RESEARCH ARTICLE
Effects of Sleep on Language and Motor
Consolidation: Evidence of Domain
General and Specific Mechanisms
Dafna Ben-Zion1,2,3,4
, Ella Gabitov5
, Anat Prior1,2
, and Tali Bitan3,4,6,7
1Department of Learning Disabilities, University of Haifa, Haifa, Israel
2Edmond J. Safra Brain Research Center for the Study of Learning Disabilities, University of Haifa, Haifa, Israel
3Institute of Information Processing and Decision Making, University of Haifa, Haifa, Israel
4The Integrated Brain and Behavior Research Center (IBBRC), University of Haifa, Haifa, Israel
5McConnell Brain Imaging Center, Montreal Neurological Institute, McGill University, Montreal, Quebec, Canada
6Department of Psychology, University of Haifa, Haifa, Israel
7Department of Speech Language Pathology, University of Toronto, Toronto, Ontario, Canada
Keywords: consolidation, language, morphology, sleep, motor, grammar
ABSTRACT
The current study explores the effects of time and sleep on the consolidation of a novel
language learning task containing both item-specific knowledge and the extraction of
grammatical regularities. We also compare consolidation effects in language and motor
sequence learning tasks, to ask whether consolidation mechanisms are domain general. Young
adults learned to apply plural inflections to novel words based on morphophonological rules
embedded in the input, and learned to type a motor sequence using a keyboard. Participants
were randomly assigned into one of two groups, practicing each task during either the morning
or evening hours. Both groups were retested 12 and 24 hours post-training. Performance on
frequent trained items in the language task stabilized only following sleep, consistent with a
hippocampal mechanism for item-specific learning. However, regularity extraction, indicated
by generalization to untrained items in the linguistic task, as well as performance on motor
sequence learning, improved 24 hours post-training, irrespective of the timing of sleep. This
consolidation process is consistent with a frontostriatal skill-learning mechanism, common
across the language and motor domains. This conclusion is further reinforced by cross-domain
correlations at the individual level between improvement across 24 hours in the motor task
and in the low-frequency trained items in the linguistic task, which involve regularity
extraction. Taken together, our results at the group and individual levels suggest that some
aspects of consolidation are shared across the motor and language domains, and more
specifically, between motor sequence learning and grammar learning.
INTRODUCTION
Creating durable and accessible knowledge is essential to second language learning, as it is for
learning other cognitive and motor skills. Memory research has identified consolidation as a
key process in establishing such long-term representations (Dudai et al., 2015). The role of
sleep in memory consolidation has been investigated in declarative / episodic memories
(Mölle et al., 2011; Plihal & Born, 1997; Wilhelm et al., 2011) and in procedural motor
sequence learning (Brashers-Krug et al., 1996; Korman et al., 2003, 2007; Robertson,
a n o p e n a c c e s s
j o u r n a l
Citation: Ben-Zion, D., Gabitov, E.,
Prior, A., & Bitan, T. (2022). Effects of
sleep on language and motor
consolidation: Evidence of domain
general and specific mechanisms.
Neurobiology of Language, 3(2),
180–213. https://doi.org/10.1162/nol_a
_00060
DOI:
https://doi.org/10.1162/nol_a_00060
Supporting Information:
https://doi.org/10.1162/nol_a_00060
Received: 15 January 2021
Accepted: 21 October 2021
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Author:
Dafna Ben-Zion
dafmail@gmail.com
Handling Editor:
Kate Watkins
Copyright: © 2021
Massachusetts Institute of Technology
Published under a Creative Commons
Attribution 4.0 International
(CC BY 4.0) license
The MIT Press
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
2
1
8
0
1
9
8
9
3
2
6
n
o
_
a
_
0
0
0
6
0
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Consolidation in language and motor learning
Consolidation:
The process by which new and
initially labile memories transform
into long-term memories.
Inflectional Morphology:
The process by which the
grammatical category of words is
changed, for example, by adding
affixes.
Pascual-Leone, & Miall, 2004; Stickgold, 2005), but the factors that determine when sleep
plays a critical role in consolidation are still under debate (Cordi & Rasch, 2021; Diekelmann
& Born, 2010; King et al., 2017; Schmidt et al., 2020). Sleep dependent consolidation in lan-
guage learning has been studied mainly in vocabulary learning (Davis & Gaskell, 2009;
Dumay & Gaskell, 2007; James et al., 2017; Schreiner & Rasch, 2017), which relies on
item-specific, declarative hippocampal mechanisms. Grammar learning, in contrast, is sug-
gested to rely to a greater extent on procedural non-hippocampal learning mechanisms, which
may be akin to motor skill learning (Hedenius et al., 2011; Ullman, 2015). In the current study,
we examine the effect of sleep on the consolidation of a morphological learning task that
enables us to study item-specific knowledge, the extraction of grammatical regularities, and
the interactions between them. The study also compares the effect of sleep on language learn-
ing to its effect on motor sequence learning, to identify the shared and distinct factors that
affect consolidation in both domains. Finally, we ask to what extent individual differences
determine consolidation across the language and motor domains.
Memory Consolidation
Memory consolidation is a process that transforms new and initially labile memories into more
stable representations that become integrated into the network of pre-existing long-term mem-
ories (Diekelmann & Born, 2010). This process is time-dependent and can also be sleep-
dependent (Dudai et al., 2015). Memory consolidation is considered sleep-dependent when
a period of post-learning sleep enhances performance or qualitatively changes the represen-
tation of information compared with a wake interval of equal length (Korman et al., 2007;
Rasch et al., 2007; Tucker et al., 2006; Walker et al., 2003). While evidence for sleep-
dependent consolidation was shown in both hippocampal and non-hippocampal-dependent
tasks, the type of learning may have an effect on the pattern of consolidation (Diekelmann
et al., 2009; Diekelmann & Born, 2010; Song, 2009).
In the context of hippocampal-based learning, the complementary learning system (CLS)
framework (McClelland, 2013; McClelland et al., 1995) and the active system consolidation
hypothesis, suggest that memories initially encoded in parallel in neocortical networks and in
the hippocampus are repeatedly reactivated in the hippocampus during sleep, and thereby
become gradually redistributed within the neocortex (Diekelmann & Born, 2010). The reorga-
nization in the neocortex during consolidation enables the generation of new associations
through the extraction of overlapping features from separate events and thereby eventually
facilitates novel inferences that afford generalization and insight (Diekelmann & Born, 2010).
Consolidation of non-hippocampal learning, including perceptual learning (Censor et al.,
2006; Fenn et al., 2003) and motor skill acquisition (described in detail below), can also ben-
efit from sleep (King et al., 2017; Korman et al., 2003; Walker & Stickgold, 2004). Circuit-level
redistribution of experience-dependent representational information over time can occur in
tasks that can be acquired independently of the hippocampus and the medial temporal lobe
(Dudai et al., 2015). For example, the synaptic homeostasis hypothesis (Tononi & Cirelli,
2006, 2014) suggests that information encoding during wakefulness leads to a net increase in
synaptic strength in the brain. Sleep then serves to globally downscale synaptic strength. As a
result of this renormalization process, weakly activated synapses become virtually ineffective
after sleep, whereas strongly activated synapses are preserved and may consolidate (Crick &
Mitchison, 1983; Tononi & Cirelli, 2006). Nevertheless, there is also evidence that corticos-
triatal and hippocampal-dependent memory systems do not act independently, but rather
interact during consolidation (Albouy et al., 2015; Coynel et al., 2010; Debas et al., 2014),
Neurobiology of Language
181
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
2
1
8
0
1
9
8
9
3
2
6
n
o
_
a
_
0
0
0
6
0
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Consolidation in language and motor learning
so that even tasks that do not depend on the hippocampal system during initial phases of
acquisition, may show hippocampal-based sleep dependent consolidation (King et al.,
2017; Klinzing et al., 2019; Sawangjit et al., 2018).
Behavioral changes following consolidation processes are expressed as enhancement in
performance or as stabilization (Censor et al., 2012; Diekelmann & Born, 2010; Ellenbogen
et al., 2007; Ellenbogen, Hulbert et al., 2006; Fischer et al., 2006; Korman et al., 2007; Rasch
& Born, 2013; Wagner et al., 2004). While there is a debate on whether sleep-related stabili-
zation and offline gains represent similar or different processes, there is evidence that both are
related to an active role of sleep in consolidation (Diekelmann & Born, 2010).
Consolidation in Language Learning
To date, most research on sleep-dependent consolidation in language learning focused on
vocabulary learning (Gais et al., 2006; Henderson et al., 2012; James et al., 2017; Landi
et al., 2018), which is typically associated with item-specific knowledge and episodic learning.
The adaptation of the CLS framework to word learning (Davis & Gaskell, 2009) suggests that a
newly learned word is initially stored as a distinct episodic trace relying on the hippocampus,
but following an offline consolidation period it becomes integrated with the existing lexicon in
neocortical long-term semantic memory. Support for this notion comes from studies showing
that the integration of new linguistic knowledge into the existing lexicon is sleep-dependent
(Dumay & Gaskell, 2007; Henderson et al., 2013; Tamminen et al., 2010). To wit, perfor-
mance on recall tests of the newly learned vocabulary, 12 to 24 hours post-training, shows
a stabilizing effect when sleep is afforded immediately after training compared to an equiva-
lent period of wakefulness (Gais et al., 2006; Tamminen et al., 2010). Moreover, other studies
even reported offline gains, measured as an increase in the number of recalled words or a
reduction in reaction time following a night’s sleep compared to performance immediately
after training (Dumay & Gaskell, 2007; Heim et al., 2017; Henderson et al., 2012).
In contrast to vocabulary, grammar learning is suggested to rely to a greater extent on pro-
cedural skill learning mechanisms associated with frontostriatal brain regions (Hedenius et al.,
2011; Ullman, 2015). The term procedural learning is often associated with implicit learning,
or lack of awareness, which is not necessarily the case for regularity extraction in the current
study, and in grammar learning more generally. Therefore, we avoid using this term here, and
instead we refer to skill learning. Given the differential role that sleep may play in the consol-
idation of hippocampal- vs. non-hippocampal-dependent learning (Diekelmann & Born,
2010), it is important to examine the role of sleep in the consolidation of grammar learning.
Studies examining the effect of sleep on statistical or serial order learning outside the language
domain report contradicting findings, as some found that sleep increased recognition of new
rule-based sequences (Durrant et al., 2011; Lerner & Gluck, 2019; Nieuwenhuis et al., 2013),
but others found no benefit (Szmalec et al., 2012).
Very few studies investigated sleep dependent consolidation following learning of grammat-
ical and morphological regularities in language (Batterink et al., 2014; Kim & Fenn, 2020;
Mirković & Gaskell, 2016; Tamminen et al., 2010), and here too there are contradicting results.
Two studies found no benefit of sleep for extracting morphological regularities in a novel lan-
guage using a short nap (Mirković & Gaskell, 2016) or nocturnal sleep deprivation (Tamminen
et al., 2020). Only one study found an association between the extraction of grammatical reg-
ularities and sleep (Batterink et al., 2014). In this study, participants’ sleep was monitored dur-
ing an afternoon nap that followed training on new syntactic rules. The results showed that the
amounts of slow-wave and rapid-eye-movement sleep predicted participants’ sensitivity to
Neurobiology of Language
182
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
2
1
8
0
1
9
8
9
3
2
6
n
o
_
a
_
0
0
0
6
0
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Consolidation in language and motor learning
Item-specific learning:
The memorization of exemplars as
whole units.
implicit regularities embedded in the stimuli (Batterink et al., 2014). However, because only
one group of participants was examined, the effect of sleep cannot be compared to the effect
of wake. A recent study examined the effect of explicit awareness on sleep-dependent consol-
idation following syntactic rule learning in a second language (Kim & Fenn, 2020). They found
that only those participants who were aware of the grammar rules at the end of training
improved their performance following sleep, suggesting that the benefit of sleep for extraction
of grammatical regularities during consolidation depends on explicit awareness.
Given these mixed findings, the current study explores the effect of sleep on consolidation
in a language task that affords both item-specific learning and extraction of morphological
regularities. These two aspects of language learning may involve different consolidation mech-
anisms and thus may be differentially affected by sleep. The effects of sleep on consolidation in
the language task will also be compared to these effects in a motor sequence learning task in
order to examine domain generality of consolidation mechanisms.
Consolidation in Motor Learning
Motor skill learning is one of the most studied domains in relation to sleep dependent consol-
idation. It is broadly accepted that motor learning takes place not only online (i.e., during task
practice) but also offline (i.e., between training sessions) in the absence of any further practice
(Doyon et al., 2018; Karni et al., 1998; King et al., 2017; Nettersheim et al., 2015; Schmidt
et al., 2020). However, the role of sleep in such offline processes is still hotly contested.
Whereas sleep is generally beneficial in consolidating skills, it might be critical only in
some motor tasks, particularly those requiring movement sequence learning and generating
new movement routines (Debas et al., 2010, 2014; Korman et al., 2007; see also recent
reviews by Cordi & Rasch, 2021; Dudai et al., 2015; Hu et al., 2020; King et al., 2017; Rasch
& Born, 2013). This beneficial effect of sleep can be reflected behaviorally either by enhance-
ment or stabilizing the skill levels that were achieved by the end of training (Maltry et al.,
2020; Robertson, 2012). For example, improved performance after post-training sleep was
documented in finger motor sequence tasks (Fischer et al., 2002, 2005; Korman et al.,
2007; Walker et al., 2002).
Recently, several key findings supporting the idea of sleep-dependent consolidation of
motor sequence knowledge have been called into question (for the results of a meta-analysis,
see Pan & Rickard, 2015, and a review by Cellini, 2017). Some studies suggest that sleep does
not improve performance of the trained motor sequence (Brawn et al., 2010; Nettersheim
et al., 2015; Rickard et al., 2008), but rather facilitates recovery of a previously attained skill
level after it has been degraded during the post-training wake interval (Brawn et al., 2010).
Others propose that the simple passage of time and time spent in sleep may facilitate different
aspects of motor sequence knowledge (King et al., 2017), and that participants’ awareness of
the learned sequence may determine the necessity of sleep in consolidating that sequence
(Robertson, Pascual-Leone, & Press, 2004).
Associations Between Language and Motor Consolidation
In the current study, we directly compare the effect of sleep on consolidation of motor and
language tasks and ask whether consolidation processes might be shared across domains.
The idea of shared learning mechanisms across domains is supported by the notion that motor
sequence learning and language grammar learning may both rely on procedural learning
(Hedenius et al., 2011; Ullman, 2015). Thus, although the elements of each task are different,
Neurobiology of Language
183
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
2
1
8
0
1
9
8
9
3
2
6
n
o
_
a
_
0
0
0
6
0
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Consolidation in language and motor learning
i.e., morphemes versus finger movements, in both tasks participants learn to link these elements
based on regular patterns. Several lines of evidence lead to the hypothesis of a shared consol-
idation process in motor sequence and grammar learning, including common neural substrates
and learning mechanisms across domains, as well as shared properties of consolidation.
First, there is evidence that both motor sequence learning and language learning rely to
some extent on similar brain areas, including the supplementary motor area (SMA) and the
basal ganglia. The SMA is involved in the integration of sequential elements into unified rep-
resentations, in both motor (Cona & Semenza, 2017; Orban et al., 2010; Wymbs & Grafton,
2013) and language tasks (Ferstl et al., 2008; Hertrich et al., 2016; Nakai et al., 1999; Needle
et al., 2015; Segaert et al., 2012), and specifically in the sequential integration of linguistic
elements into higher order representations (Cona & Semenza, 2017). The basal ganglia are
also involved in motor sequence learning (Albouy et al., 2008; Gabitov et al., 2015; Hikosaka
et al., 2002; Lehéricy et al., 2005; Penhune & Steele, 2012) and language learning tasks (Chan
et al., 2013; Nevat et al., 2017; Ullman & Pierpont, 2005).
Evidence for these shared mechanisms also comes from children with developmental lan-
guage disorder showing deficits in motor serial reaction time tasks, and other tasks associated
with procedural learning (see meta-analysis by Lum et al., 2014). These findings are the basis
for the procedural deficit hypothesis (Lum et al., 2014; Ullman & Pierpont, 2005), which sug-
gests that language and grammar deficits in developmental language disorders are linked to a
more general deficit in procedural learning mechanisms, rooted in the SMA and frontostriatal
circuits (Ullman & Pierpont, 2005). However, a recent study states that the implicit learning
tasks that are often used to test procedural learning capacities have low reliability (West et al.,
2017), raising some questions regarding this interpretation. Similarly, patients with Parkinson’s
disease, which is characterized by motor control deficits, show impairments in morphological
processing (Eyigoz et al., 2020), again suggesting a shared neural substrate for learning and
processing in the two domains.
Second, consolidation of both motor and language learning are associated with similar
sleep mechanisms. Specifically, time spent in stage 2 sleep and specific characteristics of stage
2 spindles are associated with improved performance after sleep in motor tasks (Morin et al.,
2008; Peters et al., 2008), a vocabulary learning task (Mirković & Gaskell, 2016), and other
hippocampal-dependent episodic memory tasks (Mednick et al., 2013; Schabus et al., 2004).
