RESEARCH
Creativity at rest: Exploring functional network
connectivity of creative experts
William Orwig1,2
, Roni Setton1
Diana I. Tamir4
, Ibai Diez2
, Jorge Sepulcre2
, Elisenda Bueichekú2
, Meghan L. Meyer3
,
, and Daniel L. Schacter1
1Department of Psychology, Harvard University, Cambridge, MA, USA
2Gordon Center for Medical Imaging, Department of Radiology, Massachusetts General Hospital and
Harvard Medical School, Boston, MA, USA
3Department of Psychological and Brain Sciences, Dartmouth College, Hanover, NH, USA
4Department of Psychology, Princeton University, Princeton, NJ, USA
a n o p e n a c c e s s
j o u r n a l
Keywords: Creativity, fMRI, Functional connectivity, Distal simulation, Vividness
ABSTRACT
The neuroscience of creativity seeks to disentangle the complex brain processes that underpin
the generation of novel ideas. Neuroimaging studies of functional connectivity, particularly
functional magnetic resonance imaging (fMRI), have revealed individual differences in brain
network organization associated with creative ability; however, much of the extant research is
limited to laboratory-based divergent thinking measures. To overcome these limitations, we
compare functional brain connectivity in a cohort of creative experts (n = 27) and controls
(n = 26) and examine links with creative behavior. First, we replicate prior findings showing
reduced connectivity in visual cortex related to higher creative performance. Second, we
examine whether this result is driven by integrated or segregated connectivity. Third, we
examine associations between functional connectivity and vivid distal simulation separately in
creative experts and controls. In accordance with past work, our results show reduced
connectivity to the primary visual cortex in creative experts at rest. Additionally, we observe a
negative association between distal simulation vividness and connectivity to the lateral visual
cortex in creative experts. Taken together, these results highlight connectivity profiles of highly
creative people and suggest that creative thinking may be related to, though not fully
redundant with, the ability to vividly imagine the future.
AUTHOR SUMMARY
This study sought to uncover the neurocognitive basis of creativity by comparing brain
connectivity in a cohort of creative experts and controls. Using graph theory methods, we
replicate past work showing reduced connectivity to the primary visual cortex in highly creative
people. Additionally, we observed a negative association between vividness of distal future
simulation and connectivity to the lateral visual cortex in creative experts. Taken together, these
findings highlight connectivity profiles of highly creative people and suggest that creative
thinking may be related to the ability to vividly imagine events in the distant future.
INTRODUCTION
The neuroscience of creativity seeks to disentangle the complex brain processes that underpin
the generation of novel ideas. Neuroimaging studies of functional connectivity, particularly
Citation: Orwig, W., Setton, R., Diez, I.,
Bueichekú, E., Meyer, M. L., Tamir, D. I.,
Sepulcre, J., & Schacter, D. L. (2023).
Creativity at rest: Exploring functional
network connectivity of creative
experts. Network Neuroscience, 7(3),
1022–1033. https://doi.org/10.1162/netn
_a_00317
DOI:
https://doi.org/10.1162/netn_a_00317
Supporting Information:
https://doi.org/10.1162/netn_a_00317
Received: 21 October 2022
Accepted: 31 March 2023
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Author:
William Orwig
williamorwig@g.harvard.edu
Handling Editor:
Olaf Sporns
Copyright: © 2023
Massachusetts Institute of Technology
Published under a Creative Commons
Attribution 4.0 International
(CC BY 4.0) license
The MIT Press
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
7
3
1
0
2
2
2
1
5
4
7
7
1
n
e
n
_
a
_
0
0
3
1
7
p
d
/
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Creativity and functional connectivity
Divergent thinking:
The ability to generate novel ideas to
open-ended problems.
functional magnetic resonance imaging (fMRI), have revealed individual differences in brain
network organization associated with creative ability (Beaty et al., 2014; Takeuchi et al.,
2012); however, much of the extant research is limited to laboratory measures of creative per-
formance. Divergent thinking (DT) has been a central concept in creativity research since its
first usage by Guilford (1950). The most common assessment of DT is the alternative uses task
(AUT), involving the generation of novel uses for common objects. Behavioral studies have
shown that DT, as assessed by the AUT, predicts both the quantity of self-reported creative
achievements (Jauk et al., 2014) and the quality of expert-rated creative performance (Beaty
et al., 2013), as well as the academic and creative successes of school-aged children (Plucker,
1999; Torrance, 1988). The predictive power of DT has fueled decades of empirical research
on the neurocognitive basis of creativity (Cogdell-Brooke et al., 2020; Kim, 2008; Runco &
Acar, 2012; Wu et al., 2015). Computerized methods for scoring AUT have led to widely
accessible tools for assessing individual differences in creativity (Beaty & Johnson, 2021). While
DT certainly captures some aspects of creativity, it is not all-encompassing and its psychometric
merit has been the subject of some debate in the field (Forthmann et al., 2021; Zeng et al.,
2011). To overcome these limitations, we focus our analyses on a cohort of notable creative
experts.
Current theories suggest that creativity is not attributable to a single brain region; rather,
novel ideas result from interactions between large-scale brain networks (Beaty et al., 2019).
Among the most well studied of these functional brain networks, the default network (DN)—
including midline and posterior inferior parietal regions—shows increased activation in the
absence of an external stimulus (Raichle et al., 2001). DN activity has been linked with
self-generated thought, such as mind wandering and imagination (Andrews-Hanna et al.,
2014; Buckner et al., 2008). Both task-based and resting-state fMRI studies have contributed
to current understanding of the neurocognitive basis of creativity. Task-based approaches have
helped to uncover the relationship among networks during creative task performance (Beaty
et al., 2018; Shi et al., 2018). It has been proposed that DN contributes to the generation of
candidate ideas, while executive control networks exert top-down monitoring to meet specific
task goals or constraints (Beaty et al., 2014, 2016). Resting-state functional connectivity is
thought to reflect experience-dependent patterns of connectivity that relate to individual
and group differences in behavior (Stevens & Spreng, 2014). Research on resting-state network
organization in relation to creative cognition has investigated interindividual differences, find-
ing that increased “hubness” (the presence of hubs that form connections between different
communities) of DN regions was positively associated with creativity (Kenett et al., 2020).
Additionally, a recent study from our group has identified a pattern of reduced resting-state
connectivity to the visual cortex associated with increased DT (Orwig et al., 2021). One inter-
pretation of these results is that highly creative people are more prone to engage in internally
directed cognition in the absence of an external task. A primary aim of the present study was to
replicate this finding of reduced connectivity to visual cortex in a cohort of creative experts.
While most participants in creativity research are drawn from the general population, some
recent efforts involve the collection of neuroimaging data from creative professionals. The
Big-C Project is a large-scale study seeking to identify behavioral and functional brain charac-
teristics that distinguish exceptionally creative people. For instance, Anderson et al. (2022)
found that highly creative individuals had higher local clustering coefficients during resting
state and reduced local clustering while performing the AUT compared with less creative indi-
viduals. Furthermore, findings from this dataset indicate less activation in bilateral occipital
cortex during a DT task in highly creative versus less creative individuals (Japardi et al.,
2018). Related work by Chrysikou et al. (2020) described patterns of neural activation between
Network Neuroscience
1023
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
7
3
1
0
2
2
2
1
5
4
7
7
1
n
e
n
_
a
_
0
0
3
1
7
p
d
.
t
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Creativity and functional connectivity
Vividness:
Degree of detail present in mental
imagery.
