Pragmatics in Action: Indirect Requests Engage Theory
of Mind Areas and the Cortical Motor Network
Markus J. van Ackeren1, Daniel Casasanto2, Harold Bekkering3,
Peter Hagoort3,4, and Shirley-Ann Rueschemeyer1,3
Abstract
■ Research from the past decade has shown that understanding
the meaning of words and utterances (i.e., abstracted symbols)
engages the same systems we used to perceive and interact with
the physical world in a content-specific manner. For example,
understanding the word “grasp” elicits activation in the cortical
motor network, that is, part of the neural substrate involved in
planned and executing a grasping action. In the embodied litera-
ture, cortical motor activation during language comprehension
is thought to reflect motor simulation underlying conceptual
knowledge [note that outside the embodied framework, other
explanations for the link between action and language are
offered, e.g., Mahon, B. Z., & Caramazza, A. A critical look at the
embodied cognition hypothesis and a new proposal for grouding
conceptual content. Journal of Physiology, 102, 59–70, 2008;
Hagoort, P. On Broca, brain, and binding: A new framework.
Trends in Cognitive Sciences, 9, 416–423, 2005]. Previous re-
search has supported the view that the coupling between lan-
guage and action is flexible, and reading an action-related word
form is not sufficient for cortical motor activation [Van Dam,
W. O., van Dijk, M., Bekkering, H., & Rueschemeyer, S.-A. Flexibil-
ity in embodied lexical–semantic representations. Human Brain
Mapping, doi: 10.1002/hbm.21365, 2011]. The current study goes
one step further by addressing the necessity of action-related
word forms for motor activation during language comprehension.
Subjects listened to indirect requests (IRs) for action during an
fMRI session. IRs for action are speech acts in which access to
an action concept is required, although it is not explicitly encoded
in the language. For example, the utterance “It is hot here!” in a
room with a window is likely to be interpreted as a request to
open the window. However, the same utterance in a desert will
be interpreted as a statement. The results indicate (1) that
comprehension of IR sentences activates cortical motor areas
reliably more than comprehension of sentences devoid of any
implicit motor information. This is true despite the fact that IR
sentences contain no lexical reference to action. (2) Compre-
hension of IR sentences also reliably activates substantial por-
tions of the theory of mind network, known to be involved in
making inferences about mental states of others. The implica-
tions of these findings for embodied theories of language are
discussed. ■
INTRODUCTION
Human language is the most sophisticated communication
system in the animal kingdom. Specifically, language allows
us to encode complex semantic knowledge in a very con-
cise, symbolic way. However, the relationship between
symbolic representations and our knowledge of the physi-
cal world is still debated. Embodied theories of language
postulate that understanding the meaning of words and
utterances engages the same systems we use to perceive
and interact with the physical world (Barsalou, 1999, 2008;
Fischer & Zwaan, 2008; Lakoff & Johnson, 1999). For ex-
ample, understanding the word “grasp” elicits activation in
the cortical motor network. In the embodied literature,
cortical motor activation during language comprehension is
thought to reflect motor simulation underlying conceptual
1University of York, UK, 2New School for Social Research, New
York, NY, 3Donders Institute for Brain, Cognition and Behaviour,
Nijmegen, The Netherlands, 4Max Planck Institute for Psycho-
linguistics, Nijmegen, The Netherlands
knowledge (Barsalou, 1999, 2008). Alternative perspectives,
however, such as after-effects of conceptual understanding
(Mahon & Caramazza, 2008) and decoding propositional
content (Hagoort, 2005), have also been put forth.
Although the functional relevance of the link between
language and action remains a topic of debate, there is
little doubt that the connection exists: A plethora of pre-
vious studies using a variety of experimental techniques
have reliably demonstrated that language referring ex-
plicitly to actions (e.g., object nouns, action verbs, and
action sentences) modulates the activation in the cor-
tical motor network (Glenberg & Gallese, 2011; Willems,
Ludovica, DʼEsposito, Ivry, & Casasanto, 2011; Van Dam,
Rueschemeyer, & Bekkering, 2010; Rüschemeyer, Brass, &
Friederici, 2007; Gallese & Lakoff, 2005; Tettamanti et al.,
2005; Hauk, Johnsrude, & Pulvermüller, 2004; Glenberg
& Kaschak, 2002; Glenberg, 2000). For example, Hauk and
colleagues (2004) demonstrated that regions that respond
to movement execution with the hand, foot, or mouth will
also respond to verbs denoting actions (e.g., pick, kick,
and lick). Specifically, the activation patterns revealed a
© 2012 Massachusetts Institute of Technology
Journal of Cognitive Neuroscience 24:11, pp. 2237–2247
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
4
/
1
2
1
4
/
2
1
2
1
3
/
7
2
1
2
9
3
4
7
4
/
7
1
8
6
7
7
o
8
c
6
n
3
_
8
a
/
_
j
0
o
0
c
2
n
7
4
_
a
p
_
d
0
0
b
2
y
7
g
4
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
f
t
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
somatotopic organization of action verbs in the primary
and premotor cortices. In another study, Willems and col-
leagues (2011) applied off-line theta-burst TMS to either
the left or the right premotor cortex of their subjects. Sub-
sequently, subjects made lexical decisions on verbs denot-
ing manual actions (e.g., to throw, to write) or nonmanual
actions (e.g., to earn, to wander). The results indicated
that subjects were faster in making lexical decisions on
manual action verbs than nonmanual action verbs when
TMS was applied to the left premotor cortex. This finding
suggests a functional contribution of the cortical motor
system to language understanding. Taken together, these
results have been interpreted as evidence that semantic
knowledge is instantiated on-line in modality-specific areas
in the brain.
There is still much debate regarding the importance
of the action-related word form in activating the cortical
motor network during language comprehension. Specifi-
cally, previous research has challenged the assumption
that reading an action-related word form automatically
and invariantly activates the cortical motor system. For
example, in idiomatic expressions like “kick the bucket,”
the literal meaning of the utterance denotes an action with
the foot. However, the speaker meaning denotes that a
person has passed away. Raposo, Moss, Stamatakis, and
Tyler (2009) compared idiomatic expressions like these
with literal sentences denoting actions and found that
the former did not activate the cortical motor network in
the same way as literal sentences (but see also Boulenger,
Hauk, & Pulvermüller, 2011; Desai, Binder, Conant, Mano
& Seidenberg, 2011). This is in line with other results show-
ing less cortical motor activation for nonconventional, non-
literal sentences (e.g., handling the truth) and complex
words with motor stems (Rüschemeyer et al., 2007; Aziz-
Zadeh, Wilson, Rizzolatti, & Iacoboni, 2006). These results
have supported the view that the coupling between lan-
guage and action is flexible, and reading an action-related
word form is not sufficient for cortical motor activation.
The fact that the relation between word forms referring
explicitly to actions and activation of the cortical motor
network is not static raises the question whether action
words are even necessary to elicit cortical motor activa-
tion during language comprehension. Previous research
has shown that word meaning activates the cortex in a
modality-specific way (Rüschemeyer et al., 2007; Tettamanti
et al., 2005; Hauk et al., 2004). However, during natural
communication, we do not always express meaning in a
literal way. For example, during a conversation, a speaker
might mention the fact that “it is quite warm in here.” In
the appropriate context, this utterance will be interpreted
as an indirect request (IR) to open the window, although
no explicit action-related word form has been used. Inter-
preting such a statement as an IR relies on an inference
rather than a learned association between form and mean-
ing (Holtgraves, 1994). In other words, the listener has to
be aware of the intention of the speaker to interpret the
utterance correctly.
