RESEARCH
Probing the association between resting-state
brain network dynamics and
psychological resilience
Dominik Kraft1
and Christian J. Fiebach1,2
1Department of Psychology, Goethe University Frankfurt, Frankfurt, Germany
2Brain Imaging Center, Goethe University Frankfurt, Frankfurt am Main, Germany
Keywords: Resting-state, Time-varying connectivity, Multilayer modularity, Psychological
resilience, Network reconfigurations, Node flexibility, Node promiscuity, Node degree
a n o p e n a c c e s s
j o u r n a l
ABSTRACT
This study aimed at replicating a previously reported negative correlation between node
flexibility and psychological resilience, that is, the ability to retain mental health in the face of
stress and adversity. To this end, we used multiband resting-state BOLD fMRI (TR = .675 sec)
from 52 participants who had filled out three psychological questionnaires assessing
resilience. Time-resolved functional connectivity was calculated by performing a sliding
window approach on averaged time series parcellated according to different established
atlases. Multilayer modularity detection was performed to track network reconfigurations over
time, and node flexibility was calculated as the number of times a node changes community
assignment. In addition, node promiscuity (the fraction of communities a node participates in)
and node degree (as proxy for time-varying connectivity) were calculated to extend previous
work. We found no substantial correlations between resilience and node flexibility. We
observed a small number of correlations between the two other brain measures and resilience
scores that were, however, very inconsistently distributed across brain measures, differences
in temporal sampling, and parcellation schemes. This heterogeneity calls into question the
existence of previously postulated associations between resilience and brain network
flexibility and highlights how results may be influenced by specific analysis choices.
AUTHOR SUMMARY
We tested the replicability and generalizability of a previously proposed negative association
between dynamic brain network reconfigurations derived from multilayer modularity
detection (node flexibility) and psychological resilience. Using multiband resting-state BOLD
fMRI data and exploring several parcellation schemes, sliding window approaches, and
temporal resolutions of the data, we could not replicate previously reported findings regarding
the association between node flexibility and resilience. By extending this work to other
measures of brain dynamics (node promiscuity, degree) we observe a rather inconsistent
pattern of correlations with resilience that strongly varies across analysis choices. We conclude
that further research is needed to understand the network neuroscience basis of mental health
and discuss several reasons that may account for the variability in results.
Citation: Kraft, D., & Fiebach, C. J.
(2022). Probing the association
between resting-state brain network
dynamics and psychological resilience.
Network Neuroscience, 6(1), 175–195.
https://doi.org/10.1162/netn_a_00216
DOI:
https://doi.org/10.1162/netn_a_00216
Supporting Information:
https://doi.org/10.1162/netn_a_00216
Received: 20 July 2021
Accepted: 8 November 2021
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Author:
Dominik Kraft
dominik.kraft@gmail.com
Handling Editor:
Alex Fornito
Copyright: © 2021
Massachusetts Institute of Technology
Published under a Creative Commons
Attribution 4.0 International
(CC BY 4.0) license
The MIT Press
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
6
1
1
7
5
2
0
0
2
4
4
6
n
e
n
_
a
_
0
0
2
1
6
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Resting-state brain network dynamics and psychological resilience
Functional connectivity:
Statistical dependencies among
(fMRI) time series from different
regions, nodes, or networks.
Graph theoretical framework:
Mathematical tool to model and
analyze pairwise relations between
different objects (here: time series
from brain regions and their
functional connectivity).
INTRODUCTION
Abundant literature in human clinical neuroscience has established a link between changes in
intrinsic functional connectivity of large-scale brain networks and psychological disorders such
as depression (Mulders et al., 2015) or schizophrenia (Dong et al., 2018; Li et al., 2019; see also
Menon, 2011, for a general overview). As a consequence, brain bases of preserving mental
health in the face of stress and adversity (resilience) have also become a focus of interest
(e.g., Southwick & Charney, 2012). Two recent studies (Long et al., 2019; Paban et al., 2019)
have reported associations between psychological resilience and brain network dynamics by
using multilayer modularity, a relatively new tool from the evolving field of network neurosci-
ence that integrates spatial and temporal information in a graph theoretical framework
(Muldoon & Bassett, 2016). In both studies, resilience was assessed with a frequently used ques-
tionnaire, the Connor–Davidson Resilience Scale (CD-RISC; Connor & Davidson, 2003), and
network dynamics under task-free (resting-state) conditions were examined by detecting time-
evolving patterns of nonoverlapping and coherent modules and by quantifying the frequency
with which brain nodes switched between modules (node flexibility; Bassett et al., 2011).
The link between resilience and brain network dynamics is motivated by (a) an assumed
relationship between neuronal and cognitive flexibility (e.g., Braun et al., 2015; Chen et al.,
2016) and by (b) associations of resilience with higher cognitive flexibility, both in theoret-
ical models (Southwick & Charney, 2012) and in empirical work (e.g., Genet & Siemer,
2011). While this would predict greater resilience in more (cognitively or neuronally) flexible
persons, it has also been argued that cognitive flexibility might not be universally adaptive
and that resilience may depend on an interplay between flexibility and stability (Parsons
et al., 2016). This proposal receives support
from observations of changes along
flexibility-stability dimensions in psychiatric conditions, both behaviorally (e.g., rigid behav-
iors like rumination in depression; Nolen-Hoeksema et al., 2008) and neuronally (e.g.,
increased network flexibility, variability in patients with autism or schizophrenia; Gifford
et al., 2020; Harlalka et al., 2019). These and similar results suggest that the extreme tails
of the ‘flexibility-stability’ distribution (Kashdan & Rottenberg, 2010) may indeed be related
to psychopathology and that, accordingly, adaptive behavior in the ‘normal’ (i.e., unaffected)
range depends on a balance between flexibility and stability. Whereas the above-cited work
by Long et al. (2019) and Paban et al. (2019) took a primarily neuroscientific perspective as
a starting point, the psychological perspective offered here is not fully consistent with their
empirical results. However, it may offer a valuable conceptual framework for a neurocogni-
tive model of resilience and therefore calls for further empirical research to clarify the current
inconsistencies.
Paban et al. (2019) measured task-free EEG and conducted network analyses in source
space. Negative correlations between node flexibility and resilience were observed in the
alpha, beta, and delta frequency bands, including superior parietal cortex, medial orbitofrontal
cortex, and cuneus. A subsequent resting-state functional MRI (rs-fMRI) study (Long et al.,
2019) also reports lower node flexibility in more resilient persons, primarily in visual cortices
and the left medial-orbital superior frontal gyrus. Except for a partial overlap in visual regions
(lingual gyrus), localization results differed between studies and Long et al. (2019) did not rep-
licate correlations in ‘higher order’ (superior parietal, inferior frontal) areas reported by Paban
et al. (2019). Whereas some of these inconsistencies may result from inherent differences
between methods (e.g., differences in temporal resolution between fMRI and EEG), others
may reflect specific methodological choices by the authors. For example, Long et al. (2019)
studied BOLD ‘dynamics’ using only 12 nonoverlapping time windows (each of 20 sample
points length, derived from 250 measurements of TR = 2 sec).
Network Neuroscience
176
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
6
1
1
7
5
2
0
0
2
4
4
6
n
e
n
_
a
_
0
0
2
1
6
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Resting-state brain network dynamics and psychological resilience
Even though gold standards for analyses of dynamic multilayer modularity are yet to be
established, recent studies suggest that more data are required for sensitive and reliable estima-
tion of network dynamics from BOLD fMRI (Hindriks et al., 2016; Yang et al., 2021). To ame-
liorate such methodological shortcomings and to further our understanding of the relationship
between resilience and brain network dynamics, we here replicate and extend these results by
using temporally highly resolved fMRI (multiband/MB sequence; MB factor 4; TR = .675 sec)
from 52 healthy young adults who completed three resilience questionnaires. We first repli-
cated Long et al.’s (2019) analysis pipeline as closely as possible, by downsampling data to
a TR of 2.025 sec and using the same analysis parameters. Following this, we explore effects
of specific analysis choices (like different windowing schemes) on network flexibility. Lastly,
we repeat correlation analyses with optimized denoising and the full temporal resolution of
the MB data, resulting in a time series of 753 overlapping windows (Figure 1). Node flexibility
was calculated as the number of times a node changes its community assignment between
windows, divided by the total possible number of changes (Bassett et al., 2011). Given the
results of Long et al. (2019), we expected that resilience and node flexibility should be neg-
atively correlated.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
Downsampling:
Creating a ‘slower’, i.e., less densely
sampled time series by artificially
selecting every xth time point.
Multilayer modularity detection:
Detection of highly connected nodes
(i.e., modules) in temporal, ordinal
networks.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
6
1
1
7
5
2
0
0
2
4
4
6
n
e
n
_
a
_
0
0
2
1
6
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
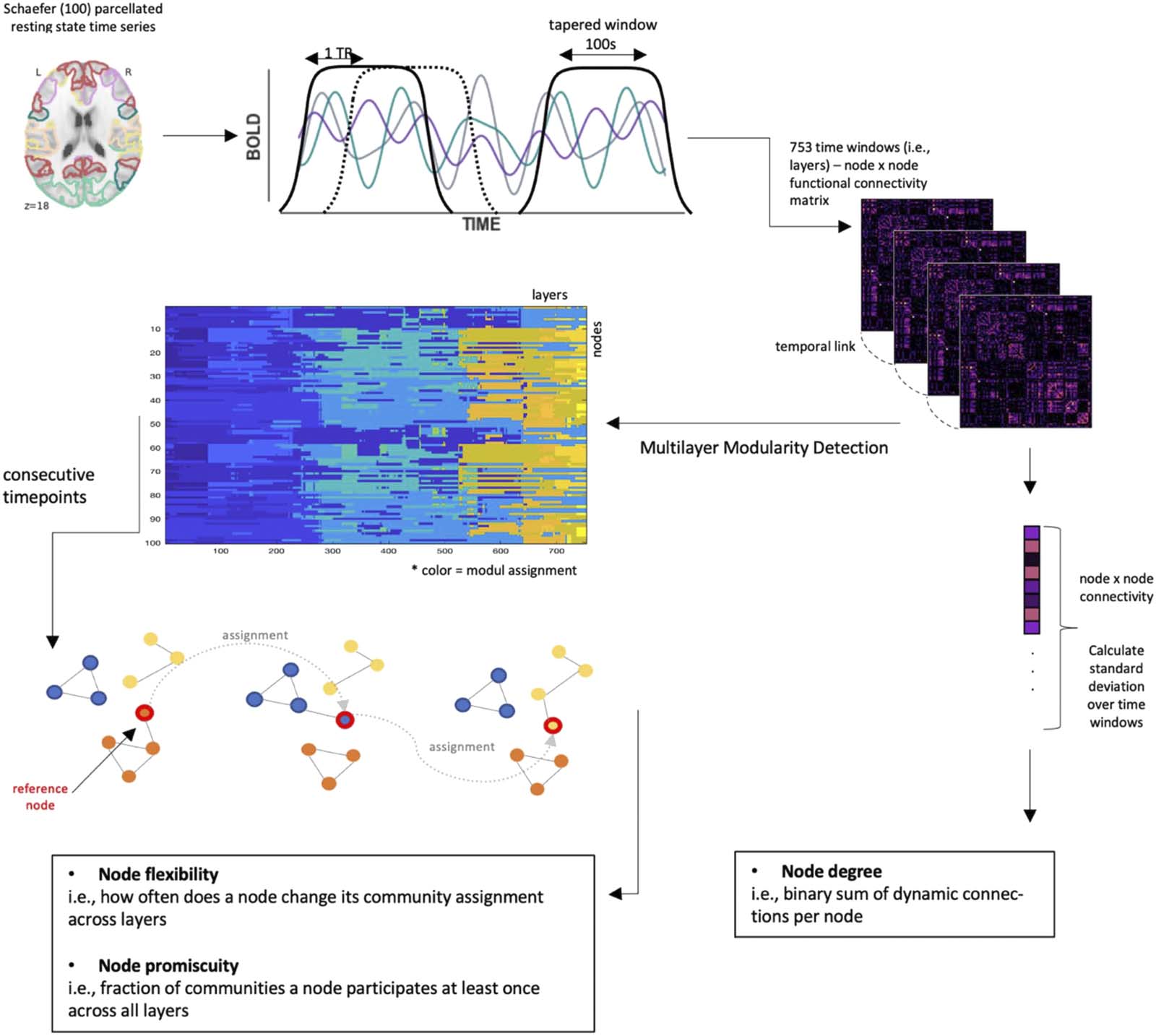
Figure 1. Schematic illustration of the workflow, here visualized for analyses with original multiband data (TR = .675 sec), the Schaefer100
atlas, and overlapping sliding windows of size 100 sec and offset 1 TR. Procedures for other parameter choices are analogous. Starting with the
mean resting-state time series, 753 functional connectivity matrices (layers) representing intercorrelations between the 100 different nodes
were calculated via a sliding window approach; multilayer modularity detection was performed on ordinal layers. Network measures were
calculated as described in the figure; see Methods section for further details.
