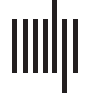RESEARCH ARTICLE
Lexical Access in Naming and Reading:
Spatiotemporal Localization of Semantic
Facilitation and Interference Using MEG
a n o p e n a c c e s s
j o u r n a l
Julien Dirani1
and Liina Pylkkänen2
1New York University
2New York University and New York University Abu Dhabi
Keywords: lexical selection, naming, reading, semantic interference, MEG
ABSTRACT
Naming an object involves quick retrieval of a target word from long-term memory. Research
using the semantic interference paradigm has shown that objects take longer to name when
they are preceded by primes in the same semantic category. This has been interpreted as
reflecting either competition during lexical selection or as an interference effect at a later,
postlexical level. Since the behavioral finding has been a core argument for the existence of
competition during lexical selection in naming, understanding its processing level is important
for models of language production. We used MEG to determine the spatiotemporal
localization of the interference effect. We also compared its neural signature to the effect of
semantic relatedness in reading, in which relatedness is expected to speed up behavioral
responses and reduce activity in the left superior temporal cortex at around 200–300 ms. This
is exactly what we found. However, in naming, we observed a more complex pattern for our
semantically related targets. First, the angular gyrus showed a facilitory pattern at 300–400 ms,
likely reflecting aspects of lexical access. This was followed by a broadly distributed and
sustained interference pattern that lasted until articulatory stages. More transient interference
effects were also observed at 395–485 ms in the left STG and at ∼100–200 ms before
articulation in the parietal cortex. Thus, our findings suggest that the semantic interference
effect originates from both early and late sources, which may explain its varying localizations
in previous literature.
INTRODUCTION
Retrieval of words from memory is a core component of language production. The most com-
mon experimental and clinical paradigm for studying this process is object naming, a task in
which a subject or patient is asked to produce the name of an object shown in a picture.
Classic models of naming hypothesize this task to involve the lexical selection stage where
related items compete with each other for selection (Bloem & La Heij, 2003; Levelt, 2001;
Levelt, Roelofs, & Meyer, 1999). The main empirical evidence for the competition is a behav-
ioral slow-down when a picture is preceded by a word prime from a related semantic category
(e.g., dog preceding an image of a cat). Whether this effect originates at a lexical or postlexical
level is, however, controversial.
Here we sought to characterize its neural sources in time and space with MEG. As a com-
parison task to naming, we chose overt reading (i.e., reading aloud), which also involves
Citation: Dirani, J., & Pylkkänen, L.
(2020). Lexical access in naming and
reading: Spatiotemporal localization of
semantic facilitation and interference
using MEG. Neurobiology of Language,
1(2) 185–207. https://doi.org/10.1162/
nol_a_00008
DOI:
https://doi.org/10.1162/nol_a_00008
Supporting Information:
https://doi.org/10.1162/nol_a_00008
Received: 28 August 2019
Accepted: 19 March 2020
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Author:
Julien Dirani
julien.dirani@nyu.edu
Handling Editor:
Matthew Lambon Ralph
Copyright: © 2020 Massachusetts
Institute of Technology. Published
under a Creative Commons Attribution
4.0 International (CC BY 4.0) license.
The MIT Press
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
2
1
8
5
1
8
6
7
7
2
2
n
o
_
a
_
0
0
0
0
8
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Lexical access in naming and reading
articulation but should not evoke the behavioral interference effect since written words spell
out the concept to be named, that is, the word selection process involves a word recognition
rather than a word retrieval. As a way to diagnose whether neural effects reflected facilitation
or interference, our design included a repetition priming condition, assumed to elicit only fa-
cilitory effects. Thus, effects in the same direction as repetition priming were assumed to be
facilitory and effects in the opposite direction were assumed to be interfering.
Our general aim was to provide a basic characterization of the effects of semantic category
relatedness in naming and overt reading, with a focus on the controversial lexical vs. postlex-
ical question about relatedness effects in naming.
The Picture-Word Interference Paradigm
On one account, lexical selection in production is considered a competitive process in which
multiple candidates compete for selection (Levelt, 2001). Crucially, the speed and ease of lex-
ical selection is affected by the activation level of the coactivated items. Core evidence for
lexical selection by competition comes from picture-word interference (PWI) paradigms
(Bloem & La Heij, 2003; Levelt et al., 1999), in which target pictures are named more slowly
when preceded or superimposed by categorically related word primes (cat-dog) as compared
with unrelated primes (chair-dog). The effect has been interpreted as a lexical-level effect
where coactivated items (here “dog” and “cat”) compete for selection.
However, a number of subsequent findings have called into question the competitive na-
ture of lexical selection. It appears that manipulating the type of semantic relation between
the word prime and target could modulate priming effects. For instance, there is a reversal of
the interference into a semantic facilitation with semantically related verb distractors (e.g.,
drive-car; Mahon, Costa, Peterson, Vargas, & Caramazza, 2007), with part-whole relations
(e.g., engine-car; Costa, Alario, & Caramazza, 2005), with associates (carrot-rabbit; Heij,
Dirkx, & Kramer, 1990; Sailor, Brooks, Bruening, Seiger-Gardner, & Guterman, 2009;
Xavier, Segui, & Ferrand, 2000) and with within-category close vs. within-category far dis-
tractor words (e.g., car-truck vs. wagon-truck; Mahon et al., 2007). Further, a facilitation ef-
fect is observed when primes are masked (Dhooge & Hartsuiker, 2010; Finkbeiner &
Caramazza, 2006b).
Difficulties in reconciling these findings with the competitive account of lexical selection
have led to the formulation of the response exclusion hypothesis (REH; Janssen, Schirm,
Mahon, & Caramazza, 2008; Mahon et al., 2007), which is based on the premises that (a)
language production involves a single-channel output buffer and that (b) written and auditory
words have privileged access to this buffer over names of images (Mahon et al., 2007).
Therefore, before the name of the image can be produced, this output buffer must be cleared
of the representation of the prime. This process is regulated by semantic information (Glaser
& Glaser, 1989; La Heij, 1988; Lupker, 1979) and gives rise to the interference effect. In
other words, the REH argues that the competition originates postlexically, at the point when
a decision about a response must be made. The competition is dependent on response-relevant
criteria: for example, distractor verbs do not interfere with naming times of images because
participants expect to have to name an object, making the verbs easier to exclude as potential
responses (Mahon et al., 2007). Given the new evidence for a postlexical locus of semantic
interference, the REH contends that lexical access is a noncompetitive process. Specifically,
multiple candidates are activated during lexical access, but instead of competing for selec-
tion, the first candidate to reach a certain threshold is selected (Finkbeiner & Caramazza,
2006a, 2006b; Janssen et al., 2008). That is, according to the REH, the speed of lexical
Neurobiology of Language
186
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
2
1
8
5
1
8
6
7
7
2
2
n
o
_
a
_
0
0
0
0
8
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Lexical access in naming and reading
selection is unaffected by coactivated candidates. It is worth mentioning that the REH ex-
plains facilitation effects (such as with carrot-rabbit relations) in terms of early priming at
the lexical or conceptual level, which is outweighed by a late interference with response-
relevant distractors only. Additional evidence in support of the REH and a noncompetitive
lexical-access process comes from bimodal bilinguals who do not show cross-language se-
mantic interference effects (Giezen & Emmorey, 2016). Since signed and spoken languages
engage different articulators, this is taken as evidence for a postlexical, articulatory locus of
semantic interference.
However, more recent studies failed to replicate the effect that motivated the REH. For in-
stance, it appeared that the within-category close vs. within-category far effect could not be
replicated, rather it was found that the inference increased as semantic distance got smaller
(Fieder, Wartenburger, & Rahman, 2019; Rose, Aristei, Melinger, & Abdel Rahman, 2019). In
addition, the part-whole facilitation effect was replicated at stimulus onset asynchronies
(SOAs) of 150 ms and 300 ms, but not 0 ms (Sailor & Brooks, 2014). Other effects that were
initially shown in support of the REH showed that they were actually in line with computa-
tional models of lexical selection by competition. For instance, the WEAVER++ model
(Roelofs, 2003) supports lexical selection by competition and has shown to be able to account
for distractor-frequency effects where low-frequency primes interfere more than high-frequency
primes. Further, the model also accounted for the influence of a mask on the priming effects
(Roelofs, Piai, & Schriefers, 2011).
Stimulus Onset Asynchrony Modulates Priming Effects
Previous studies have shown that manipulating SOA can modulate the priming effects in PWI
tasks. Results from these studies show that the interference effect disappears with SOAs ap-
proaching 400 ms and longer (Bloem, van den Boogaard, & La Heij, 2004; Heij et al.,
1990), with interference appearing to be the strongest at short SOAs, when the distractor is
presented between −100 ms and 150 ms from the onset of the image (Costa et al., 2005;
Heij et al., 1990; Sailor et al., 2009; Starreveld & La Heij, 1995; Xavier et al., 2000). It is then
clear that a full account of interference and facilitation effects in priming should include a
manipulation of SOA.
Neural Correlates of the Semantic Interference and Facilitation Effects
Determining the timing of semantic interference—specifically, whether competition occurs
during lexical selection or at later stages of processing—is important for models of lexical ac-
cess in production. Lexical selection is typically reported to start at around 200 ms (Indefrey,
2011; Munding, Dubarry, & Alario, 2016). Therefore, an early effect of semantic interference
at ∼200 ms would imply a lexical-level interference and competition for lexical access, while
a later effect starting at ∼400 ms would imply a postlexical effect and no competition during
lexical access. Since electrophysiological measures have high temporal resolution, they can
provide valuable information regarding the timing of the semantic interference. Different EEG
studies have yielded variable timing estimates for the interference, showing both early (200–
500 ms; Aristei, Melinger, & Abdel Rahman, 2011) and late (325–600 ms; Blackford,
Holcomb, Grainger, & Kuperberg, 2012) onsets for the relevant effect. Additional EEG findings
have also shown much later effects at 500–750 ms, which were interpreted as cognitive con-
trol mechanisms (Janssen, Hernández-Cabrera, van der Meij, & Barber, 2014). MEG measures
showed a semantic interference effect at 350–650 ms in the left superior frontal gyrus in the
4–10 Hz range (Piai, Roelofs, Jensen, Schoffelen, & Bonnefond, 2014), however, an earlier
Neurobiology of Language
187
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
2
1
8
5
1
8
6
7
7
2
2
n
o
_
a
_
0
0
0
0
8
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Lexical access in naming and reading
effect at 150–225 ms was found using a blocked semantic interference paradigm where im-
ages belonging to the same vs. different categories were presented in a blocked design
(Maess, Friederici, Damian, Meyer, & Levelt, 2002). Further, an intracranial EEG study on
epileptic patients showed that the interference effect was associated with an increase in activity
in the supplementary motor area at both early (200–300 ms) and late (700–800 ms) time windows
(Anders et al., 2019) and were interpreted as representing the initial conflict detection and later
preresponse control during object naming. In essence, the semantic interference effect has been
reported in various time windows ranging from 150 ms to 800 ms. A recent study has suggested an
account of the semantic interference that is consistent across many different paradigms pointing
to a lexical level locus (Roelofs, 2018). Nevertheless, the electrophysiological literature still does
not show a consistent picture for the lexical locus of the semantic interference effect. It is therefore
still unclear whether the semantic interference effect localizes at an early or a late time window,
or at both.
Although the blood-oxygen-level dependent response is too slow to detect the timing of the
interference effect, fMRI studies have provided information regarding the spatial extent of the
effect. Semantic interference has modulated activity in the left posterior superior temporal gy-
rus (STG), left anterior cingulate cortex (ACC), bilateral orbitomedial prefrontal cortex, and the
left midsection of the middle temporal gyrus (MTG) (de Zubicaray, Hansen, & McMahon,
2013; de Zubicaray, Wilson, McMahon, & Muthiah, 2001).
In contrast to naming, semantic relatedness has a straightforwardly facilitory effect in
reading that is thought to originate at the lexical level via spreading activation from the
prime to neighboring representations (Hala, Pexman, & Glenwright, 2007; Katz &
Feldman, 1983; Rosson, 1983; Vinogradov, Ober, & Shenaut, 1992). To our knowledge,
no study to date has examined the neural correlates of semantic facilitation in overt reading.
In contrast, semantic priming has been extensively studied in silent reading, with lexical de-
cision as the most common experimental task. The most consistent result from this work has
been a reduction of the N400 response as a reflex of semantic priming, starting at around
250 ms and peaking at ∼400 ms post-word onset (Holcomb & Anderson, 1993; Matsumoto,
Iidaka, Haneda, Okada, & Sadato, 2005; Rossell, Price, & Nobre, 2003). The spatial extent
of the effect involves the left inferior frontal gyrus, left STG and left MTG, the middle frontal
gyri, the ACC, as well as at the inferior parietal lobe, as measured by hemodynamic methods
(Kotz, Cappa, von Cramon, & Friederici, 2002; Matsumoto et al., 2005; Rissman, Eliassen, &
Blumstein, 2003; Rossell et al., 2003).