Distal simulation:
Imagination of events in the distant
future.
eminent and non-eminent thinkers; despite showing no behavioral differences, eminent
thinkers engaged more temporoparietal and less occipital areas than did controls during a
creative generation task. Additionally, findings from Lotze et al. (2014) show reduced inter-
hemispheric connectivity in highly verbally creative individuals compared with less verbally
creative individuals.
A second aim is to link creative experts’ functional connectivity to a uniquely creative
behavior other than DT: vividness of imagination. Imagination refers to the ability to mentally
construct and manipulate images that are not directly present to the senses. Imagination is
closely associated with creativity, drawing upon semantic (Abraham & Bubic, 2015) and epi-
sodic memory processes (Devitt et al., 2017). Numerous studies conducted in the context of
research on episodic memory and future thinking have described the striking cognitive and
neural similarities between remembering past experiences and imagining future or other hypo-
thetical experiences (Schacter et al., 2012, 2017). Emerging behavioral and neuroimaging
research point to a conjunction between episodic retrieval, future imagination, and divergent
creative thinking by identifying common engagement of the hippocampus and default network
regions (Beaty et al., 2018; Madore et al., 2019; Thakral et al., 2020). Additionally, DT has been
linked with the ability to vividly imagine novel and specific future events (Addis et al., 2016;
Thakral et al., 2021). Behavioral studies have provided evidence for an association between
vividness of mental imagery and divergent thinking (Forisha, 1978; Gonzalez et al., 1997).
Together, these studies highlight the role of constructive episodic processes in creative thinking.
Meyer et al. (2019) examined whether creative experts might be distinguished in their abil-
ity to vividly imagine the future. Across two behavioral studies and one neuroimaging study,
the authors compared the vividness of proximal (i.e., considering what you might do tomor-
row) versus distal (i.e., considering what life might be like next century) simulation in creative
experts and age-matched controls. In both groups, vividness of proximal simulation involved
increased medial prefrontal cortex activity. By contrast, creative experts reported having more
vivid distal simulations than did controls, and they showed increased activity of the dorsal
medial subnetwork of the DN compared with controls while doing so. The dorsal medial sub-
network includes dorsal medial prefrontal cortex, the temporoparietal junction, along with
swaths of inferior frontal and lateral temporal cortex as key structures (Andrews-Hanna
et al., 2010) and has been linked to several processes that call on high levels of abstraction,
such as semantic processing and mentalizing. Meyer et al. (2019) therefore concluded that an
ability to construe abstract thoughts may link creativity and distal simulation. Creative experts
in the Meyer et al. (2019) study showed higher connectivity within the dorsal medial DN at
rest compared with controls; however, this atlas-based approach is limited, with only the DN
tested. This work leaves open the possibility to study whole-brain, voxel-level networks to
capture salient features of creative experts’ brains.
Building upon our recent work, we revisit the resting-state fMRI data from Meyer et al.
(2019), implementing a high-resolution graph theory approach to more precisely characterize
global network features of creative thinkers. As with previous analyses (Orwig et al., 2021), a
voxel-level cortical hub strategy was used to identify connectivity differences between creative
experts and controls without introducing any regional priors. We hypothesize that creative
experts will display brain centrality changes in visual areas relative to controls. As creatives
showed task-evoked activity differences specific to distal simulation, we also test whether
resting-state connectivity may differentially relate to vividness of distal simulation in creatives
versus controls. Linking this finding to higher vividness of distal simulations would serve as
evidence that highly creative people may more readily silo visual network connectivity and
highlight differential patterns of connectivity between creative experts and controls.
Network Neuroscience
1024
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
7
3
1
0
2
2
2
1
5
4
7
7
1
n
e
n
_
a
_
0
0
3
1
7
p
d
.
/
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Creativity and functional connectivity
METHODS
Sample
The present research uses behavioral and neuroimaging data previously published in Meyer
et al. (2019). We analyze resting-state fMRI data from 53 participants (27 creative experts, 26
controls). Creative experts are defined as any individual who either has been recognized by an
award for their creative work, held a position at a prestigious institution known for excellence
in a creative domain, or attained commercial success in a creative domain. Specifically,
writers, actors, and directors were targeted to capture a range of creative expertise, while still
ensuring that creative experts had experience imagining distant times, places, and perspec-
tives. Past research showed that writers and actors/directors demonstrated superior distal sim-
ulation skills relative to visual artists; furthermore, creative experts significantly outperformed
controls on divergent thinking measures, confirming that “real-world” experts are in fact more
creative than controls (Meyer et al., 2019). The sample also includes a control group, targeting
professionals working in the legal, medical, and financial industries, based on past work that
identified individuals in these professions as scoring in the mid-to-low range on standardized
creativity assessments (Beketayev & Runco, 2016). Groups did not vary in age (mean creative
experts = 36.08 years, SD = 9.85 years; mean controls = 33.73 years, SD = 7.32 years, t(50) =
0.98, p = 0.33) or gender (χ2(1) = 0.03, p = 0.88, w = 0.001). Participants provided informed
consent in accordance with the Princeton University Institutional Review Board (IRB). All
materials and data are available on Open Science Framework (https://osf.io/cy8wt/).
Distal Simulation Task
Prior to scanning, a subset of participants (n = 45; 23 creative experts, 22 controls) completed
a distal simulation task, in which they were shown a simulation prompt in the temporal
domain (e.g., imagine what the world will be like in 500 years). Participants were shown
the prompt for two minutes and instructed to imagine the experience and write a description
of their simulation. To derive a subjective measure of vividness, participants then rated the
quality of their distal simulations in response to four questions: (i) How vividly did you imagine
the experience? (ii) To what extent did you see what you imagined in your mind’s eye? (iii) To
what extent did you feel immersed in what you imagined? and (iv) How difficult was it for you
to imagine the experience (reverse scored). Participants responded using a 1 (not at all) to 100
(extremely) sliding scale. A composite score of distal simulation vividness was computed as
the average response value across these questions.
MRI Acquisition and Preprocessing
Scanning was conducted at the Princeton Neuroscience Institute on a 3T Siemens Prisma MRI
system with a 64-channel head coil. High-resolution T1 scans (MP-RAGE; TR/TE = 2,300/2.27,
flip angle = 8°, 256 × 256 matrix, 1 mm thick, 25 0mm FoV) were acquired for anatomical
normalization. Resting-state functional scans were acquired with a T2*-weighted echo-planar
plus sequence with 69 interleaved slices (TR/TE = 1,500/27ms, flip angle = 75°, 96 × 48
matrix, 2 mm thick, 192 mm FoV; multiband acceleration factor = 3).
MRI data for both anatomical and functional images were preprocessed using FMRIB Soft-
ware Library v5.0.7 (FSL) and MATLAB 2017a (MathWorks Inc., Natick, MA). The anatomical
and functional preprocessing pipelines were adapted from previous work (Diez et al., 2019).
