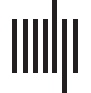RESEARCH
Dynamic large-scale network synchronization
from perception to action
Jonni Hirvonen
1,2
, Simo Monto
1,†
1
, Sheng H. Wang
1
, J. Matias Palva
1
, and Satu Palva
1Helsinki Institute for Life Sciences, Neuroscience Center, University of Helsinki, Finland
2BioMag Laboratory, HUS Medical Imaging Center, Helsinki University Central Hospital, Finland
†Present affiliation: Centre for Interdisciplinary Brain Research, Department of Psychology, University of Jyväskylä, Finland
Keywords: MEG, Synchronization, Somatosensory, Communication, Perception, Action
a n o p e n a c c e s s
j o u r n a l
ABSTRACT
Sensory-guided actions entail the processing of sensory information, generation of perceptual
decisions, and the generation of appropriate actions. Neuronal activity underlying these
processes is distributed into sensory, fronto-parietal, and motor brain areas, respectively.
How the neuronal processing is coordinated across these brain areas to support functions
from perception to action remains unknown. We investigated whether phase synchronization
in large-scale networks coordinate these processes. We recorded human cortical activity
with magnetoencephalography (MEG) during a task in which weak somatosensory stimuli
remained unperceived or were perceived. We then assessed dynamic evolution of phase
synchronization in large-scale networks from source-reconstructed MEG data by using
advanced analysis approaches combined with graph theory. Here we show that perceiving
and reporting of weak somatosensory stimuli is correlated with sustained strengthening of
large-scale synchrony concurrently in delta/theta (3–7 Hz) and gamma (40–60 Hz) frequency
bands. In a data-driven network localization, we found this synchronization to dynamically
connect the task-relevant, that is, the fronto-parietal, sensory, and motor systems. The
strength and temporal pattern of interareal synchronization were also correlated with the
response times. These data thus show that key brain areas underlying perception,
decision-making, and actions are transiently connected by large-scale dynamic phase
synchronization in the delta/theta and gamma bands.
AUTHOR SUMMARY
We studied the functional role of interareal phase synchronization in the
perception-to-action cascade. To this end we recorded human cortical activity with
magnetoencephalography (MEG) during a task where weak somatosensory stimuli were
perceived or remained unperceived. We identified time-resolved large-scale networks of
synchronization by using source-reconstructed MEG data with advanced analysis tools and
graph theory. We found that perceiving and reporting of weak somatosensory stimuli was
correlated with sustained strengthening of large-scale phase synchronization concurrently in
delta/theta (3–7 Hz) and gamma (40–60 Hz) frequency bands. Synchronization
characterizing the perceived stimuli connected the sensorimotor system with frontoparietal
attentional systems. These data suggest that large-scale network synchronization may
coordinate neuronal processing across brain regions during perception, decision-making,
and when responding to weak somatosensory stimuli.
Citation: Hirvonen, J., Monto, S., Wang,
S. H., Palva, J. M., & Palva, S. (2018).
Dynamic large-scale network
synchronization from perception to
action. Network Neuroscience, 2(4),
442–463 https://doi.org/10.1162/
netn_a_00039
DOI:
https://doi.org/10.1162/netn_a_00039
Supporting Information:
https://doi.org/10.1162/netn_a_00039
Received: 11 August 2017
Accepted: 19 December 2017
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Author:
Satu Palva
satu.palva@helsinki.fi
Handling Editor:
Olaf Sporns
Copyright: © 2018
Massachusetts Institute of Technology
Published under a Creative Commons
Attribution 4.0 International
(CC BY 4.0) license
The MIT Press
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
2
4
4
4
2
1
0
9
2
2
4
9
n
e
n
_
a
_
0
0
0
3
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Large-scale synchronization from perception to action
Magnetoencephalography (MEG):
Measures magnetic fields produced
by coherent current flows in
neuronal populations.
Source reconstruction:
Inverse modeling of MEG sensor data
to estimate the underlying cortical
source current distribution and time
series.
Phase synchronization:
Indicates a consistent phase
relationship between oscillations,
here between different brain areas.
INTRODUCTION
Understanding the neuronal mechanisms that govern the translation of conscious perception
of sensory information into perceptual decisions and generation of appropriate actions is an
important challenge in neuroscience. Several studies using functional magnetic resonance
imaging (fMRI; Blankenburg et al., 2003; Dehaene and Changeux, 2011; Hegner et al., 2016;
Li Hegner et al., 2015), human intracranial electroencephalography (iEEG; Fisch et al., 2009;
Gaillard et al., 2009), and noninvasive source-reconstructed electro- and magnetoencephalog-
raphy (EEG and MEG; Hirvonen & Palva, 2016; King et al., 2016; Salti et al., 2015) show that
neuronal activity in prefrontal (PFC) and posterior parietal cortex (PPC) together with that in
sensory areas is correlated positively with conscious perception. King et al. (2016) recently
used a decoding approach of MEG data to show that brain regions as well as temporal evolu-
tion of neuronal processing were distinct for perception, maintenance, and visibility of seen
and unseen visual stimuli.
Also, perceptual decision-making, for deciding to report the presence or absence of a sen-
sory stimuli or a target, involves activity in PPC and PFC in human (Donner et al., 2009b;
Kaplan et al., 2017; Tosoni et al., 2008), monkey (Siegel et al., 2015), and mouse (Goard
et al., 2016) local field potential data. More specifically, it has been suggested that the sen-
sorimotor decision process engages neuronal structures that are involved in preparing the as-
sociated actions, such as the oculomotor regions of parietal and prefrontal cortex in monkeys
(Schall, 2001; Yang & Shadlen, 2007) and the response-related PPC regions and motor cortex
in humans (Donner et al., 2009b; Gould et al., 2012; Kaplan et al., 2017; Tosoni et al., 2008)
(Medendorp et al., 2011). Motor actions, conversely, dynamically modulate the processing of
sensory information (Gutteling et al., 2011; Wohlschlager, 2000), and at the behavioral level,
rhythmicity of visual perception is coupled with that of action (Tomassini et al., 2015).
These lines of evidence thus indicate multiple levels of coupling between perception and
action, but how this cooperation is achieved at the neuronal level has remained incompletely
understood. One candidate mechanism to achieve such coordination is neuronal synchroni-
zation, which has been shown to regulate neuronal communication (Fries, 2015; Gregoriou
et al., 2009; Maris et al., 2016; Siegel et al., 2012; Singer, 2009) and provide a temporal
correlation–based framework for integrating information processing in neuronal circuits at
multiple scales (Fries, 2015; Schroeder & Lakatos, 2009; Siegel et al., 2012). In humans, neu-
ronal activity in a large fraction of the cerebral cortex can be noninvasively recorded with EEG
and MEG, but using these methods for estimating long-range synchronization demands both
a source reconstruction–based approach and methods to address the challenges of residual
signal leakage (Brookes et al., 2012; Palva & Palva, 2012; Schoffelen & Gross, 2009). Prelim-
inary evidence from the visual modality suggests that long-range synchronization among the
visual and frontal brain areas is indeed correlated with visual perception (Hipp et al., 2011)
and that between motor cortical areas and peripheral nervous system with the execution of
motor actions (Schoffelen & Gross, 2009). Although these studies provide partial support for
the idea that perception-action cycle could be implemented via phase synchronization, the
complete network architectures bridging perception and action have remained unaddressed.
We hypothesized that dynamic large-scale network phase synchronization would underlie
the coordination of neuronal processing from perception to perceptual decisions and actions
by regulating the neuronal processing between PFC, PPC, and sensory and motor systems.
To test this hypothesis, we recorded MEG during a somatosensory stimulus detection task
in which the subjects were presented constant-current somatosensory stimuli at the thresh-
old of perception that were perceived or remained unperceived (Hirvonen & Palva, 2016).
Network Neuroscience
443
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
2
4
4
4
2
1
0
9
2
2
4
9
n
e
n
_
a
_
0
0
0
3
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Large-scale synchronization from perception to action
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
2
4
4
4
2
1
0
9
2
2
4
9
n
e
n
_
a
_
0
0
0
3
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 1. A schematic overview of the analysis pipeline. This figure shows the different analysis
steps and outputs of the results (for a–m, see Methods).
We then used MEG source modeling–based all-to-all phase coupling analysis (Figure 1) to
identify the cortical networks characterizing perception-action sequence–associated neuronal
processing. We found concurrent delta/theta- and gamma-band synchronization to connect
the somatosensory, fronto-parietal, and motor cortical regions for stimuli that were perceived
and an absence of this coupling for stimuli that remained undetected. These findings pro-
vide evidence for neuronal phase synchronization to play a functionally significant role in the
cooperation of sensory, decision, and motor stages from perception to action (Figure 1).
RESULTS
Behavioral Performance
The subjects’ task during the MEG recordings was to detect weak, constant-intensity electrical
stimuli given to the tip of the right index finger during two 30-min sessions at random intervals
between 1.5 and 4.5 s. The strength of the stimuli was calibrated to the threshold of detection
prior to recordings and maintained constant during the MEG recordings. Thus, during MEG
recordings, these stimuli were variably either detected (Hits) or remained undetected (Misses)
(Figure 2A). The hit rate (HR) was 38.32 ± 15.26% [mean ± standard deviation (SD)], and
Network Neuroscience
444
Large-scale synchronization from perception to action
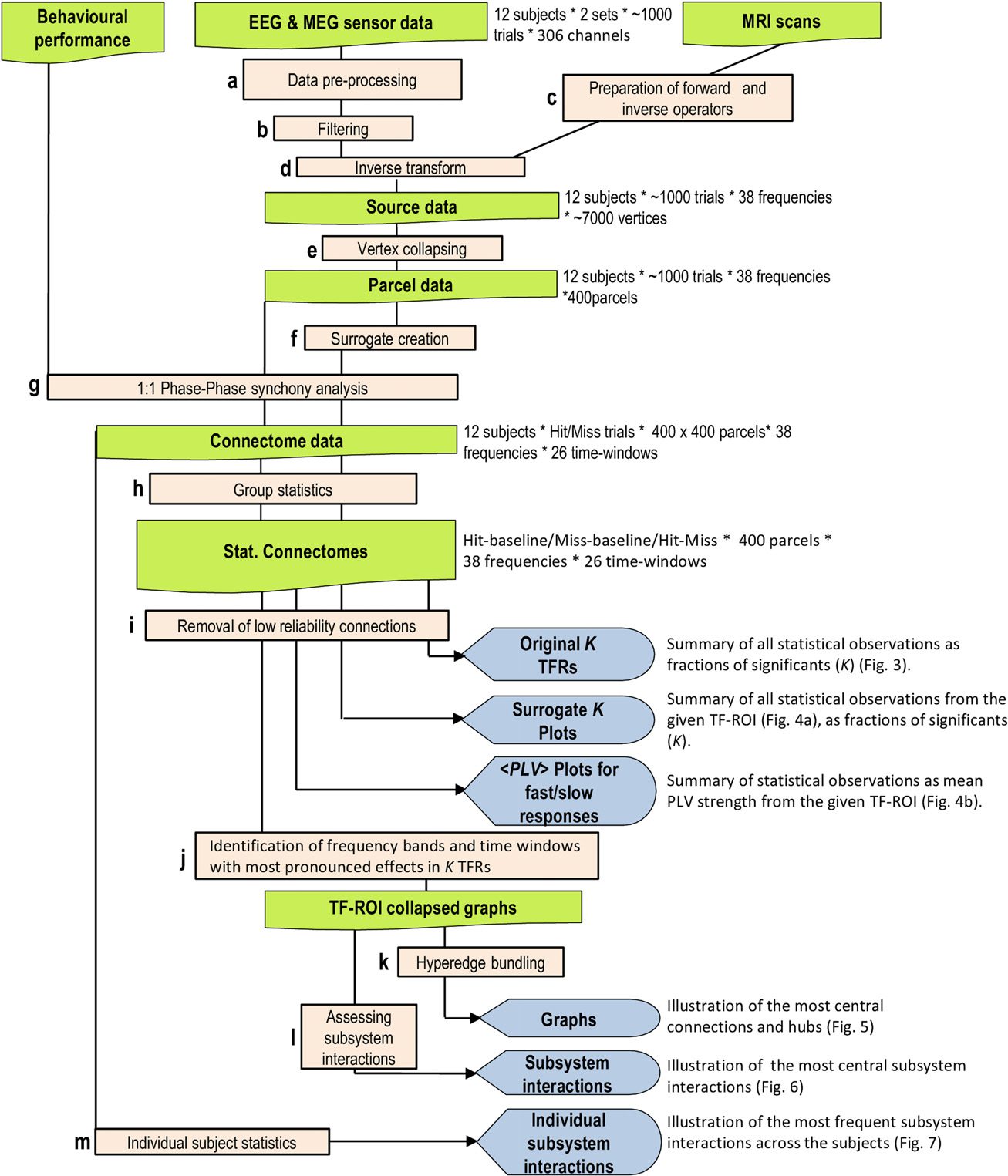
Figure 2. Task and behavioral performance. (A) Schematic illustration of the experimental para-
digm with a stream of constant-intensity somatosensory stimuli presented at 1.5- to 4.5-s intervals
and the corresponding behavioral Hit-Miss time series for a representative subject. Blue bars denote
consciously perceived stimuli, and the red bars denote unperceived stimuli. (B) Individual (black
lines) and group (red line) response time (RT) distributions.
the reaction time (RT) was 399 ± 135 ms (mean ± SD). We estimated the fraction of false
alarms (FA), that is, responses when no stimulus was presented, from the time window preced-
ing electrical stimuli (Figure 2B, see Methods section). On the basis of the finite-state model
approach, we estimated FA rate to be 0.2 ± 0.3% (mean ± SD) of all trials, which gives a true
stimulus detection rate of 38.12%.
