Overrecruitment in the Aging Brain as a Function
of Task Demands: Evidence for
a Compensatory View
Antonino Vallesi1,2, Anthony R. McIntosh2,3, and Donald T. Stuss2,3
Abstract
■ This study used fMRI to investigate the neural effects of in-
creasing cognitive demands in normal aging and their role for per-
formance. Simple and complex go/no-go tasks were used with two
versus eight colored letters as go stimuli, respectively. In both
tasks, no-go stimuli could produce high conflict (same letter, dif-
ferent color) or low conflict (colored numbers) with go stimuli.
Multivariate partial least square analysis of fMRI data showed that
older adults overengaged a cohesive pattern of fronto-parietal re-
gions with no-go stimuli under the specific combination of factors
which progressively amplified task demands: high conflict no-go
trials in the first phase of the complex task. This early neural over-
recruitment was positively correlated with a lower error rate in the
older group. Thus, the present data suggest that age-related extra-
recruitment of neural resources can be beneficial for performance
under taxing task conditions, such as when novel, weak, and com-
plex rules have to be acquired. ■
INTRODUCTION
Cognitive changes with aging are accompanied by modifi-
cations in brain functioning, as shown by neuroimag-
ing evidence (e.g., Grady, 2008; Park, Polk, Mikels, Taylor,
& Marshuetz, 2001). Although some studies report an under-
recruitment of brain regions with age (e.g., Rypma &
DʼEsposito, 2000), different patterns of age-related neural
overrecruitment have also been often described in the lit-
erature, especially under demanding task conditions.
Those patterns include activations in similar areas as those
engaged by young adults but with a greater magnitude of
activation, a more symmetric pattern of brain activity with
additional activation in homologous areas of the opposite
hemisphere in the older, or additional activation of com-
pletely different areas (Morcom, Li, & Rugg, 2007; Hedden
& Gabrieli, 2004; Cabeza, 2001, 2002; Logan, Sanders,
Snyder, Morris, & Buckner, 2002; Park et al., 2001).
Changes in the use of cognitive and neural resources
have been associated with aging in a number of different
contexts. An age-related cognitive decline often occurs
in tasks involving suppression of information interfering
with the present goals (Sweeney, Rosano, Berman, & Luna,
2001; Hasher, Zacks, & May, 1999). Mirroring this age-
specific inhibitory deficit in cognition (Hasher & Zacks,
1988), brain imaging and electrophysiological evidence
shows an increased neural activity (overrecruitment) asso-
ciated with processing of interfering information in older
adults (Gazzaley et al., 2008; Zysset, Schroeter, Neumann,
1SISSA (International School for Advanced Studies), Trieste, Italy,
2Rotman Research Institute at Baycrest, Toronto, Canada, 3Univer-
sity of Toronto, Canada
& Yves von Cramon, 2007; Gazzaley, Cooney, Rissman, &
DʼEsposito, 2005; Nielson, Langenecker, & Garavan, 2002;
but see Grady et al., 1995).
Moreover, age-related differences in the capacity to cir-
cumvent interference from distracting nontarget informa-
tion are amplified by increasing task complexity (Gazzaley,
Sheridan, Cooney, & DʼEsposito, 2007; Viskontas, Morrison,
Holyoak, Hummel, & Knowlton, 2004). Although common
factors such as generalized slowing have been proposed
to explain the disrupting effects of task complexity with
age (Salthouse, 1996), significant age effects remain after
adequately controlling for differences in processing speed
( Verhaeghen, Cerella, & Basak, 2006; Keys & White, 2000),
suggesting that a number of different mechanisms may
underlie an age-related decline in information processing.
Again, at the neural level, age-related overrecruitment of
brain regions often accompanies more complex and de-
manding task conditions, such as dual versus single tasks
(Smith et al., 2001), source versus item memory retrieval
(Morcom et al., 2007), or high versus low selection de-
mands in word generation tasks (Persson et al., 2004).
Poor performance under nonroutine contingencies
in aging (Craik & Byrd, 1982), such as during conflicting
and complex task conditions, suggests an age-related
decline in task setting, a hypothesized frontally based func-
tion thought to be required to establish weak stimulus–
response associations or rules (Alexander, Stuss, Shallice,
Picton, & Gillingham, 2005; Stuss, Shallice, Alexander, &
Picton, 1995), especially when those rules compete with
more prepotent ones ( Vallesi, McIntosh, Alexander, &
Stuss, 2009; Alexander, Stuss, Picton, Shallice, & Gillingham,
2007; Stuss & Alexander, 2007).
© 2010 Massachusetts Institute of Technology
Journal of Cognitive Neuroscience 23:4, pp. 801–815
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
–
p
r
d
t
i
2
c
3
l
4
e
–
8
p
0
d
1
f
1
/
9
2
4
3
0
/
9
4
7
/
0
8
o
0
c
1
n
/
1
2
0
7
1
7
0
5
2
2
1
1
3
4
9
/
0
j
o
p
c
d
n
.
b
2
y
0
g
1
u
0
e
.
s
t
2
o
1
n
4
9
0
0
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
f
t
.
.
/
.
o
n
1
8
M
a
y
2
0
2
1
As briefly reviewed above, neural overrecruitment with
aging has been associated with both suppression and task
complexity, although, to the best of our knowledge, no
study has combined the two factors in a single experimen-
tal design. To test whether a combination of those factors
amplifies neural overrecruitment in aging (i.e., when both
cognitive conflict and task complexity are high), a factorial
design crossing two complexity levels with two suppres-
sion levels was adopted. Specifically, two versions of a
go/no-go task were used. In the simpler version, the sub-
jects responded to red X and blue O (go stimuli) while with-
holding responses to the blue X and red O (high-conflict
no-go stimuli) and to numbers of either color (low-conflict
no-go stimuli). Go responses were prepotent for letters
because they were twice more frequent than no-go re-
sponses to letters (50% vs. 25%) and because of Stroop-
like effects (Stroop, 1935). A classical Stroop interference
effect arises when subjects are required to name the ink
color of incongruent color words (e.g., the word “BLUE”
written in red ink) because it is faster to read a word than
to name a color. In the present task, likewise, participants
were faster in identifying the letter than its ink color and
then in combining these two kinds of information in order
to make a no-go decision, when appropriate.
Thus, we assumed that there was a need to suppress a
prepotent go response in the presence of high-conflict
no-go letters, whereas suppression was less required for
low-conflict no-go stimuli, which were much faster to dis-
tinguish from go responses because they belonged to a
different category (numbers vs. letters; see Vallesi, Stuss,
McIntosh, & Picton, 2009 for event-related potential sup-
port to this assumption). In the more complex version, four
vowels and four consonants replaced Os and Xs. A predic-
tion was that neural overrecruitment would occur in older
adults when task demands were maximal, that is, for the
high-conflict no-go condition of the complex task.
Regarding specific predictions about the functional anat-
omy underlying the necessity to deal with increasing task
demands, we expected an activation of left lateral pre-
frontal cortex, as lesions in this region are associated with
an increase in false alarms to nontarget information in dif-
ferent domains and tasks (Alexander et al., 2007; Stuss &
Alexander, 2007; see also Bunge, 2004; Fletcher, Shallice,
& Dolan, 2000, for neuroimaging evidence). However, it
is likely that a more extensive fronto-parietal network will
be involved in acquiring weak and complex associations
not only between stimuli and responses (e.g., Dosenbach,
Fair, Cohen, Schlaggar, & Petersen, 2008) but also between
stimuli and a nonresponse (Vallesi, McIntosh, Alexander,
et al., 2009).
To partially anticipate the findings of the present study,
older adults did engage an extensive set of fronto-parietal
regions for the high-conflict no-go stimuli similarly to the
young adults but, as expected, overrecruited these regions
in the complex task, suggesting a relation between over-
recruitment and task complexity. There is some evidence
indicating that age-related deficits with complex tasks es-
pecially emerge in the early stages of task performance
( Willis & Nesselroade, 1990), and age differences are sub-
stantially reduced after practice (Kramer, Hahn, & Gopher,
1999). Thus, to further test how this network behaves in
aging when task demands are further taxed, we capitalized
on the assumption that task difficulty is high in novel sit-
uations and decreases with learning, further dividing the
trials of the complex task into a first and a second run.
The prediction was made that if overrecruitment is asso-
ciated with increasing task demands, it should mostly occur
in the first run.
When increased neural activity has been found with
aging, two opposite accounts have been proposed to ex-
plain it (Grady, 2008). The compensatory view posits that
neural overrecruitment reflects adaptive plasticity to im-
prove or maintain performance despite age-related neuro-
degenerative modifications (Gutchess et al., 2005; Buckner,
2004; Reuter-Lorenz, 2002; Cabeza et al., 1997; Grady et al.,
1994). An alternative view posits that this additional neural
recruitment reflects a loss of neural specificity and efficiency
with aging, especially when it is present despite worse per-
formance in the older than in the young adults (Rypma,
Eldreth, & Rebbechi, 2007; Zarahn, Rakitin, Abela, Flynn,
& Stern, 2007; Colcombe, Kramer, Erickson, & Scalf, 2005;
Park et al., 2001).
