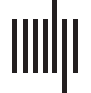RESEARCH
The topology, stability, and instability of
learning-induced brain network
repertoires in schizophrenia
Emmanuel D. Meram, Shahira Baajour, Asadur Chowdury, John Kopchick, Patricia Thomas,
Usha Rajan, Dalal Khatib, Caroline Zajac-Benitez, Luay Haddad, Alireza Amirsadri,
Jeffrey A. Stanley, and Vaibhav A. Diwadkar
Department of Psychiatry and Behavioral Neurosciences, Brain Imaging Research Division,
Wayne State University School of Medicine, Detroit, MI, USA
a n o p e n a c c e s s
j o u r n a l
Keywords: Schizophrenia, Associative learning, fMRI, Graph theory, Betweenness centrality
ABSTRACT
There is a paucity of graph theoretic methods applied to task-based data in schizophrenia
(SCZ). Tasks are useful for modulating brain network dynamics, and topology. Understanding
how changes in task conditions impact inter-group differences in topology can elucidate
unstable network characteristics in SCZ. Here, in a group of patients and healthy controls
(n = 59 total, 32 SCZ), we used an associative learning task with four distinct conditions
(Memory Formation, Post-Encoding Consolidation, Memory Retrieval, and Post-Retrieval
Consolidation) to induce network dynamics. From the acquired fMRI time series data,
betweenness centrality (BC), a metric of a node’s integrative value was used to summarize
network topology in each condition. Patients showed (a) differences in BC across multiple
nodes and conditions; (b) decreased BC in more integrative nodes, but increased BC in less
integrative nodes; (c) discordant node ranks in each of the conditions; and (d) complex
patterns of stability and instability of node ranks across conditions. These analyses reveal that
task conditions induce highly variegated patterns of network dys-organization in SCZ. We
suggest that the dys-connection syndrome that is schizophrenia, is a contextually evoked
process, and that the tools of network neuroscience should be oriented toward elucidating
the limits of this dys-connection.
AUTHOR SUMMARY
This paper demonstrates disordered network repertoires in schizophrenia during learning and
memory. Graph theoretic analysis was applied to fMRI data collected during four distinct
phases of a learning paradigm, to capture network repertoires. We show that the network
topology changes across conditions, and that brain nodes peripheral to learning assumed
greater importance in schizophrenia. Moreover, nodes with highly stable contributions to
network topology were aberrantly localized to early sensory cortex in schizophrenia, but
distributed across the brain in healthy participants. These results highlight the dynamic nature
of the dys-connection syndrome, emphasizing the value of studying task-induced network
topology in schizophrenia in conjunction with graph theoretic metrics.
Citation: Meram, E. D., Baajour, S.,
Chowdury, A., Kopchick, J., Thomas,
P., Rajan, U., Khatib, D., Zajac-Benitez,
C., Haddad, L., Amirsadri, A., Stanley,
J. A., & Diwadkar, V. A. (2023). The
topology, stability, and instability
of learning-induced brain network
repertoires in schizophrenia. Network
Neuroscience, 7(1), 184–212. https://
doi.org/10.1162/netn_a_00278
DOI:
https://doi.org/10.1162/netn_a_00278
Supporting Information:
https://doi.org/10.1162/netn_a_00278
Received: 27 April 2022
Accepted: 5 September 2022
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Author:
Vaibhav A. Diwadkar
vdiwadka@med.wayne.edu
Handling Editor:
Alex Fornito
Copyright: © 2022
Massachusetts Institute of Technology
Published under a Creative Commons
Attribution 4.0 International
(CC BY 4.0) license
The MIT Press
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
7
1
1
8
4
2
0
7
2
1
3
3
n
e
n
_
a
_
0
0
2
7
8
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Learning-induced brain network repertoires in schizophrenia
Network repertoire:
A property of a network as (in our
case) summarized by graph theoretic
measures.
Learning:
The process of associating pairs of
arbitrary memoranda during an
experimentally titrated paradigm.
Betweenness centrality:
Our employed graph theoretic metric
indexing a node’s integrative value
or “hubness” (i.e., relative
importance within a network).
Memory Formation/encoding:
A learning condition during which
pairs of items are visually presented
for forming associations.
Post-Encoding Consolidation:
An instruction-free covert condition
that immediately follows Memory
Formation.
Memory Retrieval:
A condition during which
participants must recall (from
memory) the item associated with the
retrieval cue.
Post-Retrieval Consolidation:
An instruction-free covert condition
that immediately follows Memory
Retrieval.
INTRODUCTION
Schizophrenia is a complex and debilitating neuropsychiatric disorder that has long been
characterized as a “dys-connection syndrome” of the brain (Friston, Brown, Siemerkus, &
Stephan, 2016; Silverstein, Bressler, & Diwadkar, 2016). Disordered brain network repertoires
are characteristic signatures of dys-connection (Spronk et al., 2021), indicative of a profound
loss of functional integrity that the disorder induces. The current investigation was motivated
by two interrelated goals: (a) to characterize altered brain network repertoires in schizophrenia
evoked during each of four distinct conditions of an associative learning paradigm (Baajour
et al., 2020; Stanley et al., 2017), by (b) estimating the betweenness centrality (BC) (Rubinov
& Sporns, 2010; Telesford, Joyce, Hayasaka, Burdette, & Laurienti, 2011) of each cerebral
node. BC, a graph theoretic measure, is well suited to capture the integrative profile of a node
in any directed or undirected graph. In our case, nodes represent brain regions and vertices
represent the condition-evoked “distance” (i.e., estimated functional connectivity) between
those regions.
In schizophrenia, graph theoretic measures have been widely used to capture altered net-
work repertoires derived from resting-state fMRI signals (J. Chen et al., 2021; Ji et al., 2019; Lei
et al., 2020; Rolls et al., 2020). More recent applications have used graph theory to investigate
altered task-driven connectomics. These investigations have been conducted in domains
including working memory (Deng et al., 2022; Stolz, Emerson, Nahkuri, Porter, & Harrington,
2021; Yang et al., 2020), executive function (Zhao, Cheng, Li, & Yu, 2018), social cognition
(Oliver et al., 2021), and theory of mind (Bitsch, Berger, Nagels, Falkenberg, & Straube, 2021).
Task-driven fMRI is valuable in explicitly driving network dynamics, thus promoting discovery
of contextually driven network differences (Baajour et al., 2020; Thakkar, Diwadkar, & Rolfs,
2017; Tso et al., 2021). Contextual modulation may be particularly valuable in understanding
schizophrenia, because functional dys-connection is like to be contextually evoked, rather
than invariant across behavioral contexts (Logothetis, 2008; Stephan & Roebroeck, 2012).
Our own reliance on an object-location associative learning paradigm (Büchel, Coull, &
Friston, 1999; Diwadkar et al., 2008) was motivated by (a) the knowledge that impaired
learning interjects with the schizophrenia phenotype (Brambilla et al., 2011; Brambilla, Riva,
Melcangi, & Diwadkar, 2007; Stephan, Baldeweg, & Friston, 2006; Wannan et al., 2018), and
(b) the ability to configure the learning paradigm into distinct conditions related to Memory
Formation (Encoding), Post-Encoding Consolidation (requiring recapitulation of associations
presented), cued Memory Retrieval (retrieval of object name at the cued location), and
Post-Retrieval Consolidation. While the process of learning is distributed across conditions,
each condition provides unique characteristics to the learning process. Thus, Memory Forma-
tion and Retrieval are performance driven, but Consolidation while devoid of sensorimotor
stimulation is an active rest state, requiring memories to be rehearsed and recapitulated
(Ravishankar et al., 2019). Accordingly, we expect network repertoires summarized by BC
(Bullmore & Sporns, 2009) to reflect condition-driven processing. Inter-group differences
are unlikely to be condition independent.
Graph theoretic approaches can efficiently summarize swathes of spatiotemporal fMRI data
(Farahani, Karwowski, & Lighthall, 2019), and multiple metrics have been proposed for cap-
turing the different roles played by a node in modulating the “flow” of information in directed
or undirected graphs (van den Heuvel & Sporns, 2013). These metrics range from “basic” mea-
sures of degree centrality including in- and out-degree centrality (that respectively capture the
number of vertices entering or exiting a node), to more “integrative” measures such as close-
ness centrality (CC) (Bavelas, 1950) and BC. Integrative metrics are designed to capture a
Network Neuroscience
185
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
7
1
1
8
4
2
0
7
2
1
3
3
n
e
n
_
a
_
0
0
2
7
8
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Learning-induced brain network repertoires in schizophrenia
node’s role based on its cumulative relationships with other nodes in a network. CC is calcu-
lated as the reciprocal of the sum of the length of the shortest paths between the node and all
other nodes in the graph, and while similar to BC, has typically been employed with binary as
opposed to weighted graphs. BC, however, is more sensitive to the integrative value of a node
precisely because it quantifies how a node acts as a bridge along the shortest path between
any two other nodes, a measure that is a central element of a node’s functional role. Thus, BC
provides an excellent representation of a node’s relative importance within a network and has
been used as an index of a node’s “hubness” (Rubinov & Sporns, 2010) or integrative value
(Kivimaki, Lebichot, Saramaki, & Saerens, 2016). Understanding how a highly integrative task
like associative learning impacts the integrative roles of nodes was central to our motivations,
providing the rationale for our choice of BC in characterizing network repertoires.
Across participants and conditions, the edges between nodes in each graph were repre-
sented by a typical distance metric (using bivariate correlation models applied to summarize
time series data) (Wang et al., 2015) (resulting in a weighted, undirected graph). Then, BC was
estimated for each node in each participant and condition. Subsequent analyses were directed
toward understanding (a) how each of the four task conditions modulated the topology of net-
work repertoires and inter-group differences, that is, healthy controls ≠ patients with schizo-
phrenia (HC ≠ SCZ), (b) how the integrative importance of nodes changed as a function of task
condition, and (c) inter-group similarities and differences in the stability and instability of the
observed integrative importance of nodes. In Supporting Information we also report relation-
ships between task proficiency and BC in both groups, and in patients, we report on how
well BC was predicted by variables including antipsychotic dosage (Nosé & Barbui, 2008;
Nosé et al., 2008), duration of illness, and clinical state (estimated by the Positive and Neg-
ative Syndrome Scale (PANSS)) (Kay, Fiszbein, & Opler, 1987). As a complement to existing
investigations of resting-state fMRI in SCZ (Alexander-Bloch et al., 2010; van den Heuvel,
Mandl, Stam, Kahn, & Hulshoff Pol, 2010), we suggest that this work along with other noted
graph theoretic applications to task-based data, accentuates the role of contextual process-
ing in revealing the dynamics of dys-connection in schizophrenia.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
7
1
1
8
4
2
0
7
2
1
3
3
n
e
n
_
a
_
0
0
2
7
8
p
d
.
t
RESULTS
We organize the presentation of results in the following logical order. First, we first provide a
comprehensive accounting of inter-group differences in BC across each of the four experimen-
tal conditions (Figure 1A, Figure 2A, Figure 3A, and Figure 4A). For ease of access, nodes are
classified by lobe (frontal, basal ganglia, temporal, parietal, visual, others; Supporting Informa-
tion Table S1 provides a listing of regional classifications). Next, in Figure 5, Figure 6, Figure 7,
and Figure 8, which are based on rank ordering by BC, we examine the relative intranetwork
importance of nodes. Third, these emerging results led to further explorations of the relationship
between node rankings across groups in each of the four conditions (Figure 9A, Figure 10A,
Figure 11A, and Figure 12A). Fourth, in a final assessment, in each group, we identified nodes
with stable and high ranks across all four memory conditions (Figure 13 and Figure 14).
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Betweenness Centrality Analysis
In Figures 1A–4A, the bar graphs represent the mean (±SEM ) BC for each group for that node
(SCZ are represented by opaque colors, whereas HC are represented by transparent colors).
Nodes with significant inter-group differences (pFDR < 0.05, SCZ ≠ HC) are identified by insets
(and labels). In Figures 1B–4B, nodes with significant inter-group differences are presented on
lateral or medial cortical surfaces (for a succinct visual summary).
Stability:
How much a node’s rank by
betweenness centrality changes
across conditions (derived from the
standard deviation of ranks).
Rank:
The order of nodes based on its
betweenness centrality; a lower
number reflects higher rank (i.e.,
greater integrative importance).
Network Neuroscience
186
Learning-induced brain network repertoires in schizophrenia
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
1
8
4
2
0
7
2
1
3
3
n
e
n
_
a
_
0
0
2
7
8
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 1. BC during Memory Formation. (A) BC values during Memory Formation are depicted for each of the 90 cerebral parcels (“nodes”). The
nodes are organized (and color coded) by lobe (with the color scheme maintained in all subsequent figures). The slightly transparent bars rep-
resent healthy controls (HC) and the opaque bars represent patients (SCZ). Significant differences in BC (pFDR < 0.05, HC ≠ SCZ), are
clearly identified (insets and asterisks). As is evident, Memory Formation induced significant decreases in BC in multiple nodes, including the
middle temporal, superior frontal, precuneus, thalamus, and putamen. Conversely, increases in BC were observed in a complement of nodes
including the hippocampus, parahippocampal gyrus, superior temporal, superior frontal, inferior frontal, paracentral lobule, supramarginal,
cuneus, superior occipital, and middle occipital gyri. (B) For ease of visualization, nodes showing significant inter-group differences are rendered
on lateral or medial cortical surfaces where each subfigure denotes nodes with lower BC (left) or higher BC (right) in patients.
Network Neuroscience
187
Learning-induced brain network repertoires in schizophrenia
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
1
8
4
2
0
7
2
1
3
3
n
e
n
_
a
_
0
0
2
7
8
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
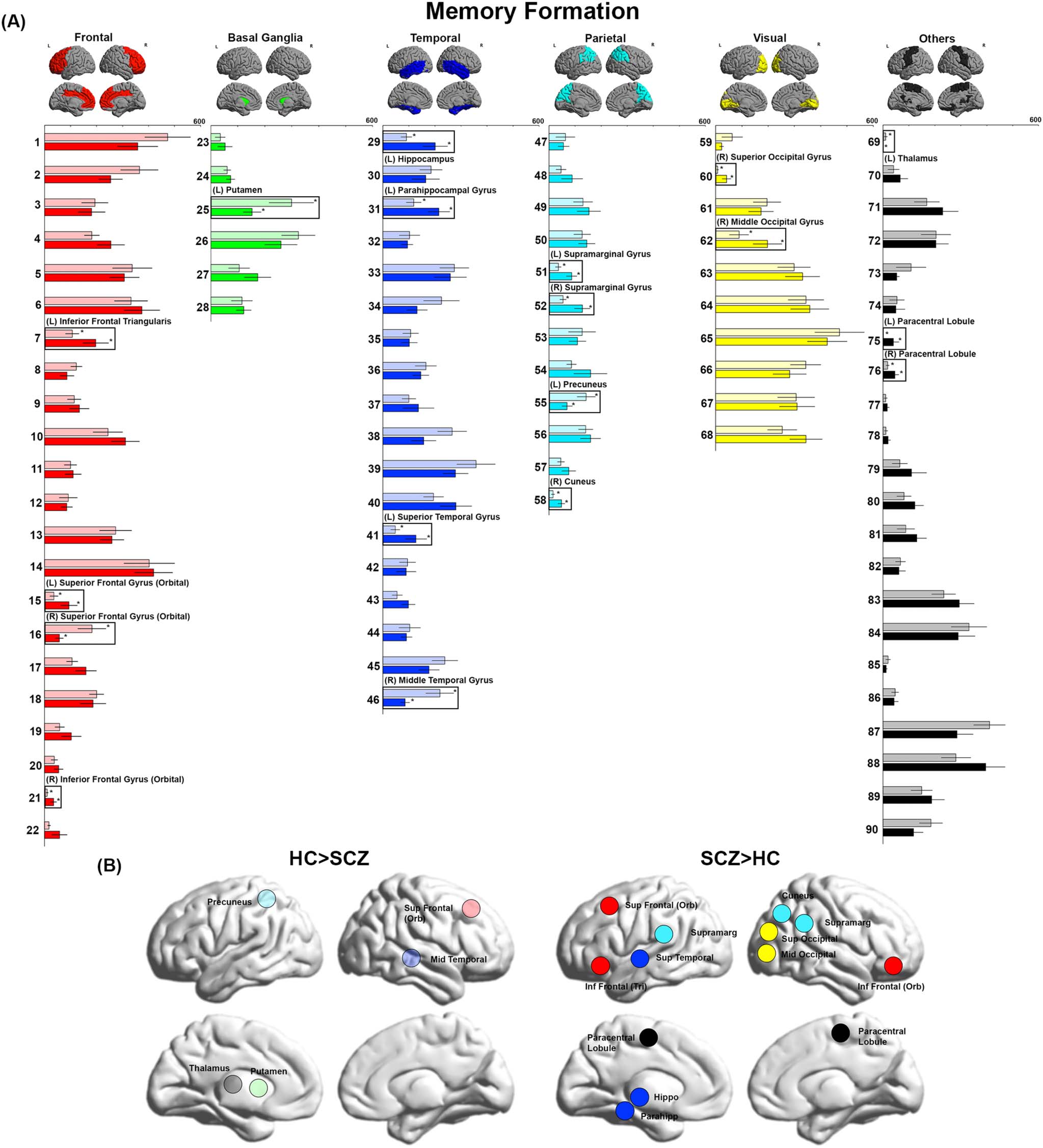
Figure 2. BC during Post-Encoding Consolidation. (A) BC values during Post-Encoding Consolidation are depicted. Even in the absence of
sensorimotor stimulation, significant inter-group differences were observed. As seen, we observed decreases in BC in the precuneus, inferior
occipital cortex, inferior temporal cortex, calcarine, Heschl’s gyrus, posterior cingulate, superior frontal, putamen, and thalamus. Conversely,
we observed increases in BC in the left hippocampus and right parahippocampal gyrus as well as multiple nodes across frontal, temporal, and
basal ganglia regions. (B) The locations of nodes with significant inter-group differences are visualized on lateral and medial cortical surfaces.
