RESEARCH ARTICLE
Analogy-Related Information Can Be Accessed
by Simple Addition and Subtraction of fMRI
Activation Patterns, Without Participants
Performing any Analogy Task
Meng-Huan Wu1
, Andrew J. Anderson2,3
, Robert A. Jacobs1
, and Rajeev D. S. Raizada1
1Department of Brain & Cognitive Sciences, University of Rochester, Rochester, New York, USA
2Department of Neuroscience, University of Rochester, Rochester, New York, USA
3Del Monte Institute for Neuroscience, University of Rochester, Rochester, New York, USA
Keywords: fMRI, language, word analogy, word2vec
ABSTRACT
Analogical reasoning, for example, inferring that teacher is to chalk as mechanic is to wrench,
plays a fundamental role in human cognition. However, whether brain activity patterns of
individual words are encoded in a way that could facilitate analogical reasoning is unclear.
Recent advances in computational linguistics have shown that information about analogical
problems can be accessed by simple addition and subtraction of word embeddings (e.g.,
wrench = mechanic + chalk − teacher). Critically, this property emerges in artificial neural
networks that were not trained to produce analogies but instead were trained to produce
general-purpose semantic representations. Here, we test whether such emergent property can
be observed in representations in human brains, as well as in artificial neural networks. fMRI
activation patterns were recorded while participants viewed isolated words but did not
perform analogical reasoning tasks. Analogy relations were constructed from word pairs that
were categorically or thematically related, and we tested whether the predicted fMRI pattern
calculated with simple arithmetic was more correlated with the pattern of the target word than
other words. We observed that the predicted fMRI patterns contain information about not only
the identity of the target word but also its category and theme (e.g., teaching-related). In
summary, this study demonstrated that information about analogy questions can be reliably
accessed with the addition and subtraction of fMRI patterns, and that, similar to word
embeddings, this property holds for task-general patterns elicited when participants were not
explicitly told to perform analogical reasoning.
INTRODUCTION
Analogical reasoning is a fundamental component of human cognition. Despite extensive re-
search in psychology (Gentner, 1983), cognitive neuroscience (Chiang et al., 2021; Waltz
et al., 1999), and artificial intelligence (Turney, 2006), scientific understanding of the neural
bases of analogical problems is limited. Recent advances in computational linguistics have
shown that information about analogical problems can be accessed by simple addition and
subtraction of word embeddings, which are numeric feature vectors reflecting textual contexts
a n o p e n a c c e s s
j o u r n a l
Citation: Wu, M.-H., Anderson, A. J.,
Jacobs, R. A., & Raizada, R. D. S.
(2022). Analogy-related information can
be accessed by simple addition and
subtraction of fMRI activation patterns,
without participants performing any
analogy task. Neurobiology of
Language, 3(1), 1–17. https://doi.org
/10.1162/nol_a_00045
DOI:
https://doi.org/10.1162/nol_a_00045
Supporting Information:
https://doi.org/10.1162/nol_a_00045
Received: 2 September 2020
Accepted: 15 June 2021
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Author:
Meng-Huan Wu
menghuanwu@rochester.edu
Handling Editor:
Steven Small
Copyright: © 2021
Massachusetts Institute of Technology
Published under a Creative Commons
Attribution 4.0 International
(CC BY 4.0) license
The MIT Press
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
1
1
9
8
6
8
4
4
n
o
_
a
_
0
0
0
4
5
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Analogy-related information accessed by simple arithmetic in fMRI patterns
or word co-occurrences (e.g., word2vec; Mikolov et al., 2013). Critically, the ability to access
information about analogy problems is emergent in artificial neural networks, because word
embeddings are extracted from networks that were trained to produce general-purpose seman-
tic representations, not word analogy relations. While this ability is far from solving analogy
problems, it is remarkable that such trivial operations can extract analogy-related information
at all.
Given those findings in computational linguistics, it is therefore intriguing to consider
whether similar emergent properties can also be observed with biological neural networks,
i.e., neural representations in the human brain. In this study, we tested whether the semantic
representations of individual words are sufficiently rich such that analogy-related information
can be accessed using simple arithmetic operations. Crucially, these patterns were elicited
when participants simply read isolated words and were not performing any analogical reason-
ing task.
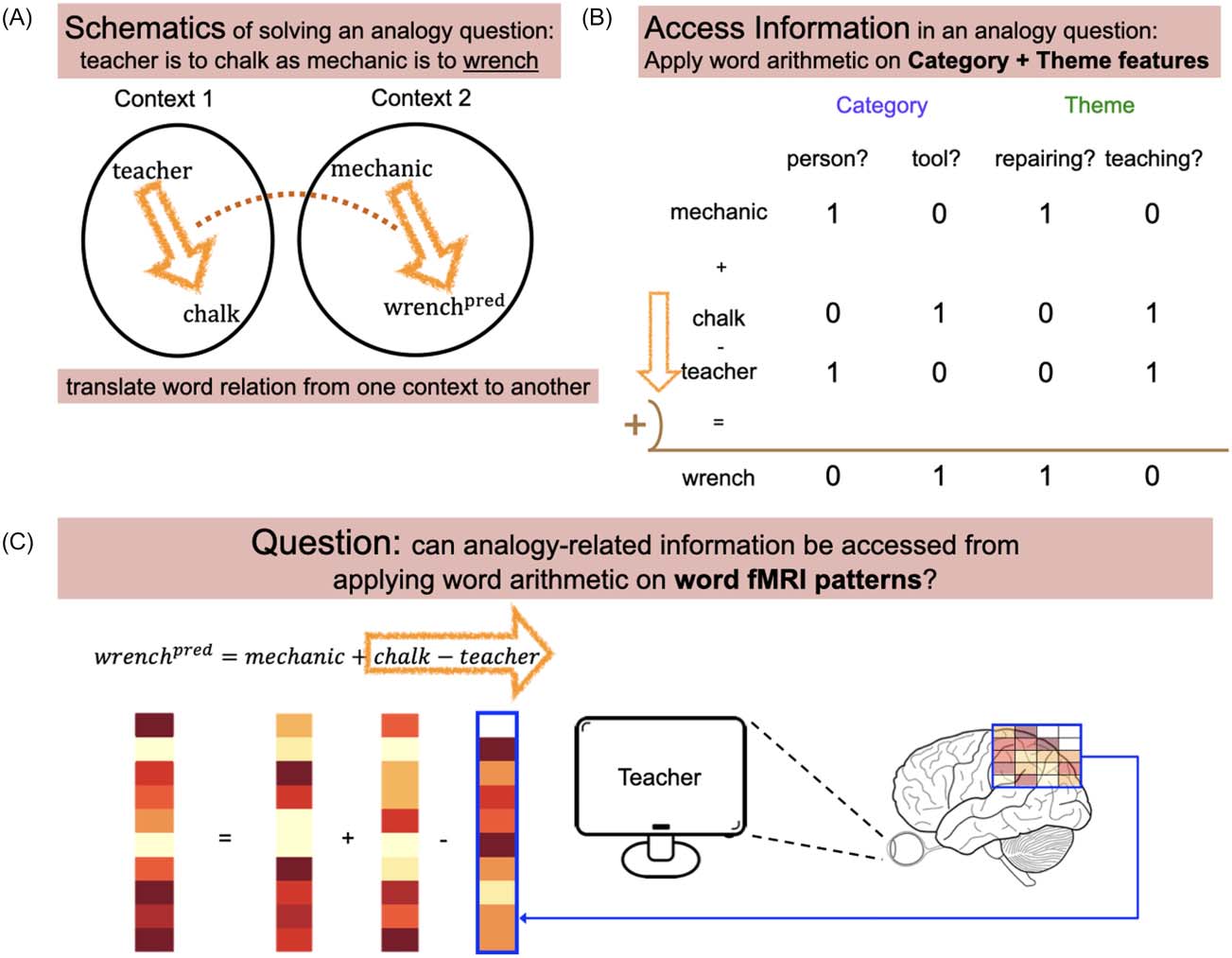
Consider an example analogy question: teacher is to chalk as mechanic is to (wrench). To
solve this question, the relationship between teacher and chalk must be translated and applied
from teacher to mechanic (Figure 1). We first assume that approximate solutions to analogical
questions can be derived geometrically, with word meaning approximated as a numeric fea-
ture vector (e.g., word2vec). Then, the relation between teacher and chalk can be computed
by the feature-wise subtraction of teacher from chalk. Adding the result onto mechanic
(Figure 1A) will produce a feature vector reasonably close to wrench if distinct semantic cat-
egories (e.g., tools and people) and contexts (also known as themes, e.g., teaching and
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
1
1
9
8
6
8
4
4
n
o
_
a
_
0
0
0
4
5
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 1.
