and questions;
(3) Katarina Begus and Victoria Southgate for the loan of their adapted CBCD fNIRS headgear;
(4) Brigitte Francois, Susan Booth, Rhiannon Armstrong, and Ingrid Notarrigo for modeling the
stimuli; and (5) Anna Blasi for her help with data analysis.
FUNDING INFORMATION
Evelyne Mercure, Economic and Social Research Council (https://dx.doi.org/10.13039/
501100000269), Award ID: ES/K001329/1. Mark H. Johnson, Medical Research Council
(https://dx.doi.org/10.13039/501100000265), Award ID: G0701484. Mairéad MacSweeney,
Wellcome Trust (https://dx.doi.org/10.13039/100004440), Award ID: 100229/Z/12/Z.
REFERENCES
Abutalebi, J., Cappa, S. F., & Perani, D. (2001). The bilingual brain
as revealed by functional neuroimaging. Bilingualism: Language
and Cognition, 4(2), 179–190.
Altvater-Mackensen, N., & Grossmann, T. (2016). The role of left
inferior frontal cortex during audiovisual speech perception in in-
fants. NeuroImage, 133, 14–20.
Aslin, R. N., Shukla, M., & Emberson, L. L. (2015). Hemodynamic cor-
relates of cognition in human infants. Annual Review of Psychology,
66, 349–379.
Benjamini, Y., & Hochberg, Y. (1995). Controlling the false discov-
ery rate: A practical and powerful approach to multiple testing.
Journal of the Royal Statistical Society B, 57(1), 289–300.
Birnholz, J. C., & Benacerraf, B. R. (1983). The development of hu-
man fetal hearing. Science, 222, 516–519.
Bishop, M., & Hicks, S. (2005). Orange eyes: Bimodal bilingualism
in hearing adults from deaf families. Sign Language Studies, 5(2),
188–230.
Bogler, C., Mehnert, J., Steinbrink, J., & Haynes, J.-D. (2014).
Decoding vigilance with NIRS. PLoS One, 9(7), e101729. https://
doi.org/10.1371/journal.pone.0101729
Brackenbury, T., Ryan, T., & Messenheimer, T. (2006). Incidental
word learning in a hearing child of deaf adults. Journal of Deaf
Studies and Deaf Education, 11(1), 76–93.
Capek, C. M., Waters, D., Woll, B., MacSweeney, M., Brammer,
M. J., McGuire, P. K., … Campbell, R. (2008). Hand and mouth:
Cortical correlates of lexical processing in British Sign Language
and speechreading English. Journal of Cognitive Neuroscience,
20(7), 1220–1234.
Capirci, O., Iverson, J. M., Montanari, S., & Voltera, V. (2002).
Gestural, signed and spoken modalities in early language devel-
opment: The role of linguistic input. Bilingualism: Language and
Cognition, 5(1), 25–37.
Costa, A., & Sebastián-Gallés, N. (2014). How does the bilingual
experience sculpt the brain? Nature Reviews Neuroscience,
15(5), 336–345.
Cristia, A., Dupoux, E., Hakuno, Y., Lloyd-Fox, S., Schuetze, M.,
Kivits, J., … Minagawa-Kawai, Y. (2013). An online database of
infant functional near infrared spectroscopy studies: A community-
augmented systematic review. PLoS One, 8(3), e58906.
Cutini, S., Moro, S. B., & Bisconti, S. (2012). Functional near infra-
red optical imaging in cognitive neuroscience: An introductory
review. Journal of Near Infrared Spectroscopy, 20(1), 75–92.
Daniels, M. (1993). ASL as a factor in acquiring English. Sign Language
Studies, 78, 23–29.
DeCasper, A. J., & Fifer, W. P. (1980). Of human bonding: Newborns
prefer their mothers’ voices. Science, 208, 1174–1176.
Dehaene-Lambertz, G., Dehaene, S., & Hertz-Pannier, L. (2002).
