RESEARCH ARTICLE
The Precentral Gyrus Contributions
to the Early Time-Course of
Grapheme-to-Phoneme Conversion
Erik Kaestner1
Chad Carlson3
, Xiaojing Wu2, Daniel Friedman2, Patricia Dugan2, Orrin Devinsky2
, Werner Doyle2,4, Thomas Thesen2, and Eric Halgren5,6
,
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
1
8
1
9
8
6
8
7
7
n
o
_
a
_
0
0
0
4
7
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
1Center for Multimodal Imaging and Genetics, University of California, San Diego, USA
2Department of Neurology, NYU Langone School of Medicine, New York, USA
3Department of Neurology, Medical College of Wisconsin, Milwaukee, USA
4Department of Neurosurgery, NYU Langone School of Medicine, New York, USA
5Department of Neurosciences, University of California at San Diego, La Jolla, USA
6Department of Radiology, University of California at San Diego, La Jolla, USA
Keywords: electrocorticography, reading, graphemes, phonemes, audiovisual integration
ABSTRACT
As part of silent reading models, visual orthographic information is transduced into an auditory
phonological code in a process of grapheme-to-phoneme conversion (GPC). This process is
often identified with lateral temporal-parietal regions associated with auditory phoneme
encoding. However, the role of articulatory phonemic representations and the precentral gyrus
in GPC is ambiguous. Though the precentral gyrus is implicated in many functional MRI
studies of reading, it is not clear if the time course of activity in this region is consistent with the
precentral gyrus being involved in GPC. We recorded cortical electrophysiology during a
bimodal match/mismatch task from eight patients with perisylvian subdural electrodes to
examine the time course of neural activity during a task that necessitated GPC. Patients made a
match/mismatch decision between a 3-letter string and the following auditory bi-phoneme.
We characterized the distribution and timing of evoked broadband high gamma (70–170 Hz)
as well as phase-locking between electrodes. The precentral gyrus emerged with a high
concentration of broadband high gamma responses to visual and auditory language as well as
mismatch effects. The pars opercularis, supramarginal gyrus, and superior temporal gyrus were
also involved. The precentral gyrus showed strong phase-locking with the caudal fusiform
gyrus during letter-string presentation and with surrounding perisylvian cortex during the
bimodal visual-auditory comparison period. These findings hint at a role for precentral cortex
in transducing visual into auditory codes during silent reading.
INTRODUCTION
The interaction of an initially auditory-only perisylvian language network and a later develop-
ing reading network has been studied since the late 1800s (Dejerine, 1892). Decades of intri-
cate behavioral work (Frost, 1998; Rastle & Brysbaert, 2006) combined with lesion studies
(Coltheart, 1980; Geschwind, 1974) led to the development of cognitive (Coltheart et al.,
2001; Diependaele et al., 2010; Harm & Seidenberg, 2004; Perry et al., 2007) and
a n o p e n a c c e s s
j o u r n a l
Citation: Kaestner, E., Wu, X.,
Friedman, D., Dugan, P., Devinsky, O.,
Carlson, C., Doyle, W., Thesen, T., &
Halgren, E. (2022). The precentral gyrus
contributions to the early time-course
of grapheme-to-phoneme conversion.
Neurobiology of Language, 3(1), 18–45.
https://doi.org/10.1162/nol_a_00047
DOI:
https://doi.org/10.1162/nol_a_00047
Supporting Information:
https://doi.org/10.1162/nol_a_00047
Received: 29 July 2020
Accepted: 16 June 2021
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Authors:
Erik Kaestner
ekaestne@health.ucsd.edu
Eric Halgren
ehalgren@health.ucsd.edu
Handling Editor:
Edward Chang
Copyright: © 2021
Massachusetts Institute of Technology
Published under a Creative Commons
Attribution 4.0 International
(CC BY 4.0) license
The MIT Press
Precentral gyrus and grapheme-phoneme conversion
Grapheme-to-phoneme conversion:
The conversion from a visual
orthographic code into an auditory
phonological code during reading.
neuroanatomical (Carreiras et al., 2014; Fiez & Petersen, 1998; Jobard et al., 2003; Price,
2012; Taylor et al., 2013) models which include interactive processing of both the graphemic
and phonological information contained in visual text. This necessitates the transduction of
graphemic into auditory phonological codes, followed by a period of lexical-semantic pro-
cessing in which visual and auditory processing interacts and influences one another. But ba-
sic questions of how a visual stimulus is transduced into an auditory code, known as
grapheme-to-phoneme conversion (GPC), remain unresolved. GPC has been associated with
both phonological encoding, classically located with Wernicke’s area in lateral temporal-
parietal regions, and phonological articulation, associated with frontal regions. However,
the possible role of the articulatory phonological representations in the precentral gyrus in
GPC remains ambiguous.
Early lesion studies focused attention on temporal-parietal regions as the entryway for visual
text into the wider auditory perisylvian lexical-semantic network via GPC (Geschwind, 1974).
Blood oxygen level-dependent (BOLD) fMRI studies provided early support for this localiza-
tion, with phonological decisions evoking greater activation than other decisions in the angu-
lar gyrus (Binder et al., 2005; Booth, 2002; McDermott et al., 2003) with several
neuroanatomical models of reading subsequently incorporating this locus (Carreiras et al.,
2014; Taylor et al., 2013). Also implicated is the nearby superior temporal gyrus (STG): It is
a critical processing area for auditory phonemes (Leonard et al., 2015; Mesgarani et al., 2014;
Travis et al., 2013) with phonologically related activity also evoked by visual language stimuli
(Booth, 2002). Intracranial electrophysiology studies have also found overlapping (Perrone-
Bertolotti et al., 2012) and correlated (Chan et al., 2014) activity to auditory and visual lan-
guage in the STG.
However, early psychological theory associated GPC with articulatory phonological cog-
nitive operations (Allport, 1979; Barron & Baron, 1977; Burani et al., 1991; Klapp, 1971;
Kleiman, 1975; Peterson & Johnson, 1971), presumably in articulatory motor cortex. This
was based on data from the articulatory suppression paradigm in which participants repeated
a nonsense phrase to occupy the articulatory cognitive operations while performing a reading
task. Results showed suppressed phonological effects (Barron & Baron, 1977; Burani et al.,
1991; Kleiman, 1975; Sun & Peperkamp, 2016) but not if mouth movements were non-
articulatory (Burani et al., 1991) or if the words were presented auditorily (Peterson &
Johnson, 1971). A variety of BOLD neuroimaging and lesion studies support precentral in-
volvement in silent reading. Masked phonological priming evokes activity in the left precentral
gyrus even when the words are not consciously perceived (Dehaene et al., 2001). Studies also
find greater activity in the precentral gyrus when making phonological judgements for visual
words (Price et al., 1997; Yen et al., 2019), with differential activation based on spelling-sound
consistency (Fiez et al., 1999) and difficultly of GPC (Binder et al., 2005). A perirolandic lesion
reduced a patient’s ability to make phonological judgements about words (Vallar et al., 1997).
Another patient with a perirolandic lesion similarly retained comprehension of visual words
but was unable to make rhyming judgements or manipulate pseudowords (Vallar & Cappa,
1987). In a patient with phonological alexia and agraphia, increased activation in the precen-
tral gyrus was observed during successful cognitive rehabilitation of phonological processing
(DeMarco et al., 2018). Finally, in transcranial magnetic stimulation, greater excitability in
motor cortex during GPC is reported (Lauro et al., 2020). The neighboring inferior frontal gyrus
cortical region pars opercularis has been similarly tied to phonological effects (Cornelissen
et al., 2009; Pammer et al., 2004; Wheat et al., 2010). Therefore, in addition to the lateral
temporal-parietal contributions to GPC, there is a strong case that the precentral gyrus is in-
volved in GPC as well.
Neurobiology of Language
19
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
1
8
1
9
8
6
8
7
7
n
o
_
a
_
0
0
0
4
7
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Precentral gyrus and grapheme-phoneme conversion
The Present Study
Here, we test the theory that the precentral gyrus is a contributor to GPC using a silent
match/mismatch task in which a 3-letter string containing sublexical phonological information
(e.g., “GUH”) was presented first, followed by auditory presentation of an auditory bi-
phoneme. During this task, intracranial electrophysiological (iEEG) activity was recorded from
the cortex. In cognitive models of reading, activation of letter/grapheme representations begins
first, and then is passed along to the graphemes’ phonological analogues by GPC (Coltheart
et al., 2001; Diependaele et al., 2010; Harm & Seidenberg, 2004). By using a bimodal task
with iEEG, which possesses high spatial-temporal resolution, we will observe the evolution of
the putative phonological-based processing during visual language encoding and then how
this activity is modified by incoming phonological information derived from auditory language
encoding. We will assess three hypotheses: (1) that a task focused on GPC will evoke visual
language activity in the precentral gyrus, (2) that this visual language evoked activity occurs at
a time consistent with precentral gyrus contributions to GPC, and (3) whether visual language
evoked activity is overlapping with auditory language evoked activity in the precentral gyrus.
The first hypothesis will be addressed by examining the distribution of increased activity to
linguistic stimuli compared to their sensory controls in perisylvian regions. Our hypothesis pre-
dicts that we will identify activity in the precentral gyrus at least as often as in the surrounding
frontal (e.g., pars opercularis) and temporal-parietal (e.g., STG and supramarginal) regions
which are highlighted in neurobiological models of reading (Carreiras et al., 2014; Fiez &
Petersen, 1998; Jobard et al., 2003; Price, 2012; Taylor et al., 2013), providing evidence that
the precentral gyrus is also active during GPC. Further, we will examine whether the precen-
tral gyrus demonstrates connectivity with the ventral visual language processing regions such
as the fusiform (Dehaene & Cohen, 2011; Lochy et al., 2018; Vinckier et al., 2007) as well as
the surrounding perisylvian regions implicated in visual language processing.
For the second hypothesis, we will characterize the time window of the observed activity in
the precentral gyrus. As mentioned, cognitive models of reading start with activation of
letter/grapheme representations followed by activation of phonemes via GPC (Coltheart
et al., 2001; Diependaele et al., 2010; Harm & Seidenberg, 2004). Neurobiological evidence
confirms this sequence. Letter/grapheme encoding occurs in posterior occipital-temporal re-
gions beginning at ~160–180 ms (Allison et al., 1994, 1999; Hirshorn et al., 2016; Thesen
et al., 2012). This is followed by widespread onset of visual language evoked activity across
large portions of the brain at around the same time (Halgren, 1990). For GPC, extracranial
electrophysiology identifies visual language evoked phonological effects beginning during a
similar time window of ~250–350 ms (Rugg, 1984; Grainger et al., 2006; Holcomb &
Anderson, 1993). Therefore, language evoked activity in the precentral gyrus during this time
window would be consistent with GPC (a question the spatial-temporal precision of iEEG is
well-placed to observe). Further, through network-level analyses we can examine whether
the precentral gyrus is significantly coupled with occipital-temporal regions during this time
window. Noninvasive imaging studies of resting-state connectivity (Stevens et al., 2017) and
diffusion tensor imaging (Bouhali et al., 2014) demonstrate that the precentral gyrus has connec-
tivity with these occipital-temporal regions, but they lack the temporal specificity to understand
when this connectivity may occur during visual language processing.
For the third hypothesis, we will seek to observe whether perisylvian visual language processing
is overlapping with auditory language processing in the same areas. Extracranial EEG evidence
demonstrates that visually presented language primes auditorily presented language, recorded over
broad brain regions within several 100 ms of onset (Holcomb et al., 2005; Kiyonaga et al., 2007).
Neurobiology of Language
20
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
1
8
1
9
8
6
8
7
7
n
o
_
a
_
0
0
0
4
7
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Precentral gyrus and grapheme-phoneme conversion
Using our match/mismatch task and the increased spatial precision of iEEG, we will assess whether
we can detect specific cortical patches which show evidence of auditory phonological represen-
tations being primed by visual phonological representations. Second, using this bimodal task will
allow us to observe whether phonological representations for auditorily encoded phonemes in
the STG (Mesgarani et al., 2014) and the precentral gyrus (Cheung et al., 2016) are also activated
by visual language stimuli. If the phonological representations activated during visual language
processing are the same phonological representations activated during auditory language
processing, we will be able to observe both overlap (i.e., visual and auditory effects in the same
electrode) and priming (i.e., differences in activity between matching and mismatching
visual/auditory phonemes in the same electrode) of these phonological representations.
Previous studies have found evidence that visually and auditorily evoked language activity
overlap in the STG when presented at separate times (Chan et al., 2014; Perrone-Bertolotti
et al., 2012), but the relationship is unknown for the precentral gyrus. Further, reports of
phoneme-specific activity in the STG (Mesgarani et al., 2014) raise the question of whether
GPC phoneme-specific activity (i.e., letter-specific activity) can be identified. Taken together,
these three hypotheses will inform whether visual language evoked activity in the precentral gy-
rus during silent visual language encoding is consistent with contributions to GPC.
MATERIALS AND METHODS
Participants and Recordings
Electrocorticographic recordings were obtained from 8 patients (5 males, mean age 35.6, age
range 17–56; Table 1) undergoing intracranial EEG monitoring as part of treatment for phar-
macologically resistant epilepsy. All procedures were approved by the Institutional Review
Board at New York University, and written informed consent was obtained from all partici-
pants. Electrode placement was determined by clinical criteria to identify seizure activity
and eloquent tissue. Each patient was implanted with subdural platinum-iridium electrode
arrays embedded in silastic sheets (AdTech Medical Instrument Corp.). Data included arrays of
grids (8 × 8 contacts) and strips (1 × 4 to 1 × 12 contacts). Contacts had a diameter of 4 mm with
2.3 mm exposure. Center-to-center spacing between contacts was 10 mm for grids and 5 mm for
microgrids. Recordings were acquired using a NicoletOne EEG system (https://neuro.natus.com/)
sampled at 512 Hz and bandpass filtered between 0.5 and 250 Hz. In total, there were 5
Table 1.
Patient clinical information, neuropsychological, and task performance
Age Onset
36
12
Sex Hand Wada
F
R
L
Implantation VCI POI WMI
114
102 113
L
47
24
56
25
26
54
17
35
22
30
M
F
M
0.1 M
14
35
13
M
F
M
R
R
R
R
R
R
R
L
L
–
–
–
L
L
L
R
R
R
L
L
L
72 102
100
–
84
–
110 100
–
–
96 102
91
92
92
83
–
83
–
86
89
P1
P2
P3
P4
P5
P6
P7
P8
PSI
102
77
86
–
94
–
94
86
Match
correct
95%
Mismatch
correct
98%
Match
RT (ms)
588
Mismatch
RT (ms)
604
84%
81%
94%
91%
86%
94%
89%
86%
95%
84%
98%
88%
92%
98%
704
601
611
587
755
738
698
740
595
746
640
757
719
627
Note. RT = response time; VCI = Verbal Comprehension Index; WMI = Working Memory Index; POI = Perceptual Organization Index; PSI = Processing Speed Index.
Neurobiology of Language
21
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
1
8
1
9
8
6
8
7
7
n
o
_
a
_
0
0
0
4
7
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Precentral gyrus and grapheme-phoneme conversion
implantations focused on the left hemisphere and 3 implantations focused on the right hemi-
sphere. Patient language lateralization based on the Wada procedure is noted in Table 1 when
it is available, with all patients with a Wada showing left lateralization. Three patients (P4, P5,
and P6) did not have Wada information available; however, all were right-handed and there-
fore believed to have typical (i.e., left) language lateralization.
Electrode Localization
Electrode localization was done through co-registration of pre- and postimplant MRI images,
followed by manual and automatic localization of electrodes (Yang et al., 2012). Coordinates
were co-registered to a standard MNI template and anatomical parcellations were determined
using a Desikan atlas (Desikan et al., 2006). For display purposes the atlas was slightly modified
by splitting long gyri into 3 equal segments, either inferior/middle/superior (precentral, post-
central parcellations) or caudal/middle/rostral (fusiform, inferior temporal, middle temporal,
super temporal, middle-frontal, superior-frontal parcellations). Three-dimensional reconstruc-
tions of cortical surfaces were created using FreeSurfer (Dale et al., 1999). Electrode localization
into a parcellation region was performed in each subject’s native brain space. Average electrode
locations, used only for display purposes, were obtained using FreeSurfer surface-to-surface
calculations with the fsaverage brain. Regions with <5 electrodes were excluded from analysis.
