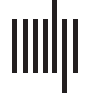RESEARCH
A novel dynamic network imaging analysis method
reveals aging-related fragmentation of
cortical networks in mouse
Daniel A. Llano1,3,4
, Chihua Ma2, Umberto Di Fabrizio2, Aynaz Taheri2, Kevin A. Stebbings3,4,
Georgiy Yudintsev3,4, Gang Xiao1,4, Robert V. Kenyon2, and Tanya Y. Berger-Wolf2,5
1Department of Molecular and Integrative Physiology, University of Illinois at Urbana-Champaign, Champaign, IL, USA
2Department of Computer Science, University of Illinois at Chicago, Chicago, IL, USA
3Neuroscience Program, University of Illinois at Urbana-Champaign, Champaign, IL, USA
4Beckman Institute for Advanced Science and Technology, Urbana, IL, USA
5Current affiliation: Department of Computer Science and Engineering, The Ohio State University, Columbus, OH, USA
Keywords: Network analysis, Aging, Auditory cortex, Motor cortex, NMDA, Random forest,
Presbycusis
ABSTRACT
Network analysis of large-scale neuroimaging data is a particularly challenging computational
problem. Here, we adapt a novel analytical tool, the community dynamic inference method
(CommDy), for brain imaging data from young and aged mice. CommDy, which was inspired
by social network theory, has been successfully used in other domains in biology; this report
represents its first use in neuroscience. We used CommDy to investigate aging-related changes
in network metrics in the auditory and motor cortices by using flavoprotein autofluorescence
imaging in brain slices and in vivo. We observed that auditory cortical networks in slices taken
from aged brains were highly fragmented compared to networks observed in young animals.
CommDy network metrics were then used to build a random-forests classifier based on NMDA
receptor blockade data, which successfully reproduced the aging findings, suggesting that the
excitatory cortical connections may be altered during aging. A similar aging-related decline in
network connectivity was also observed in spontaneous activity in the awake motor cortex,
suggesting that the findings in the auditory cortex reflect general mechanisms during aging.
These data suggest that CommDy provides a new dynamic network analytical tool to study the
brain and that aging is associated with fragmentation of intracortical networks.
AUTHOR SUMMARY
In this work, we introduce a novel technique (the community dynamic interference method,
CommDy) which is based on social network theory, to analyze neural data. We use CommDy
to analyze imaging data from the cerebral cortex of awake mice and mouse brain slices. In
each case, data were obtained from young and aged mice. Using CommDy, we found that in
both types of data, aging is associated with a fragmentation of cortical circuitry. Specifically,
the strength of intracortical connectivity diminishes with aging, leaving fragmented cortical
networks. Thus, CommDy is a novel tool now available to the neuroscience community, and
has revealed aging-related pathology of cortical networks.
a n o p e n a c c e s s
j o u r n a l
Citation: Llano, D. A., Ma, C., Di
Fabrizio, U., Taheri, A., Stebbings,
K. A., Yudintsev, G., … Berger-Wolf,
T. Y. (2021). A novel dynamic network
imaging analysis method reveals
aging-related fragmentation of cortical
networks in mouse. Network
Neuroscience, 5(2), 569–590. https://doi
.org/10.1162/netn_a_00191
DOI:
https://doi.org/10.1162/netn_a_00191
Supporting Information:
https://doi.org/10.1162/netn_a_00191
Received: 24 July 2020
Accepted: 11 March 2021
Corresponding Author:
Daniel A. Llano
d-llano@illinois.edu
Handling Editor:
Marcus Kaiser
Copyright: © 2021
Massachusetts Institute of Technology
Published under a Creative Commons
Attribution 4.0 International
(CC BY 4.0) license
The MIT Press
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
5
2
5
6
9
1
9
2
6
5
8
9
n
e
n
_
a
_
0
0
1
9
1
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Fragmentation of aging mouse cortex revealed by network analysis
INTRODUCTION
Normal aging is associated with a gradual loss of cognitive function (Glisky, 2007; Horn, 1982;
Horn & Cattell, 1967; Salthouse, 2003; Todd, Andrews, & Conlon, 2019). The mechanisms re-
sponsible for this cognitive loss are not yet known, but given the aging of the population (Beard,
Officer, & Cassels, 2016), it will be important to understand how brain networks fail with aging.
Structural changes in the aging brain have been investigated and are characterized by changes in
cortical thickness (Salat et al., 2004; Thambisetty et al., 2010), synaptic density (Huttenlocher,
1979; Liu, Erikson, & Brun, 1996; Masliah, Mallory, Hansen, DeTeresa, & Terry, 1993), and se-
lective loss of inhibitory interneurons (Caspary, Ling, Turner, & Hughes, 2008; Rozycka & Liguz-
Lecznar, 2017). Less well characterized are functional changes in cortical physiology with aging,
such as changes in functional connectivity. Functional connectivity between brain regions can
change rapidly over time (Hutchison et al., 2013; Robinson, Atlas, & Wager, 2015), is not easily
predictable from anatomical connectivity (Vanduffel, Payne, Lomber, & Orban, 1997), and is
altered in several different pathological states (Damaraju et al., 2014; Damoiseaux, Prater,
Miller, & Greicius, 2012; Schmidt, Carpenter-Thompson, & Husain, 2017; Supekar, Menon,
Rubin, Musen, & Greicius, 2008). In addition, cortical networks appear particularly vulnerable
to aging and demonstrate diminished network-level functional connectivity over the lifespan
(Andrews-Hanna et al., 2007; Ash et al., 2016; Battaglia et al., 2020; Tomasi & Volkow,
2012; Wen et al., 2020). Furthermore, aging-related disruptions in functional connectivity
correlate with declines in cognitive performance (Onoda, Ishihara, & Yamaguchi, 2012).
However, current approaches to examine functional connectivity typically average data over
long periods of time relative to timescales relevant for cognitive functions and do not provide
a clear mechanism to characterize the dynamics of functional connectivity over time. Therefore,
techniques that are able to extract the dynamics of functional connectivity from brain imaging
data will be of great value to those studying the impact of aging on the brain and to the broader
neuroscience community.
Network analysis tools are emerging approaches to understand functional connectivity of the
brain (Ash et al., 2016; Bullmore & Sporns, 2009; Chen, Cai, Ryali, Supekar, & Menon, 2016;
Karwowski, Vasheghani Farahani, & Lighthall, 2019; Rossini et al., 2019; Sporns, 2013).
Unfortunately, network analysis of brain imaging data has proven to be a particularly challeng-
ing computational problem. Brain activity is intrinsically highly dynamic, whereby functional
associations between neurons and brain regions ebb and flow as the organism’s level of arousal
or focus of attention changes. Many previous network methodologies, particularly those based
on blood flow signals, relied on collecting time series over long periods of time to build maps
(Beckmann, DeLuca, Devlin, & Smith, 2005; V. Calhoun et al., 2001; Vincent et al., 2007), and
the dynamic information extracted from these networks is lost. There have been recent attempts to
ameliorate this problem by examining functional connectivity over time. These approaches often,
though not always (W. H. Thompson, Richter, Plavén-Sigray, & Fransson, 2018), consisted of
choosing a window of time for analysis and sliding this window along the period of data acquisi-
tion (V. D. Calhoun, Miller, Pearlson, & Adalı, 2014; Pedreschi et al., 2020; G. J. Thompson et al.,
2013). Some of the problems with these approaches have been (a) the analysis windows (typically
about 30–60 s) are still long relative to the cognitive processes of interest that evolve over hundreds
of milliseconds; (b) choosing smaller windows tends to diminish signal-to-noise (SNR) ratios or
are employed on datasets with lower spatial resolution, such as magnetoencephalography
(Brookes et al., 2016); and (c) noise may be nonstationary and therefore introduce spurious “net-
works” along the time course of analysis (Hutchison et al., 2013).
Previous investigators have suggested that complex dynamic networks, be they populations
of neurons, large ecosystems, or social networks, share common underlying organizational
Network Neuroscience
570
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
5
2
5
6
9
1
9
2
6
5
8
9
n
e
n
_
a
_
0
0
1
9
1
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Fragmentation of aging mouse cortex revealed by network analysis
motifs (Bullmore & Sporns, 2009; Holme & Saramäki, 2012; Hopfield, 2008; Milo et al., 2002;
Watts & Strogatz, 1998). Therefore, common methods may be used to describe and under-
stand these networks. One dynamic network analysis method, known as the community dy-
namic inference method (CommDy) (Berger-Wolf, Tantipathananandh, & Kempe, 2010),
which is grounded in social network theory, groups nodes into communities (functional clus-
ters) in a way that minimizes the overall changes in the community affiliations of individuals
over time. CommDy was developed specifically to characterize dynamics and was originally
applied to the study of social networks, which is an area of study that has been plagued by the
same conceptual and computational bottlenecks seen in brain imaging data analysis. For ex-
ample, CommDy has successfully characterized group behavior of sheep (Barale, Kulahci,
Sulo, Berger-Wolf, & Rubenstein, 2010) and equids (Rubenstein, Sundaresan, Fischhoff,
Tantipathananand, & Berger-Wolf, 2015), as well as social interactions among groups of peo-
ple (Chayant Tantipathananandh, Berger-Wolf, & Kempe, 2007), although it has not yet been
applied to neuroscience. A useful property of CommDy is that it allows networks to be char-
acterized by a series of unitless metrics that correspond to the descriptions corresponding to
metrics in social networks (e.g., “community size,” “visiting,” etc. See Table 1 for a listing).
Such metrics provide additional information beyond more commonly applied static metrics.
In the current report, we adapt CommDy for the analysis of flavoprotein autofluorescence
imaging data from the mouse cerebral cortex in young and aged animals obtained using both
brain slice and whole-animal approaches. Flavoprotein autofluorescence imaging capitalizes
on intrinsic fluorescence that occurs in mitochondrial flavoproteins as neurons become active.
We and others have found that such signals are stable over time and are highly sensitive to
neuronal activity (D. A. Llano, Theyel, Mallik, Sherman, & Issa, 2009; Reinert, Gao, Chen, &
Ebner, 2007; Shibuki et al., 2003). Here, we examine changes in the auditory and motor cor-
tices. These brain regions were chosen based on ease of data acquisition and have both been
Flavoprotein autofluorescence:
An imaging method that uses
changes in the redox state of
mitochondria as an indicator of
neuronal activity. Activated
mitochondria fluoresce green when
stimulated with blue light.
Observed (OBS)
Number of time steps a node (pixel) was active or observed
Table 1. Descriptions of network metrics
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
5
2
5
6
9
1
9
2
6
5
8
9
n
e
n
_
a
_
0
0
1
9
1
p
d
.
t
Switching cost (SW)
Number of community switches made by an individual (normalized by the number of time
steps an individual was observed)
Visiting cost ( VIS)
Number of visits made by an individual to another community (normalized by the number
of time steps an individual was observed)
Absence cost (ABS)
Number of absences of an individual from a community (normalized by the number of time
steps an individual was observed)
Community stay (AS, MS)
Average (maximum) number of consecutive time steps an individual stayed as a member of
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Homing (HOM)
the same community over the time steps the individual was observed
Fraction of individual’s current community comembers, at each time step, who were
comembers in the previous time step (normalized by the number of time steps an
individual was observed)
Ave. group size (GS)
Average size of the groups (number of members and absents but not visitors) of which an
individual was a member
Community time span (TS)
Average span of the communities (the last time step minus the first time step of the
community’s existence) with which an individual was affiliated (as a member or absent)
Ave. community size (CS)
Average size of the communities (number of members and absents, but not visitors) with
which an individual was affiliated (as a member or absent)
Network Neuroscience
571
Fragmentation of aging mouse cortex revealed by network analysis
Paroxysmal depolarization:
A change in voltage in a neuron that
represents a brief period of extreme
excitability. These depolarizations
are typically seen during seizures
or events that predict the later
development of seizures.
GABA:
Gamma-aminobutyric acid, which is
the main inhibitory neurotransmitter
in the brain.
Degree:
The number of other nodes with
which one node shares a functional
connection, as determined by
correlation coefficient.
shown to show significant vulnerability to aging (Engle & Recanzone, 2012; Hirai et al., 1996;
Mendelson & Ricketts, 2001; Oliviero et al., 2006; Turner, Hughes, & Caspary, 2005; Willott,
Aitkin, & McFadden, 1993). Despite our growing understanding of the changes that occur in
the cortex with aging, we lack a network-level description of how aging impacts the cerebral
cortex and what synaptic changes may give rise to these network-level differences. Therefore,
in this study, we adapt CommDy for the analysis of neuroimaging data and compare network-
level changes in the auditory and motor cortices of young and aged mice.
RESULTS
Two datasets were examined in this study: brain slice imaging data from the auditory cortex during
paroxysmal depolarizations triggered by GABA blockade, and in vivo imaging data from the
awake mouse motor cortex (see Figure 1). These regions and types of preparations were used
for reasons of experimental convenience and illustrate the flexibility of the CommDy technique
to handle different types of datasets. Initial development of CommDy was done on slice data,
where the SNR is high, and then adapted for in vivo spontaneous activity data.
