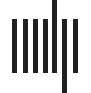RESEARCH
Brain connectivity during Alzheimer’s disease
progression and its cognitive impact
in a transgenic rat model
Emma Muñoz-Moreno 1,2, Raúl Tudela1,3, Xavier López-Gil1, and Guadalupe Soria1,3
1Experimental 7T MRI Unit, Institut d’Investigacions Bimediques August Pi i Sunyer (IDIBAPS), Barcelona, Spain
2Magnetic Resonance Image Core Facility, Institut d’Investigacions Biomdiques August Pi i Sunyer (IDIBAPS), Barcelona, Spain
3Consorcio Centro de Investigacin Biomdica en Red (CIBER) de Bioingeniera, Biomateriales y Nanomedicina (CIBER-BBN),
Group of Biomedical Imaging, University of Barcelona, Barcelona, Spain
a n o p e n a c c e s s
j o u r n a l
Keywords: Alzheimer’s disease continuum, Animal models, TgF344-AD, Connectomics, Brain
networks, Longitudinal evolution
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
4
2
3
9
7
1
9
6
6
9
0
9
n
e
n
_
a
_
0
0
1
2
6
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
ABSTRACT
The research of Alzheimer’s disease (AD) in its early stages and its progression till
symptomatic onset is essential to understand the pathology and investigate new treatments.
Animal models provide a helpful approach to this research, since they allow for controlled
follow-up during the disease evolution. In this work, transgenic TgF344-AD rats were
longitudinally evaluated starting at 6 months of age. Every 3 months, cognitive abilities
were assessed by a memory-related task and magnetic resonance imaging (MRI) was
acquired. Structural and functional brain networks were estimated and characterized by
graph metrics to identify differences between the groups in connectivity, its evolution with
age, and its influence on cognition. Structural networks of transgenic animals were altered
since the earliest stage. Likewise, aging significantly affected network metrics in TgF344-AD,
but not in the control group. In addition, while the structural brain network influenced
cognitive outcome in transgenic animals, functional network impacted how control subjects
performed. TgF344-AD brain network alterations were present from very early stages,
difficult to identify in clinical research. Likewise, the characterization of aging in these
animals, involving structural network reorganization and its effects on cognition, opens a
window to evaluate new treatments for the disease.
AUTHOR SUMMARY
We have applied magnetic resonance image–based connectomics to characterize
TgF344-AD rats, a transgenic model of Alzheimer’s disease (AD). This represents a highly
translational approach, what is essential to investigate potential treatments. TgF344-AD
animals were evaluated from early to advanced ages to describe alterations in brain
connectivity and how brain networks are affected by age. Results showed that aging had a
bigger impact in the structural connectivity of the TgF344-AD than in control animals,
and that changes in the structural network, already observed at early ages, significantly
influenced cognitive outcome of transgenic animals. Alterations in connectivity were similar
to that described in AD human studies and complement them by providing insights into
earlier stages and a plot of AD effects throughout the whole life span.
Citation: Muñoz-Moreno, E., Tudela, R.,
López-Gil, X., & Soria, G. (2020). Brain
connectivity during Alzheimer’s
disease progression and its cognitive
impact in a transgenic rat model.
Network Neuroscience, 4(2), 397–415.
https://doi.org/10.1162/netn_a_00126
DOI:
https://doi.org/10.1162/netn_a_00126
Supporting Information:
https://doi.org/10.1162/netn_a_00126
Received: 9 July 2019
Accepted: 10 January 2020
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Author:
Emma Muñoz-Moreno
emunozm@clinic.cat
Handling Editor:
Daniele Marinazzo
Copyright: © 2020
Massachusetts Institute of Technology
Published under a Creative Commons
Attribution 4.0 International
(CC BY 4.0) license
The MIT Press
Brain connectivity and cognition in a rat model of Alzheimer’s disease
INTRODUCTION
Alzheimer’s disease (AD) is a neurodegenerative disease related to most cases of dementia
in the elderly population. Brain damage associated with AD starts decades before the symp-
tomatic onset and clinical diagnosis, which has led to consideration of AD as a continuum
(Dubois et al., 2016; Jack et al., 2018). This fact makes the research of disease progression from
very early stages a key point to understand AD and develop potential pharmacological treat-
ments or other types of interventions. Consequently, it is essential that the cohorts of the at-risk
population be evaluated before the diagnosis stage and that this population be followed up on
during the years until AD symptoms appear. Recently, studies performed on AD risk population
such as carriers of apolipoprotein E (APOE)-ε4 or rs405509 alleles have detected brain differ-
ences between these subjects and control subjects in elderly (Chen et al., 2015; Reiter et al.,
2017; Shu et al., 2015) and middle-age population (Cacciaglia et al., 2018; Habib et al., 2017;
Mak et al., 2017; ten Kate et al., 2016). However, the identification of at-risk cohorts is chal-
lenging and the time required to follow up on them from middle age to the eventual advanced
phase of AD hinders the characterization of the disease progression in patient cohorts. In this
sense, animal models provide a helpful approach to evaluate the development of AD from
early to advanced stages (Do Carmo & Cuello, 2013; Galeano et al., 2014; Leon et al., 2010;
Sabbagh, Kinney, & Cummings, 2013). These models allow the study of earlier stages of the
disease, as well as the follow-up of the same subjects during the whole extent of the disease
in a relatively short period. TgF344-AD rats are an example of AD animal models. They pro-
gressively manifest most pathological hallmarks of the disease including amyloid plaques, tau
pathology, oligomeric amyloid β (Aβ), neuronal loss, and behavioral impairment (Cohen et al.,
2013; Drummond & Wisniewski, 2017). Therefore, it is a very suitable model to evaluate AD
progression during aging.
Along with the choice of proper animal models, the use of replicable techniques in ex-
perimental and clinical research can improve translationality (Sabbagh et al., 2013). This is
a crucial point given the high failure rates in the translation between preclinical and clinical
trials reported in drug research for AD (Drummond & Wisniewski, 2017; Windisch, 2014).
Neuroimaging techniques have been extensively used to identify alterations associated with
the disease in a noninvasive way and can be applied in both animal and human cohorts
(Sabbagh et al., 2013). In the search for AD biomarkers, magnetic resonance imaging (MRI)
has represented a helpful technique to characterize in vivo brain changes during AD progres-
sion (Jack et al., 2015), such as atrophy (Frisoni, Fox, Jack, Scheltens, & Thompson, 2010)
or tissue changes (Weston, Simpson, Ryan, Ourselin, & Fox, 2015). In addition, MRI can be
used to identify and describe structural and functional brain networks. For this reason it has
been used to investigate and support the hypothesis of AD as a disconnection syndrome (Brier
et al., 2014; Gomez-Ramirez & Wu, 2014; Palesi et al., 2016; Xie & He, 2012), suggesting that
cognitive decline in AD is related to functional or structural disconnection between regions
rather than localized changes in specific isolated brain areas. Thus, impairment in structural
connectivity associated with AD has been described based on gray matter patterns evaluated in
structural MRI or, mainly, based on the fiber tract estimations obtained from diffusion-weighted
MRI (Daianu et al., 2013; Fischer, Wolf, Scheurich, & Fellgiebel, 2015; Lo et al., 2010; Wee
et al., 2011). Likewise, functional disconnection has been evaluated using resting-state func-
tional MRI (rs-fMRI) and graph theory to estimate and quantify functional networks (Brier et al.,
2014; Sanz-Arigita et al., 2010; Supekar, Menon, Rubin, Musen, & Greicius, 2008); to iden-
tify resting-state networks using independent component analysis (Badhwar et al., 2017); or
to characterize connectivity between a specific region and the rest of the brain (Gour et al.,
2014). Indeed, alterations in structural and functional brain network properties have been
TgF344-AD rats:
A rat whose DNA is modified to
express genes associated with
Alzheimer’s disease (AD). It shows
age-dependent pathological AD
hallmarks.
Disconnection syndrome:
A syndrome related to alterations in
the connectivity between regions
rather than in specific areas.
Network Neuroscience
398
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
4
2
3
9
7
1
9
6
6
9
0
9
n
e
n
_
a
_
0
0
1
2
6
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Brain connectivity and cognition in a rat model of Alzheimer’s disease
described in TgF344-AD animals at early ages (5–6 months; Muñoz-Moreno, Tudela, López-
Gil, & Soria, 2018) as well as differences in functional connectivity of specific regions or
networks at several time points from 6 to 18 months (Anckaerts et al., 2019; Tudela, Muñoz-
Moreno, Sala-Llonch, López-Gil, & Soria, 2019).
Among the MRI-based studies of AD as a disconnection syndrome, graph theory metrics
have become one of the most applied methods to investigate the organization at a global
level of both structural (Daianu et al., 2013; Fischer et al., 2015; Lo et al., 2010; Muñoz-
Moreno et al., 2018; Wee et al., 2011) and functional (Brier et al., 2014; Muñoz-Moreno et al.,
2018; Sanz-Arigita et al., 2010; Supekar et al., 2008) brain networks. Graph metrics provide a
quantitative description of different aspects of the network such as integration, segregation,
or strength, and they allow performance of similar and comparable analyses in structural and
functional networks. Since global graph metrics quantify the whole brain structure, they are
more sensitive to global reorganization of the networks rather than isolated alterations in indi-
vidual connections (Rubinov & Sporns, 2010).
Therefore, in the present study, we use graph theory to investigate how the disease progres-
sion affects both structural and functional brain networks and its effects in cognitive abilities.
In this line, we evaluated how the connectivity impairments observed in young TgF344-AD
animals in our previous work (Muñoz-Moreno et al., 2018) evolve during aging. Structural
and functional MRI acquisitions were acquired every 3 months from 6 to 18 months of age
in a cohort of TgF344-AD and control rats to perform a longitudinal analysis of brain connec-
tivity. Cognitive skills were also evaluated every 3 months to test the impact of connectivity
alterations in cognition. Hence, this work aims to contribute to the understanding of AD pro-
gression and its association with cognitive decline from the perspective of the disease as a
disconnection syndrome.
