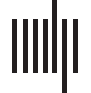RESEARCH ARTICLE
Early Development of Neural Speech Encoding
Depends on Age but Not Native Language Status:
Evidence From Lexical Tone
, Ching Man Lai1
, Akshay R. Maggu1,2
Nikolay Novitskiy1
Peggy H. Y. Chan1,3
, Hugh Simon Lam3
, Kay H. Y. Wong1
Tak Yeung Leung4, Ting Fan Leung3, and Patrick C. M. Wong1
,
,
1Department of Linguistics and Modern Languages, Brain and Mind Institute, The Chinese University of Hong Kong,
Hong Kong SAR, China
2O-lab, Duke Psychology and Neuroscience, Duke University, Durham, NC, USA
3Department of Paediatrics, The Chinese University of Hong Kong, Hong Kong SAR, China
4Department of Obsterics and Gynaecology, The Chinese University of Hong Kong, Hong Kong SAR, China
Keywords: infants, tone language, electroencephalography, frequency-following response, perceptual
narrowing
ABSTRACT
We investigated the development of early-latency and long-latency brain responses to native
and non-native speech to shed light on the neurophysiological underpinnings of perceptual
narrowing and early language development. Specifically, we postulated a two-level process to
explain the decrease in sensitivity to non-native phonemes toward the end of infancy. Neurons
at the earlier stages of the ascending auditory pathway mature rapidly during infancy
facilitating the encoding of both native and non-native sounds. This growth enables neurons at
the later stages of the auditory pathway to assign phonological status to speech according to
the infant’s native language environment. To test this hypothesis, we collected early-latency
and long-latency neural responses to native and non-native lexical tones from 85 Cantonese-
learning children aged between 23 days and 24 months, 16 days. As expected, a broad
range of presumably subcortical early-latency neural encoding measures grew rapidly and
substantially during the first two years for both native and non-native tones. By contrast,
long-latency cortical electrophysiological changes occurred on a much slower scale and
showed sensitivity to nativeness at around six months. Our study provided a comprehensive
understanding of early language development by revealing the complementary roles of earlier
and later stages of speech processing in the developing brain.
INTRODUCTION
During early infancy, infants with normal hearing are able to perceive a large inventory of
speech sounds that are both within and outside of their native language environments.
Toward the end of infancy, this broad perceptual ability is gradually reduced to a smaller
inventory of sounds that are mostly confined to the infants’ native language environments
(Werker & Tees, 1984). The term perceptual narrowing or attunement has been used to de-
scribe this phenomenon, and it is generally viewed as the basis of early language acquisition
a n o p e n a c c e s s
j o u r n a l
Citation: Novitskiy, N., Maggu, A. R.,
Lai, C. M., Chan, P. H. Y., Wong, K. H. Y.,
Lam, H. S., Leung, T. Y., Leung, T. F., &
Wong, P. C. M. (2022). Early
development of neural speech
encoding depends on age but not
native language status: Evidence from
lexical tone. Neurobiology of
Language, 3(1), 67–86. https://doi.org
/10.1162/nol_a_00049
DOI:
https://doi.org/10.1162/nol_a_00049
Supporting Information:
https://doi.org/10.1162/nol_a_00049
Received: 21 October 2020
Accepted: 22 June 2021
Competing Interests: The authors
declare competing interests.
Corresponding Author:
Patrick Wong
p.wong@cuhk.edu.hk
Handling Editor:
Janet F. Werker
Copyright: © 2021 Massachusetts
Institute of Technology Published
under a Creative Commons Attribution
4.0 International (CC BY 4.0) license
The MIT Press
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
6
7
1
9
8
6
8
5
2
n
o
_
a
_
0
0
0
4
9
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Early development of neural speech does not depend on native language
(Kuhl, 2014; Werker et al., 2012). This perceptual narrowing has a neurophysiological basis.
Kuhl postulated that exposure to a particular language results in the development of dedi-
cated (committed) neural networks which code the patterns of the language in question, and
also make the learning of other languages more difficult (Kuhl, 2004). This native language
neural commitment (NLNC) hypothesis has been influential in shaping our current under-
standing of how the nervous system supports perceptual narrowing and early language
acquisition.
While the NLNC is able to explain a variety of behavioral and neurophysiological (in
particular, cortical) data, its current rendition does not account for two important facts
about speech sound processing in the developing nervous system. First, although infants
in the first six months of life are able to perceive a large inventory of sounds, they are
unable to perceive all sounds. In fact, it seems to be the case that infants, regardless of
their language environment, have difficulty in discriminating speech sounds that are less
acoustically salient (Best et al., 1995, 2016). In order for infants to distinguish whether
or not a speech sound belongs to its native language and to use it contrastively in lexical
contexts, they must first encode and recognize it. Second, speech sound development (and
language acquisition) depends on a nervous system that is also developing, regardless of the
specific language input, and this helps to determine how well infants can encode speech.
Therefore, a comprehensive theory about early language acquisition must be situated in the
context of neural development as well as language experience. The aim of the present
study is to examine the development of speech perception in the context of both cortical
and subcortical neural development during the first two years of life. Our main question is
whether perceptual attunement can be explained by developmental changes in neural
functions.
The auditory neural system that supports hearing and spoken language consists of both sub-
cortical (including the auditory brain stem) and cortical structures that develop at different
rates. In terms of anatomical growth, the connections in the auditory brain stem acquire their
mature pattern by the 29th week of gestation and their myelination is completed within the
first postnatal year (Hartley et al., 2010; Moore & Linthicum, 2001, 2007; Moore et al., 1995,
1997; Romand & Romand, 1982; Romand et al., 1976; Sano et al., 2007). The 6th postnatal
month sees the maturation of the brain stem dendritic trees (Moore et al., 1998). The
subcortical-cortical connections, on the other hand, only mature when children are between
3 and 5 years old, and the myelination of cortical layers continues up to the age of 11 or 12
(Moore & Linthicum, 2007).
In terms of functional growth, research on the human auditory neural systems has tradition-
ally relied on scalp-recorded electroencephalography (EEG). Cortical and subcortical EEG
components can traditionally be separated by their timing and frequency composition.
High-frequency early-latency frequency-following response (FFR) is phase-locked to the spec-
tral envelope of an auditory stimulus (Moushegian et al., 1973). Although much more research
is needed to understand the sources of the FFR at different frequency ranges (Coffey et al.,
2019), the available evidence has suggested that the FFR has stronger (though not exclusive)
subcortical generators, including the inferior colliculi, especially for frequencies above 150 Hz
(Bidelman, 2018; Tichko & Skoe, 2017). The field seems to be more certain that the auditory
long-latency response (LLR) is generated exclusively in the cortex, within superior temporal
gyri in the vicinity of the Heschl’s gyrus (Godey et al., 2001) where intracranial recordings
have shown the presence of phoneme categorical representations (Chang et al., 2010;
Mesgarani et al., 2014). The adult LLRs in the time range between 50 and 240 ms are sensitive
to the nativeness of phonemes in both active discrimination and passive listening (Gansonre
Frequency-following response (FFR):
Scalp recordings of human phase-
locked neural activity that are
synchronized to periodic and
transient aspects of sound.
Neurobiology of Language
68
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
6
7
1
9
8
6
8
5
2
n
o
_
a
_
0
0
0
4
9
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Early development of neural speech does not depend on native language
et al., 2018; Kuuluvainen et al., 2014; Näätänen et al., 1997; Ross & Tremblay, 2009; Shtyrov
et al., 2000; Tremblay & Kraus, 2002; Tremblay et al., 1998, 2001, 2014).
The FFRs to speech are clearly detectable at birth (Ribas-Prats et al., 2019). The time and
frequency-domain parameters of FFR to consonants and vowels improve steadily from birth to
11 years (Anderson et al., 2012, 2015; Skoe et al., 2015). Neonates display a much less ma-
tured FFR to lexical tones, pitch patterns that are used to contrast word meaning, than do
adults (Jeng et al., 2011, 2016). Although not much is known about the development of
FFR to lexical tones in the first few years of life beyond the neonatal period, quite a few re-
search studies have examined FFR to speech more generally in adulthood. Lifelong experience
of lexical tones results in enhanced FFR encoding to lexical tones in tone language speakers
relative to non-tone language speakers (Krishnan et al., 2005, 2009). Experience with other
tone languages, e.g., Thai, may benefit the neural representation of lexical tones of another
tone language (e.g., Mandarin; Krishnan et al., 2010). With short-term training, FFR to lexical
tones can be improved in native speakers of a non-tone language (Reetzke et al., 2018; Song
et al., 2008). Experience-dependent FFR responses may be partially mediated by cortico-fugal
projections to the auditory brain stem (Intartaglia et al., 2016; Krishnan et al., 2005; Suga,
2008; Suga et al., 2000; Zhao & Kuhl, 2018) but may also manifest an online adaptation to
ongoing statistics in the brain stem (Alho et al., 2019; Escera, 2017; Slabu et al., 2012). These
results clearly demonstrate that the FFRs may be a useful tool for studying the neural devel-
opment of speech. As far as we are aware, studies have yet to investigate the early develop-
ment of FFR to native and non-native sounds during the period of rapid speech development.
Relative to FFR, a larger body of research has examined the LLR in relation to early
speech development. LLR at term birth is dominated by a wide and late positive deflection
peaking in the 200–300 ms interval (Barnet & Lodge, 1967; Barnet et al., 1975; Kushnerenko
et al., 2002; Novak et al., 1989; Wunderlich & Cone-Wesson, 2006). Later in life the orig-
inal positivity splits into P1 and P2, the trough between the positive peaks deepens and
eventually by 12–16 years old, it becomes the N1 wave of the adult auditory event-related
potential (ERP) (Cone & Whitaker, 2013; Lippé et al., 2009; Novak et al., 1989; Ponton
et al., 2002; Shafer et al., 2015; Small et al., 2018). (Throughout this article, we use the term
P1 for the largest infant positivity and consider it the homologue of the whole P1-N1-P2
complex in the adult LLRs). More specific to speech, Kuhl and colleagues (Kuhl, 2010) con-
ducted a series of studies to examine the cortical development of native and non-native
speech perception. They employed mismatch negativity (MMN), a pre-attentive component
of the LLR, that serves as an electrophysiological index of sound discrimination (Cheour-
Luhtanen et al., 1995; Partanen et al., 2013). The MMN to various vowel contrasts is present
in newborns (Cheour-Luhtanen et al., 1995), but it reduces in response to non-native vowel
contrast after 6 months of age (Cheour et al., 1998; Ortiz-Mantilla et al., 2013). Neural com-
mitment to the native language at the age of 7.5 months can predict language development
up to two years following initial neural measurements (Kuhl et al., 2008; Rivera-Gaxiola,
Klarman, et al., 2005; Rivera-Gaxiola, Silva-Pereyra, & Kuhl, 2005). A recent study of acous-
tic change complex in the infant LLR demonstrated a transformation from a merely
acoustical-driven pattern to native phoneme-category sensitivity around the age of 6 months
(McCarthy et al., 2019). Though the findings from these pioneering LLR-based (mostly
MMN) studies provide an initial neural explanation for perceptual narrowing, these studies
are focused largely on the role of the cortex. In order to obtain a more comprehensive un-
derstanding of the development of speech perception in the context of neural development,
further research is needed that examines both cortical and subcortical levels of the auditory
nervous system.
Neurobiology of Language
69
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
6
7
1
9
8
6
8
5
2
n
o
_
a
_
0
0
0
4
9
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Early development of neural speech does not depend on native language
The Current Study
In the present study, we examined the functional development of neural responses with gen-
erators in the brain stem and cortex in relation to native and non-native speech sounds. As the
anatomical and neurophysiological development of the brain stem generally occurs earlier
than that of the cortex, we postulate that the function of the brain stem in the first years of life
is solely to increase the encoding accuracy of the acoustic input, regardless of its nativeness
status. Thus, we predict a beneficial effect of age on the early-latency FFR with no interaction
between age and nativeness status during the first two years of life. An increase in encoding
fidelity would then allow the cortex to differentiate between native and non-native sounds,
enabling greater neural resources to be devoted to native sounds to facilitate infants’ phonol-
ogical and lexical development. By contrast, long-latency cortical LLR to speech develop more
slowly, and the responses show sensitivity to the nativeness status of the speech stimulus. We
expect that the already known cortical sensitivity to the nativeness status of a language extends
also to the P1 component of the infant LLR as they reflect interacting cortical processes.
