RESEARCH
Accelerated intermittent theta burst stimulation
in major depression induces decreases in
modularity: A connectome analysis
Karen Caeyenberghs1, Romain Duprat2, Alexander Leemans3, Hadi Hosseini4,
Peter H. Wilson1, Debby Klooster5,6, and Chris Baeken7
1School of Psychology, Faculty of Health Sciences, Australian Catholic University, Sydney, Australia
2Department of Psychiatry and Medical Psychology, Ghent University, Ghent, Belgium
3Image Sciences Institute, University Medical Center Utrecht, Utrecht, The Netherlands
4Department of Psychiatry and Behavioral Sciences, School of Medicine, Stanford University, Stanford, CA, USA
5Eindhoven University of Technology, Department of Electrical Engineering, Eindhoven, The Netherlands
6Academic Center for Epileptology Kempenhaeghe, Heeze, The Netherlands
7Department of Psychiatry and Medical Psychology, Ghent University, Ghent, Belgium; Department of Psychiatry, University
Hospital UZBrussel, Brussels, Belgium; and Ghent Experimental Psychiatry (GHEP) Lab, Ghent, Belgium
Keywords: Structural connectivity, Brain stimulation, Depression, Diffusion MRI, graph theory
ABSTRACT
Accelerated intermittent theta burst stimulation (aiTBS) is a noninvasive neurostimulation
technique that shows promise for improving clinical outcome in patients suffering from
treatment-resistant depression (TRD). Although it has been suggested that aiTBS may evoke
beneficial neuroplasticity effects in neuronal circuits, the effects of aiTBS on brain networks
have not been investigated until now. Fifty TRD patients were enrolled in a randomized
double-blind sham-controlled crossover trial involving aiTBS, applied to the left dorsolateral
prefrontal cortex. Diffusion-weighted MRI data were acquired at each of three time points
(T1 at baseline; T2 after the first week of real/sham aiTBS stimulation; and T3 after the second
week of treatment). Graph analysis was performed on the structural connectivity to examine
treatment-related changes in the organization of brain networks. Changes in depression
severity were assessed using the Hamilton Depression Rating Scale (HDRS). Baseline data
were compared with 60 healthy controls. We observed a significant reduction in depression
symptoms over time (p < 0.001). At T1, both TRD patients and controls exhibited a
small-world topology in their white matter networks. More importantly, the TRD patients
demonstrated a significantly shorter normalized path length (pAUC = 0.01), and decreased
assortativity (pAUC = 0.035) of the structural networks, compared with the healthy control
group. Within the TRD group, graph analysis revealed a less modular network configuration
between T1 and T2 in the TRD group who received real aiTBS stimulation in the first week
(p < 0.013). Finally, there were no significant correlations between changes on HDRS scores
and reduced modularity. Application of aiTBS in TRD is characterized by reduced
modularity, already evident 4 days after treatment. These findings support the potential
clinical application of such noninvasive brain stimulation in TRD.
AUTHOR SUMMARY
Accelerated noninvasive neurostimulation has shown promise to rapidly improve clinical
symptoms in patients suffering from treatment-resistant depression. However, the stimulation
effects on brain networks have not been well investigated but may be necessary to improve
clinical outcome. To examine treatment-related changes in the organization of brain
networks, graph analysis was performed on structural connectivity in 50 treatment-resistant
a n o p e n a c c e s s
j o u r n a l
Citation: Caeyenberghs, K., Duprat, R.,
Leemans, A., Hosseini, H., Wilson,
P. H., Klooster, D., & Baeken, C. (2019).
Accelerated intermittent theta burst
stimulation in major depression
induces decreases in modularity:
A connectome analysis. Network
Neuroscience, 3 (1), 157–172.
https://doi.org/10.1162/netn_a_00060
DOI:
https://doi.org/10.1162/netn_a_00060
Supporting Information:
https://doi.org/10.1162/netn_a_00060
Received: 7 December 2017
Accepted: 11 May 2018
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Author:
Karen Caeyenberghs
karen.caeyenberghs@acu.edu.au
Handling Editor:
Xi-Nian Zuo
Copyright: © 2018
Massachusetts Institute of Technology
Published under a Creative Commons
Attribution 4.0 International
(CC BY 4.0) license
The MIT Press
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
1
1
5
7
1
0
9
2
3
4
4
n
e
n
_
a
_
0
0
0
6
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Connectome analysis in major depression
Treatment-resistant depression:
Depressed patients who do not
respond to mainstream treatment,
e.g., medication or therapy.
Accelerated intermittent theta
burst stimulation:
Noninvasive neurostimulation
technique that uses repeated levels
of high-frequency stimulation to
excitate neurons.
depressed patients which underwent such a stimulation protocol. Compared to nondepressed
individuals, depressed patients displayed less structural integration, especially in more distal
networks of the brain. More densely interconnected regions, especially when actively
stimulated, may be of essence to explain the clinical improvement, already present after
4 days of accelerated neurostimulation.
INTRODUCTION
Major depressive disorder (MDD) is a worldwide mental health problem (WHO) and is charac-
terized by affective, cognitive, and somatic symptoms impeding the daily life and activities of
the patient. MDD typically manifests as a chronic condition, characterized by a relapsing/
remitting course and by severe impairment that persists even during periods of remission
(Conradi et al., 2011). Moreover, 20–30% of patients with MDD fail to respond to anti-
depressant medication and/or psychotherapy, a condition referred to as treatment-resistant de-
pression (TRD) (Vieta & Colom, 2011; van Randenborgh et al., 2012; Trevino et al., 2014).
As such, TRD is associated with significantly greater medical costs and productivity loss than
treatment-responsive forms, highlighting the need for more effective strategies.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
1
1
5
7
1
0
9
2
3
4
4
n
e
n
_
a
_
0
0
0
6
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
theta burst
Noninvasive brain stimulation methods have shown promise for improving clinical out-
comes in patients suffering from TRD (for reviews, see Lefaucheur et al., 2014; Brunoni et al.,
2017). The majority of studies using repetitive transcranial magnetic techniques (rTMS) for clin-
ically depressed patients have already shown that a series of daily sessions of high-frequency
(HF)-rTMS delivered on the left dorsolateral prefrontal cortex (DLPFC) or low-frequency
(LF)-rTMS applied to the right DLPFC are effective in reducing depressive symptoms (Lefaucheur
et al., 2014). Furthermore, it has been stated that theta burst stimulation (TBS)—a specific rTMS
protocol that uses bursts of high-frequency stimulation at repeated intervals—may result in
superior clinical outcomes (Huang et al., 2005; Plewnia et al., 2014; Li et al., 2014). More
recent studies have examined whether accelerated stimulation paradigms can not only yield
higher response rates but also reduce the total time of treatment, with promising results
(Holtzheimer et al., 2010; Baeken et al., 2013) For example, in our recent randomized, sham-
controlled crossover accelerated intermittent
(aiTBS) study in TRD patients
(Desmyter et al., 2016; Duprat et al., 2016), we showed significant (acute) reductions in depres-
sion severity symptoms and suicide ideation. Despite these promising results, the underlying
neurobiological mechanisms supporting these treatment-related changes remain unclear. In-
sight into these mechanisms may inform our understanding of the neurobiological bases of
depression and our ability to target those brain systems to optimize treatment. Indeed, brain
stimulation methods hold promise in selectively modulating the activity of neuronal networks
that may be implicated in depression, improving clinical outcomes (Huang et al., 2005; Sale
et al., 2015).
In an influential resting-state functional connectivity paper, Fox et al. ( 2012)
showed specific connectivity patterns in relation to clinical rTMS treatment outcome between
the (subgenual) anterior cingulate and prefrontal cortices. Globally, these observations have
been replicated by others, stimulating the left DLPFC (Baeken et al., 2014, 2017; Liston et al.,
2014; Philip et al., 2018) as well as targeting the dorsomedial prefrontal cortex (DMPFC)
(Salomons et al., 2014). Furthermore, stimulating the DLPFC causes network-specific increases
in functional connectivity in similar regions also in healthy individuals (Tik et al., 2017). In an
effort to personalize rTMS treatment, Drysdale et al. ( 2017) defined four neurophysiological de-
pression subtypes (“biotypes”) characterized by distinct patterns of dysfunctional connectivity
in limbic and frontostriatal networks, responsive or not to DMPFC rTMS treatment. Although
functional connectivity alterations are associated with the pathophysiology of MDD, future
Network Neuroscience
158
Connectome analysis in major depression
Connectome:
Map of neural connections in
the brain.
research is needed to investigate how changes in such abnormal patterns of fluctuating com-
munication may contribute to successful treatment of this severe psychiatric illness (Brakowski
et al., 2017; Kaiser et al., 2015).