MATERIALS AND METHODS
Subjects
The experiments were performed in a cohort of 18 male Fisher rats including TgF344-AD
animals (Cohen et al., 2013) and their wild-type littermates, which were evaluated at five time
points. Table 1 shows the details on the resulting sample size and average age per time point
after MRI experiments.
The animals were housed in cages under controlled temperature (21 ± 1 °C) and humid-
ity (55 ± 10%) with a 12-hr light/12-hr dark cycle. Food and water were available ad libi-
tum during the entire experiment, except during behavioral test periods as explained below.
At 2 months of age, animals start a cognitive training to perform delay nonmatched sample
Sample size and age (median ± interquartile range) at each of the five acquisitions. Sample
Table 1.
size in time point 1 is different in the structural and functional analysis (* corresponds to the number
of rs-fMRI acquisitions). Variability in the age of acquisition at the first time point is due to differences
in the cognitive training duration. Age difference between the groups was not significant.
Control
TgF344-AD
Time point 1
Time point 2
Time point 3
Time point 4
Time point 5
N
8/5*
9
7
8
8
Age (months)
5.33 ± 0.3
9.2 ± 0.5
11.3 ± 0.22
14.9 ± 0.425
17.92 ± 0.52
N
8/7*
8
9
9
6
Age (months)
6.33 ± 1.22
8.67 ± 0.2
11.3 ± 0.03
14.87 ± 0.03
18.03 ± 0.62
399
Graph metrics:
A set of measures that allow network
characterization such as efficiency,
connectivity strength, or hierarchical
organization.
Network Neuroscience
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
4
2
3
9
7
1
9
6
6
9
0
9
n
e
n
_
a
_
0
0
1
2
6
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Brain connectivity and cognition in a rat model of Alzheimer’s disease
Delay nonmatched sample
(DNMS) test:
Cognitive test to evaluate working
memory. Animals must press levers in
a given sequence to obtain a reward.
(DNMS) task. Once the learning criteria were achieved, their performance in DNMS was eval-
uated and the first MRI scan was acquired. Afterwards, 2 weeks of DNMS sessions followed
by MRI acquisition were repeated every 3 months, resulting in five evaluated time points.
Cognitive Function Evaluation
Every 3 months, working memory was evaluated by DNMS test, following the procedure de-
scribed in Muñoz-Moreno et al. (2018). DNMS was carried out in isolated operant chambers
(Med Associates, USA), equipped with a pellet dispenser and three retractable levers, two of
them in the wall where the pellet is (right and left levers) and the other in the opposite side
(center lever). During the testing weeks, rats were food-deprived, receiving 75% of the usual
food intake. In brief, DNMS requires the animal to press the levers following a specific se-
quence to obtain a pellet. First, in the sample phase, right or left lever appeared and after the
animal pressed it, a delay randomly timed between 1 and 30 s started, after which the center
lever appeared. When the animal pressed it, both right and left levers were extended again.
Correct response required a press on the lever opposite to lever that was presented in the sam-
ple phase. Each DNMS session finished after 90 min or when 90 trials were completed. The
number of trials and percentage of correct responses were recorded.
Before the first test, animals underwent a habituation and training phase to acquire the
required skills. This phase started when the animals were 2 months old and finished when they
achieved acquisition criteria as explained in Muñoz-Moreno et al. (2018). After that, animals
underwent 15 DNMS sessions (five sessions per week) to evaluate and consolidate the learned
task. At each of the following four time points, animals performed 10 DNMS sessions (2 weeks).
Magnetic Resonance Imaging
MRI acquisitions were performed on a 7.0T BioSpec 70/30 horizontal animal scanner (Bruker
BioSpin, Ettlingen, Germany). Animals were placed in the supine position in a Plexiglas holder
with a nose cone for administering anesthetic gases (1.5% isoflurane in a mixture of 30% O2
and 70% CO) and were fixed using tooth and ear bars and adhesive tape. The rat received
a 0.5 ml bolus of medetomidine (0.05 mg/kg; s.c.), and a catheter was implanted in its back
for continuous perfusion of medetomidine. Isoflurane was gradually decreased until 0%, and
15 min after the bolus the medetomidine perfusion (0.05 mg/kg; s.c.) started at rate 1 ml/hr.
The acquisition protocol included the following:
T2-weighted images, acquired using a RARE sequence with effective echo time TE =
35.3 ms, repetition time TR = 6,000 ms and RARE factor = 8, voxel size = 0.12 ×
0.12 mm2, 40 slices, slice thickness = 0.8 mm, and field of view FoV = 30 × 30 ×
32 mm3.
T1-weighted images, acquired using an MDEFT protocol with TE = 2 ms, TR = 4,000 ms,
voxel size = 0.14 × 0.14 × 0.5 mm3, and FoV= 35 × 35 × 18 mm3.
Diffusion-weighted images (DWI) using a spin-echo EPI sequence with TE = 24.86 ms,
TR = 15,000 ms, four segments, 60 gradient directions with b-value = 1,000 s/mm2,
and five volumes with b-value = 0 s/mm2; voxel size = 0.31 × 0.31 × 0.31 mm3 and
FoV = 22.23 × 22.23 × 18.54 mm3.
rs-fMRI using a gradient echo T2* acquisition, with the following parameters: TE =
10.75 ms, TR = 2,000 ms, 600 volumes (20 min), voxel size = 0.4 × 0.4 × 0.6 mm3,
FoV = 25.6 × 25.6 × 20.4 mm3. rs-fMRI acquisition is scheduled after anatomical
and diffusion MRI to ensure that isoflurane dose has been removed and the animals are
sedated only by medetomidine.
Network Neuroscience
400
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
4
2
3
9
7
1
9
6
6
9
0
9
n
e
n
_
a
_
0
0
1
2
6
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Brain connectivity and cognition in a rat model of Alzheimer’s disease
To illustrate the acquisition quality, Supplementary Figure 1 shows a series of slices of
colored fractional anisotropy computed in a randomly selected subject of the cohort; and
Supplementary Figure 2 displays a selection of functional networks extracted from the whole
cohort using independent component analysis (ICA).
Image Processing and Connectome Definition
Structural connectome:
A matrix that describes connectivity
between brain regions based on
anatomical (fiber tract) connections.
Functional connectome:
A matrix that describes connectivity
between brain regions based on the
correlation between regional
activation patterns.
The acquired images were processed to obtain the structural and functional connectomes fol-
lowing the methodology described in Muñoz-Moreno et al. (2018). Briefly, a rat brain atlas was
registered to the T2-weighted images to obtain brain masks and region parcellations (Schwarz
et al., 2006). T1-weighted images were used to segment the brain into white matter (WM), gray
matter (GM), and cerebrospinal fluid (CSF) based on tissue probability maps registered from
an atlas (Valdés-Hernández et al., 2011) to each subject brain. Parcellation and segmentation
were registered from T2/T1-weighted volumes to DWI and rs-fMRI spaces to define the regions
between which connectivity was assessed.
Fiber tract trajectories were estimated from DWI volumes using deterministic tractography
based on constrained spherical deconvolution model, considering WM voxels as seed points.
Dipy was used to process DWI volumes and estimate the fiber tracts (Garyfallidis et al., 2014).
The resulting number of streamlines generated per subject are of the order of 105 (6.18·105±
0.67·105). As defined in Muñoz-Moreno et al. (2018), the structural connectome included 76
regions. Connection between two regions was defined if at least one streamline started in one
region and ended in the other. The resulting structural connectomes have an average density
of 62.22 ± 2.78. Three connectomes were considered according to the connection weight
definition:
Fractional anisotropy (FA) weighted connectome (FA-w): The connection weight between
two regions is defined as the average FA in the streamlines connecting them.
Fiber density (FD) weighted connectome (FD-w): The connection weight between two
regions is computed as the number of streamlines normalized by the region volumes and
the streamline length (Muñoz-Moreno et al., 2018).
Structural binary connectome: Connection weight is 1 between connected regions and
0 otherwise.
Resting-state fMRI was processed to obtain the average time series in the GM voxels of each
of the regions of interest. Preprocessing includes slice timing, motion correction by spatial
realignment using SPM8, and correction of EPI distortion by elastic registration to the T2-
weighted volume using ANTs (Avants, Epstein, Grossman, & Gee, 2008). Afterwards, NiTime
(http://nipy.org/nitime/) was used for z-score normalization and detrending of the time series,
smoothing with an FWHM (full width at half maximum) of 1.2 mm, frequency filtering between
0.01 and 0.1 Hz, and regression by motion parameters and WM and CSF average signals. Since
brain activity identified by rs-fMRI has been constrained to GM (Power, Plitt, Laumann, &
Martin, 2017), only regions comprising GM tissue were considered as nodes in the functional
connectome (54 regions). Both weighted and binary functional connectomes were defined.
The connection weight was the partial correlation between the pair of regional time series,
transformed by Fisher’s z-transformation. Negative correlation coefficients were excluded since
the proposed analysis is based on graph theory metrics that are not defined for signed edges
(Fornito, Zalesky, & Breakspear, 2013). All the connections with positive weight (z > 0) were
considered. Binary functional connectome was defined setting to 1 connections where z > 0
Network Neuroscience
401
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
4
2
3
9
7
1
9
6
6
9
0
9
n
e
n
_
a
_
0
0
1
2
6
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Brain connectivity and cognition in a rat model of Alzheimer’s disease
and 0 otherwise. The resulting functional connectomes have an average density of 27.72 ±
0.53.
Brain Network Analysis
Brain network organization was described using graph theory metrics, including degree, strength,
clustering coefficient, and local and global efficiency. These metrics provide a description of
different aspects of brain networks at a global level: network density, integration, and segrega-
tion (Rubinov & Sporns, 2010).
