RESEARCH
Psychological resilience correlates with EEG
source-space brain network flexibility
Véronique Paban
1
, Julien Modolo
2
, Ahmad Mheich
2
, and Mahmoud Hassan
2
1Aix Marseille University, CNRS, LNSC, Marseille, France
2University of Rennes, INSERM, LTSI-U1099, F-35000 Rennes, France
Keywords: EEG source connectivity, Psychological resilience, Resting state, Flexibility
ABSTRACT
We aimed at identifying the potential relationship between the dynamical properties of the
human functional network at rest and one of the most prominent traits of personality, namely
resilience. To tackle this issue, we used resting-state EEG data recorded from 45 healthy
subjects. Resilience was quantified using the 10-item Connor-Davidson Resilience Scale
(CD-RISC). By using a sliding windows approach, brain networks in each EEG frequency
band (delta, theta, alpha, and beta) were constructed using the EEG source-space
connectivity method. Brain networks dynamics were evaluated using the network flexibility,
linked with the tendency of a given node to change its modular affiliation over time. The
results revealed a negative correlation between the psychological resilience and the brain
network flexibility for a limited number of brain regions within the delta, alpha, and beta
bands. This study provides evidence that network flexibility, a metric of dynamic functional
networks, is strongly correlated with psychological resilience as assessed from personality
testing. Beyond this proof-of-principle that reliable EEG-based quantities representative of
personality traits can be identified, this motivates further investigation regarding the full
spectrum of personality aspects and their relationship with functional networks.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
2
5
3
9
1
0
9
2
6
4
0
n
e
n
_
a
_
0
0
0
7
9
p
d
.
t
AUTHOR SUMMARY
In this study, we investigated the possible correlation between one of the most important
personality traits, resilience, with a metric of dynamic functional networks called flexibility.
From EEG resting-state recordings in N = 45 volunteers, we unveiled such a correlation and
identified the brain regions involved in psychological resilience, from frequency-specific
networks.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
INTRODUCTION
An evolving field of neuroscience aims to reveal the neural substrates of personality, referring
to the relatively stable character of an individual that influences her or his long-term behavioral
style (Dubois et al., 2018). A key personality character is resilience, defined as the ability to
adapt to stress, adversity, and negative events and cope actively with life challenges (Fletcher
& Sarkar, 2013; Luthar, 2003; Rutter, 2006). Recently, a multisystem model of resilience has
been proposed by Liu et al. (2017), in which resilience comprises three structures: (1) the
innermost layer, which comprises the physiological, biological, and demographic profiles of
an individual; (2) the intermediate layer, which includes internal factors such as family, friends,
and personal experiences; and (3) the outermost layer, which corresponds to external resilience
such as access to healthcare and social services. However, except a few efforts (Kong et al.,
a n o p e n a c c e s s
j o u r n a l
Citation: Paban, V., Modolo, J., Mheich,
A., & Hassan, M. (2019). Psychological
resilience correlates with EEG
source-space brain network flexibility.
Network Neuroscience, 3(2), 539–550.
https://doi.org/10.1162/netn_a_00079
DOI:
https://doi.org/10.1162/netn_a_00079
Supporting Information:
https://doi.org/10.1162/netn_a_00079
Received: 8 October 2018
Accepted: 17 January 2019
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Author:
Julien Modolo
julien.modolo@inserm.fr
Handling Editor:
Olaf Sporns
Copyright: © 2019
Massachusetts Institute of Technology
Published under a Creative Commons
Attribution 4.0 International
(CC BY 4.0) license
The MIT Press
Psychological resilience and functional networks flexibility
Resilience:
One of the main personality traits,
describing the capability of
individuals to cope with stress and
life changes.
EEG:
Electroencephalography is a
neuroimaging technique consisting
of recording with scalp electrodes
the small currents induced by
neuronal activity at the cortex level.
Source-space networks:
Networks formed by interacting
anatomical brain regions, as opposed
to sensor-level networks.
Flexibility:
Metric from graph theory quantifying
how often one node of a network
changes its affiliation with a module
over time.
2015; Reynaud et al., 2013; Waugh & Koster, 2015), the neural subtracts of this complex
personality trait remain unclear.
The last decade has witnessed an increase of studies that consider the human brain as a
large-scale network. It is thus unsurprising that network neuroscience, which uses tools from
graph theory to better understand neural systems (Bassett & Sporns, 2017), has become one of
the most promising approaches to link behavior to brain function, including personality traits
(Markett et al., 2018). In the network neuroscience model, the human brain is summarized
by a set of nodes representing brain regions and a set of edges representing the connections
between these brain regions. The magneto/electro-encephalography (M/EEG) source-space
networks provide a unique direct and noninvasive access to those electrophysiological brain
networks, at the milliseconds temporal scale (Hassan & Wendling, 2018; Mheich et al., 2018).
The excellent time precision of this method allows the tracking of the brain networks dynam-
ics at an unprecedented timescale. In this paper, we aimed to test the hypothesis that metrics
derived from brain networks dynamics can be correlated to resilience scores.
To test this hypothesis, we used resting-state EEG data recorded from N = 45 healthy
subjects. Resilience was quantified using the Connor-Davidson Resilience Scale (CD-RISC)
(Campbell-Sills & Stein, 2007), where higher scores correspond to greater resilience. Brain
networks in each frequency band (delta, theta, alpha, and beta) were constructed using the
EEG source-space connectivity method. By applying multislice modularity algorithms on the
dynamic networks, the reconfiguration of EEG source-space networks was quantified using
flexibility, defined as how often a given node changes its modular affiliation over time, com-
puted at the level of each brain region. The choice of flexibility as an appropriate metric was
made following a recent study discussing the relationship between functional networks and
personality traits (Tompson et al., 2018). Indeed, flexibility captures, in an intuitive manner, the
dynamics of functional networks. Results revealed essentially a negative correlation between
psychological resilience and EEG-based brain functional network flexibility.
