RESEARCH ARTICLE
Brain Structures and Cognitive Abilities Important
for the Self-Monitoring of Speech Errors
Ayan S. Mandal1,2
Andrew T. DeMarco2
, Mackenzie E. Fama2,3
, Elizabeth H. Lacey2,4
, Laura M. Skipper-Kallal2
,
, and Peter E. Turkeltaub2,4
1University of Cambridge, Department of Psychiatry, Cambridge, UK
2Georgetown University Medical Center, Center for Brain Plasticity and Recovery and Department of Neurology,
Washington, DC
3Towson University, Department of Audiology, Speech-Language Pathology, and Deaf Studies, Towson, MD
4MedStar National Rehabilitation Hospital, Research Division, Washington, DC
Keywords: aphasia, self-monitoring, speech production, conflict monitoring, executive function,
frontal white matter
ABSTRACT
The brain structures and cognitive abilities necessary for successful monitoring of one’s own
speech errors remain unknown. We aimed to inform self-monitoring models by examining the
neural and behavioral correlates of phonological and semantic error detection in individuals
with post-stroke aphasia. First, we determined whether detection related to other abilities
proposed to contribute to monitoring according to various theories, including naming ability,
fluency, word-level auditory comprehension, sentence-level auditory comprehension, and
executive function. Regression analyses revealed that fluency and executive scores were
independent predictors of phonological error detection, while a measure of word-level
comprehension related to semantic error detection. Next, we used multivariate lesion-
symptom mapping to determine lesion locations associated with reduced error detection.
Reduced overall error detection related to damage to a region of frontal white matter extending
into dorsolateral prefrontal cortex. Detection of phonological errors related to damage to the
same areas, but the lesion-behavior association was stronger, suggesting that the localization
for overall error detection was driven primarily by phonological error detection. These findings
demonstrate that monitoring of different error types relies on distinct cognitive functions, and
provide causal evidence for the importance of frontal white matter tracts and the dorsolateral
prefrontal cortex for self-monitoring of speech.
INTRODUCTION
Although fluent speech is littered with errors, healthy speakers can identify and repair these
mistakes. Successful communication depends on this ability to self-correct. Previous studies of
error detection have shown that it predicts positive therapeutic outcomes for both production
and comprehension in aphasia (Marshall, Neuburger, & Phillips, 1994). Schwartz, Middleton,
Brecher, Gagliardi, and Garvey (2016) provided evidence that the correction of semantic errors
promotes an adaptive change that allows aphasic patients to learn from their mistakes. These
results imply that self-monitoring has important consequences for aphasia recovery.
Despite the evidence that self-monitoring plays a role in language relearning, little is known
about the processes underlying it. There are currently two broad categories of theories
a n o p e n a c c e s s
j o u r n a l
Citation: Mandal, A. S., Fama, M. E.,
Skipper-Kallal, L. M., DeMarco, A. T.,
Lacey, E. H., & Turkeltaub, P. E. (2020).
Brain structures and cognitive abilities
important for the self-monitoring of
speech errors. Neurobiology of
Language, 1(3), 319–338. https://doi.
org/10.1162/nol_a_00015
DOI:
https://doi.org/10.1162/nol_a_00015
Supporting Information:
https://doi.org/10.1162/nol_a_00015
Received: 30 July 2019
Accepted: 20 May 2020
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Author:
Peter E. Turkeltaub
turkeltp@georgetown.edu
Handling Editor:
Steven Small
Copyright: © 2020 Massachusetts
Institute of Technology. Published
under a Creative Commons Attribution
4.0 International (CC BY 4.0) license.
The MIT Press
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
1
3
0
1
8
6
7
7
3
3
n
o
_
a
_
0
0
0
1
5
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Lesion mapping of speech error detection
regarding self-monitoring. The first category consists of comprehension-based models, where
persons detect errors by listening to their own speech (Postma, 2000). When speakers hear
themselves say something different from what was intended, then they can identify and correct
the error. Chief among comprehension-based models for error detection is the perceptual loop
model initially posed by Levelt (1983). Under the perceptual loop model, an outer auditory
loop monitors overt speech using speech comprehension systems. Pre-articulatory inner
speech is monitored as well via an inner loop that also relies on speech comprehension
(Indefrey & Levelt, 2004). This model is parsimonious because it does not assume the exis-
tence of a system dedicated to self-detecting errors. Rather, it suggests that the same mecha-
nism that allows people to comprehend the speech of others also allows them to detect their
own errors.
Comprehension-based models for self-monitoring predict that poor error detection will cor-
relate with poor comprehension abilities. If one detects an error by comprehending one’s own
overt speech, then a person who has difficulties comprehending the speech of others should
also have difficulties with self-monitoring. However, Nickels and Howard (1995) found no
correlation between error detection and any of three measures of auditory comprehension.
Furthermore, comprehension and error detection doubly dissociate: There have been case
studies of patients with aphasia who accurately detect their own errors but demonstrate poor
comprehension (Marshall, Rappaport, & Garcia-Bunuel, 1985), as well as patients with poor
error detection yet intact comprehension (Butterworth & Howard, 1987; Liss, 1998;
Marshall, Robson, Pring, & Chiat, 1998). It is worth noting, however, that while these
studies provide substantial evidence against an overt-speech monitoring loop, it has been more
difficult to test the functioning of an inner-speech monitoring loop.
In contrast to comprehension-based speech monitors, some authors have proposed production-
based self-monitoring systems, where information from the speech production process itself can
be used to detect an error. Particularly notable amongst production-based models for error
detection is the conflict-based monitor proposed by Nozari, Dell, and Schwartz (2011). In
Nozari et al.’s model, the language system emits conflict signals that arise from competition
between various semantic features or phonological units activated during naming. These
signals are received by the anterior cingulate cortex, which processes the conflict and alerts
the lateral prefrontal cortex (LPFC) to exert top-down control to resolve conflict (Botvinick,
Braver, Barch, Carter, & Cohen, 2001; Yeung, 2015; Yeung, Botvinick, & Cohen, 2004). The
conflict-based model has recently gained support from a behavioral study in neurotypical
children (Hanley, Cortis, Budd, & Nozari, 2016) and a functional MRI study in healthy adults
(Gauvin, De Baene, Brass, & Hartsuiker, 2015).
The self-monitoring models mentioned above make different predictions regarding the neu-
ral and behavioral correlates of error detection. The perceptual loop model predicts that de-
tection should depend on comprehension. Consequently, the perceptual loop model also
predicts detection to rely on brain structures that subserve comprehension, that is, primarily
regions within the temporal lobe (Hillis, Rorden, & Fridriksson, 2017). Production monitors in
general predict that lesions to areas important for word production, broadly within the frontal
lobe, should impair error detection capabilities (Catani et al., 2013; Mandelli et al., 2014). The
conflict-based model in particular also predicts that regions involved in domain-general cog-
nitive processing, such as the anterior cingulate cortex (ACC) or LPFC, should also be involved
(Yeung, 2015).
People with aphasia sometimes exhibit impairments in monitoring that are specific to one
type of error over another, such as detecting each of their phonological errors but failing to
Neurobiology of Language
320
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
1
3
0
1
8
6
7
7
3
3
n
o
_
a
_
0
0
0
1
5
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Lesion mapping of speech error detection
notice any of their semantic errors (Marshall et al., 1985; Stark, 1988). Such observations
suggest that different cognitive processes may be involved in the monitoring of different types
of errors. This notion has received empirical support in recent years from studies that have
found differences in the timing of and the learning from semantic versus phonological error
monitoring (Schuchard, Middleton, & Schwartz, 2017; Schwartz et al., 2016).
Comprehension-based models do not explicitly account for differential monitoring of phono-
logical versus semantic errors, although one may expect a selective deficit in the monitoring of
one type of error if the comprehension system has incurred a selective impairment in either
phonological or semantic processing. However, a case study of a patient with impaired pho-
nological auditory processing (in the form of auditory agnosia) yet preserved reading compre-
hension demonstrated the opposite of this expectation: The subject detected almost all of her
phonological errors but ignored her semantic errors (Marshall et al., 1985). Production-based
monitors for error detection have been proposed for virtually every stage of word production,
from lemma selection to tactile feedback following word articulation (Postma, 2000), and
therefore could support differential monitoring of errors that arise at different stages. Nozari
et al. (2011) found that measures of the lexical-semantic and lexical-phonological stages of
naming predicted detection of semantic and phonological errors, respectively. The authors
suggested that damage to the lexical-semantic or lexical-phonologic stages of naming causes
noise to predominate in the activations of representations by these systems, which obscures
the ability of a monitor to detect conflict related to errors.
An examination of the brain structures and cognitive abilities associated with detection of
different types of naming errors could help not only distinguish between current self-monitoring
accounts but also extend existing theories into new domains.
The Current Study
In the present study, we tested the anatomical and behavioral predictions of the perceptual
loop and conflict-based models for error detection in a group of participants with post-stroke
aphasia. First, we determined whether error detection within the context of a picture-naming
task related to cognitive abilities proposed to contribute to monitoring, including naming abil-
ity, fluency, word-level auditory comprehension, sentence-level auditory comprehension, and
executive function. Then, we used support vector regression lesion-symptom mapping (SVR-
LSM) to map the brain areas necessary for error detection, and probed the interrelationships
between the behavioral and neural correlates of detection ability.
MATERIALS AND METHODS
Participants
Data for the current study were pooled from cohorts of left hemisphere stroke survivors recruited
for two different studies at Georgetown University and MedStar National Rehabilitation
Hospital. Forty-nine patients (Cohort 1) were participating in a battery of tasks to determine
baseline language abilities in a transcranial direct current stimulation clinical trial. Fifty-four
patients (Cohort 2) were participating in a study designed to probe the subjective experience of
inner speech in aphasia. Twenty-three patients participated in both studies, yielding a total of
80 potential participants after combining the two cohorts. All participants were native English
speakers, had no history of other brain disorder or damage, had anterior circulation strokes,
and were tested at least six months after the stroke. Participants were excluded from analyses
if they produced too few errors of the type being examined to assess error detection (see
Dependent Variables below). Demographic details of the participants included in each
Neurobiology of Language
321
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
1
3
0
1
8
6
7
7
3
3
n
o
_
a
_
0
0
0
1
5
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Lesion mapping of speech error detection
analysis are listed in Table 1. The study was approved by the Georgetown University
Institutional Review Board, and written informed consent was obtained from all study partic-
ipants prior to enrollment in the study.
