RESEARCH
Cooperating yet distinct brain networks engaged
during naturalistic paradigms: A meta-analysis
of functional MRI results
Katherine L. Bottenhorn
1
Simon B. Eickhoff
3,4
1
, Jessica S. Flannery
1
, Emily R. Boeving
, Michael C. Riedel
2
,
, Matthew T. Sutherland
, and Angela R. Laird
1
2
1Department of Psychology, Florida International University, Miami, FL, USA
2Department of Physics, Florida International University, Miami, FL, USA
3Institute of Systems Neuroscience, Medical Faculty, Heinrich Heine University Düsseldorf, Düsseldorf, Germany
4Institute of Neuroscience and Medicine, Brain & Behaviour (INM-7), Research Centre Jülich, Jülich, Germany
Keywords: Neuroimaging meta-analysis, Naturalistic paradigms, Clustering analysis, Neuro-
informatics
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
1
2
7
1
0
9
2
2
9
0
n
e
n
_
a
_
0
0
0
5
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
ABSTRACT
Cognitive processes do not occur by pure insertion and instead depend on the full
complement of co-occurring mental processes, including perceptual and motor functions.
As such, there is limited ecological validity to human neuroimaging experiments that use
highly controlled tasks to isolate mental processes of interest. However, a growing literature
shows how dynamic, interactive tasks have allowed researchers to study cognition as it more
naturally occurs. Collective analysis across such neuroimaging experiments may answer
broader questions regarding how naturalistic cognition is biologically distributed throughout
the brain. We applied an unbiased, data-driven, meta-analytic approach that uses k-means
clustering to identify core brain networks engaged across the naturalistic functional
neuroimaging literature. Functional decoding allowed us to, then, delineate how information
is distributed between these networks throughout the execution of dynamical cognition in
realistic settings. This analysis revealed six recurrent patterns of brain activation, representing
sensory, domain-specific, and attentional neural networks that support the cognitive
demands of naturalistic paradigms. Although gaps in the literature remain, these results
suggest that naturalistic fMRI paradigms recruit a common set of networks that allow both
separate processing of different streams of information and integration of relevant information
to enable flexible cognition and complex behavior.
AUTHOR SUMMARY
Naturalistic fMRI paradigms offer increased ecological validity over traditional paradigms,
addressing the gap left by studying highly interactive cognitive processes as isolated neural
phenomena. This study identifies the connectional architecture supporting dynamic
cognition in naturalistic fMRI paradigms, the first meta-analysis of a wide range of more
realistic neuroimaging experiments. Here we identify and characterize six core patterns
of neural activity that support functional segregation and integration in large-scale brain
networks. This study provides a unique investigation of the cooperating neural systems
that enable complex behavior.
a n o p e n a c c e s s
j o u r n a l
Citation: Bottenhorn, K. L., Flannery,
J. S., Boeving, E. R., Riedel, M. C.,
Eickhoff, S. B., Sutherland, M. T., &
Laird, A. R. (2019). Cooperating yet
distinct brain networks engaged
during naturalistic paradigms:
A meta-analysis of functional MRI
results. Network Neuroscience,
3(1), 27–48. https://doi.org/10.1162/
netn_a_00050
DOI:
https://doi.org/10.1162/netn_a_00050
Supporting Information:
https://doi.org/10.1162/netn_a_00050
Received: 21 July 2017
Accepted: 2 March 2018
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Author:
Angela R. Laird
alaird@fiu.edu
Handling Editor:
Edward Bullmore
Copyright: © 2018
Massachusetts Institute of Technology
Published under a Creative Commons
Attribution 4.0 International
(CC BY 4.0) license
The MIT Press
Brain networks and naturalistic paradigms
Cognitive subtraction:
Method of isolating specific aspects
of cognition that assumes cognitive
processes do not interact when they
co-occur.
INTRODUCTION
Across the life sciences, researchers often seek a balance between ecological validity and
careful laboratory control when making experimental design decisions. This entails weigh-
ing the value of creating realistic stimuli representative of real-world, interactive experiences
versus artificial, reductionist stimuli facilitating precise assessment of “isolated” mental pro-
cess of interest via cognitive subtraction. Cognitive subtraction assumes that a single added
cognitive process does not alter the other, co-occurring processes, both neutrally and cogni-
tively. As such, task-based fMRI has traditionally utilized precisely controlled tasks to study the
neurobiological substrates of cognition. However, cognition does not occur by pure insertion;
the functioning of any cognitive process is not wholly independent from other co-occurring
processes (Friston et al., 1996). Instead, cognition is highly interactive, encompassing measur-
able changes in neural activity that are dependent on the full amalgamation of relevant social,
cognitive, perceptual, and motor processes. Thus, it is perhaps unreasonable to expect find-
ings from a highly restricted assessment of a psychological construct in the scanner to fully
generalize to real-world behaviors and settings.
With advances in technology and a desire to study cognition with greater ecological valid-
ity, increasing numbers of studies are utilizing realistic, interactive, and rich stimuli in more
ecologically valid experimental designs that fit within the scanner’s confines (Hasson & Honey,
2012; Maguire, 2012; Wang et al., 2016). “Naturalistic” paradigms employ dynamic and com-
plex stimuli (Fehr et al., 2014; Kauttonen et al., 2015; Burunat et al., 2014), in terms of multi-
modal demands (Lahnakoski et al., 2012; Maguire, 2012; Nardo et al., 2014; Dick et al., 2014;
Reed et al., 2004; Bishop & Miller, 2009), or in relation to the length of the stimulus presen-
tation (Maguire, 2012; Cong et al., 2014). Specifically, the use of video games, film clips,
and virtual reality, among others, has brought a new dimension to cognitive neuroimaging
experiments, permitting researchers to study brain activity as participants engage in tasks that
more closely represent real-life demands on attention and multimodal sensory integration. Ap-
preciation of such attention and integration processes necessitates more complex stimuli than
simple static images presented on a screen. For example, researchers have studied spatial nav-
igation with virtual reality environments as complex as the city of London (Spiers & Maguire,
2006) and as classic as a virtual radial arm maze (Marsh et al., 2010). Similarly, social cog-
nition has been probed with displays of human social interactions from a dramatic, social
television drama (Spunt & Lieberman, 2012) to clips of facial expressions with little context (Li
et al., 2015).
Everyday activities, such as navigation or social observation, involve the integration of pro-
cesses associated with object recognition, speech comprehension, motor control, and spatial
orienting, which all require the interpretation of dynamic signals often from more than one
sensory modality (e.g., audiovisual film watching or visuotactile image tracing) and neces-
sitate different attentional demands compared with the simplistic stimuli used in traditional
fMRI experiments (Giard & Peronnet, 1999; McGurk & MacDonald, 1976; Sailer et al., 2000;
Spence, 2010). Recently, this trend has produced open-source efforts such as studyforrest, a
freely available dataset of MRI scans, eye-tracking, and extensive annotations, using the movie
Forrest Gump as a rich, multimodal stimulus (studyforrest.org; Hanke et al., 2016, 2015, 2014).
Although studies of participants freely viewing films or navigating virtual environments have
been used since the early days of fMRI, the naturalistic studies represent a small portion of
the overall task-based fMRI literature (Beauregard et al., 2001; Burgess et al., 2001; Maguire,
2012). Despite offering advantages, the growing body of naturalistic fMRI research has yet to
Network Neuroscience
28
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
1
2
7
1
0
9
2
2
9
0
n
e
n
_
a
_
0
0
0
5
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Brain networks and naturalistic paradigms
be quantitatively assessed, and little is known of how the neural bases of these tasks support
complex information processing and behavioral demands.
Here, we applied an unbiased, data-driven, meta-analytic approach to quantitatively explore
and classify knowledge embedded in the naturalistic fMRI literature. Using an approach devel-
oped by Laird et al. (2015), we capitalized on the wealth and flexibility of published naturalistic
paradigms and investigated recurrent patterns of brain activation reported across a wide variety
of tasks and behaviors of interest. This method is based on the premise that functionally similar
tasks engage spatially similar patterns of brain activity and that by clustering activation patterns
from experimental contrasts, similar experimental paradigms can be identified. Naturalistic
paradigms are uniquely rich here because of the multitude of component processes contribut-
ing to realistic behavior that can be illuminated by modeling strategies in data analysis. To this
end, we extracted relevant information about the stimuli and task demands of these paradigms
and assessed motifs in the arrangement of this information, with respect the data-driven clus-
tering analysis, to determine which paradigm aspects elicited activation patterns that subserve
common and dissociable cognitive processes. Although naturalistic paradigms vary greatly and
are designed to probe a wide range of psychological constructs and behaviors, we hypothe-
sized that complex, multisensory processing are associated with a set of core neural networks
engaged by similar content domains and task demands.The objectives of this study were to first
elucidate core brain networks engaged by the myriad processes that underlie behavior during
naturalistic fMRI paradigms and then to characterize how information processing is potentially
distributed between these networks to facilitate complex behaviors in realistic settings.
METHODS
Naturalistic fMRI Paradigms
Here, “naturalistic” paradigms were operationally defined as tasks employing any stimulus that
demanded continuous, real-time integration of dynamic streams of information. This defini-
tion excludes any paradigms based on still-frame stimulus presentation, which intrinsically
impose static constraints that are rarely present in the world and, thus, limit ecologically
Importantly, a key distinction of naturalistic tasks is that stimuli are continu-
their validity.
ously presented across the duration of the task, whereas other tasks in the literature rely on
repeated trials of stimuli. As real-world behavior contextually involves all sensory modali-
ties, we included naturalistic tasks in which such stimuli were presented via the visual, audi-
tory, or tactile modalities or any combination thereof. Visual naturalistic tasks require either
a real-time interaction with visual stimuli, in the case of video games and virtual reality, or the
continuous integration of real-time information, such as during film viewing. Auditory tasks,
including the perception of music and spoken stories, similarly require the continuous inte-
gration of, and often interaction with, real-time information. Our operational definition also
included tactile naturalistic paradigms, which involve the manipulation and recognition of
physical objects. During these tactile tasks, participants gather and integrate sensory infor-
mation to create a mental representation of the object and, if necessary, form an appropriate
behavioral response. Lastly, we note the inclusion of multisensory tasks. As in life, many natu-
ralistic experiments simultaneously present auditory, visual, and tactile information, and such
tasks demand the real-time integration of information from multiple sensory modalities.
Literature Search, Filtering, and Annotation
An extensive literature search was performed to amass a corpus of naturalistic fMRI studies
that were published since the emergence of fMRI in 1992. To identify published naturalistic
Network Neuroscience
29
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
1
2
7
1
0
9
2
2
9
0
n
e
n
_
a
_
0
0
0
5
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Brain networks and naturalistic paradigms
fMRI studies, PubMed searches were carried out by focusing on stimulus types common to
naturalistic research (e.g., video games, film, virtual reality). The first search string, performed
on January 13, 2016, used the following string to identify relevant studies by their titles and
((“naturalistic”[Title/Abstract] OR “real-world”[Title/Abstract] OR “ecologically
abstracts:
valid”[Title/Abstract] OR “true-to-life”[Title/Abstract] OR “realistic”[Title/Abstract] OR “video
game”[Title/Abstract] OR “film”[Title/Abstract] OR “movie”[Title/Abstract] OR “virtual reality”
[Title/Abstract]) AND (“fMRI”[Title/Abstract] OR “functional magnetic resonance imaging”
[Title/Abstract]) AND (“Humans”[MeSH])). This search yielded 679 studies ( January 2016),
some of which utilized stimulus types that we had not included in our initial query, includ-
ing music, speech, and tactile objects. To identify any studies using these tasks that may
not have been returned by the initial query, a second search was performed on January 20,
2016, using the string ((“music”[Title/Abstract] OR “speech”[Title/Abstract] OR “spoken”[Title/
Abstract] OR “tactile object”[Title/Abstract]) AND (“naturalistic”[Title/Abstract] OR “real-
world”[Title/Abstract] OR “ecologically valid”[Title/Abstract] OR “true-to-life”[Title/Abstract]
OR “realistic”[Title/Abstract]) AND (“fMRI”[Title/Abstract] OR “functional magnetic resonance
imaging”[Title/Abstract]) AND “Humans”[MeSH]). This secondary search returned 48 studies,
some of which were included in the results of the first search. The two sets of search results
were pooled to identify 754 unique studies, which were then reviewed and filtered to identify
studies utilizing naturalistic paradigms as defined above.