RESULTS
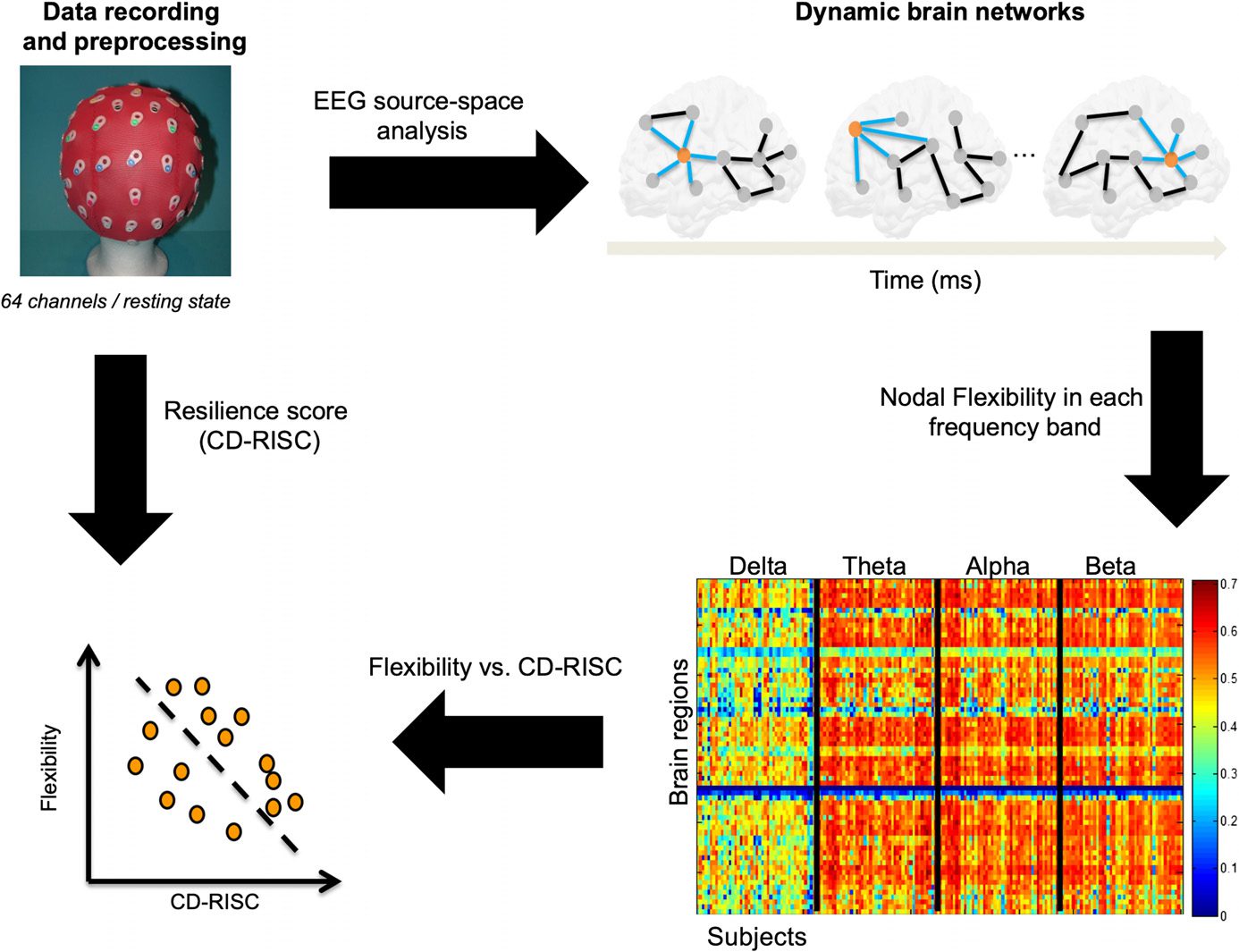
The study is summarized in Figure 1. First, EEG data were recorded and preprocessed. Second,
the dynamic networks were estimated using the EEG source-space approach, giving a set of
brain network at the given time period for each frequency band. Third, the flexibility of each
brain region in each frequency band were computed for each subject. Finally, the correlation
between the brain regions flexibility and the resilience score (CD-RISC) was computed.
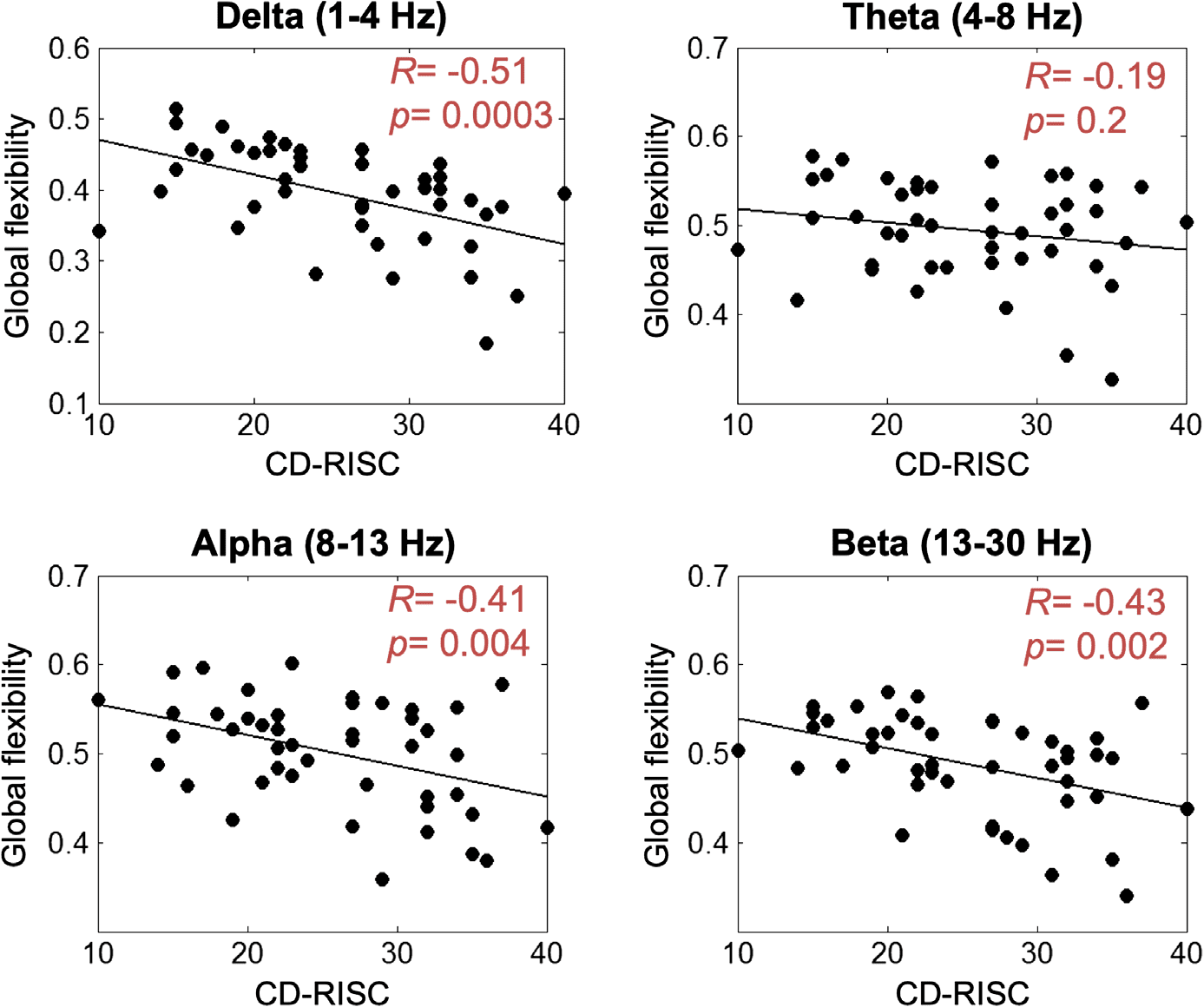
Then, we computed the Pearson correlation between the resilience score and the global
flexibility (averaged over all brain regions for each subject in each frequency band). The corre-
sponding results are presented in Figure 2. For all frequency bands, a negative correlation was
observed. This negative correlation was significant for delta (R = −0.51, p = 0.0003), alpha
(R = −0.41, p = 0.004), and beta (R = −0.43, p = 0.002) bands, but nonsignificant for the
theta band (R = −0.19, p = 0.2).