Behavioral Tasks
All relevant tasks that had been administered to both Cohort 1 and Cohort 2 were selected for the
study. These included tasks that measure confrontation picture-naming performance and error
detection, as well as several functions proposed to be important for error monitoring, including
word-level auditory comprehension, sentence-level speech comprehension, short-term
Table 1. Demographic data and performance on measures of interest for study participants
Age (years)
Sex (male/female)
Time since stroke (months)
Education (years)
Handedness (left/right/ambidextrous)
Lesion size (cm3)
Naming accuracy
Phonological errors
Semantic errors
Total error detection
Phonological error detection
Semantic error detection
WAB-R auditory comprehension
Word-to-picture matching
MLU
Digit span forwards
Digit span backwards
Digit span difference
Spatial span forwards
Spatial span backwards
Spatial span difference
Total error
detection group
(N = 64)
60.4 (9.3)
42/22
47.6 (48.8)
16.0 (2.9)
56/5/3
Phonological error
detection group
(N = 58)
60.0 (9.2)
38/20
43.4 (44.6)
15.9 (3.0)
51/4/3
Semantic error
detection group
(N = 32)
60.6 (8.4)
19/13
49.3 (57.2)
16.3 (2.6)
27/4/3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
1
3
0
1
8
6
7
7
3
3
n
o
_
a
_
0
0
0
1
5
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
117.9 (81.0)
117.8 (81.4)
121.6 (80.5)
0.52 (0.31)
0.48 (0.21)
0.16 (0.12)
0.40 (0.24)
0.40 (0.28)
0.38 (0.25)
0.93 (0.065)
0.92 (0.11)
4.2 (2.3)
4.7 (3.2)
2.2 (2.0)
2.5 (2.4)
6.3 (2.2)
5.1 (2.4)
1.1 (1.7)
0.51 (0.31)
0.51 (0.19)
0.15 (0.10)
0.40 (0.25)
0.39 (0.28)
0.38 (0.25)
0.92 (0.065)
0.91 (0.11)
4.1 (2.3)
4.5 (3.2)
2.0 (1.9)
2.5 (2.4)
6.3 (2.2)
5.3 (2.3)
0.48 (0.29)
0.39 (0.19)
0.24 (0.11)
0.42 (0.21)
0.42 (0.24)
0.38 (0.25)
0.92 (0.074)
0.92 (0.10)
4.4 (2.2)
5.3 (3.6)
2.3 (2.2)
3.0 (2.3)
6.1 (2.5)
5.3 (2.4)
0.95 (1.7)
0.81 (1.4)
Note. Standard deviations are presented in parentheses. Each behavioral measure has been divided by its maximum possible score, except MLU, digit span, and
spatial span tasks, which have no maximum score. Phonological error detection and semantic error detection groups are subsets of the total error detection
group. Twenty-nine participants are in both the phonological error detection and semantic error detection groups. WAB-R = Western Aphasia Battery-Revised,
MLU = mean length of utterance.
Neurobiology of Language
322
Lesion mapping of speech error detection
memory, working memory, executive function, and fluency. In participants for whom scores
were available from both prior studies, the scores of the two equivalent tests were averaged.
Philadelphia Naming Test
Participants from Cohort 1 were administered a 60-item version of the Philadelphia Naming
Test (PNT; Roach et al., 1996). The PNT is a picture-naming task where participants must
name a series of black and white drawings. Participants from Cohort 2 were administered a
120-item naming task, which included the 60-item PNT plus an additional 60 items. For pa-
tients who participated in both prior studies, only the 60 PNT items that matched across both
groups were used to provide an overall naming accuracy score, but all available trials were
pooled across both tasks for the purpose of coding error types and error detection. To deter-
mine whether this pooling substantially impacted the results, analyses were also conducted
using the error detection performance from only the 60-item PNT administered to both cohorts
(see the online supporting information located at https://www.mitpressjournals.org/doi/suppl/
10.1162/nol_a_00015).
Western Aphasia Battery-Revised auditory comprehension
The Western Aphasia Battery-Revised ( WAB-R) Yes/No Questions task was administered. This
task requires a yes/no response to 20 items including questions that are biographical, environ-
mental, and noncontextual/grammatically complex in nature (Kertesz, 2007) and assesses
sentence-level comprehension ability.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
1
3
0
1
8
6
7
7
3
3
n
o
_
a
_
0
0
0
1
5
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Digit span forwards
Participants were asked to repeat strings of numbers of increasing length in the same order in
which they heard the sequence. Two strings were presented at each length, and testing
stopped after both strings at a given length were recited incorrectly. The total number of strings
recited correctly was taken, in which a low number indicates poor verbal short-term memory.
Word-to-picture matching
Participants heard a word and pointed to the target item in a field of semantically related pic-
tures. The version of the task used in Cohort 1 included a field of six pictures, whereas in
Cohort 2 the field included four pictures. Both tasks included 48 trials. Accuracies on the
two tasks were not different for the 23 patients who performed both, paired t(22) = 0.776,
p = 0.45, so the tasks were treated as equivalent. Poor performance on this task is interpreted
to indicate word-level comprehension impairment, but could reflect other factors (see
Discussion below).
Mean length of utterance
Mean length of utterance (MLU) during a picture description task was used as a measure of
speech fluency. This quantity is derived by calculating the mean number of words used in each
utterance. Participants from Cohort 1 described the picnic scene from the WAB–R, while those
from Cohort 2 described the “Cookie Theft” picture from the Boston Diagnostic Aphasia
Examination (Goodglass, Kaplan & Barresi, 2001). The scores from the two pictures were not
different for the patients for whom both scores were available, paired t(19) = 1.20, p = 0.244,
so the tasks were treated as equivalent. Five subjects were missing data and so are excluded
from analyses using MLU.
Neurobiology of Language
323
Lesion mapping of speech error detection
Digit span backwards
The same procedures were followed as for digit span forwards, except that participants were
asked to recite the number strings in reverse order, a manipulation that is typically thought to
tax executive control of working memory.
Digit span difference
The difference between the forwards and backwards digit span scores was calculated. A high
number on the measure is interpreted here to indicate poor executive control, but could also
reflect other factors (see Discussion).
Spatial span forwards
The Corsi-Block tapping task was used (Corsi, 1972). Participants were asked to tap a se-
quence of blocks of increasing length in the same order in which they saw an examiner tap
the blocks. Two sequences were presented at each length, and testing stopped after both se-
quences of a given length were repeated incorrectly. The total number of sequences repeated
correctly was taken, in which a low number indicates poor nonverbal short-term memory.
Spatial span backwards
The same procedures were followed as for spatial span forwards, except that participants were
asked to tap the sequence of blocks in reverse order, a manipulation that is typically thought to
tax executive control of working memory.
Spatial span difference
The difference between the forwards and backwards spatial span scores was calculated. A
high number on this measure is interpreted to indicate poor executive control, but could also
reflect other factors (see Discussion).
Coding Naming Responses for Error Type and Error Detection
Error type
Videos of naming responses were transcribed into the International Phonetic Alphabet and
scored offline for accuracy, error type, and error detection. Error coding was based on the
PNT rules (Roach et al., 1996). Only the first naming attempt for each item was coded.
False starts and fragments were not considered as first naming attempts as per PNT scoring
rules. Errors, whether words or nonwords, were coded as phonological if they shared the
stressed vowel, at least two phonemes, or the first or last phoneme as the target. Errors that
were semantically related to the target were coded as semantic errors. Errors that were both
phonologically and semantically related to the target were coded with mixed errors and were
thus considered as neither phonological nor semantic. The other error types specified in the
PNT scoring rules were coded but are not considered here. Seventeen subjects (four of whom
received the 120-item naming task) were graded by two independent scorers to determine
interrater reliability. Errors received the same error code from different scorers on 91.7% of
trials. This is comparable to the interrater reliability observed in similar studies (Schwartz
et al., 2016).
Neurobiology of Language
324
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
1
3
0
1
8
6
7
7
3
3
n
o
_
a
_
0
0
0
1
5
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Lesion mapping of speech error detection
Error detection
Error detection was assessed within the context of the picture-naming task using an adapted
version of the coding protocol developed by Schwartz and colleagues (Schwartz et al., 2016).
A participant was said to have detected an error if they uttered a statement indicating aware-
ness that an error was made or if they attempted to self-correct the error. In other words,
when a participant made an error, it would count as detected if the person either followed
the errant response with a statement like “no, not that,” or made an attempt to correct their
first response (e.g., “cat … I mean dog”), indicating recognition that the first response was in-
correct. Attempts at self-correction were coded as detected errors regardless of whether the
second attempt was correct or incorrect because either provides evidence that the individual
is aware of the error. Only the first naming attempt for each item was scored for error detec-
tion; detection or correction of errors in attempts at self-correction were not scored (e.g., for
“cat … I mean dog … I mean bird” only the first self-correction is scored). This ensured that
there was only one error detection score per item, which reduced bias toward individuals who
made many attempts at self-correction, and made it simpler to interpret detection of specific
error types. Nonverbal indicators of error acknowledgement, such as head shakes, were not
coded. Repetition of the initial naming attempt (e.g., “cat … cat”) was also not coded as an
error detection. The accuracy of attempts at self-correction was recorded, but those data are
not considered here because there was not adequate power to further divide the types of de-
tections and corrections. Consistent with protocols in prior studies (Nozari et al., 2011;
Schuchard et al., 2017; Schwartz et al., 2016), participants were not given instructions to in-
dicate awareness of an error or to self-correct, so all error detection was spontaneous.