Network Neuroscience
177
Resting-state brain network dynamics and psychological resilience
To extend previous work, we also assessed node promiscuity and node degree, and their
relation to resilience. Node promiscuity, the fraction of communities a node participates in at
least once, and node flexibility are complementary measures that inform us whether brain
dynamics per se (i.e., assessed via node flexibility) or the diversity of brain systems with which
a node interacts (assessed via its promiscuity) may be associated with resilience. Node degree,
in turn, is a proxy for dynamic connectivity that does not rely on modularity detection algo-
rithms (and is thus invariant to potential algorithmic idiosyncrasy or parameter choices). How-
ever, previous work demonstrates that dynamic connectivity is related to various cognitive and
behavioral traits, as well as clinical conditions like posttraumatic stress disorder (PTSD, often
used as proxy for studies of resilience; Bolsinger et al., 2018; Jin et al., 2017; Lurie et al., 2020),
which makes it a candidate marker for analyzing the neurobiology of resilience. To the best of
our knowledge, relationships between resilience and promiscuity as well as degree have so far
not been studied. Accordingly, we here aim at a more complete characterization of putative
relationships between brain network dynamics and resilience.
RESULTS
Behavioral Results: Subjective Resilience Ratings
Descriptive statistics for resilience scales are listed in Table 1. Psychometric properties were
satisfactory, with internal consistencies (Cronbach’s alpha) between .7 and .85 (Table 1). Resil-
ience scores in our sample were comparable to the respective original publications for the
German versions (CD-RISC: 30.6, Sarubin et al., 2015; RS-13: 70, Leppert et al., 2008; Brief
Resilience Scale (BRS): 3.58 and 3.37, Chmitorz et al., 2018). Resilience ratings were signif-
icantly intercorrelated: CD-RISC − RS, r = .60; CD-RISC − BRS, r = .61; BRS − RS, r = .45,
all p < .001 (FDR-corrected for multiple comparisons).
Replication of Long et al. (2019)
We replicated the analysis pipeline of Long et al. (2019) as closely as possible, involving
downsampling of multiband data by a factor of 3 to a virtual TR of 2.025 sec, use of the
two parcellation schemes used by Long et al. (2019), that is, AAL90 and Power264 atlases,
and the same denoising strategy (with 26 parameters) as used in that study. Age, gender,
and framewise displacement were included as covariates of no interest (see Methods). Given
that recent evidence implies that functionally derived parcellations outperform anatomical
atlases like the automated anatomical atlas (AAL) (Dadi et al., 2019), we also conducted all
analyses with a further atlas, the functionally defined parcellation scheme of Schaefer (100
nodes), which approximately matches the number of nodes in the AAL. Following Long
et al. (2019), the downsampled time series was segmented into nonoverlapping windows of
20 TRs length, resulting in 15 time windows (as opposed to 12 in the original study). We did
Table 1. Descriptive statistics of resilience questionnaires (N = 52)
M (SD)
min–max
α (95% CI)
CD-RISC
28.88 (4.73)
18–38
RS
71.37 (10.06)
21–87
BRS
3.72 (.73)
1.83–5.00
.70 (.56–.81)
.85 (.79–.91)
.82 (.73–.89)
Note: CD-RISC = Connor–Davidson Resilience Scale; RS = Resilience Scale; BRS = Brief Resilience Scale; M =
mean; SD = standard deviation; min = minimum; max = maximum; α = Cronbach’s alpha; CI = confidence
interval.
Network Neuroscience
178
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
1
1
7
5
2
0
0
2
4
4
6
n
e
n
_
a
_
0
0
2
1
6
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Resting-state brain network dynamics and psychological resilience
Table 2. Correlations between global flexibility and resilience scales according to the pipeline
used in Long et al. (2019)
Atlas
AAL 90
CD-RISC
r = −.14, p = .33
Resilience scales
BRS
r = −.33, p = .05
RS
r = −.20, p = .23
Power 264
r = .06, p = .87
r = −.02, p = .87
r = −.03, p = .87
Schaefer 100
r = −.01, p = .94
r = −.12, p = .58
r = −.16, p = .58
not replicate the correlation between global flexibility and CD-RISC reported by Long et al.
(2019; all p > = .33), but observe a borderline significant correlation between global flexibility
(AAL atlas) and the BRS resilience score (r = −.33, p = .05). Across the three parcellation
schemes and resilience questionnaires, we did not observe any further significant effects, nei-
ther at global, subnetwork, or node level (Table 2).
Effects of Overlapping Versus Nonoverlapping Time Windows
Segmenting the time series into nonoverlapping windows results in a low number of sample
points, generally not considered sufficient for analyzing network dynamics. Many studies
today segment BOLD time series into overlapping windows, as this allows for an estimation
of network dynamics with greater sensitivity and reliability (Hindriks et al., 2016; Lurie et al.,
2020; Yang et al., 2021). When analyzing our downsampled data with a high number of over-
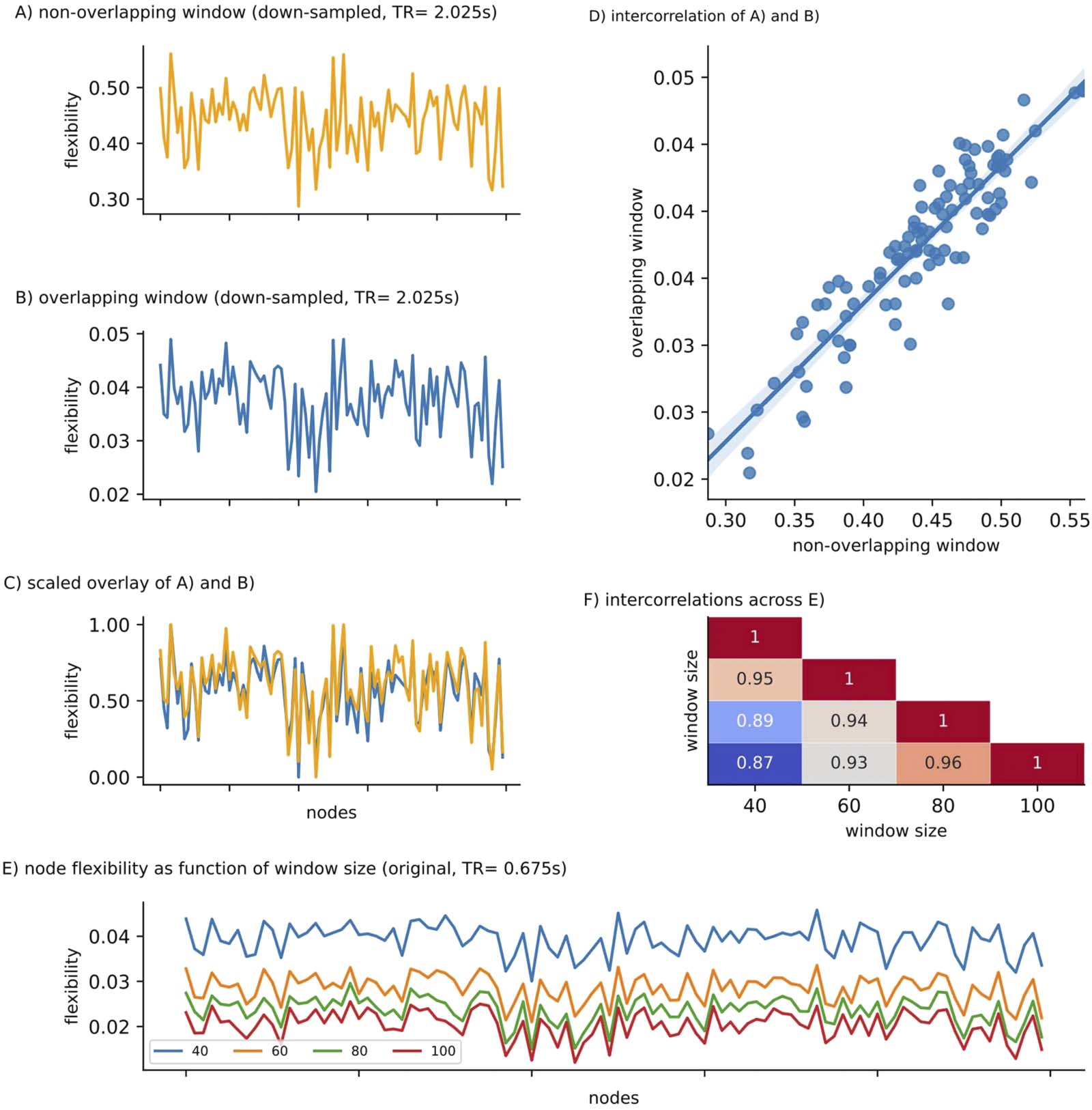
lapping windows (i.e., 251; see Methods), nodal flexibility was scaled by a factor of ∼1/10; cp.
y-axis scaling of Figures 2A and 2B), as expected given the higher overlap between consecu-
tive windows. However, high spatial similarity was preserved between node flexibility derived
from downsampled data with overlapping versus nonoverlapping windows (r = .90, p < .0001;
Figure 2C and 2D), indicating that there are no qualitative differences between windowing
schemes. We thus use overlapping windows for all further analyses.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
1
1
7
5
2
0
0
2
4
4
6
n
e
n
_
a
_
0
0
2
1
6
p
d
t
.
Dynamic Network Measures in Original Versus Downsampled Functional Connectivity Data
Analyses of network dynamics may be limited by lower temporal resolutions (which was here
simulated by downsampling to allow for a direct replication of the results by Long et al., 2019).
As compared to the original data (TR = .675 sec), downsampling shifted node flexibility (calcu-
lated using overlapping windows; see section Replication of Long et al. (2019)) toward higher
values. However, high spatial similarity (i.e., correlation) exists between the distribution of nodal
flexibility in original versus downsampled data (r = .95, p < .0001). As outlined above, we will in
the following also investigate node promiscuity and degree as further measures of brain network
flexibility. We thus also explore how these measures are affected by downsampling of the BOLD
data: as for flexibility, downsampling led to higher promiscuity values, but a high spatial simi-
larity was preserved (r = .93, p < .0001). We observed a significantly lower nodal degree in the
downsampled data (0.06–0.61% significant connections; M = 0.20) compared to the original
data (0.12–0.75%; M = 0.33), t(99) = 13.93, p < .0001, which is consistent with a recent report
by Pedersen et al. (2018) that fewer data result in lower nodal degree values.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Correlation Between Dynamic Functional Connectivity Measures and Resilience
Given that no previous data exist to derive specific hypotheses concerning the relationship
between resilience and node promiscuity as well as degree, all subsequently reported associ-
ations were tested two-sided using Spearman correlations. Correlations were calculated for all
Network Neuroscience
179
Resting-state brain network dynamics and psychological resilience
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
1
1
7
5
2
0
0
2
4
4
6
n
e
n
_
a
_
0
0
2
1
6
p
d
.
t
Influence of methodological differences in dynamic network analysis on node flexibility. Node flexibility for the downsampled data
Figure 2.
(TR = 2.025 sec) with (A) nonoverlapping (window size = 20 TR) versus (B) overlapping windows (window size = 50 TR). Overlaying rescaled
variants of both arrays indicates (C) only minor differences between node flexibility values derived from different window schemes and
(D) high spatial similarity, that is, a high correlation coefficient, between (A) and (B). (E) Node flexibility for the original data (multiband,
TR = .675 sec), across nodes (x-axis) as a function of window size (40- to 100-sec length) using overlapping windows (F) The distribution
of flexibility values across nodes shows high spatial similarity (all correlations p < .0001). For the analyses shown in (E) and (F), flexibility
values were derived after repeating the modularity detection algorithm analyses 10 times and choosing the run yielding the highest modularity
value Q (see Methods for details).
