RESEARCH
Static and dynamic aspects of cerebro-cerebellar
functional connectivity are associated with
self-reported measures of impulsivity:
A resting-state fMRI study
Majd Abdallah1, Nicolas Farrugia
2, Valentine Chirokoff1, and Sandra Chanraud 1,3
1Aquitaine Institute of Cognitive and Integrative Neuroscience, UMR CNRS 5287, University of Bordeaux, France
2Electronics Department Lab STICC, IMT Atlantique, UMR CNRS 6285, Brest, France
3Laboratory of Neuroimaging and Daily Life, EPHE, PSL Research University, Bordeaux, France
a n o p e n a c c e s s
j o u r n a l
Keywords: Cerebellum, Cerebro-cerebellar system, Impulsivity, Resting-state fMRI, Static functional
connectivity, Dynamic functional connectivity
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
4
3
8
9
1
1
8
6
7
4
1
3
n
e
n
_
a
_
0
0
1
4
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
ABSTRACT
Human and animal brain studies bring converging evidence of a possible role for the
cerebellum and the cerebro-cerebellar system in impulsivity. However, the precise nature of
the relation between cerebro-cerebellar coupling and impulsivity is far from understood.
Characterizing functional connectivity (FC) patterns between large-scale brain networks that
mediate different forms of impulsivity, and the cerebellum may improve our understanding of
this relation. Here, we analyzed static and dynamic features of cerebro-cerebellar FC using a
highly sampled resting-state functional magnetic resonance imaging (rs-fMRI) dataset and
tested their association with two widely used self-reports of impulsivity: the UPPS-P
impulsive behavior scale and the behavioral inhibition/approach systems (BIS/BAS) in a large
group of healthy subjects (N = 134, ≈ 1 hr of rs-fMRI/subject). We employed robust
data-driven techniques to identify cerebral and cerebellar resting-state networks and extract
descriptive summary measures of static and dynamic cerebro-cerebellar FC. We observed
evidence linking BIS, BAS, sensation seeking, and lack of premeditation to the total strength
and temporal variability of FC within networks connecting regions of the prefrontal cortex,
precuneus, posterior cingulate cortex, basal ganglia, and thalamus with the cerebellum.
Overall, our findings improve the existing knowledge of the neural correlates of impulsivity
and the behavioral correlates of the cerebro-cerebellar system.
AUTHOR SUMMARY
Accumulating evidence from preclinical and neuroimaging studies proposes that the
cerebellum regulates impulsive behavior through its interactions with brain regions that
subserve control and reward processes. To further explore this proposal, we analyzed static
and dynamic aspects of resting-state functional connectivity between the cerebellum and
distinct large-scale brain networks using robust methods and evaluated them against a set of
self-reported measures of impulsivity. We found compelling new evidence linking multiple
forms of impulsivity to strength and temporal variability of FC between large-scale cerebral
networks, involved in top-down control and reward, and the cerebellum. Our findings
highlight the utility of combining static and dynamic FC approaches in furthering current
understanding of cerebro-cerebellar coupling and the neurobiology of complex behaviors.
Citation: Abdallah, M., Farrugia, N.,
Chirokoff, V., & Chanraud, S. (2020).
Static and dynamic aspects of
cerebro-cerebellar functional
connectivity are associated with
self-reported measures of impulsivity:
A resting-state fMRI study. Network
Neuorscience, 4(3), 891–909.
https://doi.org/10.1162/netn_a_00149
DOI:
https://doi.org/10.1162/netn_a_00149
Supporting Information:
https://doi.org/10.1162/netn_a_00149
Received: 19 January 2020
Accepted: 13 May 2020
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Author:
Majd Abdallah
majd.abdallah@u-bordeaux.fr
Handling Editor:
Lucina Uddin
Copyright: © 2020
Massachusetts Institute of Technology
Published under a Creative Commons
Attribution 4.0 International
(CC BY 4.0) license
The MIT Press
Cerebro-cerebellar functional connectivity and impulsivity
Inhibitory control:
The ability to inhibit unwanted
habitual behaviors in order to
perform a task or complete
a certain goal.
Reward sensitivity:
The tendency to detect, seek, and
derive pleasure from rewarding
and desirable stimuli.
Behavioral disinhibition:
The lack of restraint or control over
one’s actions or behaviors commonly
triggered by brain injury.
INTRODUCTION
Impulsivity is a multidimensional personality construct present to varying degrees in healthy
individuals as well as diverse neuropsychiatric populations (Bakhshani, 2014). Psychologists
define impulsivity as the tendency to initiate actions dominated by spontaneity and urgency
with little to no consideration of consequences (Bakhshani, 2014). Self-report questionnaires
are often used to measure two broad constructs highly related to impulsivity: inhibitory con-
trol and reward sensitivity (Jauregi, Kessler, & Hassel, 2018). Interindividual differences in
these processes are believed to arise from genetic and neural origins that are not fully under-
stood (Khadka et al., 2014). However, extensive evidence from neuroimaging studies suggests
that monoaminergic cortico-striatal systems underlie the different constructs of impulsivity
(Dalley, Everitt, & Robbins, 2011; Fineberg et al., 2014; Mitchell & Potenza, 2014). Interest-
ingly, a hypothesis for the involvement of the cerebellum and the cerebro-cerebellar system in
impulsivity has recently been advanced by Miquel, Nicola, Gil-Miravet, Guarque-Chabrera,
and Sanchez-Hernandez (2019) based on numerous findings from human and animal brain
studies. These findings propose that the cerebellum regulates different forms of impulsivity
by interacting with and modulating regions of the prefrontal cortex and the basal ganglia
(Miquel et al., 2019).
Originally thought of as a sensorimotor structure, the cerebellum is now known to be func-
tionally diverse and involved in higher cognitive processes (Buckner, 2013; Strick, Dum, &
Fiez, 2009). Studies have confirmed that the majority of the cerebellum maps onto associa-
tion regions in a manner that mirrors the cerebral asymmetries for cognition, language, and
attention (Buckner, 2013; Habas et al., 2009). In this context, consistent findings from many
studies indicate that the cerebellum may be involved in higher cognitive processes related
to impulsivity (Miquel et al., 2019). Neuroimaging studies have identified structural and func-
tional connections between the cerebellum and brain regions that subserve control and reward
brain processes, such as the prefrontal cortex, anterior cingulate cortex, insula, ventral tegmen-
tal area, thalamus, and basal ganglia (Caligiore et al., 2017; Carta, Chen, Schott, Dorizan, &
Khodakhah, 2019; Moreno-Rius & Miquel, 2017). Moreover, patients with posterior cerebellar
damage have been observed to exhibit difficulties in controlling behavior and emotions, and
show signs of impulsiveness and behavioral disinhibition (Schmahmann & Sherman, 1997).
In addition, certain brain disorders that exhibit alterations in the cerebro-cerebellar circuitry,
such as alcohol use disorder, also feature impulsivity as a key component of the disease pat-
tern (Jung et al., 2014; Miquel et al., 2019). Although supported by accumulating evidence,
the link between cerebro-cerebellar coupling and impulsivity has not been thoroughly inves-
tigated within the framework of functional connectivity (FC). Characterizing the patterns of FC
between cerebral networks that mediate cognitive processes such as inhibitory control and
reward processing, and the cerebellum, may improve our understanding of the hypothesized
cerebellar role in impulsivity.
Resting-state functional magnetic resonance imaging (rs-fMRI) has received attention for the
comprehensive evaluation of interregional FC in the absence of tasks (Sporns, 2018; Van den
Heuvel & Pol, 2010). It has proven to be suitable for the exploration of the neural corre-
lates of certain behaviors, cognitive abilities, and personality traits in both healthy and clini-
cal populations. Resting-state fMRI studies have uncovered important principles of cerebellar
functioning pertaining to the spatiotemporal organization of networks in the cerebellum
(Buckner, Krienen, Castellanos, Diaz, & Yeo, 2011; Guell, Gabrieli, & Schmahmann, 2018).
Importantly, studies have shown that the cerebellum contains individual-specific represen-
tations of most well-known cortical networks including an overrepresented executive con-
trol network (Marek et al., 2018). Until recently, most of the rs-fMRI literature have assessed
Network Neuroscience
892
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
4
3
8
9
1
1
8
6
7
4
1
3
n
e
n
_
a
_
0
0
1
4
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cerebro-cerebellar functional connectivity and impulsivity
Cerebro-cerebellar system:
The set of highly functionally and
anatomically interconnected cerebral
and cerebellar regions.
Domain-general function:
The uniform function of adaptive
control or error-based learning
attributed to the cerebellum across
motor and cognitive domains.
Large-scale brain network:
A collection of widespread brain
regions having strong functional
connectivity and performing the
same function.
Hidden Markov models:
A probabilistic method to analyze
sequential data (e.g., time series) and
find hidden patterns (i.e., states) that
underlie them.
interregional coupling under the assumption of stationary FC, but this “static” approach is
believed to miss out on valuable information embedded in the dynamic nature of the brain
(Calhoun, Miller, Pearlson, & Adalı, 2014; Lurie et al., 2020). Accordingly, recent studies have
begun exploring the temporal dynamics of brain activity and connectivity, and have pointed to
the presence of ongoing temporal reconfiguration of FC strength that supports cognition and
exhibits alterations in several brain disorders (Allen et al., 2014; Lurie et al., 2020). Measures
of dynamic FC complement and, in some cases, outperform measures of static FC in explaining
certain behavioral factors (Liégeois et al., 2019). However, joint information from both cap-
tures more variance in behavior than either alone (Liégeois et al., 2019; Ramos-Nuñez et al.,
2017). Major advances have been made in understanding how cerebral networks dynamically
interact and impact behavior, but little is known about the dynamics of cerebro-cerebellar net-
works and their behavioral correlates. That being said, we believe that exploiting static as well
as dynamic aspects of cerebro-cerebellar FC can provide further insight into the functional
repertoire of the cerebro-cerebellar system.
In this study, we aimed to characterize cross-sectional differences in static and dynamic
cerebro-cerebellar FC, and test for potential associations with self-reported impulsivity. The
uniform cytoarchitecture of the cerebellum suggests that cerebellar modules differ in the pro-
jection of their afferent and efferent connections to sensorimotor and higher cognitive networks
while serving a unitary domain-general function (Marek et al., 2018; Voogd & Glickstein,
1998). Therefore, we sought to estimate the total strength and temporal variability of FC be-
tween cerebral networks of interest and the cerebellum. These measures reflected different
aspects of the total cerebellar influence within distinct large-scale brain networks. We hypoth-
esized that FC between cerebral networks, which are involved in control and reward brain
processes, and the cerebellum could be associated with impulsivity. To test this hypothesis,
we employed an open dataset comprising highly sampled resting-state fMRI data (four runs,
≈ 15 min/run) and self-reports assessing different elements of impulsivity, from a large group
of healthy participants (N = 134, 62 females). We decomposed the rs-fMRI data into separate
cerebral and cerebellar resting-state networks (RSNs), using data-driven techniques, to account
for the functional heterogeneity present in both structures. Then, we estimated static FC among
the identified RSNs and used hidden Markov models (HMMs) to model whole-brain dynamics
and estimate subject-specific dynamic FC matrices (brain states; Vidaurre, Smith, & Woolrich,
2017). Finally, we calculated summary measures of the total strength and temporal variabil-
ity of FC between distinct cerebral RSNs of interest and the cerebellum and evaluated them
against self-reported measures of impulsivity using multivariate general linear models.
