RESEARCH
Theta-burst TMS to the posterior superior
temporal sulcus decreases resting-state fMRI
connectivity across the face processing network
1, Geena Ianni2, Benjamin Gutierrez1,
Daniel A. Handwerker
Vinai Roopchansingh3, Javier Gonzalez-Castillo 1, Gang Chen4,
1,3, Leslie G. Ungerleider
2, and David Pitcher5
Peter A. Bandettini
a n o p e n a c c e s s
j o u r n a l
Citation: Handwerker, D. A., Ianni, G.,
Gutierrez, B., Roopchansingh, V.,
Gonzalez-Castillo, J., Chen, G.,
Bandettini, P. A., Ungerleider, L. G., &
Pitcher, D. (2020). Theta-burst TMS to
the posterior superior temporal sulcus
decreases resting-state fMRI
connectivity across the face
processing network. Network
Neuroscience, 4(3), 746–760.
https://doi.org/10.1162/netn_a_00145
DOI:
https://doi.org/10.1162/netn_a_00145
Supporting Information:
https://doi.org/10.1162/netn_a_00145
Received: 8 October 2019
Accepted: 8 May 2020
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Authors:
Daniel A. Handwerker
handwerkerd@mail.nih.gov
David Pitcher
david.pitcher@york.ac.uk
Handling Editor:
Lucina Uddin
Copyright: No rights reserved.
This work was authored as part of the
Contributor’s official duties as an
Employee of the United States
Government and is therefore the work
of the United States Government. In
accordance with 17 U.S.C. 105, no
copyright protection is available for
such works under U.S. law.
The MIT Press
1Section on Functional Imaging Methods, Laboratory of Brain and Cognition, National Institute of Mental Health,
Bethesda, MD, USA
2Section on Neurocircuitry, Laboratory of Brain and Cognition, National Institute of Mental Health, Bethesda, MD, USA
3Functional MRI Facility, National Institute of Mental Health, Bethesda, MD, USA
4Scientific and Statistical Computing Core, National Institute of Mental Health, Bethesda, MD, USA
5Department of Psychology, University of York, Heslington, York, UK
Keywords: Resting-state fMRI, Theta-burst TMS (TBS), Face network, Superior temporal sulcus (STS),
Multi-echo fMRI
ABSTRACT
Humans process faces by using a network of face-selective regions distributed across the
brain. Neuropsychological patient studies demonstrate that focal damage to nodes in this
network can impair face recognition, but such patients are rare. We approximated the effects
of damage to the face network in neurologically normal human participants by using theta
burst transcranial magnetic stimulation (TBS). Multi-echo functional magnetic resonance
imaging (fMRI) resting-state data were collected pre- and post-TBS delivery over the
face-selective right superior temporal sulcus (rpSTS), or a control site in the right motor
cortex. Results showed that TBS delivered over the rpSTS reduced resting-state connectivity
across the extended face processing network. This connectivity reduction was observed not
only between the rpSTS and other face-selective areas, but also between nonstimulated
face-selective areas across the ventral, medial, and lateral brain surfaces (e.g., between the
right amygdala and bilateral fusiform face areas and occipital face areas). TBS delivered over
the motor cortex did not produce significant changes in resting-state connectivity across the
face processing network. These results demonstrate that, even without task-induced fMRI
signal changes, disrupting a single node in a brain network can decrease the functional
connectivity between nodes in that network that have not been directly stimulated.
AUTHOR SUMMARY
Human behavior is dependent on brain networks that perform different cognitive functions.
We combined theta burst transcranial magnetic stimulation (TBS) with resting-state fMRI to
study the face processing network. Disruption of the face-selective right posterior superior
temporal sulcus (rpSTS) reduced fMRI connectivity across the face network. This impairment
in connectivity was observed not only between the rpSTS and other face-selective areas, but
also between nonstimulated face-selective areas on the ventral and medial brain surfaces
(e.g., between the right amygdala and bilateral fusiform face areas and occipital face areas).
Thus, combined TBS/fMRI can be used to approximate and measure the effects of focal brain
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
4
3
7
4
6
1
8
6
7
3
3
9
n
e
n
_
a
_
0
0
1
4
5
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
TMS to the STS disrupts the face network
damage on brain networks, and suggests such an approach may be useful for mapping
intrinsic network organization.
INTRODUCTION
The ubiquitous presence of faces in our environment makes them a salient stimulus for studying
the cognitive functions of the human brain. Functional magnetic resonance imaging studies
have identified regions across the brain that exhibit a stronger neural response to faces than
to objects (Gauthier et al., 2000; Kanwisher, McDermott, & Chun, 1997; McCarthy, Puce,
Gore, & Allison, 1997; Phillips et al., 1997; Puce, Allison, Bentin, Gore, & McCarthy, 1998).
These face-selective regions are linked to form distributed nodes of a face processing network
(Calder & Young, 2005; Haxby, Hoffman, & Gobbini, 2000). Neuropsychological patients with
damage to these face-selective areas exhibit face-selective recognition impairments (Barton,
2008; Bouvier & Engel, 2006; Landis, Cummings, Christen, Bogen, & Imhof, 1986; Rezlescu,
Barton, Pitcher, & Duchaine, 2014; Rossion et al., 2003), providing strong evidence that faces
are processed in a specialized network. Despite the importance they have afforded the study of
brain function, such patients are rare. In addition, the interpretation of the data they produce is
limited by individual differences in premorbid ability (Farah, 2004), and compensatory effects
of plasticity that may have occurred after the incident (Robertson & Murre, 1999).
In the present study we used theta burst transcranial magnetic stimulation (TBS) in combi-
nation with multi-echo fMRI acquisition (Posse et al., 1999). Resting-state fMRI was used as a
proxy for the impact of disruption on one node in the face network of neurologically normal ex-
perimental participants. Over two sessions, these participants were scanned using multi-echo
fMRI pre- and post-TBS delivery over the face-selective right superior temporal sulcus (rpSTS),
the stimulation site of interest, or the hand area of the right motor cortex, the control site. Multi-
echo fMRI denoising is a relatively new method that quantitatively identifies and removes non-
blood oxygen-weighted noise from fMRI data (Kundu et al., 2013). Prior to TBS, participants
viewed 3-second videos of moving faces and objects (Pitcher, Dilks, Saxe, Triantafyllou, & Kan-
wisher, 2011) during fMRI scanning to functionally localize face-selective regions of interest
(ROIs).
The face-selective areas in the rpSTS, the fusiform face area (FFA) (Kanwisher et al., 1997;
McCarthy et al., 1997) and the occipital face area (OFA) (Gauthier et al., 2000) comprise the
core nodes of the face processing network (Calder & Young, 2005; Haxby et al., 2000). In
addition to these core nodes, fMRI studies have also identified face-selective voxels in the
amygdala (Phillips et al., 1997). Face processing models propose these areas perform different
cognitive functions (e.g., recognizing identity, facial expression, or eye gaze direction), but
share task-relevant information. The existence of functional connections between face areas
is supported by evidence showing that disruption to one of these areas can impair a range of
face recognition tasks. For example, transcranial magnetic stimulation (TMS) delivered over the
rpSTS disrupts the McGurk effect (Beauchamp, Nath, & Pasalar, 2010) as well as performance
on face processing tasks involving eye gaze (Pourtois et al., 2004), trustworthiness judgments
(Dzhelyova, Ellison, & Atkinson, 2011), and discriminating facial expressions (Pitcher, 2014;
Sliwinska & Pitcher, 2018). Neuropsychological patients with lesions to the rpSTS also show
face processing impairments. A patient with a lesion to the rpSTS and angular gyrus was im-
paired on an unfamiliar face matching task (Sakurai, Hamada, Tsugawa, & Sugimoto, 2016),
while another patient with a lesion to right superior temporal gyrus was impaired at discrimi-
nating gaze detection (Akiyama et al., 2006).
747
Theta burst TMS (TBS):
A type of transcranial magnetic
stimulation where the same
stimulation pattern fluctuates at
around a 5-Hz cycle.
Face network:
A group of brain regions that show
significant activity changes in
response to visual face stimuli. While
these regions have been defined
using univariate analyses with
task-based fMRI, they often
significantly correlate with each
other at rest. In this manuscript, the
following regions were a priori
defined as part of the face network:
posterior superior temporal sulcus
(pSTS), amygdala, fusiform face area
(FFA), and occipital face area (OFA).