The anatomical T1 preprocessing included the following: reorientation to right-posterior-
inferior (RPI) with fslreorient2std; alignment to anterior and posterior commissures with a cus-
tom script; skull stripping using MNI brain mask template projected to individual space; gray
Network Neuroscience
1025
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
7
3
1
0
2
2
2
1
5
4
7
7
1
n
e
n
_
a
_
0
0
3
1
7
p
d
.
/
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Creativity and functional connectivity
matter, white matter, and cerebrospinal fluid segmentation using FMRIB’s Automated Segmen-
tation Tool (FAST) (Zhang et al., 2001); and computation of nonlinear transformation between
individual skull-stripped T1 and 2-mm resolution MNI152 template images using the FSL fnirt
tool. The functional MRI preprocessing pipeline included the following: slice time correction
using Slicetimer; reorientation to RPI with fslreorient2std; realigning functional volumes within
runs with rigid body transformations (six-parameter linear transformation); computation of the
transformation between individual skull-stripped T1 and mean functional images using
boundary-based registration with flirt; intensity normalization; and removal of confounding
factors from the data using linear regression, including 12 motion-related covariates (rigid
motion parameters and its derivatives), linear and quadratic terms, and five components each
from the lateral ventricles and white matter. Global signal regression was not applied owing to
the spurious correlations this approach can introduce (Murphy et al., 2009). Transformation of
resting-state data to MNI space was performed applying the resulting transformation of
concatenating the transformation from functional to structural and from structural to MNI. Spa-
tial smoothing with an isotropic Gaussian kernel of 6-mm FWHM and band-pass filtering
(0.01–0.08 Hz) to reduce low-frequency drift and high-frequency noise were also applied.
Using in-house MATLAB scripts, head motion was quantified using realignment parameters
obtained during image preprocessing, including three translation and three rotation estimates.
Scrubbing of time points with excess head motion and interpolation of all time points with
Jenkinson framewise displacement greater than 0.2 mm was applied. No participants demon-
strated excessive head motion; thus, none were removed from the study based on these
criteria. The distributions of the correlations across time series were reviewed for possible con-
tamination. No outliers were observed.
Weighted Degree Functional Connectivity Analysis
Functional connectivity was computed at the individual level using whole-brain voxel-level
weighted degree ( WD) analysis. WD is a measure of centrality, computed as the sum of the
strengths of functional connections between each voxel and the rest of the brain (Bullmore &
Sporns, 2009). This centrality measure captures the global features of the networks, which are
thought to be relevant in exceptionally creative people (Anderson et al., 2022). WD is one of
many possible measures to capture centrality in functional connectivity. Building upon recent
findings of WD connectivity associated with individual differences in creativity (Orwig et al.,
2021), we applied the same analysis in this context to detect cortical hubs associated with
creative expertise. Pearson correlation coefficients were used to calculate the connectivity
matrices of each participant using the time series of all cortical gray matter voxels. An r-to-z
Fisher transformation was applied to the resulting correlation matrix, and negative values were
removed because of their controversial interpretation (Qian et al., 2018). To minimize noise,
we considered only the most significant links using a false discovery rate (FDR) at q-level less
than 0.005 (Benjamini & Hochberg, 1995). After obtaining a high-resolution 39,080 × 39,080
connectivity matrix for each participant, we summed all the weighted connections of each
voxel to generate the WD adjacency matrix. This adjacency matrix was transformed into a
brain map and projected on cortical surfaces showing the extent to which each voxel is func-
tionally connected to the rest of the brain.
Integration and Segregation
Research into the functional composition of network assembly has revealed a modular
organization—networks are composed of densely connected modules, or communities, asso-
ciated with specific cognitive functions, and cortical hubs that integrate information across
Weighted degree:
Graph theory measure that quantifies
the relative prominence of each node
within the whole network
architecture.
Network Neuroscience
1026
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
7
3
1
0
2
2
2
1
5
4
7
7
1
n
e
n
_
a
_
0
0
3
1
7
p
d
t
.
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Creativity and functional connectivity
communities (Bassett & Bullmore, 2006; Bullmore & Sporns, 2009; Diez et al., 2015).
Measures of segregation quantify the presence of communities within the overall brain net-
work, whereas integration captures the brain’s ability to rapidly combine information between
communities (Rubinov & Sporns, 2010). To examine group differences in segregation and inte-
gration, we organized gray matter voxels with the 17-network solution from Yeo et al. (2011). If
the link’s start and end voxels belonged to the same resting-state network, the link was then
classified as a segregated link; otherwise, the link was classified as an integrated link. For each
participant, we computed the WD of segregated and integrated links separately obtaining two
connectivity maps. Higher values in segregated connectivity maps indicate that a given voxel
has a higher number of strong functionally connected links to other voxels within the same
functional network, whereas higher values in integrated functional connectivity maps indicate
that the voxel is an important hub for integrating information between networks.
Statistical Analysis
At the behavioral level, we conducted univariate linear regression to test for group differ-
ences in distal simulation vividness between creative experts and controls. We report the
regression parameters (t and p statistics) for this analysis, with a significance threshold of
α = 0.05. At the neuroimaging level, general linear models were used to compute the group
difference between creative experts and controls for whole-brain WD maps, segregation
maps, and integration maps. Additionally, general linear models were used to compute the
association between WD and distal simulation vividness within creative experts and control
groups. Whole-brain correction for multiple comparisons was computed using Monte Carlo
simulation with 10,000 iterations to estimate the probability of false positive clusters with a
two-tailed p value less than 0.05 (3dClustSim, afni.nimh.nih.gov). First, we generated the
residual of the statistics and used it to estimate the spatial autocorrelation present in our data.
Then, we computed null models by generating 10,000 random maps with the estimated spa-
tial autocorrelation (3dClustSim). These null models were used to evaluate the likelihood of
getting a brain cluster of a particular size for a p value less than 0.05 by chance. These data
were then used to remove all clusters smaller than the estimated size. Cortical surfaces were
visualized using the population-average landmark and surface-based projections of CARET
software ( Van Essen, 2005). Surface images were displayed using a color scale based on
T-scores.