Network Neuroscience
188
Learning-induced brain network repertoires in schizophrenia
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
1
8
4
2
0
7
2
1
3
3
n
e
n
_
a
_
0
0
2
7
8
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
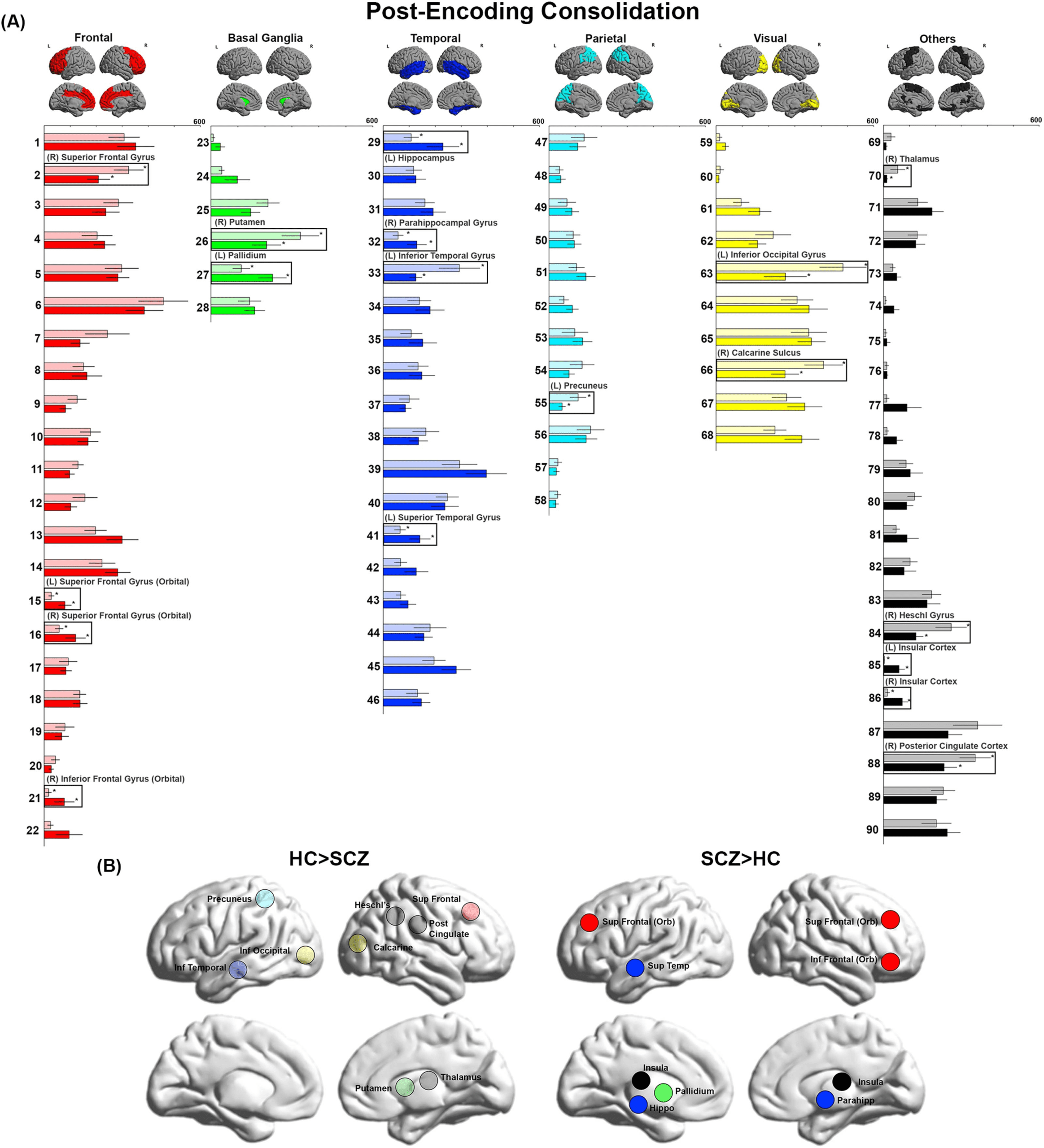
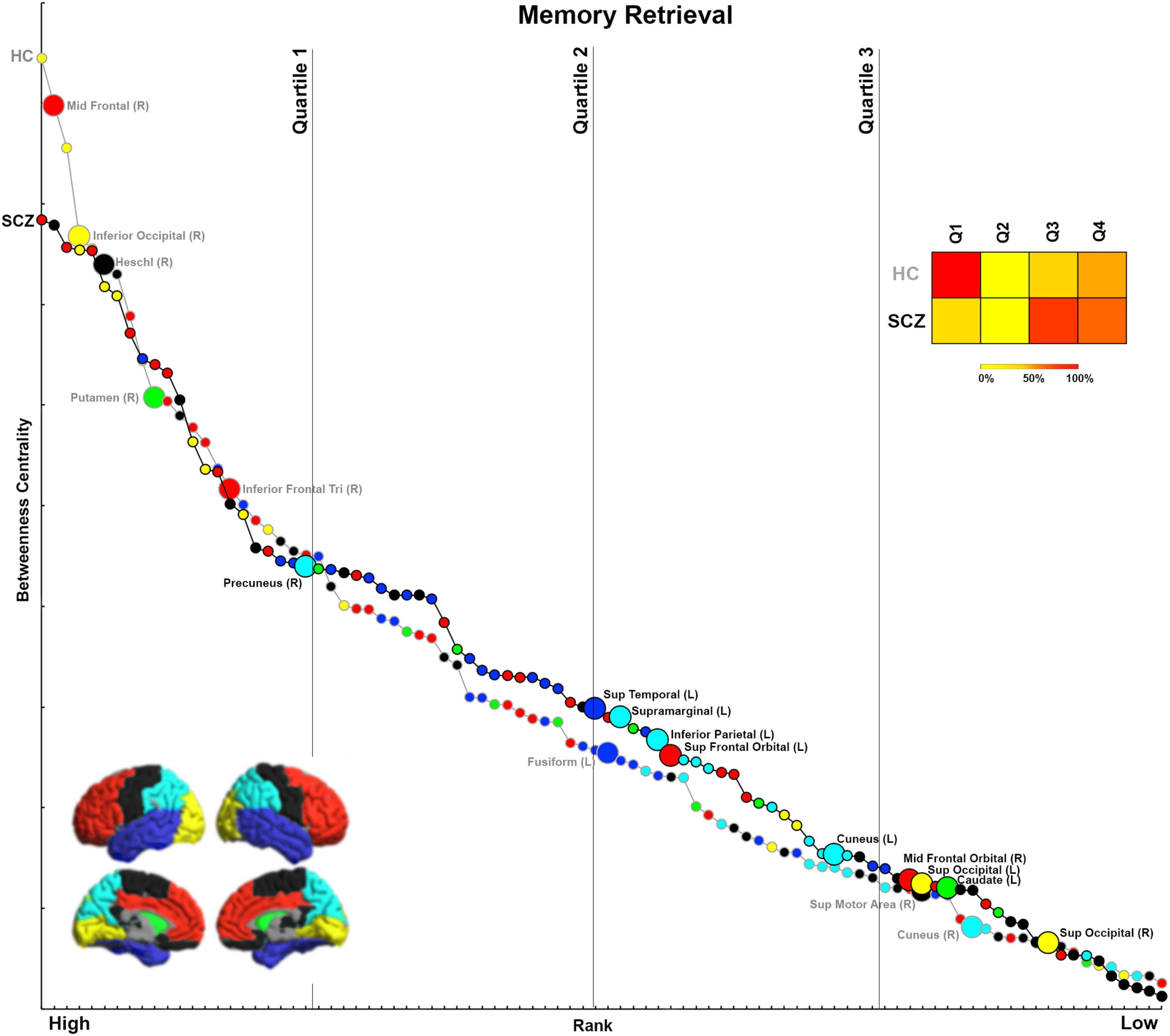
Figure 3. BC during Memory Retrieval. (A) BC values during Memory Retrieval are depicted. We observed decreases in BC in the cuneus,
Heschl’s gyrus, mid frontal, inferior frontal, inferior occipital, supplementary motor cortex, putamen, and fusiform. Conversely, we observed
increases in BC in the precuneus, superior occipital, mid frontal, cuneus, inferior parietal, supramarginal, superior frontal, superior temporal,
and caudate. (B) The locations of nodes with significant inter-group differences are visualized on lateral and medial cortical surfaces.
Network Neuroscience
189
Learning-induced brain network repertoires in schizophrenia
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
1
8
4
2
0
7
2
1
3
3
n
e
n
_
a
_
0
0
2
7
8
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
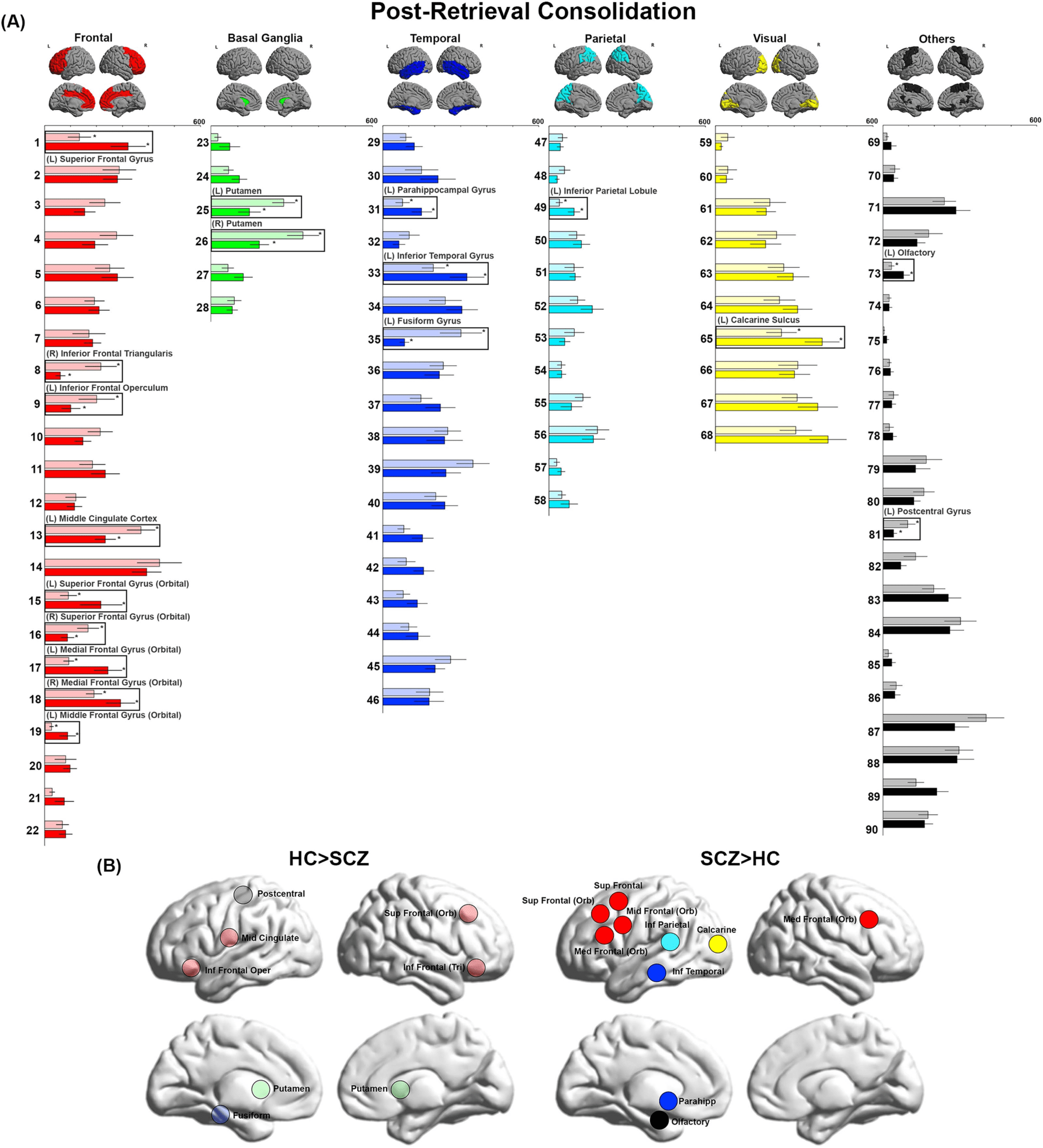
Figure 4. BC during Post-Retrieval Consolidation. (A) BC values during Post-Retrieval Consolidation are depicted. We observed decreases in
BC in the postcentral gyrus, fusiform, putamen, and multiple nodes in the frontal lobe. Conversely, we observed increases in BC in the olfactory
bulb, as well as multiple frontal, parietal, visual, and temporal lobe nodes. (B) The locations of nodes with significant inter-group differences
are visualized on lateral and medial cortical surfaces.