Task Design
Figure 1A displays a schematic of the task. Patients performed silent match>) for 450 ms, then immediately returning to the fixation. The
bi-phoneme was played next, which lasted for 450 ms, followed by a response period. In total,
1,000 ms from bi-phoneme onset were allowed for a participant response. Stimulus onset
asynchrony was varied between 700 and 1,000 ms randomly.
This was a silent task with participants responding with their hand, ipsilateral to the hemi-
sphere being recorded from to avoid hand-motor movement activity contaminating the record-
ings. The vocal silence of the task ensured that any activity observed in putative articulatory
cortex was sub-articulatory and not related to overt motor movement.
In total, four types of trials were presented. The first two trial types were Match and
Mismatch, in which the letter-string and the bi-phoneme either phonetically matched or did
not. The final two trial types provided visual and auditory sensory controls. In Visual Control
trials a 3-symbol false-font was displayed, followed by a normal bi-phoneme. These false-font
stimuli share the same basic visual sensory features matched to regular letters. Each false-font
character was matched to a real letter in the English alphabet in size, number of strokes, total
line length, and curvature (for example see Figure 1A; Thesen et al., 2012). In Auditory Control
trials the normal letter-string was displayed followed by a noise-vocoded stimulus. Noise-
vocoded stimuli preserve temporal envelope cues in broad frequency bands but restrict the
listener to degraded information on the distribution of spectral energy. Therefore, temporal
and amplitude cues of speech were preserved in each spectral band, providing a control for
the sensory characteristics of speech, but the spectral detail within each band was degraded. A
study of noise-vocoded speech processing using iEEG found that the number of bands
Neurobiology of Language
22
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
1
8
1
9
8
6
8
7
7
n
o
_
a
_
0
0
0
4
7
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Precentral gyrus and grapheme-phoneme conversion
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
1
8
1
9
8
6
8
7
7
n
o
_
a
_
0
0
0
4
7
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
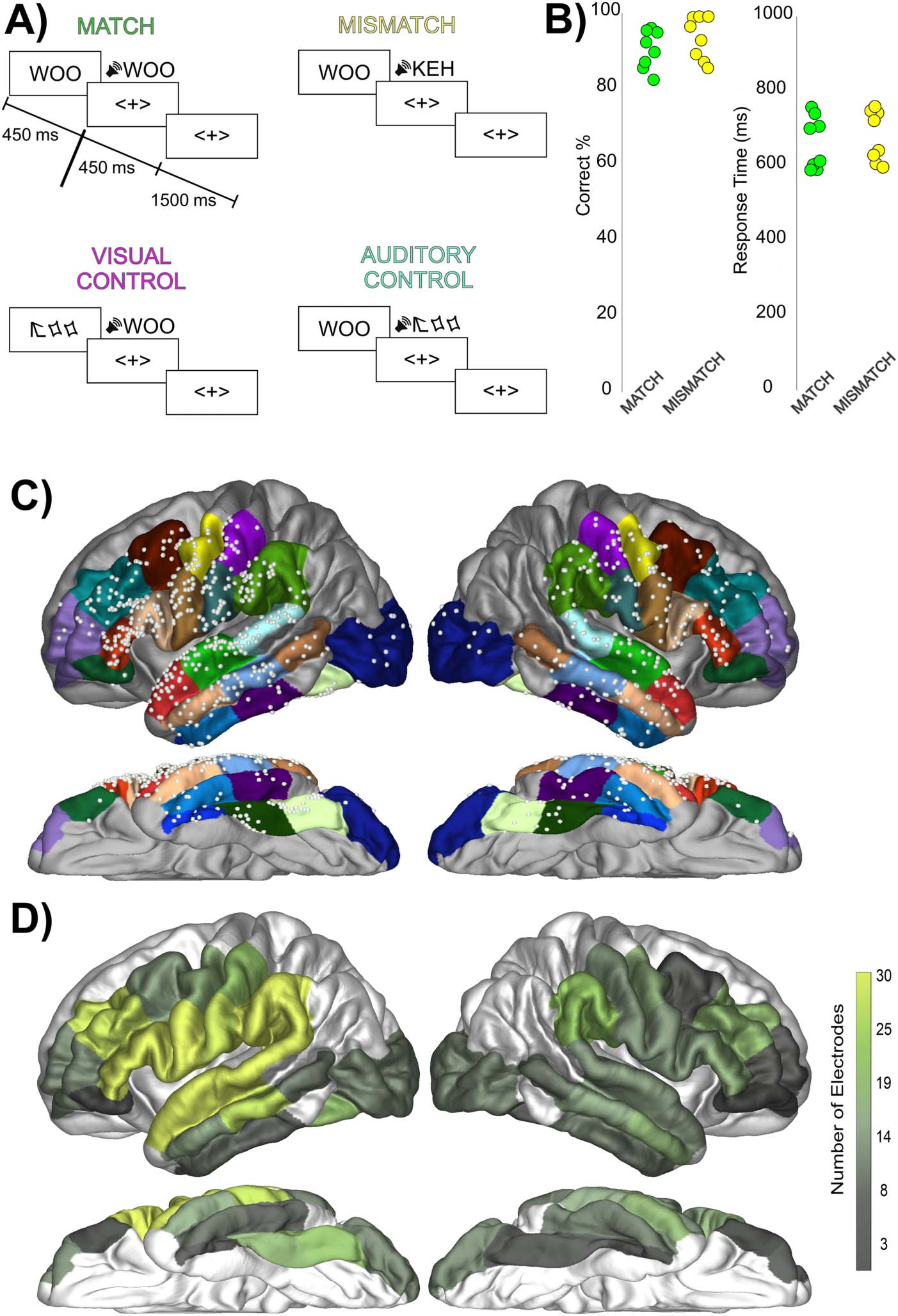
Figure 1. Task design, performance, and electrode coverage. (A) The sequence of stimulus presen-
tation in the four trial types. (B) Patient performance on the Match and Mismatch trials as expressed
by percentage correct and response time, demonstrating similar performance across the two trial
types. Each dot represents the performance of one patient. (C) Electrode coverage highlighting
the coverage of perisylvian electrodes. Electrode coverage across the included regions of interest
(ROIs) presented on an average brain for illustration purposes. Colors on brain highlight the ROIs
involved in the study from the Desikan atlas. Grey regions were not included in analyses due to lack
of electrode coverage in ROIs. (D) Electrode coverage expressed as the total number of electrodes
within each ROI.
Neurobiology of Language
23
Precentral gyrus and grapheme-phoneme conversion
determined how widespread processing of the stimuli was in perisylvian regions. At lower
numbers of bands (1–2 bands), speech was unintelligible and evidence of processing was
largely restricted to Heschl’s gyrus. However, at higher numbers of bands (≥3 bands for good
performers, at least >4 bands for bad performers), intelligibility increased above chance and
activity was found in regions such as the STG, though activity evoked by noise-vocoded stim-
uli was still less than clear speech (Nourski et al., 2019). Because we were interested in pro-
cessing beyond Heschl’s gyrus in regions such as the superior temporal and precentral gyri, we
chose to have a higher number of bands (i.e., to include some intelligibility in our noise-
vocoded stimuli). Therefore, noise-vocoded stimuli were created by taking the existing bi-
phoneme stimuli and creating a 6-band stimulus in which white-noise was multiplied by power
in each of the bands to create a matched set of auditory stimuli with identical time-varying
spectral acoustics (Chan et al., 2014; Chen & Loizou, 2011; Horowitz, 2014; Souza &
Rosen, 2009; Travis et al., 2013). On both types of control trials, participants were instructed
to respond with a Mismatch response. In total, there were 768 trials, with 192 of each of the four
trial types. These trials were broken down into 3 runs. Within each run, letter-string/bi-phoneme
stimuli were created by crossing 4 consonants and 4 vowels (i.e., 16 bi-phoneme combinations
per run) to facilitate balanced presentations of each letter and phoneme.
Data Processing
Data were preprocessed using MATLAB (MathWorks), the Fieldtrip toolbox (Oostenveld et al.,
2011), and custom scripts. We used an average subtraction reference for each patient to remove
global artifacts and noise, followed by a bandstop around line-noise and its harmonics (60, 120,
180 Hz). Data were epoched to the onset of the letter-string, from −1,500 to 2,500 ms, to avoid
epoch-related edge artifacts introduced by converting from the time domain to the frequency
domain. Temporal padding was removed at the end of preprocessing for finalized −500 to
1,500 ms epochs. To calculate broadband high gamma (BHG), epochs were transformed to the
time–frequency domain using the wavelet transform from 70 to 170 Hz in 10 Hz increments.
Constant temporal and frequency resolution across target frequencies were obtained by adjusting
the wavelet widths according to the target frequency. The wavelet widths increase linearly from 14
to 38 as frequency increased from 70 to 170 Hz, resulting in a constant temporal resolution with a
standard deviation of 16 ms and frequency resolution of 10 Hz. For each epoch, spectral power was
calculated from the wavelet spectra, normalized by the inverse square frequency to adjust for the
rapid drop-off in the EEG power spectrum with frequency, and averaged from 70 to 170 Hz,
excluding line noise harmonics. This data was smoothed by a moving window matching the tem-
poral characteristics of the wavelet (i.e., a normal distribution with 16 ms standard deviation). Each
trial epoch was demeaned with a baseline from −250 to 0 ms. Trials containing artifacts were iden-
tified by amplitude and variance, visually inspected for artifacts, and removed from further analysis.
Analysis
Behavior
We compared patient performance and response speed on Match and Mismatch trials with a
t test to gauge if electrophysiological differences between these two trial types could be attrib-
uted to differences in difficulty.
Task-modulation
Our first goal was to identify electrodes that were responsive and modulated by our task
manipulations. Electrodes that had significantly increased activity from a baseline of 0 to any
of the four trial types between 50 and 900 ms were identified using a timepoint-by-timepoint
Broadband gamma power:
Gives an indirect measure of the
aggregate surrounding neuronal
population firing, here calculated by
measuring the power in the
frequency range 70–170 Hz.
Neurobiology of Language
24
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
1
8
1
9
8
6
8
7
7
n
o
_
a
_
0
0
0
4
7
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Precentral gyrus and grapheme-phoneme conversion
t test corrected for temporal false-discovery rate at p < 0.05 (Benjamini & Hochberg, 1995).
Next, a one-way ANOVA was run between the four trial types from 50–900 ms at p < 0.01 tem-
porally corrected using a bootstrapped shuffling of trial identity 1,000 times (Maris & Oostenveld,
2007). Only electrodes which were significant in both these tests (i.e., with a significant increase
from baseline in BHG and a significant difference between trial types during this increase) were
included in further analysis. We refer to these as Task-Modulated electrodes.
Language-preference
Next we sought to understand if the evoked activity was related to language processing by
comparing visual and auditory language to their sensory controls. Task-Modulated electrodes
were assessed for whether they were responding preferentially to either visual or auditory
linguistic stimuli as evidenced by an increased response to letter-strings relative to false-font
stimuli from 50–450 ms (Text-Preference) or an increased response to bi-phonemes relative to
noise-vocoded stimuli from 450–900 ms (Phoneme-Preference). ANOVAs were run timepoint-
by-timepoint, once again corrected using the bootstrapped shuffling method. The Task
Modulation ANOVA results were used to mask significant Text-Preference and Phoneme-
Preference time-periods to ensure differences found between stimulus types were part of the
originally identified Task-Modulated temporal period.
Individual letter and phoneme identity sensitivity
A key question is the location of the cortical representation of sublexical linguistic units for
letters and the overlap of these letter representations with phoneme representations. An area
which contains such representations would be expected to have differential neural responses
based on letter/phoneme identity. To assess such sublexical representations, a 1-way ANOVA
was run timepoint-by-timepoint on Task-Modulated electrodes between consonant identity for
either letter-strings from 0–450 ms (Letter-Sensitive) or bi-phonemes from 450–900 ms
(Phoneme-Sensitive), temporally corrected using the bootstrapped shuffling method. The
“Task Modulated” ANOVA results were used to mask significant time-periods.
Mismatch effect
Successfully performing our match/mismatch task necessitates encoding phonemes presented
both visually and auditorily. If the same phonological representations are used by both sensory
modalities (i.e., strong overlap of phonological processing), then cortical patches containing
these representations should show differential processing depending on whether the visual
and auditory phonemes match or mismatch, due to repetition priming. In BHG, repetition
priming (i.e., re-encoding a recently encoded stimulus) evokes reduced power (McDonald
et al., 2010). This is likely because re-encoding a recently encoded stimulus evokes less neu-
ronal activity than the initial encoding (Gotts et al., 2012). Mismatch-sensitive electrodes were
therefore defined as having a larger BHG response to mismatch trials (i.e., non-primed trials)
than to matched trials (i.e., primed trials) during presentation of the bi-phoneme (i.e., from
500–900 ms) identified using a 1-way ANOVA temporally corrected using the bootstrapped
shuffling method (Mismatch). The Task Modulated ANOVA results were used to mask signif-
icant time-periods.
Preference for degraded speech
Behavioral studies have demonstrated text can improve the encoding of degraded speech, pre-
sumably through GPC (Frost, 1991; Frost & Kampf, 1993). To identify areas that may partici-
pate in this process, we sought to investigate how noise-vocoded preference overlaps with
Neurobiology of Language
25
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
3
1
1
8
1
9
8
6
8
7
7
n
o
_
a
_
0
0
0
4
7
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Precentral gyrus and grapheme-phoneme conversion
Text-Preference by identifying electrodes with greater responses to noise-vocoded stimuli ver-
sus bi-phonemes from 500–900 ms (Noise-Preference), using the same procedure detailed for
Phoneme-Preference electrodes. The Task Modulated ANOVA results were used to mask sig-
nificant time-periods.
Overlap Comparisons
We also sought to characterize whether visual and auditory effects within a region were
observed in separate or in overlapping electrodes. This was operationalized as an examination
of the overlap between Text-Preference effects and auditory effects (Phoneme-Preference,
Mismatch, Noise-Preference) using the binomial test of overlap percentages compared to
chance. Because overlap within a region could happen by chance, we took the percentage
of Text-Selective electrodes within a parcellation region as the baseline chance. For example,
in the Precentral parcellation, 16 out of 54 electrodes (~30%) displayed Text-Preference
effects, and therefore, randomly distributed auditory effects in this same region would be
expected to overlap with 30% of the Text-Preference effects by chance. A significant (i.e.,
p < 0.05 on the binomial test) increase in overlap above this baseline percentage would be
evidence of overlap. This comparison was made across all included electrodes across a hemi-
sphere as well as within the regions of interest.
Regional Comparisons
Comparisons between regions are difficult in iEEG due to sparse coverage with variation due
to clinical considerations. However, studies with large numbers of patients note that responses
occur in similar regions relative to neuroanatomical landmarks (Ojemann et al., 1989). Our
study made use of nonparametric statistics to compare both proportion of electrodes and tim-
ing of effect onsets grouped into the broad Desikan atlas (Desikan et al., 2006) parcellations.
To compare distribution across putative articulatory or putative encoding cortex we per-
formed planned a priori comparisons between the precentral parcellation and the STG, supra-
marginal, and pars opercularis parcellations. A Fisher’s Exact test was used to compare
proportions across regions. With three comparisons (i.e., precentral gyrus versus STG, supra-
marginal gyrus, and pars opercularis), the Bonferroni corrected p value threshold was p <
0.016. We also compared effect onset timings between regions using a Ranksum nonparamet-
ric test, when possible. The effect onset was defined as the first timepoint displaying a signif-
icant effect after correction for temporal multiple comparisons. The variable number of effects
per parcellation does cause differences in power between comparisons; for example, a critical
question is the timing of effects in the precentral gyrus versus the STG and pars opercularis, but
while the precentral gyrus had 16 Text-Preference electrodes and the pars opercularis had 7,
the STG contained only 3. However, despite these difficulties some temporal regularities
emerged. These timing analyses will mirror the a priori structure of the regional distribution
analyses and share their p value corrections.
Broadband High Gamma Amplitude
Broadband high gamma amplitude gives an indirect measure of the aggregate surrounding
neuronal population firing (Ray et al., 2008). To understand whether amplitude differed
between regions, we calculated max amplitude of language trials for letter-string presentation
and bi-phoneme presentation within Task-Modulated electrodes. To assess amplitude the
average waveform for language was z scored relative to its baseline (the same time window
Neurobiology of Language
26
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
3
1
1
8
1
9
8
6
8
7
7
n
o
_
a
_
0
0
0
4
7
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Precentral gyrus and grapheme-phoneme conversion
Phase-locking value:
Measures the consistency of the
relative phase of frequencies within
the electrophysiological signal
between two electrodes.
as used for baselining in data analysis). Then the highest value was found for each electrode
for the letter-string time window (0–450 ms) and the bi-phoneme time window (450–900 ms).