RESULTS
Connectivity Patterns Associated With Creative Expertise
We performed whole-brain WD analysis to identify group-level connectivity differences
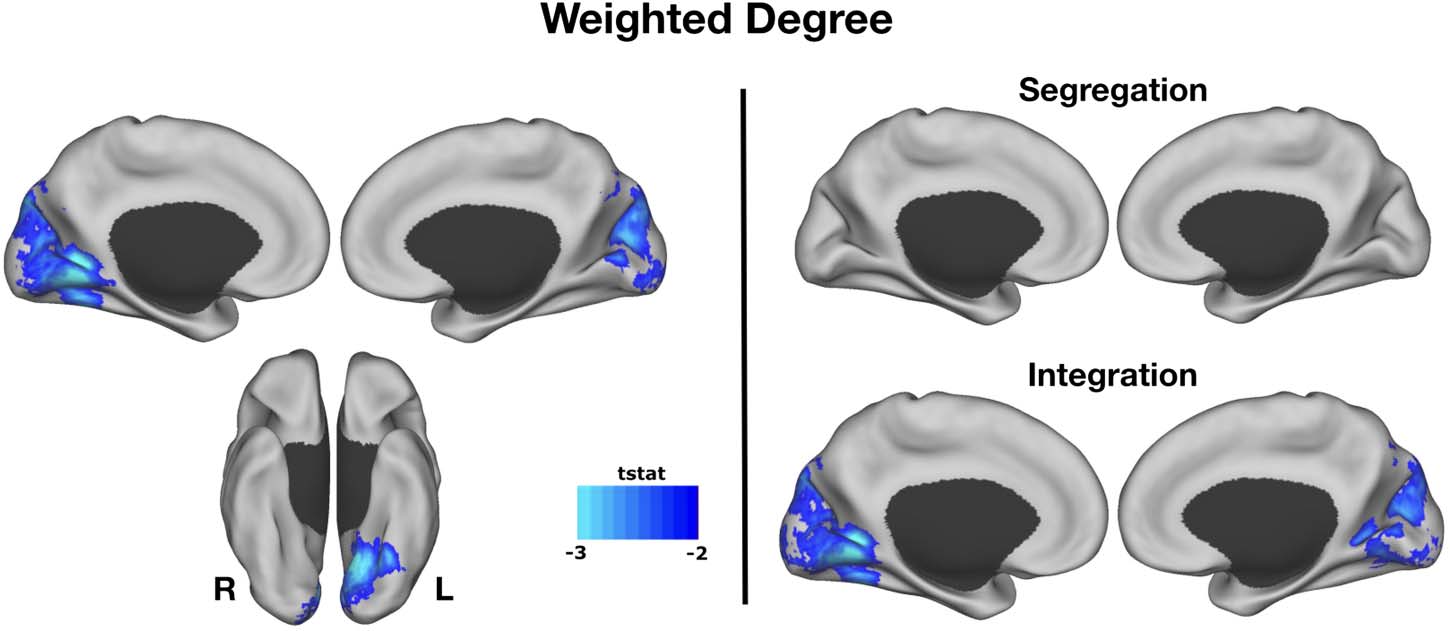
between creative experts and controls. Creative experts showed lower WD connectivity in
the medial visual cortex compared with controls (Figure 1). Next, we examined whether seg-
regation or integration of networks was driving this effect. Segregation maps did not differ
between creative experts and controls; however, integration maps reveal a negative associa-
tion between connectivity of medial visual cortex and creative expertise. In sum, these results
indicate that, relative to controls, creative experts have reduced connectivity between visual
cortex and the rest of the brain during resting state.
Connectivity Patterns Associated With Distal Simulation Vividness
Behavioral analysis revealed no significant differences between creative experts and controls
in relation to vividness of distal simulation (t = 1.09, p = 0.28). Given that Meyer et al.
Network Neuroscience
1027
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
7
3
1
0
2
2
2
1
5
4
7
7
1
n
e
n
_
a
_
0
0
3
1
7
p
d
/
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Creativity and functional connectivity
Figure 1. Weighted degree. WD connectivity across the medial visual cortex was lower in creative expertise compared with controls.
Segregation maps did not differ between creative experts and controls; however, integration maps reveal a negative association between
connectivity of medial visual cortex and creative expertise.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
(2019) report significant differences in a larger behavioral sample, it is likely that the present
sample lacked sufficient statistical power to detect these behavioral differences. To examine
the relationship between functional connectivity and distal simulation vividness, we per-
formed two independent linear regression analyses within groups of creative experts and
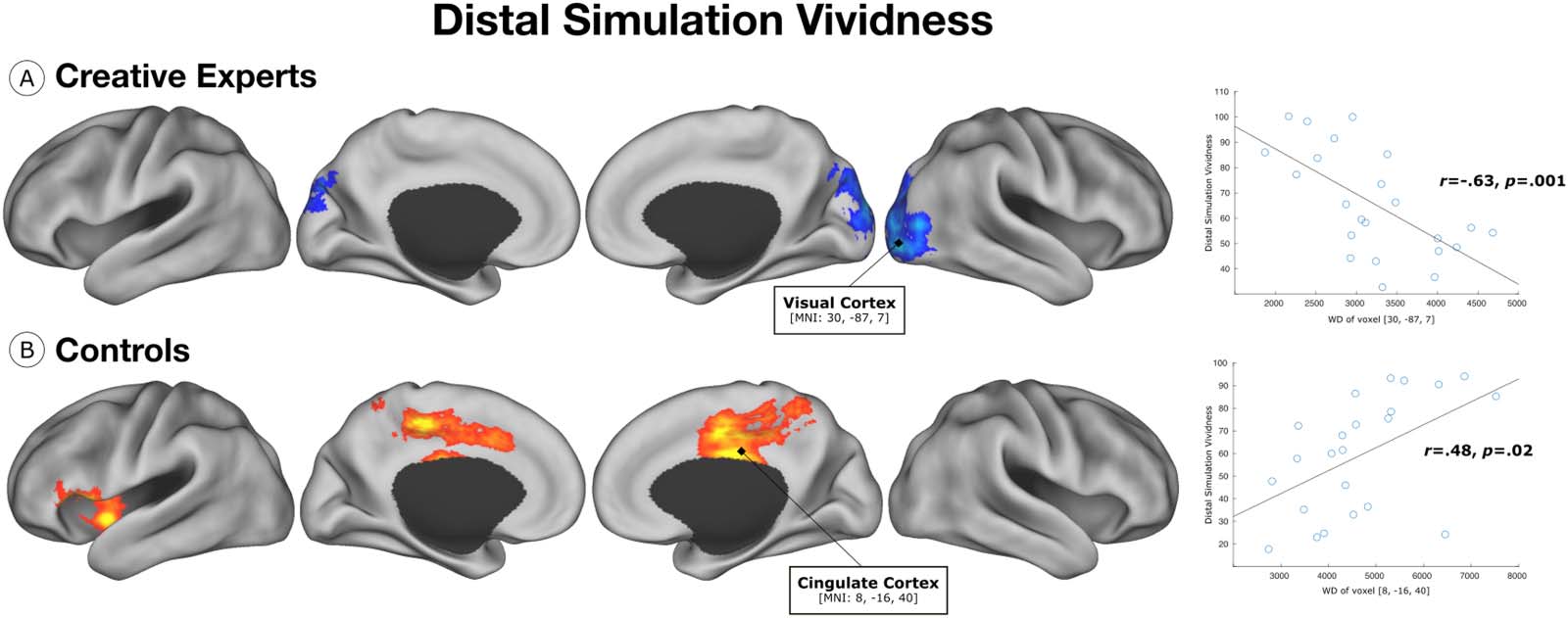
controls. Results indicate a negative association between WD in the lateral visual cortex
and distal simulation vividness in creative experts (Figure 2A). Conversely, we find a positive
association between WD of voxels in the cingulate cortex and left anterior insula and distal
simulation vividness in controls (Figure 2B). These findings reveal differential patterns of
connectivity, between creative experts and controls, associated with vividness of distal
simulation.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
7
3
1
0
2
2
2
1
5
4
7
7
1
n
e
n
_
a
_
0
0
3
1
7
p
d
/
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 2. Distal simulation vividness. (A) Within the group of creative experts, WD of voxels in the lateral visual cortex were negatively
associated with distal simulation vividness. (B) Within the control group, WD of voxels in the cingulate cortex and left anterior insula were
positively associated with distal simulation vividness.