A connectome framework may further improve our understanding of the biological mech-
anisms of therapeutic effects. Recently, graph theoretical analyses of structural and functional
brain connectivity in humans have contributed to new conceptualizations of the pathogenesis
of MDD (for a review, see Gong & He, 2015). For example, using resting-state fMRI, Zhang
et al. ( 2011) found that the drug-naive, first-episode MDD patients (N = 30) showed lower
path length, higher global efficiency, and increased nodal centralities. In another functional
connectome study, Chen et al. ( 2017) found decreased clustering coefficient, local efficiency,
and transitivity in MDD patients (N = 16). The results of Guo et al. ( 2012) revealed abnormal
nodal centralities in resting-state functional brain networks of 38 MDD patients compared with
healthy controls. Using gray matter covariance networks, Singh et al. ( 2013) demonstrated sig-
nificantly decreased clustering coefficient and nodal alterations in patients with MDD (N = 93)
compared with healthy controls. Twenty-nine MDD participants showed changes (mainly in
cognitive-emotional circuitry and fronto-parietal circuitry) in eigenvector centrality, local clus-
tering coefficient, and nodal efficiency in the study by Qin et al. ( 2014) using white matter
networks. In early stage MDD patients, a significant decrease in small-worldness and a signif-
icantly decreased strength in the frontal-subcortical and limbic regions was found by Lu et al.
( 2017). Korgaonkar et al. ( 2014) found no significant group differences for the graph theory
measures, despite the fact that their network-based statistics revealed lowered structural con-
nectivity in two subnetworks in a cohort of 95 MDD outpatients. In sum, these connectome
studies have revealed that patients with MDD are associated with anomalies in the topological
organization of large-scale functional and structural brain networks (involving global integra-
tion, local segregation, modular structure, and network hubs).
To date, only a few studies have examined the effects of stimulation on brain networks in
humans, but these studies mainly used transcranial direct-current stimulation or deep brain
stimulation treatments. Specifically, two studies have used a connectome approach to exam-
ine the effects of brain stimulation by using transcranial direct-current stimulation in healthy
volunteers (Polanía et al., 2011; Peña-Gómez et al., 2012), albeit studies of functional
connectivity. In addition, a recent structural connectivity study in 11 TRD patients (Riva-Posse
et al., 2017) examined the effects of subcallosal cingulate deep brain stimulation. Their results
supported the advantage of using an individualized tractography map that is based on a group
“connectome blueprint” of past responders to prospectively identify the implantation target,
surpassing traditional approaches that rely on anatomical landmarks or stereotactic coordi-
nates. However, their connectome analysis included only four white matter bundles (i.e., for-
ceps minor, uncinate fasciculus, cingulum, and fronto-striatal fibers). Furthermore, whether
and how aiTBS might produce measurable and durable changes in structural brain integra-
tion and segregation in TRD is unclear. Building on our previous clinical studies of aiTBS
(Desmyter et al., 2016; Duprat et al., 2016), our current exploratory study examined whether
an aiTBS protocol could induce changes in the organizational properties of brain networks
and whether such changes are associated with amelioration of depressive symptoms.
METHODS
Participants
This study was part of a larger project investigating the effects of aiTBS on depressive symptoms
and suicide risk (http://clinicaltrials.gov/show/NCT01832805). The study was carried out in
Network Neuroscience
159
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
1
1
5
7
1
0
9
2
3
4
4
n
e
n
_
a
_
0
0
0
6
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Connectome analysis in major depression
accordance with the principles of the Declaration of Helsinki and approved by the local ethics
committee of the University Hospital Ghent. Written consent was obtained from all subjects.
In the present study, a total of 106 adults aged 18 to 65 years (mean age = 39.9 years,
SD = 12.2 years, 40 men and 66 women) were included. Sixty healthy controls (mean age =
38.6 years, SD = 12.5 years, 26 men) were recruited from the general population with flyers.
Volunteers received payment for their participation. By using the structured Mini-International
Neuropsychiatric Interview (MINI), 46 patients were diagnosed with major depression (mean
age = 41.6 years, SD = 11.7 years, 14 men and 32 women); the MINI is a short, accurate
structured interview for DSM-IV and ICD-10 psychiatric disorders for clinical trials. Patients
were at least stage I therapy-resistant depressed according to the Thase and Rush staging model
(Rush, Thase, & Dubé, 2003), that is, patients had not responded to at least one antidepres-
sant pharmacotherapy trial. Participants with contraindications to MRI scanning (e.g., ferrous
implant, claustrophobia, and pacemaker), bipolar or psychotic symptoms, history of epilep-
tic insult, cerebral surgery, alcohol dependence, or a suicidal attempt within 6 months were
excluded. Antidepressant and antipsychotic medication and mood stabilizers were gradually
tapered off and fully stopped 2 weeks before the start and during the whole period of the aiTBS
treatment. Healthy volunteers were free of mental diagnosis (also assessed with the MINI) and
any psychotropic agent.
Brain Stimulation Protocol
All patients were enrolled in a randomized, double-blind, sham-controlled crossover study
(for an overview of the design, see Supporting Information Figure S1, Caeyenberghs, Duprat,
Leemans, Hosseini, Wilson, Klooster, & Chris Baeken, 2019). Patients were randomly allocated
to two groups: during the first week, one group received the active (verum) stimulation and
the other group started with the sham condition. The treatment conditions (sham, real) were
reversed during the second week. The healthy control group only underwent baseline mea-
surements and did not receive any stimulation. The interested reader is also referred to our
previous studies (Desmyter et al., 2016; Duprat et al., 2016).
Intermittent TBS stimulation was applied using a Magstim Rapid2 Plus1 magnetic stimula-
tor (Magstim Company Limited, Wales, UK) with a figure-of-eight-shaped coil. A stimulation
intensity of 110% of the patient’s resting motor threshold was administered during treatment.
We used the Brainsight neuronavigation system (BrainsightTM, Rogue Research) to identify the
site of stimulation (i.e., the center part of the midprefrontal gyrus [Brodmann 9/46]) based on
the anatomical MRI scan of each individual in order to accurately target the left dorsolateral
prefrontal cortex (DLPFC).
aiTBS was delivered at five sessions per day during 4 days. Between the daily sessions
there was a pause of approximately 15 min. Each aiTBS session consisted of 54 trains of 10
the bursts were
bursts of three stimuli. These stimuli were applied in a 50 Hz frequency:
repeated every 200 ms. This resulted in 2 s of stimulation with a cycling period of 8 s, yielding
1,620 stimuli per session. With a total of 20 sessions, this yielded a sum of 32,400 stimuli per
complete treatment. For the sham condition, a specially designed sham coil, identical in form
and sound to the active coil but without delivering any active stimulation, was placed on the
same target site. The aiTBS administrators could not be blinded to assignment as the physical
coils needed to be changed. Throughout the whole treatment (aiTBS and sham), patients were
blindfolded, wore earplugs, and were kept unaware of the type of stimulation.
Network Neuroscience
160
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
1
1
5
7
1
0
9
2
3
4
4
n
e
n
_
a
_
0
0
0
6
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Connectome analysis in major depression
To examine depression severity changes, the Hamilton Depression Rating Scale (HDRS;
Hamilton, 1967) was administered at three time points (i.e., at baseline [T1], after the first
week of stimulation [T2], and after the second week of stimulation [T3]) by an independent
rater, blind to the treatment condition.