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
three parcellations, for original (TR = .675 sec) and downsampled (TR = 2.025) data, using an
optimized denoising strategy (36 parameters; see Methods), overlapping time windows, and
the same covariates as during the replication attempt reported above.
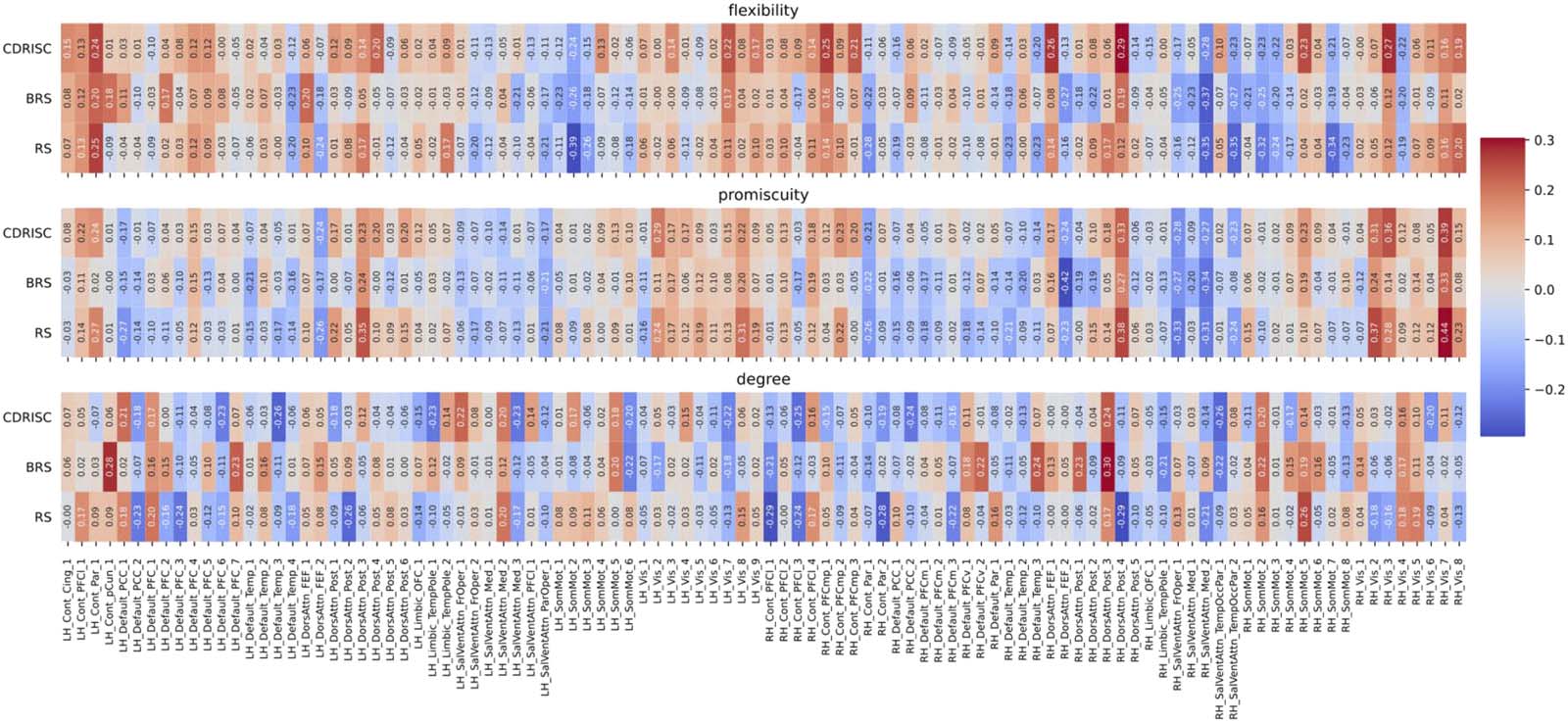
In the original data, we observed two significant positive correlations at the nodal level of
the Power264 atlas between flexibility of a cingulo-opercular node (#54) and the CD-RISC (r =
.52, p = .02), and between promiscuity of a visual node (#165) and the BRS resilience score
(r = .57, p = .004). After downsampling (but now exploiting higher temporal resolution due to
the overlapping windowing scheme), the AAL-based analysis yielded a significant association
between the promiscuity of the left pallidum (node level, AAL) and BRS resilience (r = −.49,
p = .02). In the Schaefer100 analysis, significant negative correlations emerged between the
Network Neuroscience
180
Resting-state brain network dynamics and psychological resilience
Table 3. Significant correlations between measures of brain dynamics (node flexibility, node promiscuity, and node degree) and psychological
resilience as measured with three different scales
Atlas
Original data
AAL90
Power264
Schaefer100
Downsampled data
AAL90
Power264
Node
Level of analysis
RSN
Global
flexibility node #54:
CD-RISC (r = .52, p = .02)
promiscuity node #165:
BRS (r = .57, p = .004)
promiscuity pallidum (L):
BRS (r = −.49, p = .02)
degree subcortical:
BRS (r = −.37, p = .05)
degree cerebellar:
BRS (r = −.37, p = .05)
Schaefer100
degree Vis_7 (L):
RS (r = −.48, p = .03)
global degree:
BRS (r = −.35, p = .03)
Note: Scales include CD-RISC, BRS, and RS (see Methods for details). All combinations of atlases and levels of analyses were computed, but only significant
results are described in the table. RSN, resting-state network: analyses with node-level measures averaged within (functionally) defined subnetworks. Global:
analyses based on average across values of all nodes. Original Data, multiband BOLD echo planar imaging (TR = .675 sec). Downsample Data, virtual down-
sampling of multiband data to a TR of 2.025 sec. AAL90, Power264, SCHAEFER100, different parcellation schemes as described in the Methods section. Note
that eight nodes were excluded from analyses with the Power264 atlas due to BOLD signal dropout. L = left hemisphere.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
1
1
7
5
2
0
0
2
4
4
6
n
e
n
_
a
_
0
0
2
1
6
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
degree of a visual node (left Vis_7) and the RS score (r = −.48, p = .03), as well as between
global degree and BRS resilience (r = −.35, p = .03). In the Power264 atlas we observed two
borderline significant results, between the BRS resilience score and the degree of the subcor-
tical resting-state network (RSN) (r = −.37, p = .05) and the cerebellar RSN (r = −.37, p = .05).
No correlation was observed for any additional tested combination (see Table 3). Figure 3,
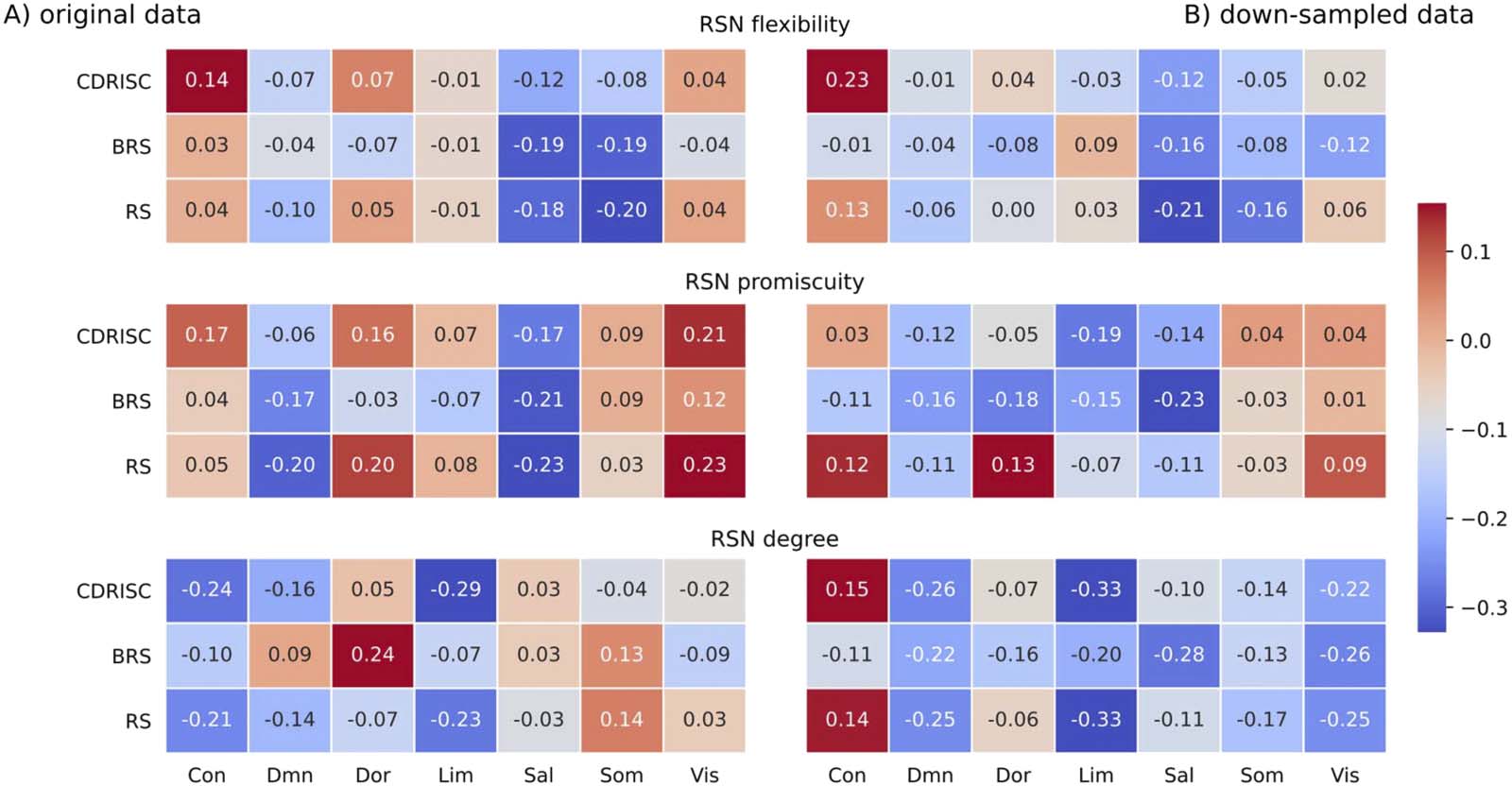
Figure 4, and Figure 5 visualize nodal and RSN results in an exemplary manner for the
Schaefer100 parcellation.
Follow-Up Analyses
To further investigate putative factors that may have influenced reported results, we conducted
several post hoc analyses on data derived from the Schaefer100 parcellation.
Effects of covariates. Recalculation of all node-level correlations without covariates yielded
results comparable to the above-reported findings (cf. Supporting Information File 1, Figures S1
and S2). Correlations for original (MB) resolution were −.41 < r < .44, all p > .11. For
downsampled data, the above-reported significant correlation for one visual node disappeared
(−.43 < r < .30, all p > .14).
Network Neuroscience
181
Resting-state brain network dynamics and psychological resilience
Figure 3. Correlations between network measures and resilience at the level of resting-state networks (RSNs) in the Schaefer100 atlas for the
(A) original data (multiband; TR = .675 sec) and (B) downsampled data (TR = 2.025 sec; see Methods for details). The number in each cell
represents the respective correlation coefficient; all p > .12. Network labels: Con = fronto-parietal-control; Sal = salience/ventral attention;
Lim = limbic; Dor = dorsal attention; Som = somatomotor; DMN = default mode; Vis = visual. Resilience questionnaires: CD-RISC = Connor–
Davidson Resilience Scale; BRS = Brief Resilience Scale, RS = Resilience Scale.
Effects of window length. To explore how node flexibility changes as a function of window
length, node flexibility was calculated for window lengths between 40 and 100 sec in steps
of 20 (original/MB data). Similar to the effect of downsampling (see above), we observed an
increase in node flexibility with decreasing window size (Figure 2E), however, with high spa-
tial similarity of flexibility values across window sizes, all r > .87, p < .0001 (Figure 2F). We did
not observe any significant correlations between node flexibility and resilience when varying
the window size between 40 and 80 sec (all p > .13).
