RESEARCH
Classification and prediction of cognitive
performance differences in older age based
on brain network patterns using a
machine learning approach
Camilla Krämer1,2
, Johanna Stumme1,2, Lucas da Costa Campos1,2, Christian Rubbert3,
Julian Caspers3, Svenja Caspers1,2*, and Christiane Jockwitz1,2*
1Institute of Neuroscience and Medicine (INM-1), Research Centre Jülich, Jülich, Germany
2Institute for Anatomy I, Medical Faculty & University Hospital Düsseldorf, Heinrich Heine University Düsseldorf,
Düsseldorf, Germany
3Department of Diagnostic and Interventional Radiology, Medical Faculty & University Hospital Düsseldorf,
Heinrich Heine University Düsseldorf, Düsseldorf, Germany
*These authors contributed equally.
Keywords: Cognition, Aging, Resting-state functional connectivity, Graph-theoretical analyses,
Machine learning
ABSTRACT
Age-related cognitive decline varies greatly in healthy older adults, which may partly be
explained by differences in the functional architecture of brain networks. Resting-state
functional connectivity (RSFC) derived network parameters as widely used markers describing
this architecture have even been successfully used to support diagnosis of neurodegenerative
diseases. The current study aimed at examining whether these parameters may also be useful
in classifying and predicting cognitive performance differences in the normally aging brain by
using machine learning (ML). Classifiability and predictability of global and domain-specific
cognitive performance differences from nodal and network-level RSFC strength measures were
examined in healthy older adults from the 1000BRAINS study (age range: 55–85 years). ML
performance was systematically evaluated across different analytic choices in a robust cross-
validation scheme. Across these analyses, classification performance did not exceed 60%
accuracy for global and domain-specific cognition. Prediction performance was equally low
with high mean absolute errors (MAEs ≥ 0.75) and low to none explained variance (R2 ≤ 0.07)
for different cognitive targets, feature sets, and pipeline configurations. Current results highlight
limited potential of functional network parameters to serve as sole biomarker for cognitive aging
and emphasize that predicting cognition from functional network patterns may be challenging.
AUTHOR SUMMARY
In recent years, new insights into brain network communication related to cognitive
performance differences in older age have been gained. Simultaneously, an increasing number
of studies has turned to machine learning (ML) approaches for the development of biomarkers
in health and disease. Given the increasing aging population and the impact cognition has on
the quality of life of older adults, automated markers for cognitive aging gain importance. This
study addressed the classification and prediction power of resting-state functional connectivity
(RSFC) strength measures for cognitive performance in healthy older adults using a battery of
a n o p e n a c c e s s
j o u r n a l
Citation: Krämer, C., Stumme, J., da
Costa Campos, L., Rubbert, C.,
Caspers, J., Caspers, S., & Jockwitz, C.
(2023). Classification and prediction of
cognitive performance differences in
older age based on brain network
patterns using a machine learning
approach. Network Neuroscience,
7(1), 122–147. https://doi.org/10.1162
/netn_a_00275
DOI:
https://doi.org/10.1162/netn_a_00275
Supporting Information:
https://doi.org/10.1162/netn_a_00275
Received: 29 April 2022
Accepted: 22 August 2022
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Author:
Christiane Jockwitz
c.jockwitz@fz-juelich.de
Handling Editor:
Olaf Sporns
Copyright: © 2022
Massachusetts Institute of Technology
Published under a Creative Commons
Attribution 4.0 International
(CC BY 4.0) license
The MIT Press
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
7
1
1
2
2
2
0
7
2
1
6
8
n
e
n
_
a
_
0
0
2
7
5
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cognitive performance differences in older age
standard ML approaches. Classifiability and predictability of cognitive abilities was found to be
low across analytic choices. Results emphasize limited potential of these metrics as sole
biomarker for cognitive aging.
INTRODUCTION
Healthy older adults vary greatly in the extent to which they experience age-related cognitive
decline (Habib et al., 2007). While some older adults seem to maintain their cognitive abilities until
old age, others show higher rates of cognitive decline during the aging process (Cabeza, 2001;
Damoiseaux et al., 2008; Hedden & Gabrieli, 2004; Raz, 2000; Raz & Rodrigue, 2006). In light
of the continuously growing aging population, the impact of cognitive decline on everyday func-
tioning of older adults has gained momentum in research (Avery et al., 2020; Deary et al., 2009;
Depp & Jeste, 2006; Fountain-Zaragoza et al., 2019; Luciano et al., 2009; Vieira et al., 2022).
In this context, differences in the functional architecture of brain networks have been iden-
tified as a potential source of variance explaining cognitive performance differences during
aging (Chan et al., 2014; Stumme et al., 2020). Age-related differences have been linked to
changes in resting-state functional connectivity (RSFC) of major resting-state networks, for exam-
ple, the default mode network (DMN), the sensorimotor network (SMN), and the fronto-parietal
and visual networks (Andrews-Hanna et al., 2007; Chong et al., 2019; Ng et al., 2016; Stumme
et al., 2020). In detail, age-related cognitive decline is associated with both decreases in the
functional specialization of brain networks (reduced network segregation) and increasingly
shared coactivation patterns between functional brain networks (increased network integration)
(Andrews-Hanna et al., 2007; Chan et al., 2014; Chong et al., 2019; Fjell et al., 2015; Grady
et al., 2016; Ng et al., 2016; Onoda et al., 2012; Stumme et al., 2020). Furthermore, RSFC dif-
ferences in older age may differentiate between healthy older adults and individuals suffering
from mild cognitive impairment (MCI) or Alzheimer’s disease (AD). For instance, both MCI and
AD have been related to reduced RSFC within the DMN and SMN, the degeneration of specific
brain hubs, and aberrant functional brain network organization (Dai et al., 2015; Farahani et al.,
2019; Sanz-Arigita et al., 2010; Supekar et al., 2008; Wang et al., 2013).
Given the role of RSFC network patterns in cognition in healthy and pathological aging,
research on neurodegenerative diseases has started to embark on the development of diagnos-
tic biomarker for automatic patient classification based on RSFC. For the development of diag-
nostic biomarkers, machine learning (ML) methods may be particularly suited. This is due to
their ability to deal with high-dimensional data and to detect spatially distributed effects in the
brain that might otherwise not be detected using univariate approaches (Dadi et al., 2019;
Orrù et al., 2012; Woo et al., 2017; Zarogianni et al., 2013). In this context, RSFC-derived
metrics capturing network integration and segregation have already been successfully used
as diagnostic markers for MCI and AD, using ML approaches (Hojjati et al., 2017; Khazaee
et al., 2016). In healthy older populations, functional network measures have also provided
new insights into brain network communication related to cognitive performance differences
(Chan et al., 2014; Chong et al., 2019; Stumme et al., 2020). Specifically, a previous study has
demonstrated that shifts in within- and inter-network connectivity may be linked to differences
in cognitive performance in older age (Stumme et al., 2020). Thus, RSFC network properties
may also constitute potential meaningful candidates in search for a marker for nonpathological
age-related cognitive decline (Chan et al., 2014; Stumme et al., 2020).
Previous studies have mainly used RSFC matrices, either containing information across the
whole-brain or within specific networks, as input features to ML revealing initial promising
results in the prediction of different cognitive facets in older adults (Avery et al., 2020; He
Machine learning (ML):
Set of methods used to automatically
find patterns in data that allow
classification and prediction.
Network Neuroscience
123
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
7
1
1
2
2
2
0
7
2
1
6
8
n
e
n
_
a
_
0
0
2
7
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cognitive performance differences in older age
et al., 2020; Kwak et al., 2021; Pläschke et al., 2020). For instance, it has been shown that
working memory performance could be predicted by specific RSFC patterns in meta-analytically
defined brain networks in an older but not younger age group by using relevance vector regres-
sion (RVR) (Pläschke et al., 2020). Furthermore, a variety of neuropsychological test scores and
fluid intelligence could be successfully predicted from RSFC in large older samples using ML (He
et al., 2020; Kwak et al., 2021). Nevertheless, it remains unclear if RSFC strength measures tar-
geting network integration and segregation may provide additional useful information in classi-
fying and predicting global and domain-specific cognitive performance in older adults (Avery
et al., 2020; Dubois et al., 2018; He et al., 2020; Kwak et al., 2021; Pläschke et al., 2020). Further
knowledge in this context may be helpful on the road to building a reliable and accurate
biomarker for cognitive performance in healthy older adults that could ultimately be used to pre-
dict prospective cognitive decline. The current investigation, therefore, aims to systematically
examine whether RSFC strength parameters, capturing within- and inter-network connectivity,
may reliably classify and predict cognitive performance differences in a large sample of older
adults (age: 55–85) from the 1000BRAINS study by using a battery of standard ML approaches.
MATERIALS AND METHODS
Participants
Data for the current investigation stems from the 1000BRAINS project (Caspers et al., 2014), an
epidemiologic population-based study examining variability of brain structure and function
during aging in relation to behavioral, environmental, and genetic factors. The 1000BRAINS
sample was drawn from the 10-year follow-up cohort of the Heinz Nixdorf Recall Study and
the associated MultiGeneration study (Schmermund et al., 2002). As 1000BRAINS aims at the
characterization of the aging process in the general population, no exclusion criteria other than
eligibility for MR measurements (Caspers et al., 2014) were applied. In the current study, 966
participants were included within the age range 55 to 85 years. From this initial sample, 99
participants were excluded due to missing resting-state functional magnetic resonance imaging
(fMRI) data or failed preprocessing. Furthermore, 25 participants were excluded due to insuf-
ficient quality of the preprocessed functional data described in further detail below (see Data
Acquisition and Preprocessing section). Another 27 participants with missing scores on the
DemTect, a dementia screening test, or those scoring smaller or equal to 8 were excluded
due to the possibility of substantial cognitive impairment (Kalbe et al., 2004). Finally, two par-
ticipants were excluded due to more than three missing values within the neuropsychological
assessment (see Cognitive Performance section). This resulted in an initial (unmatched) sample
of 813 participants (372 females, Mage = 66.99, SDage = 6.70; see Table 1A and Figure 1:
Sample). All subjects provided written consent prior to inclusion and the study protocol of
1000BRAINS was approved by the Ethics Committee of the University of Essen, Germany.
Table 1. Demographic information for unmatched and matched samples regarding age, educational level, and risk of dementia
Female
Male
Total
N
372
441
813
A. Unmatched sample
Age
66.38 (6.53)
Education
5.93 (1.84)
DemTect
15.42 (2.29)
67.5 (6.8)
6.95 (1.91)
14.38 (2.33)
66.99 (6.70)
6.48 (1.94)
14.86 (2.37)
N
232
286
518
B. Matched sample
Education
5.88 (1.7)
Age
65.33 (5.48)
DemTect
15.43 (2.22)
67.81 (6.44)
6.96 (1.87)
14.45 (2.25)
66.7 (6.15)
6.48 (1.87)
14.89 (2.29)
Note. Mean displayed with standard deviation (SD) appearing in parentheses.
Network Neuroscience
124
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
7
1
1
2
2
2
0
7
2
1
6
8
n
e
n
_
a
_
0
0
2
7
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cognitive performance differences in older age
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
7
1
1
2
2
2
0
7
2
1
6
8
n
e
n
_
a
_
0
0
2
7
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
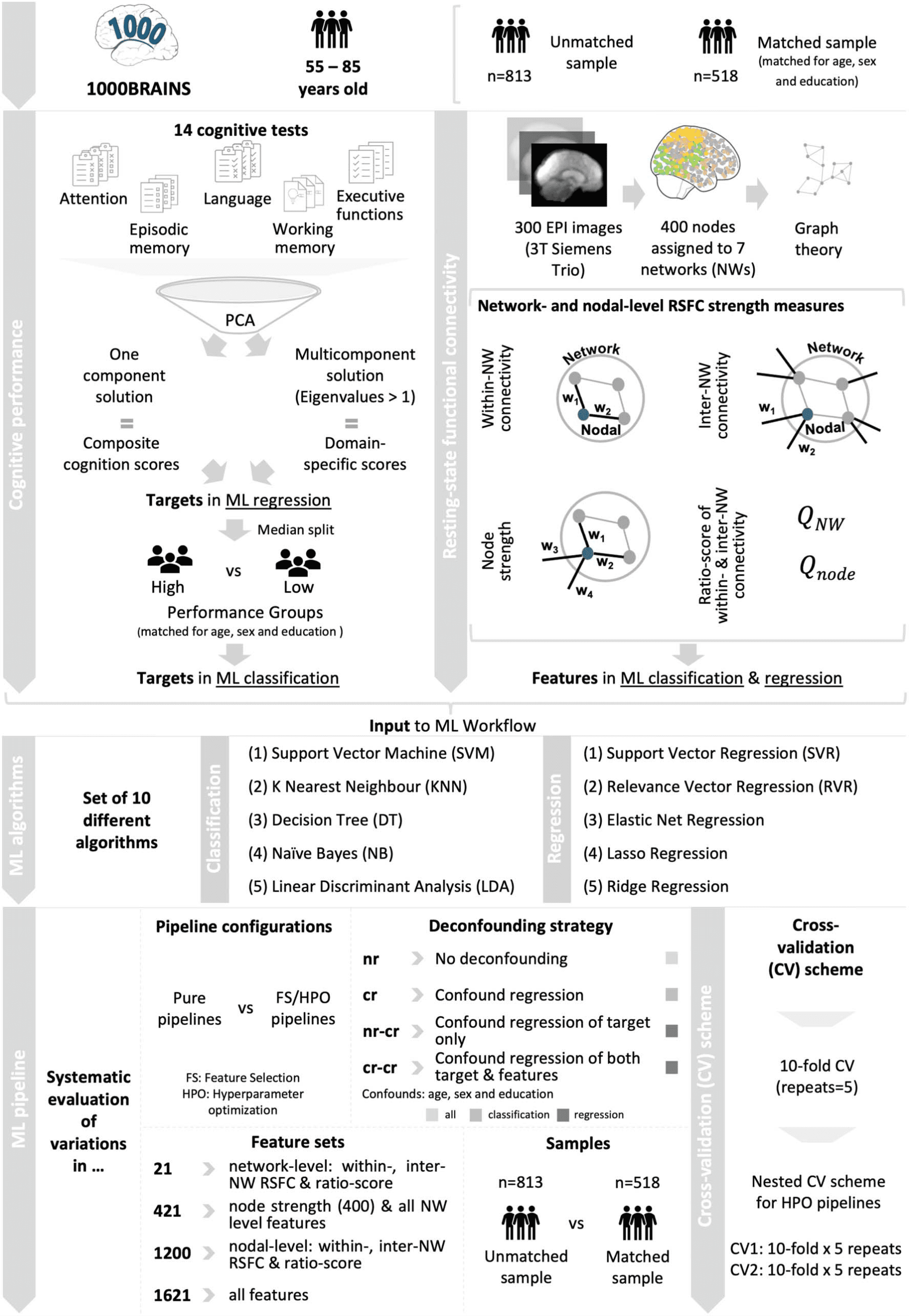
Figure 1. Schematic overview of workflow.
Network Neuroscience
125
Cognitive performance differences in older age
Global cognition:
General cognitive ability that
encompasses cognitive functioning
across different domains.