In social psychology, making inferences about mental
states of others is referred to as having a theory of mind
(ToM). Recent research has indicated that areas that are
activated when we think about mental states of others in
general are also sensitive to pragmatic aspects of language
understanding and production (Willems et al., 2010; Sassa
et al., 2007). In summary, IRs may refer to an action con-
cept in the absence of an explicit action-related word.
Therefore, these utterances allow us to investigate the
possibility of triggering activation of the cortical motor
network in the absence of action words.
Previous research has shown that action-related word
forms are not sufficient for cortical motor activations.
The goal of this study goes a step further by investigat-
ing whether explicit word forms are a necessary condition
for cortical motor activations during language compre-
hension. Hemodynamic changes in the brain were mea-
sured while subjects listened to IRs for actions (e.g., “It
is very hot here”). It was hypothesized that (a) the im-
plied meaning of an action should elicit a cortical motor
activation. Specifically, we expected activation in areas
that are also involved in movement planning, object ma-
nipulation and action goals, such as BA 6 and the inferior
parietal lobule (IPL; Fogassi et al., 2005). Importantly,
these areas have also been associated with understand-
ing actions through language in a large number of stud-
ies ( Van Dam et al., 2010; Postle, McMahon, Ashton, &
de Zubicaray, 2008; Rüschemeyer et al., 2007). Sensitivity
of these regions to IRs would suggest that cortical motor
activation during language comprehension is not depen-
dent on the presence of action words but could be purely
the outcome of a pragmatic inference. In addition, we ex-
pected that (b) this inferential step will be reflected by a
higher level of activation in classical ToM areas such as
the medial pFC (mPFC) and the TPJ (Gallagher & Frith,
2003).
METHODS
Participants
The participants were 16 students from the local uni-
versity. Three volunteers had to be excluded from the
data because of excessive movement, response errors,
and health-related problems that were not known to
the experimenter before the experiment. The remain-
ing 13 participants were all healthy women between the
ages of 18 and 24 years with normal or corrected-to-normal
vision and no hearing impairments (n = 13; mean age =
21.39 years). All participants were native speakers of Dutch
and right-handed. In addition, none of the participants
reported any known neurological impairment. Before the
experiment, participants gave written informed consent in
accordance with the Declaration of Helsinki. For their par-
ticipation, subjects either received a financial compensa-
tion or course credits. The study fell under the legal ethical
approval procedure in the Netherlands.
2238
Journal of Cognitive Neuroscience
Volume 24, Number 11
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
4
/
1
2
1
4
/
2
1
2
1
3
/
7
2
1
2
9
3
4
7
4
/
7
1
8
6
7
7
o
8
c
6
n
3
_
8
a
/
_
j
0
o
0
c
2
n
7
4
_
a
p
_
d
0
0
b
2
y
7
g
4
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
.
/
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Stimuli
The stimuli consisted of 128 images/visual scenes and
128 spoken sentences. The spoken sentences were re-
cordings from a native speaker of Dutch. The visual scenes
were assembled from multiple image search engines on
the Web, which are publicly available (e.g., flickr.com;
images/google.com).
The stimuli were segregated into 64 unique item sets.
One item set comprised four distinct sentence–scene
combinations. The four conditions were IR, picture con-
trol (PC), utterance control (UC), and picture–utterance
control (PUC). Within each item set, only one of these
combinations was associated with an IR. Importantly,
each request implied a manual action only. The other
three sentence–scene combinations could be interpreted
as a plausible statement but not as requests. However,
the three control conditions were included to control for
the effect of the visual scene and the utterance alone.
This assumption was tested in an on-line survey before
MRI data acquisition. Participants in the questionnaire
study (n = 20) viewed randomized combinations of one
visual scene and one sentence from a single item set.
Their task was to decide whether they thought that the
interlocutor wanted something from them. The items in
the IR condition were interpreted as requests much more
often than in the other three conditions (IR: M = 71%;
SE = 4; PC: M = 26%; SE = 3; UC: M = 18%; SE = 2;
PUC: M = 14%; SE = 2). This difference was significant
at an alpha level of .05, F(1.92, 36.33) = 110.31, p < .001,
partial η2 = .85.1
In addition, a localizer task was used to identify (a) re-
gions that were specifically activated when thinking
about another personʼs believes and desires (ToM) and
(b) regions that were involved in simple hand actions.
The ToM network was identified with the localizer during
time windows at which the subjects read a story and sub-
sequently judged statements about the story. The stories
required either an inference on the physical state of an
object (false photograph) or another personʼs belief (false
belief ). Each participant saw 24 physical state and 24 men-
tal state stories. The stimuli were introduced by the Saxe
lab2 (Dodell-Feder, Koster-Hale, Bedny, & Saxe, 2010; Saxe
& Kanwisher, 2003) and translated into Dutch for the
present experiment. Regions that were involved in simple
hand actions were localized during time windows in which
subjects produced button presses with their right index
and middle finger. Hand regions were targeted because
the indirect requests required simple hand actions as well
(Figure 1).
Stimulus Presentation
For both the main task and the functional localizer, sub-
jects lay supine in the scanner. All visual materials were
presented via a projector outside the scanner. Participants
viewed the screen via a nonmagnetic mirror. The auditory
Figure 1. Illustration of the fMRI paradigm for IRs. The top half
shows the time course of the experiment. On each trial, a fixation
cross was presented for 500 msec, followed by a visual scene. The
utterance was presented auditorily, 200 msec after picture onset.
Each trial lasted exactly 3000 msec. On 15% of the trials, the subject
was prompted to indicate whether the utterance was a request. The
bottom half depicts the design of the study with a single item set.
The conditions are IR, PC, UC, and PUC.
stimuli were presented via nonmagnetic headphones that
also dampened the noise from the scanner. Before the
start of the experiment, the volume of the headphones
was adjusted to the subjects convenience. Participantsʼ
responses to the tasks were recorded via a nonmagnetic
button box inside the scanner.
The implicature task was an epoch-related design in
which each participant saw two independent sentence–
scene combinations from each item set, resulting in a
total number of 128 stimuli per participant (32 items
per condition). The items were individually pseudo-
randomized in such a way that the same condition was
never presented more than twice in a row. Additionally,
16 null events were included in the design. To maintain
participantsʼ vigilance and ensure that they processed the
stimuli more deeply, 15% of the experimental trials were
accompanied by a catch question (“Do you think that the
person made a request?”), to which participants could
respond with a button press indicating either a “yes” or
“no” response. To make the trial onset unpredictable for
the subject and to enhance the resolution of the time
window within a trial, the intertrial interval was randomly
jittered in a range of 4000–6000 msec (M = 5000 msec). The
trial began with the presentation of a fixation cross for
500 msec, followed by the visual scene. After 200 msec,
the sentence stimulus was presented (mean duration =
1357 msec). At the sentence offset, a variable interval filled
the remaining time so that every picture presentation
lasted 2300 msec. Thus, every trial lasted exactly 3000 msec.
van Ackeren et al.