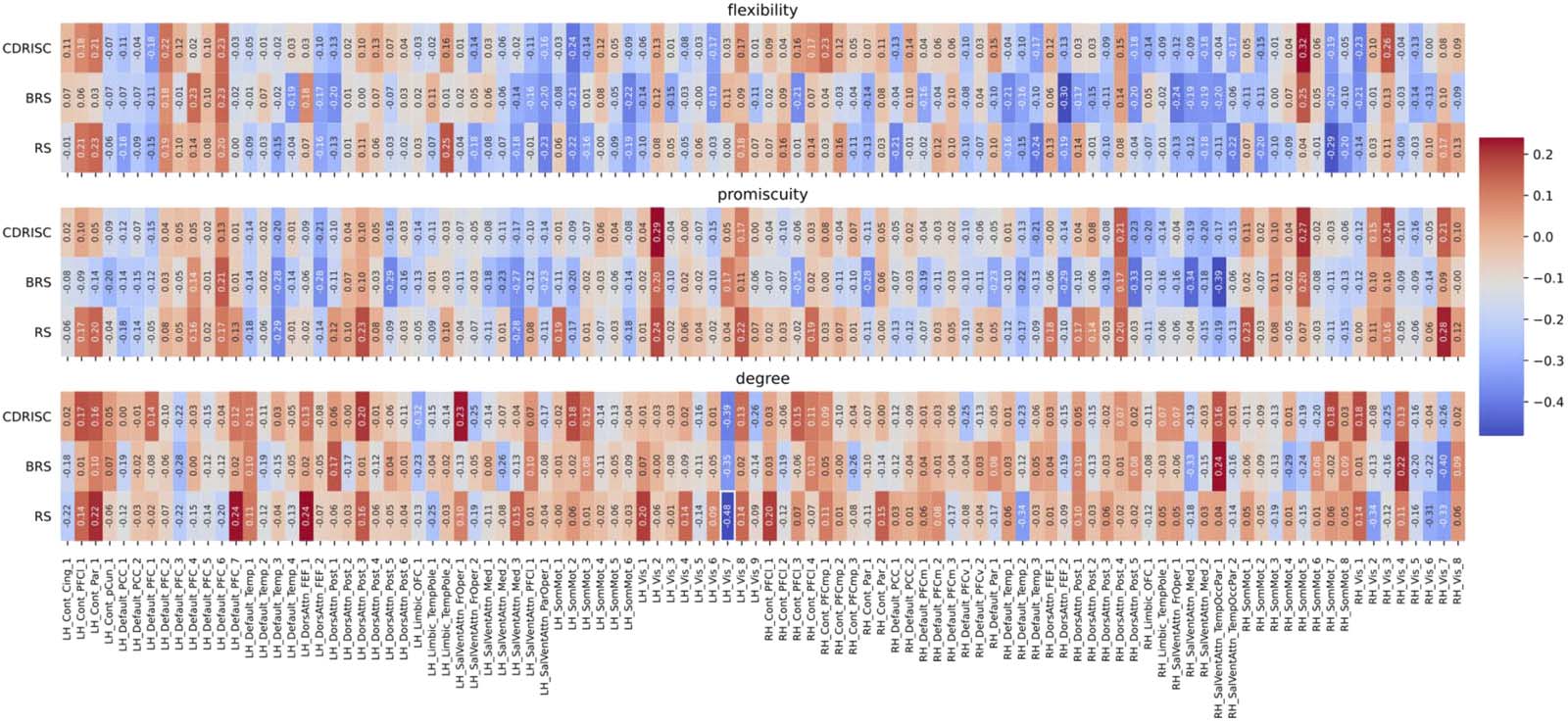
Figure 4. Correlations between network measures and resilience at the nodal level for the original (multiband) data (TR = .675 sec) in the
Schaefer100 atlas. Node names are depicted on the x-axis ticks; for a detailed list see https://bit.ly/3yvOBwz. LH = left hemisphere; RH = right
hemisphere. Figure S1 in the Supporting Information depicts the same correlations, but without including covariates (see Follow-Up Analyses
in the Results section).
Network Neuroscience
182
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
6
1
1
7
5
2
0
0
2
4
4
6
n
e
n
_
a
_
0
0
2
1
6
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Resting-state brain network dynamics and psychological resilience
Figure 5. Correlations between network measures and resilience at the nodal level for the downsampled data (TR = 2.025 sec) in the
Schaefer100 atlas (all p > .10). Node names are depicted on the x-axis ticks; for a detailed list see https://bit.ly/3yvOBwz. LH = left hemisphere;
RH = right hemisphere. The significant correlation between the degree of node Vis_7 and the RS scale is highlighted by white rectangle.
Figure S2 in the Supporting Information depicts the same correlations, but without including covariates (see Follow-Up Analyses in the Results
section).
Effects of denoising. While repeating the correlation analyses (Schaefer100 parcellation, over-
lapping windows) after denoising with a 26-parameter regression model (following Long et al.,
2019) as opposed to the 36-parameter model used above, no correlations with resilience were
found at global (multiband: all p > .54; downsampled: all p > .74) or nodal level (multiband:
all p > .15; downsampled: all p > .53). In the RSN analysis, significant negative correlations
were found between degree of the limbic RSN and all resilience scales (multiband; CD-RISC:
r = −.39, p = .03; BRS: r = −.44, p = .008; RS: r = −.40, p = .03; downsampled: all p > .16).
Effects of spatial resolution. As the Schaefer atlas is available in multiple parcellations, it is best
suited to investigate the influence of spatial resolution. Using 200 (vs. 100) nodes, we did not
observe any correlation on the global level, neither for original (all p > .50), nor downsampled
(all p > .13) data. While there were also no significant correlations on the RSN level for original
data (all p > .29), degree of the limbic RSN was negatively correlated to both CD-RISC (r =
−.57, p < .001) and RS (r = −.38, p = .04) in downsampled data. No significant effects emerged
at nodal level (original: all p > .36; downsampled: all p > .10).
Effects of motion. For the 26-parameter denoised data, we observed a significant correlation
between global flexibility and mean framewise displacement (FD) (r = .38, p = .02, FDR-
corrected) derived from the original (multiband) data, whereas no other brain measure was
correlated with FD for original or downsampled data (all p > .11) and no correlations emerged
after optimized (36-parameter plus despiking) denoising (original: all p > .63; downsampled:
all p > .05).
DISCUSSION
The present study investigated associations between intrinsic functional brain connectivity
dynamics and psychological resilience. More specifically, we explored whether a previously
reported negative correlation between resilience and node flexibility as determined using
resting-state BOLD fMRI (Long et al., 2019) can be replicated in multiband BOLD data with
higher temporal resolution (TR = .675 sec). Additionally, we aimed at extending previous
Network Neuroscience
183
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
6
1
1
7
5
2
0
0
2
4
4
6
n
e
n
_
a
_
0
0
2
1
6
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Resting-state brain network dynamics and psychological resilience
findings by including two further metrics of network dynamics, as well as two further question-
naires measuring slightly different theoretical conceptions of resilience. Correlation analyses
were performed at different topological scales, that is, for whole brain, (functional) subnet-
works, and nodes. Results did not support the previously reported negative association
between node flexibility and resilience, neither in a direct replication matching as closely
as possible methodological parameters of the original study, nor when extending the method-
ology in terms of higher temporal resolution, finer grained sliding window approach, or further
network measures. In these analyses, we found a small number of significant correlations that
were distributed rather inconsistently across different measures of brain dynamics and resil-
ience, across atlases, and across sampling schemes: for multiband data (original time resolution;
Power264 parcellation), flexibility of a cingulo-opercular node correlated with resilience measured
using the CD-RISC questionnaire and promiscuity of a visual node correlated with BRS resilience,
however, both in the opposite direction relative to Long et al. (2019). For downsampled data, we
observed negative correlations between global degree and BRS and between the degree of a visual
node and the RS score (Schaefer100 parcellation). After increasing the spatial resolution to 200
nodes (follow-up analysis), these effects disappeared, and negative resilience correlations emerged
for the limbic network (CD-RSIC, RS). When using less stringent denoising (as in the original study),
we also observed negative correlations with limbic network degree (all three resilience scales), for
multiband but not downsampled data. When applying the AAL atlas, a negative correlation emerged
with promiscuity for a subcortical node (pallidum; BRS). Of note, we also observed a borderline sig-
nificant (p = .05) negative correlation between global flexibility and resilience (BRS) in the direct rep-
lication. We do not consider this strong support in favor of a replication since we found no similar
effects using the Power264 or Schaefer100 atlases or with the other two questionnaires. In sum,
these heterogeneous results cannot be considered a successful replication. The fact that the lim-
bic network was involved in these correlations multiple times suggests that its role for resilience
should be investigated further. On the other hand, these effects were not consistent, so that they
should be treated with caution. Lastly, post hoc analyses conducted with multiband data and the
Schaefer100 parcellation suggest invariance of results against changes in windowing schemes
(size, overlap of windows). In the following, we will discuss potential limitations of the present
study, which may be important when considering reasons for differences between present and
previous results. We then discuss in more depth factors that may account for the failed
replication of the results of Long et al. (2019).
Potential Limitations
Potential limitations of investigating network dynamics in BOLD data with low temporal res-
olution, as in the original study, have already been discussed in the Introduction. When ana-
lyzing temporally highly resolved multiband data, we chose the window size to be equal to
1/fmin (Leonardi & Van De Ville, 2015). We cannot rule out that such long window sizes might
hamper the sensitivity to small changes, as calculating connectivity matrices over longer
periods serves as a smoothing kernel minimizing the ability to resolve time-varying functional
connectivity (TVFC) (Vergara et al., 2019). However, this disadvantage of MB-echo planar
imaging (EPI) is more than compensated by the ability to acquire more data in the same time,
the possibility to investigate faster dynamics, and to thus better disentangle neural and phys-
iological signals (Yang & Lewis, 2021). Furthermore, we showed empirically that node flexi-
bility was largely robust against changes in window size.
Potential limitations may also stem from the questionnaires used: short versions of two
scales (CD-RISC: 10 items; RS: 13 items) were used due to timing constraints, whereas the long
(25-items) CD-RISC was used by Long et al. (2019). However, we deem this unproblematic as
Time-varying functional
connectivity:
Functional connectivity that varies
over time, often labeled as ‘dynamic
connectivity’.
Network Neuroscience
184
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
6
1
1
7
5
2
0
0
2
4
4
6
n
e
n
_
a
_
0
0
2
1
6
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Resting-state brain network dynamics and psychological resilience
validity studies show that the short versions can be considered valid and reliable instruments
for measuring trait resilience (Chmitorz et al., 2018; Leppert et al., 2008; Sarubin et al., 2015).
Our data confirm this, as all questionnaires showed satisfying levels of internal consistency.
Methodological Considerations
Even though the overall direction of correlations between node flexibility and resilience was
the same in two previous publications using fMRI (Long et al., 2019) and EEG (Paban et al.,
2019), neural localizations partly differed. Adding to this heterogeneity, correlation results in
the present study were highly inconsistent (see above) and results thus provide no strong evi-
dence for an association between network dynamics and resilience. This demonstrates the
importance of replication attempts also in network neuroscience studies of brain-behavior cor-
relations. To support such work in the future, we have compiled a list of methodological fea-
tures that we have found important in the present replication study (Table 4).
Physiological noise and preprocessing. When applying less stringent denoising, a positive corre-
lation between global flexibility and mean FD emerged (in multiband time resolution, over-
lapping windows), suggesting that increased motion inside the scanner may artificially alter
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
6
1
1
7
5
2
0
0
2
4
4
6
n
e
n
_
a
_
0
0
2
1
6
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Table 4. Recommendations for individual difference research using dynamic functional connectivity
Area
Methodological
Keyword
Recommendation
Parcellations
Test different parcellation schemes and atlases
Test different node resolutions to explore stability of effects
(currently only for Schaefer atlas possible)
Preprocessing, denoising
Optimize preprocessing and denoising strategies to different types
of functional connectivity indices
Test influence of different denoising pipelines to identify possible
relationship between motion and measure of interest
Exclude high motion subjects (rather strict than lenient if amount
of data allows for)
Sliding window technique,
Test different windowing schemes (e.g., size of windows,
dynamic functional connectivity
amount of overlap)
Use sufficient amount of data, if possible, e.g., via multiband
fMRI (but take into account that acceleration decreases
signal-to-noise ratio) or longer measurements
Multilayer modularity
Test different parameter settings
Psychological
Construct of interest
Incorporate different measures for (e.g., two resilience scales) or
aim for a complete characterization (i.e., all possible metrics)
for the construct of interest, if possible
Brain: construct of interest
Motivate in/exclusion of covariates (test both if applicable)
relationship/confounding variables
Provide descriptive statistics for measures of interest
Provide reliability measures (if applicable)
Replications
Method section
Provide enough detail to allow for replication attempts
Note: This brief list of recommendations does not claim to be complete, but rather advocates to always aim at incorporating the most recent findings and
empirical evidence from methodological studies.