Cognitive Performance
All subjects underwent a large neuropsychological assessment testing the cognitive domains atten-
tion, executive functions, episodic memory, working memory ( WM), and language (for further
details, see Caspers et al., 2014). Fourteen cognitive variables targeting selective attention, process-
ing speed, figural and verbal fluency, problem solving, vocabulary, WM, and episodic memory
were selected for the purpose of the current study (see Figure 1: Cognitive performance). Further
information on the tests and variables chosen in the current investigation are found in Supporting
Information Table S1. In case of missing values (more than three missing values led to exclusion) in
the neuropsychological assessment, missing values were replaced by the median for respective sex
(males, females) and age groups (55–64 years, 65–74 years, 75–85 years). Imputation of missing
values was performed to avoid further loss of information and power. In a next step, raw scores from
all 14 neuropsychological tests used in the analysis were transformed into z-scores. For interpret-
ability purposes, scores for neuropsychological tests with higher values meaning lower perfor-
mance (i.e., time to complete the tasks or number of errors made) were inverted.
Neuropsychological test performance was reduced to cognitive composite scores using
principal component analysis (PCA). To disentangle effects specific to certain cognitive
facets, global and domain-specific cognitive performance were examined (Tucker-Drob,
2011). PCA was used to extract a one-component solution for global cognition and a multi-
component solution for cognitive subdomains based on eigenvalues >1. Lastly, varimax rota-
tion was applied to enhance the interpretability of extracted components. Individual global
and domain-specific component scores obtained from the PCA were used as targets in ML
prediction of cognitive performance differences.
For classification of cognitive performance differences, the initial (unmatched) sample was
separated into high- and low-performing groups. To do so, a median split was performed based
on each of the three cognitive component scores (as extracted in the PCA). To remove the effect of
potential confounders, the high- and low-performance groups derived from global cognition
were additionally matched with respect to age, sex, and educational level by using propensity
score matching, which constitutes a statistical approach to match participants based on their pro-
pensity scores (McDermott et al., 2016; Randolph et al., 2014; Stern et al., 1994; Vemuri et al.,
2014). This led to a matched sample with N = 518 (232 females, Mage = 66.7, SDage = 6.15; see
Table 1B and Figure 1: Sample and Cognitive performance). Further demographic information
regarding age, educational level, and sex distribution between high- and low-performance
groups in the unmatched and matched sample can be found in Table 2. All cognitive analyses
were performed using IBM SPSS Statistics 26 (https://www.ibm.com/de-de/analytics/spss
-statistics-software) and customized Python ( Version 3.7.6) and R scripts ( Version 4.00).
Functional Imaging
Imaging data was acquired using a 3T Siemens Tim-TRIO MR
Data acquisition and preprocessing.
scanner with a 32-channel head coil. Out of the whole MR imaging protocol (for details, see
Caspers et al. 2014), the current study used for surface reconstruction the 3D high-resolution
T1-weighted magnetization-prepared rapid acquisition gradient-echo (MPRAGE) (176 slices,
slice thickness = 1 mm, TR = 2,250 ms, TE = 3.03 ms, FoV = 256 × 256 mm2, flip angle = 9°,
voxel resolution = 1 × 1 × 1 mm3); and for resting-state analyses, the 11:30 minutes resting-state
fMRI with 300 EPI (gradient-echo planar imaging) volumes (slices 36, slice thickness = 3.1 mm,
TR = 2,200 msec, TE = 30 msec, FoV = 200 × 200 mm2, voxel resolution = 3.1 × 3.1 × 3.1 mm3).
During the resting-state scan, participants were instructed to keep their eyes closed, to relax and
let their mind wander, but not to fall asleep. This was checked during a postscan debriefing.
Network Neuroscience
126
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
7
1
1
2
2
2
0
7
2
1
6
8
n
e
n
_
a
_
0
0
2
7
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
C
o
g
n
i
t
i
v
e
p
e
r
f
o
r
m
a
n
c
e
d
i
f
f
e
r
e
n
c
e
s
i
n
o
l
d
e
r
a
g
e
t
N
e
w
o
r
k
N
e
u
r
o
s
c
e
n
c
e
i
Table 2. Differences in cognitive scores, age, educational level, and sex distribution between high- and low-performance groups in the unmatched and matched sample
COGNITIVE COMPOSITE
NON-VERBAL MEMORY & EXECUTIVE
VERBAL MEMORY & LANGUAGE
Low
−.79
(0.72)
69.49
(6.43)
5.84
(1.76)
206
200
−.66
(0.63)
67.06
(6.1)
6.39
(1.82)
143
116
Group
t
−37.17
p
df
<0.001
697.9
11.48
<0.001
811
−10.51
<0.001
805.0
–
–
–
–
–
–
−28.67
<0.001
460.2
1.32
0.19
516
−.1.06
0.29
516
–
–
–
–
–
–
High
.79
(0.47)
64.49
(5.99)
7.13
(1.91)
235
172
.71
(0.44)
66.34
(6.2)
6.56
(1.92)
143
116
Low
−.78
(0.68)
69.24
(6.58)
6.03
(1.88)
187
220
−.68
(0.61)
67.69
(6.20)
6.31
(1.85)
127
128
High
.78
(0.56)
64.72
(6.02)
6.94
(1.9)
254
152
.75
(0.54)
65.74
(5.95)
6.64
(1.88)
159
104
Group
t
−36.02
p
df
<0.001
784.8
Low
−.81
(0.60)
10.28
<0.001
805.1
68.09
−6.87
<0.001
810.8
–
–
–
–
–
–
−28.35
<0.001
516
3.65
<0.001
516
−2.01
<0.05
516
–
–
–
–
–
–
(6.72)
5.97
(1.76)
245
161
−.74
(0.54)
66.63
(6.01)
6.3
(1.77)
165
99
Group
t
−36.67
p
df
<0.001
811
4.74
<0.001
811
−7.81
<0.001
800
–
–
–
–
–
–
−31.24
<0.001
516
−.25
.81
516
−2.25
<0.05
506.1
–
–
–
–
–
–
High
.80
(0.59)
65.89
(6.5)
7.00
(1.99)
196
211
.74
(0.53)
66.77
(6.29)
6.67
(1.96)
121
133
Unmatched
Sample
Cog.
Score
Matched
Sample
Age
Education
Males
Females
Cog.
Score
Age
Education
Males
Females
Note. Standard deviation (SD) appears in parentheses. Cog. Score = cognitive score. Unmatched sample: global: X 2(1) = 4.01, p < .05; memory and executive: X 2(1) = 22.61, p < .001; language: X 2(1) = 12.16, p <
.001; Matched Sample: global: X 2(1) = 0, p = 1; memory and executive: X 2(1) = 5.94, p < .05; language: X 2(1) = 11.56, p < .001.
1
2
7
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
1
2
2
2
0
7
2
1
6
8
n
e
n
_
a
_
0
0
2
7
5
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cognitive performance differences in older age
Preprocessing steps closely followed those from Stumme and colleagues (2020). During pre-
processing, the first four volumes from the 300 EPI were removed for each participant. All func-
tional images were corrected for head movement using a two-pass procedure. First, all volumes
were aligned to the first image and then to the mean image using affine registration. Spatial
normalization to the MNI152 template (2-mm-voxel size) of all functional images was achieved
by using a “unified segmentation” approach as previous studies have shown increased registra-
tion accuracies compared to normalization based on T1-weighted images (Ashburner & Friston,
2005; Calhoun et al., 2017; Dohmatob et al., 2018). Furthermore, ICA-AROMA, that is, ICA-
based automatic removal of motion artifacts (Pruim et al., 2015), which constitutes a data-
driven method for the identification and removal of motion-related components from MRI data,
was applied. Additionally, global signal regression (GSR) was performed in order to minimize
the association between motion and RSFC (Burgess et al., 2016; Ciric et al., 2017; Parkes et al.,
2018). Moreover, GSR has been found to improve behavioral prediction performance and to
enhance the link between RSFC and behavior (Li et al., 2019). In a final step, a band-pass filter
was applied (0.01–0.1 Hz). As a quality check for our preprocessing, further steps were imple-
mented. Initially, we checked for potential misalignments in the mean functional AROMA data
with the check sample homogeneity option in the Computational Anatomy Toolbox (CAT 12)
(Gaser et al., 2022). Participants detected as outliers with >2 SD away from the mean were
excluded. Additionally, we checked for volume-wise severe intensity dropouts (DVARS) in the
preprocessed data by using an algorithm by Afyouni and Nichols (2018). For each participant,
p values for spikes are generated, and participants with more than 10% of the 300 volumes
detected as dropouts were excluded from further analyses. To check the quality control applied,
we assessed the correlation between age and motion after the application of AROMA and the
exclusion of deviating participants and found it to be nonsignificant (percentage (%) of corrupted
volumes * age, r = .03, p = .39).
Functional connectivity analyses. For connectivity analyses, the 400-node cortical parcellation
by Schaefer and colleagues (2018) was adopted. The 400 regions of interest from the parcella-
tion scheme can be allocated to seven network parcels of known functional resting-state net-
works (Yeo et al., 2011). These include the visual, sensorimotor, limbic, fronto-parietal, default
mode, dorsal, and ventral attention network.
A whole-brain graph was established from functional data (Rubinov & Sporns, 2010). This
included, (i) a mean time series extraction for each node using fslmeants (Smith et al., 2004),
(ii) individual edge definition as the Pearson’s correlation of respective average time series of
two nodes, (iii) a statistical significance test of each correlation coefficient using the Fourier trans-
form and permutation testing (repeats = 1,000) with nonsignificant edges at p ≥ 0.05 being set
to zero (Stumme et al., 2020; Zalesky et al., 2012), and (iv) Fisher’s r-to-z-transformation applied
to the 400 × 400 adjacency matrix. Furthermore, since there is still debate about the true nature
of anticorrelations in the brain, only positive correlations were considered in subsequent analyses
(negative correlations were set to zero) (Murphy et al., 2009; Murphy & Fox, 2017; Saad et al.,
2012). Finally, no further thresholding related to network density or network size was applied to
the brain graph as it may, in addition to controlling the absolute number of edges, also increase
the number of false positives and induce systematic differences in overall RSFC (Stumme et al.,
2020; van den Heuvel et al., 2017; van Wijk et al., 2010). For the estimation of strength measures,
the final network used, thus, may be described as a positively weighted network.
In a next step, connectivity estimates were calculated using the software bctpy with net-
work parameters defined as in Rubinov and Sporns (2010) (https://pypi.org/project/bctpy/).
All metrics estimated in the current investigation are based on the estimation of strength
Network Neuroscience
128
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
7
1
1
2
2
2
0
7
2
1
6
8
n
e
n
_
a
_
0
0
2
7
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cognitive performance differences in older age
Inter-network RSFC:
Connectivity strength estimate of one
node (nodal) or all nodes (network)
within a network to all nodes outside
its network.
Ratio-score:
A metric capturing within-network
RSFC of one node (nodal) or all
nodes (network) within a network in
relation to its inter-network RSFC.
Within-network RSFC:
Connectivity strength estimate of one
node (nodal) or all nodes (network)
within a network to all nodes within
its network.
Feature set:
The specific combination of input
features used in ML.
values, which do not appear to be distorted by varying amounts of edges and have been
shown to reliably quantify networks (Finn et al., 2015). In total, seven parameters were com-
puted for later use in ML. Within- and inter-network RSFC as well as a ratio-score indicating
network segregation were obtained at both network and nodal level (see Figure 1: RSFC; for
further details on network parameters, see Stumme et al., 2020). Within-network RSFC was
defined as the sum of strength values from all nodes (network) or one node (nodal) within a
network to all nodes within its related network divided by the number of existing edges in the
network (network: 7 features; nodal: 400 features). Inter-network RSFC referred to the sum of
strength values from all nodes (network) or one node (nodal) within a network to all nodes
outside its network divided by the number of all edges in the network (network: 7 features;
nodal: 400 features). The ratio-score captured within-network RSFC of all nodes (network)
or one node (nodal) in relation to its inter-network RSFC (network: 7 features; nodal: 400 fea-
tures). Additionally, the strength of each node was calculated as the sum of all connectivity
weights attached to a node (i.e., 400 features). In total, the feature vector for each subject
consisted of 1,621 features (4 × 400 = 1,600 nodal features and 3 × 7 = 21 network-level
features). From this, four different feature sets were derived and used in ML (21 features: all
network-level features; 421 features: node strength and all network-level features; 1,200 fea-
tures: nodal within- and inter-network and ratio of within/inter-network RSFC; 1,621 features:
all features).
Systematic Application of a Battery of Standard Machine Learning Approaches
ML was used to assess whether RSFC strength measures can be used to distinguish (i.e.,
classification) and predict (i.e., regression) cognitive performance differences in older adults.
As there is currently no agreement on a standard ML pipeline using neuroimaging data given
the high variability in dataset properties, we systematically evaluated different analytical
choices (see Figure 1: ML algorithms and pipeline). Performance of different ML algorithms,
pipeline compositions, extents of deconfounding, and variations in feature set and sample
sizes were assessed (Arbabshirani et al., 2017; Cui & Gong, 2018; Khazaee et al., 2016;
Mwangi et al., 2014; Paulus & Thompson, 2021; Pervaiz et al., 2020). As such, we tested
a total of 556 unique pipelines in the classification (406 pipelines) and regression (150 pipe-
lines) setting. The scikit-learn library (version: 0.22.1) in Python ( Version 3.7.6) (Pedregosa
et al., 2011; https://scikit-learn.org/stable/index.html) was used for all ML analyses unless
specified.
ML algorithms. For classification, Five different algorithms were examined: support vector
machine (SVM), K-nearest while (KNN), decision tree (DT), naïve Bayes (NB) and linear dis-
criminant analysis (LDA). Further information on the algorithms can be found in the Supporting
Information Methods.
For regression, five different algorithms were assessed: support vector regression (SVR),
RVR, Ridge regression (Ridge), least absolute shrinkage and selection operator regression
(LASSO), and elastic net regression (Elastic Net) (Cui & Gong, 2018). The package scikit-
rvm compatible with scikit-learn by James Ritchie (https://github.com/ JamesRitchie/scikit
-rvm) was used for RVR computation. Further information on the regression algorithms can
be found in the Supporting Information Methods.
Basic ML pipeline. The basic ML pipeline was constructed as follows: the previously calculated
connectivity estimates were used as input features for the ML workflow. Targets varied
between classification (high vs. low cognitive performance group; matched sample) and
regression (global and domain-specific cognitive scores; unmatched sample) (see Cognitive
Network Neuroscience
129
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
7
1
1
2
2
2
0
7
2
1
6
8
n
e
n
_
a
_
0
0
2
7
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cognitive performance differences in older age
Pipeline configuration:
A specific setup of an ML pipeline to
be tested in the analysis.
Domain-specific cognition:
Cognitive processes that are linked
and dedicated to specific mental
abilities, e.g., executive and memory
functions.
Performance section in Materials and Methods). Input features were scaled to unit variance in
a first step in all pipeline configurations within the cross-validation setting. All models were
evaluated using a repeated 10-fold cross-validation (CV) (five repeats). In case of an additional
hyperparameter optimization (HPO) step, a repeated nested CV scheme was implemented for
selecting optimal parameters (outer and inner loop: 10 folds × 5 repeats) (see Figure 1: CV
scheme; Lemm et al., 2011). This was done to avoid data leakage and to obtain an unbiased
estimate of the generalization performance of complete models (Lemm et al., 2011). Balanced
accuracy (BAC) was used to assess classification performance. It was chosen to account for
potential group size differences in domain-specific cognition. Sensitivity and specificity were
also calculated to provide a more complete picture and can be found in the Supporting Infor-
mation. Mean absolute error (MAE ) and coefficient of determination (R2) were computed in the
prediction setting.