MATERIALS AND METHODS
Participants
A total of 134 healthy participants (62 females, ages 20–40 years) from the Neuroanatomy
and Connectivity protocol (N&C), which is part of the Max Planck Institute Leipzig Mind-
Brain-Body (MPILMBB) dataset, were included in this study. All included participants were
healthy with no past or present signs of any neuropsychiatric condition, fulfilled the MRI safety
requirements, and provided written informed consent prior to their participation (Mendes et al.,
2019). Originally, the dataset included fully preprocessed MRI data and a battery of behavioral
assessments from 188 participants. However, because of a gap in the age distribution, we
excluded 26 subjects that were older than 55, and 28 subjects for missing data from source.
Details on the inclusion and exclusion criteria can be found in Mendes et al. (2019).
Network Neuroscience
893
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
4
3
8
9
1
1
8
6
7
4
1
3
n
e
n
_
a
_
0
0
1
4
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cerebro-cerebellar functional connectivity and impulsivity
Self-Reports of Impulsivity
Two widely used self-assessments of impulsivity were included in this study: the UPPS-P im-
pulsive behavior scale and the behavioral inhibition/approach systems scales, widely known
as BIS/BAS. The normality of the different variables was tested using the Shapiro-Wilk test, and
departures from normality were counteracted using rank-based inverse Gaussian transform.
The two self-reports are described in detail in the following paragraphs.
The UPPS-P impulsive behavior scale is a self-report questionnaire designed to measure
impulsive behavior across the five-factor model of personality: negative urgency, positive ur-
gency, sensation seeking, lack of premeditation, and lack of perseverance (Whiteside & Lynam,
2001). Higher scores on the negative and positive urgency subscales indicate a higher tendency
to act rashly under the effect of negative and positive emotions, respectively. A higher score
on the sensation-seeking subscale indicates a higher tendency to seek novel experiences. Fur-
thermore, a higher score on the lack of premeditation subscale indicates a higher tendency
to act rashly without planning or thinking. Finally, a higher score on the lack of perseverance
subscale indicates an increased inability to remain focused and engaged in a possibly arduous
and boring task (Whiteside & Lynam, 2001).
The BIS/BAS scale is also a self-report questionnaire that measures two general motivational
systems argued by theorists to underlie behavior: a behavioral inhibition system (BIS) that reg-
ulates sensitivity to punishment and negative cues, and a behavioral approach system (BAS)
that regulates sensitivity to desirable cues and nonpunishment (i.e., rewards). A higher score
on the BIS scale indicates an increased sensitivity to negative outcomes of anticipated actions
and hence a higher tendency to avoid them, whereas a higher score on the BAS scale indicates
an increased sensitivity to rewards and desirable outcomes and hence a higher tendency to en-
gage in goal-directed behaviors (Gray, 1991). The BAS scale included in this study is the sum of
three subscales: BAS drive, BAS fun seeking, and BAS reward responsiveness. Taken together,
these measures represent a sufficient set of variables that reflect interindividual differences in
inhibitory control and reward sensitivity.
MRI Data Acquisition
The resting-state fMRI acquisition parameters are described in full detail in Mendes et al.
(2019). In summary, four resting-state fMRI scans were acquired for each individual in axial
orientation using T2*-weighted gradient-echo echo planar imaging (GE-EPI) with multiband
acceleration. Sequences were identical across the four runs, with the exception of varying
slice orientation and phase-encoding direction. The phase-encoding direction was anterior–
posterior (AP) for Runs 1 and 3, and posterior–anterior (PA) for Runs 2 and 4. The complete
set of parameters was set as follows: voxel size = 2.3-mm isotropic, FOV = 202 × 202 mm2,
imaging matrix = 88 × 88, 64 slices with 2.3-mm thickness, TR = 1,400 ms, TE = 39.4 ms, flip
angle = 69
, echo spacing = 0.67 ms, bandwidth = 1,776 Hz/Px, partial Fourier 7/8, no pre
scan normalization, multiband acceleration factor = 4,657 volumes, duration = 15 min 30 s per
run. Individuals were instructed to remain awake, during the resting-state scan, with their eyes
open and to fixate on a crosshair. The use of four 15-min rs-fMRI scans per subject enhances
the temporal signal-to-noise ratio (tSNR) in the data, improves the estimation of resting-state
networks, and permits a more reliable modeling of functional connectivity dynamics in the
brain.
◦
Network Neuroscience
894
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
4
3
8
9
1
1
8
6
7
4
1
3
n
e
n
_
a
_
0
0
1
4
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cerebro-cerebellar functional connectivity and impulsivity
Resting-State fMRI Preprocessing
The preprocessing pipeline is described in full detail in Mendes et al. (2019). In summary, the pre-
processing steps included (a) removal of the first five volumes from each of the four resting-state
fMRI runs, (b) rigid-body alignment to the first volume using FSL MCFLIRT to obtain transfor-
mation parameters for motion correction; (c) fieldmap unwarping using FSL-FLIRT and FSL-
FUGUE to estimate transformation parameters for distortion correction (Jenkinson, Beckmann,
Behrens, Woolrich, & Smith, 2012); (d) coregistration to each subject’s structural scan via
FreeSurfer’s boundary-based registration to estimate transformation parameters for coregistra-
tion; (e) normalization of structural scans to MNI152 2-mm space using diffeomorphic nonlin-
ear registration as implemented in ANTsSyN algorithm to estimate transformation parameters
for spatial normalization (Avants et al., 2011) (f) application of all transformation parameters
to each volume in the four resting-state runs in one interpolation step; (g) inclusion of six mo-
tion parameters, their first-order derivatives, and outliers from Nipype’s rapidart algorithm as
nuisance regressors in a general linear model (GLM); (h) The aCompCor method to remove
physiological noise from residual data from the previous denoising step (Behzadi, Restom,
Liau, & Liu, 2007); and finally (i) band-pass filtering [0.01–0.1 Hz]. All included subjects ex-
hibited relatively low in-scanner motion—mean framewise displacement <0.5-mm across all
resting-state scans (Power, Barnes, Snyder, Schlaggar, & Petersen, 2012). Preprocessed data
were obtained from https://ftp.gwdg.de/pub/misc/MPI-Leipzig_Mind-Brain-Body/derivatives/.
Group Independent Component Analysis (GICA)
Preprocessed data from all subjects were analyzed using group independent component anal-
ysis (GICA) as implemented in the GIFT toolbox software http://mialab.mrn.org/software/gift/.
GICA decomposes the rs-fMRI data into linear mixtures of spatially independent components
(ICs) that exhibit unique time course profiles (Allen et al., 2014). In order to investigate cerebro-
cerebellar FC, we decomposed the cerebrum and cerebellum, separately, into spatially indepen-
dent components. By applying a “cerebellum-only” GICA approach, we can extract cerebellar
ICs and signals that are usually overpowered by signals of cortical and subcortical origin when
performing a whole-brain GICA (Dobromyslin et al., 2012). Moreover, parcellating the cere-
bellum permits modeling the patterns of static and dynamic FC among functionally diverse
cerebellar networks and their cerebral counterparts with enhanced accuracy, relative to what
is possible when considering the cerebellum as one homogeneous region of interest. Cerebral
and cerebellar GICA analyses are explained in detail below.
To isolate the cerebellum from the brain, we generated a cerebellar
Cerebellum-only GICA.
mask in MNI152 space using the standard cerebellar MNI152 anatomical template from FSL.
Concatenated cerebellar rs-fMRI data across all subjects and scans were demeaned and ana-
lyzed using principal components analysis (PCA) to reduce the dimensionality to 100 subject-
level PCs (retaining > 99% of the variance in the data) and 25 group-level PCs. Then, we
applied the Infomax algorithm 20 times using the ICASSO toolbox to automatically estimate
and select the most reliable set of 25 independent components in the cerebellum. We then
used the group information guided ICA, or GIG-ICA, to estimate subject-specific ICs and time
series. GIG-ICA extracts subject-specific ICs and time series with better accuracy and corre-
spondence than dual regression (Salman et al., 2019). The choice of number of components
was in accordance with previous studies that identified between 7 and 20 cerebellar RSNs
using different data-driven techniques (Bernard et al., 2012; Buckner et al., 2011; Kipping,
Tuan, Fortier, & Qiu, 2016; Wang, Kipping, Bao, Ji, & Qiu, 2016). However, since noise may
Network Neuroscience
895
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
4
3
8
9
1
1
8
6
7
4
1
3
n
e
n
_
a
_
0
0
1
4
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cerebro-cerebellar functional connectivity and impulsivity
still be present in the data even after preprocessing, we assumed a slightly higher number of
ICs than the putative number of cerebellar RSNs to allow for better disentanglement of signals
from each other and from noise. ICs that exhibited spatial activation near the gray matter/white
matter/cerebro-spinal fluid borders or exhibited irregular patterns with no functional relevance
were discarded as noise, whereas ICs that exhibited unilateral/bilateral spatial activation in the
gray matter or had relevance to well-known cerebellar functional clusters were retained as
RSNs (Buckner et al., 2011). Finally, the time series of cerebellar RSNs were standardized to
have a mean equal to 0 and a standard deviation equal to 1 for each subject in each resting-
state run.
A similar approach to the cerebellum-only GICA was performed in order to
Cerebral GICA.
estimate cerebral RSNs. A brain mask in MNI152 space was generated from a standard FSL
MNI152 brain anatomical template after setting all cerebellar voxels to 0 (Jenkinson et al.,
2012). Concatenated, demeaned data from all subjects and runs were analyzed using PCA to
estimate 120 subject-level PCs (retaining > 99% of the variance in the data) and, subsequently,
30 group-level PCs. We applied the Infomax algorithm 20 times using ICASSO to estimate and
automatically select the most reliable set of 30 ICs, and GIG-ICA to estimate subject-specific
ICs and time series. The choice of the number of components was driven by our interest in large-
scale brain networks that were suitable for subsequent FC analysis in terms of dimensionality,
complexity, and interpretability. ICs that exhibited spatial activation near the edges and in the
white matter were discarded. Finally, the time series of cerebral RSNs were standardized to
have a mean equal to 0 and a standard deviation equal to 1 for each subject in each resting-
state run.
Functional Connectivity Analysis
To construct static FC matrices, we computed pairwise Pearson’s
Static functional connectivity.
full and partial correlation coefficients in each of the four resting-state fMRI runs using the
Ledoit-Wolf estimator as implemented in the nilearn and scikit-learn Python packages
(Abraham et al., 2014; Ledoit & Wolf, 2004). Pearson’s full and partial correlation coefficients
quantify and reflect different types of FC: Full correlations measure direct and indirect func-
tional connections,whereas partial correlations measure direct functional connections only
(Varoquaux & Craddock, 2013). This is informative because cerebro-cerebellar networks form
closed-loop circuits, and it is likely that cerebellar networks directly connect to singular brain
networks rather than affecting large-scale complex processes. However, indirect connections
are also believed to expand the influence of the cerebellum, according to Sokolov, Miall, and
Ivry (2017).