MRI Data Acquisition
MR examination was performed on a Siemens 3T TrioTim MRI scanner (Siemens, Erlangen,
Germany) by using a 32-channel head coil at the Ghent Institute of Functional Imaging (Uni-
versity of Ghent). Diffusion images were acquired using a single-shot echo planar imaging (EPI)
sequence. The major acquisition parameters included the following: repetition time (TR) =
8,500 ms, echo time (TE) = 85 ms, voxel size = 2.0 × 2.0 × 2.0 mm3, slice thickness = 2 mm,
field of view (FOV) = 244 × 244 mm2, matrix size = 122 × 122, 68 contiguous sagittal slices,
no gap, scan time = 9:14 min). For each participant, a total of 62 diffusion-weighted images
(DWI) were acquired, including two non-diffusion-weighted images (b = 0 s/mm2) and 60
diffusion-weighted images (b = 800 s/mm2) with 62 noncollinear gradient directions. In addi-
tion, we acquired anatomical scans using a 3D-TFE sequence (TR/TE = 2,530 ms/2.58 ms; flip
angle = 7 deg; FOV = 220 × 220 mm2; resolution = 0.9 × 0.9 × 0.9 mm3; number of slices =
176; TA = 6 min). Of note, scans of the patients were administered at three time points (T1 at
baseline, T2 after the first week of real/sham aiTBS stimulation, and T3 after the second week
of treatment), while only baseline scans were collected from the healthy control group.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
1
1
5
7
1
0
9
2
3
4
4
n
e
n
_
a
_
0
0
0
6
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
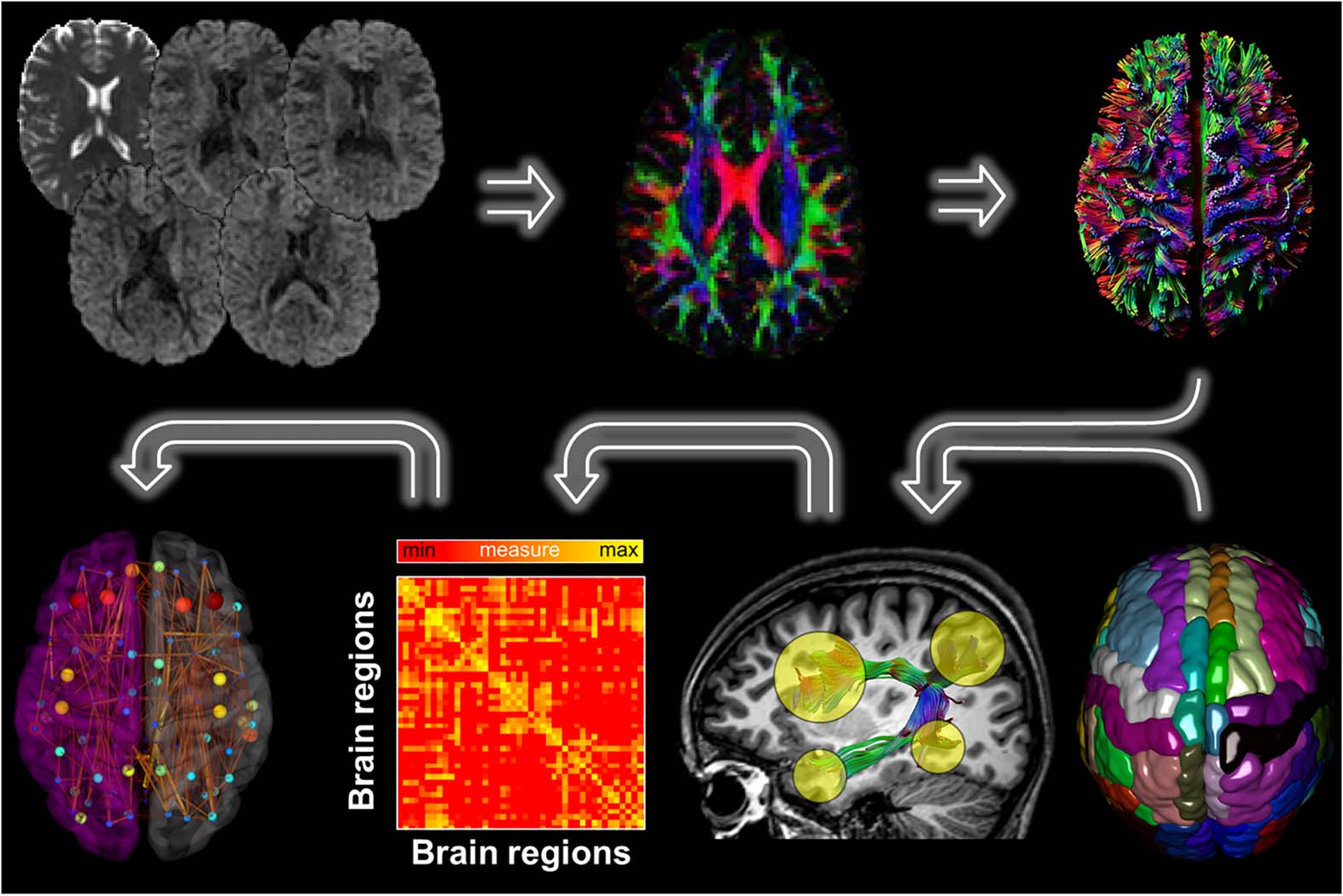
Figure 1. Overview of the MRI data processing pipeline. First, for each DWI dataset a whole brain
deterministic tractography was performed using ExploreDTI. The Desikan-Killiany atlas, consisting
of 89 brain regions, was then used to segment the fiber bundles between each pair of ROIs. We
next determined the density weight between each pair of regions, resulting in 89 × 89 connectivity
matrices. Finally, from the resulting brain network graph metrics were computed.
Network Neuroscience
161
Connectome analysis in major depression
MRI Preprocessing
Figure 1 shows the DWI and T1 processing pipeline. FreeSurfer (http://surfer.nmr.mgh.harvard.
edu) was used for cortical reconstruction and volumetric segmentation reconstruction of the
brain’s surface by using a semiautomated approach described in detail elsewhere (Fischl et al.,
2002; Jovicich et al., 2009), with the use of additional computing resources from the Multi-
modal Australian ScienceS Imaging and Visualisation Environment (MASSIVE) cluster at
Monash University (https://www.massive.org.au/; Goscinski et al., 2014).
Images were pro-
cessed automatically using the FreeSurfer longitudinal stream (Reuter & Fischl, 2011). Default
parameters were used for all processing steps. The results for each subject at each time point
were carefully inspected to ensure the accuracy of the skull stripping, segmentation, and corti-
cal surface reconstruction. Poor data quality, such as inclusion of dura in the pial surface after
skull stripping, and surface deformations, was revealed in two TRD patients. These T1 datasets
were excluded from all further analyses. Finally, the T1.mgz (i.e., the FreeSurfer T1 image) and
aparc+aseg.mgz (i.e., image containing ROIs constructed by the FreeSurfer pipeline) files were
converted to NIfTI format (T1.nii and aparc+aseg.nii) to be used in further diffusion analyses.
ExploreDTI (v4.8.6) (Leemans et al., 2009) was used to process each DWI dataset by using
the following multistep-procedure: first, the FreeSurfer T1.nii files were processed using the
mask function from ExploreDTI, applying a kernel size of morphological operators of 5 and a
threshold of 0.05. Subsequently, diffusion data were corrected for signal drift, subject motion,
eddy current-induced distortions, and susceptibility artifacts (Irfanoglu et al., 2012; Leemans
& Jones, 2009; Vos et al., 2017), with the masked T1.nii files as undistorted (target) scans.
The corrected diffusion results were quality checked in every subject. Poor data quality was
observed in one patient because of severe head motion (exceeding the size of 1 voxel), and
two patients because of artifacts. These DWI data were excluded from further analyses.
The diffusion tensor was estimated from the corrected images with the robust fitting routine
REKINDLE (Veraart et al., 2013; Tax et al., 2015). To correct for EPI distortions, the DWI were
nonrigidly aligned (image contrast during registration is the FA) to the subjects’ individual high-
resolution T1-weighted image, with the deformation field constrained along the phase encoded
A-P axis. Whole-brain tractography was reconstructed in the individual T1 space (Basser et al.,
2000) with a uniform seed point resolution of 2 mm3, step size of 1 mm, an angle threshold of
30
, and FA threshold of 0.2.
◦
Connectome Analyses
The Desikan–Killiany atlas was also used to derive the nodes for our graph theoretical anal-
yses, resulting in 89 ROIs in each subject (see Supporting Information Table S1, Caeyenberghs
et al., 2019). These comprised all cortical ROIs from the Desikan–Killiany atlas (60 cortical
areas), plus cerebellum cortex, thalamus proper, caudate, putamen, pallidum, hippocampus,
amygdala, accumbens-area and ventral diencephalon (all of them bilateral), and brainstem.
Interregional connectivity was then examined by determining the connection density (number
of fiber connections per unit surface and normalized for fiber bundle length) between any
two nodes (i.e., any two regions of the Desikan–Killiany template; Hagmann et al., 2008). The
resulting density weight was converted to symmetrical connectivity matrices (89 × 89 ROIs)
and the main diagonal was set to zeros. These matrices were subsequently used for graph
theoretical analysis, as discussed in the next section.
162
Graph theoretical analysis:
A mathematical framework for
quantifying topological properties
of networks.
Network Neuroscience
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
1
1
5
7
1
0
9
2
3
4
4
n
e
n
_
a
_
0
0
0
6
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Connectome analysis in major depression
Modularity:
Subdivisions of the network in
groups of nodes with many links to
nodes within the group and few links
to nodes outside the group.
Graph Theoretical Analyses
Analyses of network properties were performed using the Graph Analytical Toolbox version
1.4.1 (GAT; Hosseini et al., 2012a), which uses routines of the Brain Connectivity Toolbox for
network metrics calculation (Rubinov & Sporns, 2010).
Cross-Sectional Graph Theoretical Analysis
To allow comparison of global network properties between groups and
Threshold selection.
avoid biases associated with using a single threshold (van Wijk et al., 2010), the matrices
were thresholded at a range of network densities (Dmin: Dmax) (Bassett et al., 2008; Bernhardt
et al., 2011; He et al., 2008; Hosseini et al., 2012a, 2012b). Where Dmin was defined as the
minimum density above which both of the networks were not fragmented (0.10 for this study),
and Dmax was set at 0.20 as after this threshold the graphs became increasingly random.
For each threshold, the following global network metrics were extracted:
Network metrics.
small-worldness, normalized clustering coefficient, normalized shortest path length, global
efficiency, clustering coefficient, modularity, and assortativity. Additionally, the following four
regional network metrics were calculated for each threshold: degree, local efficiency, node
betweenness centrality, and clustering coefficient. An explanation for each global and regional
network metric can be found in Supporting Information Table S2 (Caeyenberghs et al., 2019).
