RESEARCH ARTICLE
On the Connection Between Language Control
and Executive Control—An ERP Study
Mathieu Declerck1,2,3
, Gabriela Meade4
, Katherine J. Midgley5, Phillip J. Holcomb5
Ardi Roelofs2, and Karen Emmorey1
1School of Speech, Language, and Hearing Sciences, San Diego State University, San Diego, USA
2Donders Institute for Brain, Cognition and Behaviour, Radboud University, Nijmegen, Netherlands
3Department of Linguistics and Literary Studies, Vrije Universiteit Brussel, Brussels, Belgium
4Joint Doctoral Program in Language and Communicative Disorders, San Diego State University &
University of California, San Diego, USA
5Department of Psychology, San Diego State University, San Diego, USA
Keywords: language control, executive control, language switching, task switching, ERPs
ABSTRACT
Models vary in the extent to which language control processes are domain general. Those that
posit that language control is at least partially domain general insist on an overlap between
language control and executive control at the goal level. To further probe whether or not language
control is domain general, we conducted the first event-related potential (ERP) study that directly
compares language-switch costs, as an index of language control, and task-switch costs, as an
index of executive control. The language switching and task switching methodologies were
identical, except that the former required switching between languages (English or Spanish)
whereas the latter required switching between tasks (color naming or category naming). This
design allowed us to directly compare control processes at the goal level (cue-locked ERPs) and
at the task performance level (picture-locked ERPs). We found no significant differences in the
switch-related cue-locked and picture-locked ERP patterns across the language and task switching
paradigms. These results support models of domain-general language control.
INTRODUCTION
A contested claim in the bilingual literature is that language control, which involves reducing
cross-language interference and increasing the chances of selecting a word from the target
language, is part of a more general executive control process (e.g., de Bruin et al., 2014; Green,
1998; Weissberger et al., 2015; however, see Calabria et al., 2015; Declerck et al., 2015; Jylkkä
et al., 2018). The absence of conclusive evidence for domain-general language control could be
due to prior research typically not focusing on the processing level at which language control and
executive control are assumed to overlap, namely the goal level (e.g., Green, 1998; Roelofs, 2003;
Thomas & Allport, 2000). In the present study, we investigated the claim that language control is
domain general by using event-related potentials (ERPs) to examine whether or not there is overlap
between language control and executive control during goal activation and selection, and/or
during later stimulus processing.
The literature on whether or not language control is domain general is quite divided. For instance,
Allport and colleagues (Meuter & Allport, 1999; Thomas & Allport, 2000) proposed that language
control occurs entirely between language task schemas. Task schemas are domain-general mental
a n o p e n a c c e s s
j o u r n a l
Citation: Declerck, M., Meade, G.,
Midgley, K. J., Holcomb, P. J., Roelofs,
A., & Emmorey, K. (2021). On the
connection between language control
and executive control—An ERP study.
Neurobiology of Language, 2(4),
628–646. https://doi.org/10.1162
/nol_a_00032
DOI:
https://doi.org/10.1162/nol_a_00032
Supporting Information:
https://doi.org/10.1162/nol_a_00032
Received: 7 October 2020
Accepted: 25 January 2021
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Author:
Mathieu Declerck
mathieu.declerck@vub.be
Handling Editor:
Anthony Steven Dick
Copyright: © 2021 Massachusetts
Institute of Technology. Published
under a Creative Commons Attribution
4.0 International (CC BY 4.0) license.
The MIT Press
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
4
6
2
8
1
9
7
9
7
1
5
n
o
_
a
_
0
0
0
3
2
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Language versus task switching
Switch costs:
The difference between switch and
repetition trials.
Posterior switch positivity:
Switch trials elicit a larger positivity
across posterior electrodes than
repetition trials.
processes used to achieve a specific goal (e.g., produce in a specific language or perform a specific
task). Put differently, these authors assumed that language control occurs entirely at the goal level,
and that it is part of executive control. Others have proposed that this task schema mechanism is
complemented by a language-specific control process that occurs at the lemma level between
translation-equivalent representations (e.g., Green, 1998). Still others have assumed that language
control occurs entirely at the lemma level rather than at the goal level (Declerck et al., 2015;
Grainger et al., 2010). More specifically, Declerck et al. (2015) proposed that language control
occurs between language nodes, which are the mental representations of each language at the
lemma level, and translation-equivalent representations at the lemma level. Thus, the mechanisms
that underlie language control and the extent to which they are domain general versus language
specific continue to be debated.
To examine the possibility of domain-general language control, most studies have compared
performance in language switching, to examine language control processes, and task switching, to
examine executive control processes (for other methodologies used to examine domain-general
language control see, e.g., Declerck et al., 2019; Jylkkä et al., 2018; Linck et al., 2020; Struys et al.,
2019). In a language switching paradigm, bilinguals typically name digits or pictures based on a
visually presented language cue (e.g., frames in a different color for each language). This setup
requires bilinguals to either switch from one language to another across trials (switch trials) or
repeat the same language (repetition trials). The cost in response time and/or accuracy of switching
languages—relative to staying in the same language—has been used as an index of language
control (e.g., Declerck & Philipp, 2015; Green, 1998). Task-switch costs are a measure of execu-
tive control that are obtained using a similar setup as the language-switch costs (e.g., Kiesel et al.,
2010). However, instead of switching between different languages, participants switch between
distinct tasks (e.g., switching between parity and magnitude judgments with number stimuli).
Even though some studies comparing language- and task-switch costs have shown a similar
pattern across the two paradigms (e.g., Declerck et al., 2017; Weissberger et al., 2015), others have
not (e.g., Branzi et al., 2016; Calabria et al., 2015). It could be that language control is only partly
domain general, as assumed by the inhibitory control model (Green, 1998). If so, some studies may
have failed to provide evidence for domain-general language control because they did not
sufficiently capture the main locus of the overlap between language and executive control. As
discussed, this overlap is assumed to occur at the goal level (e.g., Green, 1998; Roelofs, 2003;
Thomas & Allport, 2000). Several studies allow for insight into whether there is an overlap between
language and executive control at the goal level by examining the cue-to-stimulus interval in
language and task switching. During this interval, participants activate and, given sufficient time,
select the goal to speak a specific language during language switching or the goal to perform a
specific task during task switching. This makes the cue-to-stimulus interval particularly interesting
to examine possible links between language and executive control.
Two ERP language-switching studies that examined cue processing are relevant here. Lavric
et al. (2019) asked German-English bilinguals to perform a picture naming task in mixed language
blocks. The cue-to-stimulus interval was either 100 or 1,500 ms. When focusing on the cue-locked
ERPs with a cue-to-stimulus interval of 1,500 ms, they observed a posterior switch positivity (time
window: 300–700 ms). That is, cues in switch trials elicited a larger positivity across posterior
electrodes than cues in repetition trials. The posterior switch positivity is interpreted as an index
of task rule activation and is often observed in the task switching literature (e.g., Barceló & Cooper,
2018; Chevalier et al., 2015; Manzi et al., 2011; for reviews, see De Baene & Brass, 2014;
Karayanidis et al., 2010). This finding led Lavric et al. (2019) to conclude that there is an overlap
between control processes implemented on task schemas, and thus at the goal level, during
bilingual language production and non-linguistic processing.
Neurobiology of Language
629
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
4
6
2
8
1
9
7
9
7
1
5
n
o
_
a
_
0
0
0
3
2
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Language versus task switching
Anterior switch negativity:
Switch trials elicit a larger negativity
across anterior electrodes than
repetition trials.
However, this finding of Lavric et al. (2019) was not supported by the ERP language-switching
study of Verhoef et al. (2010). With a cue-to-stimulus interval of 750 ms, Verhoef and colleagues
found an anterior switch negativity around the same time window as Lavric et al. observed a pos-
terior switch positivity. Cues in switch trials elicited a larger negativity across anterior electrodes
than cues in repetition trials. Lavric et al. also observed an anterior negativity that occurred after
their posterior positivity and was relatively more protracted in time. These differences in timing
led to different accounts of the anterior switch negativity: Verhoef et al. proposed that the earlier
anterior switch negativity was an index of goal engagement, whereas Lavric et al. assumed that it
was an index of goal maintenance.
Yet the findings of Lavric et al. (2019) and Verhoef et al. (2010) might not be as different as they
initially appear. In an earlier task switching study, Lavric et al. (2008) found a dipolar ERP compo-
nent related to cue processing, with an anterior switch negativity and a posterior switch positivity
that occurred in parallel and were interdependent. Hence, the posterior switch positivity observed
by Lavric et al. (2019) and the anterior switch negativity observed by Verhoef et al. (2010) might
actually be two parts of the same dipolar ERP component.
Verhoef et al. (2010) also observed a negative peak in an earlier time window (200–350 ms)
with cue-locked ERPs that was larger for switch than repetition trials over posterior sites. According
to the authors, this component is an index of disengagement of the non-target language. In the cue-
locked task switching literature, there is typically an early posterior switch positivity, which can
occur as early as 200 ms but shows large temporal variability across studies (e.g., Eppinger
et al., 2007; Jamadar et al., 2010; Manzi et al., 2011). This early posterior switch positivity has been
linked to task goal activation, and can be dissociated from the later posterior switch positivity (e.g.,
Eppinger et al., 2007; Manzi et al., 2011; for reviews, see De Baene & Brass, 2014; Karayanidis
et al., 2010).
We have mainly focused thus far on cue processing since the cue-locked ERPs provide
theoretically interesting insights into domain-general language control. However, the literature
indicates that there might also be some overlapping processes across language and executive
control that occur after the target stimulus has been presented. Most prominently, a similar
stimulus-locked N2 component has been observed in both language and task switching studies,
which has generally been taken as an index of inhibitory control of the non-target language/task
and conflict monitoring (e.g., Gaál & Czigler, 2015; Finke et al., 2012; Jackson et al., 2001; Kang
et al., 2020). This component entails a larger negativity for switch trials than repetition trials and
is usually found around 200–350 ms after stimulus presentation. Unlike the cue-locked, early
posterior negativity observed in Verhoef et al. (2010), the stimulus-locked N2 is typically more
pronounced over anterior sites (however, see Zheng et al., 2020).
Stimulus-locked ERPs in the language switching paradigm are less straightforward beyond the
N2 component. Some studies have observed a late positive complex (LPC), characterized by a
larger positivity over posterior sites for switch trials relative to repetition trials around 400–650 ms
after stimulus presentation (e.g., Jackson et al., 2001; Liu et al., 2016; Martin et al., 2013), whereas
others have found a widespread late switch-related negativity (Kang et al., 2020; Peeters, 2020;
Peeters & Dijkstra, 2018). Task switching studies, on the other hand, tend to find evidence for a
P3b component around 400–600 ms after stimulus onset (e.g., Barceló, 2003; Provost et al.,
2018; Tieges et al., 2007; for a review, see Gajewski et al., 2018). This component is generally char-
acterized by a larger positivity for repetition trials than switch trials over posterior sites. In sum, across
the language and task switching literatures, there are three different post-N2 stimulus-locked ERP
patterns that temporally overlap and that might be shared by language and executive control: the
LPC, a late switch-related negativity, and the P3b component.