Network Neuroscience
1028
Creativity and functional connectivity
Perceptual decoupling:
The capacity to disengage attention
from perception.
DISCUSSION
This study aimed to characterize WD connectivity associated with creativity. Our findings
indicate that creative experts show reduced WD connectivity of primary visual areas to the
rest of the brain at rest compared with controls. This finding extends past work describing indi-
vidual differences in continuous creative performance within the general population to a
group difference present in a highly expert sample of creatives (Orwig et al., 2021). Moreover,
we found that reduced WD connectivity to lateral visual cortex was associated with more vivid
distal simulations in creative experts. Taken together, these findings resemble a similar pattern
of connectivity associated with creative expertise and distal imagination.
One interpretation of these findings is that in the absence of an external task—namely, dur-
ing resting state—highly creative people are more prone to engage in internally directed cog-
nition, tuning out sensory information from the present environment. Perceptual decoupling,
first described by Schooler et al. (2011), refers to the capacity to disengage attention from per-
ception during mind wandering. Previous studies of creative cognition have found extended
deactivation in occipital cortex associated with internal attention (Benedek et al., 2016). The
present findings highlight patterns of reduced WD connectivity to visual cortex in highly cre-
ative people. It may be the case that creative experts are engaged in more internally directed
cognitive processes during resting state and this manifests in reduced connectivity to the pri-
mary visual areas, compared with controls. An alternative explanation of these results is that
involvement in creative pursuits somehow alters the functional organization of resting-state
networks, resulting in the observed reduction in connectivity. It is a well-established phenom-
enon that repeated exposure to the same stimulus results in diminished response in the human
visual cortex. Repetition suppression, the relative attenuation of neural signal evoked by
repeated presentation of a stimulus, has been investigated extensively using fMRI (Fritsche
et al., 2020; Kourtzi & Kanwisher, 2001; Larsson & Smith, 2012). In the context of the present
findings, perhaps the observed negative association between creative expertise and visual cor-
tex connectivity is a consequence of training in a creative domain. According to this view,
years of experience with writing, acting, or directing may lead to functional changes in the
resting-state network organization. These explanations need not be mutually exclusive: It is
possible that creative experts engage in more internally driven thought at rest and, in doing
so, modify connectivity of visual cortex. Future work aimed at disentangling the two—either
by probing thought content at rest or by introducing a novel visual paradigm to examine rates
of repetition suppression—will be fruitful in understanding why this functional property is
unique to creatives.
Despite the long-standing and intuitive connection between imagination and creative
thought, the precise roles of imagery and distal simulation in the creative process remains
an open question. While some studies have found a correlation between subjective imagery
vividness and creativity (Campos & Pérez, 1989; Shaw & Belmore, 1982), others have failed to
observe a reliable association (LeBoutillier & Marks, 2003). Vividness ratings did not differ
between the creative experts and controls on which we report here; however, Meyer et al.
(2019) did report a difference in their larger behavioral sample. It is likely, then, that the pres-
ent sample lacked sufficient statistical power. Using this sample, Meyer et al. (2019) reported
that creative experts engage regions of the dorsal medial default network during distal simu-
lation more than controls and show higher connectivity within this subnetwork at rest. While
Meyer et al. (2019) only tested for group differences within the default network, their results
suggested altered brain activity subserving distal simulation across groups that could be
detected within functional connectivity patterns at rest. One could argue that failure to observe
Network Neuroscience
1029
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
7
3
1
0
2
2
2
1
5
4
7
7
1
n
e
n
_
a
_
0
0
3
1
7
p
d
t
.
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Creativity and functional connectivity
a behavioral difference in this subsample undermines the logic of comparing functional con-
nectivity between creative experts and controls. It is possible, however, that separable neural
resources result in comparable behavior. A prime example of this can be seen in the healthy
aging literature, where high-performing older adults recruit additional brain resources to reach
comparable performance as young adults on working memory tasks (Cabeza et al., 2002,
2004; Park & Reuter-Lorenz, 2009; Reuter-Lorenz et al., 2000). For this reason, it is essential
to look at the correlations within the entire sample as well as separately within each group.
Although we acknowledge that the absence of a behavioral difference in our subsample limits
to some extent the conclusions that we can draw, by describing associations with distal sim-
ulation vividness, within groups of creatives and controls, we provide novel insights into the
relationship between imagination and creative expertise.
We therefore examined whether vividness would differentially associate with resting-state
functional connectivity across groups. We find that vividness for distal future simulation was
negatively associated with connectivity to lateral occipital cortex, but only in creative experts.
Numerous studies have reported that visual imagery evokes activation in early visual cortex,
though others fail to observe this effect (Kosslyn & Thompson, 2003). Here we find that a dis-
tinguishing characteristic of the creative brain, namely lower WD connectivity of visual cortex,
may support more vivid distal simulation. This is in contrast to healthy controls, wherein
higher vividness ratings were associated with greater WD connectivity between regions of
the ventral attention network and the rest of the brain. We focused our analysis exclusively
on distal simulation, rather than proximal simulation vividness (which was collected with dif-
ferent procedures, while participants were in the scanner) because past work indicates that
imagination of distal future events is directly relevant to creativity (Meyer et al., 2019).
Although these findings should be replicated in a larger sample, they suggest diverging brain
patterns that may be readily recruited for imagination of the far future. If it is the case that
creative people more vividly imagine events in the distant future, it could be speculated that,
in order to do so, they must attenuate sensory input from their immediate surroundings.
The present research uses resting-state fMRI data to describe WD connectivity of the brain at
rest. It should be noted that there is a considerable amount of noise inherent in working with
resting-state data, given that we are not able to control for participant mood or state of mind dur-
ing scanning. With these limitations in mind, we utilize graph theory metrics in weighted net-
works, which have been shown to have robust within-subject reproducibility (Ran et al.,
2020). Resting-state functional connectivity is thought to reflect the repeated use of circuits during
different tasks and has strong correspondence to task-based connectivity (Stevens & Spreng,
2014). It may be the case that differences in connectivity at rest speak to group differences in
how creatives and controls engage in simulation tasks, which may in turn be reflected in
resting-state architectures. While future studies may seek to replicate these findings in larger sam-
ples of creative experts, this data-driven approach offers initial evidence that connectivity differ-
ences at rest may reflect brain-related changes associated with creative expertise. Additionally,
assessment of more laboratory measures (e.g., trait-level mind wandering, openness to experi-
ence) in future samples of creative experts could advance understanding of the behavioral cor-
relates of creativity. Further analysis of the complex interplay between imagination, vividness,
and visual cortex promises to offer new insights into the neurocognitive basis of creative thinking.
AUTHOR CONTRIBUTIONS
William Orwig: Conceptualization; Formal analysis; Investigation; Project administration;
Visualization; Writing – original draft; Writing – review & editing. Roni Setton: Methodology;
Network Neuroscience
1030
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
7
3
1
0
2
2
2
1
5
4
7
7
1
n
e
n
_
a
_
0
0
3
1
7
p
d
/
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Creativity and functional connectivity
Writing – review & editing. Ibai Diez: Formal analysis; Methodology; Writing – review &
editing. Elisenda Bueichekú: Writing – review & editing. Meghan L. Meyer: Data curation.