Finally, individual differences in various participant characteristics have been linked to con-
solidation in different domains, raising the possibility of a shared underlying mechanism. For
example, age is a factor influencing consolidation—adolescents showed a smaller consoli-
dation effect than young adults in vocabulary learning (Landi et al., 2018), and older adults
showed reduced consolidation of motor sequence learning following sleep than did younger
adults (Spencer et al., 2007; see also Wilhelm et al., 2008, 2013). Similarly, general cogni-
tive abilities such as working memory or intelligence affect consolidation in episodic mem-
ory tasks (Fenn & Hambrick, 2012, 2015). Finally, individual differences in non-motor
domains, such as those reflected in attention deficit disorder or dyslexia, are also linked
to reduced consolidation in motor learning (Adi-Japha et al., 2011; Needle et al., 2015;
Wilhelm et al., 2012).
Taken together, these findings suggest that consolidation processes in motor sequence
learning and language learning might be governed by similar underlying mechanisms. How-
ever, a recent study that directly examined consolidation of a Hebb repetition task (which
bears similarities to language learning) and of a motor serial reaction time task, though without
manipulation of sleep, did not find any cross-domain correlations (Henderson & Warmington,
Neurobiology of Language
184
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
2
1
8
0
1
9
8
9
3
2
6
n
o
_
a
_
0
0
0
6
0
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Consolidation in language and motor learning
Phonology:
The sound system of a language.
2017). This finding calls into question the idea of a general consolidation ability. The current
study directly addresses the questions of sleep dependency and domain generality of consol-
idation, by comparing the effect of sleep on morphological learning and on motor sequence
learning and examining the correlations between the two.
Objectives of the Current Study
In the current study, we investigate the effect of sleep on consolidation of learning morpho-
logical regularities in a novel language and compare it to the effect of sleep on motor sequence
learning, in a typical adult population. For the motor learning, we use a computerized version
(Gabitov et al., 2019) of a finger sequence learning task that has previously been shown to be
affected by sleep (Fischer et al., 2002, 2005; Karni et al., 1998; Korman et al., 2007). For the
language task, we use an artificial language paradigm in which participants learn to apply
plural inflections to novel words based on morphophonological rules embedded in the input,
adapted from previous studies (Ben Zion et al., 2019; Nevat et al., 2017, 2018). Performance
in early stages of learning in a similar task was previously shown to rely on frontostriatal areas
(Nevat et al., 2017).
We manipulated the timing of training so that one group was trained in the morning (wake-
first) and the other group was trained in the evening (sleep-first), and both groups were tested
12 and 24 hours post-training, to determine the effect of sleep versus the passage of time. Par-
ticipants performed the language and motor tasks with the same schedule. This design allowed
us to probe the existence and characteristics of the posited general consolidation ability in two
complementary ways. First, at the group level, we examined the effect of sleep on consolida-
tion in both the motor and language domains in the same group of participants. Second, we
examined whether individuals’ consolidation gains in the two tasks are correlated, to test
whether the ability to consolidate newly encoded knowledge relies on learning mechanisms
that are common to both domains.
The current version of the artificial language task was constructed such that it allows us to
differentiate, to some extent, between item-specific learning and the extraction of regularities,
i.e., the learning of the grammar. To this end, we manipulated the frequency of presentations
of items during training. Hence, performance on high frequency items reflects processes of
item-specific learning more than performance on low frequency items, which relies on both
item-specific and regularity extraction components. Previous studies also support this differ-
entiation between high and low frequency presentation, by demonstrating that the consoli-
dation of items tends to benefit differently from sleep, dependent on the strength of the initial
encoding (Denis et al., 2020; Diekelmann et al., 2009; Stickgold, 2009). In contrast to per-
formance on trained items, which partially reflects item specific learning, the generalization
to untrained items was solely dependent on the ability to extract the regularity from the
trained input.
We predicted that sleep will promote consolidation in both the language and motor tasks,
but the benefit of sleep may be reflected differently for each task. In the language task, we
predicted that sleep will result in the stabilization of item-specific knowledge (evident by per-
formance on high frequency items), as was previously evident in vocabulary learning and epi-
sodic memory tasks (Gais et al., 2006; Payne et al., 2012; Peiffer et al., 2020; Tamminen et al.,
2010). Based on findings of sleep related offline gains in motor sequence learning tasks
(Fischer et al., 2002, 2005; Karni et al., 1998; Korman et al., 2007) and the effect of sleep
on grammar learning (Batterink et al., 2014), we predicted that sleep will improve performance
in the motor task and in linguistic tasks that rely on the extraction of regularities, namely low
Neurobiology of Language
185
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
2
1
8
0
1
9
8
9
3
2
6
n
o
_
a
_
0
0
0
6
0
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Consolidation in language and motor learning
frequency trained items and generalization to untrained items. We also expected this aspect of
grammar learning to be correlated with motor sequence learning.
MATERIALS AND METHODS
Participants
Forty-four healthy young adults participated in the study. Of these, 38 (18–33 years old;
mean = 24.295, SD = 3.32; 29 women) are included in the analysis of the language learning
task, and of them 36 participants are also included in the analysis of the motor sequence
learning task. Three participants dropped out due to technical and personal issues, and two
participants were excluded due to low performance (1.5 SD below the mean) in two reading
screening tests. Each participant was randomly assigned to either the morning wake-first
group (n = 20) or the evening sleep-first group (n = 18). The group assignment for each
participant was the same for the motor and language tasks. Nineteen and 17 participants
from the wake-first and sleep-first groups, respectively, completed both the linguistic and
motor learning tasks with a minimum of 2 weeks apart. The order of the tasks was counter-
balanced across participants so that half of the participants in each group participated in the
language task first, and half in the motor task first.
All participants were native Hebrew speakers and spoke at least one other language
(English) as a foreign language. They were right-handed (self-report) and had normal or
corrected-to-normal vision and no hearing deficits. The screening procedure included self-
report of no history of neurological or psychiatric illness, no learning disability or attention
disorder, no addiction to alcohol (no more than one alcoholic beverage a day), and being a
non-smoker. In addition, to ensure good quality of sleep, exclusionary criteria included the use
of medication that affects sleep, mid-day naps, pregnancy, working night shifts, trans-Atlantic
trips within 3 months prior to the study, drinking more than 3 caffeinated beverages per day,
and obesity (BMI > 30; group mean = 22.58; SD = 2.45). Sleep disorders were ruled out by the
Mini Sleep Questionnaire (Natale et al., 2014; Zoomer et al., 1985), and circadian tendency
was measured using the Hebrew version of the Morningness-Eveningness Questionnaire
(Horne & Ostberg, 1976); both showed no significant differences between the groups. Partic-
ipants maintained between 6 and 9 hours of proper nocturnal sleep and abstained from
caffeinated and alcoholic drinks 24 hours prior to the experiment and during the experiment
itself (which lasted 36 hours), as reported in a sleep log.
Participants’ normal reading level was confirmed by two screening tests: the one-minute
word reading test and the one-minute pseudoword reading test (Shatil, 1995, 1997). In these
tests, participants read a list of pointed words or pseudowords as quickly and as accurately as
possible within 1 minute, and the number of correct items was counted. Only two participants
met the exclusion criterion of more than 1.5 SD below the average of our local norms (Weiss
et al., 2015) in both tests, and were excluded.
In order to make sure the two groups did not differ on phonological and morphological
awareness as well as on working memory ability, the following tests were conducted: (1)
the Phoneme Deletion Test for Pseudowords (Ben-Dror & Shani, 1996); (2) morphological
awareness was tested using two production tests: a word-inflection task (Cohen-Mimran,
2009) and a word-derivation task (Raz-Salzburg & Ravid, 2009); and (3) short term and work-
ing memory were assessed using the digit span task from the Wechsler Adult Intelligence Scale
(Wechsler, 1955) and were collapsed to a normalized working memory z score. The two
groups did not differ significantly on any of these measures (Table 1).
Neurobiology of Language
186
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
2
1
8
0
1
9
8
9
3
2
6
n
o
_
a
_
0
0
0
6
0
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Consolidation in language and motor learning
Table 1. Mean score and standard deviation per group in screening tests
Screening test
Word reading test
Pseudoword reading test
Phoneme Deletion Test for Pseudowords
Inflectional morphology
Derivational morphology
Working memory
Words per minute
Nonwords per minute
Accuracy
Accuracy
Accuracy
z score
Wake-first group
SD
18.6
mean
107.1
Sleep-first group
SD
17.6
mean
104.5
56.5
92.2%
87.7%
96.2%
10.4
11.9
11.7%
9.3%
3.2%
2.7
54.7
92.7%
88.0%
96.6%
11.6
12.4
8.0%
11.1%
3.8%
2.9
This study was reviewed and approved by an ethics committee. All participants gave writ-
ten informed consent and received financial compensation.
Design and Experimental Procedure
The study consisted of two experimental tasks: (1) language learning, and (2) motor sequence
learning, conducted on separate days with at least 2 weeks apart. The experimental protocol
for each task was composed of three sessions separated by 12 hours. First, a screening session
was administered (usually conducted concurrently with the first experimental session), in which
baseline parameters were measured. Next, participants learned either to generate a motor
sequence or to inflect items according to a new linguistic regularity. For both tasks, the first
session included training and the second and third sessions included only retests. For the motor
task, generalization of the learned skill was measured only at the end of the last session,
whereas for the linguistic task, generalization was tested at the end of each session. Morning
and evening sessions were conducted between 7–9 a.m. and 7–9 p.m., respectively (Figure 1A).
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
2
1
8
0
1
9
8
9
3
2
6
n
o
_
a
_
0
0
0
6
0
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Language learning task
The task and stimuli were adapted from previous studies (Ben Zion et al., 2019; Nevat et al.,
2017, 2018). The trained items consisted of 36 novel words, which were aurally presented
together with pictures of the objects they refer to (e.g., real objects like an apple). All items
consisted of two syllables (CVCVC) in their singular form (the stem). Plural forms were created
by applying one of three possible ( VC) suffixes to the stem (-an, -esh, -ur); each suffix was
applied to one third of the items (12 items).
Pairings of stems and suffixes were determined by the last two phonemes of the stem, the
phonological cue, such that each suffix was associated with two phonological cues. For exam-
ple, stems ending with /oz/ and with /ap/ took the suffix -an; thus, the plural for kutoz was
kutozan and the plural for nifap was nifapan (see full details in Table 2). However, 6 trained
items (2 of each suffix) did not follow these rules and were included in order to increase the
difficulty of learning the regularities, thus mimicking the inconsistency of natural languages.
These exception words took a different suffix from the one predicted by the words’ phonolo-
gical cue. For example, although most words ending with the cue /oz/ received the suffix -an,
the stem nuboz received the suffix -esh forming the word nubozesh, which did not adhere to
the general inflection rule. These exception words were removed from the analysis because
previous studies found that these words show lower accuracy than consistent words (Nevat
et al., 2017), and their small number does not allow for a separate analysis.
Neurobiology of Language
187
Consolidation in language and motor learning
Figure 1. Overall design of the experiment. (A) The wake-first group performed training (T) of either the motor or the linguistic learning task,
during the morning between 7 and 9 a.m., retested (R) after 12 hours of wakefulness, and retested again after a period of sleep. The sleep-first
group preformed training during the evening, retested following a period of sleep, and retested again after an additional 12 hours. (B) Language
learning task design, consisting of three sessions. (C) Motor sequence learning task consisting of three sessions. T-S refers to Trained Sequence
while U-S refers to Untrained Sequence; T-H refers to Trained Hand while U-H refers to Untrained Hand.
During training, half of the items (18; 6 of each suffix, all with consistent cues) were pre-
sented 3 times per block (high-frequency items), while the other half were presented only once
per block (low-frequency items). One third (6) of the low-frequency items were exception
words (Table 2). Since the exception words were excluded, the analysis included fewer low
frequency than high frequency words (12 and 18 respectively). Frequency was manipulated in
order to differentiate between item-specific learning and regularity extraction. Specifically, in
the current design, since both high and low frequency items share the same regularities, the
differences between them can only be attributed to item-specific learning, which plays a more
prominent role in high frequency items.
Table 2.
Trained words
Suffix -an
Suffix -esh
Suffix -ur
High frequency
laloz
refoz
nifap
tuvap
bomoz
datsap
Low frequency
gishoz
Exceptions
kutoz
gukid
venap
sogap
lagis
motsid
rilid
zutid
lebid
panid
nuboz
sibaf
tegaf
libok
torok
dumaf
shuzok
bodaf
gazaf
zelok
navok
delok
potaf
pikis
fazis
reshis
molis
vusis
filap
Note. List of trained items, presented by suffix and frequency (high and low). Exception items, presented at a low
frequency, received unpredictable suffixes and were removed from the analysis.
Neurobiology of Language
188
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
2
1
8
0
1
9
8
9
3
2
6
n
o
_
a
_
0
0
0
6
0
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Consolidation in language and motor learning
Generalization of the morphological regularity was tested by inflecting untrained items. At
the end of each session, participants were also tested on the inflection of 30 unique untrained
(new) items, resulting in a total of 90 untrained items across all sessions. In each generalization
test of 30 items, there were 5 words containing each of the 6 phonological cues, resulting in 10
items receiving each of the three suffixes.
The language task was performed over three sessions: (1) the first session, which included
the instruction block, the pre-training trained items test, three blocks of training including feed-
back, the post-training trained items test, and finally a generalization test on untrained items;
(2) the 12-hr retest session, which consisted of a trained items test and a generalization test;
and (3) the 24-hr retest session, which consisted of trained items and generalization tests, and
an explicit rule knowledge questionnaire (Figure 1B).
In the instruction block, each of the 36 training items was presented once. Upon key press,
the singular form was presented aurally together with an image of a real object to be learned as
its referent on the screen. The singular form was followed by a visual cue consisting of two
asterisks (**), indicating the plural form of the word would soon be presented. The plural form
was then presented aurally, followed by the presentation of a question mark, indicating that
participants were to repeat the plural form they had just heard. The question mark remained
on the screen for a maximal duration of 5 sec, or until a vocal response was detected
(Figure 2A).
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
2
1
8
0
1
9
8
9
3
2
6
n
o
_
a
_
0
0
0
6
0
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 2. Design of trials. (A) Instruction block: each item was presented once, together with the picture that refers to its meaning. (B) Trained-
items tests: each item was tested once and required the judgment of correctly and incorrectly inflected plural forms. (C) Training: participants
produced the inflected forms of the trained items, receiving feedback. (D) Untrained-items tests: participants inflected untrained items from
their singular to their plural form.
Neurobiology of Language
189
Consolidation in language and motor learning
The trained-item test required the judgment of correctly and incorrectly inflected plural
forms that were presented both before and after training in the first session, and in each of
the following two sessions (4 trained-item tests overall). Each of the 36 trained items was pre-
sented once in each test. During the test, the singular form was aurally presented (without its
picture) followed by an aurally presented plural form of the same word. Participants were
instructed to press “1” on a standard keyboard if the plural form was correct, and “2” other-
wise. They were given 3 sec to respond (Figure 2B). In each test, half of the presented 36 plural
forms were correct, and half were incorrect. Incorrect inflections were created by adding one
of the other suffixes to the stem. Across sessions, each participant was presented with all dif-
ferent incorrectly affixed forms in a random order.
Training took place in the first session (Figure 1B). During training, participants heard the
singular form accompanied by its picture and attempted to produce the plural form. The cor-
rect plural form of the word was then presented aurally, as feedback (Figure 2C). The training
session consisted of three blocks, separated by breaks. In each block half of the items, low-
frequency items, were presented once (18 trials), and half, high frequency items, were pre-
sented 3 times (54 trials), resulting in a total of 3 or 9 presentations per word, respectively
(Table 2). The order of items within each block was randomized.
In the untrained-items tests (generalization task), participants were asked to inflect 30
untrained items from their singular to their plural form (Figure 2D). Words were presented
in a randomized order. The untrained-items test was presented at the end of each session
(Figure 1B), for a total of three times.
At the end of the third linguistic session (in the 24-hr retest), participants answered a ques-
tionnaire assessing their explicit knowledge of the inflection regularity. Participants were asked
to write down which suffixes they remember, and to explicate what rule guided them in
inflecting singular stems to their plural form.
Motor sequence learning task
Participants were trained to perform accurately and repeatedly a 5-element sequence by tap-
ping a 4-key response pad with their left (non-dominant) hand (Figure 3A). This computerized
version of the sequential finger-tapping task, adapted from Karni et al. (Karni, 1995; Karni
et al., 1998), has been widely used to study mnemonic processes underlying learning of a
Figure 3. Motor sequence learning task setup and trials. (A) Illustration depicting the mapping of
digits to numbers. (B) Each session began with an explicit introduction of the sequence, followed by
14 training blocks (indicated by a green cross). The blocks comprise 60 key presses—the code and
the time of each pressed key are registered—and are separated by rest periods of 25 sec (red cross).
Neurobiology of Language
190
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
2
1
8
0
1
9
8
9
3
2
6
n
o
_
a
_
0
0
0
6
0
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Consolidation in language and motor learning
new motor skill (Albouy et al., 2015; Censor et al., 2010; Gabitov et al., 2019; Gal et al., 2019;
King et al., 2017; Walker et al., 2002; for a recent review, see Doyon et al., 2018).
