RESEARCH ARTICLE
Neural Mechanisms Underlying the Dynamic
Updating of Native Language
a n o p e n a c c e s s
j o u r n a l
Kelly Sharer
and Malathi Thothathiri
Department of Speech, Language and Hearing Sciences, The George Washington University, Washington, DC, USA
Keywords: sentence comprehension, syntax, adaptation, left inferior frontal gyrus (LIFG), multiple-
demand (MD) network, cognitive control
ABSTRACT
Language users encounter different sentence structures from different people in different contexts.
Although syntactic variability and adults’ ability to adapt to it are both widely acknowledged,
the relevant mechanisms and neural substrates are unknown. We hypothesized that syntactic
updating might rely on cognitive control, which can help detect and resolve mismatch between
prior linguistic expectations and new language experiences that countervail those expectations
and thereby assist in accurately encoding new input. Using functional neuroimaging (fMRI),
we investigated updating in garden-path sentence comprehension to test the prediction that
regions within the left inferior frontal cortex might be relevant neural substrates, and additionally,
explored the role of regions within the multiple demand network. Participants read ambiguous and
unambiguous main-verb and relative-clause sentences. Ambiguous relative-clause sentences led
to a garden-path effect in the left pars opercularis within the lateral frontal cortex and the left
anterior insula/frontal operculum within the multiple demand network. This effect decreased upon
repeated exposure to relative-clause sentences, consistent with updating. The two regions showed
several contrastive patterns, including different activation relative to baseline, correlation with
performance in a cognitive control task (the Stroop task), and verb-specificity versus generality in
adaptation. Together, these results offer new insight into how the brain updates native language.
They demonstrate the involvement of left frontal brain regions in helping the language system
adjust to new experiences, with different areas playing distinct functional roles.
INTRODUCTION
Humans begin to learn and appear to master the fundamental aspects of their native language
during childhood (e.g., Fernald, Thorpe, & Marchman, 2010; Gómez, 2002; Mehler et al.,
1988; Newport, 1990; Scott & Fisher, 2009; Smith & Yu, 2008; Werker & Tees, 2005). But they
also need to continually adapt this language throughout the lifespan. Adults face considerable
variation in how any given language is used—they have to interact with and understand different
people who have different lexical and syntactic preferences that vary depending on dialect,
education, and social context (Tagliamonte, 2011). In situations where a reader or a listener
encounters a distribution of syntactic structures that is different from their own language experi-
ence (e.g., a different dialect or register), mechanisms that enable the brain to update syntactic
expectations would be advantageous (Fine, Jaeger, Farmer, & Qian, 2013). Observational and
experimental evidence suggests that language users are indeed capable of adjusting their native
language comprehension in different environments. But the neural mechanisms that support such
dynamic updating over the course of syntactic experience are neither known nor fully explored.
Citation: Sharer, K., & Thothathiri, M.
(2020). Neural mechanisms underlying
the dynamic updating of native
language. Neurobiology of Language,
1(4), 492–522. https://doi.org/10.1162
/nol_a_00023
DOI:
https://doi.org/10.1162/nol_a_00023
Supporting Information:
https://doi.org/10.1162/nol_a_00023
Received: 9 September 2019
Accepted: 25 August 2020
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Author:
Malathi Thothathiri
malathi@gwu.edu
Handling Editor:
Evelina Fedorenko
Copyright: © 2020 Massachusetts
Institute of Technology. Published
under a Creative Commons Attribution
4.0 International (CC BY 4.0) license.
The MIT Press
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
4
4
9
2
1
8
6
7
8
1
1
n
o
_
a
_
0
0
0
2
3
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Dynamic updating of native language
(For related evidence from priming studies, see Devauchelle, Oppenheim, Rizzi, Dehaene, &
Pallier, 2009; Ledoux, Traxler, & Swaab, 2007; Noppeney & Price, 2004; Segaert, Kempen,
Petersson, & Hagoort, 2013; Segaert, Menenti, Weber, Petersson, & Hagoort, 2011; Tooley,
Traxler, & Swaab, 2009.) The present article seeks to address this question. We develop and test
neural predictions arising from an integrative framework that combines ideas from different
strands of the neurolinguistics and psycholinguistics literature. To our knowledge, the results
offer some of the first neural evidence on syntactic updating.
Sentence Processing Is Incremental and Sensitive to Syntactic Probabilities
One relevant set of findings and conclusions underlying the current framework pertains to incre-
mental processing within language. Sentences unfold over time. Therefore, listeners and readers
could either wait until the end of the sentence to determine its structure and interpretation, or they
could use incremental evidence available at any given moment to interpret the sentence on an
ongoing basis. The former ensures a correct interpretation but could be inefficient. In contrast,
the latter is more efficient but could, on occasion, lead to a wrong interpretation that would have
to be corrected.
Considerable prior evidence examining ambiguous, garden-path sentences suggests that sen-
tence processing is incremental. In these types of sentences, a sequence of words is temporarily
consistent with multiple syntactic structures. For example, a well-studied ambiguity involves the
choice between main verb (MV) and relative clause (RC) structures, as demonstrated in (1) and (2).
(1) MV Ambiguous: The experienced soldiers warned about the dangers before the midnight
raid.
(2) RC Ambiguous: The experienced soldiers warned about the dangers conducted the
midnight raid.
The initial segments of (1) and (2) are identical and involve the verb warned. The two sen-
tences differ in whether this verb is to be ultimately interpreted as the main verb that describes
the primary action enacted by the soldiers (who did the warning, as in [1] or as part of a relative
clause that provides secondary information about the soldiers (who were warned, as in [2]).
Thus, these sentences are temporarily ambiguous between two possible syntactic analyses
until the critical disambiguating information (e.g., conducted the … in [2]) arrives. Several
studies examining this and other syntactic ambiguities have found a garden-path effect
whereby unexpected, less common completions (e.g., RC) increase reaction times (RTs),
especially during the disambiguating portion of the sentence, as compared to expected,
more common completions (e.g., MV; Frazier & Rayner, 1982; MacDonald, 1994;
MacDonald, Just, & Carpenter, 1992; Trueswell, Tanenhaus, & Kello, 1993). This suggests
that the sentence processing system generates relatively quick syntactic analyses and
interpretations that keep pace with (or even jump ahead of ) what is heard or read instead
of waiting for the end of the sentence and all relevant information (Friederici & Kotz, 2003;
Osterhout & Holcomb, 1992; Osterhout, Holcomb, & Swinney, 1994; Trueswell &
Tanenhaus, 1994; see also Altmann & Kamide, 1999; Bernolet & Hartsuiker, 2010;
Federmeier & Kutas, 1999; Ferretti, McRae, & Hatherell, 2001; Kamide, Altmann, &
Haywood, 2003; Tanenhaus, Carlson, & Trueswell, 1989). Note that throughout this paper,
we refer to syntactic probabilities, analyses, expectations, and updating for the sake of
simplicity. Syntactic probabilities are intertwined with semantic and lexical factors in natural
language. Thus, factors other than syntax could guide expectations and interpretations during
sentence processing (see, e.g., Kim & Osterhout, 2005; Levy, 2008; Snedeker & Trueswell,
Neurobiology of Language
493
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
4
4
9
2
1
8
6
7
8
1
1
n
o
_
a
_
0
0
0
2
3
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Dynamic updating of native language
2004; Thothathiri, Kim, Trueswell, & Thompson-Schill, 2012). The present hypotheses about
updating expectations are agnostic about whether those expectations arise from syntax alone
or from a combination of cues.
Prior evidence suggests that probability is a key factor that drives expectations during sentence
processing. Under a probabilistic parsing model (e.g., Hale, 2001; Levy, 2008), multiple possible
syntactic structures are activated during incremental sentence processing and preferentially ranked
according to their probability derived from past experience. Processing difficulty arises whenever a
word is unexpected or surprising given the previous words in the sentence. These accounts can
explain the garden-path effects described above: Sentences like (2) lead to processing difficulty
in the disambiguating region (e.g., conducted the…) because those words are not commonly
encountered in the context of the previous words (Hale, 2001; Levy, 2008).
Past evidence also suggests that the brain is sensitive to syntactic probability on different levels,
including a verb-general level that tracks how often a structure is experienced independent of the
verb, and a verb-specific level that encodes how often a particular verb appears in a particular
structure. Sentences that are less common overall lead to greater processing difficulty (Frazier &
Rayner, 1982; MacDonald et al., 1992). In addition, the probability distributions used to make
predictions during incremental processing can vary according to the verb. For example,
Osterhout et al. (1994) demonstrated that reading less common sentential complement (SC)
sentences only elicited a P600 (indicating syntactic difficulty and reanalysis) when the sentences
contained verbs biased towards an alternative direct-object structure and not when they con-
tained verbs that appeared in SC sentences more frequently (see also Trueswell et al., 1993). In
fact, similar verb-specific effects are found even for frequently encountered, less complex struc-
tures like the MV. Wilson and Garnsey (2009) reported slower reading times for ambiguous MV
sentences containing RC-biased verbs than for those containing MV-biased verbs, indicating the
influence of individual verbs’ statistical preferences independent of syntactic complexity.
Adults Adapt to Changing Syntactic Probabilities
Syntactic probabilities are not static but dynamic, changing with new language input. They also
vary depending on modality (e.g., spoken vs. written language), formality (e.g., formal vs.
informal speech), the speaker’s dialect, and other factors. Thus, it might be useful for the brain
not only to track syntactic probabilities when learning a language, but also to dynamically adapt
syntactic expectations in situations where the probabilities differ from prior experience. Emerging
evidence suggests that adults do adjust their expectations in this way. For example, one eye-
tracking study showed that having heard different speakers express different kinds of meanings,
listeners came to anticipate the meanings that were most associated with each speaker (Kamide,
2012). Similarly, Kroczek and Gunter (2017) showed that when listeners were exposed to two
German speakers, one biased towards using a subject-initial structure and the other biased
towards using an object-initial structure, their subsequent parsing of an ambiguous structure
varied systematically based on the speaker. This speaker effect was maintained one day after
exposure and facilitated quick speaker-based adjustment in a follow-up session nine months
later, demonstrating that speaker-dependent updating of syntactic predictions can be main-
tained over time.
Using a different manipulation, Ryskin and colleagues showed that when exposed to verbs
with different biases for different sentence structures, listeners’ interpretations came to be guided
by those verb biases, which were manipulated during the course of the experiment (Ryskin, Qi,
Duff, & Brown-Schmidt, 2017). More broadly, exposure to uncommon sentence structures
appears to facilitate subsequent processing of those sentence structures in both behavioral
Neurobiology of Language
494
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
4
4
9
2
1
8
6
7
8
1
1
n
o
_
a
_
0
0
0
2
3
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Dynamic updating of native language
(Farmer, Fine, Yan, Cheimariou, & Jaeger, 2014; Fine et al., 2013; Wells, Christiansen, Race,
Acheson, & MacDonald, 2009) and electrophysiological (Coulson, King, & Kutas, 1998) studies.
(Note that there is debate about some behavioral findings and measures—specifically on whether
faster reading times could reflect task-level adaptation instead of syntactic updating (Prasad &
Linzen, 2019; Stack, James, & Watson, 2018). We address task-based adaptation versus syntactic
updating in the Discussion section. For the present, we note that the most recent study on this
topic concluded that syntactic updating (going beyond task practice) is detectable if a study is
adequately powered (Prasad & Linzen, 2019). In all of these cases, the updated comprehension
patterns were observed in adults who were mature native speakers of the language, indicating
that the updating of syntactic probabilities continues beyond the acquisition that occurs in
childhood.
Potential Mechanisms: A Hypothesized Role for Cognitive Control
Updating syntactic probabilities minimally requires the brain to encode the frequency of occur-
rence of different syntactic structures in the current linguistic environment. For example, in a sit-
uation where a reader encounters many RC sentences, the brain must (a) know when it is
experiencing an RC (and not an MV) structure; and (b) update the probability of an RC such that
it becomes relatively more expected in the given situation. In cases of syntactic ambiguity where
the actual sentence structure goes against the statistically prevalent pattern in a language, step (a)
might require the detection and resolution of conflict between the more expected structure (e.g.,
MV) and the less expected structure (e.g., RC). Put another way, to update probabilities based on
exposure to an RC Ambiguous sentence (like [2] above), the more likely but incorrect MV analysis
needs to be discarded in favor of the less likely but correct RC analysis. Following the original
proposal of Novick, Trueswell, and Thompson-Schill (2005), growing evidence has corroborated
the view that cognitive control—the brain’s ability to regulate behavior and handle conflict
between incompatible representations and responses—is a plausible mechanism for such
selection between competing alternatives. Behaviorally, cognitive control ability correlates
with and causally influences the comprehension of conflict-inducing sentences (Hsu &
Novick, 2016; Novick, Kan, Trueswell, & Thompson-Schill, 2009; Thothathiri, Asaro, Hsu, &
Novick, 2018). Neurally, similar regions are activated by cognitive control tasks and language
processing that involves conflict between incompatible interpretations (Hsu, Jaeggi, & Novick,
2017; January, Trueswell, & Thompson-Schill, 2009; Ye & Zhou, 2009; see the detailed discus-
sion below). Thus, while the precise conditions that necessitate the involvement of cognitive con-
trol are sometimes debated, there is near-consensus that this function is relevant for sentence
processing, especially when the structure and/or content of the sentence contravenes prior
expectations.
The present study extends previous work on cognitive control and the processing of individual
garden-path sentences to ask if cognitive control might also be related to how well readers
dynamically adapt their syntactic expectations in an environment where such sentences are
encountered often. We lay out the specific hypothesis and predictions later after a review of
the potential neural substrates below.
Potential Neural Substrates: A Hypothesized Role for Left Inferior Frontal Cortex Regions
As a starting point, we hypothesized that regions known to be involved in both sentence process-
ing and cognitive control might be plausible substrates for syntactic updating. Areas within the left
inferior frontal cortex are obvious candidates because they have been consistently linked to
Neurobiology of Language
495
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
4
4
9
2
1
8
6
7
8
1
1
n
o
_
a
_
0
0
0
2
3
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Dynamic updating of native language
sentence processing in different populations and paradigms. In healthy adults, they show in-
creased activation under higher sentence processing demands (January et al., 2009; Keller,
Carpenter, & Just, 2001; Mason, Just, Keller, & Carpenter, 2003; Rodd, Longe, Randall, &
Tyler, 2010). In priming paradigms, they show repetition suppression when the sentence structure
is repeated (Santi & Grodzinsky, 2010; Segaert et al., 2013; Segaert, Menenti, Weber, Petersson,
& Hagoort, 2011; Weber & Indefrey, 2009). Neuropsychological studies further indicate that pa-
tients with frontal damage are impaired in sentence comprehension (see, e.g., Novick et al., 2005;
Tyler et al., 2011, and references therein). Overall, there is widespread agreement that this patch
of cortex is relevant for sentence comprehension.
Beyond this consensus, there are two recurrent issues that are still debated. First, there is
anatomical and functional heterogeneity within the left inferior cortex. Many previous studies
have suggested explicitly or implicitly that the pars opercularis and pars triangularis are especially
relevant for syntactic processing (Ben-Shachar, Hendler, Kahn, Ben-Bashat, & Grodzinsky, 2003;
Caplan, Alpert, & Waters, 1998; Caplan, Chen, & Waters, 2008; Grodzinsky & Friederici, 2006;
Just, Carpenter, Keller, Eddy, & Thulborn, 1996). However, these findings are also quite variable,
leading to some suggestions that the functional segregation might be weak rather than strong
(Tyler et al., 2011). Therefore, in the present study, we separately analyzed activation patterns
within the pars opercularis, the pars triangularis, and the frontal orbital cortex with the expec-
tation that one or more of those regions would show the predicted effects. We did not take a
strong a priori position on which of these subregions might be the most relevant.
Relatedly, the types of operations supported by the different subregions present a second
issue. One of the earliest proposals was that different regions were specialized for different
aspects of language, such as syntax, phonology, and semantics. For syntax, a longstanding claim
is that Broca’s area (roughly BA 44/45) is involved in specific syntactic operations. Support for
theories under this umbrella has generally taken the form of showing that higher syntactic com-
plexity leads to greater activation (e.g., Ben-Shachar et al., 2003; Santi & Grodzinsky, 2010). A
contrasting perspective has challenged this syntactic specialization view by showing that factors
other than syntactic complexity can explain activation within Broca’s area. For example, one
study showed that conflict between syntactic and semantic cues explained the activation pat-
terns within this region better than did syntactic structure (e.g., passives vs. actives; Thothathiri
et al., 2012). Others have shown that the same Broca’s area subregion within each participant is
recruited for nonsentential as well as sentence processing tasks (January et al., 2009; Ye & Zhou,
2009). Such evidence has been used to argue that region(s) within the left frontal cortex support
the processing of complex sentence structures via their role in selecting between conflicting
representations.
A third class of theories lies between the above two perspectives. For example, the memory,
unification, and control model put forward by Hagoort (2016) suggests that the left frontal cortex
is involved in unification operations—building larger structures from smaller structures stored in
the posterior cortex—but this unification is not limited to syntax. It can also include other domains
(e.g., semantics) that require combinatorial processes. At the same time, this model proposes that
there might be some spatial specialization such that syntactic, semantic, and phonological uni-
fication might especially recruit BA 44/45, BA 45/47, and BA 44/6, respectively (Hagoort, 2016).