The constructed FC matrices were Fisher r-to-z transformed to stabilize the variance of cor-
relation coefficients and corrected for the effective number of degrees of freedom according to
Bartlett’s theory, which controls for the effect of serial autocorrelation on the estimation of FC
(Afyouni, Smith, & Nichols, 2019; Bartlett, 1946). Then, for each resting-state run, we extracted
the cerebro-cerebellar FC subnetwork and calculated the weighted degree or strength of cere-
bral RSNs of interest as the sum of their positively weighted cerebellar edges, and averaged
the values across all resting-state runs for each subject. This metric reflected the total static
influence of the cerebellum within distinct cerebral RSNs. The cerebro-cerebellar FC strength
metric was calculated using the following formula:
=
Si
J
∑
j=1
wij, wij > 0,
(1)
896
Direct functional connections:
Functional connectivity between any
two brain regions after statistically
removing the effect of all other
regions.
Closed-loops circuits:
The set of neuronal connections that
carry signals from cerebral regions to
the cerebellum and back to the same
regions.
Network Neuroscience
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
4
3
8
9
1
1
8
6
7
4
1
3
n
e
n
_
a
_
0
0
1
4
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cerebro-cerebellar functional connectivity and impulsivity
where Si is the cerebro-cerebellar FC strength of cerebral RSN i, and wij is the weight of the
edge linking cerebral RSN i and cerebellar RSN j. Negatively weighted edges were discarded
because of the lack of consensus and ambiguity surrounding their nature, interpretation, and
means of analysis (Hallquist & Hillary, 2018).
To model whole-brain FC dynamics among the identified
Dynamic functional connectivity.
cerebro-cerebellar RSNs, we applied the hidden Markov models (HMMs) on temporally con-
catenated BOLD time series from all runs and subjects as implemented in the hidden Markov
model multivariate autoregression (HMM-MAR) toolbox https://github.com/OHBA-analysis/
HMM-MAR. The HMMs method is a windowless dynamic FC approach that bypasses the
limitations of sliding-windows and k-means clustering by being directly applied to the BOLD
time series. The method uses variational Bayesian inference to estimate a set intermittently
recurring brain states at the group level, each described as a multivariate Gaussian distribu-
tion with a mean representing a spatial activation pattern and a covariance matrix represent-
ing a FC pattern (Vidaurre et al., 2017). We were mostly interested in FC changes; hence,
brain states were only defined by their Pearson’s full or partial correlation matrices (i.e., FC
patterns) rather than by changes in absolute signal (i.e., spatial activation). We refer to brain
states as dynamic FC states in the remaining sections and subsections of the article. Partial
correlation matrices were obtained by inverting the covariance matrices which were auto-
matically regularized within the Bayesian framework (Ryali et al., 2016). Furthermore, the
Bayesian inference process also permits the estimation of the probability of occurrence of
each state at each time point, along with the Viterbi path that represents the most likely se-
quence of states (Quinn et al., 2018). These were used to estimate the subject-specific state
frequency of occurrence, defined as the number of times a state is visited across all scans.
The HMMs method requires a prespecified number of states, so we assumed a fixed number
of 6 states as a compromise between a lower model order (5 states) and higher model orders
(8, 10, and 12 states) after performing a stability analysis using all configurations (see Supple-
mentary Figure S2). Results obtained using the 5- and 8-state configurations are presented in
Supplementary Tables 2 and 3.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
4
3
8
9
1
1
8
6
7
4
1
3
n
e
n
_
a
_
0
0
1
4
9
p
d
.
t
In order to estimate a descriptive
Subject-specific dynamic FC matrices and temporal variability.
summary measure of the dynamics of cerebro-cerebellar FC, we explored the manifestation of
the states at the subject level. Particularly, we performed an additional iteration of the Bayesian
inference process for each subject, given the initial group-level estimates, states probability of
occurrence at each time point, and the RSNs time series as prior information when updating
and reinferring the states for each subject. This yielded a maximum of 6 dynamic FC states
per subject, each represented by full and partial correlation matrices. The subject-specific dy-
namic FC matrices were Fisher r-to-z transformed and used to calculate the state-wise strength
of cerebral RSNs of interest as the sum of their positively weighted cerebellar edges, in a similar
fashion to the static FC analysis. Then, we calculated the temporal variability of the strength
values for each cerebral RSN i, denoted Vi, as the unbiased frequency-weighted standard de-
viation across the states, in which the subject-specific state frequency of occurrence values
were used as weight factors. This way more frequently visited states contributed more to tem-
poral variability of FC strength. The temporal variability of FC strength was calculated using
the following formula:
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network Neuroscience
Vi
= v
u
u
t
∑6
k=1 fk (cid:0)
∑6
Sik
k=1 fk
− Si(cid:1)
− 1
2
,
(2)
897
Cerebro-cerebellar functional connectivity and impulsivity
where Sik is the cerebro-cerebellar FC strength of cerebral RSN i in state k, and the weight
factor fk is the frequency of occurrence of state k. The rationale behind computing temporal
variability of cerebro-cerebellar FC using the subject-specific dynamic FC states was based
on the observation that static FC matrices highly resembled the frequency-weighted mean of
the dynamic FC matrices: cosine similarity > 0.98 on average for full correlation matrices and
> 0.94 on average for partial correlation matrices (see Supplementary Figure S3). Therefore,
static FC could be considered as a superposition of dynamic FC states identified via HMMs,
as was also observed by Karapanagiotidis et al. (2018). In other words, dynamic FC states
could be considered as nonrandom transient deflections from the static FC pattern at short
timescales.
In order to confirm the presence of robust and genuine
Assessing the robustness of FC dynamics.
dynamic FC in the rs-fMRI data, we generated 100 null datasets from a multivariate Gaussian
distribution fitted to the rs-fMRI data of each individual subject as in Vidaurre et al. (2017).
According to the authors, the correlations between brain regions in the null data are similar
to those in the rs-fMRI data but are presumed to be stationary and exhibit no dynamic struc-
ture. We applied the HMMs method, with unchanged parameters, on each set of null data and
extracted a metric that allowed us to compare the resultant dynamics with to those obtained
in the rs-fMRI data. One important metric that has been used in a previous study is the maxi-
mum fractional occupancy (Vidaurre et al., 2017). Generally speaking, fractional occupancy
(FO) is defined as the proportion of time each state is visited by each subject, whereas maxi-
mum fractional occupancy (maxFO) is the maximum proportion of time spent by each subject
visiting the most occurring state (Vidaurre et al., 2017). High values of maximum fractional
occupancy close to 1 indicate that a single state describes the entirety of the data and hence
the absence of dynamics in FC. On the contrary, low values of maxFO indicate that multiple
recurring states describe the data and hence the presence of dynamics. In this context, we
compared the distributions of maximum fractional occupancy values in the rs-fMRI data with
those in the null data to assess the presence/absence of genuine FC dynamics. More detailed
theoretical and practical information on the use of HMMs to study brain FC dynamics can be
found in Baker et al. (2014), Ryali et al. (2016), Vidaurre et al. (2017), Quinn et al. (2018), and
https://github.com/OHBA-analysis/HMM-MAR/wiki/User-Guide.
Statistical Analysis
To test for associations between cerebro-cerebellar FC and impulsivity, we used multivariate
general linear models (GLMs) that included the impulsivity scales as predictors and the static
and dynamic FC measures as response variables in a multiple linear regression framework. In
addition, age, gender, and mean framewise displacement were included as nuisance covari-
ates. To test for significance and correct for multiple comparisons, we applied nonparametric
permutation testing with 10,000 permutations and a maximum z-statistic procedure to ob-
tain family-wise error adjusted p values across all tests. Particularly, we performed random
rearrangements of the labels in the observed data and estimated an empirical distribution of
maximum z-statistics obtained from all tests and permutations. Then, a family-wise error ad-
justed p value for each test was obtained by computing the proportion of maximum z-scores
that is above the observed z0 for each test in the nonpermuted data. This method provides
strong control of Type I errors without being too conservative, as is the case with conventional
techniques that correct for multiple comparisons (e.g., Bonferroni; Winkler, Ridgway, Webster,
Smith, & Nichols, 2014). We reported significant associations with family-wise error adjusted
p < 0.05.
Network Neuroscience
898
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
3
8
9
1
1
8
6
7
4
1
3
n
e
n
_
a
_
0
0
1
4
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cerebro-cerebellar functional connectivity and impulsivity
Furthermore, to evaluate the replicability of our inference framework, we used repeated
stratified fivefold cross-validation to split the initial sample into training (80% of data, 107
subjects) and testing (20% of data, 27 subjects) subsamples where the proportions of males and
females were preserved in each split. Cross-validation was repeated 100 times with a different
randomization in each repetition. This ensured that all subjects took part in the training and
testing phases across all folds and repetitions. GLMs were refitted to the training data and
then used to predict the outcome in the testing data. We only included the variables that
exhibited significant associations in the previous step (i.e., GLMs fitted to the entire data) while
controlling for age, gender, and mean framewise displacement. Finally, we reported the median
values of the explained variance obtained in the training data, denoted R2
train, and testing data,
denoted R2
test, across all folds and repetitions.
RESULTS
Behavioral and Demographic Data
Summary statistics of demographic and behavioral data are provided in Table 1, whereas the
partial correlations between the different self-reports of impulsivity are provided in Supple-
mentary Table 1 in the Supporting information. Variables were standardized to have a mean
equal to zero and standard deviation equal to one. We assessed for possible multicollinear-
ity using the variance inflation factor (VIF) approach. Most variables were found to have a
VIF < 2 except for the UPPS-P negative and positive urgency subscales. Accordingly, in or-
der to avoid potential multicollinearity effect due to the strong association between the two
subscales (Student’s t = 11.34, r = 0.68, p < 10−15), we used factor analysis to obtain one
urgency factor while preserving a sufficient amount of variance. The final set of self-reported
impulsivity measures included six variables: UPPS-P urgency, UPPS-P lack of premeditation,
UPPS-P lack of perseverance, UPPS-P sensation seeking, behavioral inhibition system (BIS)
scale, and behavioral approach system (BAS) scale.
Group Independent Component Analysis (GICA)
Resting-state fMRI data from the cerebral cortex and subcortex were decom-
Cerebral GICA.
posed into 30 ICs out of which 25 ICs were identified as RSNs based on visual inspection of
the localization of spatial activation in the gray matter (GM) and illustrated in see Figure 1.
Table 1. Behavioral and demographic summary statistics.