Network Neuroscience
185
Resting-state brain network dynamics and psychological resilience
functional connectivity and network flexibility. We did not observe any other correlation
between dynamic brain measures and motion, suggesting that denoising approaches and their
effectiveness may interact with temporal sampling schemes (i.e., MB vs. conventional EPI).
This underscores the importance of incorporating recent empirical insights regarding denois-
ing strategies for module detection and analyses of network reconfigurations (Lydon-Staley
et al., 2019).
Sliding window technique. Both windowing schemes used in our study (overlapping vs. non-
overlapping) are among the most used approaches within the sliding window framework (Iraji
et al., 2020). For window-based TVFC analyses, window length is a critical aspect, as it has to
balance the ability of robustly estimating TVFC (which benefits from higher numbers of
timepoints) against susceptibility to noise (Damaraju et al., 2020; Iraji et al., 2020). Extensive
studies have evaluated the impact of different window types, lengths, and overlap on TVFC,
however, without yet identifying a gold standard (e.g., Abrol et al., 2017; Shakil et al., 2016).
At present, lengths between 30 sec and 100 sec are commonly used (see Figure S1 in Preti
et al., 2017). Within this range, one would, however, assume that strong effects should be
robust against specific analysis choices (compare, e.g., Damaraju et al., 2020, vs. Haimovici
et al., 2017). This is supported by our follow-up analyses showing that results did not vary
between different window schemes or sizes.
Reliability. Recent research suggests that multilayer modularity in particular (Yang et al., 2021)
and TVFC in general (Hindriks et al., 2016) are a function of scanning duration and that
rs-fMRI sequences around 10 min with standard TRs (as used by Long et al., 2019) are not
sufficient for reliable parameter estimation. As reliability strongly influences which correlations
are detectable (Hedge et al., 2018), short measurements may be prone to false positive corre-
lations. Yang et al. (2021) propose that at least 20 min of rs-fMRI is needed for reliable
multilayer modularity analysis—in that study based on a TR of 1.45 sec and nonoverlapping
windows of size 100 sec. Here, we measured 10 minutes of rs-fMRI with a TR of about half
that length and used a substantially larger number of windows, intentionally selected to allow
for reliable multilayer analyses. Yang et al. (2021) also suggest that the standard settings for
intra- and interlayer parameters (γ = ω = 1) in dynamic network measures may not necessarily
turn out to be optimal across datasets and across different network measures in terms of
reliability. However, when varying the intra- and interlayer parameters beyond the standard
setting (in the original/MB data, Schaefer100 atlas; see Supporting Information), we did not
observe any significant correlations between node flexibility or promiscuity and resilience
scores, except for a single correlation between the flexibility of a visual node and BRS when
setting γ = 1.4 (r = .50, p = .02). This correlation, however, points in the opposite direction of
what had been reported before (Long et al., 2019). We can thus conclude that the specific
choice of intra- and interlayer parameters has no strong effect on detecting resilience-flexibility
correlations.
It is also important that the behavioral variables of interest show
Resilience questionnaires.
satisfying psychometric properties. Internal consistencies for the resilience questionnaires were
satisfactory, ranging from .70 to .85. When investigating individual differences, samples
should not be too homogeneous on the target measures, as interperson variability is a precon-
dition for detecting correlations. Even though CD-RISC data appear to be in a somewhat
narrower range and more left-skewed than in Long et al. (2019), both the RS-13 and the
BRS show sufficient variability (Table 1), which increases confidence that our results are not
driven by a lack of heterogeneity. Moreover, resilience scores in our sample were distributed
similar to the respective original publications of these measures. It is, however, difficult to
Network Neuroscience
186
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
6
1
1
7
5
2
0
0
2
4
4
6
n
e
n
_
a
_
0
0
2
1
6
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Resting-state brain network dynamics and psychological resilience
compare our behavioral results directly to the studies by Paban et al. (2019) and Long et al.
(2019), as the former did not report descriptive values (but high internal consistency:
Cronbachs α = .90), whereas the latter did report descriptive statistics but no psychometrics.
Conclusion
To summarize, our results do not provide support for the previously reported negative associ-
ation between node flexibility and psychological resilience. We extended previous studies by
including additional measures of functional connectivity dynamics and two further resilience
questionnaires and found only weak and inconsistent evidence of associations between net-
work dynamics and resilience, mostly for node degree, a proxy for TVFC, but not for node
flexibility or promiscuity. Our study highlights how specific degrees of freedom in the analysis
of functional connectivity may influence the presence or absence of effects of interest. This
underscores the need for testing the robustness and generalizability of proposed effects via
replication.
METHODS
Code and data have been deposited at https://doi.org/10.5281/zenodo.5761102 and will be
made publicly available after an embargo period that ends December 31, 2024.
Participants
In total, N = 69 right-handed university students were enrolled in the study, of whom N = 60
completed the study protocol and were included for further analyses. All participants were
native speakers of German, right-handed, and between 18 and 35 years old; absence of
current psychiatric episode was assured with a structured interview (MINI; Sheehan et al.,
1998). This sample size was based on considerations of statistical power for investigating
across-participant relationships between BOLD activation and cognitive flexibility (see prereg-
istration: https://osf.io/a64jn); the task-free (resting state) fMRI data reported here were
acquired together with the preregistered task-based fMRI experiments. During image quality
checks and preprocessing, eight datasets were excluded from further analyses (four due to low
quality of questionnaire data and four due to motion artifact; see below for details of exclusion
criteria), so that the reported analyses are based on data from 52 participants (26 females,
25 males, 1 diverse; 18–34 years; mean age 24.0 ± 3.7). Using the pwr package in
R (Champely et al., 2017), we estimated that our final sample size of N = 52 has a power
of > .99 to detect correlational effects of the size reported by Long et al. (2019), that is, of
around r = .55. All participants provided written informed consent, and all procedures were
approved by the Ethics Committee of the Department of Psychology of Goethe University
Frankfurt, Germany.
Functional MR Image Acquisition
Resting-state functional MRI (rs-fMRI) data were collected prior to task-based fMRI (which is
not part of the present report; see previous paragraph) on a 3-T Siemens Prisma MR-Scanner
equipped with a 32-channel head coil, using a multiband (MB factor = 4) EPI sequence with
the following parameters: 900 volumes (10:14 min), TR = 675 ms, TE = 30 ms, voxel size =
3 mm3, flip angle = 60°, FoV = 222 mm, acquisition matrix = 74 × 74, 40 slices. During the
rs-fMRI measurement, participants were asked to keep their eyes open and gaze at a white
fixation cross, located at the center of a screen (Nordic Neuro Lab, 40″, 1,920 × 1,080,
60 Hz), to stay relaxed and not to think about anything specific. In a separate session, a T1
Network Neuroscience
187
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
6
1
1
7
5
2
0
0
2
4
4
6
n
e
n
_
a
_
0
0
2
1
6
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Resting-state brain network dynamics and psychological resilience
weighted (T1w) 3D structural MR scan was acquired with a MPRAGE sequence (4:26 min,
voxel size = 1 mm3, TR = 1,900 ms, TE = 2.52 ms, acquisition matrix = 256 × 256, 192 slices)
for purposes of coregistration between functional and structural data.
Resilience Questionnaires
The psychological construct ‘resilience’ was quantified using German versions of three self-
report questionnaires, the 10-item version of Connor–Davidson Resilience Scale (CD-RISC –
10 items; Connor & Davidson, 2003; German version by Sarubin et al., 2015), the Brief
Resilience Scale BRS – 6 items (Smith et al., 2008; German version by Chmitorz et al.,
2018), and the Resilience Scale (RS – 13 items; Wagnild & Young, 1993; German version by
Leppert et al., 2008). Note that short versions of the questionnaires were selected, as the present
study was part of a more extensive study protocol and the short versions have been shown to
allow for a time-efficient data collection with comparable validity and reliability (Chmitorz
et al., 2018; Leppert et al., 2008; Sarubin et al., 2015). We included the CD-RISC and BRS
because of their good ratings in an evaluation study by Windle et al. (2011). The RS was addi-
tionally chosen to increase comparability with other research, as this questionnaire is frequently
used in resilience research. In addition, the selected questionnaires also differ in that CD-RISC
and RS define resilience as a personality trait, whereas resilience is understood as an outcome
in the BRS (see original publications for more details). Resilience questionnaires were filled out
during an informational preparation session for the fMRI measurements and administered
online using Unipark software (EFS Survey, Questback GmbH). As completion time of online
questionnaires has been identified as the most reliable indicator of data being meaningful or
meaningless (Leiner, 2013), we evaluated the quality of questionnaire data by using a quality
index provided by the Unipark system that compares the completion time of each participant
with the average completion time of our sample. As preregistered (https://osf.io/c94y8) partic-
ipants with an Unipark quality index of .20 or lower were excluded, resulting in exclusion of
five participants (see also Leiner, 2013, for a similar criterion). Resilience scores were calcu-
lated according to the respective manuals. For each resilience scale, we also assessed internal
consistency by calculating Cronbach’s alpha and its 95% confidence interval.
MR Data Quality Control
Quality of imaging data was assessed using both fMRIPREP’s visual reports as well as MRIQC
0.15.2rc1 (Esteban et al., 2017, 2019). T1w and functional images for each participant were
visually checked for signal artifacts, whole-brain coverage, and correct alignment between
structural and functional data. Following a procedure proposed by Faskowitz et al. (2019),
functional data were excluded if marked as an outlier (i.e., exceeding ×1.5 the interquartile-
range either from Q1 or Q3) in more than 50% of the MRIQC quality metrics: dvars, tsnr, snr,
efc, aor, aqi (see the MRIQC documentation for more information about these metrics; Esteban
et al., 2017). Given the sensitivity of resting-state analyses to movements and given that some
of the aforementioned metrics are influenced by motion, we additionally included FD as a
metric for quantifying motion artifacts (Maknojia et al., 2019). Due to the higher sampling rate
of multiband EPI, motion parameters exhibit a high-frequency (HF) component resulting from
head motion due to respiration (Power et al., 2019) that is usually not observable with standard
single-band fMRI (Williams & Van Snellenberg, 2019). With this ‘spurious’ HF motion compo-
nent, the head appears to be in constant motion and summary measures (such as FD) might be
contaminated, leading to exaggerated flagging of ‘bad’ volumes. Note that this applies less to
subsequent functional connectivity measures, as those are routinely band-pass filtered,
whereas summary measures (e.g., FD) are calculated on raw motion parameters (Gratton
188
Multiband EPI:
Fast echo planar imaging (EPI)
sequence which is characterized by
the simultaneous acquisition of
multiple slices. Multiband EPI
shortens the acquisition time of a
single brain volume and thus allows
to acquire more data at a given time.
Network Neuroscience
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
6
1
1
7
5
2
0
0
2
4
4
6
n
e
n
_
a
_
0
0
2
1
6
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Resting-state brain network dynamics and psychological resilience
et al., 2020; Power et al., 2019; Williams & Van Snellenberg, 2019). We therefore calculated
filtered FD (FDfilt) from Butterworth-filtered raw head movement traces to better isolate ‘true’
head movements (Power et al., 2019). Following Lydon-Staley et al. (2019), we excluded sub-
jects with mean FDfilt >0.2 mm or >20 volumes with FDfilt >0.25 mm to ensure high data
quality. We excluded four subjects based on their FDfilt, while no subject was excluded based
on MRIQC’s metrics.
Image Preprocessing
T1w and rfMRI images were preprocessed using fMRIPREP 20.1.1 (Esteban et al., 2019), which
is based on Nipype 1.5.0 (Gorgolewski et al., 2011). A boilerplate text released under a CC0
license describing preprocessing details can be found in the Supporting Information. For fur-
ther pipeline information, see fMRIPREP’s documentation (Esteban et al., 2019). Due to the use
of MB data and the high sampling rate, no slice time correction was applied (see, e.g., Glasser
et al., 2013). Distortion corrected functional images in T1w space were further denoised using
the XPC Engine 1.2.1 (Ciric et al., 2017). We implemented a denoising strategy that has been
shown to be relatively effective in mitigating motion artifacts in the study of dynamic func-
tional connectivity, (multilayer) subnetwork detection, and measures of module reconfigura-
tion (Lydon-Staley et al., 2019). BOLD data was first demeaned, detrended, and despiked on a
voxelwise basis, instead of using more aggressive censoring methods that may result in varying
window lengths across participants (Hutchison et al., 2013), and then temporally filtered with
a first-order Butterworth filter by using a passband of 0.01–0.08 Hz. These operations were
followed by a confound regression, that included (a) six motion estimates derived from fMRI-
PREPs realignment, (b) mean signals from white matter ( WM) and cerebrospinal fluid (CSF),
(c) mean global signal, (d) temporal derivatives of these nine regressors, as well as (e) the qua-
dratic terms of all 18 parameters, resulting in a 36-parameter model to obtain residual BOLD
time series. All regressors were also band-pass filtered to avoid reintroducing noise caused by a
frequency-dependent mismatch (Hallquist et al., 2013). For the direct replication of Long et al.