Early-latency neural encoding FFR and long-latency cortical LLR recordings were simulta-
neously collected via EEG (Figure 1) from 85 Cantonese-learning children as they heard two
native (Cantonese) and one non-native (Mandarin) speech stimuli that were different only in
their pitch (Supplementary Figure 1; Supporting Information (SI) can be found at https://doi.org
/10.1162/nol_a_00049). Both Cantonese and Mandarin are tone languages. Lexical tones are
ideal for testing our hypothesis because phase-locking to fundamental frequencies that repre-
sent lexical tones can be measured by FFR. Children’s EEG responses were collected longitu-
dinally at up to 4 time points (Supplementary Figure 2). Since not every child had a recording
at every time point, the final data set represented a mixture of longitudinal trajectories and
cross-sectional data points. We employed mixed-effects models that took account of repeated
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
6
7
1
9
8
6
8
5
2
n
o
_
a
_
0
0
0
4
9
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 1. The waveforms of the early-latency and long-latency responses differ in their timing, temporal structures, and maturational patterns.
The left panel shows the frequency-following response generated predominantly in the brain stem. The right panel shows the waveforms of the
long-latency responses generated in the cortex. Note the difference in time and amplitude scales between the two panels. The rows corre-
spond to the age groups younger and older than 6 months that are also coded with darker and lighter shades of grey correspondingly. The
three lines represent each of the three lexical tones: Native /ga2/ (solid), Native /ga4/ (dash-dotted), and Non-Native /ga3/ (dotted).
Neurobiology of Language
70
Early development of neural speech does not depend on native language
measures when the data were available within the same subjects. This design is similar to other
studies of neural development (Raznahan et al., 2014; Wierenga et al., 2014).
We expected measures of FFR to improve linearly and substantially across age during the
early stages of development (23 days to 24; 16 months), as predicted by the literature on brain-
stem development (Moore et al., 1998). Importantly, improvement in these FFR measures can
be observed in both native and non-native tones. By contrast, LLR measures are likely to improve
more slowly than those of early-latency neural encoding measures within the same develop-
mental period because the auditory cortex takes longer to develop (Moore & Linthicum,
2007). As in previous studies of speech development in the cortex, we expected to see sensitivity
to the nativeness status of speech sounds occurring at about 6 months (Ortiz-Mantilla et al.,
2013), because 6 months is when cortical responses to lexical tones are found to be adultlike
(Cheng et al., 2013). As lexical tone is a type of speech sound that occurs in every syllable in a
tone language and about 70% of the languages in the world are tone languages (Fromkin, 2014;
Yip, 2002), we believe an investigation into the cortical-subcortical development of speech
using lexical tone as a starting point is a reasonable one. Our results can lay the foundation
for future research that examines other categories of speech sounds.
MATERIALS AND METHODS
Participants
Eighty-five Cantonese-learning infants and toddlers (48 males) between 23 days and 24;
16 months of age (mean age 7; 5 months, SD 5; 9 months), with no significant medical history,
participated in this study. Most of the participants were recorded longitudinally with up to 4 visits
5; 7 months ± 7.2 days apart on an average, producing 177 data points (Supplementary Figure 2).
By self report, parents were native Cantonese speakers and spoke Cantonese at home. Note that
although English and Mandarin (Putonghua) are languages spoken by residents of and visitors to
Hong Kong and are taught in the public schools, the vast majority of the population speaks
Cantonese (88.9%) as their usual language, with English (4.3%) and Mandarin (1.9%) being
the clear minorities (Government of Hong Kong SAR, 2016).
Written informed consent was obtained from parents or guardians prior to their child’s par-
ticipation in the study. The experimental protocol was in compliance with the Declaration of
Helsinki.
Stimuli and Experimental Design
The lexical tones were Cantonese (Native) tones 2 and 4 (rising and falling) and Mandarin
(Non-Native) tone 3 (dipping) embedded in the syllable /ga/. We chose the speech stimuli
based on two principles: (1) The native tone categories must be distinctive; tone 2 and tone
4 being of different contour shapes fulfilled this requirement. (2) The native and non-native
tone categories must also be distinctive. Mandarin tone 3 is quite unique and does not overlap
with any Cantonese tones. Furthermore, these same speech stimuli were used in our previous
study (Maggu et al., 2018), attesting to their validity for obtaining robust FFR responses.
The stimuli were spoken by a phonetically-trained female speaker, and were duration and
amplitude normalized. The fundamental frequency (F0) ranges for the Native /ga2/, Native
/ga4/, and Non-Native /ga3/ were respectively 182–278 Hz, 152–187 Hz, and 142–177 Hz
(Supplementary Figure 1). These three tones are described as 25, 21, and 214, respectively,
according to Chao’s nomenclature (Chao, 1930). Those frequencies are well above the fre-
quency range for cortical generators to be the dominant sources (Bidelman et al., 2018).
Neurobiology of Language
71
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
6
7
1
9
8
6
8
5
2
n
o
_
a
_
0
0
0
4
9
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Early development of neural speech does not depend on native language
Non-native tone was acoustically more complex than the other tones (see SI). This acoustic
complexity was not a confound because our results suggested that the Native /ga2/ tone posed
the most difficulty for the listeners (see SI). The stimuli were presented to the participants via
Audio CPT module of STIM2 (Compumedics, Australia). One thousand repetitions of the tones
were delivered in separate blocks to both ears of the participant with fixed interstimulus inter-
val 500 ms (stimulus-onset-asynchrony 674 ms) and alternating polarity.
EEG Recording and Analysis
Continuous EEG was collected using a Synamps RT amplifier connected to a Curry 7.05 work-
station (Compumedics, Australia) from five Ag/AgCl electrodes at Cz, M1 (left mastoid) and
M2 (right mastoid) at a sampling rate of 20 kHz with CPz as reference and Fpz as ground.
Interelectrode impedances were maintained at less than or equal to 1 kΩ. Cz data were re-
referenced offline with the average of two mastoids. The subsequent analysis was performed
on Cz. All data analysis was performed in MATLAB 9.2 (R2017a; mathworks.com), with
custom-made scripts using built-in functions of EEGlab (Delorme & Makeig, 2004), ERPLAB
(Lopez-Calderon & Luck, 2014), Signal Processing Toolbox (mathworks.com), and Statistics
and Machine Learning Toolbox (mathworks.com). Two different analysis pipelines were used
for FFR and LLR extraction correspondingly (see below). For each of the early-latency neural
encoding FFR and long-latency cortical LLR responses, we examined a number of conventional
metrics to investigate their developmental trajectories. An important consideration was whether
the native and non-native tones differ in their developmental trajectories of the auditory early-
latency neural encoding and the long-latency cortical responses.
Our recording procedures allowed us to simultaneously collect early-latency FFR and long-
latency LLR with the same set of stimuli in the same child. As far as we are aware, this is the
first study to simultaneously record FFR and LLR in infants; only adult studies of the kind have
been performed to date (Alho et al., 2019; Shiga et al., 2015). For each of the early-latency FFR
and long-latency cortical LLR measures, we examined a number of conventional metrics
(Anderson et al., 2012, 2015; Jeng et al., 2011, 2013, 2016; Skoe et al., 2015) to investigate
their developmental trajectories. The aim was to trace all the subjects longitudinally. However,
the main challenge of a longitudinal design was subject attrition, which turned this study into a
mixed cross-sectional and longitudinal design. Additionally, the EEG recording in infants is
prone to movement artifacts and not every recording session is successful. One option would
be keeping only the subjects with a certain amount of longitudinal data points. The size of our
sample would allow us such a manipulation. However that would not only reduce statistical
power but also render the sample unrepresentative in relation to the original population. We
opted to keep all subjects who had at least one successfully recorded EEG session. To take
advantage of the fact that repeated measurements were taken from some participants, we em-
ployed mixed-effects models with subject as a factor. Our primary focus was to understand
whether the growth of FFR measures was rapid, and whether this growth could be observed
in both native and non-native tones. Our secondary focus was to understand whether the
growth of the LLR measures appeared to be less substantial, and whether sensitivity to native-
ness could be seen at around 6 months of age.
FFR Analysis
For FFR extraction, the data were filtered with an 8-order Butterworth 80–1500 Hz bandpass
filter and down-sampled to 3 kHz. The epochs were extracted around the stimulus onset time
Neurobiology of Language
72
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
6
7
1
9
8
6
8
5
2
n
o
_
a
_
0
0
0
4
9
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Early development of neural speech does not depend on native language
from −50 to 225 ms using a ±25 μV artifact rejection criterion (Skoe & Kraus, 2010). The EEG
recording blocks with more than 10% of the rejected sweeps per block (i.e., >100 rejections)
were not included in further analyses (Maggu et al., 2016, 2018). Thus, the minimum number
of trials per block was 900. One block comprised the EEG responses to one tone-type for a
particular subject at a particular age. The averaged FFR with more than 0.7 μV maximum were
excluded for suspicion of stimulus artifact based on preliminary data screening. These proce-
dures resulted in exclusion of 18% of the EEG blocks, including the whole set of data from
one participant. In addition, 13% Native /ga2/ blocks, 7% Native /ga4/ blocks, and 9% of the
Non-Native /ga4/ blocks were missing in the final data set. The final number of trials per
block was 988.8 ± 1.5 for Native /ga2/, 990.0 ± 5.17 for Native /ga4/, and 984.8 ± 2.0 for
Non-Native /ga3/. Neither the difference between the tones for the number of accepted trials
nor the correlation between the number of trials with age was significant (Native /ga2/ vs. Native
/ga4/: t(316) = −0.243, p = 0.808; Native /ga2/ vs. Non-Native /ga3/: t(313) = 1.597, p = 0.111;
Native /ga4/ vs. Non-Native /ga3/: t(323) = 0.981, p = 0.327; Native /ga2/ vs. age: r = 0.15,
p = 0.063; Native /ga4/ vs. age: r = 0.097, p = 0.215; Non-Native /ga3/ vs. age: r = 0.054,
p = 0.5).
FFR was calculated as a time-domain average of all good epochs with baseline correction
across the pre-stimulus interval (−50 to 0 ms).
FFR pitch contour was extracted with a sliding time window applied at the time-frequency
decomposition of intertrial phase coherence (a.k.a., phase-locking factor, Tallon-Baudry et al.,
1996). In comparing FFR pitch contour with auditory stimulus pitch contour, the latter was
shifted 20 ms toward the end of the time course for the pitch tracking accuracy calculation
(see Jeng et al., 2013 and SI for details).
A number of time-domain and frequency-domain metrics were extracted from FFR. For the
main report on FFR here, we selected the four measures most commonly employed in FFR
developmental studies: lower-frequency and middle-frequency spectral band-power
(Anderson et al., 2012, 2015; Skoe et al., 2015), pitch strength (Jeng et al., 2011, 2013,
2016) and signal-to-noise ratio (SNR). Kolmogorov-Smirnov tests confirmed the normality of
the four measures in question (see SI). The FFR SNR was calculated as a dB-transformed ratio
between root mean square power of post-stimulus and pre-stimulus intervals of FFR (see SI for
the formula). Whole-epoch autocorrelation was calculated on FFR waveforms. Pitch strength
was calculated as peak-to-peak distance on an autocorrelation curve between the first auto-
correlation peak and the preceding trough. Lower and middle-frequency spectral band-powers
were obtained by calculating the fast Fourier transform of FFR waveforms and extracting the
power around the F0 and its two lowest overtones (2D and 3D harmonics). See SI and
Supplementary Figure 3 for the description of the other metrics.
The aforementioned results focused mostly on the encoding of individual tones. One
important requirement of phonological development is to differentiate distinct speech sound
categories. We therefore examined classification of the three tones with a support vector
machine (SVM) for each month of age that delivered more than 10 observations, i.e., from
birth to 14 months. Averaged FFRs were down-sampled to 120 Hz in order to reduce the
number of features in the models. The interval from 0 to 150 ms was extracted, resulting in
13 time points/features per observation. The data at each time point were normalized across
observations. The data of each age were bootstrapped 10,000 times. In each bootstrap sam-
ple, we ran SVM classification with cross-validated accuracy as the outcome. The procedure
was performed for both real tone labels and randomly permuted labels labels. (See SI
for the
detailed SVM parameters.)