Each of 754 candidate studies was first screened and then reviewed according to the
following exclusion criteria (Figure 1; Moher, Liberati, Tetzlaff, Altman, & Altman, 2009). The
screening process examined the Abstracts and Methods of each paper to exclude nonnatural-
istic tasks in which static, timed blocks of stimuli were presented with a well-defined window
for participant response. In this step, we also excluded studies that assessed training or learn-
ing across multiple trials or across some period of practice (e.g., pre- vs. post contrasts), as our
focus was on neural underpinnings of the tasks themselves and not training-induced changes
thereof. In determining eligibility of each paper, studies of participants under the age of 18 or of
participants with any history of neurological or psychiatric diagnosis were excluded. After this
study-level examination, we then inspected each reported experimental contrast within each
paper. In this context, “experiment” represents each statistical parametric image presented, as
the result of some functional image data analysis, such as contrasting experimental conditions
(Fox et al., 2005). Experiments from analyses that used an a priori region(s) of interest to inves-
tigate activation or functional connectivity were omitted permitting identification of whole-
brain neural networks. We also excluded contrasts modeling ANOVA interaction-specific
activations because of the inherent complexity of such effects. In this step, any studies/contrasts
that did not meet the minimum requirements for coordinate-based meta-analysis, reporting
the brain activation locations in a three-dimensional, standardized coordinate space, were
discarded.
During inspection of each contrast, one study associate (KLB) manually annotated each
experiment with terms that described the experimental design with respect to stimulus type
utilized, sensory modality engaged, and the task nature. These terms described the salient
aspects of the stimuli and behaviors associated with each individual experimental contrast
from the corpus of naturalistic paradigms, annotating the particular aspects of the tasks high-
lighted by each modeled experimental contrast, and not the intended psychological construct
interrogated by the original report. These manual annotations were then independently re-
viewed and confirmed by a second study associate ( JSF) to assure consistency and accuracy.
Any disagreements or inconsistencies between KLB and JSF were resolved following a final
conversation between the two associates.
Network Neuroscience
30
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
1
2
7
1
0
9
2
2
9
0
n
e
n
_
a
_
0
0
0
5
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Brain networks and naturalistic paradigms
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
1
2
7
1
0
9
2
2
9
0
n
e
n
_
a
_
0
0
0
5
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
PRISMA flow chart:
Describes how papers are
systematically reviewed for
inclusion, specified by the Preferred
Reporting Items for Systematic
Review and Meta-Analysis.
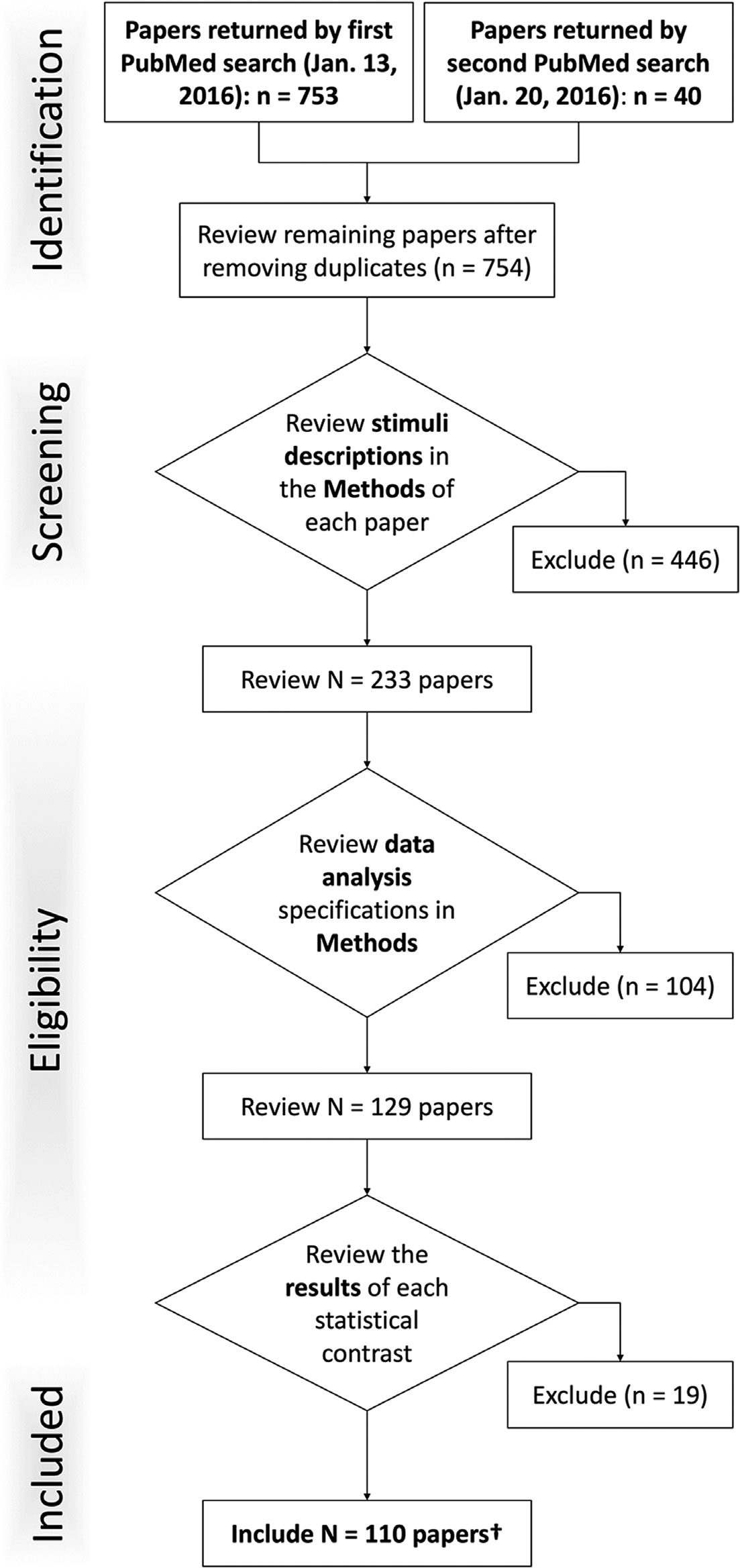
Figure 1. PRISMA flow chart of inclusion and exclusion criteria. Each of the experiments returned
by the PubMed queries were screened according to this schematic.
Network Neuroscience
31
Brain networks and naturalistic paradigms
Modeled activation (MA) maps:
Whole-brain maps recreating results
from published neuroimaging studies
as Gaussian kernels centered at each
reported x-, y-, and z-coordinate of
activation.
K-means clustering:
Method by which n experiments are
assigned to one of k clusters, based
on proximity to the cluster’s center.
Experimental Design and Statistical Analysis
Following the identification of relevant papers and experiments/
Modeled activation maps.
contrasts, reported brain activation coordinates were extracted. All Talairach atlas-based coor-
dinates (Talairach & Tournoux, 1988) were converted to Montreal Neurological Institute (MNI)
space (Collins et al., 1994; Evans et al., 1993) using the tal2icbm transformation (Lancaster
et al., 2007; Laird et al., 2010). Probabilistic modeled activation (MA) maps were created
from the foci reported in each individual contrast by modeling a spherical Gaussian blur around
each focus, with full width at half maximum determined by the number of subjects in each
experiment in order to represent the uncertainty induced by the inherent variability from indi-
vidual differences and between-lab differences (Eickhoff et al., 2009). These MA maps were
concatenated into an array of n experiments by p voxels, which was then analyzed for pairwise
correlations that reflected the degree of spatial similarity between the MA maps from each of
the n experiments and those of every other experiment. The resultant n × n correlation matrix
represented the similarity of spatial topography of MA maps between every possible pair of
experiments.
Individual naturalistic experiments (n MA maps) were then clas-
K-means clustering analysis.
sified into K groups based on their spatial topography similarities. The k-means clustering
procedure was performed in Matlab (Mathworks, R2013b for Linux), which grouped experi-
ments by pairwise similarity, calculating correlation distance by 1 minus the correlation be-
tween MA maps (from the aforementioned correlation matrix) and finding the “best” grouping
by minimizing the sum of correlation distances within each cluster (code available at
https://github.com/62442katieb/meta-analytic-kmeans). This approach begins by choosing K
arbitrary maps as representative centroids for each of the K clusters and assigning experiments
to each cluster based on the closest (most similar) centroid. This process continued iteratively
until a stable solution was reached.
Solutions were investigated for a range of K = 2–10 clusters. Once the clustering analysis
was complete for all K, we compared each solution with the neighboring solutions and as-
sessed for improvement across parcellation schemes by using four metrics describing cluster
separation and stability (Bzdok et al., 2015; Eickhoff et al., 2016a). This allowed us to objec-
tively select the number of clusters that most optimally divided the dataset. The first metric,
average cluster silhouette across clustering solutions, assessed the separation between clus-
ters and described whether clusters were distinct or overlapping. A higher silhouette value
indicates that greater separation is ideal and that each experiment fits well into its cluster,
with lower misclassification likelihood of fringe experiments into neighboring clusters. Sta-
bility is indicated by a relatively minimal change in silhouette from one solution (K) to the
next (K + 1), indicated by the positive derivative of the silhouette score closest to zero, with
greatest stability evidenced by the smallest change between two points. Second, we consid-
ered the consistency of experiment assignment by comparing the ratio of the minimum number
of experiments consistently assigned to a cluster relative to the mean number of experiments
consistently assigned to that cluster. In this case, only ratios above 0.5, in which at least half
of the experiments were consistently assigned, were considered viable solutions. Third, the
variation of information was quantified, which compared the entropy of clusters with the
mutual information shared between them for each solution K and its K − 1 and K + 1 neigh-
bors. A large decrease in variation of information from K − 1 to K and increase from K to
K + 1, a local minimum in the plot of variation of information across K, indicated a decrease
in overlap between solutions and, thus, stability of solution K. In this case, “large” is defined,
Network Neuroscience
32
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
1
2
7
1
0
9
2
2
9
0
n
e
n
_
a
_
0
0
0
5
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Brain networks and naturalistic paradigms
Activation likelihood
estimate (ALE):
Process by which the voxel-wise
union of MA maps is calculated
to create a meta-analytic,
statistical map of brain
activation.
Meta-analytic groupings (MAGs):
Each of k clusters of experiments,
cluster assignment via k-means
clustering.
Functional decoding:
The use of previous neuroimaging
results to understand the probability
of some mental process, given a
pattern of brain activation.
too, in relative terms, with the largest decrease indicating greatest stability of the solutions
considered. Finally, we computed a hierarchy index for each solution, which assessed how
clusters split from the K − 1 to K solution to form the additional cluster. A lower hierarchy
index indicated that clusters present in K stemmed from fewer of the clusters present in K − 1,
another indication of stability in groupings demonstrated by a local minimum across values of
K. An optimal clustering solution is one that demonstrated minimal overlap between clusters
(i.e., high silhouette value), while exhibiting relative stability in comparison with the previous
and next solutions (i.e., consistency > 0.5, a local minimum in variation of information, and
lower hierarchy index than previous).
From the identified optimal clustering solution, we probed the
Meta-analytic groupings.
underlying neural topography associated with each of the K groups of experiments (Laird et al.,
2015). To this end, the activation likelihood estimate (ALE) meta-analysis algorithm (Turkeltaub
et al., 2002; Laird et al., 2005) was applied to generate a map of convergent activation for
each grouping of experiments with similar topography. The ALE algorithm includes a weight-
ing of the number of subjects when computing these maps of convergent activation and
accounts for uncertainty associated with individual, template, and registration differences
between and across experiments (Eickhoff et al., 2009; Turkeltaub et al., 2012). The union of
these probability distributions was used to calculate ALE scores, a quantitative assessment
of convergence between brain activation across different experiments, which was compared
against 1,000 permutations of a null distribution of random spatial arrangements (Eickhoff
et al., 2012). These ALE values for each meta-analytic grouping of experiments were thresh-
olded at p < 0.01 (cluster-level corrected for family-wise error) with a voxel-level, cluster-
forming threshold of p < 0.001 (Eickhoff et al., 2016b; Woo et al., 2014). The resultant ALE
maps thus reflected the convergent activation patterns within each of the K clusters. The ex-
perimental K clusters are hereafter referred to as meta-analytic groupings (MAGs), representing
meta-analytic groups of experiments demonstrating similar activation patterns.