Our protocol provided the opportunity to test whether
any neural overrecruitment observed in the present study
reflects a compensatory or an inefficient use of neural re-
sources with increasing task difficulty. We correlated an
overall measure of the degree of recruitment of brain re-
gions sensitive to task demands with accuracy on the
high-conflict no-go condition of the complex task (where
age differences mostly occurred). The compensatory ac-
count would suggest that the older adults who showed
more neural overrecruitment in the first run would also
be expected to perform better on this run (positive corre-
lation). An opposite prediction can be made on the ineffi-
ciency account; that is, the older adults who overrecruited
these regions more on the first run would also be the
ones that show worse performance on this run (negative
correlation).
METHODS
Participants
Fourteen young (8 women; mean age = 27 years, range =
20–34 years) and 14 older (9 women; mean age = 70 years,
range = 60–80 years) volunteers took part in the study.
All the participants had normal or corrected-to-normal
vision. All were right-handed with an average score on
the Edinburgh Handedness Inventory (Oldfield, 1971)
of 87 and 89 for young and older, respectively. None of
the participants had any history of drug or alcohol abuse,
or history of psychiatric, neurological, or other medical ill-
ness, which might compromise cognitive function. None
reported memory or other cognitive problems noted by
802
Journal of Cognitive Neuroscience
Volume 23, Number 4
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
–
p
r
d
t
i
2
c
3
l
4
e
–
8
p
0
d
1
f
1
/
9
2
4
3
0
/
9
4
7
/
0
8
o
0
c
1
n
/
1
2
0
7
1
7
0
5
2
2
1
1
3
4
9
/
0
j
o
p
c
d
n
.
b
2
y
0
g
1
u
0
e
.
s
t
2
o
1
n
4
9
0
0
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
.
t
f
.
/
.
o
n
1
8
M
a
y
2
0
2
1
either themselves or their relatives and friends. The two
groups were also matched in their education level (17 and
16 years, for young and older, respectively). Given this
high level of education, the results of the present study
cannot be generalized to the whole aging population.
Mini Mental State Examination (Folstein, Folstein, &
McHugh, 1975) was administered to older participants
in order to screen them for dementia (score range: 28–
30/30). Each participant signed an informed consent that
was previously approved by the local Ethics Research
Board, and received a compensation of $50. Part of the
data of the young group (simple task) has already been
reported elsewhere ( Vallesi, McIntosh, Alexander, et al.,
2009).
Experimental Material and Design
Stimuli were presented centrally against a gray background.
Participants viewed the stimuli through a mirror placed on
the head coil. Go/no-go stimuli were letters and numbers
written in Times New Roman font and colored in blue or
red (50% each). In the simple task, go stimuli were “red
O” and “blue X,” and no-go stimuli were “blue O” and
“red X” (high-conflict no-go) on the one side, and red and
blue numbers 2 and 3 (low-conflict no-go) on the other
side. In the complex task, go stimuli were red vowels (A,
E, I, U) and blue consonants (L, N, P and Z), and no-go
stimuli were blue vowels and red consonants (high-conflict
no-go), or red and blue numbers 4, 5, 6, and 7 (low-conflict
no-go). In both tasks, the association between color and
go/no-go letters was counterbalanced across subjects. Par-
ticipants were briefly familiarized with the task and stimuli
before entering the scanner room to ensure that they un-
derstood the instructions and to reduce anxiety.
Each trial began with a go or no-go stimulus lasting
300 msec. The deadline for the go response was 2 sec after
stimulus onset. A blank screen followed the stimulus pre-
sentation. Interstimulus interval varied randomly and con-
tinuously between 2.2 and 4.2 sec. This manipulation was
important to sample the whole hemodynamic response
function. Participants performed two consecutive runs for
each task. Each run had 64 go (50%), 32 high-conflict no-go
(25%), and 32 low-conflict no-go (25%) stimuli. Although
there was an equal number of go and no-go trials, when
one only considers the conflicting go/no-go stimuli belong-
ing to the same category (letters), no-go letters were half
as frequent as the go letters, a typical manipulation in the
go/no-go literature meant to produce prepotent go re-
sponding (e.g., Hester, Murphy, & Garavan, 2004; Nielson
et al., 2002; Rubia et al., 2001).
The total number of test trials was 512. Participants
were instructed to press a button with the index finger of
their dominant hand as soon as they saw a go stimulus,
and refrain from responding when a no-go stimulus ap-
peared. Thus, the experiment consisted of a 2 Task (sim-
ple vs. complex) × 3 Condition ( go, high-conflict and
low-conflict no-go) × 2 Run (first and second runs)
design. Six familiarization trials preceded each run. Dur-
ing the presentation of these initial trials only, participants
received visual feedback about their performance. The or-
der of presentation of the two tasks was counterbalanced
across participants.
Analysis of the Behavioral Data
The familiarization trials and the first test trial were dis-
carded from the analyses. Moreover, the low-conflict no-go
condition was discarded from subsequent behavioral anal-
yses because performance on those conditions was at ceil-
ing in both groups and tasks (see Figure 1). Accuracy data
on the go and high-conflict no-go stimuli were analyzed
separately by means of a 2 × 2 × 2 mixed ANOVA with
task (simple vs. complex) and run (first vs. second) as the
within-subject factors, and age (young vs. older) as the
between-subjects factor.
RT analysis was limited to the go conditions only because
not enough commission errors occurred for the no-go con-
ditions. A 2 × 2 × 2 mixed ANOVA was carried out on the go
RTs, with task (simple vs. complex) and run (first vs. sec-
ond) as the within-subject factors, and age (young vs. older)
as the between-subjects factor.
Image Acquisition and Data Preprocessing
Images were acquired on a 3-T Siemens Magnetom Trio
scanner with a matrix 12-channel head coil. Functional vol-
umes were obtained using a whole-head T2*-weighted EPI
sequence (repetition time [TR] = 2 sec, echo time [TE] =
30 msec, flip angle [FA] = 70°, 28 oblique axial slices with
interleaved acquisition, 3.1 × 3.1 × 5 mm voxel resolution,
field of view [FOV] = 20 cm, acquisition matrix [AM] =
64 × 64). The first 5 volumes were discarded to allow
the magnetization to reach steady state. Physiological data
(heart and respiration rate) were acquired during the scan-
ning session. Anatomical images were acquired using an
MP-RAGE sequence (TR = 2 sec, TE = 2.63 sec, 160 oblique
axial slices, with a 1-mm3 voxel size, FOV = 25.6 cm, AM =
256 × 256), either before or after the functional images
(counterbalanced across subjects). Stimuli were presented
visually through a mirror mounted on the coil that reflected
images from a projector located at the bottom of the scan-
ner. Finger-press responses were recorded with an MRI-
compatible response pad. T2 Fluid Attenuated Inversion
Recovery sequences (TR = 9 sec, TE = 96 msec, FA =
165°, FOV = 2.2 cm, AM = 256 × 162) were acquired at
the beginning of the MR session to assess the presence of
white matter disease, especially in the older adults. No par-
ticipant was found with white matter hyperintensities out-
side the normal range.
Part of the preprocessing was performed with AFNI soft-
ware (http://afni.nimh.nih.gov/; Cox, 1996). EPI time-series
data were corrected for cardiac and respiratory parameters
(program 3dretroicor) and for difference in the timing of
slice acquisition (program 3dTshift). Six-parameter rigid
Vallesi, McIntosh, and Stuss
803
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
–
p
r
d
t
i
2
c
3
l
4
e
–
8
p
0
d
1
f
1
/
9
2
4
3
0
/
9
4
7
/
0
8
o
0
c
1
n
/
1
2
0
7
1
7
0
5
2
2
1
1
3
4
9
/
0
j
o
p
c
d
n
.
b
2
y
0
g
1
u
0
e
.
s
t
2
o
1
n
4
9
0
0
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
.
.
.
f
/
t
o
n
1
8
M
a
y
2
0
2
1
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
–
p
r
d
t
i
2
c
3
l
4
e
–
8
p
0
d
1
f
1
/
9
2
4
3
0
/
9
4
7
/
0
8
o
0
c
1
n
/
1
2
0
7
1
7
0
5
2
2
1
1
3
4
9
/
0
j
o
p
c
d
n
.
b
2
y
0
g
1
u
0
e
.
s
t
2
o
1
n
4
9
0
0
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
.
/
.