Neurobiology of Language
73
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
6
7
1
9
8
6
8
5
2
n
o
_
a
_
0
0
0
4
9
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Early development of neural speech does not depend on native language
LLR Analysis
For LLR extraction, the data were filtered with a 4-order Butterworth 0.1–30 Hz bandpass filter
and down-sampled to 3 kHz. The epochs were extracted around the stimulus latency time
from −100 to 600 ms using a ±100 μV artifact rejection criterion. All epochs with amplitude
in the artifact rejection criterion were considered good epochs. Since LLR is known to habit-
uate with the repetition of the same sound (Cortesa et al., 2019; Näätänen & Picton, 1987),
only 300 first good epochs for each participant/condition were used for the purposes of anal-
ysis. LLR was calculated as a time-domain average with baseline correction across the pre-
stimulus interval (−100 to 0 ms). SNR was calculated on an averaged LLR with a pre-stimulus
interval from −100 to 0 ms and a post-stimulus interval from 0 to 600 ms (see SI for the for-
mula). The LLR main positive peak (i.e., P1) was searched for automatically as a maximum
across the post-stimulus interval (0 to 600 ms; see SI for details). Similarly to the FFR analyses,
we employed linear mixed-effecs (LME) models with age and tone as fixed effects and partic-
ipant as a random intercept.
Previous research on the cortical development of lexical tones has demonstrated that
6 months of age is an important milestone for adultlike responses, as the polarity of MMN
switches from a positive to a negative (adultlike) response during this period (Cheng et al.,
2013). This finding converges with the studies of vowels and consonants that have shown a de-
crease in MMN responses to non-native sounds in infants older than 6 months of age (Cheour
et al., 1998; Rivera-Gaxiola, Silva-Pereyra, & Kuhl, 2005). We tested cortical development of
lexical tones by examining age-dependent effects categorically, using 6 months (165 days) as a
cutoff. We took a step further to analyze the effects of nativeness by categorizing our stimulus
into native (pooled Native /ga2/ and Native /ga4/) and non-native (Non-Native /ga3/) tones with
a focus on the latency and amplitude measures. Because P1 peak latency and amplitude did not
meet the normality assumption, we used non-parametric tests to investigate the hypothesis of
age-dependent sensitivity to speech nativeness.
FFR vs. LLR Comparison
In order to compare the magnitude of growth between the early-latency neural encoding FFR
and long-latency cortical LLR measures, we examined three measures that can be obtained in
both FFR and LLR, namely, SNR, amplitude, and latency. We examined a group of younger
(n = 22) and older (n = 17) children with an arbitrary age cutoff of younger than 42 days and
older than 400 days, as that would provide the maximum number of participants in each
age group. Extracting the effect size of age cutoff (i.e., the slope of the regression line between the
two age groups) on the normalized metrics enabled us to compare the maturation of the metrics
directly, regardless of individual means and variance, where maturation was indicated by larger
SNR, increased amplitude, and shorter latency.
RESULTS
Early-Latency Neural Encoding FFR Measures
The FFR consists of a series of transient peaks followed by a steady state response (Figure 1,
left panel). We first examined whether, on average, the infants’ FFRs showed high fidelity.
For each tone, we calculated the intertrial phase coherence extracted pitch contour, which
followed the pitch contour of the stimuli when the time lag equal to the averaged latency of
stimulus-response correlation (20 ms) was introduced. We found that the averaged pitch
contours were strongly correlated with the pitch contours of the speech stimuli, with r 2
Neurobiology of Language
74
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
6
7
1
9
8
6
8
5
2
n
o
_
a
_
0
0
0
4
9
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Early development of neural speech does not depend on native language
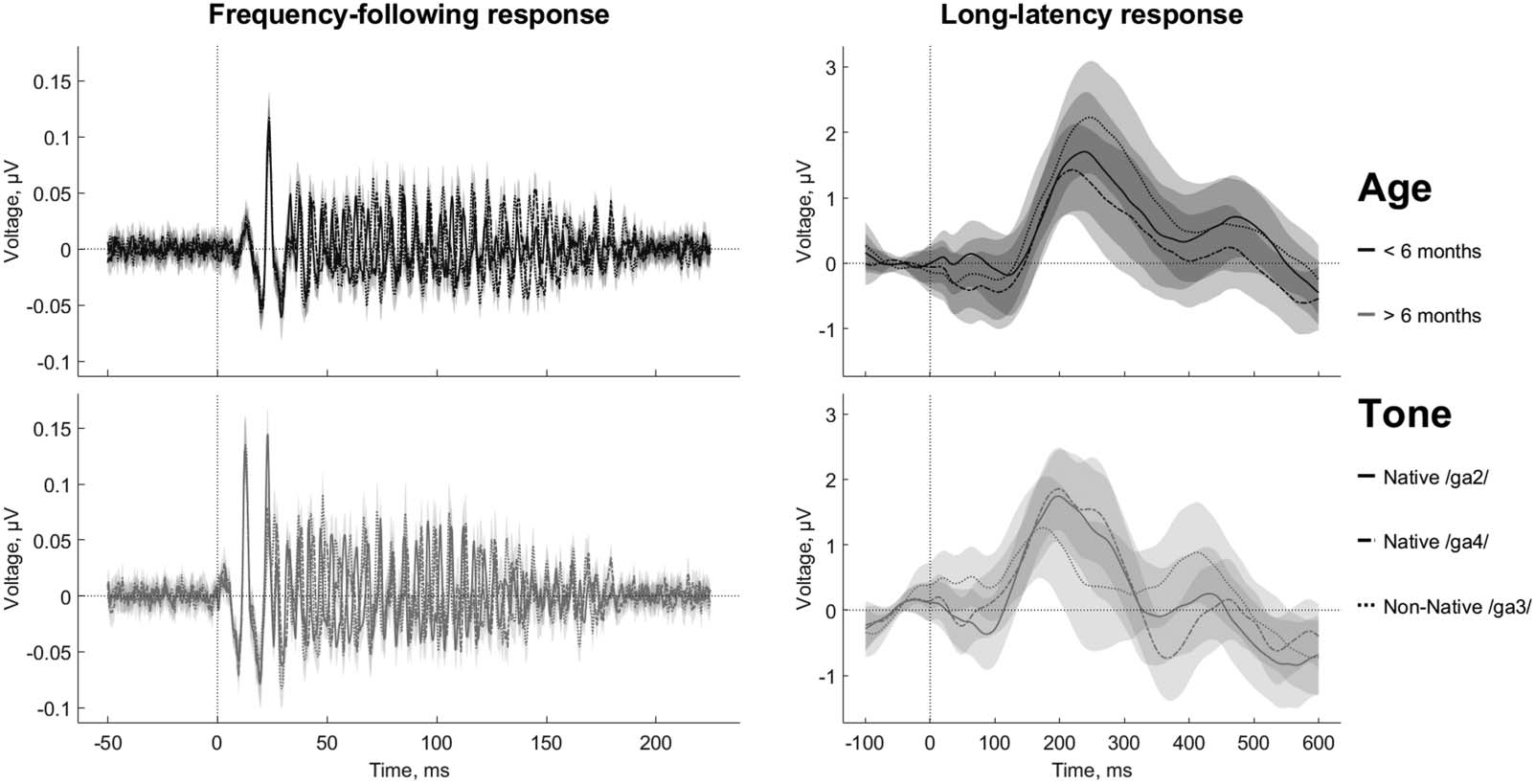
Figure 2. The pitch contour of the frequency-following response (FFR) follows the pitch contour of the auditory stimuli. The columns
correspond to the lexical tones Native /ga2/, Native /ga4/, and Non-Native /ga3/. Upper row: spectrogram of the auditory stimuli.
Middle row: the overlap of the auditory (black line) and early-latency brain-response (grey circles) pitch contours. The pitch tracking (audio
vs. brain signal correlation) is 0.91 for Native /ga2/, 0.97 for Native /ga4/, and 0.95 for Non-Native /ga3/. Bottom row: Grand-averaged
time-frequency decomposition of the intertrial phase coherence.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
6
7
1
9
8
6
8
5
2
n
o
_
a
_
0
0
0
4
9
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
values of 0.91 for Native /ga2/, 0.97 for Native /ga4/, and 0.95 for Non-Native tones (Figure 2).
This suggests that as a group, our infant participants were quite accurate in encoding the speech
stimuli.
LME analysis
F tests of the fixed effects within the mixed-effects model revealed a main effect of age for FFR
−5; pBf = 0.0002; η
2 = 0.04), lower-frequency spectral
SNR (F(1, 473) = 19.21; p = 1.44 · 10
p
band-power (F(1, 473) = 10.91; p = 0.0010; pBf = 0.0133; η
2 = 0.02), and middle-frequency
p
2 = 0.04). The main
spectral band-power (F(1, 473) = 17.48; p = 3.47 · 10
p
effect of age on pitch strength was marginally significant (F(1, 473) = 7.24; p = 0.0074; pBf =
0.0958; η
2 = 0.02). There was a marginal effect of tone for pitch strength only (F(2, 473) =
p
4.12; p = 0.0169; pBf = 0.2198; ηp
2 = 0.02), while it was not significant for FFR SNR,
lower-frequency spectral band-power and middle-frequency spectral band-power (all p >
0.05). Most importantly, none of the FFR measures revealed an interaction between age
and tone, including FFR SNR, lower-frequency spectral power, middle-frequency spectral
power, and pitch strength (all p > 0.05, Figure 3).
−5; pBf = 0.0005; η
Classification analysis
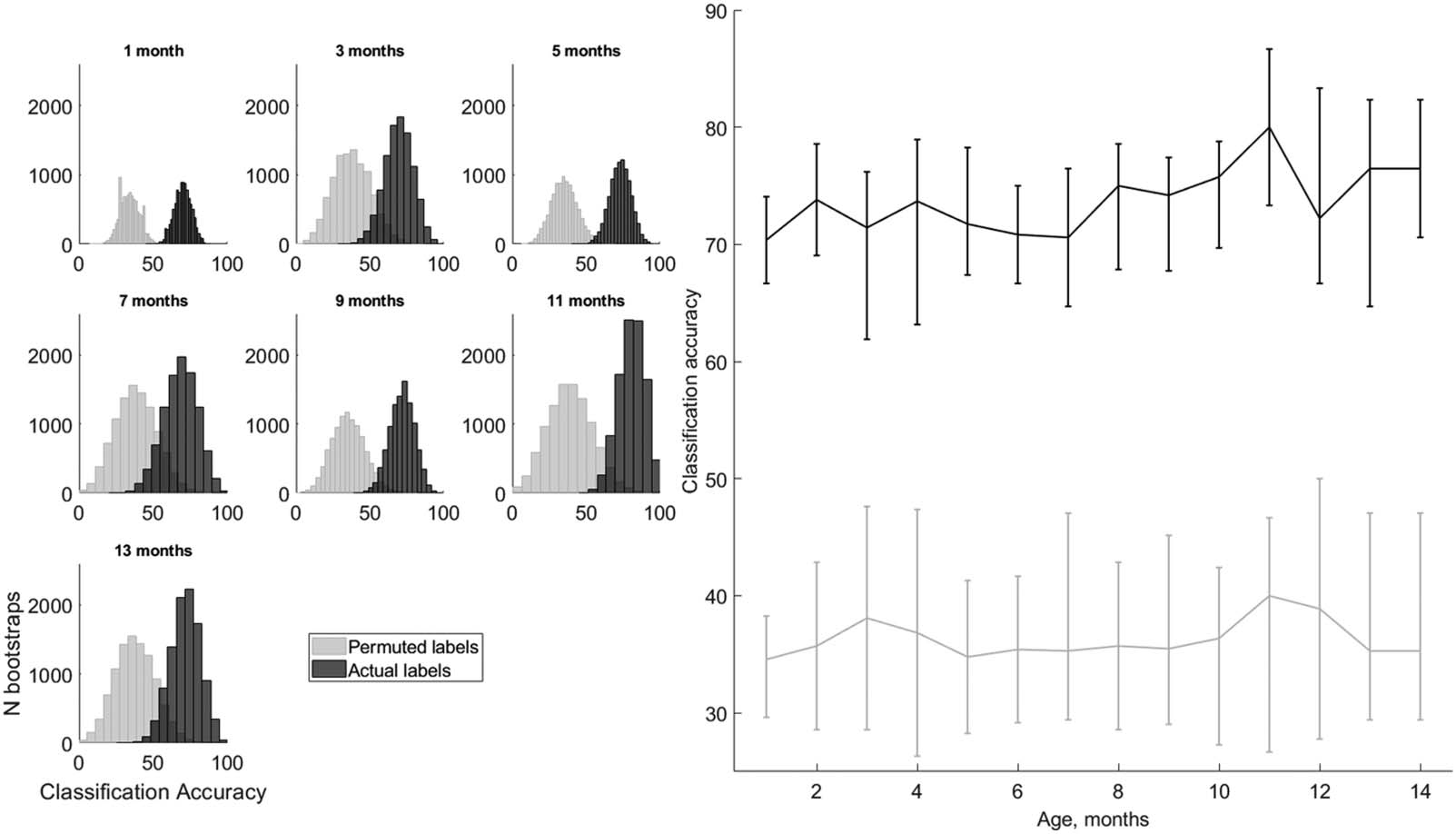
We found that SVM classification of the early-latency brain responses to the three tones im-
proved over time (Figure 4). The median classification of the tone was overall higher than the
permuted classification accuracy (mean 73.8 ± 0.75% and 36.3 ± 0.43%, t(13) = 53.48; p =
−16; d = 14.3). Importantly, classification accuracy improved with age as indicated by
1.25 · 10
the Pearson correlation (r = 0.64; p = 0.0136; pBfr = 0.0272; d = 1.67). Adding a quadratic
term did not improve the model, suggesting a linear dependency (F(1, 11) = 0.14; p = 0.71;
η
2 = 0.01). No age dependency was found for the permuted data classification (r = 0.24; p =
p
0.407; pBfr = 0.814; d = 0.49).