Nodal degree measures the number of connections of a region. Nodal strength is computed
as the sum of the region connection weights. Network degree and strength are respectively the
nodal degree and strength averaged in all the brain regions. Higher network degree/strength
indicates more or stronger connections. Global efficiency measures network integration: the
ability to combine information from different regions. It is inversely related to the shortest
path length between each pair of nodes. Higher global efficiency characterizes stronger and
faster communication through the network. Network segregation, the ability for specialized
processing within densely interconnected groups of regions, was quantified by local efficiency
and clustering coefficient. Local efficiency is the average in the whole network of the nodal
efficiencies (efficiency of the subnetwork associated with a region). Clustering coefficient is
the average of the nodal clustering coefficients, which measure the number of neighbors of a
node that are also neighbors with each other. High values of these metrics are related to highly
segregated and connected networks. In this way, alterations in the whole-brain organization
were evaluated using the same kind of measures in both structural and functional connec-
tomes. Nonetheless, the interpretation of these parameters must take into account the specific
kind of connectome and the definition of the connection weights. On the other hand, these
metrics have been commonly used in human studies of AD (Brier et al., 2014; Daianu et al.,
2013; Fischer et al., 2015; Sanz-Arigita et al., 2010) and therefore can provide comparable
and translational results.
Statistics Longitudinal Models
The main objective of our study was the research of alterations in the longitudinal evolution
of the TgF344-AD brain networks with respect to the control group and its impact on cogni-
tion. Linear mixed-effects (LME) models (Oberg & Mahoney, 2007) were used to model the
influence of age and group in the network metrics and to identify differences in the effect of
aging between the two groups. LME models include both fixed effects (parameters common
to an entire population such as age or group) and random effects (subject-specific parameters
modeling the subject deviation from the population).
LME was defined to regress each of the network metrics including group, age, and the
interaction between them as independent variables:
ys = β
0 + β
1
· group + β
· age + β
3
2
· group · age + β
4,s + ξ, s = 1, …, S,
(1)
Linear mixed-effects models:
A statistical model containing both
fixed effects (common parameters of
a population) and random effects
(subject-specific) to fit longitudinal
evolution.
where ys is the network metric in the subject s at a given age, s represents each of the S subjects;
0 is the global intercept; β
β
3 are the fixed-effect parameters, assessing the influence
4,s is the subject-specific correction. ξ is the
of group, age, and interaction respectively; and β
regression error term.
1, β
2, β
Multiple comparisons were corrected using false discovery rate (FDR; Benjamin & Hochberg,
1995). The effects of group, age, or interaction between age and group were considered
Network Neuroscience
402
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
4
2
3
9
7
1
9
6
6
9
0
9
n
e
n
_
a
_
0
0
1
2
6
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Brain connectivity and cognition in a rat model of Alzheimer’s disease
significant if the corrected p value (pFDR) was less than 0.05. When the interaction was sig-
nificant, control and transgenic groups were modeled separately to evaluate the age effect in
each group:
ysCTR = β
ysTG = β
0,CTR + β
0,TG + β
1,CTR
· age + β
2,sCTR + ξ
CTR, sCTR = 1, …, SCTR;
1,TG
· age + β
2,sTG + ξ
TG, sTG = 1, …, STG.
(2)
Network-based statistics:
Method to control for multiple
comparisons when statistical testing
is performed between every
connection in the brain network.
To complement the longitudinal analysis, differences in brain networks at each of the five ac-
quisition time points were evaluated. Kruskal-Wallis tests were applied to identify statistically
significant differences between whole-brain organization in transgenic and control groups.
Multiple comparisons were corrected using FDR (Benjamin & Hochberg, 1995). Together with
this, network-based statistics (NBS) toolbox (Zalesky, Fornito, & Bullmore, 2010) was used to
identify specific networks of connections that differ between transgenic and control animals.
NBS was performed with the following settings: t test with a threshold of 3.1, 5,000 permuta-
tions, and a significance level of p < 0.05.
Finally, the relationship between connectivity and cognitive performance was evaluated
fitting an LME model to test whether such relation is significant and whether it differs between
the groups:
DNMSs = β
0 + β
1
s = 1, ..., S,
· connectivity + β
· group + β
3
2
· connectivity · group + β
4,s + ξ,
(3)
where DNMSs represents the result in the DNMS task (number of trials and percentage of
correct responses) and connectivity refers to each of the network metrics. Thus, the relation-
ship between cognitive outcome and each of the network parameters was evaluated. Multiple
comparisons were corrected by FDR (Benjamin & Hochberg, 1995). When interaction was
significant (pFDR < 0.05), the model was fitted separately to transgenic and control groups to
test the significance of the relationship between the network metric and the cognitive outcome
in each group.
RESULTS
Longitudinal Analysis
Linear mixed-effects (LME) model was fitted to regress each of the network metrics and to
evaluate significant effects of age, group, or their interaction. Results are shown in Figure 1,
where the whole distribution of data is shown (each point represents a time point / subject), as
well as the fitting of the LME model as a function of group and age. If a significant effect of the
interaction between age and group was detected, the model was adjusted to each of the groups
independently to fit the network metric as a function of age. Significant pFDR values of each
of the terms in the model (group, age, and group-age interaction) are displayed in Figure 1.
Supplementary Table 1 compiles pFDR values and effect sizes (Cohen’s f 2) for the parameters
of the LME model fitted to the whole cohort, and Supplementary Table 2 shows pFDR values
and effect sizes (Cohen’s f 2, R2) of the model adjusted to each of the groups.
In FA-w connectome, group effect was significant in strength and clustering coefficient, but
no significant interaction between group and age was detected. Both FA-w strength and clus-
tering were clearly decreased in the transgenic group. A significant increase of FA-w strength
with age was observed, with a more notable slope in the case of transgenic animals.
Network Neuroscience
403
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
2
3
9
7
1
9
6
6
9
0
9
n
e
n
_
a
_
0
0
1
2
6
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Brain connectivity and cognition in a rat model of Alzheimer’s disease
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
2
3
9
7
1
9
6
6
9
0
9
n
e
n
_
a
_
0
0
1
2
6
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 1. Global network metrics of the structural connectome. Strength, global, and local efficiency and average clustering coefficient of
the three structural connectomes (fractional anisotropy-weighted, fiber density weighted, and binary connectomes). For each network metric,
each dot represents the value of one animal at one time point (blue: control, orange: TgF344-AD). Blue and orange lines represent the fit of the
linear mixed-effects model; pFDR values are shown for significant effects (pGroup group effect, pAge age effect, pInter effect of the interaction
between age and group). If interaction was significant group models were fitted to the data, in orange, pFDR values of the effect of age in the
LME model fitted to the transgenic group (no significant effects were observed in the control group).
Network Neuroscience
404
Brain connectivity and cognition in a rat model of Alzheimer’s disease
The interaction between group and age was significant in FD-w strength, local efficiency,
and clustering coefficient. In all these cases, an increase with age in the transgenic group—
significant in strength and clustering coefficient—was detected, opposite to the nonsignificant
decrease observed in controls. Similar behavior was observed in the binary connectome, where
significant age-group interaction was observed in all the network metrics, which were signifi-
cantly affected by age in the transgenic group.
No significant effects of age, group, or interaction were detected in either functional con-
nectivity or cognitive performance.
Group Differences by Time Points
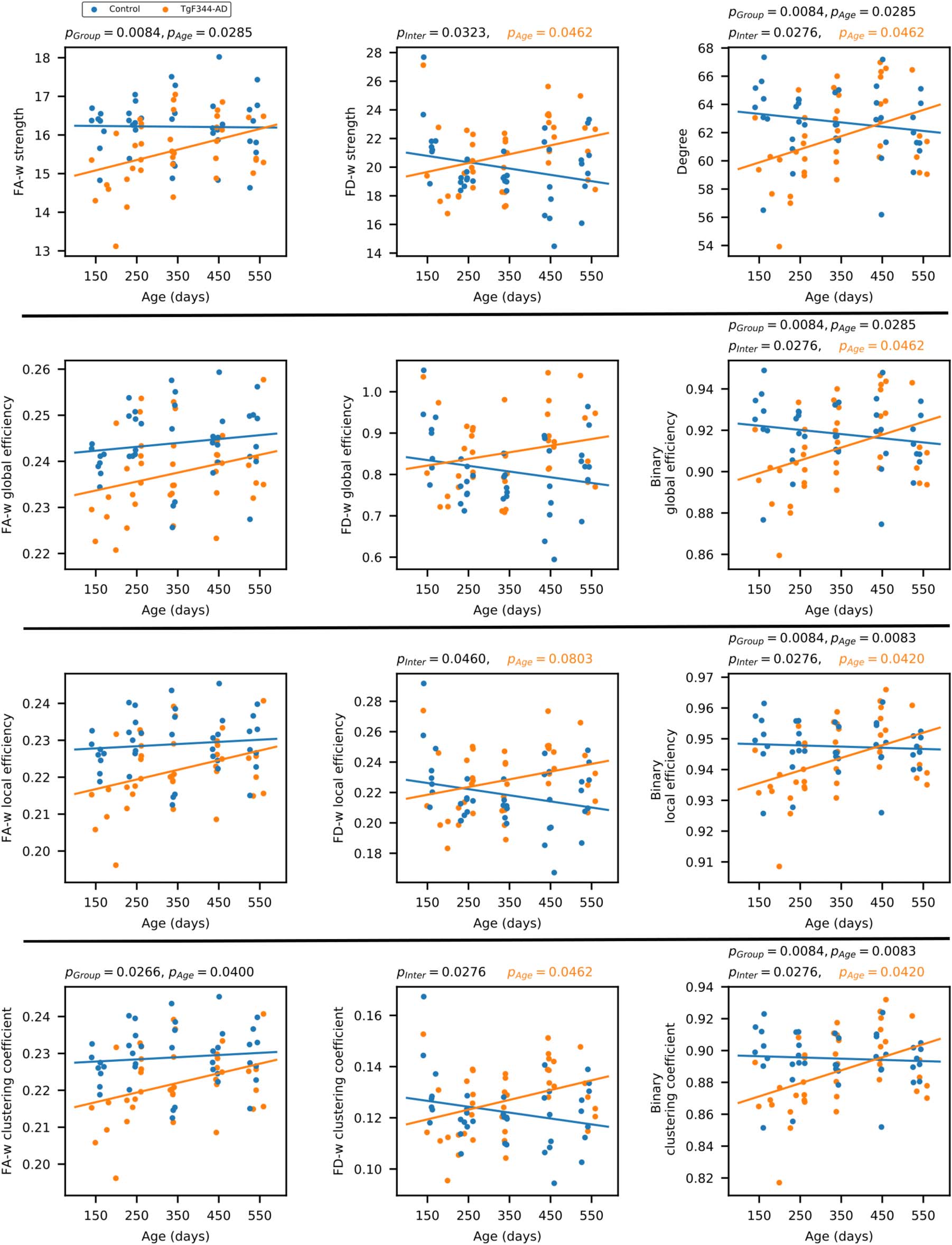
Figure 2 shows the distribution of network metrics between groups at each of the five acqui-
sition time points, and the statistically significant differences. Significance was considered as
pFDR < 0.05. Only structural network metrics are shown, since no differences were found in
functional connectomes. Supplementary Table 3 shows the pFDR values and effect sizes (η2)
of both structural and functional connectomes.