.
f
t
o
n
1
8
M
a
y
2
0
2
1
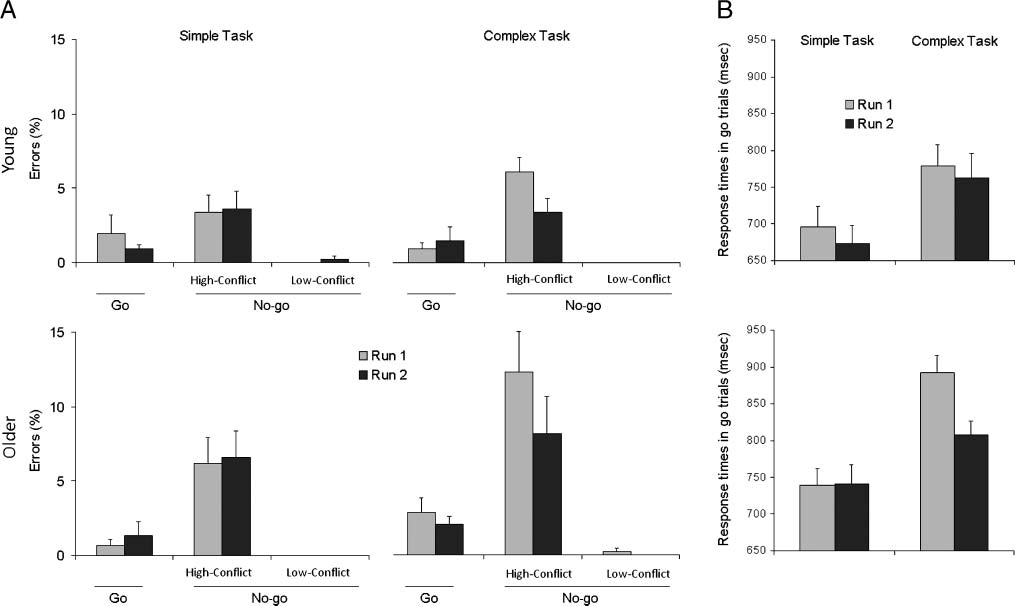
Figure 1. Behavioral data. (A) Percentage of errors (and standard errors of the mean) as a function of task, run, go/no-go conditions, and age.
(B) Mean RTs (and standard errors of the mean) of correct responses (in milliseconds) to go stimuli as a function of task, run, and age.
body inter- and intra-run motion correction was then
performed by coregistering volumes to a reference EPI
volume (AFNI program 3dvolreg). Coregistration to a
functional MNI template (EPI.nii) and spatial smoothing
(8-mm Gaussian kernel) was performed in SPM5 (Friston
et al., 1995). Group analyses were performed using Partial
Least Square software (PLS; McIntosh, Bookstein, Haxby,
& Grady, 1996). The anatomical scan was first coregistered
to the functional run that occurred closest in time in AFNI
during reconstruction (program siemenstoafni-beta2), and
then coregistered to a structural MNI template (T1.nii)
in SPM5.
Partial Least Square
We describe the conceptual details of the PLS analysis in
the following paragraphs (but see McIntosh et al., 1996,
for a more detailed explanation). PLS is a set of multivariate
statistical analyses that assesses the relations between
any set of independent measures, such as the experimental
design, and a set of dependent measures, in our case, the
whole brain hemodynamic response function (HRF). PLS
carries out the computation of the optimal partial least
squares fit to cross-block correlation between the indepen-
dent and dependent measures.
We used task PLS, an analysis that identifies patterns of
brain voxels whose signal change covaries with the experi-
mental conditions. A trial was defined as a signal segment,
beginning at the stimulus onset and lasting 7 lags, which
represents the response of each voxel averaged across
trials. Each lag corresponds to a TR (2 sec). The HRF for
each trial was expressed as the intensity difference from
trial onset, allowing investigation of changes in task-related
activity at different lags along the whole temporal segment.
No assumption was made about the shape of HRF. All task
conditions were included in this analysis. Condition aver-
ages for each voxel and lag were expressed as a deviation
from the grand mean of all conditions by lags. The devia-
tion matrix undergoes singular value decomposition to
extract a set of latent variables (LVs), which are defined
as orthogonal pairs of singular vectors. These vector pairs
reflect a symmetric relation between a cohesive pattern
of brain activity (singular image) and a particular experi-
mental effect represented as derived optimal contrast,
or design scores. The numerical weights within the sin-
gular image are called saliences and can be positive or
negative. The singular value for an LV is the covariance
between the brain and the design scores. Each LV explains
a progressively smaller percentage of the total covariance
pattern, until all the covariance has been explained. For
each LV, PLS yields also brain scores that indicate how
strongly individual subjects express the patterns on the
LV. These scores are calculated by multiplying the raw
images by the singular image on a particular LV for each
804
Journal of Cognitive Neuroscience
Volume 23, Number 4
subject. Finally, when brain scores are computed for each
time lag, a temporal brain score is obtained.
A permutation test is used to compute the overall signifi-
cance for each LV (McIntosh et al., 1996). The data matrix
rows are randomly reordered and a new set of LVs is calcu-
lated for each permutation. For the current experiment,
1000 permutations were used. If the singular value of each
new LV exceeded the original value more than 99% of the
times ( p = .01), an LV as a whole was considered signifi-
cant. To determine the reliability of the saliences identified
by the LVs, all data were submitted to a bootstrap estima-
tion of the standard errors by randomly resampling sub-
jects with replacement 200 times. PLS is recalculated for
each bootstrap sample to identify those saliences whose
value remains stable regardless of the sample chosen.
The ratio of the salience to the bootstrap standard error is
approximately equivalent to a Z score (Efron & Tibshirani,
1986).
Clusters with at least 15 contiguous voxels with a salience-
to-standard error ratio (bootstrap ratio, BSR) bigger than
5 (approximately corresponding to p < .00001) in each
lag were considered as reliable. Coordinates of the voxel
with the peak BSR within each cluster were obtained
in MNI space and converted into Talairach coordinates
to find the likely gyral locations using Matthew Brettʼs
transformation (www.mrccbu.cam.ac.uk/ Umaging /
mnispace.html). Approximate Brodmannʼs areas were
then identified using the Talairach Daemon (Lancaster
et al., 2000).
A preliminary task PLS analysis including all the six
conditions (3 go/no-go conditions × 2 tasks) and groups
(younger and older) showed a complex pattern of results,
but no difference in design scores for go stimuli in the two
age groups and tasks was observed, as can be appreciated
in Supplementary Figure S1. This pattern replicates pre-
vious results showing that most of the age-related fMRI dif-
ferences in go/no-go tasks occur in no-go trials (Nielson
et al., 2002). To focus on the most sensitive conditions
(high- vs. low-conflict no-go) and also to avoid confounds
derived from the differences between the go condition and
the other two conditions (i.e., double frequency of occur-
rence, requirement of a motor response), we conducted
a task PLS analysis (reported here) that did not include
go conditions. This analysis showed a selective overrecruit-
ment of brain regions in the older group during the com-
plex task (see Results). Two subsequent PLS analyses were
run to understand if the overrecruitment of brain regions
involved in the processing of high-conflict no-go items
in the complex task was modulated as a function of learn-
ing from Run 1 to Run 2 in the older group. A first analysis
included both tasks (simple and complex). Because this
analysis showed that critical age-related differences in
practice effects occurred exclusively in the complex task,
a second analysis focused on the complex task only (fully
reported here).
As reported below, this analysis demonstrated an over-
recruitment of an extensive set of brain regions in the older
group with high-conflict no-go trials on the first run of
the complex task. In PLS, the brain scores are an index
of how strongly each individual contributed to a given
LV. Hence, for both groups, the brain scores in this con-
dition were correlated with the accuracy data on the same
condition using a Pearson correlation analysis to get a
hint on whether overrecruitment was actually benefi-
cial or detrimental to the initial performance in the older
group. The brain scores for the high-conflict no-go con-
dition in the first run of the complex task were also cor-
related to the percentage of accuracy improvement in
the second run (i.e., percent differences with respect to
accuracy in the first run), in order to detect the nature of
the relation between initial overrecruitment and later
performance.
RESULTS
Accuracy and RT data are presented in Figure 1.
Accuracy
Older participants tended to make significantly more
commission errors (i.e., go responses) than young ones on
the high-conflict no-go trials [age main effect: F(1, 26) =
4.1, p = .053]. These types of errors were more frequent
for both groups in the first run than in the second one
[run main effect: F(1, 26) = 7.9, p < .01], and in the complex
task than in the simple one [task main effect: F(1, 26) =
5.5, p < .05]. A significant two-way interaction indicated
that commission errors to high-conflict no-go stimuli were
especially frequent in the first run of the complex task
[Run × Task interaction: F(1, 26) = 8.6, p < .01]. The
ANOVA concerning accuracy on go stimuli did not reveal
any significant effect.
Response Times
Older subjects tended to be slower than young ones [age
main effect: F(1, 26) = 4, p = .056]. RTs were longer in
the complex than in the simple task [task main effect: F(1,
26) = 121.8, p < .001], and in the first than in the second
run [run main effect: F(1, 26) = 17.8, p < .001]. RTs were
much longer in the first run of the complex task than in
the second one, whereas the difference between the two
runs was much reduced in the simple task [Run × Task
interaction: F(1, 26) = 5.9, p < .05]. This pattern was par-
ticularly pronounced in the older group [Age × Run ×
Task interaction: F(1, 26) = 8.9, p < .01]. When the raw
data for each subject were transformed to percent change
scores (i.e., mean RT in each condition divided by the
overall mean RT and then multiplied by 100), this critical
three-way interaction was still significant [F(1, 26) = 6.9,
p = .01], thus showing that it was not an artifact of gen-
eral slowing.