The motor task was performed over three sessions. Each session was initiated with verifica-
tion of the participant’s familiarity with the keypad. After successful verification, participants
were asked to reproduce the target sequence accurately three times in a row to make sure that
they understood the task. Participants were then instructed to generate the target sequence
repeatedly “as fast and as accurately as possible.” In case of occasional errors, participants
were asked “not to correct errors and instead to continue by starting from the beginning of
the sequence.”
The sequence was introduced to participants using numbers from 1 to 4 (with numbers cor-
responding to digits as is shown in Figure 3A). During the first session, participants were
trained on either sequence A (41324) or sequence B (42314) using a block design (14 blocks).
The untrained sequence was later used to measure specificity of the acquired knowledge (see
below). The next two sessions were used to assess participants’ performance, hence, were
retested on the trained sequence 12 and 24 hours post-training by performing four blocks of
the trained sequence in each session (Figure 1C).
Each block (in training and retests) consisted of 60 keypresses (equivalent to 12 repetitions
of the 5-element sequence). These performance periods were separated by short periods of rest
(25 sec). During rest periods a red cross was presented in the middle of the screen, and during
performance blocks the cross was shown in green. The color of the cross automatically
changed from red to green and from green to red indicating the beginning and the end of each
block, respectively. During the task, participants were given only these visual cues and did not
receive any feedback (Figure 3B).
To test generalization of the skill to the untrained hand, participants took part in three addi-
tional 4-block tests at the end of the third session. All participants performed these tasks in the
same order: (a) the trained sequence (T-S) performed with the untrained hand (U-H), (b) the
untrained sequence (U-S ) performed with the trained hand (T-H), and (c) the untrained
sequence (U-S) performed with the untrained hand (U-H). The latter two tests (i.e., performing
the untrained sequence with both hands) were meant to serve as a reference to measure the
specificity of the acquired knowledge to the trained sequence. Nevertheless, due to a techni-
cal error, the untrained sequence was always performed in the trained hand before the
untrained hand and was thus already familiar when performed by the untrained hand. We
therefore do not include this (c) condition in the analysis.
Statistical Analyses
We analyzed the data using mixed repeated measures ANOVAs with consolidation interval as a
within-subject factor and group as a between-subject factor. We also used one-sample t tests to
estimate the magnitude of gains, and two-sample t tests to examine differences between groups.
The distribution of measures of interest across individuals was tested for skewness and kurtosis.
Since the protocol was very demanding, including seven meetings per participant, sched-
uled in fixed hours of the day, requiring participants to abstain from caffeine and maintain
rigorous sleep hygiene for several days, participant groups were relatively small, possibly
compromising the statistical power of the study. Therefore, to examine the strength of evi-
dence, additional Bayesian analyses were carried out in parallel to the frequentist analyses
(ANOVAs and t tests), to assess the odds for the research and null hypotheses given the
obtained data. Bayes Factors (BF ) were computed using JASP (version 0.8.6.0; cf. 2017; van
Doorn et al., 2020; Wagenmakers et al., 2018). Additionally, post hoc sensitivity analysis was
Neurobiology of Language
191
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
2
1
8
0
1
9
8
9
3
2
6
n
o
_
a
_
0
0
0
6
0
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Consolidation in language and motor learning
Token frequency:
The frequency with which a whole
word (token) appears in the
language.
computed in G*power 3.0.10 (Faul et al., 2007) in order to determine the minimum reliable
effect size at 80% power level given our sample size.
Language Learning Task
Only consistent words were included in the analysis of trained items. Reaction times (RTs) for
correct responses were calculated. Percentage of correct responses was calculated for each
individual in each token frequency (high and low) at each testing point.
As our main research questions focus on the consolidation period (i.e., from the end of
training to 12 and 24 hours post-training), our analyses were conducted on three measures,
calculated separately for accuracy and RT of trained items and for accuracy in untrained items:
(1) gains during the first 12 hours after training, (2) gains during the second 12 hours post-
training, and (3) gains across the entire 24 hours from the end of training. Individual gains in
each interval were calculated as a ratio from the performance level at the beginning of that
interval, e.g., the first 12-hr interval = [(difference in performance levels from the end of
training test to the 12-hr retest) / end of training test]. The change in performance was always
calculated so that higher (more positive) values indicate improvement. Hence for RT, it was
calculated as [performance at the end of training minus the 12-hr retest] while for accuracy
it was calculated as [performance at the 12-hr retest minus performance at the end of train-
ing]. Similarly, the second 12-hr interval = [(difference in performance levels from the 12-hr
to the 24-hr retest) / the 12-hr retest]. Finally, the total 24-hr interval was calculated as [(dif-
ference in performance levels from the end of the training test to the 24-hr retest) / the end of
training test]. Normalized gains in performance were used in all analyses to control for indi-
vidual differences in absolute performance levels and allow for a between groups compar-
ison, as was used in previous studies exploring the effect of consolidation on offline gains
(Abend et al., 2013; Doyon et al., 2009; Korman et al., 2007, 2021). For each measure we
first examined whether participants improved in performance during the total 24-hr consol-
idation period (this was done across groups when there was no difference between groups).
We then examined the effect of sleep on the first and second 12-hr intervals by comparing
between intervals and groups.
Participants’ explicit awareness of the morphophonological rule as expressed in the ques-
tionnaire was scored on a scale of 1 (no knowledge or uncalculated guess) to 6 (precise
explicit regularity knowledge). (See Table 3 for scoring criteria.) Participants’ awareness score
Table 3.
Scoring criteria of awareness to the morphophonological regularity
Score
1
Uncalculated guess
Level of awareness
2
3
4
5
6
Non-phonological strategy (e.g., semantic)
Basic phonological strategy leading to knowledge of one suffix ( VC) or
consonant with suffix (CVC)
Intermediate phonological strategy leading to two suffixes ( VC) or
consonant with suffixes (CVC)
Phonological strategy leading to three suffixes ( VC) and consonant with
suffixes (CVC)
Full phonological strategy leading only to three distinguished suffixes ( VC)
Neurobiology of Language
192
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
2
1
8
0
1
9
8
9
3
2
6
n
o
_
a
_
0
0
0
6
0
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Consolidation in language and motor learning
in each group was estimated against the score of “2,” which represents a non-phonological
strategy, using a one-sample t test. These scores were also compared between groups using a
two-sample t test. The correlation of explicit knowledge with raw performance and consoli-
dation (both 12-hr intervals and the total 24-hr interval) on the language task was tested using
Spearman correlations.
Motor Learning Task
Motor skill was evaluated using a measure reflecting the tapping speed, namely the time (dura-
tion) to complete each block (i.e., 60 key presses). The percentage of correctly performed and
completed sequences, out of 12 possible sequences per block, was also calculated (accuracy).
At each time point, performance was calculated as the mean across four blocks. The normal-
ized consolidation intervals and the statistical analysis of the trained sequence were calculated
as in the language learning task, as was the analysis relating to the change in performance
across the total consolidation interval and the effect of sleep on consolidation.
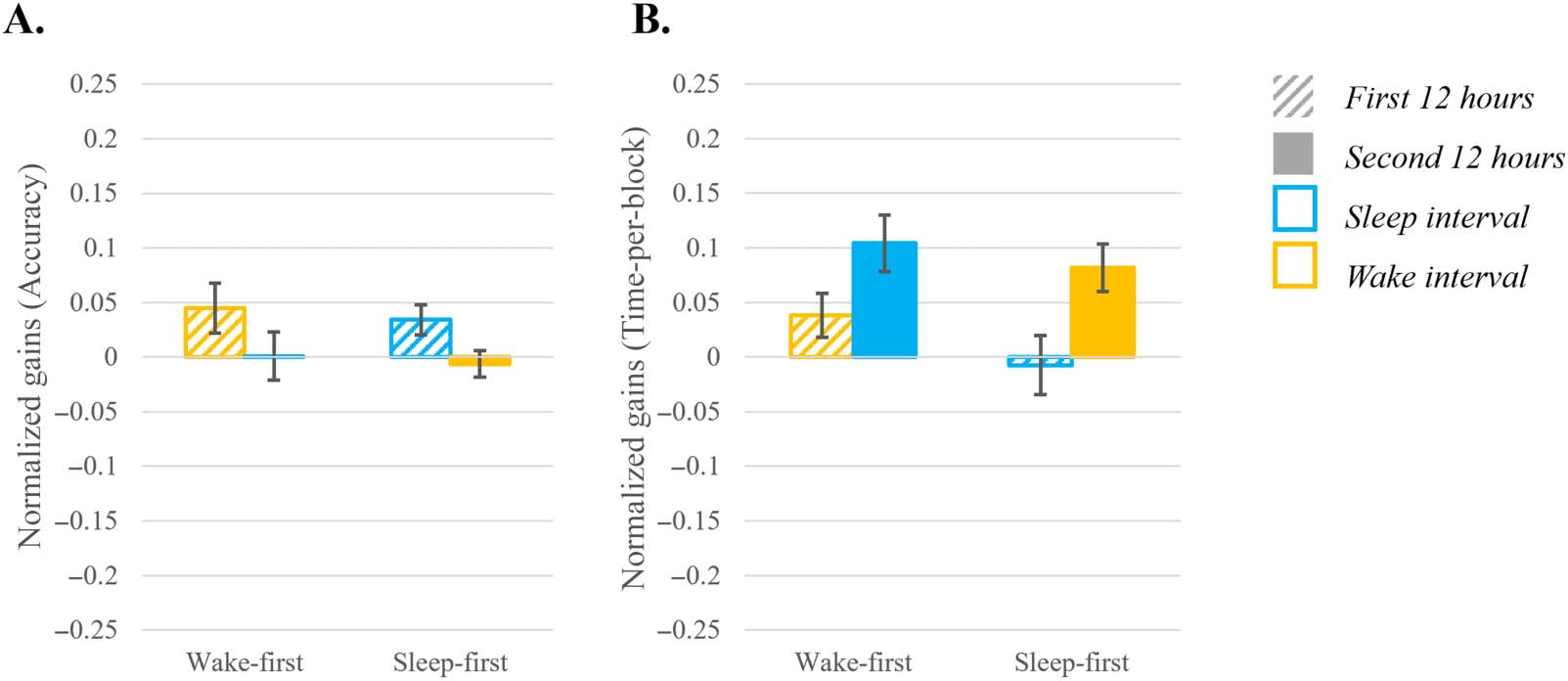
To test whether the motor skill was generalized to new conditions, we tested the specificity
of the skill to the trained sequence in comparison to a new sequence, and its transfer to the
untrained hand at the end of the third session (at the 24-hr retest). Sequence specificity in the
trained hand and transfer of the trained sequence to the untrained hand were computed for
each individual, based on the average time-per-block of the 4 blocks, measured at the 24-hr
time point, and normalized to the individual’s performance in order to account for general
individual differences in speed. Thus, Sequence Specificity = [(Untrained Sequence −
Trained Sequence) / Untrained Sequence]. Higher (positive) values reflect higher specificity
(i.e., better performance of the trained than the untrained sequence). Transfer to the
untrained hand = [(Trained Sequence in the untrained hand – Trained Sequence in the
trained hand at the 24-hr retest) / Trained Sequence in the untrained hand]. Smaller values
reflect better transfer (i.e., smaller differences between the trained and untrained hand). Both
measures were compared to zero within each group using a one-sample t test and compared
between groups using a two-sample t test.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
2
1
8
0
1
9
8
9
3
2
6
n
o
_
a
_
0
0
0
6
0
p
d
/
.
l
Correlation Analyses
To examine the extent to which performance on trained items was related to performance on
untrained items, in both linguistic and motor tasks, we conducted a correlation analysis within
each task using both raw performance and normalized consolidation intervals. Correlation
analyses were also conducted for individual consolidation gains between the linguistic and
motor tasks. The results were corrected for multiple comparisons using Bonferroni correction.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
RESULTS
Group Effect on Consolidation
Language learning task
Trained items. Figure 4 shows the raw performance on high and low frequency trained items
for descriptive purposes. Nevertheless, all statistical analyses were conducted on normalized
individual consolidation gains (as described in the Materials and Methods section), shown in
Figure 5.
Consolidation across 24 hours. Our first research question was whether participants
improved during the consolidation period from the end of training to the 24-hr retest, and
whether this was affected by group or item frequency. To this end we conducted a two-way
Neurobiology of Language
193
Consolidation in language and motor learning
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
2
1
8
0
1
9
8
9
3
2
6
n
o
_
a
_
0
0
0
6
0
p
d
.
/
l
Figure 4. Raw performance on trained items at four testing points, presented by frequency and group. (A) and (B) Accuracy rates and (C) and
(D) reaction times (RT) for the high frequency (upper panels) and low frequency (lower panels). This figure, showing raw performance, is
presented for display purposes only, as the statistical analyses were conducted on normalized consolidation intervals.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
repeated measures ANOVA on the total consolidation gain in accuracy with frequency (high
and low) as a within-subject factor and group (wake-first, sleep-first) as a between-subject
factor. There were no significant main effects or interactions and the BFs showed moderate
to strong support for H0 (0.087 ≤ BFincl ≤ 0.247). We thus collapsed the gains in the high
and low frequency items, and combined the two groups, and tested whether the 24-hr con-
solidation gain across item frequencies and groups was significantly different from zero in a
one-sample t test. This analysis revealed an overall small improvement across 24 hours [t(37) =
2.556; p = 0.015, d = 0.414; BF10 = 2.974 (anecdotal support for H1)]. These analyses were
also conducted for RTs, finding the same pattern of no difference between groups and frequen-
cies in RT gains across 24 hours, with strong to anecdotal support for H0 (0.096 ≤ BFincl ≤
0.363). The gains in RTs during the 24-hr consolidation period were not significantly different
from zero [t(37) = 114; p = 0.910, d = 0.018; BF10 = 0.176 (moderate support for H0)],
Neurobiology of Language
194
Consolidation in language and motor learning
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
2
1
8
0
1
9
8
9
3
2
6
n
o
_
a
_
0
0
0
6
0
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
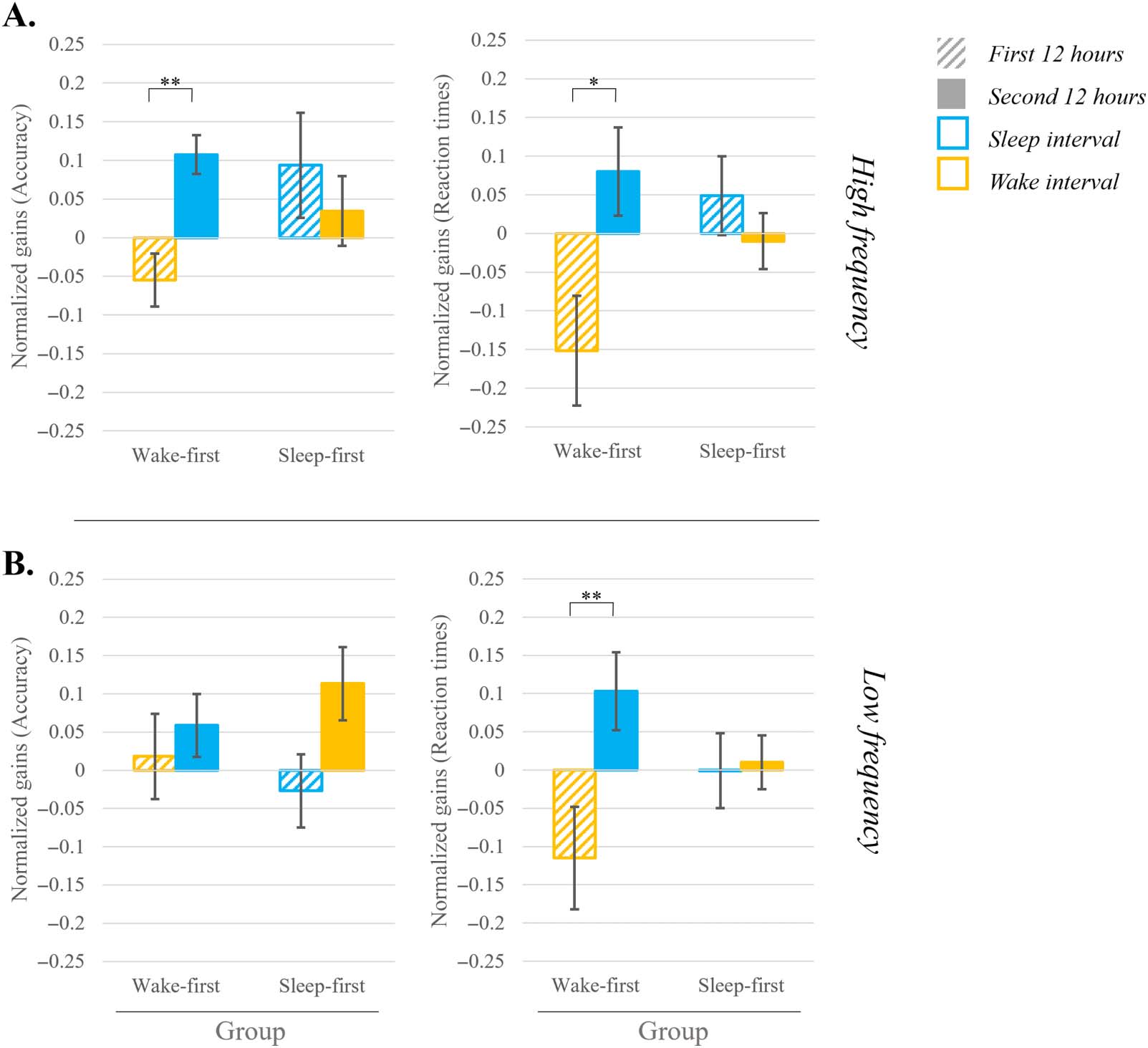
Figure 5. Gains in performance in the language task, presented by frequency and group (wake-first/sleep-first). (A) High frequency and
(B) Low frequency. 12-hr gains are presented for the two consolidation intervals (first 12 hours striped bar; second 12 hours full bar) with
the color indicating if an interval includes sleep (blue) or wake (yellow). Accuracy rate is presented on the left side and reaction times on
the right. Error bars indicate standard error. For both measures positive values indicate improvement. *indicates significance at p < 0.05;
**indicates significance at p < 0.001.
suggesting stable performance across 24 hours. Hence, across 24 hours post-training partici-
pants showed some improvement in accuracy levels and stable reaction times, both of which
did not differ across frequencies and groups.