Systematic evaluation of ML pipeline options. Regarding pipeline configurations, different pipe-
line configurations were investigated. Performance of baseline models were compared to
those from pipelines with feature selection (FS) and HPO as they have been found to greatly
impact ML performance (Brown & Hamarneh, 2016; Guyon & Elisseeff, 2003; Hua et al.,
2009; Mwangi et al., 2014). For baseline models, algorithms were run with default settings
from scikit-learn without additional FS and HPO steps (pure pipeline). If FS was not performed
in conjunction with HPO, default parameters were equally used. We investigated different FS
methods in the present study (Mwangi et al., 2014).
For classification, two univariate filters, that is, ANOVA F-test and mutual information,
were compared to L1-based (using a linear SVM) and hybrid FS. For the univariate filters,
the top 10% of features were selected. Furthermore, L1-based (i.e., regularization) FS using a
linear SVM to create sparse models in combination with the five classifiers was examined.
Finally, a hybrid FS method, which combines both filter and wrapper methods, was consid-
ered (Kazeminejad & Sotero, 2019; Khazaee et al., 2016). Initially, a univariate filter
(ANOVA F-test) was applied selecting 50% of the top performing features. On the remaining
half of the features, a sequential forward floating selection wrapper was used to determine
the top 10 features contributing to the classification using the mlxtend package for Python
(Khazaee et al., 2016; Pudil et al., 1994; Raschka, 2018). FS was always performed on the
training set.
Different FS methods were also examined in ML regression. A univariate correlation–based
filter was applied in case of SVR, RVR, and Ridge regression (Finn et al., 2015; Guyon &
Elisseeff, 2003). Again the top 10% of features were selected. In contrast, LASSO and Elastic
Net regression are embedded FS algorithms. Due to their regularization penalty, only features
with a high discriminatory power will have a nonzero weight and will contribute to the task
at hand (Zou & Hastie, 2005). Thus, they enforce sparsity and with it integrate FS in their
optimization problem (Mwangi et al., 2014).
In terms of HPO, three of the five classification algorithms had hyperparameters to be
tuned, that is, SVM, KNN, and DT. HPO was carried out for (i) regularization parameter C
for SVM (10−4 to 101, 10 steps, logarithmic scale) for linear, radial basis function (RBF) and
polynomial (poly) kernel, (ii) maximum tree depth (4, 6, 8, 10, 20, 40, None) and optimum
criterion (gini impurity vs entropy) for DT, and (iii) number of neighbors for KNN (1, 3, 5, 7, 9,
11,13, 15, 17, 19, 21, 23, 25). HPO was assessed with and without additional FS (ANOVA
F-test) in classification. The following hyperparameters were tuned in ML prediction: (i) regu-
larization parameter lambda λ for LASSO and Ridge regression (LASSO: 10−1 to 102, Ridge:
10−3 to 105, 10 steps, logarithmic scale); (ii) parameters lambda, λ, and alpha, α, for Elastic Net
Network Neuroscience
130
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
7
1
1
2
2
2
0
7
2
1
6
8
n
e
n
_
a
_
0
0
2
7
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cognitive performance differences in older age
Deconfounding strategy:
The approach of how to control for
the impact of potential confounders,
e.g., age or sex.
(λ : 10−1 to 102, 10 steps, logarithmic scale; α: 0 to 1, 10 steps); and (iii) regularization param-
eter C for SVR (10−4 to 101, 10 steps, logarithmic scale) and kernel type (linear, RBF, and poly).
HPO was assessed in conjunction with FS in prediction as some algorithms incorporated
embedded feature selection. All HPO was performed on the inner loop using grid search
assessing the performance of all parameter combinations and choosing the best one in terms
of inner loop performance. All pipeline options were explored for feature sets without (nr con-
dition) and with deconfounding (cr, nr-cr, cr-cr condition) applied.
For deconfounding strategy, if deconfounding was applied, the covariates age, sex and edu-
cational level were regressed from features/targets. To avoid data leakage, confound regression
was always carried out within the ML pipeline. Following Rasero and colleagues (2021), con-
founders were regressed from targets/features by using a linear regression model, which was fit
using only the training set and then applied to both training and test data to obtain residuals.
Different extents of deconfounding (nr = no deconfounding; classification: cr = confounders
regressed from features; regression: nr-cr = confounders regressed from targets, cr-cr = con-
founders regressed from both features and targets) were implemented to assess its impact on
ML performance (Pervaiz et al., 2020).
For ML validation analyses, we performed several further analyses to validate our ML
approach. First, we investigated the influence of a finer grained parcellation on ML perfor-
mance (Dadi et al., 2019; Khazaee et al., 2016). Therefore, we compared ML performance
results obtained from using a 400-node and 800-node parcellation (Schaefer et al., 2018).
Additionally, ML performance was explored separately in males and females, given the
well-established gender differences in RSFC and its potential impact on ML performance
(Nostro et al., 2018; Stumme et al., 2020; Weis et al., 2019). Furthermore, we examined
whether the inclusion of information from negative correlations in terms of functional connec-
tivity may alter ML performance results. In this context, we calculated our strength measures
based on (i) the absolute values from both positive and negative correlations and (ii) only on
the absolute values from negative correlations and used these separately as features to ML.
Additionally, we investigated how classification performance changes when only extreme
groups, defined as the highest and lowest 25% of individuals scoring on the global cognition
component, are included (Dadi et al., 2021; Vieira et al., 2022). Classification performance
was examined in unmatched and matched (for age, sex, and education) samples (see Support-
ing Information Tables S2–S3). In terms of validating our pipeline, we tested our ML pipelines
in the context of age, which has repeatedly been shown to be successfully predicted from
RSFC patterns (Liem et al., 2017; Meier et al., 2012; Pläschke et al., 2017; Vergun et al.,
2013). To adapt this to our classification setting, we examined the classification of extreme
age groups (old vs. young; see Supporting Information Tables S4–S5) in feature set 421 (Vieira
et al., 2022). In the prediction setting, age was predicted continuously. Prediction analyses
were carried out for extreme groups, the unmatched sample and the whole age range of the
1000BRAINS cohort (18–85 age) (see Supporting Information Tables S4–S5).
Model Comparisons and Statistical Analyses
To assess the reliability and stability of the derived principal components (PCs), we performed
two additional analyses. First, we checked for the robustness of the PCA against the imputation
of missing values on different cognitive tests. Therefore, we obtained a validation sample, in
which all participants with missing values in any of the cognitive tests were excluded from the
unmatched sample (N = 749, 343 females, Mage = 66.86, SDage = 6.62). Then, we compared
component loadings from the original PCA results to the recalculated ones in the validation
sample by calculating Pearson’s correlations. Second, we turned to the stability of the PCs
Network Neuroscience
131
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
7
1
1
2
2
2
0
7
2
1
6
8
n
e
n
_
a
_
0
0
2
7
5
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cognitive performance differences in older age
across data splits to address the dependency between training and test sets introduced by per-
forming PCA as a first step in the analysis outside of the ML framework. In case of stability of
PCs, we may assume that this dependency will not affect our results. Therefore, we addition-
ally divided the data into two subsamples (random split-half procedure was implemented;
Sripada et al., 2020b; Thompson et al., 2019) and performed a PCA on each sample sepa-
rately. Component loadings from the split halves were compared to the original loadings by
computing Pearson’s correlations (see Supporting Information Tables S9–S10).
To assess the relation between cognitive scores derived from PCA and potential confounding
factors, we calculated partial correlations between all cognitive scores (global and domain
specific) and age (corrected for education and sex) as well as education (corrected for age and
sex) in the unmatched sample. Furthermore, to examine sex differences in cognitive scores, a mul-
tivariate analysis of covariance (MANCOVA) was computed with cognitive scores as dependent
variables, sex as the independent variable, and the inclusion of age and education as covariates.
For checking the quality of the dichotomization into a high- and low-performance group,
we performed independent samples t-tests to test for significant differences in cognitive per-
formance (global and domain specific) between high- and low-performance groups in the
unmatched and matched sample. Additionally, we assessed the relation between confounding
factors and group membership. Thus, we performed independent samples t-test to examine
group differences in terms of age and education and chi-square tests for independence to
assess differences in the sex distribution across high- and low-performance groups in
unmatched and matched samples.
To contextualize ML performance and obtain a chance-level prediction equivalent, we
compared ML model estimations to those from a reference model, that is, dummy classifier
and regressor, given the low computational costs of dummy estimates and their similarity in
distribution to approaches based on permutation (Engemann et al., 2020; Vieira et al., 2022).
In this case, the percentage of folds, for which the ML models were better than the reference
model in terms of accuracy (classification) and R2 (regression), was calculated with higher per-
centages (>80%) indicating robust outperformance of the reference model.
RESULTS
We performed twofold analyses to investigate whether cognitive performance differences
could be distinguished and predicted based on RSFC strength measures. In a first step, a simple
classification setting was chosen to examine if high- and low-performance groups can be
accurately classified from RSFC strength parameters using different ML pipeline configurations,
analytic choices, and feature sets. In a second step, we sought to address if the continuous
prediction of cognitive scores leads to ML performance differences compared to the classifi-
cation. Thus, we implemented a regression framework to analyze, whether cognitive perfor-
mance differences could be predicted from RSFC strength measures.
Cognitive Performance Across Unmatched and Matched Samples
A one-component solution for global cognition and a multicomponent solution for cognitive
subdomains based on the eigenvalue criterion (eigenvalue > 1) were extracted. Data suitability
for PCA was tested with the Kaiser–Meyer–Olkin (KMO) index examining the extent of com-
mon variability. With a value of KMO = 0.91, data appeared suitable for PCA. Component
scores from the one-component solution were stored as the COGNITIVE COMPOSITE (i.e.,
global cognition) score for each individual (see Figure 2 and Supporting Information Tables S6
and S7 and Figure S8). With regards to domain-specific cognitive scores, two components could
Network Neuroscience
132
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
7
1
1
2
2
2
0
7
2
1
6
8
n
e
n
_
a
_
0
0
2
7
5
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cognitive performance differences in older age
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
7
1
1
2
2
2
0
7
2
1
6
8
n
e
n
_
a
_
0
0
2
7
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
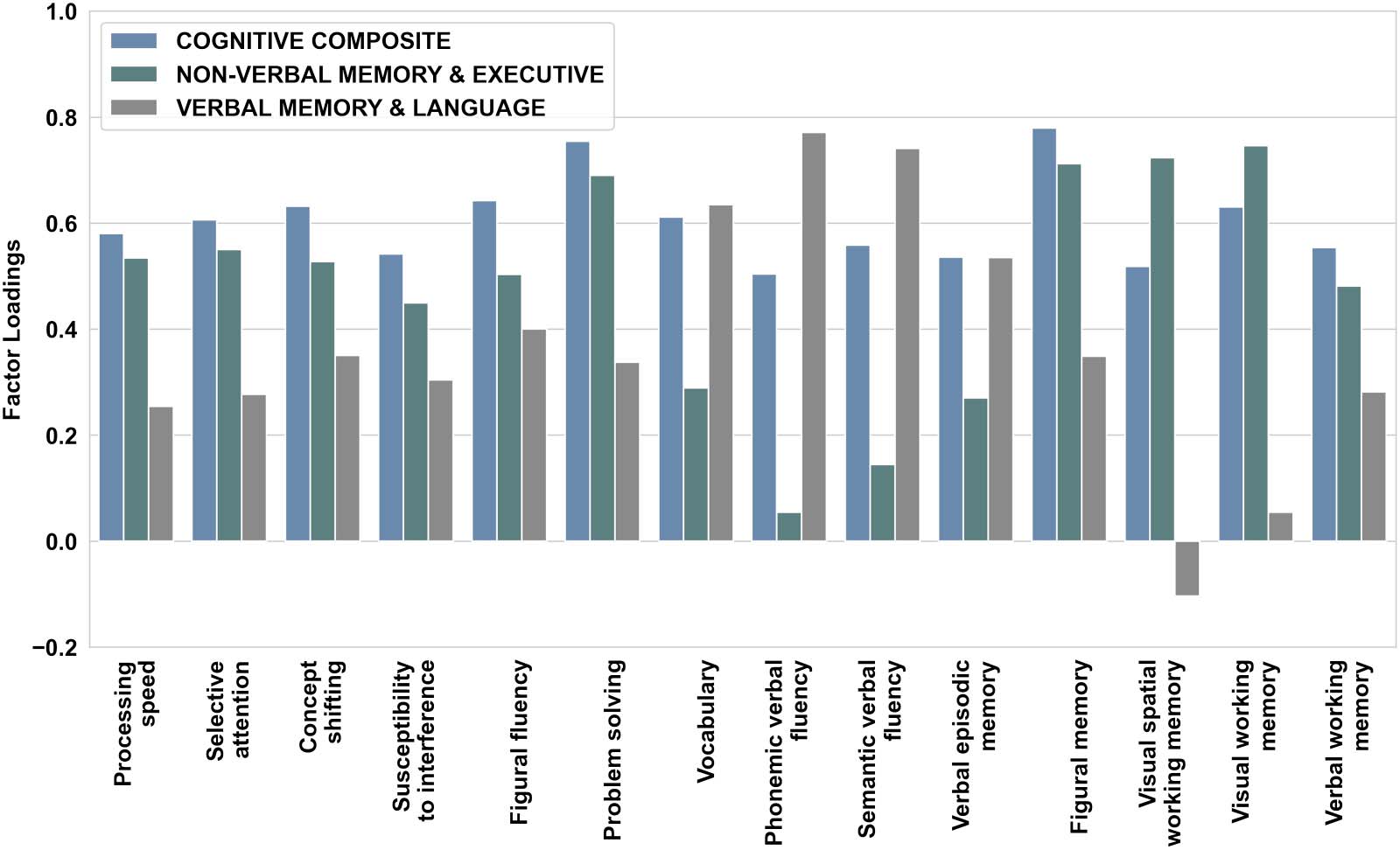
Figure 2. Factor loadings of each cognitive function on the one-component and multicomponent solution extracted from PCA analysis (after
varimax rotation).
be discovered from the PCA (see Figure 2 and Supporting Information Tables S6 and S7). The
first component mainly covered performance in visual spatial and spatial WM, figural mem-
ory, problem solving, selective attention, and processing speed (NON-VERBAL MEMORY &
EXECUTIVE component; see Figure 2 and Supporting Information Table S7). The second
component centrally reflected performance on semantic and phonemic verbal fluency, vocab-
ulary, and verbal episodic memory ( VERBAL MEMORY & LANGUAGE component; see
Figure 2 and Supporting Information Table S7). In terms of robustness and stability of PCs,
component loadings for all three extracted components were highly similar across the original
sample, the random split half samples and the validation sample (r > 0.86, p > 0.01; Supporting
Information Tables S9 and S10) indicating that PCs appear stable across subsets of data and
robust against the imputation of missing values. Age was significantly negatively correlated with
global and domain-specific cognitive performance scores (controlled for sex and educational
level; COGNITIVE COMPOSITE: r = −.48, p < .001; NON-VERBAL MEMORY & EXECUTIVE:
r = −.43, p < .001; VERBAL MEMORY & LANGUAGE: r = −.19, p < .001). Higher educational
level was significantly associated with higher global and domain-specific cognitive perfor-
mance (COGNITIVE COMPOSITE: r = .40, p < .001; NON-VERBAL MEMORY & EXECUTIVE:
r = .21, p < .001; VERBAL MEMORY & LANGUAGE: r = .35, p < .001; controlled for age and
sex). A multivariate analysis of covariance (MANCOVA) with age and education as covariates
revealed males to perform significantly better than females on the NON-VERBAL MEMORY &
EXECUTIVE component (F(1, 809) = 30.22, p < .001, ηp
2 = 0.036), while females outperformed
males on the VERBAL MEMORY & LANGUAGE component (F(1, 809) = 46.11, p < .001, ηp
2 =
0.056). In turn, no sex differences were found for global cognition (COGNITIVE COMPOSITE:
F(1, 809) = 0.024, p = .877, ηp
2 = 0.0). Component scores (global and domain-specific) obtained
from PCA were used as targets in ML prediction.