Connectivity
To test for a putative network between the electrodes displaying effects of interest, we used
phase-locking value (PLV) calculated pairwise between electrodes as described in Lachaux
et al. (1999). PLV measures the consistency of the relative phase of frequencies within the
electrophysiological signal. High PLV indicates consistent synchronization of the synaptic cur-
rents in pyramidal apical dendrites between the cortical locations underlying the intracranial
sensors. For this study we measured frequencies from 4 Hz to 12 Hz. This frequency range was
chosen based on a prior paper which found strong phase locking in this frequency range within
the reading network (Thesen et al., 2012). Neural activity in this lower frequency range is
associated with feedforward and feedback activity across coordinating neural networks during
visual language processing (Halgren et al., 2015). To understand whether the PLV values we
obtained in Task-Modulated electrodes were greater than would be expected by random
chance, we created a distribution of baseline PLV values for each individual patient. This dis-
tribution was obtained by taking the max PLV value from a baseline period (−200–0 ms) for all
electrode pairs within a patient. A PLV value in the task window for the Task-Modulated elec-
trodes of interest (100–1,300 ms) was considered significant (i.e., greater than would be ex-
pected by chance) if it was p < 0.00005 based on the subject’s own baseline PLV distribution.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
3
1
1
8
1
9
8
6
8
7
7
n
o
_
a
_
0
0
0
4
7
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
RESULTS
Behavior
Average proportion correct for Match (89%) and Mismatch (92%) trials was not significantly
different ( p > 0.05; Figure 1B). Likewise, response time for Match (660 ms) and Mismatch
(678 ms) trials was not significantly different ( p > 0.05). Therefore, there is no behavioral evidence
for difficulty differences in performing either judgement. For the control trials, behavioral perfor-
mance was similar to other trial types for False-Font (94%) and Noise-Vocoded (92%) trials.
Task-Modulation Effect Distribution
In discussing results for the following sections, we will report results during the letter-string pre-
sentation and/or during bi-phoneme presentation. Results during the letter-string presentation
relate to the first hypothesis that GPC evokes activity in the precentral gyrus and the second
hypothesis that the timing of this activity will be consistent with precentral gyrus contributions
to GPC. Results during the bi-phoneme presentation will relate to the third hypothesis regarding
whether visual and auditory phonological processing are overlapping.
First we characterized the regions responding to and modulated by the match/mismatch
task. Figure 1C–D displays the distribution and number of electrodes per parcellation.
Supplementary Figure 1 displays electrode distribution color-coded by individual patient.
(Supporting information can be found online at https://doi.org/10.1162/nol_a_00047). Figure 2
displays the distribution and proportions of Task-Modulated electrodes. Supplementary Figure 2
displays the proportions of Task-Modulated electrodes for each individual patient. Table 2 dis-
plays left hemisphere distribution of Task-Modulated electrodes. During the letter-string pre-
sentation, these effects (i.e., significant ANOVA differences between the four stimulus
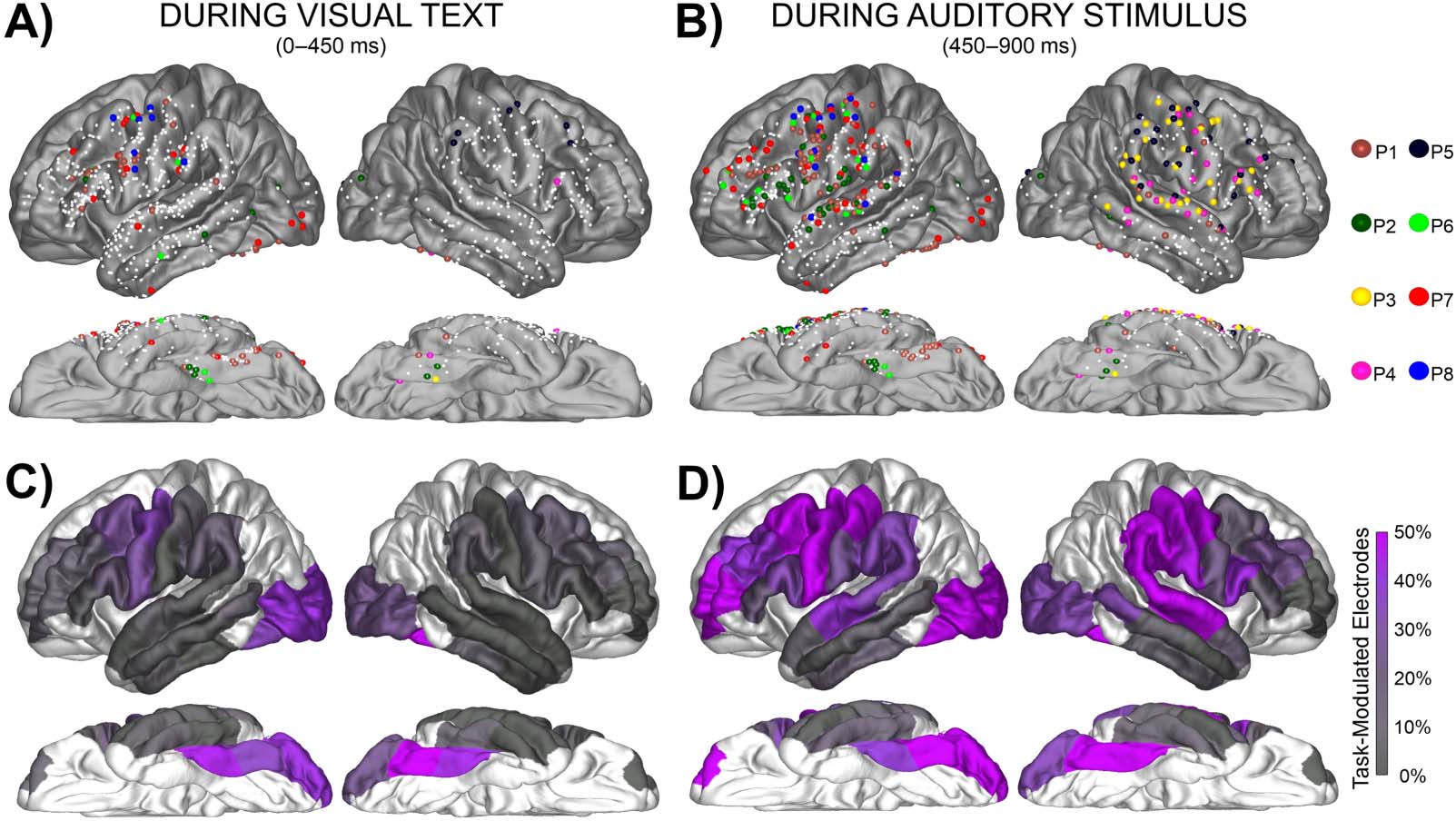
conditions as well as an overall increase above baseline) were overall left- (17%) versus right-
lateralized (8%; Fisher’s Exact Test [FET]: p < 0.005). Activity was more widespread during
Neurobiology of Language
27
Precentral gyrus and grapheme-phoneme conversion
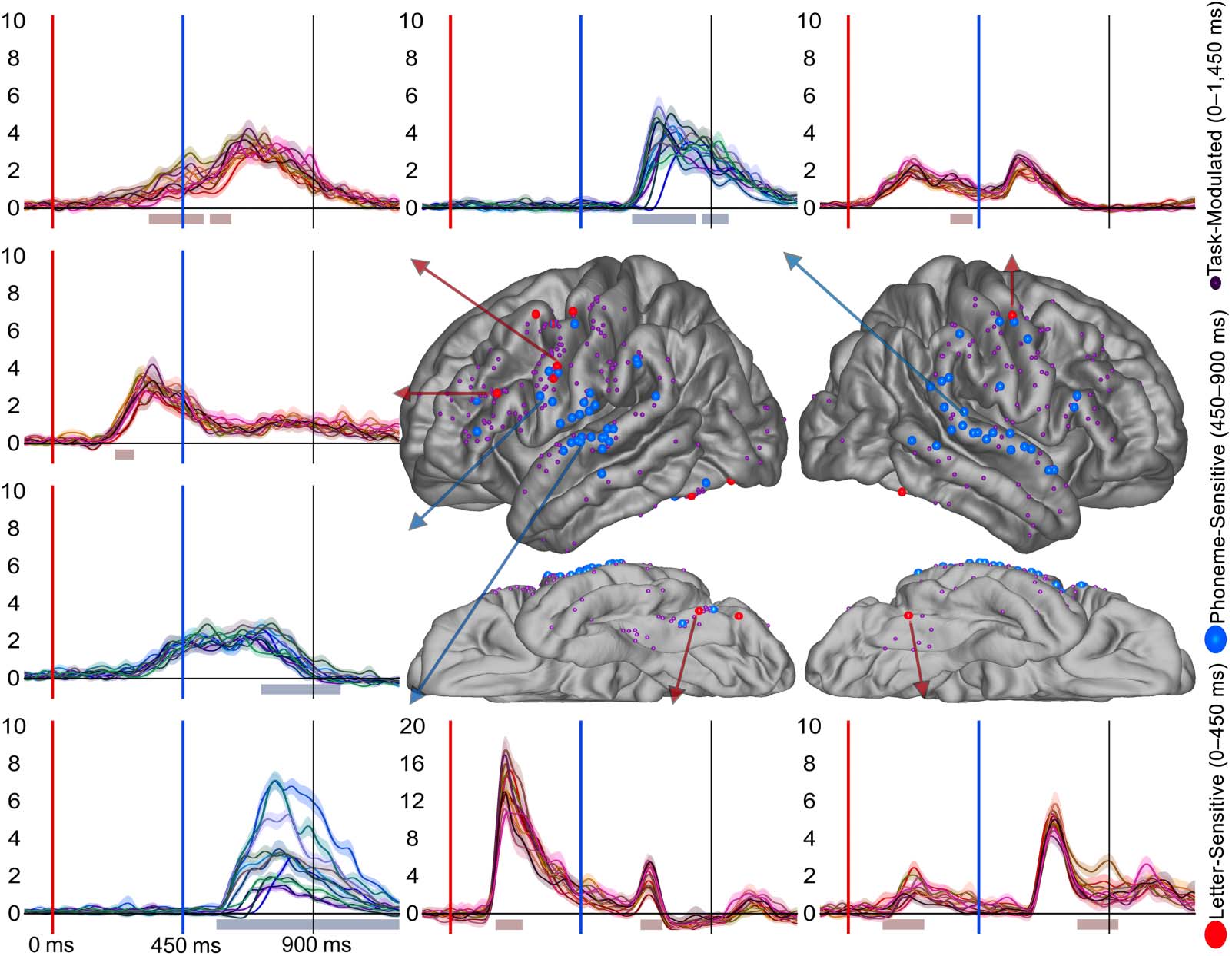
Figure 2. Task-Modulated electrode distribution across the cortex. (A–B) Electrodes demonstrating both a significant increase from baseline
and a significant difference between the four trial types (Match, Mismatch, Visual Control, Auditory Control) during text presentation from
0–450 ms and during bi-phoneme presentation from 450–900 ms. Smaller white dots represent electrodes recorded from which did not
meet criteria for being Task-Modulated (i.e., either did not show task-evoked activity or did not show modulation based on stimulus type).
Each electrode is color-coded for individual patient. (C–D) Electrodes meeting criteria for a Task-Modulated effect displayed as percentages
out of total electrodes in a ROI.
bi-phoneme presentation and there was no left- (59%) versus right-lateralization (63%; FET:
p > 0.05). Due to the left-lateralization of the reading network (Carreiras et al., 2014; Fiez
& Petersen, 1998; Jobard et al., 2003; Price, 2012; Taylor et al., 2013) and the left-lateralized
results during letter-string presentation, we will be focusing on left hemisphere comparisons
moving forward.
Related to our first hypothesis that this GPC task would evoke at least as much activity in the
precentral as the surrounding perisylvian regions, we found that during letter-string presenta-
tion, the left precentral gyrus had at least as great a proportion of Task-Modulated electrodes as
the other temporal-parietal parcellations. The precentral parcellation had a greater proportion
of Task-Modulated electrodes during letter-string presentation (31%, 4 of 5 patients) compared
to the STG (4%, 2 of 5 patients; FET: p < 0.001) and not a significantly different proportion
than the supramarginal (15%, 4 of 5 patients; FET: p < 0.05) or the pars opercularis (21%, 5 of
5 patients; FET: p > 0.05). In preparation for characterizing overlap (third hypothesis), we iden-
tified many electrodes during bi-phoneme presentation which demonstrated Task-Modulated
effects. During bi-phoneme presentation, the precentral gyrus (70%, 5 of 5 patients) had a
greater proportion of Task-Modulated electrodes than the STG (38%, 5 of 5 patients; FET:
p < 0.001), supramarginal (34%, 4 of 5 patients; FET: p < 0.001), and pars opercularis
(33%, 4 of 5 patients; FET: p < 0.001). Because proportion of electrodes displaying an effect
is just one way to compare activity across regions, we also include Supplementary Figure 3,
which displays the max BHG amplitudes for Task-Modulated electrodes across regions.
Language-Preference Effects
Having established the strong modulation of the activity in the precentral parcellation by task
stimuli, next we characterized which stimuli evoked responses in the precentral (Figure 3).
Neurobiology of Language
28
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
3
1
1
8
1
9
8
6
8
7
7
n
o
_
a
_
0
0
0
4
7
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Precentral gyrus and grapheme-phoneme conversion
Table 2. Distribution of Task-Modulated electrodes in each left-hemisphere region during letter-string presentation (0–450 ms) and bi-
phoneme presentation (450–900 ms)
Region
Lateral occipital
Fusiform
Precentral
Postcentral
MTG
STG
Supramarginal
Pars opercularis
Pars triangularis
Middle frontal
Task-Modulated (0–450 ms)
42% (5 / 12)
39% (15 / 38)
31% (17 / 54)
4% (2 / 50)
5% (2 / 43)
4% (3 / 74)
15% (6 / 41)
21% (8 / 39)
5% (2 / 38)
12% (5 / 43)
Left hemisphere
Subjects
3 / 3
4 / 4
3 / 5
1 / 5
1 / 5
2 / 5
4 / 5
5 / 5
1 / 5
2 / 5
Task-Modulated (450–900 ms)
50% (6 / 12)
50% (19 / 38)
70% (38 / 54)
40% (20 / 50)
7% (3 / 43)
38% (28 / 74)
34% (14 / 41)
33% (13 / 39)
21% (8 / 38)
40% (17 / 43)
Subjects
3 / 3
3 / 4
5 / 5
4 / 5
2 / 5
5 / 5
4 / 5
4 / 5
4 / 5
3 / 5
Note. Task-Modulated columns: #% (#/#) = proportion of electrodes with Task-Modulated effect (electrodes showing effect / total electrodes). Subjects columns:
# / # = number of patients with ≥1 electrode showing effect in region / total patients with electrodes in region.
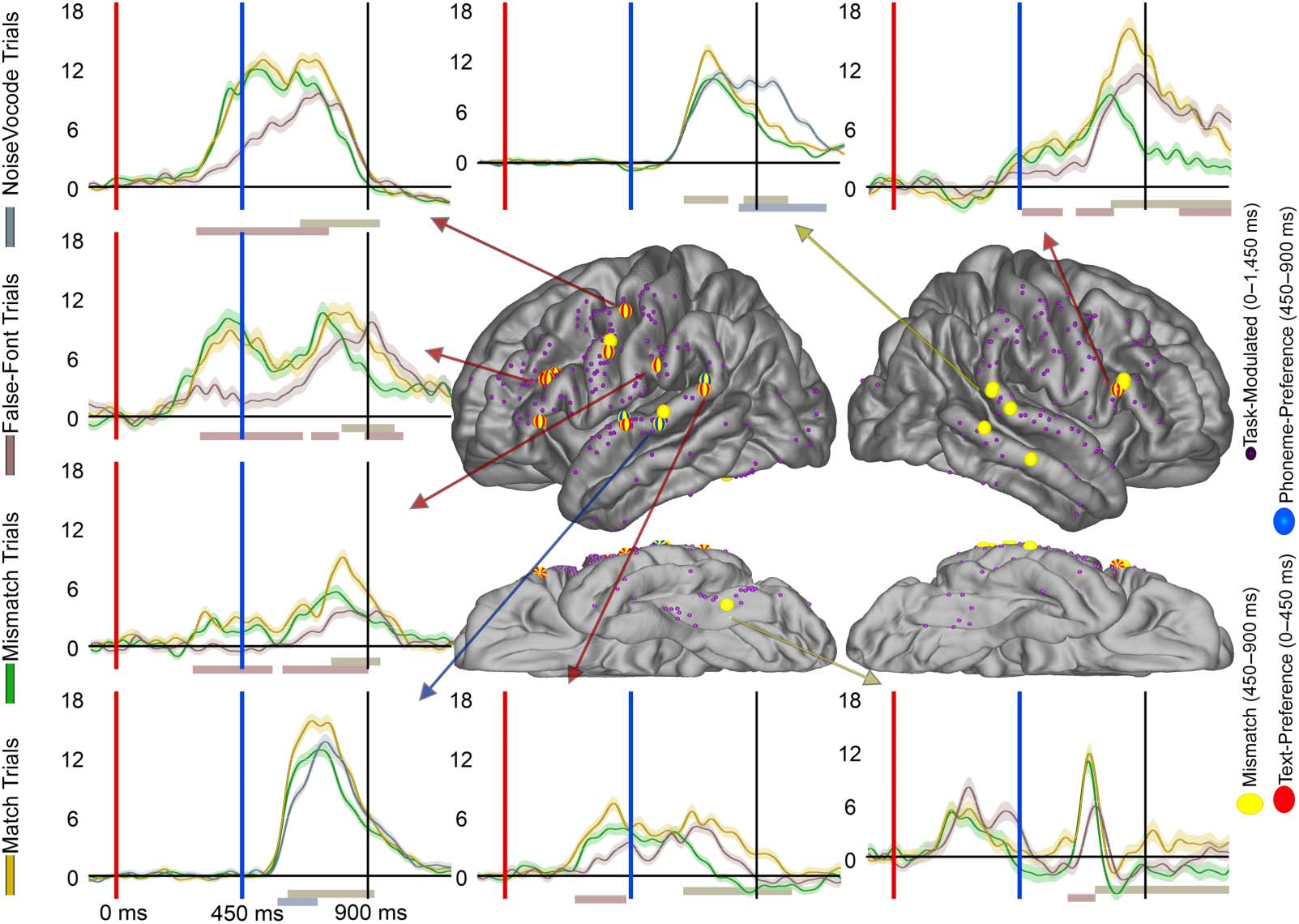
Table 3 displays the distribution electrodes displaying effects. During the letter-string presen-
tation we examined the Text-Preference effect (i.e., BHG to letter-string significantly greater
than to false fonts) to understand if these effects were at least as consistently found in the pre-
central gyrus as in other perisylvian regions (hypothesis 1) and in a time window consistent with
GPC (hypothesis 2). In the a priori left-hemisphere comparison, the precentral parcellation had
more Text-Preference electrodes (30%, 4 of 5 patients) than the STG (4%, 2 of 5 patients; FET:
p < 0.001) and was not significantly different from the supramarginal (15%, 4 of 5 patients; FET:
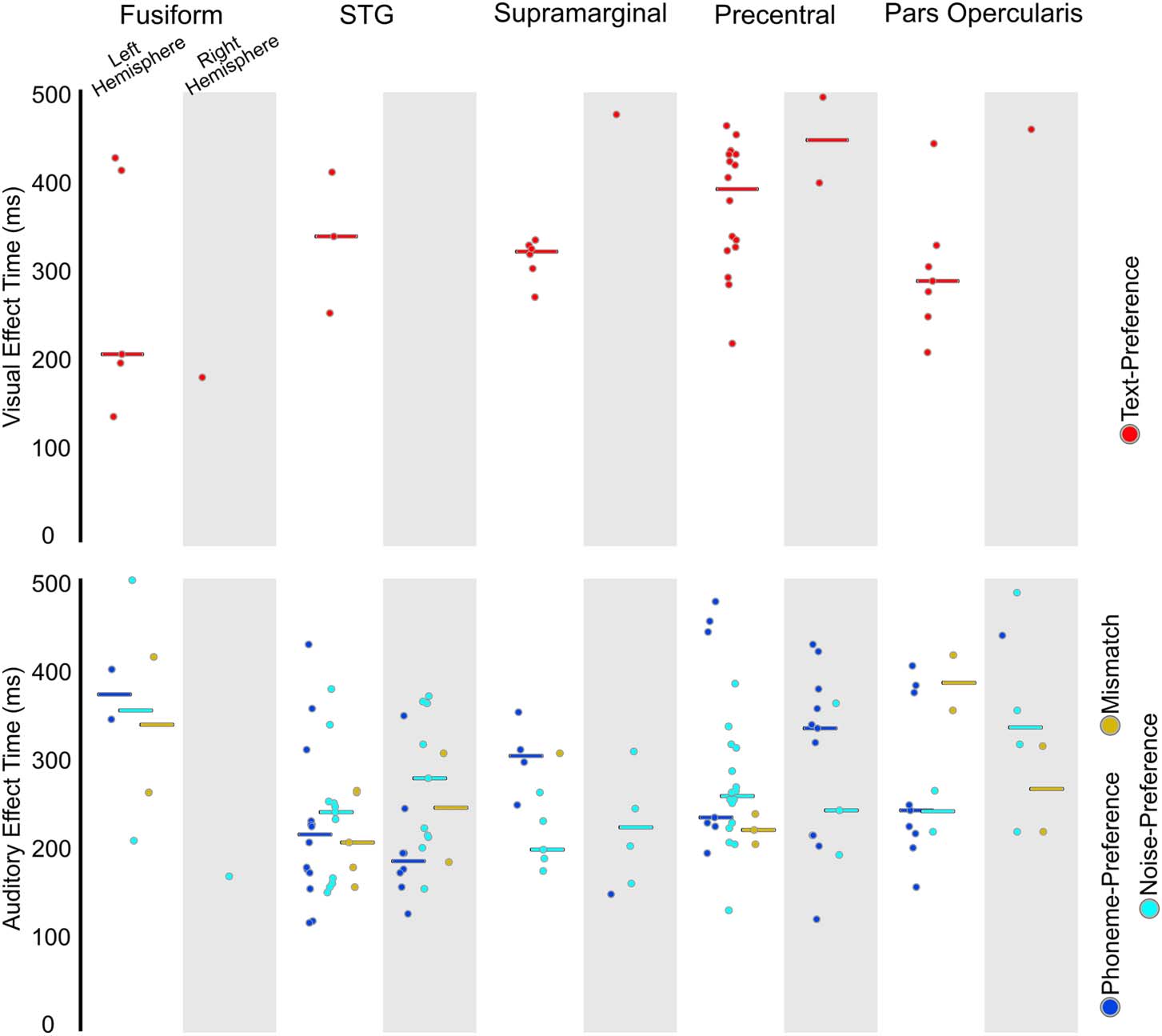
p < 0.05) or pars opercularis (18%, 4 of 5 patients; FET: p > 0.05). For effect onset timing
(Figure 4 and Table 4), there were no significant differences in Text-Preference onsets between
the precentral gyrus (Earliest (1st): ~220 ms; Mdn: ~390 ms) and the pars opercularis (1st:
~200 ms; Mdn: ~280 ms; RankSum (Rs): p > 0.05), supramarginal (1st: ~260 ms; Mdn: ~320 ms;
Rs: p > 0.05), or STG (1st: ~240 ms; Mdn: ~340 ms; Rs: p > 0.05). Therefore, there was consistent
timing across regions during the time period associated with GPC, with all regions demonstrating
effects during the GPC relevant time period.
Here we also report Phoneme-Preference electrodes identified during the bi-phoneme pre-
sentation, which will be important to later considerations of overlap (hypothesis 3; explored in
the Results section, Overlap of Text-Preference Electrodes With Auditory Effects). In the left-
sided a priori comparisons, there were no significant differences between the precentral (13%,
4 of 5 patients) and the STG (18%, 3 of 5 patients; FET: p > 0.05), supramarginal (11%, 1 of
5 patients; FET: p > 0.05), or pars opercularis (23%, 4 of 5 patients; FET: p > 0.05) parcella-
tions. Supplementary Figure 2 displays the proportions of Text-Preference and Phoneme-
Preference electrodes for each individual patient.
Neurobiology of Language
29
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
1
8
1
9
8
6
8
7
7
n
o
_
a
_
0
0
0
4
7
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Precentral gyrus and grapheme-phoneme conversion
Figure 3. Text-Preference and Phoneme-Preference electrode distribution across the cortex. Display of electrode location (approximate,
morphed to an average brain for display purposes) for Text-Preference (red), Phoneme-Preference (blue), and both effects (orange) electrodes.
Smaller electrodes represent sites that were Task-Modulated (dark purple) but did not prefer language stimuli to controls. Arrows from specific
electrodes are color-coded for the electrodes effect and point to examples illustrating typical waveforms for each region. Shaded regions
surrounding the average waveforms reflect the standard error of the mean of the averaged trials. Vertical axis for BHG is in arbitrary units.
The red shaded region at plot bottom highlights a significant Text-Preference effect period (letter-string > false-font) and the bluish bar high-
lights a significant Phoneme-Preference effect period (bi-phoneme > noise-vocoded). Analyses were temporally corrected using a boot-
strapped shuffling of trial identity 1,000 times.
Letter- & Phoneme-Sensitive Effects
Next, we sought to determine if there were any electrodes which responded differentially
based on letter identity (i.e., Letter-Sensitive electrodes) and what the distribution of the elec-
trodes was (hypothesis 1) and the timing of these effects (hypothesis 2). There were 8 Letter-
Sensitive electrodes found. Though sparse, electrodes with effects were in expected ventral
visual regions but more surprisingly also in frontal regions (Figure 5). The electrodes were
in the ventral occipital-temporal regions (1 in the left lateral occipital, 1 in the left caudal
fusiform, and 1 in the right caudal fusiform) and the lateral frontal (3 in the left precentral gyrus
across 3 of the 5 patients; 1 in the left pars opercularis). While the greatest number of Letter-
Sensitive electrodes were in the precentral parcellation it must be noted that in terms of
proportion, the greatest proportion of Letter-Sensitive electrodes were found in the occipital-
temporal regions as would be expected. Due to the limited number of electrodes, regional
statistical comparisons were not informative. The lack of Letter-Sensitive effects is not surpris-
ing as the main region implicated in graphemic encoding, the ventral visual pathway centered
on the posterior fusiform, was not well covered in this study. The earliest Letter-Sensitive effect
was in the left caudal fusiform at ~180 ms. The precentral Letter-Sensitive electrodes had a
qualitatively later onset time (1st: ~340 ms; Mdn: ~420 ms). This further demonstrates the
presence of reading-related effects in the precentral gyrus at a timing qualitatively a bit later
than Text-Preference effects, but still during the letter-string presentation.
We next sought to understand the overlap of these Letter-Sensitive electrodes with Phoneme-
Sensitive electrodes (hypothesis 3). Phoneme-Sensitive electrodes were more numerous,
Neurobiology of Language
30
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
1
8
1
9
8
6
8
7
7
n
o
_
a
_
0
0
0
4
7
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Precentral gyrus and grapheme-phoneme conversion
Region
Lateral occipital
Fusiform
Precentral
Postcentral
MTG
STG
Supramarginal
Pars opercularis
Pars triangularis
Middle frontal
Table 3. Distribution of electrodes displaying each effect divided into region in the left hemisphere
Selective
electrodes
9
Text-
Selective
0
Letter-
Sensitive
1
Voice
Selective
0
Noise-Vocoded
Selective
1
Phoneme-
Sensitive
0
Incongruent
effects
0
24
39
25
5
30
19
19
12
24
5
16
1
0
3
6
7
1
4
1
3
0
0
0
0
1
0
0
1
7
8
1
13
4
9
5
6
1
16
5
0
10
5
2
0
1
2
8
10
1
14
2
0
1
0
2
3
0
0
5
1
2
1
1
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
1
8
1
9
8
6
8
7
7
n
o
_
a
_
0
0
0
4
7
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Note. # = number of electrodes in each region displaying the relevant effect.
totaling 35 electrodes, with the highest proportion in STG. There were significantly more
Phoneme-Sensitive electrodes in the precentral (15%, 3 of 5 patients) than the pars opercularis
(0%, 0 of 5 patients; p = 0.01) but no difference between the precentral gyrus and the STG (19%,
3 of 5 patients; p > 0.05) or supramarginal (5%, 1 of 5 patients; p > 0.05). For the overlap,
we note that the only region with both a Letter-Sensitive and a Phoneme-Sensitive effect in the
same electrode was the precentral gyrus, though this was observed in only a single electrode.
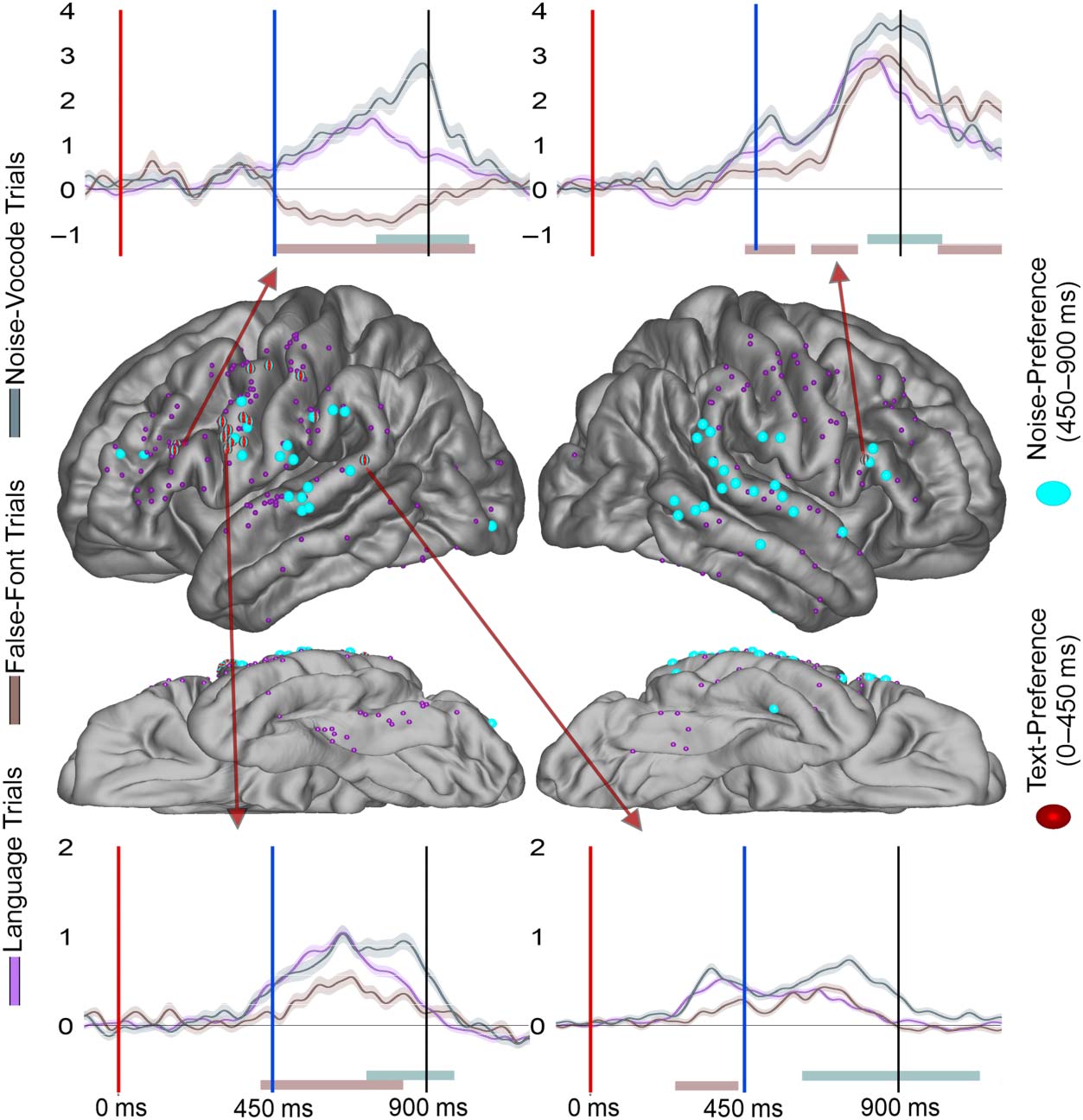
Mismatch Effects
Next, we focused on activity during the bi-phoneme presentation to understand the distribution
of electrodes which showed evidence of cross-modal phonological priming. These effects could
be evidence of overlapping visually-encoded and auditorily-encoded phonological representa-
tions (hypothesis 3). However, Mismatch effects were sparse, totaling 15 electrodes (Figure 6).