Thus, under this model, the processes occurring within the left frontal cortex are described in
broader terms (e.g., selecting and unifying different representations) that are consistent with the
cognitive control account while also allowing for the possibility that those processes might be
specialized in a graded way for different representations (e.g., syntactic vs. semantic) in different
subregions. Other researchers have proposed that functional specialization (e.g., for syntactic
processing) might arise at a network rather than the local level, whereby different networks, all
Neurobiology of Language
496
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
4
4
9
2
1
8
6
7
8
1
1
n
o
_
a
_
0
0
0
2
3
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Dynamic updating of native language
involving a given frontal region, might be important for different tasks (Fedorenko & Thompson-
Schill, 2014; Hsu et al., 2017).
Together, extant evidence does not support a strong version of syntactic modularity within
the left inferior frontal cortex. Recent accounts that argue for some subspecialization neverthe-
less reject the notion that parts of the frontal cortex are localized exclusively for syntax (see,
e.g., Fedorenko, Blank, Siegelman, & Mineroff, 2020; Fedorenko, Duncan, & Kanwisher,
2012; Hagoort, 2016). They suggest instead that these regions might be engaged in broader
linguistic processing and/or unification between different types of representations. But whether
there might be specialization for linguistic processing, more broadly, is still debated. Studies
that have analyzed individual-specific functional regions of interest (ROIs) have been
especially relevant here because group-level analysis can potentially obscure functional
differences between neighboring patches of cortex due to variability between subjects.
Using subject-specific functional ROIs, Fedorenko et al. (2012) found that nearby clusters
within Brodmann areas 44 and 45 had different functional profiles. Within each area, one
cluster that was activated more for non-words than sentences showed a domain-general pro-
file, namely activation for the harder versus easier version of a variety of verbal and nonverbal
tasks. The other cluster, which was activated more for sentences than non-words, was more
selective. It either showed no significant activation or inconsistent results for the non-sentence
tasks. The authors concluded that Broca’s area is not homogenous; it contains multiple regions
that vary in their degree of generality versus specificity (Fedorenko et al., 2012). A related
study showed that the regions activated for sentences over non-words were not significantly
activated for math, working memory, and cognitive control tasks (Fedorenko, Behr, &
Kanwisher, 2011). Synthesizing such evidence, these researchers have suggested that the left
inferior frontal cortex might contain two different kinds of regions—language selective regions
that are specifically activated for lexical and combinatorial linguistic processing, and domain-
general regions that could be involved in sentence processing, especially with a difficult task,
but are not specific to language.
However, other studies using subject-specific functional ROIs have noted significant overlap
between sentence comprehension and cognitive control (Hsu et al., 2017; January et al., 2009;
Ye & Zhou, 2009). Two studies reported that left frontal regions activated by a cognitive control
task (e.g., the Stroop task) were also activated for sentences containing conflict between incom-
patible interpretations (January et al., 2009; Ye & Zhou, 2009). Hsu et al. (2017) similarly found
overlap in the left frontal cortex for Stroop and sentence comprehension (among other tasks).
However functional connectivity of the frontal cortex with other brain regions differed for dif-
ferent tasks, supporting a network-based view of specialization.
In the present study, we hypothesize only that cognitive control plays a role in processing and
updating the statistics for sentences containing conflict between interpretations (see more below).
This claim does not preclude (a) the functional segregation of cognitive control operations for lin-
guistic versus non-linguistic representations (see, e.g., Thothathiri, Gagliardi, & Schwartz, 2012);
or (b) the existence of language-specific regions involved in non-conflict-related processes. With
respect to (a), several of the studies supporting a role for cognitive control have noted overlap
between sentence comprehension and the color-word Stroop task (e.g., January et al., 2009;
Hsu & Novick, 2016; Thothathiri et al., 2018). Fedorenko and colleagues too have reported weak
Stroop effects in some language-specific functional ROIs in some studies (Fedorenko et al., 2011;
Fedorenko et al., 2012). The Stroop task contains linguistic content, so overlap in activation is not
inconsistent with some degree of linguistic specialization. With respect to (b), Hsu et al. (2017)
note that the claim about cognitive control applies to regions identified using a contrast of conflict
versus no-conflict linguistic processing and not to other broader contrasts like sentences versus
Neurobiology of Language
497
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
4
4
9
2
1
8
6
7
8
1
1
n
o
_
a
_
0
0
0
2
3
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Dynamic updating of native language
non-words (which could identify regions engaged in lexical, syntactic, and semantic processes
that are unrelated to cognitive control). Using such a conflict-specific approach, they showed that
conflict functional ROIs in the left frontal cortex were engaged across syntactic and non-syntactic
tasks. We took a similar approach to this and related previous studies in that we examined left
inferior frontal activation for conflict-inducing sentences in particular. Additionally, we used
behavioral performance in a verbal Stroop task to index individual differences in cognitive control,
and asked if those differences predicted individual variation in how activation adapted with
exposure. We did not test and therefore do not make claims about the domain generality of the
conflict resolution processes (unlike some previous studies). Instead, we focus on whether the
cognitive control hypothesis could be extended to longer-term updating going beyond the pro-
cessing of individual conflict sentences.
Potential Neural Substrates: Exploratory Investigation of the Multiple Demand Network
The multiple demand (MD) network includes several brain regions in the frontal and parietal cor-
tices associated with domain-general cognitive control (Duncan, 2010, 2013). These regions are
sensitive to task difficulty across a wide variety of tasks, including arithmetic, spatial and verbal
working memory, and Stroop, demonstrating that their function is not restricted to specific rep-
resentational domains (Fedorenko, Duncan, & Kanwisher, 2013). The network as a whole has
been conceptualized as being important for goal-directed and task-appropriate behavior, includ-
ing organizing subgoals, directing attention, and selecting task-relevant over irrelevant informa-
tion (Duncan, 2010, 2013). Individual regions within the network are thought to be important for
specific aspects of such behavior. For example, the anterior cingulate cortex (ACC) might be
important for detecting and monitoring conflict (Botvinick, Cohen, & Carter, 2004; Ullsperger,
Fischer, Nigbur, & Endrass, 2014). The anterior insula and the frontal operculum (AI/FO) might be
important for sustained task performance, and the inferior frontal and parietal sulci (IFS and IPS)
for faster trial-by-trial adjustments (Dosenbach, Fair, et al., 2007; Dosenbach, Visscher, et al.,
2006; Nomura et al., 2010). Given this profile, it is possible that one or more regions within this
network, especially those in the frontal cortex, are involved in aiding the processing of conflict-
inducing (i.e., RC Ambiguous) sentences that are known to result in processing difficulty.
Accordingly, we conducted exploratory analyses within 10 MD ROIs derived from the prior
literature (Duncan, 2010).
An Integrative Framework and Associated Neural Predictions
The hypothesis explored in this paper derives from three features of sentence processing described
above and reiterated here:
(a) Language processing unfolds incrementally as the sentence is heard or read, and this
incremental processing is guided by the statistical properties of the language;
(b) Listeners and readers update their syntactic expectations as the statistical properties of
language change with new experiences; and
(c) Cognitive control is used to handle conflict that arises from the incremental processing
of sentences whose structures violate prior expectations and statistical properties.
Based on (a) to (c), we hypothesized that:
The dynamic updating of native language uses cognitive control, which can help with correctly
encoding new language experiences that violate prior expectations and thereby aid in the
accurate updating of syntactic probabilities.
Neurobiology of Language
498
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
4
4
9
2
1
8
6
7
8
1
1
n
o
_
a
_
0
0
0
2
3
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Dynamic updating of native language
Importantly, we are agnostic as to whether cognitive control plays a direct role in updating
probabilities per se. We propose only that one possible (indirect) role of cognitive control is to
help the parser arrive at the correct analysis of sentences that violate prior expectations, which
in turn serves as the input for updating. Prior behavioral evidence suggests that language users
do adapt to the statistics of novel language environments. In this paper, we hypothesized and
tested predictions about the neural substrates that are involved in this process.
Based on prior evidence for the involvement of left inferior frontal regions in sentence process-
ing and cognitive control, we predicted that one or more of these regions would be engaged in
syntactic updating. As a potential contrast, we also looked at the middle temporal gyrus (MTG),
which has been consistently linked to syntactic processing but not cognitive control. Frontal and
temporal cortices may be concurrently engaged during sentence processing but play different
roles. In particular, it has been suggested that the MTG might subserve the storage of syntactic
knowledge while frontal regions might be involved in more dynamic processes, including re-
solving conflict between or unifying different linguistic representations (e.g., Hagoort, 2016;
Novick et al., 2005). Therefore, we predicted that one or more of the pars opercularis, the pars
triangularis, and the frontal orbital cortex, but not the MTG, would show patterns corresponding
to dynamic adjustment, as outlined below.
Additionally, based on the association between the MD network and domain-general cogni-
tive control, we expected that one or more regions within this network could be involved in
handling conflict and/or difficulty during sentence comprehension. Below, we lay out the predic-
tions for the different ROIs after we describe the study design.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
4
4
9
2
1
8
6
7
8
1
1
n
o
_
a
_
0
0
0
2
3
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
The Present Study
We chose the MV/RC ambiguity described above because it is known to generate robust garden-
path effects. Each participant underwent three runs of functional neuroimaging (fMRI). They read
ambiguous and unambiguous MV and RC sentences, indicated reading completion, and
answered intermittent comprehension questions to verify compliance. There were four types of
sentences, (1) to (4), including the first two sentence types repeated from above.
(1) MV Ambiguous: The experienced soldiers warned about the dangers before the midnight
raid.
(2) RC Ambiguous: The experienced soldiers warned about the dangers conducted the
midnight raid.
(3) MV Unambiguous: The experienced soldiers spoke about the dangers before the midnight
raid.
(4) RC Unambiguous: The experienced soldiers who were told about the dangers conducted
the midnight raid.
Unambiguous sentences like (3) and (4) served as control conditions for comparison with the
corresponding ambiguous sentences (MV Ambiguous was compared to MV Unambiguous, and
RC Ambiguous was compared to RC Unambiguous). The unambiguous sentence types led to a
single interpretation and were not expected to generate any conflict. Ambiguous sentences like
(1) and (2) were temporarily consistent with two possible analyses. We chose verbs that were
predominantly biased towards the MV sentence structure (see the Materials and Methods section).
This meant that the MV Ambiguous condition was not expected to generate conflict despite the
presence of temporary ambiguity, because the expectation created from prior experience would
be consistent with the actual sentence structure. In contrast, the RC Ambiguous condition was
Neurobiology of Language
499
Dynamic updating of native language
predicted to involve conflict because prior experience would lead readers to expect MV, which
would be contradicted by information in the sentence that indicates an RC structure instead.
The first run preceded the exposure phase of the study. It was intended to evaluate which ROIs
showed the garden-path effect, namely increased activation specifically in the case of conflict
between the expected and actual sentence structures. Concretely, this corresponds to a larger
ambiguity effect for RC sentences (RC Ambiguous minus RC Unambiguous) than MV sentences
(MV Ambiguous minus MV Unambiguous). A priori, we predicted that one or more of the left
inferior frontal ROIs would show this pattern of activation. Contingent on the results from
Run1, we restricted all subsequent analyses to the regions showing the garden-path effect.
The second run constituted the exposure phase and consisted only of RC sentences. We
hypothesized that if a region were involved in updating expectations based on new language
experiences, it would show a decrease in the RC ambiguity effect from the beginning to the
end of the exposure phase. Thus, we would expect a negative slope over the course of the expo-
sure run. Further, we predicted that if updating were related to cognitive control, then individual
differences in this ability might correspond to how activation changes with exposure.
Specifically, those with better cognitive control could encode the RC experiences more accu-
rately and therefore show larger decreases in the ambiguity effect compared to those with poorer
cognitive control. We tested this prediction by computing the correlation between performance
on a Stroop task and the change in ambiguity-related activation (slope) from the beginning to the
end of the exposure phase.
The third and final run constituted the post-exposure period. It served as a counterpart to the
first run, which was pre-exposure. The question here was whether any updating effect observed in
the exposure phase persisted across runs, that is to say, over a gap of several minutes and whether
this updating was restricted to verbs repeatedly seen in RC sentences or generalized more broadly
to verbs not seen during the exposure phase. If updating were verb-specific, we would expect a
garden-path effect for unexposed but not exposed verbs and a significant difference between
ambiguous RC sentences with the two kinds of verbs. Conversely, if updating were verb-general,
we would expect no garden-path effect for either verb type and no difference between ambiguous
RC sentences with the two verb types.
Our predictions pertain to each run separately because the three runs had different design
characteristics. The exposure phase (Run2) presented verbs in RC structures repeatedly but the
other runs did not. MV structures were present in Run1 and Run3, but not Run2. A difference
between exposed and unexposed verbs could be investigated in Run3 but not in the other runs.
Different design matrices for the three runs could lead to differences in parameter estimates and
scale factors. Thus, the results within each run are interpretable but cannot be compared directly
across runs (Mumford, 2007; Pernet, 2014).
To summarize, we tested four predictions:
P.1: In Run1, a larger RC than MV ambiguity effect in one or more of the left inferior frontal
ROIs.
P.2: In Run2, a negative slope corresponding to a decrease in the RC ambiguity effect from
the beginning to the end of the exposure phase in the conflict-sensitive region(s) showing
a garden-path effect in Run1.
P.3: In Run2, correlation between Stroop scores and the decrease in ambiguity-related activa-
tion over Run2 in the conflict-sensitive regions.
P.4: In Run3, a garden-path effect for unexposed but not exposed verbs in the conflict-sensitive
regions, corresponding to verb-specific updating.
Neurobiology of Language
500
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
4
4
9
2
1
8
6
7
8
1
1
n
o
_
a
_
0
0
0
2
3
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Dynamic updating of native language
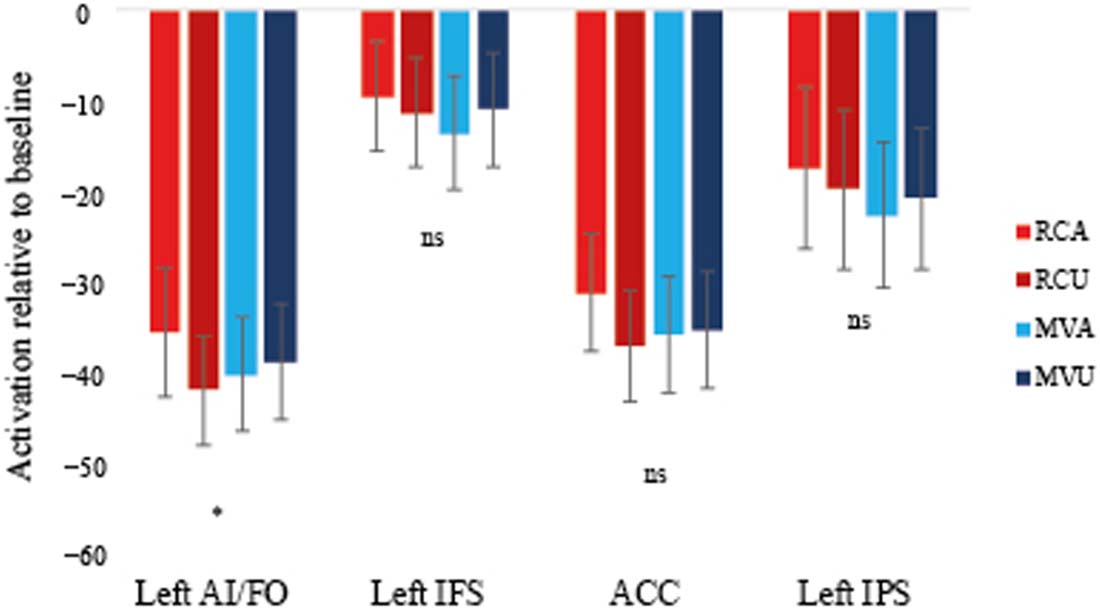
In addition to the above main predictions, we explored whether one or more frontal regions
within the MD network also showed conflict sensitivity and adaptation over time.
MATERIALS AND METHODS
Participants
Thirty-one right-handed and neurotypical native English speakers with normal or corrected to
normal vision from the Washington, DC, area participated in the study. Three were excluded
due to inability to complete all portions of the fMRI data acquisition, recording equipment
malfunction, and falling asleep during the study. Data from the remaining twenty-eight partici-
pants (18–27 years; M = 21.11; 16 female) were analyzed. All underwent an MRI safety screening
and provided informed consent under a protocol approved by the George Washington University
Institutional Review Board. Participants were paid $25 or given course credit.
Materials and Procedure
Participants completed a Stroop task outside the scanner and a sentence reading and baseline
task inside the scanner. They were familiarized with each task prior to data acquisition
(Stroop: eight practice trials; Sentence reading: eight practice trials (four comprehension ques-
tions); Baseline: four practice trials). All stimuli were presented using E-Prime 2.0 (Psychology
Software Tools, Pittsburgh, PA). For the Stroop task, participants completed a single iteration of
practice. For the scanner tasks (intermixed sentence reading, comprehension questions, and
baseline), participants repeated practice two to four times to fully grasp the different response
mappings and instructions for the different tasks. Practice ended when a participant achieved
greater than 50% accuracy. (Note that for one participant, we could not compute accuracy during
practice due to a data recording problem.)