Functional neuroimaging of speech perception in infants. Science,
298(5600), 2013–2015.
Dehaene-Lambertz, G., Hertz-Pannier, L., Dubois, J., Mériaux, S.,
Roche, A., Sigman, M., & Dehaene, S. (2006). Functional organiza-
tion of perisylvian activation during presentation of sentences in pre-
verbal infants. Proceedings of the National Academy of Sciences,
103(38), 14240–14245.
Dehaene-Lambertz, G., Montavont, A., Jobert, A., Allirol, L., Dubois,
J., Hertz-Pannier, L., & Dehaene, S. (2010). Language or music,
mother or Mozart? Structural and environmental influences on in-
fants’ language networks. Brain and Language, 114(2), 53–65.
Delpy, D. T., Cope, M., van der Zee, P., Arridge, S. R., Wray, S., &
Wyatt, J. S. (1988). Estimation of optical pathlength through tissue
from direct time of flight measurement. Physics in Medicine &
Biology, 33(12), 1433.
Di Lorenzo, R., Blasi, A., Junge, C., van den Boomen, C., van Rooijen,
R., & Kemner, C. (2019). Brain responses to faces and facial expres-
sions in 5-month-olds: An fNIRS study. Frontiers in Psychology, 10,
1240.
Di Lorenzo, R., Pirazzoli, L., Blasi, A., Bulgarelli, C., Hakuno, Y.,
Minagawa, Y., & Brigadoi, S. (2019). Recommendations for mo-
tion correction of infant fNIRS data applicable to data sets ac-
quired with a variety of experimental designs and acquisition
systems. NeuroImage, 200, 511–527.
Duncan, A., Meek, J. H., Clemence, M., Elwell, C. E., Tyszczuk, L.,
Cope, M., & Delpy, D. (1995). Optical pathlength measurements
on adult head, calf and forearm and the head of the newborn
infant using phase resolved optical spectroscopy. Physics in
Medicine & Biology, 40(2), 295.
Emberson, L. L., Zinszer, B. D., Raizada, R. D. S., & Aslin, R. N.
(2017). Decoding the infant mind: Multivariate pattern analysis
(MVPA) using fNIRS. PLoS One, 12(4), e0172500. https://doi.org/
10.1371/journal.pone.0172500
Emmorey, K. (2001). Language, cognition, and the brain: Insights
from sign language research. London: Psychology Press.
Emmorey, K., Luk, G., Pyers, J. E., & Bialystok, E. (2008). The
source of enhanced cognitive control in bilinguals: Evidence
from bimodal bilinguals. Psychological Science, 19(12),
1201–1206.
Evans, S., Kyong, J., Rosen, S., Golestani, N., Warren, J., McGettigan,
C., … Scott, S. (2013). The pathways for intelligible speech:
Neurobiology of Language
30
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
1
1
9
1
8
6
7
9
0
9
n
o
_
a
_
0
0
0
0
1
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Experience impacts brain activation for language in infancy
Multivariate and univariate perspectives. Cerebral Cortex, 24(9),
2350–2361.
Fava, E., Hull, R., & Bortfeld, H. (2014). Dissociating cortical activity
during processing of native and non-native audiovisual speech
from early to late infancy. Brain Sciences, 4(3), 471–487.
Ferjan Ramírez, N., Ramírez, R. R., Clarke, M., Taulu, S., & Kuhl,
P. K. (2017). Speech discrimination in 11-month-old bilingual
and monolingual infants: A magnetoencephalography study.
Developmental Science, 20(1), e12427. https://doi.org/10.1111/
desc.12427
Formisano, E., De Martino, F., Bonte, M., & Goebel, R. (2008).
“Who” is saying “what”? Brain-based decoding of human voice
and speech. Science, 322(5903), 970–973.