Schematics of the current study. (A) Similar to solving analogy questions, information
about word analogies was accessed by translating word relations from one context to another.
(B) The categorical and thematic membership of each word can be determined by applying simple
word arithmetic operations to such features. (C) We attempted to study the simple yet unanswered
question: Can such analogy-related information be accessed from applying addition and subtraction
to the fMRI patterns of individual words?
Neurobiology of Language
2
Analogy-related information accessed by simple arithmetic in fMRI patterns
Functional magnetic resonance
imaging (fMRI):
A technique that measures brain
activity and is used to study brain
function.
repairing) are each represented as distinct semantic dimensions (Figure 1B), such that words
with different categories and themes occupy distinct positions in the semantic feature space.
Following this reasoning, we would anticipate that the segregation of not only themes but
semantic categories is also an emergent structural property of word embeddings, which
was indeed demonstrated with word2vec (Fu et al., 2014).
There is likewise extensive evidence that information associated with semantic categories
and themes is anatomically segregated in the human brain (Kalénine et al., 2009; Lewis et al.,
2015; Sachs et al., 2008; Schwartz et al., 2011). These studies have emphasized the role of the
anterior temporal lobe (ATL) in representing categories, and the inferior parietal lobe (IPL) in
representing themes. This suggests that analogy-related information can be accessed from
brain activation patterns (Figure 1C), but only if they are sampled across anatomically sepa-
rated regions (e.g., IPL and ATL). In contrast, several studies found evidence inconsistent with
the segregation perspective; thematic structure has been documented within the ATL (Peelen
& Caramazza, 2012), and different semantic categories and themes have been distinguished
within individual brain regions (Anderson et al., 2014; Xu et al., 2018). Additionally, word
embeddings and other semantic models differentiating categories and themes can explain
functional magnetic resonance imaging (fMRI) activation patterns within multiple localized
regions of an anatomically distributed semantic network spanning temporal, parietal, and fron-
tal cortex (Anderson et al., 2015, 2019; Carota et al., 2017; Huth et al., 2012, 2016; Mitchell
et al., 2008; Pereira et al., 2016, 2018; J. Wang et al., 2017). It is therefore unclear whether
analogy-related information can be accessed using fMRI patterns extracted from within indi-
vidual brain regions, or whether it is necessary to integrate information across the semantic
network (e.g., ATL and IPL).
The Current Study
In this study, we tested two hypotheses. H1: Analogy-related information can be accessed
from adding and subtracting task-general fMRI patterns elicited by viewing isolated words
(Figure 1C). H2: Analogy-related information can be accessed from adding and subtracting
fMRI patterns within individual regions of the brain’s semantic network.
MATERIALS AND METHODS
Participants
Sixteen undergraduate students and researchers (age: 23.9 ± 6.8 years; 10 female) participated in
the current study. All participants are native English speakers, right-handed, and have normal or
corrected-to-normal vision. All participants provided written consent in accordance with the
University of Rochester Research Subjects Review Board. Two participants were excluded
due to experimental hardware or software failure, and one was excluded for excessive motion
artifact (>2 mm in x, y, z dimensions or 2 degrees in pitch, roll, yaw dimensions); therefore,
13 participants were included in the final analysis. We conducted our analysis on 13 participants
because this is a standard sample size in other contemporary fMRI studies of isolated word rep-
resentation that used 9 and 11 participants (Just et al., 2010; Mitchell et al., 2008).
Stimuli
The 45 words used as stimuli are listed in Table 1. The words were organized into three categories
(person/building/tool) and 15 themes. These three categories were selected since it was rela-
tively easy to think up a single theme that would simultaneously apply to all three of these
Neurobiology of Language
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
1
1
9
8
6
8
4
4
n
o
_
a
_
0
0
0
4
5
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Analogy-related information accessed by simple arithmetic in fMRI patterns
Table 1.
All 45 words used as stimuli in the current study
15 themes (thematic)
Person
doctor
soldier
farmer
mechanic
photographer
teacher
chef
knight
Eskimo
bodybuilder
gardener
shopper
judge
hairdresser
chemist
3 categories (taxonomic)
Place
hospital
Tool
stethoscope
fort
barn
garage
studio
school
restaurant
castle
igloo
gym
gun
plow
wrench
camera
chalk
skillet
sword
harpoon
dumbbell
greenhouse
mower
mall
court
barbershop
laboratory
cart
gavel
trimmer
beaker
categories (e.g., doctor-hospital-stethoscope). We found it difficult to add additional categories
that were universally applicable to themes. Stimuli word characteristics such as word length,
word frequency, and word imageability ratings are listed in Table 2. Stimuli words are highly
imageable overall (M = 6.26 on a 7-point scale).
Words were organized such that an analogy question can be formed by selecting quadru-
plets associated with two categories and two themes, and finally selecting one of the four
words as the target word. For each of the 1,260 distinct analogy questions, we tested whether
information about the target word in an analogy question is encoded in the fMRI patterns of the
other three words.
Table 2.
Characteristics of the 45 stimulus words, organized by each category
Characteristic
Person
Place
Tool
All categories
Word length
mean (SD)
7.40 (2.29)
6.47 (2.50)
5.93 (1.94)
6.6 (2.29)
Word frequency per
million mean (SD)
28.56 (65.85)
31.55 (48.67)
7.64 (15.64)
22.58 (48.17)
Imageability
mean (SD)
6.21 (0.41)
6.31 (0.44)
6.28 (0.48)
6.26 (0.43)
Note. Word frequency counts are retrieved from the Westbury Lab USENET Corpus (Shaoul & Westbury, 2006).
Imageability norms from the available words were retrieved from the Glasgow Norms (Scott et al., 2019).
Neurobiology of Language
4
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
1
1
9
8
6
8
4
4
n
o
_
a
_
0
0
0
4
5
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Analogy-related information accessed by simple arithmetic in fMRI patterns
Procedure
Prior to the scanning session, participants filled out the Edinburgh Handedness Inventory form
(Veale, 2014) and an adult background form; the forms were to screen whether participants
were right-handed and native English speakers respectively. Afterward, they were asked to
write down three properties for each of the 45 words on a sheet. This procedure was to ensure
that participants were familiar with each word. Note that participants were not provided with
the categories and themes throughout the whole experiment and that they were not instructed
to perform any task relevant to analogical reasoning.
At the beginning of each run, participants were instructed to form a corresponding mental
image upon seeing a word on the screen. For the 45 stimulus word trials, a word selected from
Table 1 would appear on the screen for 3 s, followed by a blank screen interstimulus jitter of 3, 4,
or 5 s. To engage participants in the task, the stimulus word trials were intermixed with 3 catch
trials, where participants saw one of three questions (“Is the previous word a person/building/
tool?”) and were instructed to press the left button for answering “No” and the right button for
“Yes.” All 45 stimuli appeared exactly once in each run (48 trials), and the trial order was shuffled
with the exception that a catch trial could not be the first trial. On average, each run took 324 s
(45 word-stimulus trials × 7 s + 3 catch trials × 3 s), which is 162 TRs (where TRs represents
repetition time). Participants performed seven runs in total; they completed one practice run
on a computer before the scanner session and six runs in the scanner.
Image Acquisition
Whole-brain images were acquired with a 3-T Siemens MAGNETOM PrismaFit scanner with a
64-channel head coil. A high-resolution structural image was acquired using a T1-weighted
MP-RAGE sequence (repetition time = 2,530 ms, echo time = 2.34 ms, flip angle = 7°, field of
view = 256 mm, matrix = 256 × 256, 1 × 1 × 1 mm sagittal left-to-right slices). Afterward, each
participant was scanned for six functional runs (five runs for one participant). T2*-weighted
functional images were acquired with an interleaved echo planar imaging (EPI) pulse se-
quence (repetition time = 2,000 ms, echo time = 30 ms, flip angle = 70°, field of view =
256 × 256 mm, matrix = 128 × 128, 90 foot-to-head slices, voxel size = 2 × 2 × 2 mm).
The first six volumes of each run were discarded to allow the signal to reach steady-state
equilibrium.
fMRI Data Preprocessing
Structural and functional images were processed using statistical parametric mapping (SPM12,
v6906; https://www.fil.ion.ucl.ac.uk/spm/software/), and the following preprocessing stages
were performed. EPI images were first corrected for head motion and slice time acquisition.