All network metrics were compared with the corresponding values obtained and averaged from
20 random networks with the same number of nodes, edges, and degree distribution (Hosseini
& Kesler, 2013).
Group comparisons global network metrics. Nonparametric permutation testing (5,000 repeti-
tions) was used to determine the statistical significance of between-group differences, control-
ling for age (He et al., 2008; Hosseini et al., 2012b). In each permutation, the connectivity
matrices of each participant were randomly reassigned to one of the two groups (TRD, con-
trols) so that each randomized group had the same number of subjects as the original groups.
Then, an association matrix was obtained for each randomized group. These association ma-
trices were then normalized, and network measures were calculated for each network at each
density and summarized using area under the curve (AUC) (Hosseini et al., 2012b). This re-
sulted in a null distribution of differences, against which the p values of the actual differences
in the curve functions obtained by comparing controls and TRD patients were computed. This
nonparametric permutation test based on AUC inherently accounts for multiple comparisons
across the range of densities (Bassett et al., 2011; Singh et al., 2013).
The same permutation procedure was used to
Group comparisons regional network metrics.
test the significance of the between-group differences in regional network measures, that is,
comparing the AUC of the regional network measures over the specified density range. The
p values reported for regional differences between groups were false discovery rate (FDR)
corrected for multiple comparisons, with a statistical threshold of p < .05.
Hub Analysis
Finally, we also performed a qualitative hub analysis. The nodes with the largest betweenness
centrality were considered to be the most important regions in the brain network (hubs). Hubs
Network Neuroscience
163
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
1
1
5
7
1
0
9
2
3
4
4
n
e
n
_
a
_
0
0
0
6
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Connectome analysis in major depression
are essential for coordinating brain functions through their connectivity with various brain
regions (Cole et al., 2010) and facilitate efficient communication across the network. In the
present study, a node was considered to be a hub if its regional betweenness centrality was
2 SD higher than the mean betweenness centrality of the network. The hubs were quantified
based on the AUC of the betweenness centrality in the specified density range.
Longitudinal Graph Theoretical Analyses
Longitudinal graph analysis was performed with the Graph Analysis Toolbox, version 1.4.1
(Amidi et al., 2017), using the following procedure: first, networks were normalized by the
mean network strength, and the following global network metrics were quantified for the
normalized networks at each time point: betweenness centrality, normalized clustering co-
efficient, normalized path length, small-worldness, global efficiency, local efficiency, and
modularity. These network metrics were then extracted for further analyses with general linear
models (see below Statistical Analyses).
Statistical Analyses
For the baseline analyses (controls vs. TRD patients, order 0 vs. order 1 TRD patients) group
comparisons on demographic variables (i.e., age) were performed with t tests for continuous
variables, and χ2 analyses for categorical variables (i.e., gender). Within the group of TRD
patients only, a repeated-measures ANOVA was conducted on HDRS scores and longitudinal
graph metrics with Order (2: aiTBS>sham or sham>aiTBS) as between-subject factor and time
(3: T1-T3) as within-subject factor. An exploratory analysis was also performed within respon-
ders versus nonresponders to examine changes in graph metrics with time, using a 2 (responder
vs. nonresponder) × 3 (time points) repeated-measures ANOVA. Finally, Pearson product cor-
relation coefficients were calculated within each Order (subgroups of TRD patients), between
(a) the change in modularity (calculated as the difference score T2-T1), for which there was a
significant order × time interaction effect, and (b) change in depression scores, that is, differ-
ence in HDRS scores between T1 and T2 (delta HDRS score T2-T1). The p values reported for
correlations were uncorrected for multiple comparisons with a statistical threshold of p < .05.
These analyses should be considered exploratory.
RESULTS
Clinical and Demographic Characteristics of the Subjects
As shown in Supporting Information Table S1 (Caeyenberghs et al., 2019), there were no sig-
nificant differences in age (t(104) = −1.231, p < 0.221) and gender (χ2 = 1.844, p < 0.226)
between TRD patients and control subjects. In addition, the results revealed no significant
differences in age (t(44) = 0.815, p < 0.419), gender (χ2 = 0.199, p < 0.754), or pretreatment
HDRS scores (t(44) = 0.626, p < 0.534) between the two groups of TRD patients.
Behavioral Results
Repeated-measures ANOVA showed a significant decrease of HDRS scores over time,
F(2, 78) = 32.21, p < 0.001. Post hoc paired samples t tests showed significant reductions in
HDRS scores between T1 and T2 (p < 0.001), and between T2 and T3 (p < 0.001), indicating
that the HDRS score at T2 (mean = 21.76, SD = 5.65) was lower than T1 (mean = 17.79,
SD = 6.26), and the HDRS score at T3 (mean = 14.56, SD = 6.87) was lower compared with
Network Neuroscience
164
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
1
1
5
7
1
0
9
2
3
4
4
n
e
n
_
a
_
0
0
0
6
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Connectome analysis in major depression
Small-world topology:
Close local clustering of connections
between neighboring nodes but a
short path length between any distant
pair of nodes caused by relatively
few long-range connections.
T2. There was no significant effect of the order of treatment (sham>aiTBS vs. aiTBS>sham;
p < 0.154) and no significant interaction effect (p < 0.308). Defining treatment response as a
reduction of 50% from baseline HDRS scores, we found 11 responders (27%) at T3 in the TRD
group.
Baseline Group Differences in the Structural Connectome
We investigated (baseline) between-group differences in global network measures, comparing
the AUC for these network measure curves (density range of 0.10:0.01:0.20). Both groups
showed a small-world organization of the structural brain network expressed by a normalized
clustering coefficient > 1, normalized path length ≈ 1, and small-world index > 1. The struc-
tural network of the TRD patients showed a significantly shorter normalized path length
(pAUC < 0.01) compared with the healthy control group. Also, the structural brain networks
in TRD patients were characterized by a lower assortativity (pAUC < 0.035) compared with
controls. Group effects were absent in the other global network metrics, including global ef-
ficiency (pAUC = 0.325), normalized clustering coefficient (pAUC = 0.943), small-worldness
(pAUC = 0.737), local efficiency (pAUC = 0.223), modularity (pAUC = 0.460), and clustering
coefficient (pAUC = 0.261).
Direct comparison of the nodal graph metrics (nodal degree, local efficiency, clustering
coefficient, and betweenness centrality) revealed no significant group differences after FDR
correction. Results of the nodal analyses using an exploratory threshold of p < 0.05 are re-
ported in Supporting Information (Caeyenberghs et al., 2019).
Finally, our qualitative analysis of the hub distribution using nodal betweenness centrality
revealed that both groups exhibited seven hubs including the bilateral precuneus, bilateral
superior frontal gyri, right superior parietal gyrus, left thalamus proper, and brainstem.
Structural Network Alterations with aiTBS Stimulation
The 2 × 3 repeated-measures ANOVAs revealed a significant order × time interaction effect
for modularity, F(2, 78) = 3.30, p < 0.042. However, no significant main effects of order,
F(1, 39) = 1.301, p < 0.261, or time F(2, 78) = 0.477, p < 0.622, were found. Post hoc t test
showed a significant reduction in modularity (i.e., less modular network configuration) from T1
to T2 in the order 1 group (aiTBS>sham) (p < 0.013), but not in the order 0 group (sham>aiTBS).
No significant main effects or significant interaction effects were found for the other global
network metrics.
Structural Network Alterations in Responders
We observed a marginal significant responder × time interaction effect on betweenness cen-
trality, F(2, 78) = 3.07, p < 0.052. The main effects of responder, F(2, 39) = 0.002, p < 0.964,
and time, F(2, 78) = 1.558, p < 0.217, were not significant. Post hoc tests revealed marginally
increased values of betweenness centrality in the group of responders at T3 compared with T1
(p < 0.072).
Correlations Between Changes in the Structural Connectome and Changes in Depression Severity Scores
The analyses of correlations between the significant change in modularity from T1 to T2 and the
changes in depression scores on the HDRS between T1 and T2 showed no direct associations
(order 1 group [aiTBS>sham]: r = 0.208, p < 0.379; order 0 group [sham> aiTBS]: r =
−0.032, p < 0.889).
Network Neuroscience
165
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
1
1
5
7
1
0
9
2
3
4
4
n
e
n
_
a
_
0
0
0
6
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Connectome analysis in major depression
DISCUSSION
This study addressed the broad issue of whether graph theory and its metrics can be used to map
the clinical effects of accelerated brain stimulation protocols on major depression. Specifically,
ours was the first study to explore aiTBS-induced changes in the structural connectome of
patients with TRD by using a well-controlled clinical trial design. Results showed immediate
reductions in depression severity symptoms. Moreover, our graph theoretical analyses revealed
modularity changes after 4 days of active stimulation, which suggests decreased functional
segregation of the patients’ structural brain networks. This is consistent with our clinical find-
ings in this cohort, where depression severity scores further declined 2 weeks after the aiTBS
treatment protocol (Duprat et al., 2016).