Sustained Large-Scale Delta and Gamma Synchrony Characterizes Perception-Action Cycle
To investigate whether large-scale synchronization would characterize the neuronal process-
ing from perception to action and to map the anatomy and frequency patterns of such putative
synchronization, we tested whether synchronization would characterize the processing of
those stimuli that were reported perceived. We thus characterized phase synchrony among
all cortical areas for each subject, time window, and frequency band separately for trials in
which near-sensory stimulation was considered as perceived (Hits) or as unperceived
(Misses). We represented statistically significant observations of interareal phase synchrony as
graphs where cortical areas were the vertices and significant connections the edges (Bullmore
& Sporns, 2009; Palva et al., 2010; Rubinov & Sporns, 2010). The extent of synchrony was
estimated with connection density (K), which is the proportion of significant interareal in-
teractions from all possible pairwise interactions among the 400 brain areas in our cortical
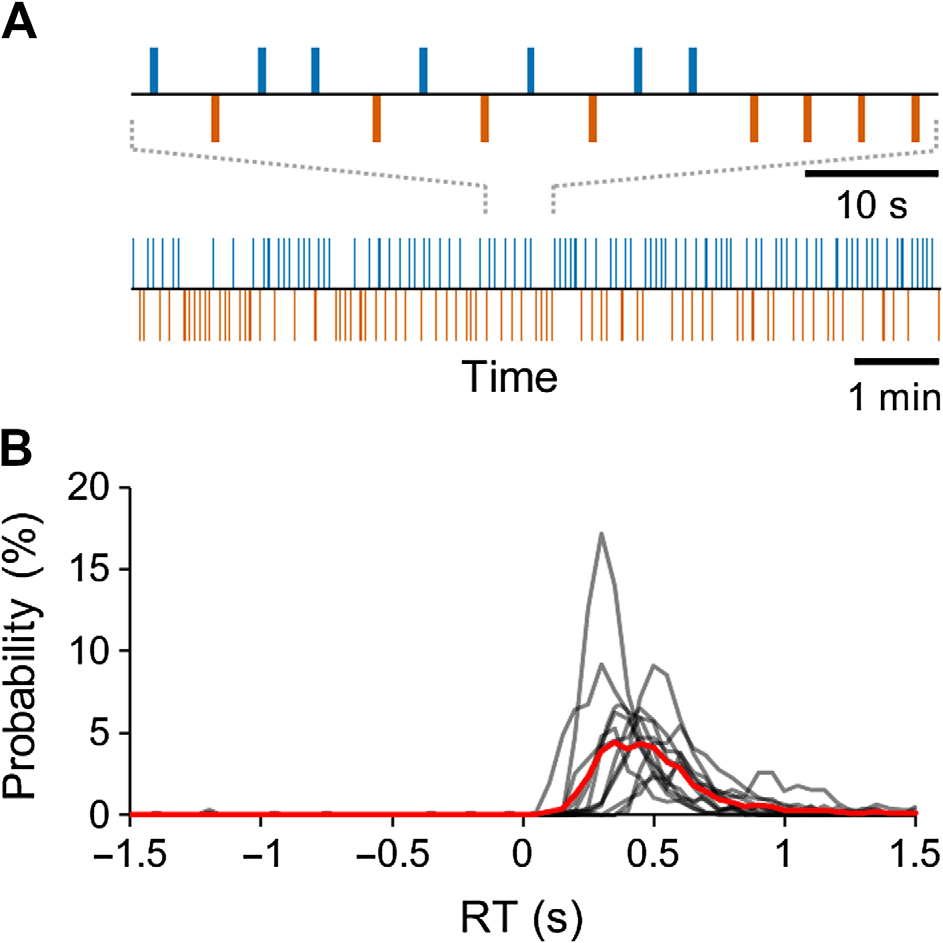
parcellation. The frequency spectrum of K showed that synchronization in the mid-gamma
frequency band (mid-γ, 40–60 Hz) at 150–500 ms and in the delta/theta (δ–θ, 3–7 Hz) band
from 0 to 500 ms after the stimulus onset was strengthened above baseline levels for Hits
(Wilcoxon signed-rank test, p < 0.05, corrected for multiple comparisons) when interareal
synchronization was measured with the phase-locking value (PLV; Figure 3A). An essentially
identical result was obtained when the imaginary part of PLV (iPLV) that is insensitive to source
mixing was used to measure synchronization (Figure 3B), which showed that delta and gamma-
band synchronization were not attributable to signal mixing and artificial synchronization.
445
Connection density:
A proportion of connections in a
graph out of all possible connections;
here connections were defined by
their statistical significance.
Parcellation:
Division of the cortical surface into
parcels that here correspond to
neuroanatomically named brain
areas.
Phase-locking value (PLV):
The absolute value of complex
PLV, quantifies the strength
of phase synchronization.
Imaginary part of complex
PLV (iPLV):
A phase synchronization measure
that is sensitive only to nonzero
and non-pi phase lag couplings
and thereby excluded the direct
effects of linear mixing.
Network Neuroscience
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
2
4
4
4
2
1
0
9
2
2
4
9
n
e
n
_
a
_
0
0
0
3
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Large-scale synchronization from perception to action
In contrast, interareal synchronization was largely insignificant (Wilcoxon signed-rank test,
p < 0.05, corrected for multiple comparisons) for Misses and in the gamma band even briefly
suppressed below the baseline level both when estimated with PLV (Figure 3A) and iPLV
(Figure 3B). Importantly, this suppression shows that the lack of synchronization for Misses was
not due to poor signal-to-noise ratio for missed stimuli, but that Hits and Misses were associ-
ated with genuinely distinct synchronization patterns. A comparison between Hits and Misses
confirmed that both large-scale delta- and gamma-band synchrony were indeed stronger for
Hits than Misses (Wilcoxon signed-rank test, p < 0.05, corrected) estimated with both PLV
(Figure 3A) and iPLV (Figure 3B).
Observations of large-scale synchronization can also arise artificially from evoked responses
or phase locking of ongoing activity to the stimulus onsets (Palva & Palva, 2012). In the present
data, early transient phase locking of ongoing oscillations to stimulus onset as well as transient
amplitude increase was observed for both Hits and Misses and was stronger for Hits than
Misses between 3 and 30 Hz (Hirvonen & Palva, 2016) (Supporting Information Figure 2,
Hirvonen, Monto, Wang, Palva, & Palva, 2018). Sustained phase locking of oscillations to
stimulus onset and sustained oscillation amplitude increases during the time windows of large-
scale delta/theta- and gamma-band phase synchronization were observed only in the low-
alpha and delta/theta frequencies. In the delta/theta band both phase locking and oscillation
amplitudes were increased, whereas in the alpha band, phase locking to stimulus onset was
increased but the oscillations amplitudes were more suppressed for the Hits than for Misses.
Evoked/stimulus-locked activity:
A cortical response with a
consistent latency after stimuli
and thus observable in
trial-averaged response.
These findings hence indicate qualitatively that large-scale gamma-band synchronization
cannot be caused by evoked or stimulus phase-locked activity. To confirm this as well as to
assess the contribution of evoked activity in the observed delta/theta band large-scale syn-
chronization, we compared synchronization data against surrogate data where non-stimulus-
locked phase relationships were eliminated but evoked/stimulus-locked components were
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
2
4
4
4
2
1
0
9
2
2
4
9
n
e
n
_
a
_
0
0
0
3
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. Large-scale delta- and gamma-band synchronization characterizes neuronal process-
ing of perceived but not unperceived stimuli. (A) Time-frequency representations of the extent of
significant interareal synchronization as estimated with PLV for perceived (Hit) stimuli, unperceived
(Miss), as well as for their difference (Hit-Miss) compared with the prestimulus baseline (N = 12)
(Wilcoxon signed-ranked test, p < 0.05, corrected for multiple comparisons). Sustained gamma-
band (40–60 Hz) synchronization and delta/theta- (δ/θ, 3–7 Hz) band synchronization were stronger
for Hits than Misses. The color indicates the connection densities of positive (K+) and negative (K−)
observations, that is, the fractions of connections with a statistically significant positive or negative
difference from the baseline level, respectively. (B) Time-frequency representations of the extent of
significant interareal synchronization as estimated with iPLV.