2239
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
1
2
1
4
/
2
1
2
1
3
/
7
2
1
2
9
3
4
7
4
/
7
1
8
6
7
7
o
8
c
6
n
3
_
8
a
/
_
j
0
o
0
c
2
n
7
4
_
a
p
_
d
0
0
b
2
y
7
g
4
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
.
f
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Participants were instructed to listen to the sentences care-
fully and decide whether they think the person wanted
something from them or not while listening. Before the
actual start of the experiment, there was a practice run
outside the scanner.
After the implicature task, subjects proceeded with the
localizer task. The procedure of this task is described in
Dodell-Feder et al. (2010). Also for this task, there was a
practice run outside the scanner, before the experiment.
fMRI Data Acquisition
MRI data acquisition was performed on a Siemens Magnetom
Trio scanner (Siemens Medical System, Erlangen, Germany)
with a magnetic field strength of 3 T. The functional scans
for the implicature task and the localizer were acquired
using a multiecho gradient pulse sequence (repetition
time = 2390 msec; echo time = 9.4, 21.17, 32.94, 44.71,
and 56.48 msec; flip angle = 90°). Each volume consisted
of 31 transversal slices with a thickness of 3 mm. The voxel
resolution was 3.5 mm × 3.5 mm × 3.5 mm.
After the collection of functional data, a structural scan
was performed for each individual participant. The im-
age was a T1-weighted 3-D MPRAGE sequence compris-
ing 192 sagittal slices (repetition time = 2300 msec; echo
time = 3.03 msec; slice thickness = 1 mm).
fMRI Data Analysis
The raw MR images were preprocessed and analyzed using
the Matlab toolbox SPM8 (Statistical Parametric Mapping,
www.fil.ion.ucl.uk/spm). Before the analysis, the first six
volumes were excluded to control for T1 equilibration
effects. Six movement parameters (three translations and
three rotations) were extracted from the first echo of
each volume and subsequently used to correct for small
head movements in all five echoes of each volume. Sub-
sequently, all five echoes were combined into a single vol-
ume using a weighted average. To correct for delays in
slice timing during image acquisition, the time courses of
each voxel were realigned toward slice 16. After segmenta-
tion into gray and white matter, images were normalized
to a standard EPI template within Montreal Neurological
Institute (MNI) space and resampled at an isotropic voxel
size of 3 mm. Lastly, the images were convolved with a
Gaussian smoothing kernel with 8-mm FWHM. To correct
for slow drifts in the signal, a high pass filter was applied
at 128 sec.
The combined and preprocessed time series of the
implicature task was analyzed as an event-related design
(epoch = 1.5 sec) on a subject-by-subject basis. Within a
general linear model framework, each condition was con-
volved with a canonical hemodynamic response function
(HRF) and used as a regressor. In addition, the movement
parameters from the realignment algorithm, time, and dis-
persion derivatives were included as effects of no interest.
A single contrast comparing IR versus three control (PC,
UC, and PUC) conditions was generated for each participant
individually. Because the images from each subject had
been aligned to standard MNI space, a second level ran-
dom effects analysis could be performed at the group
level. The critical contrasts from all participants were in-
cluded in the model, and a group analysis was performed
using a one-sample t test. To control for multiple compari-
sons, a cluster extend threshold was determined using a
Monte Carlo simulation with 1000 iterations (Slotnick,
Moo, Segal, & Hart, 2003). The simulation results indicated
that a cluster with p < .0001 (uncorrected) and a cluster size
k > 12 (324 mm3) was significant at p < .01 (corrected).
The localizer task was used to identify areas that were
selectively active during inferences about mental states
and hand movements. To extract the signal, which was
related to the ToM network, the images were analyzed as
a block design. Each block was defined as the period of
time from the onset of the story to the offset of the state-
ment. Subsequently, this time window was convolved with
a canonical HRF. Only movement parameters were in-
cluded as effects of no interest. A contrast comparing the
false belief stories versus the false photograph stories
was created for each subject individually. Subsequently,
a random effects group analysis was conducted on the
individual participant contrast images using a one-sample
t test.
Additionally, button presses with the right hand were
analyzed to identify regions that are involved in action
execution. Manual button presses were analyzed as an
event-related design. The moment a subject pressed
a button was modeled with a canonical HRF and used
as a regressor. In addition, the movement parameters
from the realignment algorithm, time, and dispersion
derivatives were used as effects of no interest.
ROI Analysis
ROI analyses were conducted to investigate whether the
condition in which the utterance and the visual context
formed an implicature exerted a stronger effect on (a) the
neural motor network that is involved in hand actions
and ( b) the ToM network. Four ROIs for the ToM
network were defined as the clusters in the whole-brain
analysis that were sensitive to the contrast false belief
versus false photograph (Table 1). With respect to the
action network, there were very strong anatomical
hypotheses. Therefore, the contrast image for hand
actions (Action > 0) from the second-level whole-brain
analysis was masked with cytoarchitectonically defined
probability maps of BA 6 (Geyer, 2004) and left and right
IPL/PF (Caspers et al., 2006, 2008; Table 2). The former
will be referred to as IPL in the remainder of this discus-
sion. Subsequently, MNI coordinates for peak values
within the largest active cluster were used to create spheres
of 6-mm radius using the ROI toolbox Marsbar (Brett,
Anton, Valabregue, & Poline, 2002).
2240
Journal of Cognitive Neuroscience
Volume 24, Number 11
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
4
/
1
2
1
4
/
2
1
2
1
3
/
7
2
1
2
9
3
4
7
4
/
7
1
8
6
7
7
o
8
c
6
n
3
_
8
a
/
_
j
0
o
0
c
2
n
7
4
_
a
p
_
d
0
0
b
2
y
7
g
4
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
t
/
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
The ROIs from the localizer task were interrogated
with respect to the four conditions (IR, PC, UC, PUC)
from the implicature task. Percent signal change was ex-
tracted and averaged within each participant. Thus, for
each of the 13 participants in our study, there were four
values. With these four conditions, ANOVAs with repeated
measures were conducted for each ROI.
RESULTS
Behavioral Results
Behavioral responses to catch trials were analyzed to test
whether subjects responded as predicted by the question-
naire study. First, a one-sample t test was conducted on
the percentage of correct responses to assess whether
participants were able to do the implicature task. This as-
sumption was confirmed, t(12) = 9.64, p < .001, M =
80%, SE = 3. Subsequently an ANOVA with repeated mea-
sures was conducted on the RTs to test for differences
in task difficulty between the conditions (IR, PC, UC,
PUC). This test was significant, F(2.14, 24.43) = 3.56,
p < .05, partial η2 = .23. Planned comparisons revealed
participants responded to the IR condition faster than
to the PC [F(1, 12) = 13.13, p < .005, partial η2 = .52;
IR: M = 045 msec, SE = 86 msec; PC: M = 1299 msec,
SE = 116 msec], the UC [F(1, 12) = 6.1, p < .05, partial
η2 = .34; UC: M = 1193 msec, SD = 91 msec], and PUC
[F(1, 12) = 5.05, p < .05, partial η2 = .3; PUC: M = 1314
msec, SE = 131 msec] condition. The fact that participants
responded faster, giving positive responses, is not surpris-
ing; however, it shows that recognizing IR was not more
difficult than recognizing statements. Lastly, an ANOVA
with repeated measures tested whether participants recog-
nized requests more often in the IR condition. The results
replicated the findings from the questionnaires, F(2.74,
32.93) = 49.76, p < .001, partial η2 = .81. Specifically,
requests were more often identified in the IR condition than
in the PC [F(1, 12) = 113.81, p < .001, partial η2 = .91; IR:
M = 83%, SE = 4; PC: M = 19%, SE = 4], UC [F(1, 12) =
71.44, p < .001, partial η2 = .86; UC: M = 34%, SE = 6],
and PUC [F(1, 12) = 101.57, p < .001, partial η2 = .9;
PUC: M = 12%, SE = 7] condition.