Neurobiology of Language
630
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
4
6
2
8
1
9
7
9
7
1
5
n
o
_
a
_
0
0
0
3
2
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Language versus task switching
As this brief review suggests, it is not yet clear whether similar control processes are imple-
mented during cue processing or subsequently during stimulus processing between language
and task switching. It is especially difficult to draw conclusions across studies from the language
and task switching literatures due to the different experimental designs and approaches to analyz-
ing and reporting results. Additionally, the relatively small number of language-switching ERP
studies, especially with regard to cue processing, makes it difficult to gauge the robustness of their
results. Hence, to further investigate the possibility of domain-general language control with ERPs,
it would be beneficial to directly compare language and task switching patterns within the same
participants and using the same experimental design (e.g., cue-to-stimulus interval, stimuli,
responses). In the current study, we did exactly that, by conducting the first ERP study that directly
compares control processes in nearly identical language and task switching paradigms. More
specifically, in the language-switching blocks, English-Spanish bilingual participants named
either the color (block X) or category (block Y) of pictures in a mixed language context (English
vs. Spanish). In the task-switching blocks, the same bilinguals used either English (block X) or
Spanish (block Y) in a mixed task context (color vs. category naming). So the only difference
between the language and task switching paradigms was that bilingual participants switched
between languages in the former and between tasks in the latter (cf. Declerck et al., 2017). By
having such similar language and task switching paradigms, we were able to directly compare
the temporal neural dynamics during language and task switching for the first time.
Since the language and task switching paradigms are methodologically identical, differences
in the ERP pattern between language- and task-switch costs could be taken as evidence of
different underlying mechanisms for language control and executive control. For instance, if
different control processes are implemented during goal processing, then differences in the
ERP pattern should be observed during cue processing. According to the limited literature, a
larger early negativity in the cue-locked ERPs might be expected during language switching
(Verhoef et al., 2010), whereas a larger early posterior positivity might be expected during task
switching (e.g., Eppinger et al., 2007; Jamadar et al., 2010; Manzi et al., 2011). Moreover, in the
later time window of the cue-locked ERPs, it could be that switching languages results in a larger
anterior negativity (Verhoef et al., 2010), whereas switching tasks results in a larger posterior
positivity (for reviews, see De Baene & Brass, 2014; Karayanidis et al., 2010) or a dipolar com-
ponent (Lavric et al., 2008). Finally, if different control mechanisms are involved for language
and task switching we might also expect a difference in a late time window of the picture-locked
ERPs. Task switching studies tend to observe a modulation of the P3b component (for a review,
see Gajewski et al., 2018), whereas the language switching literature has shown a switch-related
modulation of the LPC (e.g., Jackson et al., 2001; Liu et al., 2016; Martin et al., 2013) or a late
switch-related negativity (Kang et al., 2020; Peeters, 2020; Peeters & Dijkstra, 2018).
METHOD
Participants
Twenty-six English-Spanish bilinguals took part in the experiment. Two participants were excluded
due to experimenter error. The remaining 24 participants consisted of 22 women who were
23.2 years old on average (SD = 3.0 years). All participants were right-handed, no older than
30 years of age, and no one had a prior history of neurological dysfunctions. Prior to the experi-
ment the participants were asked to complete a language history questionnaire and an English and
Spanish vocabulary test through a lexical decision task (LexTALE; Izura et al., 2014; Lemhöfer &
Broersma, 2012; see Table 1). All participants were volunteers who were paid for their time.
Neurobiology of Language
631
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
4
6
2
8
1
9
7
9
7
1
5
n
o
_
a
_
0
0
0
3
2
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Language versus task switching
Table 1. Means for the demographic information (SD in parentheses) for each language
Age of acquisition (years)
Time currently used (%)
Speaking*
Writing*
Reading*
LexTALE (%)
English
4.6 (3.7)
63.2 (28.3)
6.6 (0.9)
6.5 (1.0)
6.6 (0.8)
88.7 (9.2)
Spanish
1.9 (3.8)
36.8 (28.3)
5.9 (1.2)
5.5 (1.3)
5.9 (1.2)
74.8 (14.0)
Note.* Self-rated scores on a scale of 1 (low proficiency) to 7 (high proficiency).
Informed consent was obtained in accordance with the Institutional Review Board at San Diego
State University.
Stimuli
The stimuli consisted of 48 pictures (line drawings) that were presented in one of four colors
(brown, green, blue, and orange), with an equal number of pictures in each color. Each of the
pictures depicted a concept from one of four semantic categories (furniture, clothing, food, and
animals), with 12 pictures designated to each category.
The participants were instructed to use one of two languages (English or Spanish) or perform
one of two tasks (color naming or category naming) based on one of four shape cues (square,
circle, pentagon, and parallelogram). The cues subtended a maximal visual angle of 2.7° in each
direction. The square and circle cues were always used together in the same block, as were the
pentagon and parallelogram, because these shapes were easily distinguishable from one
another. Furthermore, one set of cues was always used in the language switching paradigm,
and the other set in the task switching paradigm for a specific participant. The mapping be-
tween cues and languages/tasks was counterbalanced across participants.
Procedure
The experiment took place in a dimly lit room, where the participants were seated in a comfort-
able chair about 150 cm from the monitor. Prior to each of the four experimental blocks, each
consisting of 96 trials, there was a practice block of 20 trials. In the two language-switching
blocks, the cue determined whether participants should use English or Spanish. In one of the
blocks, the bilingual participants named the color in which the picture was presented and in
the other block they named the semantic category of the picture (See Figure 1 for an overview
of the different blocks). The order of these two blocks was counterbalanced across participants.
In the task-switching blocks, the cue determined whether participants should name the color in
which the picture was presented or its semantic category. One block was entirely in English and
the other in Spanish. The order of these two blocks was also counterbalanced across partici-
pants. Finally, the order of the switching paradigm that came first (i.e., language or task) was also
counterbalanced across participants.
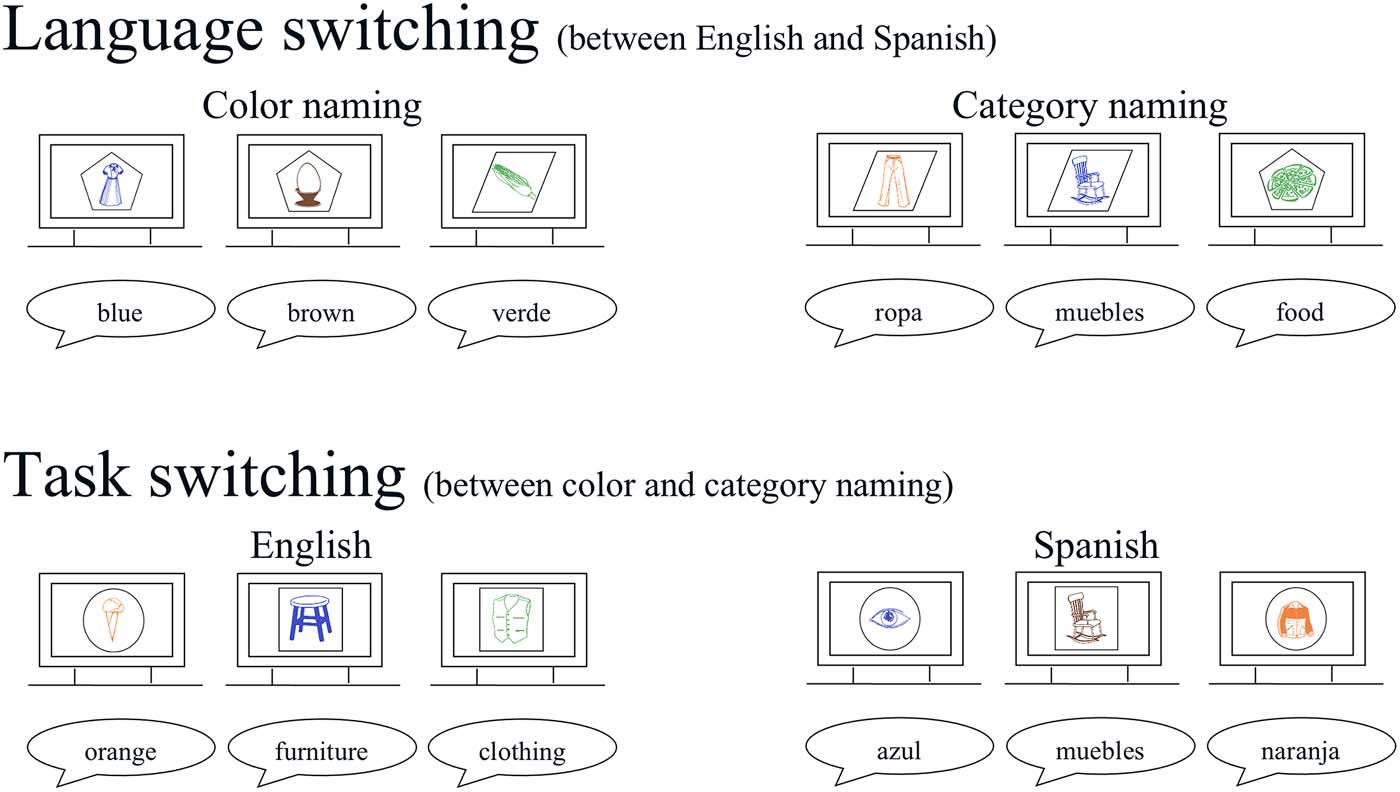
Each trial started with a central fixation cross that was presented for 500 ms. This was followed
by a blank screen for 300 ms, after which the cue shape was presented. After 800 ms of solely the
Neurobiology of Language
632
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
4
6
2
8
1
9
7
9
7
1
5
n
o
_
a
_
0
0
0
3
2
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Language versus task switching
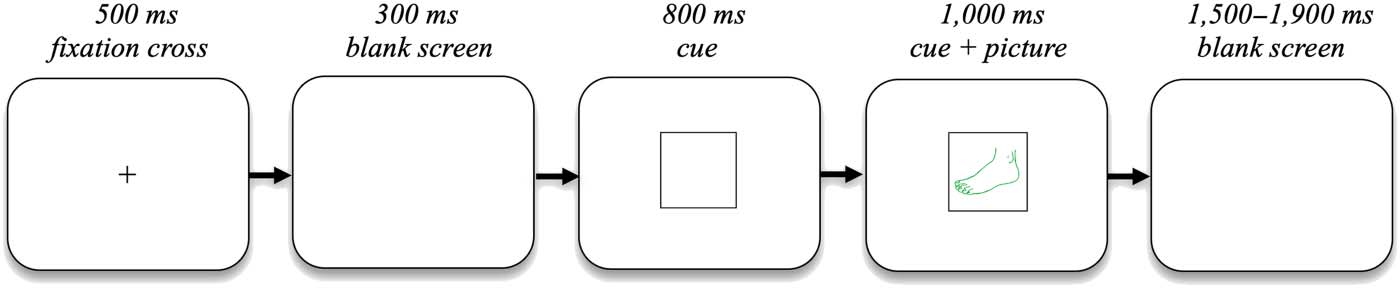
Figure 1. Overview of the different experimental conditions and blocks. In this example, the pen-
tagon was the cue for participants to respond in English and the parallelogram was the cue to re-
spond in Spanish in the language switching paradigm. The circle was the cue to name the color in
which the picture was presented, and the square was the cue to name the semantic category of the
picture in the task switching paradigm. It should also be noted that the size of the cues and pictures
relative to the screen are much larger in this figure than in the actual experiment, to make them
more visible.
cue shape being on the screen, the picture was presented inside the cue shape for 1,000 ms.