Most differences were observed at the earliest time point where FA-w and binary structural
network metrics were significantly decreased in the transgenic group with respect to controls.
Regarding the FD-w connectome, while a tendency to decreased values was observed in the
transgenic group with respect to controls at the first time point, a significant increase was de-
tected in strength, local efficiency, and clustering coefficient at 15 months of age. No significant
differences were found in the functional network metrics.
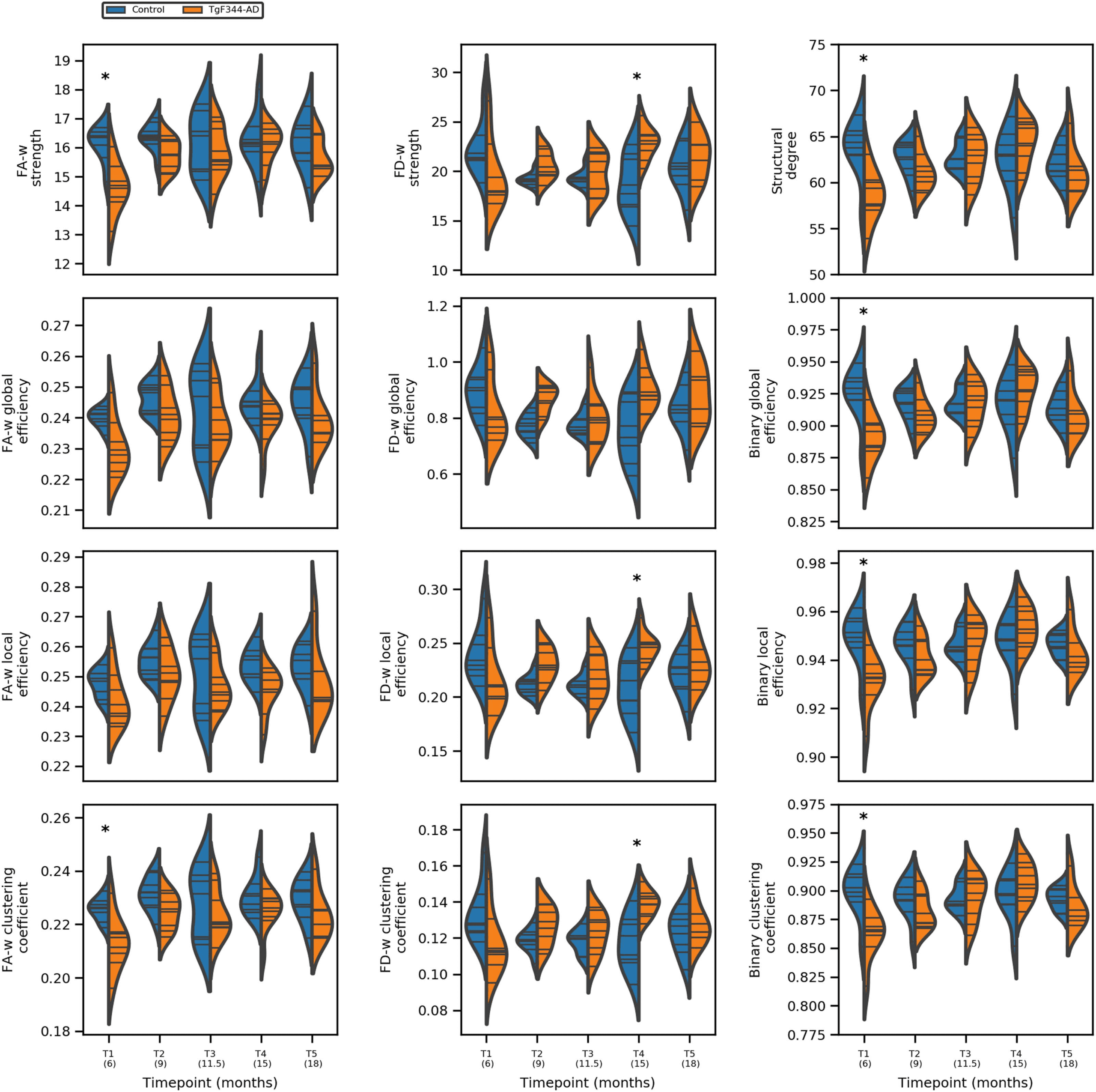
Together with graph metrics we evaluate differences in network edges. Figure 3 shows the
group average FA-w and functional weighted connectomes considering only the strongest con-
nections (FA-w> 0.3 and z > 0.05, respectively) to provide an illustrative plot of the brain net-
works. Stronger and denser FA-w connections are observed in the control group in comparison
with the transgenic group, especially in the last time point, although network metrics were not
significantly different. A decrease in the connection strength with time in the transgenic brain
can also be observed. Regarding the functional connectome it can be seen that at 18 months
of age, z > 0.05 connections in the TgF344-AD brain are less and weaker. In previous time
points connectivity is similar between both groups, and even higher strength in specific links
can be observed in the transgenic brain.
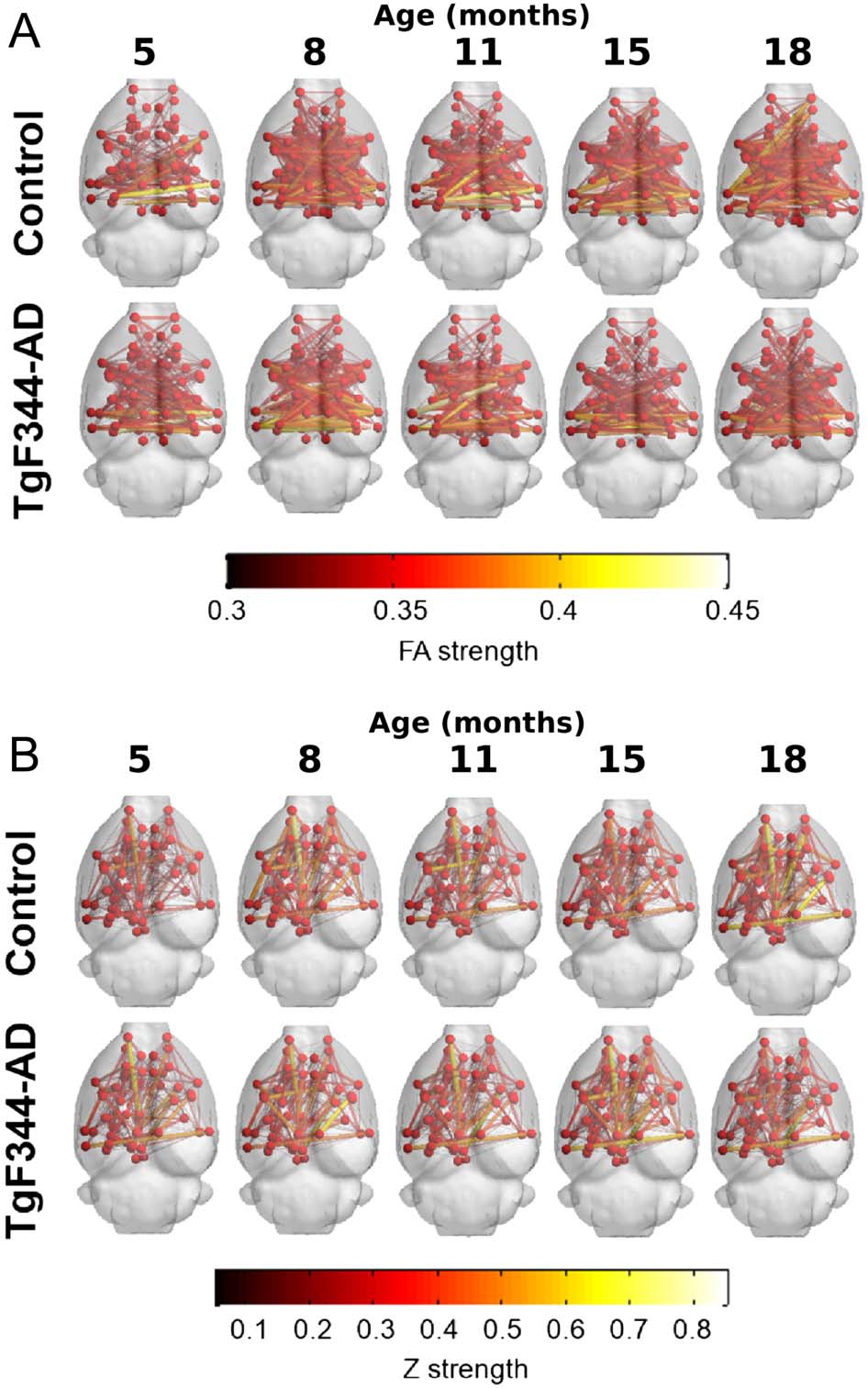
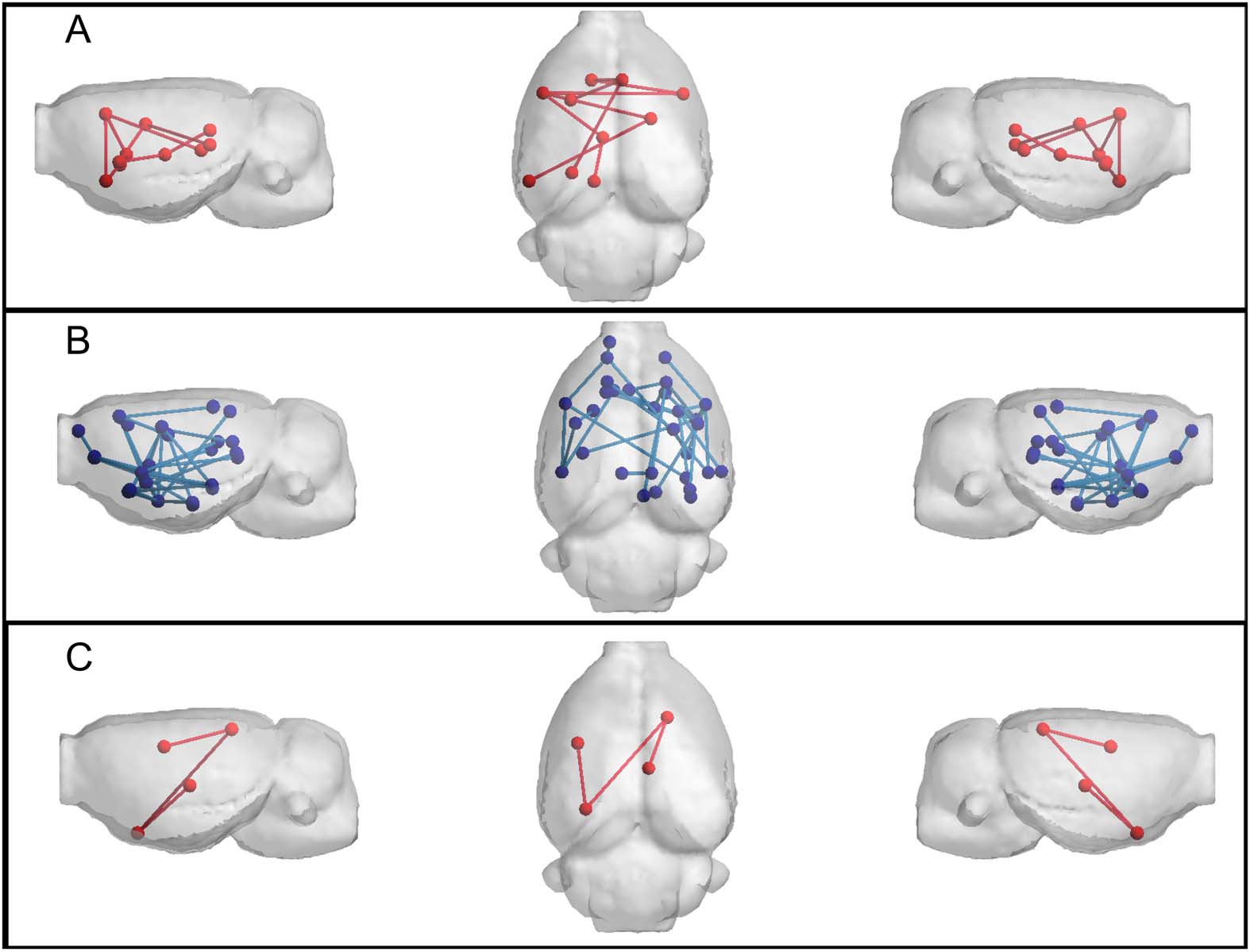
To statistically identify differences in the network edges, NBS was applied. Resulting net-
works are shown in Figure 4. Differences were detected at 8 months of age (time point 2) in
a subnetwork of the FD-w connectome (increased in transgenic animals) and at 15 months of
age (time point 4) in FA-w and functional connectomes. While the subnetwork identified in
the FA-w connectome was decreased in transgenic animals, subnetworks in the functional net-
work were increased in this group. The list of regions between which connectivity was altered
is shown in the Supporting Information.
Relationship Between Connectivity and Cognition
The effect of connectivity in cognitive results and whether this effect was different in transgenic
and control animals was evaluated using LME models.
Although no significant differences were observed in cognitive outcome between groups
(see Supplementary Figure 3), it can be noted that (a) transgenic animals performed a lower
number of trials than control animals at the first time point and (b) there is a high variability in
Network Neuroscience
405
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
4
2
3
9
7
1
9
6
6
9
0
9
n
e
n
_
a
_
0
0
1
2
6
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Brain connectivity and cognition in a rat model of Alzheimer’s disease
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
4
2
3
9
7
1
9
6
6
9
0
9
n
e
n
_
a
_
0
0
1
2
6
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 2. Network metrics of the three structural connectomes (fractional anisotropy weighted, fiber density weighted, and binary con-
nectome) in control (blue) and transgenic (orange) groups at each of the five time points. * represents statistically significant difference
(pFDR < 0.05).
Network Neuroscience
406
Brain connectivity and cognition in a rat model of Alzheimer’s disease
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
2
3
9
7
1
9
6
6
9
0
9
n
e
n
_
a
_
0
0
1
2
6