Dependent Variables
Total error detection score
The total error detection score was calculated by dividing the number of detections by the total
number of errors on initial naming attempts, and so is expressed as the proportion of errors that
were detected by each individual. Errors of all types—phonological, semantic, mixed, and so
forth—were considered for this measure. Total error detection scores were not considered
from participants who produced fewer than 10 errors, leaving 64 participants in the analyses
of total error detection.
Error detection
The phonological error detection score was calculated by dividing the number of detections of
phonological errors by the total number of phonological errors produced. Phonological error
detection scores were not considered from participants who produced fewer than five phono-
logical errors, leaving 58 participants in the analyses of phonological error detection.
The semantic error detection score was calculated by dividing the number of detections of
semantic errors by the total number of semantic errors produced. Semantic error detection
scores were not considered from participants who produced fewer than five semantic errors,
leaving 32 participants in the analyses of semantic error detection.
Behavioral Analysis
Statistical analyses were conducted in SPSS 25 (https://www.ibm.com/products/spss-statistics).
Outlier scores (greater or less than three interquartile distances from the median) were excluded
from analyses. This resulted in exclusion of two individuals’ word-picture matching scores.
Neurobiology of Language
325
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
1
3
0
1
8
6
7
7
3
3
n
o
_
a
_
0
0
0
1
5
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Lesion mapping of speech error detection
A series of one-way analyses of variance (ANOVAs) was first used to confirm that scores of
each test did not differ between the participants from only Cohort 1, only Cohort 2, and those
who were in both cohorts.
Bivariate correlations were used to screen for interrelationships between variables of inter-
est. Next, three multiple linear regression analyses were performed with backwards elimina-
tion of predictors with p > 0.1 to determine which behavioral scores related to detection of
each error type (all errors, phonological errors, semantic errors). Only scores that were univari-
ately correlated (uncorrected for multiple comparisons) with at least one of the dependent var-
iables were entered into these regression analyses. These scores included word-to-picture
matching, MLU, digit span difference, spatial span backwards, and spatial span difference.
To investigate whether the monitoring of different types of errors related to distinct cognitive
abilities, we performed two additional regression analyses for phonological and semantic error
detection, where detection of the other error type was included as predictor along with the
original cognitive measures. These analyses were necessarily limited to individuals who were
in both the phonological error detection and semantic error detection groups and had scores
for all behavioral predictors (N = 27).
Neuroimaging
MRI acquisition and preprocessing
Three-dimensional T1-weighted MRIs were acquired from participants on a 3.0 T Siemens Trio
scanner with the following parameters: TR = 1,900 ms; TE = 2.56 ms; flip angle = 9°; 160
contiguous 1 mm sagittal slices; field of view (FOV) = 250 × 250 mm; matrix size = 246 ×
256; voxel size 1 mm3. A T2-weighted sampling perfection with application optimized con-
trasts using a different flip angle evolution (SPACE) sequence was acquired with the following
parameters: 176 sagittal slices; slice thickness = 1.25 mm; FOV = 240 × 240 mm; matrix size =
384 × 384; TR = 3,200 ms; echo train length = 145, variable TE; variable flip angle; voxel size =
0.625 × 0.625 × 1.25 mm3.
Lesions were manually traced on coregistered T1-weighted and T2-weighted images in
native space using ITK-SNAP 3.6 (http://www.itksnap.org) by a board certified neurologist
(P.E.T.). Native space MPRAGEs and lesion tracings were warped to Montreal Neurological
Institute (MNI) space using the Clinical Toolbox Older Adult Template as the target template
(Rorden, Bonilha, Fridriksson, Bender, & Karnath, 2012) via a custom pipeline. First, brain
parenchyma was extracted from each native space image by applying a mask intended to
minimize the clipping of gray matter edges. The initial mask was generated by combining the
lesion tracing image (binarized) with white and gray matter tissue probability maps generated by
the unified segmentation procedure in SPM12 (http://picsl.upenn.edu/software/ants/) applied to
the original native space image, cost-function masked with the lesion tracing. The resulting mask
was blurred and inverted to remove nonbrain tissue from the image.
The resulting brain extracted image was then normalized using Advanced Normalization
Tools software (ANTs; http://picsl.upenn.edu/software/ants/; Avants, Tustison, & Song, 2009).
Lesion masking was used at each step of the ANTs process. After bias field correction was
applied, normalization proceeded using a typical ANTs procedure, including a rigid transform
step, an affine transform step, and a nonlinear symmetric normalization (SyN) step. Next, the
output of this initial ANTs warp was recursively submitted to three additional applications of
the SyN step. Finally, the resulting linear (rigid and affine) and four nonlinear warp fields were
concatenated, and the original native space MPRAGE and lesion tracings were transformed to
Neurobiology of Language
326
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
1
3
0
1
8
6
7
7
3
3
n
o
_
a
_
0
0
0
1
5
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Lesion mapping of speech error detection
the template space using BSpline interpolation. This iterative application of nonlinear warping was
intended to improve normalization of expanded ventricles and displaced deep structures in indi-
viduals with large lesions. The normalized lesion tracings were finally downsampled to 2.5 mm3.
Lesion-symptom mapping
We implemented SVR-LSM (Zhang, Kimberg, Coslett, Schwartz, & Wang, 2014) using a
MATLAB-based toolbox (DeMarco & Turkeltaub, 2018) running under MATLAB R2017a
(The MathWorks, Inc.). SVR-LSM was used to identify the left hemisphere areas associated
with impaired self-monitoring. SVR-LSM applies a machine learning based algorithm to find
lesion-symptom relationships more sensitively and specifically than traditional mass-univariate
lesion-symptom mapping approaches (Mah, Husain, Rees, & Nachev, 2014). Only voxels
damaged in at least 10% of the participants in the study were considered for each analysis.
Lesion volume confounds were controlled in all analyses by regressing the lesion volume out
of both behavioral scores and lesion masks, a method that provides rigorous control of lesion
volume and is more sensitive than alternative approaches (DeMarco & Turkeltaub, 2018).
Voxel-wise beta-values were thresholded at p < 0.005 using 10,000 permutations of the be-
havioral scores to generate voxel-wise null distributions. To correct for multiple comparisons,
a cluster threshold determined from the 10,000 permutation maps was applied to control the
familywise error rate at 0.05 (Mirman et al., 2018). SVR-LSM analyses were performed exam-
ining lesion locations associated with failure to detect each error type (total errors, phonolog-
ical errors, semantic errors). An additional SVR-LSM analysis for phonological error detection
was performed where MLU was added as a covariate. MRIs were not obtained on some
participants, so the sample sizes for the SVR-LSM analyses were N = 57 for total errors, N = 51
for phonological errors, and N = 29 for semantic errors.
Model quality was assessed in two ways. The first was prediction accuracy, which is a den-
sity of correlation coefficients between predicted scores and training scores across 10 replica-
tions of a 5-fold cross-validated model. The mean of this density (average correlation
coefficient) is used to summarize how well the predicted scores trend with the real scores.
However, previous work has observed that the quality of back-projected spatial patterns can-
not be assessed on the basis of prediction accuracy alone (Rasmussen et al., 2012). Indeed,
this work has observed a trade-off between model visualization reproducibility and prediction
accuracy. Therefore, a second metric produced by Rasmussen et al. (2012), pattern reproduc-
ibility index, was used to assess reproducibility of the back-projected pattern. Pattern repro-
ducibility index is calculated as a density of voxel-wise correlation coefficients computed
pairwise between 10 replicates of SVR-β maps, each generated using a random 80% of obser-
vations. Model quality measures were as follows: total detection accuracy 0.31 (SD 0.07),
reproducibility r = 0.88 (SD 0.05); phonological detection accuracy 0.25 (SD 0.07), reproduc-
ibility r = 0.89 (SD 0.04); phonological detection controlling MLU accuracy 0.26 (SD 0.11),
reproducibility r = 0.86 (SD 0.04); semantic detection accuracy 0.36 (SD 0.15), reproducibility
r = 0.75 (SD 0.12).
Region of interest analyses for semantic error detection
To compensate for the smaller sample size of the semantic error detection SVR-LSM analyses,
we selected theory-driven regions of interest (ROIs) from the Harvard Oxford Cortical atlas
(https://fsl.fmrib.ox.ac.uk/fsl/fslwiki/Atlases) to determine whether brain regions important for
auditory comprehension or speech production related to semantic error monitoring.
Selected ROIs included the posterior superior temporal gyrus, the anterior superior temporal
Neurobiology of Language
327
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
1
3
0
1
8
6
7
7
3
3
n
o
_
a
_
0
0
0
1
5
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Lesion mapping of speech error detection
gyrus, the posterior middle temporal gyrus, Heschl’s gyrus, the planum temporale, the angular
gyrus, the inferior frontal gyrus (IFG), the pars triangularis, and the middle frontal gyrus. Lesion
load for each ROI was calculated for each participant by dividing the number of voxels that
overlapped between the lesion mask and the ROI by the total number of voxels within the
ROI. Partial correlations were calculated between ROI lesion load and semantic error detec-
tion, controlling for total lesion volume.
Data availability
The data that support the findings of this study are available on request from the corresponding
author. The data are not publicly available due to their containing information that could com-
promise the privacy of research participants. Source code that was used to conduct the SVR-
LSM analyses in this study is available at https://github.com/atdemarco/svrlsmgui/.
RESULTS
Relationships of Behavioral Scores to Error Detection
Average scores across the groups are shown in Table 1. Since the data were derived from two
partially overlapping patient cohorts, we first performed a series of one-way ANOVAs to con-
firm that cohort differences did not introduce biases in the variables of interest (age, time since
stroke, education, PNT, word-to-picture matching, WAB-R auditory comprehension, MLU,
digit span forwards, digit span backwards, digit span difference, spatial span forwards, spatial
span backwards, spatial span difference, total error detection, phonological error detection,
semantic error detection). No effects of cohort (participants from only cohort 1 vs. participants
from only cohort 2 vs. individuals from both cohorts) were identified (all p > 0.05).