Vallesi, McIntosh, and Stuss
805
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
3
l
4
e
-
8
p
0
d
1
f
1
/
9
2
4
3
0
/
9
4
7
/
0
8
o
0
c
1
n
/
1
2
0
7
1
7
0
5
2
2
1
1
3
4
9
/
0
j
o
p
c
d
n
.
b
2
y
0
g
1
u
0
e
.
s
t
2
o
1
n
4
9
0
0
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
.
.
f
/
t
.
o
n
1
8
M
a
y
2
0
2
1
fMRI Data
PLS Results: High-conflict and Low-conflict No-go
This analysis yielded one significant LV only (observed
singular value = 29.6, explained cross-block covariance =
44.4%, p < .001). The design scores for this LV are shown
in Figure 2A.
This LV showed a contrast between high-conflict and
low-conflict no-go in both tasks. Critically, it also showed
a No-go condition × Task complexity × Age interaction.
Older participants had greater (opposite) design scores
for conflict and low-conflict no-go in the complex task
than in the simple task, and than young participants in
both tasks. The individual subjectsʼ contribution to an LV
is estimated with an overall measure called brain score
(see Methods). The brain scores for each lag (i.e., temporal
brain scores) were submitted to a mixed ANOVA with
no-go condition (high-conflict vs. low-conflict) and task
(simple vs. complex) as the within-subject factors, and
age (young vs. older) as the between-subjects factor. The
No-go condition × Task × group three-way interaction
showed a strong tendency in lag 3 [F(1, 26) = 3.9, p =
.059] and was significant in lag 4 [F(1, 26) = 4.4, p < .05],
whereas it was far from significance in the other lags (see
Figure 2B). Therefore, clusters with reliable saliences for
lags 3 and 4 are listed in Table 1 and shown in Figure 2C.
Clusters that showed greater activation for high-conflict
than for low-conflict no-go conditions, especially in the older
group and in the complex task, included bilaterally the
inferior and middle frontal gyrus, inferior parietal lobule,
posterior cerebellum; on the left, the insula, precentral gy-
rus, fusiform gyrus, putamen; and on the right, the anterior
Figure 2. Results of the first
PLS analysis. (A) Design scores
(arbitrary units) for the Latent
Variable 1, according to age,
task, and no-go condition. (B)
Temporal brain scores (arbitrary
units and sign) indicating how
the brain network in (C)
generally responded to the
task conditions. The symbols
* and ** indicate a trend and a
significant Age × Task × No-go
condition interaction in lags 3
and 4, respectively. (C) Brain
clusters (number of voxels ≥ 15,
bootstrap ratio ≥ 5), where
design and temporal scores
shown in Panels A and B were
mainly expressed. Time from
stimulus onset is indicated on
the y-axis of the singular image
and is expressed in lags (1 lag =
2 sec repetition time). The
x-axis shows the z-coordinate
of the axial slice in MNI space.
Cold colors indicate clusters
with negative bootstrap ratios,
which were differentially more
activated for experimental
conditions with negative design
scores in Panel A and negative
temporal scores in Panel B
(i.e., high-conflict no-go in
both tasks and groups). Warm
colors indicate clusters with
positive bootstrap ratios,
which were differentially more
activated for experimental
conditions with positive design
scores in Panel A and positive
temporal scores in Panel B (i.e.,
irrelevant no-go in both tasks
and groups). The bootstrap
ratio map is superimposed on
the average anatomical scans
from all 28 participants.
806
Journal of Cognitive Neuroscience
Volume 23, Number 4
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
3
l
4
e
-
8
p
0
d
1
f
1
/
9
2
4
3
0
/
9
4
7
/
0
8
o
0
c
1
n
/
1
2
0
7
1
7
0
5
2
2
1
1
3
4
9
/
0
j
o
p
c
d
n
.
b
2
y
0
g
1
u
0
e
.
s
t
2
o
1
n
4
9
0
0
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
/
.
f
.
.
t
o
n
1
8
M
a
y
2
0
2
1
Table 1. Reliable Clusters Identified for LV1 in the First Task-PLS Analysis Including Both Age Groups (Young vs. Older), Tasks
(Simple vs. Complex), and No-go Conditions (Conflict vs. Irrelevant No-go)
Negative Saliences
Lag
Cluster Region
3
3
3
3
3
3
3
3
3
4
4
4
4
4
4
4
4
4
4
4
4
R Claustrum
L Inferior Parietal Lobule
L Insula
L Precentral Gyrus
R Precuneus
R Middle Frontal Gyrus
R Cingulate Gyrus
L Fusiform Gyrus
R Inferior Frontal Gyrus
L Inferior Parietal Lobule
R Inferior Parietal Lobule
R Uvula
L Middle Frontal Gyrus
L Inferior Semilunar Lobule
R Middle Frontal Gyrus
R Cingulate Gyrus
R Inferior Temporal Gyrus
L Fusiform Gyrus
R Inferior Frontal Gyrus
L Putamen (Lentiform Nucleus)
L Fusiform Gyrus
Lag
4
Cluster Region
Bilateral Medial Frontal Gyrus
BA
–
40
13
6
19
46
32
37
9
40
40
–
46
–
46
32
20
37
47
–
37
BA
11
Talairach
y
23
−33
19
1.7
−64
36
21
−59
9
−52
−44
−75
36
−72
36
25
−47
−59
27
3.9
−44
z
−1.2
42
−4.3
33
36
20
39
−11
25
43
43
−33
20
−37
20
39
−14
−14
−8.1
−0.2
−18
x
28
−48
−28
−40
32
48
8
−48
40
−32
32
16
−44
−32
51
8
63
−51
32
−24
−40
Size
68
273
60
169
194
39
119
31
23
350
297
286
127
104
96
38
30
43
34
25
28
BSR
−10
−9.9
−9.9
−8.9
−8.8
−8.2
−7.3
−7.1
−6.1
−10
−9.7
−9.4
−9.2
−8.4
−8.1
−7.9
−7.7
−7.4
−7.2
−6.7
−6.6
Positive Salience
Talairach
y
38
x
0
z
−19
Size
63
Bootstrap
8.4
R = right hemisphere; L = left hemisphere; BA = approximate Brodmannʼs area; BSR = bootstrap ratio in the PLS analysis.
cingulate, inferior temporal gyrus and precuneus. The only
cluster that showed more activation for the opposite con-
trast, that is, low-conflict versus high-conflict no-go, mainly
in the complex task and in the older group, was located on
the rostral medial prefrontal region (Brodmannʼs area 11).
PLS Results: Practice Effects
A first analysis of practice effects from Run 1 to Run 2 in-
cluded both tasks. As the design scores suggest (Fig-
ure 3A), LV1 (observed singular value = 41.2, explained
cross-block covariance = 25%, p < .0001) expressed a
complex interaction between task complexity, run, condi-
tion, and age. In the simple task, design scores for high-
conflict and low-conflict no-go diminished from Run 1 to
Run 2, without substantial difference between the two
age groups. On the other hand, marked age differences
emerged in the complex task. To better understand this
interaction, and to focus on the conditions in which learn-
ing occurred, in terms of accuracy in both groups and in
terms of speed especially in the older adults, we focused
Vallesi, McIntosh, and Stuss
807
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
3
l
4
e
-
8
p
0
d
1
f
1
/
9
2
4
3
0
/
9
4
7
/
0
8
o
0
c
1
n
/
1
2
0
7
1
7
0
5
2
2
1
1
3
4
9
/
0
j
o
p
c
d
n
.
b
2
y
0
g
1
u
0
e
.
s
t
2
o
1
n
4
9
0
0
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
f
.
.
.
t
/
o
n
1
8
M
a
y
2
0
2
1
Figure 3. Results of the PLS
analyses on practice effects
between Run 1 and Run 2.
(A) Design scores for the Latent
Variable 1 of the PLS analysis
including task, no-go condition,
run, and age. (B) Design scores
for the Latent Variable 1 of
the PLS analysis focusing on
practice effects in the complex
task only. Experimental
conditions included no-go
condition, run, and age. (C)
Temporal brain scores for the
Latent Variable 1 of the PLS
analysis focusing on practice
effects in the complex task
only. The symbol * indicates
significant Age × No-go
condition × Run interaction
in lags 2, 3, and 4. (D) Brain
clusters (number of voxels ≥ 15,
bootstrap ratio ≥ 5), where
design and temporal scores
shown in B and C were mainly
expressed. Time from stimulus
onset is indicated on the y-axis
of the singular image and is
expressed in lags (1 lag = 2 sec
repetition time). The x-axis
shows the z-coordinate of the
axial slice in MNI space. Warm
colors indicate clusters with
positive bootstrap ratios,
which were differentially more
activated for experimental
conditions with positive (vs.
negative) design scores in
Panel B and temporal scores
in Panel C (i.e., high-conflict
no-go). The bootstrap ratio
map is superimposed on the
average anatomical scans
from all 28 participants.