Neurobiology of Language
75
Early development of neural speech does not depend on native language
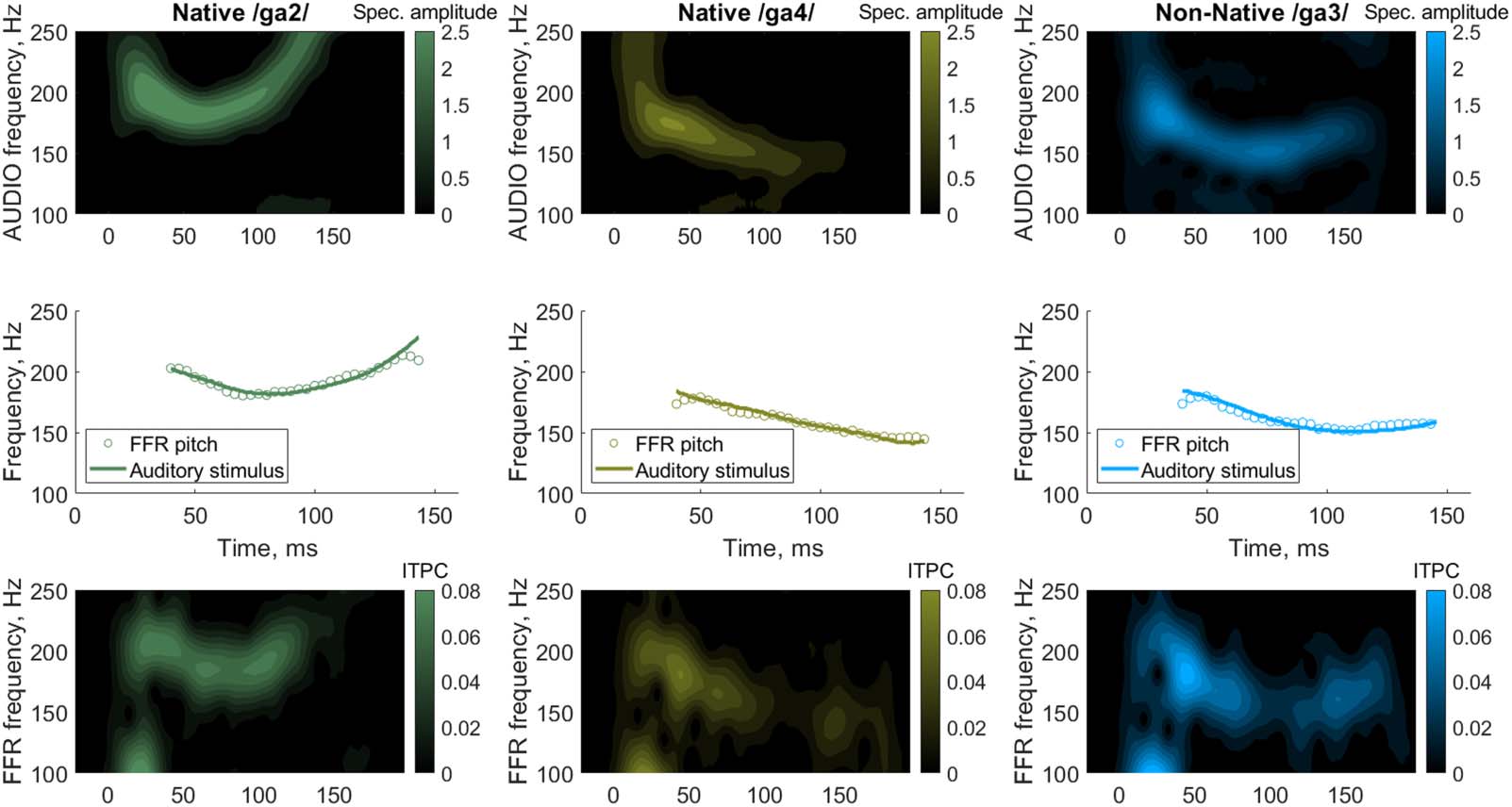
Figure 3. Developmental trajectories of early-latency brain measures show linear growth with age, regardless of speech nativeness. The top
panels show the root mean square (RMS) of the frequency-following response (FFR) (A); the FFR autocorrelogram (B); and the lower (120–260 Hz)
(C) and middle (260–750 Hz) (D) frequency bands of the FFR spectrum. The shaded area marks the standard deviation. The bottom panels plot
the output metrics as a function of age in days. The metrics are the signal-to-noise ratio (SNR) of the time-domain waveform (E); the pitch
strength (trough-to-peak magnitude of the autocorrelogram) (F); and the mean power in the lower (G) and middle (H) frequency bands.
Individual data points are plotted as grey dots with lines connecting the data of the same participant and the same tone. The three lines
represent the average age-dependent development for each of the three lexical tones Native /ga2/ (solid), Native /ga4/ (dash-dotted), and
Non-Native /ga3/ (dotted). The data are smoothed with Eilers’ technique (Eilers, 2003).
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
6
7
1
9
8
6
8
5
2
n
o
_
a
_
0
0
0
4
9
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. Tone classification with support vector machine improves with age for actual but not for permuted tone labels. The left panel shows
real (black) and permuted (grey) labels for the cross-validated classification accuracy distribution across 10,000 bootstrapped samples for
selected ages in 2-month steps from 1 to 13 months. The right panel shows the medians of those distributions as a continuous function of
age in months from 1 to 14 for real (black) and permuted (grey) labels. The error bars show the range between the first and third quartile of the
distributions. The real label classification correlates significantly with age (r = 0.64, p < 0.05, d = 1.67), while the permuted label classification
does not (r = 0.24, p > 0.1, d = 0.49).
Neurobiology of Language
76
Early development of neural speech does not depend on native language
Long-Latency Cortical LLR Measures
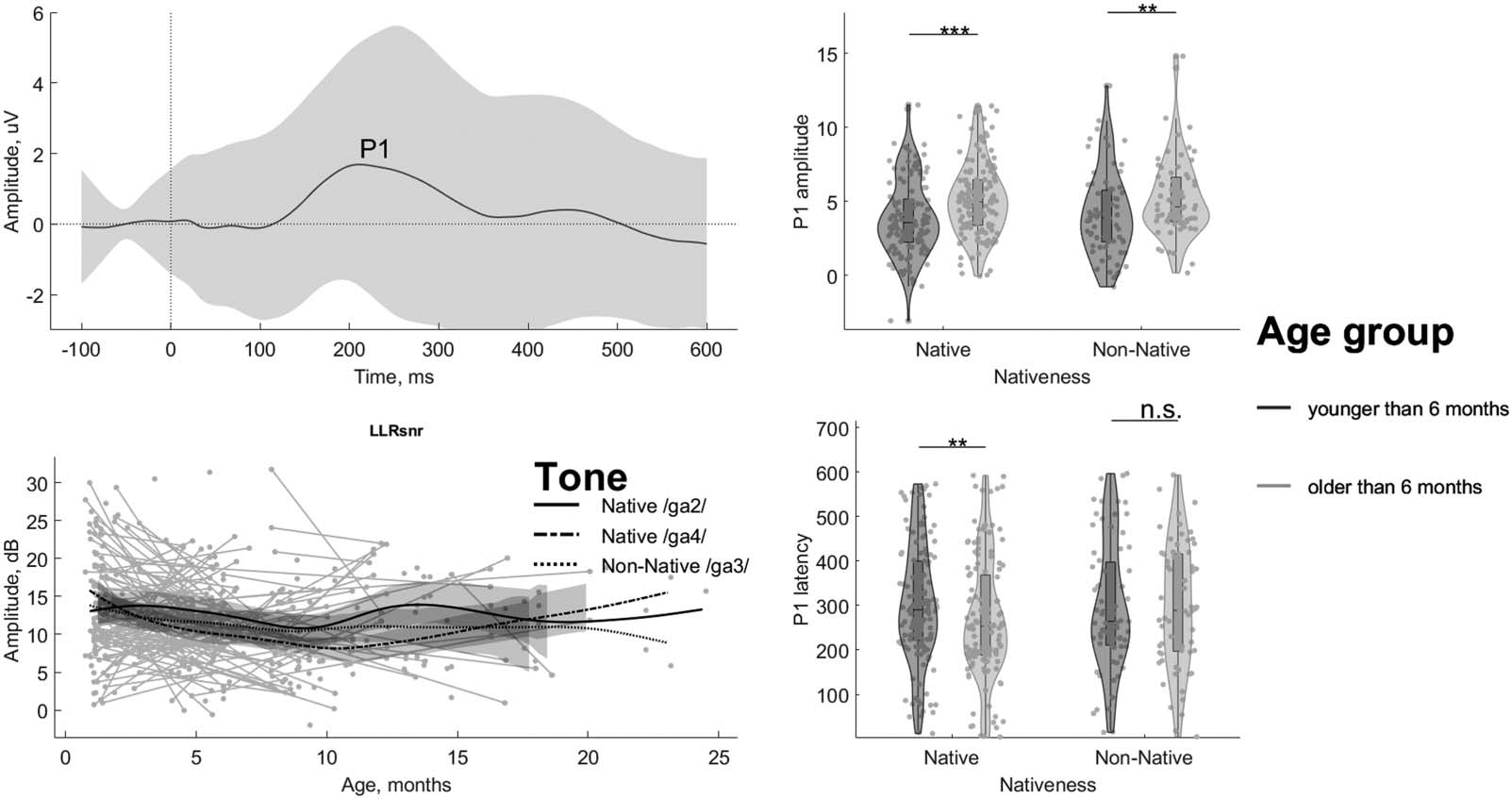
The LLR was dominated by a large and wide positive peak P1 (Figure 1, right panel). We mea-
sured the latency and amplitude of P1, as well as its SNR. As in the case of FFR, we investigated
the changes of LLR measures as a function of age (Figure 5). In the LME results, there was an
increase with age in P1 amplitude (F(1, 409) = 9.14; p = 0.0027; pBf = 0.0345; ηp
2 = 0.02), but
not in LLR SNR ( p > 0.05) or P1 latency ( p > 0.05). None of the LLR parameters were different
between tones, i.e., LLR SNR, P1 amplitude, or P1 latency (all p > 0.05). There was no inter-
action between age and tones for any of the LLR parameters either, i.e., LLR SNR, P1 amplitude,
and P1 latency (all p > 0.05). In general, the effect of age on LLR is much less robust compared
with the FFR results.
We then followed the findings of the literature concerning the cortical development of lex-
ical tones at 6 months of age (Cheng et al., 2013). For non-native speech, the age split resulted
in 63 participants (71 data points) and 51 participants (68 data points), respectively, in the
younger and older groups in the analysis. For native speech, 65 participants (135 data points
for pooled tones, 75 data points for averaged tones) and 61 participants (141 pooled tone data
points, 89 averaged tone data points) were in the younger and older groups, respectively. The
Wilcoxon rank-sum tests demonstrated a significant shortening of P1 latency and an increase
in its amplitude after the age of 6 months for the native tones (pooled Native /ga2/ and Native
−5, r = 0.24 for
/ga4/: Z = −2.45, p = 0.014, r = −0.15 for latency and Z = 4.04, p = 5.27 · 10
amplitude; averaged Native /ga2/ and Native /ga4/: Z = −2.41, p = 0.016, r = −0.19 for latency
−4, r = 0.30 for amplitude), while the change was relatively small for
and Z = 3.87, p = 1.07 · 10
the Non-Native /ga3/ tone (Z = 0.16, p = 0.56, r = 0.01 for latency and Z = 2.52, p = 0.006,
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
6
7
1
9
8
6
8
5
2
n
o
_
a
_
0
0
0
4
9
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 5. The long-latency response (LLR) signal-to-noise ratio (SNR) does not grow with age, while its peak latency reduces only for the
native tones. Upper left panel shows the LLR time-domain waveform averaged across all data points, with the shaded area marking the stan-
dard deviation. Peak P1 is labeled at maximum. Lower left panel shows LLR SNR as a function of age in days. Individual data points are plotted
as grey dots with lines connecting the data of the same participant and the same tone. The three lines represent the average age-dependent
development for each of the three lexical tones Native /ga2/ (solid), Native /ga4/ (dash-dotted), and Non-Native /ga3/ (dotted). The data are
smoothed with Eilers’ technique (Eilers, 2003). Upper and lower right panels show violin plots of P1 amplitude and latency distributions across
the 6-month cutoff. The tones are pooled into native (/ga2/ and /ga4/, dark-grey) and non-native (/ga3/, light-grey) groups. The grey circles
represent individual data and the diamonds mark the medians of the distribution. The significance of the age effect in the Wilcoxon rank-sum
test is illustrated with asterisks separately for native and non-native tones (**p < 0.01, ***p < 0.001, uncorrected, n.s. non-significant).