Bivariate correlations between behavioral variables of interest and each dependent variable
are provided in Table 2 and Table 3. Intercorrelations among behavioral variables of interest
are provided in Table 4. Phonological error detection and semantic error detection were cor-
related with one another (r = 0.570; p = 0.0012; N = 29), and this relationship persisted after
controlling for lesion volume (r = 0.502; p = 0.011; N = 23). Regression models were used to
identify predictors of error detection, first examining detection of all errors, and then semantic
and phonological errors separately. Only behavioral measures that were univariately correlated
with at least one of the dependent variables were entered into these regression models. These
measures were word-to-picture matching, MLU, digit span difference, spatial span backwards,
Table 2. Correlations between error detection and demographic variables
Age
Total error
detection
(N = 64)
0.269*
Phonological
error detection
(N = 58)
0.200
Semantic
error detection
(N = 32)
−0.183
Time since stroke
−0.035
−0.044
Education
0.211
0.101
0.080
−0.001
Note. ***p < 0.05, after Bonferroni correction for 9 comparisons, **p < 0.01 uncorrected, *p < 0.05 uncorrected.
Neurobiology of Language
328
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
1
3
0
1
8
6
7
7
3
3
n
o
_
a
_
0
0
0
1
5
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Lesion mapping of speech error detection
Table 3. Correlations between error detection and other variables
PNT
Total error detection
0.027 (N = 64)
WAB-R auditory comprehension
0.115 (N = 62)
Word-to-picture matching
0.302* (N = 62)
Phonological error detection
−0.063 (N = 58)
0.022 (N = 56)
0.244 (N = 56)
Mean length of utterance
0.514*** (N = 59)
0.502*** (N = 54)
Digit span forwards
Digit span backwards
Digit span difference
Spatial span forwards
Spatial span backwards
Spatial span difference
−0.074 (N = 59)
0.148 (N = 59)
−0.226 (N = 59)
0.096 (N = 59)
0.327** (N = 59)
−0.329** (N = 59)
−0.210 (N = 54)
0.084 (N = 54)
−0.340** (N = 54)
0.022 (N = 54)
0.276* (N = 54)
−0.352* (N = 54)
Semantic error detection
0.079 (N = 32)
0.139 (N = 32)
0.523** (N = 31)
0.365 (N = 31)
0.033 (N = 31)
0.082 (N = 31)
−0.023 (N = 31)
0.265 (N = 31)
0.377* (N = 31)
−0.171 (N = 31)
Note. Sample sizes differ due to missing scores. WAB-R = Western Aphasia Battery-Revised. ***p < 0.05 after Bonferroni correction for 33 comparisons, **p < 0.01
uncorrected, *p < 0.05 uncorrected.
and spatial span difference. Total error detection was predicted by MLU (standardized β =
0.380; p = 0.002; variance inflation factor [VIF] = 1.025) and spatial span difference (standard-
ized β = −0.333; p = 0.006; VIF = 1.025) with an overall adjusted R2 of 0.268, F(2, 54) = 11.3,
p < 0.001. Phonological error detection was predicted by MLU (standardized β = 0.428; p =
0.001; VIF = 1.031), digit span difference (standardized β = −0.249; p = 0.04; VIF = 1.061),
and spatial span difference (standardized β = −0.255; p = 0.038; VIF = 1.091) with an overall
adjusted R2 of 0.334, F(3, 48) = 9.54; p < 0.001. Semantic error detection was predicted only by
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
1
3
0
1
8
6
7
7
3
3
n
o
_
a
_
0
0
0
1
5
p
d
.
l
Table 4. Correlations between behavioral measures
PNT
WAB-R
Yes/No
0.277*
Word-to-picture
matching
0.679***
MLU
Digit
span fwd
0.491*** 0.562***
Digit
span bwd
0.529*** 0.313*
Digit
span diff
Spatial
span fwd
0.314*
Spatial
span bwd
0.253*
Spatial
span diff
0.055
WAB-R Yes/No
0.337**
0.332*
0.471***
0.304*
0.374**
0.140
0.032
0.137
Word-to-picture matching
0.456*** 0.340**
0.440*** 0.079
0.401***
0.416*** −0.069
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
MLU
Digit span fwd
Digit span bwd
Digit span diff
Spatial span fwd
Spatial span bwd
0.474***
0.626*** 0.092
0.190
0.252
−0.104
0.680*** 0.780*** 0.379**
0.261*
0.128
0.071
0.474***
0.490*** −0.065
0.110
−0.063
0.230
0.723***
0.292*
−0.450***
Note. Correlations are performed for the full sample (N = 64). Sample size varies between 57 and 64 on individual correlations because of missing values on
some tests. PNT = Philadelphia Naming Test, WAB-R = Western Aphasia Battery-Revised, MLU = mean length of utterance, fwd = forwards, bwd = backwards,
diff = difference. ***p < 0.05 after Bonferroni correction for 33 comparisons, **p < 0.01 uncorrected, *p < 0.05 uncorrected.
Neurobiology of Language
329
Lesion mapping of speech error detection
word-to-picture matching (standardized β = 0.516; p = 0.003; VIF = 1.000) with an overall ad-
justed R2 of 0.240, F(1, 28) = 10.173, p = 0.003. These results were mostly preserved when
reanalyzed using error detection performance based only on the 60 PNT items administered
to all participants (see the online supporting information for this article).
To determine whether the behavioral predictors of phonological and semantic error detec-
tion were specific to the monitoring of those respective error types, we performed two addi-
tional regression analyses for phonological and semantic error detection, where detection of
the other error type was included as a predictor along with the original cognitive measures.
Phonological error detection was predicted by semantic error detection (standardized β =
0.587; p = 0.001; VIF = 1.23), spatial span difference (standardized β = −0.321; p = 0.039;
VIF = 1.02), spatial span backwards (standardized β = −0.336; p = 0.047; VIF = 1.23), and
MLU (standardized β = 0.283; p = 0.071; VIF = 1.08) with an overall adjusted R2 of 0.461,
F(4, 22) = 6.56; p = 0.001. Semantic error detection was predicted by phonological error
detection (standardized β = 0.392; p = 0.018; VIF = 1.14), and word-to-picture matching (stan-
dardized β = 0.464; p = 0.006; VIF = 1.14) with an overall adjusted R2 of 0.455, F(2, 24) =
11.8; p < 0.001.
Lesion-Symptom Mapping
Lesion overlap maps demonstrated good coverage of the left middle cerebral artery territory for
all analyses (Figure 1). Lesion volume correlated modestly with total error detection (r =
−0.278; p = 0.04), and a trending relationship was observed with phonological error detection
(r = −0.243; p = 0.09) and semantic error detection (r = −0.31; p = 0.10). Lesion volume was
subsequently controlled for in all SVR-LSM analyses. SVR-LSM analyses demonstrated that de-
creased total error detection mapped onto lesions in a large region of frontal white matter and
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
1
3
0
1
8
6
7
7
3
3
n
o
_
a
_
0
0
0
1
5
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
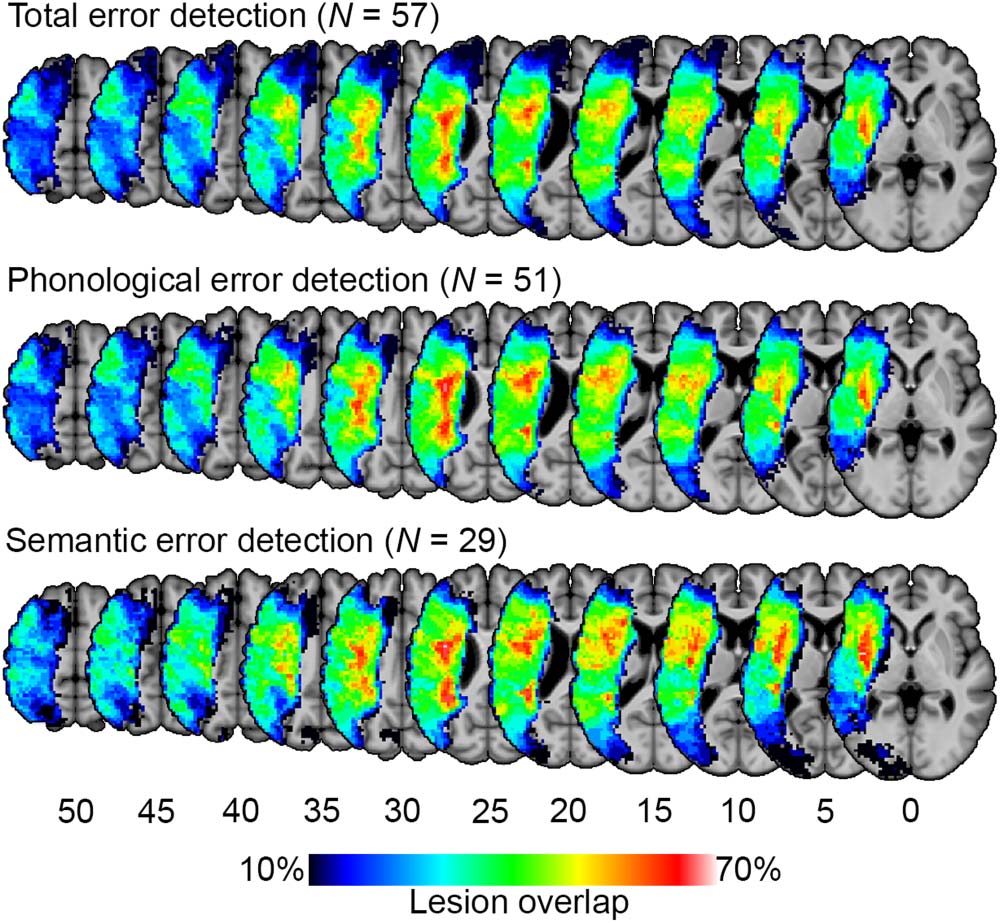
Figure 1.