Garcia-Sierra, A., Rivera-Gaxiola, M., Percaccio, C. R., Conboy,
B. T., Romo, H., Klarman, L., … Kuhl, P. K. (2011). Bilingual
language learning: An ERP study relating early brain responses
to speech, language input, and later word production. Journal of
Phonetics, 39(4), 546–557.
Gervain, J., Mehler, J., Werker, J. F., Nelson, C. A., Csibra, G.,
Lloyd-Fox, S., … Aslin, R. N. (2011). Near-infrared spectroscopy:
A report from the McDonnell Infant Methodology Consortium.
Developmental Cognitive Neuroscience, 1(1), 22–46.
Griffith, P. L. (1985). Mode-switching and mode-finding in a hearing
child of deaf parents. Sign Language Studies, 48, 195–222.
Gu, Y., Miao, S., Han, J., Liang, Z., Ouyang, G., Yang, J., & Li, X.
(2018). Identifying ADHD children using hemodynamic responses
during a working memory task measured by functional near-infrared
spectroscopy. Journal of Neural Engineering, 15(3), 035005. https://
doi.org/10.1088/1741-2552/aa9ee9
Haynes, J.-D., & Rees, G. (2006). Neuroimaging: Decoding mental
states from brain activity in humans. Nature Reviews Neuroscience,
7(7), 523.
Heger, D., Mutter, R., Herff, C., Putze, F., & Schultz, T. (2013).
Continuous recognition of affective states by functional near in-
frared spectroscopy signals. Paper presented at the Humaine
Association C onference on A ffective C ompu ting an d
Intelligent Interaction (ACII), Geneva, Switzerland, 823–837.
https://doi.org/10.1109/ACII.2013.156
Hickok, G., Bellugi, U., & Klima, E. S. (1996). The neurobiology
of sign language and its implications for the neural basis of lan-
guage. Nature, 381(6584), 699–702.
Hofmann, K., & Chilla, S. (2015). Bimodal bilingual language de-
velopment of hearing children of deaf parents. European Journal
of Special Needs Education, 30(1), 30–46.
Hosseini, S. H., Mano, Y., Rostami, M., Takahashi, M., Sugiura, M., &
Kawashima, R. (2011). Decoding what one likes or dislikes from
single-trial fNIRS measurements. Neuroreport, 22(6), 269–273.
Hull, R., & Vaid, J. (2007). Bilingual language lateralization: A meta-
analytic tale of two hemispheres. Neuropsychologia, 45(9),
1987–2008.
Ichikawa, H., Kitazono, J., Nagata, K., Manda, A., Shimamura, K.,
Sakuta, R., … Kakigi, R. (2014). Novel method to classify hemo-
dynamic response obtained using multi-channel fNIRS measure-
ments into two groups: Exploring the combinations of channels.
Frontiers in Human Neuroscience, 8, 480.
Johnson, J. M., Watkins, R. V., & Rice, M. L. (1992). Bimodal bilin-
gual language development in a hearing child of deaf parents.
Applied Psycholinguistics, 13(1), 31–52.
Lane, H., & Webster, J. W. (1991). Speech deterioration in postlin-
gually deafened adults. The Journal of the Acoustical Society of
America, 89(2), 859–866.
Liu, H., & Cao, F. (2016). L1 and L2 processing in the bilingual brain:
A meta-analysis of neuroimaging studies. Brain, 159, 60–73.
Lloyd-Fox, S., Blasi, A., & Elwell, C. (2010). Illuminating the devel-
oping brain: The past, present and future of functional near infra-
red spectroscopy. Neuroscience & Biobehavioral Reviews, 34(3),
269–284.