Neurobiology of Language
77
Early development of neural speech does not depend on native language
r = 0.21 for amplitude) (Figure 5, right panel). Overall, these results replicated earlier findings
that perceptual narrowing is supported by cortical processes (Conboy & Kuhl, 2011).
Specifically, our results here confirmed that sensitivity to the nativeness status of lexical tones
occurs at about 6 months of age, as suggested by a previous study (Cheng et al., 2013).
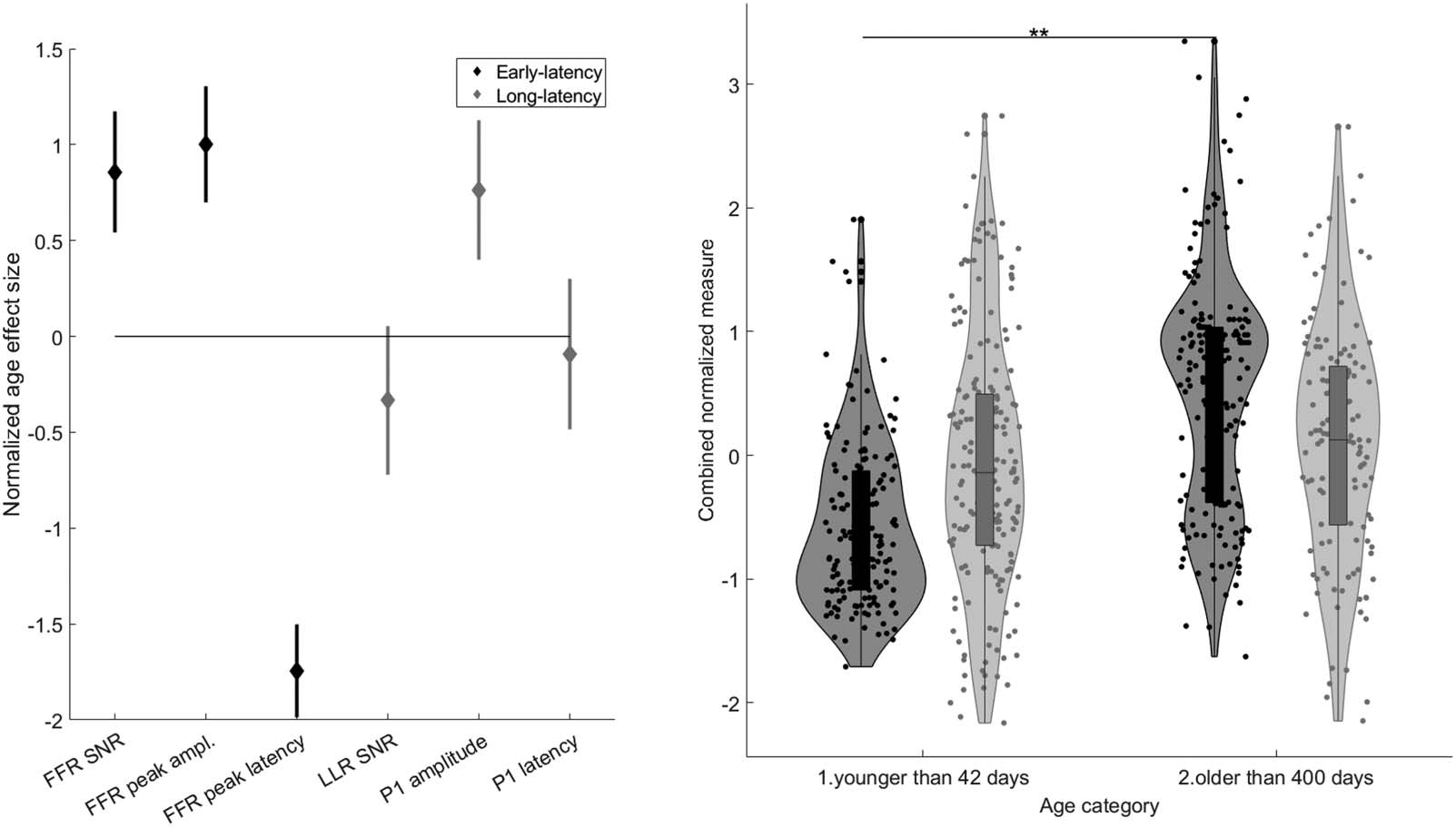
Early-Latency Neural Encoding FFR vs. Long-Latency Cortical LLR Measures
As shown in the left panel of Figure 6, the effect size of FFR SNR (0.86, 95% CI [0.54, 1.17])
was significantly larger than that of LLR SNR (−0.33, 95% CI [−0.72, 0.05]). The effect size of
FFR peak amplitude (1.00, 95% CI [0.70, 1.30]) was marginally larger than that of LLR P1
amplitude (0.76, 95% CI [0.40, 1.13]). Finally, the effect size of FFR peak latency was signif-
icantly larger (i.e., more negative, −1.74, 95% CI [−1.99, −1.50]) than that of LLR P1 latency
(−0.09, 95% CI [−0.48, 0.30]). In other words, two out of the three cortical measures showed
less change over time, and two out of the three measures showed a significantly larger age-
related improvement in the early-latency neural encoding responses.
In order to simultaneously compare the effects of age category on the multiple parameters
of early- and long-latency responses, we entered the aforementioned normalized metrics into a
combined LME analysis with Age category, Tone, ERP type (early-latency vs. long-latency re-
sponses), and ERP measure (SNR vs. latency vs. amplitude) as fixed effects and Subject as ran-
dom intercept. F tests of the fixed effects demonstrated the significant main effect of Age
category (F(1, 620) = 9.92, p = 0.0017; η
2 = 0.02) and the interaction between Age category
p
and ERP type (F(1, 620) = 5.62, p = 0.0174; η
2 = 0.01). None of the other main effects or
p
interactions was significant (see Supplementary Table 3). In the follow-up split-by-ERP-type
LMEs, the metrics of the early-latency (F(1, 317) = 10.6, p = 0.0013; η
2 = 0.03), but not of
p
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
3
1
6
7
1
9
8
6
8
5
2
n
o
_
a
_
0
0
0
4
9
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 6. The maturational effect is greater for early-latency (black) than long-latency (grey) response measures. Left panel: Both brain
subdivisions are represented by three compatible time-domain measures; signal-to-noise ratio (SNR), peak amplitude, and peak latency.
The diamonds represent effect sizes measured as the difference between the means of the youngest (younger than 42 days) and oldest (older
than 400 days), with participants’ data normalized for each measure. Lines cover the confidence intervals of those effect sizes in the linear
models. The confidence intervals for SNR and peak latency do not overlap between the frequency-following response (FFR) and the
long-latency response (LLR), indicating significant differences. Right Panel: The linear mixed effects analysis shows that a combination of
normalized measures grows with age for early-latency (black) but not for long-latency (grey) response measures (**p < 0.01).
Neurobiology of Language
78
Early development of neural speech does not depend on native language
the long-latency responses (F(1, 303) = 0.018, p = 0.8930; ηp
(right panel of Figure 6).
2 < 0.001), were improved by age
DISCUSSION
The goal of the present study was to investigate the contributions of early-latency neural en-
coding and long-latency cortical processing toward early language development in the first
two years of life. We postulated that a degree of encoding accuracy of speech sounds regard-
less of native status must occur in the early stages of life, before these speech sounds’ native
status could be determined. Consistent with this hypothesis, we found that the early-latency
neural encoding of speech grows linearly and markedly for both native and non-native tone
categories for children who are learning a tone language. In contrast, the growth in the long-
latency cortical responses is much slower, but crucially the change is sensitive to the native-
ness status, with changes occurring at about 6 months of age, as reported in previous studies
(Cheour et al., 1998; Cheng et al., 2013; Rivera-Gaxiola, Silva-Pereyra, & Kuhl, 2005). We
argue that our results reflect the complementary roles of neural structures that are associated
with earlier and later stages of processing along the ascending auditory neural pathway. For
the frequencies that we examined, these neural structures are likely located in the brain stem
and cortex for earlier and later stages of encoding and processing, respectively. While the
brain stem (and potentially other neural structures) encodes speech sounds faithfully in an ear-
lier stage of processing, the cortex assigns phonological information to them at a later stage.
These two processes together are foundational to early language development.
Behavioral studies of lexical tones suggest that perception of lexical tones emerges at or be-
fore 6 months of age for tone-learning infants (Mattock & Burnham, 2006; Yeung et al., 2013).
However, it is unclear how development of lexical tone perception is associated with early-
latency and long-latency neural responses. As far as we are aware, the present study is the first
to address this question. Our long-latency cortical findings are largely consistent with those of
the studies of vowel and consonant perception (Cheour et al., 1998; Rivera-Gaxiola, Silva-
Pereyra, & Kuhl, 2005) showing native and non-native differentiation in MMN responses after
6 months (and sometimes earlier). Although Cheng et al. (2013) did not investigate native and
non-native tones, their findings of an adultlike cortical MMN response to native Mandarin tones
in 6-month-old Mandarin-learning infants are generally consistent with the results in the present
study. The MMN paradigm favored by the majority of previous LLR studies amplifies the brain
responses to the phoneme categories by subtracting standards from deviants in oddball se-
quence (Escera, 2017; Näätänen & Winkler, 1999). However, in adults the sensitivity to pho-
nological contrasts can be found already in the so-called obligatory brain responses (Grimaldi,
2018). The amplitude of responses to native phonemic contrast was found to increase with age
in an equiprobable ERP paradigm (Key et al., 2012). We extended the existing MMN studies by
investigating the difference between the obligatory P1 response in infant LLR to stimuli in all-
standard blocks. The results converge with the MMN findings as we found a latency delay for
non-native tone that is likely to be caused by difficulty in categorization after attunement to
native phonology at around 6 months. It is clear that multiple cortical LLR components are re-
sponsive to the native status of speech sounds. We must acknowledge that the conclusions de-
rived from our cortical results were based on conducting less robust statistical analyses
separating the native and non-native data, rather than relying on findings from a significant
Age × Nativeness interaction. Notwithstanding that, the results converged with previous studies.
In terms of early-latency speech encoding processes, Jeng et al. (2013, 2016) were the first
to study FFRs in tone-learning infants. These two smaller-scale studies found that neonates
Neurobiology of Language
79
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
3
1
6
7
1
9
8
6
8
5
2
n
o
_
a
_
0
0
0
4
9
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Early development of neural speech does not depend on native language
from Mandarin-speaking families did not differ from neonates from English-speaking families
in their FFRs to Mandarin tones. This suggests the cross-language differences in FFRs to lex-
ical tones found in studies of adults (Krishnan et al., 2010) are most likely attributable to
language experience. It should be noted, however, that the developmental time course of
FFR to lexical tones and the potential within-subject differences between native and non-
native lexical tone encoding have not been investigated before. Two other studies specifi-
cally focused on the development of the FFR signal in early years and across the lifespan by
examining non-tone language-speaking listeners, but they did not examine lexical tones or
the developmental difference between native and non-native speech perception (Anderson
et al., 2015; Skoe et al., 2015).