The Stroop task
The Stroop task was completed in a quiet testing room outside the MRI scanner. Participants
saw one word per trial, displayed in blue, green, or yellow 40-point Courier New font against
a black background. They were asked to indicate the font color by pressing the appropriate
colored button. Stimuli belonged to four conditions. In the Neutral condition, the words (deal,
farmer, horse, plenty, stage, and tax) were unrelated to the font color. The Congruent condition
contained words that were the same as the font color (e.g., blue in blue font). On Incongruent–
Eligible trials, the color words were different from the font color (e.g., green in blue font). The
word meaning (e.g., green) was an available response option, thereby giving rise to conflict at
both the representational and the response levels. In contrast, Incongruent–Ineligible trials
consisted of color words (brown, orange, and red ) that were different from the font color
(e.g., red in blue font), but the word meaning (e.g., red) was not an available response option.
Therefore, these trials were expected to give rise to representational conflict only (no response
conflict).
Participants completed four blocks of 72 trials each (18 neutral, 18 congruent, 18 incongruent–
eligible, and 18 incongruent–ineligible trials per block). Each trial began with a 300 ms fixation
cross, followed by a 500 ms blank screen and then the stimulus word. Stimuli stayed visible
until the participant responded by pressing the 4 (blue), 5 (green), or 6 (yellow) key. Stimulus
order was pseudorandomized to avoid more than three repetitions of colors or stimulus words
in a row.
Neurobiology of Language
501
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
4
4
9
2
1
8
6
7
8
1
1
n
o
_
a
_
0
0
0
2
3
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Dynamic updating of native language
The sentence reading task
The main sentence reading task was completed inside the scanner. Participants silently read MV
and RC sentences presented in black 14-point Courier New font on a white background. When
they finished reading a sentence, they used the index finger of their right hand to press the leftmost
of three buttons to indicate completion. On some trials, they answered a subsequent yes/no
comprehension question by using the middle finger of their right hand to press the middle button
for Yes or the ring finger of their right hand to press the rightmost button for No. The comprehension
questions ensured that participants were attending to the task and reading the sentences.
Sentences belonged to one of four types: MV Ambiguous, RC Ambiguous, MV Unambiguous,
and RC Unambiguous (see examples [1] to [4] above). Twenty-four sentences were modified
from Fine et al. (2013). The remainder were newly created for this study. All verbs used in the
ambiguous sentences were at least 1.5 times more likely to appear in MV than RC structures
(based on average frequencies calculated from the following corpora: Brown, Wall Street
Journal, Switchboard, and British National Corpus; Roland, Dick, & Elman, 2007). Thus, readers
were expected to initially interpret the ambiguous sentences as MV, which should lead to garden-
pathing when the sentence happens to be RC. The corresponding unambiguous sentence types
did not contain any ambiguity. For MV Unambiguous sentences, ambiguity was avoided by using
either intransitive verbs (e.g., remained) or transitive verbs that have different forms in MV versus
RC structures (e.g., spoke vs. spoken). For RC Unambiguous sentences, ambiguity was avoided
by including an explicit relativizer (e.g., that, who).
Each participant completed three functional runs within the scanner. Run1 and Run3 con-
tained 120 sentences each (30 of each type). Within each of these runs, 30 verbs appeared once
in MV Ambiguous and once in RC Ambiguous (a total of 60 ambiguous sentences). Thirty unique
verbs occurred in MV Unambiguous sentences, and a different set of 30 unique verbs occurred in
RC Unambiguous sentences (a total of 60 unambiguous sentences). Run2 provided exposure to
RC sentences. Fifteen of the 30 ambiguous verbs used in Run1 were used eight times each in Run2
(four times in RC Ambiguous and four times in RC Unambiguous. Total N = 120). There were no
MV sentences. Thus, the design may be summarized as pre-RC-exposure (Run1), RC-exposure
(Run2), and post-RC-exposure (Run3).
Twenty sentences in each run were followed by yes/no comprehension questions (e.g.,
Sentence: The silly boys who were reprimanded during the play quickly left the auditorium;
Question: Did the boys leave the auditorium quickly?; Correct answer: Yes. Sentence: The
farm animals washed in their pens at the state fair; Question: Were the animals roaming free?;
Correct answer: No). The questions were randomly interspersed, followed each sentence type
an equal number of times, pertained to different parts of the sentences, and did not call atten-
tion to the syntactic ambiguity (when present). This design was chosen to mitigate the possi-
bility of explicit strategies for ambiguity resolution while also encouraging the processing of all
parts of each sentence. The correct answer was No for half of the trials.
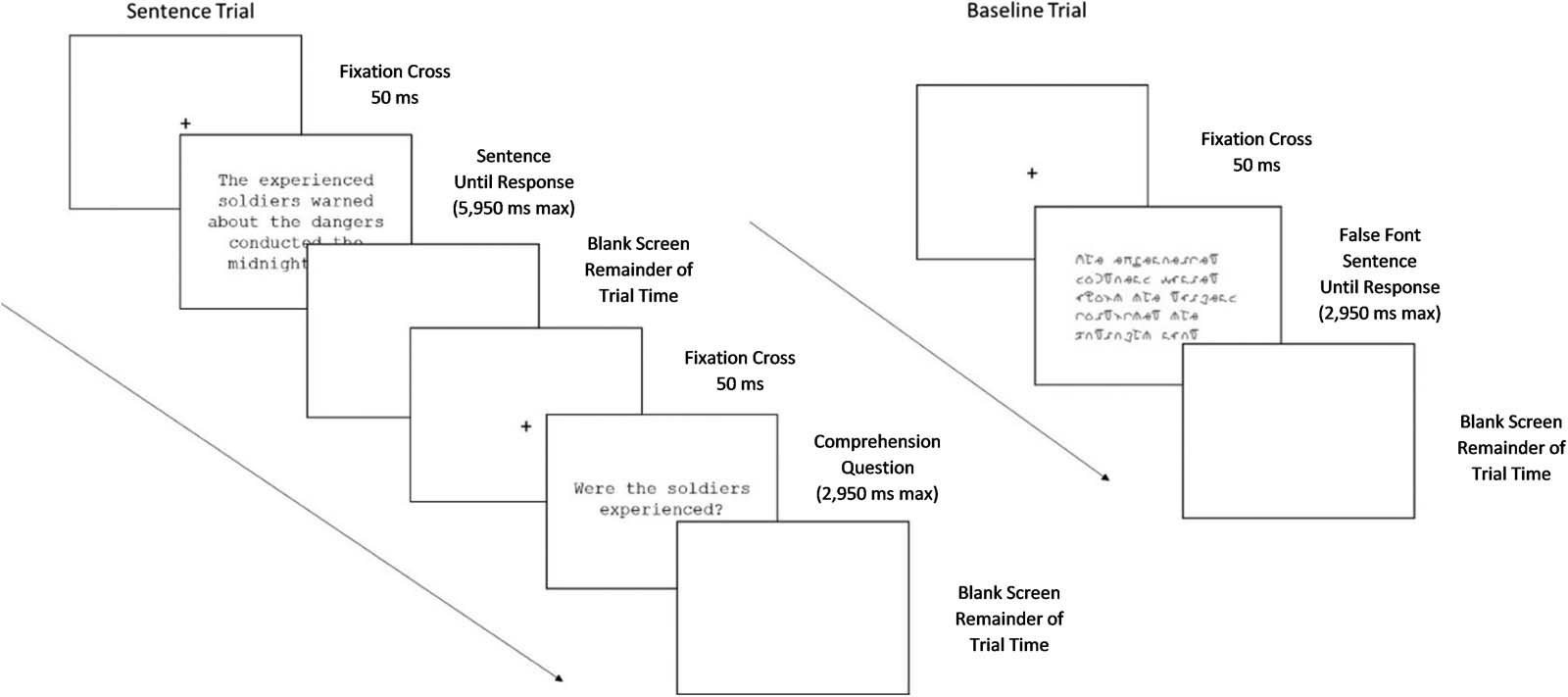
Sentence trials lasted 6 s (50 ms fixation cross followed by sentence presentation for a max-
imum of 5,950 ms). If a participant responded before the allotted time had finished, a blank
screen replaced the stimulus until the trial-time elapsed. Comprehension questions, when
present, followed the sentences (after the allotted 6 s). Comprehension question trials lasted
3 s (50 ms fixation cross followed by question presentation for a maximum of 2,950 ms). The
question remained on the screen until participants selected a response. Following a response,
a blank screen replaced the question until the trial-time elapsed (Figure 1).
Sentence stimuli were counterbalanced in a number of ways. Four counterbalanced lists were
created such that any “semantic frame” (e.g., The experienced soldiers [main verb OR relative
Neurobiology of Language
502
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
4
4
9
2
1
8
6
7
8
1
1
n
o
_
a
_
0
0
0
2
3
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Dynamic updating of native language
Figure 1.
Sequence of stimulus presentation in sentence (left) and baseline (right) trials.
clause] about the dangers [preposition OR main verb] the midnight raid.) appeared in an MV
Ambiguous structure in one list, an RC Ambiguous structure in another list, and so on. This
ensured that the types of events described by the sentences were roughly semantically equivalent
between lists. Participants were randomly assigned to a list. We also counterbalanced which
verbs were presented in Run2 (verbs presented during Run2 in lists 1 and 3 were absent in lists
2 and 4 and vice versa). Examples illustrating the counterbalancing are shown in the Appendix in
the online supporting information located at https://www.mitpressjournals.org/doi/suppl
/10.1162/nol_a_00023. Sentence length was adjusted such that there were no significant differ-
ences between MV and RC sentences overall (Run1: t < 1.5, p > 0.05; Run3: t < 1, p > 0.05).
The baseline task
Baseline trials were pseudorandomly interspersed between sentence trials. Participants visually
scanned lines of false-font (BACS2serif; Vidal, Chetail, & Content, 2016) and pressed the middle
(Yes) or the right (No) buttons to indicate whether they saw the numeral “4” within the false-font
characters. Half of the trials contained the numeral at a randomly selected location within the
sequence. The false-font stimuli were matched in the number of lines and angles to a subset of
the experimental sentences. Thus, the baseline task controlled for visual processing and manual
button presses but did not involve linguistic processing.
Run1 and Run3 each contained 30 baseline trials; Run2 contained 60 trials. The number of
baseline trials equaled the number of trials for each sentence type. Thus, the baseline task was not
rare, to help minimize the possibility of a “rare event” response that could complicate interpre-
tation (e.g., Busse & Woldorff, 2003). The order of baseline and sentence trials was randomized
using Optseq2 (Greve, 2002; http://surfer.nmr.mgh.harvard.edu/optseq/). Each baseline trial
lasted 3 s (50 ms fixation cross followed by the stimulus for a maximum of 2,950 ms). If a partic-
ipant responded before the allotted time had finished, a blank screen replaced the stimulus until
the trial-time elapsed (Figure 1).
Dependent Measures and Analyses
Stroop task
Reaction time and accuracy were recorded on each trial. Trials with inaccurate responses were
excluded from the RT analysis. For each participant, a normalized representational conflict
Neurobiology of Language
503
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
4
4
9
2
1
8
6
7
8
1
1
n
o
_
a
_
0
0
0
2
3
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Dynamic updating of native language
processing score was calculated as (mean RT on Incongruent–Ineligible trials − mean RT on neu-
tral trials) ÷ (Mean RT on neutral trials) * 100. Higher scores indicate poorer conflict processing.
We focused on Incongruent–Ineligible trials and representational conflict because this type of
conflict should be more similar to the conflict between alternative interpretations elicited by
garden-path sentences (Hsu & Novick, 2016; Thothathiri et al., 2018).
Sentence reading fMRI analyses
Structural and functional images were acquired using a 3T Siemens Trio Scanner. All scans took
place at the Center for Functional and Molecular Imaging at Georgetown University. A sagittal
T1-weighted MPRAGE sequence (TR = 1,900 ms, TE = 2.52 ms, flip angle = 9°, T1 = 900 ms, slice
thickness = 1 mm) was used to acquire structural images for each participant. An echoplanar
imaging sequence (TR = 3,000 ms, TE = 30 ms, flip angle = 90°, slice thickness = 3 mm) was used
to measure the blood oxygen level dependent (BOLD) response during functional imaging.
Image processing and analysis were completed using FSL (Jenkinson, Beckmann, Behrens,
Woolrich, & Smith, 2012). Nonbrain voxels were removed and images were LAS-oriented using
FLS tools (bet, fslswapdim, and fslorient). Image preprocessing also included motion-correction using
MCFLIRT, spatial smoothing using a Gaussian kernel (full width at half maximum = 5 mm), and
high-pass filtering (100 Hz). Images were normalized to MNI-152 space. An interleaved slice timing
correction was applied to functional activation data. A general linear model containing regressors for
each event type, which were convolved with a double gamma hemodynamic response function,
was used to analyze each participant’s BOLD response data. For Run1, event types included MV
Ambiguous, MV Unambiguous, RC Ambiguous, RC Unambiguous, comprehension questions,
and baseline. For Run2, event types included RC Ambiguous, RC Unambiguous, comprehension
questions, and baseline. Each of the two sentence event types was modeled separately for the first,
second, third, and fourth occurrence of verbs so as to track changes in activation with RC exposure.
For Run3, regressors included MV Ambiguous Exposed, MV Ambiguous Unexposed, MV
Unambiguous, RC Ambiguous Exposed, RC Ambiguous Unexposed, RC Unambiguous, compre-
hension questions, and baseline. Standard motion parameters were included.
We analyzed activation in two kinds of ROIs. Participant-specific functional ROIs in the left fron-
tal and temporal cortices were extracted from Run1 as follows. The contrast of all sentences (MV
Ambiguous, MV Unambiguous, RC Ambiguous, RC Unambiguous) versus baseline (visual search
of false-font characters) in Run1 was subjected to cluster-level error correction (Z > 3.1, corrected
cluster p < 0.05) to obtain all regions that showed more activation for sentences than baseline. Four
anatomical regions from the Harvard-Oxford atlas (https://neurovault.org/collections/262/; Desikan
et al., 2006) were extracted, three in the left frontal cortex (left pars opercularis = inferior frontal
gyrus, pars opercularis, left hemisphere; left pars triangularis = inferior frontal gyrus, pars triangu-
laris, left hemisphere; and left frontal orbital cortex = frontal orbital cortex, left hemisphere) and one
in the left temporal cortex (left MTG = posterior middle temporal gyrus, left hemisphere). The func-
tional Sentence > Baseline map was masked with each of the four anatomical regions. For the MTG,
we extracted the top 100 voxels showing the most activation for each subject. For the three subre-
gions within the left inferior frontal cortex, to minimize overlap, we extracted the top 30 voxels. Any
remaining overlapping voxels were assigned to the label with the highest probability. This proce-
dure yielded functional ROIs in the left frontal and temporal cortices that showed a Sentence >
Baseline pattern, across all sentence types (hereafter referred to as Sentence > Baseline ROIs.
See Supplementary Figure 1 in the online supporting information).
A second set of 10 group-level functional ROIs from the prior literature that are associated with
cognitive demands and thought to be a part of the MD network were defined as 5 mm spheres
Neurobiology of Language
504
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
4
4
9
2
1
8
6
7
8
1
1
n
o
_
a
_
0
0
0
2
3
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Dynamic updating of native language
around the coordinates reported in Duncan (2010). They included lateral frontal, medial frontal
and lateral parietal regions (left and right inferior frontal sulcus [IFS] AI/FO, left and right rostro-
lateral prefrontal cortex, pre-supplementary motor area, dorsal ACC, left and right IPS; hereafter
referred to as MD ROIs).
Within these regions, we extracted the median activation for each sentence type relative to
baseline for each subject using featquery (FSL). For the Sentence > Baseline ROIs, activation is
expected to be significantly above zero by virtue of how the ROIs were selected. However, the
critical analyses compared activation between different sentence conditions. These comparisons
were orthogonal to the contrast used to identify the ROIs (i.e., the contrast vectors were orthog-
onal [inner product was zero] and the design was balanced; Kriegeskorte, Simmons, Bellgowan,
& Baker, 2009. Further, the key adaptation results are from runs that are distinct from the run used
to define the ROIs. The MD ROIs were defined using coordinates from the previous literature—
activation for sentences might therefore be either greater or less than baseline. In all cases,
repeated measures analyses of variance (ANOVA) were used to determine whether there
was a significant ambiguity effect and whether ambiguity effects differed between conditions.