Lloyd-Fox, S., Blasi, A., Elwell, C., Charman, T., Murphy, D., &
Johnson, M. H. (2013). Reduced neural sensitivity to social stimuli
in infants at risk for autism. Proceedings of the Royal Society B,
280(1758), 20123026. https://doi.org/10.1098/rspb.2012.3026
Lloyd-Fox, S., Blasi, A., McCann, S., Rozhko, M., Katus, L., Mason,
L., … The BRIGHT Project Team. (2019). Habituation and novelty
detection fNIRS brain responses in 5-and 8-month-old infants: The
Gambia and UK. Developmental Science, 22, e12817. https://doi.
org/10.1111/desc.12817
Lloyd-Fox, S., Blasi, A., Mercure, E., Elwell, C., & Johnson, M. H. (2012).
The emergence of cerebral specialization for the human voice over
the first months of life. Social Neuroscience, 7(3), 317–330.
Lloyd-Fox, S., Blasi, A., Volein, A., Everdell, N., Elwell, C. E., &
Johnson, M. H. (2009). Social perception in infancy: A near infra-
red spectroscopy study. Child Development, 80(4), 986–999.
Lloyd-Fox, S., Papademetriou, M., Darboe, M. K., Everdell, N. L.,
Wegmuller, R., Prentice, A. M., … Elwell, C. E. (2014).
Functional near infrared spectroscopy (fNIRS) to assess cognitive
function in infants in rural Africa. Scientific Reports, 4, 4740.
Lloyd-Fox, S., Richards, J. E., Blasi, A., Murphy, D. G., Elwell, C. E., &
Johnson, M. H. (2014). Coregistering functional near-infrared spec-
troscopy with underlying cortical areas in infants. Neurophotonics,
1(2), 025006. https://doi.org/10.1117/1.NPh.1.2.025006
Luu, S., & Chau, T. (2008). Decoding subjective preference from
single-trial near-infrared spectroscopy signals. Journal of Neural
Engineering, 6(1), 016003. https://doi.org/10.1088/1741-2560/
6/1/016003
MacSweeney, M., Campbell, R., Woll, B., Giampietro, V., David,
A. S., McGuire, P. K., … Brammer, M. J. (2004). Dissociating lin-
guistic and nonlinguistic gestural communication in the brain.
NeuroImage, 22(4), 1605–1618.
MacSweeney, M., Capek, C. M., Campbell, R., & Woll, B. (2008).
The signing brain: The neurobiology of sign language. Trends
in Cognitive Science, 12(11), 432–440. https://doi.org/10.1016/
j.tics.2008.07.010
Mahmoudzadeh, M., Dehaene-Lambertz, G., Fournier, M.,
Kongolo, G., Goudjil, S., Dubois, J., … Wallois, F. (2013).
Syllabic discrimination in premature human infants prior to com-
plete formation of cortical layers. Proceedings of the National
Academy of Sciences, 110(12), 4846–4851.
May, L., Byers-Heinlein, K., Gervain, J., & Werker, J. F. (2011).
Language and the newborn brain: Does prenatal language expe-
rience shape the neonate neural response to speech? Frontiers in
Psychology, 2, 222.
May, L., Gervain, J., Carreiras, M., & Werker, J. F. (2018). The spec-
ificity of the neural response to speech at birth. Developmental
Science, 21(3), e12564. https://doi.org/10.1111/desc.12564
McGettigan, C., Evans, S., Rosen, S., Agnew, Z. K., Shah, P., & Scott,
S. K. (2012). An application of univariate and multivariate
approaches in fMRI to quantifying the hemispheric lateralization
of acoustic and linguistic processes. Journal of Cognitive Neuro-
science, 24(3), 636–652.
Mercure, E., Kushnerenko, E., Goldberg, L., Bowden-Howl, H.,
Coulson, K., Johnson, M. H., & MacSweeney, M. (2018).
Language experience influences audiovisual speech integration
in unimodal and bimodal bilingual infants. Developmental
Science, 22, e12701. https://doi.org/10.1111/desc.12701
Mercure, E., Quiroz, I., Goldberg, L., Bowden-Howl, H., Coulson,
K., Gliga, T., … MacSweeney, M. (2018). Impact of language
Neurobiology of Language
31
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
1
1
9
1
8
6
7
9
0
9
n
o
_
a
_
0
0
0
0
1
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Experience impacts brain activation for language in infancy
experience on attention to faces in infancy: Evidence from unimo-
dal and bimodal bilingual infants. Frontiers in Psychology, 9, 1943.