The effect of sleep on consolidation. To probe for the effect of sleep on consolidation,
we performed a three-way repeated measures ANOVA on 12-hr consolidation gains, with
interval (first, second) and frequency (high and low) as within-subject factors and sleep group
(wake-first vs. sleep-first) as a between-subject factor. For reaction times, we found a significant
interaction between interval and sleep group [F(1, 36) = 9.056; p = 0.005; η2 = 0.201; BFincl =
42.735 (very strong support for H1)], indicating that sleep affects consolidation of trained items
across item frequencies. The three-way interaction between frequency, interval, and sleep
group was not significant [F(1, 36) = 0.335; p = 0.566; η2 = 0.009; BFincl = 0.059 (strong sup-
port for H0)]. For accuracy, this analysis revealed a marginally significant three-way interaction
between frequency, interval, and sleep group [F(1, 36) = 4.031; p = 0.052; η2 = 0.101; BFincl =
0.128 (moderate support for H0)]. Because we had distinct theoretical predictions for the
Neurobiology of Language
195
Consolidation in language and motor learning
high and low frequency items, which are expected to rely to some extent on different mech-
anisms, we wanted to examine whether the effect of sleep was due to one or both frequency
levels. Therefore, although the three-way interaction with frequency was only marginal for
accuracy, we conducted follow-up analyses separately for each frequency, for both accuracy
and RTs.
For high frequency trained items a two-way repeated measures ANOVA on consolidation
gains in accuracy comparing the first and second 12-hr intervals, and the two sleep groups,
revealed no main effects of either interval (BFincl = 0.670, anecdotal support for H0) or group
(BFincl = 0.546, anecdotal support for H0). However, the interaction between interval and
group was significant [F(1, 36) = 4.344; p = 0.044; η2 = 0.108; BFincl = 1.445 (anecdotal sup-
port for H1)]. To follow-up on this interaction, we conducted a paired-sample t test comparing
consolidation intervals within each group. Participants in the wake-first group showed a sig-
nificant difference between intervals [t(19) = −3.364; p = 0.003; d = 0.752; BF10 = 13.191
(strong support for H1)]. Figure 5A (left panel) shows that this was due to deterioration during
wake and improvement during sleep. In contrast, in the sleep-first group there was no differ-
ence between intervals [t(17) = 0.602; p = 0.555; d = 0.141; BF10 = 0.286 (moderate support
for H0)]. Figure 5A shows that when sleep occurred in the first interval, performance was stable
during the subsequent wake interval.
The two-way repeated measures ANOVA performed on consolidation gains in reaction
times (first and second 12-hr intervals) showed no significant main effects of either interval
(BFincl = 1.404, anecdotal support for H1) or group (BFincl = 0.901, anecdotal support for
H0), but again revealed a significant interaction between interval and group [F(1, 36) =
5.198; p = 0.029; η2 = 0.126; BFincl = 2.958 (anecdotal support for H1)]. A follow-up analysis
on this interaction, conducted separately for each group using a paired-sample t test, revealed,
once again, a significant difference between intervals for the wake-first group only [t(19) =
−2.307; p = 0.032; d = −.516; BF10 = 1.956 (anecdotal support for H1)]. Here as well, obser-
vation of Figure 5A (right panel) shows that this was driven by deterioration in performance
during wake and an improvement during sleep. No significant difference between the intervals
was found for the sleep-first group [t(17) = 0.788; p = 0.442; d = 0.185; BF10 = 0.320
(moderate support for H0)]. Figure 5A shows again that when sleep occurred during the first
interval, performance was stable during the subsequent wake interval.
For low frequency trained items a two-way repeated measures ANOVA performed on
consolidation gains in accuracy (first 12 hours and second 12 hours) revealed no significant
main effects of interval or group, nor an interaction (and the BF showed moderate to anec-
dotal support for H0 (0.191 ≤ BFincl ≤ 0.726). However, for reaction times the two-way
repeated measures ANOVA performed on consolidation gains (first 12 hours and second
12 hours) revealed a significant main effect of interval [F(1, 36) = 6.731; p = 0.014; η2 =
0.158; BFincl = 5.535 (moderate support for H1)] with no main effect of group (BFincl =
0.723, anecdotal support for H0). This analysis also showed a significant interaction between
interval and group [F(1, 36) = 5.488; p = 0.025; η2 = 0.132; BFincl = 2.167 (anecdotal sup-
port for H1)]. Follow-up analyses, using paired-sample t tests to compare between the con-
solidation intervals within each group, revealed a significant difference between intervals for
the wake-first group [t(19) = −3.541; p = 0.002; d = 0.791; BF10 = 18.630 (strong support for
H1)]. Figure 5B (right panel) shows that this is driven by deterioration during wake and
improvement during sleep. No significant difference between the intervals was found for
the sleep-first group [t(17) = −0.176; p = 0.862; d = 0.041; BF10 = 0.246 (moderate support
for H0)]. Figure 5B shows that when sleep occurred in the first interval performance was
stable across the first and second intervals.
Neurobiology of Language
196
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
2
1
8
0
1
9
8
9
3
2
6
n
o
_
a
_
0
0
0
6
0
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Consolidation in language and motor learning
(A) Raw performance accuracy of untrained items, by group (wake-first/sleep-first) presented at three testing time points. The dashed
Figure 6.
line at 33.3% represents chance level. (B) Gains in performance for untrained items, presented by group: 12-hr gains are presented for the two
consolidation intervals (first 12 hours striped bar; second 12 hours full bar) with the color indicating if an interval includes sleep (blue) or wake
(yellow). Error bars indicate standard errors.
Untrained items. Figure 6A shows the raw performance on untrained items for the three testing
time points. Preliminary analyses on raw performance using one-sample t tests showed that
participants performed above chance level (33.3%) at all testing time points (5.93 ≤ t ≤
7.803, all p < 0.001, corrected for 3 time points; see Figure 6A). All subsequent statistical
analyses were conducted on normalized individual gains (shown in Figure 6B).
An independent-sample t test on the 24-hr interval showed no significant difference
between groups [t(36) = 0.789; p = 0.436; d = 0.26; BF10 = 0.391 (anecdotal support for
H0)]. Therefore, to examine the pattern of change during the 24-hr consolidation period, we
collapsed across both groups and tested whether the 24-hr gain was different from zero using a
one-sample t test. We found a small and significant improvement in performance across
groups during the 24-hr consolidation period [t(37) = 2.445; p = 0.019; d = 0.396; BF10 =
2.407 (anecdotal support for H1)]. To address the question of the effect of sleep on consolida-
tion, a two-way repeated measures ANOVA on the 12-hr intervals (first and second 12 hours)
revealed no significant main effects of interval or group and no interaction, (all BF showed
moderate support for H0 (0.106 ≤ BFincl ≤ 0.220). See Figure 6B.
Motor task
Figure 7 presents the raw performance in the motor task for descriptive purposes. All statistical
analyses were conducted on normalized individual intervals (see Figure 8).
Trained sequence.
Accuracy. Participants’ performance was highly accurate with 92.86% ± 4.95% (mean ±
SEM ) accuracy on average throughout the experiment, consistent with previous studies on
Neurobiology of Language
197
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
2
1
8
0
1
9
8
9
3
2
6
n
o
_
a
_
0
0
0
6
0
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Consolidation in language and motor learning
Figure 7. Performance on the trained motor sequence presented by group, both for (A) Accuracy and (B) Time per block. Each time point
represents the mean values of four blocks of 60 key-presses each. This figure of raw performance is presented for display purposes only, as the
statistical analyses were conducted on normalized consolidation intervals.
motor sequence learning (Gabitov et al., 2014, 2017; Korman et al., 2003; Walker et al., 2002).
An independent-sample t test on the 24-hr interval showed no significant difference between
groups [t(34) = 0.623; p = 0.537; d = 0.21; BF10 = 0.375 (anecdotal support for H0)]. We therefore
collapsed across groups and used a one-sample t test to test whether the overall change in per-
formance over 24 hours was different from zero. There was a significant improvement during the
24-hr consolidation period [t(35) = 2.746; p = 0.009; d = 0.457; BF10 = 4.432 (moderate support
for H1)]. To test the effect of sleep on consolidation a two-way repeated measures ANOVA
Figure 8. Gains in performance in the motor task, in Accuracy (A) and Time-per-block (B), presented for the two consolidation intervals (first
12 hours striped bar; second 12 hours full bar) with the color indicating if an interval includes sleep (blue) or wake (yellow). Error bars indicate
standard error. In both Accuracy and Time-per-block positive values indicate improvement.
Neurobiology of Language
198
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
2
1
8
0
1
9
8
9
3
2
6
n
o
_
a
_
0
0
0
6
0
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Consolidation in language and motor learning
performed on the 12-hr intervals (first and second 12 hours) revealed a marginal main effect of
interval [F(1, 34) = 3.996; p = 0.054; η2 = 0.105; BFincl = 9.248 (moderate support for H1)]
driven by greater improvement during the first 12 hours and stabilization during the second
12 hours, across groups. There was no main effect for group (BFincl = 0.071; strong support
for H0) nor an interaction between interval and group (BFincl = 0.083; strong support for H0)
(see Figure 8A).
Time per block. As performance accuracy is typically very high in this task (Gabitov
et al., 2014, 2017; Korman et al., 2003; Walker et al., 2002), the main learning outcome mea-
sure is speed measured as time per block. An independent-sample t test on the 24-hr interval
showed no significant difference between groups [t(34) = 0.585; p = 0.563; d = 0.20; BF10 =
0.368 (anecdotal support for H0)]. Hence, we collapsed across groups to examine the
improvement across 24 hours. A significant and very strong improvement in performance
was found across 24 hours [t(35) = 4.917; p < 0.001; d = 0.891; BF10 = 1053.055 (extreme
support for H1)].
In order to test the effect of sleep on consolidation, a two-way repeated measures ANOVA
on the 12-hr intervals was conducted, with consolidation intervals as a within-subject factor
and group as a between-subject factor. The analysis revealed a significant main effect for con-
solidation interval [F(1, 34) = 4.945; p = 0.033; η2 = 0.127; BFincl = 3.222 (moderate support
for H1)] indicating stable performance during the first interval and subsequent improvement
during the second interval across groups. No significant effect was found for group (BFincl =
0.378, anecdotal support for H0) nor an interaction between interval and group (BFincl = 0.761,
anecdotal support for H0), indicating that offline gains appeared only 24 hours post-training,
independent of the timing of sleep (see Figure 8B).
Generalization. The specificity of participants’ skill to the trained sequence, as mea-
sured by the normalized sequence specificity index 24 hours post-training, was not different
between the groups as was indicated by an independent-sample t test [t(34) = 0.198; p =
0.884; d = 0.06; BF10 = 0.327 (moderate support for H0)]. This indicates that the proximity
of the training session to sleep did not affect sequence specificity 24 hours post-training.
We therefore collapsed across groups, and found that sequence specificity was significantly
above zero [t(35) = 6.341, p < 0.001; d = 1.05; BF10 = 57587.32 (extreme support for H1)],
indicating that the trained sequence was performed better than the untrained sequence.
Participants’ generalization of gains to the untrained hand, as measured by the normalized
transfer to untrained hand, were not different between groups [t(34) = 1.630; p = 0.122; d =
0.55; BF10 = 0.893 (anecdotal support for H0)]. We therefore collapsed across groups and
using a one-sample t test found that this measure was not significantly different from zero
[t(35) = 1.585; p = 0.122; d = 0.26; BF10 = 0.559 (anecdotal support for H0)], indicating no
difference between the trained and untrained hands.
Individual Differences in Consolidation
Correlations within the language domain
To examine the extent to which performance on trained items (which can rely on item-specific
learning and on extraction of regularities) depends on extraction of regularities (measured by
performance on untrained items), we calculated Pearson correlations between raw accuracy
levels on the trained and untrained items at each testing time point across groups, separately
for low and high frequency trained items. These analyses revealed strong and significant pos-
itive correlations between tasks for all 3 testing time points (0.486 ≤ r ≤ 0.713, all p ≤ 0.002,
Neurobiology of Language
199
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
2
1
8
0
1
9
8
9
3
2
6
n
o
_
a
_
0
0
0
6
0
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Consolidation in language and motor learning
corrected for 6 comparisons). Hence, better ability to recall high and low frequency trained
items was positively associated with the ability to extract the phonological regularities and
implement them to untrained items immediately post-training, and throughout the other tests.
However, when examining the extent to which the consolidation of extraction of regularities is
related to the consolidation of trained items, by testing for correlations between normalized
consolidation intervals of the trained and untrained items (both 12-hr intervals and the total
24-hr interval), no significant correlations were found.
To test whether participants developed explicit awareness of the morphophonological regu-
larities by the end of the experiment, and whether this was associated with their consolidation in
the language task, we used the results of the questionnaire administered at the end of the third
session. Participants from both groups developed a basic level of awareness of the morphopho-
nological regularities, as indicated by mean awareness of 2.88 (± 1.56) for the wake-first group
and 3.611 (± 1.61) for the sleep-first group. The groups did not differ from each other in the levels
of awareness [t(34) = −1.362; p = 0.182; d = 0.46; BF10 = 0.659 (anecdotal support for H0)], so
we collapsed across groups and using a one-sample t test found that this level of awareness was
significantly above 2 (representing a non-phonological strategy; [t(35) = 4.658; p < 0.001; d =
0.776; BF10 = 516.85 (extreme support for H1)]. Explicit knowledge was significantly correlated
(Spearman rank order correlation) with raw performance accuracy on trained and untrained
items at all 3 testing points (0.441 < r < 0.658, all p ≤ 0.01, corrected for 3 comparisons).
However, explicit regularity knowledge was not associated with gains in either of the 12-hr
intervals or the total 24-hr interval, for either trained or untrained items across groups.
Correlations within the motor domain
To examine the association between consolidation in the motor task, and specificity of the
acquired skill to the trained sequence, we examined the correlation between participants’ con-
solidation in speed across 24 hours and sequence specificity. Because there were no differ-
ences between groups in any of these measures, this was done across groups. The analysis
revealed a marginal positive correlation between offline gains and sequence specificity in
the trained hand (r(37) = 0.324, p = 0.054; d = 0.10), indicating that participants who
improved more during the 24-hr interval tended to show more specificity to the trained
sequence 24 hours post-training.
In contrast, we found no association between consolidation in the motor task across
24 hours and transfer of the acquired sequence-specific skill to the untrained hand ( p =
0.074, following the exclusion of two outliers).
Correlations between the language and motor domains
To test whether there are individual differences in consolidation that are shared across the
motor and language domains, we examined the correlations between the 24-hr interval in
the two domains in terms of speed, which is the more relevant measure for the motor task.
We found no significant correlation between the overall normalized gains across 24 hours
in the motor and the high frequency trained items in the linguistic task (r(36) = 0.016, p =
0.924; d = 0.00). However, the 24-hr consolidation gains of the motor task were significantly
correlated with the 24-hr gains in the low frequency trained items in the linguistic task, though
the effect size was relatively small (r(36) = 0.329, p = 0.049; d = 0.10). Nevertheless, no sig-
nificant difference was found between the high and low frequency items in terms of their cor-
relation with the motor consolidation (z = −1.323; p = 0.093). Thus, participants who
improved more across 24 hours in the motor domain also showed greater improvement in
the linguistic domain, but only for the low frequency trained items.
Neurobiology of Language
200
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
2
1
8
0
1
9
8
9
3
2
6
n
o
_
a
_
0
0
0
6
0
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Consolidation in language and motor learning
Sensitivity Analysis
Sensitivity analysis, computed in G*power 3.0.10 (Faul et al., 2007) with alpha = 0.05,
revealed that our current sample size had 80% power to detect an effect size of d ≥ 0.466,
for one-sample t tests, which were conducted in the current study in order to test for an overall
improvement across 24 hours. Our findings of significant improvement over 24 hours in
trained and untrained items in the language task are slightly below this effect size, which
together with the BF suggests that these findings could be anecdotal. In contrast, the 24 hours
improvement in the motor task showed a larger effect size, suggesting it is reliable. In addition,
sensitivity analysis with the above parameters was also computed for a two-way repeated mea-
sures ANOVA with within-between interactions, conducted in the current study to test for the
effect of sleep on consolidation. It revealed that our sample size had enough power to detect
an effect size of f ≥ 0.234 (i.e., η2 ≥ 0.051). All significant interactions in the current study
showed larger effect sizes, adding confidence to their reliability. Finally, sensitivity analysis
with the above parameters was computed for paired-sample t tests, conducted in the current
study for testing the differences between the first and second consolidation intervals within
group. This showed there is enough power to detect an effect size of d ≥ 0.660. Two of the
three significant findings showed larger effect sizes (high frequency accuracy, and low fre-
quency RT) suggesting these findings have higher reliability.