Network Neuroscience
190
Learning-induced brain network repertoires in schizophrenia
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
1
8
4
2
0
7
2
1
3
3
n
e
n
_
a
_
0
0
2
7
8
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
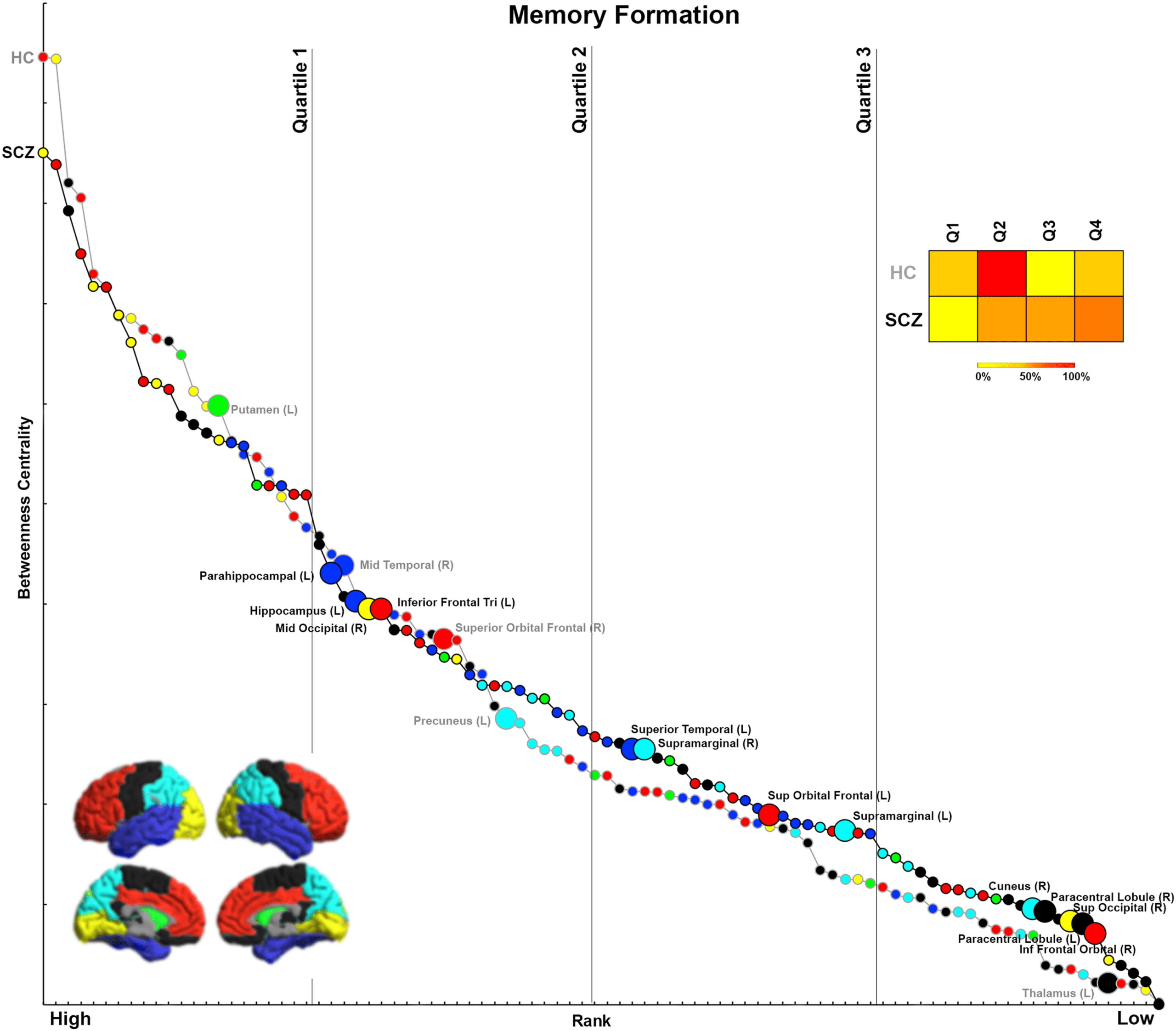
Figure 5. Ranking by BC for Memory Formation. After ranking each node within each group (HC, SCZ) by BC, data from each group
are depicted in this two-dimensional space mapping BC (vertical axis) and rank (horizontal axis). Contiguous curves connect nodes
within each group thus clearly separating data across HC (gray curve) and SCZ (black curve). Nodes with significant inter-group differ-
ences in BC (see Figure 1) are noted (enlarged marker and added node label within the group with the significant increase). Vertical lines
on the graph divide the ranking space into quartiles, allowing us to assign nodes with significant differences into rank-based quartiles.
The associated heat map represents the percentage of significantly different nodes in each rank-based quartile for each tail of the results
(HC > SCZ, SCZ > HC). It is possible to divine the relative importance of significantly different BC measures. For example, as seen
in SCZ, nodes with decreased BC tend to be somewhat more highly ranked, but nodes with increased BC tend to be somewhat more
lowly ranked.
Network Neuroscience
191
Learning-induced brain network repertoires in schizophrenia
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
7
1
1
8
4
2
0
7
2
1
3
3
n
e
n
_
a
_
0
0
2
7
8
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 6. Ranking by BC for Post-Encoding Consolidation. The figure presents data from the Post-Encoding Consolidation condition following
the convention of Figure 5. As seen, a high percentage of nodes that have decreased BC in SCZ lie in the first quartile of ranks, indicating that
nodes with significantly lower BC in the SCZ group are also highly ranked within the HC network.
Memory Formation
As seen, in SCZ significant reductions in BC were observed in the right superior frontal
gyrus (orbital), the left putamen, the right middle temporal gyrus, the left precuneus, and
the left thalamus. Conversely, increased BC was observed across several regions in the
frontal lobe (left inferior frontal triangularis, left superior frontal gyrus (orbital), right inferior
frontal gyrus (orbital)), the temporal and medial temporal lobe (left hippocampus, left para-
hippocampal gyrus, left superior temporal gyrus), the parietal lobe (bilateral supramarginal
Network Neuroscience
192
Learning-induced brain network repertoires in schizophrenia
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
7
1
1
8
4
2
0
7
2
1
3
3
n
e
n
_
a
_
0
0
2
7
8
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 7. Ranking by BC for Memory Retrieval. The figure presents BC and rank data from each group for the Memory Retrieval condition.
Again, nodes with significantly lower BC in SCZ tend to be more highly ranked within the HC network.
gyrus, right cuneus), and the visual lobe (right superior occipital gyrus, right middle occip-
ital gyrus).
Post-Encoding Consolidation
SCZ showed decreased BC across several lobes, including the frontal (right superior frontal
gyrus), temporal (left inferior temporal gyrus), parietal (left precuneus), and visual cortices (left
inferior occipital gyrus and right calcarine sulcus), and the basal ganglia (right putamen).
Decreases were also observed in the right thalamus, right Heschl’s gyrus, and right posterior
Network Neuroscience
193
Learning-induced brain network repertoires in schizophrenia
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
7
1
1
8
4
2
0
7
2
1
3
3
n
e
n
_
a
_
0
0
2
7
8
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
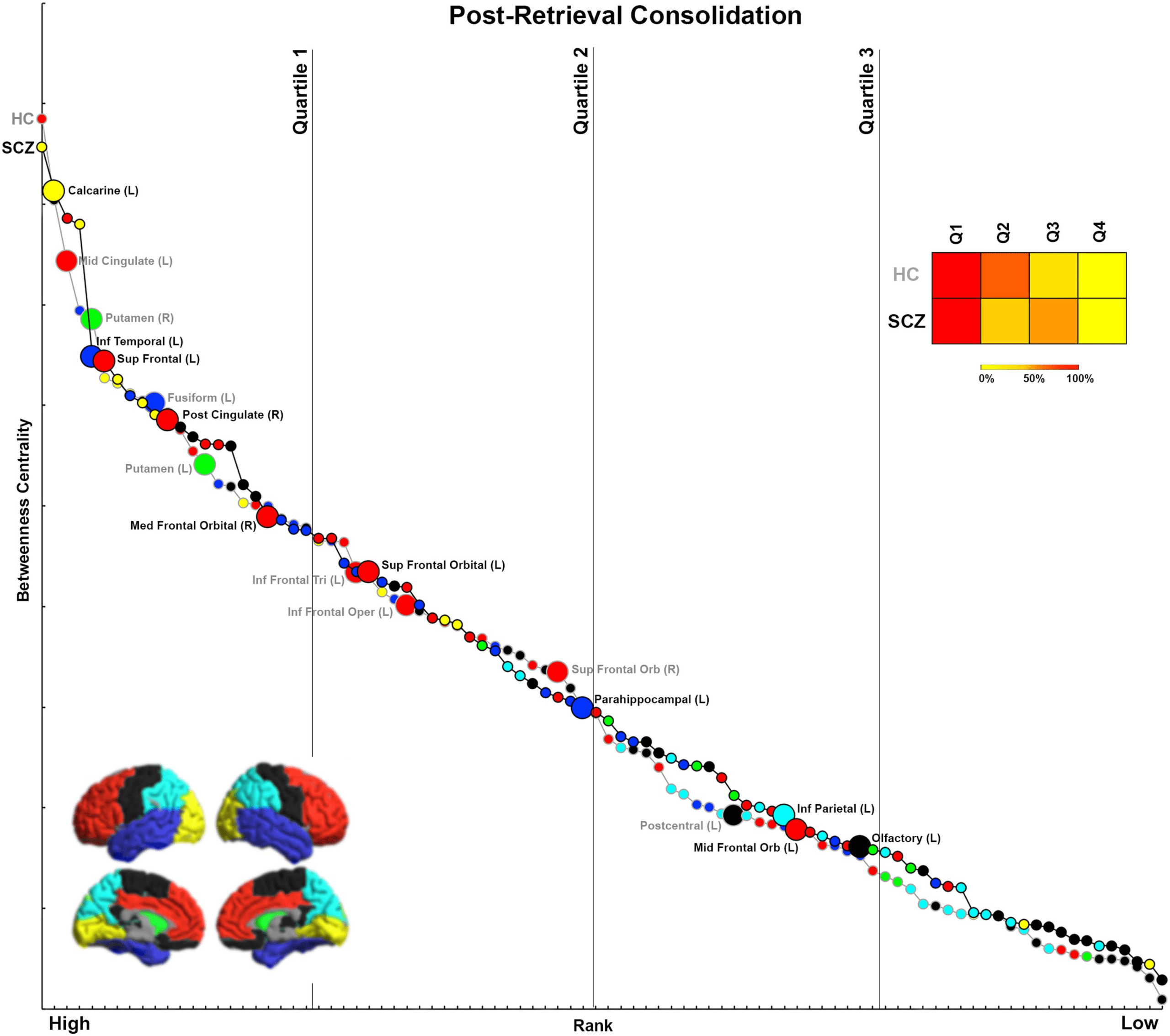
Figure 8. Ranking by BC for Post-Retrieval Consolidation. The figure presents BC and rank data from each group for the Post-Retrieval
Consolidation condition. Notably, and unlike in Figures 5–7, nodes with both decreased and increased BC in SCZ tend to be more highly
ranked.
cingulate cortex. Conversely, we observed increases in BC in multiple lobes, including the
frontal (bilateral superior frontal gyrus (orbital) and the right inferior frontal gyrus (orbital)),
basal ganglia (left pallidum), and temporal (left hippocampus, right parahippocampal gyrus,
left superior temporal gyrus) as well as the bilateral insular cortex.
Memory Retrieval
Decreased BC was observed in eight nodes across the frontal lobe (right middle frontal gyrus,
right inferior frontal triangularis), basal ganglia (right putamen), temporal lobe (left fusiform
Network Neuroscience
194
Learning-induced brain network repertoires in schizophrenia
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
7
1
1
8
4
2
0
7
2
1
3
3
n
e
n
_
a
_
0
0
2
7
8
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
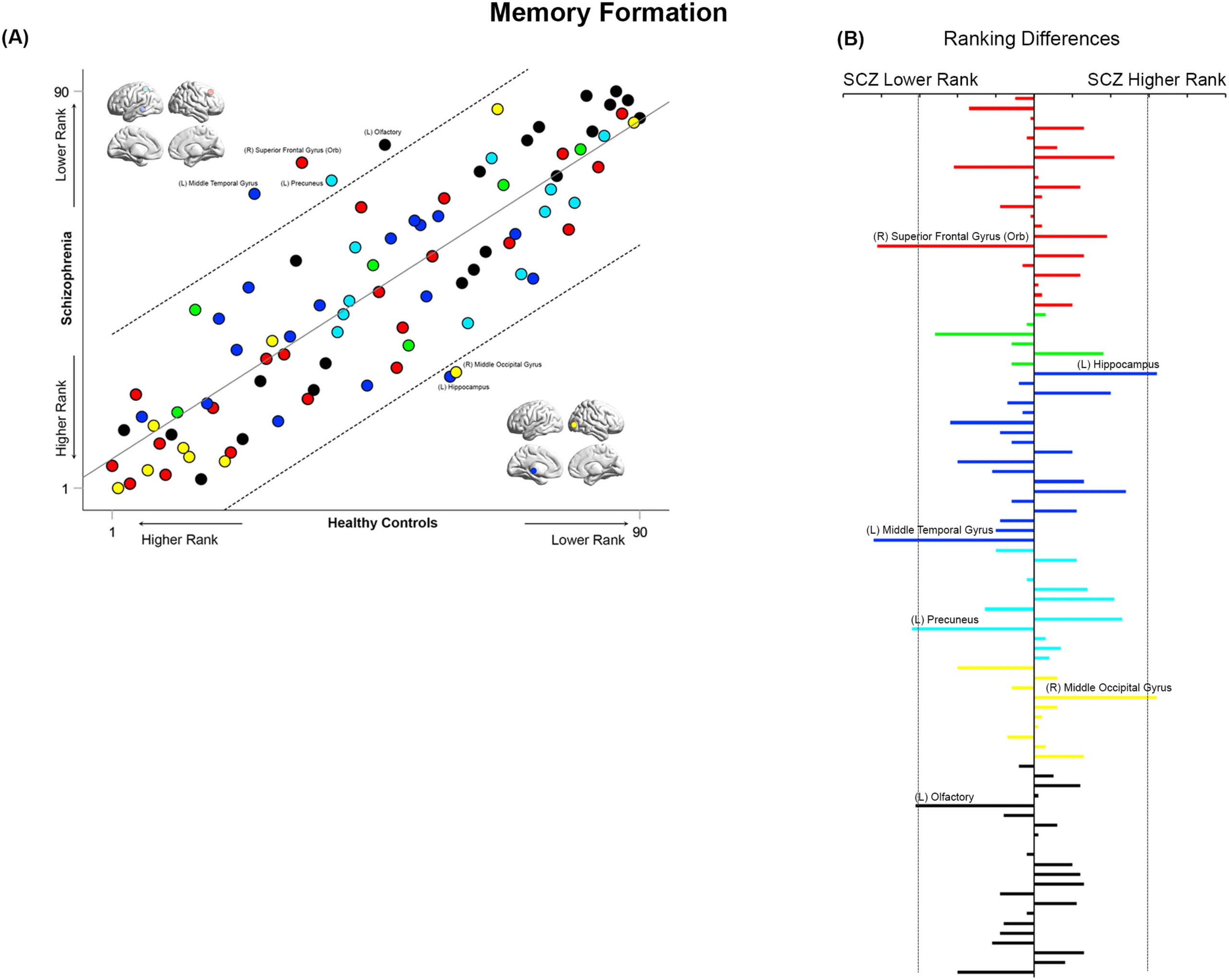
Figure 9. Cross-ranking for Memory Formation. (A) The scatter plot depicts the cross-ranking of nodes across each of the HC (horizontal
axis) and SCZ groups (vertical axis). The color coding of nodes is consistent with the general scheme used in the manuscript. The regression
line represents the best fit linear model (see Results), with the dashed lines representing the 95% confidence interval for the model. While
node ranks are highly correlated across groups, multiple nodes lay outside the confidence interval of the regression model. These include
the left middle temporal gyrus, left precuneus, right superior frontal gyrus (orbital), and left olfactory (lower rank in SCZ), and the left
hippocampus and right middle occipital gyrus (higher rank in SCZ). (B) To further accentuate the import of the identified nodes from panel
A, across all 90 nodes, we computed the absolute inter-group difference in rank. These are depicted in the bar graph (organized from top to
bottom along the lines of Supporting Information Table S1). The dropped dashed lines reflect the 95% confidence interval of the model from
panel A, and as seen, the absolute value of the difference in rank lies outside the interval for both directions (HC > SCZ and SCZ > HC).
gyrus), parietal lobe (right cuneus), and visual cortex (right inferior occipital gyrus). Addition-
ally, decreased BC was observed in the right supplementary motor area and right Heschl’s
gyrus. By comparison, increased BC was observed in 10 nodes, centered mostly around the
parietal lobe (left inferior parietal lobule, left supramarginal gyrus, right precuneus, left
cuneus), with additional effects in the frontal lobe (left superior frontal gyrus (orbital) and
Network Neuroscience
195
Learning-induced brain network repertoires in schizophrenia
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
7
1
1
8
4
2
0
7
2
1
3
3
n
e
n
_
a
_
0
0
2
7
8
p
d
.