Healthy subjects (N = 134, 62 females)
Mean SD Median Min Max VIF Correlation with age (r) Gender difference (t)
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
3
8
9
1
1
8
6
7
4
1
3
n
e
n
_
a
_
0
0
1
4
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Age
PosUrg
NegUrg
Premed
Persev
SenSeek
BIS
BAS
24
1.9
2.2
2
2
2.8
20.5
37.2
4
0.5
0.5
0.4
0.5
0.6
3.1
3.7
25
1.9
2.2
2
1.9
2.8
21
37
20
1
1.2
1.1
1
1.4
12
28
40
3
3.3
3
3.2
4
28
47
–
2.5
2.3
1.3
1.36
1.13
1.15
1.22
–
0.07
−0.01
−0.02
0.04
−0.18
−0.1
0.1
∗
−0.464
0.13
2.91
−0.74
−1.54
−3.32
4.73
0.45
∗∗
∗∗
Note. SD: standard deviation,VIF: variance inflation factor, r : Pearson’s correlation, t : student’s t,
PosUrg: positive urgency, NegUrg: negative urgency, Premed: lack of premeditation, Persev: lack
of perseverance, SenSeek: sensation seeking, BIS: behavioral inhibition system, BAS: behavioral
approach system. * p < 0.05 , ** p < 0.01.
Network Neuroscience
899
Cerebro-cerebellar functional connectivity and impulsivity
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
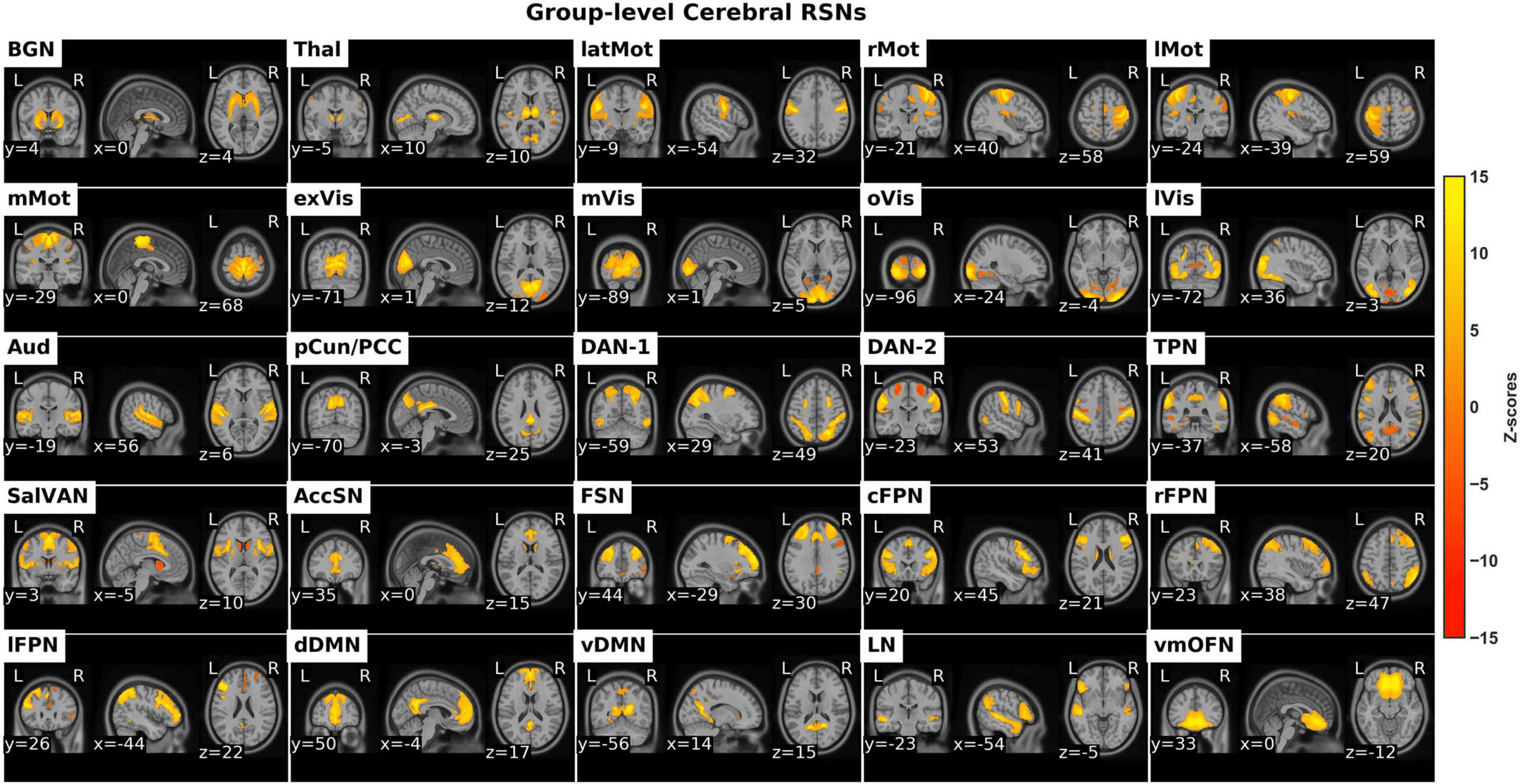
Figure 1. The identified cerebral RSNs. BGN: basal ganglia network, Thal: thalamus, latMot: lateral motor network, rMot: right motor
network, lMot: left motor network, mMot: medial motor network, exVis: extra-striate visual network, mVis: medial visual network, oVis:
occipital visual network, lVis: lateral visual network, Aud: auditory network, pCun/PCC: precuneus/posterior cingulate cortex network, DAN:
dorsal attention network, TPN: task positive network, SalVAN: salience-ventral attention network, AccSN: anterior cingulate cortex salience
network, FSN: frontal salience network, cFPN: central frontoparietal network, rFPN: right frontoparietal network, lFPN: left frontoparietal
network, dDMN: dorsal default mode network, vDMN: ventral default mode network, LN: language network, vmOFN: ventro-medial orbito-
frontal network.
/
/
/
/
4
3
8
9
1
1
8
6
7
4
1
3
n
e
n
_
a
_
0
0
1
4
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The retained RSNs were arranged into groups of putative functional groups based on their
anatomical and functional properties. The functional groups were subcortical, motor, visual,
auditory, attention, salience, frontoparietal, and default mode.
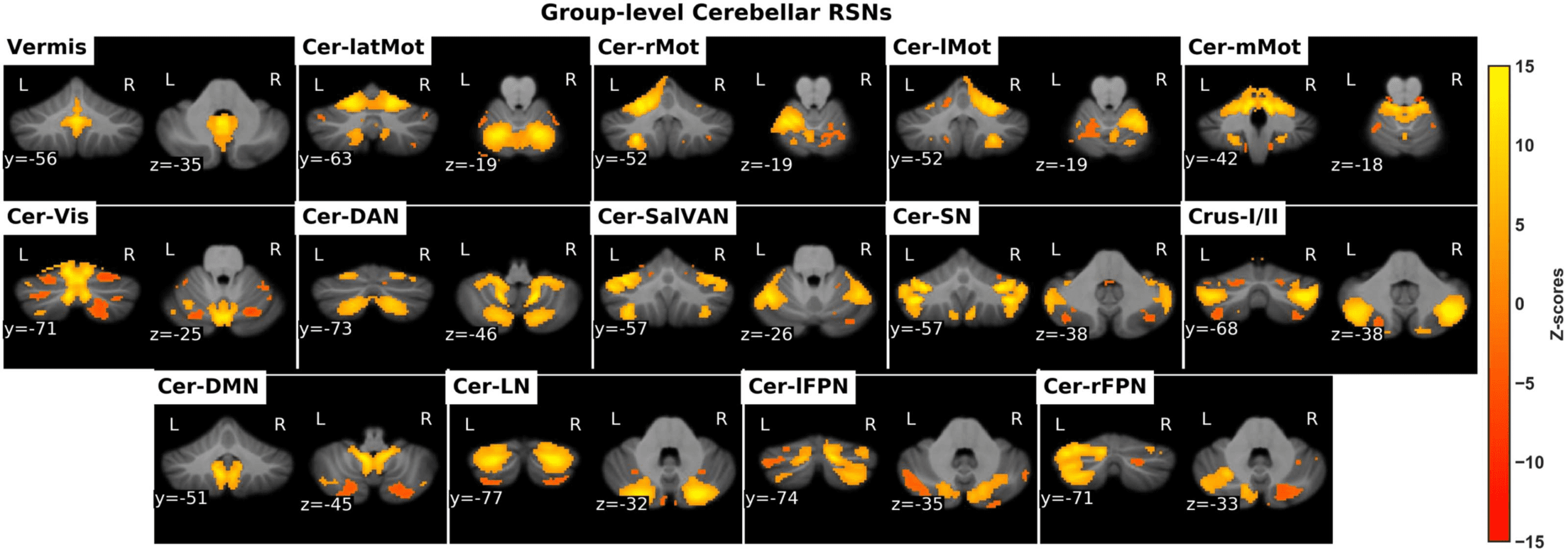
Resting-state fMRI data from the cerebellum were decomposed into
Cerebellum-only GICA.
25 ICs, out of which 14 ICs were visually identified as cerebellar RSNs and illustrated in
Figure 2, while 11 ICs were identified as noise and hence discarded. The RSNs were arranged
into groups of putative functional domains based on their anatomical or functional properties
and overlap with previously established cerebellar clusters based on a winner-takes-all ap-
proach (Buckner et al., 2011). The functional clusters were motor, visual, attention, salience,
frontoparietal, and default mode. However, two cerebellar RSNs, whose spatial activation maps
were well situated in the GM, did not overlap with well-known cerebellar clusters. These were
labelled as “Vermis” and “Crus-I/II” based on the anatomical landmarks that overlap with their
spatial activation maps. In addition, taking into consideration the contralateral representation
of large-scale networks in the cerebellum, labels of unilateral cerebellar RSNs were inverted.
For instance, if spatial activation was mostly localized in the left posterior cerebellum, the nam-
ing would be cerebellar right frontoparietal (Cer-rFPN) because of the inverted frontoparietal
map present in the posterior cerebellar lobe.
Network Neuroscience
900
Cerebro-cerebellar functional connectivity and impulsivity
Figure 2. The identified cerebellar RSNs. Cer-latMot: cerebellar lateral motor network, Cer-rMot: Cerebellar right motor network, Cer-lMot:
cerebellar left motor network, Cer-mMot: cerebellar medial motor network, Cer-Vis: cerebellar visual network, Cer-DAN: cerebellar dorsal
attention network, Cer-SalVan: cerebellar salience/ventral attention network, Cer-SN: cerebellar salience network, Cer-DMN: cereballar default
mode network, Cer-LN: cerebellar language network, Cer-lFPN: cerebellar left frontoparietal network, Cer-rFPN: cerebellar right fronto-parietal
network.
Table 2. Significant associations between cerebro-cerebellar FC and self-reported measures of
impulsivity.