We then focused on the correlation between flexibility and the resilience score at the level
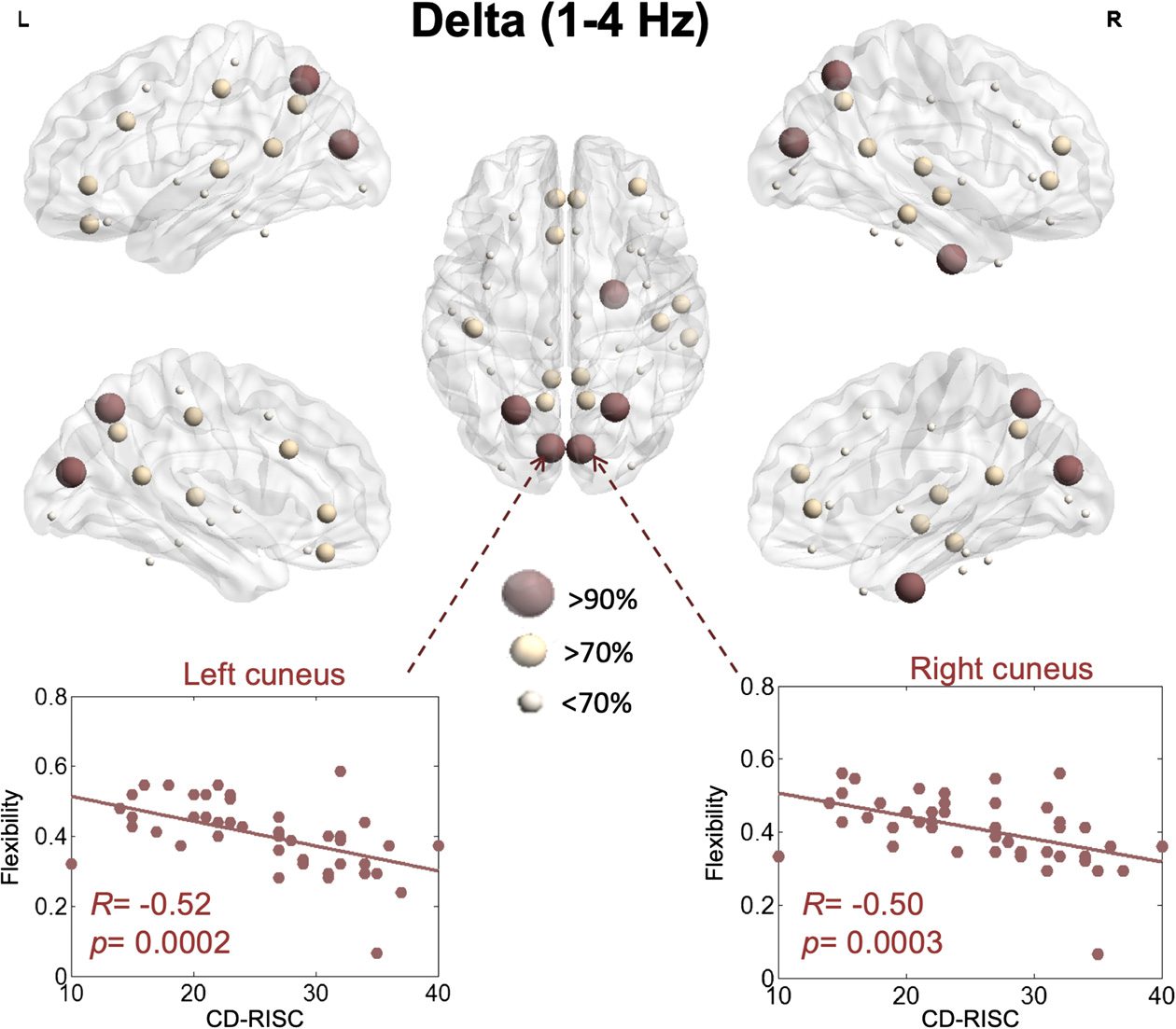
of each brain region. In the delta band, brain regions that were significant and correlated
with the CD-SCORE are illustrated in Figure 3. The flexibility of the left cuneus (R = −0.52,
p = 0.0002, false discovery rate [FDR] corrected), the right cuneus (R = −0.50, p = 0.0004,
FDR corrected), the left superior parietal (R = −0.49, p = 0.0005, FDR corrected), the right
superior parietal (R = −0.49, p = 0.0006, FDR corrected), and the right entorhinal (R = −0.45,
p = 0.0006, FDR corrected) had the highest (>90%) correlation with the resilience score
Network Neuroscience
540
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
2
5
3
9
1
0
9
2
6
4
0
n
e
n
_
a
_
0
0
0
7
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Psychological resilience and functional networks flexibility
Figure 1. Experimental protocol and data analysis pipeline.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
2
5
3
9
1
0
9
2
6
4
0
n
e
n
_
a
_
0
0
0
7
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 2. Correlation between the global flexibility (averaged over all brain regions) and the
CD-RISC score.
Network Neuroscience
541
Psychological resilience and functional networks flexibility
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
Figure 3.
resilience within the delta band.
Illustration of the brain regions for which flexibility is significantly correlated with
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
2
5
3
9
1
0
9
2
6
4
0
n
e
n
_
a
_
0
0
0
7
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
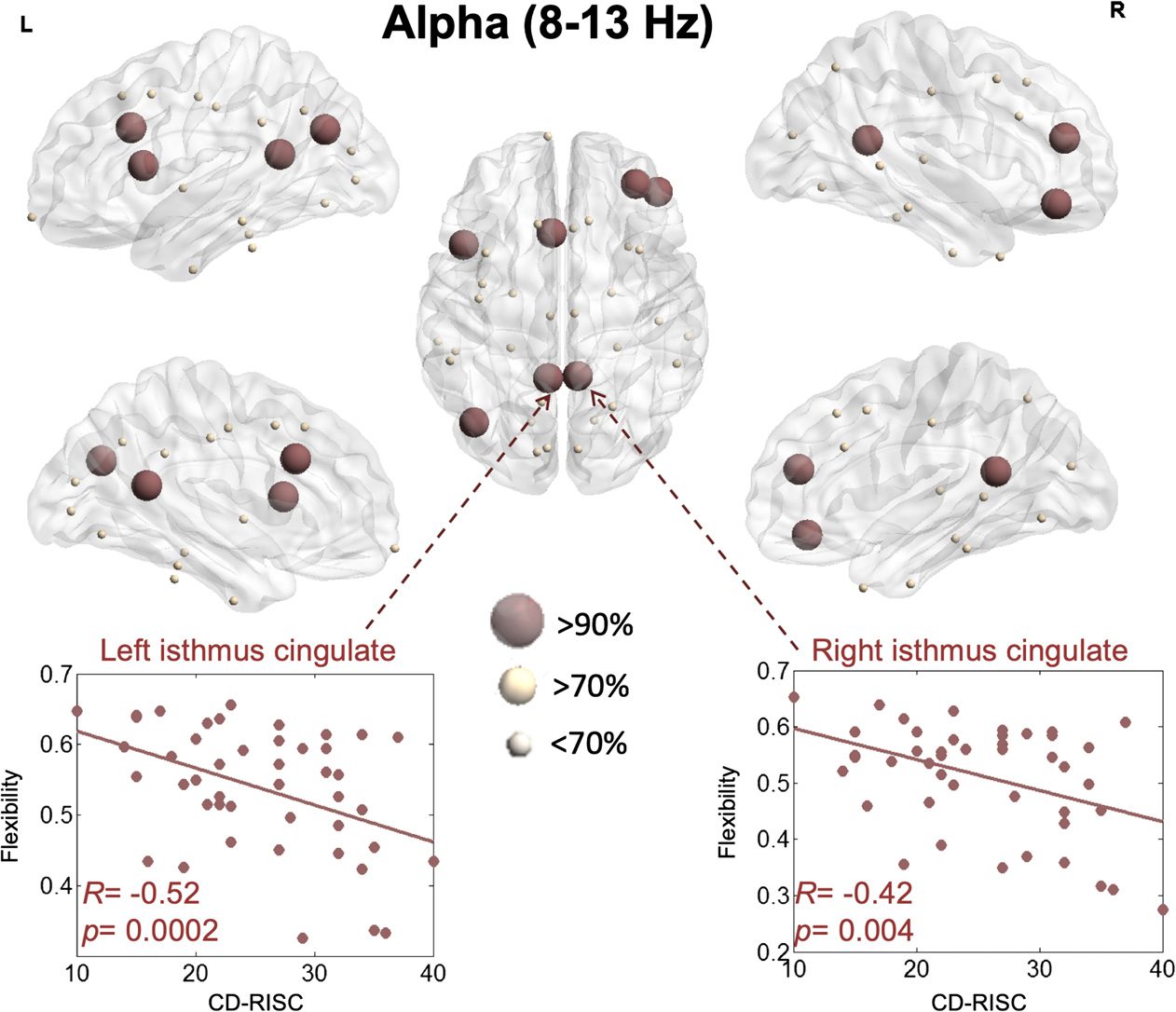
Figure 4.