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. Average structural and functional brain networks. (A) Group average of the FA-w con-
nectome, and (B) group average of the functional weighted connectome at each time point. Only the
strongest connections are plotted (FA-w> 0.3 and z > 0.05). Color and width of the links represent
connection strength.
the results of the transgenic group at the last time point, when the performance of some of the
animals sharply falls. Furthermore, brain network organization had a significant impact on the
cognitive results.
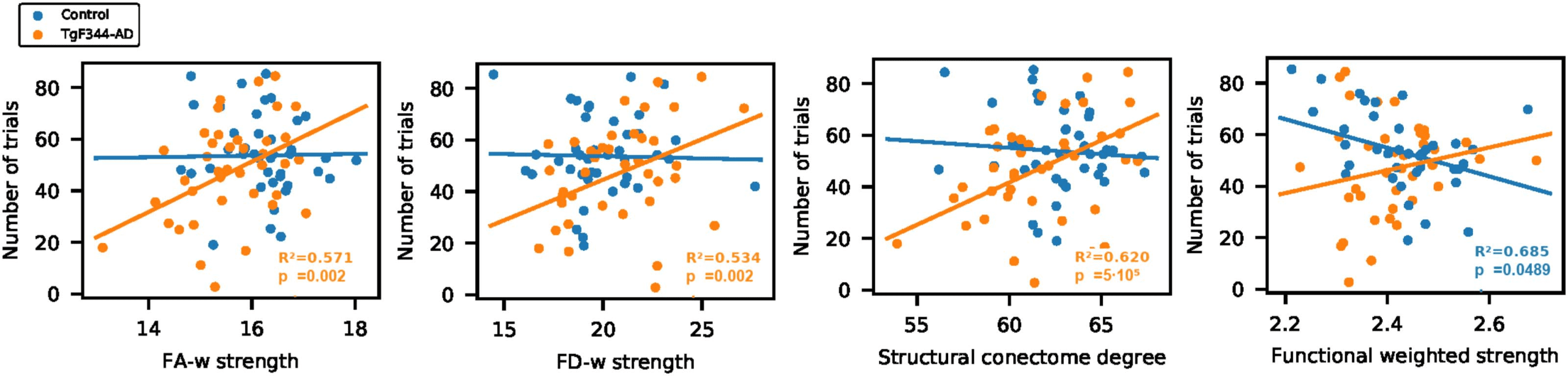
The interaction between FA-w strength and group had a significant effect in DNMS results
(pinter = 0.0223). Considering the group-specific model, this metric had a significant influence
in the number of trials performed by the transgenic animals (ptg = 0.0018), but not in controls.
Group and metric also show a significant effect in cognitive outcome (pgroup = 0.0203 and
pmetric = 0.0006, respectively).
Similar results were observed considering any of the FD-w metrics (strength: pgroup =
0.0098, pmetric = 0.0007, pinter = 0.0016; local efficiency: pgroup = 0.0098, pmetric = 0.0005,
−5, pinter = 0.0065)
pinter = 0.0167; clustering coefficient: pgroup = 0.0051, pmetric = 1.97 · 10
−5, pinter = 0.0065; global effi-
or binary metrics (degree: pgroup = 0.0051, pmetric = 1.6 · 10
−5, pinter = 0.0065; local efficiency: pgroup = 0.0098;
ciency: pgroup = 0.0051, pmetric = 1.6 · 10
−6, pinter = 0.0064; clustering coefficient: pgroup = 0.0098, pmetric = 4.4 ·
pmetric = 4.4 · 10
−6, pinter = 0.0064), except for FD-w global efficiency (pgroup = 0.0051, pmetric = 0.0035),
10
Network Neuroscience
407
Brain connectivity and cognition in a rat model of Alzheimer’s disease
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
4
2
3
9
7
1
9
6
6
9
0
9
n
e
n
_
a
_
0
0
1
2
6
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. Networks altered in TgF344-AD animals resulting from NBS analysis. (A) FD-w connec-
tome at 8 months of age; altered connections are related to limbic system. (B) FA-w connectome at
15 months of age; connections related to memory processing and executive functions. (C) functional
connectome at 15 months of age; connections related to processing of sensory inputs. Red indicates
subnetwork increased in the transgenic group; blue, subnetwork decreased in the transgenic group.
where the interaction between group and metric was not significant. In all the cases, the
higher the FD-w or structural binary network metrics, the more trials the transgenic animal
performed (FD-w strength, ptg = 0.0019; local efficiency, ptg = 0.0014; clustering coef-
−5; global efficiency, ptg =
ficient, ptg = 6.54 · 10
−5). All the
5.29 · 10
reported p values are FDR corrected. Figure 5 shows FA-w, FD-w strength, and structural degree
−5 and binary degree, ptg = 5.29 · 10
−5; clustering coefficient ptg = 1.26 · 10
−5; local efficiency, ptg = 1.26 · 10
Figure 5. Cognitive performance and network metrics. Linear mixed-effect model fit of DNMS result as a function of network metric and
group. Relationship between the structural and functional connectome metrics and number of trials performed in DNMS test. R2 and pFDR
in case of significant metric effect (orange: significance in TgF344-AD group; blue: significance in control group).
Network Neuroscience
408
Brain connectivity and cognition in a rat model of Alzheimer’s disease
(results are similar in all the mentioned metrics). More details are provided in Supplementary
Tables 4 and 5.