For Run2, we were interested in whether the RC ambiguity effect decreased with exposure. For
each subject, we computed the difference in activation between the RC Ambiguous and RC
Unambiguous conditions at the first through fourth appearance of the verbs and then used linear
regression to obtain the slope of change over time. We used a one-sample t test to evaluate whether
the mean slope across subjects was less than zero and a binomial test to evaluate whether most
participants showed a negative slope.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
4
4
9
2
1
8
6
7
8
1
1
n
o
_
a
_
0
0
0
2
3
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Correlation between neural activation and cognitive control
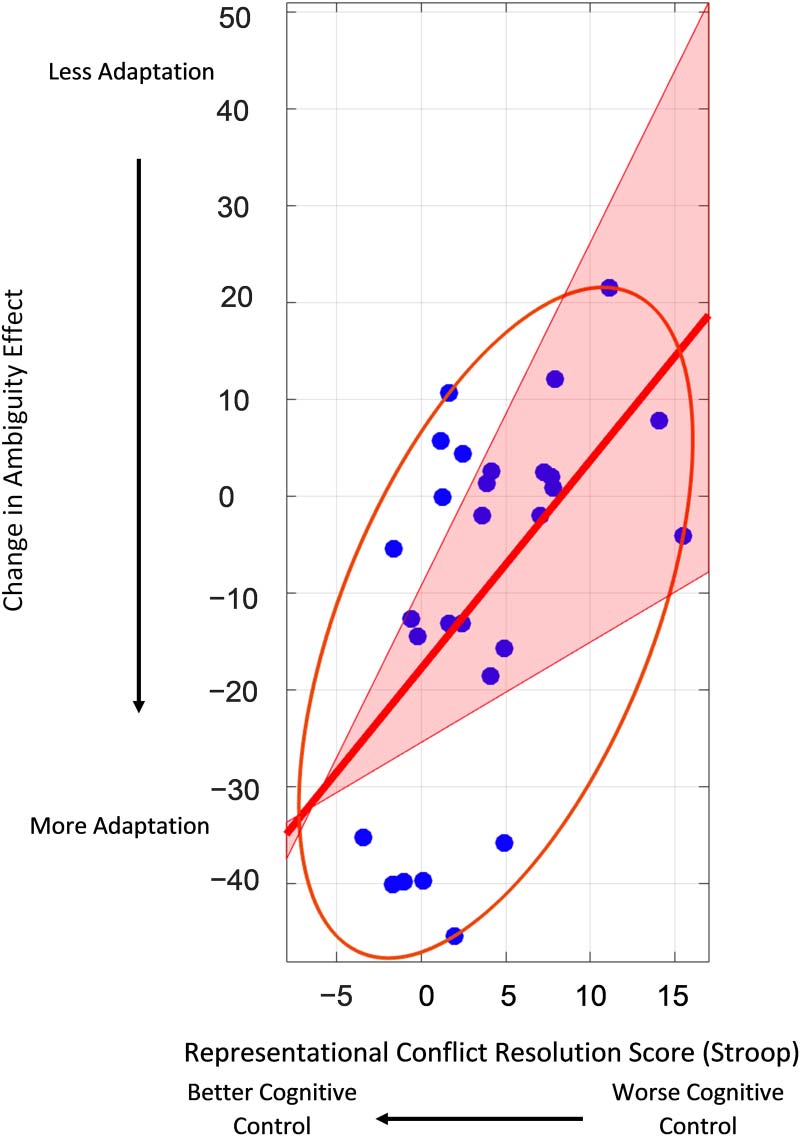
Skipped Pearson correlations (Pernet, Wilcox, & Rousselet, 2013) were used to evaluate the
correlation between each participant’s representational conflict processing score (Stroop) and
the change in their RC ambiguity effect with exposure (slope of change within Run2). A signif-
icant correlation is one for which the 95% confidence interval does not span zero. Our hypoth-
esis predicts a positive correlation. We expected that participants with better conflict resolution
would be more likely to arrive at the correct analysis of RC structures and therefore learn more
from RC exposure. Thus, better conflict processing (lower Stroop scores) is predicted to be
associated with smaller RC ambiguity effects with increasing exposure (lower, more negative
slopes).
It is worth noting that under alternative hypotheses that do not propose a relationship between
cognitive control and syntactic ambiguity resolution and updating, the expected correlation
could be in the opposite direction. For example, participants with poorer conflict resolution could
activate the relevant brain regions more and therefore have a higher likelihood of showing a
larger drop in activation over time. In this case, poorer conflict processing (higher Stroop scores)
would be associated with lower slopes (a negative correlation). Because our hypothesis explicitly
linked cognitive control to correctly encoding new language experiences and updating syntactic
probabilities (see An Integrative Framework and Associated Neural Predictions above), we had
a principled reason to expect a positive rather than a negative (or non-direction-specific)
correlation.
Sentence reading behavioral data
In addition to the neural activation data, we collected and analyzed behavioral data in the form of
comprehension question accuracy and whole-sentence reading times. The former allowed us to
Neurobiology of Language
505
Dynamic updating of native language
ascertain whether participants were reading and processing different parts of the sentences and
thereby verify task compliance.
Whole-sentence reading times were computed from trials where the participants indicated
reading completion and answered any accompanying comprehension question correctly. (Trials
with no completion response and/or incorrect answers were excluded.) Thus, we restricted the
analyses to trials where participants appeared to be following the task instructions. For each
participant and run, reading times from the eligible trials were length-adjusted as follows.
Linear regression was used to generate a predicted reading time for each sentence based on its
length (number of letters). The intercept value was set to 0 (because a sentence with 0 characters
would require 0 reading time). Length-adjusted reading times were computed as: (Actual reading
time) − (Predicted reading time). A positive number indicates that a sentence was read slower
than what would be expected based on length.
We used mixed-effects linear regression (lmer function in R version 1.1.463; Bates, Maechler,
Bolker, & Walker, 2015) to evaluate the ambiguity effect and compare the effect between
conditions. The models contained all applicable fixed effects (e.g., Ambiguity, Structure), random
intercepts, and random slopes. We only simplified the random effects structure when the full
model did not converge (described wherever applicable in the Results section).
To evaluate whether the ambiguity effect changed over the course of Run2 and whether that
change correlated with Stroop performance, we used the same procedures as for the fMRI anal-
yses. However, our predictions for the behavioral data were weaker than for the neural analyses.
The use of whole-sentence rather than word-by-word or segment-by-segment presentation
allowed us to examine relatively naturalistic reading. However, this design necessarily meant that
the reading times were not restricted to the disambiguating regions within a sentence. They likely
indexed the many cognitive operations involved in reading a sentence (Rayner, 1998; Rayner,
Kambe, & Duffy, 2000), including those that are unrelated to conflict. One consequence of this
could be that the lack of conflict-specificity of the reading time measure makes a correlation with
Stroop less likely than in the fMRI analyses. A second consequence could be that the inclusion of
non-disambiguating-region processing times in the reading time (e.g., wrap-up effects at the end
of a sentence) makes evaluation of the change in the ambiguity effect less precise and predictable.
The critical fMRI analyses, by contrast, focused on regions known to be involved in cognitive
control and could therefore prove more sensitive to the component of interest. Accordingly, we
opted a priori to focus on neural predictions and therefore discuss the corresponding results at
greater length below.
RESULTS
fMRI Results
We discuss the main results in the order of the predictions (P.1 to P.4) described above.
P.1: Brain regions showing the garden-path effect in Run1
First, we looked for a pattern of greater ambiguity effect for RC than MV sentences in Run1, which
would indicate sensitivity to conflict between the expected and actual sentence structures. We
discuss results from the subject-specific left inferior frontal and the MTG ROIs first. The left pars
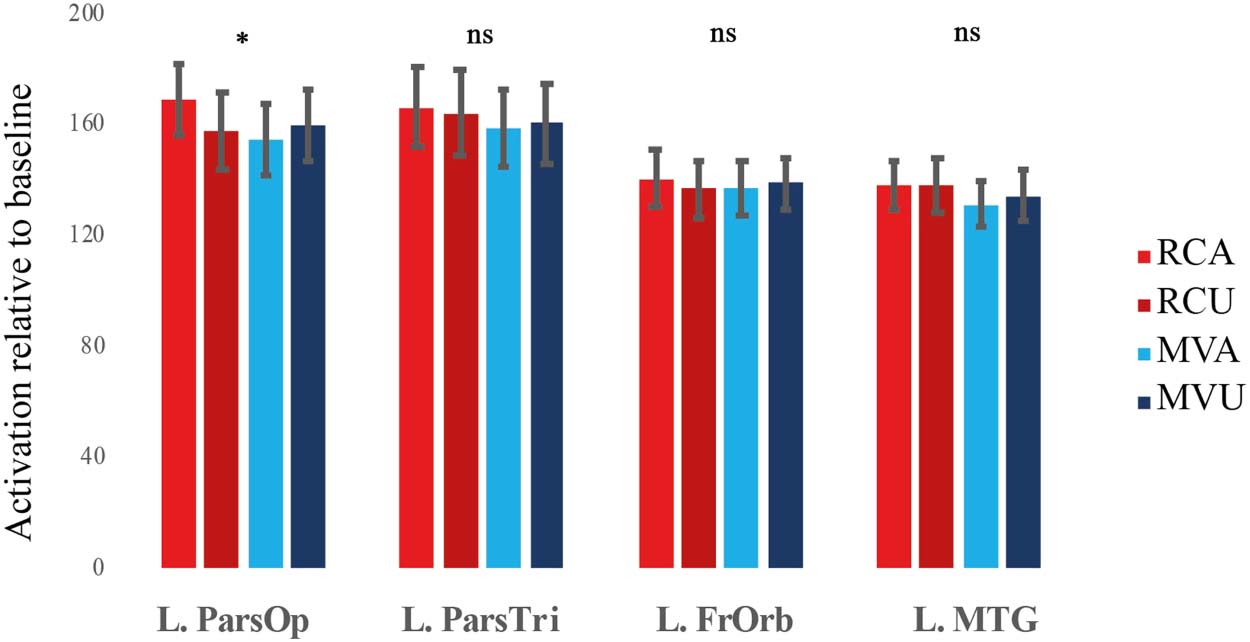
opercularis showed the predicted pattern, namely, a significant interaction between structure and
ambiguity, F(1, 27) = 10.72, p < 0.0125, Bonferroni-corrected for analysis of 4 ROIs. This inter-
action was due to a significant effect of ambiguity for RC, RC Ambiguous > RC Unambiguous;
Neurobiology of Language
506
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
4
4
9
2
1
8
6
7
8
1
1
n
o
_
a
_
0
0
0
2
3
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Dynamic updating of native language
F(1, 27) = 10.72, p < 0.05, but not MV, F(1, 27) = 2.73, p > 0.05, structures. The other frontal
regions and the MTG did not show such a conflict effect (Structure × Ambiguity interaction for the
left pars triangularis: F(1, 27) = 0.51, p > 0.05; the left frontal orbital cortex: F(1, 27) = 1, p > 0.05;
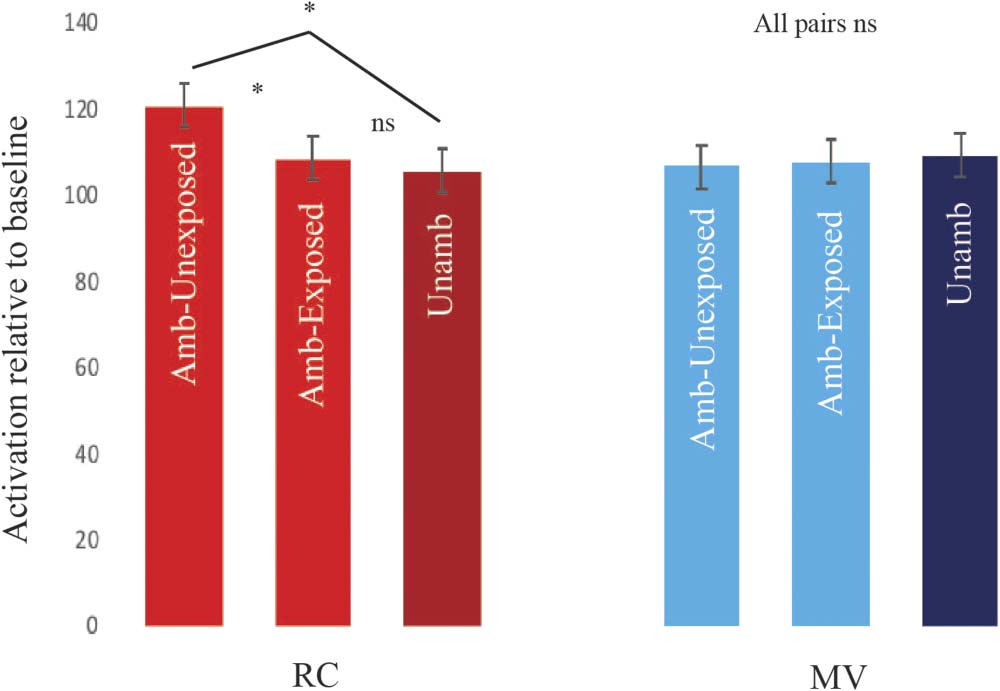
and the left MTG: F(1, 27) = 0.92, p > 0.05). Figure 2 shows the pattern in the left pars opercularis
and the other three regions for comparison. The three-way Structure × Ambiguity × Region inter-
action was significant, Huynh-Feldt corrected F(2.59, 69.92) = 3.22, p < 0.05, suggesting that the
pars opercularis subregion within the left inferior frontal cortex might be especially relevant. Note
that analysis of the top 30 voxels in the MTG (paralleling the number of voxels in the frontal ROIs)
yielded the same pattern of results. There was no significant Structure × Ambiguity effect in the
MTG, F(1, 27) = 0.36, p > 0.05), and there was a significant three-way Structure × Ambiguity × Region
interaction in the pooled analysis of all four ROIs, Huynh-Feldt corrected F(2.77, 74.79) = 3.24,
p < 0.05.
Overall, the results from Run1 conformed to our predictions in showing a garden-path effect in
the frontal (left pars opercularis) but not the temporal regions (left MTG). Whole-brain analysis of
the interaction between Ambiguity and Structure did not reveal any suprathreshold clusters
(cluster p < 0.05, z > 3.1). Uncorrected results (z > 3.1) showed activation in the left lateral
and medial frontal cortices only (Supplementary Figure 2 in the online supporting information),
consistent with our prediction that frontal regions might be the most relevant. Not all frontal
regions showed an effect, however, a point we turn to in the Discussion section. All subsequent
analyses were restricted to the regions showing an effect.