t
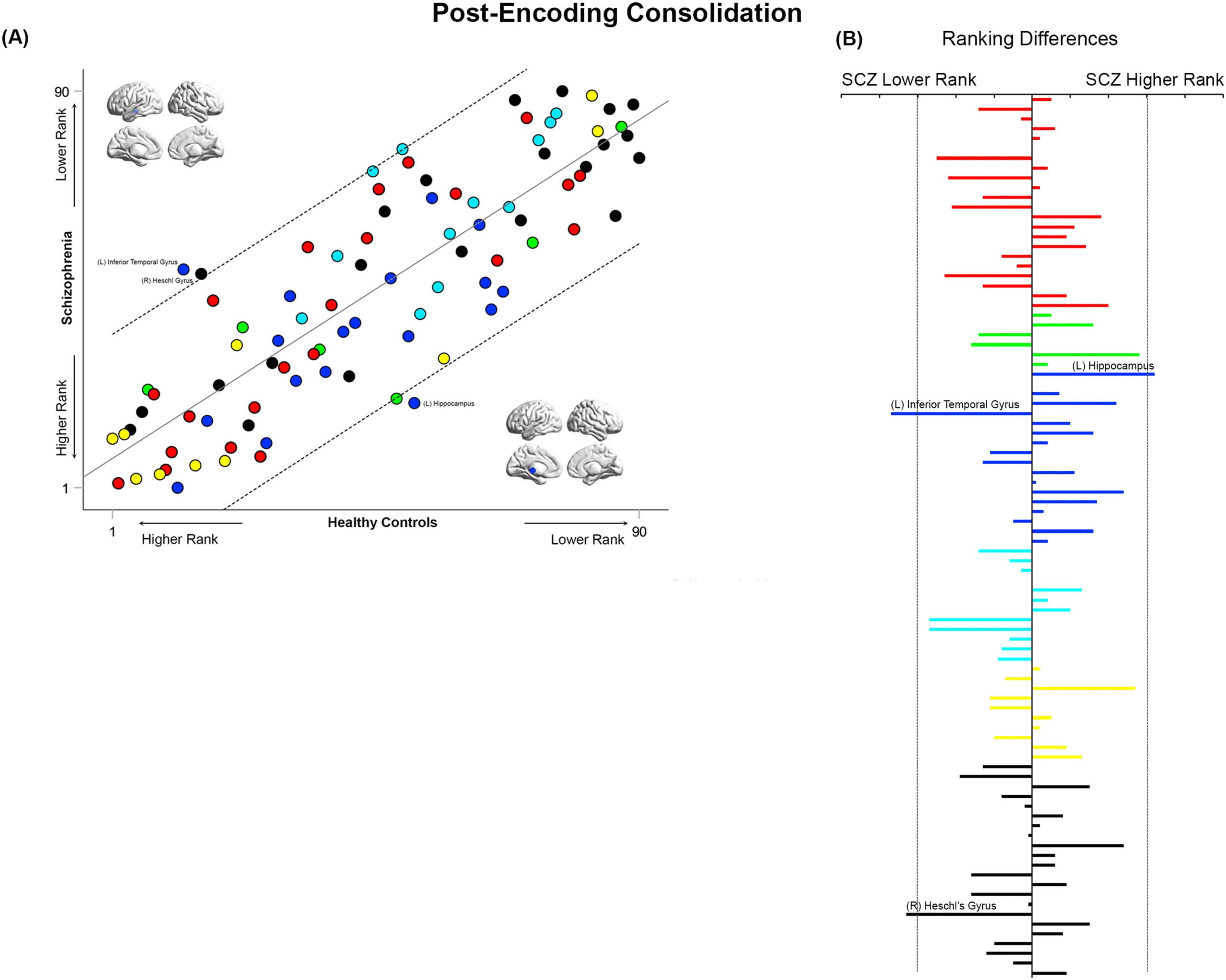
Figure 10. Cross-ranking for Post-Encoding Consolidation. (A) Cross-ranking information (consistent with the format in Figure 9) is presented
for the Post-Encoding Consolidation condition. While node ranks are highly correlated across groups, multiple nodes lay outside the confi-
dence interval of the regression model. These include the left inferior temporal gyrus and right Heschl’s gyrus (lower rank in SCZ), and the left
hippocampus (higher rank in SCZ). (B) The absolute inter-group difference in rank is presented in the same convention as Figure 9.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
the right middle frontal gyrus (orbital)), basal ganglia (left caudate nucleus), temporal lobe (left
superior temporal gyrus), and visual cortex (bilateral superior occipital gyrus).
Post-Retrieval Consolidation
Decreased BC was observed primarily in frontal nodes (right inferior frontal triangularis, left
inferior frontal operculum, left middle cingulate cortex, and right superior frontal gyrus
(orbital)). Moreover, decreases were also observed in the basal ganglia (bilateral putamen),
temporal lobe (left fusiform gyrus), and left postcentral gyrus. Increased BC was observed in
a complement of nodes in the frontal lobe (left superior frontal gyrus, left superior frontal gyrus
Network Neuroscience
196
Learning-induced brain network repertoires in schizophrenia
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
7
1
1
8
4
2
0
7
2
1
3
3
n
e
n
_
a
_
0
0
2
7
8
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
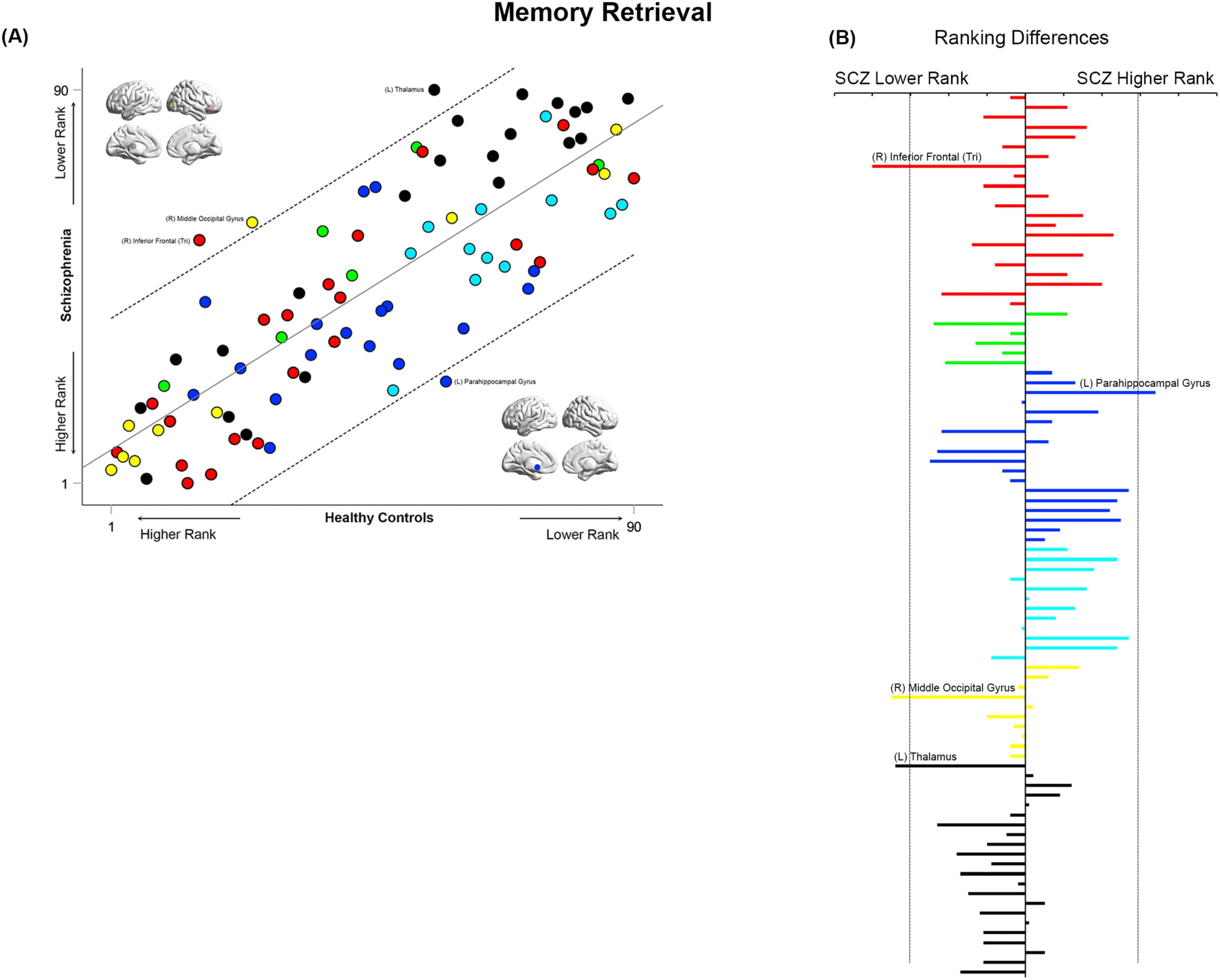
Figure 11. Cross-ranking for Memory Retrieval. (A) Cross-ranking information (consistent with the format in Figures 9–10) is presented for the
Memory Retrieval condition. While node ranks are highly correlated across groups, multiple nodes lay outside the confidence interval of the
regression model. These include the right inferior frontal triangularis, right middle occipital gyrus, and left thalamus (lower rank in SCZ), and
the left parahippocampal gyrus (higher rank in SCZ). (B) The absolute inter-group difference in rank is presented in the same convention as
Figures 9–10.
(orbital), bilateral medial frontal gyrus, and left middle frontal gyrus (orbital)). Increased BC
was also noted in the temporal lobe (left parahippocampal gyrus and left inferior temporal
gyrus), parietal lobe (left inferior parietal lobule), visual cortex (left calcarine sulcus), and
the left olfactory bulb.
Observed effect sizes (Cohen’s d ) were estimated across all analyses presented in
Figures 1–4, to assess any limiting role of the employed sample size (n = 59) on the
observed inter-group differences. A wide range of effect sizes were observed (.001 ≤
Cohen’s d ≤ .83). Notably, 55% of the values lay in the small to medium, or higher range
Network Neuroscience
197
Learning-induced brain network repertoires in schizophrenia
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
7
1
1
8
4
2
0
7
2
1
3
3
n
e
n
_
a
_
0
0
2
7
8
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
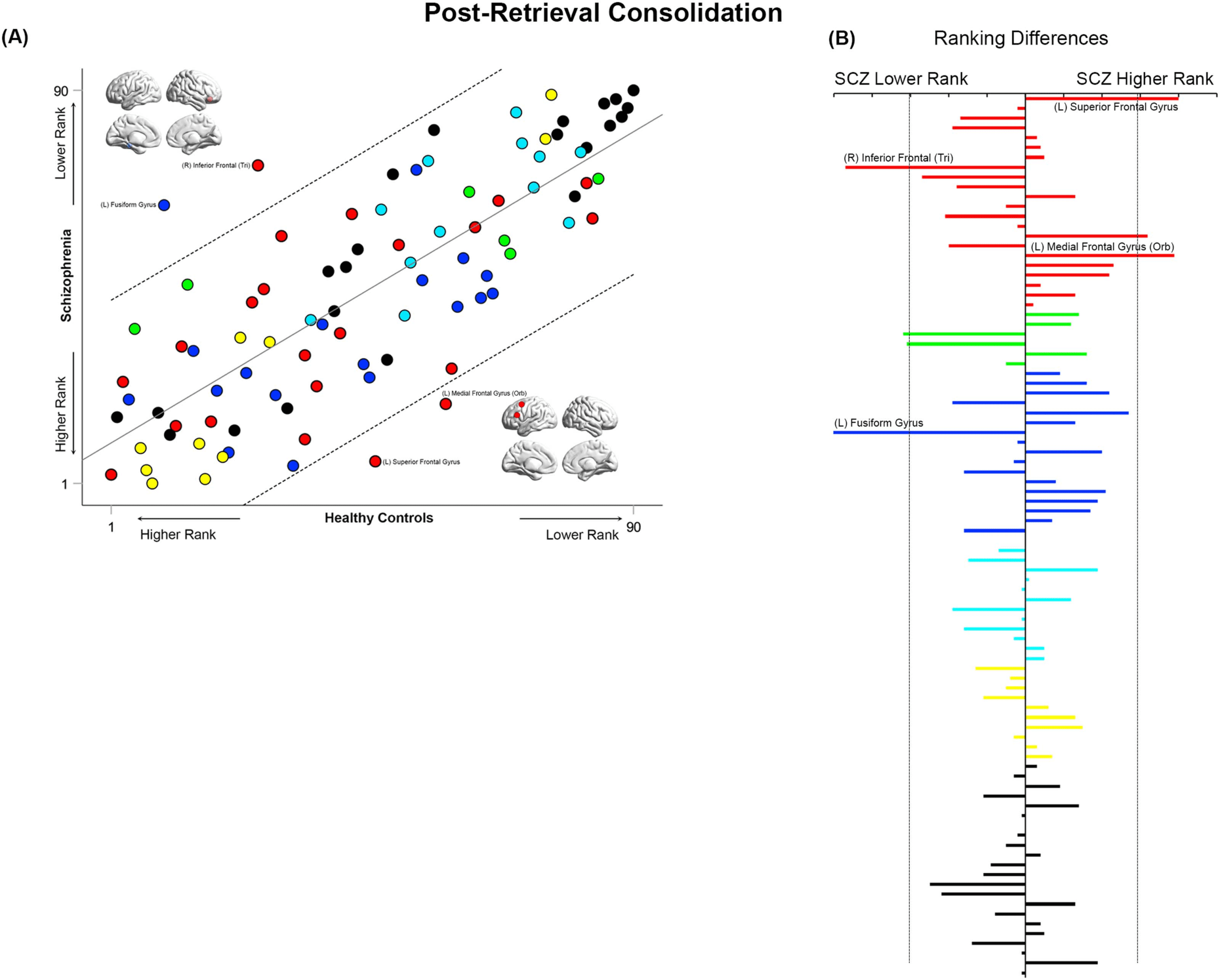
Figure 12. Cross-ranking for Post-Retrieval Consolidation. (A) Cros- ranking information (consistent with the format in Figures 9–11) is pre-
sented for the Post-Retrieval Consolidation condition. While node ranks are highly correlated across groups, multiple nodes lay outside the
confidence interval of the regression model. These include the left fusiform gyrus and right inferior frontal triangularis (lower rank in SCZ), and
the left superior frontal and medial frontal orbital (higher rank in SCZ). (B) The absolute inter-group difference in rank is presented in the same
convention as Figures 9–11.
(Cohen’s d ≥ .2), with 14% lying in the medium to large range (Cohen’s d ≥ .5) (Cohen,
1988).
In further explorations from these main results (Supporting Information Figures S6 and S7), (a)
relationships between behavioral performance and BC (across and within groups), and (b) within
SCZ, relationships between clinical variables and antipsychotic dosage, and BC were explored
using correlational analyses. These analyses were conducted node-wise in each of the four con-
ditions. We observed sparse convergence between nodes with significant relationships in
these exploratory analyses and nodes with significant inter-groups differences (Figures 1–4)
but more focused independent investigations are needed.
Network Neuroscience
198
Learning-induced brain network repertoires in schizophrenia
Analyses of Rankings Based on BC
Initial analyses (Figures 1–4) identified nodes with significant inter-group differences. How-
ever, these results are agnostic about the integrative importance of these nodes within each
participant group and condition. To investigate whether nodes with different BC were more (or
less) important within their network, BC was used to rank nodes in each group and condition.
This ranking is an ordinal measure of the integrative importance of any node. For each con-
dition (Figure 5–8), the nodes are arranged in order of rank, separately for each group. Curves
are used to distinguish between SCZ (black line) and HC data (gray line), with color coding for
the lobes maintained. Only nodes with significant differences (from Figures 1–4) are labeled
(and enlarged for visual access). To simplify appraisal, the space is divided by vertical lines.
These divide the ranking space into quartiles (based on ranking). Finally, heat maps represent
the percentage of significantly different nodes for each direction (HC > SCZ, SCZ > HC) that
fall within each of the four quartiles, and they permit assessment of the relative importance of
nodes with significant differences.
Memory Formation
In SCZ, 80% of nodes with significantly lower BC (relative to HC) were highly ranked (i.e., in
the first and second quartiles) within the respective networks. These included the putamen,
middle temporal, superior frontal orbital, and precuneus. Conversely, 70% of the nodes with
significantly higher BC (relative to HC) were lowly ranked (i.e., in the third or fourth quartiles)
within the network, including nodes such as the inferior frontal orbital, paracentral lobule,
superior occipital lobule, and cuneus.