fMRI Results
ToM Localizer
A whole-brain analysis on the ToM localizer was conducted,
in which the story and the statement were modeled as one
block. The pattern of results replicated previous findings
in English (Dodell-Feder et al., 2010). That is, regions that
are part of the ToM network showed a stronger BOLD
response for false belief stories than for false photograph
stories (Table 1). These were clusters in the left and right
TPJ, the left precuneus, and the mPFC. These clusters were
interrogated in the subsequent ROI analysis of the ToM
network.
Action Localizer
To identify regions that were sensitive to action prepara-
tion or execution, a whole-brain analysis was conducted
on the localizer. Specifically, the moment of a button
press, convolved with a canonical HRF, was used as re-
gressor. Because there were very specific anatomical pre-
dictions, the image was masked with an anatomical map
of BA 6 and bilateral IPL. Table 2 shows the peak activa-
tions within these regions. Specifically, activation peaks
were found in the left superior frontal gyrus (SFG), left
precentral gyrus, and left medial SFG. In functional terms,
these peaks are located within the left premotor cortex
and the left pre-SMA. In addition, there were peak ac-
tivations in the left and right IPL, overlapping with the
supramarginal gyrus. The peak activations in the frontal
motor regions (left SFG, left PCG, and left pre-SMA) as
well as the strongest activation peak in left and right IPL
were used to create 6-mm spheres for the subsequent
ROI analysis.
Whole-brain Analysis
An overview of significant peak activations in the whole-
brain analysis of the implicature task is depicted in Table 3.
On the medial surface of the brain, a widespread cluster
of activation was found in the mPFC and the insular cor-
tex, extending into the OFC. Additionally, regions in the
posterior and middle cingulate cortex were sensitive to
Table 1. Brain Regions from Whole-brain Analysis of the Localizer Task Showing Significantly More Activation during False
Belief versus False Photograph Stories ( p < .0001, k > 12)
Region
Precuneus
Left mPFC
Right TPJ
Left TPJ
Cluster Level Extent ( Voxels)
491
23
115
156
Only the largest peak voxel per cluster is depicted.
Peak Voxel Level
MNI Coordinates
t
14.56
8.33
7.88
8.93
equivZ
5.83
4.71
4.59
4.86
x
−3
−6
57
−51
y
−58
50
−55
−49
z
22
40
22
31
van Ackeren et al.
2241
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
4
/
1
2
1
4
/
2
1
2
1
3
/
7
2
1
2
9
3
4
7
4
/
7
1
8
6
7
7
o
8
c
6
n
3
_
8
a
/
_
j
0
o
0
c
2
n
7
4
_
a
p
_
d
0
0
b
2
y
7
g
4
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
f
t
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Table 2. Brain Regionʼs Whole-brain Analysis of the Localizer Task that Were Significantly Active during Button Presses with the
Right Hand ( p < .0001, k > 12)
Region
BA 6
Left SFG
Left precentral gyrus
Left medial SFG
Left inferior parietal cortex (PF)
Left inferior parietal lobule
Right inferior parietal cortex (PF)
Right supramarginal gyrus
Cluster Level Extent ( Voxels)
t
equivZ
x
y
Peak Voxel Level
MNI Coordinates
458
113
40
12.66
11.84
10.90
5.56
5.43
5.27
−24
−24
−6
−1
−10
23
13.21
5.64
−48
−46
7.00
4.34
66
−40
z
67
64
58
37
28
The results were restricted to anatomically defined neural motor regions (BA 6, bilateral IPL).
the manipulation. On the lateral surface, there were clus-
ters of activation around the left and right TPJs. However,
the latter was much more dominant in the left hemisphere.
Lastly, both thalami showed selective activation to IR.
ROI Analysis
In the ROI analysis, ROIs from the ToM and action localizer
were interrogated with respect to the average percent sig-
nal change in the implicature task. All inferential statistics
in this section were evaluated at an alpha level of p < .05.
ANOVAs with repeated measures were corrected for viola-
tions of sphericity using Greenhouse–Geisser correction.
ToM ROIs
ANOVAs with repeated measures were conducted for each
ROI from the ToM localizer (mPFC, precuneus, left TPJ,
and right TPJ) to test whether these regions were sensitive
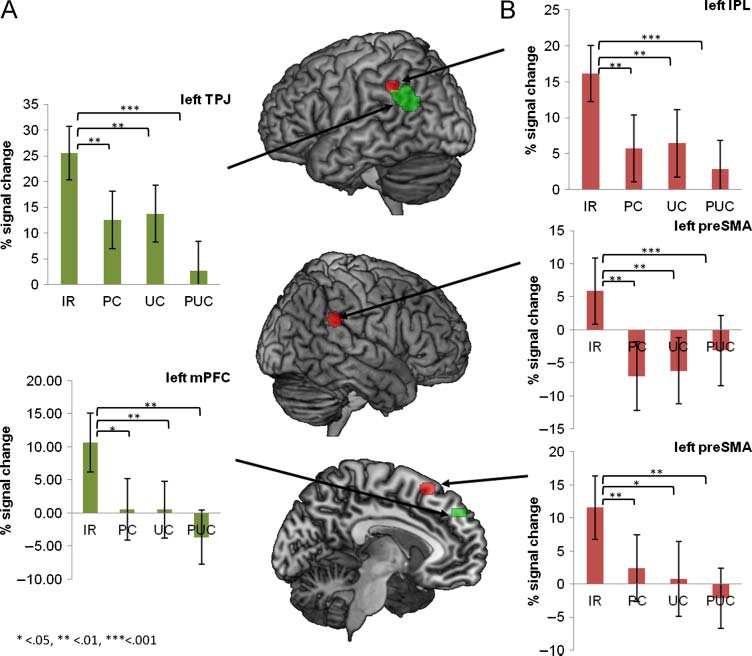
to IRs. Interrogation of the mPFC revealed a significant
main effect of Condition, F(2.39, 28.72) = 4.68, p < .05,
partial η2 = .28. Planned comparisons indicated that the
average percent signal change was higher in the IR con-
dition than in the three control conditions [IR vs. PC: F(1,
12) = 5.39, p < .05, partial η2 = .31; IR: M = .11, SE = .04;
PC: M = .01, SE = .05; IR vs. UC: F(1, 12) = 14.14, p < .005,
partial η2 = .54; UC: M = .01, SE = .04; IR vs. PUC:
F(1, 12) = 16.29, p < .005, partial η2 = .58; PUC: M =
−.04, SE = .04; Figure 2A]. ANOVAs on the left TPJ also
revealed a significant main effect of Condition, F(2.35,
28.21) = 10.47, p < .001, partial η2 = .47. Specifically,
planned comparisons showed that average percent signal
change was higher in the IR condition than in the control
conditions [IR vs. PC: F(1, 12) = 15.19, p < .005, partial
η2 = .56; IR: M = .26, SE = .05; PC: M = .13, SE = .06;
IR vs. UC: F(1, 12) = 12.91, p < .005, partial η2 = .52;
UC: M = .14, SE = .06; IR vs. PUC: F(1, 12) = 27.02, p <
.001, partial η2 = .69; PC: M = .03, SE = .06; Figure 2A]. In
addition, there was a significant main effect in the right TPJ,
F(1.92, 22.98) = 5.04, p < .05, partial η2 = .3. Planned
comparisons revealed the same effects as in the left TPJ,
albeit that the contrast with the UC condition was only mar-
ginally significant [IR vs. PC: F(1, 12) = 11.91, p < .01, partial
η2 = .5; IR: M = .17, SE = .05; PC: M = .06, SE = .1; IR vs.