P.2: Change in the RC ambiguity effect over Run2
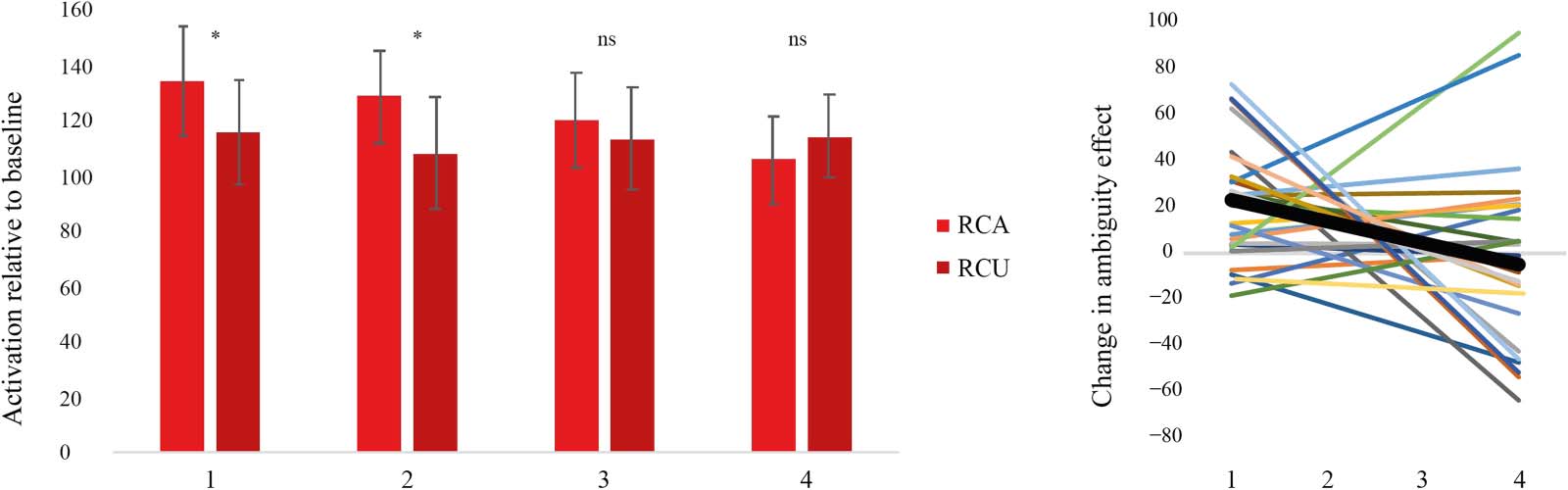
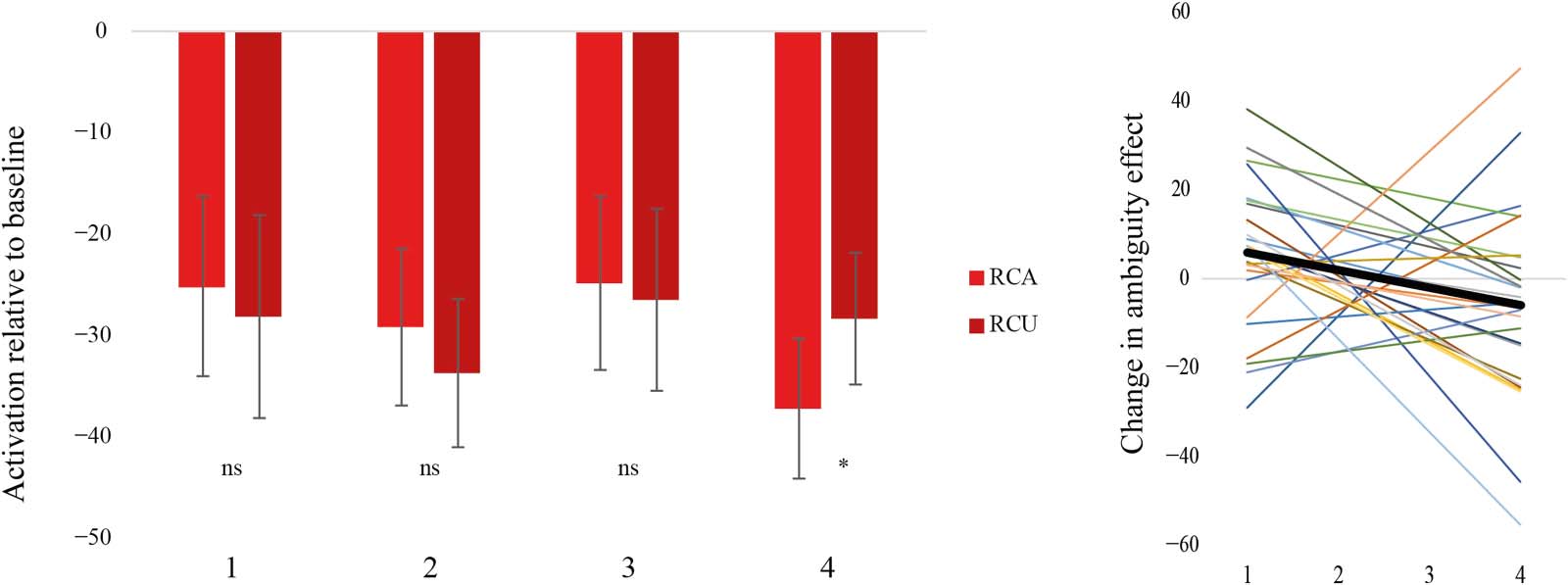
In Run2, we predicted that the RC ambiguity effect would decrease with exposure to RC sentences,
resulting in a negative slope. Within the left pars opercularis, the RC ambiguity effect was signif-
icant at the first, F(1, 27) = 12.06, p < 0.05, and second, F(1, 27) = 12.42, p < 0.05, but not the third
occurrence, F(1, 27) = 0.72, p > .05, of the verbs. By the fourth occurrence, RC Ambiguous
sentences showed numerically lower activation than RC Unambiguous sentences, but this effect
was not significant, F(1, 27) = 1.59, p > 0.05 (Figure 3 left panel). To specifically evaluate whether
Figure 2. Activation for different sentence types relative to baseline in Run1 within Sentence >
Baseline regions of interest. Only the left pars opercularis showed an interaction corresponding to a
conflict effect. Here and elsewhere, error bars denote Cousineau-Morey within-subject standard errors
(Rmisc package; Morey, 2008). L = left, ParsOp = pars opercularis, ParsTri = pars triangularis, FrOrb =
frontal orbital cortex, MTG = middle temporal gyrus, RCA = relative clause ambiguous, RCU = relative
clause unambiguous, MVA = main verb ambiguous, MVA = main verb unambiguous.
Neurobiology of Language
507
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
4
4
9
2
1
8
6
7
8
1
1
n
o
_
a
_
0
0
0
2
3
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Dynamic updating of native language
Left: Activation for relative clause ambiguous (RCA) and unambiguous (RCU) structures within the left pars opercularis in Run2.
Figure 3.
There was an ambiguity effect at the first and second but not at the third and fourth occurrence of verbs. Right: Slope of change over time for
each participant. Bold line indicates mean slope.
the RC ambiguity effect decreased with exposure, we performed a linear regression of RC
Ambiguous minus RC Unambiguous over four time points and extracted the slope for each
participant (Figure 3 right panel). The mean slope was −9.45, t(27) = −2.73, p < 0.05.
Seventeen out of twenty-eight participants had a negative slope (Binomial p > 0.05), suggesting
some variability (see also section P.3 below).
P.3: The relationship between change over Run2 and cognitive control
We computed Skipped Pearson correlations to evaluate the relationship between the decrease in
the ambiguity effect over Run2 and individual differences in cognitive control (Stroop scores). In
the left pars opercularis, we observed a significant positive correlation as predicted (Skipped
Pearson r = 0.544779, CI = [0.298778, 0.739922]). Those who had lower Stroop scores (indi-
cating better cognitive control) were more likely to show a decreasing ambiguity effect with
exposure, consistent with our hypothesis (Figure 4).
The association observed above was computed using a robust correlation method that eval-
uates and accounts for bivariate outliers (Pernet et al., 2013). In addition, we evaluated the reli-
ability of the measurements. For the Stroop conflict score, odd–even split-half reliability
(calculated using the Spearman–Brown prophesy formula) was 0.71, suggesting that the measure-
ment was reliable. For the decrease in ambiguity with exposure, we supplemented the original
analysis of the slope by calculating difference scores separately using the first and third appear-
ances (third minus first) and the second and fourth appearances (fourth minus second). The two
difference scores computed from different subsets of trials were significantly correlated with one
another (Skipped Pearson r = 0.605445, CI = [0.227441, 0.803849]). Each difference score was
also correlated with the Stroop conflict score, consistent with the original analysis (third minus
first: r = 0.492034, CI = [0.243332, 0.733144]; fourth minus second: r = 0.527813, CI =
[0.257456, 0.711858]).
P.4: Garden-path effects for exposed and unexposed verbs in Run3
In Run3, we asked whether the effect of exposure in the left pars opercularis persisted from Run2
and if so, whether it generalized to verbs other than those that appeared during the exposure
phase. For exposed verbs, there was no significant difference between the RC Ambiguous and
RC Unambiguous conditions and no interaction between ambiguity and structure ( p’s > 0.05). In
contrast, unexposed verbs showed a significant RC ambiguity effect, F(1, 27 = 16.88, p < 0.05,
Neurobiology of Language
508
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
4
4
9
2
1
8
6
7
8
1
1
n
o
_
a
_
0
0
0
2
3
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Dynamic updating of native language
Figure 4. Correlation between individuals’ representational conflict processing score (Stroop) and
adaptation of the relative clause ambiguity effect in the left pars opercularis in Run2. Individuals with better
cognitive control demonstrated greater adaptation. The ellipse represents the bounding area for included
versus excluded bivariate data points. The shaded region represents the 95% CI for the correlation.
and an interaction between ambiguity and structure, F(1, 27) = 13.73, p < 0.05. Direct compar-
ison between RC Ambiguous Unexposed and RC Ambiguous Exposed revealed a significant
difference, F(1, 27) = 9.09, p < 0.05. In sum, the left pars opercularis showed persistent effects
of RC exposure but only for verbs seen repeatedly in that structure (Figure 5).
Figure 5. Activation for different sentences relative to baseline in Run3 within the left pars opercu-
laris. Exposed verbs did not show an RC ambiguity effect. Unexposed verbs showed a significant RC
ambiguity effect and a difference from exposed verbs. RC = relative clause, MV = main verb, amb =
ambiguous, unamb = unambiguous.
Neurobiology of Language
509
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
4
4
9
2
1
8
6
7
8
1
1
n
o
_
a
_
0
0
0
2
3
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Dynamic updating of native language
Results for the Multiple Demand Network
We conducted parallel analyses in the MD network as for P.1 to P.4 above. Among the 10 MD
ROIs, only the left AI/FO showed a significant interaction between structure and ambiguity in
Run1, F(1, 27) = 5.48, p < 0.05. This interaction was due to a significant effect of ambiguity
for RC, RC Ambiguous > RC Unambiguous; F(1, 27) = 7.44, p < 0.05, but not MV, F(1, 27) =
0.32, p > 0.05, structures. No other MD region showed this conflict effect. (Please see the
Supplementary Information in the online supporting information for details.) Figure 6 shows
the pattern in the left AI/FO and for comparison, the patterns in two other frontal regions (left
IFS, ACC) and a parietal region (left IPS).
In Run2, within the left AI/FO, there was no ambiguity effect at the first, F(1, 27) = 0.51, p >
0.05, second, F(1, 27) = 1.57, p > 0.05, and third, F(1, 27) = 0.11, p > 0.05, occurrence of verbs.
By the fourth occurrence, there was a reverse ambiguity effect such that RC Ambiguous sentences
showed lower activation than RC Unambiguous sentences, F(1, 27) = 4.59, p < 0.05 (Figure 7 left
panel). The mean slope was −3.83, t(27) = −2.04, p = 0.05. Twenty out of twenty-eight partici-
pants had a negative slope (Binomial p < 0.05; Figure 7 right panel). However, in contrast to the
left pars opercularis, there was no correlation between the change in activation in the left AI/FO
and Stroop performance (Skipped Pearson r = 0.103193, CI = [−0.331871, 0.516918]).
In Run3, analyses of exposed and unexposed verbs revealed no RC ambiguity effect or inter-
action between ambiguity and structure for either verb type. The two conditions also did not differ
from one another (all p’s > 0.05). Thus, unlike the left pars opercularis, the left AI/FO showed a
generalized effect of exposure, which extended to verbs not seen repeatedly in the RC structure
(Figure 8).
Behavioral Results
Accuracy on the comprehension questions was significantly above chance (50%) in all three
runs, indicating that participants paid attention to the task and encoded the content of the sen-
tences (Run1: 81.3%, Run2: 77.9%, Run3: 73.6%. One-sample t test all p’s < 0.05). In Run1, for
the length-adjusted RT analyses, the mixed model contained the fixed effects of Ambiguity,
Structure, and their interaction along with random intercepts by participant and verb, and random
Figure 6. Activation for different sentence types relative to baseline in Run1 within the multiple-
demand regions of interest. Only the left AI/FO showed an interaction corresponding to a conflict
effect. AI/FO = anterior insula/frontal operculum, IFS = inferior frontal sulcus, ACC = anterior cin-
gulate cortex, IPS = intraparietal sulcus, RCA = relative clause ambiguous, RCU = relative clause
unambiguous, MVA = main verb ambiguous, MVA = main verb unambiguous.
Neurobiology of Language
510
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
4
4
9
2
1
8
6
7
8
1
1
n
o
_
a
_
0
0
0
2
3
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Dynamic updating of native language
Figure 7. Left: Activation for relative clause ambiguous (RCA) and unambiguous (RCU) structures within left AI/FO in Run2. There was no
ambiguity effect at the first to third occurrences and a reverse ambiguity effect at the fourth occurrence of verbs. Right: Slope of change over
time for each participant. Bold line indicates mean slope. AI/FO = anterior insula/frontal operculum.
slopes of ambiguity and structure by participant. The results revealed a significant main effect
of Structure ((cid:1) = −123.59, p < 0.05) and a significant interaction between Ambiguity and
Structure ((cid:1) = 292.55, p < 0.05). The interaction was due to a significant ambiguity effect for
RC (Unambiguous M = −99.11 ms, Ambiguous M = 166.01 ms; (cid:1) = 263.31, p < 0.05) but not
MV sentences (Unambiguous M = 27.04 ms, Ambiguous M = 0.49 ms; (cid:1) = −26.12, p > 0.05). This
is consistent with garden-pathing, but strong conclusions might be precluded by the coarse-
grained nature of the measure (see more in the Discussion section).
In Run2, we found a significant ambiguity effect at the first (Unambiguous M = 13.47 ms,
Ambiguous M = 174.65 ms; (cid:1) = 161.08, p < 0.05), second (Unambiguous M = −47.19 ms,
Ambiguous M = 166.11 ms; (cid:1) = 221.33, p < 0.05), and fourth occurrence (Unambiguous M =
−136.94 ms, Ambiguous M = 12.31 ms; (cid:1) = 151.04, p < 0.05) of the verbs. There was no effect at
the third occurrence (Unambiguous M = −68.90 ms, Ambiguous M = 15.73 ms; (cid:1) = 87.46, p >
0.05). The mean slope across participants was −17.74, t(27) = −0.92, p > 0.05. Half the partic-
ipants showed a negative slope and the other half a positive slope, indicating no consistent
Figure 8. Activation for different sentences relative to baseline in Run3 within the left anterior
insula/frontal operculum. Neither exposed nor unexposed verbs showed an RC ambiguity effect.
RC = relative clause, MV = main verb, amb = ambiguous, unamb = unambiguous.
Neurobiology of Language
511
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
4
4
9
2
1
8
6
7
8
1
1
n
o
_
a
_
0
0
0
2
3
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Dynamic updating of native language
pattern. Thus, unlike neural activation in the left pars opercularis and the left AI/FO, behavioral
analysis of whole-sentence reading times did not reveal a consistent decrease in the ambiguity
effect over RC exposure. The slope of change for the behavioral scores did not correlate with
the slope for neural activation within the left pars opercularis (Skipped Pearson r = 0.0586045,
CI = [−0.25037, 0.384576]) or the left AI/ FO (Skipped Pearson r = −0.0229688, CI =
[−0.492274, 0.445291]).
In Run3, for exposed verbs, the analysis revealed no main effects and no interaction between
ambiguity and structure (RC Unambiguous M = −45.78 ms, RC Ambiguous M = 60.49 ms, MV
Unambiguous M = 18.48 ms, MV Ambiguous M = 12.35 ms; p’s > 0.05). In contrast, for unex-
posed verbs, there was a significant interaction ((cid:1) = 151.70, p < 0.05), which arose from a sig-
nificant ambiguity effect for RC (Unambiguous M = −45.78 ms, Ambiguous M = 129.31 ms; (cid:1) =
175.77, p < 0.05) but not MV (Unambiguous M = 18.48 ms, Ambiguous M = 42.12 ms; (cid:1) = 22.63,
p > 0.05). Direct comparison between RC Ambiguous Unexposed and RC Ambiguous Exposed
did not reveal a significant difference ( p > 0.05).
DISCUSSION
Adults adapt to new input in their native language. To our knowledge, the present study is among
the first to investigate the neural mechanisms underlying this adjustment. We exposed partici-
pants to a relatively infrequent structure in English that involves RCs and asked if and how their
brains adapted to the exposure. Prior to the experimentally manipulated exposure phase, two
frontal brain regions—the left pars opercularis and the left AI/FO—showed sensitivity to conflict
during sentence processing, namely increased activation for processing ambiguous sentences
that resolved to the more infrequent RC structure. During the exposure phase, both regions
showed decreasing RC-ambiguity-related activation, suggesting that they were adapting in some
way to the current language environment. Following the exposure phase, the adjustment per-
sisted for verbs that had repeatedly appeared in RC structures. These results conformed to our
prediction that frontal brain regions would be relevant for adjusting to a novel language environ-
ment, consistent with their involvement more broadly in adaptive behavior.
Beyond this general conclusion, the study revealed three contrastive patterns between the two
regions that are informative for theorizing about the nature of syntactic updating and the under-
lying neural substrates: (1) The left pars opercularis ROI, selected using this functional criterion in
the first run, was consistently activated for processing sentences compared to baseline in all runs.
In contrast, the left AI/FO ROI, which is a part of the MD network, consistently showed the reverse
(Baseline > Sentence) pattern. (2) Individual differences in a cognitive control task (Stroop) cor-
related with adaptation of neural activation with RC exposure within the left pars opercularis but
not the left AI/FO. (3) The adaptation effect within the left pars opercularis was verb-specific,
showing a difference in adjustment between verbs that had repeatedly appeared in RC sentences
during exposure and those that had not. In contrast, the left AI/FO showed broader adaptation and
no difference between exposed and unexposed verbs.
The findings for the left pars opercularis are consistent with our proposal that regions within the
left inferior frontal cortex could support syntactic updating via their role in handling conflict
between incompatible representations. This region showed the expected conflict effect prior to
exposure, adaptation during the exposure phase, correlation of adaptation with individual differ-
ences in Stroop, and a theoretically meaningful relationship between exposure and adaptation in
the form of verb-specificity. By comparison, the results for the left AI/FO suggested involvement in
adjusting to a novel environment, but this role appeared to be broad and not specifically related to
sentence or conflict processing. This region showed greater activation for a nonsentence task than
Neurobiology of Language
512
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
4
4
9
2
1
8
6
7
8
1
1
n
o
_
a
_
0
0
0
2
3
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Dynamic updating of native language
for sentence reading, no correlation between adaptation and Stroop, and broad adjustment rather
than tuning to relevant statistical properties of the language input. Thus, although both regions
appear to play a role in adjusting to new language environments, their functions could be impor-
tantly different. Before we expand on this topic, we first consider alternative explanations for the
observed effects.