While effects of semantic relatedness have been studied with neurobiological measures
both in naming and reading (with most of the latter addressing silent reading), few studies
to date have directly compared them (Anders et al., 2019). Such a comparison would provide
crucial insight into the competitive vs. noncompetitive question in lexical access, because it
allows for directly comparing the interference and facilitation effects in tasks that are maximally
similar at the behavioral level. In the present study, we exploited the high temporal resolution
of MEG to compare the spatiotemporal neural responses associated with facilitation priming
in reading to semantic interference in naming. The relatively good spatial resolution of MEG
also allowed us to complement the temporal data with spatial information, which in turn al-
lowed us to examine the spatial extent of the interference and facilitation effects on the cor-
tical surface. MEG does not provide the spatial resolution of fMRI, since it makes statistical
estimates regarding the sources of the signal measured at the sensors. Nevertheless, given
prior hypotheses regarding the source localization of effects, the spatial resolution of MEG
offers a good estimate of the sources of the signal on the cortical surface. Here, if interference
Neurobiology of Language
188
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
2
1
8
5
1
8
6
7
7
2
2
n
o
_
a
_
0
0
0
0
8
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Lexical access in naming and reading
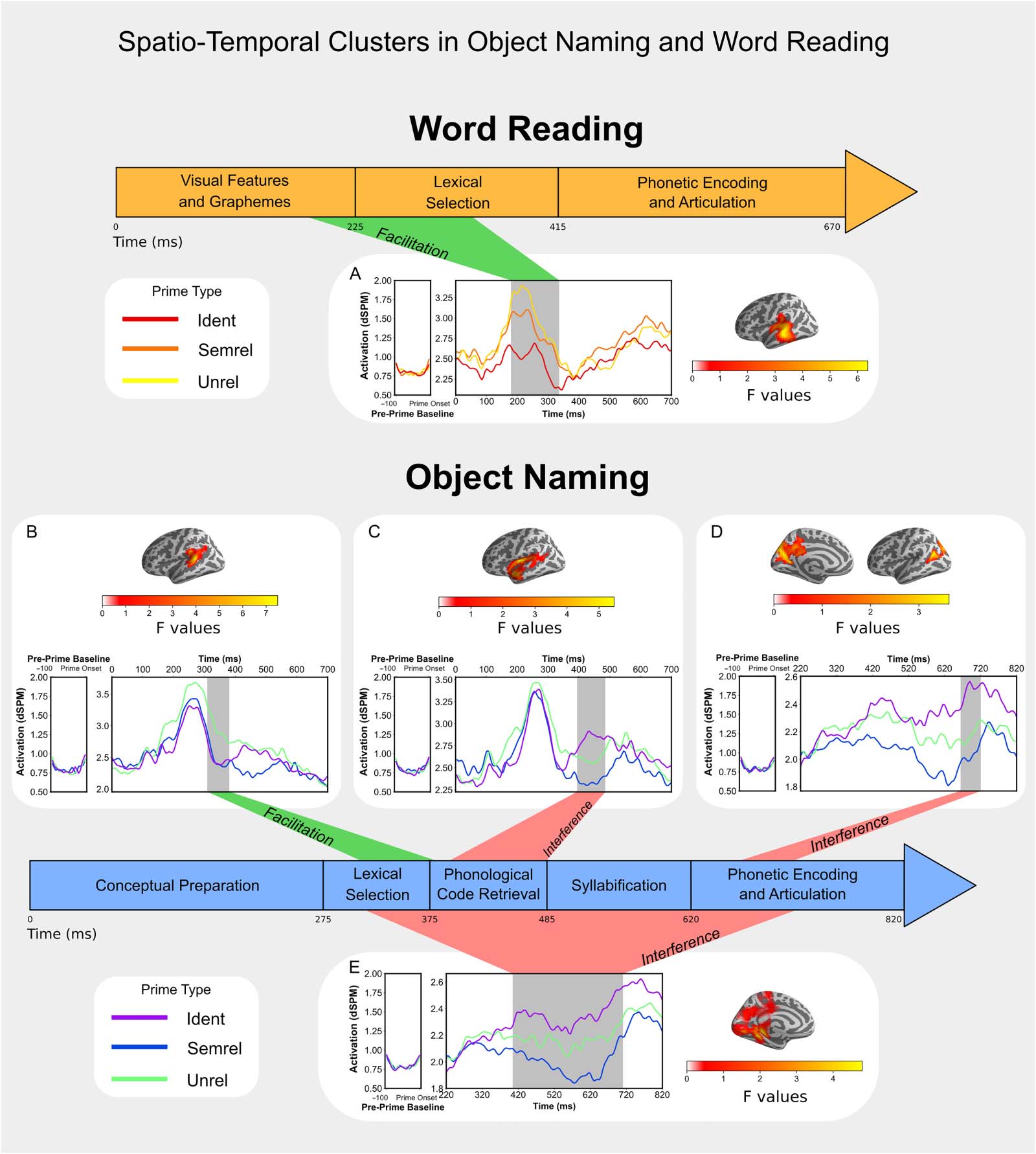
Figure 1. Three contrasting hypotheses (H1, H2, H3) regarding the localization of semantic interference in object naming. Estimated times of
processing steps are based on Indefrey and Levelt (2004).
effects in naming and facilitation effects in reading both localize in temporal areas (mainly
STG and MTG) around 200–300 ms or earlier, this would conform to a shared lexical-level
origin for the interference and facilitation effects. In contrast, if the interference effect in nam-
ing is manifested later than 400 ms after picture onset, a postlexical account of this effect is
more likely (Figure 1). A lexical level of interference would be in line with models of lexical
selection by competition, while a postlexical locus would imply that lexical selection is not
competitive. Finally, a manipulation of SOA could provide us with additional insight as to
the timing of priming effects. For instance, it is not clear how priming in reading would be
affected, with possibly a facilitation effect turning into interference at a given SOA.
Importantly, the timing estimations that we are using (Figure 1) are based on Indefrey and
Levelt’s (2004) model of word production, which assumes a naming latency of 600 ms.
Response times of participants vary from 600 ms, so these estimations can be inaccurate.
To properly interpret the timing of our results, a proportional rescaling of the processing steps
has been done, according to Roelofs and Shitova (2017). That is, each estimated processing
step in the Levelt model is made longer or shorter depending on the average response time. For
example, timing of lexical access is estimated to start at 200 ms for a naming time of 600 ms.
For an observed naming time of 750 ms, the estimated timing of lexical access would have to
be rescaled to be estimated to start at 250 ms (750/600 × 200).
MATERIALS AND METHODS
Participants
Thirty right-handed native English speakers were paid to take part in the study. Two partici-
pants were excluded due to excessive artifacts that resulted in >25% of rejected trials, and 3
participants were rejected due to equipment failure, leaving 25 good participants (14 female,
M = 22.67, SD = 5.55). All participants had normal or corrected-to-normal vision and reported
no history of neurological or language disorders. The study received ethical approval from the
institutional review board at New York University Abu Dhabi.
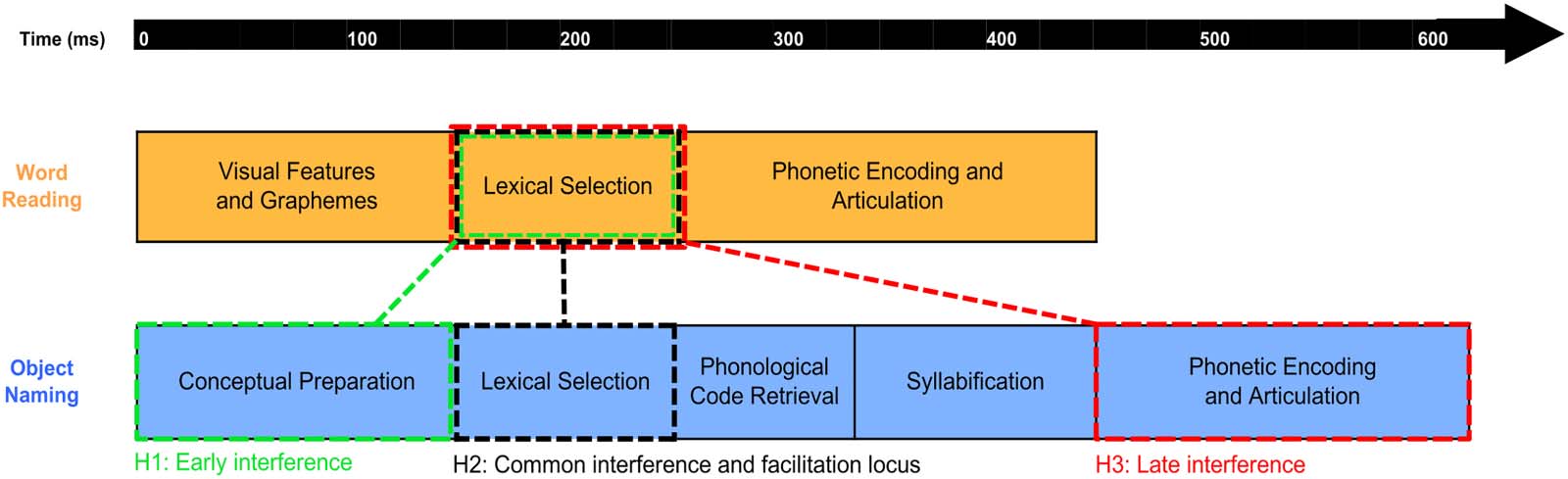
Experimental Design
The experiment consisted of an overt reading and an object naming task. The targets to be
named consisted of line drawings (Object Naming) and lowercase words (Word Reading).
In both tasks, three levels of primes were manipulated: First, semantically related primes
Neurobiology of Language
189
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
2
1
8
5
1
8
6
7
7
2
2
n
o
_
a
_
0
0
0
0
8
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Lexical access in naming and reading
(Semrel, e.g., chair-table), which were words that belonged to the same semantic category of
the targets. Categorically related primes have been shown to reliably induce interference ef-
fects in object naming. We therefore only used this type of semantic relation in order to guar-
antee we would observe the interference priming effect that we aimed to compare to
facilitation in the reading task. We also used unrelated primes (Unrel, e.g., dog-table),
which were words that differed from the target in all aspects (visual, phonology, and seman-
tics). Finally, identical primes (Ident, e.g. table-table) repeated the target word or the word that
would best name the object in the target image. We included this condition to provide us with
a clear facilitation effect for both tasks, which would allow us to interpret the remaining effects
in comparison. This is crucial for the neural data, where interpreting an increase or decrease in
activation is not always straightforward. Crucially, Ident primes serve as the basis on which to
discriminate between facilitory and interfering neural patterns. Since identical prime-target
pairs (table-table) are maximally related and expected to elicit robust facilitory repetition prim-
ing, we considered a pattern facilitory if the semantically related condition patterned between
the unrelated and identical conditions. That is, the pattern is facilitory if the semantically related
condition patterned in the same direction as the identical condition, compared with the unre-
lated condition. In contrast, in an interference pattern, the semantically related condition should
diverge from the unrelated condition in the opposite direction of the identical condition. For
example, if the identical condition showed an increase in activation compared with the unre-
lated condition, a decrease in activation for the Semrel compared with the Unrel primes would
be interpreted as an interference effect. All primes were in capital letters while all target words
were in lowercase (e.g., TABLE-table) to control for purely visual priming.
We also manipulated the SOA at four different intervals: 150, 200, 250, and 300 ms. These
SOAs were in the range that showed reliable interference effects in object naming (Heij et al.,
1990; Sailor et al., 2009) and were chosen after undergoing a behavioral pilot of our exper-
imental task aiming at selecting SOAs that would elicit a reliable interference effect in our
design. This was done in order to successfully elicit an interference effect to properly assess
the neural signature associated with it. Importantly, the potential interaction of SOA with
Prime Type and Task could provide us with additional insight as to the timing of priming
effects.
Stimuli were presented using Psychopy 1.84.2 (Peirce, 2007) on a screen positioned above
the participants’ heads while they laid back on a bed in the magnetically shielded room of the
MEG. Each trial started with the presentation of a fixation cross that appeared for 300 ms,
followed by a blank screen for 300 ms. Next, the prime appeared for 100 ms, followed by
a blank screen. The duration of the prime was held constant, but the blank screen following
it varied to create an SOA of 150, 200, 250, or 300 ms depending on the condition. Finally,
the target remained on screen until the participants named it (Figure 2). Responses were re-
corded with a microphone positioned near the participant’s mouth and the utterance times
were recorded in real time using Psychopy’s Voicekey.