Lesion overlap maps for study participants. Lesion-symptom mapping analyses were lim-
ited to voxels that were lesioned in at least 10% of participants (i.e., at least six participants for total
and phonological error detection, at least three for semantic error detection).
Neurobiology of Language
330
Lesion mapping of speech error detection
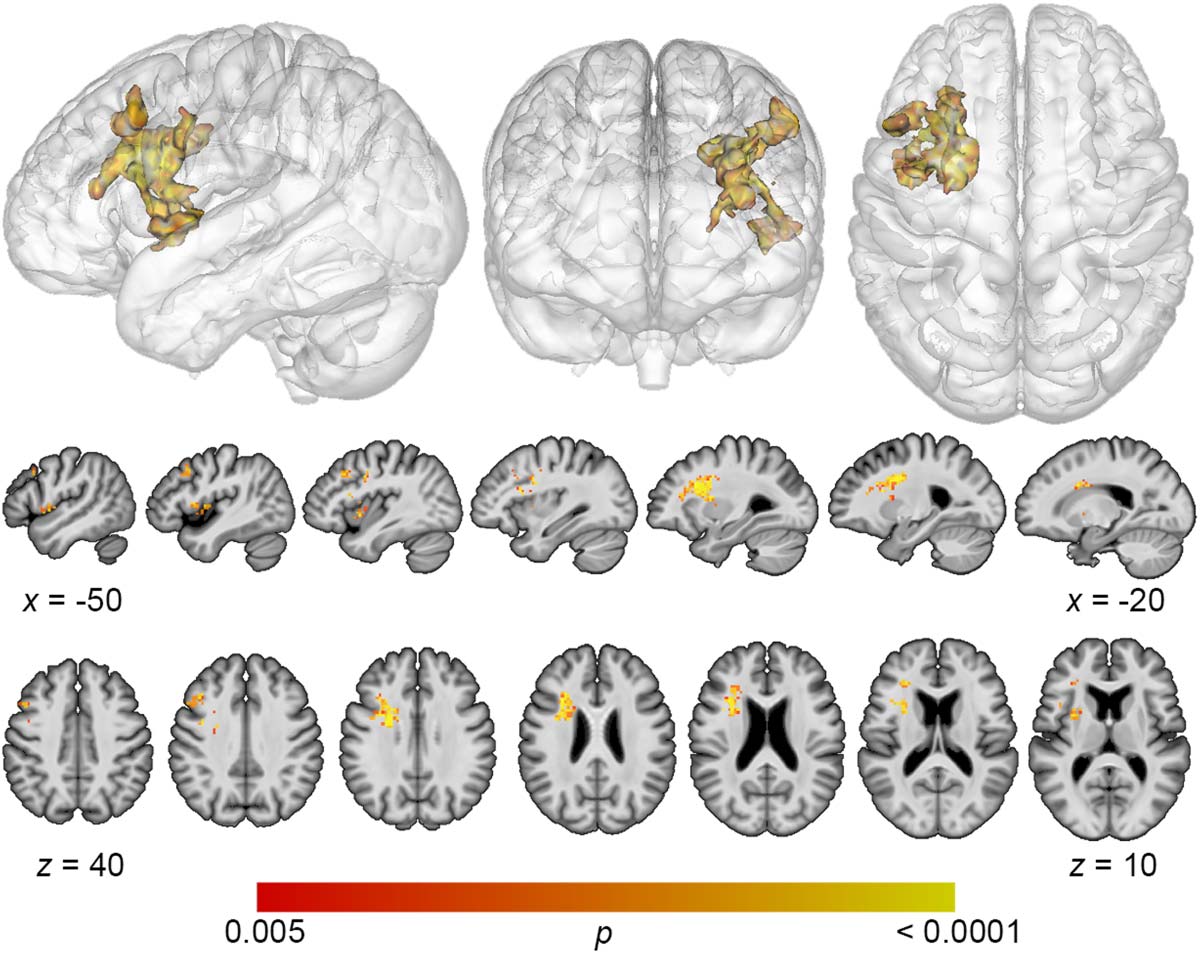
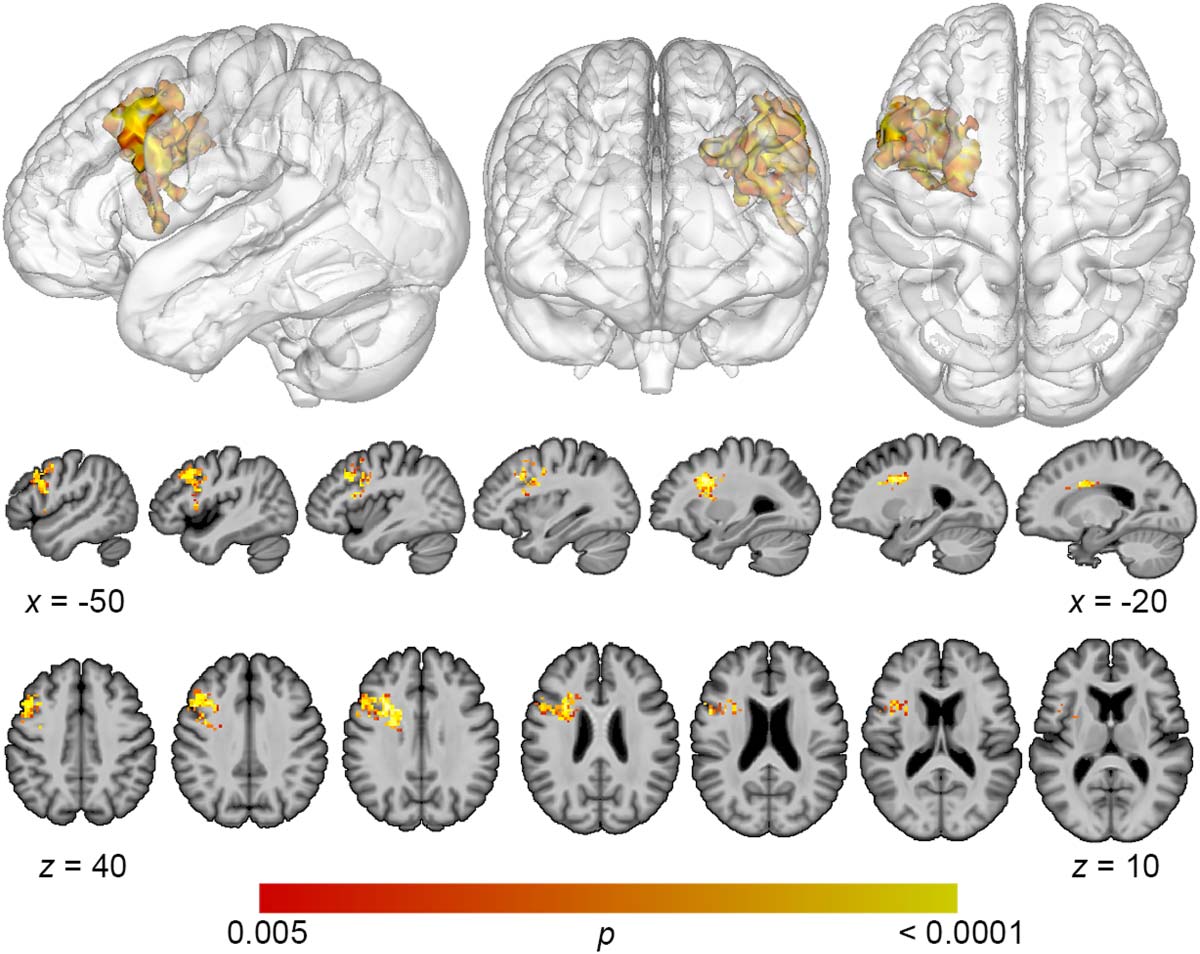
Figure 2. Multivariate lesion-symptom mapping results for total error detection. Support vector
regression lesion-symptom mapping analysis demonstrated that decreased detection of all error
types was associated with damage to a large region of frontal white matter and dorsolateral prefrontal
cortex (voxel-wise p < 0.005, cluster-level familywise error < 0.05).