As shown in Figure 5, the number of trials was significantly influenced by the interaction
between group and functional weighted strength (pinter = 0.0150), global efficiency (pinter =
0.0180), and local efficiency (pinter = 0.0167). When specific group models were considered,
the effect of functional strength in the cognitive performance was significant in the control
animals (pctr = 0.0489), but not in the transgenic cohort.
DISCUSSION
There is a growing interest in the study of early stages of AD and its progression until symp-
tomatic onset, since brain changes start decades before the clinical diagnosis (Dubois et al.,
2016; Jack et al., 2018). To contribute to the understanding of these early brain changes and
their progression during aging, the present study focuses on an animal model of the disease
and describes the longitudinal evolution of structural and functional brain network organiza-
tion from very early stages. Although recent studies have evaluated connectivity in population
at risk of AD or in its preclinical phases in human cohorts (Berlot, Metzler-Baddeley, Ikram,
Jones, & O’Sullivan, 2016; Farrar et al., 2017; Pereira et al., 2017), they focused on elderly or
middle-aged subjects, and in the case of longitudinal analysis only short periods of time with
respect to human life span have been evaluated. In the present study, the use of TgF344-AD rats
allows the investigation of earlier alterations and to follow up on subjects during their entire
life span. Therefore, it can provide new insights into the disease progression and be helpful in
the investigation of treatments and interventions.
Our results show differences between transgenic and control groups in the progression of
the structural connectivity during aging. Alterations in the structural brain network of popu-
lation at risk of AD or in its preclinical phases in human cohorts of elderly or middle-aged
subjects have been reported (Berlot et al., 2016; Chen et al., 2015; Farrar et al., 2017; Fischer
et al., 2015; Pereira et al., 2017; Shu et al., 2015; Zhao et al., 2017), which are in line with the
differences we have observed in TgF344-AD animals at equivalent ages (15 months). Further-
more, earlier alterations were observed in the animal model: Structural connectivity differences
were already present in young animals (6 months of age). Together with this, differences in the
evolution of the structural metrics were detected. While aging had no significant effect in the
evolution of network metrics in the control group, it significantly affected metrics in the trans-
genic animals. In this group, network metrics increased linearly with age, but a decrease in the
metric values at the last time point can be observed in Figures 1 and 2, although LME could
not fit this change of trend. In spite of this global increment with age, FA-w network metrics in
transgenic animals always remain lower than in controls, in line with observations in preclini-
cal or clinical phases of AD in aged patients (Chen et al., 2015; Pereira et al., 2017; Shu et al.,
2015). Binary and FD-w metrics also increased significantly in transgenic animals during ag-
ing, but they were decreased with respect to controls only at early ages. Indeed, FD-w was
increased in transgenic animals at 8 and 15 months. This could be related to the hyperconnec-
tivity effect described after brain injury and in preclinical phases of AD (Hillary & Grafman,
2017), explained as a mechanism to preserve communication in the network and minimize
the behavioral deficits. Although hyperconnectivity has been mainly identified in functional
networks, higher binary structural network properties were also described in APOE-ε4 carriers
before mild cognitive impairment (MCI) appears (Ma et al., 2017). Thus, to compensate for
the lower FA-w strength in the network, more connections would be established to preserve
behavioral performance. In this line, the increase in FA-w network metrics with age observed
Network Neuroscience
409
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
4
2
3
9
7
1
9
6
6
9
0
9
n
e
n
_
a
_
0
0
1
2
6
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Brain connectivity and cognition in a rat model of Alzheimer’s disease
in transgenic animals is probably related to the presence of more connections rather than to
FA increase in the brain. The results obtained with NBS also points to this line. They showed
hyperconnectivity in subnetworks of functional and FD-w connectomes, while decreased con-
nectivity was observed in a subnetwork of the FA-w connectome. Namely, hyperconnectivity
was detected in networks related to the processing of sensory inputs in both functional and
FD-w connectome. The altered connections observed in the FD-w network are part of the
limbic system and are responsible for the processing of different sensory inputs required to
link emotions and memories. This could try to compensate for the decrease observed in FA-w
connectome in networks related to memory processing and executive functions.
Impairments in functional connectivity have been described in at-risk, preclinical, or clini-
cal AD cohorts, but depending on methodological issues or cohort selection differences have
been reported in opposed senses (Phillips, McGlaughlin, Ruth, Jager, & Soldan, 2015). Like-
wise, in the studied animal model, TgF344-AD, differences in connectivity between specific
regions or networks have been reported (Anckaerts et al., 2019; Tudela et al., 2019) at several
ages. In the present work, we focused on the analysis of brain network globally instead of
evaluation of specific connections. From this point of view, the whole-brain network metrics
describing integration and segregation of the functional connectomes did not significantly dif-
fer between control and transgenic groups. This could be related to three factors. The first is
the hyperconnectivity effect previously mentioned (Hillary & Grafman, 2017) between specific
regions or networks to compensate damaged connections (hypoconnectivity). This effect has
been described, for instance, in young APOE-ε4 carriers (Ma et al., 2017) or amnestic MCI
patients (Kim et al., 2015). Since global network metrics involve averaging properties of all the
connections (Rubinov & Sporns, 2010), decreased connections could be compensated for by
increased connections (such as that detected by NBS at 15 months of age), resulting in similar
values of the global network metrics, as observed in our study. Second, the training and the
repetition of the cognitive task could lead to a learning effect, increasing the cognitive reserve
of these animals and therefore preserving the functional connectivity. Higher functional con-
nectivity has been related to higher cognitive reserve in both healthy elders (Arenaza-Urquijo
et al., 2013) and MCI patients (Franzmeier et al., 2017) with respect to subjects with low
cognitive reserve. This fact could also be related to the absence of significant differences in
the cognitive outcome. Nevertheless, further investigation on animals not undergoing DNMS
should be performed to confirm this hypothesis. Finally, the third factor that could influence the
absence of functional connectivity differences could be the disease timing. Brain changes as
amyloid-β concentration and neural loss in TgF344-AD have been described from 16 months
of age in Cohen et al. (2013), and significant cognitive impairment is mainly described from
15 months (Cohen et al., 2013; Tsai et al., 2014) although tendencies to impairment or differ-
ences in anxiety or learning abilities have been reported at earlier stages (Cohen et al., 2013;
Muñoz-Moreno et al., 2018; Pentkowski et al., 2018). Actually, an increase in variability of the
cognitive outcome was observed at 18 months of age, when some of the transgenic animals
performed much worse than at previous ages. This suggests that the symptomatic onset occurs
around 18 months of age, and before this age, functional connectivity might compensate for
the structural network damage resulting in similar functional network metrics than in control
animals. The observed significant relation between structural network metrics and cognitive
performance could be related to the ability of functional connectivity to cope with the struc-
tural disconnection preserving cognition, until a breakpoint when it is not able to deal with
extensive structural network damage. This relationship between structural connectivity and
cognition has also been described in MCI subjects (Berlot et al., 2016; Farrar et al., 2017) and
in cognitive normal individuals harboring amyloid pathology and neurodegeneration (Pereira
Cognitive reserve:
Ability of the brain to preserve
cognitive skills when brain damage
occurs.
Network Neuroscience
410
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
4
2
3
9
7
1
9
6
6
9
0
9
n
e
n
_
a
_
0
0
1
2
6
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Brain connectivity and cognition in a rat model of Alzheimer’s disease
et al., 2017), while no correlation was observed in control subjects. Coherently, structural
connectivity did not correlate with DNMS in our control cohort, where correlation between
DNMS and functional connectivity was observed.
Strengths and Limitations
The use of MRI-based connectomics to longitudinally analyze the brain network in an animal
model of AD provides a valuable and highly translational approach to the research on the
mechanism of the disease progression, since the applied methodology can be easily translated
to clinical investigation. Moreover, animal models allow for characterization and follow-up
of the same cohort from early stages until advanced phases of the disease. Indeed, the model
used in our experiments, the TgF344-AD rats, has been shown to develop all the AD patho-
logical hallmarks in a progressive manner, which makes it especially suitable for longitudinal
evaluation.
The use of graph metrics allows for comparison between alterations observed in the animal
model and previous results in human cohorts in the literature. This is essential to validate how
the animal model mimics the pathology in patients. It is also a critical point to allow transla-
tionality between preclinical and clinical trials in the research for AD treatments (Drummond
& Wisniewski, 2017; Sabbagh et al., 2013). Our results are coherent with those observed in
elderly or middle-aged human cohorts and provide further information about earlier brain al-
terations and the pattern of disconnection associated with AD progression. Furthermore, the
use of a rat model allows for better behavioral characterization than other animal models (Do
Carmo & Cuello, 2013). This makes it possible to perform cognitive evaluation of memory-
related functions and relate animal performance to brain connectivity. Our results revealed
that the influence of brain network organization in cognitive abilities differs between trans-
genic and control animals.
Regarding the limitations of the study, the relatively small number of subjects could limit the
statistical analysis. However, the measures were repeated at five time points, which increases
the sample size evaluated by the longitudinal models to 80 observations. Note that our main
results are based on models fitted to all these observations. A small sample could have a bigger
impact on the complementary analysis at specific time points, but even with this limitation
significant differences between the groups were detected after multiple comparison correction.
These differences were coherent with previous findings observed in bigger cohorts from the
human population. Nevertheless, the small sample size could hamper the statistical analysis
and be related, for instance, to the lack of significant differences at the last time point.
MRI protocols were optimized to achieve a compromise between sensitivity, image quality,
and acquisition time. After optimization, TE of the gradient-echo BOLD acquisition was set to
10.75 ms. Although it is slightly shorter than common echo times used to sensitize images to
BOLD variations, the analysis of the resulting BOLD signal showed patterns of connectivity
consistent with previous knowledge, as shown in Supplementary Figure 2. Resting-state ac-
quisitions were acquired using medetomidine as sedation, which was considered to preserve
connectivity networks better than isoflurane anesthesia (Kalthoff, Po, Wiedermann, & Hoehn,
2013). However, recent studies have thoroughly investigated the effect of anesthesia in brain
function during resting state, and suggested that the use of only medetomidine could hinder
brain function that has been observed in awake animals. These new findings should be taken
into account in new experimental protocols, and could lead to additional conclusions that
complement our analysis.