DISCUSSION
The current study examined patterns of improvement and stabilization, sleep dependence, and
domain generality of memory consolidation, by comparing consolidation in a language learn-
ing task and a motor sequence learning task and the correlations between the two. The struc-
ture of the language learning task allowed us to probe two varieties of learning: memorization
of exemplars, namely, item-specific learning, and the extraction of patterns embedded in the
input into grammatical regularities. This distinction between components in the language task
allowed us to specifically address the effects of consolidation of these components. In addi-
tion, consolidation patterns of the language task were compared to consolidation of motor
sequence learning to probe for common mechanisms, both at the group level, and in terms
of individual differences. Below we discuss the results in view of the current literature and in
projection to wider implications.
Consolidation in Language Learning
In the current study, we charted the consolidation and the role of sleep in learning morpho-
logical inflections over the course of 24 hours in typical adults divided into two groups, wake-
first and sleep-first. In discussing the results, we address findings relating to trained items and
untrained items separately, as each reflects, to some extent, distinct linguistic components.
Consolidation and the role of sleep in item-specific learning
Participants’ accuracy in inflecting all trained items showed a small improvement 24 hours
after training, indicating continued offline processing of the acquired information after training.
Importantly, consolidation of trained items was affected by sleep. This was shown in reaction
time by the strong evidence for an interaction between interval and sleep group in both high
and low frequency items. For reaction time of all trained items, while the wake-first group
showed deterioration during wake and recovery in the subsequent sleep interval, the sleep-
first group showed stable performance across the two intervals. Interestingly, for accuracy, the
effect of sleep on consolidation was only evident in high frequency items (although it should
Neurobiology of Language
201
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
2
1
8
0
1
9
8
9
3
2
6
n
o
_
a
_
0
0
0
6
0
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Consolidation in language and motor learning
be noted that the interaction with frequency was only marginal). For these high frequency
items, the effect of sleep was similar to reaction times: The wake-first group deteriorated during
wake and recovered during the subsequent sleep, while the sleep-first group showed no such
deterioration during wake. Altogether these results show a clear effect of sleep on consolida-
tion of trained items, but while high frequency items showed an effect of sleep in both accu-
racy and reaction time, low-frequency items show this effect only in reaction time. Because
high and low frequency items share the same regularities, the differences between them in the
effect of sleep on accuracy can most likely be attributed to item-specific learning, which
played a more prominent role in high frequency items. In contrast, for low frequency trained
items, which should rely more on the extraction of regularities, the effect of sleep on consol-
idation was only partial.
The stabilizing effect of sleep on consolidation of trained items was expressed in several
ways. (1) In the sleep-first group, during the first interval, sleep prevented degradation of mem-
ory that was evident in the comparable interval for the wake-first group, most likely due to
interference. (2) During the second interval, the sleep-first group maintained stable perfor-
mance of the learned linguistic information despite being awake. Thus, sleep immediately after
training protected against future wake interference. (3) When the wake-first group slept, during
their second interval, they managed to recover their performance, which had deteriorated in
the previous wake interval. Thus, sleep facilitated recovery after memory degradation.
While the first of these findings can also be explained by a model in which sleep provides
only a passive protection from interference (Jenkins & Dallenbach, 1924; Mednick et al.,
2011), the latter two findings (sleep protecting against later degradation and sleep facilitating
recovery) argue against such a model, and suggest that sleep plays an active role in consoli-
dation and is associated with qualitative changes to memory representations due to specific
sleep-related neural processes. This hypothesis is consistent with other studies suggesting that
during stabilization, memory traces are actively strengthened during sleep (Diekelmann &
Born, 2010), and thus accessibility to the memories improves (Dumay, 2016). Moreover, this
active information processing that occurs during sleep creates more durable memory traces,
which are then more resistant to interference introduced after the sleep period, resulting in
stabilization of performance (Brawn et al., 2010; Fenn et al., 2003; Sonni & Spencer, 2015).
This process was also specifically suggested for consolidation of episodic memories
(Ellenbogen, Payne, & Stickgold, 2006) during which spontaneous reactivation occurring in
the hippocampus during sleep improves memory maintenance (Schreiner & Rasch, 2018).
Our findings, showing a stabilizing effect of sleep on trained items, and more prominent
effects on trained items with better opportunities for item-specific knowledge (high frequency
items), are in line with previous evidence for a stabilization effect of sleep on episodic learning
in general (Ellenbogen, Hulbert et al., 2006; Schönauer et al., 2015) and specifically on vocab-
ulary learning (Dumay, 2016; Gais et al., 2006). Given the important role of hippocampal-
dependent learning in vocabulary learning (Davis & Gaskell, 2009), this suggests that the
effects of sleep on learning of trained items in the current study also reflect hippocampal-based
item-specific learning.
Consolidation and the extraction of linguistic regularities
In the current study, participants were also tested on their ability to generalize their knowledge
of morphophonological regularities embedded in the trained items to the inflection of
untrained items. Both groups presented above-chance performance on untrained items at
the end of training and a small significant overall enhancement in performance across 24 hours,
Neurobiology of Language
202
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
2
1
8
0
1
9
8
9
3
2
6
n
o
_
a
_
0
0
0
6
0
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Consolidation in language and motor learning
regardless of the timing of sleep. The benefit of the passage of time for regularity extraction,
expressed as enhancement in performance, with no evidence that the timing of sleep influ-
ences that process, is consistent with a previous study reporting that a short nap did not
enhance the extraction of grammatical regularities (Mirković & Gaskell, 2016). The current
study shows no benefit of sleep even after a full night’s sleep. These findings are in contrast
to some studies reporting a beneficial effect of sleep on regularity extraction through the asso-
ciation between sleep and grammar learning (Batterink et al., 2014; Kim & Fenn, 2020). Nev-
ertheless, Kim and Fenn (2020) suggest that sleep benefits generalization only for learners who
are aware of the regularities before sleep. In the current study, we did not measure partici-
pants’ explicit awareness of the regularities before sleep, hence variability among participants
may explain this discrepancy.
The improvement in generalization during the 24-hr consolidation period is consistent with
the notion that consolidation is associated with the abstraction of gist information from newly
encoded memories to form cognitive schemata (Lewis & Durrant, 2011). During this process,
shared information is extracted from separate exemplars resulting in the ability to generalize
these commonalities to novel experiences. In the current study, the process of regularity
extraction began during training, resulting in above-chance performance in the generalization
test immediately after training. Interestingly, grammar knowledge continued to evolve over the
next 24 hours, suggesting a qualitative change in the representations of knowledge during off-
line consolidation. While this grammar knowledge was not explicitly instructed, it had
become conscious to some degree by 24 hours post-training, and this awareness was associ-
ated with performance on untrained items.
The association between item-specific knowledge and regularity extraction is evident in
the strong positive correlations found between performance on high and low trained items on
the one hand and untrained items on the other hand, throughout all testing time points.
These correlations may suggest that even during training, performance on trained items
was based, to some degree, on grammar knowledge. Alternatively, they may suggest that
memorization of specific exemplars contributed to the evolution of grammar knowledge,
consistent with evidence from computational models suggesting that improved memorization
of exemplars can lead to better generalization of information (Khandelwal et al., 2019;
Morcos et al., 2018).
Nevertheless, despite the above-mentioned correlations between raw performance on
trained and untrained items, there were no correlations between the consolidation gains on
trained and untrained items. These findings, which replicate previous studies (Ben Zion et al.,
2019), may suggest that while grammar knowledge evolved from memorization of specific
exemplars, the consolidation of the two types of knowledge may be mediated by distinct neu-
rocognitive mechanisms. This finding is consistent with the idea that learning of trained items
and particularly high frequency items relied mainly on item-specific memorization, which is
associated with the episodic, hippocampal system and is stabilized by sleep. In contrast, the
extraction of regularities may depend on frontostriatal skill learning, suggested to be involved
in grammar learning (Hedenius et al., 2011; Ullman, 2015; but see also Kumaran &
McClelland, 2012, for an alternative model suggesting that both exemplar memorization
and generalization may be supported by the hippocampal system). Consistent with the current
results, this skill-learning system may underlie offline improvement during the consolidation
interval with no necessity for sleep (Kemény & Lukács, 2016; Meier & Cock, 2014; Song,
2009). Further support for this notion comes from a previous neuroimaging study that used
a similar language paradigm (Nevat et al., 2017) and showed activation in the frontostriatal
network during inflection of trained items.
Neurobiology of Language
203
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
2
1
8
0
1
9
8
9
3
2
6
n
o
_
a
_
0
0
0
6
0
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Consolidation in language and motor learning
Consolidation in Motor Skill Learning
In the current study we also tested the role of sleep in motor sequence learning, in the same
participants and with the same schedule as the language task. Thus, the sequence was trained
either in the morning (wake-first group) or evening (sleep-first group) and was tested 12 and
24 hours post-training in both groups. Sequence specificity in the trained hand and transfer of
the skill to the untrained hand were also tested at the 24-hr time point.
Considering speed as our main measure of interest, the results indicate that in both groups
performance of the trained sequence was maintained during the first 12 hours post-training,
and then improved during the next 12 hours. Thus, significant enhancement in performance
was observed only 24 hours post-training regardless of the timing of sleep. These results sug-
gest that sleep was not sufficient for consolidation to occur in the first 12 hours. Nevertheless,
the total 24-hr interval included both the passage of time and sleep, for both groups. Hence,
while sleep may have contributed to the evolving consolidation gains over the 24-hr interval,
our design cannot test whether it was indeed necessary. The lack of overnight gains for the
sleep-first group was unexpected, as many studies did show significant post sleep enhance-
ment, in comparison to wakefulness, even following a short nap (Brawn et al., 2010; Gregory
et al., 2014; Korman et al., 2007; Walker et al., 2003; Wilhelm et al., 2008). Nevertheless, a
recent meta-analysis of 48 motor learning studies, out of which 23 studies used the finger-
tapping paradigm, found that many of these studies did not show a significant effect of sleep
on consolidation, and concluded that the effect of sleep on motor sequence consolidation is
rather small (Hedges g = 0.47; Schmidt et al., 2020). This conclusion is in line with the results
of another meta-analysis that suggests that sleep by itself is not sufficient to induce enhanced
performance following motor learning (Pan & Rickard, 2015).
Regardless of the role of sleep, the enhanced performance after 24 hours in the motor task is
consistent with previous motor skill learning studies showing offline improvement after the end
of training (Robertson, Pascual-Leone, & Miall, 2004). Previous studies suggest that this
improvement reflects a qualitative change and reorganization of motor representations during
consolidation (Gabitov et al., 2014; Korman et al., 2007; Rasch et al., 2007; Tucker et al.,
2006; Walker et al., 2003; for reviews, see Doyon et al., 2018; Dudai, 2004; Dudai et al.,
2015). Our current findings, of a correlation between improvement in performance on the
trained sequence during the 24-hr interval and sequence specificity, support this notion.
Sequence specificity reflects the formation of a unified representation of the sequence as a
whole from the separate representations of individual movements (Dorfberger et al., 2012).
At the neural level, this transformation is supported by reorganization within the corticostriatal
loop (Doyon et al., 2018), including a shift in the connectivity between the putamen and the
motor cortex (Gabitov et al., 2015). Thus, our findings support the conclusion that the forma-
tion of this unified representation is associated with improved performance on the trained
sequence.
Finally, our results showed similar performance levels of the trained sequence in the trained
and untrained hands at the 24-hr retest. Although the untrained hand was also participants’
dominant hand, previous studies that compared the dominant and non-dominant hands before
training showed no differences between the hands in initial performance levels or learning
rates in the finger-tapping task (Wiestler et al., 2014) and other motor learning tasks (Panzer
et al., 2009; Schambra et al., 2011). Hence, the similarity in performance between the hands
at the 24-hr retest suggests that the representation of the trained skill was transferred to the
untrained hand. Due to technical problems, we could not properly assess sequence specificity
in the untrained hand. Nevertheless, these results are in line with previous studies showing
Neurobiology of Language
204
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
2
1
8
0
1
9
8
9
3
2
6
n
o
_
a
_
0
0
0
6
0
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Consolidation in language and motor learning
generalization from the trained to the untrained hand (Dorfberger et al., 2012; Korman et al.,
2003; Rozanov et al., 2010; Witt et al., 2010) indicating that the neural representation of the
skill includes cortical areas controlling the opposite effector (Dorfberger et al., 2012; Gabitov
et al., 2016).
Shared and Distinct Consolidation Mechanisms Across Domains
The design of the current study, including the motor and language learning tasks in the same
population, allowed us to examine the effect of sleep on consolidation across domains, and
probe the existence of shared mechanisms both at the group and at the individual level.
At the group level, sleep played a stabilizing role in the consolidation of item-specific
knowledge, but the consolidation of grammar knowledge (evident by generalization to
untrained items) showed similarities to the consolidation of motor sequences. In both motor
and grammar learning, performance improved 24 hours after training, and sleep was not suf-
ficient to induce enhancement 12 hours post-training. These findings are in line with the
notion that offline enhancement in skill learning is independent of the timing of sleep (Kemény
& Lukács, 2016; Meier & Cock, 2014; Song, 2009). Additionally, in both domains, consolida-
tion was associated with a qualitative change to the representation. In the language task, this
was evident directly in the increase in generalization during the consolidation interval, indi-
cating that the knowledge acquired about trained items became more abstract and generaliz-
able. In the motor task, this was evident in the correlation between sequence specificity and
consolidation of the trained sequence after 24 hours. In both cases, consolidation was asso-
ciated with a process by which a sequence of separate events (cue and suffix, or separate digit
movements) was transformed into a unified representation.
At the individual level, we examined the correlations between consolidation in the two
tasks to identify shared neurocognitive learning mechanisms. This analysis showed a positive
correlation between participants’ improvement over 24 hours in the motor task and their 24-hr
consolidation in the low frequency trained items in the linguistic task, indicating that some
aspects of consolidation are domain general, and may characterize individuals regardless of
the specific skill being learned. Another interesting aspect of this correlation is the finding that
it was evident only for low (and not high) frequency items in the linguistic task (though the
difference between the correlations was not significant). Given that learning of low-frequency
trained items relies on regularity extraction, more than the high-frequency items, this correla-
tion may be due to the skill learning component in both motor and grammar learning. One
caveat to this conclusion is that we did not find a correlation between consolidation in the
motor task and consolidation of untrained items in the linguistic task, which also relies on
grammar learning.
Taken together, our results at the group and individual levels suggest that some aspects of
consolidation are shared across the motor and language domains, and more specifically
between motor sequence learning and grammar. This conclusion is in line with the idea that
motor sequence and grammar learning recruit overlapping neural substrates (Cona &
Semenza, 2017; Ferstl et al., 2008; Hertrich et al., 2016; Nakai et al., 1999) and rely on
procedural/skill-learning mechanisms (Ullman & Pierpont, 2005). This suggestion earns
additional support from previous findings demonstrating that consolidation in the motor and
language domains is sensitive to individual differences between learners in a wide range of
abilities (Adi-Japha et al., 2011; Fenn & Hambrick, 2012, 2015; Needle et al., 2015). They
are also in line with the suggestion of shared consolidation mechanisms between other dis-
tinctive domains such as perceptual and motor learning (Censor et al., 2012). However,
Neurobiology of Language
205
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
2
1
8
0
1
9
8
9
3
2
6
n
o
_
a
_
0
0
0
6
0
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Consolidation in language and motor learning
several previous studies have failed to find evidence for shared mechanisms in consolidation
in the language and motor domains (Henderson & Warmington, 2017; Wilson et al., 2012).
We suggest that this divergence across studies might be a consequence of the specific tasks
selected for each domain in previous research (Wilson et al., 2012), or of the consolidation
intervals selected (Henderson & Warmington, 2017). This fascinating issue undoubtedly
deserves continued systematic research.
Limitations
The current study should be viewed considering several limitations. First, due to the very
demanding protocol, the number of participants in the study was relatively small (N = 38).