These effects were concentrated mainly in the perisylvian regions of interest including the STG
(5), precentral (3), pars opercularis (2), and supramarginal (1). Two electrodes with a Mismatch
effect were found in the fusiform as well. The earliest Mismatch effect was in the STG at ~160 ms
post bi-phoneme presentation, with the median of the STG and precentral both occurring at
~220 ms post bi-phoneme presentation. This was followed by the medians of the supramarginal
(Mdn: ~300 ms), fusiform (Mdn: 340 ms), and pars opercularis (Mdn: ~390 ms).
Overlap of Text-Preference Electrodes With Auditory Effects
As a second assay into the relationship of visual language encoding to auditory language en-
coding (hypothesis 3), next we assessed overlap of visual and auditory language encoding by
comparing Text-Preference electrodes’ overlap with three effects during the bi-phoneme
Neurobiology of Language
31
Precentral gyrus and grapheme-phoneme conversion
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
1
8
1
9
8
6
8
7
7
n
o
_
a
_
0
0
0
4
7
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4.
Effect onset timing from the letter-string presentation (top) and bi-phoneme presentation
(bottom). Circles represent the onset of a significant effect at an individual electrode within each
region. The top panel displays onset of effects from the start of letter-string presentation (i.e., starting
from 0 ms) for Text-Preference (red) for the left hemisphere (not shaded) and right hemisphere
(shaded). The bottom panel displays onset of effects from bi-phoneme onset (i.e., starting from
450 ms) for Phoneme-Preference (blue), Noise-Preference (cyan), and Mismatch effects (yellow)
for the left hemisphere (not shaded) and right hemisphere (shaded).
presentation: Phoneme-Preference electrodes (Figure 3), Mismatch electrodes (Figure 6), and
Noise-Preference electrodes (Figure 7). Because overlap within a region could happen by
chance, we took the proportion of Text-Selective electrodes within a parcellation region as
the baseline and compared this to the observed proportion.
For Phoneme-Preference effect electrodes, there was a significant overlap of Text-
Preference with Phoneme-Preference electrodes (observed overlap (O): 25%, expected
overlap (E): 10%; binomial test (BT): p < 0.001). However, at the regional level none of the
perisylvian regions reached significance, though all showed numerically greater overlap than
would be expected by chance: precentral (O: 43%, E: 30%; BT: p > 0.05), STG (O: 8%, E: 4%;
BT: p > 0.05), supramarginal (O: 44%, E: 15%; BT: p > 0.05), and pars opercularis (O: 50%, E:
18%; BT: p > 0.05). For Mismatch effect electrodes, there was a significant overlap of Text-
Preference and Mismatch electrodes (O: 37%, E: 10%; BT: p < 0.001). However, again no
individual parcellation reached significance; the precentral parcellation (O: 20%, E: 31%;
BT: p > 0.05), STG (O: 33%, E: 4%; BT: p < 0.05), and the pars opercularis (O: 100%, E:
18%; BT: p < 0.05) were all non-significant. For Noise-Preference electrodes, there was a
significant overlap of Text-Preference and Noise-Preference electrodes (O: 37%, E: 10%;
Neurobiology of Language
32
Table 4. Median and range of regional effect onsets in the left hemisphere
Onset from text presentation (0 ms)
Text-Selective
200 ms (140–420)
180 ms (180)
Letter-Sensitive
Voice-Selective
370 ms (340–400)
Onset from auditory presentation (450 ms)
Phoneme-Sensitive
Vocode-Preference
440 ms (440–440)
360 ms (220–500)
Incongruent
340 ms (260–420)
Region
Fusiform
Lateral occipital
MTG
STG
–
–
340 ms (240–400)
Supramarginal
320 ms (260–320)
–
–
–
260 ms (260)
–
440 ms (440)
–
200 ms (200)
–
200 ms (200)
–
–
220 ms (120–420)
240 ms (160–380)
180 ms (120–440)
220 ms (160–260)
310 ms (260–360)
200 ms (180–260)
260 ms (180–340)
300 ms (300)
Precentral
390 ms (220–460)
420 ms (340–420)
240 ms (200–480)
260 ms (140–380)
280 ms (140–360)
220 ms (220–240)
Pars triangularis
420 ms (420)
–
280 ms (180–380)
–
300 ms (300)
380 ms (380)
Pars operculatris
280 ms (200–440)
220 ms (220)
240 ms (160–400)
240 ms (220–260)
–
390 ms (360–420)
Note. #ms (#–#) = median effect onset in ms (earliest effect onset in ms − latest effect onset in ms).
P
r
e
c
e
n
t
r
a
l
g
y
r
u
s
a
n
d
g
r
a
p
h
e
m
e
-
p
h
o
n
e
m
e
c
o
n
v
e
r
s
i
o
n
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
3
1
1
8
1
9
8
6
8
7
7
n
o
_
a
_
0
0
0
4
7
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
N
e
u
r
o
b
o
o
g
y
i
l
o
f
L
a
n
g
u
a
g
e
3
3
Precentral gyrus and grapheme-phoneme conversion
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
3
1
1
8
1
9
8
6
8
7
7
n
o
_
a
_
0
0
0
4
7
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 5.
Letter-Sensitive and Phoneme-Sensitive electrode distribution across the cortex. Display of electrode location (approximate,
morphed to an average brain for display purposes) for electrodes differentially responsive to individual letters (red) or phonemes (blue).
Smaller electrodes represent sites that were Task-Modulated (dark purple) but did not demonstrate letter or phoneme identity sensitivity.
Arrows from specific electrodes are color-coded for the electrode’s effect and point to examples illustrating typical waveforms for each region.
Shaded regions surrounding the average waveforms reflect the standard error of the mean of the averaged trials. Each different line color
represents the average response to one of the 12 consonant letters (reddish colors) or 12 consonant phonemes (bluish colors). Vertical axis
for BHG is in arbitrary units. The red bar at plot bottom highlights a significant Letter-Sensitivity effect period and the blue bar highlights a
significant Phoneme-Sensitivity effect period. Analyses were temporally corrected using a bootstrapped shuffling of trial identity 1,000 times.
BT: p < 0.001; Figure 7). The perisylvian follow-up analyses identified the Precentral as having
a significant relationship (O: 63%, E: 31%; BT: p = 0.01) but not the STG (O: 10%, E: 4%; BT:
p > 0.05), supramarginal (O: 20%, E: 15%; BT: p > 0.05), or pars opercularis (O: 50%, E: 18%;
BT: p > 0.05). Taken together, Text-Preference electrodes showed significantly greater overlap
with auditory effects than would be predicted by chance across perisylvian regions, though in
individual regions this rarely reached significance. We note that while the Text-Preference
overlap was significantly greater than chance, the proportion of total overlap across the 3 audi-
tory effects (25–37%) demonstrated that many electrodes showed only Text-Preference or only
auditory effects.
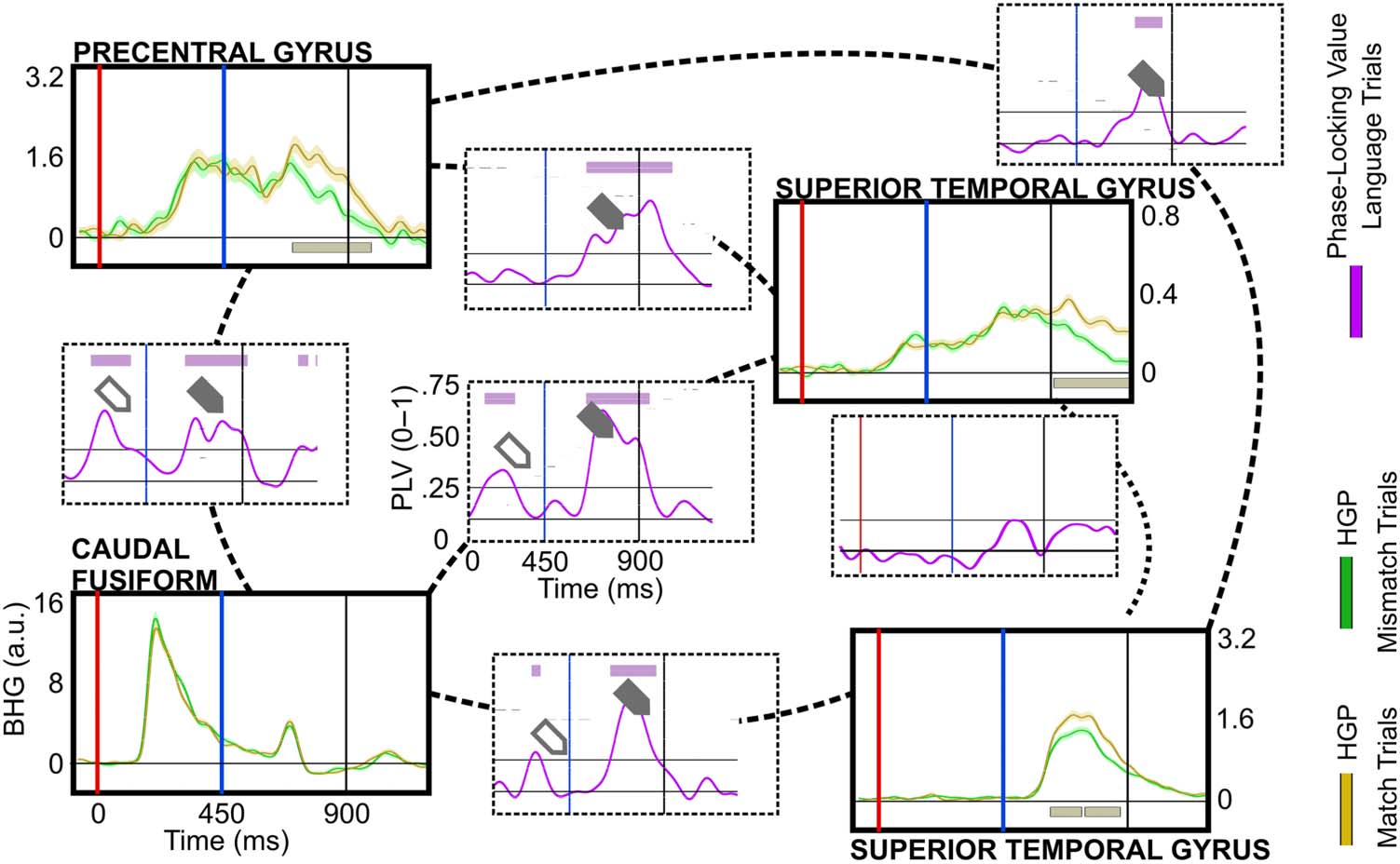
Connectivity Results
Finally, we probed the possible networks involving the dorsal route using PLV (P1’s PLV results
are displayed in Figure 8). During letter-string presentation, we characterized the distribution
(hypothesis 1) and timing (hypothesis 2) of connectivity across two subnetworks: (1) a caudal
fusiform to perisylvian subnetwork to capture connectivity between the ventral and dorsal
reading routes, and (2) a precentral gyrus to perisylvian subnetwork. If the precentral gyrus
Neurobiology of Language
34
Precentral gyrus and grapheme-phoneme conversion
Figure 6. Mismatch effect electrode distribution across the cortex. Electrodes displayed on the brain for the Mismatch effect (yellow) across
the cortex. Overlapping Text-Preference (red) and Phoneme-Preference (blue) effects are noted with stripes. Smaller electrodes represent sites
that were Task-Modulated (dark purple) but did not demonstrate a Mismatch effect. Arrows from specific electrodes are color-coded for the
electrodes effect and point to illustrating typical waveforms for each region. Shaded regions surrounding the average waveforms reflect the
standard error of the mean of the averaged trials. Vertical axis for BHG is in arbitrary units. The red bar at plot bottom highlights a significant
Letter-Preference effect period, the blue bar highlights a significant Phoneme-Preference effect period, and the yellow bar highlights a signif-
icant Mismatch effect period. Analyses were temporally corrected using a bootstrapped shuffling of trial identity 1,000 times.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
1
8
1
9
8
6
8
7
7
n
o
_
a
_
0
0
0
4
7
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
is involved in GPC during reading, then its pattern of connectivity and the timing of this
activity should include a relationship with either the fusiform (direct connection from the
ventral reading route to the precentral) or from surrounding perisylvian regions (indirect con-
nection from the ventral reading route to the precentral) during letter-string presentation.
For the caudal fusiform to perisylvian subnetwork during the letter-string presentation, 2
patients had a Task-Modulated electrode in the left caudal fusiform. Both patients had a
~180 ms onset of Text-Preference effect, a timing associated with both fusiform orthographic
processing (Hirshorn et al., 2016; Thesen et al., 2012) and widespread processing across the
cortex (Halgren, 1990). The timing and location of these Text-Preference effects mark both
patients’ electrodes as excellent assays into the temporal dynamics of the caudal fusiform
orthographic hub’s relationship with the dorsal route. During letter-string presentation both
electrodes showed significant PLV with precentral electrodes (P1 PLV-onset: ~200 ms; P7
PLV-onset: ~180 ms) and with an electrode in the supramarginal gyrus (P1: ~400 ms; P7:
~200 ms). But only 1 patient had significant PLV between the caudal fusiform and the STG
(P1: ~180 ms) and pars opercularis (P1: ~180 ms). These patients’ BHG and PLV data are illus-
trated side-by-side in Supplementary Figure 4.
For the precentral to perisylvian subnetwork, 4 patients had Task-Modulated effects in the
left precentral parcellation. These 4 patients displayed a muted connectivity between
Neurobiology of Language
35
Precentral gyrus and grapheme-phoneme conversion
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
1
8
1
9
8
6
8
7
7
n
o
_
a
_
0
0
0
4
7
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 7. The overlap of Text-Preference and Noise- Preference effects. Display of electrode lo-
cation (approximate, morphed to an average brain for display purposes) for Noise-Preference (cyan)
electrodes. Preceding overlapping Text-Preference (red) effects are noted with red stripes. Smaller
electrodes represent sites that were Task-Modulated (dark purple) but did not demonstrate a
Noise-Preference effect. Arrows from specific electrodes point to an example typical waveform
for each region. Shaded regions surrounding the average waveforms reflect the standard error of
the mean of the averaged trials. Vertical axis for BHG is in arbitrary units. The red bar at plot bottom
highlights a significant Letter-Preference effect period and the cyan bar highlights a significant
Noise-Preference effect period. Analyses were temporally corrected using a bootstrapped shuffling
of trial identity 1,000 times.
precentral electrodes and surrounding perisylvian regions during letter-string presentation,
with only 1 patient displaying late significant PLV between the precentral and an electrode
in the pars opercularis, (P1: ~380 ms) and an electrode in the STG (P1: ~420 ms). No patients
had electrodes which displayed significant PLV between the precentral and supramarginal.
Therefore, there is evidence of a direct connection between the ventral reading route and
the precentral gyrus during reading, at a timing consistent with the onset of phonological pro-
cessing in the dorsal perisylvian reading network.
Next, we examined the pattern of connectivity between perisylvian regions during the bi-
phoneme presentation to understand how this functional network evolved when processing
both visually and auditorily encoded phonemes (hypothesis 3). During the bi-phoneme pre-
sentation the precentral electrodes displayed more widespread PLV with surrounding perisyl-
vian electrodes than during letter-string presentation. There were 3 out of 4 patients with
significant PLV between electrodes in the precentral and STG (P1: ~80 ms; P2: ~180 ms;
P7: ~60 ms) and between the electrodes in the precentral and supramarginal (P1: ~60 ms;
Neurobiology of Language
36
Precentral gyrus and grapheme-phoneme conversion
Illustration of a proposed grapheme-to-phoneme reading network in 4 electrodes from
Figure 8.
patient P1. Plots of waveforms in solid-line boxes illustrate broadband high gamma differences be-
tween Match (green) and Mismatch (yellow) trials. Shaded regions surrounding the average wave-
forms reflect the standard error of the mean of the averaged trials. The yellow bar at the plot bottom
signifies periods of significant Mismatch effect. Analyses were temporally corrected using a boot-
strapped shuffling of trial identity 1,000 times. Plots in dotted-line boxes illustrate phase-locking
values (PLV) over time between electrodes for letter-string (purple) trials with the grey line the sig-
nificance threshold for PLV for this subject, and the purple bar at the top showing periods of sig-
nificant PLV. The open arrow highlights significant PLV during letter-string presentation; the filled
arrow signifies significant PLV during bi-phoneme presentation. The red line at 0 ms denotes
letter-string onset and the blue line at 450 ms denotes bi-phoneme onset.