Network Neuroscience
446
Large-scale synchronization from perception to action
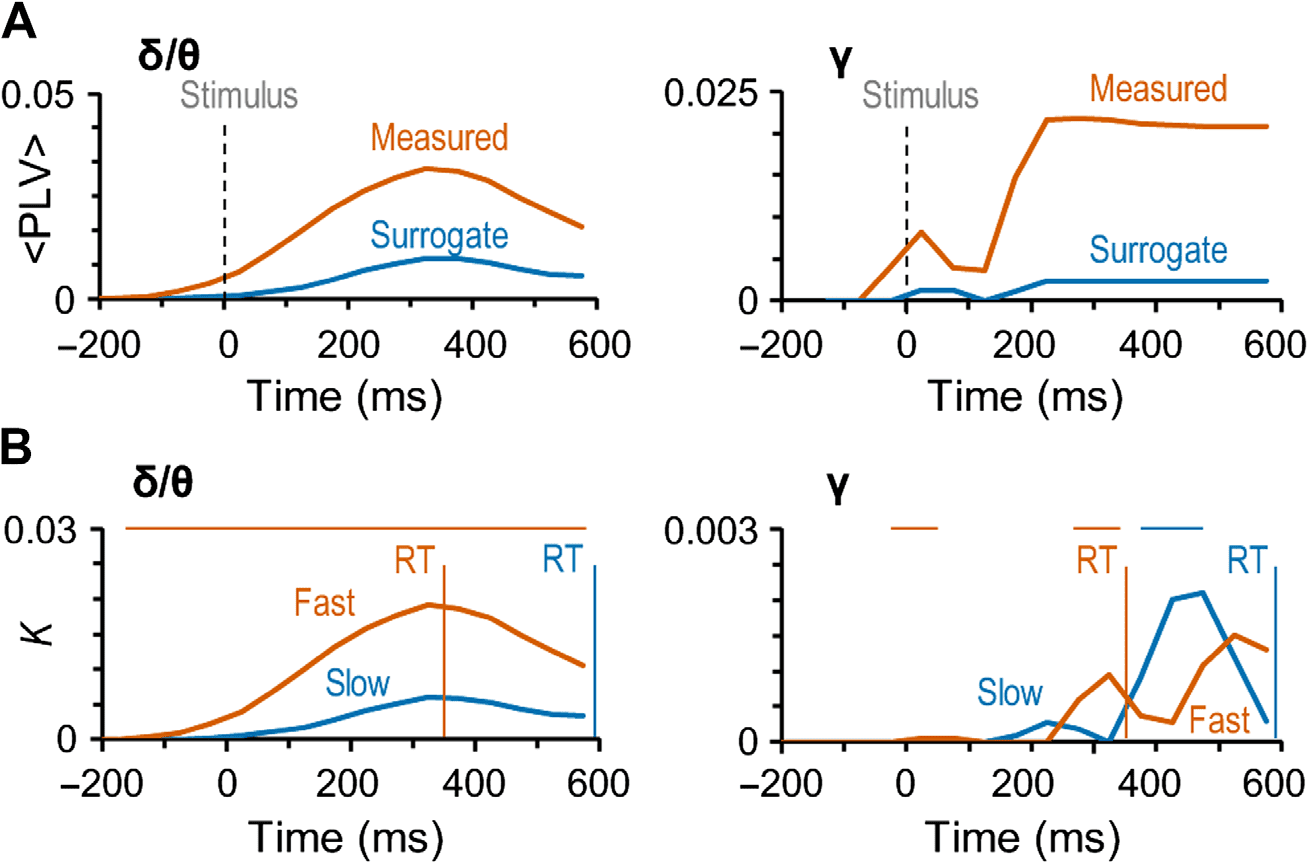
Figure 4. Gamma- and delta/theta-band synchronization is not artificial and predicts the speed
of sensorimotor decisions. (A) The mean coupling strength (
Hits in the original (red line) and surrogate data (blue line) in delta/theta- (δ/θ) and gamma- (γ)
frequency bands. (B) Connection density (K) as a function of time for the difference between Hits
and Misses separately for trials with fast (red) and slow (blue) RTs. Vertical lines show the mean
RT in these two categories: 356 ± 122 ms (mean ± SD across subjects) for the trials with fast RTs
and 594 ± 229 ms for the trials with slow RTs. The horizontal bars above the plots show the time
window in significant difference between trials with fast and slow trial RTs (Wilcoxon signed-ranked
test, p < 0.05, corrected for multiple comparisons). The trials were split at the median RT of each
subject and the mean of these median RTs was 464 ± 142 ms (mean ± SD across subjects).
preserved (see Methods). We compared the strengths of phase synchrony in original data
against the corresponding mean values of surrogate data. Importantly,
in the delta/theta
(Figure 4A, left) and specifically in the gamma-band (Figure 4A, right) synchronization was
significantly stronger in the original than surrogate data for Hits. These analyses thus unequiv-
ocally indicate that neither the large-scale delta/theta- or gamma-band synchronization ob-
served for detected stimuli can be explained by artificial synchronization caused either by
phase locking of ongoing oscillations to stimulus onset or additive evoked responses, or by
confounders such as stimulus-altered local source topographies or autocorrelations.
Correlation of Large-Scale Synchronization with the Motor Responses
Our main goal was to reveal the networks underlying the coordination of processing from per-
ception to action. To thus identify whether and when synchronization is locked to actions, that
is, the motor responses rather than related to perceptual processing, we investigated whether
the strength of the observed large-scale delta/theta and gamma-band synchronization would be
correlated with RTs. We divided the responses within each subject into “fast” (356 ± 122 ms)
and “slow” (594 ± 229 ms) response categories by the individual median RT (464 ± 142 ms,
mean ± SD across subjects). We then estimated the strength of synchronization separately for
these two categories of responses. This analysis indicated that for fast and slow responses both
delta/theta band and gamma bands had distinct synchronization patterns (Figure 4B). For in
delta/theta, synchronization was stronger for the fast than for the slow responses during the
whole response time (Wilcoxon signed-ranked test, p < 0.05, corrected for multiple compar-
isons). However, gamma-band synchronization differed for the fast and slow responses only
Network Neuroscience
447
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
2
4
4
4
2
1
0
9
2
2
4
9
n
e
n
_
a
_
0
0
0
3
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Large-scale synchronization from perception to action
Subsystem:
A set of brain areas defined by
intrinsic connectivity and typically
corresponding to areas with
associated functional roles.
during the different peak latencies, which showed that for the fast responses, gamma-band
synchronization began around 200 ms from stimulus onset, whereas for the slow responses
it began 350 ms from stimulus onset (Wilcoxon signed-ranked test, p < 0.05, corrected for
multiple comparisons). These very different patterns of correlations with the RTs suggest that
although delta/theta synchronization coordinates directly the motor actions, the functional
significance of gamma-band synchronization in coordinating motor actions is dependent on
perceptual and/or decision processes that have large temporal variability.
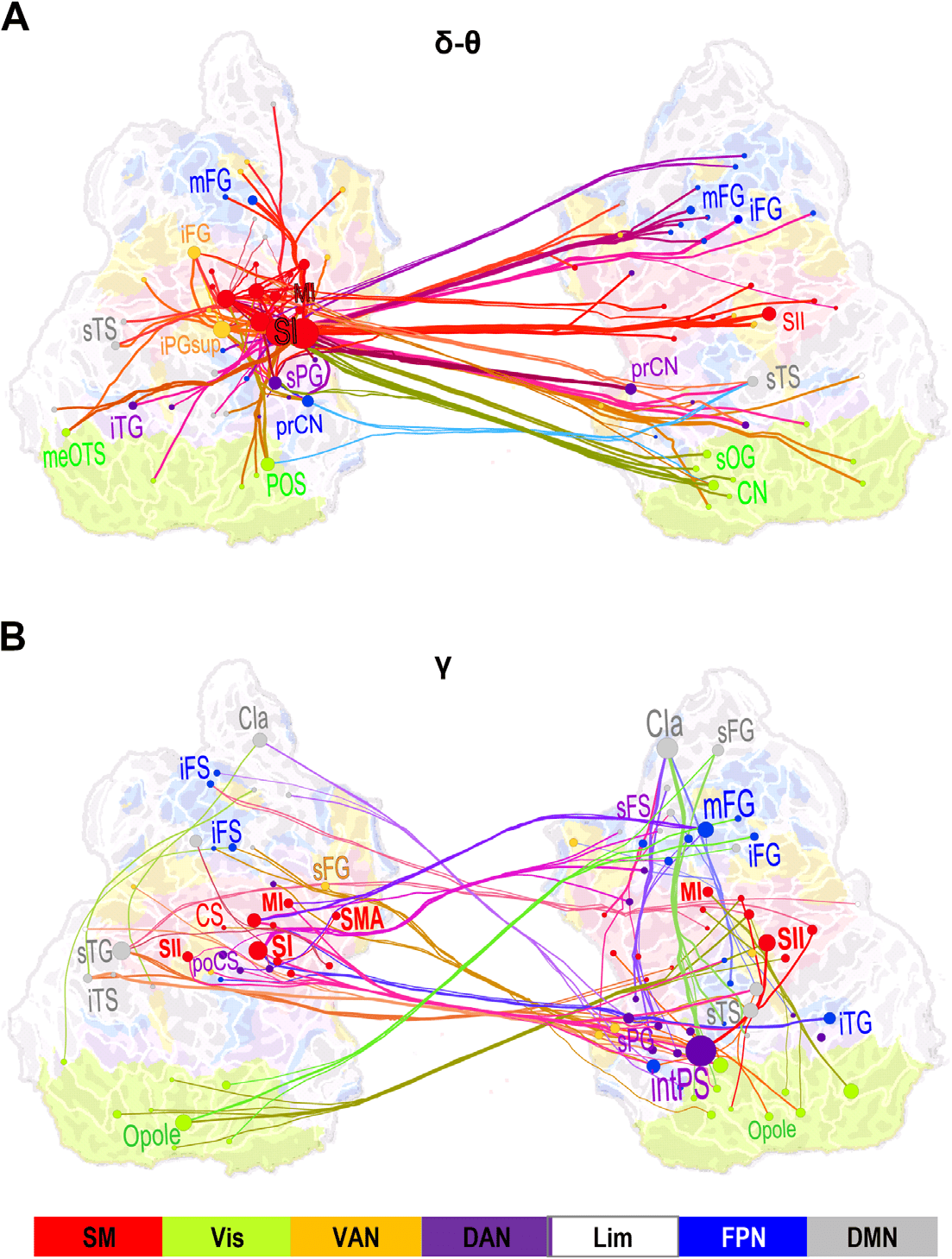
Large-Scale Synchrony Connects Sensorimotor Areas with Prefrontal and Posterior Parietal Cortices
To identify the cortical areas connected by large-scale delta/theta- and gamma-band synchro-
nization, we identified both the most central of the significant interareal connections and key
cortical areas, that is, the network hubs, by using PageRank for those time-frequency win-
dows where synchronization was significantly greater for Hits than for Misses. To obtain a
coarse localizer for the functional subsystems of the cerebral cortex, we co-localized our cor-
tical parcellations with the seven predominant systems defined by intrinsic fMRI BOLD signal
correlations (Yeo et al., 2011). In the delta/theta band, the strongest connections for the differ-
ence between Hits and Misses were observed bilaterally in the sensorimotor (SM) system in the
parcels corresponding to the contralateral primary sensory area (SI), secondary somatosensory
areas (SII), and motor area (MI). Additionally, delta/theta-band synchronization connected SM
with the bilateral middle frontal gyrus (mFG) in the dorsolateral prefrontal cortex (dlPFC) and
with nodes in the visual cortex (Figure 5A). In the mid-gamma band, the most central con-
nections were observed between contralateral SM, the most central hubs being SI, SII, and
MI, and the ipsilateral mFG and bilateral inferior frontal gyrus (iFG) of the dPLFC as well as
with the intPS of the PPC (Figure 5B). Functionally, mFG and iFG belong to the fronto-parietal
network (FPN) whereas intPS belongs to the dorsal attention network (DAN). Gamma-band
synchronization hence connected SM to FPN and DAN. In addition, mid-gamma-band syn-
chronization connected mFG and intPS ipsilaterally.