Thus, to distinguish between underpowered and more reliable findings, we also included
the results of the Bayes factor analyses and post hoc sensitivity analysis (at 80% power). Future
research should aim to replicate the results with larger sample sizes, possibly using power
analysis before data collection (as elaborated in Brysbaert, 2019). Second, the design of the
motor task in the current study did not allow us to identify the association between consoli-
dation and generalization of sequence specific knowledge to the untrained hand. This is
because the untrained sequence was introduced in a fixed order, first to the trained and then
to the untrained hand. This resulted in familiarity with this sequence when performed by the
untrained hand, so it could no longer serve as a baseline for sequence specificity. Future stud-
ies can use a unique untrained sequence for each hand, which would allow proper testing of
the ability to apply a newly learned order of movements to a new condition. Third, our study
was designed to test the effect of sleep in intervals of 12 hours. The finding that motor
sequence learning and linguistic regularity extraction were enhanced across 24 hours, which
includes sleep in both groups, does not enable us to examine the contribution of sleep to con-
solidation in a 24-hour interval. Therefore, it is possible that sleep alone or time alone are not
sufficient for offline gains, and both may be needed for consolidation. This question would
require a sleep deprivation manipulation. Fourth, the smaller number of low frequency items
compared to high frequency items included in the analysis raises a question of whether differ-
ences in the statistical power between high and low frequency items might explain the differ-
ent effect of sleep on consolidation in the accuracy measures in these conditions. Fifth, we
chose to test participants’ explicit knowledge of the regularities only at the end of the exper-
iment, in order to minimize the potential effect of the questionnaire on the performance in the
tests. As a result, we do not know the degree of explicit awareness of the regularities at the end
of training, which may have affected its consolidation, as suggested by Kim and Fenn (2020).
Finally, as in most studies measuring consolidation, it should be noted that offline improve-
ment can be a result of the effect of testing, rather than a truly offline process (Eriksson et al.,
2011; Schoch et al., 2017). Studies aiming to ameliorate this problem by including separate
groups tested in different intervals show the benefit of active consolidation in addition to the
facilitative effect of testing itself (Kroneisen & Kuepper-Tetzel, 2021).
Conclusions
By examining the role of sleep in consolidation during grammar learning in a novel language,
and comparing it to motor sequence learning, the current study was able to dissociate two
patterns of consolidation in the language learning task, and find commonalities between con-
solidation in the language and motor domains. One component we identified is item-specific
learning, which is mostly evident here in high-frequency trained items. This item-specific
learning, which is similar to new vocabulary learning and relies on hippocampal mechanisms,
shows active dependency on sleep that promotes its memory stabilization. The second
Neurobiology of Language
206
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
2
1
8
0
1
9
8
9
3
2
6
n
o
_
a
_
0
0
0
6
0
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Consolidation in language and motor learning
component of language learning is the extraction of regularities embedded in the trained input.
This component starts to emerge during training and continues to evolve during the consoli-
dation period regardless of the timing of sleep. This grammar learning component, which may
depend on frontostriatal skill-learning mechanisms, was similar to motor skill learning and as
such reflects a commonality across domains in consolidation of skill. These similarities on the
group level were further reinforced by cross-domain correlations at the individual level.
ACKNOWLEDGMENTS
We acknowledge the data collection team and specifically Palzur Alon, Hagag Adi Anna, and
Geskin Kate, research assistants, for coordinating the experiments. This work was supported by
the National Institute for Neurobiology in Israel (NiPi) grant no. 208-11-12 and the Israel
Science Foundation (ISF) grant no. 1052/16 to Tali Bitan.
FUNDING INFORMATION
Tali Bitan, National Institute for Neurobiology in Israel (NiPi), Award ID: 208-11-12. Tali Bitan,
Israel Science Foundation, Award ID: 1052/16.
AUTHOR CONTRIBUTIONS
Dafna Ben-Zion: Conceptualization: Lead; Data curation: Lead; Formal analysis: Lead;
Funding acquisition: Equal; Investigation: Lead; Methodology: Equal; Project administration:
Lead; Resources: Lead; Supervision: Lead; Visualization: Lead; Writing – original & review:
Lead. Ella Gabitov: Data curation: Support; Software: Support. *Anat Prior: Conceptualization:
Equal; Methodology: Equal; Supervision: Equal; Visualization: Equal; Writing – original &
review: Equal. *Tali Bitan: Conceptualization: Equal; Funding acquisition: Lead; Methodology:
Equal; Supervision: Equal; Visualization: Equal; Writing – original & review: Equal. (*Equal
contribution.)
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
2
1
8
0
1
9
8
9
3
2
6
n
o
_
a
_
0
0
0
6
0
p
d
.
/
l
REFERENCES
Abend, R., Karni, A., Sadeh, A., Fox, N. A., Pine, D. S., & Bar-Haim,
Y. (2013). Learning to attend to threat accelerates and enhances
memory consolidation. PLOS ONE, 8(4), Article e62501. https://
doi.org/10.1371/journal.pone.0062501, PubMed: 23638100
Adi-Japha, E., Fox, O., & Karni, A. (2011). Atypical acquisition and
atypical expression of memory consolidation gains in a motor
skill in young female adults with ADHD. Research in Develop-
mental Disabilities, 32(3), 1011–1020. https://doi.org/10.1016/j
.ridd.2011.01.048, PubMed: 21349685
Albouy, G., Fogel, S., King, B. R., Laventure, S., Benali, H., Karni,
A., Carrier, J., Robertson, E. M., & Doyon, J. (2015). Maintaining
vs. enhancing motor sequence memories: Respective roles of
striatal and hippocampal systems. NeuroImage, 108, 423–434.
https://doi.org/10.1016/j.neuroimage.2014.12.049, PubMed:
25542533
Albouy, G., Sterpenich, V., Balteau, E., Vandewalle, G., Desseilles,
M., Dang-Vu, T., Darsaud, A., Ruby, P., Luppi, P. H., Degueldre,
C., Peigneux, P., Luxen, A., & Maquet, P. (2008). Both the hippo-
campus and striatum are involved in consolidation of motor
sequence memory. Neuron, 58(2), 261–272. https://doi.org/10
.1016/j.neuron.2008.02.008, PubMed: 18439410
Batterink, L. J., Oudiette, D., Reber, P. J., & Paller, K. A. (2014). Sleep
facilitates learning a new linguistic rule. Neuropsychologia, 65,
169–179. https://doi.org/10.1016/j.neuropsychologia.2014.10
.024, PubMed: 25447376
Ben-Dror, I., & Shani, M. (1996). Phonological awareness test.
Mofet.
Ben Zion, D., Nevat, M., Prior, A., & Bitan, T. (2019). Prior knowl-
edge predicts early consolidation in second language learning.
Frontiers in Psychology, 10, Article 2312. https://doi.org/10
.3389/fpsyg.2019.02312, PubMed: 31681106
Brashers-Krug, T., Shadmehr, R., & Bizzi, E. (1996). Consolidation
in human motor memory. Nature, 382(6588), 252–255. https://
doi.org/10.1038/382252a0, PubMed: 8717039
Brawn, T. P., Fenn, K. M., Nusbaum, H. C., & Margoliash, D.
(2010). Consolidating the effects of waking and sleep on
motor-sequence learning. Journal of Neuroscience, 30(42),
13977–13982. https://doi.org/10.1523/ JNEUROSCI.3295-10
.2010, PubMed: 20962219
Brysbaert, M. (2019). How many participants do we have to
include in properly powered experiments? A tutorial of power
analysis with reference tables. Journal of Cognition, 2(1), Article
16. https://doi.org/10.5334/joc.72, PubMed: 31517234
Cellini, N. (2017). Memory consolidation in sleep disorders. Sleep
Medicine Reviews, 35, 101–112. https://doi.org/10.1016/j.smrv
.2016.09.003, PubMed: 27765468
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neurobiology of Language
207
Consolidation in language and motor learning
Censor, N., Dimyan, M. A., & Cohen, L. G. (2010). Modification of
existing human motor memories is enabled by primary cortical
processing during memory reactivation. Current Biology, 20(17),
1545–1549. https://doi.org/10.1016/j.cub.2010.07.047,
PubMed: 20817532
Censor, N., Karni, A., & Sagi, D. (2006). A link between perceptual
learning, adaptation and sleep. Vision Research, 46(23), 4071–4074.
https://doi.org/10.1016/j.visres.2006.07.022, PubMed: 16979688
Censor, N., Sagi, D., & Cohen, L. G. (2012). Common mechanisms
of human perceptual and motor learning. Nature Reviews Neu-
roscience, 13(9), 658–664. https://doi.org/10.1038/nrn3315,
PubMed: 22903222
Chan, S. H., Ryan, L., & Bever, T. G. (2013). Role of the striatum in
language: Syntactic and conceptual sequencing. Brain and
Language, 125(3), 283–294. https://doi.org/10.1016/j.bandl
.2011.11.005, PubMed: 22200490
Cohen-Mimran, R. (2009). The contribution of language skills to
reading fluency: A comparison of two orthographies for Hebrew.
Journal of Child Language, 36(3), 657–672. https://doi.org/10
.1017/S0305000908009148, PubMed: 19134231
Cona, G., & Semenza, C. (2017). Supplementary motor area as key
structure for domain-general sequence processing: A unified
account. Neuroscience and Biobehavioral Reviews, 72, 28–42.
https://doi.org/10.1016/j.neubiorev.2016.10.033, PubMed:
27856331
Cordi, M. J., & Rasch, B. (2021). How robust are sleep-mediated
memory benefits? Current Opinion in Neurobiology, 67, 1–7.
https://doi.org/10.1016/j.conb.2020.06.002, PubMed: 32711356
Coynel, D., Marrelec, G., Perlbarg, V., Pélégrini-Issac, M., Van de
Moortele, P. F., Ugurbil, K., Doyon, J., Benali, H., & Lehéricy,
S. (2010). Dynamics of motor-related functional integration dur-
ing motor sequence learning. NeuroImage, 49(1), 759–766.
https://doi.org/10.1016/j.neuroimage.2009.08.048, PubMed:
19716894
Crick, F., & Mitchison, G. (1983). The function of dream sleep.
Nature, 304(5922), 111–114. https://doi.org/10.1038/304111a0,
PubMed: 6866101
Davis, M. H., & Gaskell, M. G. (2009). A complementary systems
account of word learning: Neural and behavioural evidence.
Philosophical Transactions of the Royal Society B: Biological
Sciences, 364(1536), 3773–3800. https://doi.org/10.1098/rstb
.2009.0111, PubMed: 19933145
Debas, K., Carrier, J., Barakat, M., Marrelec, G., Bellec, P., Tahar,
A. H., Karni, A., Ungerleider, L. G., Benali, H., & Doyon, J.
(2014). Off-line consolidation of motor sequence learning results
in greater integration within a cortico-striatal functional network.
NeuroImage, 99, 50–58. https://doi.org/10.1016/j.neuroimage
.2014.05.022, PubMed: 24844748
Debas, K., Carrier, J., Orban, P., Barakat, M., Lungu, O., Vandewalle,
G., Tahar, A. H., Bellec, P., Karni, A., Ungerleider, L. G., Benali, H.,
& Doyon, J. (2010). Brain plasticity related to the consolidation of
motor sequence learning and motor adaptation. Proceedings
of the National Academy of Sciences of the United States of
America, 107(41), 17839–17844. https://doi.org/10.1073/pnas
.1013176107, PubMed: 20876115
Denis, D., Schapiro, A. C., Poskanzer, C., Bursal, V., Charon, L.,
Morgan, A., & Stickgold, R. (2020). The roles of item exposure
and visualization success in the consolidation of memories across
wake and sleep. Learning and Memory, 27(11), 451–456. https://
doi.org/10.1101/LM.051383.120, PubMed: 33060281
Diekelmann, S., & Born, J. (2010). The memory function of sleep.
Nature Reviews Neuroscience, 11(2), 114–126. https://doi.org/10
.1038/nrn2762, PubMed: 20046194
Diekelmann, S., Wilhelm, I., & Born, J. (2009). The whats and
whens of sleep-dependent memory consolidation. Sleep Medi-
cine Reviews, 13(5), 309–321. https://doi.org/10.1016/j.smrv
.2008.08.002, PubMed: 19251443
Dorfberger, S., Adi-Japha, E., & Karni, A. (2012). Sequence specific
motor performance gains after memory consolidation in children
and adolescents. PLOS ONE, 7(1), Article e28673. https://doi.org
/10.1371/journal.pone.0028673, PubMed: 22276097
Doyon, J., Gabitov, E., Vahdat, S., Lungu, O., & Boutin, A. (2018).
Current issues related to motor sequence learning in humans.
Current Opinion in Behavioral Sciences, 20, 89–97. https://doi
.org/10.1016/j.cobeha.2017.11.012
Doyon, J., Korman, M., Morin, A., Dostie, V., Tahar, A. H., Benali,
H., Karni, A., Ungerleider, L. G., & Carrier, J. (2009). Contribution
of night and day sleep vs. simple passage of time to the consoli-
dation of motor sequence and visuomotor adaptation learning.
Experimental Brain Research, 195(1), 15–26. https://doi.org/10
.1007/s00221-009-1748-y, PubMed: 19277618
Dudai, Y. (2004). The neurobiology of consolidations, or, how
stable is the engram? Annual Review of Psychology, 55(1), 51–86.
https://doi.org/10.1146/annurev.psych.55.090902.142050,
PubMed: 14744210
Dudai, Y., Karni, A., & Born, J. (2015). The consolidation and trans-
formation of memory. Neuron, 88(1), 20–32. https://doi.org/10
.1016/j.neuron.2015.09.004, PubMed: 26447570
Dumay, N. (2016). Sleep not just protects memories against forget-
ting, it also makes them more accessible. Cortex, 74, 289–296.
https://doi.org/10.1016/j.cortex.2015.06.007, PubMed:
26227582
Dumay, N., & Gaskell, M. G. (2007). Sleep-associated changes in the
mental representation of spoken words. Psychological Science,
18(1), 35–39. https://doi.org/10.1111/j.1467-9280.2007.01845.x,
PubMed: 17362375
Durrant, S. J., Taylor, C., Cairney, S., & Lewis, P. A. (2011). Sleep-
dependent consolidation of statistical learning. Neuropsychologia,
49(5), 1322–1331. https://doi.org/10.1016/j.neuropsychologia
.2011.02.015, PubMed: 21335017
Ellenbogen, J. M., Hu, P. T., Payne, J. D., Titone, D., & Walker, M. P.
(2007). Human relational memory requires time and sleep.
Proceedings of the National Academy of Sciences of the United
States of America, 104(18), 7723–7728. https://doi.org/10.1073
/pnas.0700094104, PubMed: 17449637
Ellenbogen, J. M., Hulbert, J. C., Stickgold, R., Dinges, D. F., &
Thompson-Schill, S. L. (2006). Interfering with theories of sleep
and memory: Sleep, declarative memory, and associative inter-
ference. Current Biology, 16(13), 1290–1294. https://doi.org/10
.1016/j.cub.2006.05.024, PubMed: 16824917
Ellenbogen, J. M., Payne, J. D., & Stickgold, R. (2006). The role of sleep
in declarative memory consolidation: Passive, permissive, active or
none? Current Opinion in Neurobiology, 16(6), 716–722. https://doi
.org/10.1016/j.conb.2006.10.006, PubMed: 17085038
Eriksson, J., Kalpouzos, G., & Nyberg, L. (2011). Rewiring the brain
with repeated retrieval: A parametric fMRI study of the testing
effect. Neuroscience Letters, 505(1), 36–40. https://doi.org/10
.1016/j.neulet.2011.08.061, PubMed: 21983436
Eyigoz, E., Courson, M., Sedeño, L., Rogg, K., Orozco-Arroyave,
J. R., Nöth, E., Skodda, S., Trujillo, N., Rodríguez, M., Rusz, J.,
Muñoz, E., Cardona, J. F., Herrera, E., Hesse, E., Ibáñez, A.,
Cecchi, G., & García, A. M. (2020). From discourse to pathology:
Automatic identification of Parkinson’s disease patients via
morphological measures across three languages. Cortex, 132,
191–205. https://doi.org/10.1016/j.cortex.2020.08.020,
PubMed: 32992069
Neurobiology of Language
208
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
2
1
8
0
1
9
8
9
3
2
6
n
o
_
a
_
0
0
0
6
0
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Consolidation in language and motor learning
Faul, F., Erdfelder, E., Lang, A. G., & Buchner, A. (2007). G*Power
3: A flexible statistical power analysis program for the social,
behavioral, and biomedical sciences. Behavior Research
Methods, 39(2), 175–191. https://doi.org/10.3758/BF03193146,
PubMed: 17695343
Fenn, K. M., & Hambrick, D. Z. (2012). Individual differences in
working memory capacity predict sleep-dependent memory
consolidation. Journal of Experimental Psychology: General,
141(3), 404–410. https://doi.org/10.1037/a0025268, PubMed:
21910555
Fenn, K. M., & Hambrick, D. Z. (2015). General intelligence pre-
dicts memory change across sleep. Psychonomic Bulletin and
Review, 22(3), 791–799. https://doi.org/10.3758/s13423-014
-0731-1, PubMed: 25253197
Fenn, K. M., Nusbaum, H. C., & Margoliash, D. (2003). Consolida-
tion during sleep of perceptual learning of spoken language.
Nature, 425(6958), 614–616. https://doi.org/10.1038
/nature01951, PubMed: 14534586
Ferstl, E. C., Neumann, J., Bogler, C., & Von Cramon, D. Y. (2008).