Stimuli
The lists of all primes and the list of targets were English nouns in their root form, balanced
for length (Ident: M = 4.79, SD = 1.41; Unrel: M = 4.83, SD = 1.22; Semrel: M = 5.24, SD =
1.29) and frequency (Log frequencies: Ident: M = 9.12, SD = 1.35; Unrel: M = 9.03, SD =
1.62; Semrel: M = 8.28, SD = 1.39) across all lists. After the stimuli generation was done, 50
participants rated the semantic relation between the unrelated and semantically related
primes and the targets via the Amazon Mechanical Turk platform (www.mturk.com). They
Neurobiology of Language
190
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
2
1
8
5
1
8
6
7
7
2
2
n
o
_
a
_
0
0
0
0
8
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Lexical access in naming and reading
Figure 2. Trial structure and experimental design. SOA = stimulus onset asychrony.
were instructed to rate how much they thought the words belonged to the same category
using a Likert scale ranging from 1 to 5. Any prime-target pairs that had an average rating
between 2.5 and 3.5 were excluded from the stimuli, creating two distinct groupings of se-
mantically related and unrelated prime-targets (Semrel pairs: M = 4.31, SD = 0.32; Unrel
pairs: M = 1.25, SD = 0.19).
The stimuli consisted of 82 Sets. Within each Set, there was one common target that was
repeated six times: three times as an image and three times as a word. There were unique
Semrel and Unrel primes for each of the target types. The Ident prime was repeated twice,
once with each target type. Since we also wanted to manipulate SOA at four intervals (150,
200, 250, 300 ms), we opted to present each one of our Sets of stimuli (i.e., six prime-target
pairs, with a shared target) twice. In order to control for anticipation and predictability, we
created two versions of each prime type while trying to minimize the semantic distance be-
tween the two. In other words, each Set was presented twice and the versions of the primes
were changed in each repetition. The result was that subjects saw each target 12 times (six
times as an image, and six times as a word), and each unique prime twice (once before the
image, and once before the word), with the exception of the Ident prime that was seen four
times. In order to avoid confounding any effects of SOAs with effects created by specific items,
it was necessary to avoid consistently pairing specific SOAs with specific Sets. That is, we had
to counterbalance the pairing of SOAs with Sets across subjects. This was done by first arbi-
trarily splitting the 82 Sets into two lists of 41 Sets. Each list of Sets (A and B) was then paired
with two SOAs, ensuring that each Set was presented twice (as previously stated). This pairing
was counterbalanced across every six participants in order to ensure that a specific pairing of
SOA to item did not confound the group-level results. Further, to control for the order of rep-
etitions of targets within subjects, each of the 24 cells of the design was assigned to a block
number following a Latin-squared method. Thus, the block number corresponded to the order
of trials in the experiment. This was done to ensure that within each subject, the number of
times that a given condition (e.g., Unrel, Object Naming, 150 ms SOA) appeared earlier in the
experiment than another condition (e.g., Semrel, Word Reading, 200 ms SOA) was equal, pair-
wise, across all conditions in the experiment. However, since the total number of cells in the
design was 24 (6 prime-target pairs × 4 SOAs), while the total number of items in a List was 42,
it was not possible to fully cross all of the conditions with block number (which would require
Neurobiology of Language
191
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
2
1
8
5
1
8
6
7
7
2
2
n
o
_
a
_
0
0
0
0
8
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Lexical access in naming and reading
48 Sets per List, 96 in total). Thus, we distributed each of the conditions across block number
in a manner that was as close to uniform as possible. Within the resulting 12 blocks, trial order
was randomized, which conserved the Latin-squared order over the whole trials. There was a
total of 41 trials per condition.
The downside of this randomization is that we repeated primes twice, over random pairs of
SOAs. Ideally, we would have proceeded in one of the following two ways, but both were
ultimately problematic: present every Set with all of the four SOAs (items would repeat exces-
sively, potentially allowing participants to anticipate upcoming targets); or create completely
unique Sets of stimuli for each SOA, balanced on relevant characteristics (unfeasible to
generate enough unique Sets of prime-target pairs that could be controlled in all the necessary
ways). The procedure that we used here was a compromise to minimize as much as possible
the number of repetitions across SOAs, while counterbalancing the association of specific
stimuli tokens with specific conditions.
MEG Acquisition and Processing
Continuous MEG was recorded with a 208-channel axial gradiometer system (Kanazawa
Institute of Technology) at a sampling rate of 1,000 Hz with an online band-pass filter of
0.1–200 Hz. The raw data was noise-reduced with the continuously adjusted least-squares
method (Adachi, Shimogawara, Higuchi, Haruta, & Ochiai, 2001) using the MEG
Laboratory software 2.004A ( Yokogawa Electric and Eagle Technology Corp., Japan). All
the following preprocessing was done using the MNE-Python 0.14 (Gramfort et al., 2014)
and Eelbrain 0.25.2 (Brodbeck, 2017) packages. The data was first converted to .fif format.
After visual inspection of the data, bad channels were excluded, and the data was low-pass
filtered offline at 40 Hz. An independent component analysis was then fitted to the data using
the “fastica” method, selecting components by 95 cumulative percentage of explained vari-
ance. Components related to eye-blinks, heartbeats, saccades, and dead channels were then
rejected manually. Epochs from −100 to 600 ms from target onset were extracted and baseline
correction was done using the 100 ms before the onset of the target. Epochs exceeding a max-
imum peak-to-peak threshold of ±2000 femto-tesla were removed automatically, and the re-
maining epochs were scanned for eye-blink artifacts and were removed accordingly. It was not
necessary to clean the data for articulation artifacts since correct responses never occurred
before 600 ms after target onset. Finally, wrong responses, responses in which participants
stuttered, and responses faster than 300 ms and slower than 2,000 ms were excluded from
the analysis. For the naming task, responses that were a correct match with the picture but
that were not in agreement with their word equivalent were excluded. For example, if the im-
age depicted a “dog,” and a participant said “puppy,” it was considered a wrong response. All
the remaining good epochs (Trials per condition across subjects: M = 956.5, SD = 29.5) were
down-sampled by 5, so that the sampling rate became 200 Hz, and were then averaged by
condition to form the evoked responses.
Each subject’s head-shape was created using an optical FastSCAN scanner (Polhemus) and
was co-registered with the FreeSurfer (http://surfer.nmr.mgh.harvard.edu/) average brain. To
execute a better co-registration, the average brain was scaled using three-dimensional axes
to match each subject’s head-shape. The source space was defined as a dipole grid on the
white matter surface using the topology of a recursively subdivided icosahedron (“ico-4” op-
tion). Only sources in the left hemisphere were included and were defined using the PALS-B12
atlas (Van Essen, 2005). A separate inverse solution was then computed for each subject with
the evoked responses, using the forward solution as well as the noise covariance matrix
Neurobiology of Language
192
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
2
1
8
5
1
8
6
7
7
2
2
n
o
_
a
_
0
0
0
0
8
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Lexical access in naming and reading
computed from the respective 100 ms baselines of each condition. For each subject, the noise
covariance matrix was estimated using the best estimator out of the three methods shrunk co-
variance, diagonal fixed covariance, and empirical covariance, based on log-likelihood and
cross-validation on unseen data (Engemann & Gramfort, 2015). For each source location, min-
imum norm current estimates were computed using three orthogonal dipoles, resulting in a 3D
vector. Only the lengths of the vectors were retained, resulting in orientation-free source
estimations. The resulting estimates were noise-normalized at each source using a signal-
to-noise ratio regularization factor of 3 for the regularization of the inverse operator. This re-
sulted in noise-normalized statistical parametric maps, which were then converted to dynamic
maps (dSPMs) and provided information about the statistical reliability of the estimated signal
at each source (Dale et al., 2000). Finally, source activity was morphed to the FreeSurfer aver-
age source space in order to be comparable across subjects.
Statistical Analyses
MEG stimulus-locked analyses
The initial statistical analysis was based on a mass univariate analysis with spatiotemporal
cluster-based permutation tests (Holmes, Blair, Watson, & Ford, 1996; Maris &
Oostenveld, 2007) applied to the source estimates (in dSPMs). Average source estimates
for each condition and for each subject were used in the analysis. The F value of a 2 × 3 × 4
repeated-measures ANOVA (Task × Prime Type × SOA) was computed for each source at each
time point in the full left hemisphere and limited to the 100–600 ms time window. This F map
was thresholded at an F value corresponding to an uncorrected p value of 0.01. Clusters were
formed based on direct adjacency in space and time, with the restrictions that they contain a
minimum of 10 sources and last at least 10 ms. The sum of all F values ((cid:1)F ) was computed for
each resulting cluster. This procedure was then repeated 10,000 times, each time with a ran-
dom permutation of the data, by shuffling condition labels within subjects. For each permu-
tation, the largest of the (cid:1)F was saved to create a nonparametric permutation distribution. The
Monte Carlo p value was computed for each cluster in the original F map as the proportion of
random permutations in which the observed (cid:1)F was larger than the values from the permuta-
tion distribution. We retained clusters whose Monte Carlo p value was smaller or equal to
0.05.
A secondary analysis was performed to unpack the patterns of priming effects within each
Task. The same cluster-based permutation test described above was performed, sub-setting the
data by Task and thus using a 3 × 4 repeated-measures ANOVA (Prime Type × SOA). The
Monte Carlo p-value threshold was corrected using the Bonferroni method to account for
the multiple comparisons across the two tasks.
MEG response-locked analysis
Electrophysiological data is most often analyzed time-locked to the onset of the stimulus
(Indefrey, 2011). While it has proven to be very useful, this approach is not ideal to assess
the later processing steps of language production, mainly the planning of the articulation. In
fact, activity that is related to the execution of the response tends to be reduced in stimulus-
locked analyses (Riès, Janssen, Burle, & Alario, 2013). In the current study, one of our hy-
potheses predicts that the semantic interference effect localizes at later motor preparatory
stages of processing, which makes it crucial to properly assess the stages right before the
articulation occurs. Thus, it was essential to include an analysis with epochs locked at the
response time.
Neurobiology of Language
193
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
2
1
8
5
1
8
6
7
7
2
2
n
o
_
a
_
0
0
0
0
8
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Lexical access in naming and reading
For each trial, the utterance time was taken along with the 600 ms preceding it to create
epochs of equal length. One additional subject was excluded from this analysis due to ex-
cessive noise in the MEG data (>25% of trials rejected), resulting in a sample of 24 partic-
ipants. Baseline correction was done using the 100 ms before the onset of the prime, and
source reconstruction was estimated using the similar procedure described in “MEG acqui-
sition and processing.” Finally, the same spatiotemporal within-task 3 × 4 ANOVA (Prime
Type × SOA) that was previously mentioned was run on the response-locked source-level
epochs.
Plots and assumptions of MEG results
For all analyses, significant clusters were plotted as time courses as well as bar graphs showing
their average dSPM value. Sources included in the cluster were plotted on the FsAverage brain
with the average F values for the time window of the significant cluster. In all plots, time 0
represented the onset of the target. Concerning results for the effect of Prime Type, we only
reported clusters showing a semantic priming pattern. That is, we only presented clusters
where the Semrel and Unrel conditions showed distinct time courses that separate from each
other. The reason is that pure Ident priming effects do not directly address our hypothesis,
since the Ident condition was only included as a baseline for interpreting semantic priming
effects.
It is important to mention an assumption that goes into plotting clusters found using spatio-
temporal permutation tests. In the spatiotemporal test, a significant cluster is a set of contigu-
ous points in the time-source domain. That is, not all sources contribute equally to the cluster
at all time points, which makes it difficult to visualize the spatiotemporal cluster as a contin-
uous time course. The most straightforward way to visualize the cluster is to average over all
the sources (in space) and plot the resulting average activation over time, which assumes all
sources contribute equally to the cluster at all time points. Crucially, as a consequence, we
cannot visually deduct from the plotted time course anything that is outside the cluster’s time
extent because we do not have a priori information that any effect survived the multiple com-
parison there. We can interpret the activation in the cluster time window because we confi-
dently know that there is an effect there that survived the permutation test, even though the
plot is susceptible to smearing out information in space and time.
Behavioral Analyses
Voice utterance reaction times (RTs) were analyzed with a linear mixed-effect model using the
LmerTest package (Kuznetsova, Brockhoff, & Christensen, 2017) in R (R Core Team, 2013). As
with the MEG data, wrong responses, responses in which participants stuttered, and responses
faster than 300 ms and slower than 2,000 ms were excluded from the analysis. The initial
model included all main effects of Prime Type, Task, SOA, all two-way interactions, and
the three-way interaction as fixed effects. Random intercepts were used for subjects and items.