Adaptation of CommDy for Analysis of Neural Data
Prior to applying the CommDy technique, there are several data preprocessing steps required
to transform raw brain imaging data into CommDy input, including using the time series of
each pixel to construct a simple network representation, and then static community detection
in each time step. After that, CommDy was applied to infer dynamic communities. “Static”
networks refer to temporal networks with multiple snapshots, and “dynamic” networks refer
to those networks linked over time. To create the network representation, pixels were used as
nodes and a weighted, thresholded network of pixel value correlations across an empirically
determined number of time steps (the “window size”) was then generated. The correlation
threshold was determined empirically, and correlation values above the threshold were used
to build the correlation matrix.
To determine the appropriate time window and correlation thresholds for CommDy anal-
ysis applied to flavoprotein autofluorescence imaging data for slice data, the analysis window
size was systematically changed from 25 frames (352 ms) to 200 frames (2,817 ms) and the
resulting spatial distributions of the average degree of each node were examined (see
Supporting Information Figure S1). For each analysis window (25, 50, 100, or 200 frames),
the correlation coefficient for each pair of pixels was computed for the time segment where
the analysis window straddled the peak of the response. The average degree of each pixel was
then computed as the number of other pixels with which it had correlation coefficients in the
range described above each column. The degree of each node was normalized to the maxi-
mum degree for each image. Each image, therefore, contains the spatial distribution of normal-
ized node degree for a given analysis window size and given range of correlation coefficient.
At an analysis window of 25 frames, high numbers of correlations were seen in portions of the
auditory cortex at correlation coefficient thresholds between 0.7 and 0.9, but at thresholds
above 0.9, spurious correlations were detected outside of the slice (dashed arrow, top right
image). Using a 50-frame window, nodes restricted to the auditory cortex were seen at all
thresholds above 0.7. Similar findings were seen at 100 and 200 frames. Given the increased
utility of any dynamic network analysis tool at shorter analysis windows (and therefore higher
temporal resolution), it was determined that the smallest window not producing spurious
correlations would be used for analysis. Similar findings were seen for in vivo data (data
not shown). Therefore 50 frames and a correlation threshold of 0.7 (corresponding to the
Network Neuroscience
572
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
5
2
5
6
9
1
9
2
6
5
8
9
n
e
n
_
a
_
0
0
1
9
1
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Fragmentation of aging mouse cortex revealed by network analysis
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
5
2
5
6
9
1
9
2
6
5
8
9
n
e
n
_
a
_
0
0
1
9
1
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
(A) Left: diagram of imaging setup for slice. The brain slice is illuminated with blue light and green light is collected using an
Figure 1.
epifluorescence setup. Right: a series of images during a paroxysmal depolarization, shown as change in fluorescence over baseline for each
pixel. (B) Left: diagram of imaging setup for in vivo experiments. Right: two representative regions of interest on the surface of the mouse brain,
illustrating oscillations in the raw signals. (C) Analysis pipeline showing each step of the analysis. Step 1: raw images are obtained; Step 2: time
step is defined as a small (sliding) window of time, and correlation networks are computed for each time step; Step 3: the Louvain algorithm is
applied to the correlation network to infer the community structure of each time step; and Step 4: CommDy is applied to the sequence of time
steps of communities to infer cohesive dynamic community structure. Note that the two right-most images in C show the same temporal
networks with the community labels changed from independent for each time step to cohesive over the timeline.
Louvain:
A method to detect groups of
functionally-connected nodes based
on optimization of modularity.
red box in Supporting Information Figure S1) were the window and threshold used for all sub-
sequent analyses.
By sliding the correlation window one step or frame each time following each correlation
extraction, and doing this over the entire time line, a time series of correlation networks was
obtained. CommDy can take the resulting dynamic network as the input directly or, for more
efficient processing, it takes any grouping of the time steps as input. In each time step of these
time series, we applied the Louvain static community inference method (Blondel, Guillaume,
Network Neuroscience
573
Fragmentation of aging mouse cortex revealed by network analysis
Lambiotte, & Lefebvre, 2008) to find snapshots of functional clusters, the “groups” that form
the basis of CommDy. The Louvain algorithm was chosen because among the modern static
community inference methods, its steps are highly intuitive and it is highly scalable, while
achieving results comparable to other methods. The algorithm is based on two steps that
are repeated iteratively to optimize the modularity in the network (Newman, 2006). First,
the algorithm groups nodes into communities to maximize modularity locally. Then the algo-
rithm hierarchically rebuilds the network, aggregating the newly discovered communities into
supernodes. Thus, the nodes in this new network are communities discovered in the previous
step, and the new links between those nodes are weighted by the cumulative weight of the
links between the old nodes that were aggregated into the corresponding communities. The
two steps are repeated iteratively until no further increase in modularity is possible.
To measure the stability of the Louvain algorithm, which is stochastic in nature, we examined
six paroxysmal depolarizations in young mice and six paroxysmal depolarizations in aged mice.
The Louvain algorithm was run 30 times on each paroxysmal depolarization, and we computed
the standard deviation of each modularity value. We found that all standard deviations were
−3, which is lower than previous reports (Blondel et al., 2008), and, for modularity
below 5 × 10
values above 0.5, the coefficient of variation was below 0.5%, suggesting that these modularity
values are stable and the identified static communities are consistent across repetitions.
To understand how the clusters (communities inferred by Louvain algorithm in each time
step) change over time, we use the CommDy method (Berger-Wolf et al., 2010; Chayant
Tantipathananandh & Berger-Wolf, 2009; Chayant Tantipathananandh et al., 2007; C.
Tantipathananandh & Berger-Wolf, 2011). In CommDy, dynamic communities are essentially
viewed as dynamic clusters, where the membership of the individual (represented by a node in
each time step) inside the cluster is determined by the total value of the “social cost,” as man-
ifested by the individual’s interactions over time, aiming to minimize interactions outside an
individual’s community as well as switching among communities. The inferred community
structure parsimoniously minimizes the overall social cost for all the individuals by using
the deterministic constant-factor approximation algorithm (Chayant Tantipathananandh &
Berger-Wolf, 2009). The definition of social cost is based on two explicit assumptions about
individual behavior, motivated by research in social sciences and further supported by the
definition of the dynamic community as a cluster. First, it assumes that individuals tend not
to change their “home” community affiliation too often (Backstrom, Huttenlocher, Kleinberg,
& Lan, 2006). Second, it assumes that individuals tend to interact with their respective home
communities most of the time (Wasserman & Faust, 1994). These assumptions are translated
into three cost parameters potentially incurred by an individual. First, the CommDy method
posits a cost for a switch from one community to another. Second, there is a cost of visiting a
community of which one is not a member. Third, in datasets for which not all individuals are
observed all the time, there is a cost of absence for an individual who is not observed at a
gathering of a community of which it is a member. A dynamic community is then defined
as a time series of sets of individuals among whom the overall social cost of interacting is
minimized (Berger-Wolf et al., 2010). Note that these costs have not yet been defined for brain
networks and were set to equal values for our initial studies, with subsequent exploration of
the range of the parameters. For visualization, once communities were assigned, the pixels
belonging to the top 20 communities over the entire time line were distinguished using a palette
of 20 colors (Figure 2).
CommDy can be used to quantitatively describe network activity in terms of the node and
structural network metrics based on network theory (Rubenstein et al., 2015). These metrics
permit quantitative comparisons between different networks (e.g., comparing the cortical
Network Neuroscience
574
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
5
2
5
6
9
1
9
2
6
5
8
9
n
e
n
_
a
_
0
0
1
9
1
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Fragmentation of aging mouse cortex revealed by network analysis
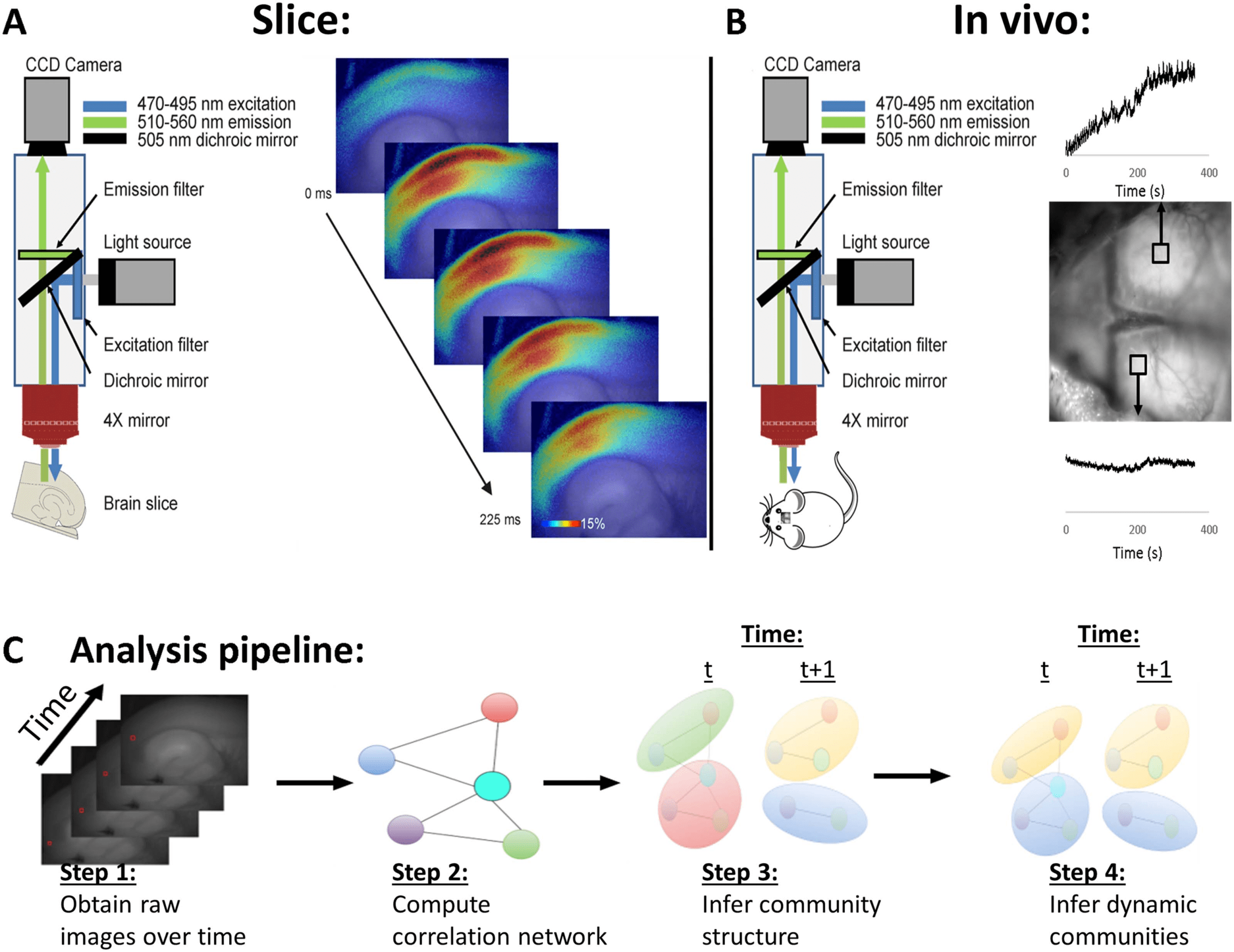
Figure 2. Young animals show greater average degree of each node than older animals. (A) Average normalized degree of each node, averaged
across all five animals, superimposed upon an averaged anatomical map. (B) Identical analysis, but for aged animals. (C) Bar graph showing that
the average degree of each node is significantly higher in the young than the aged animals. Error bars = standard deviation; **p = 0.005. See text
for details.
networks in aged vs. young animals). Therefore, quantitative analyses of CommDy activation
patterns were performed using 10 relevant community metrics that describe the interactions
between pixels (see Table 1 for a listing and definition of the metrics).
Impact of Aging on Network Dynamics in Brain Slice Preparations
Paroxysmal depolarizations were imaged in brain slices taken from five young (all were
5.5 months old) and five aged (all were 22 months old) animals (a total of 41 and 30 activations,
respectively). Initially, static analysis was performed on auditory cortical networks in young and
aging mice. To do this, the average degree of each node was computed for each pixel as the
average number of edges at each node during the 100-frame window surrounding the peak of
the paroxysmal depolarization. These values were then averaged across paroxysmal depolariza-
tions within each animal, normalized to the overall magnitude of the response, and the resulting
grand average for each group of animals is shown in Figure 2. As shown, the average normalized
degree of each node was substantially higher in younger animals than in aged animals (mean
normalized node degree in young animals = 813.0 ± 353.6 [SD] edges per active node, mean in
aged animals = 149.1 ± 162.4 [SD] edges per active node, p = 0.005, Mann–Whitney). These
data suggest that the average connectivity between nodes is higher in younger animals than in
aging animals and cannot be accounted for by any differences in the magnitude in the response
of the young versus aged animals.
Average degree of each node does not provide insights into the changing associations
between nodes. We therefore performed dynamic analysis using CommDy, which is built upon
static communities detected by the Louvain algorithm independently in each time step.