PUC: F(1, 12) = 8.6, p < .05, partial η2 = .42; PUC: M =
−.03, SE = .05; IR vs. UC: F(1, 12) = 3.33, p = .09; UC: M =
.10, SE = .06]. Lastly, the analysis of the precuneus did not
reach significance, F(1.78, 21.33) = 2.06, p > .1.
Action ROIs
ANOVAs with repeated measures were conducted for
each of the five cytoarchitectonically and functionally
defined ROIs from the action localizer (left SFG, left
PCG, left pre-SMA, left IPL, and right IPL) to estimate the
sensitivity of these areas to IRs. Interrogation of the left
pre-SMA revealed a significant effect of Condition, F(2.21,
26.5) = 5.92, p < .01, partial η2 = .33. Planned compari-
sons indicated that the level of activation in the IR condi-
tion was higher than in the three control conditions [IR vs.
PC: F(1, 12) = 9.86, p < .01, partial η2 = .45; IR: M = .12,
SE = .05; PC: M = .02, SE = .05; IR vs. UC: F(1, 12) = 5.49,
p < .05, partial η2 = .31; UC: M = .01, SE = .06; IR vs. PUC:
F(1, 12) = 16.94, p < .005, partial η2 = .59; PUC: M = −.02,
SE = .05; Figure 2B]. The analysis of the left IPL yielded
a significant effect of Condition, F(2.69, 32.29) = 9.63,
p < .001, partial η2 = .45. Planned comparisons indicated
that the average percent signal change was higher in the
IR condition than in the control conditions [IR vs. PC: F(1,
12) = 14.65, p < .005, partial η2 = .55; IR: M = .16, SE = .04;
PC: .06, SE = .05; IR vs. UC: F(1, 12) = 15.18, p < .005, par-
tial η2 = .56; UC: M = .06, SE = .05; IR vs. PUC: F(1, 12) =
30.09, p < .001, partial η2 = .72; PUC: M = .03, SE = .04;
Figure 2B]. Also, there was a main effect of Condition in
the right IPL, F(2.68, 32.17) = 5.02, p < .01, partial η2 = .3.
2242
Journal of Cognitive Neuroscience
Volume 24, Number 11
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
1
2
1
4
/
2
1
2
1
3
/
7
2
1
2
9
3
4
7
4
/
7
1
8
6
7
7
o
8
c
6
n
3
_
8
a
/
_
j
0
o
0
c
2
n
7
4
_
a
p
_
d
0
0
b
2
y
7
g
4
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
f
/
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Table 3. Brain Regions from Whole-brain Analysis of the Implicature Task Showing Significantly More Activation during IRs
versus Controls (PC, UC and PUC; p < .0001, k > 12)
Region
Medial frontal cortex
Right ACC
Left ACC
Right ACC
Right middle frontal gyrus
Right pars opercularis
Left precentral gyrus
Right insular cortex
Right insula lobe
Pars triangularis
Right putamen
Left insular cortex
Left insula lobe
Pars orbitalis
Thalamus
Right thalamus
Left thalamus
Right middle temporal gyrus
Right posterior middle temporal gyrus
Posterior cingulate cortex
Posterior cingulate cortex
Left middle cingulate cortex
Cingulate gyrus
Left TPJ
Left supramarginal gyrus
Left angular gyrus
Right TPJ
Right supramarginal gyrus
Right superior temporal gyrus
Right supramarginal gyrus
Right precuneus
Right precuneus
Right precuneus
Left precuneus
Cluster Level Extent ( Voxels)
t
equivZ
x
y
z
Peak Voxel Level
MNI Coordinates
323
52
21
13
69
139
37
12
15
232
179
19
53
21
9.08
8.40
8.02
8.76
6.92
6.76
7.77
6.70
6.02
8.08
7.59
7.04
6.54
6.53
7.46
9.03
8.17
8.09
9.07
9.07
6.46
5.81
5.49
11.16
6.42
9.90
4.89
4.73
4.63
4.82
4.32
4.26
4.56
4.24
4.01
4.65
4.51
4.35
4.19
4.19
4.48
4.88
4.67
4.65
4.89
4.89
4.16
3.94
3.81
5.31
4.15
5.07
15
−3
6
21
45
−51
39
42
24
−36
−39
9
−6
51
45
−6
−12
6
−60
−48
63
63
51
6
18
−9
32
44
41
56
17
2
26
35
23
20
23
−7
−19
−49
−31
−37
−40
−34
−46
−49
−43
−52
−40
−67
−52
−64
25
19
19
28
13
46
−5
−2
−8
−5
−14
7
13
13
−5
22
37
25
28
31
25
22
28
37
37
37
van Ackeren et al.
2243
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
4
/
1
2
1
4
/
2
1
2
1
3
/
7
2
1
2
9
3
4
7
4
/
7
1
8
6
7
7
o
8
c
6
n
3
_
8
a
/
_
j
0
o
0
c
2
n
7
4
_
a
p
_
d
0
0
b
2
y
7
g
4
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
/
f
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 2. ROIs were
interrogated with respect to
the conditions IR, PC, UC,
and PUC. The image shows
all ROIs, superimposed on
a brain template. The bar
diagrams illustrate mean
percent signal change for
each condition. The error
bars depict the standard error.
(A) Green ROIs show regions
from the ToM localizer (mPFC
and TPJ). (B) Red ROIs refer
to regions that were activated
during action execution
(pre-SMA and bilateral IPL).
Planned comparisons showed that average percent sig-
nal change was higher in the IR condition than in the con-
trol conditions [IR vs. PC: F(1, 12) = 13.77, p < .005, partial
η2 = .53; IR: M = .06, SE = .05; PC: M = −.07, SE = .05; IR
vs. UC: F(1, 12) = 10.58, p < .01, partial η2 = .47; UC: M =
−.06, SE = .05; IR vs. PUC: F(1, 12) = 6.2, p < .05, partial
η2 = .34, PUC: M = −.03, SE = .05; Figure 2B]. However,
ANOVAs with repeated measures, investigating the effect
of condition in the left SFG and the left PCG did not reach
significance, F(2.9, 34.81) = 1.34, p > .1 and F(1.77, 21.2) =
.94, p > .1, respectively.