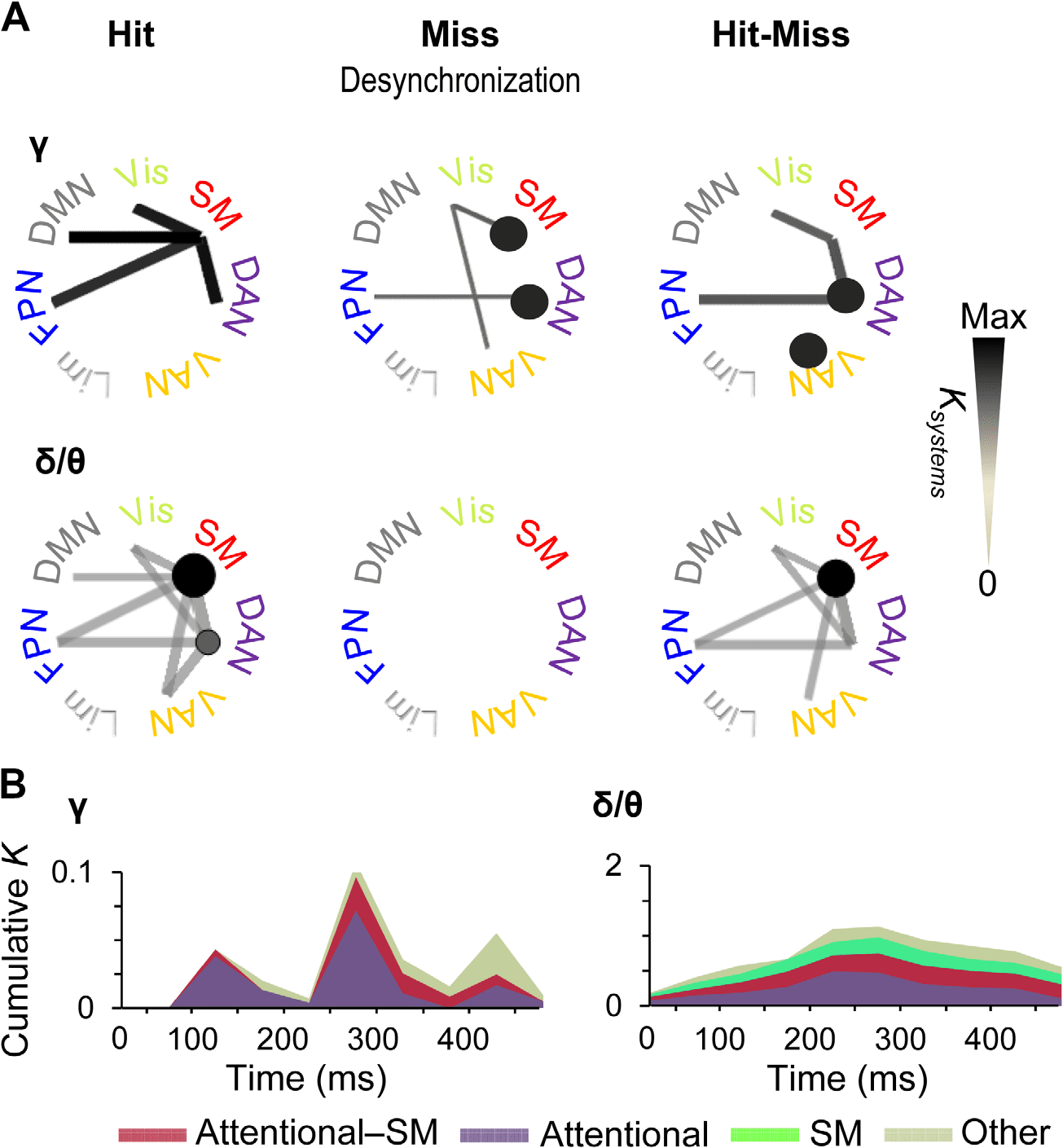
Large-Scale Synchrony Among Functional Subsystems
To complement the analyses of strongest connections, we obtained a systems-level view of all
significant observations of interareal connectivity by identifying which functional brain sys-
tems were coupled by greater fractions of significant connections (Ksystems) than expected by
chance (multiple-comparisons controlled graph permutation test, p < 0.05, see Methods).
This analysis showed that for Hits, gamma-band synchronization connected the SM system
with DAN and FPN as well as with the default mode network and visual system (Figure 6A).
Interestingly, for Misses, the suppression of synchronization below baseline levels was found
both within SM and DAN as well as among SM, ventral attention network (VAN), and the
visual system, and between DAN and FPN. Hence, when the stimuli were not detected, atten-
tional networks were uncoupled by suppression of gamma-band synchronization. In addition,
the significant differences in synchronization between Hits and Misses were more prevalent
between DAN and SM, within DAN and within VAN, and between DAN and FPN than ex-
pected for comparable random networks. Thus, these system-level results corroborate the
findings of the most central connections and showed that gamma synchronization among so-
matosensory and attentional systems predicts stimulus detection in both the delta/theta- and
gamma bands. Hit-related synchronization was found among SM and attentional systems for
both frequency bands, the key difference being that in the delta/theta band, within-system
synchronization was found in SM, whereas in the gamma band it was robust within DAN and
VAN.
Network Neuroscience
448
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
2
4
4
4
2
1
0
9
2
2
4
9
n
e
n
_
a
_
0
0
0
3
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Large-scale synchronization from perception to action
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
2
4
4
4
2
1
0
9
2
2
4
9
n
e
n
_
a
_
0
0
0
3
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 5. Large-scale synchronization connects somatosensory and attentional brain systems. (A)
Graph of the significant differences in the strength of interareal phase synchrony as estimated with
PLV between Hits and Misses in the delta/theta-frequency band (δ/θ, 3–7 Hz) and in the time win-
dow of 125–275 ms from stimulus onset (cf. Figure 2C). Lines connect the coupled parcels and line
colors are determined by the parcel brain systems (see below). Delta/theta-band synchronization
was centered on the contralateral (left hemispheric) sensorimotor (SM, red) system, and in particu-
lar, on the primary somatosensory cortex (SI) therein that was strongly and bilaterally coupled with
frontal and parietal regions. (B) Graph of gamma-band synchronization (γ, 40–60 Hz) reveal sig-
nificantly stronger connections for Hits than Misses over 225–375 ms from stimulus onset. Stronger
gamma-band synchronization for Hits than Misses was observed within SM and between SM and
the ipsilateral frontoparietal and dorsal (FP and DA, blue and purple) attention networks. Graphs
are displayed on an inflated and flattened cortical surface with 300 (A) and 200 (B) of the most
central edges based on parcel PageRank centralities selected for visualization. SI is primary and
SII is secondary somatosensory area. MI and SMA are primary and supplementary motor areas, re-
spectively. Parcel and corresponding node colors indicate the Yeo-atlas brain systems derived from
BOLD intrinsic connectivity connectome. SM = somatomotor (SM), green = visual (Vis), yellow =
ventral attention network (VAN), purple = dorsal attention network (DAN), white = limbic (Lim),
gray = default mode network (DMN).
To illustrate the temporal evolution of the system-scale gamma synchronization and specif-
ically to compare quantitatively the contributions of task-positive attention and sensory net-
works, we estimated the total connection densities of significant system-system couplings
separately for within-attentional, attentional-sensorimotor, and all other systems (Figure 6B).
Network Neuroscience
449
Large-scale synchronization from perception to action
Here, attentional systems included FPN, DAN, and VAN, whereas the “other” systems included
the default mode, visual, and limbic systems. We found system-level gamma synchronization
to emerge in three waves. In the first wave at around 100–150 ms from stimulus onset, connec-
tivity was almost completely dominated by coupling within the attentional systems, with only a
minor sensory-attentional fraction (Figure 6B, left). In the second wave at around 250–300 ms,
coupling within-attentional systems remained predominant, but sensory-attentional coupling
became much more salient. In the third wave, at around 450 ms from stimulus onset in which
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
2
4
4
4
2
1
0
9
2
2
4
9
n
e
n
_
a
_
0
0
0
3
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 6. Time-varying subsystem connectivity within and between attentional systems and SM.
(A) Connection densities of significant interareal gamma-band (40–60) Hz and delta/theta-band
(3–7 Hz) synchronization among Yeo-atlas brain systems for Hits and Misses compared with base-
line and for their difference. Only such system-system connections are shown that exhibit greater
connection densities than expected by chance in shuffled graphs (p < 0.05, permutation statistics,
see Methods). The color, line width, and radius of circles of the system-system connections indicates
the connection density of significant couplings (Ksystems) within (circles) or between (lines) the func-
tional systems in a time window of 225–375 ms. (B) Time-resolved cumulative connection densi-
ties (K) of gamma- and delta/theta-band synchronization estimated separately for within-attentional
(DAN, FNP, and VAN), between SM and attentional, and all other functional subsystems. Synchro-
nization in task-positive sensory and attentional systems predicted subsequent conscious perception
in gamma band, whereas synchronization in the delta/theta band was observed in SM along with
connections to and within-attentional systems throughout the time course of stimulus detection.
Network Neuroscience
450
Large-scale synchronization from perception to action
on average also the behavioral responses were given, connectivity in other systems was most
prevalent. In contrast, delta synchronized emerged in one wave, which peaked between 200
and 300 ms after stimulus onset. Furthermore, it predominantly comprised synchronization
within SM and attentional systems and between these two (Figure 6B, right).
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
2
4
4
4
2
1
0
9
2
2
4
9
n
e
n
_
a
_
0
0
0
3
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
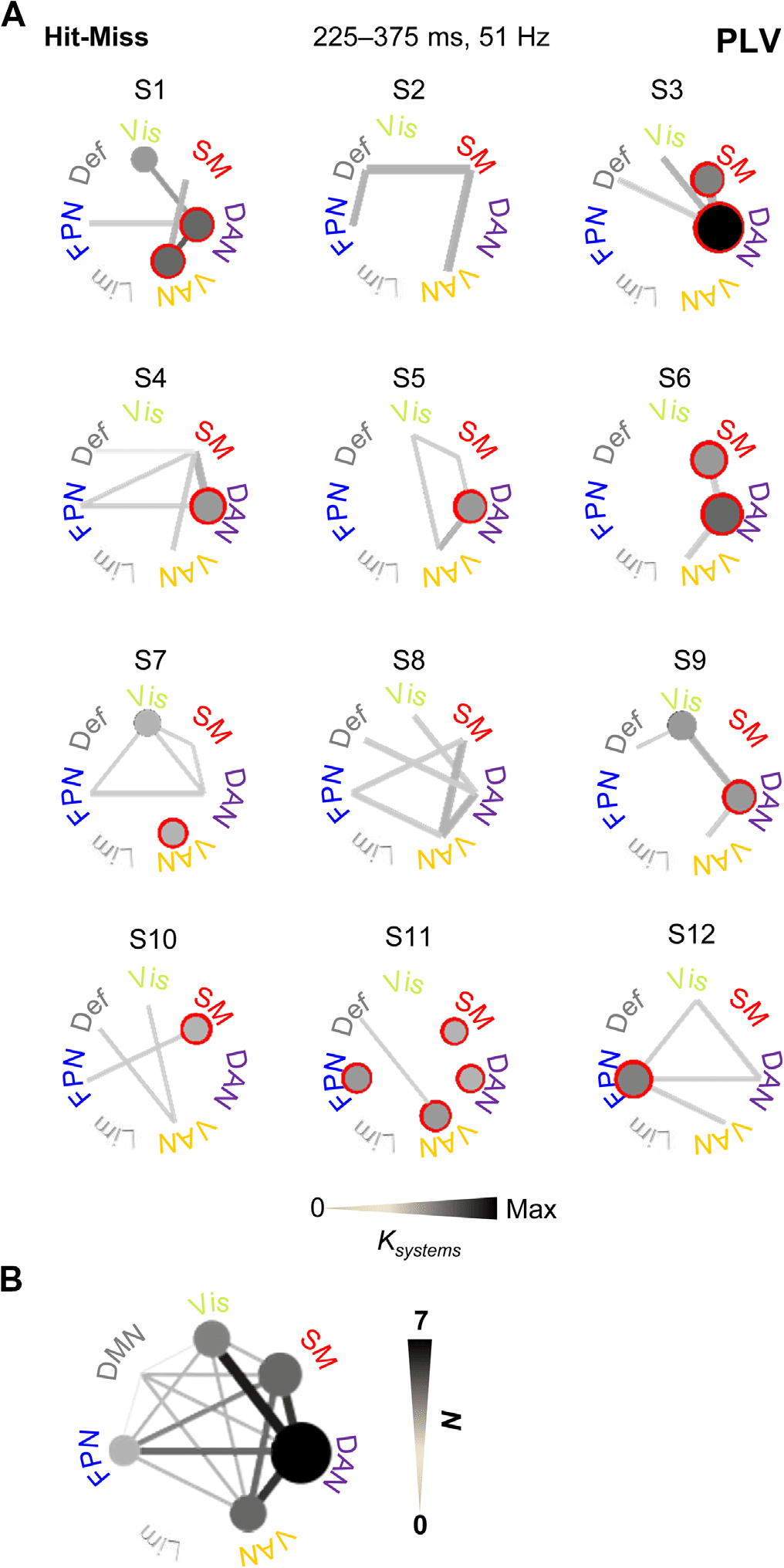
Figure 7. Synchrony within and between attentional and sensorimotor systems is frequently ob-
served in individual subject’s statistical analyses. (A) All significant connections among functional
subsystems separately for each subject. Significant connections are shown with gray tone that is
scaled relative to the connection density between (lines) or within (circles) the subsystems. Red
surroundings indicate the most common within-subsystem connections across the subjects. (B) All
significant connections across all subjects in individual subject statistical analyses. Line color indi-
cates the number of subjects (N) in which significant connections were were observed.
Network Neuroscience
451
Large-scale synchronization from perception to action
Single-Subject Statistics Corroborate The Robustness of Gamma-Band Synchronization in the
Sensorimotor Decision Process.