Multi-echo fMRI:
During typical fMRI, protons are
excited and there is a delay, the echo
time, before data are collected. That
delay is typically designed to result in
a high contrast for blood oxygenation
differences. In multi-echo fMRI, data
are collected at several echo times
each time protons are excited. This
results in data that have different
levels of contrast for blood
oxygenation differences. This added
information can be used to
empirically decrease noise.
Transcranial magnetic
stimulation (TMS):
A method that induces current in
neural tissue by using a rapidly
changing magnetic field. The pattern
of magnetic field changes can vary.
Network Neuroscience
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
4
3
7
4
6
1
8
6
7
3
3
9
n
e
n
_
a
_
0
0
1
4
5
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
TMS to the STS disrupts the face network
Previous studies have shown TMS-induced face network activation changes during tasks
(Pitcher, Duchaine, & Walsh, 2014; Pitcher, Japee, Rauth, & Ungerleider, 2017). The nodes of
the face network also show functional integration during resting-state fMRI (Li, Song, & Liu,
2019; X. Wang et al., 2016; Zhang, Tian, Liu, Li, & Lee, 2009). In this study, we tested whether
the same face network areas defined in those studies show correlation decreases in response to
TMS to the rpSTS, even in the absence of a face stimulus. If so, then this would build evidence
that behaviorally defined face processing regions are part of a network whose nodes regularly
communicate and interact with each other without this communication being induced by
a specific set of stimuli. Results demonstrated that TBS delivered over the rpSTS caused a
reduction in correlations between the stimulated node and unstimulated nodes of the face
processing network.
RESULTS
For each volunteer, face-selective ROIs were localized using voxels with larger responses to
face videos than to object videos for the left posterior superior temporal sulcus (lpSTS), bilateral
FFA, bilateral OFA, and bilateral amygdala. The right posterior superior temporal sulcus (rpSTS)
ROIs were face-selective voxels in gray matter that were also within an 18-mm-diameter sphere
centered on the stimulation site. This added restriction means the voxels in the rpSTS ROI
were likely to have been directly affected by TBS stimulation. The stimulation site was defined
using the same functional localizer scan, but collected on a preceding session. The right-hand
motor stimulation site was anatomically defined using an anatomical scan from a preceding
session. ROIs for the bilateral hand areas were manually drawn following the gray matter
anatomy of the hand knob in the precentral sulcus. The locations for the ROIs are shown in
Supporting Information Figure S1.
For ROI-based analyses, mean time series of the 10-minute resting-state fMRI data were
calculated using the voxels within each ROI. The correlations between these ROIs were cal-
culated for the resting runs pre- and post-TBS stimulation. The correlation coefficients were
Fisher Z transformed. Since the ROIs are a priori selected from a network where we expect
to see disruption (Pitcher et al., 2014; Pitcher et al., 2017)—with the hand motor regions as
a control—the statistical changes of interest will focus on the ROI analyses. A matrix-based
analysis (MBA) through Bayesian multilevel modeling was used to identify pairs of ROIs where
a decrease in correlation magnitude was larger than expected along with a measure of statis-
tical evidence (Chen et al., 2019). The advantage of this approach is that, instead of adopting
a univariate GLM with the assumption that each ROI pair is an independent entity that shares
no commonality or similarity with its peers, the magnitude estimates and uncertainties of all
ROI pairs are assessed as part of a single integrative model.
Figure 1 shows the effects of stimulation on the correlations pre- and post-TBS for all pairs
of ROIs (bilateral STS, FFA, OFA, amygdala, and hand motor cortices) by using Bayesian mul-
tilevel modeling. For rpSTS stimulation, multiple ROI pairs within the face processing network
showed decreased connectivity that would be likely to be greater than zero with a posterior
probability of at least 0.95 (Figure 1A). For pre- and post-TBS to the motor cortex (Figure 1B)
and interaction effects, there was no strong evidence that any ROI pairs were likely to be
greater than zero. All posterior probabilities were less than 0.85 and all effect sizes were less
than 0.07 (Supporting Information Figure S2).
To help visually compare the effect sizes for rpSTS versus motor stimulation, Figure 2 shows
boxplots for the correlations between the rpSTS and the other ROIs for the runs pre- and post-
TBS. The colored section of the pie charts in this figure show the number of volunteers that had a
Matrix-based analysis (MBA):
A recent approach that uses a
Bayesian multilevel modeling
framework to identify pairs of ROIs
where a decrease in correlation
magnitude was larger than expected
along with a measure of statistical
evidence. With this approach,
correlations between all pairs of
ROIs are assessed as part of a single
model rather than many independent
statistical tests.
Network Neuroscience
748
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
4
3
7
4
6
1
8
6
7
3
3
9
n
e
n
_
a
_
0
0
1
4
5
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
TMS to the STS disrupts the face network
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
Figure 1. Fisher Z transformed correlation magnitude changes from pre- and post-TMS to the (A) rpSTS and (B) right motor cortex. Magnitudes
are the matrix-based analysis (MBA) model fits across the population. Changes that are likely to be greater than 0 with a posterior probability
of at least 95% are full squares and include an *. The squares’ edge lengths decrease linearly to their minimum size for a 90% likelihood or
less. The green line marks the ROI pairs that are within the predefined face-selective network. Supporting Information Figure S2 contains an
unthresholded version of this figure and the interaction effect.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
4
3
7
4
6
1
8
6
7
3
3
9
n
e
n
_
a
_
0
0
1
4
5
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 2. Correlations between the rpSTS ROI and ROIs in a predefined face network as well as the bilateral primary motor hand regions.
These data correspond to the first row of the matrix in Figure 1. The full matrix of bar plots in in Supporting Information Figure S2. Magnitudes
are the MBA model fits across the population for each condition. Boxplots show 25%–75% of the distribution. The white line is the median.
Whiskers are the maximum and minimum values excluding outliers. MBA used to calculate posterior probabilities that a difference is greater
than 0. The correlations between the rpSTS and other face regions consistently decrease post-rpSTS stimulation, but not post-rMotor stimulation.
Network Neuroscience
749
TMS to the STS disrupts the face network
pre- and post-TBS correlation decrease. These pie charts show that 11 to 13 of the 16 volunteers
showed a correlation decrease post-rpSTS stimulation. The Supporting Information includes
boxplots for correlation magnitudes of all ROI pairs (Supporting Information Figures S3 and
S4). Whether the correlation is between a pair of ROIs that includes the directly stimulated
rpSTS or another pair of face network ROIs, the correlations to other ROIs in the face network
decreased across the majority of volunteers when the rpSTS was stimulated.