Alternative Explanations
One alternative explanation suggested in the behavioral literature is that adaptation could be
happening at the level of generic task practice rather than at the level of updating the likelihood
of particular sentence structures. This practice effect could be stronger for the more difficult,
ambiguous sentences leading to the observed decrease in the difference between the ambiguous
and unambiguous conditions (Prasad & Linzen, 2019; Stack et al., 2018). Neurally, increased
facility with the task could correspond to decreased difficulty and, correspondingly, decreased
activation in regions that are associated with general cognitive demands. In the context of the
current study, this explanation could potentially explain the findings from the left AI/FO but
not the left pars opercularis. In the former case, decreased recruitment as the experiment
progressed was generic and not restricted to the verbs that were exposed in the RC structures.
Thus, it is possible—maybe even likely given the profile of the MD network—that this region
broadly supported adjustment to task demands within the context of the experiment. In the left
pars opercularis, however, adjustment over the course of the experiment was restricted to those
verbs that were seen repeatedly in the RC structures. Thus, this region’s engagement depended on
how often individual verbs appeared in a particular structure—a known signature of the sentence
processing system. In sum, we suggest that a task-level effect accords well with prior literature on
the MD network and the observed results in the left AI/ FO but that the left pars opercularis
performs a different role (see more below).
Could participants have shifted to a shallower or less attentive (e.g., good-enough) mode of
processing for the critical RC Ambiguous sentences in Run3 compared to Run1, which could
potentially explain the pattern of lowered ambiguity-related activation as the experiment
progressed? Two aspects of the results suggest that this cannot explain the critical findings within
the left pars opercularis. First, we found verb-specific adaptation in the neural activation for RC
ambiguity in Run3. A general shift in processing mode would be expected to affect all rather than
a hypothesized subset of the RC Ambiguous sentences. Second, positive correlation between the
decrease in the ambiguity effect and Stroop scores in Run2 follows parsimoniously from the
proposed hypothesis about cognitive control and syntactic updating (which in turn was inspired
by much prior evidence that cognitive control plays a role in resolving syntactic conflict). In
contrast, an explanation based on processing mode would be ad hoc—it is unclear why individ-
uals with better cognitive control should be more likely to shift to good-enough processing over
time compared to those with poorer cognitive control. In sum, while we leave open the question
of potential shifting between processing modes to future studies with targeted behavioral mea-
sures, the activation patterns found in the left pars opercularis are more consistent with the pro-
posed hypothesis. Note that the behavioral measures employed in the present study were mainly
intended to verify task compliance and cannot be used to answer this question. Comprehension
questions did not focus on the ambiguity and therefore cannot help establish whether participants
processed the ambiguity shallowly or deeply. The questions were also not matched or counter-
balanced in any specific way (e.g., difficulty) between the two runs. More broadly, whether off-
line comprehension accuracy can be used to infer the depth of processing of garden-path
sentences is not clear (Qian, Garnsey, & Christianson, 2018). Reaction times are expected to
get faster whether due to task-level practice, increased experience with a syntactic structure,
Neurobiology of Language
513
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
4
4
9
2
1
8
6
7
8
1
1
n
o
_
a
_
0
0
0
2
3
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Dynamic updating of native language
or a shift in processing mode. These factors could also create asymmetric rather than equal effects
in different sentence conditions. Targeted design and analyses are therefore needed to tease apart
the different factors and interpret the behavioral measures appropriately.
Could the decreasing RC ambiguity effect in the left pars opercularis be a consequence of
short-term syntactic priming? We think this is unlikely for two reasons. First, we show that the
diminished RC ambiguity effect for the exposed verbs persisted from the end of Run2 going into
Run3. This gap of several minutes is not consistent with purely short-term priming. Second, grow-
ing consensus within the syntactic priming literature eschews dichotomy between priming and
longer-term adjustment. Because priming lasts across multiple intervening trials and shows sig-
natures of implicit learning (Bock & Griffin, 2000; Fine & Jaeger, 2013; Wells et al., 2009), the
dominant current view is that (long-term) syntactic priming is syntactic updating. The present
findings are consistent with results that have implicated the left inferior frontal cortex in adapta-
tion within a priming context (Segaert, Kempen, et al., 2013; Segaert, Menenti, et al., 2011).
Future studies comparing neural adaptation at different time lags following exposure could be
useful for additionally clarifying the underlying mechanism(s).
The Role of the Left Pars Opercularis
Some researchers have suggested that the left pars opercularis (and Broca’s area more broadly) is
responsible for specific operations (e.g., movement) that are associated with syntactic complexity
(e.g., Ben-Shachar et al., 2003). The current results do not fit within this perspective because (a)
RC Unambiguous sentences are just as complex as RC Ambiguous sentences, but we detected
higher activation for the former in Run1; (b) the complexity of these sentence types and their
associated operations do not change with exposure, but activation with the left pars opercularis
did; and (c) the operations do not differ based on the verb, but the observed results showed a
difference between exposed and unexposed verbs.
Prior evidence on the left inferior frontal cortex and syntactic ambiguity and conflict processing
are more directly relevant to the present findings (January et al., 2009; Mason et al., 2003; Rodd
et al., 2010). We extend previous results by showing that the involvement of the left pars opercu-
laris for conflict-inducing sentences adapts with additional exposure. Thus, this region is engaged
in updating syntactic expectations in individuals’ native language. The observed patterns are
broadly consistent with our description of this region’s role in terms of cognitive control. In line
with current thinking on how cognitive control operates, we would suggest that the left pars oper-
cularis is involved in biasing the activation of and resolving the conflict between linguistic repre-
sentations that reside elsewhere in posterior cortical regions (January et al., 2009; Novick et al.,
2005; Thothathiri et al., 2012; Ye & Zhou, 2009). In the context of the present study, this would
mean adjusting the activation levels of RC and MV structures such that the less frequent RC
representation wins out over the more prepotent MV representation when applicable. Such a
function would be most needed when the parser encounters ambiguous sentences that resolve
towards the less frequent RC structure, with that need diminishing upon increasing exposure to
that structure. Below we discuss nuanced questions within this broad perspective.
One question pertains to the domain-specificity or generality of this region’s function. On one
end of the spectrum, some have proposed that this region could be engaged in domain-general
cognitive control across linguistic and nonlinguistic tasks (e.g., Hsu et al., 2017; January et al.,
2009). Others have suggested that different subregions within the left frontal cortex could be spe-
cialized for the controlled processing of different types of representations (e.g., Hagoort, 2016).
The present study did not test a variety of linguistic and nonlinguistic tasks in the scanner and
cannot address this question directly. Correlation between adaptation within the left pars
Neurobiology of Language
514
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
4
4
9
2
1
8
6
7
8
1
1
n
o
_
a
_
0
0
0
2
3
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Dynamic updating of native language
opercularis and individuals’ behavioral performance on Stroop (a nonsyntactic task) suggests
that this region’s role is not specific to syntax. This is consistent with prior evidence for the co-
localization of sentential and nonsentential tasks within the left frontal cortex (January et al.,
2009; van de Meerendonk, Rueschemeyer, & Kolk, 2013; Ye & Zhou, 2009). Such an interpre-
tation leaves open the possibility that this region is engaged for language-specific rather than
syntax-specific representations (because the Stroop task involved linguistic content). However,
because adjacent regions within the frontal cortex can have different processing profiles
(Fedorenko et al., 2013; Hagoort, 2016), we cannot definitively rule out specialization for syntax.
We observed the expected patterns in the left pars opercularis but not the adjacent pars triangu-
laris or the frontal orbital cortex ROIs, which is potentially consistent with the suggestion that the
posterior parts of the left inferior frontal cortex might be specialized for the controlled syntactic
processing (Hagoort, 2016). For the present, we suggest that this region could be involved in
either syntactic updating or broader language updating.
A different question pertains to the processing stage at which the left pars opercularis is
involved in updating. This region could help resolve conflict between competing representations
thereby influencing the updating process (as we have suggested). Alternatively, activation within
this region could provide a read-out of prediction error, which could serve as input for updating
that recruits some other brain region. Both interpretations are consistent with the observed
results—both prediction error and the need for conflict resolution would be high for RC
Ambiguous sentences in Run1 because this structure is less expected, both would decrease over
time as the parser updates the probability of RC structures, and both could depend on character-
istics that affect syntactic expectations (e.g., verb-structure links). Prior literature on cognitive con-
trol suggests that the lateral prefrontal cortex broadly is involved in conflict resolution. In contexts
that do not involve updating or prediction error (e.g., Stroop), higher activation within these
regions is associated with improved behavioral performance on a given trial (Kerns, 2006;
Kerns et al., 2004). However, definitive evidence might require causal paradigms with sufficient
temporal resolution to determine if and how updating is disrupted without a functioning left
pars opercularis.
The Role of the Left Anterior Insula/Frontal Operculum
Evidence from resting state functional connectivity studies suggests that the left AI/FO is part of a
cingulo-opercular network that is involved in task set maintenance and attentional modulation
(Dosenbach, Fair, et al., 2007). This region is recruited at the beginning of a given task block and
remains active for the duration of a task, suggesting that it is involved in initiating and maintaining
a task set. Critically, these activation patterns are found across multiple domains, providing
evidence for a domain-general function in goal-directed behavior (Dosenbach, Visscher, et al.,
2006). The left AI/FO is part of the MD network, which shows increased recruitment for difficult
tasks and the representation of task-relevant over irrelevant information (Woolgar, Williams, &
Rich, 2015). Together, this evidence has led researchers to propose that this region could provide
domain-general task-level and attentional support to the processing of domain-specific informa-
tion that occurs in other more specialized regions (Dosenbach, Visscher, et al., 2006; Woolgar
et al., 2015).
The results from the present study are consistent with prior evidence and interpretation. In
particular, the left AI/FO (along with other MD regions) showed greater activation during the
baseline task than during sentence reading across all three runs. The former task involved false
fonts and was more novel to participants than the latter (reading being a familiar activity to our
literate participants). During practice prior to scanning, participants had a mean accuracy of
Neurobiology of Language
515
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
4
4
9
2
1
8
6
7
8
1
1
n
o
_
a
_
0
0
0
2
3
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Dynamic updating of native language
65.7% and 97.7% on the baseline and sentence reading tasks, respectively. This suggests that the
baseline task was more difficult in some respects. Engagement of the AI/FO could have supported
the maintenance of task sets, especially in the context of different interwoven tasks like in the
present study. As reviewed above, this function is expected to be sensitive to task difficulty (ex-
plaining greater recruitment for the baseline over the sentence task, and for harder over easier
sentences) and domain-general. The adaptation of left AI/FO activation in Run2 and the persis-
tence of this adaptation for both unexposed and exposed verbs in Run3 is consistent with such a
task-set maintenance function. As the experiment progressed and different tasks and sentence
types became more familiar, this region might have become less and less needed.
Why did we see sensitivity to sentence processing difficulty within the left AI/FO and no
other frontal MD regions? It is worth reiterating here that we used MD network ROIs based on
group-level coordinates from the prior literature. This could have potentially obscured patterns
that might be better detected using methods (e.g., subject-specific localizers) that take variabil-
ity between individuals into account. Given this caveat, we can only speculate post hoc about
the observed pattern. It is possible that frontal MD regions (left and right IFS) that are a part of a
frontoparietal network that usually shows short-lived activity in response to a cue (cf. sustained
activity in the cingulo-opercular network; Dosenbach, et al., 2007) were not as relevant in the
task contexts of the present study. Alternatively, sustained activation differences between hard-
er versus easier tasks might have been easier to detect than short-lived activation differences.
More challenging to explain is why other regions within the cingulo-opercular network—
namely the right AI/FO and the ACC—did not show sensitivity to sentence processing difficulty
like the left AI/FO. For the former, fine-grained functional specialization within AI/FO regions is
still an open question (Ullsperger, Harsay, Wessel & Ridderinkhof, 2010). With respect to the
ACC, this region has been linked to response rather than representational conflict and might be
more likely to be engaged in tasks requiring overt responses (Milham et al., 2001; Thothathiri,
2018) unlike sentence comprehension. Alternatively, the ACC might be especially important for
comprehension in a social pragmatic context (Hagoort, 2016). Future studies are needed to tease
apart the roles of different cingulo-opercular regions within the MD network during sentence
comprehension. Broadly, all of these roles are expected to be domain-general and sensitive
to task difficulty.
Broader Implications and Future Directions
While the current study addresses syntactic updating, processing ambiguity and updating expec-
tations also applies at other linguistic levels, including lexical semantics. Models of how the
parser addresses lexical-semantic ambiguity mirror models of syntactic processing. Spe-
cifically, multiple possible meanings for a given word are activated simultaneously and these
meanings have an associated probability based on frequency of use. The most frequent meaning
is typically preferred (e.g., Twilley & Dixon, 2000). However, when semantically ambiguous
words are repeatedly encountered in contexts that support the less frequent meaning, individuals
are more likely to activate the less likely meaning in later probes (Gilbert, Davis, Gaskell, & Rodd,
2018; Rodd et al., 2016; Rodd, Cutrin, Kirsch, Millar, & Davis, 2013). Synonyms for the ambig-
uous word do not show this effect, indicating updating of form–meaning mappings rather than
semantic priming (Rodd et al., 2013). The effects build up over time, persist for 20 min or more,
and are sensitive to surprisal (Gilbert et al., 2018; Rodd et al., 2013). These parallels to syntactic
processing suggest that the current hypothesis might also apply to lexical-semantic updating.
Namely, cognitive control could help disambiguate the meaning of ambiguous words and there-
by assist in updating the likelihood of less frequently encountered meanings. Regions within the
left inferior frontal cortex and the MD network could play a role in such adaptation, possibly with
Neurobiology of Language
516
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
4
4
9
2
1
8
6
7
8
1
1
n
o
_
a
_
0
0
0
2
3
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Dynamic updating of native language
adjacent subregions showing specialization for different linguistic processes and/or different pat-
terns of connectivity with posterior cortices. Thus, comparing different kinds of updating within
language could be a fruitful future direction for testing the hypothesis proposed here in a closely
related domain and for clarifying the networks that support different aspects of language.
The results of the present study are from written comprehension. Previous evidence suggests
that activation in frontotemporal language regions during sentence comprehension is similar
between the visual and auditory modalities (Devauchelle et al., 2009; Jobard, Vigneau,
Mazoyer, & Tzourio-Mazoyer, 2007). Further, P600 signals in response to syntactic expectation
violations are also similar between the two modalities (Hagoort & Brown, 2000). Therefore, we
expect the conclusions from the current study to apply broadly to sentence comprehension
(written or spoken). However, spoken language contains additional information, especially pro-
sodic cues, which could guide online syntactic expectations. Future studies could investigate
whether the presence or absence of disambiguating prosodic cues alters the need for cognitive
control and the recruitment of different brain regions during syntactic updating.
Previous behavioral studies have shown that adults can adjust their syntactic expectations
based on the particular speaker (Kamide, 2012; Kroczek & Gunter, 2017). One interesting exten-
sion of the current paradigm might be to investigate the neural substrates that support switching
between different sets of syntactic expectations for different speakers. A priori, control regions that
are known to support task-switching or set-shifting might be expected to play a role in such sit-
uations (Dove, Pollmann, Schubert, Wiggins, & Von Cramon, 2000; Swainson et al., 2003; Ruge,
Jamadar, Zimmermann, & Karayanidis, 2013). Further, Kroczek and Gunter (2017) showed that
participants maintained some information about speaker-specific syntactic probabilities over
the course of months. However, this was not immediately deployed (say from memory) but
re-established quickly with limited renewed exposure to the speakers. The cues that trigger quick
adjustment and the neural mechanisms that support it and any potential transfer of learning from
one context to another are open questions for future investigation.
The results from the present study could be strengthened in a number of ways. First, we ex-
amined regions linked to cognitive control because we wanted to test a particular process-
specific hypothesis. We would not claim, however, that those are the only regions relevant to
updating. In this context, it is worth discussing the null conflict effect in Run1 for the MTG, which
appears to stand in contrast to previous findings of syntactic and lexical ambiguity effects in the
posterior temporal cortex (e.g., Mason et al., 2003; Rodd, Longe, et al., 2010). The divergent out-
comes could potentially be related to methodological differences. For example, Mason et al.
(2003) examined a temporal ROI that was defined anatomically and included posterior, superior,
and middle temporal areas whereas the current study examined a functionally defined MTG re-
gion. Rodd, Longe, et al. (2010) investigated a different kind of syntactic ambiguity than was stud-
ied here. Regardless, like many other researchers, we would suggest that left frontal and temporal
areas work in close concert during sentence processing. The lack of an effect in the left MTG (in
contrast to the left pars opercularis) could have resulted from lack of power due to analyzing a
single run and/or the type and strength of the conflict manipulation. It is possible that alternative
paradigms with more sensitivity will detect effects in both left frontal and temporal areas. Further
research is needed to fully establish whether these regions play related but distinct roles (e.g., see
the discussion in Mason et al., 2003 and Rodd, Longe, et al., 2010) during syntactic adaptation.
Future studies could also explore the neural substrates related to other components such as the
encoding of syntactic probabilities and the connectivity between different brain regions that fa-
cilitates updating as a whole.