Clinical Findings
The positive impact of accelerated stimulation paradigms on depression severity is consistent
with our own earlier studies (Baeken et al., 2013; Duprat et al., 2016) and others work in re-
lated psychiatric disorders, such as unipolar depression (Hadley et al., 2011; Holtzheimer et al.,
2010) and suicidal patients (Desmyter et al., 2016; George et al., 2014). Although, in the pres-
ent study the observed reduction in depressive symptoms was unrelated to active or sham
stimulation, as revealed by a nonsignificant interaction effect between order and time for the
HDRS scores (see also limitations section). The clinical analysis of the Hamilton scale (i.e.,
defining treatment response as a reduction of 50% from baseline HDRS scores) showed for
the 41 included patients at the end of the 2-week study protocol (T3) 11 as clinical responders
(27%), with only 7 in remission (17%) (defined as a HDRS scorer ≤ 7). Importantly, 2 weeks
after the iTBS trial at T4 the amount of clinical responders (n = 17) mounted up to 41%. Twelve
patients were here also considered in remission (29%). These observations indicate delayed
clinical responses to aiTBS treatment. It is important to investigate whether the change in
depressive scores meets the criteria for a clinically important difference, besides analyzing
statistically significant differences.
Baseline Connectome Analyses
In this study, we also investigated white matter networks of TRD patients and healthy controls
by using diffusion MRI tractography and graph theoretical approaches. Although small-world
properties were present for both the control and TRD group, the topological architecture of
the structural networks was significantly altered in patients with TRD. First, normalized path
length, which is a measure of functional integration (i.e., ability to rapidly combine special-
ized information from distributed brain regions; Rubinov & Sporns, 2010) was altered in TRD.
Notably, normalized clustering coefficient, which is a measure of functional segregation (i.e.,
the ability of specialized processes to occur within highly interconnected groups of brain re-
gions), was not affected in TRD patients. Therefore, our results suggest a preservation of the
efficiency of local information transfer and processing and an impairment of global integration,
likely to reflect a reduced competence in information exchange between distant brain areas.
This is also supported by the lower assortativity we found in TRD patients. Second, compared
with controls, our TRD patients showed lower network assortativity. In assortative networks,
nodes with many connections tend to be connected to other nodes with many connections,
and nodes with low connections are linked to other low-connection nodes (Newman, 2002).
When network hubs are abnormally clustered and connected to low-degree nodes, assortativ-
ity drops and the structural network is less efficiently wired (Newman, 2002). Abnormalities
of assortativity similar to those we have found in TRD have been described in patients with
multiple sclerosis (Kocevar et al., 2016; Rocca et al., 2016).
Network Neuroscience
166
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
1
1
5
7
1
0
9
2
3
4
4
n
e
n
_
a
_
0
0
0
6
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Connectome analysis in major depression
In short, our global network analysis of TRD networks suggests a reduction in the balance
between network segregation and integration and a loss of efficiency in information exchange
between both close and distant regions. This is not in contradiction to former research where it
has been reported that network dysfunctions may contribute to cognitive and affective abnor-
malities in MDD (Kaiser et al., 2015). However, mixed findings in global network metrics have
been obtained in previous diffusion MRI studies examining whole-brain white matter networks
in depression. For example, Bai et al. ( 2012) showed comparable disrupted global properties
of structural networks, including reduced network strength and increased path length in re-
mitted geriatric depression (N = 35). Conversely, studies by Korgaonkar et al. ( 2014) and
Qin et al. ( 2014) found no significant group differences on graph measures in patients with
MDD. Differences in type of depression, parcellation scheme (the automated anatomical la-
beling atlas, Desikan–Killiany atlas), tractography algorithm (deterministic, probabilistic), and
analysis methods (network-based statistical analysis vs. graph theoretical analyses) are likely
to account for differences between ours and previous results.
aiTBS Influences Modularity in TRD Patients
Our longitudinal graph analyses revealed modularity changes after 4 days of real stimula-
tion. Specifically, we observed a less modular network configuration in the order 1 group
(aiTBS>sham) from T1 to T2. Modularity is a ubiquitous property of complex, large-scale
brain networks. Modularity implies that the network is composed of a set of modules each
comprising nodes that are densely connected to each other and sparsely connected to nodes
in other modules (Newman & Girvan, 2004). It is possible that these transient changes in
(modular) network configuration may be necessary for improvements in depressive symptoms
in TRD. Our finding here also complements other experimental studies of learning and behav-
ioral plasticity that show modularity to be predictive of cognitive effort and learning success
(Bassett et al., 2011; Bola & Sabel, 2015; Kitzbichler et al., 2011; Stevens et al., 2012). For ex-
ample, a MEG study by Kitzbichler et al. ( 2011) showed a less modular network configuration
during the performance of an effortful 2-back verbal working memory task in healthy adults
(N = 13).
Important to note, we observed the network effect in one group only (active stimulation
first). We could not replicate our modularity findings in the second group (sham stimulation
first). Although, a marginal significant interaction effect for modularity was observed between
T2 and T3, the decreased modularity was only valid for the time interval T1-T2. We suggest
that a carryover effect has biased our connectome results of our clinical trial. To overcome
this issue, one should leave sufficient time (months) between the active and the sham aiTBS
treatment or vice versa, which will be difficult to accept on ethical grounds, leaving TRD
patients without proper treatment.
Relationship Between Improvements in Depressive Symptoms and Changes in the
Structural Connectome
No correlations were found between changes on depression severity and the degree of change
of modularity. We observed nonsignificant correlations between change in the HDRS and
change in modularity in the TRD subgroups (order 0, order 1). In other words, network changes
did not correspond directly with clinical improvements, which may have been due to nonspe-
cific neural responses to brain stimulation. Another explanation is that the accelerated brain
stimulation protocol may have triggered changes in brain structure, but not necessarily in a way
Network Neuroscience
167
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
1
1
5
7
1
0
9
2
3
4
4
n
e
n
_
a
_
0
0
0
6
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Connectome analysis in major depression
that covaried significantly with ratings of depression. Negative findings may also suggest that
the network metrics affected by brain stimulation may not underpin clinical changes. Nonlin-
ear modeling techniques may further clarify relationships between connectome and clinical
changes with brain stimulation. Last, it could also be that the current scanning assessment was
too short to detect modularity changes, as additional significant clinical improvement was
observed 2 weeks after the aiTBS treatment (Duprat et al., 2016).
The observed reduction in depressive symptoms was unrelated to active or sham stimula-
tion. This result is consistent with previous studies revealing clinical improvements after sham
stimulation (Duecker, de Graaf, Jacobs, & Sack, 2013; Duecker & Sack, 2015; Opitz et al.,
2015), suggesting by some authors that this is part of its effect (Razza et al., 2018). Although
our sham procedure involved a specially designed placebo coil completely similar to the real
one, which did not induce any electric fields in the human cortex, this procedure is not a pure
reproduction of real rTMS, given the possible differences in skin sensations. However, clear-
cut sham rTMS procedures are not available yet (Baeken et al., 2014). Nonspecific effects (e.g.,
TRD patients receiving uncustomary attention during the trial) may also contribute to clinical
improvements after sham stimulation. Further research is necessary to investigate the effects
of sham stimulation protocols.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
1
1
5
7
1
0
9
2
3
4
4
n
e
n
_
a
_
0
0
0
6
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The above caveats aside, to our knowledge this is the first study providing evidence of struc-
tural connectome changes in response to brain stimulation in TRD. Indeed, earlier research
has shed some light as to how accelerated rTMS paradigms influenced local neurobiological
changes in TRD patients, with changes in subgenual functional connectivity (Baeken et al.,
2014, 2017), metabolism (Baeken et al., 2015), the reward system (Duprat et al., 2016), and in
local GABAergic inhibitory neurotransmission (Baeken, Lefaucheur, & Van Schuerbeek 2017).
Nevertheless, our current findings substantiate our former assumptions that brain changes re-
lated to clinical outcome is already present after only 4 days of stimulation. In the future, we
suggest using graph theoretical analysis not only to understand the effect of brain stimulation
on brain networks, but also to fine-tune brain stimulation protocols that can target a network
of brain areas rather than single brain regions.
AUTHOR CONTRIBUTIONS
Karen Caeyenberghs: Conceptualization; Formal analysis; Supervision; Writing – original
Investigation; Project administration;
draft; Writing – review & editing. Romain Duprat:
Writing – review & editing. Alexander Leemans: Formal analysis; Methodology; Software;
Writing – review & editing. Hadi Hosseini: Formal analysis; Methodology; Software; Writing –
review & editing. Peter Wilson: Writing – review & editing. Debby Klooster: Formal analy-
sis; Writing – review & editing. Chris Baeken: Conceptualization; Formal analysis; Funding
acquisition; Investigation; Project administration; Writing – review & editing.