To test for the significance of the predictors, we performed a sequential decomposition of the
contributions of the fixed effects using the ANOVA function from the LmerTest package, using
type-III hypothesis test. For each predictor, an F test and its corresponding p value were esti-
mated using Satterthwaite’s method (Giesbrecht & Burns, 1985; Hrong-Tai Fai & Cornelius,
1996). Post hoc pairwise comparisons of significant effects were done using differences of least
square means corrected for multiple comparisons using the Tukey method, with
Satterthwaite’s estimation for degrees of freedom. The final model was then retrieved with
backwards elimination of nonsignificant effects.
Neurobiology of Language
194
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
2
1
8
5
1
8
6
7
7
2
2
n
o
_
a
_
0
0
0
0
8
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Lexical access in naming and reading
RESULTS
Behavioral Data
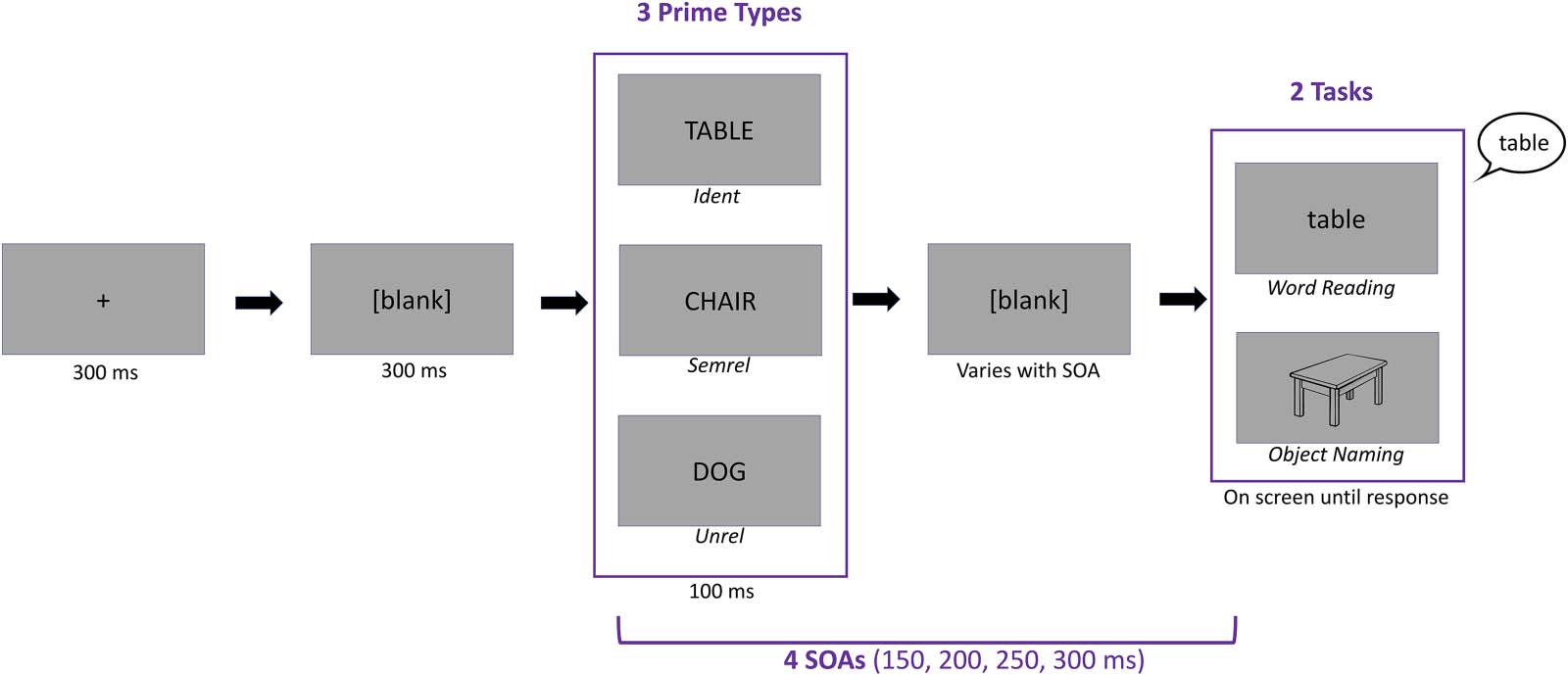
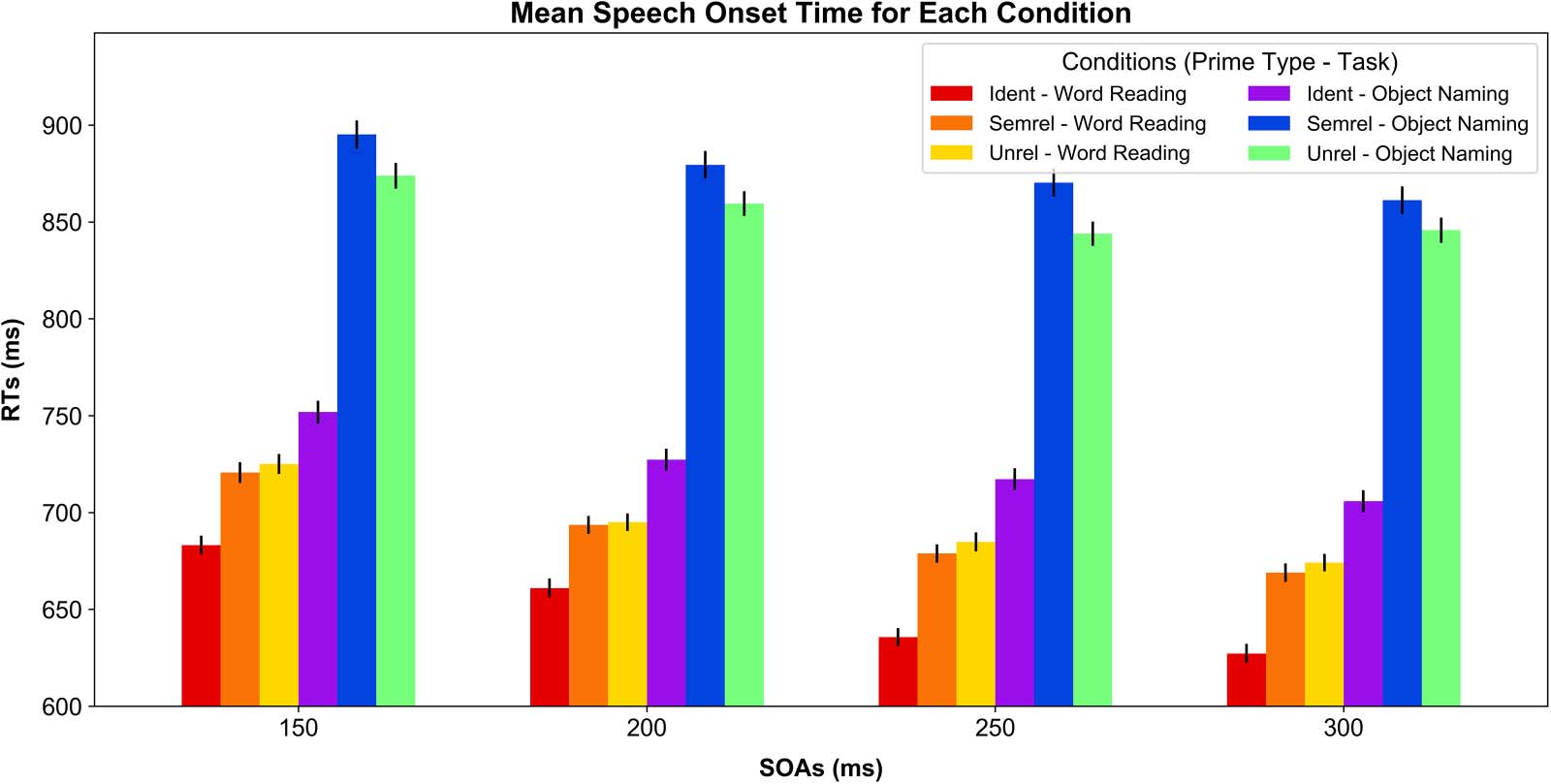
We found a main effect of Task, F(1, 23,382) = 5,585.43, p < .001, with longer RTs observed
for Object Naming (M = 817.63, SD = 211.25) compared with Word Reading (M = 679.04,
SD = 156.10; Figure 3). We also found a main effect of Prime Type, F(2, 23,378) = 1,053.39,
p < .001, showing that the Ident priming condition was the fastest (M = 688.25, SD = 172.55),
followed by Unrel (M = 773.51, SD = 194.98 and then Semrel (M = 779.79, SD = 210.62; p <
.001 for all post hoc paired tests).
Further, we found an interaction between Task and Prime Type, F(2, 23,377) = 326.16, p <
.001, which showed that, in Object Naming, RTs were shorter for Unrel (M = 855.76, SD =
200.16) compared with Semrel primes (M = 876.48, SD = 219.71; p < .001), illustrating the
predicted semantic interference effect. In contrast, for Word Reading, RTs were shorter for
Semrel (M = 690.57, SD = 155.46) compared with Unrel primes (M = 694.91, SD =
153.06), although this effect was not significant ( p = 0.21). This could be due to the fact that
in overt reading, as opposed to silent comprehension tasks, participants can rely on phonolog-
ical and orthographic information, and less on semantics, thus decreasing the strength of the
semantic facilitation effect. Further, the Ident priming condition revealed the fastest RTs in both
tasks (In Object Naming: M = 727.51, SD = 180.39; In Word Reading: M = 651.79, SD =
156.53). The main effect of SOA was a reliable predictor of RTs, F(3, 14,703) = 56.21, p <
.001; SOA 150: M = 772.26, SD = 200.35; SOA 200: M = 750.47, SD = 194.43; SOA 250:
M = 735.61, SD = 195.06; SOA 300: M = 728.31, SD = 197.71, and also interacted with the
effect of Task, F(3, 23,378) = 2.8817, p < .05. Within each task, RTs got shorter as SOAs got
longer (In Object Naming: SOA 150: M = 838.63, SD = 213.03; SOA 200: M = 820.95, SD =
209.75; SOA 250: M = 808.39, SD = 208.48; SOA 300: M = 802.77, SD = 212.01. In Word
Reading: SOA 150: M = 709.69, SD = 164.74; SOA 200: M = 683.17, SD = 150.39; SOA 250:
M = 666.43, SD = 151.96; SOA 300: M = 656.72, SD = 151.68), with the exceptions of
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
2
1
8
5
1
8
6
7
7
2
2
n
o
_
a
_
0
0
0
0
8
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. Behavioral RTs (i.e., utterance onset times) across all conditions. A stable effect of Prime Type was observed in both Word Reading
(warm colors) and Object Naming (cool colors) across all SOAs. RT = reaction time; SOA = stimulus onset asynchrony.
Neurobiology of Language
195
Lexical access in naming and reading
SOAs 250 and 300 in the Object Naming task, in which cases RTs did not significantly differ
( p = .20).
The final model that was obtained with backwards elimination of nonsignificant effects is
presented below. RTs were significantly predicted by the main effects of Prime Type and SOA,
as well as the interaction of Task with both Prime Type and SOA. Items and subjects were
included as random factors.
RT ∼ Task þ PrimeType þ SOA þ Task : PrimeType þ Task : SOA þ 1jSubject
ð
Þ þ 1jItem
ð
Þ
MEG Data
Omnibus analysis
The omnibus cluster-based permutation test revealed a robust, widespread main effect of Task.
The biggest spatial cluster expanded on most of the left hemisphere (93.72%, 2,401 sources)
and lasted for the full analysis time window (100–600 ms, p < .001; Figure 4). In addition, the
time courses of the two tasks exhibited two drastically different patterns. This indicates that
Word Reading and Object Naming were associated with strikingly different neural signatures
across the better part of the left hemisphere.