Functional Decoding
Once we elucidated convergent activation patterns within MAGs, we sought to gain insight
into what aspects of the naturalistic paradigms were most frequently associated with each MAG
via functional decoding. Functional decoding is a quantitative, data-driven method by which
researchers can infer which mental processes are related to activation in a specific brain region
(or set of brain regions) across published fMRI studies. We chose to use two complementary
functional decoding approaches, one based on our study-specific, subjective manual annota-
tions mentioned above, and another based on the objective, automated annotations provided
by the Neurosynth database for over 11,000 functional neuroimaging studies (Yarkoni et al.,
2011; Neurosynth.org). First, the manually annotated terms associated with each experiment
were grouped into the MAGs identified above and were assessed by frequency of occurrence
in each MAG. The distribution of stimulus modality, stimulus type, and salient terms across
MAGs allowed us to evaluate the relationship between activation patterns and the aspects of
naturalistic paradigms that elicited them. Second, we included an automated, data-driven an-
notation method using Neurosynth, which includes automatically extracted terms that occur
at a high frequency in the abstract of each archived study. To functionally decode our MAGs,
we compared the MAGs’ activation patterns with those reported across published neuro-
imaging papers in the Neurosynth database. To this end, we uploaded each ALE map to Neuro-
Vault, a web-based repository for 3D statistical neuroimaging maps that directly interfaces
with Neurosynth (Gorgolewski et al., 2015; NeuroVault.org,). NeuroVault enables “functional
Network Neuroscience
33
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
1
2
7
1
0
9
2
2
9
0
n
e
n
_
a
_
0
0
0
5
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Brain networks and naturalistic paradigms
decoding” by correlating unthresholded uploaded maps with term-specific meta-analytic
maps extracted from Neurosynth’s database of published functional neuroimaging studies. The
Neurosynth functional decoding results were exported as a set of terms and correlation values
representing how well the spatial distribution of activation associated with each term in the
database matched the activation pattern of the uploaded map.
Both sets of terms (i.e., obtained via manual and automated approaches) were evaluated to
assess the specific aspects of naturalistic paradigms associated with each MAG. The Neuro-
synth terms representing broad behavioral aspects across fMRI studies that elicit similar brain
activation profiles provides both an unbiased description of the experiments engaging each
MAG, as well as a comparison of our corpus of studies with the broader literature. On the other
hand, manual annotation provides a more concise, accurate description of the paradigms,
although it is predisposed to the subjective bias of human annotation. The results of this two-
pronged functional decoding approach were designed to describe the processes that engage
brain networks similar to each MAG and how these processes may be similar or different in
naturalistic fMRI studies compared with the broader functional neuroimaging literature. The
distribution of stimulus modalities and types across MAGs was assessed, too. Together, the
functional decoding results and distributions of different stimuli were interpreted to provide
insight into how information processing is functionally segregated across cooperating neural
systems during naturalistic tasks.
RESULTS
The literature search yielded a combined set of 110 studies that reported coordinates of brain
activation from naturalistic fMRI tasks among healthy adults (Figure 1; PubMed IDs avail-
able in Supporting Information Table S1, Bottenhorn, Flannery, Boeving, Riedel, Eickhoff,
Sutherland, & Laird, 2019). The final dataset included activation foci from 376 experimental
contrasts (N = 1, 817 subjects) derived from tasks using a variety of stimulus types and sensory
modalities. Across our corpus of naturalistic fMRI experiments, approximately 55% assessed
a single stimulus modality, including 40% visual stimuli, 13% auditory, and 1% tactile.
Conversely, 45% of experiments utilized multisensory stimuli, including 41% that employed
audiovisual stimuli, 2% in which a visual stimulus was paired with painful, tactile stimuli, and
1% pairing visual and nonpainful tactile stimuli (Table 1). Of the visual experiments, 69%
involved a motor response, as did 25% of the audiovisual experiments, ranging from a button
press to joystick and object manipulation. The stimulus types most frequently used across the
included experiments were films (45%), virtual reality (32%), speech (9%), and music (6%)
(Table 2).
Table 1. Distribution of stimulus modalities across the naturalistic corpus
Stimulus modality
Auditory
Audiovisual
Visual
Visual + tactile (pain)
Visual + tactile
Tactile
Note. Paradigms engaged auditory, visual, and tactile sensory modalities, both separately and
in combination.
Number of experiments
50 (13%)
154 (41%)
150 (40%)
9 (2%)
5 (1%)
4 (1%)
Network Neuroscience
34
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
1
2
7
1
0
9
2
2
9
0
n
e
n
_
a
_
0
0
0
5
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Brain networks and naturalistic paradigms
Table 2. Distribution of stimulus types across the naturalistic corpus
Stimulus type
Film
Virtual reality
Speech
Music
Video game
3D image
Tactile
Picture
Sounds
Note. Within each stimulus modality, multiple types of experimental stimuli were included across
the dataset.
Number of experiments
169 (45%)
121 (32%)
32 (9%)
21 (6%)
13 (4%)
6 (2%)
6 (2%)
4 (1%)
1 (<1%)
k-Means Clustering Solutions
MA maps were created for each contrast and then clustered to identify groups with similar ac-
tivation topographies. For completeness, the k-means clustering solutions for K = 2 – 10 clus-
ters were quantitatively evaluated across four metrics to identify an optimal solution (Figure 2).
When considering the average silhouette metric (Figure 2A), values generally increased as
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
1
2
7
1
0
9
2
2
9
0
n
e
n
_
a
_
0
0
0
5
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
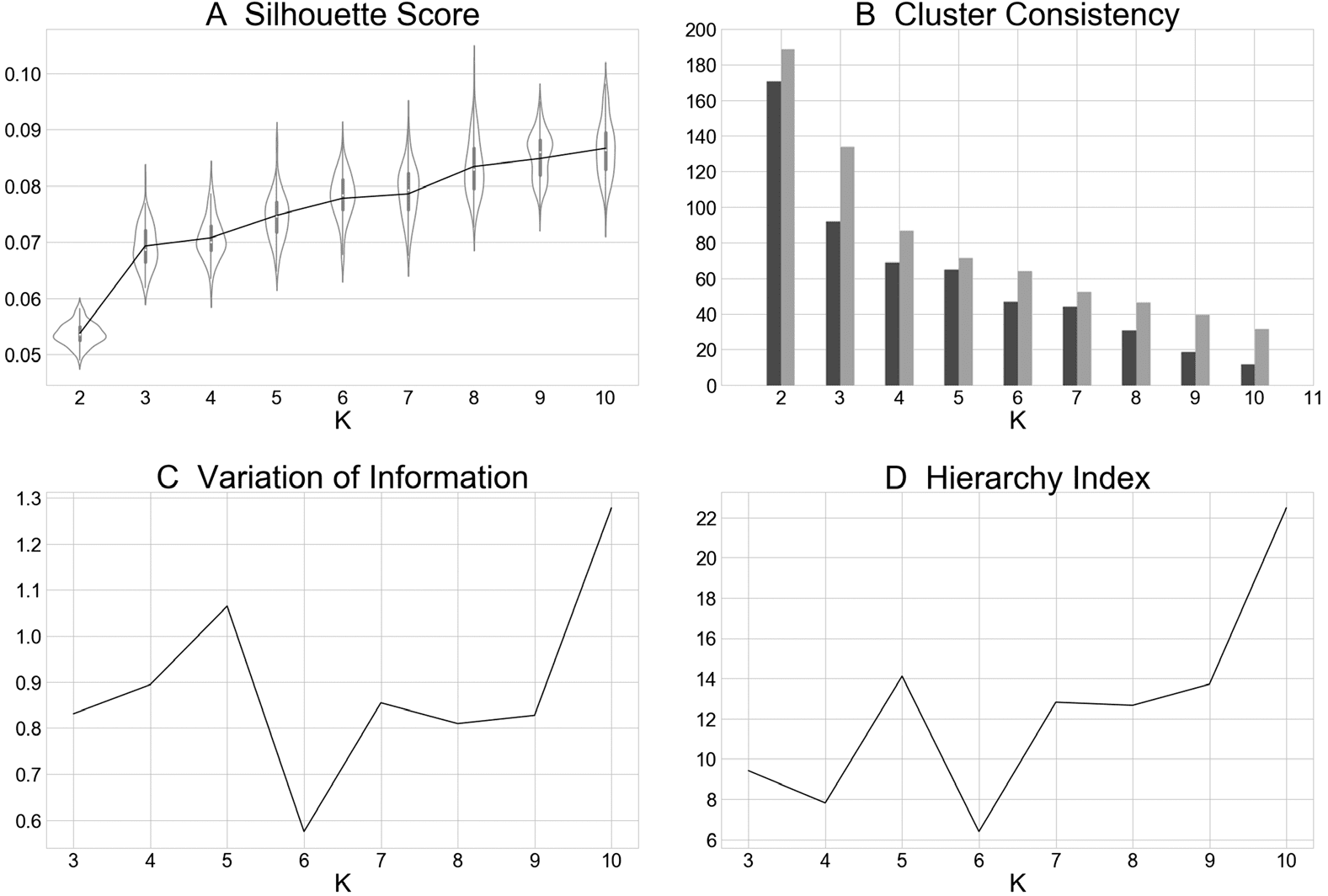
Figure 2. Metrics computed for K = 2 – 10 clustering solutions. (A) The average cluster silhouette
for each solution K from 2 to 10 clusters, showing the distribution of average silhouette values at
each value of K, resampled 100 times leaving one random experiment out each time. (B) Consis-
tency in experiments assignment to clusters, plotting the minimum consistently assigned clusters
(C) The change in variation of information, a
next to the mean of consistently assigned clusters.
distance metric, from the K − 1 to K and from K to K + 1.
(D) The hierarchy index for each of
K clustering solutions, which provides information about how clusters in the K solution stemmed
from clusters in the K − 1 solution.
Network Neuroscience
35
Brain networks and naturalistic paradigms
K increased and the smallest increase was observed between K = 6 to K = 7, indicating little
additional separation between clusters gained by moving from 6 to 7 clusters. With respect
to the consistency of assigned experiments metric (Figure 2B), each of the solutions K = 2 –
10 met the stability requirement whereby the minimum number of experiments included in
any iteration of the solution was at least 50% of the mean number of experiments included
across iterations. The variation of information metric (Figure 2C), suggested the stability of a
6-cluster solution as parameter value decreases were observed when moving from K = 5 to
K = 6, combined with parameter increases when moving from K = 6 to K = 7, indicating
that a 6-cluster solution demonstrates relative stability. The hierarchy index metric (Figure 2D)
further corroborated a 6-cluster solution, as a local minimum as observed at K = 6. Because
of agreement across these metrics, we chose to proceed with the K = 6 solution.