The extended language network: A meta-analysis of neuroimaging
studies on text comprehension. Human Brain Mapping, 29(5),
581–593. https://doi.org/10.1002/ hbm.20422, PubMed:
17557297
Fischer, S., Drosopoulos, S., Tsen, J., & Born, J. (2006). Implicit
learning-explicit knowing: A role for sleep in memory system
interaction. Journal of Cognitive Neuroscience, 18(3), 311–319.
https://doi.org/10.1162/jocn.2006.18.3.311, PubMed: 16602193
Fischer, S., Hallschmid, M., Elsner, A. L., & Born, J. (2002). Sleep
forms memory for finger skills. Proceedings of the National
Academy of Sciences of the United States of America, 99(18),
11987–11991. https://doi.org/10.1073/pnas.182178199,
PubMed: 12193650
Fischer, S., Nitschke, M. F., Melchert, U. H., Erdmann, C., & Born,
J. (2005). Motor memory consolidation in sleep shapes more
effective neuronal representations. Journal of Neuroscience,
25(49), 11248–11255. https://doi.org/10.1523/ JNEUROSCI
.1743-05.2005, PubMed: 16339020
Gabitov, E., Boutin, A., Pinsard, B., Censor, N., Fogel, S. M.,
Albouy, G., King, B. R., Benali, H., Carrier, J., Cohen, L. G.,
Karni, A., & Doyon, J. (2017). Re-stepping into the same river:
Competition problem rather than a reconsolidation failure in
an established motor skill. Scientific Reports, 7(1), 1–13. https://
doi.org/10.1038/s41598-017-09677-1, PubMed: 28839217
Gabitov, E., Lungu, O., Albouy, G., & Doyon, J. (2019). Weaker
inter-hemispheric and local functional connectivity of the soma-
tomotor cortex during a motor skill acquisition is associated with
better learning. Frontiers in Neurology, 10, Article 1242. https://
doi.org/10.3389/fneur.2019.01242, PubMed: 31827459
Gabitov, E., Manor, D., & Karni, A. (2014). Done that: Short-term
repetition related modulations of motor cortex activity as a stable
signature for overnight motor memory consolidation. Journal of
Cognitive Neuroscience, 26(12), 2716–2734. https://doi.org/10
.1162/jocn_a_00675, PubMed: 24893741
Gabitov, E., Manor, D., & Karni, A. (2015). Patterns of modulation
in the activity and connectivity of motor cortex during the
repeated generation of movement sequences. Journal of Cogni-
tive Neuroscience, 27(4), 736–751. https://doi.org/10.1162/jocn
_a_00751, PubMed: 25390206
Gabitov, E., Manor, D., & Karni, A. (2016). Learning from the other
limb’s experience: Sharing the ‘trained’ M1 representation of
the motor sequence knowledge. Journal of Physiology, 594(1),
169–188. https://doi.org/10.1113/ JP270184, PubMed:
26442464
Gais, S., Lucas, B., & Born, J. (2006). Sleep after learning aids
memory recall. Learning and Memory, 13(3), 259–262. https://
doi.org/10.1101/lm.132106, PubMed: 16741280
Gal, C., Gabitov, E., Maaravi-Hesseg, R., Karni, A., & Korman, M.
(2019). A delayed advantage: Multi-session training at evening
hours leads to better long-term retention of motor skill in the
elderly. Frontiers in Aging Neuroscience, 11, Article 321.
https://doi.org/10.3389/fnagi.2019.00321, PubMed: 31824300
Gregory, M. D., Agam, Y., Selvadurai, C., Nagy, A., Vangel, M.,
Tucker, M., Robertson, E. M., Stickgold, R., & Manoach, D. S.
(2014). Resting state connectivity immediately following learning
correlates with subsequent sleep-dependent enhancement of
motor task performance. NeuroImage, 102(2), 666–673. https://
doi.org/10.1016/j.neuroimage.2014.08.044, PubMed: 25173415
Hedenius, M., Persson, J., Tremblay, A., Adi-Japha, E., Veríssimo, J.,
Dye, C. D., Alm, P., Jennische, M., Bruce Tomblin, J., & Ullman,
M. T. (2011). Grammar predicts procedural learning and consol-
idation deficits in children with specific language impairment.
Research in Developmental Disabilities, 32(6), 2362–2375.
https://doi.org/10.1016/j.ridd.2011.07.026, PubMed: 21840165
Heim, S., Klann, J., Schattka, K. I., Bauhoff, S., Borcherding, G.,
Nosbüsch, N., Struth, L., Binkofski, F. C., & Werner, C. J.
(2017). A nap but not rest or activity consolidates language learn-
ing. Frontiers in Psychology, 8, Article 665. https://doi.org/10
.3389/fpsyg.2017.00665, PubMed: 28559856
Henderson, L.-M., & Warmington, M. (2017). A sequence learning
impairment in dyslexia? It depends on the task. Research in
Developmental Disabilities, 60, 198–210. https://doi.org/10
.1016/j.ridd.2016.11.002, PubMed: 27856107
Henderson, L.-M., Weighall, A. R., Brown, H., & Gaskell, M. G.
(2012). Consolidation of vocabulary is associated with sleep in
children. Developmental Science, 15(5), 674–687. https://doi
.org/10.1111/j.1467-7687.2012.01172.x, PubMed: 22925515
Henderson, L.[-M.], Weighall, A., & Gaskell, G. (2013). Learning
new vocabulary during childhood: Effects of semantic training
on lexical consolidation and integration. Journal of Experimental
Child Psychology, 116(3), 572–592. https://doi.org/10.1016/j
.jecp.2013.07.004, PubMed: 23981272
Hertrich, I., Dietrich, S., & Ackermann, H. (2016). The role of the
supplementary motor area for speech and language processing.
Neuroscience and Biobehavioral Reviews, 68, 602–610. https://
doi.org/10.1016/j.neubiorev.2016.06.030, PubMed: 27343998
Hikosaka, O., Nakamura, K., Sakai, K., & Nakahara, H. (2002).
Central mechanisms of motor skill learning. Current Opinion in
Neurobiology, 12(2), 217–222. https://doi.org/10.1016/S0959
-4388(02)00307-0
Horne, J. A., & Ostberg, O. (1976). A self-assessment questionnaire
to determine morningness-eveningness in human circadian
rhythms. International Journal of Chronobiology, 4(2), 97–110.
PubMed: 1027738
Hu, X., Cheng, L. Y., Chiu, M. H., & Paller, K. A. (2020). Promoting
memory consolidation during sleep: A meta-analysis of targeted
memory reactivation. Psychological Bulletin, 146(3), 218–244.
https://doi.org/10.1037/bul0000223, PubMed: 32027149
James, E., Gaskell, M. G., Weighall, A., & Henderson, L.-M. (2017).
Consolidation of vocabulary during sleep: The rich get richer?
Neuroscience and Biobehavioral Reviews, 77, 1–13. https://doi
.org/10.1016/j.neubiorev.2017.01.054, PubMed: 28274725
Jenkins, J. G., & Dallenbach, K. M. (1924). Obliviscence during
sleep and waking. American Journal of Psychology, 35(4),
605–612. https://doi.org/10.2307/1414040
Karni, A. (1995). When practice makes perfect. Lancet, 345(8946),
395. https://doi.org/10.1016/S0140-6736(95)90386-0
Neurobiology of Language
209
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
2
1
8
0
1
9
8
9
3
2
6
n
o
_
a
_
0
0
0
6
0
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Consolidation in language and motor learning
Karni, A., Meyer, G., Rey-Hipolito, C., Jezzard, P., Adams, M. M.,
Turner, R., & Ungerleider, L. G. (1998). The acquisition of
skilled motor performance: Fast and slow experience-driven
changes in primary motor cortex. Proceedings of the National
Academy of Sciences of the United States of America, 95(3),
861–868. https://doi.org/10.1073/pnas.95.3.861, PubMed:
9448252
Kemény, F., & Lukács, Á. (2016). Sleep-independent off-line
enhancement and time of the day effects in three forms of skill
learning. Cognitive Processing, 17(2), 163–174. https://doi.org/10
.1007/s10339-016-0750-0, PubMed: 26861244
Khandelwal, U., Levy, O., Jurafsky, D., Zettlemoyer, L., & Lewis, M.
(2019). Generalization through memorization: Nearest neighbor
language models. ArXiv, 1911.00172. https://arxiv.org/abs/1911
.00172
Kim, K. M., & Fenn, K. M. (2020). Sleep-dependent consolidation
of second language grammar knowledge. Studies in Second
Language Acquisition, 42(5), 1107–1120. https://doi.org/10
.1017/S0272263120000200
King, B. R., Hoedlmoser, K., Hirschauer, F., Dolfen, N., & Albouy,
G. (2017). Sleeping on the motor engram: The multifaceted
nature of sleep-related motor memory consolidation. Neurosci-
ence and Biobehavioral Reviews, 80, 1–22. https://doi.org/10
.1016/j.neubiorev.2017.04.026, PubMed: 28465166
Klinzing, J. G., Niethard, N., & Born, J. (2019). Mechanisms of sys-
tems memory consolidation during sleep. Nature Neuroscience,
22(10), 1598–1610. https://doi.org/10.1038/s41593-019-0467-3,
PubMed: 31451802
Korman, M., Doyon, J., Doljansky, J., Carrier, J., Dagan, Y., & Karni, A.
(2007). Daytime sleep condenses the time course of motor memory
consolidation. Nature Neuroscience, 10(9), 1206–1213. https://doi
.org/10.1038/nn1959, PubMed: 17694051
Korman, M., Gal, C., Gabitov, E., & Karni, A. (2021). Better later:
Evening practice is advantageous for motor skill consolidation in
the elderly. Learning and Memory, 28(3), 72–75. https://doi.org
/10.1101/LM.052522.120, PubMed: 33593924
Korman, M., Raz, N., Flash, T., & Karni, A. (2003). Multiple shifts in
the representation of a motor sequence during the acquisition of
skilled performance. Proceedings of the National Academy of
Sciences of the United States of America, 100(21), 12492–12497.
https://doi.org/10.1073/pnas.2035019100, PubMed: 14530407
Kroneisen, M., & Kuepper-Tetzel, C. E. (2021). Using day and
night – Scheduling retrieval practice and sleep. Psychology
Learning and Teaching, 20(1), 40–57. https://doi.org/10.1177
/1475725720965363
Kumaran, D., & McClelland, J. L. (2012). Generalization through
the recurrent interaction of episodic memories: A model of the
hippocampal system. Psychological Review, 119(3), 573–616.
https://doi.org/10.1037/a0028681, PubMed: 22775499
Landi, N., Malins, J. G., Frost, S. J., Magnuson, J. S., Molfese, P.,
Ryherd, K., Rueckl, J. G., Mencl, W. E., & Pugh, K. R. (2018).
Neural representations for newly learned words are modulated
by overnight consolidation, reading skill, and age. Neuropsycho-
logia, 111, 133–144. https://doi.org/10.1016/j.neuropsychologia
.2018.01.011, PubMed: 29366948
Lehéricy, S., Benali, H., Van de Moortele, P.-F., Pélégrini-Issac, M.,
Waechter, T., Ugurbil, K., & Doyon, J. (2005). Distinct basal
ganglia territories are engaged in early and advanced motor
sequence learning. Proceedings of the National Academy of Sci-
ences of the United States of America, 102(35), 12566–12571.
https://doi.org/10.1073/pnas.0502762102, PubMed: 16107540
Lerner, I., & Gluck, M. A. (2019). Sleep and the extraction of hid-
den regularities: A systematic review and the importance of
temporal rules. Sleep Medicine Reviews, 47, 39–50. https://doi
.org/10.1016/j.smrv.2019.05.004, PubMed: 31252335
Lewis, P. A., & Durrant, S. J. (2011). Overlapping memory replay
during sleep builds cognitive schemata. Trends in Cognitive
Sciences, 15(8), 343–351. https://doi.org/10.1016/j.tics.2011.06
.004, PubMed: 21764357
Lum, J. A. G., Conti-Ramsden, G., Morgan, A. T., & Ullman, M. T.
(2014). Procedural learning deficits in specific language impair-
ment (SLI): A meta-analysis of serial reaction time task perfor-
mance. Cortex, 51(1), 1–10. https://doi.org/10.1016/j.cortex
.2013.10.011, PubMed: 24315731
Maltry, L., Ohlendorf, D., Bendels, M., Groneberg, D. A., & Mickel,
C. (2020). Does sleep contribute to the consolidation of motor
memory?: A literature review. Manuelle Medizin, 59, 26–32.
https://doi.org/10.1007/s00337-020-00674-6
McClelland, J. L. (2013). Incorporating rapid neocortical learning
of new schema-consistent information into complementary
learning systems theory. Journal of Experimental Psychology:
General, 142(4), 1190–1210. https://doi.org/10.1037/a0033812,
PubMed: 23978185
McClelland, J. L., McNaughton, B. L., & O’Reilly, R. C. (1995).
Why there are complementary learning systems in the hippo-
campus and neocortex: Insights from the successes and failures
of connectionist models of learning and memory. Psychological
Review, 102(3), 419–457. https://doi.org/10.1037/0033-295X
.102.3.419, PubMed: 7624455
Mednick, S. C., Cai, D. J., Shuman, T., Anagnostaras, S., & Wixted,
J. T. (2011). An opportunistic theory of cellular and systems
consolidation. Trends in Neurosciences, 34(10), 504–514.
https://doi.org/10.1016/j.tins.2011.06.003, PubMed: 21742389
Mednick, S. C., McDevitt, E. A., Walsh, J. K., Wamsley, E., Paulus,
M., Kanady, J. C., & Drummond, S. P. A (2013). The critical role
of sleep spindles in hippocampal-dependent memory: A pharma-
cology study. Journal of Neuroscience, 33(10), 4494–4504.
https://doi.org/10.1523/ JNEUROSCI.3127-12.2013, PubMed:
23467365
Meier, B., & Cock, J. (2014). Offline consolidation in implicit
sequence learning. Cortex, 57, 156–166. https://doi.org/10
.1016/j.cortex.2014.03.009, PubMed: 24861420
Mirković, J., & Gaskell, M. (2016). Does sleep improve your gram-
mar? Preferential consolidation of arbitrary components of new
linguistic knowledge. PLOS ONE, 11(4), Article e0152489.
https://doi.org/10.1371/journal.pone.0152489, PubMed:
27046022
Mölle, M., Bergmann, T. O., Marshall, L., & Born, J. (2011). Fast and
slow spindles during the sleep slow oscillation: Disparate coales-
cence and engagement in memory processing. Sleep, 34(10),
1411–1421. https://doi.org/10.5665/SLEEP.1290, PubMed:
21966073
Morcos, A. S., Barrett, D. G. T., Rabinowitz, N. C., & Botvinick, M.
(2018). On the importance of single directions for generalization.
6th International Conference on Learning Representations (ICLR
2018), Vancouver, BC, Canada. ArXiv, 803.06959v4. https://
arxiv.org/abs/1803.06959
Morin, A., Doyon, J., Dostie, V., Barakat, M., Tahar, A. H., Korman,
M., Benali, H., Karni, A., Ungerleider, L. G., & Carrier, J. (2008).
Motor sequence learning increases sleep spindles and fast
frequencies in post-training sleep. Sleep, 31(8), 1149–1156.
https://doi.org/10.5665/sleep/31.8.1149, PubMed: 18714787
Nakai, T., Matsuo, K., Kato, C., Matsuzawa, M., Okada, T., Glover,
G. H., Moriya, T., & Inui, T. (1999). A functional magnetic reso-
nance imaging study of listening comprehension of languages in
human at 3 tesla-comprehension level and activation of the
Neurobiology of Language
210
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
2
1
8
0
1
9
8
9
3
2
6
n
o
_
a
_
0
0
0
6
0
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Consolidation in language and motor learning
language areas. Neuroscience Letters, 263(1), 33–36. https://doi
.org/10.1016/S0304-3940(99)00103-2, PubMed: 10218904
Natale, V., Fabbri, M., Tonetti, L., & Martoni, M. (2014). Psycho-
metric goodness of the Mini Sleep Questionnaire. Psychiatry
and Clinical Neurosciences, 68(7), 568–573. https://doi.org/10
.1111/pcn.12161, PubMed: 24447273
Needle, J., Nicolson, R. I., & Fawcett, A. J. (2015). Motor sequence
learning in dyslexia: Is consolidation the key? BPA - Applied
Psychology Bulletin (Bollettino Di Psicologia Applicata),
63(273), 5–15.
Nettersheim, A., Hallschmid, M., Born, J., & Diekelmann, S. (2015).
The role of sleep in motor sequence consolidation: Stabilization
rather than enhancement. Journal of Neuroscience, 35(17),
6696–6702. https://doi.org/10.1523/JNEUROSCI.1236-14.2015,
PubMed: 25926448
Nevat, M., Ullman, M. T., Eviatar, Z., & Bitan, T. (2017). The neural
bases of the learning and generalization of morphological inflec-
tion. Neuropsychologia, 98, 139–155. https://doi.org/10.1016/j
.neuropsychologia.2016.08.026, PubMed: 27575853
Nevat, M., Ullman, M. T., Eviatar, Z., & Bitan, T. (2018). The role of
distributional factors in learning and generalising affixal plural
inflection: An artificial language study. Language, Cognition
and Neuroscience, 33(9), 1184–1204. https://doi.org/10.1080
/23273798.2018.1465187
Nieuwenhuis, I. L. C., Folia, V., Forkstam, C., Jensen, O., & Petersson,
K. M. (2013). Sleep promotes the extraction of grammatical
rules. PLOS ONE, 8(6), Article e65046. https://doi.org/10.1371
/journal.pone.0065046, PubMed: 23755173
Orban, P., Peigneux, P., Lungu, O., Albouy, G., Breton, E., Laberenne,
F., Benali, H., Maquet, P., & Doyon, J. (2010). The multifaceted
nature of the relationship between performance and brain
activity in motor sequence learning. NeuroImage, 49(1), 694–702.
https://doi.org/10.1016/j.neuroimage.2009.08.055, PubMed:
19732838
Pan, S. C., & Rickard, T. C. (2015). Sleep and motor learning: Is there
room for consolidation? Psychological Bulletin, 141(4), 812–834.
https://doi.org/10.1037/bul0000009, PubMed: 25822130
Panzer, S., Krueger, M., Muehlbauer, T., Kovacs, A. J., & Shea, C. H.