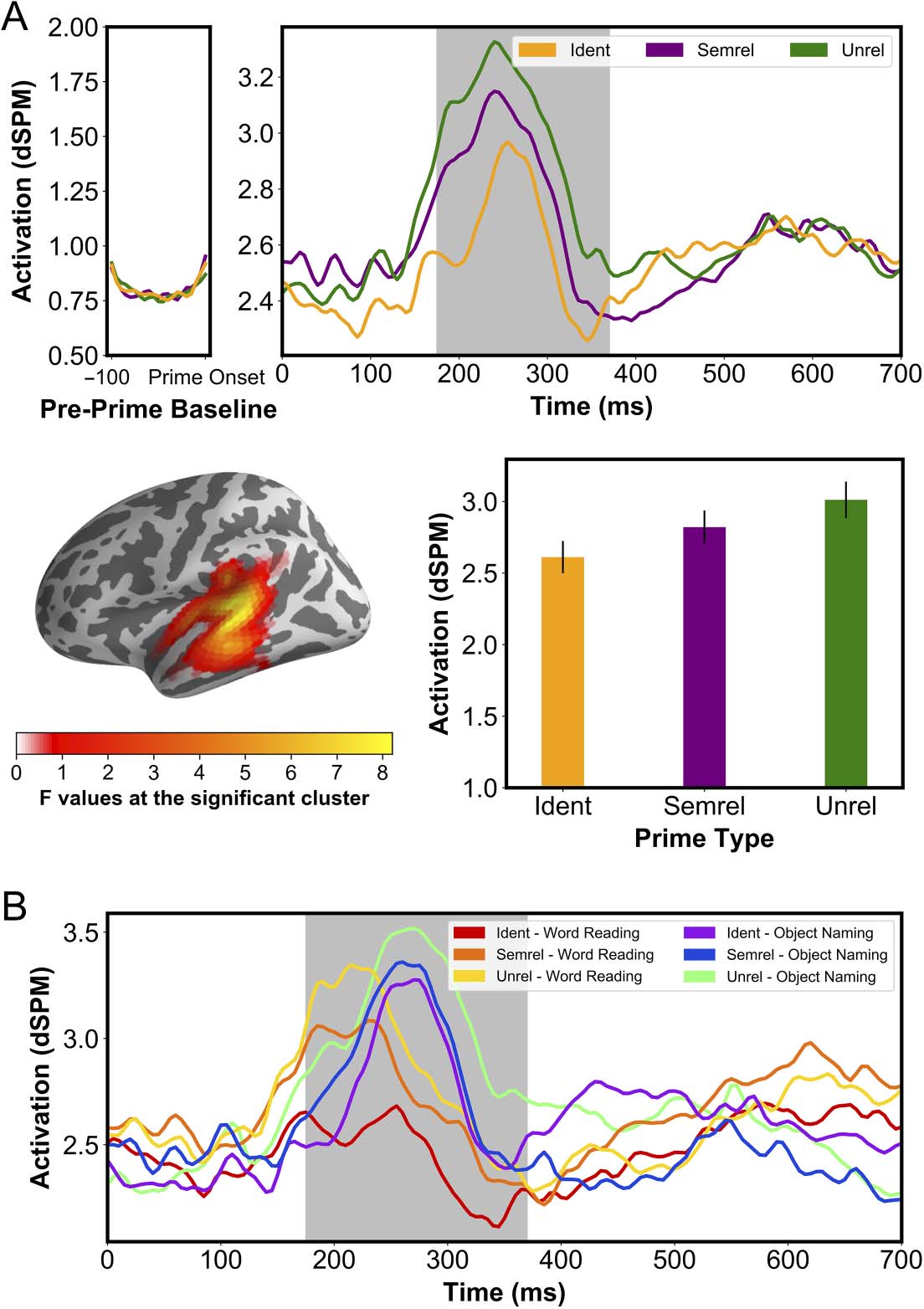
We also found a spatiotemporal cluster for the main effect of Prime Type (Figure 5) showing
a priming effect localized to the middle STG, between 175 ms and 370 ms ( p < .001, in which
activation increased stepwise as semantic distance increased. This pattern followed that of the
behavioral results collapsed over Task and SOA in which RTs were shortest for Ident primes
and longest for Unrel primes. The effect of Prime Type did, however, interact with the effect of
Task (Figure 6), indicating that the priming pattern described above might be a generalization
that is not necessarily representative of the priming patterns within each task. For Object
Naming, the priming pattern was in line with that of the behavioral results, with the highest
activation for the Ident primes, followed by the Unrel primes and then the Semrel primes. For
the Word Reading task, the activation of the Semrel condition appeared higher than that of
Unrel and Ident priming conditions, however, the latter two had similar activation levels.
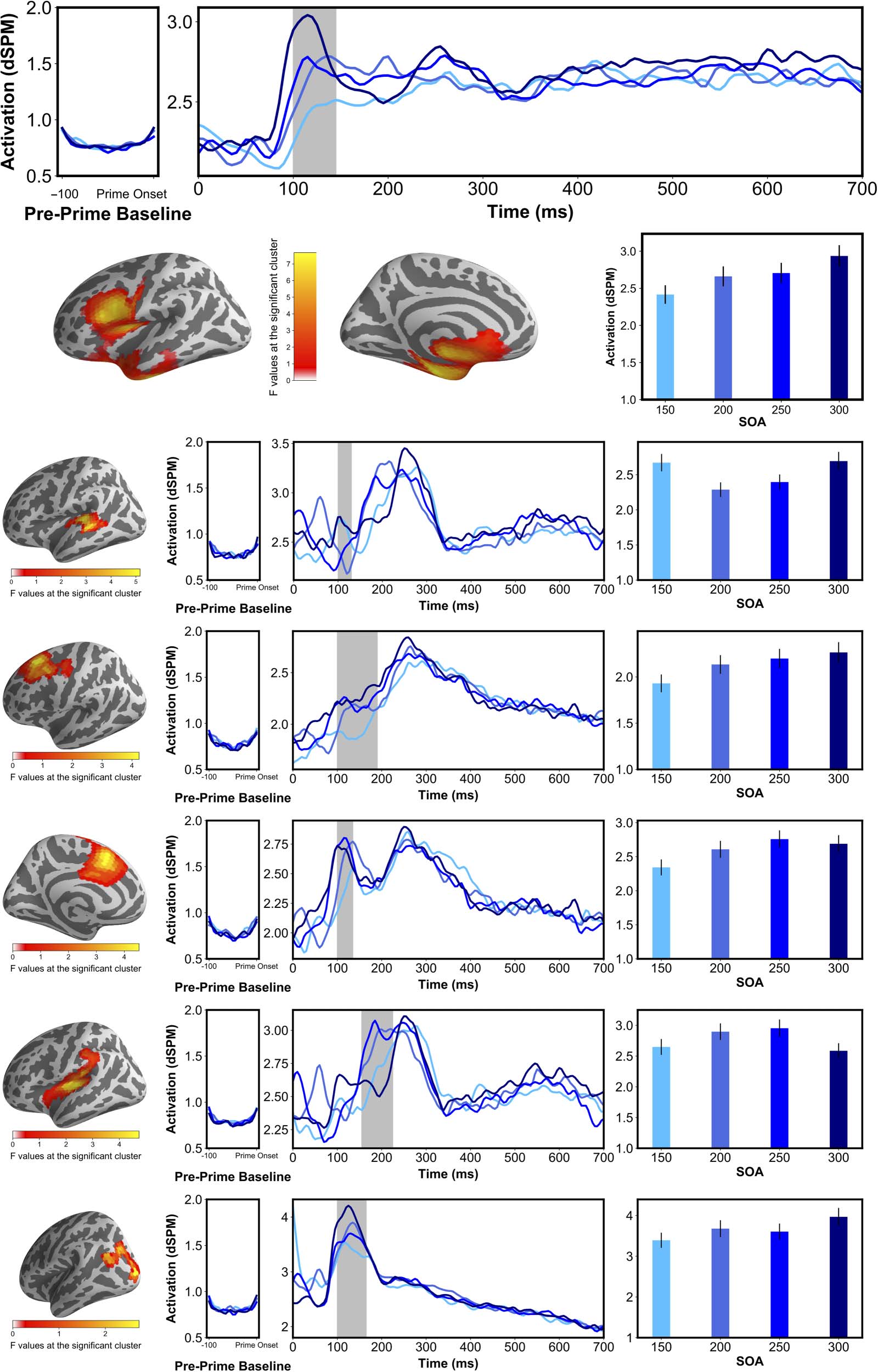
Finally, we found that SOA modulated brain responses at six different spatiotemporal clus-
ters (Figure 7). The largest cluster contained 518 sources located in frontal areas as well as
Figure 4. A widespread main effect of Task extending through the entire analysis time window and almost the entire left hemisphere (as well
as the right hemisphere, as observed in an additional post hoc test; p < 0.05, corr., with the most reliable signal centered at medial occipital
areas. The large effect of Task motivated our within-task analyses, to achieve greater sensitivity to observe priming effects. dSPM = dynamic
statistical parameter map.
Neurobiology of Language
196
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
2
1
8
5
1
8
6
7
7
2
2
n
o
_
a
_
0
0
0
0
8
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Lexical access in naming and reading
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
2
1
8
5
1
8
6
7
7
2
2
n
o
_
a
_
0
0
0
0
8
p
d
.
/
l
Figure 5. Main effect of Prime Type in the full across-tasks analysis. (A) Spatial extent of the significant cluster (left) and its time course
averaged across tasks. Bar graph shows mean amplitudes of the significant (shaded) temporal cluster across priming conditions. (B) Same data
split by task, revealing a clearer stepwise amplitude reduction for Word Reading than for Object Naming. (Grey shading on time courses
indicates p < 0.05, corr.). dSPM = dynamic statistical parameter map; Ident = identical; Semrel = semantically related; Unrel = unrelated.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
anterior medial temporal areas ( p < .001) and showed a stepwise increase in activation as
SOAs got longer.
Analysis within task
Our primary goal was to unpack the neural signatures of semantic priming for Object Naming
and Word Reading. Because the main effect of Task was so dramatic, with 93.72% of the left
hemisphere modulated by Task for the whole analysis time window, we opted for a second
analysis within Task. Our motivation was to exclude the large effect of Task in order to gain a
better understanding of semantic priming within each task.
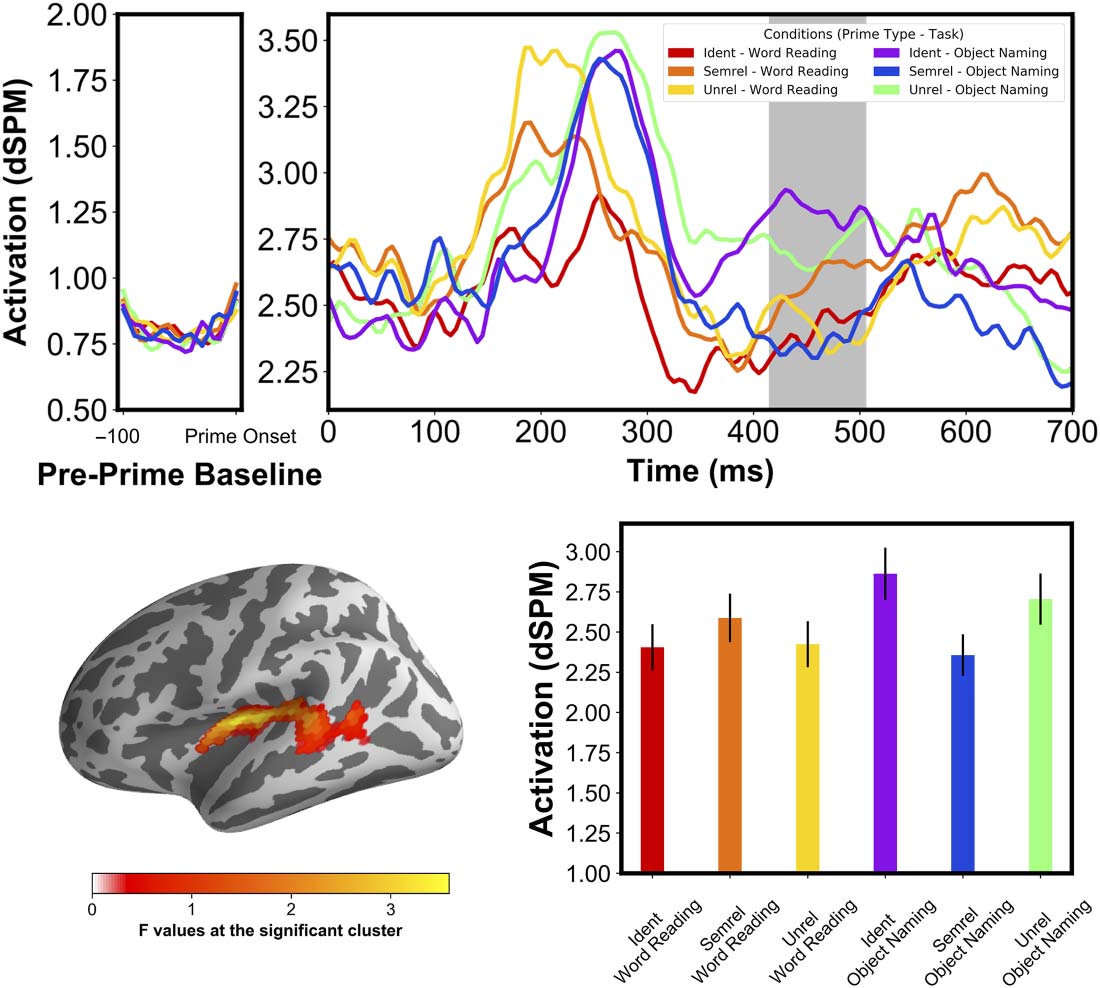
With this second analysis, we were able to observe distinct priming patterns for each task
(Figure 8A–D). For the Object Naming task, we found an early facilitation effect at 310–380 ms
followed by an interference effect at 395–485 ms. The facilitation was localized in sources
Neurobiology of Language
197
Lexical access in naming and reading
Figure 6.