Functional images were co-registered to the T1-weighted image. The T1-weighted structural
image was segmented into tissue maps, and the resulting deformation field was applied to the
functional images to spatially normalize them. No spatial smoothing or voxel spatial clustering
was further applied to the images. Preprocessed data were later analyzed with the general
linear model. We included 300 regressors in the design matrix: 288 regressors of interest
where the onset of each experiment trial (48 trials × 6 runs) was convolved with the standard
hemodynamic response function, 6 constant regressors (one for each run) to account for
run-to-run signal variations, and 6 motion nuisance regressors. Finally, the blood oxygen
level-dependent (BOLD) activation time series for each voxel was fitted to these regressors.
The fitted beta coefficients of the three catch trials for each run were discarded and not used
for further analyses. The beta coefficients across all runs within each voxel were standardized
Neurobiology of Language
5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
1
1
9
8
6
8
4
4
n
o
_
a
_
0
0
0
4
5
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Analogy-related information accessed by simple arithmetic in fMRI patterns
(i.e., rescaled to have zero mean and unit variance). Note that standardizing the beta coeffi-
cients without excluding the target word trials may cause the double-dipping problem
(Kriegeskorte et al., 2009) and lead to inflated rankings. To prevent this, we calculated the
mean and standard deviation for each voxel on all trials except the target word trials, and
we standardized all beta coefficients (including the target word trials) by subtracting out this
mean and dividing by this standard deviation. Finally, the fMRI pattern of a word was deter-
mined by concatenating the average beta coefficients over the six corresponding trials from
the selected voxels.
Voxel Selection Criterion
For all analyses in the current study, only a subset of the voxels that were relevant to the task
was used for the ranking analyses. We chose voxels with the highest stability scores (described
below) from all valid voxels in a given region of interest (ROI), i.e., whole brain and the selected
brain regions respectively to test H1 and H2. Note that the voxels selected need not be spatially
connected, and the selection criterion is not based on the results or significance statistics. For
each voxel, we calculated a stability score: a 6-by-6 (or 5-by-5) inter-run correlation matrix was
first calculated by taking the Pearson correlation between runs, and the stability score was ac-
quired by averaging the upper triangular part of this matrix (see Mitchell et al., 2008 for similar
methods). Intuitively, this criterion selected voxels that were consistently activated by the word
stimuli across multiple runs. To ensure that the analyses aren’t sensitive to the voxel selection
criterion, we selected a wide range number of stable voxels (i.e., from 100 to 6,400 voxels) for
the first analyses. For the ROI analyses, the most stable 100 voxels in each ROI were selected.
To exclude the fMRI activation patterns of the target word from the calculation of the predicted
fMRI pattern (described in the Experimental Design and Statistical Analysis section), for every
analogy question we calculated the voxel stability scores without the target word.
Experimental Design and Statistical Analysis
To test the first hypothesis (H1) of whether analogy-related information can be accessed from
adding and subtracting fMRI activation patterns associated with individual words, we devised
five ranking metrics (Figure 2) to examine whether information about the identity, category,
and theme of a word can be accessed. Intuitively, if the predicted fMRI pattern contains in-
formation about certain categories, it should be more similar to the word patterns in that cat-
egory than others (and the ranking would be higher). Note that ranking metrics instead of raw
distance metrics were used since we are predominantly interested in whether the distance
between the predicted pattern and the ground truth word is shorter than that for other words,
not the magnitude of the distance itself. Furthermore, the various analyses performed with
different numbers of voxels and brain regions can be compared directly with the ranking
metrics.
Each test metric was computed in three steps: (1) construct the predicted fMRI pattern, (2)
calculate the correlations between the predicted pattern and that of candidate words, and (3)
rank the correlations. To illustrate how the ranking metrics were calculated, we used the fol-
lowing example analogy: teacher is to chalk as mechanic is to (wrench), where wrench is the
target word. Similar to the arithmetic operations used in word2vec (Mikolov et al., 2013), the
predicted fMRI pattern of the target word was simply calculated as the addition/subtraction of
other word fMRI patterns in the question:
wrenchpred ¼ mechanic þ chalk − teacher
(1)
6
Neurobiology of Language
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
1
1
9
8
6
8
4
4
n
o
_
a
_
0
0
0
4
5
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Analogy-related information accessed by simple arithmetic in fMRI patterns
Figure 2. Description of the ranking metrics. Top: The example analogy question and the arith-
metic operations we applied to create the predicted fMRI pattern (i.e., wrenchpred). The target word
(i.e., wrench) is underlined. Bottom: The three main ranking metrics. For each metric, the Pearson
correlation coefficient between wrenchpred and the target pattern (in bold font) was ranked against
the correlation between wrenchpred and all other candidate words. The rank was scaled to [0, 1]
and averaged across all four target words in all possible analogy questions. In the identity metric the
target pattern was the fMRI pattern of the target word (i.e., wrench), and all other words in the
stimuli list, except the four words in the analogy question, were candidate words. In the category
metric, category templates, which were the averaged fMRI patterns across all remaining words in
each category, were used instead of the fMRI patterns of individual words. The target category tem-
plate was the category that the word actually belonged to, and the two other templates served as
candidates. In the theme metric, the target pattern was the word in the same theme and the final
unused category (i.e., garage), and all other words in the unused category (i.e., building words)
were candidate words.
The ranking for this target word was calculated as:
1.
Identity metric: We tested whether the predicted fMRI pattern is closer to the true word
wrench compared to other words. Specifically, we calculated and ranked the Pearson
correlation coefficients between wrenchpred and the fMRI patterns of all other 42 words
in the stimuli list (the three words used to create wrenchpred, i.e., mechanic, chalk, and
teacher, were excluded). Intuitively, the ranking of the corresponding correlation coef-
ficient between wrenchpred and the ground truth pattern wrench represents how close
the predicted pattern is to the true word compared to other words. Finally, the ranking
(value between 1 and 42) is linearly scaled to the 0 ~ 1 range to facilitate comparison
between different ranking metrics, which can have different numbers of candidate
words.
2. Category metric: To test which category (i.e., person/building/tool) wrenchpred belonged
to, we tested whether it was more similar to a template canonical representation of tool
Neurobiology of Language
7
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
1
1
9
8
6
8
4
4
n
o
_
a
_
0
0
0
4
5
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Analogy-related information accessed by simple arithmetic in fMRI patterns
words than person words and building words. First, the template of each category was
calculated by taking the voxel-wise mean of the fMRI patterns of all 13 remaining
words in that category (e.g., the tool template was the average of the tool words that
remained after the two tool words in the analogy question were excluded). Second,
we ranked the Pearson correlation coefficients between wrenchpred and the three cat-
egory templates. Finally, the rank associated with the true category template was
scaled to the 0 ~ 1 range.
4.
3. Theme metric: To test which theme (e.g., education, shopping, etc.) wrenchpred be-
longed to, we first gathered all words belonging to the last category unused in the anal-
ogy question (e.g., building words in the example question), and tested whether
wrenchpred was closer to the word in the same theme (i.e., garage) compared to the
other 13 words (e.g., mall, court). Similarly to other metrics, we extracted the rank of
the correlation between wrenchpred and garage and scaled it to the 0 ~ 1 range.
Identity (within-category) metric: This metric serves to eliminate the confound that the
identity metric can be significant when the fMRI patterns only encode category infor-
mation. Unlike the Identity metric, the Pearson correlation coefficients were only cal-
culated between wrenchpred and the fMRI patterns of all other words in the true
category. Similarly, the rank of correlation between wrenchpred and the ground truth
pattern wrench is scaled to the 0 ~ 1 range.
Identity (close foil) metric: In standard analogical reasoning paradigms, a target word is
usually probed against a close foil word that answers the analogy question partially; for
instance, garage is a close foil word as it has the same theme as but a different category
from the target word. We tested whether wrenchpred is closer to the ground truth pattern
wrench than the fMRI pattern of a close foil word, which was selected in the current
study to be the unused word in the same theme (e.g., garage).
5.
The procedure above describes how the five ranking metrics were calculated in one anal-
ogy question. To assess whether analogy-related information can be extracted from fMRI pat-
terns in a ROI, we performed a group-level statistical analysis. To facilitate this, in each of the
five tests, we reduced each individual’s rank scores to a single summary metric, by averaging
ranks (within each participant) across all analogy questions. We then performed a group-level
test of whether the mean rankings were greater than data-driven estimates of chance-level
computed for each participant. To determine the chance level, we first randomly shuffled
the correspondence between word labels and their fMRI patterns, ran the ranking metrics with
the shuffled data, and calculated the average ranking across all analogy questions. This per-
mutation process was repeated 1,000 times for each participant, each ROI, and each metric,
and we aggregated the 1,000 results to get the null mean. Finally, for each ROI and each met-
ric, we performed a one-sided paired t test between the real ranking metrics (13 points, one
from each participant) and the null mean from each participant (also 13 points). Note that
since the rankings were scaled to the 0 ~ 1 range, the null means were close to the theoretical
chance level of 0.5 (M = 0.4999, SD = 0.001). The paired t test is preferred over the standard
one since we can empirically estimate the chance level instead of assuming it to be 0.5, and
the two tests yield quantitatively similar results. For the ROI-level analysis, p values associated
with multiple ROIs were corrected for multiple comparisons using the false discovery rate
method (Benjamini & Hochberg, 1995). In light of recent concerns over statistical testing in
multivariate pattern analyses (Allefeld et al., 2016), it is important to note that the current rank
measure can be any value between 0 and 1 and should be distinguished from information-like
measures such as classification accuracy.