Post-Encoding Consolidation
As with Memory Formation, 80% of nodes with significantly lower BC in SCZ were highly
ranked (seen solely in the first quartile of ranks), including the inferior occipital, calcarine,
posterior cingulate, putamen, superior frontal, inferior temporal, and Heschl’s gyrus. Con-
versely, a relatively large percentage of nodes (∼65%) with significantly higher BC were lowly
ranked, including the parahippocampal gyrus, superior frontal orbital, inferior frontal orbital,
and insula.
Memory Retrieval
In SCZ, ∼65% of nodes with significantly lower BC were highly ranked (in the first quartile),
including the middle frontal, inferior occipital, Heschl’s gyrus, putamen, and inferior frontal
triangularis. Conversely, 90% of nodes with significantly higher BC were lowly ranked (i.e., in
the third and fourth quartiles) within the network. These include the superior temporal, supra-
marginal gyrus, inferior parietal, superior frontal orbital, cuneus, middle frontal orbital, supe-
rior occipital, and caudate.
Post-Retrieval Consolidation
Only during Post-Retrieval Consolidation was a distinct trend observed. Here, most nodes with
decreases in BC (90%) were highly ranked. These included the middle cingulate, putamen,
fusiform, inferior frontal triangularis and operculum, and superior frontal orbital. Moreover,
70% of nodes with increases in BC were also highly ranked. These included the calcarine,
inferior temporal, superior frontal, posterior cingulate, medial frontal orbital, superior frontal
orbital, and parahippocampal gyrus.
Network Neuroscience
199
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
7
1
1
8
4
2
0
7
2
1
3
3
n
e
n
_
a
_
0
0
2
7
8
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Learning-induced brain network repertoires in schizophrenia
Inter-group Similarities in General Network Topology Based on Node Ranks
How similar are the groups in terms of “general” network topology features? We operational-
ized this question by investigating the inter-group relationships between node rankings. These
rankings provide an ordinal measure of the relative integrative importance of a node within
that group (and condition). Therefore, quantifying the correlations of ranks provide a measure
of inter-group consistency in general network topology. High cross-correlations would suggest
that rank order is largely maintained, and that general network topology is more similar than
different between groups. In such case, it would be particularly important to identify nodes that
are outliers in each correlation model. These questions were investigated using linear regres-
sion models applied to the observed ranks of the 90 nodes in each of the SCZ and HC groups
in each of the four conditions (see Figures 9A–12A). Outliers in the regression models lying
outside the 95% confidence interval of each model are noted for differences in ranking.
Across all conditions, node ranks were highly correlated across groups (all r 2 ≥ 0.63,
F1,88 ≥ 151.6), suggesting that general network topology was more similar than different.
However, in each of the four experimental conditions, specific nodes lay outside the 95%
confidence interval of the model. In each of Figures 9B–12B, the bar graphs represent the
absolute difference in observed rank between groups (the dropped lines represent the 95%
confidence interval in each model).
During Memory Formation (Figure 9), the middle temporal gyrus, superior frontal gyrus
(orbital), precuneus, and olfactory bulb all lay outside the confidence interval, suggesting a
substantially lower ranking in SCZ. By comparison, the hippocampus and middle occipital
gyrus lay outside the confidence interval, suggesting a substantially higher ranking.
During Post-Encoding Consolidation (Figure 10), the inferior temporal gyrus and Heschl’s
gyrus appeared to have a substantially lower ranking in SCZ, whereas the hippocampus had a
higher ranking.
During Memory Retrieval (Figure 11), the inferior frontal triangularis, middle occipital
gyrus, and thalamus had a lower ranking in SCZ, whereas the parahippocampal gyrus had
a higher ranking.
Finally, during Post-Retrieval Consolidation (Figure 12), the fusiform gyrus and inferior fron-
tal triangularis had a lower ranking in SCZ, whereas the superior and medial frontal gyri had a
higher ranking.
Stable and Unstable Ranks Across Conditions
Within any condition and group, a node’s ranking (based on BC) reflects its relative impor-
tance to the functional organization of the network. How stable is this relative importance
across conditions? Addressing this issue allows us to capture some measure of the stability
and instability of network repertoires across task conditions in each of the SCZ and HC
groups.
To estimate this, we first calculated the standard deviation (SD) of each node’s ranking
across conditions, where a low SD represents a high degree of stability in the node’s rank.
In Figure 13A, for each of the HC and SCZ groups, nodes are presented in descending order
of stability, showing that some nodes (e.g., the right hippocampus) are equi-stable, whereas
other nodes (e.g., the left hippocampus) show greatly differing stabilities across groups. Across
all 90 nodes, we next created a relative stability map (Figure 13B) with values (absolute dif-
ference in the stability rank) representing the difference in inter-group stability of that node. For
ease of access we code the markers on the vertical axis to reflect lobe assignments (while also
Network Neuroscience
200
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
7
1
1
8
4
2
0
7
2
1
3
3
n
e
n
_
a
_
0
0
2
7
8
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Learning-induced brain network repertoires in schizophrenia
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
7
1
1
8
4
2
0
7
2
1
3
3
n
e
n
_
a
_
0
0
2
7
8
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
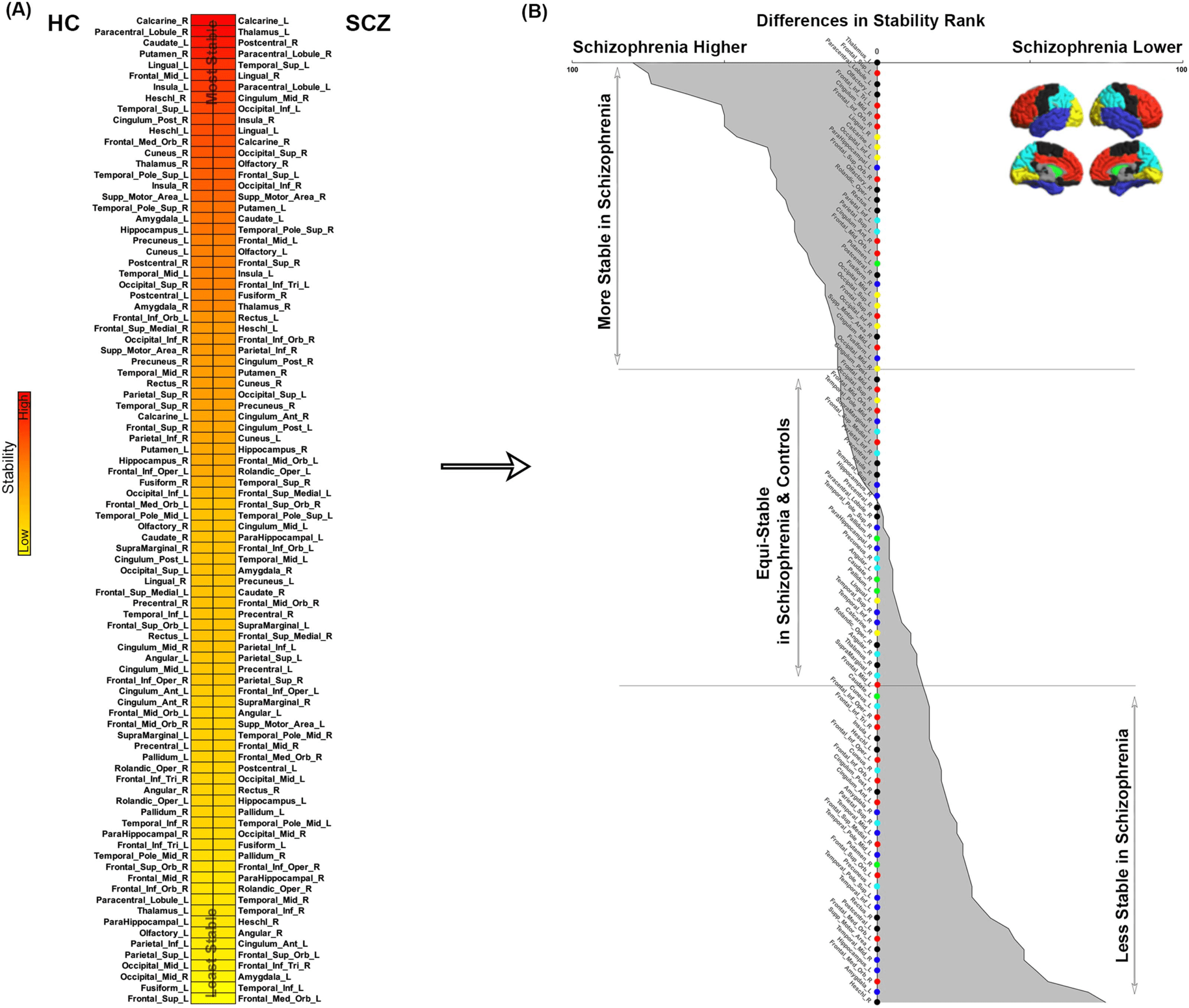
Figure 13. Network stabilities. (A) In each group, nodes are arranged in descending order of the stability of their rank across conditions. As
seen, across groups, the ordering is different suggesting that the stability of node ranks varied greatly by node. (B) To quantify this, across all 90
nodes, we calculated the difference in rank by stability (SCZ minus HC). Resultant values were reordered from most negative (most stable in
schizophrenia) to most positive (least stable in schizophrenia). We plotted the 90 nodes and shaded the area under the curve to reflect the
difference in rank by stability between SCZ and HC (naturally, the relative stability map presents symmetrically). In addition to listing node
identity, for ease of access we also code the vertical access with colors reflecting lobe assignments for each node (see lateral and medial brain
representations). Then, the 90-node space was cleaved (horizontal lines) into tertiles representing in sequence, (a) nodes more stable in SCZ,
(b) nodes that were relatively equi-stable, and (c) nodes less stable in SCZ.
listing node identity). Then, the 90-node space was cleaved (horizontal lines) into tertiles rep-
resenting in sequence, (a) nodes more stable in SCZ, (b) relatively equi-stable nodes and, (c)
nodes less stable in SCZ.
Salient to our interests were nodes that were both highly ranked and highly stable. These
nodes could reasonably be thought to underpin network organization independent of task
Network Neuroscience
201
Learning-induced brain network repertoires in schizophrenia
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
7
1
1
8
4
2
0
7
2
1
3
3
n
e
n
_
a
_
0
0
2
7
8
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
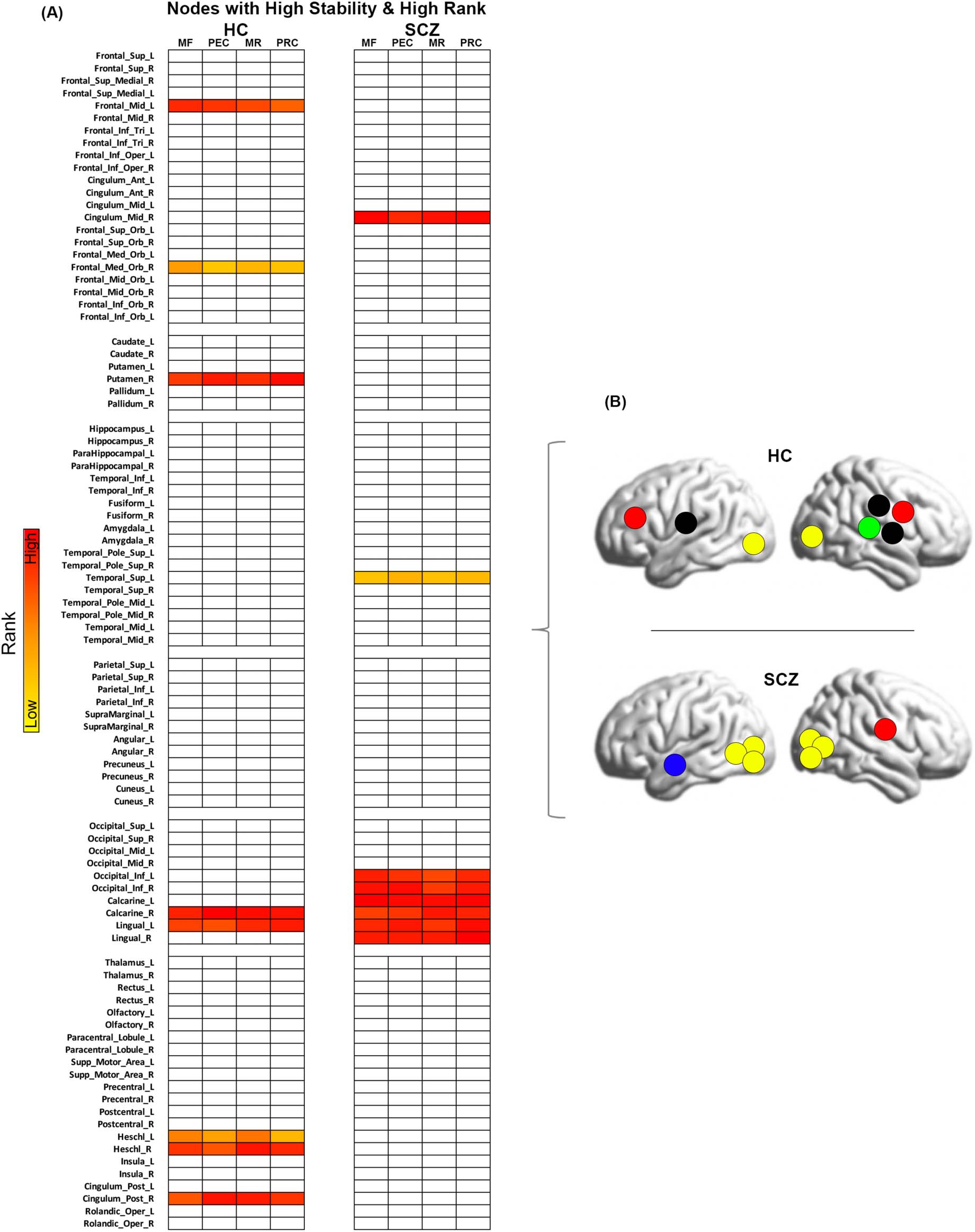
Figure 14. Nodes with high stability and high rank. (A) The figure depicts the nodes that are both highly stable and highly ranked (across the
conditions of Memory Formation (MF), Post-Encoding Consolidation (PEC), Memory Retrieval (MR), and Post-Retrieval Consolidation (PRC)) in
each of the HC and SCZ groups. These nodes are particularly salient to our analyses because they underpin network implementation inde-
pendent of task condition. (B) Lateral brain representations of the network topology (obtained from panel A) for both HC and SCZ are depicted
here. As can be clearly seen, the network stability varies between groups, with SCZ evincing a highly circumscribed organization of nodes and
HC displaying a more distributed network.
Network Neuroscience
202
Learning-induced brain network repertoires in schizophrenia
condition (therefore generally being more “hub” like). Of the 16% of nodes with the lowest SD
(i.e., the most stable in terms of rank), Figure 14 represents those which showed the highest
rank (∼9% of the total nodes). Thus, the figure represents highly stable and highly ranked
nodes in each group. As seen, in HC, these cross-condition “hubs” were distributed across
the cortex and included nodes in the frontal lobe (mid frontal, medial frontal orbital), the puta-
men, the bilateral Heschl’s gyrus, the posterior cingulate, and nodes in the visual lobe (calcar-
ine and lingual). In notable contrast, the highly circumscribed locations of the stable hubs in
schizophrenia primarily lay in the visual lobe (bilateral inferior occipital, bilateral calcarine,
and bilateral lingual gyrus), except for the mid cingulate and superior temporal.
DISCUSSION
Using an associative learning paradigm with multiple conditions, we induced brain network
dynamics in SCZ patients and controls to (a) characterize resultant network repertoires based
on betweenness centrality (BC) (Z. Chen & Calhoun, 2018; Cheng et al., 2015; Lord, Horn,
Breakspear, & Walter, 2012), (b) examine group (HC ≠ SCZ) differences in estimated BC, (c)
assess the relative importance (based on rank ordering by BC) of significantly different nodes
within each group’s network, (d) examine the relationship between the ranks of nodes across
groups, and (e) assess the relative stability of the node rankings across conditions. These inves-
tigations revealed four salient results: (a) inter-group differences in BC were observed across
experimental conditions (even during periods of passive memory consolidation) (Figures 1–4);
(b) nodes with increased BC in SCZ were lowly ranked, whereas nodes with decreased BC in
SCZ were highly ranked (Figures 5–8); (c) in each of the conditions, observed ranks were cor-
related across groups (Figures 9–12), but differed for several task-relevant nodes; (d) finally, in
SCZ, nodes with high rank stability were isolated to early sensory cortex, but in HC, were
distributed across the cerebral cortex (Figures 13–14).