Neurophysiological data suggest that the roles of the brain stem and cortex and the rela-
tionship between them may change considerably as the infant grows. Evolutionary older brain-
stem structures take shape earlier and mature faster than the neocortex (Moore & Linthicum,
2007). Postmortem fetal studies show active processes of axon growth, dendrite tree arboriza-
tion, and myelination in the brain stem, which start in the last prenatal month and are mostly
completed by the end of the first postnatal year (Hartley & King, 2010; Moore et al., 1995,
1997, 1998; Moore & Linthicum, 2007; Sano et al., 2007). It is the neocortex that empowers
humans with speech, but the development of cortical structure achieves its adult-like condi-
tion only by the age of 11–12 years (Moore & Guan, 2001). Indeed, the functioning of the
auditory cortex in the perinatal period is physiologically very different from its functioning
in adults in terms of the cortical layer domination (Moore & Linthicum, 2007). The immature
status of the cortex implies also the immature condition of the corticofugal projections to the
brain stem. Thus, the potential contribution of the corticofugal system to speech perception in
adults may be related to its maturation (Suga et al., 2000). On the other hand, recent studies
have clearly shown that the adult midbrain is not a mere relay and is capable of detecting
regularities in the auditory stream (Malmierca et al., 2019). This capability also needs time
to mature. Our data confirm previous descriptions using non-invasive EEG recordings. We
have followed several non-invasive measures of early-latency neural encoding that are likely
to reflect the spread of myelination and increase in synaptic activity in the brain stem and
structures connected to it. Moreover, we show that cortical responses grow more slowly
and that their maturation is also native-language specific. We anticipate that further studies
will combine functional methods such as EEG with non-invasive structural methods such as
diffusion MRI in order to obtain a better understanding of the neurophysiological basis of
speech acquisition.
Neural phase-locking to the stimulus envelope is the most accurate representation of the
physical properties of the stimulus. It requires accurate timing of neural firing, i.e., a strong
degree of myelination. The capability of neurons in phase-locking to the high frequencies
reduces along the auditory axis, and in the cortex the spatial coding of frequency (tonotopy)
is prevalent instead (Leaver & Rauschecker, 2016). The FFR may correspond to the phase-
locked activity of neurons of different neural centers across the ascending auditory pathway.
The empirical studies available to date seem to suggest that phase locking at higher frequencies
(>150 Hz) in the FFR most likely originates from the auditory brain stem (Bidelman, 2018;
Tichko & Skoe, 2017). The basic conceptualization proposed here is not restricted to the brain
stem or another particular neural structure per se, but rather suggests a division of labour of
different auditory neural centers during language development. It seems likely that some cen-
ters contribute to representing basic acoustic elements of speech via direct phase-locking to the
stimulus, while others assign function to those acoustic elements. They both need to be in place
in order for spoken language to develop. It is worth emphasizing that our present findings do not
Neurobiology of Language
80
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
6
7
1
9
8
6
8
5
2
n
o
_
a
_
0
0
0
4
9
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Early development of neural speech does not depend on native language
inform—nor do they depend on—our understanding of the precise neuroanatomical origins of
the early-latency neural speech encoding. Our results merely suggest that not all neural re-
sponses to speech are equal in their neurodevelopmental trajectories, and most importantly
the differences between temporally earlier FFR and later LLR are important for a child to suc-
ceed in acquiring native speech. Future research should examine the exact locations of the
neural generators of these two types of responses.
We studied lexical tone and speakers of one tone language as the starting point to inves-
tigate the development of FFR and LLR in relation to native and non-native speech sounds. The
broader applicability of these findings must be ascertained through future studies of other
speech categories and young children learning other languages. Future research must also
consider the impact of sleep on neural measurements. Although the vast majority of our par-
ticipants were sleeping naturally during the experiment, it was not clear at which exact stage of
the sleep cycle, and a few were even awake during the EEG recording. Research has demon-
strated that different sleep stages may affect the amplitude of the LLR, specifically P3 at the
latency of around 600–700 ms (Barnet et al., 1975). Although the focus of our LLR analysis
was on the P1 component, we could not completely rule out the influence of sleep stage on
our results. We expected that our large sample size and recordings at multiple time points
would reduce the impact of sleep on our findings. Nonetheless, we must acknowledge that
the lack of information about sleep stages from our participants might have affected the results.
Therefore, an important future direction of this line of research is to establish to what extent
our results are reproducible in the fully awake infants.
In summary, our findings suggest that neural centers at the earlier stages of the ascending
auditory pathway are crucial for native language development because they provide an accu-
rate representation of an incoming acoustic signal, regardless of whether or not those sounds
are native. This accurate representation would allow neural centers at later stages of the au-
ditory pathway to determine their linguistic relevance. Our study provides a critical piece of
information about language development that, to the best of our knowledge, no previous study
has considered. It opens up a new line of inquiry for future research. For example, future stud-
ies should examine the minimum required encoding accuracy in the brain stem and other
structures before the signal is sufficiently accurate for linguistic processing in the cortex.
They should investigate the age at which early-latency neural encoding contributes to language
development when the child is older, and the extent of this contribution. Given that long-term
sound exposure can modulate early-latency neural encoding in adulthood (Krishnan et al.,
2005, 2010; Wong et al., 2007), future studies should examine the age at which the influence
of auditory experience on neural encoding of native speech emerges. In terms of clinical ap-
plication, an understanding of the division of labour among different centers of the auditory
pathway would enable diagnostic protocols to be developed for investigating different subtypes
of central auditory processing and language problems (Ingvalson & Wong, 2013; Ettlinger et al.,
2014). While some disorders may be a result of poor encoding at lower-level centers, others
may stem from a failure to assign accurate phonological categories in the cortex.
ACKNOWLEDGMENTS
The authors wish to thank the families for participating in this study, many of whom were par-
ticipants in the Stanley Ho Developmental Cohort Study. We would also like to thank Lydia
Leung, Agnes Chou, Debby Ma, and Charlene Chiu for their assistance. This work was sup-
ported by the Dr. Stanley Ho Medical Development Foundation and a grant from the
Innovation and Technology Fund (Hong Kong) #ITS/067/18. Some of the results reported in
Neurobiology of Language
81
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
6
7
1
9
8
6
8
5
2
n
o
_
a
_
0
0
0
4
9
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Early development of neural speech does not depend on native language
this article were presented at the 2019 Annual Meeting of the Organization for Human Brain
Mapping.
FUNDING INFORMATION
Patrick Wong, Innovation and Technology Commission – Hong Kong (https://dx.doi.org/10
.13039/501100007156), Award ID: ITS/067/18. Patrick Wong, Dr. Stanley Ho Medical
Development Foundation.
AUTHOR CONTRIBUTIONS
Nikolay Novitskiy: Data curation, Formal analysis, Software, Validation, Visualization,
Writing – original draft. Akshay R. Maggu: Conceptualization, Methodology, Investigation,
Writing – review & editing. Ching Man Lai: Data curation, Investigation, Project administra-
tion, Writing – review & editing. Peggy H. Y. Chan: Data curation, Investigation, Writing –
review & editing. Kay H. Y. Wong: Data curation, Investigation, Writing – review & editing.
Hugh Simon Lam: Supervision, Resources, Writing – review & editing. Tak Yeung Leung:
Resources, Writing – review & editing. Ting Fan Leung: Supervision, Resources, Writing –
review & editing. Patrick C. M. Wong: Supervision, Conceptualization, Methodology,
Project administration, Funding acquisition, Resources, Writing – original draft.
COMPETING INTERESTS
Patrick C. M. Wong is the founder of a startup company supported by a Hong Kong SAR
Government startup scheme for universities.
REFERENCES
Alho, K., Z_ arnowiec, K., Gorina-Careta, N., & Escera, C. (2019).
Phonological task enhances the frequency-following response
to deviant task-irrelevant speech sounds. Frontiers in Human
Neuroscience, 13, Article 245. https://doi.org/10.3389/fnhum
.2019.00245, PubMed: 31379540
Anderson, S., Parbery-Clark, A., White-Schwoch, T., & Kraus, N.
(2012). Aging affects neural precision of speech encoding.
Journal of Neuroscience, 32(41), 14156–14164. https://doi.org
/10.1523/JNEUROSCI.2176-12.2012, PubMed: 23055485
Anderson, S., Parbery-Clark, A., White-Schwoch, T., & Kraus, N.
(2015). Development of subcortical speech representation in hu-
man infants. Journal of the Acoustical Society of America, 137(6),
3346–3355. https://doi.org/10.1121/1.4921032, PubMed:
26093424
Barnet, A. B., & Lodge, A. (1967). Click evoked EEG responses in
normal and developmentally retarded infants. Nature, 214(5085),
252–255. https://doi.org/10.1038/214252a0, PubMed:
4226768
Barnet, A. B., Ohlrich, E. S., Weiss, I. P., & Shanks, B. (1975).
Auditory evoked potentials during sleep in normal children from
ten days to three years of age. Electroencephalography and
Clinical Neurophysiology, 39(1), 29–41. https://doi.org/10.1016
/0013-4694(75)90124-8, PubMed: 50197
Best, C. T., Goldstein, L. M., Nam, H., & Tyler, M. D. (2016).
Articulating what infants attune to in native speech. Ecological
Psychology, 28(4), 216–261. https://doi.org/10.1080/10407413
.2016.1230372, PubMed: 28367052
Best, C. T., McRoberts, G. W., LaFleur, R., & Silver-Isenstadt, J.
(1995). Divergent developmental patterns for infants’ perception
of two nonnative consonant contrasts. Infant Behavior &
Development, 18(3), 339–350. https://doi.org/10.1016/0163
-6383(95)90022-5
Bidelman, G. M. (2018). Subcortical sources dominate the neuro-
electric auditory frequency-following response to speech.
NeuroImage, 175, 56–69. https://doi.org/10.1016/j.neuroimage
.2018.03.060, PubMed: 29604459
Bidelman, G. M., Davis, M. K., & Pridgen, M. H. (2018). Brainstem-
cortical functional connectivity for speech is differentially
challenged by noise and reverberation. Hearing Research, 367,
149–160. https://doi.org/10.1016/j.heares.2018.05.018,
PubMed: 29871826
Chang, E. F., Rieger, J. W., Johnson, K., Berger, M. S., Barbaro,
N. M., & Knight, R. T. (2010). Categorical speech representation
in human superior temporal gyrus. Nature Neuroscience, 13(11),
1428–1432. https://doi.org/10.1038/nn.2641, PubMed:
20890293
Chao, Y. R. (1930). ə sistim əv “toun-letəz” [A system of tone letters].
Le Maître Phonétique, 8, 24–27.
Cheng, Y. Y., Wu, H. C., Tzeng, Y. L., Yang, M. T., Zhao, L. L., &
Lee, C. Y. (2013). The development of mismatch responses to
Mandarin lexical tones in early infancy. Developmental
Neuropsychology, 38(5), 281–300. https://doi.org/10.1080
/87565641.2013.799672, PubMed: 23862633
Cheour, M., Ceponiene, R., Lehtokoski, A., Luuk, A., Allik, J., Alho,
K., & Näätänen, R. (1998). Development of language-specific
pho neme representation s in t he infant brain. Nature
Neuroscience, 1(5), 351–353. https://doi.org/10.1038/1561,
PubMed: 10196522
Neurobiology of Language
82
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
6
7
1
9
8
6
8
5
2
n
o
_
a
_
0
0
0
4
9
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Early development of neural speech does not depend on native language
Cheour-Luhtanen, M., Alho, K., Kujala, T., Sainio, K., Reinikainen,
K., Renlund, M., Aaltonen, O., Eerola, O., & Näätänen, R.
(1995). Mismatch negativity indicates vowel discrimination in
newborns. Hearing Research, 82(1), 53–58. https://doi.org/10
.1016/0378-5955(94)00164-L, PubMed: 7744713
Coffey, E. B. J., Nicol, T., White-Schwoch, T., Chandrasekaran, B.,
Krizman, J., Skoe, E., Zatorre, R. J., & Kraus, N. (2019). Evolving
perspectives on the sources of the frequency-following response.
Nature Communications, 10, Article 5036. https://doi.org/10
.1038/s41467-019-13003-w, PubMed: 31695046
Conboy, B. T., & Kuhl, P. K. (2011). Impact of second-language
experience in infancy: Brain measures of first- and second-
language speech perception. Developmental Science, 14(2),
242–248. https://doi.org/10.1111/j.1467-7687.2010.00973.x,
PubMed: 21499512
Cone, B., & Whitaker, R. (2013). Dynamics of infant cortical audi-
tory evoked potentials (CAEPs) for tone and speech tokens.
International Journal of Pediatric Otorhinolaryngology, 77(7),
1162–1173. https://doi.org/10.1016/j.ijporl.2013.04.030,
PubMed: 23722003
Cortesa, C. S., Hudac, C. M., & Molfese, D. L. (2019). Dynamic
effects of habituation and novelty detection on newborn event-
related potentials. Brain and Language, 199, Article 104695.
https://doi.org/10.1016/j.bandl.2019.104695, PubMed:
31610478
Delorme, A., & Makeig, S. (2004). EEGLAB: An open source tool-
box for analysis of single-trial EEG dynamics including indepen-
dent component analysis. Journal of Neuroscience Methods,
134(1), 9–21. https://doi.org/10.1016/j.jneumeth.2003.10.009,
PubMed: 15102499
Eilers, P. H. C. (2003). A perfect smoother. Analytical Chemistry,
75(14), 3631–3636. https://doi.org/10.1021/ac034173t,
PubMed: 14570219
Escera, C. (2017). The role of the auditory brainstem in regularity
encoding and deviance detection. In N. Kraus, S. Anderson, T.