Diana I. Tamir: Data curation; Funding acquisition. Jorge Sepulcre: Funding acquisition; Meth-
odology; Supervision; Writing – review & editing. Daniel L. Schacter: Funding acquisition;
Supervision; Writing – review & editing.
FUNDING INFORMATION
Jorge Sepulcre, National Institutes of Health, Award ID: R01AG061811. Jorge Sepulcre,
National Institutes of Health, Award ID: R01AG061445. Daniel L. Schacter, National Institute
on Aging (https://dx.doi.org/10.13039/100000049), Award ID: AG008441. Diana I. Tamir,
John Templeton Foundation (https://dx.doi.org/10.13039/100000925).
REFERENCES
Abraham, A., & Bubic, A. (2015). Semantic memory as the root of
imagination. Frontiers in Psychology, 6, 325. https://doi.org/10
.3389/fpsyg.2015.00325, PubMed: 25852626
Addis, D. R., Pan, L., Musicaro, R., & Schacter, D. L. (2016). Diver-
gent thinking and constructing episodic simulations. Memory,
24(1), 89–97. https://doi.org/10.1080/09658211.2014.985591,
PubMed: 25483132
Anderson, A., Japardi, K., Knudsen, K. S., Bookheimer, S. Y.,
Ghahremani, D. G., & Bilder, R. M. (2022). Big-C creativity in
artists and scientists is associated with more random global but
less random local fMRI functional connectivity. Psychology of
Aesthetics, Creativity, and the Arts. Advance online publication.
https://doi.org/10.1037/aca0000463
Andrews-Hanna, J. R., Reidler, J. S., Sepulcre, J., Poulin, R., &
Buckner, R. L. (2010). Functional-anatomic fractionation of the
brain’s default network. Neuron, 65(4), 550–562. https://doi.org
/10.1016/j.neuron.2010.02.005, PubMed: 20188659
Andrews-Hanna, J. R., Smallwood, J., & Spreng, R. N. (2014). The
default network and self-generated thought: Component
processes, dynamic control, and clinical relevance. Annals of
the New York Academy of Sciences, 1316(1), 29–52. https://doi
.org/10.1111/nyas.12360, PubMed: 24502540
Bassett, D. S., & Bullmore, E. (2006). Small-world brain networks.
The Neuroscientist, 12(6), 512–523. https://doi.org/10.1177
/1073858406293182, PubMed: 17079517
Beaty, R. E., Benedek, M., Silvia, P. J., & Schacter, D. L. (2016). Cre-
ative cognition and brain network dynamics. Trends in Cognitive
Sciences, 20(2), 87–95. https://doi.org/10.1016/j.tics.2015.10
.004, PubMed: 26553223
Beaty, R. E., Benedek, M., Wilkins, R. W., Jauk, E., Fink, A., Silvia,
P. J., Hodges, D. A., Koschutnig, K., & Neubauer, A. C. (2014).
Creativity and the default network: A functional connectivity
analysis of the creative brain at rest. Neuropsychologia, 64,
92–98. https://doi.org/10.1016/j.neuropsychologia.2014.09.019,
PubMed: 25245940
Beaty, R. E., & Johnson, D. R. (2021). Automating creativity assess-
ment with SemDis: An open platform for computing semantic
distance. Behavior Research Methods, 53(2), 757–780. https://
doi.org/10.3758/s13428-020-01453-w, PubMed: 32869137
Beaty, R. E., Seli, P., & Schacter, D. L. (2019). Network neurosci-
ence of creative cognition: Mapping cognitive mechanisms and
individual differences in the creative brain. Current Opinion in
Behavioral Sciences, 27, 22–30. https://doi.org/10.1016/j
.cobeha.2018.08.013, PubMed: 30906824
Beaty, R. E., Smeekens, B. A., Silvia, P. J., Hodges, D. A., & Kane,
M. J. (2013). A first look at the role of domain-general cognitive
and creative abilities in jazz improvisation. Psychomusicology:
Music, Mind, and Brain, 23(4), 262–268. https://doi.org/10
.1037/a0034968
Beaty, R. E., Thakral, P. P., Madore, K. P., Benedek, M., & Schacter,
D. L. (2018). Core network contributions to remembering the
past, imagining the future, and thinking creatively. Journal of
Cognitive Neuroscience, 30(12), 1939–1951. https://doi.org/10
.1162/jocn_a_01327, PubMed: 30125219
Beketayev, K., & Runco, M. A. (2016). Scoring divergent thinking
tests by computer with a semantics-based algorithm. Europe’s
Journal of Psychology, 12(2), 210–220. https://doi.org/10.5964
/ejop.v12i2.1127, PubMed: 27298632
Benedek, M., Jauk, E., Beaty, R. E., Fink, A., Koschutnig, K., &
Neubauer, A. C. (2016). Brain mechanisms associated with inter-
nally directed attention and self-generated thought. Scientific
Reports, 6(1), 22959. https://doi.org/10.1038/srep22959,
PubMed: 26960259
Benjamini, Y., & Hochberg, Y. (1995). Controlling the false discov-
ery rate: A practical and powerful approach to multiple testing.
Journal of the Royal Statistical Society: Series B (Methodological),
57(1), 289–300. https://doi.org/10.1111/j.2517-6161.1995
.tb02031.x
Buckner, R. L., Andrews-Hanna, J. R., & Schacter, D. L. (2008). The
brain’s default network: Anatomy, function, and relevance to dis-
ease. Annals of the New York Academy of Sciences, 1124, 1–38.
https://doi.org/10.1196/annals.1440.011, PubMed: 18400922
Bullmore, E., & Sporns, O. (2009). Complex brain networks: Graph
theoretical analysis of structural and functional systems. Nature
Reviews Neuroscience, 10(3), 186–198. https://doi.org/10.1038
/nrn2575, PubMed: 19190637
Cabeza, R., Anderson, N. D., Locantore, J. K., & McIntosh, A. R.