Finally, a blank screen appeared for 1,500 ms with a jitter of 0–400 ms (see Figure 2 for a visual
depiction of the trial procedure). The participants were asked to blink after they responded up
until the fixation cross.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
4
6
2
8
1
9
7
9
7
1
5
n
o
_
a
_
0
0
0
3
2
p
d
/
.
l
Behavioral Analyses
The independent variables of the behavioral analyses were Paradigm (language vs. task switching)
and Trial type (switch vs. repetition trials). The dependent variables for the behavioral analyses
were reaction times and error rates. (The data have been made publicly available at https://osf
.io/67mep/.)
For the error analysis, we only counted errors that were not preceded by other errors or omis-
sions; otherwise, it would be unclear whether to consider the error trial as a switch or repetition.
The errors consisted of incorrect use of language or incorrect target word, and were noted by a
trained research assistant who was fluent in both languages. Reaction times were calculated
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 2. Overview of the trial procedure.
Neurobiology of Language
633
Language versus task switching
from picture presentation to voice onset and were only included in the analysis if they were
between 200 and 2,500 ms long. The first trial of each block and error trials were excluded from
the RT analysis, as were trials following an error trial or an omission trial for the reason discussed
above. These criteria resulted in the exclusion of 11.01% of trials.
EEG Recording and Analyses
Participants were fitted with Electro-Caps using 29 active electrodes. Additional electrodes were
placed on each mastoid, under the left eye, and next to the outer canthus of the right eye. The
recording from the left mastoid was used as a reference, both during recording and for all
analyses. The recording from the right mastoid was used to measure differential mastoid activity
(there were no differences between conditions at the right mastoid site, so the left mastoid was
used as a reference for all subsequent ERP comparisons). The electrode below the left eye was
used to identify blinks, in combination with the activity of FP1, and the electrode next to the right
eye was used to identify any horizontal eye movements. Impedances were maintained below
2.5 kΩ. EEG was amplified using SynAmps RT amplifiers (Neuroscan-Compumedics) with a
bandpass of DC to 100 Hz and was sampled throughout at 500 Hz.
For the cue-locked ERP data, each epoch was time-locked to cue onset and was 900 ms long,
including 100 ms prior to the cue as a baseline. For the picture-locked ERP data, each epoch was
time-locked to stimulus onset and was 700 ms long, including 100 ms prior to the stimulus as a
baseline. The length of the picture-locked epoch was determined based on the shortest reaction
times to minimize speech-related artifacts. Any artifacts, including blinks or other eye move-
ments, that were detected resulted in exclusion of that specific trial from the corresponding
analyses. Additionally, the first trial of each block, error trials, and trials immediately following
these errors were excluded from analyses. These criteria resulted in the exclusion of an average
of 18.45% of trials from the cue-locked analyses and 20.19% of trials from the picture-locked
analyses. More specifically, there was an average of 75.79 (SD = 12.34) language-switch trials,
80.33 (SD = 10.80) language-repetition trials, 78.75 (SD = 9.00) task-switch trials, and 78.29
(SD = 12.21) task-repetition trials per participant in the cue-locked ERP analyses. In the picture-
locked ERP analyses, there was an average of 74.54 (SD = 10.99) language-switch trials, 77.79
(SD = 9.09) language-repetition trials, 76.79 (SD = 8.63) task-switch trials, and 77.33 (SD = 9.85)
task-repetition trials per participant.
Separate ERPs were averaged for each participant and condition at each electrode and low-
pass filtered at 15 Hz. Mean amplitudes of the early time window in the cue-locked data were
calculated for each participant between 200 and 350 ms (cf. Verhoef et al., 2010). We were also
interested to see whether a posterior switch positivity, an anterior switch negativity, or a dipolar
component would be observed in a later time window in the cue-locked data, which we mea-
sured between 350–700 ms (cf. Lavric et al., 2019). Regarding the picture-locked time windows,
we also calculated the mean amplitudes of the early time window between 200 and 350 ms, in
line with the N2 literature (e.g., Gaál & Czigler, 2015; Zheng et al., 2020). A late picture-locked
time window was calculated between 400–600 ms, which overlaps with the time windows of
the LPC, late switch-related negativity, and the P3b (e.g., Gajewski et al., 2018; Jackson et al.,
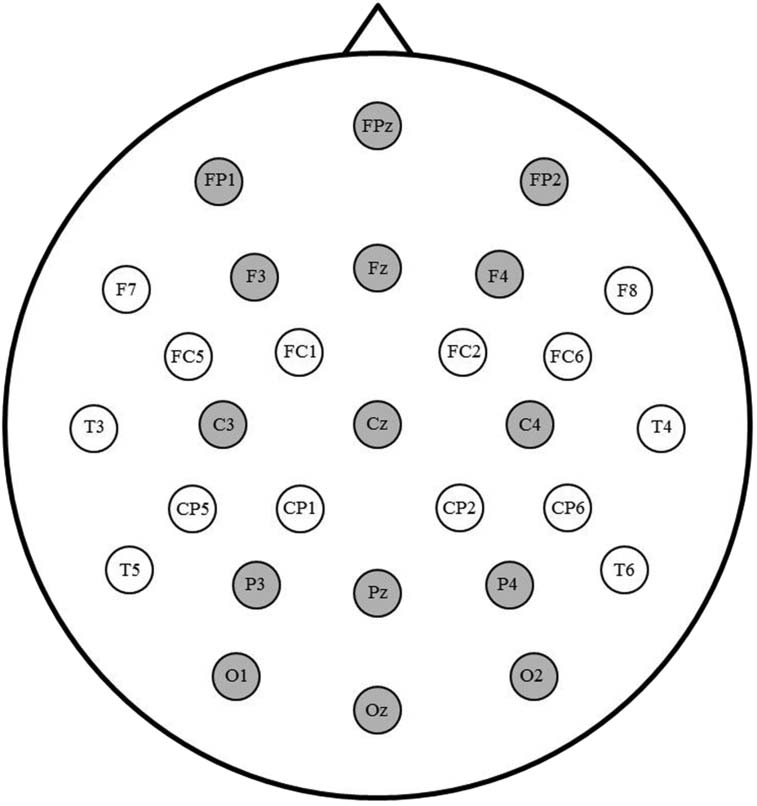
2001; Kang et al., 2020). To capture patterns at both anterior and posterior sites, we relied on a
broad grid of 15 electrodes for the analyses, as illustrated in Figure 3. The omnibus ANOVA
consisted of the following factors: Paradigm (language vs. task switching), Trial type (switch
vs. repetition trials), Laterality (left, midline, right), and Anterior/Posterior (prefrontal, frontal,
central, parietal, occipital). For all measures containing more than one degree of freedom in
the numerator, we applied Greenhouse-Geisser correction.
Neurobiology of Language
634
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
4
6
2
8
1
9
7
9
7
1
5
n
o
_
a
_
0
0
0
3
2
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Language versus task switching
Figure 3. Sites highlighted in grey were included in the analyses.
RESULTS
Behavioral Results
The reaction time analysis showed a significant main effect of Trial type, with slower responses
in switch trials (1,062 ms) than in repetition trials (1,019 ms), F(1, 23) = 33.17, p < 0.001, η
2 =
p
0.590 (see Table 2).
No significant difference was observed between the size of language- and task-switch costs
(95% CI = [−33, 36]; for further evidence along these lines obtained with an ex-Gaussian analysis
of reaction time distributions, see Supplementary Materials; supporting information can be found
online at https://www.mitpressjournals.org/doi/suppl/10.1162/nol_a_00032; the interaction
between Paradigm and Trial type was not significantly influenced by the order of the paradigms,
which was also the case for all other analyses.) To quantify this null effect, we relied on Bayesian
Null Hypothesis Testing (e.g., Aczel et al., 2018; Rouder et al., 2009; Wagenmakers et al., 2018),
which is a statistical test that shows the degree to which the Null hypothesis (H0) should be
accepted over the Alternative hypothesis (H1). To this end, we compared a model that only
includes main effects of Paradigm and Trial type against a model that includes both the main
effects and their interaction. The results confirmed that a model that only includes both main
effects accounts for the data better than a model that also includes the interaction (BF01 = 3.58;
Kass & Raftery, 1995). In other words, we have statistical evidence that the language- and task-
switch costs are about three and a half times more likely to be similar than they are to be different.
Table 2. Overall reaction times in ms and percentage of errors (SD in parentheses), as a function of Paradigm (language vs. task switching)
and Trial type (switch vs. repetition trials)
Dependent variables
Reaction time
Switch
1,068 (203)
Language switching
Repetition
1,024 (184)
Switch costs
44
Switch
1,056 (201)
Task switching
Repetition
1,014 (206)
Error rate
5.22 (3.76)
2.11 (2.28)
3.11
3.62 (2.28)
2.70 (2.15)
Neurobiology of Language
Switch costs
42
0.92
635
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
4
6
2
8
1
9
7
9
7
1
5
n
o
_
a
_
0
0
0
3
2
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Language versus task switching
The error rate analysis showed a significant main effect of Trial type, with more errors observed
in switch trials (4.39%) than in repetition trials (2.41%), F(1, 23) = 28.49, p < 0.001, η
2 = 0.553 (see
p
Table 2). Furthermore, the interaction between Paradigm and Trial type was significant, F(1, 23) =
13.98, p = 0.001, η
2 = 0.378, with larger language-switch costs (3.11%) than task-switch costs
p
(0.92%; 95% CI = [1.18, 3.56]).