For classification of cognitive performance differences, high- and low-performance groups
were created by a median split after the extraction of participants’ component scores (as
extracted in the PCA). High- and low-performance groups in the initial (unmatched) sample
Network Neuroscience
133
Cognitive performance differences in older age
differed significantly in global and domain-specific cognitive performance, as well as in terms
of age, educational level, and sex (see Table 2). The high-performing group was found to be
significantly younger and better educated than the low-performing group (see Table 2). More
males than females were represented in the high-performance group for the COGNITIVE
COMPOSITE and the NON-VERBAL MEMORY & EXECUTIVE component (see Table 2).
The reversed pattern was found for the VERBAL MEMORY & LANGUAGE component (see
Table 2).
To control for the impact of confounding factors, high- and low-performance groups of the
COGNITIVE COMPOSITE component were matched on age, educational level, and sex. This
led to a matched subsample (N = 518; see Figure 1: Sample and Table 1B). High- and low-
performance groups again differed significantly in their global and domain-specific cognitive
performance (see Table 2). No significant group differences were encountered in terms of
age, educational level and sex distribution for the COGNITIVE COMPOSITE component
(see Table 2). Participants in the low-performance group on the NON-VERBAL MEMORY &
EXECUTIVE and VERBAL MEMORY & LANGUAGE component were found to be significantly
less educated than participants in the high-performance group. A similar significant pattern for
differences in the sex distribution was encountered as in the unmatched sample (see Table 2).
Group memberships (high vs. low) were used as targets in ML classification.
Classification Results
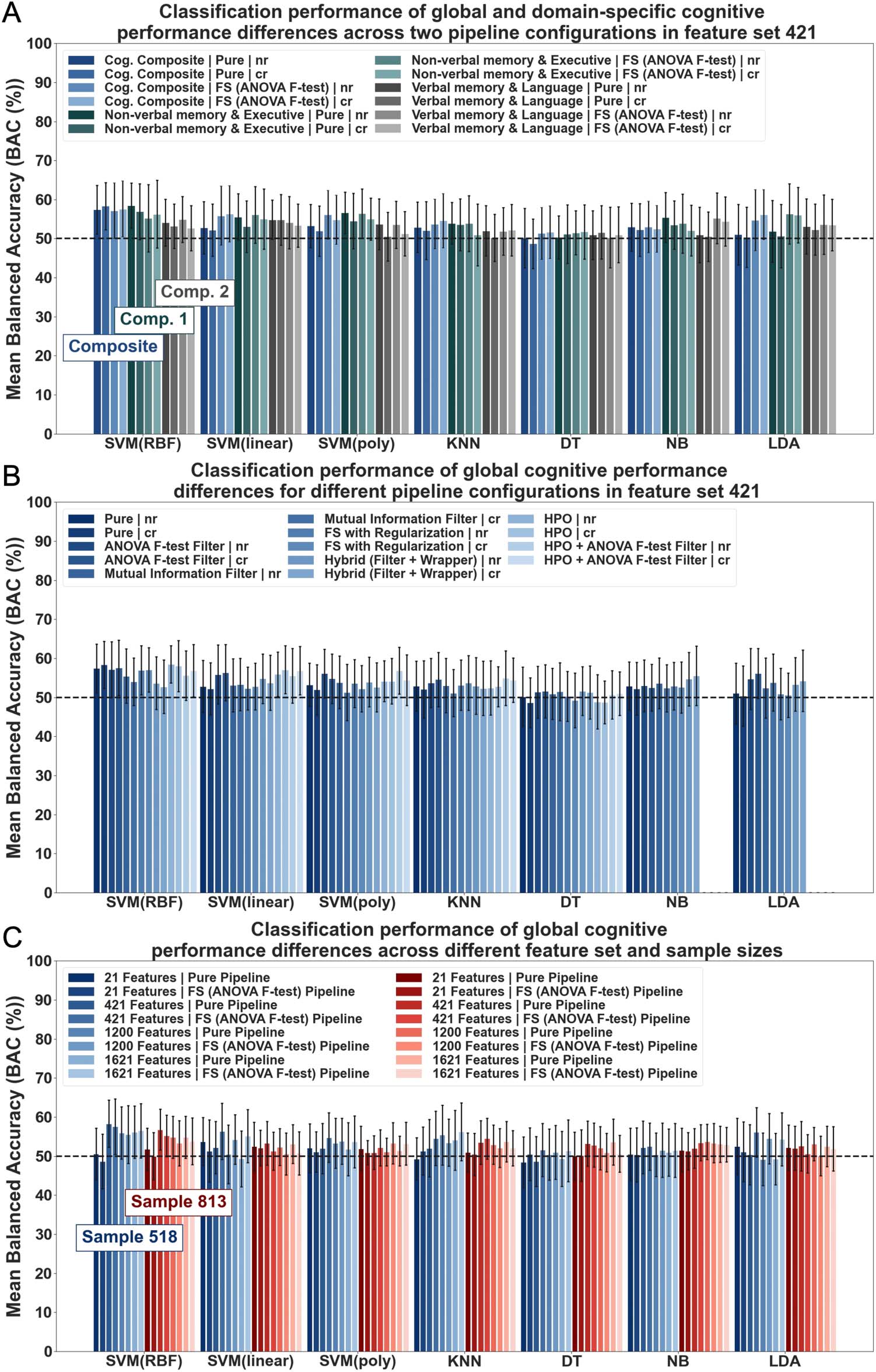
Classification performance across global cognition and cognitive domains. ML was used in a first
step to assess the usefulness of RSFC strength measures to distinguish cognitive performance
differences in older adults. All algorithms were first implemented in a feature set with 421 fea-
tures to examine classification performance of global and domain-specific cognitive perfor-
mance differences in the matched sample. Across all implemented ML pipelines with and
without univariate feature selection (FS), performance did not exceed 60% accuracy (see
Figure 3A and Supporting Information Table S11). Mean BACs ranged between 48.68% to
58.33% for global cognition and 50.21% to 58.44% for domain-specific cognition. These
results were further supported by the comparison to the dummy classifier. The majority of
models did not outperform the dummy classifier in more than 80% of folds. Higher accuracies
compared to the dummy were achieved mainly in no more than 50% to 80% of folds, sug-
gesting rather modest overall performance and limitations in reliability (see Supporting Infor-
mation Table S12). Classification accuracies for the NON-VERBAL MEMORY & EXECUTIVE
component were marginally higher than for the VERBAL MEMORY & LANGUAGE compo-
nent, which was also supported by results from comparisons to the dummy estimate (see
Figure 3A and Supporting Information Tables S11–S13). No systematic differences between
models based on features with (cr) or without (nr) deconfounding, that is, controlling for the
effects of age, sex, and education on features, could be observed (Figure 3A). Initial results
suggested poor discriminatory power of RSFC strength measures for global and domain-
specific cognitive performance differences in a large population-based older sample.
Classification performance across different pipeline configurations for global cognition. To examine
the impact of different pipeline configurations, we investigated ML performance in a pure
pipeline, that is, without FS, and in FS/hyperparameter optimization (HPO) pipelines, that
is, additional step of feature selection (FS) and HPO, for global cognition. All algorithms were
first implemented in a pure pipeline using 421 features. Baseline results revealed classification
accuracies between 48.68% to 58.33% (see Figure 3B). Baseline results were then compared
to those from different FS/HPO pipelines. Estimations from FS/HPO pipelines were found to be
Network Neuroscience
134
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
1
2
2
2
0
7
2
1
6
8
n
e
n
_
a
_
0
0
2
7
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cognitive performance differences in older age
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
1
2
2
2
0
7
2
1
6
8
n
e
n
_
a
_
0
0
2
7
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. Classification performance results of cognitive performance differences (based on global and domain-specific scores) from RSFC
strength measures. Classification results across algorithms: Support Vector Machine (SVM) with Radial Basis Function (RBF), linear and polyno-
mial (poly) kernel, K-Nearest Neighbour (KNN), Decision Tree (DT), Naïve Bayes (NB), Linear Discriminant Analysis (LDA). Results shown for (A)
different targets (cognitive composite and cognitive components), (B) pipeline configurations (pure (no FS/HPO) vs. FS/HPO pipelines), (C) sam-
ples (matched vs. unmatched sample) and feature set sizes (21, 421, 1,200, 1,621). Error bars correspond to standard deviation (SD); nr = no
confound regression applied to features; cr = age, sex, and education regressed from features; unless otherwise specified, cr condition showed.
Network Neuroscience
135
Cognitive performance differences in older age
similar to baseline estimations (MBAC range: 48.77–58.46%; in 42–96 % of folds BAC >
dummy classifier; see Figure 3B and Supporting Information Tables S14–S16). Thus, additional
pipeline steps, that is, FS and HPO, which are commonly found to enhance performance, did
not substantially increase classification accuracies in the current study (Brown & Hamarneh,
2016; Mwangi et al., 2014).
feature sets and sample sizes for global cognition.
Classification performance across different
Classification performance for global cognition was also examined for varying feature sets (i.e.,
21, 421, 1,200, 1,621) and sample sizes (matched vs. unmatched). No performance improve-
ments could be observed for greater feature set sizes (Feature sets 21 and 421: MBAC range:
48.42–59.31%, in 34–98% of folds BAC > dummy classifier; feature sets 1,200 and 1,621:
MBAC range: 48.96–58.72%, in 38–94% of folds BAC > dummy classifier) in both samples
across pipeline configurations and algorithms (see Figure 3C and Supporting Information
Tables S17–S20). A small difference between samples emerged in the nr condition. Relatively
higher accuracies across feature sets were found in the nr condition of the unmatched sample
than in the matched sample (Unmatched sample: MBAC range nr: 49.33–59.31%, in 44–98%
of folds BAC > dummy classifier; Matched sample: MBAC range nr: 48.96–57.41%, in 40–86%
of folds BAC > dummy classifier; see Supporting Information Tables S17–S20). This effect was
no longer found in the cr condition (Unmatched sample: MBAC range cr: 50.00–56.81%, in
42–94% of folds BAC > dummy classifier; Matched sample: MBAC range cr: 48.42–58.33%, in
34–94% of folds BAC > dummy classifier; see Figure 3C and Supporting Information Tables
S17–S20). ML performance in this specific case (nr condition/unmatched sample), however, is
most likely influenced by confounds. Overall, findings suggest that increasing feature set and
sample size may not systematically aid classification performance in our study. It, however,
further underlines the relatively low discriminatory power of the specific RSFC strength mea-
sures for the research question at stake.
Regression
In a
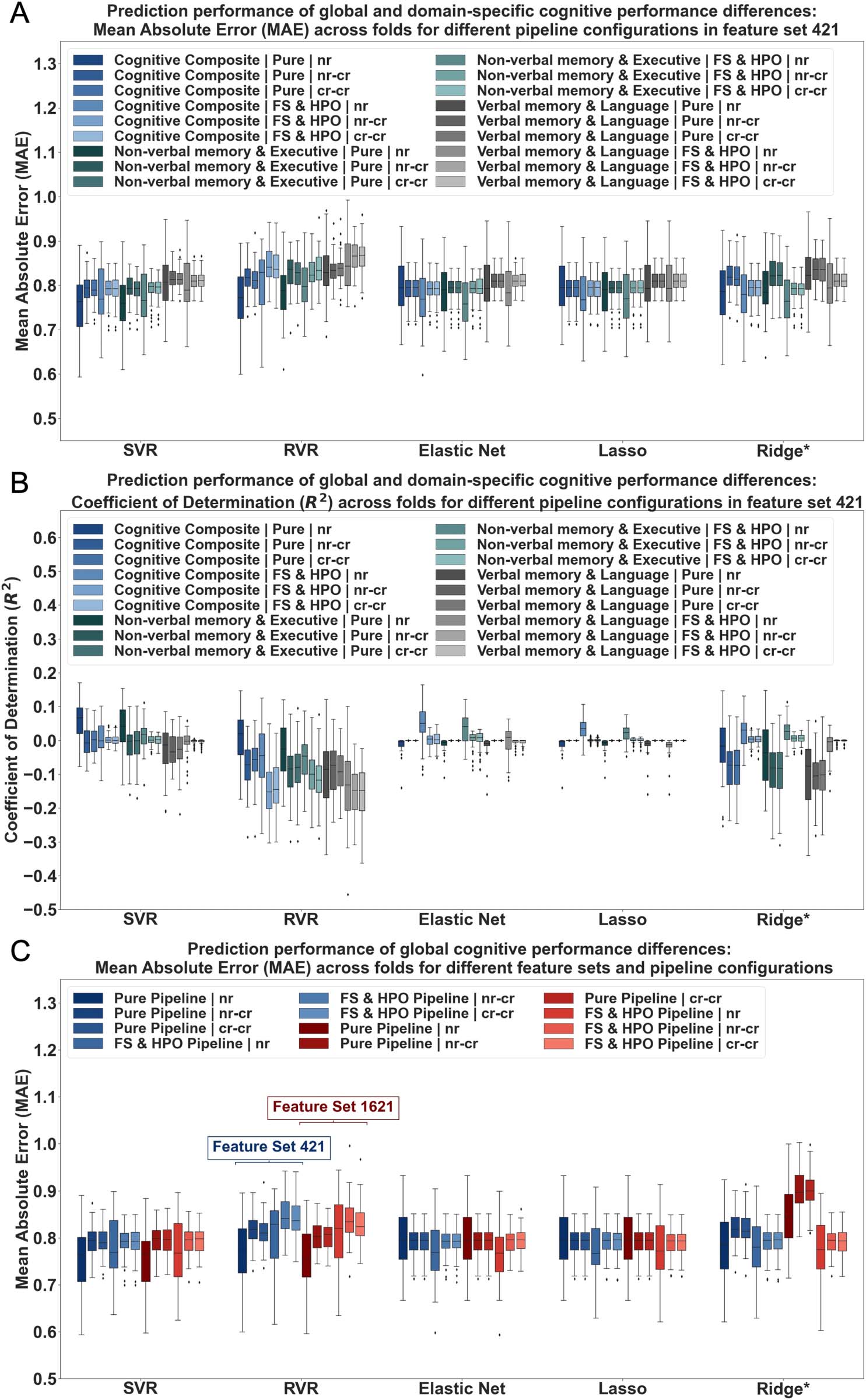
Prediction performance of global cognition and cognitive domains across pipeline configurations.
second step, ML was used to assess whether RSFC strength measures can be used to contin-
uously predict cognitive performance in older adults. ML prediction performance of global and
domain-specific cognition from RSFC strength measures was initially evaluated in feature set
421 in the unmatched sample. Across pipeline configurations and deconfounding strategies,
MAEs obtained for global and domain-specific cognition were high, ranging between 0.76 and
1.14 (see Figure 4A). Simultaneously, the coefficient of determination (R2) was found to be low
(≤0.06) or even negative, indicating that predicting the mean of cognitive scores would have
yielded better results than our model’s predictions (see Figure 4B and Supporting Information
Tables S21 and S22). The NON-VERBAL MEMORY & EXECUTIVE component revealed
slightly lower MAE and higher R2 than the VERBAL MEMORY & LANGUAGE component
across conditions (see Figure 4A and B and Supporting Information Tables S21 and S22). Nev-
ertheless, predictability compared to global cognition was similar in range. Furthermore,
results were comparable for different algorithms except for Ridge regression in pure pipelines,
which showed markedly elevated MAE, and reduced explained variance for all targets for
default values of the hyperparameter lambda (see Supporting Information Table S21). Manual
adjustment of the hyperparameter led to similar performance to other algorithms (see
Figure 4A and B and Supporting Information Table S21). No systematic predictive performance
differences were found for FS and HPO pipelines (see Figure 4A and B and Supporting Infor-
mation Tables S21 and S22). In terms of different extents of deconfounding, the nr condition
Network Neuroscience
136
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
7
1
1
2
2
2
0
7
2
1
6
8
n
e
n
_
a
_
0
0
2
7
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cognitive performance differences in older age
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
7
1
1
2
2
2
0
7
2
1
6
8
n
e
n
_
a
_
0
0
2
7
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. Regression performance results of cognitive performance differences (based on global and domain-specific cognitive scores) from RSFC
strength measures. Regression performance across algorithms: Support Vector Regression (SVR), Relevance Vector Regression (RVR), Elastic Net, LASSO
and Ridge Regression. Results shown for (A and B) cognitive composite and cognitive component scores, (A and C) different pipeline configurations
(pure (no FS/HPO) vs. FS and HPO pipelines), and (C) feature set sizes (421, 1621) (C). Ridge*: default values in pure pipeline manually adjusted; nr = no
confound regression; nr-cr = age, sex, and education regressed from target; cr-cr = age, sex, and education regressed from target and features.