the dorsolateral prefrontal cortex (DLPFC) (Figure 2). A single cluster was identified with a
volume of 7,156 mm3, with center of mass at MNI coordinates −32.7, 8.7, 20.5. Decreased
detection of phonological errors related to damage to the same regions as total error detection,
but the relationship was stronger, resulting in a larger cluster of significant lesion-deficit asso-
ciation (8,046 mm3), centered at −31.8, 9.4, 26.1 (Figure 3). The stronger lesion-behavior re-
lationship observed for phonological error detection suggests that the localization of total error
detection was driven primarily by phonological error detection. To confirm that the results did
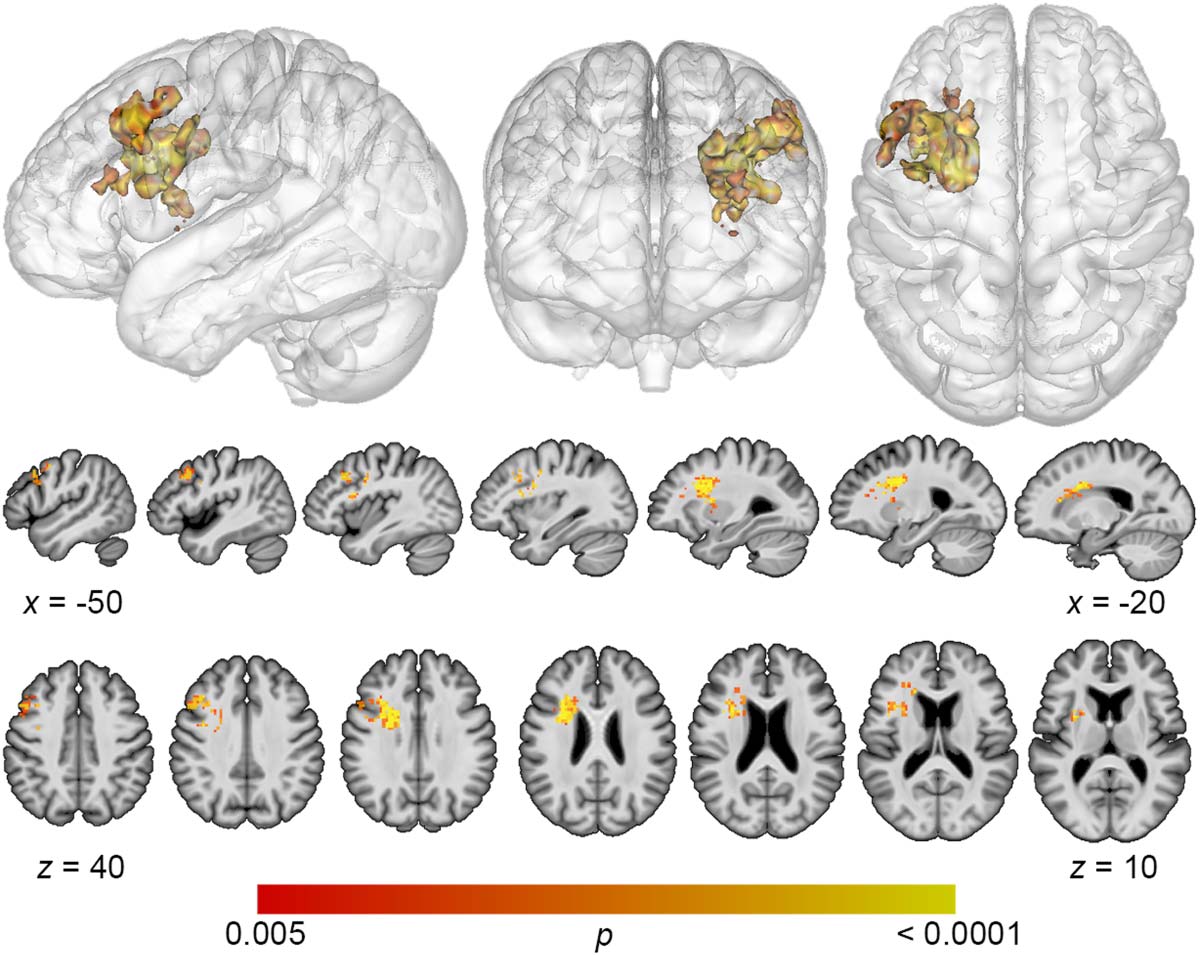
not simply reflect reduced speech output among individuals with frontal white matter damage,
we added MLU as a covariate to our SVR-LSM analysis of phonological error detection. The
result remained significant after controlling for fluency and centered more closely to the
DLPFC. A single cluster was identified with a volume of 9,641 mm3, and center of mass at
MNI coordinates −37.3, 9.3, 28.7 (Figure 4). No significant clusters were identified in the
SVR-LSM analysis of semantic error detection (see Figure 1 in the online supporting informa-
tion). These results did not change substantially when reanalyzed using error detection perfor-
mance based only on the 60 PNT items administered to all participants (see the online
supporting information).
ROI Analyses for Semantic Error Detection
In light of the reduced sample size for the SVR-LSM analysis of semantic error detection, theory-
driven ROIs were selected to determine the association of semantic error detection with lesions
in specific brain regions involved in auditory comprehension, lexical processing, word produc-
tion, and executive function (Table 5). The results of all ROI analyses were null, even at an
uncorrected statistical threshold.
Neurobiology of Language
331
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
1
3
0
1
8
6
7
7
3
3
n
o
_
a
_
0
0
0
1
5
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Lesion mapping of speech error detection
Figure 3. Multivariate lesion-symptom mapping results for phonological error detection. Reduced
detection of phonological errors related to damage to frontal white matter and dorsolateral prefron-
tal cortex (voxel-wise p < 0.005, cluster-level familywise error < 0.05).
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
1
3
0
1
8
6
7
7
3
3
n
o
_
a
_
0
0
0
1
5
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. Multivariate lesion-symptom mapping results for phonological error detection, controlling
for fluency. After adding mean length of utterance as a covariate, reduced detection of phonological
errors related to damage to dorsolateral prefrontal cortex (voxel-wise p < 0.005, cluster-level familywise
error < 0.05).
Neurobiology of Language
332
Lesion mapping of speech error detection
Table 5. Results of region of interest analyses
Anterior superior temporal gyrus
Posterior superior temporal gyrus
Posterior middle temporal gyrus
Heschl’s gyrus
Planum temporale
Angular gyrus
Inferior frontal gyrus
Middle frontal gyrus
Semantic error detection (N = 29)
0.035
0.298
0.153
0.126
0.208
0.297
−0.065
−0.167
Note. Partial correlations between region of interest (ROI) lesion load and semantic error detection, controlling for
total lesion volume. Negative values indicate a relationship between ROI damage and behavioral impairment.
***p < 0.05 after Bonferroni correction for 8 comparisons, **p < 0.01 uncorrected, *p < 0.05 uncorrected.
DISCUSSION
In this study, we examined the behavioral and lesion correlates of impaired detection of
speech errors in people with aphasia. We found that reduced detection of phonological errors
was associated with low fluency and low performance on measures of executive control.
Reduced semantic error detection related to low performance on a test of word-level compre-
hension ability. Finally, we found that damage to frontal white matter and the DLPFC was
associated with poor error detection. Overall, these findings provide valuable evidence regard-
ing the neural and behavioral underpinnings of speech error monitoring, and suggest that def-
icits in monitoring of phonological and semantic errors relate to somewhat different mental
processes. Findings are consistent with aspects of both production- and comprehension-based
models of error monitoring.
Support for Production-Based Error Monitoring Models
Fluency marks successful speech production, and in this sense, an association between fluency
and phonological error detection supports production-based monitors in general. A more pre-
cise account of this relationship is less clear. The conflict model of Nozari and colleagues (2011)
predicts phonological error monitoring to decrease as a result of higher noise in the system re-
sponsible for the activation of phonological nodes. Low fluency could reflect increased noise in
the phonological system causing delays in retrieval, but this interpretation is not consistent with
our lesion-symptom mapping result, given that phonological error production has been asso-
ciated with lesions of the parietal lobe (Mirman et al., 2015; Schwartz, Faseyitan, Kim, &
Coslett, 2012). Alternatively, since both fluency and phonological errors are associated with
deficits in postlexical speech production (Nickels & Howard, 1995; Romani, Olson,
Semenza, & Granà, 2002; Schwartz, Wilshire, Gagnon, & Polansky, 2004; Wilshire, 2002),
the association between fluency and detection of phonological errors may imply that these
errors are detected during postlexical speech production. Although the Nozari conflict
model does not include postlexical speech production stages, many production-based
models of error monitoring suggest that monitoring also occurs at these stages (see Postma,
2000 for a review).
Neurobiology of Language
333
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
1
3
0
1
8
6
7
7
3
3
n
o
_
a
_
0
0
0
1
5
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Lesion mapping of speech error detection
Our lesion-symptom mapping results are also consistent with models suggesting that error
detection relies on speech production processes. Reduced error detection related to damage to
a large region of frontal white matter underlying areas associated with speech production
(Price, 2010). Prior studies have indicated that white matter tracts in this region are responsible
for supporting fluent speech (Catani et al., 2013; Mandelli et al., 2014), which is consistent
with our behavioral findings. A recent fMRI study in healthy participants also provided support
for a speech monitor dependent on frontal regions. Gauvin and colleagues (2015) found that
self-monitoring elicits activity across several areas involved in speech production, such as the
presupplementary motor area (pre-SMA) and IFG (Gauvin et al., 2015). Our results add causal
evidence that frontal white matter tracts are involved in successful detection of speech errors.
Recent work has implicated the left frontal aslant tract (FAT), which connects left IFG to
pre-SMA and SMA, in the initiating, sequencing, and stopping of language production
(Catani et al., 2013; Dick, Garic, Graziano, & Tremblay, 2018). Given that the white matter
implicated in our study was adjacent to the IFG, it is logical to ask whether the FAT is involved
in self-monitoring. However, a number of white matter tracts run through the areas implicated
here, including tracts connecting medial and lateral frontal regions such as the FAT, tracts con-
necting subcortical structures and frontal regions, and tracts connecting frontal regions with
temporoparietal areas. A future diffusion tensor imaging study could clarify the specific white
matter connections that are essential for proper speech error monitoring.
The Role of Executive Control in Speech Error Monitoring
Considerable debate centers on whether the monitor used to detect one’s own speech errors is
domain-general or language-specific. The perceptual loop model predicts the monitor to be
specific to language since it should depend on the speech comprehension system (Levelt,
1983; Postma, 2000). The conflict-based model of Nozari and colleagues (2011) is a domain-
general model in that conflict signals arising during naming are received by a frontal brain
structure (namely, the ACC), which monitors errors produced in other cognitive domains as
well (Nozari et al., 2011). Domain-general theories of error monitoring predict successful
detection to rely on executive functioning, regardless of the type of error being detected
(Botvinick et al., 2001; Yeung et al., 2004). In support of this prediction, we identified a rela-
tionship between two measures of executive function (digit span difference and spatial span
difference) and the monitoring of phonological errors. However, in contrast to the predictions
of a domain-general theory, we did not find an association between these measures and semantic
error detection. The finding that the monitoring of different types of errors relates to different
behavioral measures may be interpreted as evidence against a domain-general model. However,
it is also worth noting that we found a correlation between phonological error detection and
semantic error detection, implying that some overlapping processes underlie monitoring of
both error types. We also found some support for the anatomical predictions of a domain-
general model for error monitoring, at least in the analysis of phonological errors. Reduced
error detection related to damage to the DLPFC, a region known to be associated with working
memory and domain-general executive function (Miller, 2001; Miller & Cohen, 2001).