(2019) and to directly test the influence of different denoising schemes, we also implemented a
more lenient 26-paramter denoising approach that included 24 motion parameters following
Friston et al. (1996) together with mean signals from WM and CSF. Within the XCP engine, the
Schaefer (resolution: 100, 200; Schaefer et al., 2018), AAL90 (Tzourio-Mazoyer et al., 2002),
and Power264 (Power et al., 2011) atlases were transformed to native T1 space and resampled
to match the BOLD images (see Supporting Information for details). For each subject the whole
brain was parcellated into distinct regions and a functional time series was extracted for each
region, corresponding to the average across all voxels within that region.
Time-Varying Functional Connectivity
All subsequent TVFC analyses were performed with the numpy (1.18.5; Harris et al., 2020)
and scipy (1.5.0; SciPy 1.0 Contributors et al., 2020) packages in python 3.8.3.
Down-Sampling and Windowing Scheme
To directly replicate findings described in Long et al. (2019) and to investigate if different
sampling rates (i.e., MB-EPI vs. conventional EPI) yield different results, we downsampled
our original resting-state BOLD time series by a factor of three to resemble a TR of 2 as closely
as possible (3 × .675 sec = 2.025 sec). Raw motion traces were also downsampled accordingly,
and FD was calculated without filtering. For the direct replication, we segmented the time series
into 15 nonoverlapping windows with length of 20 TR (40.5 sec), analogous to the window-
ing scheme used in Long et al. (2019). As this low number of windows may hamper the ability
Network Neuroscience
189
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
6
1
1
7
5
2
0
0
2
4
4
6
n
e
n
_
a
_
0
0
2
1
6
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Resting-state brain network dynamics and psychological resilience
Sliding window correlation:
FC calculation over a defined
window, which is shorter than the
whole time series. Window type may
vary in its appearance. Sliding the
window over the time series can
produce time resolved functional
connectivity (one matrix per
window).
to investigate brain network dynamics (Hindriks et al., 2016; Lurie et al., 2020; Yang et al.,
2021), we additionally used overlapping windows that were shifted by a single time point
(TR). All windows were tapered with a Hamming filter to reduce potential edge artifacts and
to suppress spurious correlations (Shakil et al., 2016; Zalesky et al., 2014), and pairwise Pearson
correlation coefficients between all nodes were calculated within each window.
For analyses of the ‘original’ MB data with high temporal resolution, we used a fixed win-
dow length of 148 time points (100 sec), which satisfies the frequency criterion that the length
should be at least be equal to 1/fmin (Leonardi & Van De Ville, 2015) and allows for a full
oscillation of the slowest frequency in the range of 0.01–0.08Hz. After downsampling, sliding
window correlations with an adjusted window length of 50 time points (101.25 sec) and sub-
sequent TVFC and modularity analyses were calculated as described above. To investigate
whether the calculated network measures differ between original and downsampled data
and to rule out any idiosyncratic algorithmic behavior (i.e., from multilayer modularity), we
compared all three metrics at the nodal level, as these were the starting points for all subse-
quent measures. For node flexibility and node promiscuity we calculated the spatial similarity
(i.e., Spearman correlation coefficient) between mean values per node (average over partici-
pants) in the original and downsampled data to explore whether nodes behave in comparable
manner in both time series. As the downsampling creates ‘sparser’ data and disrupts the
smoothness of the original data (which might result in more abrupt changes in community
assignments), we anticipated a tendency toward higher values for both metrics in the down-
sampled data. For node degree, we expected lower values in the downsampled data, as a
recent study by Pedersen et al. (2018) showed that nodal degree decreased with less available
data and thus tested this hypothesis using a paired t test.
We calculated the standard deviation (SD) of each node × node correlation (i.e., node-node
connection or edge) over time as a proxy for TVFC (often also referred to as ‘dynamic’ connectivity;
Lurie et al., 2020). To test whether these SDs likely reflect ‘true’ dynamics, we benchmarked these
estimates against phase randomized surrogate (null) data that preserved autocorrelation, power
spectral density, and stationary cross correlation of the observed data (Lurie et al., 2020; Prichard
& Theiler, 1994; Savva et al., 2019). More precisely, we created 500 surrogates for each subject by
phase randomizing the empirical time series obtained from the 100 nodes. To preserve the correl-
ative structure between node time series, all signals were multiplied by the same uniformly random
phase and the SD for each edge was calculated, respectively (see Savva et al., 2019, for a detailed
description of phase randomization). The empirically observed SDs were ranked against this
null distribution and p values were obtained by dividing the number of times SDsurr >= SDreal
by the number of surrogates. Given that each subject has 4,950 unique connections, FDR
correction ( p < 0.05) was applied to reduce type I errors.
To describe ‘dynamics’ in nodal space, we calculated the nodal degree metric as the binary
sum of significant (i.e., ‘dynamic’) connections for each node (Pedersen et al., 2018). Of note,
when testing TVFC with surrogate null data, one needs to be cautious as the absence of sig-
nificance (relative to the null model) does not necessarily indicate the absence of dynamic
connectivity. This interpretation depends heavily on the null model applied, and the data
may contain meaningful fluctuations relative to other definitions of the null model (see Lurie
et al., 2020, for a comprehensive overview).
Multilayer Modularity
For multilayer modularity, negative correlations in each node × node matrix (i.e., per subject
and time window) were set to zero and correlations were z-transformed as done in previous
Network Neuroscience
190
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
1
1
7
5
2
0
0
2
4
4
6
n
e
n
_
a
_
0
0
2
1
6
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Resting-state brain network dynamics and psychological resilience
studies (e.g., Finc et al., 2020; Pedersen et al., 2018). We constructed an ordinal multilayer
network in which each layer (i.e., time window) represents a weighted adjacency matrix. To
assess the spatiotemporal community structure and to track network reconfigurations, each
node was linked to itself across layers. To detect communities (i.e., groups of nodes that are
more densely connected to one another than to the rest of the network), we used the multilayer
counterpart (Mucha et al., 2010) of the modularity function proposed by Newman and Girvan
(2004). To optimize the multilayer modularity function, we used an iterative and generative
Louvain like algorithm (implemented with code from Jeub et al., 2011–2019; see Supporting
Information for details). The tunable parameters γ and ω were held constant across layers and
set to unity, as had been done in previous studies (e.g., Braun et al., 2015; Yin et al., 2020). As
the modularity approach is not deterministic, we repeated it 100 times for each participant and
chose the run yielding the highest modularity value as, for example, done in Finc et al. (2020).
We also tested the influence of varying the intra- and interlayer parameters beyond the stan-
dard setting in the original data (see Supporting Information) but found no substantial influence
on the reported results.
Time-Resolved Analyses of Brain Network Reconfiguration
Within the time-varying community framework, we assessed node flexibility as the number of
times a node changes its community assignment between adjacent layers, normalized by the
total number of possible changes. Node flexibility can be interpreted as a metric that allows
one to quantify reconfigurations of functional connectivity patterns that a brain region
undergoes over time (Braun et al., 2015). To enrich the spatiotemporal description of a node,
we further calculated node promiscuity, defined as the fraction of communities a node partic-
ipates in at least once, across all layers. This metric allows one to quantify the distribution of a
node’s connections over time, for example, whether high flexibility stems from a switching
between two communities or an evenly distributed allegiance to a larger number of different
modules (Garcia et al., 2018; Papadopoulos et al., 2016). The higher the nodal promiscuity,
the more modules a node participates in at least once across time. For each of the three node-
specific measures of dynamic network reconfiguration, that is, flexibility, promiscuity, and
degree, we additionally calculated the respective global measure as the average across all
nodes, as well as network-specific measures by averaging across all nodes belonging to the
respective RSNs, which varied across atlases.
Correlation Analyses
All correlational analyses were performed with the pingouin statistics package (0.3.11; Vallat,
2018) in python 3.8.3. To test for associations between resilience and time-varying brain net-
work measures, we calculated partial (Spearman) correlations with age, gender, and FD
included as covariates of no interest. Note that unlike Long et al. (2019), years of education
was not included as covariate, as all participants were students and years of education were
thus not acquired in our study. Correlations were performed at the global, nodal, and network-
specific levels for all pairwise combinations between resilience questionnaires (CD-RISC, BRS,
RS) and network measures (flexibility, promiscuity, degree). Results were corrected for multiple
statistical comparisons by using the false discovery rate ( p < .05; Benjamini & Hochberg,
1995).
Follow-Up Analyses
Lastly, to more directly assess the effect of specific analysis choices on observed correlations
between network measures and resilience, we varied a number of such factors systematically,
Network Neuroscience
191
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
1
1
7
5
2
0
0
2
4
4
6
n
e
n
_
a
_
0
0
2
1
6
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Resting-state brain network dynamics and psychological resilience
using as ‘standard’ for comparison the analysis pipeline based on MB data parcellated using
the Schaefer100 atlas, 36-parameter denoising, and overlapping sliding window scheme. We
investigated the effects of including versus excluding covariates, varying sliding window size
(in a frequently used range between 40 and 100 sec in steps of 20 sec; Preti et al., 2017), and
how the two different denoising pipelines affect resilience correlations. We also explored
within the same parcellation scheme (the Schaefer atlas) the effects of increasing the spatial
resolution (i.e., from 100 to 200 nodes).
ACKNOWLEDGMENTS
We thank Dr. Cindy Eckart for providing valuable comments on an earlier version of this
manuscript and our research assistants for their help in data collection.
SUPPORTING INFORMATION
Supporting information for this article is available at https://doi.org/10.1162/netn_a_00216.
AUTHOR CONTRIBUTIONS
Dominik Kraft: Conceptualization; Data curation; Formal analysis; Investigation; Methodology;
Project administration; Software; Visualization; Writing – original draft; Writing – review &
editing. Christian J. Fiebach: Conceptualization; Project administration; Writing – original draft;
Writing – review & editing.
FUNDING INFORMATION
This research was funded by the German Research Foundation (CRC 1193 grant no. INST
247/859-1 awarded to C.J.F.). D.K. is supported by the German Academic Scholarship
Foundation.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
1
1
7
5
2
0
0
2
4
4
6
n
e
n
_
a
_
0
0
2
1
6
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
REFERENCES
Abrol, A., Damaraju, E., Miller, R. L., Stephen, J. M., Claus, E. D.,
Mayer, A. R., & Calhoun, V. D. (2017). Replicability of
time-varying connectivity patterns in large resting state fMRI
samples. NeuroImage, 163, 160–176. https://doi.org/10.1016/j
.neuroimage.2017.09.020, PubMed: 28916181
Bassett, D. S., Wymbs, N. F., Porter, M. A., Mucha, P. J., Carlson,
J. M., & Grafton, S. T. (2011). Dynamic reconfiguration of human
brain networks during learning. Proceedings of the National
Academy of Sciences, 108(18), 7641–7646. https://doi.org/10
.1073/pnas.1018985108, PubMed: 21502525
Benjamini, Y., & Hochberg, Y. (1995). Controlling the false discovery
rate: A practical and powerful approach to multiple testing.