Dynamic analysis of paroxysmal depolarizations in young and aged animals using CommDy
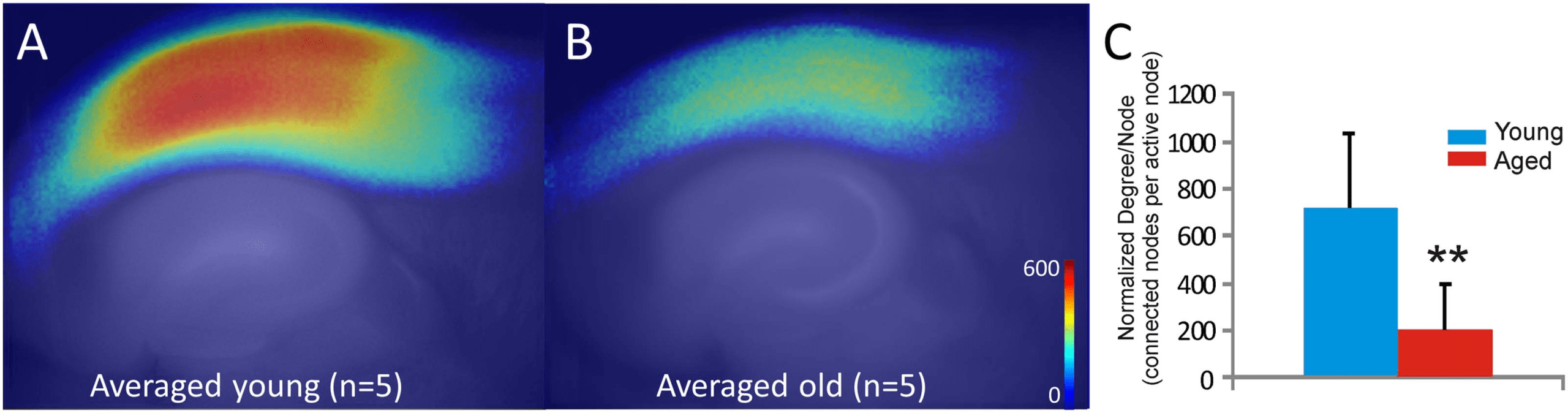
revealed that network activity differed between these two groups. Young animals showed
distinct patterns of activity across the auditory cortex during paroxysmal depolarizations. See
Figure 3A–3E for an example from one representative young mouse. Early during the response,
multiple small clusters (communities) were observed (Figure 3B). As the response evolves, a
distinct cluster (colored red) emerges in the upper layers of the auditory cortex (Figure 3C), which
then spreads to produce two broad red networks – one in the upper layers and another in the
lower layers – which persists to at least 500 msec after onset (Figure 3E; see Supporting
Information Movie 1). In the aged animals, there is also broad activation across the auditory
Network Neuroscience
575
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
5
2
5
6
9
1
9
2
6
5
8
9
n
e
n
_
a
_
0
0
1
9
1
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Fragmentation of aging mouse cortex revealed by network analysis
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
Figure 3. Representative CommDy population dynamics across time for a paroxysmal depolarization in a young animal (A–E) and an aged
animal (F–J). Each color represents a different community. (K) Bar graph of average CommDy metrics for young (n = 40 activations) and aged
(n = 31 activations) animals, normalized to the average value for young animals. Group size, community size, average stay, and maximum stay
are normalized for the total number of activated pixels. OBS = observed; TS = community time span; SW = switching cost; ABS = absence cost;
VIS = visiting cost; HOM = homing; GS = average group size; CS = average community size; AS = average community stay; MS = maximum
community stay. Fuller explanation of these metrics is found in Table 1. Error bars = standard error; *p < 0.05.
f
/
/
/
/
/
5
2
5
6
9
1
9
2
6
5
8
9
n
e
n
_
a
_
0
0
1
9
1
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
cortex. However, the activation is much more heterogeneous than that seen in the young animal
and comprises nodes of many different communities and never coalesces into coherent clusters
seen in the young animal (see Figure 3F–3J for a representative aged mouse; see Supporting
Information Movie 2).
CommDy network metrics were compared between young and aged mice. To ensure that
differences in network measures were not due to changes in the absolute magnitude of activation,
individual group and community sizes were normalized by the total number of nodes in each
activation. Using a mixed-effects model, age was found to be a significant predictor for both
normalized group size (mean values = 0.211 ± 0.113 [SD] for young vs. 0.058 ± 0.046 [SD] for
aged; p = 0.0001) and normalized community size (mean values = 0.235 ± 0.142 [SD] for young
vs. 0.046 ± 0.044 [SD] for aged; p = 0.0002). The differences in homing, a measure of network
cohesion, trended toward significance but did not survive correction for multiple comparisons
(mean values = 0.756 ± 0.663 [SD] for young vs. 0.125 ± 0.081 [SD] for aged; p = 0.0168).
See Figure 3K for a comparison of all 10 network metrics.
Impact of Hearing Loss and Cortical Thickness on CommDy Parameters
Age, cortical thinning and hearing loss are known to be correlated (Ha et al., 2020; Li et al.,
2012; Salat et al., 2004; Stebbings et al., 2016). Therefore, to disentangle these influences on
Network Neuroscience
576
Fragmentation of aging mouse cortex revealed by network analysis
Auditory brainstem response:
A surface electrophysiological tool to
measure peripheral hearing. When
sounds reach the brain, they cause a
series of small voltage deflections
that can be detected with a surface
electrode, and the minimal sound
amplitude that elicits this response is
known as the hearing threshold.
NMDA Receptor:
A subtype of glutamate receptor that
is in part responsible for excitatory
connections between neurons and is
involved when synaptic strength
changes over time.
APV:
A drug that blocks NMDA receptors.
cortical network activity, we asked whether the aging-related changes in group and commu-
nity size could be better explained by cortical thickness or hearing loss. We first measured the
differences in hearing threshold between the groups, and, as expected, differences in aged and
young animals were noted in hearing thresholds (33.9 ± 2.2 [SD] dB SPL [sound pressure level]
for young and 59.9 ± 27.6 [SD] dB SPL for aged, p = 0.032, Mann–Whitney; Supporting
Information Figure S2A). Activations were examined from animals with high auditory brainstem
response (ABR) thresholds (≥45 dB SPL, n = 3 mice) and compared to those with low ABR
thresholds (<45 dB SPL, n = 7 mice). Similar to separation of groups based on age (n = 5 per
group), using a mixed-effects model, normalized group size, and normalized community size
were both larger in the better hearing animals. The values for normalized group size = 0.189 ±
0.070 (SD) for low ABR threshold versus 0.120 ± 0.052 (SD) for high ABR threshold, ( p =
0.0002, Supporting Information Figure S2B). The values for normalized community size =
0.215 ± 0.058 (SD) for young vs. 0.146 ± 0.052 (SD) for aged ( p = 0.0001, Supporting
Information Figure S2C). In addition, as previously described in a related dataset (Stebbings
et al., 2016), we observed differences in cortical thickness between the two groups (1.20 ±
0.04 [SD] mm for young and 1.04 ± 0.04 [SD] mm for aged, p = 0.012, Mann–Whitney;
Supporting Information Figure S3A). Partial correlations between cortical thickness and group
size as well as between cortical thickness and community size were computed. Partial corre-
lations were used to account for the impact of different numbers of activations in different
animals. Significant correlations were observed in both cortical thickness versus group size
(r = 0.734, p < 0.0001; Supporting Information Figure S3B) and cortical thickness versus com-
munity size (r = 0.722, p < 0.0001; Supporting Information Figure S3C). Given the multiple
potential related predictors (age, hearing loss, cortical thickness) of group and community size,
a linear mixed model multivariate regression was conducted using all three predictors. For
community size, age was the only significant predictor ( p = 0.011), whereas for group size,
cortical thickness was the only significant predictor ( p = 0.046; see Supporting Information
Table S1).
Aging-Related Changes Are Related to Declines in Intracortical Connectivity
To test whether the aging-related changes in group and community size described above may
be related to changes in intracortical connectivity, diminished intracortical connectivity was
pharmacologically simulated in a coronal brain slice from a young animal by blocking NMDA
receptors, which are enriched in cortico-cortical synapses (Fleidervish, Binshtok, & Gutnick,
1998; Gil, Connors, & Amitai, 1999). Unlike the thalamocortical slice described above, this
preparation does not contain connectivity between subcortical nuclei and the cortex and thus
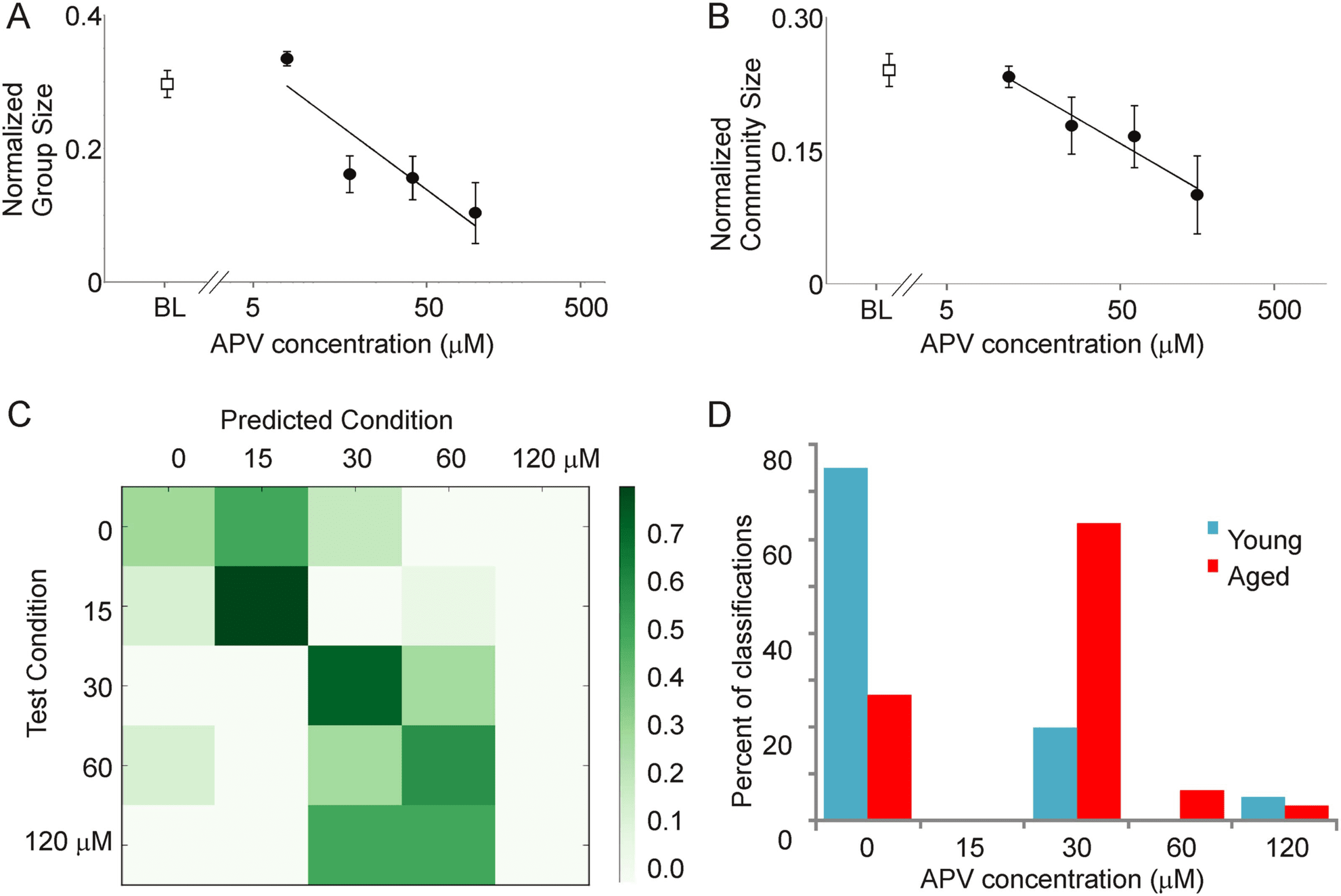
provides an indicator of the impact of intracortical connections. A concentration-dependent
drop in both normalized group and community size was seen with bath application of the
−9 for group size, p = 0.004 for community
NMDA blocker APV (1-way ANOVA, p = 5 × 10
size; Figure 4A and 4B). To further determine whether network changes observed during
NMDA blockade resembled network changes that occur during aging, a classifier was built
using the averaged metrics from Table 1 obtained for the NMDA blockade data as input fea-
tures, and the ability of this classifier to distinguish patterns of activity in young versus aged
was measured. A random forest method (Breiman, 2001) was used to build a classifier by using
the five different classes of NMDA blockade data (baseline, 15, 30, 60, and 120 μM APV).
Given the five dose categories, chance performance of the classifier = 0.2 (or 0.35 using
the majority class as the default prediction). Using 50 trees and leave-one-out validation,
the accuracy of the classifier for APV data = 0.575, which is greater than that expected by
chance (see confusion matrix, Figure 4C). When applied to all data obtained from young
Network Neuroscience
577
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
2
5
6
9
1
9
2
6
5
8
9
n
e
n
_
a
_
0
0
1
9
1
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Fragmentation of aging mouse cortex revealed by network analysis
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
2
5
6
9
1
9
2
6
5
8
9
n
e
n
_
a
_
0
0
1
9
1
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Impact of increasing concentrations of bath-applied APV at concentrations of 15 (n = 14 activations), 30 (n = 9), 60 (n = 9), and 120 μM
Figure 4.