FUNDING INFORMATION
Chris Baeken, Concerted Research Action of Ghent University, Award ID: BOF16/GOA/017. Karen
Caeyenberghs, Australian Catholic University (http://dx.doi.org/10.13039/501100000990),
supported by an National Health and Medical Research Council Career Development Fellow-
ship and an ACURF Program grant by the Australian Catholic University (ACU). Chris Baeken,
Ghent University Multidisciplinary Research Partnership, Award ID: The integrative neuro-
science of behavioral control. Peter Wilson, Australian Catholic University (http://dx.doi.org/
10.13039/501100000990), Award ID: ACURF Program grant.
Network Neuroscience
168
Connectome analysis in major depression
REFERENCES
Amidi, A., Hosseini, S. M. H., Leemans, A., Kesler, S. R., Agerbæk,
M., Wu, L. M., & Zachariae, R.
(2017). Changes in brain
structural networks and cognitive functions in testicular cancer
patients receiving cisplatin-based chemotherapy. Journal of the
National Cancer Institute, 109(12).
Baeken, C., Lefaucheur, J. P., & Van Schuerbeek, P. (2017). The
impact of accelerated high frequency rTMS on brain neuro-
chemicals in treatment-resistant depression:
Insights from 1H
MR spectroscopy. Clinical Neurophysiology, 128(9), 1664–1672.
https://doi.org/10.1016/j.clinph.2017.06.243
Baeken, C., Marinazzo, D., Everaert, H., Wu, G. R., Van Hove, C.,
Audenaert, K., . . . De Raedt, R. (2015). The impact of accelerated
HF-rTMS on the subgenual anterior cingulate cortex in refractory
unipolar major depression: Insights from 18FDG PET brain imag-
ing. Brain Stimulation, 8(4), 808–815. https://doi.org/10.1016/
j.brs.2015.01.415
Baeken, C., Marinazzo, D., Wu, G. R., Van Schuerbeek, P., De Mey,
J., Marchetti, I., . . . De Raedt, R. (2014). Accelerated HF-rTMS in
treatment-resistant unipolar depression: Insights from subgenual
anterior cingulate functional connectivity. The World Journal of
Biological Psychiatry, 15(4), 286–297. https://doi.org/10.3109/
15622975.2013.872295
Baeken, C., Vanderhasselt, M. A., Remue,
J., Herremans, S.,
Vanderbruggen, N., Zeeuws, D., . . . De Raedt, R.
(2013). In-
tensive HF-rTMS treatment in refractory medication-resistant uni-
polar depressed patients. Journal of Affective Disorders, 151(2),
625–631. https://doi.org/10.1016/j.jad.2013.07.008
Bai, F., Shu, N., Yuan, Y., Shi, Y., Yu, H., Wu, D., . . . Zhang, Z.
(2012). Topologically convergent and divergent structural con-
nectivity patterns between patients with remitted geriatric de-
pression and amnestic mild cognitive impairment. The Journal
of Neuroscience, 32(12), 4307–4318. https://doi.org/10.1523/
JNEUROSCI.5061-11.2012
Basser, P. J., Pajevic, S., Pierpaoli, C., Duda, J., & Aldroubi, A.
(2000). In vivo fiber tractography using DT-MRI data. Magnetic
Resonance in Medicine, 44(4), 625–632.
Bassett, D. S., Bullmore, E., Verchinski, B. A., Mattay, V. S.,
Weinberger, D. R., & Meyer-Lindenberg, A.
(2008). Hierar-
chical organization of human cortical networks in health and
schizophrenia. The Journal of Neuroscience, 28(37), 9239–9248.
https://doi.org/10.1523/JNEUROSCI.1929-08.2008
Bassett, D. S., Wymbs, N. F., Porter, M. A., Mucha, P. J., Carlson,
(2011). Dynamic reconfiguration of
the
J. M., & Grafton, S.
human brain networks during learning. Proceedings of
National Academy of Sciences of
America, 108(18), 7641–7646. https://doi.org/10.1073/pnas.
1018985108
the United States of
Bernhardt, B. C., Chen, Z., He, Y., Evans, A. C., & Bernasconi, N.
(2011). Graph-theoretical analysis reveals disrupted small-world
organization of cortical thickness correlation networks in tempo-
ral lobe epilepsy. Cerebral Cortex, 21(9), 2147–2157. https://doi.
org/10.1093/cercor/bhq291
Bola, M., & Sabel, B. A.
(2015). Dynamic reorganization of
brain functional networks during cognition. NeuroImage, 114,
398–413. https://doi.org/10.1016/j.neuroimage.2015.03.057
Brakowski, J., Spinelli, S., Dorig, N., Bosch, O. G., Manoliu, A.,
Holtforth, M. G., & Seifritz, E. (2017). Resting state brain net-
work function in major depression—Depression symptomatol-
ogy, antidepressant treatment effects, future research. Journal
of Psychiatric Research, 92, 147–159. https://doi.org/10.1016/
j.jpsychires.2017.04.007
Brunoni, A. R., Chaimani, A., Moffa, A. H., Razza, L. B., Gattaz,
W. F., Daskalakis, Z. J., & Carvalho, A. F. (2017). Repetitive
transcranial magnetic stimulation for the acute treatment of ma-
jor depressive episodes: A systematic review with network meta-
JAMA Psychiatry, 74(2), 143–152. https://doi.org/10.
analysis.
1001/jamapsychiatry.2016.3644
Caeyenberghs, K., Duprat, R., Leemans, A., Hosseini, H., Wilson,
P. H., Klooster, D., & Baeken, C. (2019). Supporting information
for “Accelerated intermittent theta burst stimulation in major
depression induces decreases in modularity: A connectome
analysis.” Network Neuroscience, 3(1), 157–172. https://doi.org/
10.1162/netn_a_00060
Chen, V. C., Shen, C. Y., Liang, S. H., Li, Z. H., Hsieh, M. H., Tyan,
Y. S., . . . Weng, J. C. (2017). Assessment of brain functional con-
nectome alternations and correlation with depression and anxi-
ety in major depressive disorders. PeerJ, 5, e3147. https://doi.org/
10.7717/peerj.3147
Conradi, H. J., Ormel,
Cole, M. W., Pathak, S., & Schneider, W. (2010). Identifying the
brain’s most globally connected regions. NeuroImage, 49(4),
3132–3148. https://doi.org/10.1016/j.neuroimage.2009.11.001
J., & de Jonge, P. (2011). Presence of
individual
(residual) symptoms during depressive episodes
and periods of remission: A 3-year prospective study. Psycho-
logical Medicine, 41(6), 1165–1174. https://doi.org/10.1017/
S0033291710001911
Desmyter, S., Duprat, R., Baeken, C., Van Autreve, S., Audenaert,
K., & van Heeringen, K.
(2016). Accelerated intermittent theta
burst stimulation for suicide risk in therapy-resistant depressed
patients: A randomized, sham-controlled trial. Frontiers in Human
Neuroscience, 10, 480.
.
.
Drysdale, A. T., Grosenick, L., Downar, J., Dunlop, K., Mansouri, F.,
(2017). Resting-state connectiv-
Meng, Y.,
ity biomarkers define neurophysiological subtypes of depres-
sion. Nature Medicine, 23(1), 28–38. https://doi.org/10.1038/
nm.4246
. Liston, C.
Duecker, F., de Graaf, T. A., Jacobs, C., & Sack, A. T.
(2013).
Time- and task-dependent non-neural effects of real and sham
TMS. PLoS One, 8(9), 1–9. https://doi.org/10.1371/journal.pone.
0073813
Duecker, F., & Sack, A. T. (2015). Rethinking the role of sham TMS.
Frontiers in Psychology, 6, 1–5. https://doi.org/10.3389/fpsyg.2015.
00210
Duprat, R., Desmyter, S., Rudi de, R., van Heeringen, K., Van den
(2016). Accelerated
Abbeele, D., Tandt, H., . . . Baeken, C.
Network Neuroscience
169
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
1
1
5
7
1
0
9
2
3
4
4
n
e
n
_
a
_
0
0
0
6
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Connectome analysis in major depression
theta burst stimulation treatment
intermittent
in medication-
resistant major depression: A fast road to remission? Journal of
Affective Disorders, 200, 6–14. https://doi.org/10.1016/j.jad.
2016.04.015
Fischl, B., Salat, D. H., Busa, E., Albert, M., Dieterich, M.,
Haselgrove, C.,
. . . Dale, A. M. (2002). Whole brain seg-
mentation: Automated labeling of neuroanatomical structures in
the human brain. Neuron, 33, 341–355.
(2012). Efficacy of
Fox, M. D., Buckner, R. L., White, M. P., Greicius, M. D., & Pascual-
Leone, A.
transcranial magnetic stimula-
tion targets for depression is related to intrinsic functional
connectivity with the subgenual cingulate. Biological Psychi-
atry, 72(7), 595–603. https://doi.org/10.1016/j.biopsych.2012.