silience within the alpha band.
Illustration of the brain regions for which flexibility is significantly correlated with re-
Network Neuroscience
542
Psychological resilience and functional networks flexibility
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
2
5
3
9
1
0
9
2
6
4
0
n
e
n
_
a
_
0
0
0
7
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
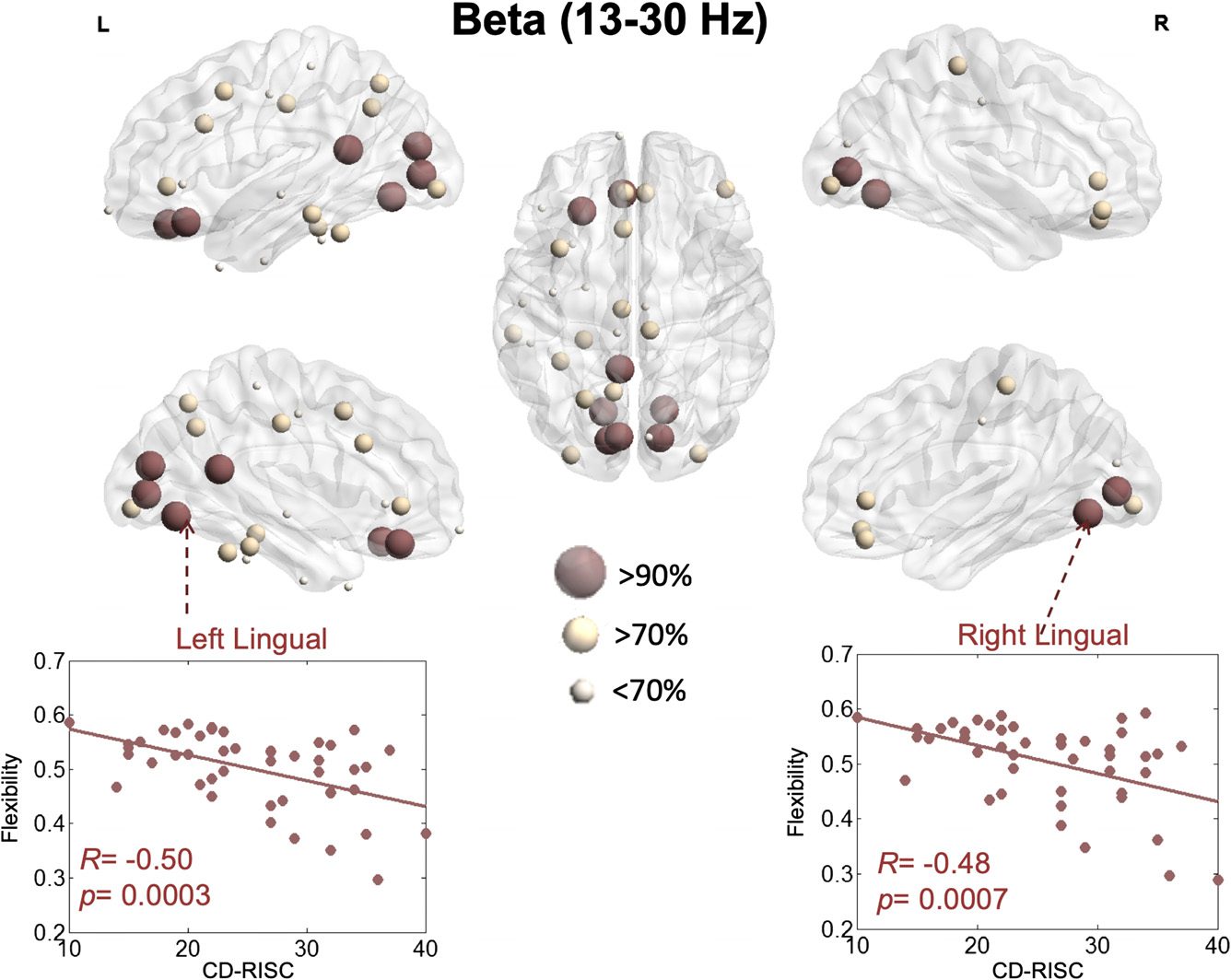
Figure 5.
resilience within the beta band.
Illustration of the brain regions for which flexibility is significantly correlated with
(e.g., the brain regions that had the highest 90% correlation between the resilience score and
the network-based score).
The results for the alpha band are presented in Figure 4. The flexibility of the left caudal
anterior cingulate (R = −0.46, p = 0.001, FDR corrected), the right rostral middle frontal
(R = −0.46, p = 0.001, FDR corrected), the right pars orbitalis (R = −0.45, p = 0.001, FDR
corrected), the left inferior parietal (R = −0.44, p = 0.002, FDR corrected), the left isthmus
cingulate (R = −0.43, p = 0.003, FDR corrected), the right isthmus cingulate (R = −0.42, p =
0.004, FDR corrected), and the right pars opercularis (R = −0.41, p = 0.004, FDR corrected)
showed the highest correlations with the resilience score.
Results regarding the beta band revealed a more pronounced implication of visual networks
(Figure 5). Specifically, the flexibility of the left lingual (R = −0.50, p = 0.0004, FDR corrected),
right lingual (R = −0.48, p = 0.0007, FDR corrected), left pericalcarine (R = −0.48, p =
0.0007, FDR corrected), right pericalcarine (R = −0.47, p = 0.0009, FDR corrected), left
cuneus (R = −0.46, p = 0.001, FDR corrected), left isthmus cingulate (R = −0.46, p = 0.001,
FDR corrected), left medial orbitofrontal (R = −0.45, p = 0.001, FDR corrected), and left
lateral orbitofrontal (R = −0.45, p = 0.001, FDR corrected) had the highest correlation with
the resilience score.