Cue-Locked ERP Results
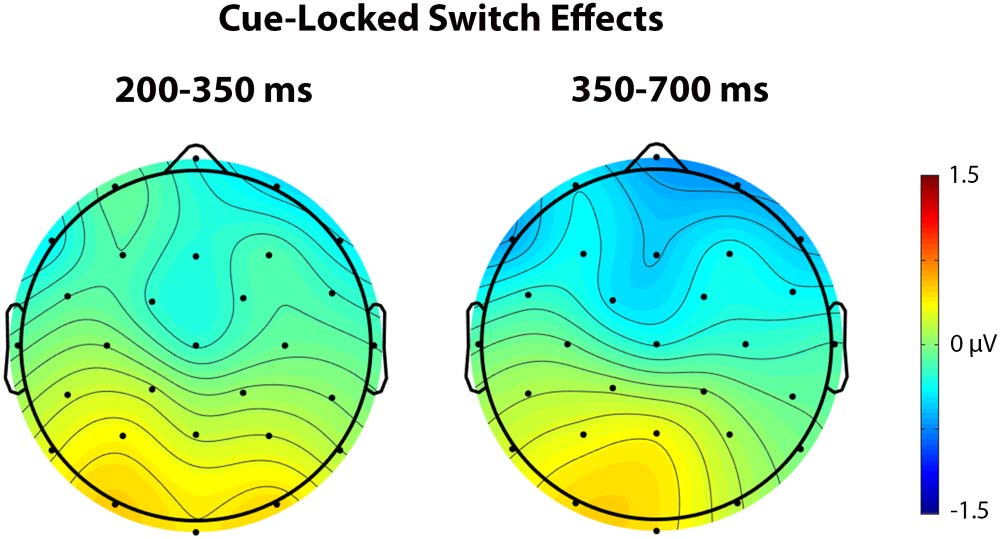
In the early time window (200–350 ms), a significant interaction was observed between Trial
type and Anterior/Posterior, F(4, 92) = 4.76, p = 0.018, η
2 = 0.172, indicating a larger negativity
p
during cue processing in switch trials than repetition trials at anterior sites and a larger positivity
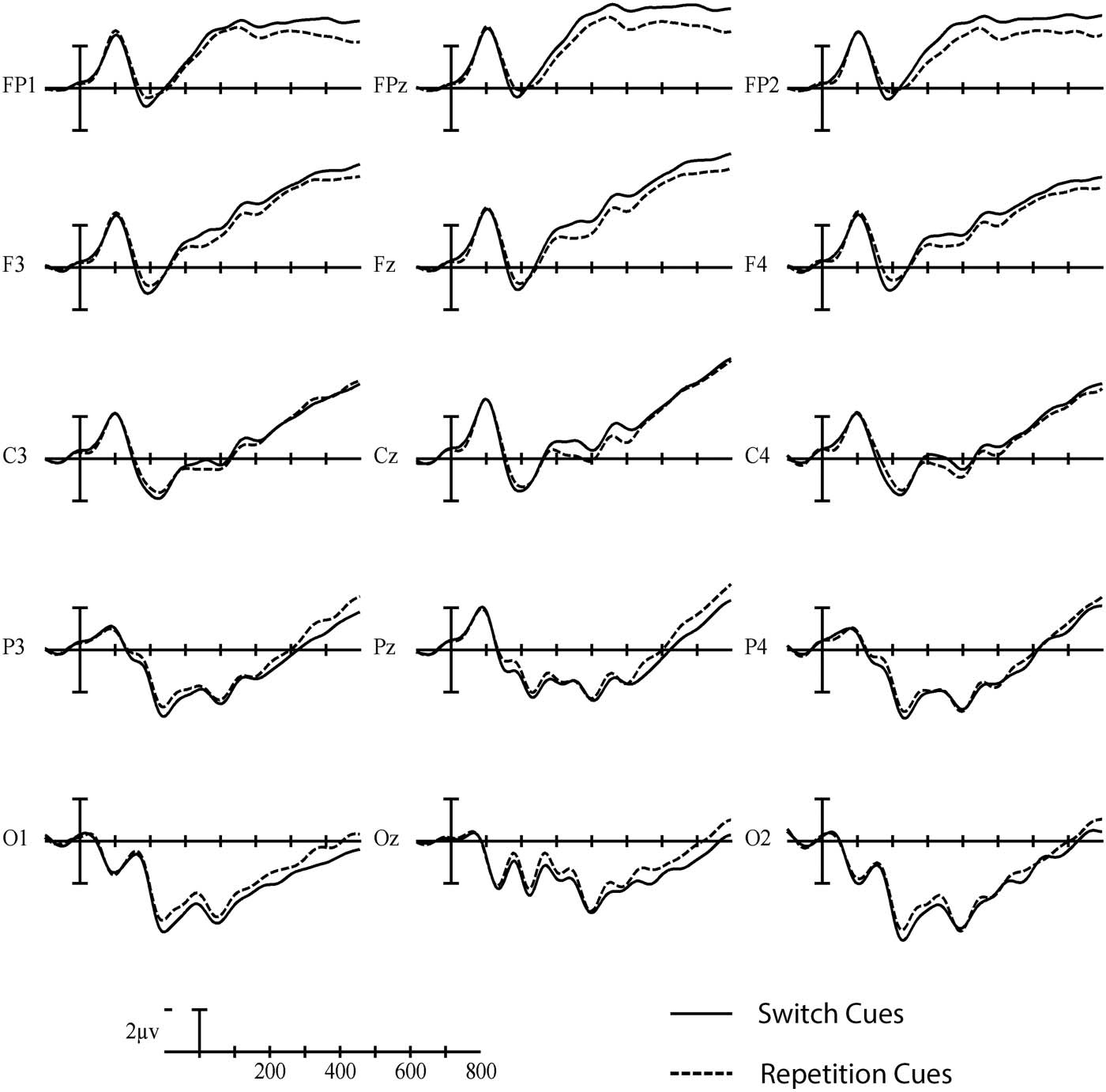
in switch than repetition trials at more posterior sites (see Figures 4 and 5).
No significant switch cost differences were observed between language and task switching
(95% CI = [−3.42, 1.08]). Using Bayesian Null Hypothesis Testing, we confirmed that a model
that does not include the interaction between Paradigm and Trial type, but does include both
main effects, accounts for the data better than a model that includes the interaction. This held
true for both Fz (BF01 = 3.38; 95% CI = [−0.83, 4.81]) and Pz (BF01 = 3.18; 95% CI = [−5.73,
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
4
6
2
8
1
9
7
9
7
1
5
n
o
_
a
_
0
0
0
3
2
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. Grand average cue-locked ERP waveforms elicited across language and task switching
with switch trials (solid line) and repetition trials (dashed line). Each vertical tick marks 100 ms, and
negative is plotted up. The calibration bar marks 2 μV.
Neurobiology of Language
636
Language versus task switching
Figure 5. These scalp voltage maps show the distribution of the cue-locked Trial type effect
(switch trials – repetition trials) in the 200–350 ms (left) and 350–700 ms (right) time windows col-
lapsed across language and task switching paradigms. Cool colors indicate a larger negativity for
switch trials relative to repetition trials.
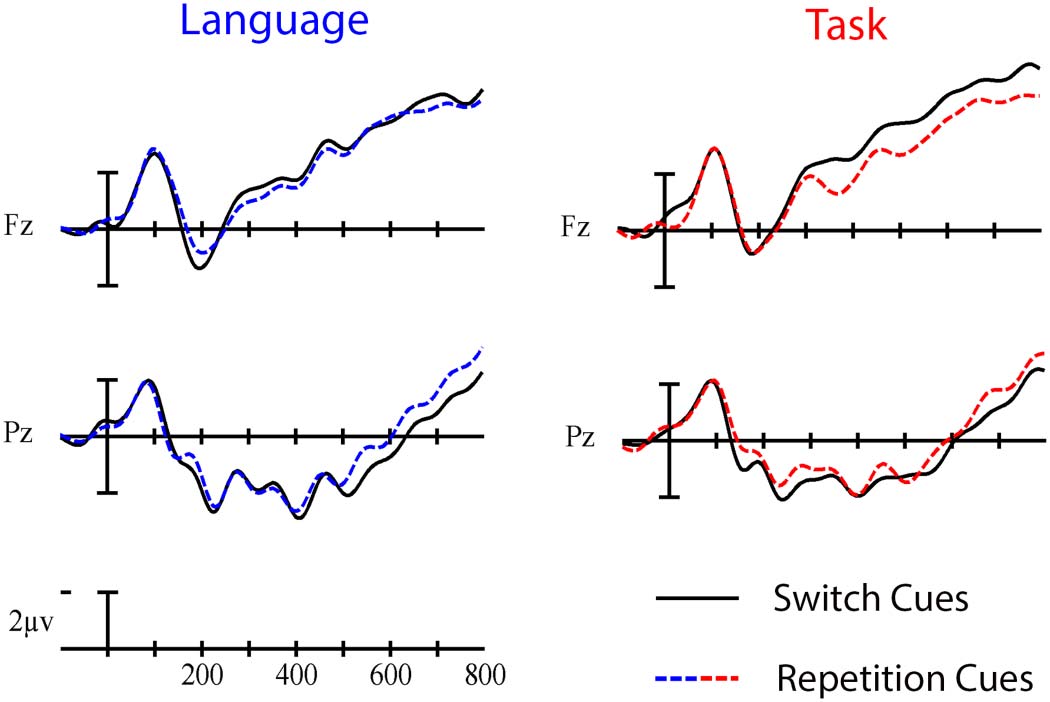
0.35]), which were chosen as representative anterior and posterior electrodes where the Trial
type effects appeared largest across all cue- and picture-locked analyses (see the waveforms
and voltage maps in this article). That is, we have statistical evidence that the language- and
task-switch costs are about three times more likely to be similar than they are to be different
in this time window (for a visual depiction, see Figure 6).
In the late time window (350–700 ms), a significant main effect was found for Paradigm, F(1,
23) = 6.01, p = 0.022, η
2 = 0.207, indicating that cues elicited a larger negativity in the task
p
switching paradigm than in the language switching paradigm. There was also an interaction
between Trial type and Anterior/Posterior, F(4, 92) = 4.59, p = 0.029, η
2 = 0.167, indicating that
p
cues in switch trials elicited larger negativities than those in repetition trials over anterior sites,
but larger positivities over posterior sites.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
4
6
2
8
1
9
7
9
7
1
5
n
o
_
a
_
0
0
0
3
2
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 6. Grand average cue-locked ERP waveforms at representative electrodes Fz and Pz elic-
ited by switch trials (solid line) and repetition trials (dashed line) in the language (left) and task
switching (right) paradigms. Each vertical tick marks 100 ms, and negative is plotted up. The cal-
ibration bar marks 2 μV.
Neurobiology of Language
637
Language versus task switching
Again, no significant switch cost differences were found between language and task switching
(95% CI = [−0.31, 1.08]). Using Bayesian Null Hypothesis Testing, we confirmed that a model that
does not include the interaction between Trial type and Paradigm, but does include both main
effects, accounts for the data better than a model that includes the interaction and both main effects
for Pz (BF01 = 3.65; 95% CI = [−0.30, 1.73]). However, no definitive conclusions could be made
for Fz (BF01 = 1.95; 95% CI = [−0.65, 0.99]) based on the Bayesian Null Hypothesis Testing
(cf. Kass & Raftery, 1995). Hence, it is difficult to indicate one way or another whether there
was a substantial difference in this window with respect to Trial type between language and task
switching over anterior sites.