P7: ~60 ms; P8: ~220 ms). Between the precentral and pars opercularis, 2 out of the 4 patients
displayed significant PLV (P1: ~160 ms; P2: ~160 ms).
DISCUSSION
Though theories of GPC have implicated articulatory phonemes, the place of the precentral
gyrus in neuroanatomical models of reading is ambiguous (Carreiras et al., 2014; Fiez &
Petersen, 1998; Taylor et al., 2013). Here we present evidence from a bimodal phonological
match/mismatch task supporting a role for the precentral gyrus in mediating visual and audi-
tory phonology. The presence of Text-Preference, Letter-Sensitive, and Mismatch effects oc-
curred at rates at least as frequently in the precentral gyrus compared to other perisylvian
regions cited by neurobiological models as involved in GPC. Further, the precentral had sig-
nificant connectivity with the caudal fusiform during letter-string presentation and significant
connectivity with adjoining temporal-parietal regions during bi-phoneme presentation.
Further, the timing of the visual language evoked activity and connectivity in the precentral
gyrus is consistent with the time window associated with GPC from extracranial electrophys-
iological research. The relationship of visual and auditory language evoked activity was
mixed. Though there was significant overlap in evoked activity, there were also many elec-
trodes in the perisylvian regions which were responsive to only a single language modality.
Overall, our study is consistent with a role for the precentral gyrus in GPC.
Neurobiology of Language
37
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
1
8
1
9
8
6
8
7
7
n
o
_
a
_
0
0
0
4
7
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Precentral gyrus and grapheme-phoneme conversion
The Presence of Visual Language Processing in the Precentral Gyrus
The precentral gyrus’ relationship to articulatory phonological representations and STG’s rela-
tionship to encoding phonological representations is important to long-standing debates re-
garding the relative contributions of articulatory versus encoding phonemic representations
to silent reading (Besner, 1987; Frost, 1998). fMRI studies have reported evoked BOLD acti-
vation during reading tasks in the STG and other temporal-parietal regions (Booth, 2002;
Rumsey et al., 1997; Simos et al., 2002) as well as the precentral gyrus and inferior frontal
regions (Binder et al., 2005; Dehaene et al., 2001; Fiez et al., 1999; Price et al., 1997; Yen
et al., 2019). We replicate these studies, with a significant proportion of precentral gyrus
electrodes (30%) showing Text-Preference effects, as well as Text-Preference effects in the
STG (4%), supramarginal (15%), and pars opercularis (18%). In addition, the precentral gyrus
also had 3 electrodes across 3 patients in which individual Letter-Sensitive electrodes were
found. Finally, in the 2 patients that had a Task-Modulated electrode in the left caudal fusiform,
significant connectivity between the electrodes in the fusiform and electrodes in the precentral
gyrus was found. Taken together, these findings offer strong replications of previous research
findings of precentral involvement during reading, here during a GPC specific task.
The nature of visual phonological processing in the precentral gyrus will require additional
study to better understand the possible functional response profiles. In this study, electrodes
responding to visual text appeared spatially distributed in both the inferior (close to the inferior
frontal sulcus) and more dorsal (close to the superior frontal sulcus) precentral gyrus. The infe-
rior region is adjacent to the inferior frontal gyrus, and this larger region including the inferior
precentral gyrus and pars opercularis has been reliably found to be more activated by phonol-
ogical processing tasks rather than semantic processing tasks during reading (Gitelman et al.,
2005; McDermott et al., 2003; Price et al., 1997). This inferior precentral region may be broadly
overlapping with mouth articulatory movements and has been hypothesized to be involved in
speech processing (Pulvermüller et al., 2006). However, evidence has also emerged of a more
dorsal region in the precentral gyrus separate from articulatory areas that is also involved in
speech perception (Berezutskaya et al., 2020; Cheung et al., 2016). The relationship of these
two areas, the inferior and more dorsal precentral, during speech perception is still under study.
In our study focusing on visual language, electrodes from both the more inferior and the more
superior regions shared a similar response waveform during our task (Supplementary Figure 5).
The timing of onsets to visual text, the onset and duration of Text-Preference effects, and the
sustained nature of activity throughout the presentation of the bi-phoneme were consistent
across both the inferior and the more dorsal regions. Future study will be necessary to understand
how phonological representations across the precentral gyrus are encoded during reading.
The Time-Course of the Precentral Gyrus’ Involvement in Visual Text Processing
The time-course of GPC is important to understanding the precentral gyrus’ potential role in
silent reading. Visual word encoding proceeds along the ventral temporal reading route in a
feedforward sweep (Dehaene & Cohen, 2011; Lochy et al., 2018; Vinckier et al., 2007). Visual
processing begins at 60 ms in posterior visual cortex (Foxe & Simpson, 2002) with Letter-
Sensitive encoding onsets in posterior occipital-temporal regions ~160–180 ms (Allison
et al., 1994, 1999; Hirshorn et al., 2016; Thesen et al., 2012). Lexical-semantic effects begin
in the anterior-ventral temporal lobe at ~200–250 ms (Chan et al., 2011; Lochy et al., 2018;
Nobre et al., 1994; Nobre & McCarthy, 1995; Thesen et al., 2012). These lexical-semantic
onset times align well with the onset of the rough time period from ~250–500 ms associated
with the N400 complex, taken to index lexical-semantic integration (Marinkovic(cid:1), 2004).
Neurobiology of Language
38
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
1
8
1
9
8
6
8
7
7
n
o
_
a
_
0
0
0
4
7
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Precentral gyrus and grapheme-phoneme conversion
During this extended N400 complex time period, prolonged feedforward/feedback interaction
in the language network has been observed (Halgren et al., 2006, 2015). In dorsal regions
simultaneous and widespread activity begins at ~180 ms (Halgren, 1990), and in the inferior
frontal gyrus (Sahin et al., 2009), latencies consistent with Letter-Sensitive effects begin in
caudal occipital-fusiform regions. It is during the widespread and simultaneous integrative
period of feedforward/feedback interaction that GPC and phonemic integration from the dorsal
regions would be expected.
During letter-string presentation we found two subjects with both a Text-Preference onset at
~160 ms in the caudal fusiform and significant connectivity between caudal fusiform and dor-
sal route regions. This supports the caudal fusiform, strongly associated with orthographic pro-
cessing (Cohen et al., 2000; Dehaene et al., 2001; Gaillard et al., 2006; Lüders et al., 1991), as
a hub connecting the ventral reading route with the widespread dorsal reading route activity.
Indeed, a resting-state fMRI study found the fusiform preferentially linked with frontal regions
including the precentral and pars opercularis as well as posterior temporal-parietal regions
(Stevens et al., 2017). These dorsal regions’ earliest Text-Preference effects were in pars oper-
cularis at ~200 ms (Mdn: 280 ms), precentral at ~220 ms (Mdn: 390 ms), STG at ~240 ms
(Mdn: 340 ms), and supramarginal at ~260 ms (Mdn: 320 ms) with no statistically significant
differences in onset times. This pattern is consistent with widespread early and sustained pro-
cessing in the dorsal (i.e., putative phonological) reading network during visual language
encoding. Effects indexing individual letter identity sensitivity (i.e., Letter-Sensitive effects)
emerged later in the precentral gyrus at ~340 ms (Mdn: ~420 ms) compared to earlier
Letter-Sensitive effects in the caudal fusiform at ~180 ms (Mdn: ~230 ms), demonstrating
quicker onsets in visual orthographic processing coupled with later dorsal processing.
This observed timing aligns with extracranial EEG studies investigating the timing of pho-
nological priming. Pseudohomophone priming effects (e.g., “BRANE” primes “BRAIN”) are
found during evoked components starting at ~250 ms (Grainger et al., 2006). Simultaneous
presentation of visual and auditory words that either match or mismatch shows differences
beginning at ~300 ms (Holcomb & Anderson, 1993). Rhyme judgement evoked differences
at ~300–350 ms (Bentin et al., 1999; Rugg, 1984; Rugg & Barrett, 1987). These extracranial
EEG studies provide evidence that phonological effects begin from ~250–350 ms, depending
on the paradigm used, the same time window as visual language evoked activity onset across
our perisylvian dorsal (i.e., putative phonological) route.
The Relationship Between Visual and Auditory Language Processing in Perisylvian Regions
Cognitive models of reading implicitly or explicitly theorize that GPC links orthographic pro-
cessing to the existing auditory language system. Here we directly test this theory by exam-
ining the overlap and separation as well as the priming of phonological representations
activated first by visual language input then second by auditory language input. The STG’s
role in encoding auditorily presented phonemes (Leonard et al., 2015; Mesgarani et al., 2014;
Travis et al., 2013) suggests that it may also help encode visually presented phonemes.
Previous intracranial studies have found overlapping activation in the STG to visually and
auditorily presented words (Perrone-Bertolotti et al., 2012) and individual neurons with cor-
related firing between phonemes and graphemes (Chan et al., 2014) and noted resting state
connectivity between the STG and fusiform (Stevens et al., 2017). We confirm this role for the
STG in phonemic encoding, with strong STG responsivity during the bi-phoneme presentation
and many Phoneme-Preference and Phoneme-Sensitive effects. Previous research has also
found involvement of the precentral gyrus in auditory language processing (Cheung et al.,
Neurobiology of Language
39
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
1
8
1
9
8
6
8
7
7
n
o
_
a
_
0
0
0
4
7
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Precentral gyrus and grapheme-phoneme conversion
2016), which we replicate here with the precentral gyrus showing Phoneme-Preference and
Phoneme-Sensitive effects.
To understand if phonological processing during reading makes use of the existing auditory
language system, we examined whether visually evoked effects overlapped with auditorily
evoked effects and whether visually encoded phonemes prime the auditory encoding of pho-
nemes. We did find significant overlap of Text-Preference with Phoneme-Preference elec-
trodes. However, while this overlap was significantly greater than chance it was not
comprehensive (25% of Text-Preference electrodes were also Phoneme-Preference), with
many effects in perisylvian regions which were only evoked by visual language. Perhaps sur-
prisingly, only limited evidence of visual phonological priming of auditory phonemes was
found, evidenced by the electrodes with Mismatch Effects. If the exact same phonological rep-
resentations in the perisylvian language system are activated by both visual and auditory lan-
guage input, then widespread priming effects would be expected (Gotts et al., 2012). While we
did find mismatch effects in the STG, precentral, pars opercularis, and supramarginal gyrus,
they were overall sparse, totaling just 15 electrodes. The median onset time of Mismatch ef-
fects in both the precentral gyrus and STG was ~220 ms post bi-phoneme presentation, which
is right around the time the first phoneme of the bi-phoneme pair was complete and in line
with timing in reports of visual/auditory priming in the extracranial EEG literature (Holcomb
et al., 2005; Kiyonaga et al., 2007). The STG and precentral showed significant PLV during the
bimodal period of comparison, evidence for integration of grapheme and phoneme identity.
Indeed, during bi-phoneme presentation both regions displayed significant PLV with each. The
earliest Mismatch effect was found in the STG at ~160 ms, in a region which was completely
unresponsive during the preceding letter-string presentation implying that this patch of STG
was involved in the integration between grapheme and phoneme but not the processing of
the grapheme. In total, the distribution of visual and auditory language effects suggests both
overlapping and separate language representations for visual language processing in perisyl-
vian cortex.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
1
8
1
9
8
6
8
7
7
n
o
_
a
_
0
0
0
4
7
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Limitations
The data we present is from a small number of patients who had electrophysiology recorded
directly from the cortical surface. Though this data is valuable for understanding cognitive
function due to possessing high spatial and temporal precision, qualifications are necessary
due to the rarity of this data and therefore the low number of patients included in this study.
Of the 8 patients recorded from, 5 had implantations covering the left hemisphere. As GPC is a
predominantly left-lateralized function and our patients were all presumed to have left-
lateralized language, attention must be paid to our small sample size. Further, electrode place-
ment is based on clinical rather than research considerations, and surface-based electrodes
lack access to sulcal cortex. Therefore, it is difficult to make strong inferences about interre-
gional comparisons. Further, though we found interesting effects, the limited number of elec-
trodes necessitates further replication. For example, Letter-Sensitive electrodes we found in the
precentral gyrus, a total of 3 electrodes in 3 patients. The Letter-Sensitive effects in the pre-
central gyrus differed in waveform, a sharp waveform in the fusiform versus a boxier elongated
waveform in the precentral, and differed in timing, early (Mdn: ~180 ms) for the fusiform and
later (Mdn: ~420 ms) for the precentral. However, it is not possible to conclude a unique role
for the precentral gyrus in GPC from these electrodes but they do offer a target for further
exploration. Finally, 3 subjects did not have definitive language lateralization available via
Wada; however, these patients were all right-handed and therefore have a high likelihood
of having left-lateralized language.
Neurobiology of Language
40
Precentral gyrus and grapheme-phoneme conversion
Conclusion and Future Directions
Early psychological theories emphasized a place for subvocal articulation in silent reading
(Allport, 1979; Barron & Baron, 1977; Burani et al., 1991; Klapp, 1971; Kleiman, 1975;
Peterson & Johnson, 1971; Sun & Peperkamp, 2016). In the Self-Learning Hypothesis model,
articulation is a key determinant in learning to read (Share, 1995). Articulating during reading
facilitates learning (Cunningham et al., 2002; Kyte & Johnson, 2006) and motor cortex disrup-
tion in childhood by seizures in rolandic epilepsy causes long-term reading impairments
(Clarke et al., 2007; Piccinelli et al., 2008; Staden et al., 1998). A neuroimaging review of
reading disorders linked increased precentral cortex BOLD activation to compensatory mech-
anisms centered on increased reliance on GPC in disordered reading (Hancock et al., 2017).
Computational modeling of learning to read (Harm & Seidenberg, 2004), as well as empirical
evidence (Grainger et al., 2012), shows a shift from reliance on phonological to orthographic
information as reading skill increases; however, peak performance requires both types of
information (Harm & Seidenberg, 2004), and phonological information continues to influence
silent reading (Frost, 1998; Perfetti et al., 1988; Rastle & Brysbaert, 2006). Therefore, we pro-
pose that the putative functional relationship between precentral gyrus and caudal fusiform
suggested by this study is likely formed when articulating during learning to read and remains
a contributing factor to silent reading throughout the lifespan. The empirical evidence presented
here does not conclusively establish an articulatory mechanistic understanding of GPC. Indeed,
auditory phonological processing in the more dorsal aspect of the precentral gyrus is reported to
be related to acoustic rather than articulatory representations (Cheung et al., 2016), raising the
possibility of multiple response profiles in precentral phonological processing. During reading, a
finer spatial resolution is needed to identify cortical patches, and potentially individual neurons,
representing overlapping letter/phoneme identities and overlapping encoding/articulation. The
greatly increased spatial resolution permitted by next-generation electrodes (Ganji et al., 2017;
Khodagholy et al., 2015) will allow for the measuring and stimulating down to a width of 50 μm
to test the articulatory GPC hypothesis.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
1
8
1
9
8
6
8
7
7
n
o
_
a
_
0
0
0
4
7
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
ACKNOWLEDGMENTS
Supported by NIH R01 NS018741, Kavli Institute for Brain and Mind, and Chancellor’s
Collaboratories Award (University of California San Diego).
FUNDING INFORMATION
Eric Halgren, National Institute of Neurological Disorders and Stroke (https://dx.doi.org/10
.13039/100000065), Award ID: NS018741.
AUTHOR CONTRIBUTIONS
Erik Kaestner: Conceptualization; Project administration; Software; Formal analysis; Writing –
original draft; Visualization. Xiaojing Wu: Conceptualization; Data curation; Investigation;
Writing – review & editing. Daniel Friedman: Investigation; Writing – review & editing.
Patricia Dugan: Investigation; Writing – review & editing. Orrin Devinsky: Investigation;
Writing – review & editing. Chad Carlson: Investigation; Writing – review & editing. Werner
Doyle: Investigation; Writing – review & editing. Thomas Thesen: Investigation; Writing –
review & editing. Eric Halgren: Conceptualization; Project administration; Resources; Writing –
original draft; Funding acquisition.