DISCUSSION
Previous research has demonstrated that language refer-
ring explicitly to actions (e.g., action verbs, nouns refer-
ring to tools, action sentences) reliably activates cortical
motor areas in the brain ( Willems et al., 2011; Van Dam
et al., 2010; Rüschemeyer et al., 2007; Tettamanti et al.,
2005; Hauk et al., 2004). In the current study, we asked
whether language that is used to refer implicitly to ac-
tions (i.e., without any explicit lexical reference to action)
shows a similar pattern of activation. In other words, can
cortical motor activation be triggered by expressions
without any lexical item that refers to an action? To this
end, participants were presented with spoken utterances,
some of which could be understood as IRs for actions
(e.g., “it is very hot here,” as a request that one open
the window) and some of which were simply descriptions
of visual scenes (e.g., “it is very hot here,” in the context
of a desert scene). The results indicate (1) that compre-
hension of IR sentences activates cortical motor areas
reliably more than comprehension of sentences devoid
of any implicit motor information. This is true despite
the fact that IR utterances contain no lexical reference to
action. (2) Comprehension of IR sentences also reliably acti-
vates substantial portions of the ToM network, known to be
involved in making inferences about mental states of others
(Frith & Frith, 2005, 2010; Gallagher & Frith, 2003; Saxe &
Kanwisher, 2003). The implications of these findings for
embodied theories of language are discussed below.
IRs and the Cortical Motor System
IR sentences activated areas within the larger cortical
motor system significantly more than sentences in any of
the three control conditions (Figure 2B). This activation
pattern was assessed in two ways: (1) in an ROI analysis
and (2) in a whole-brain analysis.
In the ROI analysis, voxels in BA 6 and bilateral IPL that
were also sensitive to finger movements during the local-
izer task (button presses) were identified as ROIs. These
regions comprised voxels in the left premotor cortex,
bilateral IPL, and pre-SMA. Interrogation of these ROIs
2244
Journal of Cognitive Neuroscience
Volume 24, Number 11
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
4
/
1
2
1
4
/
2
1
2
1
3
/
7
2
1
2
9
3
4
7
4
/
7
1
8
6
7
7
o
8
c
6
n
3
_
8
a
/
_
j
0
o
0
c
2
n
7
4
_
a
p
_
d
0
0
b
2
y
7
g
4
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
.
f
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
with regard to the four language conditions showed that
the bilateral IPL and pre-SMA were sensitive to the implicit
motor content in IR sentences. In other words, bilateral
IPL and pre-SMA showed significantly greater activation
for IR sentences than for sentences in any of the three
control conditions. In the following paragraphs, we dis-
cuss the potential role of the areas targeted by the ROI
analysis to processing IR sentences.
The inferior parietal lobe is a sensorimotor area that is
often associated with the representation of action goals
(Fogassi et al., 2005). Fogassi and colleagues (2005) ad-
dressed this hypothesis using single-cell recordings in
nonhuman primates. The authors found that a different
set of neurons fired when a monkey grasped food to
put it in a container than when it was going to eat the
food. In addition, some neurons showed the same pattern
during action observation. These results provide strong
evidence that the neurons in the IPL are sensitive to the
goal of an action. Recently, Aziz-Zadeh and Damasio
(2008) have argued that the IPL encodes the set of sen-
sorimotor events that coincide with action execution. For
example, a movement such as grasping a cup will elicit
somatosensory, proprioceptive, and visual feedback. The
signals from these three different sources are aligned in
time and therefore more likely to be associated in the
brain (Aziz-Zadeh & Damasio, 2008). The integration of
sensory information and action is particularly important
for the functional manipulation of tools. Numerous neuro-
psychological studies have associated lesions in the IPL
with a disability in manipulating objects and tools in a
meaningful way (apraxia; for a review, see Wheaton &
Hallett, 2007). The IPL is also consistently activated in
studies investigating the comprehension of action lan-
guage (Rueschemeyer, van Rooij, Lindemann, Willems, &
Bekkering, 2010). In a study using functional imaging,
Rueschemeyer, van Rooij, et al. (2010) found that the IPL
was more sensitive to words denoting functionally manip-
ulable objects (e.g., cup) as compared with volumetrically
manipulable words (e.g., bookend). Thus, the type of ac-
tion information that is instantiated in the IPL (i.e., infor-
mation about complex action plan and how to manipulate
objects) appears to be relevant both for executing actions
and for processing conceptual information about tools and
actions through language.
The pre-SMA is usually associated with executive as-
pects of motor control (Rushworth, Walton, Kennerley,
& Bannerman, 2004; Picard & Strick, 2001). Specifically,
Rushworth and colleagues (2004) have suggested that
the pre-SMA is involved in selecting and changing between
task-relevant action sets. That is, the selection of a specific
response from a set of possible responses to a sensory
stimulus. This idea is supported by the finding that chang-
ing an action set is perturbed if rTMS is applied to the
medial SFG (Willems et al., 2011). Although the pre-SMA is
not consistently activated in studies investigating the com-
prehension of action language, this is certainly not the
first time that this area has been observed (Rueschemeyer,
van Rooij, et al., 2010; Postle et al., 2008). For example,
Postle and colleagues investigated the sensitivity of the
cortical motor system (BA 6 and BA 4) to action verbs and
found that the pre-SMA in BA 6, but not M1, is sensitive
to action verbs. In the current study, we argue that once
a participant has understood that a request for action is
being made, he or she must evaluate what action is being
requested. The pre-SMA could potentially reflect the at-
tempt of the listener to select the best action alternative
from the set of possible actions one could perform in any
given situation.
In the whole-brain analysis, activation elicited by IR sen-
tences compared with the three control conditions was
assessed. The results demonstrate that IR sentences acti-
vated a fronto-parietal network, comprising the posterior
middle frontal gyrus, left precentral gyrus, and several re-
gions in the bilateral inferior parietal lobe, most notably
the supramarginal gyrus. These results are largely consis-
tent with the pattern observed in the ROI analysis and thus
provide converging evidence for the involvement of neural
motor areas in the processing of IR sentences compared
with sentences devoid of any motor content.
Our results indicate that language material devoid of
explicit action content activates neural motor areas in the
brain if presented in a situation in which reference to an
action is communicatively implied. This result marginalizes
the role of specific lexical items for cortical motor activation
in showing that action-related words are not necessary for
activating the cortical motor system during language com-
prehension. Yet, this does not mean that the motor system
is not involved in language processing. Rather, the present
findings suggest that motor areas might be involved in lan-
guage comprehension in a much more complex way than
theorists imagined just a decade ago (Pulvermüller, 1999,
2005). However, this idea is in line with more recent ap-
proaches, suggesting that the cortical motor system might
contribute to language understanding, but other sources
of information are also important (Kiefer & Pulvermüller,
2012; Meteyard, Rodriguez Cuardrado, Bahrami, & Vigliocco,
2012; Van Dam, van Dijk, Bekkering, & Rueschemeyer,
2011).
IRs and ToM
To understand the speaker meaning of an utterance, the
listener needs to infer the communicative intent of the
speaker (Holtgraves, 1994; Grice, Cole, & Morgan, 1975).