Previous studies of schizophrenia have characterized network topology of resting-state
fMRI data (Cheng et al., 2015; Nijhuis, van Cappellen van Walsum, & Norris, 2013; Oldham
et al., 2019), but increasingly, studies are now attempting to understand the impact of task-
driven contextual processing on disordered functional network topology in schizophrenia.
This task-driven approach is valuable because fMRI signals are maximally responsive to
task-driven modulation (Logothetis, 2008), and tasks amplify expressions of clinical pathology
in networks (Diwadkar & Eickhoff, 2021a). We specifically relied on BC because it can be
estimated from undirected graphs (as in our case) (Dablander & Hinne, 2019), and combines
both degree and path length in its estimation (Bullmore & Sporns, 2009; Zuo et al., 2012).
Finally, in estimating the integrative properties of nodes, BC is particularly well suited for char-
acterizing a dys-connection syndrome like schizophrenia (Medaglia, Lynall, & Bassett, 2015).
In the remainder of the Discussion, we unpack the import of the results, initially focusing on
group differences within each task condition, before discussing observed stability and instabil-
ity of repertoires across task conditions.
Inter-group Differences and Node Rankings
Memory formation is a dynamic process of sensing or perceiving novel information or memo-
randa, that are initiated into the preliminary process of learned associations (Bero et al., 2014). It
has widely distributed cerebral correlates, with nodes in the medial temporal lobe, the prefron-
tal cortex, the striatum, and the thalamus playing highly integrative roles (Pardi et al., 2020;
Squire, Stark, & Clark, 2004). While we observed an admixture of differences in inter-group
effects (SCZ ≠ HC) (Figure 1B), many nodes with increased BC in SCZ were relatively lowly
Network Neuroscience
203
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
7
1
1
8
4
2
0
7
2
1
3
3
n
e
n
_
a
_
0
0
2
7
8
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Learning-induced brain network repertoires in schizophrenia
ranked (in the third and fourth quartiles; see Figure 5), suggesting that nodes with increased
integrative roles were less important. Two notable exceptions were the hippocampus and para-
hippocampal gyrus. All inter-group differences are aberrant, so if the integrative role of these
nodes is greater in patients, why might such a finding result? “Compensatory” increases in fMRI
responses typify the schizophrenia literature (Kim et al., 2010) and may reflect inefficient rather
than adaptive responses (such as those seen in normal aging) (Reuter-Lorenz & Cappell, 2008).
Indeed, increased hippocampal activity has been observed both at rest (Tregellas et al., 2014)
and in tasks ranging from learning (Wadehra, Pruitt, Murphy, & Diwadkar, 2013) to sensory
gating (Tregellas et al., 2007). As Supporting Information Figure S1 shows, patient performance
lagged that of their healthy counterparts, reinforcing this inference.
Effortful Memory Retrieval is initiated by cues from the prefrontal cortex (Simons & Spiers,
2003; Woodcock, White, & Diwadkar, 2015), supplemented by the dorsal striatum. The
retrieval trace interjects with distributed memory representations in regions including the hip-
pocampus (Kragel & Polyn, 2015). Accordingly, the significant decrease in BC in the middle
frontal gyrus (along with the inferior frontal triangularis, the inferior occipital gyrus, Heschl’s
gyrus, and the putamen) (Figure 3) suggests a decreased integrative role during retrieval. More-
over, these nodes were highly ranked, confirming that schizophrenia is compromised by a loss
of the integrative role of regions in the prefrontal cortex that underpins the successful retrieval
of memory traces (Takehara-Nishiuchi, 2021).
A notable observation was that disordered network repertoires in schizophrenia were
evoked even during periods free of overt sensorimotor stimulation or processing (Figures 2,
4, 6, and 8). Rest states within task-driven studies have ongoing and active processes that
are related to the foreground tasks that these states are a part of (Diwadkar, Asemi, Burgess,
Chowdury, & Bressler, 2017; Ravishankar et al., 2019). However, as is known, these covert
repertoires evince unique properties that are inherited from task-active states.
Consolidation drives the recapitulation of as-yet weak memory traces through covert
rehearsal (Lilienthal, Myerson, Abrams, & Hale, 2018) and/or spontaneous brain network reor-
ganization (Malerba & Bazhenov, 2019). Two aspects of our observed group differences are
notable. First, although both consolidation conditions were superficially identical, the evoked
disordered network repertoires were different (Figures 2B vs. Figure 4B). During Post-Encoding
Consolidation, reduced BC was observed in highly ranked nodes, but increased BC was pri-
marily seen in lowly ranked nodes. However, during Post-Retrieval Consolidation nodes with
inter-group differences (Figure 4) were highly ranked (Figure 8), and the locations of these
nodes were (by definition) complementary. In SCZ, the integrative role of multiple nodes in
the frontal lobe was higher, suggesting an exaggerated role in attempting to recapitulate and
integrate inadequately formed memory traces that rely on transient working memory (He et al.,
2012; Yang et al., 2020). Second, disordered repertoires during recapitulation conditions
were only partially overlapping with those observed during preceding task-active conditions
(Figures 1B and 3B, respectively), further evidence that constructive covert psychological
processes drive unique network repertoires (Diwadkar et al., 2017; Jacobacci et al., 2020;
Ravishankar et al., 2019).
Cross-Rankings: Similarities and Differences in Network Repertoires
Analyzing cross-rankings of individual nodes (Figures 9–12) provides a window into consis-
tencies and differences in network repertoires observed in each condition. Node ranks were
highly correlated across groups in each of the conditions. This consistency indicates that even
highly debilitating conditions like schizophrenia are marked by only subtle differences in
Network Neuroscience
204
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
7
1
1
8
4
2
0
7
2
1
3
3
n
e
n
_
a
_
0
0
2
7
8
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Learning-induced brain network repertoires in schizophrenia
network repertoires (Lam, Li, Ke, & Yung, 2022; Sprooten et al., 2017), This subtlety may arise
because (a) unlike long-standing neurologic conditions, neuropsychiatric diseases affect brain
function at its “margins” (Spronk et al., 2021); and (b) thus any observed impacts are contex-
tually evoked (Robison, Thakkar, & Diwadkar, 2020). Nevertheless, in these analyses, the
hippocampus and parahippocampal gyrus showed higher ranks in patients during Memory
Formation and Retrieval, but nodes such as the superior frontal gyrus and the inferior frontal
triangularis showed lower ranks. These effects reemphasize the exaggerated centrality of
medial temporal lobe structures in schizophrenia during active task conditions, and they
highlight the dynamic nature of inter-group differences in network repertoires as driven by
task context. We note that an overwhelming focus on positive results (i.e., significant inter-
group differences), frequently obscures appreciation of the subtle nature of effects, even in
conditions as debilitating as schizophrenia (Bouttier, Duttagupta, Denève, & Jardri, 2022).
The Stability of Node Ranks Across Conditions
The final analyses revealed transitions in response to changing task conditions (Najafi,
McMenamin, Simon, & Pessoa, 2016). When ordered by stability of ranks (Figure 13), equal
numbers of (complimentary) nodes in the prefrontal cortex are more or less stable in patients
(Figure 13B). More salient is evidence (Figure 14) that in healthy controls, highly stable and
highly ranked nodes are distributed across the cortex (Mattar, Cole, Thompson-Schill, & Bassett,
2015). This finding is further evidence that while stable and flexible repertoires are a highlight of
functional adaptation and efficiency in the healthy brain (Liu, Kohn, & Fernandez, 2021;
Morin, Chang, Ma, McGuire, & Stern, 2021; Sporns & Kotter, 2004), this adaptability is imple-
mented at alternative sites in schizophrenia.
Limitations and Conclusions
Capturing network repertoires from an inherently dynamical system like the human brain is
challenging (Park & Friston, 2013), yet the use of well-titrated tasks coupled with the applica-
tion of graph theoretic measures like BC can be enormously useful (Rubinov & Sporns, 2010).
In addition to assessing differences in BC (as we initially did), the measure provides an ordinal
representation of the integrative importance (“hubness”) of nodes in a network. From a rich set
of analytic targets, we arrived at several insights. Generally, cerebral nodes with higher BC in
patients were more lowly ranked (with the opposite being true in healthy controls). This obser-
vation suggests that task implementation drives alternative network topology in schizophrenia.
Remarkably, passive memory consolidation drove altered network topology even in the
absence of overt stimulation or processing. While some aspects of network topology (based
on the cross-ranking of nodes) was similar across groups, meaningful exceptions emerged. And
finally, in controls a cross-cerebral network of nodes showed high rank stability and high rank-
ing, but in SCZ, this similarly classified network was highly localized to early sensory regions.
Such localization is suggestive of an altered reliance on sensorial as opposed to integrative
processing in the illness.
Are the observed effects specific to schizophrenia, and if not, how likely are they to be
observed across other conditions with disordered long-term memorial processing? Impair-
ments in learning and episodic memory cut across disorders of mood, anxiety, and personality
(Brambilla et al., 2011; Carcone, Lee, & Ruocco, 2020; Quraishi & Frangou, 2002; Ruocco &
Bahl, 2014; Sauro, Jorgensen, & Pedlow, 2003), though similarities and/or differences in net-
work repertoires have not been systematically addressed. As with other studies in this domain,
a lack of a clinical control group limits our ability to draw definitive conclusions about the
diagnostic specificity of our results. On a separate point of note, we also cannot definitively
Network Neuroscience
205
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
7
1
1
8
4
2
0
7
2
1
3
3
n
e
n
_
a
_
0
0
2
7
8
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Learning-induced brain network repertoires in schizophrenia
conclude that disordered network topology in schizophrenia is tied to the specific task used.
Generally, brain network repertoires reflect task-evoked profiles, one reason for why structural
brain network connectivity underpredicts functional connectivity (Park & Friston, 2013).
Indeed, the contextually (i.e., condition) driven bases of our results suggest that schizophrenia
may be better characterized as a syndrome with a propensity for task-evoked dys-connection,
rather than simply as a dys-connection syndrome. However, within-participant comparisons
using a multiplicity of datasets would be needed to reach such conclusions. Across nodes, BC
estimates (regardless of task condition), were not particularly well predicted by behavioral per-
formance (Supporting Information Figure S6) or medication dosage (patients) (Supporting
Information Figure S7). This sparsity suggests that estimates of the integrative value of nodes
were uncoupled from participant’s task proficiency or “how” medicated patients were. These
interrelationships will probably need to be investigated in more focused and systematic anal-
yses. Finally, our estimates of effect size suggest that our sample size was viable for identifying
inter-group differences, our sample was admittedly smaller than several other task-based stud-
ies in schizophrenia (and certainly smaller than resting-state studies).
The road toward long-term memory consolidation is “long and winding” (Eichenbaum,
2001). Because such consolidation is undeniably compromised in schizophrenia, learning
and memory is the subject of multiple intervention strategies (Manoach, Mylonas, & Baxter,
2020). Laboratory tasks are of limited ecological validity in addressing such fundamental ques-
tions. However, concurrently with functional neuroimaging, such tasks can be deployed to
provide insights into disordered network repertoires that underlie associative learning and
memory in schizophrenia. More than activation-based measures, complex graph theoretic
metrics that summarize contextually driven network repertoires may be more useful biomark-
ers of the illness and/or of treatment efficacy (Diwadkar & Eickhoff, 2021b).
METHODS
Participants
Wayne State University’s institutional review board approved all procedures. We collected
functional-MRI (fMRI) data from 59 participants (32 stable SCZ, 39 male, 20 female; mean
age: HC, 28.02 ± 6.69; SCZ, 29.98 ± 8.38) recruited from the greater Detroit area through
local advertisements. Participants provided informed consent and subsequently received
remuneration for their involvement. SCZ patients were identified through their treating physi-
cians, and the diagnosis was confirmed by a research psychologist using DSM-V criteria for
SCZ (American Psychiatric Association, 2013). All patients were maintained on a regimen of
atypical antipsychotics (risperidone, olanzapine, or aripiprazole). Clinical symptom severity
ratings were assessed using the PANSS (Kay et al., 1987). General intelligence was assessed
using the Wechsler Abbreviated Scale of Intelligence (Psychological Corporation, 1999). The
duration of illness was estimate from the most likely date of onset of psychotic symptoms
(hallucinations, delusions, or disorganization of thinking; bizarre or catatonic behavior) and
date of diagnosis for SCZ, using all available information (medical records, reports by family
members or significant others, and the Structured Clinical Interview for DSM Disorders
interview). HC participants were free of Axis-I psychopathology (past/present). Participants
were screened prior to entering the study to exclude any significant past/current medical
and/or neurological illness (e.g., hypertension, thyroid disease, diabetes, asthma requiring
prophylaxis, seizures, or significant head injury with loss of consciousness). The two groups
did not differ in age or gender distribution. Demographic data, clinical characterization, and
medication lists (patients) are shown in Table 1.
Network Neuroscience
206
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
7
1
1
8
4
2
0
7
2
1
3
3
n
e
n
_
a
_
0
0
2
7
8
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Learning-induced brain network repertoires in schizophrenia
Table 1. Demographics, medication, and clinical characteristics
SCZ (n = 32)
HC (n = 27)
Demographics
Age (years)
Sex (% female)
IQ
Clinical information
Duration of illness (years)
PANSS
29.98 ± 8.4
10 (31%)
89.9 ± 10.7
7.8 ± 6.1
51.2 ± 9.7
28.02 ± 6.7
10 (37%)
91.9 ± 13.0
Note. The table provides demographic and clinical information for the group of 59 participants. All patients
were stabilized on a regime of antipsychotics. In addition, two (6%) were prescribed antidepressants, eight
(25%) anxiolytics, and seven (22%) mood stabilizers. Clinical symptom severity was assessed using the Positive
and Negative Syndrome Scale (PANSS) (Kay et al., 1987) (total PANSS is reported). The duration of illness was
derived from the most likely date of onset of psychotic symptoms (hallucinations, delusions, or disorganization
of thinking; bizarre or catatonic behavior) using all clinical information, including medical records, reports by
family members or significant others, and the Structured Clinical Interview for DSM Disorders interview.
MRI Acquisition
Data (3 T Siemens Verio scanner, 32-channel volume head coil) were acquired using a
multiband gradient EPI sequence (TR = 3 s, TE = 24.6 s, multiband factor = 3, FOV = 192 ×
192 mm2, matrix = 96 × 96, 64 axial slices, resolution = 2 mm3). T1-weighted MRI images were
collected for normalization and coregistration with the EPI scan (3D magnetization-prepared
rapid gradient-echo sequence, TR = 2,150ms, TE = 3.5 ms, TI = 1,100ms, flip angle = 8°,
FOV = 256 × 256 × 160 mm3, 160 axial slices, resolution = 1mm3).