There is a possibility that TBS to the rpSTS would cause correlation decreases between
the rpSTS and the entire brain. To test for this possibility, all subjects’ data were aligned to
each other, and a group map was calculated with an ANOVA by using multivariate modeling
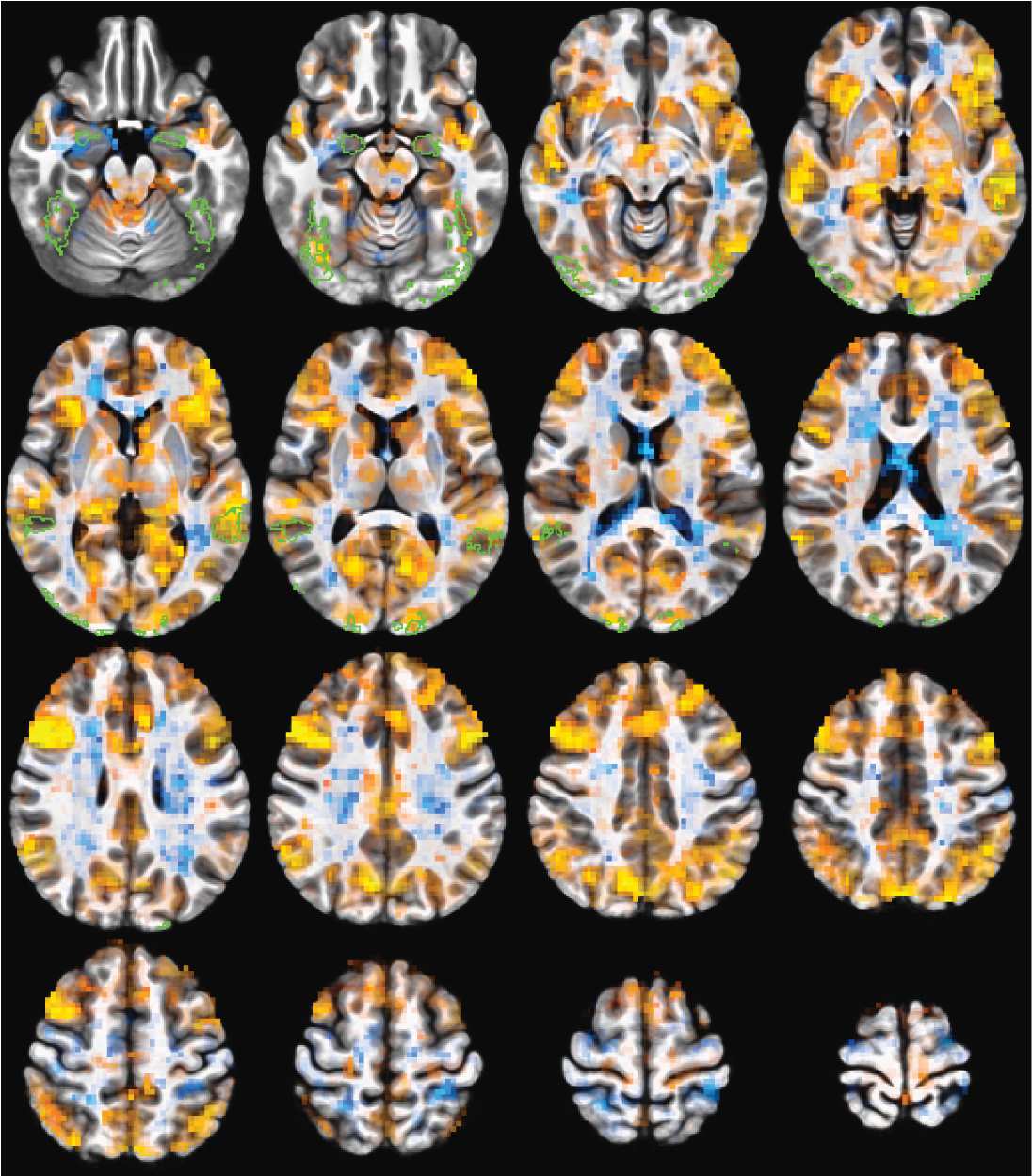
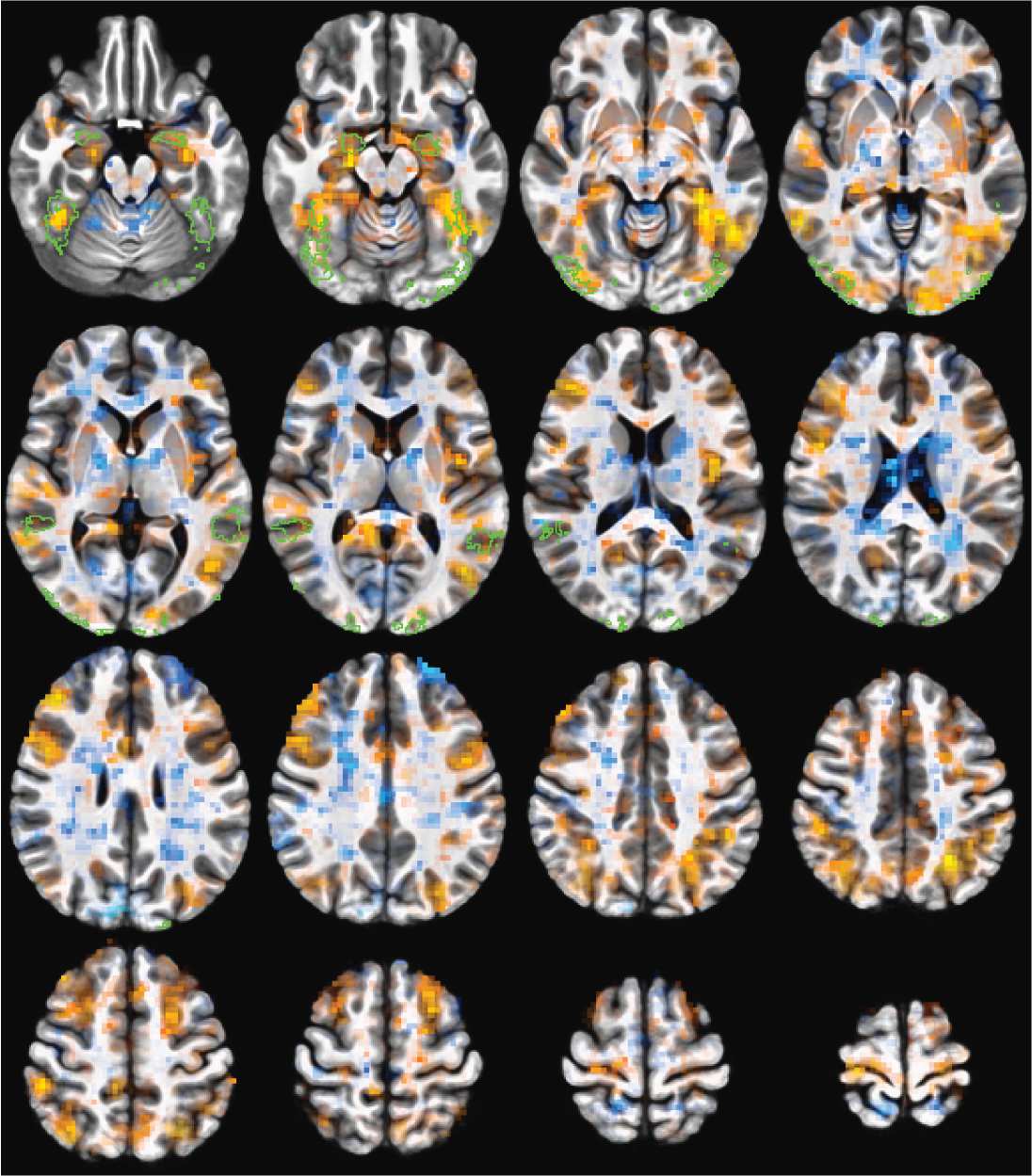
(Figure 3). While there are decreases in activity that do not all cross meaningful statistical
thresholds both within and outside the face network, the correlation decreases post-rpSTS
stimulation are clearly not a global effect.
DISCUSSION
Models of the human face processing network propose that face-selective areas perform dif-
ferent functional roles but share task-relevant information when processing faces (Calder &
Young, 2005; Haxby et al., 2000). In the current study, we approximated the impact of disrup-
tion to the face network by delivering TBS over the face-selective area in the right posterior
STS (rpSTS). The impact of the TBS stimulation was measured with resting-state fMRI, with the
goal of examining whether the relationship between face network regions is altered even in
the absence of a face-selective cognitive task. Our results demonstrated that disruption of the
rpSTS reduced resting-state connectivity, measured by correlated activity, across the nodes of
the face processing network. Figure 1 shows a reduction in connectivity between the rpSTS
and bilateral FFA, OFA, and amygdala as well as between 14 of 21 pairs of ROIs in the face
network that were not directly stimulated. This result demonstrates that disrupting a single face-
selective area causes widespread connectivity decreases across the extended face processing
network.
In addition, functional connectivity between different nodes of the face network has been
shown to correlate with behavioral measures of face recognition. Zhu and colleagues demon-
strated that connectivity between the OFA and FFA correlated with performance on face, but
not object, matching discrimination tasks (Zhu, Zhang, Luo, Dilks, & Liu, 2011). O’Neill and
colleagues demonstrated that connectivity between the FFA and perirhinal cortex correlated
with an upright, but not inverted, face matching task (O’Neil, Hutchison, McLean, & Kohler,
2014). Most recently, Ramot and colleagues demonstrated that face recognition memory was
correlated with connectivity between the face network and regions of the medial temporal
lobe (including the hippocampus) (Ramot, Walsh, & Martin, 2019). Taken together, these find-
ings suggest that synchronized spontaneous neural activity between face regions might be
responsible for the different behavioral aspects of face recognition. The current study builds
on and extends these prior findings to show that transient disruption of a single node in the
face network can disrupt functional connectivity across the wider network.