(2009). Inter-manual transfer and practice: Coding of simple
motor sequences. Acta Psychologica, 131(2), 99–109. https://
doi.org/10.1016/j.actpsy.2009.03.004, PubMed: 19389659
Payne, J. D., Tucker, M. A., Ellenbogen, J. M., Wamsley, E. J.,
Walker, M. P., Schacter, D. L., & Stickgold, R. (2012). Memory
for semantically related and unrelated declarative information:
The benefit of sleep, the cost of wake. PLOS ONE, 7(3), Article
e33079. https://doi.org/10.1371/journal.pone.0033079,
PubMed: 22457736
Peiffer, A., Brichet, M., De Tiège, X., Peigneux, P., & Urbain, C.
(2020). The power of children’s sleep - Improved declarative
memory consolidation in children compared with adults. Scien-
tific Reports, 10, Article 9979. https://doi.org/10.1038/s41598
-020-66880-3, PubMed: 32561803
Penhune, V. B., & Steele, C. J. (2012). Parallel contributions of cer-
ebellar, striatal and M1 mechanisms to motor sequence learning.
Behavioural Brain Research, 226(2), 579–591. https://doi.org/10
.1016/j.bbr.2011.09.044, PubMed: 22004979
Peters, K. R., Ray, L., Smith, V., & Smith, C. (2008). Changes in the
density of stage 2 sleep spindles following motor learning in
young and older adults. Journal of Sleep Research, 17(1), 23–33.
https://doi.org/10.1111/j.1365-2869.2008.00634.x, PubMed:
18275552
Plihal, W., & Born, J. (1997). Effects of early and late nocturnal
sleep on declarative and procedural memory. Journal of
Cognitive Neuroscience, 9(4), 534–547. https://doi.org/10.1162
/jocn.1997.9.4.534, PubMed: 23968216
Rasch, B., & Born, J. (2013). About sleep’s role in memory. Physi-
ological Reviews, 93(2), 681–766. https://doi.org/10.1152
/physrev.00032.2012, PubMed: 23589831
Rasch, B., Büchel, C., Gais, S., & Born, J. (2007). Odor cues during
slow-wave sleep prompt declarative memory consolidation. Sci-
ence, 315(5817), 1426–1429. https://doi.org/10.1126/science
.1138581, PubMed: 17347444
Raz-Salzburg, E., & Ravid, D. (2009). Verb-patterns acquisition in
Hebrew from kindergarten age through adulthood [Unpublished
manuscript]. Department of Communications Disorders, Tel Aviv
University.
Rickard, T. C., Cai, D. J., Rieth, C. A., Jones, J., & Ard, M. C. (2008).
Sleep does not enhance motor sequence learning. Journal of
Experimental Psychology: Learning Memory and Cognition,
34(4), 834–842. https://doi.org/10.1037/0278-7393.34.4.834,
PubMed: 18605872
Robertson, E. M. (2012). New insights in human memory interfer-
ence and consolidation. Current Biology, 22(2), R66–R71. https://
doi.org/10.1016/j.cub.2011.11.051, PubMed: 22280913
Robertson, E. M., Pascual-Leone, A., & Miall, R. C. (2004). Current
concepts in procedural consolidation. Nature Reviews Neurosci-
ence, 5(7), 576–582. https://doi.org/10.1038/nrn1426, PubMed:
15208699
Robertson, E. M., Pascual-Leone, A., & Press, D. Z. (2004). Aware-
ness modifies the skill-learning benefits of sleep. Current Biology,
14(3), 208–212. https://doi.org/10.1016/j.cub.2004.01.027,
PubMed: 14761652
Rozanov, S., Keren, O., & Karni, A. (2010). The specificity of
memory for a highly trained finger movement sequence:
Change the ending, change all. Brain Research, 1331, 80–87.
https://doi.org/10.1016/j.brainres.2010.03.019, PubMed:
20298683
Sawangjit, A., Oyanedel, C. N., Niethard, N., Salazar, C., Born, J.,
& Inostroza, M. (2018). The hippocampus is crucial for forming
non-hippocampal long-term memory during sleep. Nature, 564,
109–113. https://doi.org/10.1038/s41586-018-0716-8, PubMed:
30429612
Schabus, M., Gruber, G., Parapatics, S., Sauter, C., Klösch, G.,
Anderer, P., Klimesch, W., Saletu, B., & Zeitlhofer, J. (2004).
Sleep spindles and their significance for declarative memory
consolidation. Sleep, 27(8), 1479–1485. https://doi.org/10.1093
/sleep/27.7.1479, PubMed: 15683137
Schambra, H. M., Abe, M., Luckenbaugh, D. A., Reis, J., Krakauer,
J. W., & Cohen, L. G. (2011). Probing for hemispheric specializa-
tion for motor skill learning: A transcranial direct current stimu-
lation study. Journal of Neurophysiology, 106(2), 652–661.
https://doi.org/10.1152/jn.00210.2011, PubMed: 21613597
Schmidt, D., Erlacher, D., Klostermann, A., Kredel, R., & Hossner,
E.-J. (2020). Sleep-dependent motor memory consolidation in
healthy adults: A meta-analysis. Neuroscience and Biobehavioral
Reviews, 118, 270–281. https://doi.org/10.1016/j.neubiorev
.2020.07.028, PubMed: 32730847
Schoch, S. F., Cordi, M. J., & Rasch, B. (2017). Modulating influ-
ences of memory strength and sensitivity of the retrieval test on
the detectability of the sleep consolidation effect. Neurobiology
of Learning and Memory, 145, 181–189. https://doi.org/10.1016/j
.nlm.2017.10.009, PubMed: 29030296
Schönauer, M., Grätsch, M., & Gais, S. (2015). Evidence for two
distinct sleep-related long-term memory consolidation processes.
Cortex, 63, 68–78. https://doi.org/10.1016/j.cortex.2014.08.005,
PubMed: 25243990
Neurobiology of Language
211
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
2
1
8
0
1
9
8
9
3
2
6
n
o
_
a
_
0
0
0
6
0
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Consolidation in language and motor learning
Schreiner, T., & Rasch, B. (2017). The beneficial role of memory
reactivation for language learning during sleep: A review. Brain
and Language, 167, 94–105. https://doi.org/10.1016/j.bandl
.2016.02.005, PubMed: 27036946
Schreiner, T., & Rasch, B. (2018). To gain or not to gain – The
complex role of sleep for memory: Comment on Dumay
(2016). Cortex, 101, 282–287. https://doi.org/10.1016/j.cortex
.2016.06.011, PubMed: 27423210
Segaert, K., Menenti, L., Weber, K., Petersson, K. M., & Hagoort, P.
(2012). Shared syntax in language production and language
comprehension—An fMRI study. Cerebral Cortex, 22(7),
1662–1670. https://doi.org/10.1093/cercor/ bhr249, PubMed:
21934094
Shatil, E. (1995). One-minute test for words [Unpublished test].
Department of Learning Disabilities, University of Haifa.
Shatil, E. (1997). One-minute test for pseudowords [Unpublished
test]. Department of Learning Disabilities, University of Haifa.
Song, S. (2009). Consciousness and the consolidation of motor
learning. Behavioural Brain Research, 196(2), 180–186. https://
doi.org/10.1016/j.bbr.2008.09.034, PubMed: 18951924
Sonni, A., & Spencer, R. M. C. (2015). Sleep protects memories
from interference in older adults. Neurobiology of Aging, 36(7),
2272–2281. https://doi.org/10.1016/j.neurobiolaging.2015.03
.010, PubMed: 25890819
Spencer, R. M. C., Gouw, A. M., & Ivry, R. B. (2007). Age-related
decline of sleep-dependent consolidation. Learning and Memory,
14(7), 480–484. https://doi.org/10.1101/ lm.569407, PubMed:
17622650
Stickgold, R. (2005). Sleep-dependent memory consolidation.
Nature, 437, 1272–1278. https://doi.org/10.1038/nature04286,
PubMed: 16251952
Stickgold, R. (2009). How do I remember? Let me count the ways.
Sleep Medicine Reviews, 13(5), 305–308. https://doi.org/10.1016
/j.smrv.2009.05.004, PubMed: 19699665
Szmalec, A., Page, M. P. A., & Duyck, W. (2012). The development
of long-term lexical representations through Hebb repetition
learning. Journal of Memory and Language, 67(3), 342–354.
https://doi.org/10.1016/j.jml.2012.07.001
Tamminen, J., Newbury, C. R., Crowley, R., Vinals, L., Cevoli, B., &
Rastle, K. (2020). Generalisation in language learning can with-
stand total sleep deprivation. Neurobiology of Learning and
Memory, 173, Article 107274. https://doi.org/10.1016/j.nlm
.2020.107274, PubMed: 32653634
Tamminen, J., Payne, J. D., Stickgold, R., Wamsley, E. J., & Gaskell,
M. G. (2010). Sleep spindle activity is associated with the inte-
gration of new memories and existing knowledge. Journal of
Neuroscience, 30(43), 14356–14360. https://doi.org/10.1523
/JNEUROSCI.3028-10.2010, PubMed: 20980591
Tononi, G., & Cirelli, C. (2006). Sleep function and synaptic
homeostasis. Sleep Medicine Reviews, 10(1), 49–62. https://doi
.org/10.1016/j.smrv.2005.05.002, PubMed: 16376591
Tononi, G., & Cirelli, C. (2014). Sleep and the price of plasticity:
From synaptic and cellular homeostasis to memory consolidation
and integration. Neuron, 81(1), 12–34. https://doi.org/10.1016/j
.neuron.2013.12.025, PubMed: 24411729
Tucker, M. A., Hirota, Y., Wamsley, E. J., Lau, H., Chaklader, A., &
Fishbein, W. (2006). A daytime nap containing solely non-REM
sleep enhances declarative but not procedural memory. Neuro-
biology of Learning and Memory, 86(2), 241–247. https://doi.org
/10.1016/j.nlm.2006.03.005, PubMed: 16647282
Ullman, M. T. (2015). The declarative/procedural model: A neuro-
biological model of language learning, knowledge, and use. In
G. Hickok & S. L. Small (Eds.), Neurobiology of Language
(pp. 953–968). Academic Press. https://doi.org/10.1016/B978-0
-12-407794-2.00076-6
Ullman, M. T., & Pierpont, E. I. (2005). Specific language impair-
ment is not specific to language: The procedural deficit
hypothesis. Cortex, 41(3), 399–433. https://doi.org/10.1016
/S0010-9452(08)70276-4, PubMed: 15871604
van Doorn, J., van den Bergh, D., Böhm, U., Dablander, F., Derks,
K., Draws, T., Etz, A., Evans, N. J., Gronau, Q. F., Haaf, J. M.,
Hinne, M., Kucharský, Š., Ly, A., Marsman, M., Matzke, D.,
Gupta, A. R. K. N., Sarafoglou, A., Stefan, A., Voelkel, J. G., &
Wagenmakers, E.-J. (2020). The JASP guidelines for conducting
and reporting a Bayesian analysis. Psychonomic Bulletin and
Review, 28, 813–826. https://doi.org/10.3758/s13423-020
-01798-5, PubMed: 33037582
Wagenmakers, E. J., Love, J., Marsman, M., Jamil, T., Ly, A., Verhagen,
J., Selker, R., Gronau, Q. F., Dropmann, D., Boutin, B., Meerhoff, F.,
Knight, P., Raj, A., van Kesteren, E.-J., van Doorn, J., Šmíra, M.,
Epskamp, S., Etz, A., Matzke, D., … Morey, R. D. (2018). Bayesian
inference for psychology. Part II: Example applications with JASP.
Psychonomic Bulletin and Review, 25(1), 58–76. https://doi.org
/10.3758/s13423-017-1323-7, PubMed: 28685272
Wagner, U., Gais, S., Haider, H., Verleger, R., & Born, J. (2004).
Sleep inspires insight. Nature, 427, 352–355. https://doi.org/10
.1038/nature02223, PubMed: 14737168
Walker, M. P., Brakefield, T., Morgan, A., Hobson, J. A., & Stickgold,
R. (2002). Practice with sleep makes perfect: Sleep-dependent
motor skill learning. Neuron, 35(1), 206–211. https://doi.org/10
.1016/S0896-6273(02)00746-8, PubMed: 12123620
Walker, M. P., Brakefield, T., Seidman, J., Morgan, A., Hobson, J. A.,
& Stickgold, R. (2003). Sleep and the time course of motor skill
learning. Learning and Memory, 10(4), 275–284. https://doi.org
/10.1101/lm.58503, PubMed: 12888546
Walker, M. P., & Stickgold, R. (2004). Sleep-dependent learning
and memory consolidation. Neuron, 44(1), 121–133. https://doi
.org/10.1016/j.neuron.2004.08.031, PubMed: 15450165
Wechsler, D. (1955). Manual for the Wechsler Adult Intelligence
Scale. Psychological Corp. https://psycnet.apa.org/record/1955
-07334-000
Weiss, Y., Katzir, T., & Bitan, T. (2015). Many ways to read your
vowels: Neural processing of diacritics and vowel letters in
Hebrew. NeuroImage, 121, 10–19. https://doi.org/10.1016/j
.neuroimage.2015.07.029, PubMed: 26188258
West, G., Vadillo, M. A., Shanks, D. R., & Hulme, C. (2017). The
procedural learning deficit hypothesis of language learning
disorders: We see some problems. Developmental Science, 21(2),
Article e12552. https://doi.org/10.1111/desc.12552, PubMed:
28256101
Wiestler, T., Waters-Metenier, S., & Diedrichsen, J. (2014). Effector-
independent motor sequence representations exist in extrinsic
and intrinsic reference frames. Journal of Neuroscience, 34(14),
5054–5064. https://doi.org/10.1523/JNEUROSCI.5363-13.2014,
PubMed: 24695723
Wilhelm, I., Diekelmann, S., & Born, J. (2008). Sleep in children
improves memory performance on declarative but not proce-
dural tasks. Learning and Memory, 15(5), 373–377. https://doi
.org/10.1101/lm.803708, PubMed: 18441295
Wilhelm, I., Diekelmann, S., Molzow, I., Ayoub, A., Mölle, M., &
Born, J. (2011). Sleep selectively enhances memory expected
to be of future relevance. Journal of Neuroscience, 31(5),
1563–1569. https://doi.org/10.1523/JNEUROSCI.3575-10.2011,
PubMed: 21289163
Wilhelm, I., Prehn-Kristensen, A., & Born, J. (2012). Sleep-depen-
dent memory consolidation - What can be learnt from children?
Neurobiology of Language
212
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
2
1
8
0
1
9
8
9
3
2
6
n
o
_
a
_
0
0
0
6
0
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Consolidation in language and motor learning
Neuroscience and Biobehavioral Reviews, 36(7), 1718–1728.
https://doi.org/10.1016/j.neubiorev.2012.03.002, PubMed:
22430027
Wilhelm, I., Rose, M., Imhof, K. I., Rasch, B., Büchel, C., & Born, J.
(2013). The sleeping child outplays the adult’s capacity to
convert implicit into explicit knowledge. Nature Neuroscience,
16(4), 391–393. https://doi.org/10.1038/nn.3343, PubMed:
23434910
Wilson, J. K., Baran, B., Pace-Schott, E. F., Ivry, R. B., & Spencer,
R. M. C. (2012). Sleep modulates word-pair learning but not
motor sequence learning in healthy older adults. Neurobiology
o f A gi n g , 3 3( 5) , 99 1 –1 00 0 . h t t p s : / / d o i . o rg/ 1 0 . 1 0 1 6 / j
.neurobiolaging.2011.06.029, PubMed: 22244090
Witt, K., Margraf, N., Bieber, C., Born, J., & Deuschl, G. (2010).
Sleep consolidates the effector-independent representation of a
motor skill. Neuroscience, 171(1), 227–234. https://doi.org/10
.1016/j.neuroscience.2010.07.062, PubMed: 20691764
Wymbs, N. F., & Grafton, S. T. (2013). Contributions from the left
PMd and the SMA during sequence retrieval as determined by
depth of training. Experimental Brain Research, 224(1), 49–58.
https://doi.org/10.1007/s00221-012-3287-1, PubMed:
23283418
Zoomer, J., Peder, R., Rubin, A., & Lavie, P. (1985). Mini Sleep
Questionnaire for screening large populations for EDS com-
plaints. In W. P. Koella, E. Ruther, & H. Schulz (Eds.), Sleep ’84
(pp. 467–470). Gustav Fisher.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
2
1
8
0
1
9
8
9
3
2
6
n
o
_
a
_
0
0
0
6
0
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neurobiology of Language
213