Network Neuroscience
137
Cognitive performance differences in older age
resulted in slightly better prediction results compared to the other two conditions (nr: MAEs ≥
0.76; R2 ≤ 0.06; nr-cr and cr-cr: MAEs ≥ 0.79; R2 ≤ 0.00; see Supporting Information
Table S21). This was also reflected in an improved robustness against the dummy regressor
(see Figure 4C and Supporting Information Table S22). Nevertheless, it should be kept in mind
that still only a limited number of models were consistently outperforming the dummy esti-
mates in more than 80% of folds. Jointly, these results suggest that RSFC strength measures
may not contain sufficient information to reliably predict global and domain-specific cognitive
performance in older adults from a population-based cohort.
Prediction performance across varying feature set sizes for global cognition. Feature set size did
only have minimal impact in the classification setting. To verify the impact of varying feature
combinations and number of features in ML prediction, feature set 421, which was used for
comparability purposes throughout the analyses, and 1,621, which contains all possible fea-
tures, were chosen for closer examination in the regression setting. Thus, ML performance esti-
mations were examined in different pipeline configurations for global cognition. Across feature
sets and deconfounding strategies, the MAE was again found to be high (≥0.75) and the coef-
ficient of determination to be low (≤0.07) (see Supporting Information Tables S23 and S24).
The impact of different algorithms, pipeline configurations, and extents of deconfounding on
ML performance was again found to be minimal and to follow a similar pattern as before (see
Figure 4C). No significant performance differences in terms of MAE and R2 emerged for differ-
ent feature set sizes (see Figure 4C and Supporting Information Tables S23 and S24). Thus,
findings suggest in addition to minimal discriminatory power also low predictive potential
of cognitive performance differences in healthy older adults across feature sets, deconfounding
strategies, and pipeline configurations from RSFC strength measures.
Validation Analyses
Finally, we investigated the impact of a finer grained parcellation on ML performance. Results
suggest that a higher granularity has only little impact on ML performance. Classification accu-
racies ranged between 47.79% and 56.53% across feature sets and pipeline configurations for
the 800-node parcellation (see Supporting Information Tables S25 and S26 and Figure S28A),
compared to the 48.42% to 58.33% range obtained for the 400-node parcellation. Prediction
performance was found to be equally low as in the initial parcellation with high MAEs (≥0.75)
and low to none explained variance (R2 ≤ 0.07) for different feature sets and pipeline config-
urations (see Supporting Information Table S27 and Figure S28B). Thus, no benefit of a higher
granularity was observed. Furthermore, ML performance was examined in males and females
separately. Classification performance in male and female samples equally did not exceed
60% accuracy for global cognition (MBAC: 49.69–55.57%; see Supporting Information
Tables S29 and S30 and Figure S32A). Prediction performance in male and female samples
revealed comparable high MAEs (≥0.73) and low R2 (≤0.04) (see Supporting Information
Table S31 and Figure S32B). Findings, hence, further emphasize results found in the main anal-
ysis. Moreover, classification and prediction performance was assessed using connectivity esti-
mates based on (i) positive and negative correlations and (ii) only negative correlations. For
connectivity estimates based on positive and negative correlation values, classification perfor-
mance varied between 47.91% to 56.25% BAC for global cognition across algorithms, feature
sets and pipeline configurations (see Supporting Information Table S33 and Figure S35A). Pre-
diction performance equally resembled results from the main analysis (MAEs ≥ 0.75; R2 ≤
0.08; see Supporting Information Table S34 and Figure S35B). A similar pattern of results
emerged for strength measures derived from negative correlations. Classification performance
varied between 48.42% to 54.73% BAC for global cognition across algorithms, feature sets,
Network Neuroscience
138
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
7
1
1
2
2
2
0
7
2
1
6
8
n
e
n
_
a
_
0
0
2
7
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cognitive performance differences in older age
and pipeline configurations (see Supporting Information Table S36). In turn, prediction perfor-
mance was found to be equally low (MAEs ≥ 0.77; R2 ≤ 0.05; see Supporting Information
Table S37). Adding further information from anticorrelations, thus, did not appear to improve
ML performance. Furthermore, we investigated classification performance in extreme cogni-
tive groups. Across samples, pipelines, feature sets, and algorithms, classification performance
ranged between 49.70% to 62.50% BAC (see Supporting Information Tables S38 and S39).
Although slightly better classification results were achieved for extreme cognitive groups, over-
all performance remained limited. This suggests that low classification results may not be pri-
marily driven by difficulties in identifying participants close to the median and provides further
sustenance to our findings from the main analyses. An age prediction and classification frame-
work was chosen for validating our ML pipeline. In the classification of extreme age groups,
highest classification performance was obtained for linear SVM in the pure and HPO pipeline
with 85.13% and 83.13% accuracy, respectively (see Supporting Information Table S40). For
the continuous prediction of age, RSFC strength measures were found to overall predict age
reasonably well with R2 in the best cases ranging between 0.3 and 0.4 (extreme and whole
sample across age spectrum; see Supporting Information Table S41). In comparison to dummy
estimates, these models also showed reliably higher performance (see Supporting Information
Table S42). While the obtained MAEs across samples were not competitive with those reported
in the literature, results from the validation analyses, nevertheless, generally support the view
that the current pipeline may yield reasonable prediction and classification performances
(Liem et al., 2017; Pläschke et al., 2017; Vergun et al., 2013; Vieira et al., 2022). Thus, the
low ML performance estimates may be specific to the setting of classifying and predicting cog-
nitive performance differences from RSFC strength measures in healthy older adults rather than
a general finding pertained to the ML setup, parcellation granularity, sampling, or features.
DISCUSSION
The aim of the current investigation was to examine whether global and domain-specific cog-
nitive performance differences may be successfully distinguished and predicted from RSFC
strength measures in a large sample of older adults by using a systematic assessment of stan-
dard ML approaches. Results showed that classification and regression performance failed to
reach adequate discriminatory and predictive power at the individual level. Importantly, these
results persisted across different feature sets, algorithms, and pipeline configurations.
The present findings add to the notion that predicting cognition from the functional network
architecture may yield heterogeneous findings (Dubois et al., 2018; Finn et al., 2015; Rasero
et al., 2021; Vieira et al., 2022). For instance, RSFC patterns expressed in functional connec-
tivity matrices have been shown to explain up to 20% of variance in a composite cognition
score (NIH Cognitive Battery) and in a general intelligence factor (factor analysis) in two sam-
ples of the Human Connectome Project (HCP) S1200 young adult release (Dhamala et al.,
2021; Dubois et al., 2018). In contrast, global cognition (NIH Cognitive Battery; cf. Dhamala
et al., 2021) was predicted to a notably smaller degree from RSFC in young adults (median
R2 = 0.016) (Rasero et al., 2021). In older adults, Vieira et al., (2022) reported RSFC to not
predict prospective global cognitive decline, that is, change in two clinical assessments
(OASIS-3 project; median R2
CDR = 0.01). Our results further emphasize
that across different analytic choices RSFC strength measures may not reliably capture cogni-
tive performance variations in older aged adults. In light of our goal of robust and accurate
classification and prediction at the individual level, the minimum acceptable prediction accu-
racy is achieved only if the model outperforms the dummy estimate in more than 80% of the
MMSE = 0; median R2
Network Neuroscience
139
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
7
1
1
2
2
2
0
7
2
1
6
8
n
e
n
_
a
_
0
0
2
7
5
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cognitive performance differences in older age
folds. This threshold is not met by the majority of our classification and prediction models,
hinting at a limited potential as biomarker for age-related cognitive decline. Validation anal-
yses further highlight the specificity of our results to cognitive abilities. RSFC strength measures
could be used to successfully classify extreme age groups and moderately predict age (Meier
et al., 2012; Pläschke et al., 2017; Vergun et al., 2013). RSFC patterns underlying cognition,
however, may be more difficult to discern with current analytic tools, leading to mixed or null
results. It should be stressed that null results may be highly informative as they provide impor-
tant insights for future research, support a more realistic and unbiased view on brain-behavior
relations, and allow for learning from experiences across the field (Janssen et al., 2018;
Masouleh et al., 2019). Nevertheless, they tend to be underreported in the literature, leading
to a potential publication bias (Janssen et al., 2018).
Successful prediction or classification of cognitive functioning from RSFC patterns has been
reported previously (Dhamala et al., 2021; Dubois et al., 2018; Hojjati et al., 2017; Khazaee
et al., 2016; Rosenberg et al., 2016; Yoo et al., 2018). One possible explanation for the fact
that the results could not be replicated is related to the composition of the sample. Most pre-
vious studies reporting satisfactory ML performance focused on younger cohorts or patient
populations (Dhamala et al., 2021; Dubois et al., 2018; Hojjati et al., 2017; Khazaee et al.,
2016; Rosenberg et al., 2016; Yoo et al., 2018). In comparison to younger samples (Mage < 30),
low discriminatory and predictive power in the current study may be attributable to a more
complex link between RSFC and cognition evolving during the aging process (Dhamala et al.,
2021; Dubois et al., 2018; Rosenberg et al., 2016; Yoo et al., 2018). Aging is not only asso-
ciated with cognitive decline and functional network reorganization, but also with an increas-
ing interindividual variability (Andrews-Hanna et al., 2007; Chan et al., 2014; Chong et al.,
2019; Fjell et al., 2015; Grady et al., 2016; Habib et al., 2007; Hartshorne & Germine, 2015;
Hedden & Gabrieli, 2004; Mowinckel et al., 2012; Ng et al., 2016; Onoda et al., 2012;
Stumme et al., 2020). Consequently, the RSFC patterns that explain cognitive performance
levels in older adults are more difficult to identify (Scarpazza et al., 2020).
When comparing promising patient classification results to the current results, effect sizes
might be responsible for the unsatisfactory ML performance (Amaefule et al., 2021; Cui &
Gong, 2018; Kwak et al., 2021). For example, patients with MCI and AD show markedly
altered functional network organization compared to cognitively normal older adults (Badhwar
et al., 2017; Brier et al., 2014; Buckner et al., 2009; Greicius et al., 2004; Sanz-Arigita et al.,
2010; Wang et al., 2013). The sizable alterations related to pathological aging are reflected in
encouraging results in patient classification (de Vos et al., 2018; Dyrba et al., 2015; Hojjati
et al., 2017; Khazaee et al., 2016; Teipel et al., 2017). For instance, ML performance in patient
classification (HC vs. MCI vs. AD) based on RSFC graph metrics reached above 88% accuracy
(Hojjati et al., 2017; Khazaee et al., 2016). Nevertheless, these effect sizes may not be found
for healthy older populations. For instance, cognition could be significantly predicted in sam-
ples of cognitive normal and clinically impaired older adults from whole-brain RSFC patterns (r
= 0.08–0.44) (Kwak et al., 2021). However, prediction accuracy dropped substantially once
models were trained only on clinically unimpaired older adults (r = −0.04–0.24) (Kwak et al.,
2021). Accurate cognitive performance prediction from RSFC patterns in older aged adults
without the inclusion of clinical populations may, hence, be impeded by small effect sizes.
Another aspect that needs to be addressed when discussing the low ML performance con-
cerns the cognitive parameters used. Most studies including older cohorts have focused on
specific cognitive abilities (Avery et al., 2020; Fountain-Zaragoza et al., 2019; Gao et al.,
2020; Kwak et al., 2021; Pläschke et al., 2020). For instance, WM capacity could be success-
fully predicted from meta-analytically defined RSFC networks in older individuals (Pläschke
Network Neuroscience
140
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
1
2
2
2
0
7
2
1
6
8
n
e
n
_
a
_
0
0
2
7
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cognitive performance differences in older age
et al., 2020). This may be due to a more explicit mapping of RSFC patterns to specific cognitive
abilities than for general or clustered cognitive abilities, which we were interested in (Avery
et al., 2020; Gao et al., 2020; Kwak et al., 2021).
Furthermore, most prior studies have used pair-wise functional connectivity as input fea-
tures (Avery et al., 2020; Dhamala et al., 2021; Dubois et al., 2018; Gao et al., 2020; He
et al., 2020; Pläschke et al., 2020). We used functional connectivity estimates linked to cog-
nitive performance differences in aging and with promising classification performance in neu-
rodegenerative diseases (Chan et al., 2014; Hausman et al., 2020; Hojjati et al., 2017; Iordan
et al., 2018; Khazaee et al., 2016; Malagurski et al., 2020; Ng et al., 2016; Stumme et al.,
2020). Findings highlight that for reliably detecting cognitive performance differences in nor-
mally aging individuals, the additional dimensionality reduction inherent to the calculation of
RSFC strength values may be too extensive, that is, relevant information for ML was lost during
the computation (Cui & Gong, 2018; Lei et al., 2020). Also, redundancy of feature information,
that is, within- and inter-network connectivity, may have resulted in poorer ML performance,
especially in larger feature sets (Mwangi et al., 2014).
Methodological Considerations and Future Outlook
While the current investigation concentrated on RSFC strength measures, future studies might
use other imaging features, that is, more complex graph metrics, such as betweenness centrality
or modularity, multimodal or task-based fMRI data, to improve the prediction of cognitive per-
formance in older age (Draganski et al., 2013; Gbadeyan et al., 2022; McConathy & Sheline,
2015; Pacheco et al., 2015; Sripada et al., 2020b; Vieira et al., 2022). For example, prior
research has shown that global cognitive abilities could be better predicted from task-based than
resting-state fMRI data in large samples of younger adults from the HCP dataset (Greene et al.,
2018; Sripada et al., 2020a). Along these lines, it may be interesting to investigate whether task-
based fMRI data in these circumstances also outperforms RSFC in older adults. Likewise, it is also
warranted to keep a distinction between basic research and clinical applicability. Classification
and prediction results might already be informative, if they are statistically significant in
healthy subjects; however, they may not be practically relevant for the clinical context.
Furthermore, only cross-sectional data has been used in the current investigation. Although
important insights can be gained cross-sectionally, the investigation of longitudinal data
becomes indispensable in the biomarker development for prospective age-related cognitive
decline (Davatzikos et al., 2009; Liem et al., 2021). Initial efforts to predict future cognitive
decline from imaging and nonimaging data have revealed promising results (Vieira et al., 2022).