Associative Learning and Memory Paradigm
The specifically curated associative learning task cycled through four successive conditions:
Memory Formation (Encoding), Post-Encoding Consolidation, Memory Retrieval, and Post-
Retrieval Consolidation (27-s conditions each). During Memory Formation, nine objects each
uniquely associated with a location within a 3 × 3 spatial grid were presented for naming and
encoding (3 s/pair). A stimulus-free (and instruction-free) Post-Encoding Consolidation condition
followed (Ravishankar et al., 2019). Following Post-Encoding Consolidation, memory for the
nine object-location pairs was tested in a Memory Retrieval condition during which each of
the nine grid locations were cued in random order. Participants were required to name the object
associated with the location (or utter “no” if they could not recall the object). Finally, a Post-
Retrieval Consolidation period followed (which was identical in its form to the Post-Encoding
Consolidation period). The task was conducted without feedback, and eight epochs of this
sequence of conditions were employed to promote asymptotic performance (a schematic of
the task along with observed behavioral data is provided in Supporting Information Figure S1).
fMRI Data Processing and Time Series
fMRI data were processed in SPM 12 using standard temporal (slice-time correction) and spa-
tial preprocessing methods. For spatial preprocessing, EPI images were oriented to the AC-PC
line, corrected for head movement through realignment to a reference image in the sequence,
and coregistered to the anatomical high-resolution T1 image. Analyses of the displacement
parameters indicated that estimated head movement did not differ between groups (.14 ≤
Network Neuroscience
207
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
7
1
1
8
4
2
0
7
2
1
3
3
n
e
n
_
a
_
0
0
2
7
8
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Learning-induced brain network repertoires in schizophrenia
p ≤ .98). The deformations from normalizing the high-resolution T1 image were applied to the
coregistered EPI images to thus normalize the volumes to stereotactic space. A low-pass filter
(128 s) corrected for low-frequency components. At the first level, epochs were modeled with
boxcar stimulus functions convolved with a canonical hemodynamic response function to
form regressors of interest. In each first level model, the six motion parameters (3 for translation
and 3 for rotation) from the coregistration were modeled as covariates of no interest. Images
were resliced (2 mm3) and a Gaussian filter (8 mm FWHM) applied. Images exceeding 4 mm
of movement (<1% of all images) were excised from analyses. The Automated Anatomical
Labeling (AAL) atlas was used to identify 90 cerebral regions (nodes) in the a priori network
(Rolls et al., 2020).
Undirected Functional Connectivity
Mean time series were extracted from the 90 cerebral regions for each participant and for each
of the four conditions of the task. The bivariate correlations across the full matrix were com-
puted from the time series data in each condition. For each participant, resultant symmetric
90 × 90 adjacency matrices of correlation coefficients (one for each condition of the task)
were obtained. These coefficients (Pearson’s r) were normalized with the Fisher Z-
transformation (Equation 1) to ensure the variance is independent of the magnitude of the
correlation coefficients (Thompson & Fransson, 2016). The Z-values were used for the calcula-
tion of graph theoretic metrics.
(cid:1)
(cid:3)
z ¼
1
2
ln
1 þ r
1 − r
(1)
Supporting Information Figures S2–S5 provide the distance connectomes for each of the
four conditions (separately for each of the HC and SCZ groups).
Betweenness Centrality Analyses and Computation
BC estimates the number of shortest functional paths that traverse through a node. The length
between nodes is reflected in the weighted measure of the Fisher Z-transformation, which
calculates the functional proximity between nodes. Through these estimates, BC represents
a node’s role in transmitting and facilitating interactions (van den Heuvel et al., 2010). The
BC of a node in the network was computed based on the following formula (Equation 2).
BC weighted ¼
1
Þ
Þ n − 2
ð
ð
n − 1
Σh≠i
sphj ið Þ
sphj
(2)
The expression sphj(i) indicates the number of shortest paths between nodes h and j that passes
through node i. BC was estimated in each participant across the 90 cerebral nodes and for
each condition. The resultant BC values were forwarded for statistical analyses. Two-sample
t tests were conducted on the BC values for each node in each condition to study inter-group
differences. All patient-control comparisons were thresholded (pFDR < 0.05) (Benjamini &
Hochberg, 1995) across the 360 conducted tests, to identify nodes with significant inter-group
differences.
ACKNOWLEDGMENTS
We acknowledge the help of all the participants (patients and controls) and of participant
families in the conduct of this research.
Network Neuroscience
208
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
1
8
4
2
0
7
2
1
3
3
n
e
n
_
a
_
0
0
2
7
8
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Learning-induced brain network repertoires in schizophrenia
SUPPORTING INFORMATION
Supporting Information for this article is available at https://doi.org/10.1162/netn_a_00278.
AUTHOR CONTRIBUTIONS
Emmanuel D. Meram: Conceptualization; Formal analysis; Investigation; Visualization;
Writing – original draft; Writing – review & editing. Shahira Baajour: Conceptualization;
Methodology; Resources; Software. Asadur Chowdury: Conceptualization; Formal analysis;
Methodology; Resources; Software. John Kopchick: Methodology; Software; Writing – review
& editing. Patricia Thomas: Investigation; Project administration. Usha Rajan: Investigation;
Project administration; Validation. Dalal Khatib: Data curation. Caroline Zajac-Benitez:
Project administration. Luay Haddad: Data curation. Alireza Amirsadri: Data curation;
Resources. Jeffrey A. Stanley: Funding acquisition; Investigation; Methodology; Software.
Vaibhav A. Diwadkar: Conceptualization; Formal analysis; Funding acquisition; Investigation;
Methodology; Project administration; Resources; Software; Supervision; Writing – original
draft; Writing – review & editing.
FUNDING INFORMATION
Vaibhav A. Diwadkar, National Institute of Mental Health (https://dx.doi.org/10.13039
/100000025), Award ID: MH111177. Vaibhav A. Diwadkar, Ethel and James Flinn Foundation
(https://dx.doi.org/10.13039/100005161). Vaibhav A. Diwadkar, DMC Foundation (https://dx
.doi.org/10.13039/100017538). Vaibhav A. Diwadkar, Cohen Neuroscience Endowment.
Vaibhav A. Diwadkar, Jack Dorsey Endowment. Vaibhav A. Diwadkar, Lycaki-Young Funds
from the State of Michigan.
REFERENCES
Alexander-Bloch, A. F., Gogtay, N., Meunier, D., Birn, R., Clasen,
L., Lalonde, F., … Bullmore, E. T. (2010). Disrupted modularity
and local connectivity of brain functional networks in
childhood-onset schizophrenia. Frontiers in Systems Neurosci-
ence, 4, 147. https://doi.org/10.3389/fnsys.2010.00147,
PubMed: 21031030
American Psychiatric Association. (2013). Diagnostic and statistical
manual of mental disorders disorders (5th ed.). Arlington, VA:
American Psychiatric Press.
Baajour, S. J., Chowdury, A., Thomas, P., Rajan, U., Khatib, D.,
Zajac-Benitez, C., … Diwadkar, V. A. (2020). Disordered direc-
tional brain network interactions during learning dynamics in
schizophrenia revealed by multivariate autoregressive models.
Human Brain Mapping, 41(13), 3594–3607. https://doi.org/10
.1002/hbm.25032, PubMed: 32436639
Bavelas, A. (1950). Communication patterns in task-oriented
groups. Journal of the Acoustical Society of America, 22,
725–730. https://doi.org/10.1121/1.1906679
Benjamini, Y., & Hochberg, Y. (1995). Controlling the false discov-
ery rate: A practical and powerful approach to multiple testing.
Journal of the Royal Statistical Society, 57(1), 289–300. https://doi
.org/10.1111/j.2517-6161.1995.tb02031.x
Bero, A. W., Meng, J., Cho, S., Shen, A. H., Canter, R. G., Ericsson, M.,
& Tsai, L. H. (2014). Early remodeling of the neocortex upon
episodic memory encoding. Proceedings of the National Academy
of Sciences of the United States of America, 111(32), 11852–11857.
https://doi.org/10.1073/pnas.1408378111, PubMed: 25071187
Bitsch, F., Berger, P., Nagels, A., Falkenberg, I., & Straube, B.
(2021). Characterizing the theory of mind network in schizophre-
nia reveals a sparser network structure. Schizophrenia Research,
228, 581–589. https://doi.org/10.1016/j.schres.2020.11.026,
PubMed: 33229225
Bouttier, V., Duttagupta, S., Denève, S., & Jardri, R. (2022). Circular
inference predicts nonuniform overactivation and dysconnectivity
in brain-wide connectomes. Schizophrenia Research, 245, 59–67.
https://doi.org/10.1016/j.schres.2020.12.045, PubMed: 33618940
Brambilla, P., Cerruti, S., Bellani, M., Ferro, A., Marinelli, V.,
Giusto, D., … Diwadkar, V. A. (2011). Shared impairment in asso-
ciative learning in schizophrenia and bipolar disorder. Progress
in Neuro-Psychopharmacology & Biological Psychiatry, 35,
1093–1099. https://doi.org/10.1016/j.pnpbp.2011.03.007,
PubMed: 21420463
Brambilla, P., Riva, M. A., Melcangi, R., & Diwadkar, V. A. (2007).
The role of glutamatergic pathways in schizophrenia: From ani-
mal models to human imaging studies. Clinical Neuropsychiatry,
4, 199–207.
Büchel, C., Coull, J. T., & Friston, K. J. (1999). The predictive value
of changes in effective connectivity for human learning. Science,
Network Neuroscience
209
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
1
8
4
2
0
7
2
1
3
3
n
e
n
_
a
_
0
0
2
7
8
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Learning-induced brain network repertoires in schizophrenia
283(5407), 1538–1541. https://doi.org/10.1126/science.283
.5407.1538, PubMed: 10066177
Bullmore, E., & Sporns, O. (2009). Complex brain networks: Graph
theoretical analysis of structural and functional systems. Nature
Reviews Neuroscience, 10(3), 186–198. https://doi.org/10.1038
/nrn2575, PubMed: 19190637
Carcone, D., Lee, A. C. H., & Ruocco, A. C. (2020). Disrupted rela-
tionship between hippocampal activation and subsequent mem-
ory performance in borderline personality disorder. Journal of
Affective Disorders, 274, 1041–1048. https://doi.org/10.1016/j
.jad.2020.05.050, PubMed: 32663930
Chen, J., Muller, V. I., Dukart, J., Hoffstaedter, F., Baker, J. T.,
Holmes, A. J., … Patil, K. R. (2021). Intrinsic connectivity patterns
of task-defined brain networks allow individual prediction of
cognitive symptom dimension of schizophrenia and are linked
to molecular architecture. Biological Psychiatry, 89(3), 308–319.
https://doi.org/10.1016/j.biopsych.2020.09.024, PubMed:
33357631
Chen, Z., & Calhoun, V. (2018). Effect of spatial smoothing on task
fMRI ICA and functional connectivity. Frontiers in Neuroscience,
12, 15. https://doi.org/10.3389/fnins.2018.00015, PubMed:
29456485
Cheng, H., Newman, S., Goni, J., Kent, J. S., Howell, J., Bolbecker,
A., … Hetrick, W. P. (2015). Nodal centrality of functional net-
work in the differentiation of schizophrenia. Schizophrenia
Research, 168(1–2), 345–352. https://doi.org/10.1016/j.schres
.2015.08.011, PubMed: 26299706
Cohen, J. (1988). Statistical power analysis for behavioral sciences
(2nd ed.). Hillsdale, NJ: Laurence Erlbaum Associates.
Dablander, F., & Hinne, M. (2019). Node centrality measures are
a poor substitute for causal inference. Scientific Reports, 9(1),
6846. https://doi.org/10.1038/s41598-019-43033-9, PubMed:
31048731
Deng, M., Liu, Z., Zhang, W., Wu, Z., Cao, H., Yang, J., &
Palaniyappan, L. (2022). Associations between polygenic risk,
negative symptoms, and functional connectome topology during
a working memory task in early-onset schizophrenia. Schizo-
phrenia, 8(1), 54. https://doi.org/10.1038/s41537-022-00260-w,
PubMed: 35853905
Diwadkar, V. A., Asemi, A., Burgess, A., Chowdury, A., & Bressler,
S. L. (2017). Potentiation of motor sub-networks for motor control
but not working memory: Interaction of dACC and SMA revealed
by resting-state directed functional connectivity. PLoS One,
12(3), e0172531. https://doi.org/10.1371/journal.pone
.0172531, PubMed: 28278267
Diwadkar, V. A., & Eickhoff, S. B. (2021a). Brain network dyscon-
nection in neuropsychiatric disorders: The practice of “normal
science”. In V. A. Diwadkar & S. B. Eickhoff (Eds.), Brain network
dysfunction in neuropsychiatric illness: Methods, applications,
implications (pp. 1–15). New York, NY: Springer International
Publishing. https://doi.org/10.1007/978-3-030-59797-9_1
Diwadkar, V. A., & Eickhoff, S. B. (2021b). Brain network dysfunc-
tion in neuropsychiatric illness: Methods, applications and impli-
cations. New York, NY: Springer Nature. https://doi.org/10.1007
/978-3-030-59797-9
Diwadkar, V. A., Flaugher, B., Jones, T., Zalanyi, L., Ujfalussy, B.,
Keshavan, M. S., & Erdi, P. (2008). Impaired associative learning
in schizophrenia: Behavioral and computational studies.
Cognitive Neurodynamics, 2(3), 207–219. https://doi.org/10
.1007/s11571-008-9054-0, PubMed: 19003486
Eichenbaum, H. (2001). The long and winding road to memory
consolidation. Nature Neuroscience, 4(11), 1057–1058. https://
doi.org/10.1038/nn1101-1057, PubMed: 11687811
Farahani, F. V., Karwowski, W., & Lighthall, N. R. (2019). Applica-
tion of graph theory for identifying connectivity patterns in
human brain networks: A systematic review. Frontiers in Neuro-
science, 13, 585. https://doi.org/10.3389/fnins.2019.00585,
PubMed: 31249501
Friston, K. J., Brown, H. R., Siemerkus, J., & Stephan, K. E. (2016).
The dysconnection hypothesis (2016). Schizophrenia Research,
176(2–3), 83–94. https://doi.org/10.1016/j.schres.2016.07.014,
PubMed: 27450778
He, H., Sui, J., Yu, Q., Turner, J. A., Ho, B. C., Sponheim, S. R., …
Calhoun, V. D. (2012). Altered small-world brain networks in
schizophrenia patients during working memory performance.
PLoS One, 7(6), e38195. https://doi.org/10.1371/journal.pone
.0038195, PubMed: 22701611
Jacobacci, F., Armony, J. L., Yeffal, A., Lerner, G., Amaro, E., Jr.,
Jovicich, J., … Della-Maggiore, V. (2020). Rapid hippocampal
plasticity supports motor sequence learning. Proceedings of the
National Academy of Sciences of the United States of America,
11 7 ( 3 8 ) , 2 3 89 8 – 2 3 9 0 3 . h t t p s : / / d o i . o rg / 1 0 . 1 0 7 3 / p n a s
.2009576117, PubMed: 32900965
Ji, J. L., Diehl, C., Schleifer, C., Tamminga, C. A., Keshavan, M. S.,
Sweeney, J. A., … Anticevic, A. (2019). Schizophrenia exhibits
bi-directional brain-wide alterations in cortico-striato-cerebellar
circuits. Cerebral Cortex, 29(11), 4463–4487. https://doi.org/10
.1093/cercor/bhy306, PubMed: 31157363
Kay, S. R., Fiszbein, A., & Opler, L. A. (1987). The positive and neg-
ative syndrome scale (PANSS) for schizophrenia. Schizophrenia
Bulletin, 13(2), 261–276. https://doi.org/10.1093/schbul/13.2
.261, PubMed: 3616518
Kim, M. A., Tura, E., Potkin, S. G., Fallon, J. H., Manoach, D. S.,
Calhoun, V. D., … Turner, J. A. (2010). Working memory circuitry
in schizophrenia shows widespread cortical inefficiency and
compensation. Schizophrenia Research, 117(1), 42–51. https://
doi.org/10.1016/j.schres.2009.12.014, PubMed: 20096539
Kivimaki, I., Lebichot, B., Saramaki, J., & Saerens, M. (2016). Two
betweenness centrality measures based on randomized shortest
paths. Scientific Reports, 6, 19668. https://doi.org/10.1038
/srep19668, PubMed: 26838176
Kragel, J. E., & Polyn, S. M. (2015). Functional interactions
between large-scale networks during memory search. Cerebral
Cortex, 25(3), 667–679. https://doi.org/10.1093/cercor/bht258,
PubMed: 24084128
Lam, Y.-S., Li, J., Ke, Y., & Yung, W.-H. (2022). Variational dimen-
sions of cingulate cortex functional connectivity and implications
in neuropsychiatric disorders. Cerebral Cortex, 32(24), 5682–5697.
https://doi.org/10.1093/cercor/bhac045, PubMed: 35193144
Lei, D., Pinaya, W. H. L., van Amelsvoort, T., Marcelis, M., Donohoe,
G., Mothersill, D. O., … Mechelli, A. (2020). Detecting schizo-
phrenia at the level of the individual: Relative diagnostic value
of whole-brain images, connectome-wide functional connec-
tivity and graph-based metrics. Psychological Medicine, 50(11),
1852–1861. https://doi.org/10.1017/S0033291719001934,
PubMed: 31391132
Network Neuroscience
210
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
1
8
4
2
0
7
2
1
3
3
n
e
n
_
a
_
0
0
2
7
8
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Learning-induced brain network repertoires in schizophrenia
Lilienthal, L., Myerson, J., Abrams, R. A., & Hale, S. (2018). Effects
of environmental support on overt and covert visuospatial
rehearsal. Memory, 26(8), 1042–1052. https://doi.org/10.1080
/09658211.2018.1462390, PubMed: 29667476
Liu, W., Kohn, N., & Fernandez, G. (2021). Dynamic transitions
between neural states are associated with flexible task switching
during a memory task. Journal of Cognitive Neuroscience, 33(12),
2559–2588. https://doi.org/10.1162/jocn_a_01779, PubMed:
34644388
Logothetis, N. K. (2008). What we can do and what we cannot do
with fMRI. Nature, 453(7197), 869–878. https://doi.org/10.1038
/nature06976, PubMed: 18548064
Lord, A., Horn, D., Breakspear, M., & Walter, M. (2012). Changes in
community structure of resting state functional connectivity in
unipolar depression. PLoS One, 7(8), e41282. https://doi.org/10
.1371/journal.pone.0041282, PubMed: 22916105
Malerba, P., & Bazhenov, M. (2019). Circuit mechanisms of hippo-
campal reactivation during sleep. Neurobiology of Learning and
Memory, 160, 98–107. https://doi.org/10.1016/j.nlm.2018.04
.018, PubMed: 29723670
Manoach, D. S., Mylonas, D., & Baxter, B. (2020). Targeting sleep
oscillations to improve memory in schizophrenia. Schizophrenia
Research, 221, 63–70. https://doi.org/10.1016/j.schres.2020.01
.010, PubMed: 32014359
Mattar, M. G., Cole, M. W., Thompson-Schill, S. L., & Bassett, D. S.