White-Schwoch, R. R. Fay, & A. N. Popper (Eds.), The
frequency-following response: A window into human communi-
cation (pp. 101–120). Springer. https://doi.org/10.1007/978-3
-319-47944-6_5
Ettlinger, M., Bradlow, A. R., & Wong, P. C. M. (2014). Variability
in the learning of complex morphophonology. Applied
Psycholinguistics, 35(4), 807–831. https://doi.org/10.1017
/S0142716412000586
Fromkin, V. (2014). Tone: A linguistic survey. Academic Press.
Gansonre, C., Højlund, A., Leminen, A., Bailey, C., & Shtyrov, Y.
(2018). Task-free auditory EEG paradigm for probing multiple
levels of speech processing in the brain. Psychophysiology,
55(11), Article e13216. https://doi.org/10.1111/psyp.13216,
PubMed: 30101984
Godey, B., Schwartz, D., De Graaf, J. B., Chauvel, P., & Liégeois-
Chauvel, C. (2001). Neuromagnetic source localization of
auditory evoked fields and intracerebral evoked potentials: A
comparison of data in the same patients. Clinical Neurophysiology,
112(10), 1850–1859. https://doi.org/10.1016/S1388-2457(01)
00636-8, PubMed: 11595143
Government of Hong Kong SAR. (2016). Hong Kong: The Facts.
https://www.gov.hk/en/about/abouthk/facts.htm
Grimaldi, M. (2018). The phonetics-phonology relationship in the
neurobiology of language. In R. Petrosino, P. Cerrone, & H. van
der Hulst (Eds.), From sounds to structures (pp. 65–103). De
Gruyter. https://doi.org/10.1515/9781501506734-003
Hartley, D. E. H., & King, A. J. (2010). Development of the auditory
pathway. In A. Rees & A. R. Palmer (Eds.), The Oxford handbook
of auditory science: The auditory brain (pp. 361–386). Oxford
University Press.
Hartley, D. E. H., Vongpaisal, T., Xu, J., Shepherd, R. K., King, A. J.,
& Isaiah, A. (2010). Bilateral cochlear implantation in the ferret:
A novel animal model for behavioral studies. Journal of
Neuroscience Methods, 190(2), 214–228. https://doi.org/10
.1016/j.jneumeth.2010.05.014, PubMed: 20576507
Ingvalson, E. M., & Wong, P. C. M. (2013). Training to improve
language outcomes in cochlear implant recipients. Frontiers in
Psychology, 4, Article 263. https://10.3389/fpsyg.2013.00263,
PubMed: 23675364
Intartaglia, B., White-Schwoch, T., Meunier, C., Roman, S., Kraus,
N., & Schön, D. (2016). Native language shapes automatic neu-
ral processing of speech. Neuropsychologia, 89, 57–65. https://
doi.org/10.1016/j.neuropsychologia.2016.05.033, PubMed:
27263123
Jeng, F.-C., Hu, J., Dickman, B., Montgomery-Reagan, K., Tong, M.,
Wu, G., & Lin, C.-D. (2011). Cross-linguistic comparison of
frequency-following responses to voice pitch in American and
Chinese neonates and adults. Ear and Hearing, 32(6), 699–707.
https://doi.org/10.1097/AUD.0b013e31821cc0df, PubMed:
21543983
Jeng, F.-C., Lin, C.-D., Chou, M. S., Hollister, G. R., Sabol, J. T.,
Mayhugh, G. N., Wang, T.-C., & Wang, C.-Y. (2016).
Development of subcortical pitch representation in three-
month-old Chinese infants. Perceptual and Motor Skills, 122,
123–135. https://doi.org/10.1177/0031512516631054,
PubMed: 27420311
Jeng, F.-C., Peris, K. S., Hu, J., & Lin, C.-D. (2013). Evaluation of an
automated procedure for detecting frequency-following
responses in American and Chinese neonates. Perceptual and
Motor Skills, 116, 456–465. https://doi.org/10.2466/24.10.PMS
.116.2.456-465, PubMed: 24032322
Key, A. P., Lambert, E. W., Aschner, J. L., & Maitre, N. L. (2012).
Influence of gestational age and postnatal age on speech sound
processing in NICU infants. Psychophysiology, 49(5), 720–731.
https://doi.org/10.1111/j.1469-8986.2011.01353.x, PubMed:
22332725
Krishnan, A., Gandour, J. T., & Bidelman, G. M. (2010). The effects
of tone language experience on pitch processing in the brain-
stem. Journal of Neurolinguistics, 23(1), 81–95. https://doi.org
/10.1016/j.jneuroling.2009.09.001, PubMed: 20161561
Krishnan, A., Swaminathan, J., & Gandour, J. T. (2009). Experience-
dependent enhancement of linguistic pitch representation in the
brainstem is not specific to a speech context. Journal of Cognitive
Neuroscience, 21(6), 1092–1105. https://doi.org/10.1162/jocn
.2009.21077, PubMed: 18702588
Krishnan, A., Xu, Y., Gandour, J., & Cariani, P. (2005). Encoding of
pitch in the human brainstem is sensitive to language experience.
Cognitive Brain Research, 25(1), 161–168. https://doi.org/10
.1016/j.cogbrainres.2005.05.004, PubMed: 15935624
Kuhl, P. K. (2004). Early language acquisition: Cracking the speech
code. Nature Reviews Neuroscience, 5(11), 831–843. https://doi
.org/10.1038/nrn1533, PubMed: 15496861
Kuhl, P. K. (2010). Brain mechanisms in early language acquisition.
Neuron, 67(5), 713–727. https://doi.org/10.1016/j.neuron.2010
.08.038, PubMed: 20826304
Kuhl, P. K. (2014). Early language learning and the social brain.
Cold Spring Harbor Symposia on Quantitative Biology, 79,
211–220. https://doi.org/10.1101/sqb.2014.79.024802,
PubMed: 25943768
Kuhl, P. K., Conboy, B. T., Coffey-Corina, S., Padden, D., Rivera-
Gaxiola, M., & Nelson, T. (2008). Phonetic learning as a pathway
Neurobiology of Language
83
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
6
7
1
9
8
6
8
5
2
n
o
_
a
_
0
0
0
4
9
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Early development of neural speech does not depend on native language
(cid:2)
Kushnerenko, E., C
to language: New data and native language magnet theory
expanded (NLM-e). Philosophical Transactions of the Royal
Society B: Biological Sciences, 363(1493), 979–1000. https://
doi.org/10.1098/rstb.2007.2154, PubMed: 17846016
eponiene_ , R., Balan, P., Fellman, V.,
Huotilainen, M., & Näätänen, R. (2002). Maturation of the audi-
tory event-related potentials during the first year of life.
NeuroReport: For Rapid Communication of Neuroscience
Research, 13(1), 47–51. https://doi.org/10.1097/00001756
-200201210-00014, PubMed: 11924892
Kuuluvainen, S., Nevalainen, P., Sorokin, A., Mittag, M., Partanen,
E., Putkinen, V., Seppänen, M., Kähkönen, S., & Kujala, T.
(2014). The neural basis of sublexical speech and corresponding
nonspeech processing: A combined EEG-MEG study. Brain and
Language, 130, 19–32. https://doi.org/10.1016/j.bandl.2014.01
.008, PubMed: 24576806
Leaver, A. M., & Rauschecker, J. P. (2016). Functional topography
of human auditory cortex. Journal of Neuroscience, 36(4),
1416–1428. https://doi.org/10.1523/JNEUROSCI.0226-15.2016,
PubMed: 26818527
Lippé, S., Martinez-Montes, E., Arcand, C., & Lassonde, M. (2009).
E l e c t r o p h y s i o l o g i c a l s t u d y o f au d i t o r y d e v el o p m e n t .
Neuroscience, 164(3), 1108–1118. https://doi.org/10.1016/j
.neuroscience.2009.07.066, PubMed: 19665050
Lopez-Calderon, J., & Luck, S. J. (2014). ERPLAB: An open-source
toolbox for the analysis of event-related potentials. Frontiers in
Human Neuroscience, 8, Article 213. https://doi.org/10.3389
/fnhum.2014.00213, PubMed: 24782741
Maggu, A. R., Liu, F., Antoniou, M., & Wong, P. C. M. (2016).
Neural correlates of indicators of sound change in Cantonese:
Evidence from cortical and subcortical processes. Frontiers in
Human Neuroscience, 10, Article 652. https://doi.org/10.3389
/fnhum.2016.00652, PubMed: 28066218
Maggu, A. R., Zong, W., Law, V., & Wong, P. C. M. (2018).
Learning two tone languages enhances the brainstem encoding
of lexical tones. In 19th annual conference of the international
speech communication association, INTERSPEECH 2018
(Hyderabad, India, pp. 1437–1441). Curran Associates. https://
doi.org/10.21437/Interspeech.2018-2130
Malmierca, M. S., Carbajal, G. V., & Escera, C. (2019). Deviance
detection and encoding acoustic regularity in the auditory mid-
brain. In K. Kandler (Ed.), Oxford handbook of the auditory brain-
stem (pp. 706–740). Oxford University Press. https://doi.org/10
.1093/oxfordhb/9780190849061.013.19
Mattock, K., & Burnham, D. (2006). Chinese and English infants’
tone perception: Evidence for perceptual reorganization.
I n f a n c y , 1 0 ( 3 ) , 2 4 1 – 2 6 5 . h t t p s : / / d o i . o r g / 1 0 . 1 2 0 7
/s15327078in1003_3
McCarthy, K. M., Skoruppa, K., & Iverson, P. (2019). Development
of neural perceptual vowel spaces during the first year of life.
Scientific Reports, 9, Article 19592. https://doi.org/10.1038
/s41598-019-55085-y, PubMed: 31862999
Mesgarani, N., Cheung, C., Johnson, K., & Chang, E. F. (2014).
Phonetic feature encoding in human superior temporal gyrus.
Science, 343(6174), 1006–1010. https://doi.org/10.1126
/science.1245994, PubMed: 24482117
Moore, J. K., & Guan, Y. L. (2001). Cytoarchitectural and axonal
Journ al of the
mat uration in huma n a uditory cort e x.
Association for Research in Otolaryngology, 2(4), 297–311.
https://doi.org/10.1007/s101620010052, PubMed: 11833605
Moore, J. K., Guan, Y. L., & Shi, S. R. (1997). Axogenesis in the
human fetal auditory system, demonstrated by neurofilament
immunohistochemistry. Anatomy and Embryology, 195(1),
15–30. https://doi.org/10.1007/s004290050021, PubMed:
9006712
Moore, J. K., Guan, Y. L., & Shi, S. R. (1998). MAP2 expression in
developing dendrites of human brainstem auditory neurons.
Journal of Chemical Neuroanatomy, 16(1), 1–15. https://doi.org
/10.1016/S0891-0618(98)00057-X, PubMed: 9924969
Moore, J. K., & Linthicum, F. H., Jr. (2001). Myelination of the
human auditory nerve: Different time courses for Schwann cell
and glial myelin. Annals of Otology, Rhinology, & Laryngology,
1 1 0 ( 7 , P t . 1 ) , 6 5 5 – 6 6 1 . h t t p s : / / d o i . o r g / 1 0 . 1 1 7 7
/000348940111000711, PubMed: 11465825
Moore, J. K., & Linthicum, F. H., Jr. (2007). The human auditory
system: A timeline of development. International Journal of
A u d i o l o g y , 4 6 ( 9 ) , 4 6 0 – 4 7 8 . h t t p s : / / d o i . o r g / 1 0 . 1 0 8 0
/14992020701383019, PubMed: 17828663
Moore, J. K., Perazzo, L. M., & Braun, A. (1995). Time course of
axonal myelination in the human brainstem auditory pathway.