Neurobiology of Language
8
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
1
1
9
8
6
8
4
4
n
o
_
a
_
0
0
0
4
5
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Analogy-related information accessed by simple arithmetic in fMRI patterns
Finally, to claim that analogy-related information can be accessed from the fMRI patterns,
we consider that not only the identity metric but also the category and theme metrics must
yield test statistics that are significantly above chance. This is because above-chance ranks
in the identity metric do not automatically entail that information reflecting both the theme
and category of the word is represented in fMRI. Thus, if only the category but not the theme
is encoded in fMRI we might end up with computations (in place of Equation 1) that reduce to:
wrenchpred ¼ person þ tool − person ¼ tool
(2)
Since wrench is a tool, the predicted fMRI pattern would be ranked higher than the other
person and building words in the identity metric (rank = 0.66) and therefore above the 0.5
chance-level. One way to eliminate this possibility is to make sure that both the category
and the theme metrics are significantly above chance, and therefore the fMRI patterns encode
both category and theme information. Alternatively, a significant identity (within-category)
metric can also suggest that the word identity information can be accessed even among stim-
ulus words in the same category. In sum, we consider that the identity, category, and theme
ranking metrics must all be significantly above chance to claim evidence of successfully acces-
sing analogy-related information.
Regions of Interests Analysis
To test the second hypothesis (H2) of whether analogy-related information can be accessed
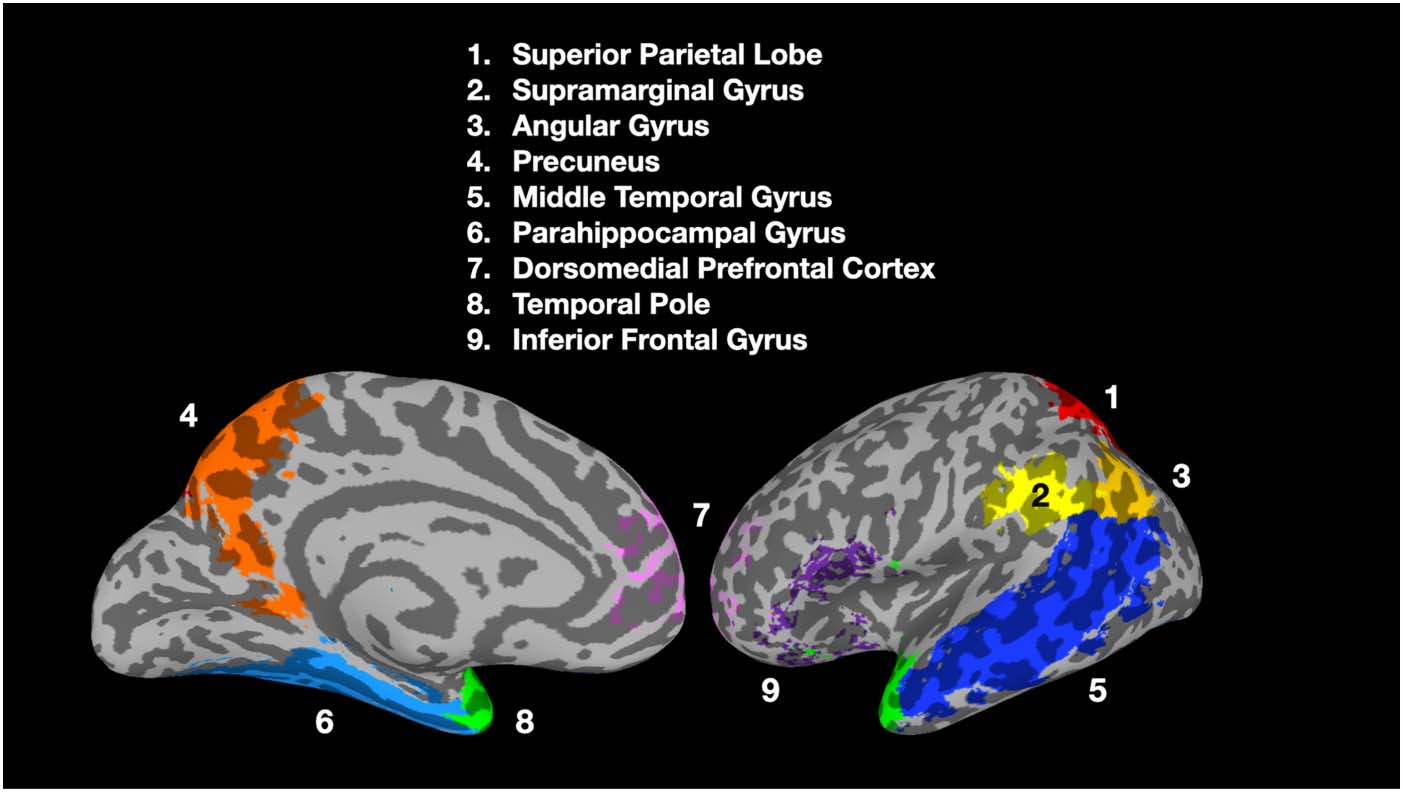
within particular brain regions (as opposed to across brain regions), nine anatomically prespe-
cified ROIs (Figure 3) were selected. These ROIs were selected for their roles in various seman-
tic processing and analogical reasoning (Carota et al., 2017; Chiang et al., 2021; Hobeika et al.,
2016) tasks, such as processing semantic category relations (Schwartz et al., 2011) and repre-
senting semantic similarity among concepts (Patterson et al., 2007). The parietal lobe ROIs were
created using the automated anatomical labeling atlas (Rolls et al., 2015; Tzourio-Mazoyer
et al., 2002). The inferior frontal gyrus and the dorsomedial prefrontal cortex were created from
the union of BA 44, 45, and 47 and the union of BA 2 and 10 respectively, where the regions
Figure 3. Nine prespecified semantic related ROIs. Colors denote different ROIs and not the sig-
nificance of results.
Neurobiology of Language
9
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
3
1
1
1
9
8
6
8
4
4
n
o
_
a
_
0
0
0
4
5
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Analogy-related information accessed by simple arithmetic in fMRI patterns
were collected from the Wake Forest University ( WFU) Pickatlas toolbox (https://school
.wakehealth.edu/Research/Labs). While these ROIs were defined on the left hemisphere due
to the left hemispheric dominance observed in the aforementioned studies, we also conducted
a secondary analysis using the right hemispheric homologs of these ROIs in light of a reviewer’s
query (see the results in Supp. Figure 2; Supporting information can be found online at https://
doi.org/10.1162/nol_a_00045). We note that this is a post-hoc analysis since the ROIs were not
selected prior to the experiment.