DISCUSSION
Our results reveal a robust and direct relationship between the network flexibility (as quan-
tified through graph theory) of human functional brain networks and resilience (as measured
with neuropsychological testing). This result has been obtained using noninvasive brain re-
cordings (EEG), in the absence of any specific task or stimulus (resting-state recordings), for
a relatively large sample size (N = 45). Specifically, the network flexibility of specific brain
regions is shown to be significantly decreased with a higher resilience score, which can appear
Network Neuroscience
543
Psychological resilience and functional networks flexibility
Rich club:
Subset of cortical regions that are
extensively connected with other
brain regions.
Static network analysis:
Analysis of the brain network
involving some form of averaging
over time.
Dynamic network analysis:
Analysis of the brain network made
using time-dependent estimates of
the network, as opposed to static
network analysis.
nonintuitive: if resilience refers to a capacity to cope with external stress and adapt, it could
imply that brain networks can reconfigure to adapt to these changes. Our results point at the
opposite: most brain regions negatively correlated with the resilience score are part of the “rich
club” (“core” functional network of brain regions [Van Den Heuvel & Sporns, 2011]) and illus-
trate that the brain core network is less flexible in resilient subjects. This suggests that a stable
core network would allow preventing excessive reconfiguration of functional brain networks
following external stress factors, for example. From a fundamental perspective, these results
also suggest that there exists a brain network of critical importance, which underlies one of
the most prominent personality traits (resilience). This raises the possibility to probe further po-
tential brain network correlates of other personality traits in the future. Furthermore, this link
between core network structures and resilience is fundamental, since it can be detected non-
invasively even in resting-state recordings. Another insight from our analysis is that dynamic
but not static network analysis was able to reveal this association between resilience and a
graph theory metric. Static analyses were realized using clustering coefficient and participa-
tion coefficient, and did not reveal any significant correlation with the resilience score (see
Supporting Information Figure S1 and Figure S2; Paban, Modolo, Mheich, & Hassan, 2019).
Therefore, this highlights the importance of the dynamic reconfiguration of brain networks in
shaping behavior. Recent studies have emphasized the interest to characterize the architecture
underlying these dynamics, for example, by identifying the “life duration” and transition rates
of these frequency-specific networks (Vidaurre et al., 2018). Overall, as emphasized in a recent
perspective paper by Tompson et al. (2018), there is a significant potential regarding the use
of network neuroscience tools to unveil the brain networks underlying personality traits, with
flexibility and integration being especially promising.
Neural Substrates of Resilience
Functional neuroimaging studies examining brain networks associated with resilience are lim-
ited. Most of them mainly focused on patient populations, such as depressive, traumatized,
and posttraumatic stress disorders, in which alterations where described in regions of the brain
involved in emotion and stress regulation circuitry (see van der Werff et al., 2013, for review).
In healthy adults, to our knowledge, only one resting-state fMRI (rs-fMRI) experiment has been
reported to date. Interestingly, Kong et al. (2015) reported that psychological resilience had
significant negative correlations with the rs-fMRI signals, but in regions such as the bilateral
insula, right dorsal, and rostral anterior cingulate cortex. Our study did not highlight the same
brain regions. Our dynamic network analysis identified regions belonging to the “core” func-
tional network described by Van Den Heuvel and Sporns (2011). Those regions are supposed
to play a central role in establishing and maintaining efficient global brain communication.
They have been shown to be involved in cognitive processes related to top-down attentional
control (superior parietal cortex [Sestieri et al., 2017]), decision-making (subdivision of the
orbitofrontal cortex [Schuck et al., 2018]), cognitive regulation of behavior (caudal anterior
cingulate [Bush et al., 2000]), and spatial location (entorhinal cortex [Kim & Maguire, 2018]).
Our data also highlighted the regions participating in the visual network (e.g., cuneus, the lin-
gual gyrus, and the pericalcarine cortex). This network oscillated in fast frequency band (beta),
which is thought to reflect different aspects of sensory information processing (Hong et al.,
2008). Another region negatively correlated with the resilience score comprised the inferior
parietal cortex. It has been shown to be implicated in major cognitive functions, including
visuo-spatial attention and visual memory (Egner et al., 2008).
The present study provided evidence of neuronal substrates of resilience. In healthy adults,
it seems to make sense that the ability of individuals to cope actively with life’s challenges
Network Neuroscience
544
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
2
5
3
9
1
0
9
2
6
4
0
n
e
n
_
a
_
0
0
0
7
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Psychological resilience and functional networks flexibility
(Fletcher & Sarkar, 2013) requests brain regions involved in high-level cognitive processes. To
be efficient, these regions should have a modular organization stable over time. As suggested
by Betzel et al. (2016), such network architecture limits information exchange across mod-
ules, which allows a specialized information processing. Such modular organization seemed
to be an important factor in maintaining a stable equilibrium of psychological and physical
functioning when facing adversity, ranging from daily problems to major life events (Bonanno,
2004). When this is not the case, that is, when few brain regions showed high modular affili-
ation exchanges, one may assume that they multiply their contributions and so participate in
a multitude of brain functioning. In this context, subjects had low-resilient scores, meaning
that they exhibit passive coping strategies, high emotional load, ruminative and depressogenic
thinking, and low life satisfaction (Davydov et al., 2010).
Methodological Considerations
First, in the present study we used a template source-space, instead of a subject-specific one,
that is, the same structural MRIs of healthy subjects were used in our EEG functional con-
nectivity analysis. In the case of healthy subjects, it was reported that coregistration with the
template yielded largely consistent connectivity and network estimates compared with native
MRI (Douw et al., 2018). However, in the case of patient-related studies, it is more preferred
to use patient-specific MRIs.
Second, it is important to keep in mind that computing the functional connectivity at the
scalp level is severely corrupted by the volume conduction problem (Brookes et al., 2014;
Lai et al., 2018; Schoffelen & Gross, 2009). M/EEG connectivity analysis at the source-space
was shown to reduce this effect as connectivity is applied to “local” time series generated by
cortical neuronal assemblies modeled, for instance, as current dipole sources. Yet, the “mixing
effects” can also occur in the source-space. To address this issue, a number of methods were de-
veloped based mostly on the rejection of zero-lag correlation. In particular, “unmixing” meth-
ods, called “leakage correction” (such as the orthogonalization approach [Colclough et al.,
2015]), have been reported, which force the reconstructed signals to have zero cross-
correlation at lag zero. Here we preferred to use the phase locking value (PLV) metric and
keep these zero-lag correlations, as several experimental studies reported the importance (in
many conditions) of the zero-lag synchronizations in the human brain. Nevertheless, we be-
lieve that there is no ideal solution yet for this methdological issue and that further efforts are
needed to completely solve the spatial leakage problem.
Third, we have performed the whole analysis in all EEG frequency bands in order to avoid
making any a priori judgments about the involved rhythms. The reported effects were the same
when other thresholds were used. However, it should be kept in mind that standard EEG fre-
quency bands are increasingly associated with specific functional roles in brain-scale infor-
mation processing. As an exemple, alpha activity appears to mediate the so-called “gating by
inhibition” mechanism involved in information routing (Bonnefond et al., 2017) through spe-
cific nodes of the cortical network while inhibiting irrelevant regions. On the other hand, theta
oscillations appear to be involved in locking distant brain regions to enable further processing
(Lisman and Jensen, 2013). Since different frequency bands appear linked with specific func-
tional roles, there is no reason a priori that psychological resilience results from an interplay
between all of those frequency bands. Therefore, we believe that our results point at those os-
cillations that are functionally linked with resilience, and that theta oscillations do not appear
to have a specific role in this case.