04.028
Geerts, P. J., Lemmens, G. M., & Baeken, C. (2015). The occur-
rence of depersonalization symptoms after accelerated HF-rTMS
of the left DLPFC in a patient with treatment-resistant depression:
A case report. Brain Stimulation, 8(3), 681–682. https://doi.org/
10.1016/j.brs.2015.02.010
George, M. S., Raman, R., Benedek, D. M., Pelic, C. G., Grammer,
G. G., Stokes, K. T., . . . Stein, M. B. (2014). A two-site pi-
lot randomized 3-D trial of high dose left prefrontal repetitive
transcranial magnetic stimulation (rTMS) for suicidal inpatients.
Brain Stimulation, 7, 421–431.
Gong, Q., & He, Y. (2015). Depression, neuroimaging and con-
nectomics: A selective overview. Biological Psychiatry, 77(3),
223–235. https://doi.org/10.1016/j.biopsych.2014.08.009
Goscinski, W. J., McIntosh, P., Felzmann, U., Maksimenko, A.,
Hall, C. J., Gureyev, T., . . . Egan, G. F. (2014). The multi-
modal Australian ScienceS Imaging and Visualization Environ-
(MASSIVE) high performance computing infrastructure:
ment
Applications in neuroscience and neuroinformatics research.
Frontiers in Neuroinformatics, 8, 1–13. https://doi.org/10.3389/
fninf.2014.00030
Guo, H., Cao, X., Liu, Z., Li, H., Chen, J., & Zhang, K.
(2012).
Machine learning classifier using abnormal brain network topo-
logicalmetrics in major depressive disorder. Neuroreport, 23(17),
1006–1011. https://doi.org/10.1097/WNR.0b013e32835a650c
Guo, H., Cheng, C., Cao, X., Xiang, J., Chen, J., & Zhang, K. (2014).
Resting-state functional connectivity abnormalities in first-onset
unmedicated depression. Neural Regeneration Research, 9(2),
153–163. https://doi.org/10.4103/1673-5374.125344
Hadley, D., Anderson, B. S., Borckardt, J. J., Arana, A., Li, X., Nahas,
Z., & George, M. S. (2011). Safety, tolerability, and effectiveness
of high doses of adjunctive daily left prefrontal repetitive trans-
cranial magnetic stimulation for treatment-resistant depression in
a clinical setting. Journal of ECT, 27(1), 18–25. https://doi.org/
10.1097/YCT.0b013e3181ce1a8c
Hagmann, P., Cammoun, L., Gigandet, X., Meuli, R., Honey, C. J.,
Wedeen, V. J., & Sporns, O. (2008). Mapping the structural
core of human cerebral cortex. PLoS Biology, 6(7), 1479–1493.
https://doi.org/10.1371/journal.pbio.0060159
Hamilton, M. (1967). Development of a rating scale for primary
depressive illness. The British Journal of Social and Clinical
Psychology, 6(4), 278–296.
He, Y., Chen, Z., & Evans, A. (2008). Structural insights into aber-
large-scale cortical networks in
topological patterns of
rant
Alzheimer’s disease. Journal of Neuroscience, 28(18), 4756–4766.
https://doi.org/10.1523/jneurosci.0141-08.2008
Holtzheimer, P. E., McDonald, W. M., Mufti, M., Kelley, M. E.,
Quinn, S., Corso, G., & Epstein, C. M. (2010). Accelerated repet-
itive transcranial magnetic stimulation for treatment-resistant
depression. Depression and Anxiety, 27(10), 960–963. https://
doi.org/10.1002/da.20731
Hosseini, S. M., Hoeft, F., & Kesler, S. R. (2012). GAT: A graph-
theoretical analysis toolbox for analyzing between-group differ-
ences in large-scale structural and functional brain networks.
PLoS One, 7(7), 1–15.
Hosseini, S. M., & Kesler, S. R. (2013). Influence of choice of null
network on small-world parameters of structural correlation net-
works. PLoS One, 8(6), e67354. https://doi.org/10.1371/journal.
pone.0067354
Hosseini, S. M., Koovakkattu, D., & Kesler, S. R. (2012). Altered
small-world properties of gray matter networks in breast cancer.
BMCNeurology, 12, 28. https://doi.org/10.1186/1471-2377-12-28
Huang, C. C., Su, T. P., & Wei, I. H. (2005). Repetitive transcranial
magnetic stimulation for treating medication-resistant depression
Journal of the Chinese Medical
in Taiwan: a preliminary study.
Association, 68(5), 210–215.
Huang, Y. Z., Cammoun, L., Gigandet, X., Meuli, R., Honey, C. J.,
Wedeen, V. J., & Sporns, O. (2005). Theta burst stimulation of
the human motor cortex. Neuron, 45(2), 201–206.
Irfanoglu, M. O., Walker, L., Sarlls, J., Marenco, S., & Pierpaoli, C.
(2012). Effects of image distortions originating from susceptibility
variations and concomitant fields on diffusion MRI tractography
results. NeuroImage, 61(1), 275–288. https://doi.org/10.1016/
j.neuroimage.2012.02.054
Jovicich,
J., Czanner, S., Han, X., Salat, D., van der Kouwe,
A., Quinn, B.,
. . . Fischl, B. (2009). MRI-derived measure-
ments of human subcortical, ventricular and intracranial brain
volumes: Reliability effects of scan sessions, acquisition se-
quences, data analyses, scanner upgrade, scanner vendors and
field strengths. NeuroImage, 46(1), 177–192. https://doi.org/10.
1016/j.neuroimage.2009.02.010
Kaiser, R. H., Andrews-Hanna, J. R., Wager, T. D., & Pizzagalli,
D. A. (2015). Large-scale network dysfunction in major depres-
sive disorder: A meta-analysis of resting-state functional connec-
tivity. JAMA Psychiatry, 72(6), 603–611. https://doi.org/10.1001/
jamapsychiatry.2015.0071
Kesler, S., Hadi Hosseini, S. M., Heckler, C., Janelsins, M., Palesh,
(2013). Cognitive training for
O., Mustian, K., & Morrow, G.
improving executive function in chemotherapy-treated breast
cancer survivors. Clinical Breast Cancer, 13(4), 299–306. https://
doi.org/10.1016/j.clbc.2013.02.004
Kitzbichler, M. G., Henson, R. N., Smith, M. L., Nathan, P. J.,
& Bullmore, E. T. (2011). Cognitive effort drives workspace
configuration of human brain functional networks. Journal
of Neuroscience, 31(22), 8259–8270. https://doi.org/10.1523/
JNEUROSCI.0440-11.2011
Kocevar, G., Stamile, C., Hannoun, S., Cotton, F., Vukusic, S.,
Durand-Dubief, F., & Sappey-Marinier, D. (2016). Graph theory-
based brain connectivity for automatic classification of multiple
sclerosis clinical courses. Frontiers in Neuroscience, 10. https://
doi.org/10.3389/fnins.2016.00478
Network Neuroscience
170
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
1
1
5
7
1
0
9
2
3
4
4
n
e
n
_
a
_
0
0
0
6
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Connectome analysis in major depression
Korgaonkar, M. S., Fornito, A., Williams, L. M., & Grieve,
S. M. (2014). Abnormal structural networks characterize major
depressive disorder: A connectome analysis. Biological Psychi-
atry, 76(7), 567–574. https://doi.org/10.1016/j.biopsych.2014.
02.018
Leemans, A.,
J., & Jones, D. K. (2009).
ExploreDTI: A graphical toolbox for processing, analyzing, and
visualizing diffusion MR data. Paper presented at the 17th Annual
Meeting of the International Society of Magnetic Resononance
Medicine, Hawaii, USA.
Jeurissen, B., Sijbers,
Leemans, A., & Jones, D. K. (2009). The B-matrix must be rotated
when correcting for subject motion in DTI data. Magnetic Res-
onance in Medicine, 61(6), 1336–1349. https://doi.org/10.1002/
mrm.21890
Lefaucheur,
J. P., André-Obadia, N., Antal, A., Ayache, S. S.,
Baeken, C., Benninger, D. H., . . . Garcia-Larrea, L. (2014).
Evidence-based guidelines on the therapeutic use of repeti-
tive transcranial magnetic stimulation (rTMS). Clinical Neuro-
physiology, 125(11), 2150–2206. https://doi.org/10.1016/j.clinph.
2014.05.021
Li, C. T., Chen, M. H., Juan, C. H., Huang, H. H., Chen, L. F.,
Hsieh, J. C., . . . Su, T. P. (2014). Efficacy of prefrontal theta-
burst stimulation in refractory depression: A randomized sham-
controlled study. Brain, 137(Pt 7), 2088–2098. https://doi.org/
10.1093/brain/awu109
Liston, C., Chen, A. C., Zebley, B. D., Drysdale, A. T., Gordon, R.,
Leuchter, B., . . . Dubin, M. J.
(2014). Default mode network
mechanisms of transcranial magnetic stimulation in depression.
Biological Psychiatry, 76(7), 517–526. https://doi.org/10.1016/
j.biopsych.2014.01.023
Lu, Y., Shen, Z., Cheng, Y., Yang, H., He, B., Xie, Y., . . . Han,
D. (2017). Alternations of white matter structural networks
in first episode untreated major depressive disorder with short
duration. Frontiers in Psychiatry, 8, 205. https://doi.org/10.3389/
fpsyt.2017.00205
Newman, M. E.