To corroborate the group-level analyses and to assess the robustness of gamma-band synchro-
nization in single subjects, we investigated individual interareal synchrony with single-subject
permutation statistics for the mid-gamma band (51 Hz) in the time window of 225–375 ms
where synchronization was most extensive in the group data. We estimated subsystem-level
connectivity as above separately for significant connections across subjects. This approach re-
vealed that synchronization was most prominent within SM, FPN, DAN, VAN, and the visual
system, and most repeatedly connected SM with DAN as well as DAN with VAN and the visual
system (Figure 7A). Even with considerable interindividual variability, in 9 out of 12 participants
the SM system was connected with an attentional system, and in all subjects synchronization
was observed within and/or between these functional systems (Figure 7B). These findings thus
confirmed that the correlation of large-scale gamma-band synchronization among SM, FPN,
DAN, and VAN with perception is a robust phenomenon even in individual subjects.
DISCUSSION
We aimed in this study to identify the large-scale networks of phase-locked cortical regions
supporting the neuronal processing from perception to perceptual decisions and to actions.
Building on accumulating evidence for the putative essential roles of large-scale neuronal inter-
action networks in cognitive integration (Baker et al., 2014; de Pasquale et al., 2015; Deco
et al., 2015; Petersen & Sporns, 2015; Siegel et al., 2012) and of neuronal phase coupling
in regulating the information transfer (Fries, 2015), we assessed the role of dynamic large-
scale phase synchronization in the emergence of coordinated neuronal processing that ac-
complishes the transition from sensory perception to action generation.
This is the first study to show, using human source-reconstructed noninvasive MEG or EEG
data, that dynamic interareal synchronization characterizes the neuronal activity cascade from
perception of sensory stimuli, to perceptual decisions, and to actions. We revealed the spa-
tiotemporal structure of this synchronization by using time-resolved synchrony analyses and
localized the key cortical areas of synchronization networks. Importantly, we found that the
strength of this synchronization was correlated with both detection performance (HR) and RTs,
and hence is functionally significant.
We observed that two concurrent networks of transient synchronization, one in the delta/
theta- and another in the gamma-frequency band, connected the sensory, fronto-parietal, and
motor systems from perception to action. Delta/theta-band synchronization was pronounced
within the SM system and between SM and attentional systems. In contrast, gamma-band syn-
chronization was first observed within the fronto-parietal attention networks, and subsequently
also between attention networks and the SM system, but it was not found to connect nodes
within SM. These couplings connecting the key task-relevant cortical structures suggest that
they could be functionally significant in the perception-action cycle. In line with their func-
tional significance, the strength of delta-band and the timing of gamma-band synchronization
were associated with the intertrial variability of the subsequent response times. These data thus
demonstrate that the large-scale network interaction mechanisms achieving perception, per-
ceptual decisions, and generation of actions may be achieved by phase coupling of neuronal
oscillations in delta/theta and gamma bands.
These data now show that in addition to evoked responses and local modulations of
oscillation amplitudes (Blankenburg et al., 2003; Hirvonen & Palva, 2016; Jones et al., 2007;
Network Neuroscience
452
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
2
4
4
4
2
1
0
9
2
2
4
9
n
e
n
_
a
_
0
0
0
3
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Large-scale synchronization from perception to action
King et al., 2016; Nierhaus et al., 2015; Taskin et al., 2008), somatosensory perception is also
characterized by concurrent induced neuronal synchronization connecting the task-relevant
cortical areas. In summary, these data provide evidence for the overarching hypothesis that
neuronal synchronization may coordinate communication during cognitive operations (Fries,
2015) and that long-range synchronization may play a key role in the emergence of conscious
sensory perception (Dehaene & Changeux, 2011; Engel & Singer, 2001; Tallon-Baudry, 2012)
and sensorimotor decisions (Donner et al., 2009a; Engel & Fries, 2010).
Long-Range Synchrony May Coordinate Processing from Perception to Action
The interest toward large-scale phase coupling as a putative mechanism for the emergent
coordination of distributed processing into coherent cognitive operations stems from two
established lines of evidence. First, phase coupling—synchronization—in a given neuronal
assembly endows greater postsynaptic impact than internally uncoupled, incoherent assem-
blies (Konig et al., 1995; Singer, 1999). Second, interareally organized phase relationships or
phase coupling may play a powerful role in opening or closing “channels” for communication
among brain areas by facilitating communication during the high-excitability phases of neu-
ronal oscillations (Fries, 2015). Cellular-level and animal model studies thus provided a strong
mechanistic framework for the likely functional roles of interareal phase coupling. None-
theless, because of significant methodological challenges, many of the resulting hypotheses
have remained unaddressed at the level of large-scale cortical interaction networks and their
relationship with human cognitive operations.
Prior fMRI-EEG studies have shown that activity in sensory and fronto-parietal brain areas is
related to both conscious perception and generation of actions in response to somatosensory
stimuli (Blankenburg et al., 2003; Nierhaus et al., 2015; Taskin et al., 2008), whereas EEG and
MEG studies have shown that early evoked responses are stronger for detected somatosensory
stimuli (Hirvonen & Palva, 2016; Jones et al., 2007; Nierhaus et al., 2015). Yet, the role of large-
scale network synchronization in coordinating the perception and actions has remained poorly
understood. A seminal study using intracranial EEG (iEEG) recordings from epileptic patients
found that long-distance beta synchronization between frontal, parietal, and visual areas char-
acterized visually presented words when these were consciously perceived but not if they re-
mained undetected (Gaillard et al., 2009). Whether similar long-range synchronization would
characterize also the perception of sensory stimuli in other modalities, in different experimen-
tal paradigms, and in MEG/EEG data obtained over the whole cortex has remained elusive.
Our central aim was to investigate whether large-scale networks of neuronal phase syn-
chronization would be essential in the transformation of sensory processing to sensorimotor
decision and actions. To this end, we used MEG recordings combined with several advanced
data analysis approaches and graph theory (Bullmore & Sporns, 2009) to identify the spa-
tiotemporal patterns of network synchronization from the perception of weak somatosensory
stimuli presented at a threshold of detection to the action. We found that as hypothesized,
dynamic large-scale synchronization characterized this process. Large-scale synchronization
was observed concurrently in delta/theta and gamma bands showing that even the simplest
perceptual process involves dynamic synchronization in multiple frequencies. Using a novel
parcellation scheme (Korhonen et al., 2014), combined with the novel edge-bundling
approach (Wang et al., 2018), we also identified the network structures of these networks.
Delta/theta-band synchronization connected the contralateral SI with ipsilateral SI and SII bi-
laterally and thereby the brain regions underlying the processing of somatosensory information
in monkeys (Haegens et al., 2014; Tauste Campo et al., 2015) and humans (Hegner et al., 2016;
Network Neuroscience
453
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
2
4
4
4
2
1
0
9
2
2
4
9
n
e
n
_
a
_
0
0
0
3
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Large-scale synchronization from perception to action
Hirvonen & Palva, 2016; Jones et al., 2007; Jones et al., 2009; Li Hegner et al., 2015; Papadelis
et al., 2016). This connectivity pattern positions delta/theta-band synchronization to directly
support the processing of somatosensory information. These nodes were also connected to the
motor cortex (MI) as well as to the contralateral PFC supporting a close link and dynamic inter-
actions between perceptual (Gutteling et al., 2011; Wohlschlager, 2000) and decision-making
processes (Donner et al., 2009b; Gould et al., 2012; Kaplan et al., 2017; Tosoni et al., 2008)
with the coordination of motor actions. Delta/theta-band synchronization could thus underlie
the coordination of neuronal processing achieved collectively in motor and sensory cortices.
In contrast, gamma-band synchronization did not connect cortical areas within SM but
coupled the contralateral SI and SII with ipsilateral sFS, mFG, and intPS. These areas are the
key nodes in the fronto-parietal and DAN, FPN, and DAN, as indicated both by the whole-
brain as well as by the subsystem analyses (Murakami & Okada, 2006; Power et al., 2011;
Spadone et al., 2015), respectively. These FPN/DAN nodes were also mutually connected
in both hemispheres, suggesting that gamma-band synchronization could here coordinate
neuronal communication specifically within the attentional system and between the atten-
tional and sensory systems. In summary, these data revealed many subnetworks among the key
cortical areas that were correlated with conscious somatosensory perception, decisions, and
actions in great anatomical and temporal precision. Unexpectedly, the data-driven all-to-all
data analysis approach also revealed the visual system to be consistently connected with the
sensorimotor system both in delta/theta and gamma bands. This may reflect the responsive-
ness of the visual cortex also to the somatosensory stimulation (Nordmark et al., 2012) and/or
complementary representation of the perceived somatosensory stimuli in the visual cortex
(Orlov et al., 2010), which is also plausible as the subjects had eyes closed during the task.
Suppression of Gamma-Band Synchronization for Misses
Intriguingly, we found gamma-band synchronization to be suppressed below baseline levels
for stimuli that remained undetected. These stimuli were hence subliminal in the terms that
they modulated brain activity but in an inadequate manner to reach conscious perception
(Dehaene et al., 2006; Dehaene & Changeux, 2011). Prior studies have reported that sublimi-
nal stimuli evoke neuronal activity in primary sensory cortices but not later in sensory process-
ing hierarchy for visual stimuli (Dehaene et al., 2001; Del Cul et al., 2007; Grill-Spector et al.,
2000). Our data showed that not only is the synchronization stronger for the consciously per-
ceived and reported than for unperceived and unreported stimuli, but also that synchronization
patterns between Hits and Misses are qualitatively and genuinely distinct. The suppression of
gamma activity for Misses is, however, well in line with prior fMRI studies reporting negative
BOLD fMRI signal responses in contralateral SI, SII, and SMA for subthreshold somatosen-
sory stimuli (Blankenburg et al., 2003; Nierhaus et al., 2015; Taskin et al., 2008). Suppres-
sion of synchronization may reflect active uncoupling of task-relevant cortical areas to block
conscious access from subliminal stimuli or it may reflect non-conscious stop-signals to inter-
rupt motor responses (van Gaal et al., 2008).
Temporal Pattern of Large-Scale Synchronization and Correlation with RT Supports Distinct Functions
for Delta/Theta and Gamma-Band Synchronization in Perceptual Decisions and Actions
Finally, to reveal whether network synchronization would be stable or show dynamic changes
as a function of time, we used time-resolved analysis of network synchronization. Interestingly,
despite the stable profile of gamma-band synchronization in the time-frequency representa-
tions, this analysis showed that the gamma-band connectivity patterns changed as a function
Network Neuroscience
454
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
2
4
4
4
2
1
0
9
2
2
4
9
n
e
n
_
a
_
0
0
0
3
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Large-scale synchronization from perception to action
of time while those of delta-band synchronization pattern were more stable. During the early
time windows, gamma-band synchronization connected nodes only within the attention net-
works, after which the gamma-band network comprised also the SM system, including motor
cortex (M1). This temporal pattern suggests that in the present task, attentional systems drive
sensory perception in line with prior studies, which suggest that perception is related to differ-
ences in the prestimulus network properties (Weisz et al., 2014; Frey et al., 2016) and predicted
by infraslow fluctuations of electrical activity (Monto et al., 2008). Furthermore, fMRI BOLD
signal fluctuations in attentional but not in sensory system are also correlated with percep-
tual performance in a threshold-stimulus detection task like ours (Sadaghiani et al., 2009). As
PFC and PPC coordinate both perceptual decisions (Donner et al., 2009b; Goard et al., 2016;
Hegner et al., 2016; Kaplan et al., 2017; Siegel et al., 2015; Tosoni et al., 2008) and conscious
perception (Dehaene & Changeux, 2011; Gaillard et al., 2009; Hirvonen & Palva, 2016;
Li Hegner et al., 2015; Salti et al., 2015), we speculate that gamma-band synchronization
during the later time windows, however, could underlie conscious perceptual decisions of the
weak somatosensory stimuli. The role of gamma-band synchronization in coordinating percep-
tual decisions was also supported by the distinct temporal profiles of synchronization for fast
and slow actions. Interestingly, fast and slow responses were associated with distinct gamma-
band patterns, which illustrates the presence of large intertrial variability in the functionally
significant gamma-band synchronization patterns. More specifically, the neuroanatomical and
temporal patterns of gamma-band synchronization could plausibly reflect a sequence of net-
work reconfigurations from conscious sensory decisions to the coordination of motor actions.