(n = 5) on normalized group size (A) and normalized community size (B) in a coronal slice preparation from a young adult mouse. BL = baseline
values prior to adding APV (n = 10). (C) Confusion matrix showing the performance of the random forest classifier. True categories are shown in
the rows, and predicted category is shown in the columns. Most data are seen on or near the diagonal, yielding an accuracy of 0.575. (D) Using the
same model to classify data from young versus aged mice, most activations from young animals (30/40) fall correctly into the baseline (no drug)
category, while most of the activations from aged animals (22/30) fall into a drug category.
and aged animals, this classifier identified 30 out of 40 young as belonging to the baseline
group and 22 out of 30 aged as belonging to one of the APV groups, thus “correctly” identi-
fying 74% of the activations (Figure 4D). When comparing the 30-μM dose group (the cate-
gory into which most of the aged data fell) to the baseline group, the overall accuracy of the
model was 0.70. Thus, using entirely different datasets for model development and model
validation, these data suggest that the aged data, when looked at from a network perspective,
bear strong commonalities to the effect of bathing a young slice in 30 μM APV. These data also
suggest that more complete blockade, as is present with 120 μM APV, does not replicate
aging, suggesting that aging is associated with a partial loss of cortico-cortical connectivity.
Impact of Aging on Network Dynamics In Vivo
Nine young (average age = 4.7 months, range = 4.4 to 5.0 months) and nine aged (average age =
23.2 months, range = 23.0 to 23.5 months) mice were used for these experiments. Because
Network Neuroscience
578
Fragmentation of aging mouse cortex revealed by network analysis
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
2
5
6
9
1
9
2
6
5
8
9
n
e
n
_
a
_
0
0
1
9
1
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
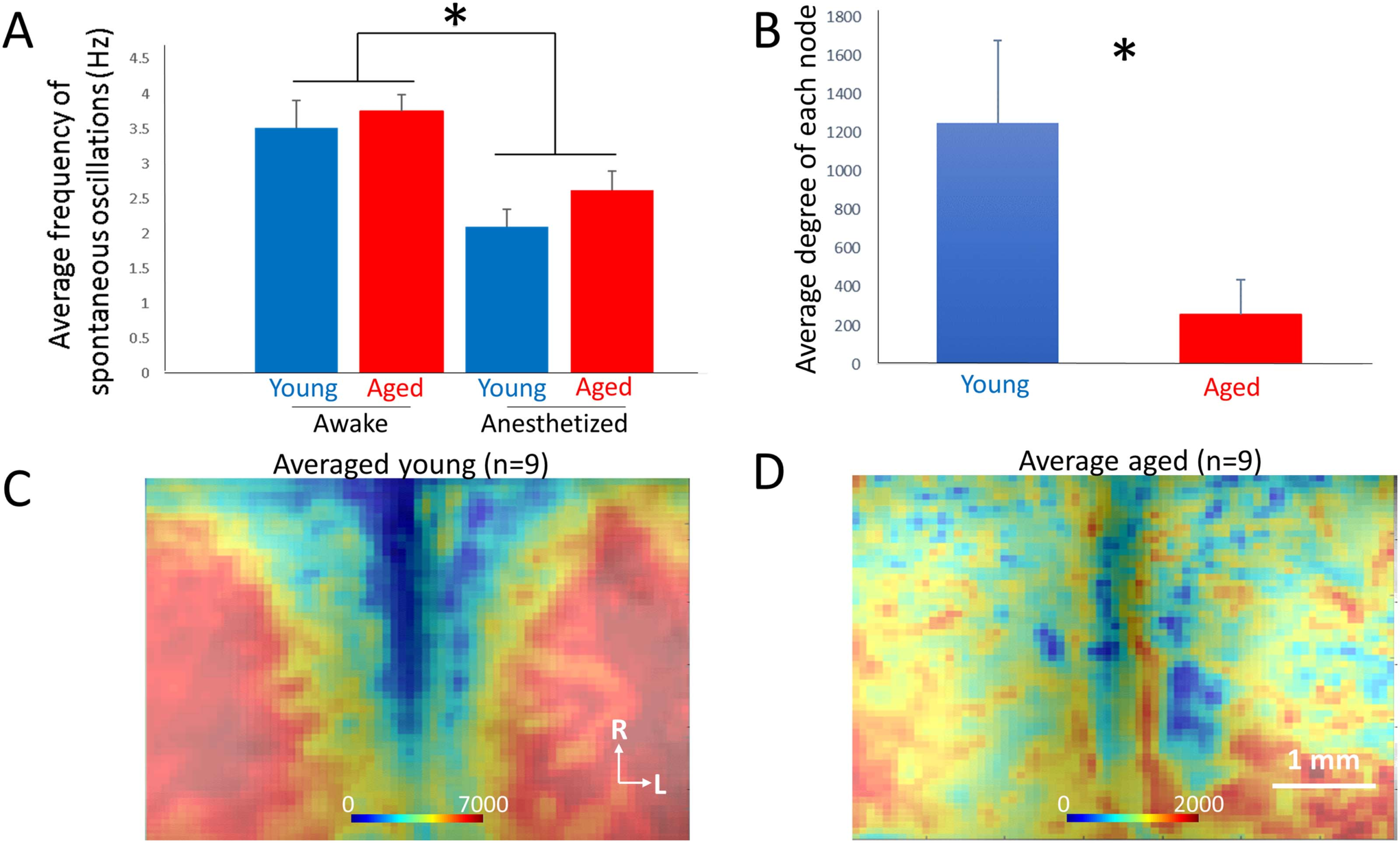
(A) The average frequency of spontaneous activity was roughly 3.5 Hz in awake mice and similar between aged and young mice.
Figure 5.
The average frequency declined significantly to approximately 2.5 Hz in the presence of ketamine/xylazine anesthesia. (B) Comparison of
average degree per node (measured as number of edges per node) in young versus aged mice (n = 9 per group). (C) Average map of degree of
each node of n = 9 young mice. (D) Average map of degree of each node of n = 9 aged mice. In both C and D, heat map corresponds to the
number of edges of each node; *p < 0.05.
flavoprotein autofluorescence has not been used previously to measure spontaneous neural
activity in vivo, we initially characterized the properties of the signals. It was observed that
the majority of the power is in the delta range (approximately 3 Hz) in both young and aged
animals (Figure 5A). To validate that the spectral properties are modulated as expected by level
of arousal, mice were anesthetized with ketamine/xylazine (after spontaneous signals were mea-
sured for CommDy analysis) and we observed the expected overall decrease in frequency in
both groups ( p = 0.006), as has been seen previously (Dworak, McCarley, Kim, & Basheer,
2011; Schüttler et al., 1987), with no differences between age groups ( p = 0.3, Figure 5A). To
determine the robustness of the spontaneous flavoprotein oscillatory signals in young compared
to aged mice, the SNR of the spontaneous activity was compared and found to be similar in the
two groups (average young SNR = 1.64 [SD 5.1] dB, average aged SNR = 1.99 [SD 4.34] dB, p =
0.79, Mann–Whitney). These data suggest that spontaneous flavoprotein autofluorescence sig-
nals track with more commonly measured indicators of spontaneous activity, such as EEG, and
show similar oscillatory properties at in young and aged mice, but given the slow nature of the
signal are limited to lower frequency spectral bands than EEG.
To compare the overall degree of connectedness between nodes in aged versus young an-
imals, the degree of each node was computed in young and aged animals. Similar to the
EEG:
Electroencephalography, which is
a technique to measure electrical
activity of the brain by placing
electrodes on the scalp.
Network Neuroscience
579
Fragmentation of aging mouse cortex revealed by network analysis
findings in brain slices from the auditory cortex, the degree of connectivity, as measured by
average number of edges per node, was significantly decreased in the aged compared to the
young animals (1,250 ± 1,214 [SD] edges/node vs. 257 ± 476 [SD] edges/node, p = 0.03,
Mann–Whitney; Figure 5B). When plotted as average 2-D maps, in young animals there are
large regions with high connectivity that are seen symmetrically on either side of the midline
(Figure 6C). In contrast, 2-D maps in older animals show essentially no organization, with
scattered islands of connectivity with rough symmetry across the midline (Figure 5D).
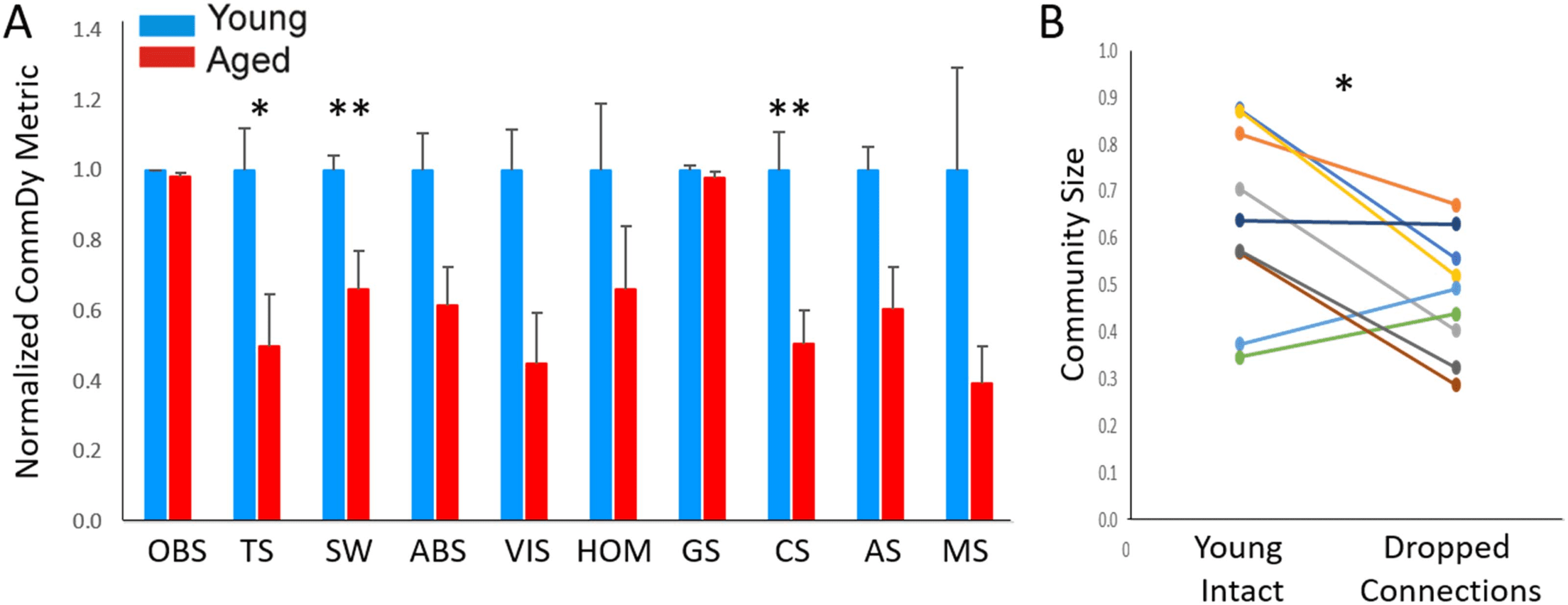
Characterization of network structure using CommDy revealed aged versus young differ-
ences for three different network metrics: time span (young = 0.686 ± 0.229 [SD], aged =
0.342 ± 0.289 [SD], p = 0.014, Mann–Whitney), switching (young = 0.920 ± 0.111 [SD], aged =
0.609 ± 0.283, p = 0.006, Mann–Whitney), and normalized community size (young = 0.642 ±
0.198 [SD], aged = 0.325 ± 0.173 [SD], p = 0.0062, Mann–Whitney; Figure 6A). Several other
network metrics also showed differences that trended toward significance: absence (young =
0.323 ± 0.098 [SD], aged = 0.199 ± 0.099 [SD], p = 0.027, Mann–Whitney), visiting (young =
0.256 ± 0.083 [SD], aged = 0.115 ± 0.105 [SD], p = 0.017, Mann–Whitney), and average stay
(young = 0.680 ± 0.128 [SD], aged = 0.411 ± 0.231 [SD], p = 0.017, Mann–Whitney); but these
did not survive multiple comparisons correction. To determine if the drop in community size with
aging is related to diminished intracortical connectivity, analogous to that seen in vitro (see
Figure 4B), connections between nodes were randomly dropped so that the average degree per
node in the young mice matched the aged mice and network metrics were recomputed. This
manipulation was intended to mimic the NMDA blockade done in vitro. Similar to the findings
in vitro, community size was found to decline with decreasing connectivity between nodes (young
intact community size = 0.642 ± 0.198 [SD], young after dropped connections community size =
0.480 ± 0.130 [SD], p = 0.03, Wilcoxon signed rank test; Figure 6B). These data suggest that in
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
2
5
6
9
1
9
2
6
5
8
9
n
e
n
_
a
_
0
0
1
9
1
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
(A) Bar graph of average CommDy metrics for spontaneous in vivo data young (n = 9) and aged (n = 9) animals, normalized to the
Figure 6.
average value for young animals. Group size, community size, average stay, and maximum stay are normalized for the total number of activated
pixels. Error bars = standard error. (B) Comparison of community size (a unitless value) in young mice either before (left) or after (right) randomly
dropping edges so that the number of edges equaled the number of edges seen in aged animals. Each line corresponds to a particular animal
before and after dropping connections. OBS = observed; TS = community time span; SW = switching cost; ABS = absence cost; VIS = visiting
cost; HOM = homing; GS = average group size; CS = average community size; AS = average community stay; MS = maximum community stay.