Network Neuroscience
411
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
4
2
3
9
7
1
9
6
6
9
0
9
n
e
n
_
a
_
0
0
1
2
6
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Brain connectivity and cognition in a rat model of Alzheimer’s disease
On the other hand, we would like to mention that all animals in the study repeated the
DNMS task every 3 months, which allows for evaluation of cognitive skills. Results point to
a learning effect and increase in cognitive reserve due to such repetition. However, further
experiments including animals that do not perform DNMS should be carried out for a more
thorough evaluation of such an effect.
Finally, the last time point evaluated in our study was 18 months of age. Brain changes and
cognitive impairment in the TgF344-AD have been described mainly from 15 months of age,
which, as previously discussed, could be related to the absence of differences in cognition or
functional connectivity. Therefore, further investigation describing connectivity at later time
points would be of great interest to characterize more advanced stages of the disease.
CONCLUSIONS
Aging had more notable impact on the structural connectivity of the TgF344-AD rats, which is
altered from early ages, than in control animals. In addition, differences in anatomical networks
directly affected the cognitive outcome of the transgenic animals, even before the symptomatic
onset. These findings are in line with results observed in middle-aged or elderly human pop-
ulation at risk of AD, and complement them with insights into earlier stages and a plot of the
effects of the disease along the whole life span. The results support the idea of AD as a discon-
nection syndrome and AD as a continuum, suggesting that brain damage is already present at
early stages, long before the symptomatic onset.
The impact of the altered anatomical connectivity in cognitive skills could be moderated
by functional network reorganization until advanced stages of the disease. This suggests the
relevance of cognitive reserve to prevent or mitigate the symptomatic onset in subjects affected
by the disease. The TgF344-AD model could therefore be a convenient model to perform
translational research of the impact of cognitive interventions in AD.
ACKNOWLEDGMENTS
TgF344-AD rats were obtained through the InMind Consortium following a kind donation by
Dr. T. Town.
SUPPORTING INFORMATION
Supporting information for this article is available at https://doi.org/10.1162/netn_a_00126.
AUTHOR CONTRIBUTIONS
Emma Muñoz-Moreno: Conceptualization; Formal analysis;
Investigation; Methodology;
Writing – Original Draft; Writing – Review & Editing. Raúl Tudela: Methodology; Writing –
Review & Editing. Xavier López-Gil: Data curation; Methodology; Writing – Review & Edit-
ing. Guadalupe Soria: Conceptualization; Data curation; Funding acquisition; Investigation;
Supervision; Writing – Review & Editing.
FUNDING INFORMATION
Instituto de Salud Carlos III (http://dx.doi.org/10.13039/501100004587), Award ID: PI14/00595.
Emma Muñoz-Moreno, Fundació la Marató de TV3 (http://dx.doi.org/10.13039/100008666),
Award ID: 201441 10. FP7 Health, Award ID: FP7-HEALTH-2011.2.2.1-2. Instituto de Salud
Network Neuroscience
412
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
4
2
3
9
7
1
9
6
6
9
0
9
n
e
n
_
a
_
0
0
1
2
6
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Brain connectivity and cognition in a rat model of Alzheimer’s disease
Carlos III (http://dx.doi.org/10.13039/501100004587), Award ID: PI18/00893, co-funded by
ERDF, “A way to make Europe”. Secretaria d’Universitats i Recerca del Departament d’Empresa
I Coneixement de la Generalitat de Catalunya, Award ID: AGAUR 2017 SGR 01003. European
Community, Award ID: FP7-HEALTH-2011.2.2.1-2, n 278850.
REFERENCES
Anckaerts, C., Blockx, I., Summer, P., Michael, J., Hamaide, J.,
Kreutzer, C., . . . Van der Linden, A. (2019). Early functional
connectivity deficits and progressive microstructural alterations
in the TgF344-AD rat model of Alzheimer’s disease: A longitudi-
nal MRI study. Neurobiology of Disease, 124, 93–107.
Arenaza-Urquijo, E. M., Landeau, B., La Joie, R., Mevel, K.,
Mézenge, F., Perrotin, A., . . . Chételat, G. (2013). Relationships
between years of education and gray matter volume, metabolism
and functional connectivity in healthy elders. NeuroImage, 83,
450–457.
Avants, B. B., Epstein, C. L., Grossman, M., & Gee, J. C. (2008). Sym-
metric diffeomorphic image registration with cross-correlation:
Evaluating automated labeling of elderly and neurodegenerative
brain. Medical Image Analysis, 12, 26–41.
Badhwar, A. P., Tam, A., Dansereau, C., Orban, P., Hoffstaedter,
F., & Bellec, P. (2017). Resting-state network dysfunction in
Alzheimer’s disease: A systematic review and meta-analysis.
Alzheimer’s & Dementia: Diagnosis, Assessment & Disease Mon-
itoring, 8, 73–85.
Benjamin, Y., & Hochberg, Y. (1995). Controlling the false discov-
ery rate: A practical and powerful approach to multiple testing.
Journal of the Royal Statistical Society. Series B (Methodological),
57(1), 289–300.
Berlot, R., Metzler-Baddeley, C., Ikram, M. A., Jones, D. K., &
O’Sullivan, M. J. (2016). Global efficiency of structural networks
mediates cognitive control in mild cognitive impairment. Fron-
tiers in Aging Neuroscience, 8, 292.
Brier, M. R., Thomas, J. B., Fagan, A. M., Hassenstab, J., Holtzman,
D. M., Benzinger, T. L., . . . Ances, B. M. (2014). Functional
connectivity and graph theory in preclinical Alzheimer’s disease.
Neurobiology of Aging, 35(4), 757–768.
Cacciaglia, R., Molinuevo, J. L., Falcón, C., Brugulat-Serrat, A.,
Sánchez-Benavides, G., Gramunt, N., . . . Gispert, J. D. (2018).
Effects of APOE-(cid:6)4 allele load on brain morphology in a cohort
of middle-aged healthy individuals with enriched genetic risk for
Alzheimer’s disease. Alzheimer’s & Dementia, 14(7), 902–912.
Chen, Y., Chen, K., Zhang, J., Li, X., Shu, N., Wang, J., . . . Reiman,
E. M. (2015). Disrupted functional and structural networks in
cognitively normal elderly subjects with the APOE ε4 allele.
Neuropsychopharmacology, 40, 1181–1191.
Cohen, R. M., Rezai-Zadeh, K., Weitz, T. M., Rentsendorj, A., Gate,
D., Spivak, I., . . . Town, T. C. (2013). A transgenic Alzheimer rat
with plaques, tau pathology, behavioral impairment, oligomeric
Aβ and frank neuronal loss. Journal of Neuroscience, 33(15),
6245–6256.
(2013). Breakdown of brain connectivity between normal aging
and Alzheimer’s disease: A structural k-core network analysis.
Brain connectivity, 3(4), 407–422.
Do Carmo, S., & Cuello, A. C. (2013). Modeling Alzheimer’s disease
in transgenic rats. Molecular Neurodegeneration, 8(1), 37.
Drummond, E., & Wisniewski, T. (2017). Alzheimer’s disease: Ex-
perimental models and reality. Acta Neuropathologica, 133(2),
155–175.
Dubois, B., Hampel, H., Feldman, H. H., Scheltens, P., Aisen, P.,
Andrieu, S., . . . Jack, C. R. (2016). Preclinical Alzheimer’s
disease: Definition, natural history, and diagnostic criteria.
Alzheimer’s & Dementia, 12(3), 292–323.
Farrar, D. C., Mian, A. Z., Budson, A. E., Moss, M. B., Koo, B. B., &
Killiany, R. J. (2017). Retained executive abilities in mild cogni-
tive impairment are associated with increased white matter net-
work connectivity. European Radiology, 28, 340–347.
Fischer, F. U., Wolf, D., Scheurich, A., & Fellgiebel, A. (2015).
in preclinical
Altered whole-brain white matter networks
Alzheimer’s disease. NeuroImage: Clinical, 8, 660–666.
Fornito, A., Zalesky, A., & Breakspear, M. (2013). Graph analysis of
the human connectome: Promise, progress, and pitfalls. Neuro-
Image, 80, 426–44.
Franzmeier, N., Caballero, M. Á. A., Taylor, A. N. W., Simon-
Vermot, L., Buerger, K., Ertl-Wagner, B., . . . Ewers, M. (2017).
Resting-state global functional connectivity as a biomarker of
cognitive reserve in mild cognitive impairment. Brain Imaging
and Behavior, 11(2), 368–382.
Frisoni, G. B., Fox, N. C., Jack, C. R., Scheltens, P., & Thompson,
P. M. (2010). The clinical use of structural MRI in Alzheimer dis-
ease. Nature Reviews Neurology, 6(2), 67–77.
Galeano, P., Martino Adami, P. V., Do Carmo, S., Blanco, E., Rotondaro,
C., Capani, F., . . . Morelli, L. (2014). Longitudinal analysis of
the behavioral phenotype in a novel transgenic rat model of
early stages of Alzheimer’s disease. Frontiers in Behavioral Neuro-
science, 8, 321.
Garyfallidis, E., Brett, M., Amirbekian, B., Rokem, A., van der Walt,
S., Descoteaux, M., & Nimmo-Smith, I. (2014). Dipy, a library for
the analysis of diffusion MRI data. Frontiers in Neuroinformatics,
8, 8.
Gomez-Ramirez, J., & Wu, J. (2014). Network-based biomarkers in
Alzheimer’s disease: Review and future directions. Frontiers in
Aging Neuroscience, 6, 12.
Gour, N., Felician, O., Didic, M., Koric, L., Gueriot, C., Chanoine,
V., . . . Ranjeva, J. P. (2014). Functional connectivity changes
differ in early and late-onset Alzheimer’s disease. Human Brain
Mapping, 35(7), 2978–2994.
Daianu, M., Jahanshad, N., Nir, T. M., Toga, A. W., Jack, C. R.,
Weiner, M. W., . . . Alzheimer’s Disease Neuroimaging Initiative.