A further methodological consideration pertains to the choice of data preparation steps, for
example, the parcellation scheme and choice of network assignment (Dubois et al., 2018). In
the current investigation, a functional network parcellation derived from younger brains was
used, which directly links brain networks to behavioral processing and is commonly used in
lifespan studies (Schaefer et al., 2018; Yeo et al., 2011). Although ML performance in the
current study was low regardless of data preparation, that is, parcellation granularity, and
ML model choices, future studies are warranted to examine generalizability to other
population-based cohorts of older aged adults and other functional network divisions.
Conclusions
The present study addressed the biomarker potential of RSFC strength measures for cognitive
performance differences in normal aging in a systematic evaluation of standard ML
approaches. Present results across different analytic choices emphasize that the potential of
Network Neuroscience
141
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
1
2
2
2
0
7
2
1
6
8
n
e
n
_
a
_
0
0
2
7
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cognitive performance differences in older age
RSFC strength measures as sole biomarker for age-related cognitive decline may be limited.
Findings add to past research demonstrating that reliable cognitive performance prediction
and distinction in healthy older adults based on RSFC strength measures may be challenging
due to small effects, high heterogeneity, and the removal of relevant information during the
computation of these parameters. Although current results are far from promising, they still
may prove useful in providing guidance on future research targets. Specifically, multimodal
and longitudinal approaches appear warranted in future studies developing a robust biomarker
for cognitive performance in healthy aging.
ACKNOWLEDGMENTS
This project was partially funded by the German National Cohort and the 1000BRAINS-Study
of the Institute of Neuroscience and Medicine, Research Centre Jülich, Germany. We thank the
Heinz Nixdorf Foundation (Germany) for the generous support of the Heinz Nixdorf Study. We
thank the investigative group and the study staff of the Heinz Nixdorf Recall Study and
1000BRAINS. This research was supported by the Joint Lab Supercomputing and Modeling
for the Human Brain. The authors gratefully acknowledge the computing time granted through
JARA on the supercomputer JURECA ( Jülich Supercomputing Centre, 2021) at Forschungszen-
trum Jülich.
SUPPORTING INFORMATION
Supporting information for this article is available at https://doi.org/10.1162/netn_a_00275.
AUTHOR CONTRIBUTIONS
Camilla Krämer: Conceptualization; Formal analysis; Methodology; Visualization; Writing –
original draft; Writing – review & editing. Johanna Stumme: Formal analysis; Methodology;
Writing – review & editing. Lucas da Costa Campos: Formal analysis; Methodology; Writing –
review & editing. Christian Rubbert: Methodology; Writing – review & editing. Julian Caspers:
Conceptualization; Methodology; Writing – review & editing. Svenja Caspers: Conceptualiza-
tion; Funding acquisition; Resources; Supervision; Writing – review & editing. Christiane
Jockwitz: Conceptualization; Methodology; Supervision; Writing – review & editing.
FUNDING INFORMATION
Svenja Caspers, European Union’s Horizon 2020 Research and Innovation Programme (HBP
SGA3), Award ID: Grant Agreement No. 945539.
REFERENCES
Afyouni, S., & Nichols, T. E. (2018). Insight and inference for
DVARS. NeuroImage, 172, 291–312. https://doi.org/10.1016/j
.neuroimage.2017.12.098, PubMed: 29307608
Amaefule, C. O., Dyrba, M., Wolfsgruber, S., Polcher, A., Schneider,
A., Fliessbach, K., Spottke, A., Meiberth, D., Preis, L., Peters, O.,
Incesoy, E. I., Spruth, E. J., Priller, J., Altenstein, S., Bartels, C.,
Wiltfang, J., Janowitz, D., Bürger, K., Laske, C., … Teipel, S. J.
(2021). Association between composite scores of domain-specific
cognitive functions and regional patterns of atrophy and
functional connectivity in the Alzheimer’s disease spectrum.
NeuroImage: Clinical, 29, 102533. https://doi.org/10.1016/j.nicl
.2020.102533, PubMed: 33360018
Andrews-Hanna, J. R., Snyder, A. Z., Vincent, J. L., Lustig, C., Head,
D., Raichle, M. E., & Buckner, R. L. (2007). Disruption of large-scale
brain systems in advanced aging. Neuron, 56(5), 924–935. https://
doi.org/10.1016/j.neuron.2007.10.038, PubMed: 18054866
Arbabshirani, M. R., Plis, S., Sui, J., & Calhoun, V. D. (2017). Single
subject prediction of brain disorders in neuroimaging: Promises
and pitfalls. NeuroImage, 145, 137–165. https://doi.org/10.1016
/j.neuroimage.2016.02.079, PubMed: 27012503
Network Neuroscience
142
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
1
2
2
2
0
7
2
1
6
8
n
e
n
_
a
_
0
0
2
7
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cognitive performance differences in older age
Ashburner, J., & Friston, K. J. (2005). Unified segmentation. Neuro-
Image, 26(3), 839–851. https://doi.org/10.1016/j.neuroimage
.2005.02.018, PubMed: 15955494
Avery, E. W., Yoo, K., Rosenberg, M. D., Greene, A. S., Gao, S., Na,
D. L., Scheinost, D., Constable, T. R., & Chun, M. M. (2020). Dis-
tributed patterns of functional connectivity predict working
memory performance in novel healthy and memory-impaired
individuals. Journal of Cognitive Neuroscience, 32(2), 241–255.
https://doi.org/10.1162/jocn_a_01487, PubMed: 31659926
Badhwar, A., Tam, A., Dansereau, C., Orban, P., Hoffstaedter, F., &
Bellec, P. (2017). Resting-state network dysfunction in
Alzheimer’s disease: A systematic review and meta-analysis.
Alzheimer’s & Dementia: Diagnosis, Assessment & Disease
Monitoring, 8(1), 73–85. https://doi.org/10.1016/j.dadm.2017
.03.007, PubMed: 28560308
Brier, M. R., Thomas, J. B., Fagan, A. M., Hassenstab, J., Holtzman,
D. M., Benzinger, T. L., Morris, J. C., & Ances, B. M. (2014). Func-
tional connectivity and graph theory in preclinical Alzheimer’s
disease. Neurobiology of Aging, 35(4), 757–768. https://doi.org
/10.1016/j.neurobiolaging.2013.10.081, PubMed: 24216223
Brown, C. J., & Hamarneh, G. (2016). Machine learning on human
connectome data from MRI. ArXiv:1611.08699. https://doi.org
/10.48550/arXiv.1611.08699
Buckner, R. L., Sepulcre, J., Talukdar, T., Krienen, F. M., Liu, H.,
Hedden, T., Andrews-Hanna, J. R., Sperling, R. A., & Johnson,
K. A. (2009). Cortical hubs revealed by intrinsic functional
connectivity: Mapping, assessment of stability, and relation to Alzhei-
mer’s disease. Journal of Neuroscience, 29(6), 1860–1873. https://
doi.org/10.1523/JNEUROSCI.5062-08.2009, PubMed: 19211893
Burgess, G. C., Kandala, S., Nolan, D., Laumann, T. O., Power,
J. D., Adeyemo, B., Harms, M. P., Petersen, S. E., & Barch,
D. M. (2016). Evaluation of denoising strategies to address
motion-correlated artifacts in resting-state functional magnetic
resonance imaging data from the Human Connectome Project.
Brain Connectivity, 6(9), 669–680. https://doi.org/10.1089/brain
.2016.0435, PubMed: 27571276
Cabeza, R. (2001). Cognitive neuroscience of aging: Contributions
of functional neuroimaging. Scandinavian Journal of Psychology,
42(3), 277–286. https://doi.org/10.1111/1467-9450.00237,
PubMed: 11501741
Calhoun, V. D., Wager, T. D., Krishnan, A., Rosch, K. S., Seymour,
K. E., Nebel, M. B., Mostofsky, S. H., Nyalakanai, P., & Kiehl, K.
(2017). The impact of T1 versus EPI spatial normalization templates
for fMRI data analyses. Human Brain Mapping, 38(11), 5331–
5342. https://doi.org/10.1002/hbm.23737, PubMed: 28745021
Caspers, S., Moebus, S., Lux, S., Pundt, N., Schütz, H., Mühleisen,
T. W., Gras, V., Eickhoff, S. B., Romanzetti, S., Stöcker, T.,
Stirnberg, R., Kirlangic, M. E., Minnerop, M., Pieperhoff, P.,
Mödder, U., Das, S., Evans, A. C., Jöckel, K.-H., Erbel, R., …
Amunts, K. (2014). Studying variability in human brain aging in
a population-based German cohort-rationale and design of
1000BRAINS. Frontiers in Aging Neuroscience, 6, 149. https://
doi.org/10.3389/fnagi.2014.00149, PubMed: 25071558
Chan, M. Y., Park, D. C., Savalia, N. K., Petersen, S. E., & Wig, G. S.
(2014). Decreased segregation of brain systems across the
healthy adult lifespan. Proceedings of the National Academy of
Sciences, 111(46), E4997–E5006. https://doi.org/10.1073/pnas
.1415122111, PubMed: 25368199
Chong, J. S. X., Ng, K. K., Tandi, J., Wang, C., Poh, J.-H., Lo, J. C.,
Chee, M. W. L., & Zhou, J. H. (2019). Longitudinal changes in the
cerebral cortex functional organization of healthy elderly. Journal
of Neuroscience, 39(28), 5534–5550. https://doi.org/10.1523
/JNEUROSCI.1451-18.2019, PubMed: 31109962
Ciric, R., Wolf, D. H., Power, J. D., Roalf, D. R., Baum, G. L.,
Ruparel, K., Shinohara, R. T., Elliott, M. A., Eickhoff, S. B.,
Davatzikos, C., Gur, R. C., Gur, R. E., Bassett, D. S., & Satterthwaite,
T. D. (2017). Benchmarking of participant-level confound regres-
sion strategies for the control of motion artifact in studies of func-
tional connectivity. NeuroImage, 154, 174–187. https://doi.org/10
.1016/j.neuroimage.2017.03.020, PubMed: 28302591
Cui, Z., & Gong, G. (2018). The effect of machine learning regres-
sion algorithms and sample size on individualized behavioral
prediction with functional connectivity features. NeuroImage,
178, 622–637. https://doi.org/10.1016/j.neuroimage.2018.06
.001, PubMed: 29870817
Dadi, K., Rahim, M., Abraham, A., Chyzhyk, D., Milham, M.,
Thirion, B., & Varoquaux, G. (2019). Benchmarking functional
connectome-based predictive models for resting-state fMRI.
NeuroImage, 192, 115–134. https://doi.org/10.1016/j.neuroimage
.2019.02.062, PubMed: 30836146
Dadi, K., Varoquaux, G., Houenou, J., Bzdok, D., Thirion, B., & Engemann,
D. (2021). Population modeling with machine learning can enhance
measures of mental health. GigaScience, 10(10), giab071.
https://doi.org/10.1093/gigascience/giab071, PubMed: 34651172
Dai, Z., Yan, C., Li, K., Wang, Z., Wang, J., Cao, M., Lin, Q., Shu,
N., Xia, M., Bi, Y., & He, Y. (2015). Identifying and mapping con-
nectivity patterns of brain network hubs in Alzheimer’s disease.
Cerebral Cortex, 25(10), 3723–3742. https://doi.org/10.1093
/cercor/bhu246, PubMed: 25331602
Damoiseaux, J. S., Beckmann, C. F., Arigita, E. J. S, Barkhof, F.,
Scheltens, P., Stam, C. J., Smith, S. M., & Rombouts, S. A. R. B.
(2008). Reduced resting-state brain activity in the “default net-
work” in normal aging. Cerebral Cortex, 18(8), 1856–1864.
https://doi.org/10.1093/cercor/bhm207, PubMed: 18063564
Davatzikos, C., Xu, F., An, Y., Fan, Y., & Resnick, S. M. (2009). Longitudinal
progression of Alzheimer’s-like patterns of atrophy in normal
older adults: The SPARE-AD index. Brain, 132(8), 2026–2035.
https://doi.org/10.1093/brain/awp091, PubMed: 19416949
de Vos, F., Koini, M., Schouten, T. M., Seiler, S., van der Grond, J.,
Lechner, A., Schmidt, R., de Rooij, M., & Rombouts, S. A. R. B.
(2018). A comprehensive analysis of resting state fMRI measures
to classify individual patients with Alzheimer’s disease. Neuro-
Image, 167, 62–72. https://doi.org/10.1016/j.neuroimage.2017
.11.025, PubMed: 29155080
Deary, I. J., Corley, J., Gow, A. J., Harris, S. E., Houlihan, L. M., Marioni,
R. E., Penke, L., Rafnsson, S. B., & Starr, J. M. (2009). Age-associated
cognitive decline. British Medical Bulletin, 92(1), 135–152.
https://doi.org/10.1093/bmb/ldp033, PubMed: 19776035
Depp, C. A., & Jeste, D. V. (2006). Definitions and predictors of suc-
cessful aging: A comprehensive review of larger quantitative
studies. The American Journal of Geriatric Psychiatry, 14(1),
6–20. https://doi.org/10.1097/01.JGP.0000192501.03069.bc,
PubMed: 16407577
Dhamala, E., Jamison, K. W., Jaywant, A., Dennis, S., & Kuceyeski,
A. (2021). Distinct functional and structural connections predict
Network Neuroscience
143
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
1
2
2
2
0
7
2
1
6
8
n
e
n
_
a
_
0
0
2
7
5
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cognitive performance differences in older age
crystallised and fluid cognition in healthy adults. Human Brain
Mapping, 42(10), 3102–3118. https://doi.org/10.1002/ hbm
.25420, PubMed: 33830577
Dohmatob, E., Varoquaux, G., & Thirion, B. (2018). Inter-subject
registration of functional images: Do we need anatomical
images? Frontiers in Neuroscience, 12, 64. https://doi.org/10
.3389/fnins.2018.00064, PubMed: 29497357
Draganski, B., Lutti, A., & Kherif, F. (2013). Impact of brain aging
and neurodegeneration on cognition: Evidence from MRI.
Current Opinion in Neurology, 26(6), 640–645. https://doi.org
/10.1097/ WCO.0000000000000029, PubMed: 24184970
Dubois, J., Galdi, P., Paul, L. K., & Adolphs, R. (2018). A distributed
brain network predicts general intelligence from resting-state
human neuroimaging data. Philosophical Transactions of the
Royal Society B: Biological Sciences, 373(1756), 20170284.
https://doi.org/10.1098/rstb.2017.0284, PubMed: 30104429
Dyrba, M., Grothe, M., Kirste, T., & Teipel, S. J. (2015). Multimodal
analysis of functional and structural disconnection in
Alzheimer’s disease using multiple kernel SVM: Functional and
structural disconnection in AD. Human Brain Mapping, 36(6),
2118–2131. https://doi.org/10.1002/ hbm.22759, PubMed:
25664619
Engemann, D. A., Kozynets, O., Sabbagh, D., Lemaître, G.,
Varoquaux, G., Liem, F., & Gramfort, A. (2020). Combining
magnetoencephalography with magnetic resonance imaging
enhances learning of surrogate-biomarkers. ELife, 9, e54055.
https://doi.org/10.7554/eLife.54055, PubMed: 32423528
Farahani, F. V., Karwowski, W., & Lighthall, N. R. (2019). Applica-
tion of graph theory for identifying connectivity patterns in
human brain networks: A systematic review. Frontiers in Neuro-
science, 13, 585. https://doi.org/10.3389/fnins.2019.00585,
PubMed: 31249501
Finn, E. S., Shen, X., Scheinost, D., Rosenberg, M. D., Huang, J.,
Chun, M. M., Papademetris, X., & Constable, R. T. (2015). Func-
tional connectome fingerprinting: Identifying individuals using
patterns of brain connectivity. Nature Neuroscience, 18(11),
1664–1671. https://doi.org/10.1038/nn.4135, PubMed:
26457551
Fjell, A. M., Sneve, M. H., Grydeland, H., Storsve, A. B., de Lange,
A.-M. G., Amlien, I. K., Røgeberg, O. J., & Walhovd, K. B. (2015).