Fuller explanation of these metrics is found in Table 1; *p < 0.05, **p < 0.01.
Network Neuroscience
580
Fragmentation of aging mouse cortex revealed by network analysis
both datasets (slice and in vivo), the effect of aging is to decrease cortical network community size
by diminishing intracortical connectivity.
DISCUSSION
In this study, we applied the CommDy network analysis algorithm to investigate changes in
dynamic network structure that occur in cortical networks as they age. After optimizing
CommDy to detect communities in a brain slice preparation, CommDy was used to compute
a series of metrics that reflect the “social” organization of the cortical networks. The common
finding between brain slice paroxysmal activations and in vivo awake spontaneous activity
was a decline in both degree of each node and cortical network community size with aging.
Community size is a reflection of temporary associations between nodes in a network. The
decline in community size seen with aging was mimicked by weakening cortical connections
pharmacologically (with the NMDA blocker APV; Figure 4) or computationally (by randomly
dropping edges; Figure 6), suggesting that aging is associated with weakening of intracortical
connections. Given the differences in the two datasets used in this study (slice vs. in vivo,
auditory vs. motor cortex, paroxysmal activity vs. routine spontaneous activity), it is remarkable
that the data converged to show diminished community size with aging. Below we describe
the limitations and the potential utilities of the CommDy technique in brain imaging studies
and how they relate to changes occurring during aging.
Technical Considerations
The current work is based on flavoprotein autofluorescence imaging, which has the advantages
of providing a stable intrinsic signal with high sensitivity and good spatial resolution (100–200
microns; D. A. Llano et al., 2009), but has a relatively sluggish time course (~0.5–1 s for time to
peak; D. A. Llano et al., 2009; Reinert et al., 2007; Shibuki et al., 2003). Given this slow time
course, one might expect that small differences in the time to activate of different groups of neu-
rons would be washed out by delays induced by neurometabolic coupling. However, despite
these slow responses, CommDy did reveal distinct populations, suggesting that more subtle het-
erogeneities than could be appreciated using traditional analytical approaches were detectable
using CommDy in the time courses of the imaging responses. Another potential methodological
consideration about the current analysis is the use of pixels as individual nodes, rather than using
defined regions or individual neurons. At the binning and magnification used in this study, an
individual pixel occupies an area of approximately 800–1,600 μm2. To estimate the number of
neurons represented in this pixel, an upper estimate of the volume will be used, corresponding to
the full thickness of the slice or the cortex, though it is likely that the blue excitation light incom-
pletely penetrated the sample. The mouse cerebral cortex contains about 9.2 × 104
neurons/mm3 (Schüz & Palm, 1989), giving a rough estimate that each pixel in our preparation
contained signal emanating from on the order of ~50–100 neurons. Compared to techniques
that image spiking activity of individual cells, such as calcium imaging, flavoprotein autofluo-
rescence offers lower spatial resolution but offers a broad field of view (multiple millimeters),
permitting large-scale networks to be identified. Compared to the most commonly used tech-
nique for large-scale brain imaging, fMRI with BOLD signals, where the voxel sizes range from
millimeters in humans to hundreds of microns in rats, the current analysis provides substantially
higher spatial resolution. In addition, we have recently applied CommDy to calcium imaging
data from cortical slices and observed that CommDy is able to detect known features of cortical
connectivity (Naik et al., 2019). Therefore, it is likely that the current analytical approach may be
adapted to any of the imaging techniques mentioned above.
Network Neuroscience
581
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
2
5
6
9
1
9
2
6
5
8
9
n
e
n
_
a
_
0
0
1
9
1
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Fragmentation of aging mouse cortex revealed by network analysis
Implications for Aging
Pathological changes associated with aging have been described at virtually every level of the
central nervous system, including the cortex. Here, we observed aging-related disruptions in
network-level function of the auditory and motor cortices. In the slice preparation, these changes
are independent of the magnitude of the activation and are likely not related to differences in
slice viability, since we have previously found that the auditory cortices in slices from young
versus aged animals do not differ in their baseline redox state or response to metabolic stress
(Stebbings et al., 2016). We note that these data are not likely to be explained by any potential
viability differences in the preparations because simple loss of connectivity does not by itself
directly imply a lack of cohesion over time, as the small groups can still coalesce into functional
units. For example, even at low densities, the presence of well-connected clusters has been
shown to result in synchronization in static networks (Du & Li, 2014; Ponce-Alvarez et al.,
2015; Zhao, Bryce Beverlin, Netoff, & Nykamp, 2011). In addition, differences between young
and aged animals in the brain slice studies are also likely not related to differences in GABAergic
synaptic inhibition, since saturating doses of a GABAA antagonist were used in the slice study.
The current data suggest that aging causes changes in the underlying local excitatory sub-
structure in the cerebral cortex (Figures 4 and 6). The specific substrate for these changes is un-
known, but could relate to changes in local dendritic branching and integration, changes in
intrinsic excitability of pyramidal cells, or changes in synaptic properties, all known to be altered
in the aging brain (Cupp & Uemura, 1980; Disterhoft & Oh, 2007; Feldman & Dowd, 1975;
Leuba, 1983). Regarding the latter hypothesis, we observed that a classifier built only using
dose-dependent changes in CommDy network metrics induced by APV was able to classify data
from young versus aged animals with a high degree of accuracy (Figure 4). These findings are
remarkable in that the cross-validation was done using entirely different datasets obtained from
different types of slices (auditory cortex slices for aging data and coronal slice for APV data).
Furthermore, these findings are consistent with a body of literature demonstrating NMDA recep-
tor hypofunction in the cerebral cortex of the aging brain (Magnusson & Cotman, 1993; Piggott,
Perry, Perry, & Court, 1992; Wenk, Walker, Price, & Cork, 1991). Therefore, these data suggest
that the aging-related changes in network-level interactions observed in the cortex of this and
previous studies (Andrews-Hanna et al., 2007; Battaglia et al., 2020; Tomasi & Volkow, 2012)
may be, at least in part, caused by changes in NMDA receptor function.
Hearing loss and cortical thickness were both associated with declines in group and commu-
nity size in aging auditory cortical networks (Supporting Information Figures S2 and S3). Hearing
threshold is known to increase with age in mice, including the CBA/CaJ mice studied here (Zheng,
Johnson, & Erway, 1999). The relationship between hearing loss and auditory cortical network
structure in the current data is likely confounded by age as the relationships between hearing
loss and community and group size were found not to be significant in a multivariate regression
when age and cortical thickness were included as predictors. However, cortical thickness was
also found to be an independent predictor of normalized group size (Supporting Information
Table S1). This relationship is consistent with the mechanisms of aging-associated thinning of
the cortex. Cortical thinning is related to the loss of synapses between cortical neurons and/or
demyelination of local cortical axons, with retention of the number of neurons (Dumitriu et al.,
2010; Haug, Kühl, Mecke, Sass, & Wasner, 1983; Masliah et al., 1993; Terry, DeTeresa, &
Hansen, 1987). As such, cortical thinning would be predicted to be associated with drops in
cortical connectivity, consistent with the current data. This idea is supported by previous findings
from network theory showing that random or uniform loss of connectivity, translated into uni-
formly lower network density, results in smaller clusters (Erdos & Rényi, 1959, 1960). Hence,
our results showing diminished group and community size with aging, combined with the
CBA/CaJ:
A mouse strain commonly used in
auditory aging studies because it
retains relatively good hearing until
older ages.
Network Neuroscience
582
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
2
5
6
9
1
9
2
6
5
8
9
n
e
n
_
a
_
0
0
1
9
1
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Fragmentation of aging mouse cortex revealed by network analysis
known drop in synaptic connectivity seen in aging, are consistent with predictions from net-
work theory.
Potential Utilities of CommDy
The notion that neuronal cell assemblies are critical for perception and may shift dynamically over
time is a foundational principle in neuroscience (Abeles, Prut, Bergman, & Vaadia, 1994; Singer,
1998), but quantifying such network dynamics has been challenging. CommDy may prove to be a
useful tool to quantitatively study brain networks. Although the current study is focused on flavo-
protein autofluorescence imaging, CommDy can be adapted to other forms of brain imaging as
well. Such adaptation would permit CommDy to be used to refine network-level hypotheses about
brain function, for example, in the study of pathological states in humans. It is speculated that
many neurological and psychiatric disorders are caused by functional disruptions in large-scale
brain networks (Filippi et al., 2012; Rubinov et al., 2009; Seeley, Crawford, Zhou, Miller, &
Greicius, 2009). Most of the current work in this area has focused on using network measures
either as a tool to classify different disease states, or as a means to better understand network dis-
ruptions associated with disease states. In both cases, much of the work has been focused on the
generation of static maps. However, in brain diseases, fluctuations in clinical symptomatology are
the rule rather than the exception. For example, seizures in epilepsy, “off” states in Parkinson’s
disease, and hallucinations in schizophrenia all occur paroxysmally on the backdrop of stable
brain structures, such that it will only be possible to understand them using dynamic assessment
tools. CommDy may add to the growing toolbox of network assessment tools and will contribute
to the understanding of brain network dynamics.
METHODS
Animal Use
CBA/CaJ mice from 4.4 to 23.5 months of age of both sexes were used. All procedures were
approved by the Institutional Animal Care and Use Committee at the University of Illinois. All
animals were housed in animal care facilities approved by the American Association for
Assessment and Accreditation of Laboratory Animal Care.
Auditory Brainstem Responses
As previously described (Stebbings et al., 2016), to measure hearing for the aging studies, ABRs
were obtained in response to tones at frequencies of 8, 16, and 32 kHz, and average thresholds
were reported. Animals were anesthetized with 100 mg/kg ketamine + 3 mg/kg xylazine intra-
peritoneally before the insertion of two subdermal electrodes, one at the vertex and one behind
the left ear. Stimuli were presented using a Tucker-Davis (TDT) system 3, ES1 free field speaker,
with waveforms being generated by RPvdsEx software. The output of the TDT speaker was
calibrated at all the relevant frequencies, using a Bruel and Kjaer type 4135 microphone and
a Bruel and Kjaer measuring amplifier (Model 2610). Each frequency was presented for 5 ms
(3 ms flat with 1 ms for both rise and fall times), at a rate of 2–6 Hz with a 100 ms analysis
window. Raw potentials were obtained with a Dagan 2400A amplifier and preamplifier head-
stage combination, and filtered between 100 Hz and 3000 Hz. An ADInstruments PowerLab
4/30 system was used to average these waveforms 500 times. Significant deflections, assessed
via visual inspection, within 10 ms after the end of the stimulus were deemed to be a response.
Blinding of ABR assessments was not done given the conspicuous physical differences in young
versus aged mice (i.e., aged animals are heavier and had less hair on snout).
583
Artificial cerebrospinal fluid:
A laboratory solution used to mimic
the fluid that normally surrounds the
brain.
SR95531:
A compound used to block GABA
receptors in the brain. Blocking these
receptors leads to hyperexcitability in
the tissue.
Network Neuroscience
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
2
5
6
9
1
9
2
6
5
8
9
n
e
n
_
a
_
0
0
1
9
1
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Fragmentation of aging mouse cortex revealed by network analysis
Brain Slice Preparation
The CBA/CaJ strain was used because of its gradual aging-related hearing loss (Frisina & Zhu,
2010; Ohlemiller, Dahl, & Gagnon, 2010; Zheng et al., 1999). Five young mice (5.5 months of
age) and five aged mice (22 months of age) were used. Mice were initially anesthetized with
ketamine (100 mg/kg) and xylazine (3 mg/kg) and then transcardially perfused with an ice-
cold sucrose saline solution (in mM: 206 sucrose, 10.0 MgCl2, 11.0 glucose, 1.25
NaH2PO4, 26 NaHCO3, 0.5 CaCl2, 2.5 KCl, pH 7.4). Slices containing the auditory cortex
were cut using a modification of the method developed by Cruikshank et al. (Cruikshank,
Rose, & Metherate, 2002), modified for the aging brain, as described previously (Stebbings
et al., 2016; Takesian, Kotak, & Sanes, 2012), see Figure 1B for brain image. Brains were
blocked by removing the olfactory bulbs and the anterior 2 mm of frontal cortex with a razor
blade. The brain was then tipped onto the coronal cut and an off-horizontal cut was made on
the dorsal surface, removing a sliver of brain angled at 20° from the horizontal plane. The
brain was then glued onto the cut angled surface, and sections were then taken. All slices were
then transferred to a holding chamber containing oxygenated incubation artificial cerebrospi-
nal fluid (ACSF) (in mM: 126 NaCl, 3.0 MgCl2, 10.0 glucose, 1.25 NaH2PO4, 26 NaHCO3, 1.0
CaCl2, 2.5 KCl, pH 7.4) and incubated at 32°C for 1 hr prior to experimentation. The ACSF
used for the experiments use equimolar MgCl2 and CaCl2 and contained 1 μM of the GABAA
antagonist SR95531 to enhance spontaneous activity. Without the addition of SR95531, we
found that the amount of activity present in the slice was insufficient for either CommDy or
traditional analyses. During imaging, slices were placed on a stainless steel mesh for two-sided
perfusion, as we have described previously (Daniel A. Llano, Slater, Lesicko, & Stebbings, 2014).