Habib, M., Mak, E., Gabel, S., Su, L., Williams, G. B., Waldman,
A., . . . O’Brien, J. T. (2017). Functional neuroimaging findings in
Network Neuroscience
413
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
4
2
3
9
7
1
9
6
6
9
0
9
n
e
n
_
a
_
0
0
1
2
6
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Brain connectivity and cognition in a rat model of Alzheimer’s disease
healthy middle-aged adults at risk of Alzheimer’s disease. Ageing
Research Reviews, 36, 88–104.
Hillary, F. G., & Grafman, J. H. (2017). Injured brains and adaptive
networks: The benefits and costs of hyperconnectivity. Trends in
Cognitive Sciences, 21(5), 385–401.
Jack, C. R., Barnes, J., Bernstein, M. A., Borowski, B. J., Brewer, J.,
Clegg, S., . . . Weiner, M. W. (2015). Magnetic resonance imaging
in Alzheimer’s Disease Neuroimaging Initiative 2. Alzheimer’s &
Dementia, 11(7), 740–756.
Jack, C. R., Bennett, D. A., Blennow, K., Carrillo, M. C., Dunn, B.,
Haeberlein, S. B., . . . Silverberg, N. (2018). NIA-AA Research
Framework: Toward a biological definition of Alzheimer’s dis-
ease. Alzheimer’s & Dementia, 14(4), 535–562.
Kalthoff, D., Po, C., Wiedermann, D., & Hoehn, M. (2013). Re-
lihability and spatial specificity of
rat brain sensorimotor
functional connectivity networks are superior under sedation
compared with general anesthesia. NMR in Biomedicine, 26(6),
638–650.
Kim, H., Yoo, K., Na, D. L., Seo, S. W., Jeong, J., & Jeong, Y.
(2015). Non-monotonic reorganization of brain networks with
Alzheimer’s disease progression. Frontiers in Aging Neuro-
science, 7, 1–10.
Leon, W. C., Canneva, F., Partridge, V., Allard, S., Ferretti, M. T.,
DeWilde, A., . . . Cuello, A. C. (2010). A novel transgenic rat
model with a full Alzheimer’s-like amyloid pathology displays
pre-plaque intracellular amyloid-beta-associated cognitive im-
pairment. Journal of Alzheimer’s Disease, 20(1), 113–126.
Lo, C.-Y., Wang, P.-N., Chou, K.-H., Wang, J., He, Y., & Lin, C.-P.
(2010). Diffusion tensor tractography reveals abnormal topolog-
ical organization in structural cortical networks in Alzheimer’s
disease. Journal of Neuroscience, 30(50), 16876–16885.
Ma, C., Wang, J., Zhang, J., Chen, K., Li, X., Shu, N., . . . Zhang, Z.
(2017). Disrupted brain structural connectivity: Pathological in-
teractions between genetic APOE (cid:6)4 status and developed MCI
condition. Molecular Neurobiology, 54(9), 6999–7007.
Mak, E., Gabel, S., Mirette, H., Su, L., Williams, G. B., Waldman,
A., . . . O’Brien, J. (2017). Structural neuroimaging in preclinical
dementia: From microstructural deficits and grey matter atrophy
to macroscale connectomic changes. Ageing Research Reviews,
35, 250–264.
Muñoz-Moreno, E., Tudela, R., López-Gil, X., & Soria, G. (2018).
Early brain connectivity alterations and cognitive impairment
in a rat model of Alzheimer’s disease. Alzheimer’s Research &
Therapy, 10, 16.
Oberg, A. L., & Mahoney, D. W. (2007). Linear mixed effects
models. In Methods in molecular biology: Topics in biostatistics
(Vol. 404, pp. 213–234). Totowa, NJ: Humana Press.
Palesi, F., Castellazzi, G., Casiraghi, L., Sinforiani, E., Vitali, P.,
Wheeler-Kingshott, C. A. G., & Angelo, E. D. (2016). Explor-
ing patterns of alteration in Alzheimer’s disease brain networks:
A combined structural and functional connectomics analysis.
Frontiers in Neuroscience, 10, 380.
Pentkowski, N. S., Berkowitz, L. E., Thompson, S. M., Drake, E. N.,
Olguin, C. R., & Clark, B. J. (2018). Anxiety-like behavior as an
early endophenotype in the TgF344-AD rat model of Alzheimer’s
disease. Neurobiology of Aging, 61, 169–176.
Pereira, J. B., van Westen, D., Stomrud, E., Strandberg, T. O., Volpe,
G., Westman, E., & Hansson, O. (2017). Abnormal structural
brain connectome in individuals with preclinical Alzheimer’s dis-
ease. Cerebral Cortex, 28(10), 3638–3649.
Phillips, D. J., McGlaughlin, A., Ruth, D., Jager, L. R., & Soldan, A.
(2015). Graph theoretic analysis of structural connectivity across
the spectrum of Alzheimer’s disease: The importance of graph
creation methods. NeuroImage: Clinical, 7, 377–390.
Power, J. D., Plitt, M., Laumann, T. O., & Martin, A. (2017). Sources
and implications of whole-brain fMRI signals in humans. Neuro-
Image, 146, 609–625.
Reiter, K., Nielson, K. A., Durgerian, S., Woodard, J. L., Smith,
J. C., Seidenberg, M., . . . Rao, S. M. (2017). Five-year longitu-
dinal brain volume change in healthy elders at genetic risk
for Alzheimer’s disease. Journal of Alzheimer’s Disease, 55(4),
1363–1377.
Rubinov, M., & Sporns, O. (2010). Complex network measures of
brain connectivity: Uses and interpretations. NeuroImage, 52(3),
1059–1069.
Sabbagh, J. J., Kinney, J. W., & Cummings, J. L. (2013). Alzhei-
mer’s disease biomarkers in animal models: Closing the transla-
tional gap. American Journal of Neurodegenerative Disease, 2(2),
108–120.
Sanz-Arigita, E. J., Schoonheim, M. M., Damoiseaux, J. S., Rombouts,
S. A. R. B., Maris, E., Barkhof, F., . . . Stam, C. J. (2010). Loss of
“small-world” networks in Alzheimer’s disease: Graph analysis
of fMRI resting-state functional connectivity. PLoS ONE, 5(11),
e13788.
Schwarz, A. J., Danckaert, A., Reese, T., Gozzi, A., Paxinos, G.,
Watson, C., . . . Bifone, A. (2006). A stereotaxic MRI template
set for the rat brain with tissue class distribution maps and co-
registered anatomical atlas: Application to pharmacological MRI.
NeuroImage, 32(2), 538–550.
Shu, N., Li, X., Ma, C., Zhang, J., Chen, K., Liang, Y., . . . Zhang, Z.
(2015). Effects of APOE promoter polymorphism on the topologi-
cal organization of brain structural connectome in nondemented
elderly. Human Brain Mapping, 36(12), 4847–4858.
Supekar, K., Menon, V., Rubin, D., Musen, M., & Greicius, M. D.
(2008). Network analysis of intrinsic functional brain connec-
tivity in Alzheimer’s disease. PLoS Computational Biology, 4(6),
e1000100.
ten Kate, M., Sanz-Arigita, E. J., Tijms, B. M., Wink, A. M., Clerigue,
M., Garcia-Sebastian, M., . . . Barkhof, F. (2016). Impact of
APOE-ε4 and family history of dementia on gray matter atrophy
in cognitively healthy middle-aged adults. Neurobiology of Ag-
ing, 38, 14–20.
Tsai, Y., Lu, B., Ljubimov, A. V., Girman, S., Ross-Cisneros, F. N.,
Sadun, A. A., . . . Wang, S. (2014). Ocular changes in TgF344-AD
rat model of Alzheimer’s disease. Investigative Ophthalmology
and Visual Science, 55(1), 523–534.
Tudela, R., Muñoz-Moreno, E., Sala-Llonch, R., López-Gil, X., &
Soria, G. (2019). Resting state networks in the TgF344-AD rat
model of Alzheimer’s disease are altered from early stages. Fron-
tiers in Aging Neuroscience, 11, 213.
Valdés-Hernández, P. A., Sumiyoshi, A., Nonaka, H., Haga,
R., Aubert-Vásquez, E., Ogawa, T., . . . Kawashima, R. (2011).
Network Neuroscience
414
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
4
2
3
9
7
1
9
6
6
9
0
9
n
e
n
_
a
_
0
0
1
2
6
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Brain connectivity and cognition in a rat model of Alzheimer’s disease
An in vivo MRI template set for morphometry, tissue segmenta-
tion, and fMRI localization in rats. Frontiers in Neuroinformatics,
5, 26.
Wee, C.-Y., Yap, P.-T., Li, W., Denny, K., Browndyke, J. N., Potter,
G. G., . . . Shen, D. (2011). Enriched white matter connectiv-
ity networks for accurate identification of MCI patients. Neuro-
Image, 54(3), 1812–1822.
Weston, P. S. J., Simpson, I. J. A., Ryan, N. S., Ourselin, S., &
Fox, N. C. (2015). Diffusion imaging changes in grey matter in
Alzheimer’s disease: A potential marker of early neurodegenera-
tion. Alzheimer’s Research & Therapy, 7, 47.
Windisch, M. (2014). We can treat Alzheimer’s disease success-
fully in mice but not in men: Failure in translation? A perspec-
tive. Neurodegenerative Diseases, 13(2–3), 147–150.
Xie, T., & He, Y. (2012). Mapping the Alzheimer’s brain with con-
nectomics. Frontiers in Psychiatry, 2, 77.
Zalesky, A., Fornito, A., & Bullmore, E. T. (2010). Network-based
statistic: Identifying differences in brain networks. NeuroImage,
53(4), 1197–1207.
Zhao, T., Sheng, C., Bi, Q., Niu, W., Shu, N., & Han, Y. (2017).
Age-related differences in the topological efficiency of the brain
structural connectome in amnestic mild cognitive impairment.
Neurobiology of Aging, 59, 144–155.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
4
2
3
9
7
1
9
6
6
9
0
9
n
e
n
_
a
_
0
0
1
2
6
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Network Neuroscience
415