Nozari and colleagues' conflict-based model (2011) predicts the ACC to be involved in error
monitoring, and prior evidence in healthy participants indicates that internal speech monitoring
recruits ACC activity (Gauvin et al., 2015). Frontal white matter lesions may disrupt connectivity
to the ACC, resulting in impaired error detection (Hogan, Vargha-Khadem, Saunders, Kirkham,
& Baldeweg, 2006), but since we lacked lesion coverage of the ACC in our participant group,
we cannot provide evidence for or against a causal role for this region in speech error
Neurobiology of Language
334
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
1
3
0
1
8
6
7
7
3
3
n
o
_
a
_
0
0
0
1
5
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Lesion mapping of speech error detection
monitoring. Future work using connectome-based lesion methods that can detect disconnected
regions outside the lesioned area (Gleichgerrcht, Fridriksson, Rorden, & Bonilha, 2017;
Yourganov, Fridriksson, Rorden, Gleichgerrcht, & Bonilha, 2016) will be useful to examine this
issue further.
It is worth considering whether our tasks for executive control–that is, those designed to
detect differences between forwards and backwards digit/spatial span–appropriately measure
the type of control required to resolve conflict within the framework of conflict monitoring
theory. Within the conflict monitoring framework, top-down control is required to override
a habitual response (e.g., the habitual response to read the word instead of name the color
in the Stroop task) and is provided by the DLPFC (MacDonald, Cohen, Stenger, & Carter,
2000; Yeung, 2015). This type of control is likely at play when completing the backwards digit
or spatial span task, since the habitual response of repeating a sequence in the same order as it
was presented must be overridden and replaced with a manipulation of that sequence in the
reverse direction. Subtracting the forwards digit/spatial span score from the backwards score
adjusts for the demands on maintenance in short-term memory required for both tasks, isolat-
ing the executive control component in the resulting measure. However, other factors could
also contribute to a difference in performance on the forwards versus backwards digit/spatial
span tasks. Some evidence indicates that separate mechanisms, one based on refreshing and
another based on rehearsal, are responsible for maintaining short-term memory during low
demand tasks like forwards digit span on the one hand and high demand tasks like backwards
digit span on the other (Baddeley, 2003; Camos, Lagner, & Barrouillet, 2009; Camos, Mora, &
Oberauer, 2011; Ghaleh et al., 2019). Therefore, a selective deficit in backwards but not for-
wards digit span could reflect an impairment in systems that support the articulatory rehearsal
of phonological information (i.e., the phonological loop; Ghaleh et al., 2019), and it could be
this aspect of the digit span difference score that is associated with poor error detection. The
finding that the difference between backwards and forwards spatial span also related to pho-
nological error detection makes this interpretation less likely, assuming that rehearsal for back-
wards spatial span relies on a visuospatial sketchpad rather than the phonological loop.
Another area of less clarity is the null relationship between semantic error detection and mea-
sures of executive function. The null finding is inconsistent with a domain-general model, but
it may simply reflect reduced power for the semantic error detection analyses. Alternatively,
our behavioral measures could be less sensitive to the type of control necessary for manipu-
lation of semantic information. Future work comparing error detection ability with perfor-
mance on a wide range of executive function and working memory tasks could clarify
which cognitive abilities are required to successfully monitor speech errors.
Mechanisms of Semantic Error Detection
Our finding that the detection of semantic errors relies on word-level comprehension is con-
sistent with comprehension-based models of monitoring that suggest that monitoring impair-
ments follow an inability to comprehend one’s own speech (Levelt, 1983). While prior studies
did not find a relationship between auditory comprehension and error monitoring (Marshall
et al., 1998; Marshall et al., 1985; Nickels & Howard, 1995), those studies investigated
phonological, not semantic errors. A role for comprehension in semantic error monitoring
would account for evidence of poor semantic error detection among patients with severe
comprehension deficits (Marshall et al., 1985).
However, some evidence suggests that both word comprehension and word production
rely on the same lexical-semantic representations (Foygel, 2000; Hanley & Nickels, 2009).
Neurobiology of Language
335
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
1
3
0
1
8
6
7
7
3
3
n
o
_
a
_
0
0
0
1
5
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Lesion mapping of speech error detection
An alternative interpretation of the word-to-picture matching measure is as an indicator of
lexical-semantic access ability, which is required for naming as well as comprehension
(Hanley & Nickels, 2009). Consistent with this interpretation, word-picture matching scores
and naming (PNT) scores correlated more strongly with each other than either did with any
other measure examined in our participants (Table 4). Semantic errors occur primarily during
the lexical-semantic stage of word production due to a failure to activate the correct lemma
from semantics (Foygel, 2000; Schwartz et al., 2009). As noted above, according to the
conflict-based model, damage to lexical-semantic access is expected to result in high levels
of conflict that cause difficulty detecting semantic errors (Nozari et al., 2011). Thus, the relation-
ship between word-picture matching and semantic error detection could be viewed as con-
sistent with this model.
While lesion-behavior associations in the SVR-LSM or ROI analyses might have helped
clarify the mechanisms by which semantic errors are monitored, the null findings of these anal-
yses proved unhelpful. These analyses were limited by the small sample size available.
Additional studies with larger populations and more specific behavioral measures will be
needed to identify the neural and behavioral correlates of semantic error detection.
Limitations
In order to capture the reflexive aspect of natural error detection, we did not give participants
explicit instructions to comment on the accuracy of their naming attempts, consistent with past
studies of self-monitoring in people with aphasia (Nozari et al., 2011; Schuchard et al., 2017;
Schwartz et al., 2016). However, it is possible that some patients were aware of the errors they
were making, but chose not to comment on them. While in theory this issue could be ad-
dressed using a paradigm in which participants are required to report their accuracy following
each naming trial, this has the potential to alter the way they approach error monitoring. We
also note that combining the two cohorts of patients was necessary to achieve an adequate
sample size, and behavioral measures examined here were selected as the best measures
available in both cohorts. Additional measures, for instance examining nonverbal semantics,
speech perception, or motor speech production, would have been informative but were not
available in both cohorts. Future prospective studies of error monitoring should select mea-
sures that allow a more precise and comprehensive delineation of the behavioral correlates
of error detection.
Conclusion
Gaining a more detailed understanding of the brain and behavioral basis of speech self-monitoring
may lead to new treatments aimed at improving awareness of errors and self-correction in
aphasia. These results demonstrate that monitoring of different error types relies on distinct
cognitive functions, and provide causal evidence for the importance of frontal white matter
tracts and DLPFC for self-monitoring of speech. These findings substantially inform the debate
regarding the neural and behavioral underpinnings of speech error monitoring.
FUNDING INFORMATION
Peter E. Turkeltaub, National Center for Advancing Translational Sciences (http://dx.doi.org/
10.13039/100006108), Award ID: KL2TR000102. Peter E. Turkeltaub, Doris Duke
Charitable Foundation (http://dx.doi.org/10.13039/100000862), Award ID: 2012062. Peter
E. Turkeltaub, National Institute on Deafness and Other Communication Disorders (http://
dx.doi.org/10.13039/100000055), Award ID: R03DC014310. Peter E. Turkeltaub, National
Neurobiology of Language
336
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
1
3
0
1
8
6
7
7
3
3
n
o
_
a
_
0
0
0
1
5
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Lesion mapping of speech error detection
Institute on Deafness and Other Communication Disorders (http://dx.doi.org/10.13039/
100000055), Award ID: R01DC014960. Mackenzie E. Fama, National Institute on Deafness
and Other Communication Disorders (http://dx.doi.org/10.13039/100000055), Award ID:
F31DC014875. Andrew T. DeMarco, National Center for Advancing Translational Sciences
(http://dx.doi.org/10.13039/100006108), Award ID: TL1TR001431.
AUTHOR CONTRIBUTIONS
Ayan S. Mandal contributed to the study concept, acquisition, and analysis, and the interpre-
tation of data, and drafted the manuscript. Mackenzie E. Fama contributed to the acquisition of
data, and the study concept and design. Laura M. Skipper-Kallal contributed to the interpre-
tation of data and edited the manuscript. Andrew T. DeMarco contributed to the analysis of
data and edited the manuscript. Elizabeth H. Lacey contributed to the acquisition of data, and
the study concept and design. Peter E. Turkeltaub contributed to the study concept and design,
and the interpretation of data, and edited the manuscript. All authors reviewed and commented
on the final version of the manuscript.
REFERENCES
Avants, B. B., Tustison, N., & Song, G. (2009). Advanced normali-
zation tools (ANTS). Insight Journal, 1–35.
Baddeley, A. (2003). Working memory: Looking back and looking
forward. Nature Reviews Neuroscience, 4, 829–839.
Botvinick, M. M., Braver, T. S., Barch, D. M., Carter, C. S., &
Cohen, J. D. (2001). Conflict monitoring and cognitive control.
Psychological Review, 108(3), 624–652. https://doi.org/
10.1037/0033-295X.108.3.624
Butterworth, B., & Howard, D. (1987). Paragrammatisms. Cognition,
26(1), 1–37.
Camos, V., Lagner, P., & Barrouillet, P. (2009). Two maintenance
mechanisms of verbal information in working memory. Journal of
Memory and Language, 61(3), 457–469.
Camos, V., Mora, G., & Oberauer, K. (2011). Adaptive choice
between articulatory rehearsal and attentional refreshing in verbal
working memory. Memory and Cognition, 39(2), 231–44.
Catani, M., Mesulam, M. M., Jakobsen, E., Malik, F., Martersteck,
A., Wieneke, C., … Rogalski, E. (2013). A novel frontal pathway
underlies verbal fluency in primary progressive aphasia. Brain,
136(8), 2619–2628.