Minagawa-Kawai, Y., van der Lely, H., Ramus, F., Sato, Y., Mazuka,
R., & Dupoux, E. (2010). Optical brain imaging reveals general
auditory and language-specific processing in early infant devel-
opment. Cerebral Cortex, 21(2), 254–261.
Misaki, M., Kim, Y., Bandettini, P. A., & Kriegeskorte, N. (2010).
Comparison of multivariate classifiers and response normalizations
for pattern-information fMRI. NeuroImage, 53(1), 103–118.
Misawa, T., Shimokawa, T., & Hirobayashi, S. (2014). Possibility for
predicting the evaluation of product price in the prefrontal cortex:
A NIRS study. International Journal of Engineering and Innovative
Technology, 4, 153–160.
Moon, C., Cooper, R. P., & Fifer, W. P. (1993). Two-day-olds prefer their
native language. Infant Behavior and Development, 16, 495–500.
Mullen, E. M. (1995). Mullen Scales of Early Learning. Circle Pines,
MN: American Guidance Service.
Murphy, J., & Slorach, N. (1983). The language development of pre-
preschool hearing children of deaf parents. International Journal of
Language & Communication Disorders, 18(2), 118–127.
Norman, K. A., Polyn, S. M., Detre, G. J., & Haxby, J. V. (2006).
Beyond mind-reading: Multi-voxel pattern analysis of fMRI data.
Trends in Cognitive Sciences, 10(9), 424–430.
Obrig, H., & Villringer, A. (2003). Beyond the visible—imaging the hu-
man brain with light. Journal of Cerebral Blood Flow & Metabolism,
23(1), 1–18.
Pena, M., Maki, A., Kovac(cid:1)ic´ , D., Dehaene-Lambertz, G., Koizumi, H.,
Bouquet, F., & Mehler, J. (2003). Sounds and silence: An optical
topography study of language recognition at birth. Proceedings of
the National Academy of Sciences, 100(20), 11702–11705.
Perani, D., Paulesu, E., Galles, N. S., Dupoux, E., Dehaene, S., Bettinardi,
V., … Mehler, J. (1998). The bilingual brain: Proficiency and age of
acquisition of the second language. Brain, 121(10), 1841–1852.
Perani, D., Saccuman, M. C., Scifo, P., Anwander, A., Spada, D.,
Baldoli, C., … Friederici, A. D. (2011). Neural language networks
at birth. Proceedings of the National Academy of Sciences, 108(38),
16056–16061.
Pereira, F., Mitchell, T., & Botvinick, M. (2009). Machine learning clas-
sifiers and fMRI: A tutorial overview. NeuroImage, 45(1), S199–S209.
Petitto, L. A., Berens, M., Kovelman, I., Dubins, M. H., Jasinska, K., &
Shalinsky, M. (2012). The “Perceptual Wedge Hypothesis” as the ba-
sis for bilingual babies’ phonetic processing advantage: New insights
from fNIRS brain imaging. Brain and Language, 121(2), 130–143.
Petitto, L. A., Katerelos, M., Levy, B. G., Gauna, K., Tétreault, K., &
Ferraro, V. (2001). Bilingual signed and spoken language acquisi-
tion from birth: Implications for the mechanisms underlying early
bilingual language acquisition. Journal of Child Language, 28(2),
453–496.
Petitto, L. A., Zatorre, R. J., Gauna, K., Nikelski, E. J., Dostie, D., &
Evans, A. C. (2000). Speech-like cerebral activity in profoundly
deaf people processing signed languages: Implications for the
neural basis of human language. Proceedings of the National
Academy of Sciences, 97(25), 13961–13966.