Functional connectivity change across multiple cortical networks
relates to episodic memory changes in aging. Neurobiology of
Aging, 36(12), 3255–3268. https://doi.org/10.1016/j.neurobiolaging
.2015.08.020, PubMed: 26363813
Fountain-Zaragoza, S., Samimy, S., Rosenberg, M. D., & Prakash,
R. S. (2019). Connectome-based models predict attentional con-
trol in aging adults. NeuroImage, 186, 1–13. https://doi.org/10
.1016/j.neuroimage.2018.10.074, PubMed: 30394324
Gao, M., Wong, C. H. Y., Huang, H., Shao, R., Huang, R., Chan,
C. C. H., & Lee, T. M. C. (2020). Connectome-based models
can predict processing speed in older adults. NeuroImage, 223,
117290. https://doi.org/10.1016/j.neuroimage.2020.117290,
PubMed: 32871259
Gaser, C., Dahnke, R., Thompson, P. M., Kurth, F., Luders, E., &
Alzheimer’s Disease Neuroimaging Initiative. (2022). CAT—A
computational anatomy toolbox for the analysis of structural
MRI data. bioRxiv. https://doi.org/10.1101/2022.06.11.495736
Gbadeyan, O., Teng, J., & Prakash, R. S. (2022). Predicting
response time variability from task and resting-state functional
connectivity in the aging brain. NeuroImage, 250, 118890.
https://doi.org/10.1016/j.neuroimage.2022.118890, PubMed:
35007719
Grady, C., Sarraf, S., Saverino, C., & Campbell, K. (2016). Age dif-
ferences in the functional interactions among the default, fronto-
parietal control, and dorsal attention networks. Neurobiology of
Aging, 41, 159–172. https://doi.org/10.1016/j.neurobiolaging
.2016.02.020, PubMed: 27103529
Greene, A. S., Gao, S., Scheinost, D., & Constable, R. T. (2018).
Task-induced brain state manipulation improves prediction of
individual traits. Nature Communications, 9(1), 2807. https://
doi.org/10.1038/s41467-018-04920-3, PubMed: 30022026
Greicius, M. D., Srivastava, G., Reiss, A. L., & Menon, V. (2004).
Default-mode network activity distinguishes Alzheimer’s disease
from healthy aging: Evidence from functional MRI. Proceedings
of the National Academy of Sciences, 101(13), 4637–4642.
https://doi.org/10.1073/pnas.0308627101, PubMed: 15070770
Guyon, I., & Elisseeff, A. (2003). An introduction to variable and
feature selection. Journal of Machine Learning Research, 3,
1157–1182.
Habib, R., Nyberg, L., & Nilsson, L.-G. (2007). Cognitive and
non-cognitive factors contributing to the longitudinal identifica-
tion of successful older adults in the Betula study. Aging, Neuro-
psychology, and Cognition, 14(3), 257–273. https://doi.org/10
.1080/13825580600582412, PubMed: 17453560
Hartshorne, J. K., & Germine, L. T. (2015). When does cognitive
functioning peak? The asynchronous rise and fall of different cog-
nitive abilities across the life span. Psychological Science, 26(4),
433–443. https://doi.org/10.1177/0956797614567339, PubMed:
25770099
Hausman, H. K., O’Shea, A., Kraft, J. N., Boutzoukas, E. M.,
Evangelista, N. D., Van Etten, E. J., Bharadwaj, P. K., Smith,
S. G., Porges, E., Hishaw, G. A., Wu, S., DeKosky, S., Alexander,
G. E., Marsiske, M., Cohen, R., & Woods, A. J. (2020). The role of
resting-state network functional connectivity in cognitive aging.
Frontiers in Aging Neuroscience, 12, 177. https://doi.org/10.3389
/fnagi.2020.00177, PubMed: 32595490
He, T., Kong, R., Holmes, A. J., Nguyen, M., Sabuncu, M. R., Eickhoff,
S. B., Bzdok, D., Feng, J., & Yeo, B. T. T. (2020). Deep neural
networks and kernel regression achieve comparable accuracies for
functional connectivity prediction of behavior and demographics.
NeuroImage, 206, 116276. https://doi.org/10.1016/j.neuroimage
.2019.116276, PubMed: 31610298
Hedden, T., & Gabrieli, J. D. E. (2004). Insights into the ageing mind:
A view from cognitive neuroscience. Nature Reviews Neurosci-
ence, 5(2), 87–96. https://doi.org/10.1038/nrn1323, PubMed:
14735112
Hojjati, S. H., Ebrahimzadeh, A., Khazaee, A., & Babajani-Feremi,
A. (2017). Predicting conversion from MCI to AD using
resting-state fMRI, graph theoretical approach and SVM. Journal
of Neuroscience Methods, 282, 69–80. https://doi.org/10.1016/j
.jneumeth.2017.03.006, PubMed: 28286064
Hua, J., Tembe, W. D., & Dougherty, E. R. (2009). Performance of
feature-selection methods in the classification of high-dimension
data. Pattern Recognition, 42(3), 409–424. https://doi.org/10
.1016/j.patcog.2008.08.001
Network Neuroscience
144
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
1
2
2
2
0
7
2
1
6
8
n
e
n
_
a
_
0
0
2
7
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cognitive performance differences in older age
Iordan, A. D., Cooke, K. A., Moored, K. D., Katz, B., Buschkuehl,
M., Jaeggi, S. M., Jonides, J., Peltier, S. J., Polk, T. A., & Reuter-
Lorenz, P. A. (2018). Aging and network properties: Stability over
time and links with learning during working memory training.
Frontiers in Aging Neuroscience, 9, 419. https://doi.org/10.3389
/fnagi.2017.00419, PubMed: 29354048
Janssen, R. J., Mourão-Miranda, J., & Schnack, H. G. (2018). Mak-
ing individual prognoses in psychiatry using neuroimaging and
machine learning. Biological Psychiatry: Cognitive Neuroscience
and Neuroimaging, 3(9), 798–808. https://doi.org/10.1016/j.bpsc
.2018.04.004, PubMed: 29789268
Kalbe, E., Kessler, J., Calabrese, P., Smith, R., Passmore, A. P.,
Brand, M., & Bullock, R. (2004). DemTect: A new, sensitive cog-
nitive screening test to support the diagnosis of mild cognitive
impairment and early dementia. International Journal of Geriatric
Psychiatry, 19(2), 136–143. https://doi.org/10.1002/gps.1042,
PubMed: 14758579
Kazeminejad, A., & Sotero, R. C. (2019). Topological properties of
resting-state fMRI functional networks improve machine learning-
based autism classification. Frontiers in Neuroscience, 12, 1018.
https://doi.org/10.3389/fnins.2018.01018, PubMed: 30686984
Khazaee, A., Ebrahimzadeh, A., & Babajani-Feremi, A. (2016).
Application of advanced machine learning methods on resting-
state fMRI network for identification of mild cognitive impairment
and Alzheimer’s disease. Brain Imaging and Behavior, 10(3),
799–817. https://doi.org/10.1007/s11682-015-9448-7, PubMed:
26363784
Kwak, S., Kim, H., Kim, H., Youm, Y., & Chey, J. (2021). Distributed
functional connectivity predicts neuropsychological test performance
among older adults. Human Brain Mapping, 42(10), 3305–3325.
https://doi.org/10.1002/hbm.25436, PubMed: 33960591
Lei, D., Pinaya, W. H. L., van Amelsvoort, T., Marcelis, M.,
Donohoe, G., Mothersill, D. O., Corvin, A., Gill, M., Vieira, S.,
Huang, X., Lui, S., Scarpazza, C., Young, J., Arango, C., Bullmore,
E., Qiyong, G., McGuire, P., & Mechelli, A. (2020). Detecting
schizophrenia at the level of the individual: Relative diagnostic
value of whole-brain images, connectome-wide functional
connectivity and graph-based metrics. Psychological Medicine,
50(11), 1852–1861. https://doi.org/10.1017/S0033291719001934,
PubMed: 31391132
Lemm, S., Blankertz, B., Dickhaus, T., & Müller, K.-R. (2011). Introduc-
tion to machine learning for brain imaging. NeuroImage, 56(2),
387–399. https://doi.org/10.1016/j.neuroimage.2010.11.004,
PubMed: 21172442
Li, J., Kong, R., Liégeois, R., Orban, C., Tan, Y., Sun, N., Holmes,
A. J., Sabuncu, M. R., Ge, T., & Yeo, B. T. T. (2019). Global signal
regression strengthens association between resting-state functional
connectivity and behavior. NeuroImage, 196, 126–141. https://doi
.org/10.1016/j.neuroimage.2019.04.016, PubMed: 30974241
Liem, F., Geerligs, L., Damoiseaux, J. S., & Margulies, D. S. (2021).
Functional connectivity in aging. In Handbook of the psychology
of aging (pp. 37–51). Elsevier. https://doi.org/10.1016/B978-0-12
-816094-7.00010-6
Liem, F., Varoquaux, G., Kynast, J., Beyer, F., Kharabian Masouleh,
S., Huntenburg, J. M., Lampe, L., Rahim, M., Abraham, A.,
Craddock, R. C., Riedel-Heller, S., Luck, T., Loeffler, M., Schroeter,
M. L., Witte, A. V., Villringer, A., & Margulies, D. S. (2017). Pre-
dicting brain-age from multimodal imaging data captures
cognitive impairment. NeuroImage, 148, 179–188. https://doi
.org/10.1016/j.neuroimage.2016.11.005, PubMed: 27890805
Luciano, M., Gow, A. J., Harris, S. E., Hayward, C., Allerhand, M.,
Starr, J. M., Visscher, P. M., & Deary, I. J. (2009). Cognitive ability
at age 11 and 70 years, information processing speed, and APOE
variation: The Lothian Birth Cohort 1936 study. Psychology and
Aging, 24(1), 129–138. https://doi.org/10.1037/a0014780,
PubMed: 19290744
Malagurski, B., Liem, F., Oschwald, J., Mérillat, S., & Jäncke, L.
(2020). Functional dedifferentiation of associative resting state
networks in older adults—A longitudinal study. NeuroImage,
214, 116680. https://doi.org/10.1016/j.neuroimage.2020
.116680, PubMed: 32105885
Masouleh, S. K., Eickhoff, S. B., Hoffstaedter, F., Genon, S., &
Alzheimer’s Disease Neuroimaging Initiative. (2019). Empirical
examination of the replicability of associations between brain
structure and psychological variables. ELife, 8, e43464. https://
doi.org/10.7554/eLife.43464, PubMed: 30864950
McConathy, J., & Sheline, Y. I. (2015). Imaging biomarkers associ-
ated with cognitive decline: A review. Biological Psychiatry,
77(8), 685–692. https://doi.org/10.1016/j.biopsych.2014.08
.024, PubMed: 25442005
McDermott, K. L., McFall, G. P., Andrews, S. J., Anstey, K. J., & Dixon,
R. A. (2016). Memory resilience to Alzheimer’s genetic risk: Sex
effects in predictor profiles. The Journals of Gerontology Series B:
Psychological Sciences and Social Sciences, 72(6), 937–946.
https://doi.org/10.1093/geronb/gbw161, PubMed: 28025282
Meier, T. B., Desphande, A. S., Vergun, S., Nair, V. A., Song, J.,
Biswal, B. B., Meyerand, M. E., Birn, R. M., & Prabhakaran, V.
(2012). Support vector machine classification and characteriza-
tion of age-related reorganization of functional brain networks.
NeuroImage, 60(1), 601–613. https://doi.org/10.1016/j.neuroimage
.2011.12.052, PubMed: 22227886
Mowinckel, A. M., Espeseth, T., & Westlye, L. T. (2012). Network-
specific effects of age and in-scanner subject motion: A
resting-state fMRI study of 238 healthy adults. NeuroImage,
63(3), 1364–1373. https://doi.org/10.1016/j.neuroimage.2012
.08.004, PubMed: 22992492
Murphy, K., Birn, R. M., Handwerker, D. A., Jones, T. B., & Bandettini,
P. A. (2009). The impact of global signal regression on resting state
correlations: Are anti-correlated networks introduced? Neuro-
Image, 44(3), 893–905. https://doi.org/10.1016/j.neuroimage
.2008.09.036, PubMed: 18976716
Murphy, K., & Fox, M. D. (2017). Towards a consensus regarding
global signal regression for resting state functional connectivity
MRI. NeuroImage, 154, 169–173. https://doi.org/10.1016/j
.neuroimage.2016.11.052, PubMed: 27888059
Mwangi, B., Tian, T. S., & Soares, J. C. (2014). A review of feature reduc-
tion techniques in neuroimaging. Neuroinformatics, 12(2), 229–244.
https://doi.org/10.1007/s12021-013-9204-3, PubMed: 24013948
Ng, K. K., Lo, J. C., Lim, J. K. W., Chee, M. W. L., & Zhou, J. (2016).
Reduced functional segregation between the default mode net-
work and the executive control network in healthy older adults:
A longitudinal study. NeuroImage, 133, 321–330. https://doi.org
/10.1016/j.neuroimage.2016.03.029, PubMed: 27001500
Nostro, A. D., Müller, V. I., Varikuti, D. P., Pläschke, R. N.,
Hoffstaedter, F., Langner, R., Patil, K. R., & Eickhoff, S. B. (2018).
Predicting personality from network-based resting-state functional
Network Neuroscience
145
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
1
2
2
2
0
7
2
1
6
8
n
e
n
_
a
_
0
0
2
7
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cognitive performance differences in older age
connectivity. Brain Structure and Function, 223(6), 2699–2719.
https://doi.org/10.1007/s00429-018-1651-z, PubMed: 29572625
Onoda, K., Ishihara, M., & Yamaguchi, S. (2012). Decreased func-
tional connectivity by aging is associated with cognitive decline.
Journal of Cognitive Neuroscience, 24(11), 2186–2198. https://
doi.org/10.1162/jocn_a_00269, PubMed: 22784277
Orrù, G., Pettersson-Yeo, W., Marquand, A. F., Sartori, G., & Mechelli,
A. (2012). Using support vector machine to identify imaging bio-
markers of neurological and psychiatric disease: A critical review.
Neuroscience & Biobehavioral Reviews, 36(4), 1140–1152. https://
doi.org/10.1016/j.neubiorev.2012.01.004, PubMed: 22305994
Pacheco, J., Goh, J. O., Kraut, M. A., Ferrucci, L., & Resnick, S. M.