In contrast, the temporal profile of delta/theta-band synchronization was more stable. This
may be caused by more stable network configuration of synchronization or the poorer temporal
resolution of wavelets compared with the gamma band. Synchronization was confined to SM
system across time. This together with an increase in the strength of synchronization but not
a change in temporal profile in response to faster RTs suggest that delta-band synchronization
is related to evidence accumulation of sensory information and coordination of motor actions
but not the achieving conscious sensory perception per se. Overall, in line with prior stud-
ies of local neuronal processing in humans (Donner et al., 2009b; Gould et al., 2012; Kaplan
et al., 2017; Medendorp et al., 2011; Tosoni et al., 2008) our data suggest that the sensorimotor
decision process and the associated actions involve overlapping neuronal circuits also at the
large-scale network level.
Conclusions
In summary, these findings constitute evidence for the hypothesis that dynamic large-scale
network synchronization plays an essential role in the coordination of neuronal processing to
achieve conscious somatosensory perception, perceptual decisions, and initiation of a motor
response.
METHODS
An overview of the workflow is given in Figure 1. All analyses, if not stated otherwise, were
analyzed with LabVIEW software (National Instruments), available on request.
Subjects and Recordings
MEG was recorded from 12 healthy right-handed subjects (27.5 ± 4.5 years of age, mean ±
standard deviation; 7 women) with a 306 channel MEG (Elekta Neuromag, Helsinki, Finland)
Network Neuroscience
455
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
2
4
4
4
2
1
0
9
2
2
4
9
n
e
n
_
a
_
0
0
0
3
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Large-scale synchronization from perception to action
at 600-Hz sampling rate as described earlier in (Hirvonen & Palva, 2016). Electromyogram
(EMG) was recorded to detect the thumb movement responses and T1-weighted anatomical
MRI scans were obtained for source localization. The study was approved by the Coordinat-
ing Ethical Committee of the Helsinki University Hospital, and the subjects gave a written,
informed consent prior to their participation in the experiment.
Task
We used a continuous stimulus detection task in which 0.2-ms electrical stimuli were given
with an intensity (mean 4.3 ± 0.6 mA) at the threshold of detection (Hirvonen and Palva 2016;
Monto et al., 2008; Palva et al., 2005). The intensity was individually calibrated before the
recordings to a level that yielded a ∼50% detection rate and then maintained constant during
the recordings. The stimuli were delivered at uniformly random 1.5- to 4.5-s intervals to the
right index finger in two separate approximately 30-min blocks. The subject’s task was to report
with a right thumb twitch whenever he or she perceived the stimulus. On average, 1,073 ±
207 (mean ± SD, n = 12) trials were acquired per subject. No trials were rejected.
Behavioral Performance
HR was defined to be the proportion of detected stimuli of all stimuli. The stimuli were clas-
sified as consciously perceived, or “Hits,” if the subject performed the right thumb twitch
response within 0.1–1.5 s after the stimulus onset and “Misses” if stimulus was not associated
with response (Figure 2A). The responses were identified from filtered (low- and high-pass fi-
nite impulse-response filter with pass band 30–190 Hz) and absolute valued EMG recordings
using 10 baseline standard deviations as the threshold for a response and the first crossing
of this threshold as the RT (Hirvonen & Palva, 2016; Monto et al., 2008; Palva et al., 2005).
Stimuli to which no response was observed were categorized as unperceived, or “Misses.” To
estimate the false alarm (FA) rate, we estimated the number of responses between −1.5 s to 0 s
before stimulus onset, during which there should not have been any responses to somatosen-
sory stimuli. For this estimation, only the trials with a distance at least 3 s to previous and next
trial were selected. FA estimate was then computed as the proportion of these responses from
all the stimulus events (Figure 2B). Delayed responses and FA were not analyzed because of
the exceedingly small number of trials in these categories.
MEG Data Preprocessing, Filtering, and Source Analysis
The signal space separation method (tSSS) was used to remove extracranial noise from the
raw MEG recordings, and independent component analysis was used to identify and exclude
components associated with eye movements and cardiac artifacts (Figure 1, a). In short, pre-
processed MEG sensor time series were filtered using Morlet wavelets into 38 frequency bands
covering 3–120 Hz with equal distances between neighboring frequencies on the log scale
by using time-frequency compromise parameter m = 5 (Figure 1, b). We used FreeSurfer
software (http://surfer.nmr.mgh.harvard.edu/ ) and minimum-norm estimate (MNE) toolkit for
volumetric segmentation of MRI images, and reconstruction of anatomical surfaces and cor-
tical parcellation (Destrieux et al., 2010; Fisch et al., 2002) (Figure 1, c). The MNE toolkit
(http://www.nmr.mgh.harvard.edu/martinos/userInfo/data/sofMNE.php) was used to create
three-layer boundary element models, cortically constrained source models, MEG-MRI co-
localization and for preparation of the forward model and MNE inverse operators (Hamalainen
& Ilmoniemi, 1994) (Figure 1, d). The source models had dipole orientations fixed to the pial
Network Neuroscience
456
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
2
4
4
4
2
1
0
9
2
2
4
9
n
e
n
_
a
_
0
0
0
3
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Large-scale synchronization from perception to action
surface normals and a 7-mm source-to-source separation throughout the cortex, which yielded
models containing 6,000–8,000 source vertices.
We used the MNE inverse operators in the form of dynamic statistical parametric map
(dSPM) operators (Dale et al., 2000) so that the noise-covariance matrices (NCM) were ob-
tained from the baseline data 0.75–0.2 s prior to stimulus onsets and by using 0.05 as the
regularization constant. The NCMs and hence the inverse operators were prepared separately
for each Morlet-wavelet filter frequency by using lambda as 0.05. We used an atlas-based
analysis strategy (Hillebrand et al., 2012; Palva et al., 2010), in which the preprocessed and
Morlet-filtered MEG sensor time series were inverse modeled sample-by-sample into source
time series (Figure 1, d) that were then collapsed into time series of 400 cortical parcels by
using individually optimized collapse operators (Figure 1, e) (Korhonen et al., 2014). In these
collapse operators, the source vertices were weighted with parcel-signal reconstruction accu-
racy, and only the set of vertices that yielded best reconstruction accuracy were used
(Korhonen et al., 2014). For creating these weighted collapse operators, and more precisely, to
assess parcel-signal reconstruction accuracy, another set of NCM and inverse operators were
computed. To obtain NCM only for this “fidelity optimization” inverse-forward modeling,
we applied 0.1–45 Hz pass-band filtering to MEG time series by using low- and high-pass
finite impulse-response filters. The 400-parcel parcellation was obtained from a precursor
“Destrieux” atlas of 148 parcels (Destrieux et al., 2010) by iteratively splitting the largest parcels
along their most elongated axis and using the same parcel-wise splits for all subjects. Using
neuroanatomical labeling as the anatomical “coordinate system” eliminates the need for inter-
subject morphing in group-level analyses, which would have compromised individual anatom-
ical accuracy.
Forward and Inverse-Modeled Trial-Shuffled Surrogates for Evoked Component Estimation
To account for the artificial synchronization attributable to evoked responses and/or phase
locking of ongoing activity to the stimuli, we created trial-shuffled surrogate data (Figure 1, f).
To reconstruct the effects of signal mixing at MEG acquisition and inverse modeling, we ap-
plied a forward-inverse modeling–based surrogate construction approach that reconstructs the
evoked components and their spatial spread caused by the signal mixing inherent to MEG/EEG
and preserves local source topography changes, amplitude dynamics, and changes in autocor-
relation structures that may constitute significant confounders (Palva & Palva, 2012). This ap-
proach hence provides a good surrogate for selectively identifying the true induced interareal
interactions for which the conventional trial shuffling (Lachaux et al., 1999) is insufficient in
the presence of signal mixing.
We used the source-modeled single-trial data in the 400-parcel parcellation as a new parcel
time series so that first in forward modeling the activity time series of each source vertex of
a given parcel for each trial was simulated with the parcel time series of a random trial.
In
each new “trial” of the surrogate data, the cortical parcels were simulated with random-trial
time series of the original data. We then source reconstructed these sensor-level surrogate
data with procedures identical to those used for real data. This procedure thus abolishes true
non-stimulus-locked phase correlations between parcels but reconstructs both the evoked and
stimulus-phase-locked components as well as the spread of the signals caused by MEG data
acquisition and inverse modeling. Phase correlation analyses (Figure 1, g) were then performed
with these surrogate source data identically to those of real data for 10 independent realizations
of the surrogate data. The means of surrogate data were compared against the corresponding
real data in Fig. 3 (Figure 1, f).
Network Neuroscience
457
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
2
4
4
4
2
1
0
9
2
2
4
9
n
e
n
_
a
_
0
0
0
3
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Large-scale synchronization from perception to action
Analysis of Phase Synchrony
All-to-all phase coupling–based functional connectivity, that is, phase synchrony, between all
parcel pairs, was estimated in reconstructed source space for all 38 narrow-band time series.
We first estimated the complex-valued phase-locking value (cPLV) between all parcel pairs.
The cPLV is defined as
cPLV =
1
N ∑N
n=1
(cid:2)
ei(θX(n)−θY(n))
(cid:3)
(1)
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
2
4
4
4
2
1
0
9
2
2
4
9
n
e
n
_
a
_
0
0
0
3
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
where N denotes sample number, θX and θY are phases of narrow-band time series of X and
Y, and I denotes the imaginary unit (Lachaux et al., 1999; Palva et al., 2005). We assess here
phase synchrony both by using the PLV, PLV = |cPLV|, and the imaginary part of cPLV (iPLV),
iPLV = |im(cPLV)|, where im indicates the imaginary value operator. PLV, a commonly used
measure for phase correlations, is essentially equally sensitive to phase coupling at all phase
lags and thus is sensitive to artificial zero-phase-lag correlations caused by MEG signal mixing
and spatial leakage in inverse modeling. iPLV, on the other hand, is insensitive to near-zero
and near-pi phase lags and thus does not yield false positives attributable exclusively to signal
mixing (Brookes et al., 2014) (Figure 1, g).
To compensate for the fact that both PLV and iPLV are biased by the number of samples, the
numbers of Hit and Miss trials were balanced within subjects before the PLV/iPLV analyses by
keeping only those events of the larger conditions, which are closest to the onset latencies of
the smallest condition. For each frequency, the cPLV values were obtained across samples in
100-ms time windows and across trials. Using 50% overlap, we obtained 26 time windows
from −725 ms to 525 ms around the stimulus.
Group Statistical Analyses
Group statistical contrasts between conditions or between prestimulus baseline and post-
stimulus periods were performed separately for each frequency, time window, and parcel pair.
Before group statistics, mean baseline values from 125 to 225 ms prior to stimulus onset were
subtracted. The significance of the difference between stimulus processing and baseline or
between Hits and Misses was estimated with the Wilcoxon signed-rank test (p < 0.05). To re-
duce false discoveries arising from multiple comparisons, we pooled significant observations
for each contrast across all cortical parcels and time windows but separately for each frequency
band. Then we discarded as many least significant observations across parcels and across time
windows as were predicted by the alpha level (α) and the total number of comparisons (Nc),
α × Nc (Figure 1, h). This false discovery reduction was done separately for each frequency
band.
Removal of Parcels and Connections with Low Reliability
The poor source reconstruction accuracy of some cortical parcels limits the overall quality of
all-to-all interaction analyses. In this study, we utilized a simulation-based method to assess
the reliability of local source time-series reconstruction and interareal interaction estimates
(Korhonen et al., 2014). We used the subject cohort’s source models, that is, forward and
inverse operators that were derived from real MEG recordings, simulated independent parcel
time series as the ground-truth data, and then simulated a virtual MEG recording and source
reconstruction by forward and inverse modeling the ground-truth time series. Thus, assessing
the correlations between simulated and reconstructed data yields quantitative estimates of the
Network Neuroscience
458
Large-scale synchronization from perception to action
reconstruction accuracy and signal mixing between parcels. We used parcel f idelity ( fu) to
quantify the source reconstruction accuracy of a parcel u; it is defined as follows:
fu = |re(cPLV(xu, ˆxu))|
(2)
where xu is the simulated original time series of parcel u, and ˆxu is the source-reconstructed
time series of xu.