Journal of the Royal Statistical Society. Series B (Methodological),
57(1), 289–300. https://doi.org/10.1111/j.2517-6161.1995
.tb02031.x
Bolsinger, J., Seifritz, E., Kleim, B., & Manoliu, A. (2018). Neuroim-
aging correlates of resilience to traumatic events—A comprehen-
sive review. Frontiers in Psychiatry, 9, 693. https://doi.org/10
.3389/fpsyt.2018.00693, PubMed: 30631288
Braun, U., Schäfer, A., Walter, H., Erk, S., Romanczuk-Seiferth, N.,
Haddad, L., Schweiger, J. I., Grimm, O., Heinz, A., Tost, H.,
Meyer-Lindenberg, A., & Bassett, D. S. (2015). Dynamic recon-
figuration of frontal brain networks during executive cognition in
humans. Proceedings of the National Academy of Sciences,
11 2 ( 3 7 ) , 11 6 7 8 – 11 6 8 3 . h t t p s : / / d o i . o rg / 1 0 . 1 0 7 3 / p n a s
.1422487112, PubMed: 26324898
Champely, S., Ekstrom, C., Dalgaard, P., Gill, J., Weibelzahl, S.,
Anandkumar, A., Ford, C., Volvic, R., & De Rosario, H. (2017).
pwr: Basic functions for power analysis. https://cran.r-project.org
/web/packages/pwr/
Chen, T., Cai, W., Ryali, S., Supekar, K., & Menon, V. (2016). Dis-
tinct global brain dynamics and spatiotemporal organization of
the salience network. PLOS Biology, 14(6), e1002469. https://
doi.org/10.1371/journal.pbio.1002469, PubMed: 27270215
Chmitorz, A., Wenzel, M., Stieglitz, R.-D., Kunzler, A., Bagusat, C.,
Helmreich, I., Gerlicher, A., Kampa, M., Kubiak, T., Kalisch, R.,
Lieb, K., & Tüscher, O. (2018). Population-based validation of a
German version of the Brief Resilience Scale. PLOS One, 13(2),
Network Neuroscience
192
Resting-state brain network dynamics and psychological resilience
e0192761. https://doi.org/10.1371/journal.pone.0192761,
PubMed: 29438435
Ciric, R., Wolf, D. H., Power, J. D., Roalf, D. R., Baum, G. L., Ruparel,
K., Shinohara, R. T., Elliott, M. A., Eickhoff, S. B., Davatzikos, C.,
Gur, R. C., Gur, R. E., Bassett, D. S., & Satterthwaite, T. D. (2017).
Benchmarking of participant-level confound regression strategies
for the control of motion artifact in studies of functional connec-
tivity. NeuroImage, 154, 174–187. https://doi.org/10.1016/j
.neuroimage.2017.03.020, PubMed: 28302591
Connor, K. M., & Davidson, J. R. T (2003). Development of a new
resilience scale: The Connor-Davidson Resilience Scale
(CD-RISC). Depression and Anxiety, 18(2), 76–82. https://doi
.org/10.1002/da.10113, PubMed: 12964174
Dadi, K., Rahim, M., Abraham, A., Chyzhyk, D., Milham, M.,
Thirion, B., & Varoquaux, G. (2019). Benchmarking functional
connectome-based predictive models for resting-state fMRI.
NeuroImage, 192, 115–134. https://doi.org/10.1016/j.neuroimage
.2019.02.062, PubMed: 30836146
Damaraju, E., Tagliazucchi, E., Laufs, H., & Calhoun, V. D. (2020).
Connectivity dynamics from wakefulness to sleep. NeuroImage,
220, 117047. https://doi.org/10.1016/j.neuroimage.2020
.117047, PubMed: 32562782
Dong, D., Wang, Y., Chang, X., Luo, C., & Yao, D. (2018). Dysfunc-
tion of large-scale brain networks in schizophrenia: A
meta-analysis of resting-state functional connectivity. Schizo-
phrenia Bulletin, 44(1), 168–181. https://doi.org/10.1093/schbul
/sbx034, PubMed: 28338943
Esteban, O., Birman, D., Schaer, M., Koyejo, O. O., Poldrack, R. A.,
& Gorgolewski, K. J. (2017). MRIQC: Advancing the automatic
prediction of image quality in MRI from unseen sites. PLoS
One, 12(9). https://doi.org/10.1371/journal.pone.0184661,
PubMed: 28945803
Esteban, O., Markiewicz, C. J., Blair, R. W., Moodie, C. A., Isik,
A. I., Erramuzpe, A., Kent, J. D., Goncalves, M., DuPre, E.,
Snyder, M., Oya, H., Ghosh, S. S., Wright, J., Durnez, J., Poldrack,
R. A., & Gorgolewski, K. J. (2019). fMRIPrep: A robust preprocess-
ing pipeline for functional MRI. Nature Methods, 16(1), 111–116.
https://doi.org/10.1038/s41592-018-0235-4, PubMed: 30532080
Faskowitz, J., Esfahlani, F. Z., Jo, Y., Sporns, O., & Betzel, R. F.
(2019). Edge-centric functional network representations of
human cerebral cortex reveal overlapping system-level architec-
ture. BioRxiv, 799924. https://doi.org/10.1101/799924
Finc, K., Bonna, K., He, X., Lydon-Staley, D. M., Kühn, S., Duch,
W., & Bassett, D. S. (2020). Dynamic reconfiguration of func-
tional brain networks during working memory training. Nature
Communications, 11(1), 2435. https://doi.org/10.1038/s41467
-020-15631-z, PubMed: 32415206
Friston, K. J., Williams, S., Howard, R., Frackowiak, R. S. J., &
Turner, R. (1996). Movement-related effects in fMRI time-series:
Movement artifacts in fMRI. Magnetic Resonance in Medicine,
35(3). https://doi.org/10.1002/mrm.1910350312, PubMed:
8699946
Garcia, J. O., Ashourvan, A., Muldoon, S. F., Vettel, J. M., & Bassett,
D. S. (2018). Applications of community detection techniques to
brain graphs: Algorithmic considerations and implications for
neural function. Proceedings of the IEEE. Institute of Electrical
and Electronics Engineers, 106(5), 846–867. https://doi.org/10
.1109/JPROC.2017.2786710, PubMed: 30559531
Genet, J. J., & Siemer, M. (2011). Flexible control in processing
affective and non-affective material predicts individual differ-
ences in trait resilience. Cognition & Emotion, 25(2), 380–388.
https://doi.org/10.1080/02699931.2010.491647, PubMed:
21432680
Gifford, G., Crossley, N., Kempton, M. J., Morgan, S., Dazzan, P.,
Young, J., & McGuire, P. (2020). Resting state fMRI based multi-
layer network configuration in patients with schizophrenia. Neu-
roImage: Clinical, 25, 102169. https://doi.org/10.1016/j.nicl
.2020.102169, PubMed: 32032819
Glasser, M. F., Sotiropoulos, S. N., Wilson, J. A., Coalson, T. S.,
Fischl, B., Andersson, J. L., Xu, J., Jbabdi, S., Webster, M.,
Polimeni, J. R., Van Essen, D. C., & Jenkinson, M. (2013). The
minimal preprocessing pipelines for the Human Connectome
Project. NeuroImage, 80, 105–124. https://doi.org/10.1016/j
.neuroimage.2013.04.127, PubMed: 23668970
Gorgolewski, K., Burns, C. D., Madison, C., Clark, D., Halchenko,
Y. O., Waskom, M. L., & Ghosh, S. S. (2011). Nipype: A flexible,
lightweight and extensible neuroimaging data processing frame-
work in Python. Frontiers in Neuroinformatics, 5. https://doi.org
/10.3389/fninf.2011.00013, PubMed: 21897815
Gratton, C., Dworetsky, A., Coalson, R. S., Adeyemo, B., Laumann,
T. O., Wig, G. S., Kong, T. S., Gratton, G., Fabiani, M., Barch,
D. M., Tranel, D., Miranda-Dominguez, O., Fair, D. A.,
Dosenbach, N. U. F., Snyder, A. Z., Perlmutter, J. S., Petersen,
S. E., & Campbell, M. C. (2020). Removal of high frequency con-
tamination from motion estimates in single-band fMRI saves data
without biasing functional connectivity. bioRxiv, 837161. https://
doi.org/10.1101/837161
Haimovici, A., Tagliazucchi, E., Balenzuela, P., & Laufs, H. (2017).
On wakefulness fluctuations as a source of BOLD functional
connectivity dynamics. Scientific Reports, 7(1), 5908. https://doi
.org/10.1038/s41598-017-06389-4, PubMed: 28724928
Hallquist, M. N., Hwang, K., & Luna, B. (2013). The nuisance of
nuisance regression: Spectral misspecification in a common
approach to resting-state fMRI preprocessing reintroduces noise
and obscures functional connectivity. NeuroImage, 82, 208–225.
https://doi.org/10.1016/j.neuroimage.2013.05.116, PubMed:
23747457
Harlalka, V., Bapi, R. S., Vinod, P. K., & Roy, D. (2019). Atypical
flexibility in dynamic functional connectivity quantifies the
severity in autism spectrum disorder. Frontiers in Human Neuro-
science, 13, 6. https://doi.org/10.3389/fnhum.2019.00006,
PubMed: 30774589
Harris, C. R., Millman, K. J., van der Walt, S. J., Gommers, R.,
Virtanen, P., Cournapeau, D., Wieser, E., Taylor, J., Berg, S.,
Smith, N. J., Kern, R., Picus, M., Hoyer, S., van Kerkwijk,
M. H., Brett, M., Haldane, A., del Río, J. F., Wiebe, M., Peterson,
P., … Oliphant, T. E. (2020). Array programming with NumPy.
Nature, 585(7825), 357–362. https://doi.org/10.1038/s41586
-020-2649-2, PubMed: 32939066
Hedge, C., Powell, G., & Sumner, P. (2018). The reliability paradox:
Why robust cognitive tasks do not produce reliable individual
differences. Behavior Research Methods, 50(3), 1166–1186.
https://doi.org/10.3758/s13428-017-0935-1, PubMed:
28726177
Hindriks, R., Adhikari, M. H., Murayama, Y., Ganzetti, M., Mantini,
D., Logothetis, N. K., & Deco, G. (2016). Can sliding-window
Network Neuroscience
193
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
1
1
7
5
2
0
0
2
4
4
6
n
e
n
_
a
_
0
0
2
1
6
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Resting-state brain network dynamics and psychological resilience
correlations reveal dynamic functional connectivity in
resting-state fMRI? NeuroImage, 127, 242–256. https://doi.org
/10.1016/j.neuroimage.2015.11.055, PubMed: 26631813
Hutchison, R. M., Womelsdorf, T., Allen, E. A., Bandettini, P. A.,
Calhoun, V. D., Corbetta, M., Della Penna, S., Duyn, J. H.,
Glover, G. H., Gonzalez-Castillo, J., Handwerker, D. A.,
Keilholz, S., Kiviniemi, V., Leopold, D. A., de Pasquale, F.,
Sporns, O., Walter, M., & Chang, C. (2013). Dynamic functional
connectivity: Promise, issues, and interpretations. NeuroImage,
80, 360–378. https://doi.org/10.1016/j.neuroimage.2013.05
.079, PubMed: 23707587
Iraji, A., Faghiri, A., Lewis, N., Fu, Z., Rachakonda, S., & Calhoun,
V. D. (2020). Tools of the trade: Estimating time-varying connec-
tivity patterns from fMRI data. Social Cognitive and Affective
Neuroscience, nsaa114. https://doi.org/10.1093/scan/nsaa114,
PubMed: 32785604
Jeub, L. G. S., Bazzi, M., Jutla, I. S., & Mucha, P. J. (2011–2019). A
generalized Louvain method for community detection imple-
mented in MATLAB, GitHub, https://github.com/GenLouvain
/GenLouvain
Jin, C., Jia, H., Lanka, P., Rangaprakash, D., Li, L., Liu, T., Hu, X., &
Deshpande, G. (2017). Dynamic brain connectivity is a better
predictor of PTSD than static connectivity. Human Brain
Mapping, 38(9), 4479–4496. https://doi.org/10.1002/ hbm
.23676, PubMed: 28603919
Kashdan, T. B., & Rottenberg, J. (2010). Psychological flexibility as
a fundamental aspect of health. Clinical Psychology Review,
30(7), 865–878. https://doi.org/10.1016/j.cpr.2010.03.001,
PubMed: 21151705
Leiner, D. J. (2013). Too fast, too straight, too weird: Post hoc iden-
tification of meaningless data in internet surveys. SSRN Electronic
Journal. https://doi.org/10.2139/ssrn.2361661
Leonardi, N., & Van De Ville, D. (2015). On spurious and real fluc-
tuations of dynamic functional connectivity during rest. Neuro-
Image, 104, 430–436. https://doi.org/10.1016/j.neuroimage
.2014.09.007, PubMed: 25234118
Leppert, K., Koch, B., Brähler, E., & Strauß, B. (2008). Die Resi-
lienzskala (RS) – Überprüfung der Langform RS-25 und einer
Kurzform RS-13.