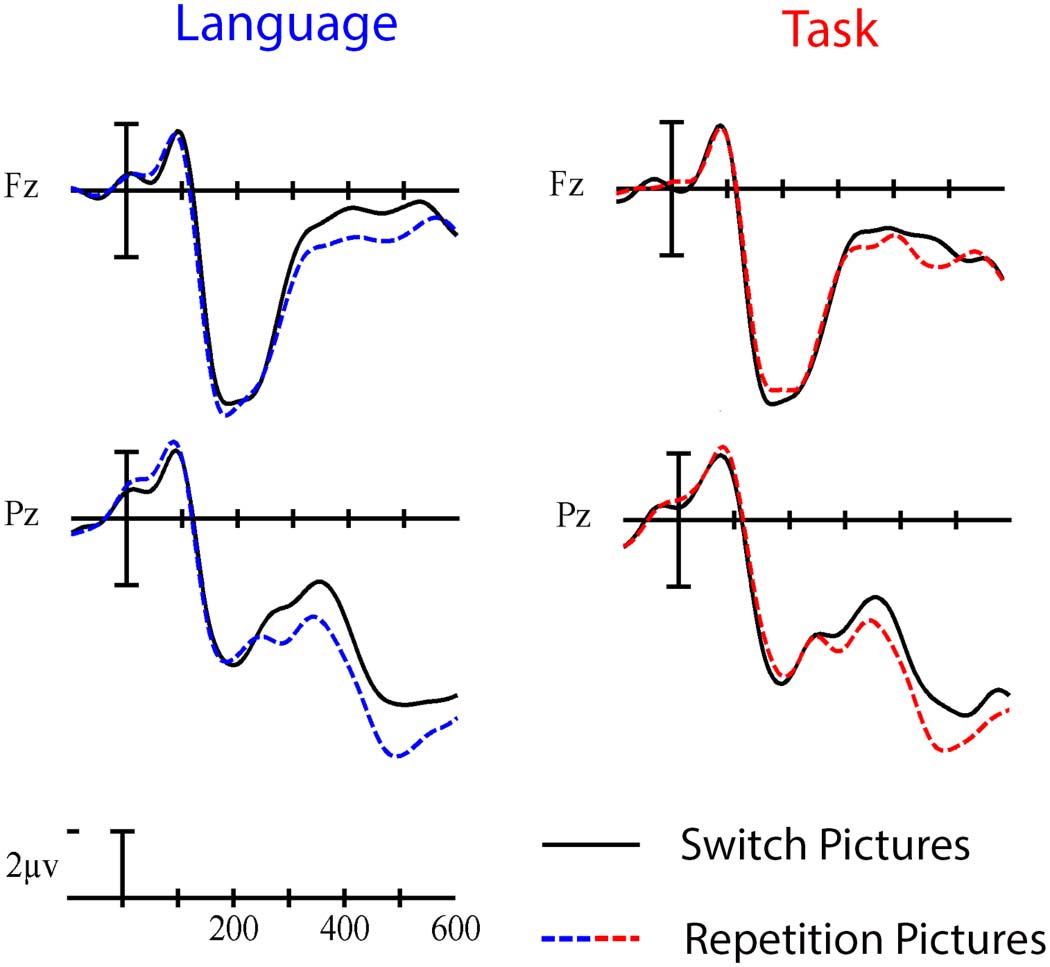
Picture-Locked ERP Results
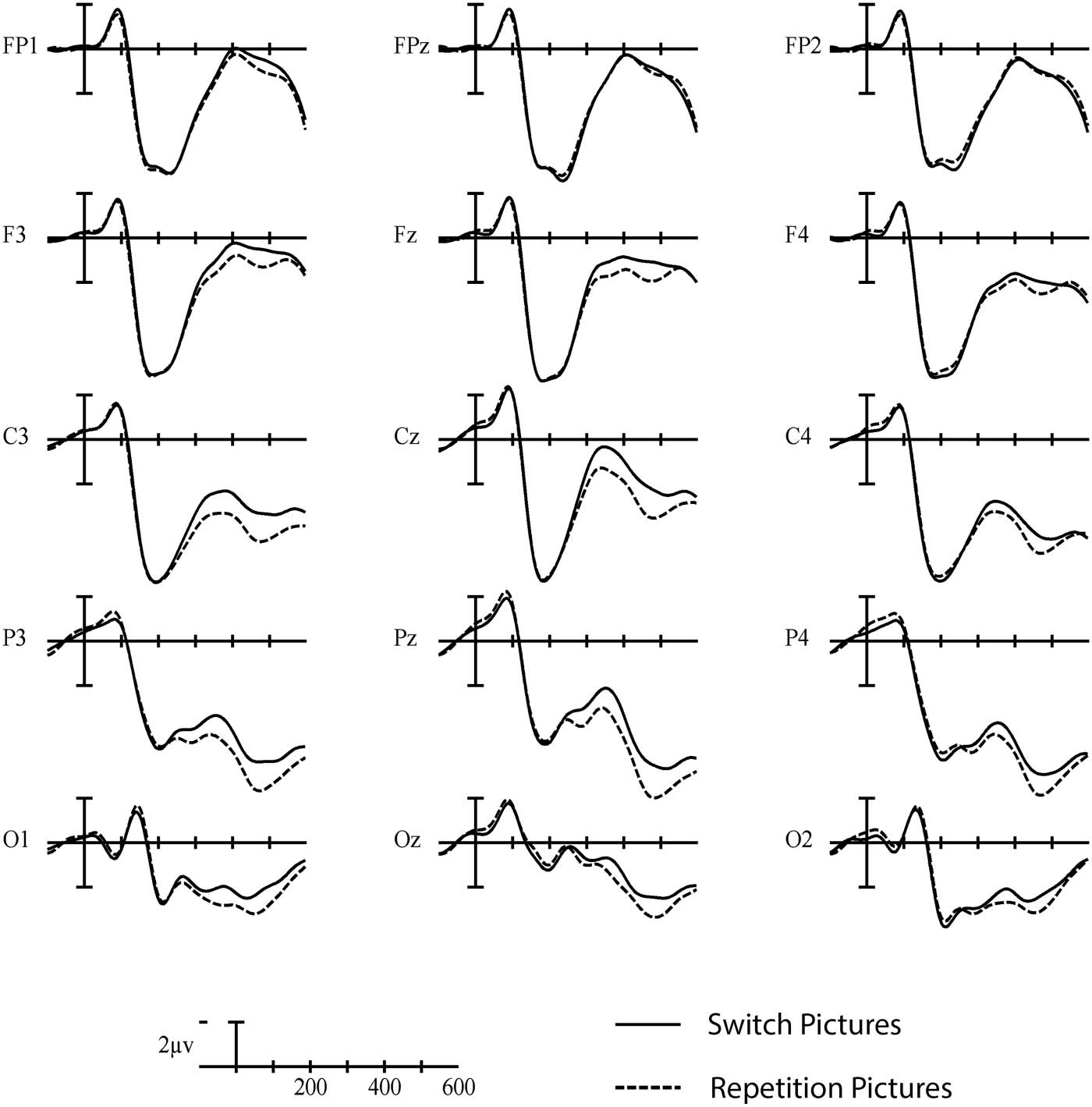
In the early time window (200–350 ms), a significant interaction was observed between Trial
type and Laterality, F(2, 46) = 6.57, p = 0.009, η
2 = 0.222, indicating a larger negativity in switch
p
compared to repetition trials, especially over the left hemisphere (see Figures 7 and 8).
Similar to the cue-locked ERPs, no significant switch cost differences were observed between
language and task switching (95% CI = [−1.01, 0.37]). Using Bayesian Null Hypothesis Testing,
we confirmed that a model that does not include the interaction between Paradigm and Trial
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
4
6
2
8
1
9
7
9
7
1
5
n
o
_
a
_
0
0
0
3
2
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 7. Grand average picture-locked ERP waveforms elicited across language and task switch-
ing with switch trials (solid line) and repetition trials (dashed line). Each vertical tick marks 100 ms
and negative is plotted up. The calibration bar marks 2 μV.
Neurobiology of Language
638
Language versus task switching
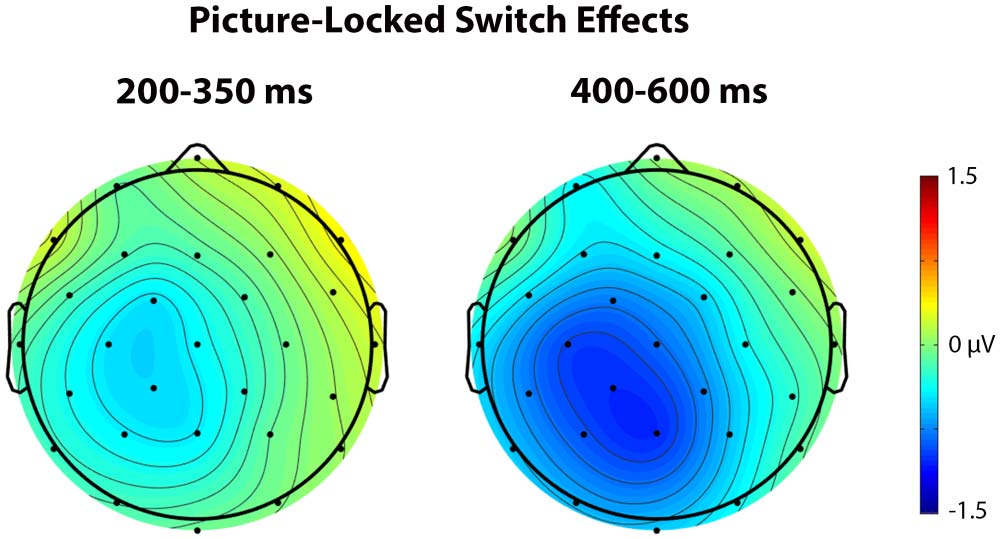
Figure 8. These scalp voltage maps show the distribution of the picture-locked Trial type effect
(switch trials – repetition trials) in the 200–350 ms (left) and 400–600 ms (right) time windows col-
lapsed across language and task switching paradigms. Cool colors indicate a larger negativity for
switch trials relative to repetition trials.
type, but does include both main effects, accounts for the data better than a model that includes
the interaction at both Fz (BF01 = 3.19; 95% CI = [−1.31, 0.57]) and Pz (BF01 = 3.24; 95% CI =
[−1.17, 0.60]) (for a visual depiction, see Figure 9).