We used parcel-to-parcel infidelity (iuv) to quantify the amount of mixing from parcels u to
another parcel v, and it is defined as follows:
iuv = |re(cPLV(xu, ˆxv))|
(3)
where xu is the original time series of parcel u, and ˆxv is the forward- and inverse-modeled time
series of another parcel v. Thereby iuv quantifies the amount of signal from u that is observed in
the reconstructed signal of another parcel v, thus describing degree of signal leak from parcel u
to parcel v. To decrease the probability of reporting artificial and spurious synchronization due
to poor source reconstruction accuracy, we first removed parcels with fidelity lower than 0.11.
These parcels were located mostly in deep and/or inferior parts of the cortex and are known to
generate weak signals in M/EEG (Supporting Information Figure 1, Hirvonen, Monto, Wang,
Palva, & Palva, 2018). Next, we excluded cortical parcels close to the eyes because they are
known to include oculomotor artifacts in MEG. These parcels are mostly located afar (e.g.,
>5 cm) from the sensors such as the orbital frontal, anterior, and inferior temporal and me-
dial structures. In total, 20% of such parcels were excluded from analyses. In addition, we
also excluded parcel-parcel interactions that connected any parcels with low fidelity, that is,
interactions that would be very unlikely to be observable with MEG. In analyzed data, these
couplings would be thus much more likely to mirror signal-mixed interactions from other
sources and thus yield false positives. Taken together, 38% of interactions were eliminated
as being likely to be contaminated by poor reconstruction accuracy and linear mixing.
(Figure 1, i).
Visualization
To summarize our results, we used connection density, K, as a function of frequency bands to
indicate the fraction of statistically significant edges in a given experimental-condition contrast
(Figure 3). These data were used to identify frequency bands of interest (Figure 1, j). The graphs
(Figure 5) corresponding to these data were then visualized so that most central connections
were identified by the mean PageRank centralities of the connected nodes (Rubinov & Sporns,
2010). For graph visualization, we first evaluated the centrality of each edge ei(u, v) in the
graph by summing PageRank centrality of the source and target nodes:
Cei = sum(PRu + PRv)
(4)
where PR denotes the PageRank centrality. The PageRank is a measure of the importance of
a node, and therefore the Cei measure weights more to edges that connect important nodes
in a graph. Subsequently, we applied a novel hyperedge bundling approach to group edges
into bundles by their adjacency in signal mixing (Wang et al., 2018), which was derived from
the parcel-to-parcel infidelity function iuv described earlier (Figure 1, k). Briefly, this approach
bundles together edges that have large-signal mixing and hence which are spurious and arti-
ficially induced by the true neuronal connection. Thus, the hyperedge represents the under-
lying true interaction and its spurious reflections caused by signal spatial leakage. We excluded
Network Neuroscience
459
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
2
4
4
4
2
1
0
9
2
2
4
9
n
e
n
_
a
_
0
0
0
3
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Large-scale synchronization from perception to action
bundles of less than four edges, because true interareal synchrony is likely to be reflected in
large numbers of signal spread caused by spurious connections around each true connection. To
improve the neuroanatomical resolution of the bundles, we discarded edges with low central-
ity (< 50%) within each bundle. The starting number of edges was selected so that the final
edge number became 300 and 200 for graphs in Figure 5 so that the largest number of edges
was selected for graphs with largest K and smallest for smallest K.
To estimate interactions between the seven functional subsystems (Yeo et al., 2011), we
morphed the original 400 × 400 adjacency matrices into 7 × 7 subsystem interaction matri-
ces (Figures 6A and 7), and evaluated K of connections among each subsystem (Figure 1, l). To
test whether these K were greater than expected by chance for a random graph, we computed
5,000 randomizations of the same 400 × 400 matrices, keeping the numbers of significant
edges constant. K values of the original subsystems interaction matrix were reported as signif-
icant if they exceeded the 95th percentile of the K values in the randomized graphs.
Individual Subject Statistics
To estimate statistical significance of synchronization of the center frequency that was identi-
fied in the group-level analysis in single subjects, we used nonparametric permutation statistics
(Figure 1, m). We computed the difference in mean PLV between Hits and Misses, δI, within
each time window and for each pair of parcels. To obtain the permutation-based statistics, we
randomly redistributed the trials into Hitperm and Missperm and computed δIperm, the mean
PLV between the permuted conditions. The random permutation and PLV recomputation was
repeated for Nperm = 5,000 times. To obtain the subject-wise permutation test statistic, pperm,
we counted the portion of permutations that resulted in higher absolute difference in PLV than
the unpermuted conditions:
pperm = #(|δIperm| > |δI|)/Nperm,
(5)
where #() denotes the counting operation. The pperm statistic was then corrected for predicted
false discovery rate as explained above using α = 0.05 and FDR = 0.05.
ACKNOWLEDGMENTS
We thank Dr. Alexander Zhigalov and MSc Santeri Rouhinen for contributing analysis and
visualization tools.
AUTHOR CONTRIBUTIONS
Jonni Hirvonen: Data analysis; Visualization; Funding aquisition; Manuscript writing. Sheng
Wang: Software design. Simo Monto: Data collection; Software design; Data analysis; Vi-
sualization; Manuscript writing. Satu Palva: Experimental design; Data collections; Funding
aquisition; Manuscript writing. Matias Palva: Experimental design: Software design; Funding
aquisition; Manuscript writing.
FUNDING INFORMATION
This work was supported by the University of Helsinki Research Grants, Academy of Finland
(SA 267030, SA 266402 and SA 273807 to SP and SA 253130 and 256472 MP), Research
Foundation of the University of Helsinki to SP and JH, and Sigrid Juselius Foundation to SP and
MP for supporting this study.
Network Neuroscience
460
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
2
4
4
4
2
1
0
9
2
2
4
9
n
e
n
_
a
_
0
0
0
3
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Large-scale synchronization from perception to action
REFERENCES
Baker, A. P., Brookes, M. J., Rezek, I. A., Smith, S. M., Behrens,
T., Probert Smith, & P. J., Woolrich, M.
(2014). Fast transient
networks in spontaneous human brain activity. ELife, 3, e01867.
Blankenburg, F., Taskin, B., Ruben, J., Moosmann, M., Ritter, P.,
Curio, G., & Villringer, A. (2003). Imperceptible stimuli and
sensory processing impediment. Science (New York, N.Y.), 299,
1864.
Brookes, M. J., Woolrich, M. W., & Barnes, G. R. (2012). Measuring
functional connectivity in MEG: A multivariate approach insen-
sitive to linear source leakage. NeuroImage, 63, 910–920.
Brookes, M. J., O’Neill, G. C., Hall, E. L., Woolrich, M. W., Baker,
A., Palazzo Corner, S., Robson, S. E., Morris, P.G., & Barnes,
G. R. (2014). Measuring temporal, spectral and spatial changes
in electrophysiological brain network connectivity. NeuroImage,
91, 282–299.
Bullmore, E., & Sporns, O. (2009). Complex brain networks: Graph
theoretical analysis of structural and functional systems. Nature
Reviews Neuroscience, 10, 186–198.
Dale, A. M., Liu, A. K., Fischl, B. R., Buckner, R. L., Belliveau, J. W.,
Lewine, J. D., & Halgren, E.
(2000). Dynamic statistical para-
metric mapping: Combining fMRI and MEG for high-resolution
imaging of cortical activity. Neuron, 26, 55–67.
de Pasquale, F., Della Penna, S., Sporns, O., Romani, G. L.,
& Corbetta, M. (2015). A dynamic core network and global
efficiency in the resting human brain. Cerebral Cortex, 26,
4015–4033.
Deco, G., Tononi, G., Boly, M., & Kringelbach, M. L. (2015).
Contributions of
Rethinking segregation and integration:
whole-brain modelling. Nature Reviews. Neuroscience, 16,
430–439.
Dehaene, S., & Changeux, J. P. (2011). Experimental and theoreti-
cal approaches to conscious processing. Neuron, 70, 200–227.
Dehaene, S., Changeux, J. P., Naccache, L., Sackur, J., & Sergent, C.
(2006). Conscious, preconscious, and subliminal processing: A
testable taxonomy. Trends in Cognitive Sciences, 10, 204–211.
Dehaene, S., Naccache, L., Cohen, L., Bihan, D. L., Mangin,
J. F., Poline, J. B., & Riviere, D. (2001). Cerebral mechanisms
of word masking and unconscious repetition priming. Nature
Neuroscience, 4, 752–758.
Del Cul, A., Baillet, S., & Dehaene, S. (2007). Brain dynamics
underlying the nonlinear threshold for access to consciousness.
PLoS Biology, 5, e260.
Destrieux, C., Fischl, B., Dale, A., & Halgren, E. (2010). Auto-
matic parcellation of human cortical gyri and sulci using standard
anatomical nomenclature. NeuroImage, 53, 1–15.
Donner, T. H., Siegel, M., Fries, P., & Engel, A. K. (2009a). Buildup
of choice-predictive activity in human motor cortex during
perceptual decision making. Current Biology, 19, 1581–1585.
Donner, T. H., Siegel, M., Fries, P., & Engel, A. K. (2009b). Buildup
of choice-predictive activity in human motor cortex during
perceptual decision making. Current Biology, 19, 1581–1585.
Engel, A. K., & Fries, P. (2010). Beta-band oscillations—Signalling
the status quo? Current Opinion in Neurobiology, 20, 156–165.
Engel, A. K., & Singer, W. (2001). Temporal binding and the neural
correlates of sensory awareness. Trends in Cognitive Sciences, 5,
16–25.
Fisch, L., Privman, E., Ramot, M., Harel, M., Nir, Y., Kipervasser, S.,
Andelman, F., Neufeld, M. Y., Kramer, U., Fried, I., & Malach, R.
(2009). Neural “ignition”: Enhanced activation linked to percep-
tual awareness in human ventral stream visual cortex. Neuron,
64, 562–574.
Fischl, B., Salat, D. H., Busa, E., Albert, M., Dieterich, M.,
Haselgrove, C., van der Kouwe, A., Killiany, R., Kennedy, D.,
Klaveness, S., Montillo, A., Makris, N., Rosen, B., & Dale, A. M.
(2002). Whole brain segmentation: Automated labeling of neuro-
anatomical structures in the human brain. Neuron, 33, 341–355.
Frey, J. N., Ruhnau, P., Leske, S., Siegel, M., Braun, C., & Weisz,
N. (2016). The tactile window to consciousness is characterized
by frequency-specific integration and segregation of the primary
somatosensory cortex. Scientific Reports, 6, 20805.
Fries, P.
(2015). Rhythms for cognition: Communication through
coherence. Neuron, 88, 220–235.
Gaillard, R., Dehaene, S., Adam, C., Clemenceau, S., Hasboun,
D., Baulac, M., Cohen, L., & Naccache, L. (2009). Converging
intracranial markers of conscious access. PLoS Biology, 7, e61.
Goard, M. J., Pho, G. N., Woodson, J., & Sur, M. (2016). Distinct
roles of visual, parietal, and frontal motor cortices in memory-
guided sensorimotor decisions. ELife, 5, 10.7554/eLife.13764.
Gould, I. C., Nobre, A. C., Wyart, V., & Rushworth, M. F. (2012).
Effects of decision variables and intraparietal stimulation on
sensorimotor oscillatory activity in the human brain. Journal of
Neuroscience, 32, 13805–13818.
Gregoriou, G. G., Gotts, S. J., Zhou, H., & Desimone, R.
(2009).
Long-range neural coupling through synchronization with atten-
tion. Progress in Brain Research, 176, 35–45.
Grill-Spector, K., Kushnir, T., Hendler, T., & Malach, R. (2000). The
dynamics of object-selective activation correlate with recogni-
tion performance in humans. Nature Neuroscience, 3, 837–843.