Interaction cluster between Task and Prime Type in the full across-tasks analysis, show-
ing an amplitude reduction for the semantically related condition in the Object Naming task only ( p <
0.05, corr.). dSPM = dynamic statistical parameter map; Ident = identical; Semrel = semantically
related; Unrel = unrelated.
in the angular gyrus area and posterior parts of the STG. Surprisingly, this facilitation effect
did not show any significant distinction between the Semrel and Ident priming conditions.
That is, this priming effect did not distinguish between related pairs and identical pairs.
Crucially, we also found in Object Naming a late cluster at 395–485 ms ( p < .001), local-
izing at the anterior STG and the insular cortex. We observed that the identical and seman-
tically related conditions separated in opposite directions from the unrelated condition, with
higher activation for Ident and lower for Semrel primes. Specifically, there was a decrease in
activation for the Semrel priming condition compared with the Unrel condition and, impor-
tantly, an increase in activation with Ident primes that elicited a strong facilitation effect in
the behavioral data, representing overall a semantic interference pattern. This pattern is in
line with RT results where Ident primes elicited the fastest responses, followed by Unrel and
then finally Semrel primes. Crucially, the priming pattern observed was different in the Word
Reading task, where we found an early facilitation priming pattern between 180–335 ms ( p <
.001) at the middle STG and expanding to the middle MTG (Figure 8A). We observed a stepwise
increase in activation as semantic distance increased, which is typical of facilitatory priming
(Bentin, McCarthy, & Wood, 1985; Holcomb, 1988). This pattern is once again in line with
the observed RTs. This cluster appeared to be very similar to the cluster in Figure 5A, which
indicates that the stepwise priming effect found in the first analysis was likely driven by the
Word Reading task alone, especially since the second analysis did not find a similar pattern
in Object Naming.
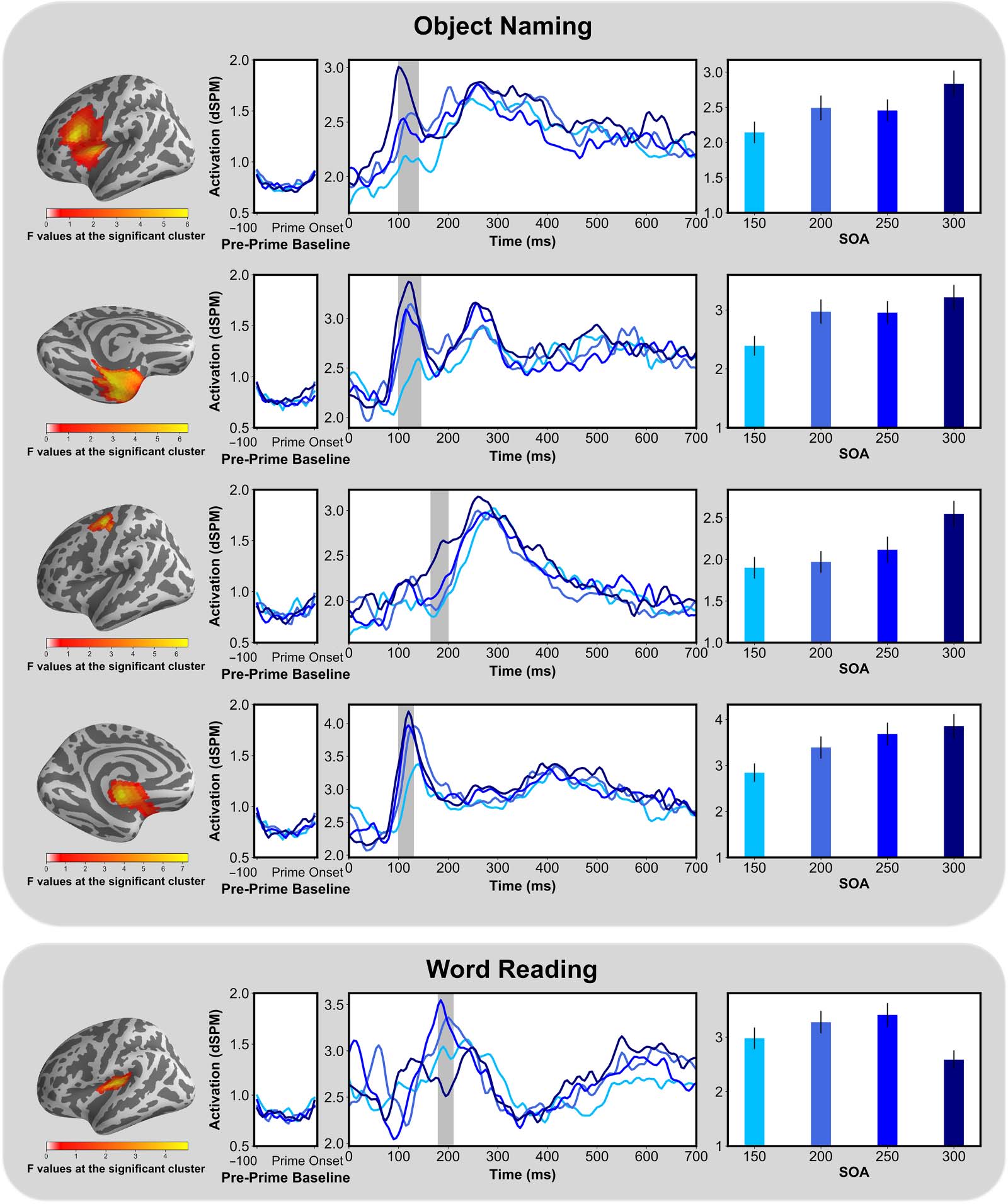
Finally, there was a main effect of SOA for both tasks (Figure 9). In Object Naming, we
found four clusters modulated by SOA, all occurring early, before 200 ms. All of the clusters
seemed to exhibit the pattern that was observed in the behavioral data, with a stepwise in-
crease in activation as SOAs got longer. In Word Reading, the posterior part of the insular
Neurobiology of Language
198
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
2
1
8
5
1
8
6
7
7
2
2
n
o
_
a
_
0
0
0
0
8
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Lexical access in naming and reading
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
2
1
8
5
1
8
6
7
7
2
2
n
o
_
a
_
0
0
0
0
8
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 7. Omnibus analysis: Spatiotemporal clusters for the main effect of SOA. The shaded regions indicate that the difference in activity
between the conditions was significant at p < 0.05. dSPM = dynamic statistical parameter map; SOA = stimulus onset asynchrony.
Neurobiology of Language
199
Lexical access in naming and reading
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
2
1
8
5
1
8
6
7
7
2
2
n
o
_
a
_
0
0
0
0
8
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 8. Within-task analysis of priming effects, showing a stepwise amplitude reduction (Ident < Semrel < Unrel) in the Word Reading task
(A) centered around the left STG ( p < 0.05, corr.). In Object Naming (bottom) we see an earlier amplitude reduction for the semantically
related and identical conditions in the angular gyrus (B) followed by a later interference pattern (Semrel < Unrel < Ident) around the STG
(C). The response-locked effects show an amplitude reduction for the semantically related condition localized at the anterior part of the in-
traparietal sulcus at −100 ms (D) and in the posterior part of the parahippocampal gyrus starting at −400 ms until −100 ms, with the seman-
tically related condition peaking at −200 ms (E). Estimated times of processing steps are based on Indefrey and Levelt (2004) and are scaled
proportionally to our observed average reaction times for each task, according to Roelofs and Shitova (2017). dSPM = dynamic statistical
parameter map; Ident = identical; Semrel = semantically related; Unrel = unrelated; STG = superior temporal gyrus.
cortex appeared to be modulated by SOA at 180–210 ms. The activations at this cluster did not
seem to follow a straightforward pattern.
Response-locked analysis within task
For the Word Reading task, we did not find any semantic facilitation effects in the speech-
locked analysis. In Object Naming, we found two clusters with distinct time courses for all
Neurobiology of Language
200
Lexical access in naming and reading
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
2
1
8
5
1
8
6
7
7
2
2
n
o
_
a
_
0
0
0
0
8
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 9. Analysis within task: Spatiotemporal clusters for the main effect of SOA, within task. The shaded regions indicate that the difference
in activity between the conditions was significant at p < 0.05. dSPM = dynamic statistical parameter map; SOA = stimulus onset asynchrony.
three priming conditions. The first cluster was located at the anterior part of the intraparietal
sulcus at −150 to −100 ms prior to the utterance time. The second cluster started at −400 ms
until −100 ms and peaked at −200 ms before utterance and spread on the posterior part of the
parahippocampal gyrus (Figure 8D–E).
DISCUSSION
Here we took advantage of the spatiotemporal resolution of MEG to disambiguate the sources
of the semantic interference and facilitation effects, which have critically shaped our under-
standing of how words are accessed in naming and reading. Our behavioral results replicated
Neurobiology of Language
201
Lexical access in naming and reading
the interference effect in naming and the facilitation effect in reading. Specifically, the fact that
semantic relatedness delays naming times has led to the hypothesis that in object naming,
semantically related words compete during lexical access. Here, we found mixed support
for this hypothesis. Our comparison of naming and reading revealed that while the semantic
facilitory effect of word reading localized in the left STG in a time window consistent with
lexical access, the interfering effect of object naming revealed a more complicated pattern.
In fact, we observed a facilitation effect at 310–390 ms in the angular gyrus and posterior
part of the STG, closely followed by a pattern consistent with interference at 395–485 ms in
the anterior STG. Our speech-locked analysis also revealed two later effects of interference
between 400–100 ms prior to word production, which is in line with motor preparatory ac-
counts of interference. Importantly, we did not find any interference effects occurring earlier
than the facilitation effect. In what follows we discuss the theoretical consequences and
empirical limitations of these findings.
Facilitation in Word Reading at 180–335 ms in Left Superior Temporal Cortex
A classic semantic priming effect exhibits a reduction of MEG signal amplitude as a function of
semantic similarity between prime and target. Thus, in our overt reading task, we expected
facilitation to manifest as lower amplitudes for the semantically related versus the unrelated
condition, followed by the lowest amplitudes for the identical condition, which involves rep-
etition of the same word. Exactly this pattern was observed in the left superior temporal cortex
at 180–335 ms, which is consistent with the localization of lexical access both in space
(Hickok & Poeppel, 2007; Hillis, Rorden, & Fridriksson, 2017; Lau, Phillips, & Poeppel,
2008) and time (Bentin et al., 1985; Indefrey & Levelt, 2004). Thus, this effect has a straight-
forward interpretation in terms of spreading activation between semantically related lexical
representations. Research on reading aloud has also shown connections between the STG
and other areas relevant for semantic processing, such as the angular gyrus and the inferior
temporal sulcus, forming a broader semantic network for reading aloud (Boukrina & Graves,
2013).
Early Facilitation and Late Interference Patterns Characterize the Behavioral Interference Effect
In contrast to word reading, our MEG data for object naming revealed a less straightforward
pattern. In the stimulus-locked analysis, we observed an early facilitation effect followed by a
later interference effect. The early facilitation effect was associated with decreased activation
in the angular gyrus and posterior STG for the identical and semantically related primes as
compared with the unrelated primes, but no distinction between the identical and semanti-
cally related primes emerged. The angular gyrus has been associated with the semantic system
and specifically with conceptual retrieval and conceptual integration (Binder, Desai, Graves,
& Conant, 2009), as well as with decreased activity for cross-modal priming effects
(Badgaiyan, Schacter, & Alpert, 1999). As previously mentioned, the STG has been associated
with lexical access (Hickok & Poeppel, 2007; Hillis et al., 2017; Lau et al., 2008). Further,
while the early effect showed a pattern where both the semantically related and identical con-
ditions separated in the same direction from the unrelated condition, the late effect patterned
such that the semantically related and identical conditions separated in opposite direction
from the unrelated condition. That is, the facilitation-eliciting identical priming showed an
increase in activation relative to the unrelated condition, while the interfering semantic primes
showed the opposite pattern: a decrease in activation compared with the unrelated condition.