To assess the impact of NMDA blockade on network activity, the NMDA blocker D-APV (Tocris,
catalog no. 0106) was dissolved in ACSF and bath applied at a series of concentrations ranging
from 15 to 120 μM, with 20-min wash-ins between each dose escalation. Cortical thickness was
evaluated by drawing a line tangent to the rostral-most extent of the hippocampus, from the
white/gray matter border of the cortex to the pia, as we have described previously (Stebbings
et al., 2016).
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
2
5
6
9
1
9
2
6
5
8
9
n
e
n
_
a
_
0
0
1
9
1
p
d
.
t
In Vivo Preparation
An initial stereotactic surgery under ketamine/xylazine (100 mg/kg and 3 mg/kg IP, respectively)
was done to glue a threaded headbolt onto the skull using OptiBond XTR kit (catalog no. 35106)
cement. After at least 3 days to recover, mice were gradually acclimated to an imaging chamber
within a soundproof booth. Their headbolts were affixed to a holder, and the body was suspended
while under isoflurane (4%) anesthesia. The mice were then allowed to emerge from anesthesia,
initially for 5 min, and gradually up to 15–20 min. Awake data were obtained once isoflurane was
removed for at least 10 min. Once awake data were obtained, the animal was reanesthetized with
ketamine/xylazine (100 mg/kg and 3 mg/kg IP, respectively) for additional imaging.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Imaging
For the slice work, flavoprotein autofluorescence imaging was done with a fluorescence illu-
minator (Prior Lumen 200) and a UMNIB Olympus filter cube (470–490 nm excitation, 505 nm
dichroic, 515 nm emission long pass). A coverslip was placed over the slice to provide a stable
imaging plane, and data were collected using an infinity-corrected 4X macro objective (NA
0.28) and a Retiga EXi camera and StreamPix software (see Figure 1A for diagram of experi-
mental setup). Data were obtained with 8 × 8 hardware binning, producing images of 130 ×
174 pixels. Acquisition rates were 71 frames per second and 1,000 frames were used for
analysis.
584
Hippocampus:
A brain region important for learning
and memory. In this case, the
hippocampus serves as a landmark
for the location of the auditory
cortex.
Dichroic:
A mirror that reflects certain
wavelengths of light, but not all, used
to direct light in a fluorescence
imaging system.
Network Neuroscience
Fragmentation of aging mouse cortex revealed by network analysis
Macroscope:
An imaging system using tandem
photography lenses used to
maximize brightness of signal using
wide-field imaging at relatively low
magnifications.
In vivo imaging was done using a macroscope system outfitted with 85 mm f/1.4 and f/1.2
Nikon lenses and an Adimec 1,000 m CCD camera (7.4 × 7.4 μm pixel size, 1,004 × 1,004 pixels).
Blue light (450 nm, 30 nm band-pass) was used for excitation and green light (515 nm, long pass)
was collected, and a 495-nm dichroic mirror was used. Images were collected at 25 frames per
second. One thousand frames were used for the analysis. Pixels were binned 4 × 4, and a region of
interest containing 60 × 70 pixels over both motor cortices and symmetric with respect to the
midline was selected for analysis. No filtering was done on the signals. SNR was computed using
the MATLAB function snr on the time series obtained by taking the time course of the average
of all pixels in the image.
Paroxysmal Depolarizations
In the presence of SR95531, spontaneous activations were observed with flavoprotein auto-
fluorescence imaging, consisting of a relatively sharp rise in fluorescence, peaking at about
150 ms after baseline, with a slow decline, gradually returning to baseline seconds. We rou-
tinely observe these spontaneous activations in all brain slices that contain the cerebral cortex
and are bathed in concentrations of SR95531 above 0.5 μM, and generally such activations
occur roughly every 30–60 s with no obvious periodicity. These electrical events strongly re-
semble the intra- and extracellular manifestations of paroxysmal depolarizing shifts, respec-
tively (Johnston & Brown, 1981; Steriade & Amzica, 1999; Vanzetta, Flynn, Ivanov,
Bernard, & Bénar, 2010; Yaron-Jakoubovitch, Koch, Segev, & Yarom, 2013), and we have
previously found that they correlate with paroxysmal depolarizing shifts (Ibrahim et al.,
2017). Therefore, we refer to them as paroxysmal depolarizations. All slice analyses in this report
are done on activity occurring during the paroxysmal depolarizations.
Statistical Analysis
To compare activations in slices from young versus aged mice, a linear mixed model analysis
was run, incorporating random effects because multiple paroxysmal depolarizations occurred
in some slices. The 10 CommDy network metrics were used As regression variables and young
versus aged was used as treatment groups. Other analysis was done without the assumption of
normality, thus Mann–Whitney testing was used to compare groups, Spearman’s correlations
were used for bivariate data, and Wilcoxon testing was used for paired data. The Holm–
Bonferroni method (Holm, 1979) was used to adjust for multiple comparisons.
ACKNOWLEDGMENTS
The authors thank David Trinco, Syed Haider and the University of Illinois Statistical
Consultation Service for their assistance with data analysis.
SUPPORTING INFORMATION
Supporting information for this article is available at https://doi.org/10.1162/netn_a_00191.
AUTHOR CONTRIBUTIONS
Daniel Adolfo Llano: Conceptualization; Data curation; Formal analysis; Funding acquisition;
Investigation; Methodology; Project administration; Resources; Supervision; Validation; Writing –
original draft; Writing – review & editing. Chihua Ma: Conceptualization; Formal analysis;
Network Neuroscience
585
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
2
5
6
9
1
9
2
6
5
8
9
n
e
n
_
a
_
0
0
1
9
1
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Fragmentation of aging mouse cortex revealed by network analysis
Investigation; Methodology; Writing – original draft; Writing – review & editing. Umberto Di
Fabrizio: Conceptualization; Formal analysis; Investigation; Methodology; Writing – original draft;
Writing – review & editing. Aynaz Taheri: Conceptualization; Formal analysis; Investigation;
Methodology. Kevin A. Stebbings: Investigation; Methodology. Georgiy Yudintsev: Data curation;
Investigation; Methodology. Gang Xiao: Formal analysis. Robert V. Kenyon: Conceptualization;
Funding acquisition; Investigation; Methodology; Project administration; Visualization; Writing –
original draft; Writing – review & editing. Tanya Y. Berger-Wolf: Conceptualization; Data curation;
Formal analysis; Funding acquisition; Investigation; Methodology; Project administration;
Resources; Software; Supervision; Visualization; Writing – original draft; Writing – review &
editing.
FUNDING INFORMATION
Daniel Adolfo Llano, National Science Foundation (https://dx.doi.org/10.13039/100000001),
Award ID: 1515587. Daniel Adolfo Llano, National Institutes of Health (https://dx.doi.org/10
.13039/100000002), Award ID: DC012125. Daniel Adolfo Llano, National Institutes of Health
(https://dx.doi.org/10.13039/100000002), Award ID: AG059103. Daniel Adolfo Llano,
American Federation for Aging Research (https://dx.doi.org/10.13039/100000965), Award
ID: 2011-03147. Daniel Adolfo Llano, Alzheimer’s Association (https://dx.doi.org/10.13039
/100000957), Award ID: NIRG-12-242848. Daniel Adolfo Llano, Kiwanis Neuroscience
Research Foundation.
REFERENCES
Abeles, M., Prut, Y., Bergman, H., & Vaadia, E. (1994).
Synchronization in neuronal transmission and its importance for
information processing. In J. Van Pelt, M. A. Corner, H. B. M.
Uylings, & F. H. Lopez da Silva (Eds.), Progress in brain research
(Vol. 102, pp. 395–404). Elsevier. https://doi.org/10.1007/978-3
-642-85148-3_3
Andrews-Hanna, J. R., Snyder, A. Z., Vincent, J. L., Lustig, C., Head,
D., Raichle, M. E., & Buckner, R. L. (2007). Disruption of large-
scale brain systems in advanced aging. Neuron, 56(5), 924–935.
https://doi.org/10.1016/j.neuron.2007.10.038, PubMed:
18054866
Ash, J. A., Lu, H., Taxier, L. R., Long, J. M., Yang, Y., Stein, E. A., &
Rapp, P. R. (2016). Functional connectivity with the retrosplenial
cortex predicts cognitive aging in rats. Proceedings of the
National Academy of Sciences of the United States of America,
1 1 3( 4 3 ) , 1 2 28 6 –1 22 9 1. ht t p s : / / d o i . or g / 1 0 . 10 7 3/ p n a s
.1525309113, PubMed: 27791017
Backstrom, L., Huttenlocher, D., Kleinberg, J., & Lan, X. (2006).
Group formation in large social networks: Membership, growth,
and evolution. Paper presented at the Proceedings of the 12th
ACM SIGKDD international conference on Knowledge discovery
and data mining, Philadelphia, PA, USA. https://doi.org/10.1145
/1150402.1150412
Barale, C., Kulahci, I., Sulo, R., Berger-Wolf, T., & Rubenstein, D.
(2010). A social networks approach to sheep movement and
leadership. Paper presented at the In 7th International
Conference on Applications of Social Network Analysis (ASNA
2010), ETH Zurich, Switzerland.
Battaglia, D., Boudou, T., Hansen, E. C. A., Lombardo, D.,
Chettouf, S., Daffertshofer, A., … Jirsa, V. (2020). Dynamic
Functional Connectivity between order and randomness and its
evolution across the human adult lifespan. Neuroimage, 222,
117156. https://doi.org/10.1016/j.neuroimage.2020.117156,
PubMed: 32698027
Beard, J. R., Officer, A. M., & Cassels, A. K. (2016). The world report on
ageing and health. The Gerontologist, 56, Suppl_2, S163–S166.
https://doi.org/10.1093/geront/gnw037, PubMed: 26994257
Beckmann, C. F., DeLuca, M., Devlin, J. T., & Smith, S. M. (2005).
Investigations into resting-state connectivity using independent
component analysis. Philosophical Transactions of the Royal
Society B: Biological Sciences, 360(1457), 1001–1013. https://
doi.org/10.1098/rstb.2005.1634, PubMed: 16087444
Berger-Wolf, T., Tantipathananandh, C., & Kempe, D. (2010).
Community identification in dynamic social networks. In C.
Philip, S. Yu, & J. Han (Eds.), Link Mining: Models, algorithms,
and applications (pp. 307–336). Springer. https://doi.org/10
.1007/978-1-4419-6515-8_12
Blondel, V. D., Guillaume, J.-L., Lambiotte, R., & Lefebvre, E. (2008).
Fast unfolding of communities in large networks. Journal of
Statistical Mechanics: Theory and Experiment, 2008(10), P10008.
https://doi.org/10.1088/1742-5468/2008/10/P10008
Breiman, L. (2001). Random forests. Machine Learning, 45(1), 5–32.
https://doi.org/10.1023/A:1010933404324
Brookes, M. J., Tewarie, P. K., Hunt, B. A. E., Robson, S. E.,
Gascoyne, L. E., Liddle, E. B., … Morris, P. G. (2016). A
multi-layer network approach to MEG connectivity analysis.
Network Neuroscience
586
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
2
5
6
9
1
9
2
6
5
8
9
n
e
n
_
a
_
0
0
1
9
1
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Fragmentation of aging mouse cortex revealed by network analysis
Neuroimage, 132, 425–438. https://doi.org/10.1016/j.neuroimage
.2016.02.045, PubMed: 26908313
Bullmore, E., & Sporns, O. (2009). Complex brain networks: Graph
theoretical analysis of structural and functional systems. Nature
Reviews Neuroscience, 10(3), 186–198. https://doi.org/10.1038
/nrn2575, PubMed: 19190637
Calhoun, V., Adali, T., McGinty, V., Pekar, J., Watson, T., &
Pearlson, G. (2001). fMRI activation in a visual-perception task:
Network of areas detected using the general linear model and
independent components analysis. Neuroimage, 14(5), 1080–1088.
https://doi.org/10.1006/nimg.2001.0921, PubMed: 11697939
Calhoun, V. D., Miller, R., Pearlson, G., & Adalı, T. (2014). The
chronnectome: Time-varying connectivity networks as the next
frontier in fMRI data discovery. Neuron, 84(2), 262–274. https://
doi.org/10.1016/j.neuron.2014.10.015, PubMed: 25374354
Caspary, D. M., Ling, L., Turner, J. G., & Hughes, L. F. (2008). Inhibitory
neurotransmission, plasticity and aging in the mammalian central
auditory system. Journal of Experimental Biology, 211(11), 1781–1791.
https://doi.org/10.1242/jeb.013581, PubMed: 18490394
Chen, T., Cai, W., Ryali, S., Supekar, K., & Menon, V. (2016).