Gutteling, T. P., Kenemans, J. L., & Neggers, S. F. (2011). Grasping
preparation enhances orientation change detection. PloS One, 6,
e17675.
Haegens, S., Vazquez, Y., Zainos, A., Alvarez, M., Jensen, O., &
Romo, R. (2014). Thalamocortical rhythms during a vibrotactile
detection task. Proceedings of the National Academy of Sciences
of the United States of America, 111, E1797–E1805.
Hamalainen, M. S., & Ilmoniemi, R. J. (1994). Interpreting mag-
netic fields of the brain: Minimum norm estimates. Medical &
Biological Engineering & Computing, 32, 35–42.
Hegner, Y. L., Lindner, A., & Braun, C. (2016). A somatosensory-
to-motor cascade of cortical areas engaged in perceptual deci-
sion making during tactile pattern discrimination. Human Brain
Mapping, 38, 1172–1181.
Hillebrand, A., Barnes, G. R., Bosboom, J. L., Berendse, H. W., &
Stam, C. J. (2012). Frequency-dependent functional connectivity
within resting-state networks: An atlas-based MEG beamformer
solution. NeuroImage, 59, 3909–3921.
Network Neuroscience
461
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
2
4
4
4
2
1
0
9
2
2
4
9
n
e
n
_
a
_
0
0
0
3
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Large-scale synchronization from perception to action
Hipp, J. F., Engel, A. K., & Siegel, M.
(2011). Oscillatory syn-
chronization in large-scale cortical networks predicts perception.
Neuron, 69, 387–396.
Hirvonen, J., Monto, S., Wang, S. H., Palva, J. M., & Palva, S. (2018).
Supporting Information for “Dynamic large-scale network syn-
chronization from perception to action.” Network Neuroscience,
2, 442–463. https://doi.org/10.1162/netn_a_00039
Hirvonen, J., & Palva, S. (2016). Cortical localization of phase and
amplitude dynamics predicting access to somatosensory aware-
ness. Human Brain Mapping, 37, 311–326.
Jones, S. R., Pritchett, D. L., Stufflebeam, S. M., Hamalainen,
M., & Moore, C. I. (2007). Neural correlates of tactile detection:
A combined magnetoencephalography and biophysically based
computational modeling study. Journal of Neuroscience, 27,
10751–10764.
Jones, S. R., Pritchett, D. L., Sikora, M. A., Stufflebeam, S. M.,
(2009). Quantitative analysis
Hamalainen, M., & Moore, C. I.
and biophysically realistic neural modeling of the MEG μ rhythm:
Rhythmogenesis and modulation of sensory-evoked responses.
Journal of Neurophysiology, 102, 3554–3572.
Kaplan, R., King, J., Koster, R., Penny, W. D., Burgess, N., &
Friston, K. J. (2017). The neural representation of prospective
choice during spatial planning and decisions. PLoS Biology, 15,
e1002588.
King, J. R., Pescetelli, N., & Dehaene, S. (2016). Brain mechanisms
underlying the brief maintenance of seen and unseen sensory
information. Neuron, 92, 1122–1134.
Konig, P., Engel, A. K., Roelfsema, P. R., & Singer, W. (1995).
How precise is neuronal synchronization. Neural Computation,
7, 469–485.
Korhonen, O., Palva, S., & Palva, J. M. (2014). Sparse weightings
for collapsing inverse solutions to cortical parcellations optimize
M/EEG source reconstruction accuracy. Journal of Neuroscience
Methods, 226C, 147–160.
Lachaux, J. P., Rodriguez, E., Martinerie, J., & Varela, F. J. (1999).
Measuring phase synchrony in brain signals. Human Brain Map-
ping, 8, 194–208.
Li Hegner, Y., Lindner, A., & Braun, C. (2015). Cortical correlates
of perceptual decision making during tactile spatial pattern dis-
crimination. Human Brain Mapping, 36, 3339–3350.
Maris, E., Fries, P., & van Ede, F.
(2016). Diverse phase relations
among neuronal rhythms and their potential function. Trends in
Neurosciences, 39, 86–99.
Medendorp, W. P., Buchholz, V. N., Van Der Werf, J., & Leone,
(2011). Parietofrontal circuits in goal-oriented behaviour.
F. T.
European Journal of Neuroscience, 33, 2017–2027.
Monto, S., Palva, S., Voipio, J., & Palva, J. M. (2008). Very slow
EEG fluctuations predict the dynamics of stimulus detection and
oscillation amplitudes in humans. Journal of Neuroscience, 28,
8268–8272.
Murakami, S., & Okada, Y. (2006). Contributions of principal
neocortical neurons to magnetoencephalography and electro-
encephalography signals. Journal of Physiology, 575, 925–936.
Nierhaus, T., Forschack, N., Piper, S. K., Holtze, S., Krause, T.,
Taskin, B., Long, X., Stelzer, J., Margulies, D. S., Steinbrink,
J., & Villringer, A. (2015). Imperceptible somatosensory stimu-
lation alters sensorimotor background rhythm and connectivity.
Journal of Neuroscience, 35, 5917–5925.
Nordmark, P. F., Pruszynski, J. A., & Johansson, R. S. (2012). BOLD
responses to tactile stimuli in visual and auditory cortex depend
on the frequency content of stimulation. Journal of Cognitive
Neuroscience, 24, 2120–2134.
Orlov, T., Makin, T. R., & Zohary, E. (2010). Topographic representa-
tion of the human body in the occipitotemporal cortex. Neuron,
68, 586–600.
Palva, J. M., Palva, S., & Kavila, K. (2005). Phase synchrony among neu-
ronal oscillations in the human cortex. Journal of Neuroscience,
25, 3962–3972.
Palva, J. M., Monto, S., Kulashekhar, S., & Palva, S. (2010). Neu-
ronal synchrony reveals working memory networks and predicts
individual memory capacity. Proceedings of
the National
Academy of Sciences of the United States of America, 107,
7580–7585.
Palva, S., & Palva, J. M. (2012). Discovering oscillatory interaction
networks with M/EEG: Challenges and breakthroughs. Trends in
Cognitive Sciences, 16, 219–230.
Palva, S., Linkenkaer-Hansen, K., Naatanen, R., & Palva, J. M. (2010).
Neuronal synchrony reveals working memory networks and
predicts individual memory capacity. Proceedings of the National
Academy of Sciences of the United States of America, 107,
7580–7585.
Papadelis, C., Arfeller, C., Erla, S., Nollo, G., Cattaneo, L., & Braun,
C. (2016). Inferior frontal gyrus links visual and motor cortices
during a visuomotor precision grip force task. Brain Research,
1650, 252–266.
Petersen, S., & Sporns, O.
(2015). Brain networks and cognitive
architectures. Neuron, 88, 207–219.
Power, J. D., Cohen, A. L., Nelson, S. M., Wig, G. S., Barnes,
K. A., Church, J. A., Vogel, A. C., Laumann, T. O., Miezin, F. M.,
Schlaggar, B. L., Petersen, S. E.
(2011). Functional network
organization of the human brain. Neuron, 72, 665–678.
Rubinov, M., & Sporns, O. (2010). Complex network measures of
brain connectivity: Uses and interpretations. NeuroImage, 52,
1059–1069.
Sadaghiani, S., Hesselmann, G., & Kleinschmidt, A. (2009). Dis-
tributed and antagonistic contributions of ongoing activity fluc-
tuations to auditory stimulus detection. Journal of Neuroscience,
29, 13410–13417.
Salti, M., Monto, S., Charles, L., King, J. R., Parkkonen, L., & Dehaene,
(2015). Distinct cortical codes and temporal dynamics
S.
for conscious and unconscious percepts. ELife, 4, 10.7554/
eLife.05652.
Schall, J. D. (2001). Neural basis of deciding, choosing and acting.
Nature Reviews Neuroscience, 2, 33–42.
Schoffelen, J. M., & Gross, J. (2009). Source connectivity analysis
with MEG and EEG. Human Brain Mapping, 30, 1857–1865.
Schroeder, C. E., & Lakatos, P.
(2009). Low-frequency neuronal
oscillations as instruments of sensory selection. Trends in Neuro-
sciences, 32, 9–18.
Siegel, M., Buschman, T. J., & Miller, E. K. (2015). Cortical informa-
tion flow during flexible sensorimotor decisions. Science, 348,
1352–1355.
Network Neuroscience
462
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
2
4
4
4
2
1
0
9
2
2
4
9
n
e
n
_
a
_
0
0
0
3
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Large-scale synchronization from perception to action
Siegel, M., Donner, T. H., & Engel, A. K. (2012). Spectral finger-
interactions. Nature Reviews
large-scale neuronal
prints of
Neuroscience, 13, 121–134.
Singer, W. (2009). Distributed processing and temporal codes in
neuronal networks. Cognitive Neurodynamics, 3, 189–196.
Singer, W.
(1999). Neuronal synchrony: A versatile code for the
definition of relations? Neuron, 24, 49–65, 111–25.
Spadone, S., Della Penna, S., Sestieri, C., Betti, V., Tosoni, A.,
Perrucci, M. G., Romani, G. L., & Corbetta, M. (2015). Dynamic
reorganization of human resting-state networks during visuo-
spatial attention. Proceedings of the National Academy of Sci-
ences of the United States of America, 112, 8112–8117.
Tallon-Baudry, C.
(2012). On the neural mechanisms subserving
consciousness and attention. Frontiers in Psychology, 2, 397.
Taskin, B., Holtze, S., Krause, T., & Villringer, A. (2008). Inhib-
itory impact of subliminal electrical finger stimulation on SI
representation and perceptual sensitivity of an adjacent finger.
NeuroImage, 39, 1307–1313.
Tauste Campo, A., Martinez-Garcia, M., Nacher, V., Luna, R.,
Romo, R., & Deco, G. (2015). Task-driven intra- and interarea
communications in primate cerebral cortex. Proceedings of the
National Academy of Sciences of the United States of America,
112, 4761–4766.
Tomassini, A., Spinelli, D., Jacono, M., Sandini, G., & Morrone,
M. C. (2015). Rhythmic oscillations of visual contrast sensitivity
synchronized with action. Journal of Neuroscience, 35, 7019–7029.
Tosoni, A., Galati, G., Romani, G. L., & Corbetta, M. (2008).
Sensory-motor mechanisms in human parietal cortex underlie
arbitrary visual decisions. Nature Neuroscience, 11, 1446–1453.
van Gaal, S., Ridderinkhof, K. R., Fahrenfort, J. J., Scholte, H. S.,
& Lamme, V. A. (2008). Frontal cortex mediates unconsciously
Journal of Neuroscience, 28,
triggered inhibitory control.
8053–8062.
Wang, S., Lobier, L., Siebenhühner, F., Palva, S., & Palva, J. M. (2018).
Hyperedge bundling: A practical solution to spurious interac-
tions in MEG/EEG source connectivity analyses. 173, 610–622.
Weisz, N., Wuhle, A., Monittola, G., Demarchi, G., Frey, J., Popov,
T., & Braun, C. (2014). Prestimulus oscillatory power and con-
nectivity patterns predispose conscious somatosensory percep-
tion. Proceedings of the National Academy of Sciences of the
United States of America, 111, 4: E417–E425.
Wohlschlager, A. (2000). Visual motion priming by invisible ac-
tions. Vision Research, 40, 925–930.
Yang, T., & Shadlen, M. N. (2007). Probabilistic reasoning by neu-
rons. Nature, 447, 1075–1080.
Yeo, B. T., Krienen, F. M., Sepulcre, J., Sabuncu, M. R., Lashkari, D.,
Hollinshead, M., Roffman, J. L., Smoller, J. W., Zollei, L., Polimeni,
(2011). The
J. R., Fischl, B., Liu, H., & Buckner, R. L.
organization of the human cerebral cortex estimated by intrin-
sic functional connectivity. Journal of Neurophysiology, 106,
1125–1165.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
2
4
4
4
2
1
0
9
2
2
4
9
n
e
n
_
a
_
0
0
0
3
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Network Neuroscience
463