Meta-Analytic Groupings
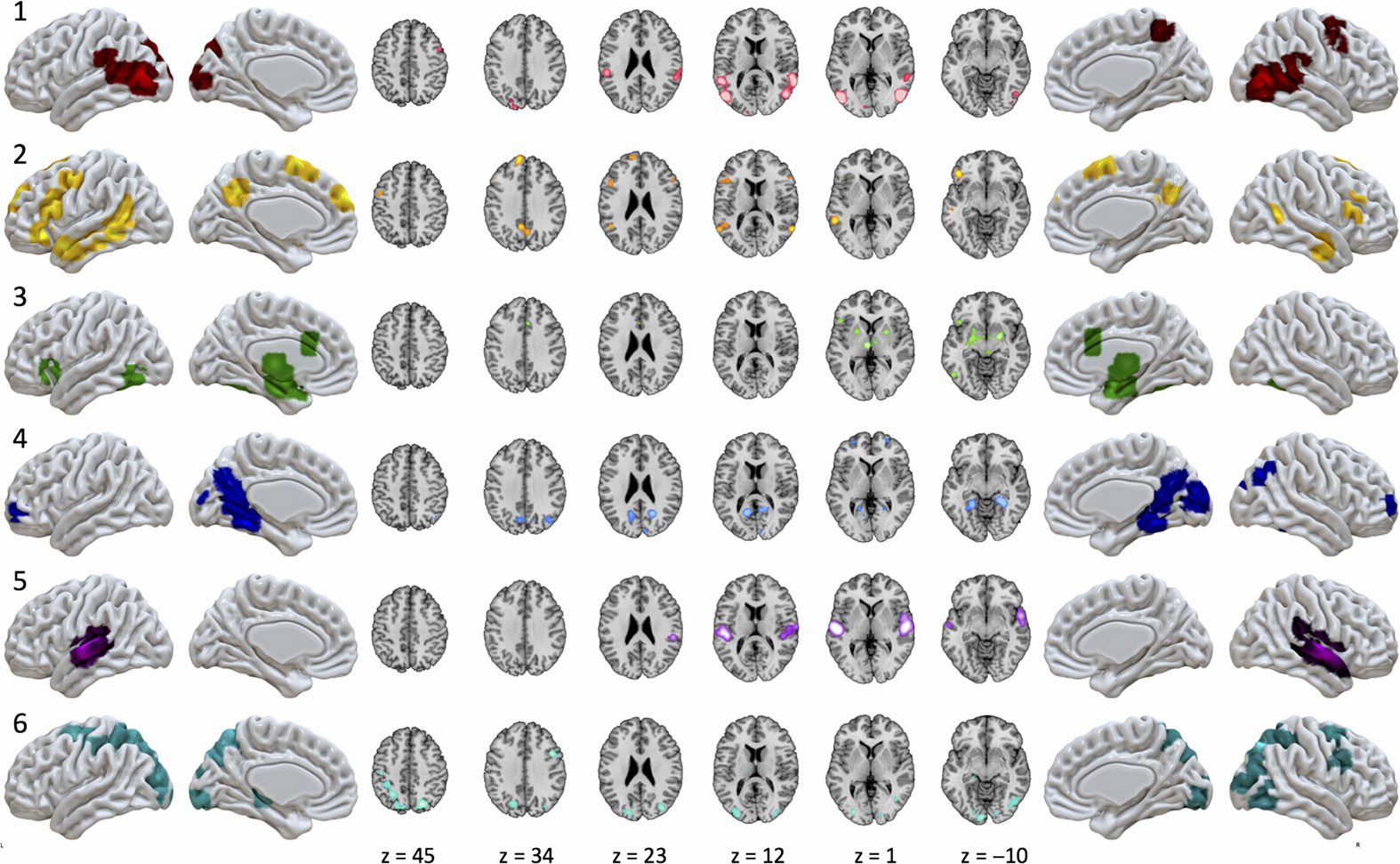
The optimal clustering solution yielded six MAGs of experiments in our corpus, suggesting
similarities in brain activation across this sample of the naturalistic literature coalesce into
six distinct patterns. The number of experiments that were clustered into each MAG ranged
from 50 to 83 experiments (mean = 62.67; SD = 12.46). ALE maps of the six MAGs were
generated and demonstrated little overlap in activation patterns, suggesting distinct patterns of
recurrent activation across our set of naturalistic experiments (Figure 3; Supporting Information
Table S2, Bottenhorn et al., 2019). Whereas some of the MAGs exhibited focal patterns of
convergent activation, restricted to a single or neighboring gyri (e.g., MAG 1 and 5), others
presented with distributed convergence across multiple lobes (e.g., MAG 2 and 6). Most of
the resulting MAGs were restricted to cortical activation patterns, although MAG 3 exhibited
convergent activation in subcortical and brainstem regions (results available on NeuroVault at
https://neurovault.org/collections/3179/ ).
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
1
2
7
1
0
9
2
2
9
0
n
e
n
_
a
_
0
0
0
5
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. Convergent activation patterns of MAGs from the naturalistic corpus. ALE meta-analysis
of experiments in each MAG yielded six patterns of convergent activation.
Network Neuroscience
36
Brain networks and naturalistic paradigms
MAG 1 included convergent activation in the bilateral posterior temporal areas, including
portions of the inferior, middle, and superior temporal gyri, extending into the inferior pari-
etal lobule and into the middle occipital gyrus, as well as in the left supramarginal gyrus,
right precentral and middle frontal gyri, and in the bilateral precuneus. MAG 2 exhibited con-
vergence in left inferior frontal gyrus, left precentral gyrus, anterior and posterior aspects of
the middle temporal gyrus, precuneus, in addition to both the left and right superior frontal
gyri. MAG 3 demonstrated a largely symmetric convergence pattern across multiple sub-
cortical structures including bilateral amygdalae, putamen, thalamus, parahippocampal gyrus,
and periaqueductal gray, with cortical clusters observed in the left inferior frontal sulcus and
inferior frontal gyrus, bilateral anterior cingulate cortex, and bilateral fusiform gyri. MAG 4
exhibited convergent activation in bilateral medial temporal lobes, parahippocampal regions,
bilateral precuneus, retrospenial posterior cingulate cortex, occipital regions including the
lingual gyrus, right calcarine sulcus, and cuneus, in addition to a small, bilateral portion of
the middle frontal gyri. MAG 5 showed convergence in the bilateral superior temporal gyri.
MAG 6 demonstrated convergence in the bilateral superior frontal sulci, intraparietal sulci,
and superior parietal lobules as well as convergence in higher order visual processing areas in
the middle occipital and lingual gyri.
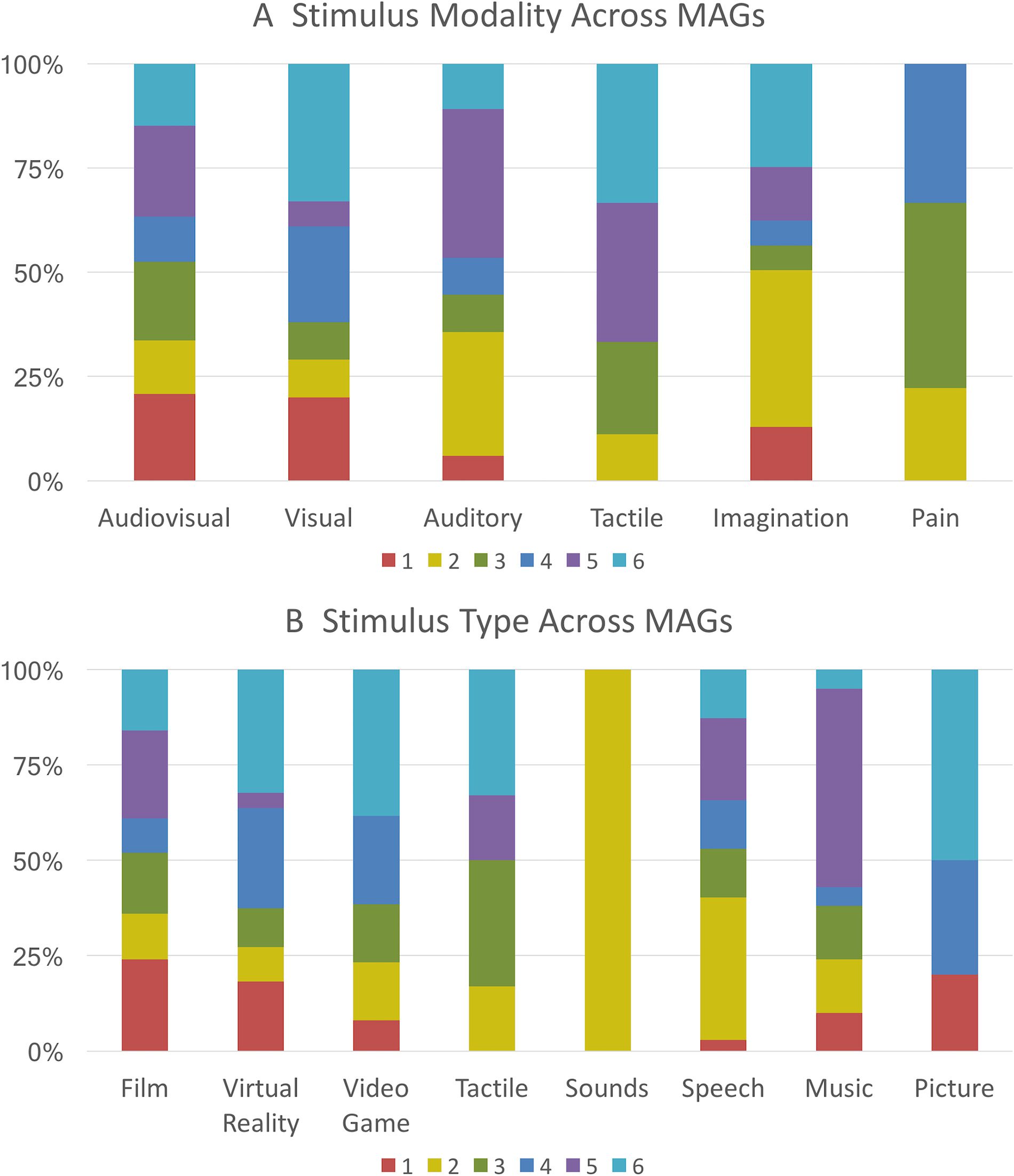
Stimulus Distribution Across MAGs
Each stimulus modality was represented in multiple MAGs, but modalities were not evenly
distributed across MAGs (Figure 4A). Experiments utilizing audiovisual tasks were somewhat
uniformly distributed across the MAGs, with a slightly higher proportion of audiovisual tasks
in MAGs 1, 3, and 5. In contrast, more than half of the experiments using auditory tasks were
grouped into MAGs 2 and 6. Notably, more experiments based on auditory and audiovisual
stimuli were clustered into MAG 5 than any other MAG. Experiments in which participants
experienced physical pain were not present in MAGs 1, 5, and 6, but distributed nearly evenly
among MAGs 2 through 4, with a slightly higher portion in MAG 3. More than half of experi-
ments that used tactile stimuli were grouped into MAG 5 and 6. Visual experiments were more
evenly distributed across clusters, although there was a markedly smaller proportion in MAG 5
than any other MAG. One stimulus type, “sounds,” was represented only once across the cor-
pus and was, thus, excluded from Figure 4. The complete distribution of stimulus modalities
across MAGs is provided in Supporting Information Table S3 (Bottenhorn et al., 2019).
As with stimulus modality, most stimulus types showed unequal, but not necessarily selec-
tive, distribution across MAGs (Figure 4B). Film-based experiments were uniformly distributed
across MAGs, and tasks utilizing spoken stimuli were more frequently grouped into MAGs 2
and 5. Again, auditory stimuli were highly associated with MAG 5, as more than 50% of
music experiments and 20% of speech experiments were clustered into MAG 5. Experiments
that required subjects to play video games were most often grouped into MAGs 4 and 6. Experi-
mental contrasts, which included a condition in which participants received tactile stimulation
or manipulated tactile objects, were most prevalent in MAGs 3 and 6. A detailed distribution
of stimulus types across MAGs is shown in Supporting Information Table S4 (Bottenhorn et al.,
2019).
Functional Decoding
Two approaches for functionally decoding each MAG, manual and automated annotations,
were performed to develop a functional interpretation of each MAG’s association with aspects
of naturalistic paradigms.
Network Neuroscience
37
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
1
2
7
1
0
9
2
2
9
0
n
e
n
_
a
_
0
0
0
5
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Brain networks and naturalistic paradigms
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
1
2
7
1
0
9
2
2
9
0
n
e
n
_
a
_
0
0
0
5
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. Distribution of stimulus modalities and types across MAGs. (A) The presence of each
sensory modality across the corpus that is associated with each MAG. (B) The proportion of each
stimulus type present within the corpus that is associated with each MAG. These percentages rep-
resent the proportion modality or stimulus type present in each MAG, compared with the total count
of that modality or stimulus type across all MAGs.