This pattern is consistent with similarity-based interference between related but not identical
Neurobiology of Language
202
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
2
1
8
5
1
8
6
7
7
2
2
n
o
_
a
_
0
0
0
0
8
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Lexical access in naming and reading
meanings and is inversely related to the behavioral RTs, meaning that as activation at the
interference cluster increased, RTs decreased.
Crucially, the time resolution of our measurement allowed us to determine whether a pos-
sible interference pattern occurs early, during conceptual preparation or lexical access, or at
later, postlexical stages of processing. The stimulus-locked results conformed to the latter hy-
pothesis, showing first a facilitation at 310–390 ms in the angular gyrus and posterior part of
the STG, possibly representing lexical-level processes, followed by a later interference pattern
at 395–485 ms. Our scaled estimates of the underlying processes occurring during the 395–
485 ms time window suggest that the later interference effect occurs during phonological code
retrieval. In addition, the interference effect localized in the anterior parts of the STG and the
insular cortex. Further, the presence of an early facilitatory effect is in line with competitive
accounts of lexical access that describe a phenomenon of reverse priming that is associated
with the semantic interference effect (Piai et al., 2014; Roelofs, Piai, Rodriguez, & Chwilla,
2016), although it is also predicted by the REH (Janssen et al., 2008). While it is not possible
to determine the exact underlying processes associated with the later interference cluster, the
late timing of the cluster as well as the fact that it occurs after the facilitation effect suggests that
it is too late to reflect lexical or prelexical stages of processing. Further, the activity in the in-
sular cortex cannot be confidently interpreted since the localization of deep sources with MEG
is still controversial (Krishnaswamy et al., 2017).
The response-locked analysis revealed two different effects. The first effect was observed at
−200 ms to −100 ms prior to naming time and localized at the anterior part of the intraparietal
sulcus. This extremely late effect is estimated to occur during the very last stages of object nam-
ing, when the motor response is being planned. This is exactly in line with the response exclusion
hypothesis, which postulates that the sematic interference effect occurs as a result of a decision
that has to be made at the articulators. A core principle of the REH is that words receive privileged
access to the motor preparation system over images. As a result, the primes must be excluded
from the single-channel motor preparation buffer before the target image can be named. Still, the
mechanism by which the response buffer excludes responses according to the REH is still not
fully specified. For instance, more details regarding the mechanism of the decision process at
the buffer are required. It is worth mentioning that the spatial localization of the late effect that
we found does not occur in frontal areas such as the premotor and motor cortices, which are
usually associated with phonetic encoding and articulatory plans (Indefrey, 2011).
In contrast to this late result, the second response-locked analysis revealed a second, more
sustained effect in the posterior part of the parahippocampal gyrus starting at −400 ms until
−100 ms, with the semantically related condition peaking at −200 ms before utterance time.
After scaling the timing of this cluster to be proportional to the RTs that we observed, the effect
appears to start during lexical access stages and to sustain until motor preparation.
As opposed to the facilitation effect in reading, which localized at a single time window,
the results for the object naming task revealed that the semantic interference has multiple loci.
The semantic interference effect seems to be characterized by an early facilitation, possibly at
the lexical level, followed by two focal interference effects at the phonological retrieval and
motor preparation stages. The interference also appeared as a sustained effect starting at the
lexical level until motor preparation. Previous literature has shown variable timings for the
localization of semantic interference findings, with some effects pointing to an early, lexical
stage of processing (Aristei et al., 2011; Maess et al., 2002; Piai et al., 2014; Roelofs, 2018),
while other findings support a postlexical locus (Anders et al., 2019; Blackford et al., 2012;
Janssen et al., 2014). Evidence for both an early and a late locus of interference have also been
Neurobiology of Language
203
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
2
1
8
5
1
8
6
7
7
2
2
n
o
_
a
_
0
0
0
0
8
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Lexical access in naming and reading
found in a blocked semantic interference paradigm using intracranial EEG with eight epileptic
patients (Anders et al., 2019) where the authors found increased activity in the presupplemen-
tary motor area at 200–300 ms and at 700–800 ms, which was interpreted, respectively, as an
initial conflict resolution and as preresponse control processes. One possibility for the incon-
sistencies in the literature might come from variabilities in paradigms and neuroimaging
methods. Nevertheless, some findings point for a common lexical level locus for semantic in-
terference, which appears across many variations of the classical picture-word interference
paradigm when proportional scaling is done to account for RTs (Roelofs & Shitova, 2017).
However, some findings, including what is presented here, still found very late effects that
seem to be postlexical even after scaling (Janssen et al., 2014). If the effect indeed has multiple
loci, then it is possible that variations in the experimental paradigms and in the neuroimaging
methods might bias one of the loci of the interference effect to be picked up in some cases but
not others. What we show here is that indeed, the semantic interference effect has a complex
neural signature, consisting of both early and late effects. Specifically, the single early lexical-
level effect that we found here revealed a faciliatory pattern, while the later effects all showed
an interference pattern, including a very late motor-preparation effect.
Priming Effects Constant across SOAs
In both the omnibus and the within-task analyses of the MEG data, we found main effects of
SOA, but surprisingly, no interaction of SOA with any other factors. Crucially, SOA did not
interact with Prime Type in either the MEG or the behavioral data. We therefore conclude that
on average our priming results generalize across all the SOAs that we tested. However, in the
context of the current study, the effect of SOA is only meaningful if it interacts with the other
factors, mainly Prime Type. Because the effect of SOA did not modulate that of Prime Type, we
will not further discuss the implications of this factor. Note that there is evidence showing that
SOA manipulation can change the size and even the direction of priming, specifically in
Object Naming (Heij et al., 1990; Xavier et al., 2000), and it is likely that SOAs longer than
400 ms might interact with Prime Type and/or Task.
Widespread Effect of Task
We observed a widespread effect of Task that covered almost the entire left hemisphere,
throughout the whole analysis time window, with the most reliable signal centered at medial
occipital areas. This large effect might be driven not only by the contrasting tasks, but also by
the contrasting modalities of the primes. In fact, in Word Reading, the prime and the target are
both written words, whereas in Object Naming, the prime is a word while the target is an
image. Therefore, our widespread effect could have been driven by the tasks themselves, or
by matching vs. mismatching modalities of the prime and target. We speculate that the effect
was likely driven by both of these contrasts, given its extensive coverage in both time and
space.
Limitations and Future Directions
It is worth mentioning that one of the challenges in trying to answer the question of early vs.
late effects is in objectively defining what is late and what is early. Models of word production
lay out processing steps differently, the main distinction being serial vs. parallel models
(Hickok, 2012; Price, 2012). Here our findings are compared to estimated timings from the
serial model of Indefrey and Levelt (2004), which has received support but has also been crit-
icized for not being backed enough by neural data to justify the pure seriality of the model
Neurobiology of Language
204
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
2
1
8
5
1
8
6
7
7
2
2
n
o
_
a
_
0
0
0
0
8
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Lexical access in naming and reading
(Strijkers & Costa, 2011, 2016). For instance, there is MEG evidence of semantic and phono-
logical information beginning in parallel as early as 100–250 ms after picture onset (Blanco-
Elorrieta, Ferreira, Del Prato, & Pylkkänen, 2018; Miozzo, Pulvermüller, & Hauk, 2014;
Pulvermüller, Shtyrov, & Hauk, 2009). Furthermore, other top-down processes, such as feed-
back, have to be taken into consideration to properly make a one-to-one match between the
neural data and the models of word production (Strijkers & Costa, 2016). Nevertheless, lexical
access has been supported consistently to occur about 200 ms after stimulus onset (Strijkers &
Costa, 2011), and the debate regarding the extent of seriality and parallelism of word produc-
tion is still open. Here we lay out the estimated spatiotemporal signature of the semantic in-
terference effect based on what is currently known of the processing stages of both reading and
naming, based on a well-established model. As models of naming and reading are further de-
veloped and a consensus regarding the seriality or parallelism of processing is reached, the
understanding of electrophysiological results will hopefully also improve.
Conclusion
We used MEG to provide a spatiotemporal characterization of the neural signatures associated
with two well-established behavioral effects during lexical processing: semantic interference
in object naming and semantic facilitation in overt reading. While the facilitation in reading
was characterized only by an early lexical-level effect in the STG, the behavioral interference
effect came with a more complex neural pattern. It consisted of both an early facilitory effect in
the angular gyrus, estimated to occur during lexical processing, and a later, sustained interfer-
ence effect in multiple regions, likely occurring during both phonological retrieval and motor
preparation. These results suggest that the semantic interference effect in naming is a multi-
locus effect, which may explain the discrepancies in prior literature regarding whether it oc-
curs at the early lexical level, or at a later postlexical level.
FUNDING INFORMATION
Liina Pylkkänen, NYUAD Research Institute, Award ID: G1001.
AUTHOR CONTRIBUTIONS
Both Julien Dirani and Liina Pylkkänen designed the research and wrote the paper. Julien
Dirani performed the research and analyzed the data.
REFERENCES
Adachi, Y., Shimogawara, M., Higuchi, M., Haruta, Y., & Ochiai, M.
(2001). Reduction of non-periodic environmental magnetic noise
in MEG measurement by continuously adjusted least squares
method. IEEE Transactions on Applied Superconductivity, 11(1),
669–672.
Anders, R., Llorens, A., Dubarry, A.-S., Trébuchon, A., Liégeois-
Chauvel, C., & Alario, F.-X. (2019). Cortical dynamics of semantic
priming and interference during word production: An intracere-
bral study. Journal of Cognitive Neuroscience, 31(7), 978–1001.
http://doi.org/10.1162/jocn_a_01406
Aristei, S., Melinger, A., & Abdel Rahman, R.
(2011).
Electrophysiological chronometry of semantic context effects
in language production. Journal of Cognitive Neuroscience,
23(7), 1567–1586.
Badgaiyan, R. D., Schacter, D. L., & Alpert, N. M. (1999). Auditory
priming within and across modalities: Evidence from positron
emission tomography. Journal of Cognitive Neuroscience, 11(4),
337–348.
Bentin, S., McCarthy, G., & Wood, C. C. (1985). Event-related po-
tentials, lexical decision and semantic priming. Electroen-
cephalography and Clinical Neurophysiology, 60(4), 343–355.
Binder, J. R., Desai, R. H., Graves, W. W., & Conant, L. L. (2009).
Where is the semantic system? A critical review and meta-
analysis of 120 functional neuroimaging studies. Cerebral
Cortex, 19(12), 2767–2796.
Blackford, T., Holcomb, P. J., Grainger, J., & Kuperberg, G. R.
(2012). A funny thing happened on the way to articulation:
N400 attenuation despite behavioral interference in picture nam-
ing. Cognition, 123(1), 84–99.
Blanco-Elorrieta, E., Ferreira, V. S., Del Prato, P., & Pylkkänen, L.
(2018). The priming of basic combinatory responses in MEG.
Cognition, 170, 49–63.
Neurobiology of Language
205
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
2
1
8
5
1
8
6
7
7
2
2
n
o
_
a
_
0
0
0
0
8
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Lexical access in naming and reading
Bloem, I., & La Heij, W. (2003). Semantic facilitation and semantic
interference in word translation: Implications for models of lexi-
cal access in language production. Journal of Memory and
Language, 48(3), 468–488.
Bloem, I., van den Boogaard, S., & La Heij, W. (2004). Semantic
facilitation and semantic interference in language production:
Further evidence for the conceptual selection model of lexical
access. Journal of Memory and Language, 51(2), 307–323.
Boukrina, O., & Graves, W. W. (2013). Neural networks underlying
contributions from semantics in reading aloud. Frontiers in
Human Neuroscience, 7, 518.
Brodbeck, C. (2017). Eelbrain ( Version v0.25). Zenodo. http://doi.
org/10.5281/zenodo.438193
Costa, A., Alario, F.-X., & Caramazza, A. (2005). On the categorical
nature of the semantic interference effect in the picture-word in-
terference paradigm. Psychonomic Bulletin & Review, 12(1),
125–131.
Dale, A. M., Liu, A. K., Fischl, B. R., Buckner, R. L., Belliveau, J. W.,
Lewine, J. D., & Halgren, E. (2000). Dynamic statistical paramet-
ric mapping: Combining fMRI and MEG for high-resolution im-
aging of cortical activity. Neuron, 26(1), 55–67.
de Zubicaray, G. I., Hansen, S., & McMahon, K. L. (2013).
Differential processing of thematic and categorical conceptual
relations in spoken word production. Journal of Experimental
Psychology: General, 142(1), 131–142.
de Zubicaray, G. I., Wilson, S. J., McMahon, K. L., & Muthiah, S.