Second, our Run3 results showed no RC ambiguity effect for verbs that were frequently expe-
rienced in those structures. For the left pars opercularis, this pattern was verb-specific and did not
Neurobiology of Language
517
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
4
4
9
2
1
8
6
7
8
1
1
n
o
_
a
_
0
0
0
2
3
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Dynamic updating of native language
extend to unexposed verbs, which we interpreted as demonstrating a specific effect of RC expo-
sure rather than a generic effect of task practice. Future studies could explicitly manipulate
exposure type (e.g., RC sentences versus other kinds of fillers; see Prasad & Linzen, 2019;
Stack et al., 2018) in order to establish more definitively whether it is repeated experience with
RC structures in particular that causes neural adaptation. Finally, replication of the Stroop corre-
lation in a larger sample, differential correlations with different cognitive control tasks, and con-
comitant neuroimaging of multiple language and cognitive control tasks would allow for stronger
inferences about the particular aspects of cognitive control that are related to syntactic updating
and the extent to which those processes are domain-specific or general.
In conclusion, emerging research indicates that adults, not just children, continue to update
their native language based on new experiences. The current study shows that left frontal cortical
regions implicated in cognitive control play a role in updating syntactic expectations. The utility
of these regions could encompass broad support for attentional demands and task maintenance
during sentence processing as well as more specific resolution of conflict that arises from encoun-
tering unexpected sentences. Together, these areas could help ensure that updating is accurate
and efficient, thereby enabling language users to better navigate diverse and variable language
environments.
ACKNOWLEDGMENTS
We would like to thank Maria Braiuca for her assistance in data collection. We would also like to
thank Dr. John Van Meter and the research assistants at the Center for Functional and Molecular
Imaging (CFMI) at Georgetown University.
FUNDING INFORMATION
This study was funded in part by a University Facilitating Fund grant from the George Washington
University to Malathi Thothathiri.
AUTHOR CONTRIBUTIONS
Kelly Sharer and Malathi Thothathiri designed the experiment. Kelly Sharer created the materials
and collected data. Kelly Sharer and Malathi Thothathiri analyzed the data, designed the figures,
and wrote the manuscript.
REFERENCES
Altmann, G. T., & Kamide, Y. (1999). Incremental interpretation at
verbs: Restricting the domain of subsequent reference. Cognition,
73(3), 247–264. DOI: https://doi.org/10.1016/S0010-0277(99)
00059-1
Bates, D., Maechler, M., Bolker, B., & Walker, S. (2015). Fitting linear
mixed-effects models using lme4. Journal of Statistical Software,
67(1), 1–48. DOI: https://doi.org/10.18637/jss.v067.i01
Ben-Shachar, M., Hendler, T., Kahn, I., Ben-Bashat, D., &
Grodzinsky, Y. (2003). The neural reality of syntactic transforma-
tions: Evidence from functional magnetic resonance imaging.
Psychological Science, 14(5), 433–440. DOI: https://doi.org
/10.1111/1467-9280.01459, PMID: 12930473
Bernolet, S., & Hartsuiker, R. J. (2010). Does verb bias modulate
syntactic priming? Cognition, 114(3), 455–461. DOI: https://
doi.org/10.1016/j.cognition.2009.11.005, PMID: 20034624
Bock, K., & Griffin, Z. M. (2000). The persistence of structural priming:
Transient activation or implicit learning? Journal of Experimental
Psychology: General, 129(2), 177. DOI: https://doi.org/10.1037
/0096-3445.129.2.177, PMID: 10868333
Botvinick, M. M., Cohen, J. D., & Carter, C. S. (2004). Conflict
monitoring and anterior cingulate cortex: An update. Trends in
Cognitive Sciences, 8(12), 539–546. DOI: https://doi.org/10.1016
/j.tics.2004.10.003, PMID: 15556023
Busse, L., & Woldorff, M. G. (2003). The ERP omitted stimulus re-
sponse to “no-stim” events and its implications for fast-rate event-
related fMRI designs. NeuroImage, 18(4), 856–864. DOI: https://
doi.org/10.1016/S1053-8119(03)00012-0
Caplan, D., Alpert, N., & Waters, G. (1998). Effects of syntactic
structure and propositional number on patterns of regional cerebral
blood flow. Journal of Cognitive Neuroscience, 10(4), 541–552.
DOI: https://doi.org/10.1162/089892998562843, PMID:
9712683
Caplan, D., Chen, E., & Waters, G. (2008). Task-dependent and task-
independent neurovascular responses to syntactic processing.
Neurobiology of Language
518
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
4
4
9
2
1
8
6
7
8
1
1
n
o
_
a
_
0
0
0
2
3
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Dynamic updating of native language
Cortex, 44(3), 257–275. DOI: https://doi.org/10.1016/j.cortex
.2006.06.005, PMID: 18387556, PMCID: PMC2427191
Coulson, S., King, J. W., & Kutas, M. (1998). Expect the unexpected:
Event-related brain response to morphosyntactic violations.
Language and Cognitive Processes, 13(1), 21–58. DOI: https://
doi.org/10.1080/016909698386582
Desikan, R. S., Ségonne, F., Fischl, B., Quinn, B. T., Dickerson, B. C.,
Blacker, D., … Albert, M. S. (2006). An automated labeling system for
subdividing the human cerebral cortex on MRI scans into gyral based
regions of interest. NeuroImage, 31(3), 968–980. DOI: https://doi
.org/10.1016/j.neuroimage.2006.01.021, PMID: 16530430
Devauchelle, A. D., Oppenheim, C., Rizzi, L., Dehaene, S., & Pallier,
C. (2009). Sentence syntax and content in the human temporal
lobe: An fMRI adaptation study in auditory and visual modalities.
Journal of Cognitive Neuroscience, 21(5), 1000–1012. DOI: https://
doi.org/10.1162/jocn.2009.21070, PMID: 18702594
Dosenbach, N. U., Fair, D. A., Miezin, F. M., Cohen, A. L.,
Wenger, K. K., Dosenbach, R. A., … Schlaggar, B. L. (2007).
Distinct brain networks for adaptive and stable task control in
humans. Proceedings of the National Academy of Sciences, 104(26),
11073–11078. DOI: https://doi.org/10.1073/pnas.0704320104,
PMID: 17576922, PMCID: PMC1904171
Dosenbach, N. U., Visscher, K. M., Palmer, E. D., Miezin, F. M.,
Wenger, K. K., Kang, H. C., … Petersen, S. E. (2006). A core sys-
tem for the implementation of task sets. Neuron, 50(5), 799–812.
DOI: https://doi.org/10.1016/j.neuron.2006.04.031, PMID:
16731517, PMCID: PMC3621133
Dove, A., Pollmann, S., Schubert, T., Wiggins, C. J., & Von Cramon,
D. Y. (2000). Prefrontal cortex activation in task switching: An
event-related fMRI study. Cognitive Brain Research, 9(1), 103–109.
DOI: https://doi.org/10.1016/S0926-6410(99)00029-4
Duncan, J. (2010). The multiple-demand (MD) system of the primate
brain: Mental programs for intelligent behaviour. Trends in
Cognitive Sciences, 14(4), 172–179. DOI: https://doi.org/10.1016
/j.tics.2010.01.004, PMID: 20171926
Duncan, J. (2013). The structure of cognition: Attentional episodes
in mind and brain. Neuron, 80(1), 35–50. DOI: https://doi.org
/10.1016/j.neuron.2013.09.015, PMID: 24094101, PMCID:
PMC3791406
Farmer, T., Fine, A., Yan, S., Cheimariou, S., & Jaeger, F. (2014).
Error-driven adaptation of higher-level expectations during reading.
In Proceedings of the Annual Meeting of the Cognitive Science
Society (pp. 2181–2186).
Federmeier, K. D., & Kutas, M. (1999). A rose by any other name:
Long-term memory structure and sentence processing. Journal of
Memory and Language, 41(4), 469–495. DOI: https://doi.org/10
.1006/jmla.1999.2660
Fedorenko, E., Behr, M. K., & Kanwisher, N. (2011). Functional
specificity for high-level linguistic processing in the human
brain. Proceedings of the National Academy of Sciences, 108(39),
16428–16433. DOI: https://doi.org/10.1073/pnas.1112937108,
PMID: 21885736, PMCID: PMC3182706
Fedorenko, E., Blank, I. A., Siegelman, M., & Mineroff, Z. (2020).
Lack of selectivity for syntax relative to word meanings throughout
the language network. Cognition, 203, 104348. DOI: https://doi
.org/10.1016/j.cognition.2020.104348, PMID: 32569894
Fedorenko, E., Duncan, J., & Kanwisher, N. (2012). Language-selective
and domain-general regions lie side by side within Broca’s area.
Current Biology, 22(21), 2059–2062. DOI: https://doi.org/10.1016
/j.cub.2012.09.011, PMID: 23063434, PMCID: PMC3494832
Fedorenko, E., Duncan, J., & Kanwisher, N. (2013). Broad domain
generality in focal regions of frontal and parietal cortex. Proceedings
of the National Academy of Sciences, 110(41), 16616–16621. DOI:
https://doi.org/10.1073/pnas.1315235110, PMID: 24062451,
PMCID: PMC3799302
Fedorenko, E., & Thompson-Schill, S. L. (2014). Reworking the language
network. Trends in Cognitive Sciences, 18(3), 120–126. DOI: https://
doi.org/10.1016/j.tics.2013.12.006, PMID: 24440115, PMCID:
PMC4091770
Fernald, A., Thorpe, K., & Marchman, V. A. (2010). Blue car, red car:
Developing efficiency in online interpretation of adjective–noun
phrases. Cognitive Psychology, 60(3), 190–217. DOI: https://doi.org
/10.1016/j.cogpsych.2009.12.002, PMID: 20189552, PMCID:
PMC2835804
Ferretti, T. R., McRae, K., & Hatherell, A. (2001). Integrating verbs,
situation schemas, and thematic role concepts. Journal of Memory
and Language, 44(4), 516–547. DOI: https://doi.org/10.1006
/jmla.2000.2728
Fine, A. B., & Jaeger, T. F. (2013). Evidence for implicit learning
in syntactic comprehension. Cognitive Science, 37(3), 578–591.
DOI: https://doi.org/10.1111/cogs.12022, PMID: 23363004
Fine, A. B., Jaeger, T. F., Farmer, T. A., & Qian, T. (2013). Rapid
expectation adaptation during syntactic comprehension. PlOS
One, 8(10), e77661. DOI: https://doi.org/10.1371/journal.pone
.0077661, PMID: 24204909, PMCID: PMC3813674
Frazier, L., & Rayner, K. (1982). Making and correcting errors during
sentence comprehension: Eye movements in the analysis of structur-
ally ambiguous sentences. Cognitive Psychology, 14(2), 178–210.
DOI: https://doi.org/10.1016/0010-0285(82)90008-1
Friederici, A. D., & Kotz, S. A. (2003). The brain basis of syntactic
processes: Functional imaging and lesion studies. NeuroImage, 20,
S8–S17. DOI: https://doi.org/10.1016/j.neuroimage.2003.09.003,
PMID: 14597292
Gilbert, R. A., Davis, M. H., Gaskell, M. G., & Rodd, J. M. (2018).
Listeners and readers generalize their experience with word
meanings across modalities. Journal of Experimental Psychology:
Learning, Memory, and Cognition, 44(10), 1533. DOI: https://doi
.org/10.1037/xlm0000532, PMID: 29389181, PMCID:
PMC6179175
Gómez, R. L. (2002). Variability and detection of invariant structure.
Psychological Science, 13(5), 431–436. DOI: https://doi.org/10.1111
/1467-9280.00476, PMID: 12219809
Greve, D. N. (2002). Optseq2. Computer software. https://surfer
.nmr.mgh.harvard.edu/optseq
Grodzinsky, Y., & Friederici, A. D. (2006). Neuroimaging of syntax
and syntactic processing. Current Opinion in Neurobiology, 16(2),
240–246. DOI: https://doi.org/10.1016/j.conb.2006.03.007,
PMID: 16563739
Hagoort, P. (2016). MUC (Memory, Unification, Control): A model
on the neurobiology of language beyond single word processing.
In G. Hickok & S. L. Small (Eds.), Neurobiology of language
(pp. 339–347). San Diego, CA: Academic Press. DOI: https://
doi.org/10.1016/B978-0-12-407794-2.00028-6
Hagoort, P., & Brown, C. M. (2000). ERP effects of listening to
speech compared to reading: The P600/SPS to syntactic violations
in spoken sentences and rapid serial visual presentation.
Neuropsychologia, 38(11), 1531–1549. DOI: https://doi.org/10.1016
/S0028-3932(00)00053-1
Hale, J. (2001). A probabilistic Earley parser as a psycholinguistic
model. In Proceedings of the second meeting of the North
American Chapter of the Association for Computational
Linguistics on Language Technologies (pp. 1–8). New York:
ACM. DOI: https://doi.org/10.3115/1073336.1073357
Hsu, N. S., Jaeggi, S. M., & Novick, J. M. (2017). A common neural
hub resolves syntactic and non-syntactic conflict through coop-
eration with task-specific networks. Brain and Language, 166,
Neurobiology of Language
519
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
4
4
9
2
1
8
6
7
8
1
1
n
o
_
a
_
0
0
0
2
3
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Dynamic updating of native language
63–77. DOI: https://doi.org/10.1016/j.bandl.2016.12.006,
PMID: 28110105, PMCID: PMC5293615
Hsu, N. S., & Novick, J. M. (2016). Dynamic engagement of cognitive
control modulates recovery from misinterpretation during real-time
language processing. Psychological Science, 27(4), 572–582. DOI:
https://doi.org/10.1177/0956797615625223, PMID: 26957521,
PMCID: PMC4833548
January, D., Trueswell, J. C., & Thompson-Schill, S. L. (2009). Co-
localization of Stroop and syntactic ambiguity resolution in Broca’s
area: Implications for the neural basis of sentence processing.
Journal of Cognitive Neuroscience, 21(12), 2434–2444. DOI:
https://doi.org/10.1162/jocn.2008.21179, PMID: 19199402,
PMCID: PMC2762484
Jenkinson, M., Beckmann, C. F., Behrens, T. E., Woolrich, M. W., &
Smith, S. M. (2012). FSL. NeuroImage, 62(2), 782–790. DOI:
https://doi.org/10.1016/j.neuroimage.2011.09.015, PMID:
21979382
Jobard, G., Vigneau, M., Mazoyer, B., & Tzourio-Mazoyer, N.
(2007). Impact of modality and linguistic complexity during reading
and listening tasks. NeuroImage, 34(2), 784–800. DOI: https://doi
.org/10.1016/j.neuroimage.2006.06.067, PMID: 17110132
Just, M. A., Carpenter, P. A., Keller, T. A., Eddy, W. F., & Thulborn,
K. R. (1996). Brain activation modulated by sentence compre-
hension. Science, 274(5284), 114–116. DOI: https://doi.org/10
.1126/science.274.5284.114, PMID: 8810246
Kamide, Y. (2012). Learning individual talkers’ structural prefer-
ences. Cognition, 124(1), 66–71. DOI: https://doi.org/10.1016/j
.cognition.2012.03.001, PMID: 22498776
Kamide, Y., Altmann, G. T., & Haywood, S. L. (2003). The time-
course of prediction in incremental sentence processing:
Evidence from anticipatory eye movements. Journal of Memory
and Language, 49(1), 133–156. DOI: https://doi.org/10.1016
/S0749-596X(03)00023-8
Keller, T. A., Carpenter, P. A., & Just, M. A. (2001). The neural bases
of sentence comprehension: A fMRI examination of syntactic and
lexical processing. Cerebral Cortex, 11(3), 223–237. DOI: https://
doi.org/10.1093/cercor/11.3.223, PMID: 11230094
Kerns, J. G. (2006). Anterior cingulate and prefrontal cortex activity
in an FMRI study of trial-to-trial adjustments on the Simon task.
NeuroImage, 33(1), 399–405. DOI: https://doi.org/10.1016/j
.neuroimage.2006.06.012, PMID: 16876434
Kerns, J. G., Cohen, J. D., MacDonald, A. W., Cho, R. Y., Stenger,
V. A., & Carter, C. S. (2004). Anterior cingulate conflict monitoring
and adjustments in control. Science, 303(5660), 1023–1026. DOI:
https://doi.org/10.1126/science.1089910, PMID: 14963333
Kim, A., & Osterhout, L. (2005). The independence of combinatory
semantic processing: Evidence from event-related potentials.
Journal of Memory and Language, 52(2), 205–225. DOI: https://
doi.org/10.1016/j.jml.2004.10.002
Kriegeskorte, N., Simmons, W. K., Bellgowan, P. S., & Baker, C. I.
(2009). Circular analysis in systems neuroscience: The dangers of
double dipping. Nature Neuroscience, 12(5), 535–540. DOI:
https://doi.org/10.1038/nn.2303, PMID: 19396166, PMCID:
PMC2841687
Kroczek, L. O., & Gunter, T. C. (2017). Communicative predictions
can overrule linguistic priors. Scientific Reports, 7(1), 1–9. DOI:
https://doi.org/10.1038/s41598-017-17907-9, PMID: 29242511,
PMCID: PMC5730607
Ledoux, K., Traxler, M. J., & Swaab, T. Y. (2007). Syntactic priming
in comprehension: Evidence from event-related potentials.