(2015). Greater cortical thinning in normal older adults predicts
later cognitive impairment. Neurobiology of Aging, 36(2),
903–908. https://doi.org/10.1016/j.neurobiolaging.2014.08.031,
PubMed: 25311277
Parkes, L., Fulcher, B., Yücel, M., & Fornito, A. (2018). An evalua-
tion of the efficacy, reliability, and sensitivity of motion correc-
tion strategies for resting-state functional MRI. NeuroImage,
171, 415–436. https://doi.org/10.1016/j.neuroimage.2017.12
.073, PubMed: 29278773
Paulus, M. P., & Thompson, W. K. (2021). Computational
approaches and machine learning for individual-level treatment
predictions. Psychopharmacology, 238, 1231–1239. https://doi
.org/10.1007/s00213-019-05282-4, PubMed: 31134293
Pedregosa, F., Varoquaux, G., Gramfort, A., Michel, V., Thirion, B.,
Grisel, O., Blondel, M., Prettenhofer, P., Weiss, R., Dubourg, V.,
Vanderplas, J., Passos, A., Cournapeau, D., Brucher, M., Perrot,
M., & Duchesnay, E. (2011). Scikit-learn: Machine learning in
Python. Journal of Machine Learning Research, 12, 2825–2830.
Pervaiz, U., Vidaurre, D., Woolrich, M. W., & Smith, S. M. (2020).
Optimising network modelling methods for fMRI. NeuroImage,
211, 116604. https://doi.org/10.1016/j.neuroimage.2020
.116604, PubMed: 32062083
Pläschke, R. N., Cieslik, E. C., Müller, V. I., Hoffstaedter, F., Plachti,
A., Varikuti, D. P., Goosses, M., Latz, A., Caspers, S., Jockwitz,
C., Moebus, S., Gruber, O., Eickhoff, C. R., Reetz, K., Heller, J.,
Südmeyer, M., Mathys, C., Caspers, J., Grefkes, C., … Eickhoff,
S. B. (2017). On the integrity of functional brain networks in
schizophrenia, Parkinson’s disease, and advanced age: Evidence
from connectivity-based single-subject classification: Schizo-
phrenia, Parkinson’s disease and aging classification. Human
Brain Mapping, 38(12), 5845–5858. https://doi.org/10.1002
/hbm.23763, PubMed: 28876500
Pläschke, R. N., Patil, K. R., Cieslik, E. C., Nostro, A. D., Varikuti,
D. P., Plachti, A., Lösche, P., Hoffstaedter, F., Kalenscher, T.,
Langner, R., & Eickhoff, S. B. (2020). Age differences in predict-
ing working memory performance from network-based func-
tional connectivity. Cortex, 132, 441–459. https://doi.org/10
.1016/j.cortex.2020.08.012, PubMed: 33065515
Pruim, R. H. R., Mennes, M., van Rooij, D., Llera, A., Buitelaar, J. K.,
& Beckmann, C. F. (2015). ICA-AROMA: A robust ICA-based
strategy for removing motion artifacts from fMRI data. Neuro-
Image, 112, 267–277. https://doi.org/10.1016/j.neuroimage
.2015.02.064, PubMed: 25770991
Pudil, P., Novovičová, J., & Kittler, J. (1994). Floating search
methods in feature selection. Pattern Recognition Letters, 15(11),
1119–1125. https://doi.org/10.1016/0167-8655(94)90127-9
Randolph, J. J., Falbe, K., Manuel, A. K., & Balloun, J. L. (2014). A
step-by-step guide to propensity score matching in R. Practical
Assessment, Research & Evaluation, 19(18), 1–6.
Raschka, S. (2018). MLxtend: Providing machine learning and data
science utilities and extensions to Python’s scientific computing
stack. Journal of Open Source Software, 3(24), 638. https://doi
.org/10.21105/joss.00638
Rasero, J., Sentis, A. I., Yeh, F.-C., & Verstynen, T. (2021). Integrating
across neuroimaging modalities boosts prediction accuracy of
cognitive ability. PLOS Computational Biology, 17(3), e1008347.
https://doi.org/10.1371/journal.pcbi.1008347, PubMed: 33667224
Raz, N. (2000). Aging of the brain and its impact on cognitive per-
formance: Integration of structural and functional findings. In The
handbook of aging and cognition (2nd ed., pp. 1–90). Mahwah,
NJ: Lawrence Erlbaum Associates Publishers.
Raz, N., & Rodrigue, K. M. (2006). Differential aging of the brain:
Patterns, cognitive correlates and modifiers. Neuroscience & Bio-
behavioral Reviews, 30(6), 730–748. https://doi.org/10.1016/j
.neubiorev.2006.07.001, PubMed: 16919333
Rosenberg, M. D., Finn, E. S., Scheinost, D., Papademetris, X.,
Shen, X., Constable, R. T., & Chun, M. M. (2016). A neuromarker
of sustained attention from whole-brain functional connectivity.
Nature Neuroscience, 19(1), 165–171. https://doi.org/10.1038/nn
.4179, PubMed: 26595653
Rubinov, M., & Sporns, O. (2010). Complex network measures of brain
connectivity: Uses and interpretations. NeuroImage, 52(3), 1059–1069.
https://doi.org/10.1016/j.neuroimage.2009.10.003, PubMed: 19819337
Saad, Z. S., Gotts, S. J., Murphy, K., Chen, G., Jo, H. J., Martin, A., &
Cox, R. W. (2012). Trouble at rest: How correlation patterns and
group differences become distorted after global signal regression.
Brain Connectivity, 2(1), 25–32. https://doi.org/10.1089/ brain
.2012.0080, PubMed: 22432927
Sanz-Arigita, E. J., Schoonheim, M. M., Damoiseaux, J. S., Rombouts,
S. A. R. B., Maris, E., Barkhof, F., Scheltens, P., & Stam, C. J. (2010).
Loss of ‘small-world’ networks in Alzheimer’s disease: Graph
analysis of fMRI resting-state functional connectivity. PLoS One,
5(11), e13788. https://doi.org/10.1371/journal.pone.0013788,
PubMed: 21072180
Scarpazza, C., Ha, M., Baecker, L., Garcia-Dias, R., Pinaya,
W. H. L., Vieira, S., & Mechelli, A. (2020). Translating research
findings into clinical practice: A systematic and critical review of
neuroimaging-based clinical tools for brain disorders. Transla-
tional Psychiatry, 10(1), 107. https://doi.org/10.1038/s41398
-020-0798-6, PubMed: 32313006
Schaefer, A., Kong, R., Gordon, E. M., Laumann, T. O., Zuo, X.-N.,
Holmes, A. J., Eickhoff, S. B., & Yeo, B. T. T. (2018). Local-global
parcellation of the human cerebral cortex from intrinsic func-
tional connectivity MRI. Cerebral Cortex, 28(9), 3095–3114.
https://doi.org/10.1093/cercor/bhx179, PubMed: 28981612
Schmermund, A., Möhlenkamp, S., Stang, A., Grönemeyer, D.,
Seibel, R., Hirche, H., Mann, K., Siffert, W., Lauterbach, K., Siegrist,
J., Jöckel, K.-H., & Erbel, R. (2002). Assessment of clinically silent
atherosclerotic disease and established and novel risk factors for
predicting myocardial infarction and cardiac death in healthy
middle-aged subjects: Rationale and design of the Heinz Nixdorf
RECALL Study. American Heart Journal, 144(2), 212–218. https://
doi.org/10.1067/mhj.2002.123579, PubMed: 12177636
Network Neuroscience
146
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
1
2
2
2
0
7
2
1
6
8
n
e
n
_
a
_
0
0
2
7
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cognitive performance differences in older age
Smith, S. M., Jenkinson, M., Woolrich, M. W., Beckmann, C. F.,
Behrens, T. E. J., Johansen-Berg, H., Bannister, P. R., De Luca,
M., Drobnjak, I., Flitney, D. E., Niazy, R. K., Saunders, J., Vickers,
J., Zhang, Y., De Stefano, N., Brady, J. M., & Matthews, P. M.
(2004). Advances in functional and structural MR image analysis
and implementation as FSL. NeuroImage, 23, S208–S219. https://
doi.org/10.1016/j.neuroimage.2004.07.051, PubMed: 15501092
Sripada, C., Angstadt, M., Rutherford, S., Taxali, A., & Shedden, K.
(2020a). Toward a “treadmill test” for cognition: Improved pre-
diction of general cognitive ability from the task activated brain.
Human Brain Mapping, 41(12), 3186–3197. https://doi.org/10
.1002/hbm.25007, PubMed: 32364670
Sripada, C., Rutherford, S., Angstadt, M., Thompson, W. K.,
Luciana, M., Weigard, A., Hyde, L. H., & Heitzeg, M. (2020b).
Prediction of neurocognition in youth from resting state fMRI.
Molecular Psychiatry, 25(12), 3413–3421. https://doi.org/10
.1038/s41380-019-0481-6, PubMed: 31427753
Stern, Y., Gurland, B., Tatemichi, T. K., Tang, M. X., Wilder, D., &
Mayeux, R. (1994). Influence of education and occupation on
the incidence of Alzheimer’s disease. JAMA: The Journal of the
American Medical Association, 271(13), 1004–1010. https://doi
.org/10.1001/jama.1994.03510370056032, PubMed: 8139057
Stumme, J., Jockwitz, C., Hoffstaedter, F., Amunts, K., & Caspers, S.
(2020). Functional network reorganization in older adults:
Graph-theoretical analyses of age, cognition and sex. Neuro-
Image, 214, 116756. https://doi.org/10.1016/j.neuroimage.2020
.116756, PubMed: 32201326
Supekar, K., Menon, V., Rubin, D., Musen, M., & Greicius, M. D.
(2008). Network analysis of intrinsic functional brain connectiv-
ity in Alzheimer’s disease. PLoS Computational Biology, 4(6),
e1000100. https://doi.org/10.1371/journal.pcbi.1000100,
PubMed: 18584043
Teipel, S. J., Grothe, M. J., Metzger, C. D., Grimmer, T., Sorg, C., Ewers,
M., Franzmeier, N., Meisenzahl, E., Klöppel, S., Borchardt, V., Walter,
M., & Dyrba, M. (2017). Robust detection of impaired resting state
functional connectivity networks in Alzheimer’s disease using elastic
net regularized regression. Frontiers in Aging Neuroscience, 8, 318.
https://doi.org/10.3389/fnagi.2016.00318, PubMed: 28101051
Thompson, W. K., Barch, D. M., Bjork, J. M., Gonzalez, R., Nagel,
B. J., Nixon, S. J., & Luciana, M. (2019). The structure of cognition
in 9 and 10 year-old children and associations with problem behav-
iors: Findings from the ABCD study’s baseline neurocognitive
battery. Developmental Cognitive Neuroscience, 36, 100606.
https://doi.org/10.1016/j.dcn.2018.12.004, PubMed: 30595399
Tucker-Drob, E. M. (2011). Global and domain-specific changes in
cognition throughout adulthood. Developmental Psychology, 47(2),
331–343. https://doi.org/10.1037/a0021361, PubMed: 21244145
van den Heuvel, M. P., de Lange, S. C., Zalesky, A., Seguin, C., Yeo,
B. T. T., & Schmidt, R. (2017). Proportional thresholding in
resting-state fMRI functional connectivity networks and conse-
quences for patient-control connectome studies: Issues and rec-
ommendations. NeuroImage, 152, 437–449. https://doi.org/10
.1016/j.neuroimage.2017.02.005, PubMed: 28167349
van Wijk, B. C. M., Stam, C. J., & Daffertshofer, A. (2010). Compar-
ing brain networks of different size and connectivity density
using graph theory. PLoS One, 5(10), e13701. https://doi.org/10
.1371/journal.pone.0013701, PubMed: 21060892
Vemuri, P., Lesnick, T. G., Przybelski, S. A., Machulda, M., Knopman,
D. S., Mielke, M. M., Roberts, R. O., Geda, Y. E., Rocca, W. A.,
Petersen, R. C., & Jack, C. R. (2014). Association of lifetime intel-
lectual enrichment with cognitive decline in the older popula-
tion. JAMA Neurology, 71(8), 1017–1024. https://doi.org/10
.1001/jamaneurol.2014.963, PubMed: 25054282
Vergun, S., Deshpande, A. S., Meier, T. B., Song, J., Tudorascu, D. L.,
Nair, V. A., Singh, V., Biswal, B. B., Meyerand, M. E., Birn, R. M.,
& Prabhakaran, V. (2013). Characterizing functional connectivity
differences in aging adults using machine learning on resting state
fMRI data. Frontiers in Computational Neuroscience, 7, 38.
https://doi.org/10.3389/fncom.2013.00038, PubMed: 23630491
Vieira, B. H., Liem, F., Dadi, K., Engemann, D. A., Gramfort, A.,
Bellec, P., Craddock, R. C., Damoiseaux, J. S., Steele, C. J.,
Yarkoni, T., Langer, N., Margulies, D. S., & Varoquaux, G.
(2022). Predicting future cognitive decline from non-brain and
multimodal brain imaging data in healthy and pathological
aging. Neurobiology of Aging, 118, 55–65. https://doi.org/10
.1016/j.neurobiolaging.2022.06.008, PubMed: 35878565
Wang, J., Zuo, X., Dai, Z., Xia, M., Zhao, Z., Zhao, X., Jia, J., Han,
Y., & He, Y. (2013). Disrupted functional brain connectome in
individuals at risk for Alzheimer’s disease. Biological Psychiatry,
73(5), 472–481. https://doi.org/10.1016/j.biopsych.2012.03.026,
PubMed: 22537793
Weis, S., Hodgetts, S., & Hausmann, M. (2019). Sex differences and
menstrual cycle effects in cognitive and sensory resting state net-
works. Brain and Cognition, 131, 66–73. https://doi.org/10.1016
/j.bandc.2017.09.003, PubMed: 29030069
Woo, C.-W., Chang, L. J., Lindquist, M. A., & Wager, T. D. (2017).
Building better biomarkers: Brain models in translational neuro-
imaging. Nature Neuroscience, 20(3), 365–377. https://doi.org
/10.1038/nn.4478, PubMed: 28230847
Yeo, B. T., Krienen, F. M., Sepulcre, J., Sabuncu, M. R., Lashkari, D.,
Hollinshead, M., Roffman, J. L., Smoller, J. W., Zöllei, L., Polimeni,
J. R., Fischl, B., Liu, H., & Buckner, R. L. (2011). The organization
of the human cerebral cortex estimated by intrinsic functional con-
nectivity. Journal of Neurophysiology, 106(3), 1125–1165. https://
doi.org/10.1152/jn.00338.2011, PubMed: 21653723
Yoo, K., Rosenberg, M. D., Hsu, W.-T., Zhang, S., Li, C.-S. R., Scheinost,
D., Constable, R. T., & Chun, M. M. (2018). Connectome-based
predictive modeling of attention: Comparing different functional
connectivity features and prediction methods across datasets.
NeuroImage, 167, 11–22. https://doi.org/10.1016/j.neuroimage
.2017.11.010, PubMed: 29122720
Zalesky, A., Fornito, A., & Bullmore, E. (2012). On the use of cor-
relation as a measure of network connectivity. NeuroImage,
60(4), 2096–2106. https://doi.org/10.1016/j.neuroimage.2012
.02.001, PubMed: 22343126
Zarogianni, E., Moorhead, T. W. J., & Lawrie, S. M. (2013). Towards
the identification of imaging biomarkers in schizophrenia, using
multivariate pattern classification at a single-subject level. Neu-
roImage: Clinical, 3, 279–289. https://doi.org/10.1016/j.nicl
.2013.09.003, PubMed: 24273713
Zou, H., & Hastie, T. (2005). Regularization and variable selection
via the elastic net. Journal of the Royal Statistical Society: Series B
(Statistical Methodology), 67(2), 301–320. https://doi.org/10
.1111/j.1467-9868.2005.00503.x
Network Neuroscience
147
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
1
2
2
2
0
7
2
1
6
8
n
e
n
_
a
_
0
0
2
7
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3