Manual annotations. Our manual annotations utilized a list of 26 corpus-specific metadata
terms, which captured salient features of the naturalistic design, rather than the psychological
constructs assumed to be involved. Table 3 displays each of these terms and their frequency
of occurrence across MAGs and across the entire corpus (Column = “Total”), highlighting
which terms described the largest number of experiments (e.g., “navigation,” “visual features,”
“emotional film,” “attention”), as well as those that accounted for a minimal number of experi-
ments (e.g., “violence,” “tactile,” “pain”). Values in Table 3 indicate the percent of experiments
labeled with each term, or the base rate of each term throughout the dataset, keeping in
mind that each experiment was labeled with only one or two terms. Once the experiments
Network Neuroscience
38
Brain networks and naturalistic paradigms
Table 3. Manual functional decoding results across meta-analytic groupings
Term
Total
MAG 1
MAG 2
Frequency per MAG
MAG 3
MAG 4
MAG 5
MAG 6
Anthropomorphic
Attention
Auditory features
Congruence
Emotional film
Encoding
Erotic
Faces
Imagination
Inference
Language
Movement
Music
Narrative
Navigation
Negative valence
Pain
Positive valence
Recognition
Retrieval
Social
Spatial memory
Tactile
Video game
21
50
17
22
61
24
15
21
23
11
47
14
21
30
81
27
9
11
12
23
26
10
9
15
3% 10†
7% 18*†
3%
3%
0
7
9% 17*
4%
2%
3%
3%
2%
7%
2%
3%
1
1
5
4
4
9
4
2
4%
12%
5
8*
4%
1%
2%
2%
3%
4%
1%
1%
2%
8
0
2
0
1
9
0
0
1
48%
36%
0%
32%
28%
4%
7%
24%
0
3
1
4
8
3
0
2
17%
6
6†
36%
19% 11*
29%
10%
17%
10%
30%
0%
18%
0%
4%
35%
0%
0%
7%
0
3
5
7
3
2
4
4
5
8
2
1
2
0%
6%
2
2
1
0
6%
18%
13% 17*†
13%
1
8†
2
2
0
3
1
3
0%
10%
26%
55%
23%
0%
14%
17%
1
9% 10*
11%
22%
36%
33%
22%
31%
20%
11%
13%
9
4†
2
2
2
2
0
1
2
10%
4%
6%
0%
28%
4%
53%
10%
9%
0%
6%
7%
14%
2
8
2
2
4
6
1
2
2
1
4
2
1
4
3%
12% 26*†
33%
1
44%
18%
17%
9%
8%
0%
11%
13%
3
2
1
4
3
7†
0
4
10%
4
16% 10*
12% 12*†
9%
3
7% 11*
25%
7%
10%
9%
0
0
8†
4
2
0
9%
9% 14*†
14%
5% 11*†
13% 11*†
32%
2
4%
33%
18%
8%
17%
12%
70%
0%
27%
4
0
1
1
1
1
0
3
0
19%
20%
3
9
71%
14%
1
6
4†
18%
0% 13*†
0%
38%
5
2
17%
0%
30%
14%
52%
5
0
6
5
1
37%
4
2% 28*
15%
0%
9%
8%
2
0
0
4
4% 10
4%
0%
33%
0%
3
1
4
6
14%
18%
6%
27%
7%
54%
33%
10%
22%
0%
13%
36%
5%
13%
35%
7%
0%
0%
33%
43%
12%
10%
44%
40%
1%
8
65
13%
35%
1
10% 23*
Violence
Visual features
Note. The relative contributions of each manually derived metadata term (e.g., term frequencies) were computed for all MAGs,
controlling for the base rate by dividing each term’s per-MAG count by that term’s total count across the corpus. Base rates are
provided as the total count for each term.
*Significant forward inference at pcorrected < 0.05.
†Significant reverse inference at pcorrected < 0.05 (corrected for false discovery rate).
0%
15% 18*†
0
25%
15% 10*
2
0% 10
13%
28%
25%
6%
25%
2
0
2
4
1
were clustered into six MAGs, we evaluated the relative contributions of each term per MAG,
controlling for base rate by dividing each term’s per-MAG count by that term’s total count
across the corpus (Table 3). We assessed, too, the ability of each term to predict whether an
experiment labeled with that term will be clustered into each MAG, (P(MAG|term)) or “forward
inference,” and the ability of belongingness to each MAG to predict whether an experiment
will be labeled with a particular term, (P(term|MAG)) or “reverse inference.” These outcomes
provide the association of each term with each MAG (Table 3). Some of the terms in the manual
annotation analysis corresponded to stimulus types in Figure 4B (e.g., per-MAG distribution
for “music” and “video game”). However, many of the manually derived terms highlighted
experimental aspects that reflect the unique and salient features of the naturalistic corpus
Network Neuroscience
39
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
1
2
7
1
0
9
2
2
9
0
n
e
n
_
a
_
0
0
0
5
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Brain networks and naturalistic paradigms
(e.g., “anthropomorphic,” “violence”) and are not included in standard neuroimaging paradigm
ontologies such as BrainMap (Fox et al., 2005) or CogPO (Turner & Laird, 2012).
To complement the manual annotation analysis, we used
Automated Neurosynth annotations.
Neurosynth’s automated annotations, which describes experiments that engage each MAG
based on published neuroimaging data, allowing comparison of our corpus with the broader
literature. MAG results were decoded in Neurosynth, yielding correlation values indicating the
similarity of the input map (i.e., each MAG’s ALE map) and maps associated with each term
from the Neurosynth database. To facilitate interpretation, the top 10 terms with the high-
est correlation values for each MAG are presented (Table 4). Terms that were near-duplicates
of terms already included in the list were removed, such as “emotion” and “emotions” if
“emotional” was higher on the list. Noncontent terms (e.g., “abstract,” “reliable”) and terms
that described brain regions, such as “insula” or “mt,” were also excluded.
The combined knowledge gained from the MAGs topog-
Functional interpretation of MAGs.
raphy, as well as the manual and automated metadata decoding analyses, provided insight
into the functional interpretation of the clustering results. Overall, the terms yielded by Neuro-
synth decoding generally agreed with the manual annotation terms in characterizing the MAGs.
Below is a summary of the six MAGs; note that reported labels do not refer to the defini-
tive function of these regions, but rather indicate how each MAG reflects differential network
contributions during naturalistic fMRI paradigms.
Manual annotations indicated that MAG 1 experiments involved attention and the process-
ing of dynamic visual features, in addition to visually presented anthropomorphic forms and
faces. Most of the stimuli in these experiments were films (Figure 4B), especially affective
films. Neurosynth results largely converged with these manual annotations, as terms includ-
ing “videos,” “body”, “observation,” and “visual motion” (Table 4) were associated with activa-
tions in MAG 1 regions. These annotations, together with the presence of convergent activation
across regions commonly associated with higher level visual processing, suggest that MAG 1
was associated with the observation of body and biological motion (Figure 3.1).
Manual annotations indicated that MAG 2 experiments involved language processing,
inference, and judgments about congruence. This MAG included relatively large proportions
of the experiments using speech, video games, and tactile stimulation (Figure 2B). Neurosynth
results supported the manual annotations’ indication that this MAG was associated with
language processing and comprehension, as terms such as “sentence,” “comprehension,”
MAG 1
MAG 2
MAG 3
MAG 4
MAG 5
MAG 6
Table 4. Automated functional decoding results from Neurosynth
corr.
NS term
0.431
Motion
0.414
Body
0.342
Static
0.300
Moving
0.299
Viewed
0.286
Visual
0.279
Visual motion
0.274
Videos
0.256
Perception
Observation
0.250
Note. The top ten Neurosynth (NS) terms are provided for each MAG, along with the corresponding Pearson’s correlation coefficient (corr.) that indicates the strength of similarity
between Neurosynth maps and each MAG.
NS term
comprehension
sentence
language
semantic
linguistic
theory mind
mental state
mind
mentalizing
language comprehension
NS term
neutral
fearful
facial
emotion
expressions
happy
angry
affective
facial expressions
neutral faces
NS term
navigation
Scenes
episodic
virtual
memory
retrieval
episodic memory
place
autobiographical
remembering
NS term
visual
spatial
attention
eye movements
execution
task
visuospatial
movements
spatial attention
hand
NS term
sounds
auditory
listening
acoustic
speech
music
pitch
spoken
tones
voice
corr.
0.74
0.732
0.711
0.675
0.669
0.625
0.612
0.590
0.572
0.568
corr.
0.417
0.408
0.375
0.351
0.336
0.318
0.309
0.306
0.304
0.289
corr.
0.324
0.316
0.294
0.278
0.276
0.270
0.258
0.208
0.201
0.201
corr.
0.446
0.437
0.435
0.434
0.431
0.404
0.401
0.397
0.395
0.385
corr.
0.555
0.451
0.441
0.415
0.406
0.403
0.381
0.360
0.359
0.350
Network Neuroscience
40
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
1
2
7
1
0
9
2
2
9
0
n
e
n
_
a
_
0
0
0
5
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Brain networks and naturalistic paradigms
“semantic,” and “mentalizing” (Table 4) were returned. These annotations and the presence of
convergent activation in predominately left lateralized regions typically associated with higher
order cognition and language suggest that MAG 2 related to language processing (Figure 3.2).
Manual annotations indicated that MAG 3 experiments involved human interactions or
affective displays, including emotional and erotic films. Films were the predominantly used
stimuli across these experiments, while most paradigms using painful stimuli were grouped
into this MAG (Figure 4B). Neurosynth results corroborated these manual annotation inter-
pretations regarding affective, aversive, and social processing, with terms such as “emotion,”
“facial expressions,” “fearful,” and “affective” (Table 4). Together, these annotations and a
convergent activation pattern involving bilateral amygdalae suggest that MAG 3 was associ-
ated with emotional processing (Figure 3.3).
Manual annotations indicated that MAG 4 heavily represented experiments involving navi-
gation through virtual reality environments, with spatial memory demands related to encoding
unfamiliar virtual landscapes for future use. A few of these experiments required language pro-
cessing as well, and half of the experiments that used 3D images were grouped into MAG 5
(Figure 4B). The manual annotations were reflected in the Neurosynth results, as similar pat-
terns of activation have been associated with “navigation,” “scenes,” “memory,” and “place.”
Additional related terms added depth to our characterization, expanding on the memory de-
mands with “retrieval,” “episodic memory,” and “remembering” (Table 4). Overall, these ex-
perimental characteristics and convergent activation in medial temporal regions and along
the visual processing stream suggest that MAG 5 was associated with navigation and spatial
memory (Figure 3.4).
Manual annotations showed that MAG 5 experiments primarily involved either film or music
stimuli (Figure 4B) and engaged either audiovisual or purely auditory processing (Figure 4A).
More than half of the included experiments that used music as stimuli were grouped into this
MAG (Figure 2B), with some stimuli involving an emotional quality (Table 3). Neurosynth
corroborated these interpretations returning terms such as “auditory,” “sounds,” “listening,”
and “speech” associated with activation of the regions in this MAG. These metadata descrip-
tions combined with convergent activation in superior temporal regions suggest this MAG’s
association with auditory processing (Figure 3.5).
Manual annotations of MAG 6 experiments implicated tasks involving visual attentional
demands and the processing of visual features, as participants engaged in video games, tactile
stimulation, and virtual reality navigation (Figure 4B, Table 3). Stimuli with high visuospatial
demands (i.e., video games, virtual reality, and pictures) were represented more by this MAG
than any other, whereas stimuli with low visuospatial demands (i.e., music and speech) were
represented the least in this MAG. Some experiments involved memory encoding, and visual
processing. Neurosynth supported this characterization returning terms including “visual,”
“attention,” “eye movements,” “saccades,” and “spatial attention” associated with activation
of the regions in this MAG (Table 4). These annotations and convergent activation in regions
resembling the dorsal attention network and areas of higher level visual processing (e.g.,
superior frontal and parietal regions, extrastriate cortex) suggest this MAG’s association with
visuospatial attention (Figure 3.6).
DISCUSSION
To characterize a core set of brain networks engaged in more ecologically valid neuroimag-
ing designs, we employed a data-driven approach that meta-analytically grouped published
Network Neuroscience
41
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
1
2
7
1
0
9
2
2
9
0
n
e
n
_
a
_
0
0
0
5
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Brain networks and naturalistic paradigms
naturalistic fMRI results according to their spatial topographies. Objective metrics suggested
that a solution of K = 6 clusters provided the most stable and disparate grouping of experiments
across the naturalistic fMRI literature, and ALE meta-analysis delineated convergent activation
across spatially distinct brain regions for each meta-analytic grouping (MAG) of experiments.
We then considered how such networks subdivide information processing by assessing the
characteristics of the constituent experiments from each MAG. Utilizing both manual and
automated functional decoding approaches, enhanced interpretations of the mental processes
associated with specific constellations of brain regions were gleaned such that the outcomes
of the two approaches generally agreed, with differences highlighting domain-specific and
domain-general processes associated with naturalistic paradigms.