Figures 2 and Supporting Information Figures S3 and S4 visualize the effect of TBS across
subjects to show that correlations between face network regions have correlation decreases for
the majority of volunteers post-rpSTS stimulation. This widespread pattern of distributed cor-
relation decreases suggests that the nodes of the face network are tightly interconnected, and
that disruption of one node can affect other nodes in the network. Importantly, TBS delivered
over the right motor cortex (which acted as a control site) did not produce above chance reduc-
tions in functional connectivity across the face network. The observed network-wide drop in
Network Neuroscience
750
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
4
3
7
4
6
1
8
6
7
3
3
9
n
e
n
_
a
_
0
0
1
4
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
TMS to the STS disrupts the face network
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
4
3
7
4
6
1
8
6
7
3
3
9
n
e
n
_
a
_
0
0
1
4
5
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. Whole-brain correlation changes (Fisher Z transformed) pre- and post-TBS stimulation.
Voxel coloring shows the magnitude of the correlation difference. Voxels are opaque for p > = 0.01
to transparent for p = 0 The green outlines are voxels in the ROIs of the predefined face network in
at least three volunteers. For TMS to the rpSTS, many of the larger signal drops are within the face
network ROIs. TMS to right motor shows decreases across more of the brain that is not localized to
the face network.
Network Neuroscience
751
TMS to the STS disrupts the face network
resting-state connectivity is consistent with models proposing that the rpSTS is a core compo-
nent of the face processing network (Calder & Young, 2005; Haxby et al., 2000). While we do
show an effect of rpSTS stimulation and not an effect for motor stimulation, we do not observe
a significant effect of motor cortex stimulation between motor ROIs, nor an interaction effect.
Our results do not completely match our predictions, possibly due to the combined effect
of two inherently noisy techniques, namely, resting-state fMRI and TBS. Resting-state fMRI is
known to be more strongly affected than task fMRI by many noise sources (e.g., intersubject
cognitive variation, physiological noise, head motion, scanner instabilities). Also, there is still
not a full understanding of all factors that modulate TBS effect size, including how much spatial
precision matters, whether different brain regions are more sensitive to TBS than others, and
the causes of intersubject variability in TBS sensitivity. Specifically in this study, we note our
poststimulation observation of spatial variation between the hand motor ROI and the hand mo-
tor stimulation site (Supporting Information Figure S1). This variation in stimulation site may
have led to an observed increase in variance of correlation changes post–motor stimulation
versus rpSTS stimulation (Supporting Information Figure S3 and S4). That added variance may
have reduced the posterior probabilities involving motor stimulation data. Although we did
our best to account for known sources of noise, residual artifacts can be expected to exist, and
those may have a multiplicative effect when combining the techniques, further hindering our
statistical power. Additionally, Figure 3 shows large, but not statistically significant, correlation
decreases in response to TBS to the rpSTS. These occur both within and outside the a priori
defined face network regions. While we will not attempt to interpret these below-threshold
correlation decreases, it is important to note that rpSTS function is not limited to face process-
ing. Additionally, the stimulation of an approximately 1-cm3 volume of cortex may affect brain
regions outside of a face network.
In this study, we could not directly test whether TBS to the rpSTS caused alterations in
face processing because we cannot both collect resting-state fMRI and also present face-
selective stimuli during the post-TBS time window when the TBS effect is observable. However,
in previous studies using face-selective stimulus that evoked BOLD measures and the same
TBS stimulation protocol as used here, changes were reported in responses to face stimuli
(Pitcher et al., 2014; Pitcher et al., 2017). The reduction in network connectivity we observed
between nonstimulated face-selective regions on the medial and ventral brain surfaces ex-
tends the findings of this prior work of Pitcher et al. Our previous combined TBS/fMRI studies
examined the impact of disruption in face-selective regions while participants viewed face
videos. These studies showed that TBS delivered over the rpSTS reduced the neural response
to faces in the rpSTS itself, as well as in other face-selective areas including the right fusiform
face area (rFFA) (Pitcher, 2014) and face-selective voxels in the right amygdala (Pitcher et al.,
2017). However, although these studies reported a reduction in the neural response to faces
in remote face-selective areas, neither study was capable of detecting changes in functional
connectivity between face-selective areas. Whereas prior studies demonstrated functional con-
nectivity between the FFA and the amygdala (Fairhall & Ishai, 2007; Herrington, Taylor, Grupe,
Curby, & Schultz, 2011; Vuilleumier, Armony, Driver, & Dolan, 2001, 2003), our study shows
that correlations between these areas are reduced post-TBS delivery over a remote node in the
network, in this case the rpSTS.
Nonhuman primate neuroanatomical studies in macaques report a cortical pathway pro-
jecting from the STS to dorsal subregions of the amygdala (Aggleton, Burton, & Passingham,
1980; Stefanacci & Amaral, 2000, 2002). In addition, neuroimaging and lesion studies of
the amygdala is engaged in facial expression
humans and macaques demonstrate that
Network Neuroscience
752
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
4
3
7
4
6
1
8
6
7
3
3
9
n
e
n
_
a
_
0
0
1
4
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
TMS to the STS disrupts the face network
recognition (Adolphs, Tranel, Damasio, & Damasio, 1994; Adolphs et al., 1999; Calder et al.,
1996; Hadj-Bouziane et al., 2012; Hoffman, Gothard, Schmid, & Logothetis, 2007; Morris
et al., 1996). We previously proposed that humans have a cortical pathway projecting along
the STS into the amygdala that processes changeable facial aspects (e.g., expression), which
was supported using combined TBS/fMRI during face, body, and object viewing (Pitcher et al.,
2017). By replicating this effect using functional connectivity and without a viewing task, we
provide further support for a pathway between these regions.
Neuropsychological patients exhibiting face-selective recognition impairments were essen-
tial to the development of cognitive and brain models of face processing (Bruce & Young, 1986;
Haxby et al., 2000). Studies of patients with right lateral lesions (in the area of the pSTS) show-
ing impairments with eye gaze direction detection (Akiyama et al., 2006; Campbell, Heywood,
Cowey, Regard, & Landis, 1990) and unfamiliar face identity matching (Sakurai et al., 2016)
have been reported. However, prosopagnosic patients with lateral lesions are less common
than those with ventral lesions. Our study further demonstrates that transiently disrupting the
brains of neurologically normal experimental participants with TBS offers an alternative and
safe proxy for modeling the effects of focal cortical disruption on cognitive networks. TBS can
be combined with functional magnetic resonance imaging (fMRI) to study the networks that
process scenes (Mullin & Steeves, 2013), faces (Rafique, Solomon-Harris, & Steeves, 2015;
Solomon-Harris, Rafique, & Steeves, 2016), decision-making (Rahnev, Nee, Riddle, Larson, &
D’Esposito, 2016), and memory (J. X. Wang et al., 2014). Crucially, while we do see some
decreases between a few face network ROIs and the motor ROIs (Figure 1), we examined the
voxel-wise correlations across the brain to show that the largest disruptions of TBS to the rpSTS
were in the face-selective network. Still, the decreases in some connections to motor cortex
ROIs hint at how altering a single node can potentially disrupt multiple distinct networks. Each
brain region does not contribute to only one self-contained network of brain regions.
Neurologists have studied the impact of focal brain lesions in patients for over 200 years.
The study of these patients has been highly influential and has produced insights into human
cognition. However, patients with focal lesions in brain ROIs are extremely rare. In addition,
the interpretation of the data they produce is tempered by factors like individual differences
in premorbid ability (Robertson & Murre, 1999) and the unknown effects of neural plasticity
that may have occurred after their incident (Farah, 2004). TBS combined with resting-state
fMRI has been used to study the default mode network (Abellaneda-Perez et al., 2019; Shang
et al., 2019), attention (Anderkova et al., 2018), visuospatial neglect (Fu et al., 2017), cere-
bellar connectivity (Rastogi et al., 2017), mental illness, (Baeken, Duprat, Wu, De Raedt, &
van Heeringen, 2017), and memory in humans (Mancini et al., 2017). These studies show that
measuring transient TBS-induced disruption with resting-state fMRI offers a systematic experi-
mental methodology for extending more than 200 years of neuropsychological research.
In conclusion, models propose that face-selective brain areas can be linked together to
form the distributed components of a face processing network (Calder & Young, 2005; Haxby
et al., 2000). The current findings support these models by showing that transient disruption
to one of these components, the rpSTS, results in widespread correlation decreases across the
distributed nodes of this network that persists for 10 minutes.