In the late time window (400–600 ms), a significant interaction was observed between
Paradigm and Anterior/Posterior, F(4, 92) = 5.91, p = 0.009, η
2 = 0.204, indicating a larger neg-
p
ativity when performing in the task switching paradigm compared to the language switching
paradigm, especially over anterior sites. There was also a significant interaction between Trial
type and Laterality, F(2, 46) = 4.99, p = 0.026, η
2 = 0.178, indicating a larger negativity in switch
p
trials compared to repetition trials, especially over left hemisphere sites.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
4
6
2
8
1
9
7
9
7
1
5
n
o
_
a
_
0
0
0
3
2
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 9. Grand average picture-locked ERP waveforms at representative electrodes Fz and Pz
elicited by switch trials (solid line) and repetition trials (dashed line) in the language (left) and task
switching (right) paradigms. Each vertical tick marks 100 ms, and negative is plotted up. The cal-
ibration bar marks 2 μV.
Neurobiology of Language
639
Language versus task switching
This analysis also showed no significant switch cost differences between language and task
switching (95% CI = [−1.28, 0.71]). Using Bayesian Null Hypothesis Testing, we confirmed that
a model that does not include the interaction between Paradigm and Trial type, but does include
both main effects, accounts for the data better than a model that includes the interaction and
both main effects at both Fz (BF01 = 3.32; 95% CI = [−1.68, 1.01]) and Pz (BF01 = 3.37; 95%
CI = [−1.41, 0.96]).
DISCUSSION
In the current ERP study, we examined whether or not the neurocognitive mechanisms under-
lying language control (for possible neural loci, see Kim et al., 2012; Sulpizio et al., 2020) are
domain general by comparing switch costs between carefully matched language and task
switching paradigms. We observed similar costs in reaction times for language and task
switches, but there was a difference in error rate. Most importantly, we obtained similar cue-
locked and picture-locked ERP patterns across the language and task switching paradigms. In
what follows, we first discuss the behavioral results, followed by the cue-locked and picture-
locked results. Finally, we discuss the limitations of this study.
Behavioral Data
We did not find any significant switch cost differences in reaction time between the language
switching and task switching paradigms (see also Supplementary Materials). This result was fur-
ther supported by Bayesian Null Hypothesis Testing, which provided statistical evidence to sup-
port the conclusion that language- and task-switch costs were similar. Such a finding is in line
with the hypothesis that language control is domain general. The error rate data were more dif-
ficult to interpret. Although the switch costs were larger during language switching relative to
task switching, this result must be qualified by the fact that the error rates were quite low overall
(3.3% of all trials).
If reliable, the error rate pattern could be interpreted as evidence for differences in the pro-
cesses that are engaged in language versus executive control. Another possibility is that the
quantitative difference that we observed could reflect a difference in the extent to which the
same control process is engaged across paradigms. For instance, language switching might re-
quire more control processes than task switching (e.g., more inhibition of the non-target lan-
guage than of the non-target task), which could have resulted in larger language-switch costs.
This is plausible since the mental representations of the first and second languages are used on
a more regular basis and thus should have a larger base activation than the mental represen-
tations of category naming and color naming. In turn, more interference, and thus more control
processes (cf. proportional control; Green, 1998; Meuter & Allport, 1999), should occur during
language than task switching. Hence, differences in the size of behavioral switch costs do not
necessarily indicate qualitative differences between language control and executive control.
That is one of the main reasons why we included electrophysiological recordings; they allow
us to make a more nuanced comparison between language and task switching with respect to
their time course and neural underpinnings.
Cue-Locked ERPs
In the early time window of the cue-locked ERPs, we observed a similar pattern across language
and task switching paradigms: a posterior switch positivity in parallel with an anterior switch
negativity. This pattern could partly be explained in terms of the early, posterior switch positivity
that has been observed in task switching studies (Eppinger et al., 2007; Manzi et al., 2011);
Neurobiology of Language
640
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
4
6
2
8
1
9
7
9
7
1
5
n
o
_
a
_
0
0
0
3
2
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Language versus task switching
however, this positivity is generally not accompanied by an anterior switch negativity. A more
straightforward explanation of the interaction between Trial type and Anterior/Posterior in the
early time window is that it reflects the start of the same dipolar pattern we observed in the fol-
lowing time window (350–700 ms). This explanation seems to be confirmed by the ERP wave-
forms (see Figure 4), where the same ERP pattern continues from the early time window into the
late time window. So, it might be that the early and later cue-locked time windows reflect the
same protracted underlying process.
This long-lasting dipolar pattern, with a larger negativity over anterior sites and a larger pos-
itivity over posterior sites in switch than repetition trials, is reminiscent of the pattern observed in
the task switching study of Lavric et al. (2008). This pattern could also explain the seemingly
diverging, but temporally overlapping, patterns in the language switching studies by Lavric
et al. (2019), who observed a posterior switch positivity, and Verhoef et al. (2010), who observed
an anterior switch negativity, as two parts of the same dipolar ERP pattern. Since the posterior
switch positivity and the anterior switch negativity observed in our study probably rely on the
same underlying process, it is not exactly clear whether the dipolar pattern should be explained
in terms of task rule activation (indexed by the posterior switch positivity) and/or goal mainte-
nance (indexed by the anterior switch negativity).
The fact that we observed the same dipolar pattern in our language switching data as Lavric
et al. (2008) observed in their task switching data is an indication that our pattern might represent
a domain-general process. This hypothesis was further supported by the fact that we observed
the same patterns across methodologically similar language and task switching paradigms. The
Bayesian Null Hypothesis Testing provided further evidence for a similar dipolar pattern be-
tween paradigms with the exception of Fz in the later time window, which proved inconclusive.
While this inconclusive finding leaves open the possibility that language control might not be
entirely domain general, not too much weight should be put on this finding, as we did not find a
significant switch cost difference between language and task switching in the cue-locked ERPs.
Picture-Locked ERPs
The interaction between Trial type and Laterality in the early time window of the picture-locked
ERPs (200–350 ms) could be interpreted in terms of the N2 component, which has been
observed in both language and task switching studies (e.g., Gaál & Czigler, 2015; Finke et al.,
2012; Jackson et al., 2001; Kang et al., 2020). The N2 component is characterized by a short
negative peak around 200–350 ms that is larger for switch than repetition trials, and most pro-
nounced over anterior sites. However, the early switch-related negativity observed in our study
does not appear to correspond to the typical N2 (see Figure 7). For example, there is no short
peak in the picture-locked waveforms that resembles an N2 component, and the difference
between switch and repetition trials seems to occur broadly across the scalp rather than being
constrained to anterior sites. Not observing a typical switch-related N2 over anterior sites is not
entirely surprising, since this component does not seem robust across studies (e.g., Christoffels
et al., 2007; Martin et al., 2013; Peeters, 2020; Peeters & Dijkstra, 2018; Timmer et al., 2019;
Verhoef et al., 2009). Instead, it looks like the pattern in the early time window of the picture-
locked data may reflect the beginning of the pattern that we see in the late time window.
In the late time window of the picture-locked ERPs, we also observed a switch-related neg-
ativity, which is in line with the findings of Kang et al. (2020), Peeters (2020), and Peeters and
Dijkstra (2018). According to Kang and colleagues, this pattern represents an N400-like compo-
nent such that word meaning retrieval is more difficult during switch trials. This explanation
makes sense applied to our study, since, similar to the N400 component, the pattern we
Neurobiology of Language
641
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
4
6
2
8
1
9
7
9
7
1
5
n
o
_
a
_
0
0
0
3
2
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Language versus task switching
observed was a large negativity with a broad scalp distribution. According to Peeters (2020; see
also Peeters & Dijkstra, 2018), this late, switch-related negativity reflects a domain-general pro-
cess, because the same pattern has been observed in task switching studies (for a review, see
Karayanidis & Jamadar, 2014). Evidence along these lines is also substantiated in our study, since
Bayesian Null Hypothesis Testing on the late time window of the picture-locked ERPs indicated
that it was more probable that language- and task-switch costs were similar than that they were
different.
Because most models assume that language and executive control overlap at the goal level, it
is not entirely clear why we should have observed a similar switch cost pattern in the picture-
locked ERPs during language and task switching. One possibility is that some goal level process-
ing might continue after picture presentation, and/or control processes related to the goal level
might spill over into processing levels that are implemented after goal processing. This would be
in line with models that assume an overlap between language and executive control at the goal
level (e.g., Green, 1998). While these claims might be true, it seems unlikely that these control
processes would affect the late time window, where we still observed the same neural pattern for
language and task switching. Another explanation might be that control processes occur at the
motor level (i.e., activation of the speech articulators), which is also shared across domains.
Several language and task switching studies have provided evidence for control processes at
the motor level (e.g., Philipp et al., 2007; Philipp & Koch, 2016; Reverberi et al., 2015).
However, this motor explanation cannot account for the earlier similarities across paradigms.
Ultimately, a combination of the explanations given here might contribute to the overall similar
language- and task-switch cost pattern observed in the picture-locked ERPs.
Limitations
In order to closely match the setup for the language and task switching paradigms, we had to use
linguistic tasks (i.e., naming a color or semantic category) in the task switching paradigm. This
allowed us to directly compare the results obtained in the language and task switching para-
digms, since the only difference was whether participants switched between languages or tasks,
respectively. However, this design means that it is possible that control processes specific to
lexical retrieval might also have been involved in the task switching paradigm. It should be noted
that the possible language-specific control processes implemented during our task switching
paradigm would be control processes implemented during single language processing, as par-
ticipants were always using the same language in the task switching blocks. Previous studies
investigating the control processes during single and mixed language processing have shown
that they can be different (Abutalebi et al., 2008; Declerck et al., 2017; Declerck et al.,
2020). So, comparing within- and between-language switch costs does not necessarily mean
that the same control processes are used. Future research may indicate whether similar ERP pat-
terns can be observed across language and task switching, in which the latter is completely void
of any linguistic processes. The downside of such a study would be that the language- and task-
switch costs would not be directly comparable due to differences other than the type of switch-
ing (e.g., different stimuli, responses, response modality, number of response alternatives, etc.).
Another possible limitation of the current study relates to power. We observed no significant
differences between language- and task-switch costs in the ERPs. Yet, it is difficult to draw strong
conclusions on the basis of null results. It might be that our study was underpowered to the point
that we could not observe any switch-cost differences between language and task switching. We
tried to address this issue by including Bayesian Null Hypothesis Testing, which allowed us
to determine to what degree the Null hypothesis is preferred over the Alternative hypothesis
Neurobiology of Language
642
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
4
6
2
8
1
9
7
9
7
1
5
n
o
_
a
_
0
0
0
3
2
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Language versus task switching
(e.g., Aczel et al., 2018; Rouder et al., 2009; Wagenmakers et al., 2018). All but one of the
Bayesian factors indicated that the Null hypothesis is at least three times more likely to explain
our data compared to the Alternative hypothesis. So, we suggest that our study can provide
meaningful insights into the claim of domain-general language control.
Finally, as with most studies that investigate bilinguals, it should be noted that the outcome of
our study could be different for other types of bilinguals. As can be seen in Table 1, the current
study relied on early bilinguals with a relatively high proficiency in both languages. Although
our results could be different for late bilinguals and/or those with lower language proficiency, no
research has provided evidence along these lines. Hence, it remains unclear whether language
history and/or language proficiency has any impact on the connection between language
control and executive control.