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
3
l
4
e
-
8
p
0
d
1
f
1
/
9
2
4
3
0
/
9
4
7
/
0
8
o
0
c
1
n
/
1
2
0
7
1
7
0
5
2
2
1
1
3
4
9
/
0
j
o
p
c
d
n
.
b
2
y
0
g
1
u
0
e
.
s
t
2
o
1
n
4
9
0
0
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
.
.
/
.
f
t
o
n
1
8
M
a
y
2
0
2
1
808
Journal of Cognitive Neuroscience
Volume 23, Number 4
on the complex task only in a subsequent task-PLS analysis
that we report below. This analysis included the following
conditions: age (young vs. older), run (first vs. second),
and no-go condition (high-conflict vs. low-conflict no-go).
The first LV only was significant (observed singular
value = 38.9, LV1 explained cross-block covariance =
42.4%, p < .0001). The design scores are reported in Fig-
ure 3B. Positive saliences indicate brain voxels differentially
more activated in processing high-conflict no-go stimuli
and negative saliences indicate voxels more activated in
processing low-conflict no-go stimuli. Moreover, as the de-
sign scores show, the overrecruitment of brain regions
present in the first run of the complex task in older sub-
jects diminished in the second run. On the other hand,
young subjects used the same network as older subjects
in the second run only. Hence, age-related differences
were observed in the learning phase of the complex task
(Run 1), and disappeared once the task has been prac-
ticed for one run.
We also examined the brain scores in each of the 7 lags
(temporal brain scores; Figure 3C), by means of mixed
ANOVAs with no-go condition (high-conflict vs. low-conflict
no-go) and run (first vs. second) as the within-subject fac-
tors, and age (young vs. older) as the between-subjects
factor. The effect of interest of these ANOVAs would be a
three-way interaction between no-go condition, run, and
age. This interaction was significant for lags 2–4 [F(1, 26)
range = 4.8–8.2, all ps < .05], and showed a weak trend
for lag 5 ( p = .1). Clusters with reliable saliences in lags
2–4 are reported in Table 2 and are shown in Figure 3D.
Table 2. Reliable Clusters Identified for LV1 in the Task-PLS Analysis Concerning Practice Effects in the Complex Task
Lag
Cluster Region
Positive Saliences
2
3
3
3
3
3
3
3
3
3
4
4
4
4
4
4
4
4
4
4
4
4
4
R Inferior Parietal Lobule
R Inferior Parietal Lobule
L Insula
L Inferior Parietal Lobule
R Middle Frontal Gyrus
R Inferior Frontal Gyrus
L Inferior Frontal Gyrus
L Middle Frontal Gyrus
Medial Frontal Gyrus
L Cuneus
R Inferior Parietal Lobule
R Inferior Frontal Gyrus
L Middle Frontal Gyrus
L Inferior Parietal Lobule
L Inferior Frontal Gyrus
R Medial Frontal Gyrus
L Fusiform Gyrus
L Insula
L Culmen
R Inferior Frontal Gyrus
R Inferior Temporal Gyrus
L Inferior Semilunar Lobule
R Middle Frontal Gyrus
BA
40
40
13/45
40
46
47
9
46
6
17
40
46
46
40
9
8
20
13
–
47
20
–
9
Talairach
x
y
z
Size
Bootstrap
36
48
−28
−32
48
32
−44
−44
0
−16
36
44
−44
−32
−44
8
−40
−32
−24
32
51
−44
36
−48
−41
23
−52
36
27
9
36
14
−77
−45
39
36
−52
9
18
−40
23
−63
27
−47
−68
9
43
43
3
43
20
−5
29
17
44
8
39
13
17
43
29
43
−18
−1
−24
−8
−14
−40
33
23
174
50
214
38
51
50
59
59
17
340
79
39
304
42
53
23
19
208
36
19
18
31
6.3
9.4
9.0
8.5
8.0
7.7
7.2
7.0
6.9
6.1
10.4
9.5
8.9
8.9
8.0
7.7
7.5
7.3
7.1
6.5
6.4
6.3
6.1
This analysis included the following experimental conditions: age group (young vs. older), run (first vs. second), and no-go condition (conflict vs.
irrelevant no-go). R = right hemisphere; L = left hemisphere; BA = approximate Brodmannʼs area; BSR = bootstrap ratio in the PLS analysis.
Vallesi, McIntosh, and Stuss
809
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
3
l
4
e
-
8
p
0
d
1
f
1
/
9
2
4
3
0
/
9
4
7
/
0
8
o
0
c
1
n
/
1
2
0
7
1
7
0
5
2
2
1
1
3
4
9
/
0
j
o
p
c
d
n
.
b
2
y
0
g
1
u
0
e
.
s
t
2
o
1
n
4
9
0
0
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
.
.
.
t
f
/
o
n
1
8
M
a
y
2
0
2
1
Figure 4. Correlation patterns
between the individual brain
scores for the high-conflict
no-go condition in the first
run of the complex task (as
obtained in the last partial
least square analysis) and
percentage of correct trials
in the same task condition
for both the young and
older groups.
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
3
l
4
e
-
8
p
0
d
1
f
1
/
9
2
4
3
0
/
9
4
7
/
0
8
o
0
c
1
n
/
1
2
0
7
1
7
0
5
2
2
1
1
3
4
9
/
0
j
o
p
c
d
n
.
b
2
y
0
g
1
u
0
e
.
s
t
2
o
1
n
4
9
0
0
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
.
t
.
/
.
f
o
n
1
8
M
a
y
2
0
2
1
Clusters that showed greater activation for high-conflict
no-go than for low-conflict no-go conditions (positive sa-
liences), especially in the older adults and in the first run
of the complex task, included bilaterally, the inferior and
middle frontal gyrus, inferior parietal lobule, and posterior
cerebellum; on the left, the insula, fusiform gyrus, cuneus,
and cerebellum; and on the right, the superior medial fron-
tal gyrus. No cluster with negative saliences (i.e., low-conflict
no-go > high-conflict no-go) survived the chosen threshold.
In the older group only, there was a positive correlation
between brain scores and accuracy in the high-conflict
no-go condition during the first run of the complex task
(r = .69, p = .007; see Figure 4), suggesting that over-
recruitment of these regions was actually beneficial to
the older individualsʼ performance. There was a trend for
a negative correlation between brain scores in this condi-
tion and the percentage of accuracy improvement in the
same high-conflict no-go condition during the second
run (r = −.51, p = .06). This trend suggests that those
older subjects that actually overrecruited the network most
for this condition on the first run did not improve much on
the second run, probably because they were already per-
forming at their best on the first run. On the other hand,
young subjects did not show any significant correlation
between brain scores in the high-conflict no-go condition
of the first run and performance on this condition during
the first run or improvement during the second run (for
both, p > .37), although ceiling effects may play a role in
this case. Finally, the brain scores for the high-conflict no-go
condition in the second run of the complex task did not
correlate significantly with performance in the second run
in either age group.
DISCUSSION
The present study investigated age-related differences in the
recruitment of brain regions as a function of progressively
increasing task demands. To this aim, three features were
systematically manipulated: cognitive conflict (high-conflict
vs. low-conflict no-go stimuli), complexity (simple vs. com-
plex task), and novelty (first vs. second run).
Previous studies in the literature have already shown that
conflict (Zysset et al., 2007; Gazzaley et al., 2005; Nielson
et al., 2002), task complexity (Gazzaley et al., 2007; Viskontas
et al., 2004), or novelty (Erickson et al., 2007b; Kramer et al.,
1999) may affect performance and brain activation in older
adults. The current study extends these previous findings
by combining, in a single experimental design, manipula-
tions concerning these three task components. As it will
appear from the following discussion, this design was suc-
cessful in showing that this combination of factors
is important for observing age-related differences in brain
activation.
At the behavioral level, participants in both age groups
performed at ceiling in the low-conflict no-go condition,
whereas they made a number of commission errors for
the high-conflict no-go stimuli, especially in the complex
task. Both age groups improved their performance to
high-conflict no-go stimuli from the first to the second run
810
Journal of Cognitive Neuroscience
Volume 23, Number 4
of the complex task. Analyses on RTs to go stimuli showed
that beneficial practice effects in speeding up the task
execution especially occurred in the older group for the
complex task. We speculate that this speeding up was simi-
lar in go and high-conflict no-go conditions, but given the
nature of the go/no-go task, we did not have a measure of
speed for the latter.