Li, S., Hu, N., Zhang, W., Tao, B., Dai, J., Gong, Y., Tan, Y., Cai, D.,
& Lui, S. (2019). Dysconnectivity of multiple brain networks in
schizophrenia: A meta-analysis of resting-state functional con-
nectivity. Frontiers in Psychiatry, 10, 482. https://doi.org/10
.3389/fpsyt.2019.00482, PubMed: 31354545
Long, Y., Chen, C., Deng, M., Huang, X., Tan, W., Zhang, L., Fan,
Z., & Liu, Z. (2019). Psychological resilience negatively corre-
lates with resting-state brain network flexibility in young healthy
adults: A dynamic functional magnetic resonance imaging study.
Annals of Translational Medicine, 7(24), 809. https://doi.org/10
.21037/atm.2019.12.45, PubMed: 32042825
Lurie, D. J., Kessler, D., Bassett, D. S., Betzel, R. F., Breakspear, M.,
Kheilholz, S., Kucyi, A., Liégeois, R., Lindquist, M. A., McIntosh,
A. R., Poldrack, R. A., Shine, J. M., Thompson, W. H., Bielczyk,
N. Z., Douw, L., Kraft, D., Miller, R. L., Muthuraman, M.,
Pasquini, L., … Calhoun, V. D. (2020). Questions and controver-
sies in the study of time-varying functional connectivity in resting
fMRI. Network Neuroscience, 4(1), 30–69. https://doi.org/10
.1162/netn_a_00116, PubMed: 32043043
Lydon-Staley, D. M., Ciric, R., Satterthwaite, T. D., & Bassett, D. S.
(2019). Evaluation of confound regression strategies for the miti-
gation of micromovement artifact in studies of dynamic
resting-state functional connectivity and multilayer network
modularity. Network Neuroscience, 3(2), 427–454. https://doi
.org/10.1162/netn_a_00071, PubMed: 30793090
Maknojia, S., Churchill, N. W., Schweizer, T. A., & Graham, S. J.
(2019). Resting state fMRI: Going through the motions. Frontiers
in Neuroscience, 13, 825. https://doi.org/10.3389/fnins.2019
.00825, PubMed: 31456656
Menon, V. (2011). Large-scale brain networks and psychopathology:
A unifying triple network model. Trends in Cognitive Sciences,
15(10), 483–506. https://doi.org/10.1016/j.tics.2011.08.003,
PubMed: 21908230
Mucha, P. J., Richardson, T., Macon, K., Porter, M. A., & Onnela,
J.-P. (2010). Community structure in time-dependent, multiscale,
and multiplex networks. Science, 328(5980), 876–878. https://
doi.org/10.1126/science.1184819, PubMed: 20466926
Mulders, P. C., van Eijndhoven, P. F., Schene, A. H., Beckmann,
C. F., & Tendolkar, I. (2015). Resting-state functional connectivity
in major depressive disorder: A review. Neuroscience & Biobe-
havioral Reviews, 56, 330–344. https://doi.org/10.1016/j
.neubiorev.2015.07.014, PubMed: 26234819
Muldoon, S. F., & Bassett, D. S. (2016). Network and multilayer
network approaches to understanding human brain dynamics.
Philosophy of Science, 83(5), 710–720. https://doi.org/10.1086
/687857
Newman, M. E. J., & Girvan, M. (2004). Finding and evaluating
community structure in networks. Physical Review E, 69(2),
026113. https://doi.org/10.1103/PhysRevE.69.026113, PubMed:
14995526
Nolen-Hoeksema, S., Wisco, B. E., & Lyubomirsky, S. (2008).
Rethinking rumination. Perspectives on Psychological Science,
3(5), 400–424. https://doi.org/10.1111/j.1745-6924.2008.00088.x,
PubMed: 26158958
Paban, V., Modolo, J., Mheich, A., & Hassan, M. (2019). Psycholog-
ical resilience correlates with EEG source-space brain network
flexibility. Network Neuroscience, 3(2), 539–550. https://doi.org
/10.1162/netn_a_00079, PubMed: 30984906
Papadopoulos, L., Puckett, J. G., Daniels, K. E., & Bassett, D. S.
(2016). Evolution of network architecture in a granular material
under compression. Physical Review E, 94(3–1), 032908. https://
doi.org/10.1103/PhysRevE.94.032908, PubMed: 27739788
Parsons, S., Kruijt, A.-W., & Fox, E. (2016). A cognitive model of
psychological resilience. Journal of Experimental Psychopathol-
ogy, 7(3), 296–310. https://doi.org/10.5127/jep.053415
Pedersen, M., Zalesky, A., Omidvarnia, A., & Jackson, G. D. (2018).
Multilayer network switching rate predicts brain performance.
Proceedings of the National Academy of Sciences, 115(52),
13376–13381. https://doi.org/10.1073/pnas.1814785115,
PubMed: 30545918
Power, J. D., Cohen, A. L., Nelson, S. M., Wig, G. S., Barnes, K. A.,
Church, J. A., Vogel, A. C., Laumann, T. O., Miezin, F. M.,
Schlaggar, B. L., & Petersen, S. E. (2011). Functional network orga-
nization of the human brain. Neuron, 72(4), 665–678. https://doi
.org/10.1016/j.neuron.2011.09.006, PubMed: 22099467
Power, J. D., Lynch, C. J., Silver, B. M., Dubin, M. J., Martin, A., &
Jones, R. M. (2019). Distinctions among real and apparent
Network Neuroscience
194
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
1
1
7
5
2
0
0
2
4
4
6
n
e
n
_
a
_
0
0
2
1
6
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Resting-state brain network dynamics and psychological resilience
respiratory motions in human fMRI data. NeuroImage, 201,
116041. https://doi.org/10.1016/j.neuroimage.2019.116041,
PubMed: 31344484
Preti, M. G., Bolton, T. A., & Van De Ville, D. (2017). The dynamic
functional connectome: State-of-the-art and perspectives. Neuro-
Image, 160, 41–54. https://doi.org/10.1016/j.neuroimage.2016
.12.061, PubMed: 28034766
Prichard, D., & Theiler, J. (1994). Generating surrogate data for time
series with several simultaneously measured variables. Physical
Review Letters, 73(7), 951–954. https://doi.org/10.1103
/PhysRevLett.73.951, PubMed: 10057582
Sarubin, N., Gutt, D., Giegling, I., Bühner, M., Hilbert, S.,
Krähenmann, O., Wolf, M., Jobst, A., Sabaß, L., Rujescu, D.,
Falkai, P., & Padberg, F. (2015). Erste Analyse der psychometrischen
Eigenschaften und Struktur der deutschsprachigen 10- und 25-Item
Version der Connor-Davidson Resilience Scale (CD-RISC).
Zeitschrift für Gesundheitspsychologie, 23(3), 112–122. https://
doi.org/10.1026/0943-8149/a000142
Savva, A. D., Mitsis, G. D., & Matsopoulos, G. K. (2019). Assess-
ment of dynamic functional connectivity in resting-state fMRI
using the sliding window technique. Brain and Behavior, 9(4),
e01255. https://doi.org/10.1002/brb3.1255, PubMed: 30884215
Schaefer, A., Kong, R., Gordon, E. M., Laumann, T. O., Zuo, X.-N.,
Holmes, A. J., Eickhoff, S. B., & Yeo, B. T. T (2018). Local-global
parcellation of the human cerebral cortex from intrinsic func-
tional connectivity MRI. Cerebral Cortex, 28(9), 3095–3114.
https://doi.org/10.1093/cercor/bhx179, PubMed: 28981612
SciPy 1.0 Contributors, Virtanen, P., Gommers, R., Oliphant, T. E.,
Haberland, M., Reddy, T., Cournapeau, D., Burovski, E.,
Peterson, P., Weckesser, W., Bright, J., van der Walt, S. J., Brett,
M., Wilson, J., Millman, K. J., Mayorov, N., Nelson, A. R. J, Jones,
E., Kern, R., … van Mulbregt, P. (2020). SciPy 1.0: Fundamental
algorithms for scientific computing in Python. Nature Methods,
17(3), 261–272. https://doi.org/10.1038/s41592-019-0686-2,
PubMed: 32015543
Shakil, S., Lee, C.-H., & Keilholz, S. D. (2016). Evaluation of sliding
window correlation performance for characterizing dynamic
functional connectivity and brain states. NeuroImage, 133,
111–128. https://doi.org/10.1016/j.neuroimage.2016.02.074,
PubMed: 26952197
Sheehan, D. V., Lecrubier, Y., Sheehan, K. H., Amorim, P., Janavs, J.,
Weiller, E., Hergueta, T., Baker, R., & Dunbar, G. C. (1998). The
Mini-International Neuropsychiatric Interview (M.I.N.I.): The
development and validation of a structured diagnostic psychiat-
ric interview for DSM-IV and ICD-10. The Journal of Clinical Psy-
chiatry, 59 Suppl 20, 22–33; quiz 34–57. PubMed: 9881538
Smith, B. W., Dalen, J., Wiggins, K., Tooley, E., Christopher, P., &
Bernard, J. (2008). The brief resilience scale: Assessing the ability
to bounce back. International Journal of Behavioral Medicine,
15(3), 194–200. https://doi.org/10.1080/10705500802222972,
PubMed: 18696313
Southwick, S. M., & Charney, D. S. (2012). The science of resil-
ience: Implications for the prevention and treatment of depres-
sion. Science, 338(6103), 79–82. https://doi.org/10.1126
/science.1222942, PubMed: 23042887
Tzourio-Mazoyer, N., Landeau, B., Papathanassiou, D., Crivello, F.,
Etard, O., Delcroix, N., Mazoyer, B., & Joliot, M. (2002). Auto-
mated anatomical labeling of activations in SPM using a macro-
scopic anatomical parcellation of the MNI MRI single-subject
brain. NeuroImage, 15(1), 273–289. https://doi.org/10.1006
/nimg.2001.0978, PubMed: 11771995
Vallat, R. (2018). Pingouin: Statistics in Python. Journal of Open
Source Software, 3(31), 1026. https://doi.org/10.21105/joss
.01026
Vergara, V. M., Abrol, A., & Calhoun, V. D. (2019). An average
sliding window correlation method for dynamic functional con-
nectivity. Human Brain Mapping, 40(7), 2089–2103. https://doi
.org/10.1002/hbm.24509, PubMed: 30659699
Wagnild, G. M., & Young, H. M. (1993). Development and psycho-
metric evaluation of the Resilience Scale. Journal of Nursing
Measurement, 1(2), 165–178. PubMed: 7850498
Williams, J. C., & Van Snellenberg, J. X. (2019). Motion denoising of
multiband resting state functional connectivity MRI data: An
improved volume censoring method [Preprint]. Neuroscience.
https://doi.org/10.1101/860635
Windle, G., Bennett, K. M., & Noyes, J. (2011). A methodological
review of resilience measurement scales. Health and Quality of
Life Outcomes, 9, 8. https://doi.org/10.1186/1477-7525-9-8,
PubMed: 21294858
Yang, Z., & Lewis, L. D. (2021). Imaging the temporal dynamics of
brain states with highly sampled fMRI. Current Opinion in Behav-
ioral Sciences, 40, 87–95. https://doi.org/10.1016/j.cobeha.2021
.02.005, PubMed: 34734109
Yang, Z., Telesford, Q. K., Franco, A. R., Lim, R., Gu, S., Xu, T., Ai,
L., Castellanos, F. X., Yan, C.-G., Colcombe, S., & Milham, M. P.
(2021). Measurement reliability for individual differences in
multilayer network dynamics: Cautions and considerations. Neu-
roImage, 225, 117489. https://doi.org/10.1016/j.neuroimage
.2020.117489, PubMed: 33130272
Yin, W., Li, T., Hung, S.-C., Zhang, H., Wang, L., Shen, D., Zhu, H.,
Mucha, P. J., Cohen, J. R., & Lin, W. (2020). The emergence of a
functionally flexible brain during early infancy. Proceedings of
the National Academy of Sciences, 202002645. https://doi.org
/10.1073/pnas.2002645117, PubMed: 32868436
Zalesky, A., Fornito, A., Cocchi, L., Gollo, L. L., & Breakspear, M.
(2014). Time-resolved resting-state brain networks. Proceedings
of the National Academy of Sciences, 111(28), 10341–10346.
https://doi.org/10.1073/pnas.1400181111, PubMed: 24982140
Network Neuroscience
195
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
1
1
7
5
2
0
0
2
4
4
6
n
e
n
_
a
_
0
0
2
1
6
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3