Recent neuroimaging studies have addressed the rela-
tionship between language and communication (Willems
et al., 2010; Sassa et al., 2007). Evidence from these stud-
ies suggests that the brain regions that are modulated by
the communicative demands of an utterance overlap with
classical ToM regions, but not language regions. For ex-
ample, Willems and colleagues (2010) asked subjects to
describe a word to another person. The authors manipu-
lated the communicative intent of the speaker by claiming
that the interlocutor either knew or did not know the
van Ackeren et al.
2245
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
4
/
1
2
1
4
/
2
1
2
1
3
/
7
2
1
2
9
3
4
7
4
/
7
1
8
6
7
7
o
8
c
6
n
3
_
8
a
/
_
j
0
o
0
c
2
n
7
4
_
a
p
_
d
0
0
b
2
y
7
g
4
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
t
/
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
target word. In addition, Willems et al. (2010) manipulated
the linguistic difficulty, by restricting the words a subject
was allowed to use in the description. Specifically, the
words had either a high or low semantic relation with
the target word. The results indicated that a region in the
mPFC was sensitive to the communicative demands of
the task while the linguistic demands were represented
in the left inferior frontal gyrus. Importantly, a region
in the pSTS was sensitive to the interaction between lin-
guistic and communicative demands of the task. A re-
lated study by Sassa and colleagues (2007) found that the
mPFC, the temporal poles, and the left TPJ were sensitive
to the communicative intention of the speaker. In sum-
mary, higher communicative task demands seem to cor-
relate with stronger activation in areas within the classical
ToM network.
In this study, IR sentences showed greater activation
than control sentences in a set of regions, known to be
involved in solving a specific ToM task, namely the false
belief task (see Apperly, 2012, for discussion on the cogni-
tive capacities underlying ToM). This was assessed using an
ROI approach. During an independent scan, participants
performed a classic ToM task (Dodell-Feder et al., 2010).
Specifically, voxels were identified, which showed greater
activation for interpreting false beliefs versus inaccurate
physical descriptions (for a more detailed description,
see Saxe & Kanwisher, 2003). As in previous studies, this
contrast elicited more activation in a set of regions in-
cluding the mPFC, the precuneus, and the bilateral TPJ.
Some of these were also sensitive to the comprehension
of IR sentences. In other words, IR sentences showed
more activation in ToM areas than any of the three control
conditions. This suggests that understanding IRs for action
requires a similar inference on the mental state of the
speaker as required for classical ToM tasks.
The results of this study show that the actual content of
the statement does not trigger activation in motor areas.
In the context of a picture of the desert, the sentence
“It is hot here” does not result in motor activation. It is
only in the case that this statement can be interpreted
as a request for action that activation of motor areas is
observed. It seems that the ToM network is needed to
interpret the statement as a request in the right context.
This suggests that the ToM system is involved in inferring
what the speaker intends to convey with the string of
words produced. In the case that the speaker intends
to convey a request for action, areas involved in motor
control get activated even when the utterance does
not contain any lexical item that refers to an action. It is
not easy to see how this could be accounted for in a
Hebbian cell assembly account (Pulvermüller, 1999, 2005),
which is based on associative connections between lexical
items and actions. However, more recent accounts have
acknowledged that different kinds of information might
contribute to language understanding (Meteyard et al.,
2012; Kiefer & Pulvermüller, 2012; Rueschemeyer,
van Rooij, et al., 2010). In our case, the motor involve-
ment seems to presuppose the compositional machinery
for decoding meaning and the inferential machinery for
deriving speaker meaning in the situational context.
Summary and Conclusion
The current study investigated whether utterances with no
explicit reference to an action activate the neural motor
system if an action is communicatively implied. Specifi-
cally, brain responses to sentence–picture combinations,
with IRs, were compared with control statements (PC,
UC, and PUC). The results indicated that some parts of
the cortical motor system were sensitive to both IR sen-
tences (pre-SMA and bilateral IPL) and action execution.
These findings extend previous research in showing that
language explicitly referring to actions is not a necessary
condition to elicit cortical motor activations. This suggests
that motor areas might be involved in language comprehen-
sion in a much more complex way than theorists imagined
just a decade ago (Pulvermüller, 1999, 2005). In addition,
areas that were involved in thinking about mental states of
others were also sensitive to IR sentences (mPFC, left TPJ).
Very likely, these regions are crucial for making an inference
about the communicative intent of the speaker.
Acknowledgments
This research was supported by the Nederlands Organisatie
voor Wetenschappelijk onderzoek Veni Grant awarded to Shirley-
Ann Rueschemeyer.
Reprint requests should be sent to Markus J. van Ackeren, Depart-
ment of Psychology, University of York, YO10 5DD, York, United
Kingdom, or via e-mail: M.vanAckeren@psych.york.ac.uk.
Notes
1. Greenhouse–Geisser correction was used to correct for viola-
tions of sphericity.
2. The collection of stimuli was provided by Rebecca Saxe and
Jessica Andrews-Hanna.
REFERENCES
Apperly, I. (2012). What is “theory of mind”? Concepts,
cognitive processes and individual differences. Quarterly
Journal of Experimental Psychology, 65, 825–839.
Aziz-Zadeh, L., & Damasio, A. (2008). Embodied semantics
for actions: Findings from functional brain imaging.
Journal of Physiology Paris, 102, 35–39.
Aziz-Zadeh, L., Wilson, S. M., Rizzolatti, G., & Iacoboni, M.
(2006). Congruent embodied representations for visually
presented actions and linguistic phrases describing
actions. Current Biology, 16, 1818–1823.
Barsalou, L. W. (1999). Perceptual symbol systems. Behavioral
Brain Science, 22, 560–577.
Barsalou, L. W. (2008). Grounded cognition. Annual Review
of Psychology, 59, 617–645.
Boulenger, V., Hauk, O., & Pulvermüller, F. (2011). Grasping
ideas with the motor system: Semantic somatotopy in
idiom comprehension. Cerebral Cortex, 19, 1905–1914.
2246
Journal of Cognitive Neuroscience
Volume 24, Number 11
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
4
/
1
2
1
4
/
2
1
2
1
3
/
7
2
1
2
9
3
4
7
4
/
7
1
8
6
7
7
o
8
c
6
n
3
_
8
a
/
_
j
0
o
0
c
2
n
7
4
_
a
p
_
d
0
0
b
2
y
7
g
4
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
/
t
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Brett, M., Anton, J., Valabregue, R., & Poline, J. (2002).
Region of interest analysis using an SPM toolbox [abstract].
8th International Conference on Functional Mapping of
the Human Brain, June 26, 2002, Sendai, Japan.
Caspers, S., Eickhoff, S. B., Geyer, S., Scheperjans, F., Mohlberg, H.,
Zilles, K., et al. (2008). The human inferior parietal lobule in
stereotaxic space. Brain Structure & Function, 212, 481–495.
Caspers, S., Geyer, S., Schleicher, A., Mohlberg, H., Amunts, K.,
& Zilles, K. (2006). The human inferior parietal lobule:
Cytoarchitectonic parcellation and interindividual
variability. Neuroimage, 33, 430–448.
Desai, R. H., Binder, J. R., Conant, L. L., Mano, Q. R., & Seidenberg,
M. S. (2011). The neural career of sensory-motor metaphors.
Journal of Cognitive Neuroscience, 23, 2376–2386.