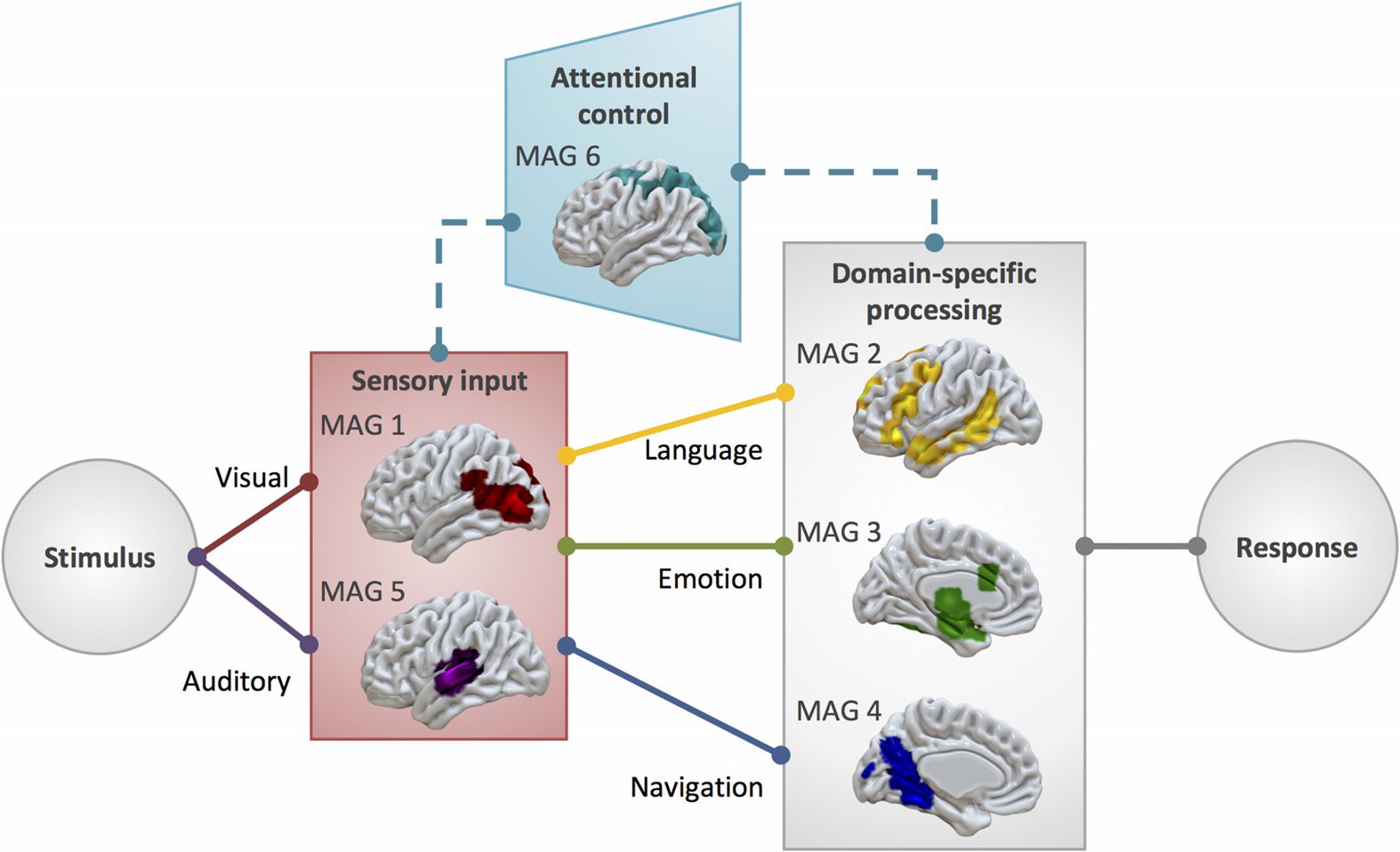
Distributed Processing for Complex Functions
Although the six identified MAGs are spatially distinct and appear to correspond with dis-
sociable mental processes, most of the included naturalistic tasks that reported more than
one statistical contrast recruited more than one MAG (66 of 86). This is consistent with func-
tional segregation and the flexible nature of the naturalistic design, demonstrating that the
manipulation of different contrasts can identify distinct networks that likely cooperate to
successfully perform a complex task. Further indicative of coordinated interactions and dis-
tributed processing, each MAG included experiments that utilized different task modalities and
task types. Overwhelmingly, the identified MAGs and the functional characterizations thereof
support the notion that complex behaviors are facilitated by coordinated interactions between
several large-scale sensory, attentional, and domain-specific networks, a position increasingly
endorsed in neuroimaging endeavors (Barrett & Satpute, 2013; Lindquist et al., 2012; Miši´c
& Sporns, 2016; Spreng et al., 2013). The characterization of identified MAGs from aspects
of the naturalistic paradigms that elicit them suggest an information processing model of co-
operating systems (Figure 5) for sensory input (MAGs 1 and 5), attentional control (MAG 6),
and domain-specific processing (MAGs 2, 3, and 4), into and from which information is segre-
gated and integrated to enable complex behaviors (e.g., language, emotion, spatial navigation).
Output relevant to the corresponding input would be relegated by motor planning and exe-
cution systems, which are notably absent from the characterization of MAGs presented here,
as experiments requiring a motor response were evenly distributed across MAGs, rather than
clustered together.
MAGs 1 and 5 primarily represent
the perceptual processing streams of
incoming
auditory and visual information, and likely cooperate to process audiovisual information.
Functional decoding suggests that MAG 1 is involved in viewing faces and anthropomor-
phic figures, which is consistent with previous research showing that posterior temporal and
temporo-occipital regions corresponding with area V5/MT are associated with the perception
of movement, specifically biological movement (Cohen Kadosh et al., 2010; Pelphrey et al.,
2005, 2004; Puce et al., 1998; Wheaton et al., 2004). Similarly, MAG 5 is associated with
listening to music and speech, as well as perceiving pitch and tone, stretching across primary
auditory cortex and into regions of higher auditory processing (Gray et al., 2009; Türe et al.,
1999). Per functional decoding of MAG 6 of both manual and automated annotations, MAG
6 is associated with visuospatial attention. This functional characterization is also supported
by corresponding fronto-parietal activations that are often associated with attending to visual
stimuli (Braga et al., 2016; Puschmann et al., 2016). MAGs 1 and 5 represent the perceptual
processing streams of audiovisual information.
Network Neuroscience
42
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
1
2
7
1
0
9
2
2
9
0
n
e
n
_
a
_
0
0
0
5
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Brain networks and naturalistic paradigms
Figure 5. Complex systems for dynamical information processing. The identified MAGs present
a framework of component systems that interact to enable complex information processing needed
for naturalistic behavior, including necessary input systems, as well as systems for modality-specific
(indicated by dashed line) visuospatial attentional gating of irrelevant information and domain-
specific processing for language-, emotion-, and navigation-related tasks.
Information processing depends on input from perceptual systems, filtered by attentional
gating, but proceeds in a functionally segregated manner, seen in domain-specific MAGs for
linguistic, emotional, and spatial processing. When considering language processing, there
is necessary input to primary auditory areas (MAG 5) that is further processed by higher level
language areas that facilitate speech perception and comprehension (MAG 2). More than a
third of contrasts from experiments that utilized speech-based paradigms contributed to the
convergent activation pattern of MAG 2, which was linked by both functional decoding tech-
niques to language-related processes. Furthermore, the regions of MAG 2 resemble a neural
“language network” (Friederici & Gierhan, 2013; Heim et al., 2003; Price, 2010; Saur et al.,
2010), including some regions associated with orofacial articulation (lip, tongue, and jaw
movements) and motor planning (SMA, pre-SMA) that allow the motor components of speech.
By presenting language in a context that is more representative of how we process language
in everyday life, such as through the use of spoken fictional narratives (AbdulSabur et al.,
2014; Wallentin et al., 2011; Xu et al., 2005) or scene descriptions (Summerfield et al., 2010),
naturalistic fMRI paradigms allow researchers to explore the multiple neural networks at work
in performing the cooperating processes that facilitate language processing. Similarly, emo-
tional processing (MAG 3) often necessitates audiovisual input (MAGs 1 and 5) and necessi-
tates attention (MAG 6). Emotional films recruited regions across these four MAGs, suggesting
a similarly diverse group of coordinated neural systems are engaged when observing affec-
tive displays. Additionally, navigation (Burgess et al., 2002; Kalpouzos et al., 2010; Wolbers
et al., 2004) depends on visual input (MAG 1), effective visuospatial attentional (MAG 6),
and spatial memory and processing (MAG 4). The functional characterization of MAG 4 from
manual and Neurosynth decoding highlights its involvement in navigation and spatial mem-
ory, supported by studies of rats and humans with brain lesions that indicate the importance of
Network Neuroscience
43
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
1
2
7
1
0
9
2
2
9
0
n
e
n
_
a
_
0
0
0
5
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Brain networks and naturalistic paradigms
medial temporal, hippocampal, and precuneus regions in processing visual scenes and spatial
information (Bird & Burgess, 2008; Epstein, 2008; Lee et al., 2005; Sailer et al., 2000; Squire
et al., 2004; Summerfield et al., 2010; Xu et al., 2005).
Finally, the characterization of MAG 6 indicates a domain-specific attentional system, as
both manual and automated Neurosynth decoding highlight its involvement in visual process-
ing in the absence of any association with other modalities. This is reflected by the distributions
of stimuli across MAGs (Figure 4), which show low numbers of auditory and pain-related stim-
uli represented in MAG 6, while rich visual stimuli that include spatial information (i.e., video
games, virtual reality, and pictures) are highly represented across the experiments in MAG 6.
Curiously, tactile object manipulation was highly represented in MAG 6, representing the
perception of spatial information in the absence of visual information (Figure 4, Table 3).
Together, these suggest that MAG 6 provides modality-specific attentional gating, depicted
by the dashed line in Figure 5.
Limitations
The present results may be limited by the k-means clustering method, which is limited by the
assumptions of the algorithm and underlying topology of the data, as it is sensitive to spherical
clusters and assumes the data are linearly separable. Furthermore, there is a potential for bias
with this method; certain parameters are specified by the researcher beforehand. To address
this potential for bias and the stability of our clustering solution, we performed duplicate
clustering analyses with both linear (hierarchical clustering using Ward’s method) and non-
linear (kernel k-means and density-based spatial clustering) methods. The results of these anal-
yses are provided in the Supporting information (Figures S1–S3, Bottenhorn et al., 2019) and
confirmed that our choice of the k-means clustering method provided optimal separation of the
data into six clusters. Experiments in our corpus were grouped using the kmeans++ algorithm
for each of K = 2 through K = 20 solutions, repeated 1,000 times to ensure that each solu-
tion minimized the point-to-centroid distance, indicative of optimal clustering (Kanungo et al.,
2004). Pearson’s correlation was selected as the distance metric, as recommended by Laird
et al. (2015). The K = 6 solution was designated as an optimal candidate solution before as-
sessing the convergent activation patterns of each MAG, based on the aforementioned metrics,
yielding a data-driven result. These results are, of course, influenced by the choice of cluster-
ing method, and should be considered accordingly. As this was a meta-analytic effort, it is
limited, too, by the initial modeling of the data. Despite this, coordinate-based meta-analyses
are considered a robust method for synthesis of previously published functional neuroimaging
literature (Eickhoff et al., 2012, 2009; Fox et al., 2005). Although the functional decoding-
based manual annotations relied on a subjective process, the results were largely confirmed
by comparison with the wider body of functional neuroimaging literature facilitated by Neuro-
synth’s automated functional decoding. It is worth noting that the naturalistic literature is some-
what limited, with an emphasis on navigation and affective processing, and continued research
and expansion of this corpus will facilitate development of a more comprehensive model of
the neural networks that support realistic behavior.
Summary and Future Work
In summary, this meta-analysis of naturalistic fMRI studies that apply dynamic, lifelike tasks
to explore the neural correlates of behavior has shown that these paradigms engage a set of
core neural networks, supporting both separate processing of different streams of information
and the integration of related information to enable flexible cognition and complex behavior.
Network Neuroscience
44
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
1
2
7
1
0
9
2
2
9
0
n
e
n
_
a
_
0
0
0
5
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Brain networks and naturalistic paradigms
We identified seven patterns of consistent activation that correspond with neural networks
that are involved in sensory input, top-down attentional control, domain-specific process-
ing, and motor planning, representing the set of behavioral processes elicited by naturalistic
paradigms in our corpus. Across the corpus, tasks provided mainly visual and auditory sensory
input that engaged regions across MAGs 1 and 5, while MAG 6 appeared to contribute to top-
down attentional control to filter out nonessential visual and/or spatial information. Salient
information can be processed by the relevant domain-specific networks, shown in MAGs 2
(language), 3 (emotion), and 4 (navigation and spatial memory), informing the appropriate re-
sponse. Most naturalistic tasks engaged multiple networks to process the relevant information
from a stimulus and generate an appropriate response. A shift in favor of utilizing naturalistic
paradigms, when possible, would greatly benefit the field, as naturalistic stimuli more closely
approximate the full complement of processing necessary for realistic behavior. Because of
the availability of naturalistic fMRI data from sources such as studyforrest.org, the Human
Connectome Project, and the Healthy Brain Network Serial Scanning Initiative (HBNSSI), an
intriguing next step in this line of work would include validating these MAGs in the primary
analysis of imaging data. Exploring how multifaceted processes interact and, ultimately, con-
tribute to behavior will allow us to better understand the brain and human behavior in the real
world. In the future, studies of this sort would greatly benefit from an automated annotation
process for an objective functional decoding of included papers, instead of subjective manual
annotation.