(2002). Aging gracefully: Compensatory brain activity in
Network Neuroscience
1031
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
7
3
1
0
2
2
2
1
5
4
7
7
1
n
e
n
_
a
_
0
0
3
1
7
p
d
t
/
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Creativity and functional connectivity
high-performing older adults. NeuroImage, 17(3), 1394–1402.
https://doi.org/10.1006/nimg.2002.1280, PubMed: 12414279
Cabeza, R., Daselaar, S. M., Dolcos, F., Prince, S. E., Budde, M., &
Nyberg, L. (2004). Task-independent and task-specific age effects
on brain activity during working memory, visual attention and
episodic retrieval. Cerebral Cortex, 14(4), 364–375. https://doi
.org/10.1093/cercor/bhg133, PubMed: 15028641
Campos, A., & Pérez, M. J. (1989). High and low imagers and their
scores on creativity. Perceptual and Motor Skills, 68(2), 403–406.
https://doi.org/10.2466/pms.1989.68.2.403
Chrysikou, E. G., Jacial, C., Yaden, D. B., van Dam, W., Kaufman,
S. B., Conklin, C. J., Wintering, N. A., Abraham, R. E., Jung, R. E.,
& Newberg, A. B. (2020). Differences in brain activity patterns
during creative idea generation between eminent and
non-eminent thinkers. NeuroImage, 220, 117011. https://doi
.org/10.1016/j.neuroimage.2020.117011, PubMed: 32504814
Cogdell-Brooke, L. S., Sowden, P. T., Violante, I. R., & Thompson,
H. E. (2020). A meta-analysis of functional magnetic resonance
imaging studies of divergent thinking using activation likelihood
estimation. Human Brain Mapping, 41(17), 5057–5077. https://
doi.org/10.1002/hbm.25170, PubMed: 32845058
Devitt, A. L., Addis, D. R., & Schacter, D. L. (2017). Episodic and
semantic content of memory and imagination: A multilevel anal-
ysis. Memory and Cognition, 45(7), 1078–1094. https://doi.org
/10.3758/s13421-017-0716-1, PubMed: 28547677
Diez, I., Bonifazi, P., Escudero, I., Mateos, B., Muñoz, M. A.,
Stramaglia, S., & Cortes, J. M. (2015). A novel brain partition
highlights the modular skeleton shared by structure and function.
Scientific Reports, 5, 10532. https://doi.org/10.1038/srep10532,
PubMed: 26037235
Diez, I., Ortiz-Terán, L., Williams, B., Jalilianhasanpour, R., Ospina,
J. P., Dickerson, B. C., Keshavan, M. S., LaFrance, W. C., Jr.,
Sepulcre, J., & Perez, D. L. (2019). Corticolimbic fast-tracking:
Enhanced multimodal integration in functional neurological dis-
order. Journal of Neurology, Neurosurgery, and Psychiatry, 90(8),
929–938. https://doi.org/10.1136/jnnp-2018-319657, PubMed:
30850473
Forisha, B. L. (1978). Mental imagery and creativity: Review and
speculations. Journal of Mental Imagery, 2(2), 209–238.
Forthmann, B., Jankowska, D. M., & Karwowski, M. (2021). How
reliable and valid are frequency-based originality scores? Evi-
dence from a sample of children and adolescents. Thinking Skills
and Creativity, 41, 100851. https://doi.org/10.1016/j.tsc.2021
.100851
Fritsche, M., Lawrence, S., & de Lange, F. P. (2020). Temporal
tuning of repetition suppression across the visual cortex. Journal
of Neurophysiology, 123(1), 224–233. https://doi.org/10.1152/jn
.00582.2019, PubMed: 31774368
Gonzalez, M. A., Campos, A., & Perez, M. J. (1997). Mental
imagery and creative thinking. Journal of Psychology, 131(4),
357–364. https://doi.org/10.1080/00223989709603521
Guilford, J. P. (1950). Creativity. American Psychologist, 5(9),
444–454. https://doi.org/10.1037/h0063487, PubMed: 14771441
Japardi, K., Bookheimer, S., Knudsen, K., Ghahremani, D. G., &
Bilder, R. M. (2018). Functional magnetic resonance imaging of
divergent and convergent thinking in Big-C creativity. Neuropsy-
chologia, 118, 59–67. https://doi.org/10.1016/j.neuropsychologia
.2018.02.017, PubMed: 29454896
Jauk, E., Benedek, M., & Neubauer, A. C. (2014). The road to cre-
ative achievement: A latent variable model of ability and person-
ality predictors. European Journal of Personality, 28(1), 95–105.
https://doi.org/10.1002/per.1941, PubMed: 24532953
Kenett, Y. N., Betzel, R. F., & Beaty, R. E. (2020). Community structure
of the creative brain at rest. NeuroImage, 210, 116578. https://doi
.org/10.1016/j.neuroimage.2020.116578, PubMed: 31982579
Kim, K. H. (2008). Meta-analyses of the relationship of creative
achievement to both IQ and divergent thinking test scores. Jour-
nal of Creative Behavior, 42(2), 106–130. https://doi.org/10.1002
/j.2162-6057.2008.tb01290.x
Kosslyn, S. M., & Thompson, W. L. (2003). When is early visual
cortex activated during visual mental imagery? Psychological
Bulletin, 129(5), 723–746. https://doi.org/10.1037/0033-2909
.129.5.723, PubMed: 12956541
Kourtzi, Z., & Kanwisher, N. (2001). Representation of perceived
object shape by the human lateral occipital complex. Science,
293(5534), 1506–1509. https://doi.org/10.1126/science
.1061133, PubMed: 11520991
Larsson, J., & Smith, A. T. (2012). fMRI repetition suppression: Neu-
ronal adaptation or stimulus expectation? Cerebral Cortex, 22(3),
567–576. https://doi.org/10.1093/cercor/ bhr119, PubMed:
21690262
LeBoutillier, N., & Marks, D. F. (2003). Mental imagery and creativ-
ity: A meta-analytic review study. British Journal of Psychology,
94(Pt. 1), 29–44. https://doi.org/10.1348/000712603762842084,
PubMed: 12648387
Lotze, M., Erhard, K., Neumann, N., Eickhoff, S. B., & Langner, R.
(2014). Neural correlates of verbal creativity: Differences in
resting-state functional connectivity associated with expertise in
creative writing. Frontiers in Human Neuroscience, 8, 516.
https://doi.org/10.3389/fnhum.2014.00516, PubMed: 25076885
Madore, K. P., Thakral, P. P., Beaty, R. E., Addis, D. R., & Schacter,
D. L. (2019). Neural mechanisms of episodic retrieval support
divergent creative thinking. Cerebral Cortex, 29(1), 150–166.
https://doi.org/10.1093/cercor/bhx312, PubMed: 29161358
Meyer, M. L., Hershfield, H. E., Waytz, A. G., Mildner, J. N., &
Tamir, D. I. (2019). Creative expertise is associated with trans-
cending the here and now. Journal of Personality and Social Psy-
chology, 116(4), 483–494. https://doi.org/10.1037/pspa0000148,
PubMed: 30714758
Murphy, K., Birn, R. M., Handwerker, D. A., Jones, T. B., & Bandettini,
P. A. (2009). The impact of global signal regression on resting state
correlations: Are anti-correlated networks introduced? Neuro-
Image, 44(3), 893–905. https://doi.org/10.1016/j.neuroimage
.2008.09.036, PubMed: 18976716
Orwig, W., Diez, I., Vannini, P., Beaty, R., & Sepulcre, J. (2021).
Creative connections: Computational semantic distance captures
individual creativity and resting-state functional connectivity.