MATERIALS AND METHODS
Participants
Seventeen right-handed participants (11 female, age mean ± SD = 27 ± 5), with no major med-
ical illness, no neurological or psychiatric illness, no serious head injury, no learning disability,
Network Neuroscience
753
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
4
3
7
4
6
1
8
6
7
3
3
9
n
e
n
_
a
_
0
0
1
4
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
TMS to the STS disrupts the face network
no history of drug or alcohol abuse in the past 3 months, no prescription drugs or supplements
affecting brain function, no serious vision or hearing problems and with normal, or corrected-
to-normal, vision gave informed consent as directed by the National Institutes of Health (NIH)
Institutional Review Board (ClinicalTrials.gov identifier: NCT01617408). One participant was
excluded due to high motion (207 of 1,200 volumes in the 4 resting runs censored due to
motion or outliers vs. 0–40 for all other volunteers).
Experimental Design
Participants completed three separate fMRI sessions, each performed on a differ-
Procedure.
ent day. The first session was an fMRI experiment designed to individually localize the TBS
stimulation sites in each participant. Participants viewed face and object videos, and a high-
resolution structural scan was also taken. The data collected in this initial session were used
for TBS target site identification only. During the two subsequent fMRI sessions, participants
were scanned pre- and post-TBS stimulation of either the right posterior STS (rpSTS) or the right
primary motor cortex hand knob (rMotor). Stimulation site order was balanced across partici-
pants. The two TBS sessions were 7–182 days apart (median = 23). All the data presented were
collected during these two TBS sessions.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
4
3
7
4
6
1
8
6
7
3
3
9
n
e
n
_
a
_
0
0
1
4
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Stimulation sites were localized using individual
Stimulation site localizer session and selection.
structural and functional images collected during an fMRI localizer task that each participant
completed prior to the combined TBS/fMRI sessions. For the independent localizer runs used
to identify face-selective ROIs, participants viewed 3-sec video clips of faces, objects, and
scrambled objects. There were 60 movie clips for each category in which distinct exemplars
appeared multiple times. Movies of faces were filmed on a black background, and framed
close-up to reveal only the faces of seven children as they danced or played with toys or
adults (who were out of frame). Fifteen different moving objects were selected that minimized
any suggestion of animacy of the object itself or of a hidden actor pushing the object (these
included mobiles, windup toys, toy planes and tractors, and balls rolling down sloped inclines).
Within each block, stimuli were randomly selected from within the entire set for that stimulus
category. This meant that the same actor or object could appear within the same block. These
stimuli were used in a previous fMRI study of face perception (Pitcher et al., 2011) and previous
combined TBS/fMRI studies (Pitcher et al., 2014; Pitcher et al., 2017).
The rpSTS was identified using a contrast of faces greater than objects. The rpSTS stimulation
target was the peak voxel in the significant cluster in the rpSTS. A T1 weighted anatomical scan
was also collected during this session, and the right motor cortex was the most superior location
on the motor cortex hand knob. A post hoc reexamination identified an instance of in which
the motor stimulation zone may not have directly stimulated the hand motor region in two
volunteers (Supporting Information Figure S1). This may have contributed to a greater variance
of responses to hand motor stimulation across the population, and thus the lower posterior
probabilities for correlation decreases post–motor stimulation and interaction effects between
the stimulation sites.
Participants were scanned using a GE 3-Tesla
Combined fMRI/TBS sessions and data acquisition.
MR 750 scanner at the NIH. fMRI images were acquired using a 32-channel head coil (36
slices, 3 × 3 × 3 mm, FOV = 21.6 cm, grid = 72 × 72, flip angle = 77
, ASSET=3, TR =
2 s, TEs = 14.8, 27.1, & 39.5 ms). In addition, high-resolution MPRAGE anatomical scans
(T1-weighted, 1 × 1 × 1 mm resolution) were acquired to anatomically localize functional
activations and register functional data between sessions.
◦
Network Neuroscience
754
TMS to the STS disrupts the face network
Before each MRI session, the stimulation site (either rpSTS or rMotor) was located using the
Brainsight TMS-MRI coregistration system (Rogue Research), the information was extracted
from the subject-specific localizer scans (see the previous “Stimulation Site Localizer Session
and Selection” section), and the proper coil locations were marked on each participant’s scalp
by using a marker pen. During the MRI session, two 10-min resting-state scans (310 volumes)
were acquired followed by functional localizer scans similar to those used for the stimulation
site localizer session and then an MPRAGE anatomical scan. The volunteers were then removed
from the scanner.
A Magstim Super Rapid Stimulator (Magstim, Whitland, UK) was used to deliver the TBS
via a figure-eight coil with a wing diameter of 70 mm. TBS was delivered at an intensity of
80% of active motor threshold or 30% of machine output (whichever was higher) over each
participant’s functionally localized rpSTS or the right motor hand knob. We used a continuous
TBS paradigm (Huang, Edwards, Rounis, Bhatia, & Rothwell, 2005) of three pulses at 50 Hz
repeated at 200-ms intervals for a 60-sec uninterrupted train of 900 pulses. This is the same
protocol that was used in our previous combined TBS/fMRI studies of the rpSTS (Pitcher et al.,
2014; Pitcher et al., 2017). We matched the protocol to aid with comparisons across studies.
The Stimulator coil handle was held pointing upward and parallel to the midline.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
4
3
7
4
6
1
8
6
7
3
3
9
n
e
n
_
a
_
0
0
1
4
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
As soon as stimulation ended, volunteers were returned to the MRI scanner where the fol-
lowing data were acquired: a brief anatomical reference scan to prescribe a slice placement
that was visually similar to the pre-TBS scan, an ASSET calibration scan, and two 10-min
resting-state scans. The resting scans began 2.5–5 min after the last TBS pulse was delivered
depending on how quickly the participant was able to return to the scanner. After the resting
scan, another MPRAGE anatomical scan was acquired. Three of the 34 MRI sessions did not
include an anatomical scan during the post-TBS part of the session due to time constraints.
The second 10-min resting scans both pre- and post-TBS had more head motion in many
subjects. For 14 of the 64 pre- or post-TBS run pairs, there was at least five more motion
censored volumes in the second run, versus one for first run to second run. Similarly the median
delta motion (frame-to-frame difference) was larger for 49 of the second runs versus first runs.
The average across runs of median delta motion difference between run pairs was 0.01 larger
in the second runs. The average median delta motion was 0.05 so this was a 19% increase in
median motion. There was only a 7% difference in median delta motion for the first and third
runs, which were used in these analyses. While these are subtle differences, the systematic
nature of this difference added a confound that made these data difficult to interpret, so the
second runs were excluded from all analyses.
Data Processing
Data were processed using AFNI (Cox, 1996). For each fMRI run, the first four presteady-state
volumes were removed and data were despiked to remove nonbiological large signal fluctua-
tions and slice time corrected. Spatial alignment transforms for motion correction within runs
and registration across runs were calculated using the middle echo (TE = 27.1 ms) time series.
Nonuniformity of image intensity for the first volume of this time series was corrected using
the bias correction code in SPM8. This reference fMRI volume was aligned to the MPRAGE
anatomical from the same pre- or post-TBS session, if available. Otherwise, it was aligned to
the anatomical scan from the same day. The anatomical scans were aligned to each other and
averaged to make a single, higher quality anatomical volume per volunteer. Registration qual-
ity for each pair of anatomical scans and for each fMRI to anatomical alignment were visually
Network Neuroscience
755
TMS to the STS disrupts the face network
inspected for accuracy. The motion correction and alignment transforms were combined into
a single affine transform matrix and applied to fMRI time series for all echoes so that fMRI data
from each subject was in a single subject-specific space across sessions.
The multi-echo data were then denoised using the MEICA algorithm. MEICA uses the ex-
pected properties of signal changes across multiple echo times to identify and remove signal
fluctuations that are unlikely to reflect the blood oxygenation differences that are central to
fMRI. By doing this, MEICA has been shown to increase the signal-to-noise ratio and remove
data artifacts (Kundu et al., 2013; Kundu, Inati, Evans, Luh, & Bandettini, 2012). The version
of the denoising code that we used is available at: https://github.com/handwerkerd/MEICA_
FaceNetworkTMS. We will refer to the output of the full MEICA algorithm as “denoised” data.
The denoised data will be used for all analyses in this manuscript.