RESULTS
Task-General fMRI Patterns of Individual Words Contain Information About Analogy Relations
Can analogy-related information be accessed from the addition and subtraction of fMRI acti-
vation patterns? We tested five ranking metrics (Figure 2) using fMRI patterns extracted from
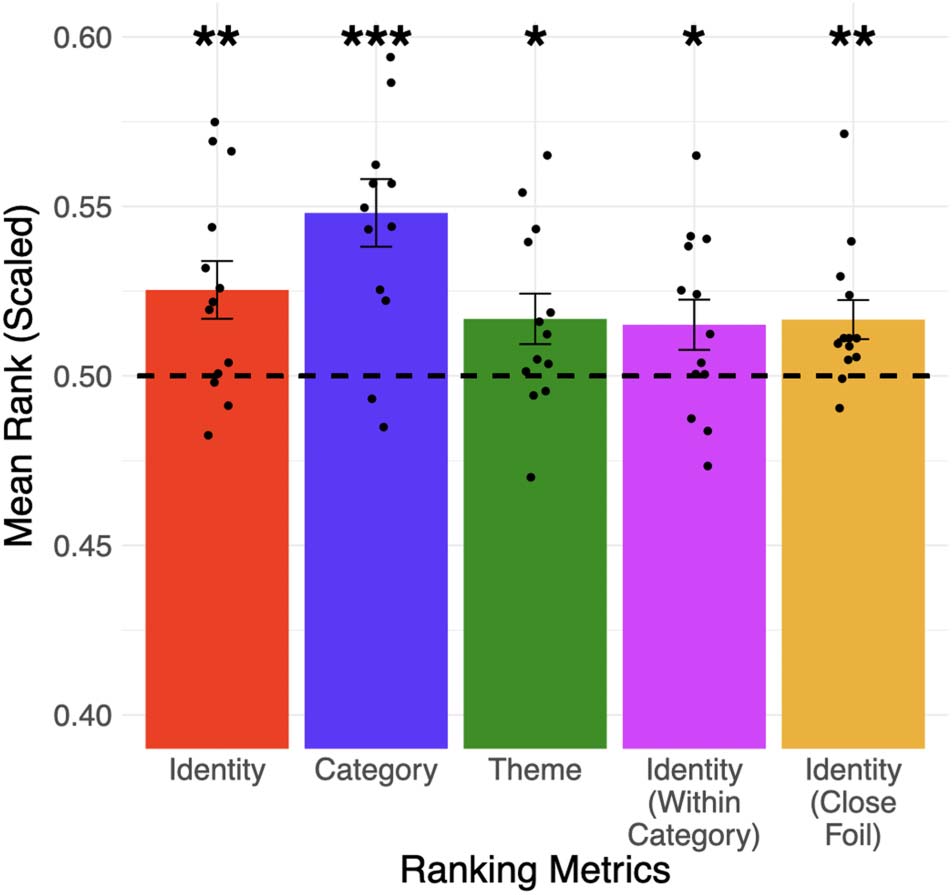
the whole brain. We found that the identity (paired t test, mean = 0.525, SD = 0.03, p = 0.006)
and category (paired t test, mean = 0.548, SD = 0.04, p < 0.001) metrics are significantly higher
than chance at the group-level (i.e., across all participants) (Figure 4), and both effects are stable
across a wide range of voxel selection criteria (up to 3,200 voxels, see Supp. Table 1). The theme
metric is also significantly above chance (paired t test, mean = 0.517, SD = 0.03, p = 0.02),
but only when up to 200 voxels are selected. We also found significant results (paired t test,
mean = 0.515, SD = 0.03, p = 0.03) with the identity (within-category) metric (i.e., we tested
whether the predicted pattern is closer to the actual patterns compared to other words in the
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
3
1
1
1
9
8
6
8
4
4
n
o
_
a
_
0
0
0
4
5
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Information about the identity (e.g., wrench vs. 41 other words), category (e.g., tool vs.
Figure 4.
two other categories), theme (e.g., repairing vs. all 13 other themes) of a word can be accessed from
the addition and subtraction of fMRI patterns of the three other words in an analogy question. The
identity of the target word can also be distinguished from words in its category (4th column) and a
foil word from its theme (5th column). Error bars represent the standard error of the mean across
participants (* p < 0.05; ** p < 0.01; *** p < 0.001). The dotted line represented chance-level per-
formance (mean rank = 0.5) derived from the participant-level permutation process. Black dots rep-
resent the mean rank of each participant. This figure only displays results from fMRI patterns where
100 voxels were selected according to the voxel selection procedure (see Materials and Methods),
and they do not need to be spatially connected; see Supp. Table 1 for the full results.
Neurobiology of Language
10
Analogy-related information accessed by simple arithmetic in fMRI patterns
same category); this eliminated the confound that the identity metric can be significant when
the fMRI patterns only encode category information. Finally, we drew inspiration from com-
mon analogical reasoning paradigms and found that the fMRI patterns of the actual word can
be reliably distinguished from that of a close foil (i.e., the identity (close-foil) metric is signif-
icant: paired t test, mean = 0.517, SD = 0.02, p = 0.007). Overall, the results support our first
hypothesis (H1) that analogy-related information from individual words can be reliably ac-
cessed, but further analysis of the ROI-level is required.
Word Categories and Themes Were Jointly Represented in the Parietal Lobe
We further investigated whether analogy-related information can be accessed within single
brain regions (as opposed to extracting fMRI patterns across multiple regions). While previous
large-scale neuroimaging studies suggested that semantic properties were widely distributed
across the whole brain (Huth et al., 2012, 2016), other studies argued that category and theme
information were localized in distinct brain regions (Kalénine et al., 2009; Sachs et al., 2008;
Schwartz et al., 2011). It is therefore of scientific interest to examine whether analogy-related
information can be accessed within a single region (i.e., both category and theme information
were represented). As a result, we ran the five ranking metrics on nine prespecified ROIs in the
left hemisphere (Figure 3); these hypothesis-driven ROIs were selected based on their role in
processing semantic and analogical relations (Carota et al., 2017; Chiang et al., 2021;
Schwartz et al., 2011), representing semantic similarity among concepts (Patterson et al.,
2007), and as a transmodal hub to generalize across various modality-specific semantic infor-
mation (Lambon Ralph et al., 2017).
As shown in Table 3, among the nine ROIs, the identity ranking of the left superior parietal
lobe, supramarginal gyrus, angular gyrus, precuneus, and middle temporal gyrus were signifi-
cantly above chance. The category metric was significantly above chance in most ROIs except
the dorsomedial prefrontal cortex and temporal pole. Regarding the theme metric, while none
Table 3.
hemisphere)
The mean rank (M ) and corrected p values of five ranking metrics in the nine prespecified semantic-related ROIs (all in the left
ROIs (left)
Superior parietal lobe
Identity
M
p
Category
p
M
Theme
M
p
0.521
0.03*
0.538
0.02*
0.516
0.08
Identity
( within category)
M
0.513
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
3
1
1
1
9
8
6
8
4
4
n
o
_
a
_
0
0
0
4
5
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Supramarginal gyrus
0.526
0.003**
0.541
0.01*
0.519
0.05 (0.006)
0.518
Angular gyrus
0.522
0.02*
0.530
0.003**
0.518
0.05 (0.01)
Precuneus
0.525
0.03*
0.532
0.02*
0.519
0.08
Middle temporal gyrus
0.521
0.03*
0.535
0.03*
0.511
0.08
Parahippocampal gyrus
0.516
0.16
0.526
0.003**
0.511
0.32
Dorsomedial prefrontal
0.505
0.26
0.528
0.07
0.501
0.43
cortex
Temporal pole
0.513
0.22
0.522
0.29
0.506
0.43
Inferior frontal gyrus
0.511
0.17
0.521
0.05*
0.507
0.32
0.517
0.519
0.512
0.511
0.500
0.507
0.506
Identity
(close foil)
p
M
0.508
0.18
0.516
0.03*
0.506
0.03*
0.510
0.07
0.514
0.03*
0.510
0.03*
0.505
0.26
0.511
0.04*
0.503
0.18
p
0.11
0.05
0.05
0.11
0.12
0.36
0.43
0.38
0.36
Note. Uncorrected p values are shown in parentheses. Significant metrics are marked in bold. While category information seems to be represented in wide-
spread regions, theme information is only present in the parietal lobe regions.
Neurobiology of Language
11
Analogy-related information accessed by simple arithmetic in fMRI patterns
Information about the identity (e.g., wrench vs. 41 other words), category (e.g., tool vs.