(2001). The semantic interference effect in the picture-word par-
adigm: An event-related fMRI study employing overt responses.
Human Brain Mapping, 14(4), 218–227.
Dhooge, E., & Hartsuiker, R. J. (2010). The distractor frequency ef-
fect in picture–word interference: Evidence for response exclu-
sion. Journal of Experimental Psychology: Learning, Memory,
and Cognition, 36(4), 878–891.
Engemann, D. A., & Gramfort, A. (2015). Automated model selec-
tion in covariance estimation and spatial whitening of MEG and
EEG signals. NeuroImage, 108, 328–342.
Fieder, N., Wartenburger, I., & Rahman, R. A. (2019). A close
call: Interference from semantic neighbourhood density and
similarity in language production. Memory & Cognition, 47(1),
145–168.
Finkbeiner, M., & Caramazza, A. (2006a). Lexical selection is not a
competitive process: A reply to La Heij et al. (2006). Cortex, 42(7),
1032–1036.
Finkbeiner, M., & Caramazza, A. (2006b). Now you see it, now you
don’t: On turning semantic interference into facilitation in a
Stroop-like task. Cortex, 42(6), 790–796.
Giesbrecht, F. G., & Burns, J. C. (1985). Two-stage analysis based
on a mixed model: Large-sample asymptotic theory and small-
sample simulation results. Biometrics, 477–486.
Giezen, M. R., & Emmorey, K. (2016). Language co-activation and
lexical selection in bimodal bilinguals: Evidence from picture–
word interference. Bilingualism: Language and Cognition, 19(2),
264–276.
Glaser, W. R., & Glaser, M. O. (1989). Context effects in stroop-like
Journal of Experimental
word and picture processing.
Psychology: General, 118(1), 13–42.
Gramfort, A., Luessi, M., Larson, E., Engemann, D. A., Strohmeier,
D., Brodbeck, C., … Hämäläinen, M. S. (2014). MNE software
for processing MEG and EEG data. NeuroImage, 86, 446–460.
Hala, S., Pexman, P. M., & Glenwright, M. (2007). Priming the
meaning of homographs in typically developing children and
children with autism. Journal of Autism and Developmental
Disorders, 37(2), 329.
Hickok, G. (2012). Computational neuroanatomy of speech pro-
duction. Nature Reviews Neuroscience, 13(2), 135.
Hickok, G., & Poeppel, D. (2007). The cortical organization of
speech processing. Nature Reviews Neuroscience, 8(5), 393.
Hillis, A. E., Rorden, C., & Fridriksson, J. (2017). Brain regions es-
sential for word comprehension: Drawing inferences from pa-
tients. Annals of Neurology, 81(6), 759–768.
Holcomb, P. J. (1988). Automatic and attentional processing: An
event-related brain potential analysis of semantic priming.
Brain and Language, 35(1), 66–85.
Holmes, A. P., Blair, R., Wat son,
Holcomb, P. J., & Anderson, J. E. (1993). Cross-modal semantic
priming: A time-course analysis using event-related brain poten-
tials. Language and Cognitive Processes, 8(4), 379–411.
I.
(1996).
Nonparametric analysis of statistic images from functional map-
ping experiments. Journal of Cerebral Blood Flow & Metabolism,
16(1), 7–22.
J., & Ford,
Hrong-Tai Fai, A., & Cornelius, P. L. (1996). Approximate F-tests of
multiple degree of freedom hypotheses in generalized least
squares analyses of unbalanced split-plot experiments. Journal
of Statistical Computation and Simulation, 54(4), 363–378.
https://doi.org/10.1080/00949659608811740
Indefrey, P. (2011). The spatial and temporal signatures of word
production components: A critical update. Frontiers in
Psychology, 2(255). https://doi.org/10.3389/fpsyg.2011.00255
Indefrey, P., & Levelt, W. J. (2004). The spatial and temporal sig-
natures of word production components. Cognition, 92(1–2),
101–144.
Janssen, N., Hernández-Cabrera, J. A., van der Meij, M., & Barber,
H. A. (2014). Tracking the time course of competition during
word production: Evidence for a post-retrieval mechanism of
conflict resolution. Cerebral Cortex, 25(9), 2960–2969.
Janssen, N., Schirm, W., Mahon, B. Z., & Caramazza, A. (2008).
Semantic interference in a delayed naming task: Evidence for
the response exclusion hypothesis. Journal of Experimental
Psychology: Learning, Memory, and Cognition, 34(1), 249–256.
Katz, L., & Feldman, L. B. (1983). Relation between pronunciation
and recognition of printed words in deep and shallow orthogra-
phies. Journal of Experimental Psychology: Learning, Memory,
and Cognition, 9(1), 157–166.
Kotz, S. A., Cappa, S. F., von Cramon, D. Y., & Friederici, A. D.
(2002). Modulation of the lexical–semantic network by auditory
semantic priming: An event-related functional MRI study.
NeuroImage, 17(4), 1761–1772.
Krishnaswamy, P., Obregon-Henao, G., Ahveninen, J., Khan, S.,
Babadi, B., Iglesias, J. E., … Purdon, P. L. (2017). Sparsity enables
estimation of both subcortical and cortical activity from MEG and
EEG. Proceedings of the National Academy of Sciences, 114(48),
E10465–E10474.
Kuznetsova, A., Brockhoff, P., & Christensen, R. (2017). lmerTest:
Tests in linear mixed effects models. R package version 2.0–33.
2016. Journal of Statistical Software, 82(13), 1–26.
La Heij, W. (1988). Components of Stroop-like interference in pic-
ture naming. Memory & Cognition, 16(5), 400–410.
La Heij, W., Dirkx, J., & Kramer, P. (1990). Categorical interference
and associative priming in picture naming. British Journal of
Psychology, 81(4), 511–525.
Lau, E. F., Phillips, C., & Poeppel, D. (2008). A cortical network for
semantics: (De)constructing the N400. Nature Reviews
Neuroscience, 9(12), 920–933.
Levelt, W. J. (2001). Spoken word production: A theory of lexical
access. Proceedings of the National Academy of Sciences, 98(23),
13464–13471.
Neurobiology of Language
206
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
2
1
8
5
1
8
6
7
7
2
2
n
o
_
a
_
0
0
0
0
8
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Lexical access in naming and reading
Levelt, W. J., Roelofs, A., & Meyer, A. S. (1999). A theory of lexical
access in speech production. Behavioral and Brain Sciences,
22(1), 1–38.
Lupker, S. J. (1979). The semantic nature of response competition in
the picture-word interference task. Memory & Cognition, 7(6),
485–495.
Maess, B., Friederici, A. D., Damian, M., Meyer, A. S., & Levelt,
W. J. (2002). Semantic category interference in overt picture nam-
ing: Sharpening current density localization by PCA. Journal of
Cognitive Neuroscience, 14(3), 455–462.
Mahon, B. Z., Costa, A., Peterson, R., Vargas, K. A., &
Caramazza, A. (2007). Lexical selection is not by competition:
A reinterpretation of semantic interference and facilitation
effects in the picture-word interference paradigm. Journal of
Experimental Psychology: Learning, Memory, and Cognition,
33(3), 503.
Maris, E., & Oostenveld, R. (2007). Nonparametric statistical testing
of EEG- and MEG-data. Journal of Neuroscience Methods, 164(1),
177–190.
Matsumoto, A., Iidaka, T., Haneda, K., Okada, T., & Sadato, N.
(2005). Linking semantic priming effect in functional MRI and
event-related potentials. NeuroImage, 24(3), 624–634. https://
doi.org/10.1016/j.neuroimage.2004.09.008
Miozzo, M., Pulvermüller, F., & Hauk, O. (2014). Early parallel ac-
tivation of semantics and phonology in picture naming: Evidence
from a multiple linear regression MEG study. Cerebral Cortex,
25(10), 3343–3355.
Munding, D., Dubarry, A.-S., & Alario, F.-X. (2016). On the cortical
dynamics of word production: A review of the MEG evidence.
Language, Cognition and Neuroscience, 31(4), 441–462.
Peirce, J. W. (2007). PsychoPy—psychophysics software in Python.
Journal of Neuroscience Methods, 162(1–2), 8–13.
Piai, V., Roelofs, A., Jensen, O., Schoffelen, J.-M., & Bonnefond, M.
(2014). Distinct patterns of brain activity characterise lexical ac-
tivation and competition in spoken word production. PLoS One,
9(2), e88674.
Price, C. J. (2012). A review and synthesis of the first 20 years of PET
and fMRI studies of heard speech, spoken language and reading.
NeuroImage, 62(2), 816–847.
Pulvermüller, F., Shtyrov, Y., & Hauk, O. (2009). Understanding
in an instant: Neurophysiological evidence for mechanistic
language circuits in the brain. Brain and Language, 110(2),
81–94.
R Core Team. (2013). R: A language and environment for statistical
computing. Vienna: R Foundation for Statistical Computing.
Riès, S., Janssen, N., Burle, B., & Alario, F.-X. (2013). Response-locked
brain dynamics of word production. PLoS One, 8(3), e58197.
Rissman, J., Eliassen, J. C., & Blumstein, S. E. (2003). An event-
related fMRI investigation of implicit semantic priming. Journal
of Cognitive Neuroscience, 15(8), 1160–1175.
Roelofs, A. (2003). Goal-referenced selection of verbal action:
Modeling attentional control in the Stroop task. Psychological
Review, 110(1), 88–125.
Roelofs, A. (2018). A unified computational account of cumulative
semantic, semantic blocking, and semantic distractor effects in
picture naming. Cognition, 172, 59–72.
Roelofs, A., Piai, V., Rodriguez, G. G., & Chwilla, D. J. (2016).
Electrophysiology of cross-language interference and facilitation
in picture naming. Cortex, 76, 1–16.
Roelofs, A., Piai, V., & Schriefers, H. (2011). Selective attention
and distractor frequency in naming performance: Comment
on Dhooge and Hartsuiker (2010). Journal of Experimental
Psychology: Learning, Memory, and Cognition, 37, 1032–
1038.
Roelofs, A., & Shitova, N. (2017). Importance of response time in
assessing the cerebral dynamics of spoken word production:
Comment on Munding et al. (2016). Language, Cognition and
Neuroscience, 32(8), 1064–1067.
Rose, S. B., Aristei, S., Melinger, A., & Abdel Rahman, R. (2019).
The closer they are, the more they interfere: Semantic similarity
of word distractors increases competition in language produc-
tion. Journal of Experimental Psychology: Learning, Memory,
and Cognition, 45(4), 753.
Rossell, S. L., Price, C. J., & Nobre, A. C. (2003). The anatomy and
time course of semantic priming investigated by fMRI and ERPs.
Neuropsychologia, 41(5), 550–564.
Rosson, M. B. (1983). From SOFA to LOUCH: Lexical contribu-
tions to pseudoword pronunciation. Memory & Cognition,
11(2), 152–160.
Sailor, K., & Brooks, P. J. (2014). Do part–whole relations produce
facilitation in the picture–word interference task? The Quarterly
Journal of Experimental Psychology, 67(9), 1768–1785.
Sailor, K., Brooks, P. J., Bruening, P. R., Seiger-Gardner, L., &
Guterman, M. (2009). Exploring the time course of semantic in-
terference and associative priming in the picture–word interfer-
ence task. Quarterly Journal of Experimental Psychology, 62(4),
789–801.
Starreveld, P. A., & La Heij, W. (1995). Semantic interference, or-
thographic facilitation, and their interaction in naming tasks.
Journal of Experimental Psychology: Learning, Memory, and
Cognition, 21(3), 686.
Strijkers, K., & Costa, A. (2011). Riding the lexical speedway: A crit-
ical review on the time course of lexical selection in speech pro-
duction. Frontiers in Psychology, 2, 356.
Strijkers, K., & Costa, A. (2016). The cortical dynamics of speaking:
Present shortcomings and future avenues. Language, Cognition
and Neuroscience, 31(4), 484–503.
Van Essen, D. C. (2005). A population-average, landmark-and
surface-based (PALS) atlas of human cerebral cortex. NeuroImage,
28(3), 635–662.
Vinogradov, S., Ober, B. A., & Shenaut, G. K. (1992). Semantic
priming of word pronunciation and lexical decision in schizo-
phrenia. Schizophrenia Research, 8(2), 171–181.
Xavier, A., Segui, J., & Ferrand, L. (2000). Semantic and associative
priming in picture naming. The Quarterly Journal of Experimental
Psychology: Section A, 53(3), 741–764.
Neurobiology of Language
207
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
2
1
8
5
1
8
6
7
7
2
2
n
o
_
a
_
0
0
0
0
8
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3