Neurobiology of Language
41
Precentral gyrus and grapheme-phoneme conversion
REFERENCES
Allison, T., McCarthy, G., Nobre, A., Puce, A., & Belger, A. (1994).
Human extrastriate visual cortex and the perception of faces,
words, numbers, and colors. Cerebral Cortex, 4, 544–554.
https://doi.org/10.1093/cercor/4.5.544, PubMed: 7833655
Allison, T., Puce, A., Spencer, D. D., & McCarthy, G. (1999).
Electrophysiological studies of human face perception. I:
Potentials generated in occipitotemporal cortex by face and
non-face stimuli. Cerebral Cortex, 9, 415–430. https://doi.org
/10.1093/cercor/9.5.415, PubMed: 10450888
Allport, A. (1979). Word recognition in reading (Tutorial paper). In
P. A. Kolers, M. E. Wrolstad, & H. Bouma (Eds.), Processing of
visible language (pp. 227–257). Springer. https://doi.org/10
.1007/978-1-4684-0994-9_14
Barron, R. W., & Baron J. (1977). How children get meaning from
printed words. Child Development, 48(2), 587–594. https://doi
.org/10.2307/1128657
Benjamini, Y., & Hochberg, Y. (1995). Controlling the false discovery
rate: A practical and powerful approach to multiple testing. Journal
of the Royal Statistical Society: Series B (Methodological), 57(1),
289–300. https://doi.org/10.1111/j.2517-6161.1995.tb02031.x
Bentin, S., Mouchetant-Rostaing, Y., Giard, M. H., Echallier, J. F., &
Pernier, J. (1999). ERP manifestations of processing printed words
at different psycholinguistic levels: Time course and scalp distri-
bution. Journal of Cognitive Neuroscience, 11, 235–260. https://
doi.org/10.1162/089892999563373, PubMed: 10402254
Berezutskaya, J., Baratin, C., Freudenburg, Z. V., & Ramsey, N. F.
(2020). High-density intracranial recordings reveal a distinct site
in anterior dorsal precentral cortex that tracks perceived speech.
Human Brain Mapping, 41, 4587–4609. https://doi.org/10.1002
/hbm.25144, PubMed: 32744403
Besner, D.
(1987). Phonology,
lexical access in reading, and
articulatory suppression: A critical review. Quarterly Journal
of Experimental Psychology, Section A: Human Experimental
Psychology, 39(3), 467–478. https://doi.org/10.1080
/14640748708401799
Binder, J. R., Medler, D. A., Desai, R., Conant, L. L., & Liebenthal,
E. (2005). Some neurophysiological constraints on models of
word naming. NeuroImage, 27(3), 677–693. https://doi.org/10
.1016/j.neuroimage.2005.04.029, PubMed: 15921937
Booth, J. (2002.) Functional anatomy of intra- and cross-modal
lexical tasks. NeuroImage, 16(1), 7–22. https://doi.org/10.1006
/nimg.2002.1081, PubMed: 11969313
Bouhali, F., Thiebaut de Schotten, M., Pinel, P., Poupon, C.,
Mangin, J.-F., Dehaene, S., & Cohen, L. (2014). Anatomical con-
nections of the visual word form area. Journal of Neuroscience,
34, 15402–15414. https://doi.org/10.1523/JNEUROSCI.4918-13
.2014, PubMed: 25392507
Burani, C., Vallar, G., & Bottini, G. (1991). Articulatory coding and
phonological judgements on written words and pictures: The
role of the phonological output buffer. European Journal of
Cognitive Psychology, 3(4), 379–398. https://doi.org/10.1080
/09541449108406235
Carreiras, M., Armstrong, B. C., Perea, M., & Frost, R. (2014). The
what, when, where, and how of visual word recognition. Trends
in Cognitive Sciences, 18, 90–98. https://doi.org/10.1016/j.tics
.2013.11.005, PubMed: 24373885
Chan, A. M., Baker, J. M., Eskandar, E., Schomer, D., Ulbert, I.,
Marinkovic, K., Cash, S. S., & Halgren, E. (2011). First-pass selec-
tivity for semantic categories in human anteroventral temporal
lobe. Journal of Neuroscience, 31(49), 18119–18129. https://doi
.org/10.1523/JNEUROSCI.3122-11.2011, PubMed: 22159123
Chan, A. M., Dykstra, A. R., Jayaram, V., Leonard, M. K., Travis,
K. E., Gygi, B., Baker, J. M., Eskandar, E., Hochberg, L. R.,
Halgren, E., & Cash, S. S. (2014). Speech-specific tuning of neu-
rons in human superior temporal gyrus. Cerebral Cortex, 24(10),
2679–2693. https://doi.org/10.1093/cercor/ bht127, PubMed:
23680841
Chen, F., & Loizou, P. C. (2011). Predicting the intelligibility of
vocoded speech. Ear and Hearing, 32(3), 331–338. https://doi
.org/10.1097/AUD.0b013e3181ff3515, PubMed: 21206363
Cheung, C., Hamilton, L. S., Johnson, K., & Chang, E. F. (2016) The
auditory representation of speech sounds in human motor cortex.
Elife, 5, Article e12577. https://doi.org/10.7554/eLife.12577,
PubMed: 27117389
Clarke, T., Strug, L. J., Murphy, P. L., Bali, B., Carvalho, J., Foster, S.,
Tremont, G., Gagnon, B. R., Dorta, N., & Pal, D. K. (2007). High risk
of reading disability and speech sound disorder in rolandic epilepsy
families: Case–control study. Epilepsia, 48(12), 2258–2265. https://
doi.org/10.1111/j.1528-1167.2007.01276.x, PubMed: 17850323
Cohen, L., Dehaene, S., Naccache, L., Lehéricy, S., Dehaene-
Lambertz, G., Hénaff, M.-A., & Michel, F. (2000). The visual
word form area: Spatial and temporal characterization of an ini-
tial stage of reading in normal subjects and posterior split-brain
patients. Brain, 123(Pt 2), 291–307. https://doi.org/10.1093/brain
/123.2.291, PubMed: 10648437
Coltheart, M. (1980). Reading, phonological recoding, and deep
dyslexia. In M. Coltheart, K. E. Patterson, & J. C. Marshall
(Eds.), Deep dyslexia (pp. 197–226). Routledge & Kegan Paul.
Coltheart, M., Rastle, K., Perry, C., Langdon, R., & Ziegler, J. (2001).
DRC: A dual route cascaded model of visual word recognition
and reading aloud. Psychological Review, 108(1), 204–256.
https://doi.org/10.1037/0033-295X.108.1.204, PubMed:
11212628
Cornelissen, P. L., Kringelbach, M. L., Ellis, A. W., Whitney, C.,
Holliday, I. E., & Hansen, P. C. (2009). Activation of the left
inferior frontal gyrus in the first 200 ms of reading: Evidence from
magnetoencephalography (MEG). PLOS ONE, 4, Article e5359.
https://doi.org/10.1371/journal.pone.0005359, PubMed:
19396362
Cunningham, A. E., Perry, K. E., Stanovich, K. E., & Share, D. L.
(2002). Orthographic learning during reading: Examining the role
of self-teaching. Journal of Experimental Child Psychology, 82(3),
185–199. https://doi.org/10.1016/S0022-0965(02)00008-5,
PubMed: 12093106
Dale, A. M., Fischl, B., & Sereno, M. I. (1999). Cortical surface-
based analysis: I. Segmentation and surface reconstruction.
NeuroImage, 9(2), 179–194. https://doi.org/10.1006/nimg.1998
.0395, PubMed: 9931268
Dehaene, S., & Cohen, L. (2011). The unique role of the visual
word form area in reading. Trends in Cognitive Sciences, 15(5),
254–262. https://doi.org/10.1016/j.tics.2011.04.003, PubMed:
21592844
Dehaene, S., Naccache, L., Cohen, L., Bihan, D. L., Mangin,
J.-F., Poline, J.-B., & Rivière, D. (2001). Cerebral mechanisms of
word masking and unconscious repetition priming. Nature
Neuroscience, 4(7), 752–758. https://doi.org/10.1038/89551,
PubMed: 11426233
Dejerine, J. (1892). Contribution à l’étude anatomopathologique
et clinique des différents variétés de cécité verbale. [Contribution
to the anatomopathological and clinical study of the various
varieties of verbal blindness]. Mémoires de la Societé de Biologie,
4, 61–90.
Neurobiology of Language
42
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
1
8
1
9
8
6
8
7
7
n
o
_
a
_
0
0
0
4
7
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Precentral gyrus and grapheme-phoneme conversion
DeMarco, A. T., Wilson, S. M., Rising, K., Rapcsak, S. Z., & Beeson,
P. M. (2018). The neural substrates of improved phonological
processing following successful treatment in a case of phonolo-
gical alexia and agraphia. Neurocase, 24(1), 31–40. https://doi
.org/10.1080/13554794.2018.1428352, PubMed: 29350575
Desikan, R. S., Ségonne, F., Fischl, B., Quinn, B. T., Dickerson,
B. C., Blacker, D., Buckner, R. L., Dale, A. M., Maguire, R. P.,
Hyman, B. T., Albert, M. S., & Killiany, R. J. (2006). An automated
labeling system for subdividing the human cerebral cortex on
MRI scans into gyral based regions of interest. NeuroImage, 31(3),
968–980. https://doi.org/10.1016/j.neuroimage.2006.01.021,
PubMed: 16530430
Diependaele, K., Ziegler, J. C., & Grainger, J. (2010). Fast phonology
and the bimodal interactive activation model. European Journal of
Cognitive Psychology, 22(5), 764–778. https://doi.org/10.1080
/09541440902834782
Fiez, J. A., Balota, D. A., Raichle, M. E., & Petersen, S. E. (1999).
Effects of lexicality, frequency, and spelling-to-sound consistency
on the functional anatomy of reading. Neuron, 24(1), 205–218.
https://doi.org/10.1016/S0896-6273(00)80833-8, PubMed:
10677038
Fiez, J. A., & Petersen, S. E. (1998). Neuroimaging studies of word
reading. Proceedings of the National Academy of Sciences of the
USA, 95(3), 914–921. https://doi.org/10.1073/pnas.95.3.914,
PubMed: 9448259
Foxe, J. J., & Simpson, G. V. (2002). Flow of activation from V1 to
frontal cortex in humans: A framework for defining “early” visual
processing. Experimental Brain Research, 142(1), 139–150.
https://doi.org/10.1007/s00221-001-0906-7, PubMed: 11797091
Frost, R. (1991). Phonetic recoding of print and its effect on the
detection of concurrent speech in amplitude-modulated noise.
Cognition, 39(3), 195–214. https://doi.org/10.1016/0010-0277
(91)90053-7, PubMed: 1841033
Frost, R. (1998). Toward a strong phonological theory of visual
word recognition: True issues and false trails. Psychological
Bulletin, 123(1), 71–99. https://doi.org/10.1037/0033-2909.123
.1.71, PubMed: 9461854
Frost, R., & Kampf, M. (1993). Phonetic recoding of phonologically
ambiguous printed words. Journal of Experimental Psychology:
Learning, Memory, and Cognition, 19(1), 23–33. https://doi.org
/10.1037/0278-7393.19.1.23
Gaillard, R., Naccache, L., Pinel, P., Clémenceau, S., Volle, E.,
Hasboun, D., Dupont, S., Baulac, M., Dehaene, S., Adam, C.,
& Cohen, L. (2006). Direct intracranial, fMRI, and lesion evi-
dence for the causal role of left inferotemporal cortex in reading.
Neuron, 50(2), 191–204. https://doi.org/10.1016/j.neuron.2006
.03.031, PubMed: 16630832
Ganji, M., Kaestner, E., Hermiz, J., Rogers, N., Tanaka, A., Cleary,
D., Lee, S. H., Snider, J., Halgren, M., & Cosgrove, G. R. (2017).
Development and translation of PEDOT: PSS microelectrodes for
intraoperative monitoring. Advanced Functional Materials, 28(12),
Article 1700232. https://doi.org/10.1002/adfm.201700232
Geschwind, N. (1974). Disconnexion syndromes in animals and
man. In N. Geschwind (Ed.), Selected papers on language and
the brain (pp. 105–236). Springer. https://doi.org/10.1007/978
-94-010-2093-0_8
Gitelman, D. R., Nobre, A. C., Sonty, S., Parrish, T. B., & Mesulam,
M.-M. (2005). Language network specializations: An analysis
with parallel task designs and functional magnetic resonance
imaging. NeuroImage, 26(4), 975–985. https://doi.org/10.1016/j
.neuroimage.2005.03.014, PubMed: 15893473
Gotts, S. J., Chow, C. C., & Martin, A. (2012). Repetition priming
and repetition suppression: A case for enhanced efficiency
through neural synchronization. Cognitive Neuroscience, 3(3–4),
227–237. https://doi.org/10.1080/17588928.2012.670617,
PubMed: 23144664
Grainger, J., Kiyonaga, K., & Holcomb, P. J. (2006). The time course
of orthographic and phonological code activation. Psychological
Science, 17(12), 1021–1026. https://doi.org/10.1111/j.1467
-9280.2006.01821.x, PubMed: 17201781
Grainger, J., Lété, B., Bertand, D., Dufau, S., & Ziegler, J. C. (2012).
Evidence for multiple routes in learning to read. Cognition, 123(2),
280–292. https://doi.org/10.1016/j.cognition.2012.01.003,
PubMed: 22357323
Halgren, E. (1990). Insights from evoked potentials into the neuro-
psychological mechanisms of reading. In A. B. Scheibel & A. F.
Wechsler (Eds.), Neurobiology of higher cognitive function
(pp. 103–150). Guilford Press.
Halgren, E., Kaestner, E., Marinkovic, K., Cash, S. S., Wang, C.,
Schomer, D. L., Madsen, J. R., & Ulbert, I. (2015). Laminar profile
of spontaneous and evoked theta: Rhythmic modulation of cor-
tical processing during word integration. Neuropsychologia, 76,
108–124. https://doi.org/10.1016/j.neuropsychologia.2015.03
.021, PubMed: 25801916
Halgren, E., Wang, C., Schomer, D. L., Knake, S., Marinkovic, K.,
Wu, J., & Ulbert, I. (2006). Processing stages underlying word
recognition in the anteroventral temporal lobe. NeuroImage,
30(5), 1401–1413. https://doi.org/10.1016/j.neuroimage.2005
.10.053, PubMed: 16488158
Hancock, R., Richlan, F., & Hoeft, F. (2017). Possible roles for
fronto-striatal circuits in reading disorder. Neuroscience &
Biobehavioral Reviews, 72, 243–260. https://doi.org/10.1016/j
.neubiorev.2016.10.025, PubMed: 27826071
Harm, M. W., & Seidenberg, M. S. (2004). Computing the meanings
of words in reading: Cooperative division of labor between visual
and phonological processes. Psychological Review, 111(3),
662–720. https://doi.org/10.1037/0033-295X.111.3.662,
PubMed: 15250780
Hirshorn, E. A., Li, Y., Ward, M. J., Richardson, R. M., Fiez, J. A., &
Ghuman, A. S. (2016). Decoding and disrupting left midfusiform
gyrus activity during word reading. Proceedings of the National
Academy of Sciences of the USA, 113(29), 8162–8167. https://
doi.org/10.1073/pnas.1604126113, PubMed: 27325763
Holcomb, P. J., & Anderson, J. E. (1993). Cross-modal semantic
priming: A time-course analysis using event-related brain poten-
tials. Language and Cognitive Processes, 8(4), 379–411. https://
doi.org/10.1080/01690969308407583
Holcomb, P. J., Anderson, J., & Grainger, J. (2005). An electrophysio-
logical study of cross-modal repetition priming. Psychophysiology,
42(5), 493–507. https://doi.org/10.1111/j.1469-8986.2005
.00348.x, PubMed: 16176372
Horowitz, L. A. (2014). Cognitive consequences of degraded
speech: The effect of background noise, noise-vocoded speech
and low-pass filtering on short story recall. Independent Studies
and Capstones. Paper 693. Program in Audiology and
Communication Sciences, Washington University School of
Medicine. https://digitalcommons.wustl.edu/pacs_capstones/693
Jobard, G., Crivello, F., & Tzourio-Mazoyer, N. (2003). Evaluation
of the dual route theory of reading: A metanalysis of 35 neuroim-
aging studies. NeuroImage, 20(2), 693–712. https://doi.org/10
.1016/S1053-8119(03)00343-4, PubMed: 14568445
Khodagholy, D., Gelinas, J. N., Thesen, T., Doyle, W., Devinsky, O.,
Malliaras, G. G., & Buzsáki, G. (2015). NeuroGrid: Recording
action potentials from the surface of the brain. Nature
Neuroscience, 18(2), 310–315. https://doi.org/10.1038/nn.3905,
PubMed: 25531570
Neurobiology of Language
43
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
1
8
1
9
8
6
8
7
7
n
o
_
a
_
0
0
0
4
7
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Precentral gyrus and grapheme-phoneme conversion
Kiyonaga, K., Grainger, J., Midgley, K., & Holcomb, P. J. (2007).