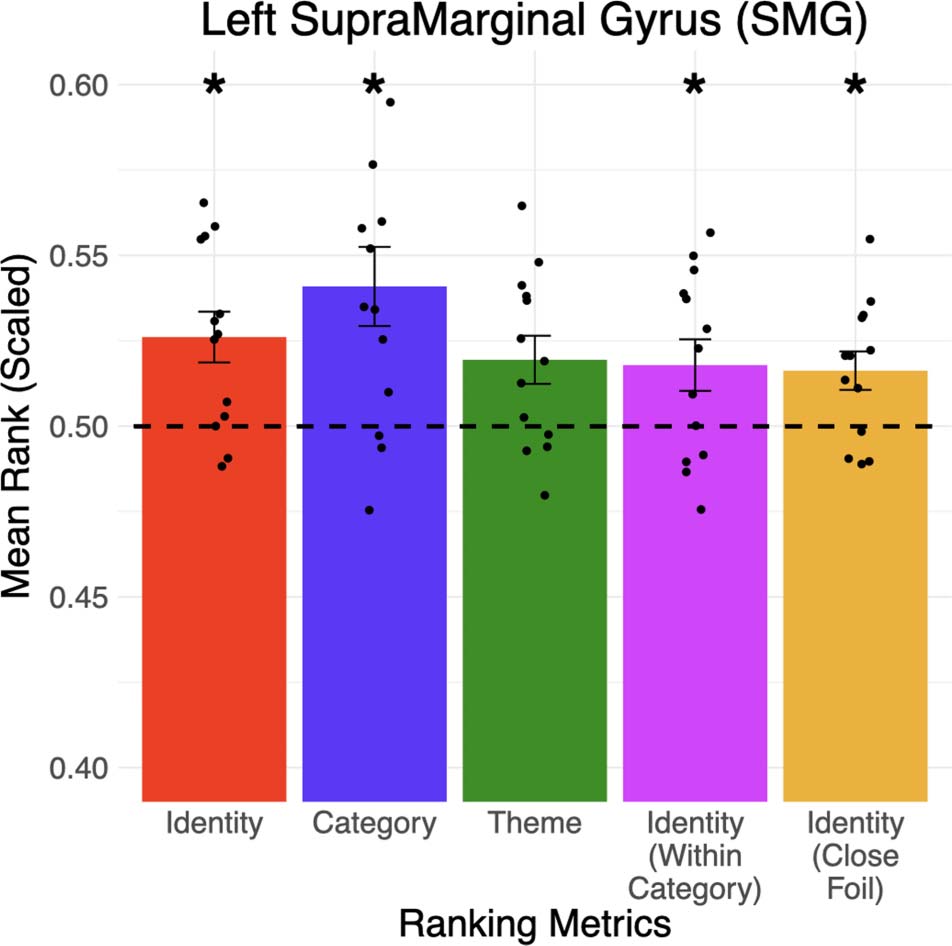
Figure 5.
two other categories), and theme (e.g., repairing vs. all 13 other themes) of a word can be accessed
from the addition and subtraction of fMRI patterns in the left supramarginal gyrus. The identity of the
target word can also be distinguished within its category (4th column) and from a close foil word in
its theme (5th column). Error bars represented the standard error of the mean across participants (* p <
0.05). The dotted line represented chance-level performance (mean rank = 0.5). Black dots represent
the mean rank of each participant.
of the ROIs are significant after multiple correction, the left supramarginal gyrus (see Figure 5)
and angular gyrus were borderline ( p = 0.05 post correction). Prompted by a reviewer, we also
conducted an analysis of the eight right-hemispheric homologs of the above left-hemisphere
ROIs (see Supp. Figure 2). The findings suggested that analogy-related information was
encoded more weakly in the right hemisphere, with only category and identity metrics signif-
icantly above chance in the right supramarginal gyrus, angular gyrus, and precuneus. Overall,
the results hinted toward our second hypothesis (H2) that the fMRI patterns of individual words
in the semantic network regions encode analogy-related information, similar to the distributed
word-embedding models (Mikolov et al., 2013).
DISCUSSION
In the current study, we first tested the hypothesis (H1) of whether analogy-related information
can be accessed from adding and subtracting fMRI activation patterns elicited by viewing in-
dividual words. We demonstrated that information about the identity, category, and theme of
the target word can be accessed from the fMRI activation patterns. Critically, it is important to
note that participants were only viewing individual words in the experiment without knowing
the structure of the stimuli list, and they were not asked to perform analogical reasoning of any
sort. This implies that like word embeddings (e.g., word2vec) extracted from artificial neural
networks, task-general semantic representations of individual words in the human brain are
sufficiently rich that simple arithmetic operations can be directly applied to them to access
analogy-related information, and that explicit analogical reasoning tasks are not necessary
to activate them.
Neurobiology of Language
12
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
3
1
1
1
9
8
6
8
4
4
n
o
_
a
_
0
0
0
4
5
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Analogy-related information accessed by simple arithmetic in fMRI patterns
We further tested the second hypothesis (H2) of whether analogy-related information can
be accessed within individual semantic network regions, or whether it is necessary to integrate
voxels from multiple areas. Our results demonstrated that while the category of a target word
could be accessed from fMRI patterns extracted from multiple parietal, temporal, and frontal
regions, theme information can only be accessed within the left parietal lobe region. This is
consistent with previous studies demonstrating that first-order analogical relations were pres-
ent in the superior parietal cortex (Chiang et al., 2021). The finding that category information
was encoded in more regions may have arisen from the category test being more powerful, or
the particular categories and themes comprising the stimuli. While this result was consistent
with previous studies demonstrating that the ATL encoded category relations and the IPL
encoded thematic relations (Schwartz et al., 2011), it newly reveals that the multiple parietal
lobe regions also contained category information. However, since tool was one of the three
categories used in the current study and task-general tool processing is associated with left
IPL activation (Ishibashi et al., 2016), it is not altogether surprising that the IPL could distin-
guish tool words from other categories. Overall, the results suggest that the intrinsic organi-
zation of semantic representations within an individual brain region contains analogy-related
information locally (by addition and subtraction).
The ranking accuracies in the current study were not high, although they were nonetheless
significantly above chance. This could have been for several reasons. First, the predicted fMRI
patterns for the target word were calculated based on integrating information across fMRI
activation patterns corresponding to only three words. This is a very small training set in com-
parison to contemporary studies that have not only used more words to correlate with fMRI
data but also applied regression-based approaches to map fMRI patterns to a semantic model
(e.g., Mitchell et al., 2008 used 58 words as a training set). Indeed, the current approach that
tests the hypotheses without parameter fitting resonates with our previous approach
(Anderson et al., 2016) to sidestep model overfitting problems and combine the strength of
both encoding analysis and representational similarity analysis (Kriegeskorte et al., 2008).
Second, while previous studies have found better model performance with richer stimuli for-
mats such as words associated with line drawings (Mitchell et al., 2008), images, sentences,
and word clouds (Pereira et al., 2018), we showed participants individual words only. While
this paradigm has seen success in previous studies (Just et al., 2010), recent studies comparing
fMRI word decoding performance between different stimuli formats hinted that presenting
stimuli in multiple modalities may activate a broader network of brain regions and lead to
higher accuracies (Pereira et al., 2018; S. Wang et al., 2020).
It is critical to restate that the fMRI patterns were elicited when participants were not per-
forming an analogical reasoning task, since this study aimed to examine whether analogy-
related information could be accessed from task-general fMRI patterns. If, contrary to what
was actually done, we had instructed participants to perform an analogical reasoning task,
then we would have risked activation specific to the analogy task obscuring and/or modifying
the task-general representations of isolated words we sought to test. For instance, in a typical
analogical reasoning task, participants might see two pairs of words and then determine
whether the relations between the two pairs were identical or not. While it has been demon-
strated that different types of relations (e.g., similarity and contrast) can be decoded from
semantic network regions (Chiang et al., 2021), it remains unclear how they can be calcu-
lated from the task-general fMRI patterns of their component words. Since the current study
demonstrated that task-general fMRI patterns were intrinsically organized and contain
analogy-related information, it serves as initial evidence that a potential mechanism to cal-
culate such relations is to simply add and subtract fMRI patterns of individual words. Future
Neurobiology of Language
13
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
3
1
1
1
9
8
6
8
4
4
n
o
_
a
_
0
0
0
4
5
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Analogy-related information accessed by simple arithmetic in fMRI patterns
studies are necessary to elucidate the relationship between task-general fMRI patterns and
the representations elicited during explicit analogical reasoning tasks, and whether the for-
mer are recruited in the construction of the latter.
The current study complements previous fMRI studies of analogical reasoning (Chiang
et al., 2021) in the following respects. First, whereas Chiang et al. (2021) examined the neural
computations underlying actual analogical reasoning, our study investigated: (1) participants
who had no knowledge that the experiment was related to analogical reasoning; (2) fMRI
activation elicited by viewing isolated words; and (3) whether fMRI activation patterns asso-
ciated with isolated words were inherently structured to reflect their analogical relationship.
Second, Chiang et al. (2021) demonstrated that fMRI activation patterns elicited during analog-
ical reasoning were correlated with patterns of features that explicitly estimated how pairs of
word2vec vectors were related, as synonyms, antonyms, cause-effect, and so on (i.e., BART; Lu
et al., 2012). Word2vec was not used in the current study; instead, the simple arithmetic oper-
ations of addition and subtraction used to access analogy-related information were directly
applied to combine fMRI activation patterns associated with individual words.
The current study has several limitations. First, this study only considered concrete nouns
that were analogically related to each other categorically or thematically. While previous stud-
ies have shed light on how abstract concepts were structured in the human brain (Anderson
et al., 2014, 2017; Pereira et al., 2018; Vargas & Just, 2019; X. Wang et al., 2018), future work
is needed to investigate whether the semantic organization of abstract nouns or adjectives in
the brain is organized such that analogy-related information can be similarly accessed.