In the following sections, we will describe the cumulative
effects of the factors influencing age-related changes in
the functional brain activity (i.e., conflict, complexity and
novelty). First, we will consider the effect of cognitive con-
flict. A cohesive set of brain regions, including bilateral
fronto-parietal regions and superior medial prefrontal cor-
tex, was similarly activated in both age groups for high-
conflict no-go stimuli and was deactivated for low-conflict
no-go stimuli in the simpler task. Some of these regions,
such as right lateral prefrontal cortex, have been related
to inhibitory processes (e.g., Aron, Robbins, & Poldrack,
2004; Hester et al., 2004; Rubia et al., 2001). Notwithstand-
ing the specific role of each of these neural nodes (Stuss
& Alexander, 2007), taken together they overlap with an
extensive fronto-parietal network that is involved during
most cognitive control tasks (Kelly, Hester, Foxe, Shpaner,
& Garavan, 2006; Fox et al., 2005; Duncan & Owen, 2000),
and shows intrinsic functional connectivity even at rest
(Vincent, Kahn, Snyder, Raichle, & Buckner, 2008).
However, marked age-specific modulation in the ac-
tivation of these regions emerged as the task difficulty
increased, as reflected in the interaction with task complex-
ity. Older participants overrecruited these regions when
conflicting no-go stimuli were embedded in the complex
task. These results corroborate previous fMRI studies of
aging reporting overrecruitment of a similar fronto-parietal
network during conditions requiring top–down control
(Grady, 2008; Morcom et al., 2007; Zysset et al., 2007). Thus,
overrecruitment especially occurs in the older group with
increasing task demands (it mostly occurs for high-conflict
vs. low-conflict no-go stimuli and during the complex vs.
simple task).
Because age-related difficulties are likely to emerge in
nonroutine situations, such as during the early phases of
task performance (e.g., Erickson et al., 2007b; Kramer
et al., 1999), the early and late runs of the complex task
were directly compared in a follow-up task-PLS analysis.
This analysis showed a set of brain regions similar to those
extracted in the previous analysis, and further character-
ized their modus operandi by revealing opposite effects
of practice for the two age groups. Older participants
showed a pronounced cohesive activation of these re-
gions in the first run, whereas activation was reduced
in the second run. In contrast, in the young group, this
set of regions did not show a consistent activation in the
first run, but its overall activation appeared in the second
run only. As a result, pronounced age-dependent differ-
ences were present in the pattern of brain activation in
the first run, whereas those differences disappeared in
the second run.
This study extends previous work by showing that the
often reported nonselective overrecruitment of brain re-
gions, including homologous fronto-parietal areas in both
hemispheres (Cabeza, 2002; Logan et al., 2002; Park et al.,
2001; Reuter-Lorenz, Stanczak, & Miller, 1999), mostly
occurs in aging with increasing task demands. More spe-
cifically, older adults need more neural resources than
younger controls to initially set up the criteria for over-
coming prepotent responding under complex task rules.
Our next question was whether using these extra neural
resources was, in fact, beneficial or detrimental for older
adultsʼ performance. Important insights on this issue came
from brain–behavior correlation analyses. Correlation anal-
yses indeed showed that neural overrecruitment mostly
occurs in those older individuals who were more success-
ful in dealing with increasing task demands, that is, those
individuals who made less commission errors (i) in the
conflicting no-go condition (ii) of the first run (iii) of the
complex task. Moreover, this initial overrecruitment tended
to occur much less in the older adults whose performance
improved during the second run only, suggesting a less
efficient capacity to initially set up the task criteria in this
subgroup.
Therefore, neural overrecruitment is beneficial for ini-
tial task performance in aging, favoring compensatory
accounts over inefficiency ones (see Grady, 2008, for a re-
view). More generally, these findings underscore the impor-
tance of looking at the between-subjects variability in the
use of spared neural resources to determine the functional
neuroanatomy that distinguish the older adults who age
gracefully from those who do not (Buckner, 2004; Cabeza,
Anderson, Locantore, & McIntosh, 2002).
A possible limit of the brain–behavior correlation anal-
ysis used in the present study is that it correlates errors
with a measure of brain activation on correct conflicting
no-go trials. Accuracy was the only behavioral measure
available for this condition in the present study, and it
is conceivable that the same subjects that produce most
commission errors on no-go trials are those that mostly
suffer from response conflict, and mostly activate partial
go responses even on correct no-go trials, although we
did not have any measure to support this assertion in
the present study. Future studies should employ a mea-
sure of partial response activation, such as electromyo-
graphic recording on the peripheral muscle used for the
go responses or electroencephalographic correlates of re-
sponse preparation (see Vallesi & Stuss, 2010), as a more
suitable dependent variable to correlate with brain activa-
tion on correct no-go trials.
An apparently surprising result is that, in the first run of
the complex task, young adults did not activate the fronto-
parietal network engaged in a cohesive manner by older
participants and, to a minor extent, by both groups in
the first run of the simple task (Figure 3A). A similar pattern
has already been described in young adults when learning
occurs specifically under highly demanding task conditions
(Erickson et al., 2007a; Sakai, Ramnani, & Passingham,
Vallesi, McIntosh, and Stuss
811
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
–
p
r
d
t
i
2
c
3
l
4
e
–
8
p
0
d
1
f
1
/
9
2
4
3
0
/
9
4
7
/
0
8
o
0
c
1
n
/
1
2
0
7
1
7
0
5
2
2
1
1
3
4
9
/
0
j
o
p
c
d
n
.
b
2
y
0
g
1
u
0
e
.
s
t
2
o
1
n
4
9
0
0
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
.
f
t
/
.
.
o
n
1
8
M
a
y
2
0
2
1
2002), suggesting a link between this increasing activity
and the emergence of a strategy that is gradually set up
in young adults. At any rate, this opposite pattern of results
shows that the relation between functional brain activity
and performance may change with age up to the point of
overturning in different age groups (e.g., Vallesi, McIntosh,
Shallice, & Stuss, 2009; Rypma & DʼEsposito, 2000), an
issue that certainly deserves further investigation.
Possible differences in the strategy used by the two age
groups should be considered. The fact that older subjects
use the regions belonging to the control network in the
learning stages of task performance (cf. Chein & Schneider,
2005) more than young controls (and reduce the need to
use it after practice) may reflect a strategic shift toward
a more reflective, deliberative cognitive style with age
(Velanova, Lustig, Jacoby, & Buckner, 2007). Although the
older adults as a group seem to use this control network to
reach a high performance level as soon as they begin to
face a complex task, young individuals might start to use
this network later on, probably to compensate for fatigue
and distraction arising as the task goes on, selectively in
the high-conflict no-go trials of the more difficult task, in
which they need to keep their focus on a task that does not
become automatic with practice due to its complexity (see
Kelly et al., 2006, for similar results).
However, it is not plausible that young people do not
use brain resources at the beginning of the complex task,
as it would appear if one only considers the analyses re-
ported here. Because the task-PLS analysis, like the princi-
pal component analysis, emphasizes the latent variables
that explain most of the variance in the data, it is possible
that it does not detect more subtle age-specific dynamics
when both groups were considered together. To overcome
this potential pitfall, additional analyses (reported in the
Supplementary Material) focused on the brain activity
in young adults while they perform the complex task. Re-
sults of these analyses suggest that, in the young group,
only a subpart of this extensive network is consistently
sensitive to practice effects (see Table S1 and Supplemen-
tary Figure S2).
Notwithstanding the fact that different and, not nec-
essarily, mutually exclusive accounts may explain the
practice-related divergence between brain activations
in the two age groups, the present results underscore
the importance of training in reducing age differences
both at the behavioral and at the neural level, especially
with high task demands. This can explain why, when par-
ticipants (partially overlapping with those tested here)
were retested 1 to 7 days later in a subsequent ERP ses-
sion with the same tasks ( Vallesi, Stuss, et al., 2009), any
Age × Task complexity interaction disappeared both
behaviorally and neurally as shown by the ERP data.
These findings corroborate and extend the existing lit-
erature. In a recent fMRI study (Erickson et al., 2007b),
younger and older adults performed two visual tasks (color
and letter detection) either separately or simultaneously.
Participants then underwent extensive training on the tasks
over several weeks. Older adults showed a decrease in
dorsal prefrontal activity after training, whereas younger
subjects showed an opposite pattern. The present study
extends these results by showing that age-related differ-
ential effects of practice do not occur with extensive train-
ing only (Erickson et al., 2007b) because even a modest
amount of practice during a single experimental session
is enough to dramatically reduce age differences in func-
tional neural activity (see also Kramer et al., 1999, for simi-
lar behavioral evidence).
A seminal neuroimaging study by Logan et al. (2002)
showed that it is possible to reduce underrecruitment of
certain brain regions when older adults are provided with
explicit instructions on effective strategies to perform a
given task. Complementing these results, the present data
show that also overrecruitment associated with age, which
was present in that study independently of the instructions
provided (Logan et al., 2002), can decrease with practice,
even without exogenous instructions. These findings sug-
gest that the aging brain is capable of functional flexibility
to a larger extent than was previously believed.
As a flip side of the extra engagement of fronto-parietal
regions in high-conflict no-go trials, older adults deacti-
vated these regions more during low-conflict no-go trials.