Corsi, P. M. (1972). Human memory and the medial temporal re-
gion of the brain (Doctoral dissertation). Dissertation Abstract
International, 34(02), 819B.
DeMarco, A. T., & Turkeltaub, P. E. (2018). A multivariate lesion-
symptom mapping toolbox and examination of lesion-volume
biases and correction methods in lesion-symptom mapping.
Human Brain Mapping, 39(11), 4169–4182.
Dick, A. S., Garic, D., Graziano, P., & Tremblay, P. (2018). The
frontal aslant tract (FAT) and its role in speech, language and ex-
ecutive function. Cortex, 111, 148–163. https://doi.org/10.1016/
j.cortex.2018.10.015.
Foygel, D. (2000). Models of impaired lexical access in speech pro-
duction. Journal of Memory and Language, 43(2), 182–216.
Gauvin, H. S., De Baene, W., Brass, M., & Hartsuiker, R. J. (2015).
Conflict monitoring in speech processing: An fMRI study of error
detection in speech production and perception. NeuroImage,
126, 96–105.
Ghaleh, M., Lacey, E. H., Fama, M. E., Anbari, Z., DeMarco, A. T.,
& Turkeltaub, P. E. (2019). Dissociable mechanisms of verbal
working memory revealed through multivariate lesion mapping.
Cerebral Cortex, 30(4), 2542–2554.
Gleichgerrcht, E., Fridriksson, J., Rorden, C., & Bonilha, L. (2017).
Connectome-based lesion-symptom mapping (CLSM): A novel
approach to map neurological function. NeuroImage: Clinical,
16, 461–467.
Goodglass, H., Kaplan, E., & Barresi, B. (2001). The assessment of
aphasia and related disorders. Philadelphia: Lippincott Williams
& Wilkins.
Hanley, J. R., Cortis, C., Budd, M. J., & Nozari, N. (2016). Did I say
dog or cat? A study of semantic error detection and correction
in children. Journal of Experimental Child Psychology, 142,
36–47.
Hanley, J. R., & Nickels, L. (2009). Are the same phoneme and lex-
ical layers used in speech production and comprehension? A
case-series test of Foygel and Dell’s (2000) model of aphasic
speech production. Cortex, 45(6), 784–790.
Hillis, A. E., Rorden, C., & Fridriksson, J. (2017). Brain regions es-
sential for word comprehension: Drawing inferences from pa-
tients. Annals of Neurology, 81(6), 759–768.
Hogan, A. M., Vargha-Khadem, F., Saunders, D. E., Kirkham, F. J.,
& Baldeweg, T. (2006). Impact of frontal white matter lesions on
performance monitoring: ERP evidence for cortical disconnec-
tion. Brain, 129(8), 2177–2188.
Indefrey, P., & Levelt, W. J. M. (2004). The spatial and temporal
signatures of word production components. Cognition, 93(1–2),
101–144.
Kertesz, A. (2007). The western aphasia battery—revised. San
Antonio, TX: Pearson.
Levelt, W. J. M. (1983). Monitoring and self-repair in speech.
Cognition, 14(1), 41–104.
Liss, J. M. (1998). Error-revision in the spontaneous speech of
apraxic speakers. Brain and Language, 62(3), 342–360.
MacDonald, A. W., Cohen, J. D., Stenger, V. A., & Carter, C. S.
(2000). Dissociating the role of the dorsolateral prefrontal and
anterior cingulate cortex in cognitive control. Science, 288(5472),
1835–1838.
Mah, Y. H., Husain, M., Rees, G., & Nachev, P. (2014). Human brain
lesion-deficit inference remapped. Brain, 137(9), 2522–2531.
Neurobiology of Language
337
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
1
3
0
1
8
6
7
7
3
3
n
o
_
a
_
0
0
0
1
5
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Lesion mapping of speech error detection
Mandelli, M. L., Caverzasi, E., Binney, R. J., Henry, M. L., Lobach,
I., Block, N., … Gorno-Tempini, M. L. (2014). Frontal white mat-
ter tracts sustaining speech production in primary progressive
aphasia. Journal of Neuroscience, 34(29), 9754–9767.
Marshall, J., Robson, J., Pring, T., & Chiat, S. (1998). Why does
monitoring fail in jargon aphasia? Comprehension, judgment,
and therapy evidence. Brain and Language, 63(1), 79–107.
Marshall, R. C., Neuburger, S. I., & Phillips, D. S. (1994). Verbal
self-correction and improvement in treated aphasic clients.
Aphasiology, 8(6), 535–547.
Marshall, R. C., Rappaport, B. Z., & Garcia-Bunuel, L. (1985). Self-
monitoring behavior in a case of severe auditory agnosia with
aphasia. Brain and Language, 24(2), 297–313.
Miller, E. K. (2001). The prefrontal cortex and cognitive control.
Nature Reviews Neuroscience, 1(1), 59–65.
Miller, E. K., & Cohen, J. D. (2001). An integrative theory of prefrontal
cortex function. Annual Review of Neuroscience, 24(1), 167–202.
Mirman, D., Chen, Q., Zhang, Y., Wang, Z., Faseyitan, O. K.,
Coslett, H. B., & Schwartz, M. F. (2015). Neural organization
of spoken language revealed by lesion–symptom mapping.
Nature Communications, 6, 6762.
Mirman, D., Landrigan, J. F., Kokolis, S., Verillo, S., Ferrara, C., &
Pustina, D. (2018). Corrections for multiple comparisons in
voxel-based lesion-symptom mapping. Neuropsychologia,
115, 112–123.
Nickels, L., & Howard, D. (1995). Phonological errors in aphasic
naming: Comprehension, monitoring and lexicality. Cortex, 31(2),
209–237.
Nozari, N., Dell, G. S., & Schwartz, M. F. (2011). Is comprehension
necessary for error detection? A conflict-based account of mon-
itoring in speech production. Cognitive Psychology, 63(1), 1–33.
Postma, A. (2000). Detection of errors during speech production: A
review of speech monitoring models. Cognition, 77(2), 97–132.
Price, C. J. (2010). The anatomy of language: A review of 100 fMRI
studies published in 2009. Annals of the New York Academy of
Sciences, 1191, 62–88.
Rasmussen, P. M., Schmah, T., Madsen, K. H., Lund, T. E.,
Yourganov, G., Strother, S. C., & Hansen, L. K. (2012).
Visualization of nonlinear classification models in neuroimaging:
Signed sensitivity maps. In BIOSIGNALS 2012 - Proceedings of
the International Conference on Bio-Inspired Systems and
Signal Processing (pp. 254–263).
Roach, A., Schwartz, M. F., Martin, N., Grewal, A. S., & Brecher, A.
(1996). The Philadelphia Naming Test: Scoring and rationale.
Clinical Aphasiology, 24, 21–133.
Romani, C., Olson, A., Semenza, C., & Granà, A. (2002). Patterns
of phonological errors as a function of a phonological versus an
articulatory locus of impairment. Cortex, 38(4), 541–567.
Rorden, C., Bonilha, L., Fridriksson, J., Bender, B., & Karnath, H. O.
(2012). Age-specific CT and MRI templates for spatial normaliza-
tion. NeuroImage, 61(4), 957–965.
Schuchard, J., Middleton, E. L., & Schwartz, M. F. (2017). The tim-
ing of spontaneous detection and repair of naming errors in
aphasia. Cortex, 93, 79–91.
Schwartz, M. F., Faseyitan, O., Kim, J., & Coslett, H. B. (2012). The
dorsal stream contribution to phonological retrieval in object
naming. Brain, 135(12), 3799–3814.
Schwartz, M. F., Kimberg, D. Y., Walker, G. M., Faseyitan, O.,
Brecher, A., Dell, G. S., & Coslett, H. B. (2009). Anterior temporal
involvement in semantic word retrieval: Voxel-based lesion-
symptom mapping evidence from aphasia. Brain, 132(12),
3411–3427.
Schwartz, M. F., Middleton, E. L., Brecher, A., Gagliardi, M., &
Garvey, K. (2016). Does naming accuracy improve through
self-monitoring of errors? Neuropsychologia, 84, 272–281.
Schwartz, M. F., Wilshire, C. E., Gagnon, D. A., & Polansky, M.
(2004). Origins of nonword phonological errors in aphasic pic-
ture naming. Cognitive Neuropsychology, 21(2), 159–186.
Stark, J. A. (1988). Aspects of automatic versus controlled process-
ing, monitoring, metalinguistic tasks, and related phenomena in
aphasia. In W. U. Dressler & J. A. Stark (Eds.), Linguistic analyses
of aphasic language (pp. 179–223). New York: Springer.
Wilshire, C. E. (2002). Where do aphasic phonological errors come
from? Evidence from phoneme movement errors in picture nam-
ing. Aphasiology, 16(1–2), 169–197.
Yeung, Nicholas. (2015). Conflict monitoring and cognitive con-
trol. In K. N. Ochsner & S. Kossslyn (Eds.), Oxford handbook
of cognitive neuroscience, Volume 2: The cutting edges.
Oxford: Oxford University Press.
Yeung, Nick, Botvinick, M. M., & Cohen, J. D. (2004). The neural
basis of error detection: Conflict monitoring and the error-related
negativity. Psychological Review, 111(4), 931–959.
Yourganov, G., Fridriksson, J., Rorden, C., Gleichgerrcht, E., &
Bonilha, L. (2016). Multivariate connectome-based symptom
mapping in post-stroke patients: Networks supporting language
and speech. Journal of Neuroscience, 36(25), 6668–6679.
Zhang, Y., Kimberg, D. Y., Coslett, H. B., Schwartz, M. F., &
Wang, Z. (2014). Multivariate lesion-symptom mapping using
support vector regression. Human Brain Mapping, 35(12),
5861–5876.
Neurobiology of Language
338
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
1
3
0
1
8
6
7
7
3
3
n
o
_
a
_
0
0
0
1
5
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3