Network Neuroscience
545
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
2
5
3
9
1
0
9
2
6
4
0
n
e
n
_
a
_
0
0
0
7
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Psychological resilience and functional networks flexibility
Finally, although EEG source connectivity has improved the spatial resolution of EEG, the
networks identified are, however, still limited to cortical gray matter. In fact, the localization of
subcortical structures remains extremely challenging using EEG technology, namely because
of anatomical and analytical reasons. For example, unlike the layered cortex, a subcortical
region does not have a sufficient organization of pyramidal cells to give rise to localizable
scalp-recorded EEG.
MATERIAL AND METHODS
Participants
A total of 45 healthy subjects were recruited (22 women). The mean age was 34.7 years old
(SD = 9.1 years, range = 18 − 55). Education ranged from 10 years of schooling to a PhD
degree. None of the volunteers reported taking any medication or drugs, nor suffering from
any past or present neurological or psychiatric disease. After receiving approval from the Aix-
Marseille university ethics committee according to the Declaration of Helsinki, participants
filled out the CD-RISC questionnaire at home approximately 1 week before the EEG experi-
ment. Written informed consent was obtained from all participants prior to study onset.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
2
5
3
9
1
0
9
2
6
4
0
n
e
n
_
a
_
0
0
0
7
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Measuring Psychological Resilience
The Connor-Davidson Resilience Scale (CD-RISC; Connor & Davidson, 2003) is a 25-item scale
that measures the ability to cope with adversity. A 10-items version (CD-RISC 10) of this scale
has been developed by Campbell-Sills and Stein (Campbell-Sills & Stein, 2007). A 10-item
version validated for French speaking populations was used in the present study (Guihard et al.,
2018; Scali et al., 2012). The 10 items are rated on a five-point Likert scale that ranges from 0
(not at all) to 4 (true nearly all of the time). Higher scores correspond to greater resilience. This
scale demonstrated good internal consistency and construct validity (Campbell-Sills & Stein,
2007). In our sample, the CD-RISC 10 exhibited a reliability of α = 0.90.
Data Acquisition and Preprocessing
Each EEG session consisted of a 10-min resting period with the participant’s eyes closed (Paban
et al., 2018). Participants were seated in a dimly lit room, were instructed to close their eyes,
and then to simply relax until they were informed that they could open their eyes. Participants
were instructed that the resting period would last approximately 10 min. The eyes-closed rest-
ing EEG recordings protocol was chosen to minimize movement and sensory input effects on
electrical brain activity. EEG data were collected using a 64-channel Biosemi ActiveTwo system
(Biosemi Instruments, Amsterdam, The Netherlands) positioned according to the standard 10–
20 system montage, one electrocardiogram, and two bilateral electro-oculogram electrodes
(EOG) for horizontal movements. Nasion-inion and preauricular anatomical measurements
were made to locate each individual’s vertex site. Electrode impedances were kept below
20 kOhm. EEG signals are frequently contaminated by several sources of artifacts, which were
addressed using the same preprocessing steps as described in several previous studies dealing
with EEG resting-state data (Kabbara et al., 2018, 2017; Rizkallah et al., 2018). Briefly, bad
channels (signals that are either completely flat or contaminated by movement artifacts) were
first identified by visual inspection, complemented by the power spectral density. These bad
channels were then recovered using an interpolation procedure implemented in Brainstorm
(Tadel et al., 2011) by using neighboring electrodes within a 5-cm radius. Epochs with voltage
fluctuations more than +80 μV and less than −80 μV were removed. Five artifact-free epochs
of 40-s lengths were selected for each participant. This epoch length was used in a previous
Network Neuroscience
546
Psychological resilience and functional networks flexibility
study, and was considered as a good compromise between the needed temporal resolution
and the results reproducibility (Kabbara et al., 2017). By using a sliding windows approach
to compute the functional connectivity, a large number of networks (depend on the analyzed
frequency band) were obtained for each 40-s epoch.
Brain Networks Construction
First, brain networks were reconstructed using the “EEG source-space connectivity” method
(Hassan et al., 2014; Hassan & Wendling, 2018), which includes two main steps: (1) recon-
struct the dynamics of the cortical sources by solving the inverse problem, and (2) measure the
statistical couplings (functional connectivity) between the reconstructed time series. In sum-
mary, EEG source connectivity links the recorded EEG signals with the functional relationship
between anatomical brain regions (e.g., networks), through the EEG inverse problem that pro-
vides the localization of the cortical sources originating these EEG signals. Let us mention that
numerous methods exist for both of these two steps (EEG inverse problem and functional con-
nectivity measures). EEGs and MRI template (ICBM152) were coregistered through the identi-
fication of anatomical landmarks by using Brainstorm (Tadel et al., 2011). A Desikan-Killiany
atlas-based segmentation approach was used, consisting of 68 cortical regions (Desikan et al.,
2006). The OpenMEEG (Gramfort et al., 2010) software was used to compute the head model.
Here, we used the weighted minimum norm estimate (wMNE) algorithm as an inverse solution.
The reconstructed regional time series were filtered in different frequency bands (delta, 1–4 Hz;
theta, 4–8 Hz; alpha, 8–13 Hz; and beta, 13–30 Hz). For each frequency band, functional
connectivity was computed between the regional time series using the PLV measure (Lachaux
et al., 1999). This combination wMNE/PLV was chosen according to a recent model-based
comparative study of different inverse/connectivity combinations (Hassan et al., 2017)
Using PLV, dynamic functional connectivity matrices were computed for each epoch using
a sliding windows technique. It consists of moving a time window of certain duration δ, and
PLV is calculated within each window. As recommended in Lachaux et al. (2000), we selected
the smallest window length equal to
central frequency , where 6 is the number of “cycles” at the
given frequency band. For instance, in the theta band, since the central frequency (Cf) equals
to 6 Hz, δ equals 1 s, and δ = 279 ms (Cf = 21.5 Hz) in beta band. Thus, for each epoch,
33 networks were obtained for the theta band, and 130 networks in the beta band. The same
calculation was adopted for other frequency bands. Finally, we kept only the strongest 10% of
connections.
6
Network Modularity and Flexibility
The obtained dynamic matrices were divided into time-dependent modules by using the multi-
slice community detection approach described in Mucha et al. (2010). It consists of introducing
a parameter that associates nodes across time, before applying the modularity procedure, and
is defined as (one modularity value is computed for each frequency band of interest; Sporns &
Betzel, 2016):
Q (γ, ω) =
(cid:2)
1
2μ ∑
ijsr
(aijs − γs pijs) + δ(σis, σjs) + δ(i, j). ωjrs
(cid:3)
δ(σis, σjr)
where nodes i and j are assigned to modules σis and σjs in window s, respectively. Aijs repre-
sents the weight of the edge between these two nodes in window s; γ is the structural resolution
parameter; ω is called the “inter-layer coupling strength,” and links the same node j within the
network evaluated at two different times (“layers”): therefore, ωjrs links the node j at times
Network Neuroscience
547
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
2
5
3
9
1
0
9
2
6
4
0
n
e
n
_
a
_
0
0
0
7
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Psychological resilience and functional networks flexibility
r and s, that is, in layer r and in layer s. Here, we chose γ = ωjrs = 1; pijs represents the
expected number of links according to a null model. As a reminder, the Kronecker δ-function
is such that δ(x, y) is 1 if x = y, and 0 otherwise.