A possible explanation is that these areas are constantly
engaged in the task in the older group and phasically de-
crease their activation only after the onset of low-conflict
no-go stimuli. On the other hand, age-specific increased ac-
tivation for the low-conflict no-go stimuli was mainly pre-
sent in the rostral medial prefrontal region (Brodmannʼs
area 11). Previous studies have shown that this region is
selectively engaged whenever stimuli markedly deviate
from previous ones, either in location or identity, even
when they do not require any overt decision (Petrides,
Alivisatos, & Frey, 2002; Nobre, Coull, Frith, & Mesulam,
1999). The present data suggest that older adults engage
this region more extensively for deviations from the con-
text (particularly within a novel and complex task) and,
more generally, that they are less able than young adults
in suppressing processing of nontarget information (cf.
Vallesi, Hasher, & Stuss, in press; Vallesi, Stuss, et al., 2009;
Gazzaley et al., 2005).
The go/no-go procedure adopted here allowed us to
investigate age-related changes in overcoming pre-
potent response tendencies. However, the use of no-go
stimuli could make it unclear as to whether the brain re-
gions that are overactivated in the older adults reflect a
greater activation of inappropriate “go” responses in
the high-conflict no-go condition (failure in response sup-
pression), engagement of control processes necessary to
suppress these inappropriate responses, or both. The acti-
vated areas were distinct from those activated for go con-
ditions when the latter were also included in the PLS
analyses (not reported here), thus making the second
alternative relatively more plausible. Furthermore, be-
cause overrecruitment of the specific cognitive control re-
gions reported here was more present in successful older
812
Journal of Cognitive Neuroscience
Volume 23, Number 4
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
–
p
r
d
t
i
2
c
3
l
4
e
–
8
p
0
d
1
f
1
/
9
2
4
3
0
/
9
4
7
/
0
8
o
0
c
1
n
/
1
2
0
7
1
7
0
5
2
2
1
1
3
4
9
/
0
j
o
p
c
d
n
.
b
2
y
0
g
1
u
0
e
.
s
t
2
o
1
n
4
9
0
0
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
.
.
.
/
f
t
o
n
1
8
M
a
y
2
0
2
1
individuals, if this overrecruitment is related to the need
to suppress partial activation of wrong go responses, it
could have a compensatory role. We have tested with a
more appropriate ERP methodology whether age-related
partial motor activations can be detected even in the ab-
sence of errors for different no-go conditions ( Vallesi &
Stuss, 2010). The results of this study showed that this
can be the case especially in those older individuals who
show faster go responses, further confirming a compensa-
tory view.
Overall, the current study suggest that older adults
need more neural resources to implement task setting,
a putative executive function necessary to establish com-
plex and nonroutine task rules that compete with prepo-
tent stimulus–response contingencies (see the first latent
variable of the PLS analysis). Many neuropsychological
and neuroimaging studies have postulated the existence
of task setting, whether they localize it in left lateral pre-
frontal cortex (e.g., Alexander et al., 2007), in fronto-polar
cortex (Sakai & Passingham, 2006), or in more extensive
fronto-parietal networks (e.g., Vallesi, McIntosh, Alexander,
et al., 2009; Dosenbach et al., 2008). Other authors have
used different terms to express the same construct accord-
ing to the domain of investigation, such as “sculpting the
response space” (Fletcher et al., 2000), cognitive associa-
tion formation (e.g., Kim, Vallesi, Picton, & Tulving, 2009),
nonroutine motor learning ( Jueptner et al., 1997), and
strategy production (Shallice, 2004). However, task setting
should be investigated more extensively in future research,
also from the cognitive point of view, before this unitary
construct can be confidently adopted as an alternative
and more parsimonious account to the explanation that
neural overrecruitment occurs in aging with increasing task
demands, as for instance, by manipulating (possibly multi-
componential) factors such as the need for suppression,
task complexity, and novelty.
Changes in the coupling between neural activity and
hemodynamic response may occur with age (Huettel,
Singerman, & McCarthy, 2001; DʼEsposito, Zarahn, Aguirre,
& Rypma, 1999). Nonetheless, these changes mainly
pertain to a decreased signal-to-noise ratio in older adults,
an effect that would bring results somewhat opposite of
those observed here. Moreover, any intrinsic difference
in the hemodynamic response per se as a function of
age could not explain the condition-specific effects ob-
served here.
In conclusion, the present study sheds light on the condi-
tions in which the neural overrecruitment usually reported
in neuroimaging studies of aging may occur and on its
possible functional significance. Compared to younger
adults, older individuals engage more extensively in a cohe-
sive set of fronto-parietal regions to successfully overcome
a prepotent and inappropriate response, but only if the
task is complex and novel. The initial engagement of these
regions has a compensatory role in aging, as it is strongly
associated to the degree of success in avoiding commis-
sion errors.
Acknowledgments
This work was supported by a postdoctoral fellowship funding
from the Canadian Institute of Health Research [CIHR, MFE-
87658] to A. V.; CIHR grants to D. T. S. [MT-12853, GR-14974];
J. S. McDonnell foundation grants to A. R. M. [220020082] and
D. T. S. [21002032]; and the Heart and Stroke Foundation Centre
for Stroke Recovery and Posluns Centre for Stroke and Cognition.
We thank Lynn Hasher for fruitful discussions.
Reprint requests should be sent to Antonino Vallesi, Cognitive
Neuroscience Sector, SISSA, Via Beirut 2–4, 34014, Trieste, Italy,
or via e-mail: vallesi@sissa.it.
REFERENCES
Alexander, M. P., Stuss, D. T., Picton, T., Shallice, T., &
Gillingham, S. (2007). Regional frontal injuries cause
distinct impairments in cognitive control. Neurology, 68,
1515–1523.
Alexander, M. P., Stuss, D. T., Shallice, T., Picton, T. W., &
Gillingham, S. (2005). Impaired concentration due to frontal
lobe damage from two distinct lesion sites. Neurology,
65, 572–579.
Aron, A. R., Robbins, T. W., & Poldrack, R. A. (2004). Inhibition
and the right inferior frontal cortex. Trends in Cognitive
Sciences, 8, 170–177.
Buckner, R. L. (2004). Memory and executive function in aging
and AD: Multiple factors that cause decline and reserve
factors that compensate. Neuron, 44, 195–208.
Bunge, S. A. (2004). How we use rules to select actions:
A review of evidence from cognitive neuroscience.
Cognitive, Affective & Behavioral Neuroscience, 4,
564–579.
Cabeza, R. (2001). Functional neuroimaging of cognitive
aging. In R. Cabeza & A. Kingstone (Eds.), Handbook of
functional neuroimaging of cognition (pp. 379–420).
Cambridge, MA: MIT Press.
Cabeza, R. (2002). Hemispheric asymmetry reduction in
older adults: The HAROLD model. Psychology and Aging,
17, 85–100.
Cabeza, R., Anderson, N. D., Locantore, J. K., & McIntosh, A. R.
(2002). Aging gracefully: Compensatory brain activity in
high-performing older adults. Neuroimage, 17, 1394–1402.
Cabeza, R., Grady, C. L., Nyberg, L., McIntosh, A. R., Tulving, E.,
Kapur, S., et al. (1997). Age-related differences in neural
activity during memory encoding and retrieval: A positron
emission tomography study. Journal of Neuroscience,
17, 391–400.
Chein, J. M., & Schneider, W. (2005). Neuroimaging studies
of practice-related change: fMRI and meta-analytic evidence
of a domain-general control network for learning. Brain
Research, Cognitive Brain Research, 25, 607–623.
Colcombe, S. J., Kramer, A. F., Erickson, K. I., & Scalf, P.
(2005). The implications of cortical recruitment and brain
morphology for individual differences in inhibitory function
in aging humans. Psychology and Aging, 20, 363–375.
Cox, R. W. (1996). AFNI: Software for analysis and visualization
of functional magnetic resonance neuroimages.
Computers and Biomedical Research, 29, 162–173.
Craik, F. I. M., & Byrd, M. (1982). Aging and cognitive
deficits: The role of attentional resources. In F. I. M. Craik
& S. Trehub (Eds.), Aging and cognitive processes
(pp. 191–211). New York: Plenum.
DʼEsposito, M., Zarahn, E., Aguirre, G. K., & Rypma, B.
(1999). The effect of normal aging on the coupling of
neural activity to the bold hemodynamic response.
Neuroimage, 10, 6–14.
Vallesi, McIntosh, and Stuss
813
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
–
p
r
d
t
i
2
c
3
l
4
e
–
8
p
0
d
1
f
1
/
9
2
4
3
0
/
9
4
7
/
0
8
o
0
c
1
n
/
1
2
0
7
1
7
0
5
2
2
1
1
3
4
9
/
0
j
o
p
c
d
n
.
b
2
y
0
g
1
u
0
e
.
s
t
2
o
1
n
4
9
0
0
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
.
.
/
t
f
.
o
n
1
8
M
a
y
2
0
2
1
Dosenbach, N. U., Fair, D. A., Cohen, A. L., Schlaggar, B. L.,
& Petersen, S. E. (2008). A dual-network architecture of
top–down control. Trends in Cognitive Sciences, 12,
99–105.