Masked cross-modal repetition priming: An event-related poten-
tial investigation. Language and Cognitive Processes, 22(3),
337–376. https://doi.org/10.1080/01690960600652471,
PubMed: 18163153
Klapp, S. T. (1971). Implicit speech inferred from response latencies
in same-different decisions. Journal of Experimental Psychology,
91(2), 262–267. https://doi.org/10.1037/ h0031852, PubMed:
5134669
Kleiman, G. M. (1975). Speech recoding in reading. Journal of
Verbal Learning & Verbal Behavior, 14(4), 323–339. https://doi
.org/10.1016/S0022-5371(75)80013-2
Kyte, C. S., & Johnson, C. J. (2006). The role of phonological recoding
in orthographic learning. Journal of Experimental Child
Psychology, 93(2), 166–185. https://doi.org/10.1016/j.jecp.2005
.09.003, PubMed: 16246358
Lachaux, J. P., Rodriguez, E., Martinerie, J., & Varela, F. J. (1999).
Measuring phase synchrony in brain signals. Human Brain
Mapping, 8(4), 194–208. https://doi.org/10.1002/(SICI)1097
-0193(1999)8:4<194::AID-HBM4>3.0.CO;2-C, PubMed:
10619414
Lauro, L. J. R., Vergallito, A., Anzani, S., & Vallar, G. (2020).
Primary motor cortex and phonological recoding: A TMS-EMG
study. Neuropsychologia, 139, Article 107368. https://doi.org
/10.1016/j.neuropsychologia.2020.107368, PubMed: 32014451
Leonard, M. K., Bouchard, K. E., Tang, C., & Chang, E. F. (2015).
Dynamic encoding of speech sequence probability in human
temporal cortex. Journal of Neuroscience, 35(18), 7203–7214.
https://doi.org/10.1523/ JNEUROSCI.4100-14.2015, PubMed:
25948269
Lochy, A., Jacques, C., Maillard, L., Colnat-Coulbois, S., Rossion, B.,
& Jonas, J. (2018). Selective visual representation of letters and
words in the left ventral occipito-temporal cortex with intracere-
bral recordings. Proceedings of the National Academy of Sciences
of the USA, 115(32), E7595–E7604. https://doi.org/10.1073/pnas
.1718987115, PubMed: 30038000
Lüders, H., Lesser, R. P., Hahn, J., Dinner, D. S., Morris, H. H.,
Wyllie, E., & Godoy, J. (1991). Basal temporal language area.
Brain, 114(Pt 2), 743–754. https://doi.org/10.1093/brain/114.2
.743, PubMed: 2043946
Marinkovic(cid:1), K. (2004). Spatiotemporal dynamics of word processing
in the human cortex. Neuroscientist, 10(2), 142–152. https://doi
.org/10.1177/1073858403261018, PubMed: 15070488
Maris, E., & Oostenveld, R. (2007). Nonparametric statistical testing
of EEG-and MEG-data. Journal of Neuroscience Methods, 164(1),
177–190. https://doi.org/10.1016/j.jneumeth.2007.03.024,
PubMed: 17517438
McDermott, K. B., Petersen, S. E., Watson, J. M., & Ojemann, J. G.
(2003). A procedure for identifying regions preferentially activated
by attention to semantic and phonological relations using func-
tional magnetic resonance imaging. Neuropsychologia, 41(3),
293–303. https://doi.org/10.1016/S0028-3932(02)00162-8,
PubMed: 12457755
McDonald, C. R., Thesen, T., Carlson, C., Blumberg, M., Girard,
H. M., Trongnetrpunya, A., Sherfey, J. S., Devinsky, O.,
Kuzniecky, R., Dolye, W. K., Cash, S. S., Leonard, M. K., Hagler,
D. J., Jr., Dale, A. M., & Halgren, E. (2010). Multimodal imaging
of repetition priming: Using fMRI, MEG, and intracranial EEG to
reveal spatiotemporal profiles of word processing. NeuroImage,
53(2), 707–717. https://doi.org/10.1016/j.neuroimage.2010.06
.069, PubMed: 20620212
Mesgarani, N., Cheung, C., Johnson, K., & Chang, E. F. (2014).
Phonetic feature encoding in human superior temporal gyrus.
Science, 343(6174), 1006–1010. https://doi.org/10.1126
/science.1245994, PubMed: 24482117
Nobre, A. C., Allison, T., & McCarthy, G. (1994). Word recognition in
the human inferior termporal lobe. Nature, 372(6503), 260–263.
https://doi.org/10.1038/372260a0, PubMed: 7969469
Nobre, A. C., & McCarthy, G. (1995). Language-related field poten-
tials in the anterior-medial temporal lobe: II. Effects of word type
and semantic priming. Journal of Neuroscience, 15(2), 1090–1098.
https://doi.org/10.1523/JNEUROSCI.15-02-01090.1995, PubMed:
7869085
Nourski, K. V., Steinschneider, M., Rhone, A. E., Kovach, C. K.,
Kawasaki, H., & Howard, M. A., III. (2019). Differential
responses to spectrally degraded speech within human auditory
cortex: An intracranial electrophysiology study. Hearing
Research, 371, 53–65. https://doi.org/10.1016/j.heares.2018.11
.009, PubMed: 30500619
Ojemann, G., Ojemann, J., Lettich, E., & Berger, M. (1989). Cortical
language localization in left, dominant hemisphere: An electrical
stimulation mapping investigation in 117 patients. Journal of
Neurosurgery, 71(3), 316–326. https://doi.org/10.3171/jns.1989
.71.3.0316, PubMed: 2769383
Oostenveld, R., Fries, P., Maris, E., & Schoffelen, J.-M. (2011).
FieldTrip: Open source software for advanced analysis of MEG,
EEG, and invasive electrophysiological data. Computational
Intelligence and Neuroscience, 2011, Article 156869. https://
doi.org/10.1155/2011/156869, PubMed: 21253357
Pammer, K., Hansen, P. C., Kringelbach, M. L., Holliday, I., Barnes,
G., Hillebrand, A., Singh, K. D., & Cornelissen, P. L. (2004). Visual
word recognition: The first half second. NeuroImage, 22(4),
1819–1825. https://doi.org/10.1016/j.neuroimage.2004.05.004,
PubMed: 15275938
Perfetti, C. A., Bell, L. C., & Delaney, S. M. (1988). Automatic
(prelexical) phonetic activation in silent word reading: Evidence
from backward masking. Journal of Memory and Language,
27(1), 59–70. https://doi.org/10.1016/0749-596X(88)90048-4
Perrone-Bertolotti, M., Kujala, J., Vidal, J. R., Hamame, C. M.,
Ossandon, T., Bertrand, O., Minotti, L., Kahane, P., Jerbi, K., &
Lachaux, J.-P. (2012). How silent is silent reading? Intracerebral
evidence for top-down activation of temporal voice areas during
reading. Journal of Neuroscience, 32(49), 17554–17562. https://
doi.org/10.1523/ JNEUROSCI.2982-12.2012, PubMed:
23223279
Perry, C., Ziegler, J. C., & Zorzi, M. (2007). Nested incremental
modeling in the development of computational theories: The
CDP+ model of reading aloud. Psychological Review, 114(2),
273–315. https://doi.org/10.1037/0033-295X.114.2.273,
PubMed: 17500628
Peterson, L. R., & Johnson, S. T. (1971). Some effects of minimizing
articulation on short-term retention. Journal of Verbal Learning &
Verbal Behavior, 10(4), 346–354. https://doi.org/10.1016/S0022
-5371(71)80033-6
Piccinelli, P., Borgatti, R., Aldini, A., Bindelli, D., Ferri, M., Perna,
S., Pitillo, G., Termine, C., Zambonin, F., & Balottin, U. (2008).
Academic performance in children with rolandic epilepsy.
Developmental Medicine & Child Neurology, 50(5), 353–356.
https://doi.org/10.1111/j.1469-8749.2008.02040.x, PubMed:
18294216
Price, C. J. (2012). A review and synthesis of the first 20 years of PET
and fMRI studies of heard speech, spoken language and reading.
NeuroImage, 62(2), 816–847. https://doi.org/10.1016/j
.neuroimage.2012.04.062, PubMed: 22584224
Price, C. J., Moore, C. J., Humphreys, G. W., & Wise, R. J. S. (1997).
Segregating semantic from phonological processes during
Neurobiology of Language
44
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
1
8
1
9
8
6
8
7
7
n
o
_
a
_
0
0
0
4
7
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Precentral gyrus and grapheme-phoneme conversion
reading. Journal of Cognitive Neuroscience, 9(12), 727–733.
https://doi.org/10.1162/jocn.1997.9.6.727, PubMed: 23964595
Pulvermüller, F., Huss, M., Kherif, F., del Prado Martin, F. M.,
Hauk, O., & Shtyrov, Y. (2006). Motor cortex maps articulatory
features of speech sounds. Proceedings of the National Academy
of Sciences of the USA, 103(20), 7865–7870. https://doi.org/10
.1073/pnas.0509989103, PubMed: 16682637
Rastle, K., & Brysbaert, M. (2006). Masked phonological priming
effects in English: Are they real? Do they matter? Cognitive
Psychology, 53(2), 97–145. https://doi.org/10.1016/j.cogpsych
.2006.01.002, PubMed: 16554045
Ray, S., Crone, N. E., Niebur, E., Franaszczuk, P. J., & Hsiao, S. S.
(2008). Neural correlates of high-gamma oscillations (60–200 Hz)
in macaque local field potentials and their potential implica-
tions in electrocorticography. Journal of Neuroscience, 28(45),
11526–11536. https://doi.org/10.1523/ JNEUROSCI.2848-08
.2008, PubMed: 18987189
Rugg, M. D. (1984). Event-related potentials in phonological
matching tasks. Brain and Language, 23(2), 225–240. https://
doi.org/10.1016/0093-934X(84)90065-8, PubMed: 6518354
Rugg, M. D., & Barrett, S. E. (1987). Event-related potentials and the
interaction between orthographic and phonological information in
a rhyme-judgment task. Brain and Language, 32(2), 336–361.
https://doi.org/10.1016/0093-934X(87)90132-5, PubMed: 3690257
Rumsey, J. M., Horwitz, B., Donohue, B. C., Nace, K., Maisog, J. M.,
& Andreason, P. (1997). Phonological and orthographic compo-
nents of word recognition: A PET-rCBF study. Brain: A Journal of
Neurology, 120(5), 739–759. https://doi.org/10.1093/brain/120.5
.739, PubMed: 9183247
Sahin, N. T., Pinker, S., Cash, S. S., Schomer, D., & Halgren, E. (2009).
Sequential processing of lexical, grammatical, and phonological
information within Broca’s area. Science, 326(5951), 445–449.
https://doi.org/10.1126/science.1174481, PubMed: 19833971
Share, D. L. (1995). Phonological recoding and self-teaching: Sine qua
non of reading acquisition. Cognition, 55(2), 151–218. https://doi
.org/10.1016/0010-0277(94)00645-2, PubMed: 7789090
Simos, P. G., Breier, J. I., Fletcher, J. M., Foorman, B. R., Castillo,
E. M., & Papanicolaou, A. C. (2002). Brain mechanisms for reading
words and pseudowords: An integrated approach. Cerebral
Cortex, 12(3), 297–305. https://doi.org/10.1093/cercor/12.3.297,
PubMed: 11839603
Souza, P., & Rosen, S. (2009). Effects of envelope bandwidth on the
intelligibility of sine- and noise-vocoded speech. Journal of the
Acoustical Society of America, 126(2), 792–805. https://doi.org
/10.1121/1.3158835, PubMed: 19640044
Staden, U., Isaacs, E., Boyd, S. C., Brandl, U., & Neville, B. G. R.
(1998). Language dysfunction in children with rolandic epilepsy.
Neuropediatrics, 29(5), 242–248. https://doi.org/10.1055/s-2007
-973569, PubMed: 9810559
Stevens, W. D., Kravitz, D. J., Peng, C. S., Tessler, M. H., & Martin, A.
(2017). Privileged functional connectivity between the visual word
form area and the language system. Journal of Neuroscience, 37(21),
5288–5297. https://doi.org/10.1523/JNEUROSCI.0138-17.2017,
PubMed: 28450544
Sun, Y., & Peperkamp, S. (2016). The role of speech production in
phonological decoding during visual word recognition: Evidence
from phonotactic repair. Language, Cognition and Neuroscience,
31(3), 391–403. https://doi.org/10.1080/23273798.2015
.1100316
Taylor, J. S. H., Rastle, K., & Davis, M. H. (2013). Can cognitive
models explain brain activation during word and pseudoword
reading? A meta-analysis of 36 neuroimaging studies.
Psychological Bulletin, 139(4), 766–791. https://doi.org/10.1037
/a0030266, PubMed: 23046391
Thesen, T., McDonald, C. R., Carlson, C., Doyle, W., Cash, S.,
Sherfey, J., Felsovalyi, O., Girard, H., Barr, W., Devinsky, O.,
Kuzniecky, R., & Halgren, E. (2012). Sequential then interactive
processing of letters and words in the left fusiform gyrus. Nature
Communications, 3, Article 1284. https://doi.org/10.1038
/ncomms2220, PubMed: 23250414
Travis, K. E., Leonard, M. K., Chan, A. M., Torres, C., Sizemore, M. L.,
Qu, Z., Eskandar, E., Dale, A. M., Elman, J. L., Cash, S. S., &
Halgren, E. (2013). Independence of early speech processing
from word meaning. Cerebral Cortex, 23(10), 2370–2379.
https://doi.org/10.1093/cercor/bhs228, PubMed: 22875868
Vallar, G., & Cappa, S. F. (1987). Articulation and verbal short-term
memory: Evidence from anarthria. Cognitive Neuropsychology,
4(1), 55–77. https://doi.org/10.1080/02643298708252035
Vallar, G., Di Betta, A. M., & Silveri, M. C. (1997). The phonolo-
gical short-term store-rehearsal system: Patterns of impairment
and neural correlates. Neuropsychologia, 35(6), 795–812. https://
doi.org/10.1016/S0028-3932(96)00127-3, PubMed: 9204486
Vinckier, F., Dehaene, S., Jobert, A., Dubus, J. P., Sigman, M., &
Cohen, L. (2007). Hierarchical coding of letter strings in the
ventral stream: Dissecting the inner organization of the visual
word-form system. Neuron, 55(1), 143–156. https://doi.org/10
.1016/j.neuron.2007.05.031, PubMed: 17610823
Wheat, K. L., Cornelissen, P. L., Frost, S. J., & Hansen, P. C. (2010).
During visual word recognition, phonology is accessed within
100 ms and may be mediated by a speech production code:
Evidence from magnetoencephalography. Journal of Neuroscience,
30(15), 5229–5233. https://doi.org/10.1523/ JNEUROSCI.4448
-09.2010, PubMed: 20392945
Yang, A. I., Wang, X., Doyle, W. K., Halgren, E., Carlson, C.,
Belcher, T. L., Cash, S. S., Devinsky, O., & Thesen, T. (2012).
Localization of dense intracranial electrode arrays using magnetic
resonance imaging. NeuroImage, 63(1), 157–165. https://doi.org
/10.1016/j.neuroimage.2012.06.039, PubMed: 22759995
Yen, M., DeMarco, A. T., & Wilson, S. M. (2019). Adaptive para-
digms for mapping phonological regions in individual partici-
pants. NeuroImage, 189, 368–379. https://doi.org/10.1016/j
.neuroimage.2019.01.040, PubMed: 30665008
Neurobiology of Language
45
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
1
8
1
9
8
6
8
7
7
n
o
_
a
_
0
0
0
4
7
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3