Conclusions
In this first ERP study to directly compare language and task switching with everything else held
constant, we observed evidence for domain-general language control. Such evidence was
observed in both the cue-locked and picture-locked ERPs, since no significant differences were
observed between the language- and task-switch costs. So, our study indicates that language
control is, to some degree at least, part of the more general executive control process. These
findings seem to best align with models that propose that language control relies (at least partly)
on domain-general processes (e.g., Green, 1998; Roelofs, 2003; Thomas & Allport, 2000).
An overlap between language control and executive control has many ramifications. For
instance, it allows for the idea that better language control through constant practice should also
influence executive control (cf. bilingual advantage; e.g., Prior & MacWhinney, 2010; however,
see, among others, Nichols et al., 2020). Beyond bilingualism, our findings also indicate that
language production encompasses processes that are not specific to language (e.g., Nozari &
Novick, 2017; Roelofs, 2003, 2021).
ACKNOWLEDGMENTS
This project has received funding from the European Union’s Horizon 2020 research and inno-
vation programme under the Marie Skłodowska-Curie grant agreement No. 840286.
FUNDING INFORMATION
Mathieu Declerck, Horizon 2020 Framework Programme (http://dx.doi.org/10.13039
/100010661), Award ID: 840286.
AUTHOR CONTRIBUTIONS
Mathieu Declerck: Conceptualization: Lead; Data curation: Lead; Formal analysis: Equal; Funding
acquisition: Lead; Investigation: Lead; Methodology: Lead; Project administration: Equal;
Visualization: Supporting; Writing – original draft: Lead. Gabriela Meade: Conceptualization:
Supporting; Formal analysis: Equal; Investigation: Supporting; Methodology: Supporting;
Visualization: Lead; Writing – review & editing: Equal. Katherine J. Midgley: Conceptualization:
Supporting; Project administration: Equal; Resources: Equal; Software: Equal; Supervision:
Supporting; Visualization: Supporting; Writing – review & editing: Equal. Phillip J. Holcomb:
Conceptualization: Supporting; Formal analysis: Supporting; Methodology: Supporting; Project
administration: Equal; Resources: Equal; Software: Equal; Supervision: Equal; Writing – review &
Neurobiology of Language
643
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
4
6
2
8
1
9
7
9
7
1
5
n
o
_
a
_
0
0
0
3
2
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Language versus task switching
editing: Equal. Ardi Roelofs: Conceptualization: Supporting; Funding acquisition: Supporting;
Investigation: Supporting; Methodology: Supporting; Supervision: Supporting; Writing – review
& editing: Equal. Karen Emmorey: Conceptualization: Supporting; Funding acquisition:
Supporting; Investigation: Supporting; Methodology: Supporting; Project administration:
Supporting; Resources: Supporting; Supervision: Lead; Writing – review & editing: Equal.
REFERENCES
Abutalebi, J., Annoni, J. M., Zimine, I., Pegna, A. J., Seghier, M. L.,
Lee-Jahnke, H., Lazeyras, F., Cappa, S. F., & Khateb, A. (2008).
Language control and lexical competition in bilinguals: An
event-related fMRI study. Cerebral Cortex, 18, 1496–1505.
DOI: https://doi.org/10.1093/cercor/bhm182, PMID: 17947346
Aczel, B., Palfi, B., Szollosi, A., Kovacs, M., Szaszi, B., Szecsi, P.,
Zrubka, M., Gronau, Q. F., van den Bergh, D., & Wagenmakers,
E.-J. (2018). Quantifying support for the null hypothesis in psy-
chology: An empirical investigation. Advances in Methods and
Practices in Psychological Science, 1, 357–366. DOI: https://
doi.org/10.1177/2515245918773742
Branzi, F. M., Calabria, M., Boscarino, M. L., & Costa, A. (2016).
On the overlap between bilingual language control and domain-
general executive control. Acta Psychologica, 166, 21–30. DOI:
https://doi.org/10.1016/j.actpsy.2016.03.001, PMID: 27043252
Barceló, F. (2003). The Madrid card sorting test (MCST): A task
switching paradigm to study executive attention with event-
related potentials. Brain Research Protocols, 11, 27–37. DOI:
https://doi.org/10.1016/S1385-299X(03)00013-8, PMID:
12697260
Barceló, F., & Cooper, P. S. (2018). An information theory account
of late frontoparietal ERP positivities in cognitive control.
Psychophysiology, 55, e12814. DOI: https://doi.org/10.1111/psyp
.12814, PMID: 28295342
Calabria, M., Branzi, F. M., Marne, P., Hernandez, M., & Costa, A.
(2015). Age-related effects over bilingual language control and
executive control. Bilingualism: Language and Cognition, 18,
65–78. DOI: https://doi.org/10.1017/S1366728913000138
Chevalier, N., Martis, S. B., Curran, T., & Munakata, Y. (2015).
Metacognitive processes in executive control development:
The case of reactive and proactive control. Journal of Cognitive
Neuroscience, 27, 1125–1136. DOI: https://doi.org/10.1162/jocn
_a_00782, PMID: 25603026, PMCID: PMC4510990
Christoffels, I. K., Firk, C., & Schiller, N. O. (2007). Bilingual language
control: An event-related brain potential study. Brain Research,
1147, 192–208. DOI: https://doi.org/10.1016/j.brainres
.2007.01.137, PMID: 17391649
De Baene, W., & Brass, M. (2014). Dissociating strategy-dependent and
independent components in task preparation. Neuropsychologia,
62, 331–340. DOI: https://doi.org/10.1016/j.neuropsychologia
.2014.04.015, PMID: 24791708
de Bruin, A., Roelofs, A., Dijkstra, T., & Fitzpatrick, I. (2014). Domain-
general inhibition areas of the brain are involved in language
switching: FMRI evidence from trilingual speakers. NeuroImage,
90, 348–359. DOI: https://doi.org/10.1016/j.neuroimage
.2013.12.049, PMID: 24384153
Declerck, M., Eben, C., & Grainger, J. (2019). A different perspec-
tive on domain-general language control using the flanker task.
Acta Psychologica, 198, 102884. DOI: https://doi.org/10.1016/j
.actpsy.2019.102884, PMID: 31306968, PMCID: PMC6718773
Declerck, M., Grainger, J., Koch, I., & Philipp, A. M. (2017). Is lan-
guage control just a form of executive control? Evidence for over-
lapping processes in language switching and task switching.
Journal of Memory and Language, 95, 138–145. DOI: https://
doi.org/10.1016/j.jml.2017.03.005
Declerck, M., Ivanova, I., Grainger, J., & Duñabeitia, J. A. (2020).
Are similar control processes implemented during single and
dual language production? Evidence from switching between
speech registers and languages. Bilingualism: Language and
Cognition, 23, 694–701. DOI: https://doi.org/10.1017
/S1366728919000695
Declerck, M., Koch, I., & Philipp, A. M. (2015). The minimum
requirements of language control: Evidence from sequential pre-
dictability effects in language switching. Journal of Experimental
Psychology: Learning, Memory, and Cognition, 41, 377–394.
DOI: https://doi.org/10.1037/xlm0000021, PMID: 24999708
Declerck, M., & Philipp, A. M. (2015). A review of control processes
and their locus in language switching. Psychonomic Bulletin &
Review, 22, 1630–1645. DOI: https://doi.org/10.3758/s13423
-015-0836-1, PMID: 25917142
Eppinger, B., Kray, J., Mecklinger, A., & John, O. (2007). Age dif-
ferences in task switching and response monitoring: Evidence
from ERPs. Biological Psychology, 75, 52–67. DOI: https://doi
.org/10.1016/j.biopsycho.2006.12.001, PMID: 17250949
Finke, M., Escera, C., & Barceló, F. (2012). The effects of fore-
knowledge and task-set shifting as mirrored in cue-and target-
locked event-related potentials. PLoS One, 7, e49486. DOI:
https://doi.org/10.1371/journal.pone.0049486, PMID:
23152912, PMCID: PMC3495879
Gaál, Z. A., & Czigler, I. (2015). Age-related processing strategies
and go–nogo effects in task-switching: An ERP study. Frontiers in
Human Neuroscience, 9, 177. DOI: https://doi.org/10.3389
/fnhum.2015.00177, PMID: 26029072, PMCID: PMC4428357
Gajewski, P. D., Ferdinand, N. K., Kray, J., & Falkenstein, M. (2018).
Understanding sources of adult age differences in task switching:
Evidence from behavioral and ERP studies. Neuroscience &
Biobehavioral Reviews, 92, 255–275. DOI: https://doi.org
/10.1016/j.neubiorev.2018.05.029, PMID: 29885425
Grainger, J., Midgley, K. J., & Holcomb, P. J. (2010). Re-thinking the
bilingual interactive-activation model from a developmental
perspective (BIA-d). In M. Kail and M. Hickman (Eds.), Language
acquisition across linguistic and cognitive systems (pp. 267–284).
John Benjamins. DOI: https://doi.org/10.1075/lald.52.18gra
Green, D. W. (1998). Mental control of the bilingual lexico-semantic
system. Bilingualism: Language and Cognition, 1, 67–81. DOI:
https://doi.org/10.1017/S1366728998000133
Izura, C., Cuetos, F., & Brysbaert, M. (2014). Lextale-Esp: A test to
rapidly and efficiently assess the Spanish vocabulary size. Psicológica,
35, 49–66.
Jackson, G. M., Swainson, R., Cunnington, R., & Jackson, S. R. (2001).
ERP correlates of executive control during repeated language
switching. Bilingualism: Language and Cognition, 4, 169–178.
DOI: https://doi.org/10.1017/S1366728901000268
Jamadar, S., Hughes, M., Fulham, W. R., Michie, P. T., &
Karayanidis, F. (2010). The spatial and temporal dynamics of
anticipatory preparation and response inhibition in task-switching.
Neurobiology of Language
644
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
4
6
2
8
1
9
7
9
7
1
5
n
o
_
a
_
0
0
0
3
2
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Language versus task switching
NeuroImage, 51, 432–449. DOI: https://doi.org/10.1016/j
.neuroimage.2010.01.090, PMID: 20123028
Jylkkä, J., Lehtonen, M., Lindholm, F., Kuusakoski, A., & Laine, M.
(2018). The relationship between general executive functions
and bilingual switching and monitoring in language production.
Bilingualism: Language and Cognition, 21, 505–522. DOI:
https://doi.org/10.1017/S1366728917000104
Kang, C., Ma, F., Li, S., Kroll, J. F., & Guo, T. (2020). Domain-
general inhibition ability predicts the intensity of inhibition on
non-target language in bilingual word production: An ERP study.
Bilingualism: Language and Cognition, 23(5), 1056–1069. DOI:
https://doi.org/10.1017/S1366728920000085
Karayanidis, F., & Jamadar, S. D. (2014). Event-related potentials re-
veal multiple components of proactive and reactive control in task
switching. In J. A. Grange & G. Houghton (Eds.), Task switching
and cognitive control (pp. 200–236). Oxford University Press.