Univariate statistical maps and correlations were calculated within subjects. For functional
localizer runs, time series were intensity normalized by dividing by the mean value in each
run. For resting-state runs, volumes when the first derivative Euclidian norm of the six motion
parameters was greater than 0.2 were censored. Volumes were also censored if more than
10% of the voxels in a volume contained large signal fluctuations that were considered out-
liers. Subject had 0–40 censored volumes, across all four pre- and post-TBS resting runs (1,200
volumes total). Censored volumes per 300 volume run were mean = 2, median = 0, and max-
imum = 22. The motion parameters and their first derivatives were regressed from the time
series. In the same step, signals from a cerebral spinal fluid (CSF) ROI and a white matter ROI
that excluded voxels within eight voxels of either stimulation site were averaged and regressed
from the data. The resting-state data were bandpass filtered (0.01–0.1 Hz) with censoring, and
the data were spatially smoothed with a 5-mm full-width-half-maximum Gaussian kernel.
The statistical tests visualized in Figures 1 and
ROI specifications and cross-subject alignment.
2 used ROIs that were defined for each volunteer. The average of each volunteer’s anatomical
scans, as described in the previous section, were processed in Freesurfer (Fischl et al., 2004) to
estimate volunteer-specific anatomical boundaries for the fusiform gyri, lateral occipital gyri,
and amygdala. Any fMRI voxel that included at least 50% the Freesurfer estimated region
was included in the region. These anatomical regions were intersected with faces>objects
functional activation maps from the localizer scans collected during the TBS sessions (false-
discovery-rate threshold q < 0.05). To make sure ROIs for each brain region were identified in
each volunteer, if 10% of the voxels in the anatomical ROI, or 10 voxels for anatomical ROIs
with less than 100 voxels did not cross this threshold, then the threshold was increased until this
threshold of faces>objects voxels was crossed. In five subjects, this liberal threshold was not
crossed in the amygdala ROIs, which has a mean of 51 voxels in the anatomical ROIs. In four of
these subjects, the same volunteers participated in a separate study that included the identical
functional localizer task with more significant faces>objects voxels in the ROIs (Pitcher et al.,
2017). Those localizer scans were used to define the functional ROIs in those four volunteers.
The voxels in the fusiform gyrus with the face>objects contrast were designated the FFA ROI
and the voxels in the lateral occipital gyrus were designated the OFA ROI. All volunteers had
functionally localized rpSTS and rFFA ROIs with q < 0.05. The threshold was q > 0.05 for one
volunteer in lFFA and rOFA, two volunteers in lpSTS, three volunteers in lOFA, four volunteers
in rAmyg, and eight volunteers in lAmyg. The left- and right-hand knob motor ROIs were hand
drawn on each volunteer’s structural scan by a single person and checked for accuracy by a
second person.
For whole-brain group maps, the within-subject aligned anatomical volumes from each
subject were registered to a standard MNI space and then nonlinearly, iteratively warped to
Network Neuroscience
756
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
4
3
7
4
6
1
8
6
7
3
3
9
n
e
n
_
a
_
0
0
1
4
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
TMS to the STS disrupts the face network
each other to make a within-study anatomical template (Figure 3 and Supporting Information
Figure S1).
ACKNOWLEDGMENTS
We thank Nancy Kanwisher for providing face localizer stimuli. We thank Shruti Japee for
reviewing functional localizer results and for identifying functional localizer scans from these
volunteers that were collected as part of other datasets.
SUPPORTING INFORMATION
Supporting Information for this article is available at https://doi.org/10.1162/netn_a_00145.
ROLE INFORMATION
Daniel A Handwerker: Conceptualization; Data curation; Formal analysis; Investigation; Method-
ology; Project administration; Software; Supervision; Validation; Visualization; Writing – Origi-
nal Draft; Writing – Review & Editing. Geena Ianni: Data curation; Formal analysis;
Investigation; Writing – Review & Editing; Validation. Benjamin Gutierrez: Data curation; For-
mal analysis; Investigation; Software. Vinai Roopchansingh: Methodology; Resources; Soft-
ware. Javier Gonzalez-Castillo: Methodology; Software; Visualization; Writing – Review &
Editing. Gang Chen: Formal analysis; Software; Writing – Review & Editing. Peter A Bandettini:
Conceptualization; Funding acquisition; Methodology; Project administration; Resources;
Supervision; Writing – Review & Editing. Leslie G Ungerleider: Conceptualization; Funding
acquisition; Methodology; Project administration; Resources; Supervision; Writing – Review &
Editing. David Pitcher: Conceptualization; Investigation; Methodology; Project administration;
Resources; Supervision; Writing – Original Draft; Writing – Review & Editing.
FUNDING INFORMATION
Peter A. Bandettini, National Institute of Mental Health (http://dx.doi.org/10.13039/100000025),
Award ID: ZIAMH002783. Leslie G. Ungerleider, National Institute of Mental Health (http://dx.
doi.org/org/10.13039/100000025), Award ID: ZIAMH002918. David Pitcher, Biotechnology
and Biological Sciences Research Council (http://dx.doi.org/10.13039/501100000268), Award
ID: BB/P006981/1.
REFERENCES
Abellaneda-Perez, K., Vaque-Alcazar, L., Vidal-Pineiro, D.,
Jannati, A., Solana, E., Bargallo, N., . . . Bartres-Faz, D. (2019).
Age-related differences in default-mode network connectivity in
response to intermittent theta-burst stimulation and its relation-
ships with maintained cognition and brain integrity in healthy
aging. NeuroImage, 188, 794–806. https://doi.org/10.1016/j.
neuroimage.2018.11.036
Adolphs, R., Tranel, D., Damasio, H., & Damasio, A. (1994). Im-
paired recognition of emotion in facial expressions following
bilateral damage to the human amygdala. Nature, 372(6507),
669–672. https://doi.org/10.1038/372669a0
Adolphs, R., Tranel, D., Hamann, S., Young, A. W., Calder, A. J.,
Phelps, E. A., Damasio, A. R.
(1999). Recognition of facial
emotion in nine individuals with bilateral amygdala damage.
Neuropsychologia, 37(10), 1111–1117. https://doi.org/10.1016/
s0028-3932(99)00039-1
Aggleton, J. P., Burton, M. J., & Passingham, R. E. (1980). Cortical
and subcortical afferents to the amygdala of the rhesus monkey
(Macaca mulatta). Brain Research, 190(2), 347–368. https://doi.
org/10.1016/0006-8993(80)90279-6
Akiyama, T., Kato, M., Muramatsu, T., Saito, F., Umeda, S., &
Kashima, H.
(2006). Gaze but not arrows: A dissociative
impairment after right superior temporal gyrus damage. Neu-
ropsychologia, 44(10), 1804–1810. https://doi.org/10.1016/j.
neuropsychologia.2006.03.007
Anderkova, L., Pizem, D., Klobusiakova, P., Gajdos, M., Koritakova,
E., & Rektorova, I. (2018). Theta burst stimulation enhances con-
nectivity of the dorsal attention network in young healthy sub-
jects: An exploratory study. Neural Plasticity, 2018(3106918).
https://doi.org/10.1155/2018/3106918
Baeken, C., Duprat, R., Wu, G. R., De Raedt, R., & van Heeringen,
(2017). Subgenual anterior cingulate-medial orbitofrontal
K.
Network Neuroscience
757
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
4
3
7
4
6
1
8
6
7
3
3
9
n
e
n
_
a
_
0
0
1
4
5
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
TMS to the STS disrupts the face network
functional connectivity in medication-resistant major depression:
A neurobiological marker for accelerated intermittent theta burst
stimulation treatment? Biological Psychiatry: Cognitive Neuro-
science and Neuroimaging, 2(7), 556–565. https://doi.org/10.
1016/j.bpsc.2017.01.001
Barton, J. J.
(2008). Prosopagnosia associated with a left occipi-
totemporal lesion. Neuropsychologia, 46(8), 2214–2224. https://
doi.org/10.1016/j.neuropsychologia.2008.02.014
Beauchamp, M. S., Nath, A. R., & Pasalar, S. (2010). fMRI-Guided
transcranial magnetic stimulation reveals that the superior tem-
Journal
poral sulcus is a cortical locus of the McGurk effect.
of Neuroscience, 30(7), 2414–2417. https://doi.org/10.1523/
JNEUROSCI.4865-09.2010
Bouvier, S. E., & Engel, S. A. (2006). Behavioral deficits and cortical
damage loci in cerebral achromatopsia. Cerebral Cortex, 16(2),
183–191. https://doi.org/10.1093/cercor/bhi096
Bruce, V., & Young, A.
(1986). Understanding face recognition.
British Journal of Psychology, 77(3), 305–327. https://doi.org/10.
1111/j.2044-8295.1986.tb02199.x
Calder, A. J., & Young, A. W.