Network
FC metric Impulsivity scale
BGN-cerebellum
Strength
Variability SenSeek
BIS
Thal-cerebellum
Variability SenSeek
FSN-cerebellum
Strength
BAS
Variability Premed
Variability SenSeek
pCun/PCC-cerebellum Variability Premed
z
−3
3.1
3.3
β
train R2
R2
p
−0.31 0.038 0.07
test
0.068
0.037 0.068 0.063
0.3
0.32 0.019 0.078 0.074
−3.1 −0.29 0.033 0.08
0.073
−3.5 −0.34 0.008 0.092 0.09
0.32 0.019 0.078 0.08
3.3
−3.7 −0.36 0.003 0.11
0.096
Variability SenSeek
3.6
0.35 0.005 0.093 0.086
Note. Strength: cerebro-cerebellar FC strength, Variability: temporal variability of cerebro-
cerebellar FC strength, z: z-statistic, β: standardized regression coefficient, p: family-wise
error adjusted p value, R2
test:
median of explained variance in the testing data.
train: median of explained variance in the training data, R2
Strength of Cerebro-Cerebellar FC and Impulsivity
The details of statistically significant findings are reported in Table 2, whereas scatterplots are
illustrated in Figure 3. Figure 5 illustrates the group-average partial correlation FC matrix and
the strongest positively weighted cerebro-cerebellar connections. It clearly shows the well-
documented topographic dichotomy of motor versus nonmotor cerebellum and the mostly
domain-specific functional connections between cerebral and cerebellar RSNs. In accordance
with our hypothesis, only static FC measures pertaining to cerebral RSNs that include regions
involved in control and reward brain processes, notably the frontoparietal, salience, atten-
tion, subcortical, and default mode networks, were evaluated against self-reported measures
of impulsivity using GLMs.
Network Neuroscience
901
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
3
8
9
1
1
8
6
7
4
1
3
n
e
n
_
a
_
0
0
1
4
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cerebro-cerebellar functional connectivity and impulsivity
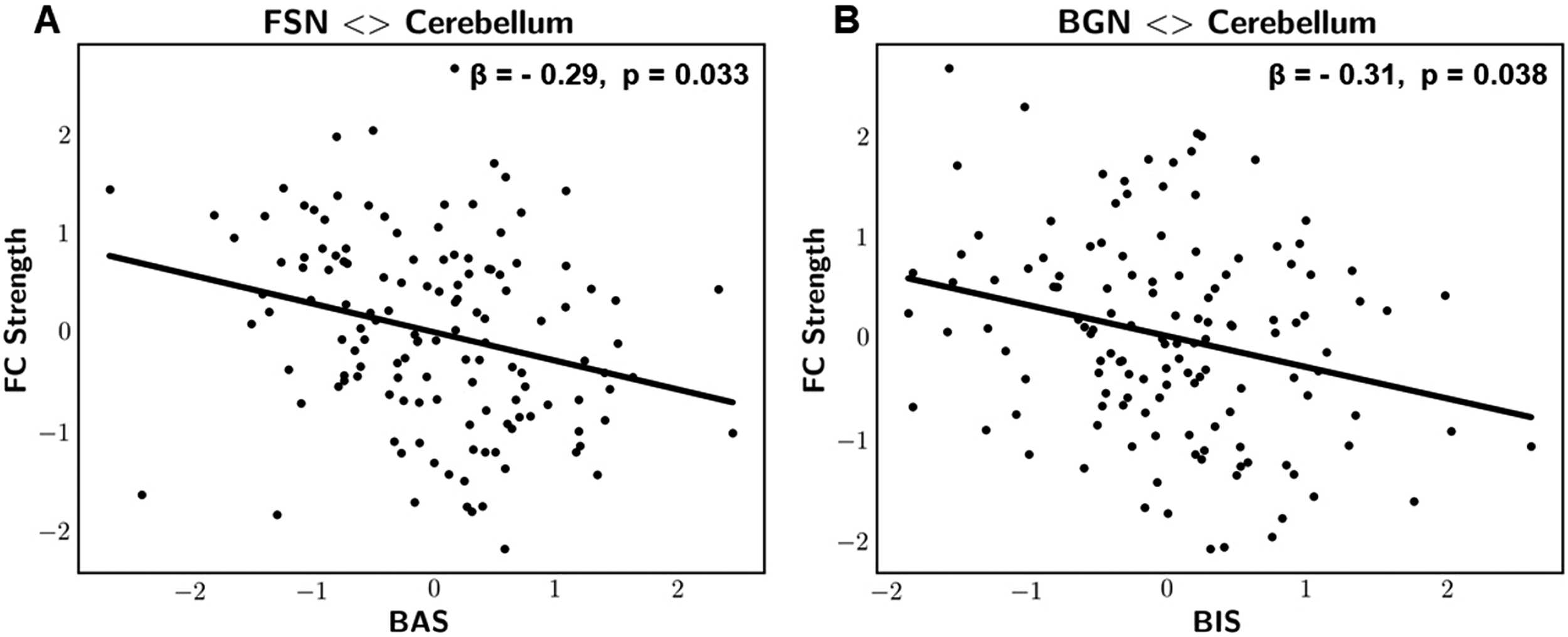
Figure 3. Scatterplot representations of significant associations between cerebro-cerebellar FC
strength and the BIS/BAS scales. (A) FSN-cerebellum FC strength vs. behavioral approach system
(BAS). (B) BGN-cerebellum FC strength vs. behavioral inhibition system (BIS). Variables were ad-
justed for age, gender, and mean framewise displacement and presented as z-scores. β: standardized
regression coefficient, p: FWE-adjusted p values.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
3
8
9
1
1
8
6
7
4
1
3
n
e
n
_
a
_
0
0
1
4
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
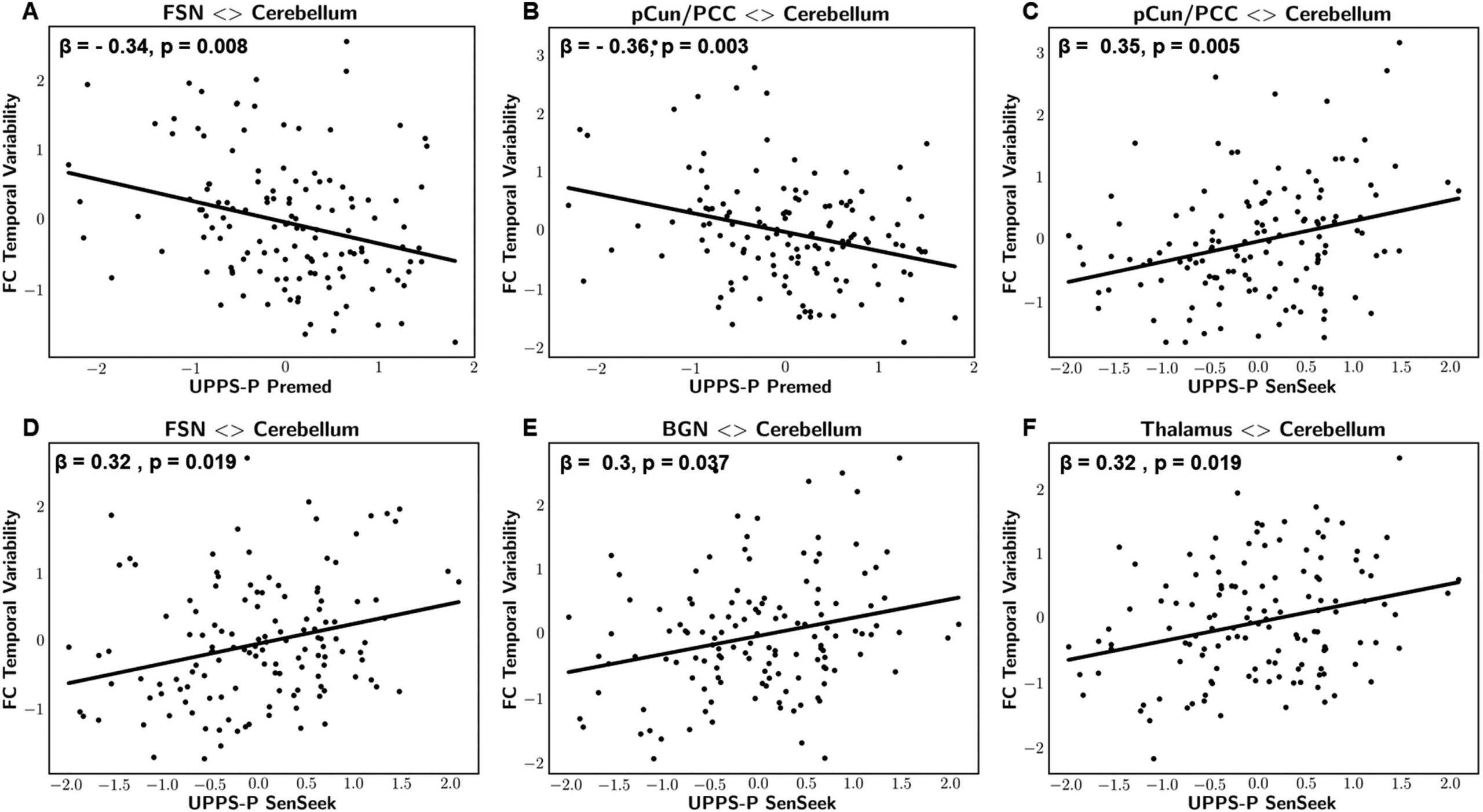
Figure 4. Scatterplot representations of statistically significant associations between dynamics of cerebro-cerebellar FC and UPPS-P mea-
sures: sensation seeking and lack of premeditation. Panels A and B represent temporal variability in FSN-cerebellum and pCun/PCC-cerebellum
FC strength vs. UPPS-P lack of premeditation, respectively. Panels C, D, E, and F represent temporal variability in FSN-cerebellum, pCun/PCC-
cerebellum, BGN-cerebellum, and thalamus-cerebellum FC strength vs. UPPS-P sensation seeking, respectively. All variables were adjusted for
age, gender, and mean framewise displacement and presented as z-scores. β: standardized regression coefficient, p: FWE-adjusted p values.
Network Neuroscience
902
Cerebro-cerebellar functional connectivity and impulsivity
Statistically significant associations were observed when quantifying static FC using partial
correlation coefficients. Particularly, results revealed a significant negative correlation between
the behavioral inhibition system (BIS) scale and the cerebro-cerebellar strength of the basal
= 0.068). In
ganglia network, BGN (z = −3, β = −0.31, p = 0.038, R2
addition, we identified a significant negative correlation between the behavioral approach
system (BAS) scale and the cerebro-cerebellar strength of the frontal salience network, FSN
= 0.073). The reported p values were
(z = −3.1, β = −0.29, p = 0.033, R2
= 0.08, R2
family-wise error adjusted.
= 0.07, R2
train
train
test
test
Temporal Variability of Cerebro-Cerebellar FC and Impulsivity
The details of statistically significant findings are reported in Table 2, whereas scatterplots
are illustrated in Figure 4. We demonstrated the inferred group-level brain FC matrices (states)
along with distribution profiles of the maximum fractional occupancy values obtained from the
rs-fMRI data and the generated null data in Supplementary Figure 1. The distribution profiles
and the group-average frequency of occurrence values (percentages) indicated the presence
of genuine FC dynamics in the rs-fMRI data as opposed to the null data, which were mostly
described by a single state. In accordance with our hypothesis, only dynamic FC measures
pertaining to cerebral RSNs that encompass regions involved in control and reward brain pro-
cesses, notably the frontoparietal, salience, attention, subcortical, and default mode networks,
were evaluated against self-reported measures of impulsivity using GLMs.