SUPPORTING INFORMATION
The authors have released all code and data associated with this manuscript. The code and
tabular data are available on GitHub (https://github.com/62442katieb/meta-analytic-kmeans),
and the unthresholded maps of each MAG are available on NeuroVault (https://neurovault.org/
collections/3179/ ). Supporting Information including tables and figures cited above is avail-
able at https://doi.org/10.1162/netn_a_00050.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
1
2
7
1
0
9
2
2
9
0
n
e
n
_
a
_
0
0
0
5
0
p
d
t
.
AUTHOR CONTRIBUTIONS
Katherine L. Bottenhorn: Formal analysis; Investigation; Methodology; Writing – original draft.
Jessica S. Flannery: Formal analysis; Writing – review & editing. Emily R. Boeving: Writing –
review & editing. Michael C. Riedel: Formal analysis; Methodology; Writing – review &
editing. Simon B. Eickhoff: Resources; Software; Writing – review & editing. Matthew T.
Sutherland: Conceptualization; Resources; Writing – review & editing. Angela R. Laird:
Conceptualization; Funding acquisition; Project administration; Resources; Software;
Writing – review & editing.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
FUNDING INFORMATION
Angela R Laird, National Institute on Drug Abuse (http://dx.doi.org/10.13039/100000026),
Award ID: U01-DA041156. Matthew T Sutherland, National Institute on Drug Abuse (http://
dx.doi.org/10.13039/100000026), Award ID: K01-DA037819. Not Applicable, National Insti-
tute on Drug Abuse (http://dx.doi.org/10.13039/100000026), Award ID: U24-DA039832. Not
Applicable, National Institute on Drug Abuse (http://dx.doi.org/10.13039/100000026), Award
ID: R01DA041353. Angela R Laird, National Institute of Mental Health (http://dx.doi.org/
10.13039/100000025), Award ID: R56-MH097870. Angela R Laird, National Science Foun-
dation (http://dx.doi.org/10.13039/100000001), Award ID: 1631325. Angela R Laird, National
Science Foundation (http://dx.doi.org/10.13039/100000001), Award ID: REAL DRL-1420627.
Network Neuroscience
45
Brain networks and naturalistic paradigms
REFERENCES
AbdulSabur, N. Y., Xu, Y., Liu, S., Chow, H. M., Baxter, M., Carson,
J., & Braun, A. R. (2014). Neural correlates and network con-
nectivity underlying narrative production and comprehension:
A combined fMRI and PET study. Cortex, 57, 107–127. https://
doi.org/10.1016/j.cortex.2014.01.017
Apps, M. A. J., Lockwood, P. L., & Balsters, J. H. (2013). The role
the midcingulate cortex in monitoring others’ decisions.
of
Frontiers in Neuroscience, 7, 251. https://doi.org/10.3389/fnins.
2013.00251
Barrett, L. F., & Satpute, A. B. (2013). Large-scale brain networks
in affective and social neuroscience: Towards an integrative
functional architecture of the brain. Current Opinion in Neuro-
biology,23,361–72. https://doi.org/10.1016/j.conb.2012.12.012
Beauregard, M., Lévesque, J., & Bourgouin, P. (2001). Neural cor-
relates of conscious self-regulation of emotion. Journal of Neuro-
science, 21, RC165.
Bird, C. M., & Burgess, N. (2008). The hippocampus and memory:
Insights from spatial processing. Nature Reviews Neuroscience,
9, 182–194. https://doi.org/10.1038/nrn2335
Bishop, C. W., & Miller, L. M. (2009). A multisensory cortical
network for understanding speech in noise. Journal of Cogni-
tive Neuroscience, 21, 1790–1805. Available at: http://www.
mitpressjournals.org/doi/10.1162/jocn.2009.21118,
[Accessed
October 26, 2017].
Bottenhorn, K. L., Flannery, J. S., Boeving, E. R., Riedel, M. C.,
Eickhoff, S. B., Sutherland, M. T., & Laird, A. R. (2019). Support-
ing Information for “Cooperating yet distinct brain networks
engaged during naturalistic paradigms: A meta-analysis of func-
tional MRI results.” Network Neuroscience, 3(1), 27–48. https://
doi.org/10.1162/netn_a_00050
Braga, R. M., Fu, R. Z., Seemungal, B. M., Wise, R. J. S., & Leech,
R. (2016). Eye movements during auditory attention predict indi-
vidual differences in dorsal attention network activity. Frontiers in
Human Neuroscience, 10, 164. https://doi.org/10.3389/fnhum.
2016.00164
Burgess, N., Maguire, E. A., & O’Keefe, J. (2002). The human hip-
pocampus and spatial and episodic memory. Neuron. https://
doi.org/10.1016/S0896-6273(02)00830-9
Burgess, N., Maguire, E. A., Spiers, H. J., & O’Keefe, J. (2001). A
temporoparietal and prefrontal network for retrieving the spatial
context of lifelike events. Neuroimage, 14, 439–453. https://doi.
org/10.1006/nimg.2001.0806
Burunat, I., Alluri, V., Toiviainen, P., Numminen, J., & Brattico, E.
(2014). Dynamics of brain activity underlying working memory
for music in a naturalistic condition. Cortex, 57, 254–269.
Bzdok, D., Heeger, A., Langner, R., Laird, A. R., Fox, P. T.,
Palomero-Gallagher, N., Vogt, B. A., Zilles, K., & Eickhoff, S. B.
(2015). Subspecialization in the human posterior medial cortex.
Neuroimage, 106, 55–71. https://doi.org/10.1016/j.neuroimage.
2014.11.009
Cohen Kadosh, K., Henson, R. N. A., Cohen Kadosh, R., Johnson,
M. H., & Dick, F.
(2010). Task-dependent activation of face-
sensitive cortex: An fMRI adaptation study. Journal of Cognitive
Neuroscience, 22, 903–917. https://doi.org/10.1162/jocn.2009.
21224
Collins, D. L., Neelin, P., Peters, T. M., & Evans, A. C. (1994).
Automatic 3D intersubject registration of MR volumetric data
Journal of Computer Assisted
in standardized Talairach space.
Tomography, 18, 192–205.
Cong, F., Puoliväli, T., Alluri, V., Sipola, T., Burunat, I., Toiviainen,
P., Nandi, A. K., Brattico, E., & Ristaniemi, T. (2014). Key issues
in decomposing fMRI during naturalistic and continuous mu-
sic experience with independent component analysis. Journal of
Neuroscience Methods, 223, 74–84.
Dick, A. S., Mok, E. H., Beharelle, A. R., Goldin-Meadow, S., &
Small, S. L. (2014). Frontal and temporal contributions to under-
standing the iconic co-speech gestures that accompany speech.
Human Brain Mapping, 35, 900–917.
Eickhoff, S. B., Bzdok, D., Laird, A. R., Kurth, F., & Fox, P. T. (2012).
Activation likelihood estimation meta-analysis revisited. Neuro-
image, 59, 2349–2361. https://doi.org/10.1016/j.neuroimage.
2011.09.017
Eickhoff, S. B., Laird, A. R., Fox, P. T., Bzdok, D., & Hensel,
L.
(2016a). Functional segregation of the human dorsomedial
prefrontal cortex. Cerebral Cortex, 26, 304–321. https://doi.org/
10.1093/cercor/bhu250
Eickhoff, S. B., Laird, A. R., Grefkes, C., Wang, L. E., Zilles, K., &
Fox, P. T.
(2009). Coordinate-based activation likelihood esti-
mation meta-analysis of neuroimaging data: A random-effects
approach based on empirical estimates of spatial uncertainty.
Human Brain Mapping, 30, 2907–2926. https://doi.org/10.1002/
hbm.20718
Eickhoff, S. B., Nichols, T. E., Laird, A. R., Hoffstaedter, F., Amunts,
K., Fox, P. T., Bzdok, D., & Eickhoff, C. R. (2016b). Behavior, sen-
sitivity, and power of activation likelihood estimation character-
ized by massive empirical simulation. Neuroimage, 137, 70–85.
https://doi.org/10.1016/j.neuroimage.2016.04.072
Epstein, R. A. (2008). Parahippocampal and retrosplenial contribu-
tions to human spatial navigation. Trends in Cognitive Sciences.
https://doi.org/10.1016/j.tics.2008.07.004
Evans, A. C., Collins, D. L., Mills, S. R., Brown, E. D., Kelly,
R. L., & Peters, T. M. (1993). 3D statistical neuroanatomical
models from 305 MRI volumes. In 1993 IEEE Conference Record
Nuclear Science Symposium and Medical Imaging Conference
IEEE, pp. 1813–1817. https://doi.org/10.1109/NSSMIC.1993.
373602
Fehr, T., Achtziger, A., Roth, G., & Struber, D. (2014). Neural corre-
lates of the empathic perceptual processing of realistic social in-
teraction scenarios displayed from a first-order perspective. Brain
Research, 1583, 141–158.
Fox, P. T., Laird, A. R., Fox, S. P., Fox, P. M., Uecker, A. M., Crank,
M., . . . Lancaster, J. L. (2005). Brainmap taxonomy of experimen-
tal design: Description and evaluation. Human Brain Mapping,
25, 185–198. https://doi.org/10.1002/hbm.20141
Friederici, A. D., & Gierhan, S. M. (2013). The language network.
Current Opinion in Neurobiology, 23, 250–254. https://doi.org/
10.1016/j.conb.2012.10.002
Friston, K. J., Price, C. J., Fletcher, P., Moore, C., Frackowiak, R. S.,
& Dolan, R. J. (1996). The trouble with cognitive subtraction.
Neuroimage,4,97–104.https://doi.org/10.1006/nimg.1996.0033
Network Neuroscience
46
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
1
2
7
1
0
9
2
2
9
0
n
e
n
_
a
_
0
0
0
5
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Brain networks and naturalistic paradigms
Gazzola, V., & Keysers, C. (2009). The observation and execution
of actions share motor and somatosensory voxels in all tested
subjects: Single-subject analyses of unsmoothed fMRI data.
Cerebral Cortex, 19, 1239–55. https://doi.org/10.1093/cercor/
bhn181
Giard, M. H., & Peronnet, F. (1999). Auditory-visual integration dur-
ing multimodal object recognition in humans: A behavioral and
Journal of Cognitive Neuroscience,
electrophysiological study.
11, 473–490. https://doi.org/10.1162/089892999563544
Gorgolewski, K. J., Varoquaux, G., Rivera, G., Schwarz, Y., Ghosh,
S. S., Maumet, . . . Margulies, D. S. (2015). NeuroVault.org:
A web-based repository for collecting and sharing unthresh-
olded statistical maps of the human brain. Frontiers in Neuro-
informatics, 9, 8. https://doi.org/10.3389/fninf.2015.00008
Gray, H., Carver, H. V., & Standring, S. (2009). Gray’s Anatomy
(40th ed.). Churchill Livingstone. https://doi.org/10.1308/
003588406X116873
Hanke, M., Adelhöfer, N., Kottke, D., Iacovella, V., Sengupta, A.,
Kaule, F. R., . . . Stadler, J. (2016). A studyforrest extension,
simultaneous fMRI and eye gaze recordings during prolonged
natural stimulation. Scientific Data, 3, 160092. https://doi.org/
10.1038/sdata.2016.92
Hanke, M., Baumgartner, F. J., Ibe, P., Kaule, F. R., Pollmann, S.,
Speck, O., Zinke, W., & Stadler, J.
(2014). A high-resolution
7-Tesla fMRI dataset from complex natural stimulation with an
audio movie. Scientific Data, 1. https://doi.org/10.1038/sdata.
2014.3
Hanke, M., Dinga, R., Häusler, C., Guntupalli, J. S., Casey, M.,
Kaule, F. R., & Stadler, J. (2015). High-resolution 7-Tesla fMRI
data on the perception of musical genres—An extension to the
studyforrest dataset. F1000Research. https://doi.org/10.12688/
f1000research.6679.1
Hasson, U., & Honey, C. J. (2012). Future trends in neuroimaging:
Neural processes as expressed within real-life contexts. Neuro-
image, 62, 1272–1278. https://doi.org/10.1016/j.neuroimage.
2012.02.004
Heim, S., Opitz, B., Müller, K., & Friederici, A. D. (2003). Phonolog-
ical processing during language production: fMRI evidence for
a shared production-comprehension network. Brain Research.