(2005). Understanding the recog-
nition of facial identity and facial expression. Nature Reviews
Neuroscience, 6(8), 641–651. https://doi.org/10.1038/nrn1724
Calder, A. J., Young, A. W., Rowland, D., Perrett, D. I., Hodges,
J. R., & Etcoff, N. L.
(1996). Facial emotion recognition after
bilateral amygdala damage: Differentially severe impairment of
fear. Cognitive Neuropsychology, 13(5), 699–745. https://doi.
org/10.1080/026432996381890
Campbell, R., Heywood, C. A., Cowey, A., Regard, M., & Landis,
T. (1990). Sensitivity to eye gaze in prosopagnosic patients and
monkeys with superior temporal sulcus ablation. Neuropsy-
chologia, 28(11), 1123–1142.
https://doi.org/10.1016/0028-
3932(90)90050-x
Chen, G., Burkner, P. C., Taylor, P. A., Li, Z., Yin, L., Glen, D. R., . . .
Pessoa, L. (2019). An integrative Bayesian approach to matrix-
based analysis in neuroimaging. Human Brain Mapping, 40(14),
4072–4090. https://doi.org/10.1002/hbm.24686
Cox, R. W.
(1996). AFNI: Software for analysis and visualization
of functional magnetic resonance neuroimages. Computers and
Biomedical Research, 29(3), 162–173. https://doi.org/10.1006/
cbmr.1996.0014
Dzhelyova, M. P., Ellison, A., & Atkinson, A. P.
(2011). Event-
related repetitive TMS reveals distinct, critical roles for right OFA
and bilateral posterior STS in judging the sex and trustworthiness
of faces. Journal of Cognitive Neuroscience, 23(10), 2782–2796.
https://doi.org/10.1162/jocn.2011.21604
Fairhall, S. L., & Ishai, A. (2007). Effective connectivity within the
distributed cortical network for face perception. Cerebral Cortex,
17(10), 2400–2406. https://doi.org/10.1093/cercor/bhl148
Farah, M. J.
(2004). Visual agnosia (2nd ed.). Cambridge, Mass.:
MIT Press.
Fischl, B., van der Kouwe, A., Destrieux, C., Halgren, E., Segonne,
F., Salat, D. H., . . . Dale, A. M. (2004). Automatically parcellating
the human cerebral cortex. Cerebral Cortex, 14(1), 11–22. Re-
trieved from https://www.ncbi.nlm.nih.gov/pubmed/14654453
Fu, W., Cao, L., Zhang, Y., Huo, S., Du, J., Zhu, L., & Song,
W.
(2017). Continuous theta-burst stimulation may improve
visuospatial neglect via modulating the attention network: A
Topics in Stroke Rehabilita-
randomized controlled study.
tion, 24(4), 236–241. https://doi.org/10.1080/10749357.2016.
1253139
Gauthier, I., Tarr, M. J., Moylan, J., Skudlarski, P., Gore, J. C., &
(2000). The fusiform “face area” is part of
Anderson, A. W.
Journal
a network that processes faces at the individual level.
of Cognitive Neuroscience, 12(3), 495–504. https://doi.org/10.
1162/089892900562165
Hadj-Bouziane, F., Liu, N., Bell, A. H., Gothard, K. M., Luh, W. M.,
Tootell, R. B., . . . Ungerleider, L. G. (2012). Amygdala lesions
disrupt modulation of functional MRI activity evoked by facial
expression in the monkey inferior temporal cortex. Proceed-
ings of the National Academy of Sciences of the United States
of America, 109(52), E3640–3648. https://doi.org/10.1073/pnas.
1218406109
Haxby, J. V., Hoffman, E. A., & Gobbini, M. I. (2000). The distrib-
uted human neural system for face perception. Trends in Cog-
nitive Sciences, 4(6), 223–233. https://doi.org/10.1016/S1364-
6613(00)01482-0
Herrington, J. D., Taylor, J. M., Grupe, D. W., Curby, K. M., &
Schultz, R. T.
(2011). Bidirectional communication between
amygdala and fusiform gyrus during facial recognition. Neuro-
Image, 56(4), 2348–2355. https://doi.org/10.1016/j.neuroimage.
2011.03.072
Hoffman, K. L., Gothard, K. M., Schmid, M. C., & Logothetis, N. K.
(2007). Facial-expression and gaze-selective responses in the
monkey amygdala. Current Biology, 17(9), 766–772. https://
doi.org/10.1016/j.cub.2007.03.040
Huang, Y. Z., Edwards, M. J., Rounis, E., Bhatia, K. P., & Rothwell,
J. C.
(2005). Theta burst stimulation of the human motor cor-
tex. Neuron, 45(2), 201–206. https://doi.org/10.1016/j.neuron.
2004.12.033
Kanwisher, N., McDermott, J., & Chun, M. M. (1997). The fusiform
face area: A module in human extrastriate cortex specialized for
Journal of Neuroscience, 17(11), 4302–4311.
face perception.
https://doi.org/10.1523/JNEUROSCI.17-11-04302.1997
Kundu, P., Brenowitz, N. D., Voon, V., Worbe, Y., Vertes, P. E., Inati,
S. J., . . . Bullmore, E. (2013). Integrated strategy for improving
functional connectivity mapping using multiecho fMRI. Proceed-
ings of the National Academy of Sciences of the United States of
America, 110(40), 16187–16192. https://doi.org/10.1073/pnas.
1301725110
Kundu, P., Inati, S. J., Evans, J. W., Luh, W. M., & Bandettini, P. A.
(2012). Differentiating BOLD and non-BOLD signals in fMRI time
series using multi-echo EPI. NeuroImage, 60(3), 1759–1770.
https://doi.org/10.1016/j.neuroimage.2011.12.028
Landis, T., Cummings, J. L., Christen, L., Bogen, J. E., & Imhof, H. G.
(1986). Are unilateral right posterior cerebral lesions sufficient
to cause prosopagnosia? Clinical and radiological findings in six
additional patients. Cortex, 22(2), 243–252. https://doi.org/10.
1016/s0010-9452(86)80048-x
(2019).
Li, J., Song, Y., & Liu, J.
Functional connectivity pat-
tern in the core face network reflects different mechanisms of
holistic face processing measured by the whole-part effect and
composite-face effect. Neuroscience, 408, 248–258. https://doi.
org/10.1016/j.neuroscience.2019.04.017
Mancini, M., Mastropasqua, C., Bonni, S., Ponzo, V., Cercignani,
M., Conforto, S., . . . Bozzali, M. (2017). Theta burst stimulation
Network Neuroscience
758
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
4
3
7
4
6
1
8
6
7
3
3
9
n
e
n
_
a
_
0
0
1
4
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
TMS to the STS disrupts the face network
of the precuneus modulates resting state connectivity in the left
temporal pole. Brain Topography, 30(3), 312–319. https://doi.
org/10.1007/s10548-017-0559-x
McCarthy, G., Puce, A., Gore, J. C., & Allison, T.
(1997). Face-
specific processing in the human fusiform gyrus. Journal of Cogni-
tive Neuroscience, 9(5), 605–610. https://doi.org/10.1162/jocn.
1997.9.5.605
Morris, J. S., Frith, C. D., Perrett, D. I., Rowland, D., Young, A. W.,
Calder, A. J., & Dolan, R. J. (1996). A differential neural response
in the human amygdala to fearful and happy facial expressions.
Nature, 383(6603), 812–815. https://doi.org/10.1038/383812a0
Mullin, C. R., & Steeves, J. K. (2013). Consecutive TMS-fMRI reveals
an inverse relationship in BOLD signal between object and scene
Journal of Neuroscience, 33(49), 19243–19249.
processing.
https://doi.org/10.1523/JNEUROSCI.2537-13.2013
O’Neil, E. B., Hutchison, R. M., McLean, D. A., & Kohler, S. (2014).
Resting-state fMRI reveals functional connectivity between face-
selective perirhinal cortex and the fusiform face area related to
face inversion. NeuroImage, 92, 349–355. https://doi.org/10.
1016/j.neuroimage.2014.02.005
Phillips, M. L., Young, A. W., Senior, C., Brammer, M., Andrew, C.,
Calder, A. J., . . . David, A. S. (1997). A specific neural substrate
for perceiving facial expressions of disgust. Nature, 389(6650),
495–498. https://doi.org/10.1038/39051
Pitcher, D. (2014). Facial expression recognition takes longer in the
posterior superior temporal sulcus than in the occipital face area.