Distinct global brain dynamics and spatiotemporal organization
of the salience network. PLoS Biology, 14(6), e1002469. https://
doi.org/10.1371/journal.pbio.1002469, PubMed: 27270215
Cruikshank, S. J., Rose, H. J., & Metherate, R. (2002). Auditory thalamo-
cortical synaptic transmission in vitro. Journal of Neurophysiology,
87(1), 361–384. https://doi.org/10.1152/jn.00549.2001, PubMed:
11784756
Cupp, C., & Uemura, E. (1980). Age-related changes in prefrontal
cortex of Macaca mulatta: Quantitative analysis of dendritic
branching patterns. Experimental Neurology, 69(1), 143–163.
https://doi.org/10.1016/0014-4886(80)90150-8, PubMed:
6771151
Damaraju, E., Allen, E., Belger, A., Ford, J., McEwen, S., Mathalon,
D., … Preda, A. (2014). Dynamic functional connectivity analysis
reveals transient states of dysconnectivity in schizophrenia.
NeuroImage: Clinical, 5, 298–308. https://doi.org/10.1016/j.nicl
.2014.07.003, PubMed: 25161896
Damoiseaux, J. S., Prater, K. E., Miller, B. L., & Greicius, M. D.
(2012). Functional connectivity tracks clinical deterioration in
Alzheimer’s disease. Neurobiology of Aging, 33(4), 828.e819–e830.
https://doi.org/10.1016/j.neurobiolaging.2011.06.024, PubMed:
21840627
Disterhoft, J. F., & Oh, M. M. (2007). Alterations in intrinsic neuronal
excitability during normal aging. Aging Cell, 6(3), 327–336.
https://doi.org/10.1111/j.1474-9726.2007.00297.x, PubMed:
17517042
Du, P., & Li, M. Y. (2014). Impact of network connectivity on the
synchronization and global dynamics of coupled systems of differ-
ential equations. Physica D: Nonlinear Phenomena, 286, 32–42.
https://doi.org/10.1016/j.physd.2014.07.008
Dumitriu, D., Hao, J., Hara, Y., Kaufmann, J., Janssen, W. G., Lou,
W., … Morrison, J. H. (2010). Selective changes in thin spine
density and morphology in monkey prefrontal cortex correlate
with aging-related cognitive impairment. The Journal of
Neuroscience, 30(22), 7507–7515. https://doi.org/10.1523
/JNEUROSCI.6410-09.2010, PubMed: 20519525
Dworak, M., McCarley, R. W., Kim, T., & Basheer, R. (2011). Delta
oscillations induced by ketamine increase energy levels in sleep-
wake related brain regions. Neuroscience, 197, 72–79. https://
doi.org/10.1016/j.neuroscience.2011.09.027, PubMed:
21958867
Engle, J. R., & Recanzone, G. H. (2012). Characterizing spatial
tuning functions of neurons in the auditory cortex of young
and aged monkeys: A new perspective on old data. Frontiers in
Aging Neuroscience, 4. https://doi.org/10.3389/fnagi.2012
.00036, PubMed: 23316160
Erdos, P., & Rényi, A. (1959). On random graphs I. Publicationes
Mathematicae Debrecen, 6, 290–297.
Erdos, P., & Rényi, A. (1960). On the evolution of random graphs.
Publications of the Mathematical Institute of the Hungarian
Academy of Sciences, 5, 17–61.
Feldman, M. L., & Dowd, C. (1975). Loss of dendritic spines in aging
cerebral cortex. Anatomy and Embryology, 148(3), 279–301.
https://doi.org/10.1007/BF00319848, PubMed: 1221885
Filippi, M., Agosta, F., Scola, E., Canu, E., Magnani, G., Marcone,
A., … Comi, G. (2012). Functional network connectivity in the
behavioral variant of frontotemporal dementia. Cortex, 49(9),
2389–2401. https://doi.org/10.1016/j.cortex.2012.09.017,
PubMed: 23164495
Fleidervish, I. A., Binshtok, A. M., & Gutnick, M. J. (1998).
Functionally distinct NMDA receptors mediate horizontal con-
nectivity within layer 4 of mouse barrel cortex. Neuron, 21(5),
1055–1065. https://doi.org/10.1016/S0896-6273(00)80623-6,
PubMed: 9856461
Frisina, R. D., & Zhu, X. (2010). Auditory sensitivity and the outer
hair cell system in the CBA mouse model of age-related hearing
loss. Open Access Animal Physiology, 2, 9. https://doi.org/10
.2147/OAAP.S7202, PubMed: 21866215
Gil, Z., Connors, B. W., & Amitai, Y. (1999). Efficacy of thalamo-
cortical and intracortical synaptic connections: Quanta, innerva-
tion, and reliability. Neuron, 23(2), 385–397. https://doi.org/10
.1016/S0896-6273(00)80788-6, PubMed: 10399943
Glisky, E. L. (2007). Changes in cognitive function in human aging.
In David R. Riddle (Ed.), Brain aging: Models, methods, and
mechanisms (pp. 3–20). CRC Press. https://doi.org/10.1201
/9781420005523-1
Ha, J., Cho, Y. S., Kim, S. J., Cho, S. H., Kim, J. P., Jung, Y. H., …
Na, D. L. (2020). Hearing loss is associated with cortical thinning in
cognitively normal older adults. European Journal of Neurology,
27(6), 1003–1009. https://doi.org/10.1111/ene.14195, PubMed:
32125747
Haug, H., Kühl, S., Mecke, E., Sass, N., & Wasner, K. (1983). The
significance of morphometric procedures in the investigation of
age changes in cytoarchitectonic structures of human brain.
Journal fur Hirnforschung, 25(4), 353–374.
Hirai, T., Korogi, Y., Sakamoto, Y., Hamatake, S., Ikushima, I., &
Takahashi, M. (1996). T2 shortening in the motor cortex: Effect of
aging and cerebrovascular diseases. Radiology, 199(3), 799–803.
https://doi.org/10.1148/radiology.199.3.8638008, PubMed:
8638008
Holm, S. (1979). A simple sequentially rejective multiple test pro-
cedure. Scandinavian Journal of Statistics, 6, 65–70.
Holme, P., & Saramäki, J. (2012). Temporal networks. Physics Reports,
519(3), 97–125. https://doi.org/10.1016/j.physrep.2012.03.001
Hopfield, J. J. (2008). Neurons, dynamics and computation. Physics
Today, 47(2), 40–46. https://doi.org/10.1063/1.881412
Network Neuroscience
587
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
2
5
6
9
1
9
2
6
5
8
9
n
e
n
_
a
_
0
0
1
9
1
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Fragmentation of aging mouse cortex revealed by network analysis
Horn, J. L. (1982). The theory of fluid and crystallized intelligence
in relation to concepts of cognitive psychology and aging in
adulthood. In Aging and Cognitive Processes (pp. 237–278).
Springer. https://doi.org/10.1007/978-1-4684-4178-9_14
Horn, J. L., & Cattell, R. B. (1967). Age differences in fluid and crys-
tallized intelligence. Acta Psychologica, 26, 107–129. https://doi
.org/10.1016/0001-6918(67)90011-X, PubMed: 6037305
Hutchison, R. M., Womelsdorf, T., Allen, E. A., Bandettini, P. A.,
Calhoun, V. D., Corbetta, M., … Gonzalez-Castillo, J. (2013).
Dynamic functional connectivity: Promise, issues, and interpre-
tations. Neuroimage, 80, 360–378. https://doi.org/10.1016/j
.neuroimage.2013.05.079, PubMed: 23707587
Huttenlocher, P. R. (1979). Synaptic density in human frontal
cortex-developmental changes and effects of aging. Brain
Research, 163(2), 195–205. https://doi.org/10.1016/0006-8993
(79)90349-4, PubMed: 427544
Ibrahim, B. A., Wang, H., Lesicko, A. M., Bucci, B., Paul, K., &
Llano, D. A. (2017). Effect of temperature on FAD and NADH-
derived signals and neurometabolic coupling in the mouse audi-
tory and motor cortex. Pflügers Archiv-European Journal of
Physiology, 469(12), 1631–1649. https://doi.org/10.1007
/s00424-017-2037-4, PubMed: 28785802
Johnston, D., & Brown, T. H. (1981). Giant synaptic potential hypoth-
esis for epileptiform activity. Science, 211(4479), 294–297. https://
doi.org/10.1126/science.7444469, PubMed: 7444469
Karwowski, W., Vasheghani Farahani, F., & Lighthall, N. (2019).
Application of graph theory for identifying connectivity patterns
in human brain networks: A systematic review. Frontiers in
Neuroscience, 13, 585. https://doi.org/10.3389/fnins.2019.00585,
PubMed: 31249501
Leuba, G. (1983). Aging of dendrites in the cerebral cortex of the
mouse. Neuropathology and Applied Neurobiology, 9(6), 467–475.
https://doi.org/10.1111/j.1365-2990.1983.tb00130.x, PubMed:
6656999
Li, J., Li, W., Xian, J., Li, Y., Liu, Z., Liu, S., … He, H. (2012).
Cortical thickness analysis and optimized voxel-based mor-
phometry in children and adolescents with prelingually profound
sensorineural hearing loss. Brain Research, 1430, 35–42. https://
doi.org/10.1016/j.brainres.2011.09.057, PubMed: 22079323
Liu, X., Erikson, C., & Brun, A. (1996). Cortical synaptic changes
and gliosis in normal aging, Alzheimer’s disease and frontal lobe
degeneration. Dementia and Geriatric Cognitive Disorders, 7(3),
128–134. https://doi.org/10.1159/000106867, PubMed:
8740626
Llano, D. A., Slater, B. J., Lesicko, A. M., & Stebbings, K. A. (2014).
An auditory colliculothalamocortical brain slice preparation in
mouse. Journal of Neurophysiology, 111(1), 197–207. https://
doi.org/10.1152/jn.00605.2013, PubMed: 24108796
Llano, D. A., Theyel, B. B., Mallik, A. K., Sherman, S. M., & Issa, N. P.
(2009). Rapid and sensitive mapping of long-range connections
in vitro using flavoprotein autofluorescence imaging combined
with laser photostimulation. Journal of Neurophysiology, 101(6),
3325–3340. https://doi.org/10.1152/jn.91291.2008, PubMed:
19321634
Magnusson, K. R., & Cotman, C. W. (1993). Effects of aging on NMDA
and MK801 binding sites in mice. Brain Research, 604(1), 334–337.
https://doi.org/10.1016/0006-8993(93)90386-2, PubMed:
8457861
Masliah, E., Mallory, M., Hansen, L., DeTeresa, R., & Terry, R.
(1993). Quantitative synaptic alterations in the human neocortex
during normal aging. Neurology, 43(1), 192–192. https://doi.org
/10.1212/ WNL.43.1_Part_1.192, PubMed: 8423884
Mendelson, J., & Ricketts, C. (2001). Age-related temporal processing
speed deterioration in auditory cortex. Hearing Research, 158(1),
84–94. https://doi.org/10.1016/S0378-5955(01)00294-5,
PubMed: 11506940
Milo, R., Shen-Orr, S., Itzkovitz, S., Kashtan, N., Chklovskii, D., &
Alon, U. (2002). Network motifs: Simple building blocks of
complex networks. Science, 298(5594), 824–827. https://doi
.org/10.1126/science.298.5594.824, PubMed: 12399590
Naik, A., Kenyon, R., Taheri, A., Berger-Wolf, T., Ibrahim, B., &
Llano, D. (2019). V-NeuroStack: 3-D time stacks for identifying
patterns in neuronal activity time-series data. Society for
Neuroscience Abstracts, 432.09/DD31. https://doi.org/10.1101
/2020.12.03.410761
Newman, M. E. (2006). Modularity and community structure in net-
works. Proceedings of the National Academy of Sciences of the
United States of America, 103(23), 8577–8582. https://doi.org/10
.1073/pnas.0601602103, PubMed: 16723398
Ohlemiller, K. K., Dahl, A. R., & Gagnon, P. M. (2010). Divergent
aging characteristics in CBA/ J and CBA/CaJ mouse cochleae.
Journal of the Association for Research in Otolaryngology, 11(4),
605–623. https://doi.org/10.1007/s10162-010-0228-1, PubMed:
20706857
Oliviero, A., Profice, P., Tonali, P., Pilato, F., Saturno, E., Dileone,
M., … Di Lazzaro, V. (2006). Effects of aging on motor cortex
excitability. Neuroscience Research, 55(1), 74–77. https://doi
.org/10.1016/j.neures.2006.02.002, PubMed: 16584795
Onoda, K., Ishihara, M., & Yamaguchi, S. (2012). Decreased func-
tional connectivity by aging is associated with cognitive decline.
Journal of Cognitive Neuroscience, 24(11), 2186–2198. https://
doi.org/10.1162/jocn_a_00269, PubMed: 22784277
Pedreschi, N., Bernard, C., Clawson, W., Quilichini, P., Barrat, A., &
Battaglia, D. (2020). Dynamic core-periphery structure of informa-
tion sharing networks in entorhinal cortex and hippocampus.