Associations were observed when describing dynamic FC states using Pearson’s full cor-
relation matrices. Particularly, results revealed significant negative correlations between the
UPPS-P lack of premeditation subscale and the temporal variability of cerebro-cerebellar FC
= 0.09)
strength of the FSN (z = −3.5, β = −0.34, p = 0.008, R2
and the precuneus/posterior cingulate cortex (pCun/PCC) network (t = −3.7, β = −0.36,
= 0.096). In addition, results revealed significant positive corre-
p = 0.003, R2
lations between the UPPS-P sensation-seeking subscale and the temporal variability of cerebro-
= 0.08),
cerebellar FC strength of the FSN (z = 3.3, β = 0.32, p = 0.019, R2
= 0.086), the
the pCun/PCC network (z = 3.6, β = 0.35, p = 0.005, R2
= 0.063), and the thalamus network
BGN (z = 3.1, β = 0.3, p = 0.037, R2
= 0.074). The reported p values were
(z = 3.3, β = 0.32, p = 0.019, R2
family-wise error adjusted.
= 0.092, R2
= 0.093, R2
= 0.078, R2
= 0.068, R2
= 0.078, R2
= 0.11, R2
train
train
train
train
train
train
test
test
test
test
test
test
Top-down control:
The process of guiding and directing
behavior towards a goal or in
response to a changing environment.
DISCUSSION
In the present study, we investigated the proposed cerebellar role in impulsivity by character-
izing different aspects of cerebro-cerebellar resting-state FC and testing their association with
self-reported measures of impulsivity. We observed evidence that linked summary measures
of static and dynamic cerebro-cerebellar resting-state FC to different forms of impulsivity in a
large group of healthy subjects. Particularly, we found that the total strength and temporal vari-
ability of FC between a set of large-scale cerebral networks, involved in top-down control and
reward brain processes, and the cerebellum were associated with the BIS, BAS, UPPS-P lack of
premeditation, and UPPS-P sensation seeking. Nonparametric permutation testing and cross-
validation based on repeated stratified fivefolds supported the significance and cross-validity
of these associations, respectively. Overall, our findings improve current knowledge on the
behavioral correlates of the cerebro-cerebellar system and the neural correlates of impulsiv-
ity. In addition, they highlight the utility of both static and dynamic FC analysis approaches
in investigating the principles of cerebro-cerebellar coupling and the neural underpinnings
Network Neuroscience
903
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
3
8
9
1
1
8
6
7
4
1
3
n
e
n
_
a
_
0
0
1
4
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cerebro-cerebellar functional connectivity and impulsivity
of impulsivity and potentially other constructs of personality. We discuss significant findings,
limitations, and future perspectives of the study in the following paragraphs.
Cerebro-Cerebellar Static FC Strength and BIS/BAS Scales
The static FC analysis results revealed a significant negative correlation between the cerebro-
cerebellar FC strength of the basal ganglia network (BGN) and the behavioral inhibition system
(BIS) scale, which measures sensitivity towards unpleasant cues and punishment. This indicates
that increased striato-cerebellar FC strength is associated with a lower tendency to avoid or
inhibit actions with possibly negative outcomes such as punishment. We believe that this is
in line with previous findings suggesting that the involvement of the cerebellum in inhibitory
control mechanisms is mediated by its interaction with the basal ganglia (Brunamonti et al.,
2014). In addition, we believe that this is consistent with accumulating evidence suggesting
that increased connectivity between the cerebellum and the basal ganglia is associated with an
overreliance on motivated “go” brain mechanisms at the expense of “no-go” inhibitory control
mechanisms, and hence with increased impulsivity (Miquel et al., 2019). Results also revealed
a significant negative correlation between the cerebro-cerebellar FC strength of the frontal
salience network (FSN) and the behavioral approach system (BAS) scale, which measures the
tendency to approach goals, rewards, nonpunishment, or escape from punishment. The FSN
included parts of the dorsolateral prefrontal cortex, superior frontal cortex, fronto-polar cortex,
and intraparietal cortex (or Brodmann areas BA7, BA8, BA9, and BA10) that are involved in
top-down control of responses to stimuli (Corbetta & Shulman, 2002). Previous studies have
proposed that the cerebellum engages with prefrontal regions in restraining ongoing actions
in response to changing environmental cues, thereby promoting prefrontal functionality in
top-down control of goal-directed behaviors (Miquel et al., 2019). We believe that the above-
mentioned finding is in line with this proposal and indicates that stronger fronto-cerebellar FC
is associated with lower activation of approach behavior towards rewarding cues, and hence
increased top-down control of goal-directed behaviors.
The associations between cerebro-cerebellar FC strength and the BIS/BAS scales were ob-
served when quantifying static FC using partial correlation coefficients. Partial correlation ma-
trices are sparse representations of FC in the brain, and they may relate to the underlying
structural connectivity better than full correlation coefficients (Varoquaux & Craddock, 2013).
In this case, the cerebro-cerebellar FC strength of a cerebral RSN is the sum of its positively
weighted direct links to the cerebellum, and hence might be related to the parallel closed-loop
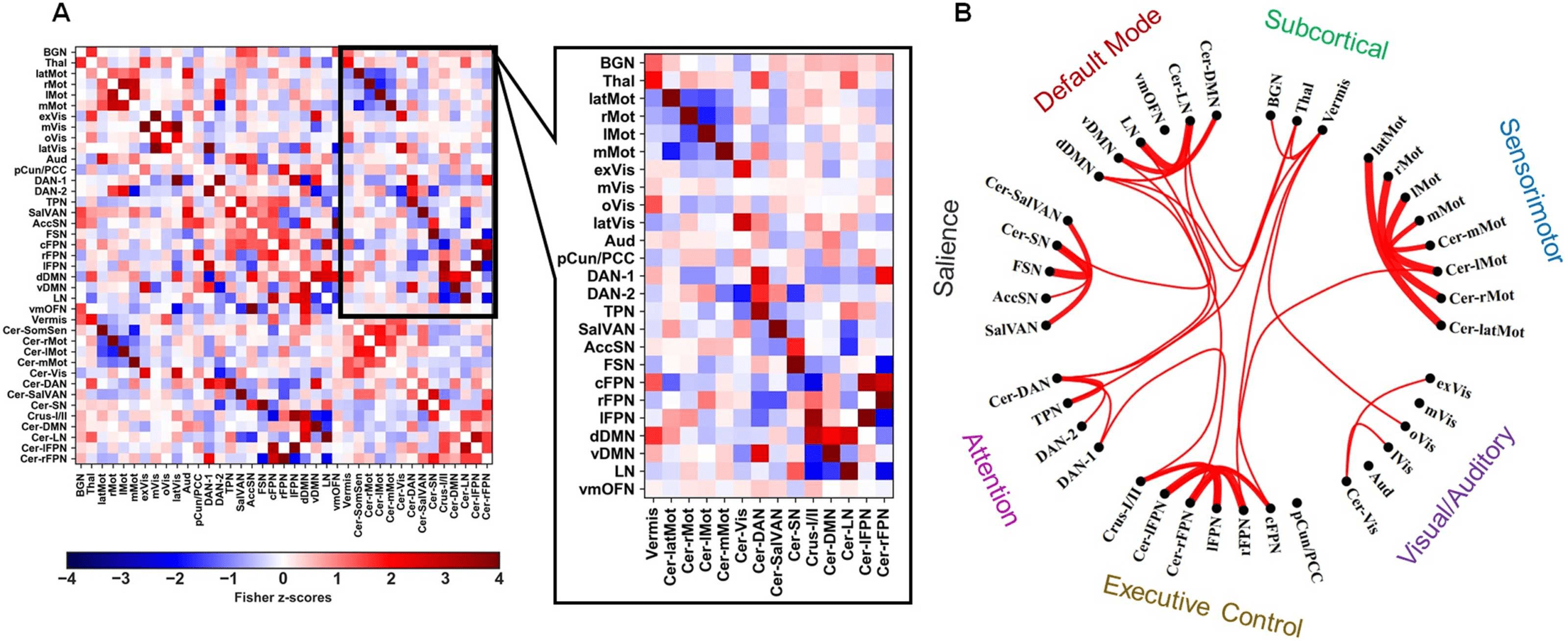
cerebro-cerebellar circuitry. This is illustrated in Figure 5, which clearly shows significantly
stronger within–functional domain cerebro-cerebellar connections than between domains at
the group level. Even though we calculated the total strength of FC between distinct cere-
bral RSNs of interest and the cerebellum, the FSN was found to be mostly connected to the
cerebellar salience network (Cer-SN), which overlaps with the lateral posterior cerebellum,
including Crus I and II, whereas the BGN was found to be mostly connected to the cerebellar
Vermis network, which mostly overlaps with the posterior lobules of the vermis. Interestingly,
previous studies have shown that the lateral posterior cerebellum is mainly involved with ex-
ecutive control networks, whereas the posterior vermis is involved with the limbic system and
connects to the ventral tegmental area (VTA), which is the center of dopaminergic cell bodies
(Buckner et al., 2011; Guell et al., 2018; Miquel et al., 2019; Stoodley & Schmahmann, 2010).
Moreover, lesions to the cerebellar vermis have been shown to induce an array of behavioral
and emotional disturbances, including impulsiveness and behavioral disinhibition (Kim, Kim,
Choi, Chung, & Moon, 2013; Miquel et al., 2019; Schmahmann, Weilburg, & Sherman, 2007).
This suggests that these cerebellar structures are likely to influence processes related to control
Network Neuroscience
904
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
3
8
9
1
1
8
6
7
4
1
3
n
e
n
_
a
_
0
0
1
4
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cerebro-cerebellar functional connectivity and impulsivity
Figure 5.
strongest 20% of direct cerebro-cerebellar connections in the group-average static FC matrix.
(A) The group-average partial correlation static FC matrix and the cerebro-cerebellar submatrix. (B) A circular graph showing the
and reward by modulating prefrontal and striatal regions (Miquel et al., 2019). We believe
that our findings are in line with these results, and we propose that the cerebellar influence
over the fronto-striatal circuit affects motivated behaviors, in accordance with previous studies
(Brunamonti et al., 2014). However, the methods used in our study are concerned with undi-
rected FC measures that do not provide information on causal influences that neural units exert
over one another. Thus, investigations using measures of directed FC, such as effective connec-
tivity, are needed for a more comprehensive understanding of the modulatory role of different
cerebellar regions in impulsivity and potentially other constructs of personality (Stephan &
Friston, 2010).