Second, the tool words in the stimuli list have lower word frequency compared to words in
other categories, and less-frequent words might elicit higher activation values in brain regions
such as the left inferior frontal gyrus (Schuster et al., 2016). However, at the same time, the
tool words were not obscure (e.g., gun, wrench, skillet) and word frequency alone could not
account for all of the ranking metrics, in particular the theme metric that compares words
within the same category. Third, the categories chosen are quite distinct, and it is challenging
to eliminate every confounder that varies between any two categories; for instance, living vs.
nonliving can also differentiate person words from the others. Again, these confounders can-
not drive the significant results in the theme metric. Further studies will be necessary to test
whether the current results can be generalized to finer-grained categories. Finally, our word
stimuli varied in length, because it was beyond our ability to devise a suitable list of words
with the same length. While word length is an important factor in semantic processing and
reading (Just et al., 2010; Schuster et al., 2016), its influence has mostly been observed in the
occipital pole and lingual/fusiform gyri (Just et al., 2010) and is unlikely to have had a sub-
stantial influence on the results of the current study (e.g., Figure 5).
Conclusions
In sum, the current study has demonstrated that analogy-related information can be accessed
by applying addition and subtraction to fMRI patterns elicited by reading isolated words. It has
further identified regions of the brain’s semantic network that represent semantic categories
and themes and revealed evidence that word representations within the left parietal lobe region
could sufficiently contain such information. For the broader literature investigating how humans
perform analogical reasoning tasks, the current study has provided initial evidence that the
relation between two words can be approximated by applying simple addition and subtrac-
tion on task-agnostic fMRI patterns of individual words.
Neurobiology of Language
14
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
3
1
1
1
9
8
6
8
4
4
n
o
_
a
_
0
0
0
4
5
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Analogy-related information accessed by simple arithmetic in fMRI patterns
ACKNOWLEDGMENTS
We thank Carol Jew for insightful comments and discussions. We also thank Dave
Kleinschmidt for providing analysis code. This work was supported by a Google Faculty
Award, NSF CAREER award 1652127, and NSF research grant DRL-1561335.
FUNDING INFORMATION
Rajeev Raizada, Google (https://dx.doi.org/10.13039/100006785). Rajeev Raizada, National
Science Foundation (https://dx.doi.org/10.13039/100000001), Award ID: 1652127. Robert A.
Jacobs, National Science Foundation (https://dx.doi.org/10.13039/100000001), Award ID:
DRL-1561335.
AUTHOR CONTRIBUTIONS
Meng-Huan Wu: Conceptualization: Equal; Data curation: Lead; Formal analysis: Lead;
Methodology: Lead; Software: Lead; Visualization: Lead; Writing – original draft: Lead. Andrew
J. Anderson: Conceptualization: Equal; Data curation: Supporting; Supervision: Supporting;
Writing – review & editing: Supporting. Robert A. Jacobs: Supervision: Lead; Writing – review
& editing: Supporting. Rajeev D. S. Raizada: Supervision: Lead; Writing – review & editing:
Supporting.
REFERENCES
Allefeld, C., Görgen, K., & Haynes, J.-D. (2016). Valid population
inference for information-based imaging: From the second-level
t-test to prevalence inference. NeuroImage, 141, 378–392.
https://doi.org/10.1016/j.neuroimage.2016.07.040, PubMed:
27450073
Anderson, A. J., Binder, J. R., Fernandino, L., Humphries, C. J.,
Conant, L. L., Raizada, R. D. S., Lin, F., & Lalor, E. C. (2019).
An integrated neural decoder of linguistic and experiential mean-
ing. Journal of Neuroscience, 39(45), 8969–8987. https://doi.org
/10.1523/JNEUROSCI.2575-18.2019, PubMed: 31570538
Anderson, A. J., Bruni, E., Lopopolo, A., Poesio, M., & Baroni, M.
(2015). Reading visually embodied meaning from the brain:
Visually grounded computational models decode visual-object
mental imagery induced by written text. NeuroImage, 120,
309–322. https://doi.org/10.1016/j.neuroimage.2015.06.093,
PubMed: 26188260
Anderson, A. J., Kiela, D., Clark, S., & Poesio, M. (2017). Visually
grounded and textual semantic models differentially decode
brain activity associated with concrete and abstract nouns.
Transactions of the Association for Computational Linguistics,
5, 17–30. https://doi.org/10.1162/tacl_a_00043
Anderson, A. J., Murphy, B., & Poesio, M. (2014). Discriminating
taxonomic categories and domains in mental simulations of con-
cepts of varying concreteness. Journal of Cognitive Neuroscience,
26(3), 658–681. https://doi.org/10.1162/jocn_a_00508, PubMed:
24168217
Anderson, A. J., Zinszer, B. D., & Raizada, R. D. S. (2016).
Representational similarity encoding for fMRI: Pattern-based syn-
thesis to predict brain activity using stimulus-model-similarities.
NeuroImage, 128, 44–53. https://doi.org/10.1016/j.neuroimage
.2015.12.035, PubMed: 26732404
Benjamini, Y., & Hochberg, Y. (1995). Controlling the false discovery
rate: A practical and powerful approach to multiple testing. Journal
of the Royal Statistical Society: Series B (Methodological), 57(1),
289–300. https://doi.org/10.1111/j.2517-6161.1995.tb02031.x
Carota, F., Kriegeskorte, N., Nili, H., & Pulvermüller, F. (2017).
Representational similarity mapping of distributional semantics
in left inferior frontal, middle temporal, and motor cortex.
Cerebral Cortex, 27(1), 294–309. https://doi.org/10.1093/cercor
/bhw379, PubMed: 28077514
Chiang, J. N., Peng, Y., Lu, H., Holyoak, K. J., & Monti, M. M.
(2021). Distributed code for semantic relations predicts neural
similarity during analogical reasoning. Journal of Cognitive
Neuroscience, 33(3), 377–389. https://doi.org/10.1162/jocn_a
_01620, PubMed: 32762520
Fu, R., Guo, J., Qin, B., Che, W., Wang, H., & Liu, T. (2014). Learning
semantic hierarchies via word embeddings. In Proceedings of the
52nd Annual Meeting of the Association for Computational
Linguistics ( Volume 1: Long Papers), (Baltimore, Maryland, June
2014, pp. 1199–1209). Association for Computational Linguistics.
https://doi.org/10.3115/v1/P14-1113
Gentner, D. (1983). Structure-mapping: A theoretical framework for
analogy. Cognitive Science, 7(2), 155–170. https://doi.org/10
.1207/s15516709cog0702_3
Hobeika, L., Diard-Detoeuf, C., Garcin, B., Levy, R., & Volle, E.
(2016). General and specialized brain correlates for analogical
reasoning: A meta-analysis of functional imaging studies.
Human Brain Mapping, 37(5), 1953–1969. https://doi.org/10
.1002/hbm.23149, PubMed: 27012301
Huth, A. G., de Heer, W. A., Griffiths, T. L., Theunissen, F. E., &
Gallant, J. L. (2016). Natural speech reveals the semantic maps
that tile human cerebral cortex. Nature, 532, 453–458. https://
doi.org/10.1038/nature17637, PubMed: 27121839
Huth, A. G., Nishimoto, S., Vu, A. T., & Gallant, J. L. (2012). A con-
tinuous semantic space describes the representation of thousands
of object and action categories across the human brain. Neuron,
Neurobiology of Language
15
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
3
1
1
1
9
8
6
8
4
4
n
o
_
a
_
0
0
0
4
5
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Analogy-related information accessed by simple arithmetic in fMRI patterns
76(6), 1210–1224. https://doi.org/10.1016/j.neuron.2012.10
.014, PubMed: 23259955
Ishibashi, R., Pobric, G., Saito, S., & Lambon Ralph, M. A. (2016).
The neural network for tool-related cognition: An activation like-
lihood estimation meta-analysis of 70 neuroimaging contrasts.
Cognitive Neuropsychology, 33(3–4), 241–256. https://doi.org
/10.1080/02643294.2016.1188798, PubMed: 27362967
Just, M. A., Cherkassky, V. L., Aryal, S., & Mitchell, T. M. (2010). A
neurosemantic theory of concrete noun representation based on
the underlying brain codes. PLOS ONE, 5(1), Article e8622.
https://doi.org/10.1371/journal.pone.0008622, PubMed:
20084104
Kalénine, S., Peyrin, C., Pichat, C., Segebarth, C., Bonthoux, F., &
Baciu, M. (2009). The sensory-motor specificity of taxonomic
and thematic conceptual relations: A behavioral and fMRI study.