Cognitive Brain Research, 16, 285–96.
Kalpouzos, G., Eriksson, J., Sjölie, D., Molin, J., & Nyberg, L.
(2010). Neurocognitive systems related to real-world prospective
memory. PLoS One, 5, e13304. https://doi.org/10.1371/journal.
pone.0013304
Kanungo, T., Mount, D. M., Netanyahu, N. S., Piatko, C. D.,
Silverman, R., & Wu, A. Y. (2004). A local search approximation
algorithm for k-means clustering. Computational Geometry, 28,
89–112. https://doi.org/10.1016/j.comgeo.2004.03.003
Kauttonen, J., Hlushchuk, Y., & Tikka, P. (2015). Optimizing meth-
ods for linking cinematic features to fMRI data. Neuroimage,
110, 136–148.
Lahnakoski, J. M., Glerean, E., Salmi, J., Jääskeläinen, I. P., Sams,
M., Hari, R., & Nummenmaa, L. (2012). Naturalistic FMRI map-
ping reveals superior temporal sulcus as the hub for the dis-
tributed brain network for social perception. Frontiers in Human
Neuroscience, 6, 233. Available at: http://www.ncbi.nlm.nih.
gov/pubmed/22905026 [Accessed October 24, 2017].
Laird, A. R., Fox, P. M., Price, C. J., Glahn, D. C., Uecker, A. M.,
Lancaster, . . . Fox, P. T. (2005). ALE meta-analysis: Control-
ling the false discovery rate and performing statistical contrasts.
Human Brain Mapping, 25, 155–164. https://doi.org/10.1002/
hbm.20136
Laird, A. R., Riedel, M. C., Sutherland, M. T., Eickhoff, S. B., Ray,
K. L., Uecker, . . . Fox, P. T. (2015). Neural architecture under-
lying classification of face perception paradigms. Neuroimage,
119, 70–80. https://doi.org/10.1016/j.neuroimage.2015.06.044
Laird, A. R., Robinson, J. L., McMillan, K. M., Tordesillas-Gutiérrez,
D., Moran, S. T., Gonzales, S. M., . . . Lancaster, J. L.
(2010).
Comparison of the disparity between Talairach and MNI co-
ordinates in functional neuroimaging data: Validation of the
Lancaster transform. Neuroimage, 51, 677–83. https://doi.org/
10.1016/j.neuroimage.2010.02.048
Lancaster, J. L., Tordesillas-Gutiérrez, D., Martinez, M., Salinas,
F., Evans, A., Zilles, . . . Fox, P. T.
(2007). Bias between MNI
and Talairach coordinates analyzed using the ICBM-152 brain
template. Human Brain Mapping, 28, 1194–205. https://doi.org/
10.1002/hbm.20345
Lee, A. C. H., Buckley, M. J., Pegman, S. J., Spiers, H., Scahill, V. L.,
Gaffan, D., . . . Graham, K. S. (2005). Specialization in the me-
dial temporal lobe for processing of objects and scenes. Hippo-
campus, 15, 782–797. https://doi.org/10.1002/hipo.20101
Li, Y., Long, J., Huang, B., Yu, T., Wu, W., Liu, C., . . . Sun, P. (2015).
Crossmodal integration enhances neural representation of task-
relevant features in audiovisual face perception. Cerebral Cortex,
25, 384–395. https://doi.org/10.1093/cercor/bht228
Lindquist, K. A., Wager, T. D., Kober, H., Bliss-Moreau, E., & Barrett,
L. F. (2012). The brain basis of emotion: A meta-analytic review.
Behavioral and Brain Sciences,
( 35) 121–143. https://doi.org/
10.1017/S0140525X11000446
Maguire, E. A. (2012). Studying the freely-behaving brain with
fMRI. Neuroimage, 62, 1170–1176. https://doi.org/10.1016/
j.neuroimage.2012.01.009
Marsh, R., Hao, X., Xu, D., Wang, Z., Duan, Y., Liu, . . . Peterson,
B. S. (2010). A virtual reality-based FMRI study of reward-based
spatial learning. Neuropsychologia, 48, 2912–2921. https://doi.
org/10.1016/j.neuropsychologia.2010.05.033
McGurk, H., & MacDonald,
(1976). Hearing lips and see-
ing voices. Nature, 264, 691–811. https://doi.org/10.1038/
264746a0
J.
Miši´c, B., & Sporns, O.
(2016). From regions to connections and
networks: New bridges between brain and behavior. Current
Opinion in Neurobiology, 40, 1–7. https://doi.org/10.1016/
j.conb.2016.05.003
Moher, D., Liberati, A., Tetzlaff, J., Altman, D. G., & Altman,
D.
(2009). Preferred reporting items for systematic reviews
and meta-analyses: The PRISMA statement. PLoS Medicine, 6,
e1000097. https://doi.org/10.1371/journal.pmed.1000097
Molenberghs, P., Cunnington, R., & Mattingley, J. B. (2009). Is
the mirror neuron system involved in imitation? A short review
and meta-analysis. Neuroscience & Biobehavioral Reviews, 33,
975–980. https://doi.org/10.1016/j.neubiorev.2009.03.010
Nardo, D., Santangelo, V., & Macaluso, E. (2014). Spatial orienting
in complex audiovisual environments. Human Brain Mapping,
35, 1597–1614.
Network Neuroscience
47
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
1
2
7
1
0
9
2
2
9
0
n
e
n
_
a
_
0
0
0
5
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Brain networks and naturalistic paradigms
Pelphrey, K. A., Morris, J. P., & McCarthy, G. (2004). Grasping the
intentions of others: The perceived intentionality of an action
influences activity in the superior temporal sulcus during social
perception. Journal of Cognitive Neuroscience, 16, 1706–1716.
https://doi.org/10.1162/0898929042947900
Pelphrey, K. A., Morris,
J. P., Michelich, C. R., Allison, T., &
McCarthy, G. (2005). Functional anatomy of biological motion
perception in posterior temporal cortex: An fMRI study of eye,
mouth and hand movements. Cerebral Cortex, 15, 1866–1876.
https://doi.org/10.1093/cercor/bhi064
Price, C. J. (2010). The anatomy of language: A review of 100 fMRI
studies published in 2009. Annals of the New York Academy
of Sciences, 1191, 62–88. https://doi.org/10.1111/j.1749-6632.
2010.05444.x
Puce, A., Allison, T., Bentin, S., Gore, J. C., & McCarthy, G. (1998).
Temporal cortex activation in humans viewing eye and mouth
movements. Journal of Neuroscience, 18, 2188–2199.
Puschmann, S., Huster, R. J., & Thiel, C. M. (2016). Mapping the
spatiotemporal dynamics of processing task-relevant and task-
irrelevant sound feature changes using concurrent EEG-fMRI.
Human Brain Mapping. https://doi.org/10.1002/hbm.23248
Reed, C. L., Shoham, S., & Halgren, E. (2004). Neural Substrates of
Tactile Object Recognition: An fMRI Study. Human Brain Map-
ping, 21, 236–246.
Sailer, U., Eggert, T., Ditterich, J., & Straube, A. (2000). Spatial and
temporal aspects of eye-hand coordination across different tasks.
Experimental Brain Research, 134, 163–173. https://doi.org/10.
1007/s002210000457
Saur, D., Schelter, B., Schnell, S., Kratochvil, D., Küpper, H.,
Kellmeyer, . . . Weiller, C. (2010). Combining functional and
anatomical connectivity reveals brain networks for auditory
language comprehension. Neuroimage, 49, 3187–3197. https://
doi.org/10.1016/j.neuroimage.2009.11.009
Spence, C.
(2010). Crossmodal spatial attention. Annals of the
New York Academy of Sciences. https://doi.org/10.1111/j.1749-
6632.2010.05440.x
Spiers, H. J., & Maguire, E. A.
Spontaneous mental-
izing during an interactive real world task: An fMRI study.
Neuropsychologia, 44, 1674–1682. https://doi.org/10.1016/
j.neuropsychologia.2006.03.028
(2006).
(2013).
Spreng, R. N., Sepulcre, J., Turner, G. R., Stevens, W. D., & Schacter,
D. L.
Intrinsic architecture underlying the relations
among the default, dorsal attention, and frontoparietal control
networks of the human brain. Journal of Cognitive Neuroscience,
25, 74–86. https://doi.org/10.1162/jocn_a_00281
Spunt, R. P., & Lieberman, M. D. (2012). An integrative model of
the neural systems supporting the comprehension of observed
emotional behavior. Neuroimage, 59, 3050–3059. https://doi.
org/10.1016/j.neuroimage.2011.10.005
Squire, L. R., Stark, C. E. L., & Clark, R. E. (2004). The medial tempo-
ral lobe. Annual Review of Neuroscience, 27, 279–306. https://
doi.org/10.1146/annurev.neuro.27.070203.144130
Summerfield, J. J., Hassabis, D., & Maguire, E. A. (2010). Differen-
tial engagement of brain regions within a “core” network during
scene construction. Neuropsychologia, 48, 1501–1509. https://
doi.org/10.1016/j.neuropsychologia.2010.01.022
Talairach, J., & Tournoux, P. (1988). Co-Planar Stereotaxic Atlas of
the Human Brain: 3-D Proportional System: An Approach to
Cerebral Imaging (1st ed.). New York: Thieme.
Türe, U., Ya¸sargil, D. C. H., Al-Mefty, O., & Ya¸sargil, M. G. (1999).
Journal of Neuro-
Topographic anatomy of the insular region.
surgery, 90, 720–733. https://doi.org/10.3171/jns.1999.90.4.0720
Turkeltaub, P. E., Eden, G. F., Jones, K. M., & Zeffiro, T. A. (2002).
Meta-analysis of the functional neuroanatomy of single-word
reading: Method and validation. Neuroimage, 16, 765–780.
https://doi.org/10.1006/nimg.2002.1131
Turkeltaub, P. E., Eickhoff, S. B., Laird, A. R., Fox, M., Wiener, M.,
& Fox, P. (2012). Minimizing within-experiment and within-
group effects in activation likelihood estimation meta-analyses.
Human Brain Mapping, 33, 1–13. https://doi.org/10.1002/hbm.
21186
Turner, J. A., & Laird, A. R. (2012). The cognitive paradigm ontology:
design and application. Neuroinformatics, 10, 57–66. https://doi.
org/10.1007/s12021-011-9126-x
Wallentin, M., Nielsen, A. H., Vuust, P., Dohn, A., Roepstorff, A., &
Lund, T. E. (2011). Amygdala and heart rate variability responses
from listening to emotionally intense parts of a story. Neuro-
image, 58, 963–973. https://doi.org/10.1016/j.neuroimage.2011.
06.077
Wang, J., Ren, Y., Hu, X., Nguyen, V. T., Guo, L., Han, J., &
Guo, C. C. (2016). Test-retest reliability of functional connectivity
networks during naturalistic fMRI paradigms. bioRxiv.
Wheaton, K. J., Thompson, J. C., Syngeniotis, A., Abbott, D. F., &
Puce, A. (2004). Viewing the motion of human body parts acti-
vates different regions of premotor, temporal, and parietal cortex.
Neuroimage, 22, 277–288. https://doi.org/10.1016/j.neuroimage.
2003.12.043
Wolbers, T., Weiller, C., & Büchel, C. (2004). Neural foundations
of emerging route knowledge in complex spatial environments.
Cognitive Brain Research, 21, 401–411. https://doi.org/10.1016/
j.cogbrainres.2004.06.013
Woo, C.-W., Krishnan, A., & Wager, T. D. (2014). Cluster-extent
based thresholding in fMRI analyses: Pitfalls and recommen-
dations. Neuroimage, 91, 412–419. https://doi.org/10.1016/j.
neuroimage.2013.12.058
Xu,
J., Kemeny, S., Park, G., Frattali, C., & Braun, A. (2005).
Language in context: Emergent features of word, sentence, and
narrative comprehension. Neuroimage, 25, 1002–1015. https://
doi.org/10.1016/j.neuroimage.2004.12.013
Yarkoni, T., Poldrack, R. A., Nichols, T. E., Van Essen, D. C., &
Wager, T. D. (2011). Large-scale automated synthesis of hu-
man functional neuroimaging data. Nature Methods, 8, 665–70.
https://doi.org/10.1038/nmeth.1635
Network Neuroscience
48
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
1
2
7
1
0
9
2
2
9
0
n
e
n
_
a
_
0
0
0
5
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3