Hearing Research, 87(1–2), 21–31. https://doi.org/10.1016
/0378-5955(95)00073-D, PubMed: 8567438
Moushegian, G., Rupert, A. L., & Stillman, R. D. (1973). Scalp-
recorded early responses in man to frequencies in the speech
range. Electroencephalography & Clinical Neurophysiology,
35(6), 665–667. https://doi.org/10.1016/0013-4694(73)90223-X,
PubMed: 4128165
Näätänen, R., Lehtokoski, A., Lennes, M., Cheour, M., Huotilainen,
M., Iivonen, A., Vainio, M., Alku, P., Ilmoniemi, R. J., Luuk, A.,
Allik, J., Sinkkonen, J., & Alho, K. (1997). Language-specific
phoneme representations revealed by electric and magnetic
brain responses. Nature, 385(6615), 432–434. https://doi.org/10
.1038/385432a0, PubMed: 9009189
Näätänen, R., & Picton, T. W. (1987). The N1 wave of the human
electric and magnetic response to sound: A review and an
analysis of the component structure. Psychophysiology, 24(4),
375–425. https://doi.org/10.1111/j.1469-8986.1987.tb00311.x,
PubMed: 3615753
Näätänen, R., & Winkler, I. (1999). The concept of auditory stimu-
lus representation in cognitive neuroscience. Psychological
Bulletin, 125(6), 826–859. https://doi.org/10.1037/0033-2909
.125.6.826, PubMed: 10589304
Novak, G. P., Kurtzberg, D., Kreuzer, J. A., & Vaughan, H. G.
(1989). Cortical responses to speech sounds and their formants
in normal infants: Maturational sequence and spatiotemporal
analysis. Electroencephalography & Clinical Neurophysiology,
73(4), 295–305. https://doi.org/10.1016/0013-4694(89)90108-9,
PubMed: 2477216
Ortiz-Mantilla, S., Hamalainen, J. A., Musacchia, G., & Benasich,
A. A. (2013). Enhancement of gamma oscillations indicates pref-
erential processing of native over foreign phonemic contrasts in
infants. Journal of Neuroscience, 33(48), 18746–18754. https://
doi.org/10.1523/ JNEUROSCI.3260-13.2013, PubMed:
24285881
Partanen, E., Kujala, T., Näätänen, R., Liitola, A., Sambeth, A., &
Huotilainen, M. (2013). Learning-induced neural plasticity of
speech processing before birth. Proceedings of the National
Academy of Sciences of the United States of America, 110(37),
15145–15150. https://doi.org/10.1073/pnas.1302159110,
PubMed: 23980148
Ponton, C. W., Eggermont, J. J., Khosla, D., Kwong, B., & Don,
M.
(2002). Maturation of human central auditory system
activity: Separating auditory evoked potentials by dipole
source modeling. Clinical Neurophysiology, 113(3), 407–420.
https://doi.org/10.1016/S1388-2457(01)00733-7, PubMed:
11897541
Neurobiology of Language
84
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
6
7
1
9
8
6
8
5
2
n
o
_
a
_
0
0
0
4
9
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Early development of neural speech does not depend on native language
Raznahan, A., Shaw, P. W., Lerch, J. P., Clasen, L. S., Greenstein,
D., Berman, R., Pipitone. J., Chakravarty, M. M., & Giedd, J. N.
(2014). Longitudinal four-dimensional mapping of subcortical
anatomy in human development. Proceedings of the National
Academy of Sciences of the United States of America, 111(4),
1592–1597. https://doi.org/10.1073/pnas.1316911111,
PubMed: 24474784
Reetzke, R., Xie, Z., Llanos, F., & Chandrasekaran, B. (2018).
Tracing the trajectory of sensory plasticity across different
stages of speech learning in adulthood. Current Biology, 28(9),
1419–1427. https://doi.org/10.1016/j.cub.2018.03.026,
PubMed: 29681473
Ribas-Prats, T., Almeida, L., Costa-Faidella, J., Plana, M., Corral,
M. J., Gómez-Roig, M. D., & Escera, C. (2019). The frequency-
following response (FFR) to speech stimuli: A normative dataset
in healthy newborns. Hearing Research, 371, 28–39. https://doi
.org/10.1016/j.heares.2018.11.001, PubMed: 30448690
Rivera-Gaxiola, M., Klarman, C. A. L., Garcia-Sierra, A., & Kuhl, P. K.
(2005). Neural patterns to speech and vocabulary growth in
American infants. Neuroreport, 16(5), 495–498. https://doi.org
/10.1097/00001756-200504040-00015, PubMed: 15770158
Rivera-Gaxiola, M., Silva-Pereyra, J., & Kuhl, P. K. (2005). Brain
potentials to native and non-native speech contrasts in 7- and
11-month-old American infants. Developmental Science, 8,
162–172. https://doi.org/10.1111/j.1467-7687.2005.00403.x,
PubMed: 15720374
Romand, R., & Romand, M.-R. (1982). Myelination kinetics of spi-
ral ganglion cells in kitten. Journal of Comparative Neurology,
204(1), 1–5. https://doi.org/10.1002/cne.902040102, PubMed:
7056884
Romand, R., Sans, A., Romand, M. R., & Marty, R. (1976). The
structural maturation of the stato-acoustic nerve in the cat.
Journal of Comparative Neurology, 170(1), 1–15. https://doi.org
/10.1002/cne.901700102, PubMed: 977813
Ross, B., & Tremblay, K. (2009). Stimulus experience modifies au-
ditory neuromagnetic responses in young and older listeners.
Hearing Research, 248(1–2), 48–59. https://doi.org/10.1016/j
.heares.2008.11.012, PubMed: 19110047
Sano, M., Kaga, K., Kuan, C. C., Ino, K., & Mima, K. (2007). Early
myelination patterns in the brainstem auditory nuclei and path-
way: MRI evaluation study. International Journal of Pediatric
Otorhinolaryngology, 71(7), 1105–1115. https://doi.org/10.1016
/j.ijporl.2007.04.002, PubMed: 17485121
Shafer, V. L., Yu, Y. H., & Wagner, M. (2015). Maturation of corti-
cal auditory evoked potentials (CAEPs) to speech recorded from
frontocentral and temporal sites: Three months to eight years of
age. International Journal of Psychophysiology, 95(2), 77–93.
https://doi.org/10.1016/j.ijpsycho.2014.08.1390, PubMed:
25219893
Shiga, T., Althen, H., Cornella, M., Zarnowiec, K., Yabe, H., &
Escera, C. (2015). Deviance-related responses along the auditory
hierarchy: Combined FFR, MLR and MMN evidence. PLOS One,
10(9), Article e0136794. https://doi.org/10.1371/journal.pone
.0136794, PubMed: 26348628
Shtyrov, Y., Kujala, T., Palva, S., Ilmoniemi, R. J., & Näätänen, R.
(2000). Discrimination of speech and of complex nonspeech
sounds of different temporal structure in the left and right cere-
bral hemispheres. NeuroImage, 12(6), 657–663. https://doi.org
/10.1006/nimg.2000.0646, PubMed: 11112397
Skoe, E., & Kraus, N. (2010). Auditory brain stem response to
complex sounds: A tutorial. Ear and Hearing, 31(3), 302–324.
https://doi.org/10.1097/AUD.0b013e3181cdb272, PubMed:
20084007
Skoe, E., Krizman, J., Anderson, S., & Kraus, N. (2015). Stability and
plasticity of auditory brainstem function across the lifespan.
Cerebral Cortex, 25(6), 1415–1426. https://doi.org/10.1093
/cercor/bht311, PubMed: 24366906
Slabu, L., Grimm, S., & Escera, C. (2012). Novelty detection in the
human auditory brainstem. Journal of Neuroscience, 32(4),
1447–1452. https://doi.org/10.1523/JNEUROSCI.2557-11.2012,
PubMed: 22279229
Small, S. A., Sharma, M., Bradford, M., & Vasuki, P. R. M. (2018).
The effect of signal to noise ratio on cortical auditory-evoked
potentials elicited to speech stimuli in infants and adults with
normal hearing. Ear and Hearing, 39(2), 305–317. https://doi
.org/10.1097/AUD.0000000000000487, PubMed: 28863034
Song, J. H., Skoe, E., Wong, P. C., & Kraus, N. (2008). Plasticity
in the adult human auditory brainstem following short-term
linguistic training. Journal of Cognitive Neuroscience, 20(10),
1892–1902. https://doi.org/10.1162/jocn.2008.20131,
PubMed: 18370594
Suga, N. (2008). Role of corticofugal feedback in hearing. Journal of
Comparative Physiology A, 194(2), 169–183. https://doi.org/10
.1007/s00359-007-0274-2, PubMed: 18228080
Suga, N., Gao, E., Zhang, Y., Ma, X., & Olsen, J. F. (2000). The cor-
ticofugal system for hearing: Recent progress. Proceedings of the
National Academy of Sciences of the United States of America,
97(22), 11807–11814. https://doi.org/10.1073/pnas.97.22
.11807, PubMed: 11050213
Tallon-Baudry, C., Bertrand, O., Delpuech, C., & Pernier, J. (1996).
Stimulus specificity of phase-locked and non-phase-locked 40 Hz
visual responses in human. Journal of Neuroscience, 16(13),
4240–4249. https://doi.org/10.1523/JNEUROSCI.16-13-04240
.1996, PubMed: 8753885
Tichko, P., & Skoe, E. (2017). Frequency-dependent fine structure
in the frequency-following response: The byproduct of multiple
generators. Hearing Research, 348, 1–15. https://doi.org/10.1016
/j.heares.2017.01.014, PubMed: 28137699
Tremblay, K. L., & Kraus, N. (2002). Auditory training induces
asymmetrical changes in cortical neural activity. Journal of
Speech, Language, and Hearing Research, 45(3), 564–572.
https://doi.org/10.1044/1092-4388(2002/045)
Tremblay, K. L., Kraus, N., & McGee, T. (1998). The time course of
auditory perceptual learning: Neurophysiological changes dur-
ing speech-sound training. Neuroreport, 9(16), 3557–3560.
https://doi.org/10.1097/00001756-199811160-00003, PubMed:
9858359
Tremblay, K. L., Kraus, N., McGee, T., Ponton, C. W., & Otis, A. B.
(2001). Central auditory plasticity: Changes in the N1-P2 complex
after speech-sound training. Ear and Hearing, 22(2), 79–90.
https://doi.org/10.1097/00003446-200104000-00001, PubMed:
11324846
Tremblay, K. L., Ross, B., Inoue, K., McClannahan, K., & Collet, G.
(2014). Is the auditory evoked P2 response a biomarker of learn-
ing? Frontiers in Systems Neuroscience, 8, Article 28. https://doi
.org/10.3389/fnsys.2014.00028, PubMed: 24600358
Werker, J. F., & Tees, R. C. (1984). Cross-language speech percep-
tion: Evidence for perceptual reorganization during the first year
of life. Infant Behavior & Development, 7(1), 49–63. https://doi
.org/10.1016/S0163-6383(84)80022-3
Werker, J. F., Yeung, H. H., & Yoshida, K. A. (2012). How do in-
fants become experts at native-speech perception? Current
Directions in Psychological Science, 21, 221–226. https://doi
.org/10.1177/0963721412449459
Wierenga, L., Langen, M., Ambrosino, S., van Dijk, S., Oranje, B.,
& Durston, S. (2014). Typical development of basal ganglia,
Neurobiology of Language
85
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
6
7
1
9
8
6
8
5
2
n
o
_
a
_
0
0
0
4
9
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Early development of neural speech does not depend on native language
hippocampus, amygdala and cerebellum from age 7 to 24.
NeuroImage, 96, 67–72. https://doi.org/10.1016/j.neuroimage
.2014.03.072, PubMed: 24705201
Wong, P. C. M., Skoe, E., Russo, N. M., Dees, T., & Kraus, N. (2007).
Musical experience shapes human brainstem encoding of linguis-
tic pitch patterns. Nature Neuroscience, 10(4), 420–422. https://
doi.org/10.1038/nn1872, PubMed: 17351633
Wunderlich, J. L., & Cone-Wesson, B. K. (2006). Maturation of CAEP
in infants and children: A review. Hearing Research, 212(1–2),
212–223. https://doi.org/10.1016/j.heares.2005.11.008, PubMed:
16480841
Yeung, H. H., Chen, K. H., & Werker, J. F. (2013). When does na-
tive language input affect phonetic perception? The precocious
case of lexical tone. Journal of Memory and Language, 68,
123–139. https://doi.org/10.1016/j.jml.2012.09.004
Yip, M. (2002). Tone. Cambridge University Press. https://doi.org
/10.1017/CBO9781139164559
Zhao, T. C., & Kuhl, P. K. (2018). Linguistic effect on speech percep-
tion observed at the brainstem. Proceedings of the National
Academy of Sciences of the United States of America, 115(35),
8716–8721. https://doi.org/10.1073/pnas.1800186115,
PubMed: 30104356
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
6
7
1
9
8
6
8
5
2
n
o
_
a
_
0
0
0
4
9
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neurobiology of Language
86