(2002). Assortative mixing in networks. Physical
Review Letters, 89(20).
Newman, M. E., & Girvan, M. (2004). Finding and evaluating com-
munity structure in networks. Physical Review E, 69. https://doi.
org/10.1103/PhysRevE.69.026113
Oberman, L., Edwards, D., Eldaief, M., & Pascual-Leone, A.
(2011). Safety of
transcranial magnetic stimula-
tion: A systematic review of the literature. Journal of Clinical
Neurophysiology, 28(1), 67–74. https://doi.org/10.1097/WNP.
0b013e318205135f
theta burst
Opitz, A., Legon, W., Mueller, J., Barbour, A., Paulus, W., & Tyler,
W. J. (2015). Is sham cTBS real cTBS? The effect on EEG dy-
namics. Frontiers in Human Neuroscience, 8. https://doi.org/10.
3389/fnhum.2014.01043
Peña-Gómez, C., Sala-Lonch, R.,
Junqué, C., Clemente,
I. C.,
Vidal, D., Bargalló, N., . . . Bartrés-Faz, D. (2012). Modulation
of
large-scale brain networks by transcranial direct current
stimulation evidenced by resting-state functional MRI. Brain
Stimulation, 5(3), 252–263. https://doi.org/10.1016/j.brs.2011.
08.006
Philip, N. S., Barredo, J., Aiken, E., & Carpenter, L. L. (2018).
Neuroimaging mechanisms of therapeutic transcranial magnetic
stimulation for major depressive disorder. Biological Psychia-
try Cognitive Neuroscience and Neuroimaging, 3(3), 211–222.
https://doi.org/10.1016/j.bpsc.2017.10.007
Plewnia, C., Pasqualetti, P., Grosse, S., Schlipf, S., Wasserka, B.,
Zwissler, B., & Fallgatter, A. (2014). Treatment of major depres-
sion with bilateral theta burst stimulation: A randomized con-
trolled pilot trial. Journal of Affective Disorders, 156, 219–223.
https://doi.org/10.1016/j.jad.2013.12.025
Polanía, R., Nitsche, M. A., & Paulus, W. (2011). Modulating
functional connectivity patterns and topological functional or-
ganization of the human brain with transcranial direct current
stimulation. Human Brain Mapping, 32(8), 1236–1249. https://
doi.org/10.1002/hbm.21104
Qin, J., Wei, M., Liu, H., Yan, R., Luo, G., Yao, Z., & Lu, Q.
(2014). Abnormal brain anatomical
topological organization
of the cognitive-emotional and the frontoparietal circuitry in
major depressive disorder. Magnetic Resonance in Medicine,
72(5), 1397–1407. https://doi.org/10.1002/mrm.25036
Razza, L. B., Moffa, A. H., Moreno, M. L., Carvalho, A. F., Padberg,
F., Fregni, F., & Brunoni, A. R. (2018). A systematic review
and meta-analysis on placebo response to repetitive transcra-
nial magnetic stimulation for depression trials. Progress in Neuro-
Psychopharmacology & Biological Psychiatry, 81, 105–113. https://
doi.org/10.1016/j.pnpbp.2017.10.016
Reuter, M., & Fischl, B. (2011). Avoiding asymmetry-induced bias
in longitudinal image processing. NeuroImage, 57(1), 19–21.
https://doi.org/10.1016/j.neuroimage.2011.02.076
Riva-Posse, P., Choi, K., Holtzheimer, P. E., Crowell, A. L., Garlow,
S. J., Rajendra, J. K., . . . Mayberg, H. S. (2017). A connec-
tomic approach for subcallosal cingulate deep brain stimulation
surgery: prospective targeting in treatment-resistant depression.
Molecular Psychiatry. https://doi.org/10.1038/mp.2017.59
Rocca, M., Choi, K. S., Holtzheimer, P. E., Crowell, A. L., Garlow,
S. J., Rajendra, J. K., . . . Mayberg, H. S. (2016). Impaired func-
tional integration in multiple sclerosis: A graph theory study.
Brain Structure and Function, 221(1), 115–131. https://doi.org/
10.1007/s00429-014-0896-4
Rubinov, M., & Sporns, O. (2010). Complex network measures of
brain connectivity: Uses and interpretations. NeuroImage, 52(3),
1059–1069. https://doi.org/10.1016/j.neuroimage.2009.10.003
Rush, A. J., Thase, M. E., & Dubé, S. (2003). Research issues in the
study of difficult-to-treat depression. Biological Psychiatry, 53(8),
743–753.
Sale, M. V., Mattingley, J. B., Zalesky, A., & Cocchi, L. (2015). Imag-
ing human brain networks to improve the clinical efficacy of
non-invasive brain stimulation. Neuroscience and Biobehav-
ioral Reviews, 57, 187–198. https://doi.org/10.1016/j.neubiorev.
2015.09.010
Salomons, T. V., Dunlop, K., Kennedy, S. H., Flint, A., Geraci, J.,
Giacobbe, P., & Downar,
(2014). Resting-state cortico-
J.
thalamic-striatal connectivity predicts response to dorsomedial
prefrontal rTMS in major depressive disorder. Neuropsycho-
pharmacology, 39(2), 488–498. https://doi.org/10.1038/npp.
2013.222
Singh, M. K., Kesler, S. R., Hadi Hosseini, S. M., Kelley, R. G.,
Amatya, D., Hamilton, J. P., . . . Gotlib, I. H. (2013). Anoma-
lous gray matter structural networks in major depressive disorder.
Network Neuroscience
171
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
1
1
5
7
1
0
9
2
3
4
4
n
e
n
_
a
_
0
0
0
6
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Connectome analysis in major depression
Biological Psychiatry, 74(10), 777–785. https://doi.org/10.1016/
j.biopsych.2013.03.005
Stevens, A. A., Tappon, S. C., Garg, A., & Fair, D. A. (2012).
Functional brain network modularity captures inter- and intra-
individual variation in working memory capacity. PLoS One, 7(1),
1–10. https://doi.org/10.1371/journal.pone.0030468
Tax, C. M., Otte, W. M., Viergever, M. A., Dijkhuizen, R. M.,
& Leemans, A. (2015). REKINDLE: Robust extraction of kur-
tosis INDices with linear estimation. Magnetic Resonance in
Medicine, 73(2), 794–808. https://doi.org/10.1002/mrm.25165
Tik, M., Hoffmann, A., Sladky, R., Tomova, L., Hummer,
A., Navarro de Lara, L., . . . Windischberger, C. (2017). To-
wards understanding rTMS mechanism of action: Stimulation
of the DLPFC causes network-specific increase in functional
connectivity. NeuroImage, 162, 289–296. https://doi.org/10.1016/
j.neuroimage.2017.09.022
Trevino, K., McClintock, S. M., McDonald Fischer, N., Vora, A.,
& Husain, M. M. (2014). Defining treatment-resistant depres-
sion: a comprehensive review of the literature. Annals of Clinical
Psychiatry, 26(3), 222–232.
van Randenborgh, A., Hüffmeier,
J., Victor, D., Klocke, K.,
Borlinghaus, J., & Pawelzik, M. (2012). Contrasting chronic with
episodic depression: An analysis of distorted socio-emotional
Journal of Affec-
information processing in chronic depression.
tive Disorders, 141(2–3), 177–184. https://doi.org/10.1016/j.jad.
2012.02.039
van Wijk, B. C., Stam, C. J., & Daffertshofer, A. (2010). Comparing
brain networks of different size and connectivity density using
graph theory. PLoS One, 5(10), e13701. https://doi.org/10.1371/
journal.pone.0013701
Veraart, J., Sijbers, J., Sunaert, S., Leemans, A., & Jeurissen, B.
(2013). Weighted linear least squares estimation of diffusion MRI
parameters: Strengths, limitations, and pitfalls. NeuroImage, 81,
335–346. https://doi.org/10.1016/j.neuroimage.2013.05.028
Vieta, E., & Colom, F. (2011). Therapeutic options in treatment-
resistant depression. Annals of Medicine, 43(7), 512–530. https://
doi.org/10.3109/07853890.2011.583675
Vos, S. B., Tax, C. M., Luijten, P. R., Ourselin, S., Leemans, A.,
& Froeling, M.
(2017). The importance of correcting for signal
drift in diffusion MRI. Magnetic Resonance in Medicine, 77(1),
285–299. https://doi.org/10.1002/mrm.26124
Zhang, J., Wang, J., Wu, Q., Kuang, W., Huang, X., He, Y., &
Gong, Q. (2011). Disrupted brain connectivity networks in drug-
naive, first-episode major depressive disorder. Biological Psychi-
atry, 70(4), 334–342. https://doi.org/10.1016/j.biopsych.2011.
05.018
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
1
1
5
7
1
0
9
2
3
4
4
n
e
n
_
a
_
0
0
0
6
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network Neuroscience
172