Journal of Cognitive Neuroscience, 33(3), 499–509. https://doi
.org/10.1162/jocn_a_01658, PubMed: 33284079
Park, D. C., & Reuter-Lorenz, P. (2009). The adaptive brain: Aging
and neurocognitive scaffolding. Annual Review Psychology, 60,
173–196. https://doi.org/10.1146/annurev.psych.59.103006
.093656, PubMed: 19035823
Plucker, J. A. (1999). Is the proof in the pudding? Reanalyses of
Torrance’s (1958 to present) longitudinal data. Creativity Research
Journal, 12, 103–114. https://doi.org/10.1207/s15326934crj1202_3
Network Neuroscience
1032
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
7
3
1
0
2
2
2
1
5
4
7
7
1
n
e
n
_
a
_
0
0
3
1
7
p
d
/
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Creativity and functional connectivity
Qian, J., Diez, I., Ortiz-Terán, L., Bonadio, C., Liddell, T., Goñi, J., &
Sepulcre, J. (2018). Positive connectivity predicts the dynamic
intrinsic topology of the human brain network. Frontiers in Sys-
tems Neuroscience, 12, 38. https://doi.org/10.3389/fnsys.2018
.00038, PubMed: 30214399
Raichle, M. E., MacLeod, A. M., Snyder, A. Z., Powers, W. J.,
Gusnard, D. A., & Shulman, G. L. (2001). A default mode of
brain function. Proceedings of the National Academy of Sciences,
98(2), 676–682. https://doi.org/10.1073/pnas.98.2.676, PubMed:
11209064
Ran, Q., Jamoulle, T., Schaeverbeke, J., Meersmans, K., Vandenberghe,
R., & Dupont, P. (2020). Reproducibility of graph measures at the
subject level using resting-state fMRI. Brain and Behavior, 10(8),
2336–2351. https://doi.org/10.1002/ brb3.1705, PubMed:
32614515
Reuter-Lorenz, P.A., Jonides, J., Smith, E. E., Hartley, A., & Miller, A.
(2000). Age differences in the frontal lateralization of verbal and
spatial working memory revealed by PET. Journal of Cognitive
Neuroscience, 12(1), 174–187. https://doi.org/10.1162
/089892900561814, PubMed: 10769314
Rubinov, M., & Sporns, O. (2010). Complex network measures of
brain connectivity: Uses and interpretations. NeuroImage, 52(3),
1059–1069. https://doi.org/10.1016/j.neuroimage.2009.10.003,
PubMed: 19819337
Runco, M. A., & Acar, S. (2012). Divergent thinking as an indicator
of creative potential. Creativity Research Journal, 24(1), 66–75.
https://doi.org/10.1080/10400419.2012.652929
Schacter, D. L., Addis, D. R., Hassabis, D., Martin, V. C., Spreng,
R. N., & Szpunar, K. K. (2012). The future of memory: Remem-
bering, imagining, and the brain. Neuron, 76(4), 677–694.
https://doi.org/10.1016/j.neuron.2012.11.001, PubMed:
23177955
Schacter, D. L., Benoit, R. G., & Szpunar, K. K. (2017). Episodic
future thinking: Mechanisms and functions. Current Opinion in
Behavioral Sciences, 17, 41–50. https://doi.org/10.1016/j
.cobeha.2017.06.002, PubMed: 29130061
Schooler, J. W., Smallwood, J., Christoff, K., Handy, T. C., Reichle,
E. D., & Sayette, M. A. (2011). Meta-awareness, perceptual
decoupling and the wandering mind. Trends in Cognitive Sci-
ences, 15(7), 319–326. https://doi.org/10.1016/j.tics.2011.05
.006, PubMed: 21684189
Shaw, G. A., & Belmore, S. M. (1982). The relationship between
imagery and creativity. Imagination, Cognition and Personality,
2(2), 115–123. https://doi.org/10.2190/4RGA-Y1A6-HEK5-LMF8
Shi, L., Sun, J., Xia, Y., Ren, Z., Chen, Q., Wei, D., Yang, W., & Qiu,
J. (2018). Large-scale brain network connectivity underlying cre-
ativity in resting-state and task fMRI: Cooperation between
default network and frontal-parietal network. Biological
Psychology, 135, 102–111. https://doi.org/10.1016/j.biopsycho
.2018.03.005, PubMed: 29548807
Stevens, W. D., & Spreng, R. N. (2014). Resting-state functional
connectivity MRI reveals active processes central to cognition.
Wiley Interdisciplinary Reviews: Cognitive Science, 5(2),
233–245. https://doi.org/10.1002/wcs.1275, PubMed: 26304310
Takeuchi, H., Taki, Y., Hashizume, H., Sassa, Y., Nagase, T.,
Nouchi, R., & Kawashima, R. (2012). The association between
resting functional connectivity and creativity. Cerebral Cortex,
22(12), 2921–2929. https://doi.org/10.1093/cercor/ bhr371,
PubMed: 22235031
Thakral, P. P., Madore, K. P., Kalinowski, S. E., & Schacter, D. L.
(2020). Modulation of hippocampal brain networks produces
changes in episodic simulation and divergent thinking. Proceedings
of the National Academy of Sciences, 117(23), 12729–12740.
https://doi.org/10.1073/pnas.2003535117, PubMed: 32457143
Thakral, P. P., Yang, A. C., Addis, D. R., & Schacter, D. L. (2021).
Divergent thinking and constructing future events: Dissociating
old from new ideas. Memory, 29(6), 729–743. https://doi.org
/10.1080/09658211.2021.1940205, PubMed: 34182887
Torrance, E. P. (1988). The nature of creativity as manifest in its testing.
In R. J. Sternberg (Ed.), The nature of creativity: Contemporary psy-
chological perspectives (pp. 43–75). Cambridge University Press.
Van Essen, D. C. (2005). A population-average, landmark- and
surface-based (PALS) atlas of human cerebral cortex. Neuro-
Image, 28(3), 635–662. https://doi.org/10.1016/j.neuroimage
.2005.06.058, PubMed: 16172003
Wu, X., Yang, W., Tong, D., Sun, J., Chen, Q., Wei, D., Zhang, Q.,
Zhang, M., & Qiu, J. (2015). A meta-analysis of neuroimaging
studies on divergent thinking using activation likelihood estima-
tion. Human Brain Mapping, 36(7), 2703–2718. https://doi.org
/10.1002/hbm.22801, PubMed: 25891081
Yeo, B. T., Krienen, F. M., Sepulcre, J., Sabuncu, M. R., Lashkari, D.,
Hollinshead, M., Roffman, J. L., Smoller, J. W., Zöllei, L.,
Polimeni, J. R., Fischl, B., Liu, H., & Buckner, R. L. (2011). The
organization of the human cerebral cortex estimated by intrinsic
functional connectivity. Journal of Neurophysiology, 106(3),
1125–1165. https://doi.org/10.1152/jn.00338.2011, PubMed:
21653723
Zeng, L., Proctor, R. W., & Salvendy, G. (2011). Can traditional
divergent thinking tests be trusted in measuring and predicting
real-world creativity? Creativity Research Journal, 23, 24–37.
https://doi.org/10.1080/10400419.2011.545713
Zhang, Y., Brady, M., & Smith, S. (2001). Segmentation of brain MR
images through a hidden Markov random field model and the
expectation-maximization algorithm. IEEE Transactions on Med-
ical Imaging, 20(1), 45–57. https://doi.org/10.1109/42.906424,
PubMed: 11293691
Network Neuroscience
1033
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
7
3
1
0
2
2
2
1
5
4
7
7
1
n
e
n
_
a
_
0
0
3
1
7
p
d
.
t
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3