Cerebro-Cerebellar Dynamic FC and Impulsivity
Dynamic FC studies have defined temporal variability of FC as a measure of how brain re-
gions are transiently integrated and segregated across time (Calhoun et al., 2014; Lord, Stevner,
Deco, & Kringelbach, 2017). Higher values of temporal variability point to increased switching
of brain FC patterns (i.e., states), which has been shown to support cognitive flexibility (Douw,
Wakeman, Tanaka, Liu, & Stufflebeam, 2016). In the context of the current study, the greater
the temporal variability, the more frequent the switching of FC strength between cerebral RSNs
and their cerebellar counterparts. Our results revealed negative correlations between the tem-
poral variability of cerebro-cerebellar FC strength of two top-down control networks, namely
the frontal salience network (FSN) and the precuneus/posterior cingulate cortex (pCun/PCC)
network, and the UPPS-P lack of premeditation subscale, which measures the tendency to act
rashly without thinking. The pCun/PCC network primarily included dorsal parts of the pre-
cuneus cortex (pCun) and the dorsal posterior cingulate cortex (PCC), which is regarded as
part of the executive control network and is also connected to the default mode and salience
networks (Leech & Sharp, 2013; Yeo et al., 2011). This suggests that the increase in the overall
switching of FC strength between the cerebellum and top-down control regions is associated
with increased control of impulses. In addition, temporal variability of cerebro-cerebellar FC
Network Neuroscience
905
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
3
8
9
1
1
8
6
7
4
1
3
n
e
n
_
a
_
0
0
1
4
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cerebro-cerebellar functional connectivity and impulsivity
strength of these two top-down control networks along with two other networks involved in
reward processing, namely the basal ganglia network (BGN) and the thalamus, were positively
correlated with the UPPS-P sensation-seeking scale, which measures the tendency to seek new
and rewarding experiences. This suggests that the increase in switching of FC strength between
the cerebellum and networks involved in top-down control and reward processing is associ-
ated with enhanced salience attribution of novel experiences.
We observed associations between temporal variability of cerebro-cerebellar FC and im-
pulsivity when using full correlation matrices to describe dynamic FC states. Pearson’s full cor-
relation coefficients measure both direct and indirect pairwise connections that may relate to
integrative brain systems involved in complex brain processes. It has been hypothesized that
the cerebellum and the cerebro-cerebellar networks are key components of the integrative
brain systems promoting the prediction, organization, and comprehension of complex se-
quences involved in higher cognitive domains (Barton, 2012; Miquel et al., 2019). In this con-
text, our findings point to a possible dynamic system that flexibly connects multiple regions of
the prefrontal cortex, precuneus, posterior cingulate cortex, basal ganglia, and thalamus with
each other and with the cerebellum, and that might play a role in different elements of im-
pulsivity. This might also be related to the postulate that the cerebellum coordinates and links
cognitive units of thought that arise from multiple brain regions in a similar fashion to coordi-
nating multimuscled movements, hence facilitating cognitive processing (Buckner, 2013). In
other words, cerebellar modules might be dynamically recruited to help ensure smooth and
coordinated information flow within and between top-down control and reward networks,
hence influencing complex processes that underlie different forms of impulsivity. However,
since we analyzed the dynamics of undirected FC and only reported correlations with impul-
sivity, the interpretations of the current findings remain speculative. Thus, a causal link between
the dynamics of cerebro-cerebellar FC and impulsivity cannot be concluded in this case, and
extensive investigations are definitely needed in the future. However, given the body of ev-
idence linking dynamic FC to cognitive flexibility and learning, and the universal principle
of cerebellar functioning in adaptive control and prediction, we believe that investigating the
dynamics of cerebro-cerebellar functional networks has the potential of enhancing our under-
standing of the principles of cerebellar functioning in the cognitive domains (Bassett et al.,
2011; Douw et al., 2016; Sokolov et al., 2017).
Limitations and Future Perspectives
There are several limitations worth noting in the current study. A first limitation, we believe,
is the use of low-dimensional GICA decomposition of the cerebrum and cerebellum that only
estimated large-scale RSNs. Although this choice served our goals and interests, it was driven
by the fact that computing partial correlation coefficients from high-dimensional data is com-
putationally challenging, and the fact that the HMMs method may not converge reliably when
the number of considered brain regions is high (Karapanagiotidis et al., 2018; Varoquaux &
Craddock, 2013). High-dimensional representations of the brain decompose large-scale RSNs
further into subnetworks and possibly aid in exploring associations between left versus right
cerebro-cerebellar networks and impulsivity. Developing new methods that can reliably pro-
cess high-dimensional data is needed in order to overcome this limitation. A second limitation
is the sole use of positively weighted edges to estimate the strength and temporal variability
of FC while discarding negatively weighted edges. Negatively weighted edges are often not
analyzed because of the ambiguity and controversy surrounding their nature and means of
analysis (Hallquist & Hillary, 2018). In addition, the importance of negative edges in cerebro-
cerebellar FC is still unknown. Thorough investigations into the nature of negative FC and its
Network Neuroscience
906
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
3
8
9
1
1
8
6
7
4
1
3
n
e
n
_
a
_
0
0
1
4
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cerebro-cerebellar functional connectivity and impulsivity
relation to cerebro-cerebellar coupling should be conducted in future research. A third lim-
itation of this study is the lack of objective measurements of impulsivity such as tasks (e.g.,
go/no-go task, stop-signal task, reward devaluation tests), which could have provided another
perspective with less bias than self-reported measures. Finally, future investigations into the role
of the cerebellum in impulsivity and other related traits should explore gender and age differ-
ences and include individuals diagnosed with neuropsychiatric disorders, such as alcohol-use
disorder and attention deficit hyperactivity disorder, that are known to feature impulsive and
compulsive symptomatology and exhibit alterations in the cerebro-cerebellar system.
ACKNOWLEDGMENTS
The authors thank Joel Swendsen, Diego Vidaurre, Alexander Schäfer, Soroush Afyouni, and
Judy Kipping for valuable advice and recommendations.
SUPPORTING INFORMATION
Supporting information for this article is available at https://doi.org/10.1162/netn_a_00149.
AUTHOR CONTRIBUTIONS
Majd Abdallah: Conceptualization; Formal analysis; Investigation; Methodology; Software;
Visualization; Writing - Original Draft; Writing - Review & Editing. Nicolas Farrugia: Data
curation; Formal analysis; Methodology; Supervision; Validation; Writing - Review & Edit-
ing. Valentine Chirokoff: Data curation; Investigation. Sandra Chanraud: Funding acquisition;
Project administration; Supervision; Validation; Writing - Review & Editing.
FUNDING INFORMATION
Sandra Chanraud, Translational Research and Advanced Imaging Laboratory (TRAIL), Award
ID: TRAIL-ANR-10-LABX-57.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
3
8
9
1
1
8
6
7
4
1
3
n
e
n
_
a
_
0
0
1
4
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
REFERENCES
Abraham, A., Pedregosa, F., Eickenberg, M., Gervais, P., Mueller, A.,
Kossaifi, J., . . . Varoquaux, G. (2014). Machine learning for neu-
roimaging with scikit-learn. Frontiers in Neuroinformatics, 8, 14.
(2019). Effective de-
grees of freedom of the Pearson’s correlation coefficient under
autocorrelation. NeuroImage, 199, 609–625.
Afyouni, S., Smith, S. M., & Nichols, T. E.
Allen, E. A., Damaraju, E., Plis, S. M., Erhardt, E. B., Eichele, T., &
(2014). Tracking whole-brain connectivity dy-
Calhoun, V. D.
namics in the resting state. Cerebral Cortex, 24(3), 663–676.
Avants, B. B., Tustison, N. J., Song, G., Cook, P. A., Klein, A., & Gee,
J. C. (2011). A reproducible evaluation of ANTs similarity met-
ric performance in brain image registration. NeuroImage, 54(3),
2033–2044.
Baker, A. P., Brookes, M. J., Rezek, I. A., Smith, S. M., Behrens, T.,
Smith, P. J. P., & Woolrich, M. (2014). Fast transient networks in
spontaneous human brain activity. eLife, 3, e01867.
Bakhshani, N.-M. (2014). Impulsivity: A predisposition toward risky
behaviors. International Journal of High Risk Behaviors and Ad-
diction, 3(2).
Bartlett, M. S.
(1946). On the theoretical specification and sam-
pling properties of autocorrelated time-series. Supplement to the
Journal of the Royal Statistical Society, 8(1), 27–41.
Barton, R. A. (2012). Embodied cognitive evolution and the cerebel-
lum. Philosophical Transactions of the Royal Society B: Biological
Sciences, 367(1599), 2097–2107.
Bassett, D. S., Wymbs, N. F., Porter, M. A., Mucha, P. J., Carlson,
J. M., & Grafton, S. T.
(2011). Dynamic reconfiguration of hu-
man brain networks during learning. Proceedings of the National
Academy of Sciences, 108(18), 7641–7646.
Behzadi, Y., Restom, K., Liau, J., & Liu, T. T.
(2007). A compo-
nent based noise correction method (CompCor) for BOLD and
perfusion based fMRI. NeuroImage, 37(1), 90–101.
Bernard, J. A., Seidler, R. D., Hassevoort, K. M., Benson, B. L.,
Welsh, R. C., Wiggins, J. L., . . . Peltier, S. J. (2012). Resting state
cortico-cerebellar functional connectivity networks: A compari-
son of anatomical and self-organizing map approaches. Frontiers
in Neuroanatomy, 6, 31.
Network Neuroscience
907
Cerebro-cerebellar functional connectivity and impulsivity
Brunamonti, E., Chiricozzi, F. R., Clausi, S., Olivito, G., Giusti,
M. A., Molinari, M., . . . Leggio, M. (2014). Cerebellar damage
impairs executive control and monitoring of movement genera-
tion. PLoS ONE, 9(1).
Buckner, R. L. (2013). The cerebellum and cognitive function: 25
years of insight from anatomy and neuroimaging. Neuron, 80(3),
807–815.
Buckner, R. L., Krienen, F. M., Castellanos, A., Diaz, J. C., & Yeo,
B. T. T. (2011). The organization of the human cerebellum esti-
mated by intrinsic functional connectivity. Journal of Neurophys-
iology, 106(5), 2322–2345. (PMID: 21795627)
Calhoun, V. D., Miller, R., Pearlson, G., & Adalı, T.
(2014). The
chronnectome: Time-varying connectivity networks as the next
frontier in fMRI data discovery. Neuron, 84(2), 262–274.
Caligiore, D., Pezzulo, G., Baldassarre, G., Bostan, A. C., Strick,
P. L., Doya, K., . . . Herreros, I.
(2017). Consensus paper:
Towards a systems-level view of cerebellar function: The inter-
play between cerebellum, basal ganglia, and cortex. Cerebellum,
16(1), 203–229.
Carta, I., Chen, C. H., Schott, A. L., Dorizan, S., & Khodakhah, K.
(2019). Cerebellar modulation of the reward circuitry and social
behavior. Science, 363(6424), eaav0581.
Corbetta, M., & Shulman, G. L.
(2002). Control of goal-directed
and stimulus-driven attention in the brain. Nature Reviews Neu-
roscience, 3(3), 201.