DOI: https://doi.org/10.1093/acprof:osobl/9780199921959
.003.0009
Karayanidis, F., Jamadar, S., Ruge, H., Phillips, N., Heathcote, A., &
Forstmann, B. U. (2010). Advance preparation in task-switching:
Converging evidence from behavioral, brain activation, and
model-based approaches. Frontiers in Psychology, 1, 25. DOI:
https://doi.org/10.3389/fpsyg.2010.00025, PMID: 21833196,
PMCID: PMC3153745
Kass, R. E., & Raftery, A. E. (1995). Bayes factors. Journal of the
American Statistical Association, 90, 773–795. DOI: https://doi
.org/10.1080/01621459.1995.10476572
Kiesel, A., Wendt, M., Jost, K., Steinhauser, M., Falkenstein, M.,
Philipp, A. M., & Koch, I. (2010). Control and interference in task
switching: A review. Psychological Bulletin, 136, 849–874. DOI:
https://doi.org/10.1037/a0019842, PMID: 20804238
Kim, C., Cilles, S. E., Johnson, N. F., & Gold, B. T. (2012). Domain
general and domain preferential brain regions associated with
different types of task switching: A meta-analysis. Human Brain
Mapping, 33, 130–142. DOI: https://doi.org/10.1002/hbm
.21199, PMID: 21391260, PMCID: PMC3421461
Lavric, A., Clapp, A., East, A., Elchlepp, H., & Monsell, S. (2019). Is
preparing for a language switch like preparing for a task switch?
Journal of Experimental Psychology: Learning, Memory, and
Cognition, 45, 1224–1233. DOI: https://doi.org/10.1037
/xlm0000636, PMID: 30024253
Lavric, A., Mizon, G. A., & Monsell, S. (2008). Neurophysiological
signature of effective anticipatory task-set control: A task-
switching investigation. European Journal of Neuroscience, 28,
1016–1029. DOI: https://doi.org/10.1111/j.1460-9568.2008
.06372.x, PMID: 18717737
Lemhöfer, K., & Broersma, M. (2012). Introducing LexTALE: A quick
and valid lexical test for advanced learners of English. Behavior
Research Methods, 44, 325–343. DOI: https://doi.org/10.3758
/s13428-011-0146-0, PMID: 21898159, PMCID: PMC3356522
Linck, J. A., Schwieter, J. W., & Sunderman, G. (2020). The differ-
ential role of executive functions in the cognitive control of
language switching. Languages, 5, 19. DOI: https://doi.org/10
.3390/languages5020019
Liu, H., Liang, L., Dunlap, S., Fan, N., & Chen, B. (2016). The effect of
domain-general inhibition-related training on language switching:
An ERP study. Cognition, 146, 264–276. DOI: https://doi.org
/10.1016/j.cognition.2015.10.004, PMID: 26491833
Manzi, A., Nessler, D., Czernochowski, D., & Friedman, D. (2011).
The development of anticipatory cognitive control processes in
task-switching: An ERP study in children, adolescents, and young
adults. Psychophysiology, 48, 1258–1275. DOI: https://doi.org
/10.1111/j.1469-8986.2011.01192.x, PMID: 21371043,
PMCID: PMC3130085
Martin, C. D., Strijkers, K., Santesteban, M., Escera, C., Hartsuiker, R. J.,
& Costa, A. (2013). The impact of early bilingualism on controlling a
language learned late: An ERP study. Frontiers in Psychology, 4, 815.
DOI: https://doi.org/10.3389/fpsyg.2013.00815, PMID: 24204355,
PMCID: PMC3817381
Meuter, R. F., & Allport, A. (1999). Bilingual language switching in
naming: Asymmetrical costs of language selection. Journal of
Memory and Language, 40, 25–40. DOI: https://doi.org/10.1006
/jmla.1998.2602
Nichols, E. S., Wild, C. J., Stojanoski, B., Battista, M. E., & Owen,
A. M. (2020). Bilingualism affords no general cognitive advan-
tages: A population study of executive function in 11,000 people.
Psychological Science, 31, 548–567, DOI: https://doi.org/10.1177
/0956797620903113, PMID: 32310712
Nozari, N., & Novick, J. (2017). Monitoring and control in lan-
guage production. Current Directions in Psychological Science,
26, 403–410. DOI: https://doi.org/10.1177/0963721417702419
Peeters, D. (2020). Bilingual switching between languages and lis-
teners: Insights from immersive virtual reality. Cognition, 195,
104107. DOI: https://doi.org/10.1016/j.cognition.2019.104107,
PMID: 31731119
Peeters, D., & Dijkstra, T. (2018). Sustained inhibition of the native
language in bilingual language production: A virtual reality ap-
proach. Bilingualism: Language and Cognition, 21, 1035–1061.
DOI: https://doi.org/10.1017/S1366728917000396
Philipp, A. M., Jolicoeur, P., Falkenstein, M., & Koch, I. (2007).
Response selection and response execution in task switching:
Evidence from a go-signal paradigm. Journal of Experimental
Psychology: Learning, Memory, and Cognition, 33, 1062–1075.
DOI: https://doi.org/10.1037/0278-7393.33.6.1062, PMID:
17983313
Philipp, A. M., & Koch, I. (2016). Action speaks louder than words,
even in speaking: The influence of (no) overt speech production
on language-switch costs. In J. W. Schwieter (Ed.), Cognitive con-
trol and consequences in the multilingual mind (pp. 127–144).
John Benjamins Publishing. DOI: https://doi.org/10.1075/bpa
.2.06phi
Prior, A., & MacWhinney, B. (2010). A bilingual advantage in task
switching. Bilingualism, 13, 253–262. DOI: https://doi.org/10
.1017/S1366728909990526
Provost, A., Jamadar, S., Heathcote, A., Brown, S. D., &
Karayanidis, F. (2018). Intertrial RT variability affects level of target-
related interference in cued task switching. Psychophysiology,
55, e12971. DOI: https://doi.org/10.1111/psyp.12971, PMID:
28776698
Reverberi, C., Kuhlen, A., Abutalebi, J., Greulich, R. S., Costa, A.,
Seyed-Allaei, S., & Haynes, J. D. (2015). Language control in
bilinguals: Intention to speak vs. execution of speech. Brain
and Language, 144, 1–9. DOI: https://doi.org/10.1016/j.bandl
.2015.03.004, PMID: 25868150
Roelofs, A. (2003). Goal-referenced selection of verbal action:
Modeling attentional control in the Stroop task. Psychological
Review, 110, 88–125. DOI: https://doi.org/10.1037/0033-295X
.110.1.88, PMID: 12529058
Roelofs, A. (2021). How attention controls naming: Lessons from
Wundt 2.0. Journal of Experimental Psychology: General. DOI:
https://doi.org/10.1037/xge0001030, PMID: 33523683
Rouder, J. N., Speckman, P. L., Sun, D., Morey, R. D., & Iverson, G.
(2009). Bayesian t tests for accepting and rejecting the null
hypothesis. Psychonomic Bulletin & Review, 16, 225–237.
DOI: https://doi.org/10.3758/PBR.16.2.225, PMID: 19293088
Struys, E., Woumans, E., Nour, S., Kepinska, O., & van den Noort,
M. (2019). A domain-general monitoring account of language
switching in recognition tasks: Evidence for adaptive control.
Neurobiology of Language
645
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
4
6
2
8
1
9
7
9
7
1
5
n
o
_
a
_
0
0
0
3
2
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Language versus task switching
Bilingualism: Language and Cognition, 22, 606–623. DOI:
https://doi.org/10.1017/S1366728918000342
Sulpizio, S., Del Maschio, N., Del Mauro, G., Fedeli, D., &
Abutalebi, J. (2020). Bilingualism as a gradient measure modulates
functional connectivity of language and control networks.
NeuroImage, 205, 116306. DOI: https://doi.org/10.1016/j.neuroimage
.2019.116306, PMID: 31654763
Thomas, M. S., & Allport, A. (2000). Language switching costs in
bilingual visual word recognition. Journal of Memory and
Language, 43, 44–66. DOI: https://doi.org/10.1006/jmla.1999.2700
Tieges, Z., Snel, J., Kok, A., Plat, N., & Ridderinkhof, R. (2007).
Effects of caffeine on anticipatory control processes: Evidence from
a cued task-switch paradigm. Psychophysiology, 44, 561–578.
DOI: https://doi.org/10.1111/j.1469-8986.2007.00534.x, PMID:
17539921
Timmer, K., Christoffels, I. K., & Costa, A. (2019). On the flexibility
of bilingual language control: The effect of language context.
Bilingualism: Language and Cognition, 22, 555–568. DOI:
https://doi.org/10.1017/S1366728918000329
Verhoef, K., Roelofs, A., & Chwilla, D. J. (2009). Role of inhibition
in language switching: Evidence from event-related brain poten-
tials in overt picture naming. Cognition, 110, 84–99. DOI: https://
doi.org/10.1016/j.cognition.2008.10.013, PMID: 19084830
Verhoef, K. M., Roelofs, A., & Chwilla, D. J. (2010). Electrophysiological
evidence for endogenous control of attention in switching be-
tween languages in overt picture naming. Journal of Cognitive
Neuroscience, 22, 1832–1843. DOI: https://doi.org/10.1162/jocn
.2009.21291, PMID: 19580393
Wagenmakers, E.-J., Love, J., Marsman, M., Jamil, T., Ly, A., Verhagen,
J., Selker, R., Gronau, Q. F., Dropmann, D., Boutin, B., Meerhoff, F.,
Knight, P., Raj, A., van Kesteren, E. J., van Doorn, J., Šmíra, M.,
Epskamp, S., Etz, A., Matzke, D., … Morey, R. D. (2018).
Bayesian inference for psychology. Part II: Example applications
with JASP. Psychonomic Bulletin & Review, 25, 58–76. DOI:
https://doi.org/10.3758/s13423-017-1323-7, PMID: 28685272,
PMCID: PMC5862926
Weissberger, G. H., Gollan, T. H., Bondi, M. W., Clark, L. R., &
Wierenga, C. E. (2015). Language and task switching in the bilingual
brain: Bilinguals are staying, not switching, experts. Neuropsychologia,
66, 193–203. DOI: https://doi.org/10.1016/j.neuropsychologia
.2014.10.037, PMID: 25446970, PMCID: PMC4596720
Zheng, X., Roelofs, A., Erkan, H., & Lemhöfer, K. (2020). Dynamics
of inhibitory control during bilingual speech production: An
electrophysiological study. Neuropsychologia, 140, 107387.
DOI: https://doi.org/10.1016/j.neuropsychologia.2020.107387,
PMID: 32057938
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
4
6
2
8
1
9
7
9
7
1
5
n
o
_
a
_
0
0
0
3
2
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neurobiology of Language
646