The multilayer network modularity was computed 100 times since Q may vary from run
to run (degeneracy). This step is mandatory precisely because the degeneracy intrinsic to the
community detection algorithm results in different networks structures depending on the run.
Classically, this issue is dealt with using the so-called “co-classification matrix” or “consensus
matrix,” whose elements indicate the ratio of each region to be in the same module with the
other regions among these 100 partitions. Only elements in the consensus matrix higher than
an appropriate random null model were considered. The randomized networks (generated with
the null model) have the same strength distribution as the true brain networks. To quantify the
dynamics of brain networks, we used the flexibility metric proposed in Bassett et al. (2011).
The flexibility of a brain region is defined as the number of times that a brain region changed
modular assignment throughout the session, normalized by the total number of changes that
were possible.
Statistical Analysis
We assessed the correlation between the two conditions (resilience score vs. network were
assessed using the Pearson correlation). We applied a FDR correction for multiple comparisons
across regions (Benjamini & Hochberg, 1995).
Software
The functional connectivity, network measures, and network visualization were performed
using BCT (Rubinov & Sporns, 2010), EEGNET (Hassan et al., 2015), and BrainNet viewer (Xia
et al., 2013), respectively. The Network Community Toolbox (http://commdetect.weebly.com/)
was used to compute the consensus matrices as well as the values provided by the flexibility
metrics.
AUTHOR CONTRIBUTIONS
Véronique Paban: Conceptualization; Data curation; Funding acquisition; Methodology;
Project administration; Resources; Writing – original draft. Julien Modolo: Formal analysis;
Investigation; Visualization; Writing – review & editing. Ahmad Mheich: Data curation; Formal
analysis; Software; Visualization. Mahmoud Hassan: Data curation; Formal analysis; Method-
ology; Software; Visualization; Writing – review & editing.
REFERENCES
Bassett, D. S., & Sporns, O. (2017). Network neuroscience. Nature
Neuroscience, 20, 353.
Bassett, D. S., Wymbs, N. F., Porter, M. A., Mucha, P. J., Carlson,
J. M., & Grafton, S. T. (2011). Dynamic reconfiguration of hu-
man brain networks during learning. Proceedings of the National
Academy of Sciences of the United States of America, 108(18),
7641–7646.
Betzel, R. F., Fukushima, M., He, Y., Zuo, X.-N., & Sporns, O.
(2016). Dynamic fluctuations coincide with periods of high and
low modularity in resting-state functional brain networks. Neuro-
Image, 127, 287–297.
Bonanno, G. A. (2004). Loss, trauma, and human resilience: Have
we underestimated the human capacity to thrive after extremely
aversive events? American Psychologist, 59, 20.
Benjamini, Y., & Hochberg, Y. (1995). Controlling the false discov-
ery rate: A practical and powerful approach to multiple testing.
Journal of the Royal Statistical Society B, 57(1), 289–300.
Bonnefond, M., Kastner, S., & Jensen, O. J. E. (2017). Communica-
tion between brain areas based on nested oscillations. ENEURO,
0153–0116.2017.
Network Neuroscience
548
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
2
5
3
9
1
0
9
2
6
4
0
n
e
n
_
a
_
0
0
0
7
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Psychological resilience and functional networks flexibility
Brookes, M. J., Woolrich, M. W., & Price, D. (2014). An introduc-
tion to MEG connectivity measurements. In Magnetoencephalog-
raphy (pp. 321–358). Berlin: Springer.
Bush, G., Luu, P., & Posner, M. I. (2000). Cognitive and emo-
tional influences in anterior cingulate cortex. Trends in Cognitive
Sciences, 4, 215–222.
Campbell-Sills, L., & Stein, M. B. (2007). Psychometric analysis and
refinement of the Connor-Davidson Resilience Scale (CD-RISC):
Validation of a 10-item measure of resilience. Journal of Trau-
matic Stress, 20, 1019–1028.
Colclough, G., Brookes, M., Smith, S., & Woolrich, M. (2015).
A symmetric multivariate leakage correction for MEG connec-
tomes. NeuroImage, 117, 439–448.
Connor, K. M., & Davidson, J. R. (2003). Development of a new re-
silience scale: The Connor-Davidson Resilience Scale (CD-RISC).
Depression and Anxiety, 18, 76–82.
Davydov, D. M., Stewart, R., Ritchie, K., & Chaudieu, I. (2010).
Resilience and mental health. Clinical Psychology Review, 30,
479–495.
Desikan, R. S., Ségonne, F., Fischl, B., Quinn, B. T., Dickerson,
B. C., Blacker, D., Buckner, R. L., Dale, A. M., Maguire, R. P.,
Hyman, B. T., Albert, M. S., & Killiany, R. J. (2006). An auto-
mated labeling system for subdividing the human cerebral cortex
on MRI scans into gyral based regions of interest. NeuroImage,
31, 968–980.
Douw, L., Nieboer, D., Stam, C. J., Tewarie, P., & Hillebrand,
A. (2018). Consistency of magnetoencephalographic functional
connectivity and network reconstruction using a template ver-
sus native MRI for co-registration. Human Brain Mapping, 39,
104–119.
Dubois, J., Galdi, P., Han, Y., Paul, L. K., & Adolphs, R. (2018).
Resting-state functional brain connectivity best predicts the
personality dimension of openness to experience. Personality
Neuroscience, 1:e6.
Egner, T., Monti, J. M., Trittschuh, E. H., Wieneke, C. A., Hirsch, J., &
Mesulam, M.-M. (2008). Neural integration of top-down spatial
and feature-based information in visual search. Journal of Neuro-
science, 28, 6141–6151.
Fletcher, D., & Sarkar, M. (2013). Psychological resilience: A review
and critique of definitions, concepts, and theory. European Psy-
chologist, 18, 12.
Gramfort, A., Papadopoulo, T., Olivi, E., & Clerc, M. (2010). Open-
MEEG: Opensource software for quasistatic bioelectromagnetics.
Biomedical Engineering Online, 9, 45.
Guihard, G., Deumier, L., Alliot-Licht, B., Bouton-Kelly, L., Michaut,
C., & Quilliot, F. (2018). Psychometric validation of the French
version of the Connor-Davidson Resilience Scale. L’Encéphale,
44, 40–45.
Hassan, M., Dufor, O., Merlet, I., Berrou, C., & Wendling, F. (2014).
EEG source connectivity analysis: From dense array recordings to
brain networks. PloS One, 9, e105041.
Hassan, M., Merlet, I., Mheich, A., Kabbara, A., Biraben, A., Nica,
A., & Wendling, F. (2017). Identification of interictal epileptic
networks from Dense-EEG. Brain Topography, 30, 60–76.
Hassan, M., Shamas, M., Khalil, M., El Falou, W., & Wendling, F.
(2015). EEGNET: An open source tool for analyzing and visual-
izing M/EEG connectome. PloS One, 10, e0138297.