(2015). A functional cartography of cognitive systems. PLoS
Computational Biology, 11(12), e1004533. https://doi.org/10
.1371/journal.pcbi.1004533, PubMed: 26629847
Medaglia, J. D., Lynall, M. E., & Bassett, D. S. (2015). Cognitive network
neuroscience. Journal of Cognitive Neuroscience, 27(8),
1471–1491. https://doi.org/10.1162/jocn_a_00810, PubMed:
25803596
Morin, T. M., Chang, A. E., Ma, W., McGuire, J. T., & Stern, C. E.
(2021). Dynamic network analysis demonstrates the formation of
stable functional networks during rule learning. Cerebral Cortex,
31(12), 5511–5525. https://doi.org/10.1093/cercor/ bhab175,
PubMed: 34313717
Najafi, M., McMenamin, B. W., Simon, J. Z., & Pessoa, L. (2016).
Overlapping communities reveal rich structure in large-scale
brain networks during rest and task conditions. NeuroImage,
135, 92–106. https://doi.org/10.1016/j.neuroimage.2016.04
.054, PubMed: 27129758
Nijhuis, E. H. J., van Cappellen van Walsum, A.-M., & Norris, D. G.
(2013). Topographic hub maps of the human structural neocorti-
cal network. PLoS One, 8(6), e65511. https://doi.org/10.1371
/journal.pone.0065511, PubMed: 23935801
Nosè, M., & Barbui, C. (2008). A simple approach to manage dosages
in drug-epidemiology research. Epidemiologia e Psichiatria Sociale,
17(3), 186–187. https://doi.org/10.1017/S1121189X00001263,
PubMed: 18924556
Nosè, M., Tansella, M., Thornicroft, G., Schene, A., Becker, T.,
Veronese, A., … Barbui, C. (2008). Is the defined daily dose sys-
tem a reliable tool for standardizing antipsychotic dosages? Inter-
national Clinical Psychopharmacology, 23(5), 287–290. https://
doi.org/10.1097/ YIC.0b013e328303ac75, PubMed: 18703938
Oldham, S., Fulcher, B., Parkes, L., Arnatkevic Iute, A., Suo, C., &
Fornito, A. (2019). Consistency and differences between central-
ity measures across distinct classes of networks. PLoS One, 14(7),
e0220061. https://doi.org/10.1371/journal.pone.0220061,
PubMed: 31348798.
Oliver, L. D., Hawco, C., Homan, P., Lee, J., Green, M. F., Gold,
J. M., … SPINS Group. (2021). Social cognitive networks and
social cognitive performance across individuals with schizophre-
nia spectrum disorders and healthy control participants. Biolog-
ical Psychiatry: Cognitive Neuroscience and Neuroimaging,
6(12), 1202–1214. https://doi.org/10.1016/j.bpsc.2020.11.014,
PubMed: 33579663
Pardi, M. B., Vogenstahl, J., Dalmay, T., Spano, T., Pu, D. L.,
Naumann, L. B., … Letzkus, J. J. (2020). A thalamocortical top-down
circuit for associative memory. Science, 370(6518), 844–848.
https://doi.org/10.1126/science.abc2399, PubMed: 33184213
Park, H.-J., & Friston, K. (2013). Structural and functional brain net-
works: From connections to cognition. Science, 342(6158),
1238411. https://doi.org/10.1126/science.1238411, PubMed:
24179229
Psychological Corporation. (1999). Wechsler Abbreviated Scale of
Intelligence ( WASI) manual. San Antonio, TX: Psychological
Corporation.
Quraishi, S., & Frangou, S. (2002). Neuropsychology of bipolar dis-
order: A review. Journal of Affective Disorders, 72(3), 209–226.
https://doi.org/10.1016/S0165-0327(02)00091-5
Ravishankar, M., Morris, A., Burgess, A., Khatib, D., Stanley, J. A., &
Diwadkar, V. A. (2019). Cortical-hippocampal functional con-
nectivity during covert consolidation sub-serves associative
learning: Evidence for an active “rest” state. Brain and Cognition,
131, 45–55. https://doi.org/10.1016/j.bandc.2017.10.003,
PubMed: 29054542
Reuter-Lorenz, P. A., & Cappell, K. A. (2008). Neurocognitive aging
and the compensation hypothesis. Current Directions in Psycho-
logical Science, 17(3), 177–182. https://doi.org/10.1111/j.1467
-8721.2008.00570.x
Robison, A. J., Thakkar, K. N., & Diwadkar, V. A. (2020). Cognition
and reward circuits in schizophrenia: Synergistic, not separate.
Biological Psychiatry, 87(3), 204–214. https://doi.org/10.1016/j
.biopsych.2019.09.021, PubMed: 31733788
Rolls, E. T., Cheng, W., Gilson, M., Gong, W., Deco, G., Lo, C. Z.,
… Feng, J. (2020). Beyond the disconnectivity hypothesis of
schizophrenia. Cerebral Cortex, 30(3), 1213–1233. https://doi
.org/10.1093/cercor/bhz161, PubMed: 31381086
Rubinov, M., & Sporns, O. (2010). Complex network measures of
brain connectivity: Uses and interpretations. NeuroImage, 52(3),
1059–1069. https://doi.org/10.1016/j.neuroimage.2009.10.003,
PubMed: 19819337
Ruocco, A. C., & Bahl, N. (2014). Material-specific discrepancies in
verbal and visual episodic memory in borderline personality dis-
order. Psychiatry Research, 220(1–2), 694–697. https://doi.org/10
.1016/j.psychres.2014.07.010, PubMed: 25064386
Sauro, M. D., Jorgensen, R. S., & Pedlow, C. T. (2003). Stress, glu-
cocorticoids, and memory: A meta-analytic review. Stress, 6(4),
235–245. https://doi.org/10.1080/10253890310001616482,
PubMed: 14660056
Silverstein, B., Bressler, S., & Diwadkar, V. A. (2016). Inferring the
dysconnection syndrome in schizophrenia: Interpretational con-
siderations on methods for the network analyses of fMRI data.
Frontiers in Psychiatry, 7, 132. https://doi.org/10.3389/fpsyt
.2016.00132, PubMed: 27536253
Network Neuroscience
211
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
1
8
4
2
0
7
2
1
3
3
n
e
n
_
a
_
0
0
2
7
8
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Learning-induced brain network repertoires in schizophrenia
Simons, J. S., & Spiers, H. J. (2003). Prefrontal and medial temporal
lobe interactions in long-term memory. Nature Reviews Neuro-
science, 4(8), 637–648. https://doi.org/10.1038/nrn1178,
PubMed: 12894239
Sporns, O., & Kotter, R. (2004). Motifs in brain networks. PLoS Biol-
ogy, 2(11), e369. https://doi.org/10.1371/journal.pbio.0020369,
PubMed: 15510229
Spronk, M., Keane, B. P., Ito, T., Kulkarni, K., Ji, J. L., Anticevic, A.,
& Cole, M. W. (2021). A Whole-brain and cross-diagnostic
perspective on functional brain network dysfunction. Cerebral
Cortex, 31(1), 547–561. https://doi.org/10.1093/cercor
/bhaa242, PubMed: 32909037
Sprooten, E., Rasgon, A., Goodman, M., Carlin, A., Leibu, E., Lee,
W. H., & Frangou, S. (2017). Addressing reverse inference in
psychiatric neuroimaging: Meta-analyses of task-related brain
activation in common mental disorders. Human Brain Mapping,
38(4), 1846–1864. https://doi.org/10.1002/hbm.23486, PubMed:
28067006
Squire, L. R., Stark, C. E. L., & Clark, R. E. (2004). The medial tem-
poral lobe. Annual Review of Neuroscience, 27, 279–306.
https://doi.org/10.1146/annurev.neuro.27.070203.144130,
PubMed: 15217334
Stanley, J. A., Burgess, A., Khatib, D., Ramaseshan, K., Arshad, M.,
Wu, H., & Diwadkar, V. A. (2017). Functional dynamics of hip-
pocampal glutamate during associative learning assessed with in
vivo 1H functional magnetic resonance spectroscopy. Neuro-
Image, 153, 189–197. https://doi.org/10.1016/j.neuroimage
.2017.03.051, PubMed: 28363835
Stephan, K. E., Baldeweg, T., & Friston, K. J. (2006). Synaptic plas-
ticity and dysconnection in schizophrenia. Biological Psychiatry,
59(10), 929–939. https://doi.org/10.1016/j.biopsych.2005.10
.005, PubMed: 16427028
Stephan, K. E., & Roebroeck, A. (2012). A short history of causal
modeling of fMRI data. NeuroImage, 62(2), 856–863. https://
doi.org/10.1016/j.neuroimage.2012.01.034, PubMed:
22248576
Stolz, B. J., Emerson, T., Nahkuri, S., Porter, M. A., & Harrington,
H. A. (2021). Topological data analysis of task-based fMRI data
from experiments on schizophrenia. Journal of Physics: Com-
plexity, 2, 035006. https://doi.org/10.1088/2632-072X/abb4c6
Takehara-Nishiuchi, K. (2021). Neurobiology of systems memory
consolidation. European Journal of Neuroscience, 54(8),
6850–6863. https://doi.org/10.1111/ejn.14694, PubMed:
32027423
Telesford, Q. K., Joyce, K. E., Hayasaka, S., Burdette, J. H., &
Laurienti, P. J. (2011). The ubiquity of small-world networks.
Brain Connectivity, 1(5), 367–375. https://doi.org/10.1089/brain
.2011.0038, PubMed: 22432451
Thakkar, K. N., Diwadkar, V. A., & Rolfs, M. (2017). Oculomotor
prediction: A window into the psychotic mind. Trends in Cogni-
tive Sciences, 21(5), 344–356. https://doi.org/10.1016/j.tics.2017
.02.001, PubMed: 28292639
Thompson, W. H., & Fransson, P. (2016). On stabilizing the vari-
ance of dynamic functional brain connectivity time series. Brain
Connectivity, 6(10), 735–746. https://doi.org/10.1089/ brain
.2016.0454, PubMed: 27784176
Tregellas, J. R., Davalos, D. B., Rojas, D. C., Waldo, M. C., Gibson,
L., Wylie, K., … Freedman, R. (2007). Increased hemodynamic
response in the hippocampus, thalamus and prefrontal cortex
during abnormal sensory gating in schizophrenia. Schizophrenia
Research, 92(1–3), 262–272. https://doi.org/10.1016/j.schres
.2006.12.033, PubMed: 17336502
Tregellas, J. R., Smucny, J., Harris, J. G., Olincy, A., Maharajh, K.,
Kronberg, E., … Freedman, R. (2014). Intrinsic hippocampal activ-
ity as a biomarker for cognition and symptoms in schizophrenia.
American Journal of Psychiatry, 171(5), 549–556. https://doi.org
/10.1176/appi.ajp.2013.13070981, PubMed: 24435071
Tso, I. F., Angstadt, M., Rutherford, S., Peltier, S., Diwadkar, V. A., &
Taylor, S. F. (2021). Dynamic causal modeling of eye gaze
processing in schizophrenia. Schizophrenia Research, 229,
112–121. https://doi.org/10.1016/j.schres.2020.11.012,
PubMed: 33229223
van den Heuvel, M. P., Mandl, R. C., Stam, C. J., Kahn, R. S., &
Hulshoff Pol, H. E. (2010). Aberrant frontal and temporal complex
network structure in schizophrenia: A graph theoretical analysis.
Journal of Neuroscience, 30(47), 15915–15926. https://doi.org
/10.1523/JNEUROSCI.2874-10.2010, PubMed: 21106830
van den Heuvel, M. P., & Sporns, O. (2013). Network hubs in the
human brain. Trends in Cognitive Sciences, 17(12), 683–696.
https://doi.org/10.1016/j.tics.2013.09.012, PubMed: 24231140
Wadehra, S., Pruitt, P., Murphy, E. R., & Diwadkar, V. A. (2013).
Network dysfunction during associative learning in schizophre-
nia: Increased activation, but decreased connectivity: An fMRI
study. Schizophrenia Research, 148(1–3), 38–49. https://doi.org
/10.1016/j.schres.2013.05.010, PubMed: 23759649
Wang, D., Buckner, R. L., Fox, M. D., Holt, D. J., Holmes, A. J.,
Stoecklein, S., … Liu, H. (2015). Parcellating cortical func-
tional networks in individuals. Nature Neuroscience, 18(12),
1853–1860. https://doi.org/10.1038/nn.4164, PubMed:
26551545
Wannan, C. M. J., Bartholomeusz, C. F., Cropley, V. L., Van
Rheenen, T. E., Panayiotou, A., Brewer, W. J., … Wood, S. J.
(2018). Deterioration of visuospatial associative memory follow-
ing a first psychotic episode: A long-term follow-up study. Psy-
chological Medicine, 48(1), 132–141. https://doi.org/10.1017
/S003329171700157X, PubMed: 28625185
Woodcock, E. A., White, R., & Diwadkar, V. A. (2015). The dorsal
prefrontal and dorsal anterior cingulate cortices exert comple-
mentary network signatures during encoding and retrieval
in associative memory. Behavioural Brain Research, 290,
152–160. https://doi.org/10.1016/j.bbr.2015.04.050, PubMed:
25960314
Yang, J., Pu, W., Wu, G., Chen, E., Lee, E., Liu, Z., & Palaniyappan,
L. (2020). Connectomic underpinnings of working memory defi-
cits in schizophrenia: Evidence from a replication fMRI study.
Schizophrenia Bulletin, 46(4), 916–926. https://doi.org/10.1093
/schbul/sbz137, PubMed: 32016430
Zhao, Z., Cheng, Y., Li, Z., & Yu, Y. (2018). Altered small-world net-
works in first-episode schizophrenia patients during cool execu-
tive function task. Behavioural Neurology, 2018, 2191208.
https://doi.org/10.1155/2018/2191208, PubMed: 30254708
Zuo, X.-N., Ehmke, R., Mennes, M., Imperati, D., Castellanos, F. X.,
Sporns, O., & Milham, M. P. (2012). Network centrality in
the human functional connectome. Cerebral Cortex, 22(8),
1862–1875. https://doi.org/10.1093/cercor/ bhr269, PubMed:
21968567
Network Neuroscience
212
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
1
8
4
2
0
7
2
1
3
3
n
e
n
_
a
_
0
0
2
7
8
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3