NeuroImage, 44(3), 1152–1162. https://doi.org/10.1016/j
.neuroimage.2008.09.043, PubMed: 18977304
Kriegeskorte, N., Mur, M., & Bandettini, P. A.
(2008).
Representational similarity analysis: Connecting the branches
of systems neuroscience. Frontiers in Systems Neuroscience,
2(4). https://doi.org/10.3389/neuro.06.004.2008, PubMed:
19104670
Kriegeskorte, N., Simmons, W. K., Bellgowan, P. S., & Baker, C. I.
(2009). Circular analysis in systems neuroscience: The dangers of
double dipping. Nature Neuroscience, 12(5), 535–540. https://
doi.org/10.1038/nn.2303, PubMed: 19396166
Lambon Ralph, M. A., Jefferies, E., Patterson, K., & Rogers, T. T.
(2017). The neural and computational bases of semantic cogni-
tion. Nature Reviews Neuroscience, 18(1), 42–55. https://doi.org
/10.1038/nrn.2016.150, PubMed: 27881854
Lewis, G. A., Poeppel, D., & Murphy, G. L. (2015). The neural ba-
ses of taxonomic and thematic conceptual relations: An MEG
study. Neuropsychologia, 68, 176–189. https://doi.org/10.1016
/j.neuropsychologia.2015.01.011, PubMed: 25582406
Lu, H., Chen, D., & Holyoak, K. J. (2012). Bayesian analogy with re-
lational transformations. Psychological Review, 119(3), 617–648.
https://doi.org/10.1037/a0028719, PubMed: 22775500
Mikolov, T., Sutskever, I., Chen, K., Corrado, G. S., & Dean, J.
(2013). Distributed representations of words and phrases and
their compositionality. In C. J. C. Burges, L. Bottou, M. Welling,
Z. Ghahramani, & K. Q. Weinberger (Eds.), NIPS ’13:
Proceedings of the 26th International Conference on Neural
Information Processing Systems (Lake Tahoe, Nevada,
December 2013, pp. 3111–3119). Curran Associates.
Mitchell, T. M., Shinkareva, S. V., Carlson, A., Chang, K.-M.,
Malave, V. L., Mason, R. A., & Just, M. A. (2008). Predicting
human brain activity associated with the meanings of nouns.
Science, 320(5880), 1191–1195. https://doi.org/10.1126
/science.1152876, PubMed: 18511683
Patterson, K., Nestor, P. J., & Rogers, T. T. (2007). Where do you
know what you know? The representation of semantic knowl-
edge in the human brain. Nature Reviews Neuroscience,
8(12), 976–987. https://doi.org/10.1038/nrn2277, PubMed:
18026167
Peelen, M. V., & Caramazza, A. (2012). Conceptual object repre-
sentations in human anterior temporal cortex. The Journal of
Neuroscience, 32(45), 15728–15736. https://doi.org/10.1523
/JNEUROSCI.1953-12.2012, PubMed: 23136412
Pereira, F., Lou, B., Pritchett, B., Kanwisher, N., Botvinick, M., &
Fedorenko, E. (2016). Decoding of generic mental representa-
tions from functional MRI data using word embeddings.
BioRxiv, 057216. https://doi.org/10.1101/057216
Pereira, F., Lou, B., Pritchett, B., Ritter, S., Gershman, S. J.,
Kanwisher, N., Botvinick, M., & Fedorenko, E. (2018). Toward
a universal decoder of linguistic meaning from brain activation.
Nature Communications, 9(1), Article 963. https://doi.org/10
.1038/s41467-018-03068-4, PubMed: 29511192
R o l l s , E. T . , J o l i o t , M . , & T z o u r i o - Ma z o y e r , N . ( 2 0 1 5 ) .
Implementation of a new parcellation of the orbitofrontal cortex
in the automated anatomical labeling atlas. NeuroImage, 122,
1–5. https://doi.org/10.1016/j.neuroimage.2015.07.075,
PubMed: 26241684
Sachs, O., Weis, S., Krings, T., Huber, W., & Kircher, T. (2008).
Categorical and thematic knowledge representation in the brain:
Neural correlates of taxonomic and thematic conceptual rela-
tions. Neuropsychologia, 46(2), 409–418. https://doi.org/10
.1016/j.neuropsychologia.2007.08.015, PubMed: 17920085
Schuster, S., Hawelka, S., Hutzler, F., Kronbichler, M., & Richlan,
F. (2016). Words in context: The effects of length, frequency, and
predictability on brain responses during natural reading. Cerebral
Cortex, 26(10), 3889–3904. https://doi.org/10.1093/cercor
/bhw184, PubMed: 27365297
Schwartz, M. F., Kimberg, D. Y., Walker, G. M., Brecher, A.,
Faseyitan, O. K., Dell, G. S., Mirman, D., & Coslett, H. B.
(2011). Neuroanatomical dissociation for taxonomic and thematic
knowledge in the human brain. Proceedings of the National
Academy of Sciences of the United States of America, 108(20),
8520–8524. https://doi.org/10.1073/pnas.1014935108,
PubMed: 21540329
Scott, G. G., Keitel, A., Becirspahic, M., Yao, B., & Sereno, S. C.
(2019). The Glasgow norms: Ratings of 5,500 words on nine
scales. Behavior Research Methods, 51(3), 1258–1270. https://
doi.org/10.3758/s13428-018-1099-3, PubMed: 30206797
Shaoul, C., & Westbury, C. (2006). USENET orthographic frequen-
cies for 111,627 English words (2005–2006). https://www.psych
.ualberta.ca/~westburylab/downloads/wlfreq.download.html
T urney, P. D.
(2006 ). Similarity o f semantic relations.
Computational Linguistics, 32(3), 379–416. https://doi.org/10
.1162/coli.2006.32.3.379
Tzourio-Mazoyer, N., Landeau, B., Papathanassiou, D., Crivello, F.,
Etard, O., Delcroix, N., Mazoyer, B., & Joliot, M. (2002).
Automated anatomical labeling of activations in SPM using a
macroscopic anatomical parcellation of the MNI MRI single-
subject brain. NeuroImage, 15(1), 273–289. https://doi.org/10
.1006/nimg.2001.0978, PubMed: 11771995
Vargas, R., & Just, M. A. (2019). Neural representations of abstract
concepts: Identifying underlying neurosemantic dimensions.
Cerebral Cortex, 30(4), 2157–2166. https://doi.org/10.1093
/cercor/bhz229, PubMed: 31665238
Veale, J. F. (2014). Edinburgh handedness inventory–short form: A
revised version based on confirmatory factor analysis. Laterality:
Asymmetries of Body, Brain and Cognition, 19(2),164–177.
https://doi.org/10.1080/1357650X.2013.783045, PubMed:
23659650
Waltz, J. A., Knowlton, B. J., Holyoak, K. J., Boone, K. B., Mishkin,
F. S., de Menezes Santos, M., Thomas, C. R., & Miller, B. L.
(1999). A system for relational reasoning in human prefrontal
cortex. Psychological Science, 10(2), 119–125. https://doi.org
/10.1111/1467-9280.00118
Wang, J., Cherkassky, V. L., & Just, M. A. (2017). Predicting the
brain activation pattern associated with the propositional content
of a sentence: Modeling neural representations of events and
states. Human Brain Mapping, 38(10), 4865–4881. https://doi
.org/10.1002/hbm.23692, PubMed: 28653794
Neurobiology of Language
16
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
3
1
1
1
9
8
6
8
4
4
n
o
_
a
_
0
0
0
4
5
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Analogy-related information accessed by simple arithmetic in fMRI patterns
Wang, S., Zhang, J., Wang, H., Lin, N., & Zong, C. (2020). Fine-
grained neural decoding with distributed word representations.
Information Sciences, 507, 256–272. https://doi.org/10.1016/j
.ins.2019.08.043
Wang, X., Wu, W., Ling, Z., Xu, Y., Fang, Y., Wang, X., Binder, J. R.,
Men, W., Gao, J. H., & Bi, Y. (2018). Organizational principles of
abstract words in the human brain. Cerebral Cortex, 28(12),
4305–4318. https://doi.org/10.1093/cercor/ bhx283, PubMed:
29186345
Xu, Y., Wang, X., Wang, X., Men, W., Gao, J.-H., & Bi, Y. (2018).
Doctor, teacher, and stethoscope: Neural representation of dif-
ferent types of semantic relations. The Journal of Neuroscience,
38(13), 3303–3317. https://doi.org/10.1523/ JNEUROSCI.2562
-17.2018, PubMed: 29476016
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
3
1
1
1
9
8
6
8
4
4
n
o
_
a
_
0
0
0
4
5
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neurobiology of Language
17