Journal of Neuroscience, 34(27), 9173–9177. https://doi.org/10.
1523/JNEUROSCI.5038-13.2014
Pitcher, D., Dilks, D. D., Saxe, R. R., Triantafyllou, C., & Kanwisher,
N.
(2011). Differential selectivity for dynamic versus static in-
formation in face-selective cortical regions. NeuroImage, 56(4),
2356–2363. https://doi.org/10.1016/j.neuroimage.2011.03.067
Pitcher, D., Duchaine, B., & Walsh, V. (2014). Combined TMS and
FMRI reveal dissociable cortical pathways for dynamic and static
face perception. Current Biology, 24(17), 2066–2070. https://
doi.org/10.1016/j.cub.2014.07.060
Pitcher, D., Japee, S., Rauth, L., & Ungerleider, L. G.
(2017). The
superior temporal sulcus is causally connected to the amyg-
Journal of Neuroscience,
dala: A combined TBS-fMRI study.
37(5), 1156–1161. https://doi.org/10.1523/JNEUROSCI.0114-
16.2016
Posse, S., Wiese, S., Gembris, D., Mathiak, K., Kessler, C., Grosse-
Ruyken, M. L., . . . Kiselev, V. G.
(1999). Enhancement of
BOLD-contrast sensitivity by single-shot multi-echo functional
MR imaging. Magnetic Resonance in Medicine, 42(1), 87–97.
https://doi.org/10.1002/(sici)1522-2594(199907)42:1<87::aid-
mrm13>3.0.co;2-o
Pourtois, G., Sander, D., Andres, M., Grandjean, D., Reveret, L.,
(2004). Dissociable roles of the
Olivier, E., & Vuilleumier, P.
human somatosensory and superior temporal cortices for pro-
cessing social face signals. European Journal of Neuroscience,
20(12), 3507–3515. https://doi.org/10.1111/j.1460-9568.2004.
03794.x
Puce, A., Allison, T., Bentin, S., Gore, J. C., & McCarthy, G. (1998).
Temporal cortex activation in humans viewing eye and mouth
movements. Journal of Neuroscience, 18(6), 2188–2199. https://
doi.org/10.1523/JNEUROSCI.18-06-02188.1998
Rafique, S. A., Solomon-Harris, L. M., & Steeves, J. K.
(2015).
TMS to object cortex affects both object and scene remote net-
works while TMS to scene cortex only affects scene networks.
Neuropsychologia, 79(Pt A), 86–96. https://doi.org/10.1016/j.
neuropsychologia.2015.10.027
Rahnev, D., Nee, D. E., Riddle, J., Larson, A. S., & D’Esposito, M.
(2016). Causal evidence for frontal cortex organization for per-
ceptual decision making. Proceedings of the National Academy
of Sciences of the United States of America, 113(21), 6059–6064.
https://doi.org/10.1073/pnas.1522551113
Ramot, M., Walsh, C., & Martin, A. (2019). Multifaceted integra-
tion: Memory for faces is subserved by widespread connections
between visual, memory, auditory, and social networks. Journal
of Neuroscience, 39(25), 4976–4985. https://doi.org/10.1523/
JNEUROSCI.0217-19.2019
Rastogi, A., Cash, R., Dunlop, K., Vesia, M., Kucyi, A., Ghahremani,
A., Chen, R. (2017). Modulation of cognitive cerebello-cerebral
functional connectivity by lateral cerebellar continuous theta
burst stimulation. NeuroImage, 158, 48–57. https://doi.org/10.
1016/j.neuroimage.2017.06.048
Rezlescu, C., Barton, J. J., Pitcher, D., & Duchaine, B. (2014). Nor-
mal acquisition of expertise with greebles in two cases of ac-
quired prosopagnosia. Proceedings of the National Academy of
Sciences of the United States of America, 111(14), 5123–5128.
https://doi.org/10.1073/pnas.1317125111
Robertson, I. H., & Murre, J. M.
(1999). Rehabilitation of brain
damage: Brain plasticity and principles of guided recovery. Psy-
chological Bulletin, 125(5), 544–575. https://doi.org/10.1037/
0033-2909.125.5.544
Rossion, B., Caldara, R., Seghier, M., Schuller, A. M., Lazeyras, F., &
Mayer, E. (2003). A network of occipito-temporal face-sensitive
areas besides the right middle fusiform gyrus is necessary for nor-
mal face processing. Brain, 126(Pt 11), 2381–2395. https://doi.
org/10.1093/brain/awg241
Sakurai, Y., Hamada, K., Tsugawa, N., & Sugimoto, I. (2016). Ven-
tral simultanagnosia and prosopagnosia for unfamiliar faces due
to a right posterior superior temporal sulcus and angular gyrus
lesion. Neurocase, 22(1), 122–129. https://doi.org/10.1080/
13554794.2015.1066827
Shang, Y., Chang, D., Zhang, J., Peng, W., Song, D., Gao, X., &
Wang, Z. (2019). Theta-burst transcranial magnetic stimulation
induced functional connectivity changes between dorsolateral
prefrontal cortex and default-mode-network. Brain Imaging and
Behavior. https://doi.org/10.1007/s11682-019-00139-y
Sliwinska, M. W., & Pitcher, D. (2018). TMS demonstrates that both
right and left superior temporal sulci are important for facial ex-
pression recognition. NeuroImage, 183, 394–400. https://doi.
org/10.1016/j.neuroimage.2018.08.025
Solomon-Harris, L. M., Rafique, S. A., & Steeves, J. K. (2016). Con-
secutive TMS-fMRI reveals remote effects of neural noise to the
“occipital face area.” Brain Research, 1650, 134–141. https://
doi.org/10.1016/j.brainres.2016.08.043
Stefanacci, L., & Amaral, D. G. (2000). Topographic organization
of cortical inputs to the lateral nucleus of the macaque monkey
Journal of Comparative
amygdala: A retrograde tracing study.
Neurology, 421(1), 52–79. https://doi.org/10.1002/(sici)1096-
9861(20000522)421:1<52::aid-cne4>3.0.co;2-o
Network Neuroscience
759
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
4
3
7
4
6
1
8
6
7
3
3
9
n
e
n
_
a
_
0
0
1
4
5
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
TMS to the STS disrupts the face network
Stefanacci, L., & Amaral, D. G.
(2002). Some observations on
cortical inputs to the macaque monkey amygdala: An antero-
grade tracing study. Journal of Comparative Neurology, 451(4),
301–323. https://doi.org/10.1002/cne.10339
Vuilleumier, P., Armony, J. L., Driver, J., & Dolan, R. J. (2001). Effects
of attention and emotion on face processing in the human brain:
An event-related fMRI study. Neuron, 30(3), 829–841. https://
doi.org/10.1016/S0896-6273(01)00328-2
Vuilleumier, P., Armony, J. L., Driver, J., & Dolan, R. J. (2003). Dis-
tinct spatial frequency sensitivities for processing faces and emo-
tional expressions. Nature Neuroscience, 6(6), 624–631. https://
doi.org/10.1038/nn1057
Wang, J. X., Rogers, L. M., Gross, E. Z., Ryals, A. J., Dokucu, M. E.,
Brandstatt, K. L., . . . Voss, J. L. (2014). Targeted enhancement
of cortical-hippocampal brain networks and associative mem-
ory. Science, 345(6200), 1054–1057. https://doi.org/10.1126/
science.1252900
Wang, X., Zhen, Z., Song, Y., Huang, L., Kong, X., & Liu, J. (2016).
The hierarchical structure of the face network revealed by its
functional connectivity pattern. Journal of Neuroscience, 36(3),
890–900. https://doi.org/10.1523/JNEUROSCI.2789-15.2016
Zhang, H., Tian, J., Liu, J., Li, J., & Lee, K. (2009). Intrinsically orga-
nized network for face perception during the resting state. Neu-
roscience Letters, 454(1), 1–5. https://doi.org/10.1016/j.neulet.
2009.02.054
Zhu, Q., Zhang, J., Luo, Y. L., Dilks, D. D., & Liu, J. (2011). Resting-
state neural activity across face-selective cortical regions is behav-
iorally relevant. Journal of Neuroscience, 31(28), 10323–10330.
https://doi.org/10.1523/JNEUROSCI.0873-11.2011
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
4
3
7
4
6
1
8
6
7
3
3
9
n
e
n
_
a
_
0
0
1
4
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network Neuroscience
760