Dodell-Feder, D., Koster-Hale, J., Bedny, M., & Saxe, R. (2010).
fMRI item analysis in a theory of mind task. Neuroimage,
55, 705–712.
Fischer, M. H., & Zwaan, R. A. (2008). Embodied language:
A review of the role of the motor system in language
comprehension. The Quarterly Journal of Experimental
Psychology, 61, 825–850.
Fogassi, L., Ferrari, P. F., Gesierich, B., Rozzi, S., Chersi, F., &
Rizzolatti, G. (2005). Parietal lobe: From action organization
to intention understanding. Science, 308, 662–667.
Frith, C., & Frith, U. (2005). Theory of mind. Current Biology,
15, 644–645.
Frith, U., & Frith, C. D. (2010). The social brain: Allowing humans
to boldly go where no other species has been. Philosophical
Transactions of the Royal Society, 365, 165–176.
Gallagher, H. L., & Frith, C. D. (2003). Functional imaging
of “theory of mind.” Trends in Cognitive Sciences, 7,
77–83.
Gallese, V., & Lakoff, G. (2005). The brainʼs conceptʼs: The
role of the sensory-motor system in conceptual knowledge.
Cognitive Neuropsychology, 21, 455–479.
Geyer, S. (2004). The microstructural border between the
motor and the cognitive domain in the human cerebral
cortex. Advances in Anatomy, Embryology and Cell
Biology, 174, 1–89.
Glenberg, A. M. (2000). Symbol grounding and meaning:
A comparison of high-dimensional and embodied theories
of meaning. Journal of Memory and Language, 43, 379–401.
Glenberg, A. M., & Gallese, V. (2011). Action-based language:
A theory of language acquisition, comprehension, and
production. Cortex, 48, 905–922.
Glenberg, A. M., & Kaschak, M. P. (2002). Grounding language
in action. Psychonomic Bulletin and Review, 9, 558–565.
Grice, H. P., Cole, P., & Morgan, J. (1975). Logic and
conversation. In P. Cole & J. L. Morgen (Eds.), Syntax
and semantics, Vol. 3, Speech acts (pp. 45–47). New York:
Academic Press.
Hagoort, P. (2005). On Broca, brain, and binding: A new
framework. Trends in Cognitive Sciences, 9, 416–423.
Hauk, O., Johnsrude, I., & Pulvermüller, F. (2004). Somatotopic
representation of action words in human motor and
premotor cortex. Neuron, 41, 301–307.
Holtgraves, T. M. (1994). Communication in context: Effects of
speaker status on the comprehension of indirect requests.
Journal of Experimental Psychology: Learning Memory
and Cognition, 20, 1205–1218.
Kiefer, M., & Pulvermüller, F. (2012). Conceptual representations
in mind and brain: Theoretical developments, current evidence
and future directions. Cortex, 48, 805–825.
Lakoff, G., & Johnson, M. (1999). Philosophy in the flesh:
The embodied mind and its challenge to western thought.
New York: Basic Books.
Mahon, B. Z., & Caramazza, A. (2008). A critical look at the
embodied cognition hypothesis and a new proposal for
grounding conceptual content. Journal of Physiology,
102, 59–70.
Meteyard, L., Rodriguez Cuardrado, S., Bahrami, B., &
Vigliocco, G. (2012). Coming of age: A review of
embodiment and the neuroscience of semantics.
Cortex, 48, 788–804.
Picard, N., & Strick, P. L. (2001). Imaging the premotor
areas. Current Opinion in Neurobiology, 11, 663–672.
Postle, N., McMahon, K., Ashton, R., & de Zubicaray, G. I. (2008).
Action word meaning representations in cytoarchitectonically
defined primary and premotor cortices. Neuroimage, 43,
634–644.
Pulvermüller, F. (1999). Words in the brainʼs language.
Behavioral and Brain Sciences, 22, 253–336.
Pulvermüller, F. (2005). Brain mechanisms linking language
and action. Nature Reviews Neuroscience, 6, 576–582.
Raposo, A., Moss, H. E., Stamatakis, E. A., & Tyler, L. K.
(2009). Modulation of motor and premotor cortices
by actions, action words and action sentences.
Neuropsychologia, 47, 388–396.
Rueschemeyer, S. A., van Rooij, D., Lindemann, O., Willems,
R. M., & Bekkering, H. (2010). The function of words:
Distinct neural correlates for words denoting differently
manipulable objects. Journal of Cognitive Neuroscience,
22, 1844–1851.
Rüschemeyer, S. A., Brass, M., & Friederici, A. D. (2007).
Comprehending prehending: Neural correlates for
processing verbs with motor stems. Journal of Cognitive
Neuroscience, 19, 855–865.
Rushworth, M. F. S., Walton, M. E., Kennerley, S. W., &
Bannerman, D. M. (2004). Action sets and decisions in
the medial frontal cortex. Trends in Cognitive Sciences,
8, 410–417.
Sassa, Y., Sugiura, M., Jeong, H., Horie, K., Sato, S., & Kawashima, R.
(2007). Cortical mechanisms of communicative speech
production. Neuroimage, 37, 985–992.
Saxe, R., & Kanwisher, N. (2003). People thinking about
thinking people—The role of the temporo-parietal
junction in “theory of mind.” Neuroimage, 19, 1835–1842.
Slotnick, S. D., Moo, L. R., Segal, J. B., & Hart, J. (2003).
Distinct prefrontal cortex activity associated with item
memory and source memory for visual shapes. Brain
Research, 17, 75–82.
Tettamanti, M., Buccino, G., Saccuman, M. C., Gallese, V.,
Danna, M., Scifo, P., et al. (2005). Listening to action-related
sentences activates fronto-parietal motor circuits. Journal
of Cognitive Neuroscience, 17, 273–281.
Van Dam, W., Rueschemeyer, S.-A., & Bekkering, H. (2010).
Action specificity reflected in embodied lexical-semantic
representations. Neuroimage, 53, 1318–1325.
Van Dam, W. O., van Dijk, M., Bekkering, H., & Rueschemeyer, S.-A.
(2011). Flexibility in embodied lexical-semantic representations.
Human Brain Mapping. doi: 10.1002/hbm.21365.
Wheaton, L. A., & Hallett, M. (2007). Ideomotor apraxia: A review.
Journal of the Neurological Sciences, 260, 1–10.
Willems, R. M., de Boer, M., de Ruiter, J. P., Noordzij, M. L.,
Hagoort, P., & Toni, I. (2010). A dissociation between
linguistic and communicative abilities in the human brain.
Psychological Science, 21, 8–14.
Willems, R. M., Ludovica, L., DʼEsposito, M., Ivry, R., &
Casasanto, D. (2011). A functional role for the motor
system in language understanding: Evidence from
theta-burst transcranial magnetic stimulation.
Psychological Science, 22, 849–854.
van Ackeren et al.
2247
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
4
/
1
2
1
4
/
2
1
2
1
3
/
7
2
1
2
9
3
4
7
4
/
7
1
8
6
7
7
o
8
c
6
n
3
_
8
a
/
_
j
0
o
0
c
2
n
7
4
_
a
p
_
d
0
0
b
2
y
7
g
4
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
.
/
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1