Dalley, J. W., Everitt, B. J., & Robbins, T. W.
Impulsivity,
compulsivity, and top-down cognitive control. Neuron, 69(4),
680–694.
(2011).
Dobromyslin, V.
I., Salat, D. H., Fortier, C. B., Leritz, E. C.,
Beckmann, C. F., Milberg, W. P., & McGlinchey, R. E.
(2012).
Distinct functional networks within the cerebellum and their re-
lation to cortical systems assessed with independent component
analysis. NeuroImage, 60(4), 2073–2085.
Douw, L., Wakeman, D. G., Tanaka, N., Liu, H., & Stufflebeam,
S. M. (2016). State-dependent variability of dynamic functional
connectivity between frontoparietal and default networks relates
to cognitive flexibility. Neuroscience, 339, 12–21.
Fineberg, N. A., Chamberlain, S. R., Goudriaan, A. E., Stein, D. J.,
Vanderschuren, L. J., Gillan, C. M., . . . Potenza, M. N. (2014).
New developments in human neurocognition: Clinical, genetic,
and brain imaging correlates of impulsivity and compulsivity.
CNS Spectrums, 19(1), 69–89.
Gray, J. A. (1991). Neural systems, emotion, and personality. In J.
Madden IV (Ed.), Neurobiology of learning, emotion, and affect
(pp. 273–306). New York, NY: Raven Press.
Guell, X., Gabrieli, J. D., & Schmahmann, J. D. (2018). Triple repre-
sentation of language, working memory, social and emotion pro-
cessing in the cerebellum: Convergent evidence from task and
seed-based resting-state fMRI analyses in a single large cohort.
NeuroImage, 172, 437–449.
Habas, C., Kamdar, N., Nguyen, D., Prater, K., Beckmann, C. F.,
(2009). Distinct cerebellar con-
Journal of Neuro-
Menon, V., & Greicius, M. D.
tributions to intrinsic connectivity networks.
science, 29(26), 8586–8594.
Hallquist, M. N., & Hillary, F. G. (2018). Graph theory approaches
to functional network organization in brain disorders: A critique
for a brave new small-world. Network Neuroscience, 3(1), 1–26.
Jauregi, A., Kessler, K., & Hassel, S. (2018). Linking cognitive mea-
sures of response inhibition and reward sensitivity to trait impul-
sivity. Frontiers in Psychology, 9, 2306.
Jenkinson, M., Beckmann, C. F., Behrens, T. E., Woolrich, M. W., &
Smith, S. M. (2012). FSL. NeuroImage, 62(2), 782–790.
Jung, Y.-C., Schulte, T., Müller-Oehring, E. M., Namkoong, K.,
Pfefferbaum, A., & Sullivan, E. V. (2014). Compromised fronto-
cerebellar circuitry contributes to nonplanning impulsivity in re-
covering alcoholics. Psychopharmacology, 231(23), 4443–4453.
Karapanagiotidis, T., Vidaurre, D., Quinn, A. J., Vatansever, D.,
Poerio, G. L., Jefferies, E., . . . Smallwood, J.
(2018). Neural
dynamics at rest associated with patterns of ongoing thought.
bioRxiv:454371.
Khadka, S., Narayanan, B., Meda, S., Gelernter, J., Han, S., Sawyer,
B., . . . Pearlson, G. D. (2014). Genetic association of impulsivity
in young adults: A multivariate study. Translational Psychiatry,
4(9), e451.
Kim, J. H., Kim, T. H., Choi, Y. C., Chung, S.-C., & Moon, S. W.
(2013). Impulsive behavior and recurrent major depression asso-
ciated with dandy-walker variant. Psychiatry Investigation, 10(3),
303.
Kipping, J. A., Tuan, T. A., Fortier, M. V., & Qiu, A. (2016). Asyn-
chronous development of cerebellar, cerebello-cortical, and
cortico-cortical functional networks in infancy, childhood, and
adulthood. Cerebral Cortex, 27(11), 5170–5184.
Ledoit, O., & Wolf, M.
(2004). A well-conditioned estimator for
Journal of Multivariate
large-dimensional covariance matrices.
Analysis, 88(2), 365–411.
Leech, R., & Sharp, D. J. (2013). The role of the posterior cingulate
cortex in cognition and disease. Brain, 137(1), 12–32.
Liégeois, R., Li, J., Kong, R., Orban, C., Van De Ville, D., Ge, T., . . .
Yeo, B. T. (2019). Resting brain dynamics at different timescales
capture distinct aspects of human behavior. Nature Communica-
tions, 10(1), 2317.
Lord, L.-D., Stevner, A. B., Deco, G., & Kringelbach, M. L. (2017).
Understanding principles of integration and segregation using
whole-brain computational connectomics: Implications for neu-
ropsychiatric disorders. Philosophical Transactions of the Royal
Society A: Mathematical, Physical and Engineering Sciences,
375(2096), 20160283.
Lurie, D. J., Kessler, D., Bassett, D. S., Betzel, R. F., Breakspear, M.,
Kheilholz, S., . . . Calhoun, V. D.
(2020). Questions and con-
troversies in the study of time-varying functional connectivity in
resting fMRI. Network Neuroscience, 4(1), 30–69.
Marek, S., Siegel, J. S., Gordon, E. M., Raut, R. V., Gratton, C.,
Newbold, D. J., . . . Dosenbach, N. U. (2018). Spatial and tem-
poral organization of the individual human cerebellum. Neuron,
100(4), 977–993.e7.
Mendes, N., Oligschlaeger, S., Lauckner, M. E., Golchert,
J.,
Huntenburg, J. M., Falkiewicz, M., . . . Margulies, D. S. (2019). A
functional connectome phenotyping dataset including cognitive
state and personality measures. Scientific Data, 6, 180307.
Miquel, M., Nicola, S. M., Gil-Miravet, I., Guarque-Chabrera, J., &
Sanchez-Hernandez, A.
(2019). A working hypothesis for the
role of the cerebellum in impulsivity and compulsivity. Frontiers
in Behavioral Neuroscience, 13.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
3
8
9
1
1
8
6
7
4
1
3
n
e
n
_
a
_
0
0
1
4
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network Neuroscience
908
Cerebro-cerebellar functional connectivity and impulsivity
Mitchell, M. R., & Potenza, M. N.
(2014). Recent insights into
the neurobiology of impulsivity. Current Addiction Reports, 1(4),
309–319.
Moreno-Rius, J., & Miquel, M.
(2017). The cerebellum in drug
craving. Drug and Alcohol Dependence, 173, 151–158.
Power, J. D., Barnes, K. A., Snyder, A. Z., Schlaggar, B. L., & Petersen,
S. E. (2012). Spurious but systematic correlations in functional
connectivity MRI networks arise from subject motion. Neuro-
Image, 59(3), 2142–2154.
Quinn, A. J., Vidaurre, D., Abeysuriya, R., Becker, R., Nobre, A. C.,
& Woolrich, M. W. (2018). Task-evoked dynamic network analy-
sis through hidden Markov modeling. Frontiers in Neuroscience,
12, 603.
Ramos-Nuñez, A. I., Fischer-Baum, S., Martin, R. C., Yue, Q., Ye, F.,
& Deem, M. W. (2017). Static and dynamic measures of human
brain connectivity predict complementary aspects of human cog-
nitive performance. Frontiers in Human Neuroscience, 11, 420.
Ryali, S., Supekar, K., Chen, T., Kochalka, J., Cai, W., Nicholas, J.,
. . . Menon, V.
(2016). Temporal dynamics and developmen-
tal maturation of salience, default and central-executive network
interactions revealed by variational Bayes hidden Markov mod-
eling. PLoS Computational Biology, 12(12).
Salman, M. S., Du, Y., Lin, D., Fu, Z., Fedorov, A., Damaraju, E., . . .
Calhoun, V. D. (2019). Group ICA for identifying biomarkers in
schizophrenia: “Adaptive” networks via spatially constrained ICA
show more sensitivity to group differences than spatio-temporal
regression. NeuroImage: Clinical, 22, 101747.
Schmahmann, J. D., & Sherman, J. C. (1997). Cerebellar cognitive
International Review of Neurobiology, 41,
affective syndrome.
433–440.
Schmahmann, J. D., Weilburg, J. B., & Sherman, J. C.
(2007).
The neuropsychiatry of the cerebellum—Insights from the clinic.
Cerebellum, 6(3), 254–267.
Sokolov, A. A., Miall, R. C., & Ivry, R. B. (2017). The cerebellum:
Adaptive prediction for movement and cognition. Trends in Cog-
nitive Sciences, 21(5), 313–332.
Sporns, O.
(2018). Graph theory methods: Applications in brain
networks. Dialogues in Clinical Neuroscience, 20(2), 111.
Stephan, K. E., & Friston, K. J. (2010). Analyzing effective connec-
tivity with functional magnetic resonance imaging. Wiley Inter-
disciplinary Reviews: Cognitive Science, 1(3), 446–459.
Stoodley, C. J. & Schmahmann, J. D.
(2010). Evidence for topo-
graphic organization in the cerebellum of motor control versus
cognitive and affective processing. Cortex, 46(7), 831–844.
Strick, P. L., Dum, R. P., & Fiez, J. A. (2009). Cerebellum and non-
motor function. Annual Review of Neuroscience, 32, 413–434.
Van den Heuvel, M., & Pol, H. (2010). Exploring the brain network:
A review on resting-state fMRI functional connectivity. European
Neuropsychopharmacology, 20, 519–534.
Varoquaux, G., & Craddock, R. C. (2013). Learning and compar-
ing functional connectomes across subjects. NeuroImage, 80,
405–415.
Vidaurre, D., Smith, S. M., & Woolrich, M. W. (2017). Brain net-
work dynamics are hierarchically organized in time. Proceedings
of the National Academy of Sciences, 114(48), 12827–12832.
Voogd, J., & Glickstein, M. (1998). The anatomy of the cerebellum.
Trends in cognitive sciences, 2(9), 307–313.
Wang, C., Kipping, J., Bao, C., Ji, H., & Qiu, A. (2016). Cerebellar
functional parcellation using sparse dictionary learning cluster-
ing. Frontiers in Neuroscience, 10, 188.
Whiteside, S. P., & Lynam, D. R.
(2001). The five factor model
and impulsivity: Using a structural model of personality to under-
stand impulsivity. Personality and Individual Differences, 30(4),
669–689.
Winkler, A. M., Ridgway, G. R., Webster, M. A., Smith, S. M., &
Nichols, T. E. (2014). Permutation inference for the general lin-
ear model. NeuroImage, 92, 381–397.
Yeo, B. T., Krienen, F. M., Sepulcre, J., Sabuncu, M. R., Lashkari,
D., Hollinshead, M., . . . Buckner, R. L. (2011). The organization
of the human cerebral cortex estimated by intrinsic functional
Journal of Neurophysiology, 106(3), 1125–1165.
connectivity.
(PMID: 21653723)
Network Neuroscience
909
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
3
8
9
1
1
8
6
7
4
1
3
n
e
n
_
a
_
0
0
1
4
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3