Hassan, M., & Wendling, F. (2018). Electroencephalography source
connectivity: Aiming for high resolution of brain networks in time
and space. IEEE Signal Processing Magazine, 35, 81–96.
Hong, L. E., Summerfelt, A., Mitchell, B. D., McMahon, R. P.,
Wonodi, I., Buchanan, R. W., & Thaker, G. K. (2008). Sensory
gating endophenotype based on its neural oscillatory pattern
and heritability estimate. Archives of General Psychiatry, 65,
1008–1016.
Kabbara, A., Eid, H., El Falou, W., Khalil, M., Wendling, F., &
Hassan, M. (2018). Reduced integration and improved segrega-
tion of functional brain networks in Alzheimer’s disease. Journal
of Neural Engineering, 15, 026023.
Kabbara, A., Falou, W. E., Khalil, M., Wendling, F., & Hassan, M.
(2017). The dynamic functional core network of the human brain
at rest. Scientific Reports, 7, 2936.
Kim, M., & Maguire, E. A. (2018). Thalamus, subiculum and retro-
splenial cortex encode 3D head direction information in volu-
metric space. bioRxiv, 335976.
Kong, F., Wang, X., Hu, S., & Liu, J. (2015). Neural correlates of
psychological resilience and their relation to life satisfaction in a
sample of healthy young adults. NeuroImage, 123, 165–172.
Lachaux, J. P., Rodriguez, E., Martinerie, J., & Varela, F. J. (1999).
Measuring phase synchrony in brain signals. Human Brain Mapping,
8, 194–208.
Lachaux,
J.-P., Rodriguez, E., Le Van Quyen, M., Lutz, A.,
Martinerie, J., & Varela, F. J. (2000). Studying single-trials of
phase synchronous activity in the brain. International Journal of
Bifurcation and Chaos, 10, 2429–2439.
Lai, M., Demuru, M., Hillebrand, A., & Fraschini, M. (2018). A com-
parison between scalp- and source-reconstructed EEG networks.
Scientific Reports, 8, 12269.
Lisman, J. E., & Jensen, O. J. N. (2013). The theta-gamma neural
code. Neuron, 77(6), 1002–1016.
Liu, J. J., Reed, M., & Girard, T. A. (2017). Advancing resilience:
An integrative, multi-system model of resilience. Personality and
Individual Differences, 111, 111–118.
Luthar, S. S. (2003). Resilience and Vulnerability: Adaptation in the
Context of Childhood Adversities. Cambridge, UK: Cambridge
University Press.
Markett, S., Montag, C., & Reuter, M. (2018). Network neuroscience
and personality. Personality Neuroscience, 1.
Mheich, A., Hassan, M., Khalil, M., Gripon, V., Dufor, O., &
Wendling, F. (2018). SimiNet: A novel method for quantifying
brain network similarity. IEEE Transactions on Pattern Analysis
and Machine Intelligence, 40, 2238–2249.
Mucha, P. J., Richardson, T., Macon, K., Porter, M. A., & Onnela,
J.-P. (2010). Community structure in time-dependent, multiscale,
and multiplex networks. Science, 328, 876–878.
Paban, V., Deshayes, C., Ferrer, M.-H., Weill, A., & Alescio-Lautier,
B. (2018). Resting brain functional networks and trait coping.
Brain Connectivity, 8(8), 475–486.
Paban, V., Modolo, J., Mheich, A., & Hassan, M. (2019). Support-
ing Information for “Psychological resilience correlates with EEG
source-space brain network flexibility.” Network Neuroscience,
3(2), 539–550. https://doi.org/10.1162/netn_a_00079
Reynaud, E., Guedj, E., Souville, M., Trousselard, M., Zendjidjian,
X., El Khoury-Malhame, . . . Canini, F. (2013). Relationship
Network Neuroscience
549
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
2
5
3
9
1
0
9
2
6
4
0
n
e
n
_
a
_
0
0
0
7
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Psychological resilience and functional networks flexibility
between emotional experience and resilience: An fMRI study in
fire-fighters. Neuropsychologia, 51, 845–849.
Rizkallah, J., Benquet, P., Kabbara, A., Dufor, O., Wendling, F.,
& Hassan, M. (2018). Dynamic reshaping of functional brain
networks during visual object recognition. Journal of Neural
Engineering, 15(5).
Rubinov, M., & Sporns, O. (2010). Complex network measures of
brain connectivity: Uses and interpretations. NeuroImage, 52,
1059–1069.
Rutter, M. (2006). Implications of resilience concepts for scientific
understanding. Annals of the New York Academy of Sciences,
1094, 1–12.
Scali, J., Gandubert, C., Ritchie, K., Soulier, M., Ancelin, M.-L.,
& Chaudieu, I. (2012). Measuring resilience in adult women
using the 10-items Connor-Davidson Resilience Scale (CD-RISC).
Role of trauma exposure and anxiety disorders. PloS One, 7,
e39879.
Schoffelen, J. M., & Gross, J. (2009). Source connectivity analysis
with MEG and EEG. Human Brain Mapping, 30, 1857–1865.
Schuck, N. W., Wilson, R., & Niv, Y. (2018). A state representation
for reinforcement learning and decision-making in the orbito-
frontal cortex. In Goal-Directed Decision Making (pp. 259–278).
Amsterdam: Elsevier.
Sestieri, C., Shulman, G. L., & Corbetta, M. (2017). The contribu-
tion of the human posterior parietal cortex to episodic memory.
Nature Reviews Neuroscience, 18, 183.
Sporns, O., & Betzel, R. F. (2016). Modular brain networks. Annual
Review of Psychology, 67, 613–640.
Tadel, F., Baillet, S., Mosher, J. C., Pantazis, D., & Leahy, R. M.
(2011). Brainstorm: A user-friendly application for MEG/EEG
analysis. Computational Intelligence and Neuroscience, 2011, 8.
Tompson, S., Falk, E. B., Vettel, J. M., & Bassett, D. S. (2018).
Network approaches to understand individual differences in
brain connectivity: Opportunities for personality neuroscience.
Personality Neuroscience, 1:e5.
Van Den Heuvel, M. P., & Sporns, O. (2011). Rich-club organiza-
tion of the human connectome. Journal of Neuroscience, 31,
15775–15786.
van der Werff, S.
J. N.,
Elzinga, B. M., & Van Der Wee, N. J. (2013). Neuroimaging
resilience to stress: A review. Frontiers in Behavioral Neuro-
science, 7, 39.
J., van den Berg, S. M., Pannekoek,
Vidaurre, D., Hunt, L. T., Quinn, A. J., Hunt, B. A., Brookes, M. J.,
Nobre, A. C., & Woolrich, M. W. (2018). Spontaneous corti-
cal activity transiently organises into frequency specific phase-
coupling networks. Nature Communications, 9, 2987.
Waugh, C. E., & Koster, E. H. (2015). A resilience framework for
promoting stable remission from depression. Clinical Psychology
Review, 41, 49–60.
Xia, M., Wang, J., & He, Y. (2013). BrainNet Viewer: A network
visualization tool for human brain connectomics. PloS One, 8,
e68910.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
2
5
3
9
1
0
9
2
6
4
0
n
e
n
_
a
_
0
0
0
7
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network Neuroscience
550