Network Neuroscience, 1–30. https://doi.org/10.1162/netn_a
_00142, PubMed: 33615098
Piggott, M. A., Perry, E. K., Perry, R. H., & Court, J. A. (1992). [3 H]
MK-801 binding to the NMDA receptor complex, and its modu-
lation in human frontal cortex during development and aging.
Brain Research, 588(2), 277–286. https://doi.org/10.1016/0006
-8993(92)91586-4, PubMed: 1393579
Ponce-Alvarez, A., Deco, G., Hagmann, P., Romani, G. L.,
Mantini, D., & Corbetta, M. (2015). Resting-state temporal syn-
chronization networks emerge from connectivity topology and
heterogeneity. PLoS Computational Biology, 11(2), e1004100.
https://doi.org/10.1371/journal.pcbi.1004100, PubMed: 25692996
Reinert, K. C., Gao, W., Chen, G., & Ebner, T. J. (2007).
Flavoprotein autofluorescence imaging in the cerebellar cortex
in vivo. Journal of Neuroscience Research, 85(15), 3221–3232.
https://doi.org/10.1002/jnr.21348, PubMed: 17520745
Robinson, L. F., Atlas, L. Y., & Wager, T. D. (2015). Dynamic func-
tional connectivity using state-based dynamic community struc-
ture: Method and application to opioid analgesia. Neuroimage,
108, 274–291. https://doi.org/10.1016/j.neuroimage.2014.12
.034, PubMed: 25534114
Network Neuroscience
588
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
2
5
6
9
1
9
2
6
5
8
9
n
e
n
_
a
_
0
0
1
9
1
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Fragmentation of aging mouse cortex revealed by network analysis
Rossini, P., Di Iorio, R., Bentivoglio, M., Bertini, G., Ferreri, F.,
Gerloff, C., … Pestilli, F. (2019). Methods for analysis of brain con-
nectivity: An IFCN-sponsored review. Clinical Neurophysiology.
https://doi.org/10.1016/j.clinph.2019.06.006, PubMed:
31401492
Rozycka, A., & Liguz-Lecznar, M. (2017). The space where aging acts:
Focus on the GABA ergic synapse. Aging Cell, 16(4), 634–643.
https://doi.org/10.1111/acel.12605, PubMed: 28497576
Rubenstein, D., Sundaresan, S., Fischhoff, I., Tantipathananand, C.,
& Berger-Wolf, T. (2015). Similar but different: Dynamic social
network analysis highlights fundamental differences between
the fission-fusion societies of two equid species, the onager
and Grevy’s zebra. PloS One, 10(10), e0138645. https://doi.org
/10.1371/journal.pone.0138645, PubMed: 26488598
Rubinov, M., Knock, S. A., Stam, C. J., Micheloyannis, S., Harris, A. W.,
Williams, L. M., & Breakspear, M. (2009). Small-world properties of
nonlinear brain activity in schizophrenia. Human Brain Mapping,
30(2), 403–416. https://doi.org/10.1002/ hbm.20517, PubMed:
18072237
Salat, D. H., Buckner, R. L., Snyder, A. Z., Greve, D. N., Desikan,
R. S., Busa, E., … Fischl, B. (2004). Thinning of the cerebral cor-
tex in aging. Cerebral Cortex, 14(7), 721–730. https://doi.org/10
.1093/cercor/bhh032, PubMed: 15054051
Salthouse, T. A. (2003). Memory aging from 18 to 80. Alzheimer
Disease & Associated Disorders, 17(3), 162–167. https://doi.org
/10.1097/00002093-200307000-00008, PubMed: 14512830
Schmidt, S. A., Carpenter-Thompson, J., & Husain, F. T. (2017).
Connectivity of precuneus to the default mode and dorsal atten-
tion networks: A possible invariant marker of long-term tinnitus.
Neuroimage: Clinical, 16, 196–204. https://doi.org/10.1016/j
.nicl.2017.07.015, PubMed: 28794980
Schüttler, J., Stanski, D. R., White, P. F., Trevor, A. J., Horai, Y.,
Verotta, D., & Sheiner, L. B. (1987). Pharmacodynamic modeling
of the EEG effects of ketamine and its enantiomers in man.
Journal of Pharmacokinetics and Biopharmaceutics, 15, 241–253.
https://doi.org/10.1007/BF01066320, PubMed: 3668802
Schüz, A., & Palm, G. (1989). Density of neurons and synapses in the
cerebral cortex of the mouse. Journal of Comparative Neurology,
286(4), 442–455. https://doi.org/10.1002/cne.902860404,
PubMed: 2778101
Seeley, W. W., Crawford, R. K., Zhou, J., Miller, B. L., & Greicius,
M. D. (2009). Neurodegenerative diseases target large-scale
human brain networks. Neuron, 62(1), 42–52. https://doi.org/10
.1016/j.neuron.2009.03.024, PubMed: 19376066
Shibuki, K., Hishida, R., Murakami, H., Kudoh, M., Kawaguchi, T.,
Watanabe, M., … Tanaka, R. (2003). Dynamic imaging of somato-
sensory cortical activity in the rat visualized by flavoprotein auto-
fluorescence. The Journal of Physiology, 549(3), 919–927. https://
doi.org/10.1113/jphysiol.2003.040709, PubMed: 12730344
Singer, W. (1998). Consciousness and the structure of neuronal rep-
resentations. Philosophical Transactions of the Royal Society of
London. Series B: Biological Sciences, 353(1377), 1829–1840.
https://doi.org/10.1098/rstb.1998.0335, PubMed: 9854255
Sporns, O. (2013). Structure and function of complex brain networks.
Dialogues in Clinical Neuroscience, 15(3), 247. https://doi.org/10
.31887/DCNS.2013.15.3/osporns, PubMed: 24174898
Stebbings, K., Choi, H., Ravindra, A., Caspary, D., Turner, J., &
Llano, D. (2016). Ageing-related changes in GABAergic inhibition
in mouse auditory cortex, measured using in vitro flavoprotein
autofluorescence imaging. The Journal of Physiology, 594(1),
207–221. https://doi.org/10.1113/ JP271221, PubMed:
26503482
Steriade, M., & Amzica, F. (1999). Intracellular study of excitability in
the seizure-prone neocortex in vivo. Journal of Neurophysiology,
82(6), 3108–3122. https://doi.org/10.1152/jn.1999.82.6.3108,
PubMed: 10601445
Supekar, K., Menon, V., Rubin, D., Musen, M., & Greicius, M. D. (2008).
Network analysis of intrinsic functional brain connectivity in
Alzheimer’s disease. PLoS Computational Biology, 4(6), e1000100.
https://doi.org/10.1371/journal.pcbi.1000100, PubMed: 18584043
Takesian, A. E., Kotak, V. C., & Sanes, D. H. (2012). Age-dependent
effect of hearing loss on cortical inhibitory synapse function.
Journal of Neurophysiology, 107(3), 937–947. https://doi.org/10
.1152/jn.00515.2011, PubMed: 22090457
Tantipathananandh, C., & Berger-Wolf, T. (2009). Constant-factor
approximation algorithms for identifying dynamic communities.
Paper presented at the Proceedings of the 15th ACM SIGKDD
International Conference on Knowledge Discovery and Data
Mining, Paris, France. https://doi.org/10.1145/1557019.1557110
Tantipathananandh, C., Berger-Wolf, T., & Kempe, D. (2007). A
framework for community identification in dynamic social net-
works. Paper presented at the Proceedings of the 13th ACM
SIGKDD International Conference on Knowledge Discovery
and Data Mining, San Jose, CA, USA. https://doi.org/10.1145
/1281192.1281269
Tantipathananandh, C., & Berger-Wolf, T. Y. (2011). Finding com-
munities in dynamic social networks. Paper presented at the
2011 IEEE 11th International Conference on Data Mining,
Vancouver, Canada. https://doi.org/10.1109/ICDM.2011.67
Terry, R. D., DeTeresa, R., & Hansen, L. A. (1987). Neocortical cell
counts in normal human adult aging. Annals of Neurology, 21(6),
530–539. https://doi.org/10.1002/ana.410210603, PubMed:
3606042
Thambisetty, M., Wan, J., Carass, A., An, Y., Prince, J. L., &
Resnick, S. M. (2010). Longitudinal changes in cortical thickness
associated with normal aging. Neuroimage, 52(4), 1215–1223.
https://doi.org/10.1016/j.neuroimage.2010.04.258, PubMed:
20441796
Thompson, G. J., Merritt, M. D., Pan, W.-J., Magnuson, M. E.,
Grooms, J. K., Jaeger, D., & Keilholz, S. D. (2013). Neural corre-
lates of time-varying functional connectivity in the rat.
NeuroImage, 83, 826–836. https://doi.org/10.1016/j.neuroimage
.2013.07.036, PubMed: 23876248
Thompson, W. H., Richter, C. G., Plavén-Sigray, P., & Fransson, P.
(2018). Simulations to benchmark time-varying connectivity
methods for fMRI. PLoS Computational Biology, 14(5),
e1006196. https://doi.org/10.1371/journal.pcbi.1006196,
PubMed: 29813064
Todd, J.-A. M., Andrews, G., & Conlon, E. G. (2019). Relational
thinking in later adulthood. Psychology and Aging, 34(4), 486.
https://doi.org/10.1037/pag0000346, PubMed: 30973240
Tomasi, D., & Volkow, N. D. (2012). Aging and functional brain
networks. Molecular Psychiatry, 17(5), 549–558. https://doi.org
/10.1038/mp.2011.81, PubMed: 21727896
Turner, J. G., Hughes, L. F., & Caspary, D. M. (2005). Effects of aging
on receptive fields in rat primary auditory cortex layer V neurons.
Network Neuroscience
589
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
2
5
6
9
1
9
2
6
5
8
9
n
e
n
_
a
_
0
0
1
9
1
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Fragmentation of aging mouse cortex revealed by network analysis
Journal of Neurophysiology, 94(4), 2738–2747. https://doi.org/10
.1152/jn.00362.2005, PubMed: 16000522
Vanduffel, W., Payne, B. R., Lomber, S. G., & Orban, G. A. (1997).
Functional impact of cerebralconnections. Proceedings of the
National Academy of Sciences of the United States of America,
94(14), 7617–7620. https://doi.org/10.1073/pnas.94.14.7617,
PubMed: 9207141
Vanzetta, I., Flynn, C., Ivanov, A. I., Bernard, C., & Bénar, C. G.
(2010). Investigation of linear coupling between single-event blood
flow responses and interictal discharges in a model of experimental
epilepsy. Journal of Neurophysiology, 103(6), 3139–3152. https://
doi.org/10.1152/jn.01048.2009, PubMed: 20457851
Vincent, J., Patel, G., Fox, M., Snyder, A., Baker, J., Van Essen, D.,
… Raichle, M. (2007). Intrinsic functional architecture in the
anaesthetized monkey brain. Nature, 447(7140), 83–86. https://
doi.org/10.1038/nature05758, PubMed: 17476267
Wasserman, S., & Faust, K. (1994). Social network analysis. Cambridge
University Press. https://doi.org/10.1017/CBO9780511815478
Watts, D. J., & Strogatz, S. H. (1998). Collective dynamics of
‘small-world’ networks. Nature, 393(6684), 440–442. https://
doi.org/10.1038/30918, PubMed: 9623998
Wen, X., He, H., Dong, L., Chen, J., Yang, J., Guo, H., … Yao, D.
(2020). Alterations of local functional connectivity in lifespan: A
resting-state fMRI study. Brain Behavior, e01652. https://doi.org
/10.1002/brb3.1652, PubMed: 32462815
Wenk, G. L., Walker, L. C., Price, D. L., & Cork, L. C. (1991). Loss
of NMDA, but not GABA-A, binding in the brains of aged rats
and monkeys. Neurobiology of Aging, 12(2), 93–98. https://doi
.org/10.1016/0197-4580(91)90047-n, PubMed: 1646968
Willott, J., Aitkin, L., & McFadden, S. (1993). Plasticity of auditory
cortex associated with sensorineural hearing loss in adult
C57BL/6J mice. Journal of Comparative Neurology, 329, 402–
411. https://doi.org/10.1002/cne.903290310, PubMed:
8459051
Yaron-Jakoubovitch, A., Koch, C., Segev, I., & Yarom, Y. (2013).
The unimodal distribution of sub-threshold, ongoing activity in
cortical networks. Frontiers in Neural Circuits, 7. https://doi.org
/10.3389/fncir.2013.00116, PubMed: 23874270
Zhao, L., Bryce Beverlin, I., Netoff, T., & Nykamp, D. Q. (2011).
Synchronization from second order network connectivity statistics.
Frontiers in Computational Neuroscience, 5. https://doi.org/10
.3389/fncom.2011.00028, PubMed: 21779239
Zheng, Q. Y., Johnson, K. R., & Erway, L. C. (1999). Assessment of
hearing in 80 inbred strains of mice by ABR threshold analyses.
Hearing Research, 130(1–2), 94–107. https://doi.org/10.1016
/s0378-5955(99)00003-9, PubMed: 10320101
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
2
5
6
9
1
9
2
6
5
8
9
n
e
n
_
a
_
0
0
1
9
1
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Network Neuroscience
590