Połczyn´ ska, M. M., Japardi, K., & Bookheimer, S. Y. (2017).
Lateralizing language function with pre-operative functional mag-
netic resonance imaging in early proficient bilingual patients. Brain
and Language, 170, 1–11.
Price, C. J. (2010). The anatomy of language: A review of 100 fMRI
studies published in 2009. Annals of the New York Academy of
Sciences, 1191(1), 62–88.
Riecker, A., Wildgruber, D., Dogil, G., Grodd, W., & Ackermann,
H. (2002). Hemispheric lateralization effects of rhythm imple-
mentation during syllable repetitions: An fMRI study. Neuro-
Image, 16(1), 169–176.
Ruxton, G. D., & Neuhäuser, M. (2013). Improving the reporting of
P-values generated by randomization methods. Methods in
Ecology and Evolution, 4(11), 1033–1036.
Sandler, W., & Lillo-Martin, D. (2006). Sign language and linguistic
universals. Cambridge, UK: Cambridge University Press.
Sato, H., Hirabayashi, Y., Tsubokura, H., Kanai, M., Ashida, T.,
Konishi, I., … Maki, A. (2012). Cerebral hemodynamics in new-
born infants exposed to speech sounds: A whole-head optical to-
pography study. Human Brain Mapping, 33(9), 2092–2103.
Schiff, N. B., & Ventry, I. M. (1976). Communication problems in
hearing children of deaf parents. Journal of Speech Hearing
Disorders, 41(3), 348–358.
Schiff-Meyers, N. (1993). Hearing children of deaf parents. In D.
Bishop & K. Mogford (Eds.), Language development in excep-
tional circumstances. Hove, UK: Psychology Press.
Sebastián-Gallés, N., Albareda-Castellot, B., Weikum, W. M., &
Werker, J. F. (2012). A bilingual advantage in visual language dis-
crimination in infancy. Psychological Science, 23(9), 994–999.
Senju, A., Tucker, L., Pasco, G., Hudry, K., Elsabbagh, M., Charman,
T., & Johnson, M. H. (2013). The importance of the eyes:
Communication skills in infants of blind parents. Proceedings of
the Royal Society B, 280(1760), 20130436. https://doi.org/10.1098/
rspb.2013.0436
Shultz, S., Vouloumanos, A., Bennett, R. H., & Pelphrey, K. (2014).
Neural specialization for speech in the first months of life. Develop-
mental Science, 17(5), 766–774.
Vannasing, P., Florea, O., González-Frankenberger, B., Tremblay,
J., Paquette, N., Safi, D., … Lassonde, M. (2016). Distinct hemi-
spheric specializations for native and non-native languages in
one-day-old newborns identified by fNIRS. Neuropsychologia,
84, 63–69.
Waldstein, R. S. (1990). Effects of postlingual deafness on speech pro-
duction: Implications for the role of auditory feedback. The Journal
of the Acoustical Society of America, 88(5), 2099–2114.
Werker, J. (2012). Perceptual foundations of bilingual acquisition in
infancy. Annals of the New York Academy of Sciences, 1251(1),
50–61.
Woll, B., & Kyle, J. G. (1989). Communication and language devel-
opment in children of deaf parents. In S. von Tetzchner,
L. S. Siegel, & L. Smith (Eds.), The social and cognitive aspects
of normal and atypical language development (pp. 129–144).
New York: Springer-Verlag.
Zinszer, B. D., Bayet, L., Emberson, L. L., Raizada, R. D., & Aslin,
R. N. (2017). Decoding semantic representations from functional
near-infrared spectroscopy signals. Neurophotonics, 5(1),
011003. https://doi.org/10.1117/1.NPh.5.1.011003
Neurobiology of Language
32
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
1
1
9
1
8
6
7
9
0
9
n
o
_
a
_
0
0
0
0
1
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3