Psychological Science, 18(2), 135–143. DOI: https://doi.org
/10.1111/j.1467-9280.2007.01863.x, PMID: 17425534,
PMCID: PMC1852484
Levy, R. (2008). Expectation-based syntactic comprehension.
Cognition, 106(3), 1126–1177. DOI: https://doi.org/10.1016/j
.cognition.2007.05.006, PMID: 17662975
MacDonald, M. C. (1994). Probabilistic constraints and syntactic
ambiguity resolution. Language and Cognitive Processes, 9(2),
157–201. DOI: https://doi.org/10.1080/01690969408402115
MacDonald, M. C., Just, M. A., & Carpenter, P. A. (1992). Working
memory constraints on the processing of syntactic ambiguity.
Cognitive Psychology, 24(1), 56–98. DOI: https://doi.org/10
.1016/0010-0285(92)90003-K
Mason, R. A., Just, M. A., Keller, T. A., & Carpenter, P. A. (2003).
Ambiguity in the brain: What brain imaging reveals about the
processing of syntactically ambiguous sentences. Journal of
Experimental Psychology: Learning, Memory, and Cognition, 29(6),
1319–1338. DOI: https://doi.org/10.1037/0278-7393.29.6.1319,
PMID: 14622064
Mehler, J., Jusczyk, P., Lambertz, G., Halsted, N., Bertoncini, J., &
Amiel-Tison, C. (1988). A precursor of language acquisition in young
infants. Cognition, 29(2), 143–178. DOI: https://doi.org/10.1016
/0010-0277(88)90035-2
Milham, M. P., Banich, M. T., Webb, A., Barad, V., Cohen, N. J.,
Wszalek, T., & Kramer, A. F. (2001). The relative involvement
of anterior cingulate and prefrontal cortex in attentional control
depends on nature of conflict. Cognitive Brain Research, 12(3),
467–473. DOI: https://doi.org/10.1016/S0926-6410(01)00076-3
Morey, R. D. (2008) Confidence intervals from normalized data: A
correction to Cousineau (2005). Tutorials in Quantitative Methods
for Psychology 4(2), 61–64. DOI: https://doi.org/10.20982/tqmp
.04.2.p061
Mumford, J. (2007). A guide to calculating percent change with
featquery. Unpublished Technical Report. http://mumford.fmripower
.org/perchange_guide.pdf
Newport, E. L. (1990). Maturational constraints on language learning.
Cognitive Science, 14(1), 11–28. DOI: https://doi.org/10.1207
/s15516709cog1401_2
Nomura, E. M., Gratton, C., Visser, R. M., Kayser, A., Perez, F., &
D’Esposito, M. (2010). Double dissociation of two cognitive con-
trol networks in patients with focal brain lesions. Proceedings of
the National Academy of Sciences, 107(26), 12017–12022. DOI:
https://doi.org/10.1073/pnas.1002431107, PMID: 20547857,
PMCID: PMC2900657
Noppeney, U., & Price, C. J. (2004). An fMRI study of syntactic
adaptation. Journal of Cognitive Neuroscience, 16(4), 702–713.
DOI: https://doi.org/10.1162/089892904323057399, PMID:
15165357
Novick, J. M., Kan, I. P., Trueswell, J. C., & Thompson-Schill, S. L.
(2009). A case for conflict across multiple domains: Memory and
language impairments following damage to ventrolateral prefrontal
cortex. Cognitive Neuropsychology, 26(6), 527–567. DOI: https://
doi.org/10.1080/02643290903519367, PMID: 20183014,
PMCID: PMC3791076
Novick, J. M., Trueswell, J. C., & Thompson-Schill, S. L. (2005).
Cognitive control and parsing: Reexamining the role of Broca’s
area in sentence comprehension. Cognitive, Affective, & Behavioral
Neuroscience, 5(3), 263–281. DOI: https://doi.org/10.3758/CABN
.5.3.263, PMID: 16396089
Osterhout, L., & Holcomb, P. J. (1992). Event-related brain potentials
elicited by syntactic anomaly. Journal of Memory and Language,
31(6), 785–806. DOI: https://doi.org/10.1016/0749-596X(92)
90039-Z
Osterhout, L., Holcomb, P. J., & Swinney, D. A. (1994). Brain poten-
tials elicited by garden-path sentences: Evidence of the application
of verb information during parsing. Journal of Experimental
Neurobiology of Language
520
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
4
4
9
2
1
8
6
7
8
1
1
n
o
_
a
_
0
0
0
2
3
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Dynamic updating of native language
Psychology: Learning, Memory, and Cognition, 20(4), 786–803.
DOI: https://doi.org/10.1037/0278-7393.20.4.786
Pernet, C. R. (2014). Misconceptions in the use of the General
Linear Model applied to functional MRI: A tutorial for junior neuro-
imagers. Frontiers in Neuroscience, 8, 1. DOI: https://doi.org
/10.3389/fnins.2014.00001, PMID: 24478622, PMCID:
PMC3896880
Pernet, C. R., Wilcox, R. R., & Rousselet, G. A. (2013). Robust cor-
relation analyses: False positive and power validation using a
new open source Matlab toolbox. Frontiers in Psychology, 3, 606.
DOI: https://doi.org/10.3389/fpsyg.2012.00606, PMID:
23335907, PMCID: PMC3541537
Prasad, G., & Linzen, T. (2019, March 16). Rapid syntactic adaptation
in self-paced reading: Detectable, but requires many participants.
PsyArXiv. DOI: https://doi.org/10.31234/osf.io/9ptg4
Qian, Z., Garnsey, S., & Christianson, K. (2018). A comparison of
online and offline measures of good-enough processing in garden-
path sentences. Language, Cognition and Neuroscience, 33(2),
227–254. DOI: https://doi.org/10.1080/23273798.2017.1379606
Rayner, K. (1998). Eye movements in reading and information pro-
cessing: 20 years of research. Psychological Bulletin, 124(3), 372.
DOI: https://doi.org/10.1037/0033-2909.124.3.372, PMID: 9849112
Rayner, K., Kambe, G., & Duffy, S. A. (2000). The effect of clause
wrap-up on eye movements during reading. The Quarterly Journal
of Experimental Psychology Section A, 53(4), 1061–1080. DOI:
https://doi.org/10.1080/713755934
Rodd, J. M., Cai, Z. G., Betts, H. N., Hanby, B., Hutchinson, C., &
Adler, A. (2016). The impact of recent and long-term experience
on access to word meanings: Evidence from large-scale internet-
based experiments. Journal of Memory and Language, 87, 16–37.
DOI: https://doi.org/10.1016/j.jml.2015.10.006
Rodd, J. M., Cutrin, B. L., Kirsch, H., Millar, A., & Davis, M. H.
(2013). Long-term priming of the meanings of ambiguous words.
Journal of Memory and Language, 68(2), 180–198. DOI: https://
doi.org/10.1016/j.jml.2012.08.002
Rodd, J. M., Longe, O. A., Randall, B., & Tyler, L. K. (2010).
The functional organisation of the fronto-temporal language
system: Evidence from syntactic and semantic ambiguity. Neu-
ropsychologia, 48(5), 1324–1335. DOI: https://doi.org/10.1016
/j.neuropsychologia.2009.12.035, PMID: 20038434
Roland, D., Dick, F., & Elman, J. L. (2007). Frequency of basic
English grammatical structures: A corpus analysis. Journal of
Memory and Language, 57(3), 348–379. DOI: https://doi.org
/10.1016/j.jml.2007.03.002, PMID: 19668599, PMCID:
PMC2722756
Ruge, H., Jamadar, S., Zimmermann, U., & Karayanidis, F. (2013).
The many faces of preparatory control in task switching: Reviewing
a decade of fMRI research. Human Brain Mapping, 34(1), 12–35.
DOI: https://doi.org/10.1002/ hbm.21420, PMID: 21998090,
PMCID: PMC6870098
Ryskin, R. A., Qi, Z., Duff, M. C., & Brown-Schmidt, S. (2017). Verb
biases are shaped through lifelong learning. Journal of Experimental
Psychology: Learning, Memory, and Cognition, 43(5), 781–794.
DOI: https://doi.org/10.1037/xlm0000341, PMID: 27762578,
PMCID: PMC5398958
Santi, A., & Grodzinsky, Y. (2010). fMRI adaptation dissociates syn-
tactic complexity dimensions. NeuroImage, 51(4), 1285–1293.
DOI: https://doi.org/10.1016/j.neuroimage.2010.03.034, PMID:
20338244, PMCID: PMC2909752
Scott, R. M., & Fisher, C. (2009). Two-year-olds use distributional
cues to interpret transitivity-alternating verbs. Language and
Cognitive Processes, 24(6), 777–803. DOI: https://doi.org/10.1080
/01690960802573236, PMID: 20046985, PMCID: PMC2701745
Segaert, K., Kempen, G., Petersson, K. M., & Hagoort, P. (2013).
Syntactic priming and the lexical boost effect during sentence
production and sentence comprehension: An fMRI study. Brain
and Language, 124(2), 174–183. DOI: https://doi.org/10.1016/j
.bandl.2012.12.003, PMID: 23376214
Segaert, K., Menenti, L., Weber, K., Petersson, K. M., & Hagoort, P.
(2011). Shared syntax in language production and language
comprehension—an fMRI study. Cerebral Cortex, 22(7), 1662–1670.
DOI: https://doi.org/10.1093/cercor/bhr249, PMID: 21934094,
PMCID: PMC3377967
Smith, L., & Yu, C. (2008). Infants rapidly learn word-referent map-
pings via cross-situational statistics. Cognition, 106(3), 1558–1568.
DOI: https://doi.org/10.1016/j.cognition.2007.06.010, PMID:
17692305, PMCID: PMC2271000
Snedeker, J., & Trueswell, J. C. (2004). The developing constraints on
parsing decisions: The role of lexical-biases and referential scenes
in child and adult sentence processing. Cognitive Psychology, 49(3),
238–299. DOI: https://doi.org/10.1016/j.cogpsych.2004.03.001,
PMID: 15342261
Stack, C. M. H., James, A. N., & Watson, D. G. (2018). A failure to
replicate rapid syntactic adaptation in comprehension. Memory
& Cognition, 46(6), 864–877. DOI: https://doi.org/10.3758/s13421
-018-0808-6, PMID: 29651687
Swainson, R., Cunnington, R., Jackson, G. M., Rorden, C., Peters,
A. M., Morris, P. G., & Jackson, S. R. (2003). Cognitive control
mechanisms revealed by ERP and fMRI: Evidence from repeated
task-switching. Journal of Cognitive Neuroscience, 15(6), 785–799.
DOI: https://doi.org/10.1162/089892903322370717, PMID:
14511532
Tagliamonte, S. (2011). Variationist sociolinguistics: Change, obser-
vation, interpretation. Hoboken, NJ: John Wiley & Sons.
Tanenhaus, M. K., Carlson, G., & Trueswell, J. C. (1989). The role
of thematic structures in interpretation and parsing. Language
and Cognitive Processes, 4(3–4), SI211–SI234. DOI: https://doi.org
/10.1080/01690968908406368
Tooley, K. M., Traxler, M. J., & Swaab, T. Y. (2009). Elec-
trophysiological and behavioral evidence of syntactic priming in
sentence comprehension. Journal of Experimental Psychology:
Learning, Memory, and Cognition, 35(1), 19–45. DOI: https://doi.org
/10.1037/a0013984, PMID: 19210079
Thothathiri, M. (2018). Statistical experience and individual cognitive
differences modulate neural activity during sentence production.
Brain and Language, 183, 47–53. DOI: https://doi.org/10.1016/j
.bandl.2018.06.005, PMID: 29913355
Thothathiri, M., Asaro, C. T., Hsu, N. S., & Novick, J. M. (2018). Who did
what? A causal role for cognitive control in thematic role assignment
during sentence comprehension. Cognition, 178, 162–177. DOI:
https://doi.org/10.1016/j.cognition.2018.05.014, PMID: 29860176
Thothathiri, M., Gagliardi, M., & Schwartz, M. F. (2012). Subdivision
of frontal cortex mechanisms for language production in aphasia.
Neuropsychologia, 50(14), 3284–3294. DOI: https://doi.org
/10.1016/j.neuropsychologia.2012.09.021, PMID: 23022077,
PMCID: PMC3518563
Thothathiri, M., Kim, A., Trueswell, J. C., & Thompson-Schill, S. L.
(2012). Parametric effects of syntactic–semantic conflict in
Broca’s area during sentence processing. Brain and Language,
120(3), 259–264. DOI: https://doi.org/10.1016/j.bandl.2011
.12.004, PMID: 22225974, PMCID: PMC3299904
Trueswell, J. C., & Tanenhaus, M. K. (1994). Toward a lexicalist
framework of constraint-based syntactic ambiguity resolution.
In C. Clifton, Jr., L. Frazier, & K. Rayner (Eds.), Perspectives on
sentence processing (pp. 155–179). Hilldale, NJ: Lawrence
Erlbaum Associates, Inc.
Neurobiology of Language
521
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
4
4
9
2
1
8
6
7
8
1
1
n
o
_
a
_
0
0
0
2
3
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Dynamic updating of native language
Trueswell, J. C., Tanenhaus, M. K., & Kello, C. (1993). Verb-specific
constraints in sentence processing: Separating effects of lexical
preference from garden-paths. Journal of Experimental Psychology:
Learning, Memory, and Cognition, 19(3), 528–553. DOI: https://
doi.org/10.1037/0278-7393.19.3.528
Twilley, L. C., & Dixon, P. (2000). Meaning resolution processes for
words: A parallel independent model. Psychonomic Bulletin &
Review, 7(1), 49–82. DOI: https://doi.org/10.3758/BF03210725,
PMID: 10780020
Tyler, L. K., Marslen-Wilson, W. D., Randall, B., Wright, P.,
Devereux, B. J., Zhuang, J., … Stamatakis, E. A. (2011). Left in-
ferior frontal cortex and syntax: Function, structure and behaviour
in patients with left hemisphere damage. Brain, 134(2), 415–431.
DOI: https://doi.org/10.1093/brain/awq369, PMID: 21278407,
PMCID: PMC3030769
Ullsperger, M., Fischer, A. G., Nigbur, R., & Endrass, T. (2014).
Neural mechanisms and temporal dynamics of performance mon-
itoring. Trends in Cognitive Sciences, 18(5), 259–267. DOI: https://
doi.org/10.1016/j.tics.2014.02.009, PMID: 24656460
Ullsperger, M., Harsay, H. A., Wessel, J. R., & Ridderinkhof, K. R.
(2010). Conscious perception of errors and its relation to the an-
terior insula. Brain Structure and Function, 214(5–6), 629–643.
DOI: https://doi.org/10.1007/s00429-010-0261-1, PMID:
20512371, PMCID: PMC2886909
van de Meerendonk, N., Rueschemeyer, S. A., & Kolk, H. H.
(2013). Language comprehension interrupted: Both language
errors and word degradation activate Broca’s area. Brain and
Language, 126(3), 291–301. DOI: https://doi.org/10.1016/j
.bandl.2013.07.004, PMID: 23933469
Vidal, C., Chetail, F., & Content, A. (2016). BACS: The Brussels
Artificial Characters Sets for studies in cognitive psychology
and neuroscience. Retrieved from osf.io/dj8qm
Weber, K., & Indefrey, P. (2009). Syntactic priming in German–
English bilinguals during sentence comprehension. NeuroImage,
46(4), 1164–1172. DOI: https://doi.org/10.1016/j.neuroimage
.2009.03.040, PMID: 19328233
Wells, J. B., Christiansen, M. H., Race, D. S., Acheson, D. J., &
MacDonald, M. C. (2009). Experience and sentence processing:
Statistical learning and relative clause comprehension. Cognitive
Psychology, 58(2), 250–271. DOI: https://doi.org/10.1016/j
.cogpsych.2008.08.002, PMID: 18922516, PMCID: PMC2621112
Werker, J. F., & Tees, R. C. (2005). Speech perception as a window
for understanding plasticity and commitment in language systems
of the brain. Developmental Psychobiology: The Journal of
the International Society for Developmental Psychobiology, 46(3),
233–251. DOI: https://doi.org/10.1002/dev.20060, PMID: 15772961
Wilson, M. P., & Garnsey, S. M. (2009). Making simple sentences
hard: Verb bias effects in simple direct object sentences. Journal
of Memory and Language, 60(3), 368–392. DOI: https://doi
.org/10.1016/j.jml.2008.09.005, PMID: 20160997, PMCID:
PMC2756706
Woolgar, A., Williams, M. A., & Rich, A. N. (2015). Attention enhances
multi-voxel representation of novel objects in frontal, parietal and
visual cortices. NeuroImage, 109, 429–437. DOI: https://doi.org/10
.1016/j.neuroimage.2014.12.083, PMID: 25583612
Ye, Z., & Zhou, X. (2009). Conflict control during sentence compre-
hension: fMRI evidence. NeuroImage, 48(1), 280–290. DOI:
https://doi.org/10.1016/j.neuroimage.2009.06.032, PMID: 19540923
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
4
4
9
2
1
8
6
7
8
1
1
n
o
_
a
_
0
0
0
2
3
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Neurobiology of Language
522