Duncan, J., & Owen, A. M. (2000). Common regions of
the human frontal lobe recruited by diverse cognitive
demands. Trends in Neurosciences, 23, 475–483.
Efron, B., & Tibshirani, R. (1986). Bootstrap methods for
standard errors, confidence intervals and other measures
of statistical accuracy. Statistical Science, 1, 54–77.
Erickson, K. I., Colcombe, S. J., Wadhwa, R., Bherer, L.,
Hasher, L., Zacks, R. T., & May, C. P. (1999). Inhibitory control,
circadian arousal, and age. In D. Gopher & A. Koriat (Eds.),
Attention and performance: XVII. Cognitive regulation
of performance: Interaction of theory and application
(pp. 653–675). Cambridge, MA: MIT Press.
Hedden, T., & Gabrieli, J. D. (2004). Insights into the ageing
mind: A view from cognitive neuroscience. Nature Reviews
Neuroscience, 5, 87–96.
Hester, R., Murphy, K., & Garavan, H. (2004). Beyond common
resources: The cortical basis for resolving task interference.
Neuroimage, 23, 202–212.
Peterson, M. S., Scalf, P. E., et al. (2007a). Training-induced
functional activation changes in dual-task processing:
An fMRI study. Cerebral Cortex, 17, 192–204.
Erickson, K. I., Colcombe, S. J., Wadhwa, R., Bherer, L.,
Huettel, S. A., Singerman, J. D., & McCarthy, G. (2001).
The effects of aging upon the hemodynamic response
measured by functional MRI. Neuroimage, 13, 161–175.
Jueptner, M., Stephan, K. M., Frith, C. D., Brooks, D. J.,
D
o
w
n
l
o
a
d
e
d
Peterson, M. S., Scalf, P. E., et al. (2007b). Training-induced
plasticity in older adults: Effects of training on hemispheric
asymmetry. Neurobiology of Aging, 28, 272–283.
Fletcher, P. C., Shallice, T., & Dolan, R. J. (2000). “Sculpting
the response space”: An account of left prefrontal activation
at encoding. Neuroimage, 12, 404–417.
Folstein, M. F., Folstein, S. E., & McHugh, P. R. (1975).
“Mini-mental state”. A practical method for grading the
cognitive state of patients for the clinician. Journal of
Psychiatric Research, 12, 189–198.
Fox, M. D., Snyder, A. Z., Vincent, J. L., Corbetta, M., Van Essen,
D. C., & Raichle, M. E. (2005). The human brain is intrinsically
organized into dynamic, anticorrelated functional networks.
Proceedings of the National Academy of Sciences, U.S.A.,
102, 9673–9678.
Friston, K. J., Ashburner, J., Frith, C. D., Pline, J. B., Heather, J. D.,
& Frackowiak, R. S. (1995). Spatial registration and
normalization of images. Human Brain Mapping, 2,
165–189.
Gazzaley, A., Clapp, W., Kelley, J., McEvoy, K., Knight, R. T.,
& DʼEsposito, M. (2008). Age-related top–down suppression
deficit in the early stages of cortical visual memory
processing. Proceedings of the National Academy of
Sciences, U.S.A., 105, 13122–13126.
Gazzaley, A., Cooney, J. W., Rissman, J., & DʼEsposito, M.
(2005). Top–down suppression deficit underlies working
memory impairment in normal aging. Nature Neuroscience,
8, 1298–1300.
Gazzaley, A., Sheridan, M. A., Cooney, J. W., & DʼEsposito, M.
(2007). Age-related deficits in component processes
of working memory. Neuropsychology, 21, 532–539.
Grady, C. L. (2008). Cognitive neuroscience of aging. Annals
of the New York Academy of Sciences, 1124,
127–144.
Grady, C. L., Maisog, J. M., Horwitz, B., Ungerleider, L. G.,
Mentis, M. J., Salerno, J. A., et al. (1994). Age-related
changes in cortical blood flow activation during visual
processing of faces and location. Journal of Neuroscience,
14, 1450–1462.
Grady, C. L., McIntosh, A. R., Horwitz, B., Maisog, J. M.,
Ungerleider, L. G., Mentis, M. J., et al. (1995). Age-related
reductions in human recognition memory due to impaired
encoding. Science, 269, 218–221.
Gutchess, A. H., Welsh, R. C., Hedden, T., Bangert, A., Minear, M.,
Liu, L. L., et al. (2005). Aging and the neural correlates of
successful picture encoding: Frontal activations compensate
for decreased medial-temporal activity. Journal of Cognitive
Neuroscience, 17, 84–96.
Hasher, L., & Zacks, R. T. (1988). Working memory,
comprehension, and aging: A review and a new view.
In G. H. Bower (Ed.), The psychology of learning and
motivation ( Vol. 22, pp. 193–225). New York: Academic
Press.
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
–
p
r
d
t
i
2
c
3
l
4
e
–
8
p
0
d
1
f
1
/
9
2
4
3
0
/
9
4
7
/
0
8
o
0
c
1
n
/
1
2
0
7
1
7
0
5
2
2
1
1
3
4
9
/
0
j
o
p
c
d
n
.
b
2
y
0
g
1
u
0
e
.
s
t
2
o
1
n
4
9
0
0
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
.
/
t
.
.
f
o
n
1
8
M
a
y
2
0
2
1
Frackowiak, R. S., & Passingham, R. E. (1997). Anatomy
of motor learning. I. Frontal cortex and attention to action.
Journal of Neurophysiology, 77, 1313–1324.
Kelly, A. M., Hester, R., Foxe, J. J., Shpaner, M., & Garavan, H.
(2006). Flexible cognitive control: Effects of individual
differences and brief practice on a complex cognitive
task. Neuroimage, 31, 866–886.
Keys, B. A., & White, D. A. (2000). Exploring the relationship
between age, executive abilities, and psychomotor speed.
Journal of the International Neuropsychological Society,
6, 76–82.
Kim, A. S., Vallesi, A., Picton, T. W., & Tulving, E. (2009).
Cognitive association formation in episodic memory:
Evidence from event-related potentials. Neuropsychologia,
47, 3162–3173.
Kramer, A. F., Hahn, S., & Gopher, D. (1999). Task coordination
and aging: Explorations of executive control processes in
the task switching paradigm. Acta Psychologica
(Amsterdam), 101, 339–378.
Lancaster, J. L., Woldorff, M. G., Parsons, L. M., Liotti, M.,
Freitas, C. S., Rainey, L., et al. (2000). Automated Talairach
atlas labels for functional brain mapping. Human Brain
Mapping, 10, 120–131.
Logan, J. M., Sanders, A. L., Snyder, A. Z., Morris, J. C., &
Buckner, R. L. (2002). Under-recruitment and nonselective
recruitment: Dissociable neural mechanisms associated
with aging. Neuron, 33, 827–840.
McIntosh, A. R., Bookstein, F. L., Haxby, J. V., & Grady, C. L.
(1996). Spatial pattern analysis of functional brain images
using Partial Least Squares. Neuroimage, 3, 143–157.
Morcom, A. M., Li, J., & Rugg, M. D. (2007). Age effects on
the neural correlates of episodic retrieval: Increased cortical
recruitment with matched performance. Cerebral Cortex,
17, 2491–2506.
Nielson, K. A., Langenecker, S. A., & Garavan, H. (2002).
Differences in the functional neuroanatomy of inhibitory
control across the adult life span. Psychology and Aging,
17, 56–71.
Nobre, A. C., Coull, J. T., Frith, C. D., & Mesulam, M. M. (1999).
Orbitofrontal cortex is activated during breaches of
expectation in tasks of visual attention. Nature
Neuroscience, 2, 11–12.
Oldfield, R. C. (1971). The assessment and analysis of
handedness: The Edinburgh inventory. Neuropsychologia,
9, 97–113.
Park, D. C., Polk, T. A., Mikels, J. A., Taylor, S. F., & Marshuetz, C.
(2001). Cerebral aging: Integration of brain and behavioral
models of cognitive function. Dialogue in Clinical
NeuroSciences, 3, 151–165.
Persson, J., Sylvester, C. Y., Nelson, J. K., Welsh, K. M., Jonides, J.,
& Reuter-Lorenz, P. A. (2004). Selection requirements during
verb generation: Differential recruitment in older and
younger adults. Neuroimage, 23, 1382–1390.
814
Journal of Cognitive Neuroscience
Volume 23, Number 4
Petrides, M., Alivisatos, B., & Frey, S. (2002). Differential
activation of the human orbital, mid-ventrolateral, and
mid-dorsolateral prefrontal cortex during the processing
of visual stimuli. Proceedings of the National Academy
of Sciences, U.S.A., 99, 5649–5654.
Reuter-Lorenz, P. (2002). New visions in the aging mind and
brain. Trends in Cognitive Sciences, 6, 394.
Reuter-Lorenz, P. A., Stanczak, L., & Miller, A. C. (1999).
Neural recruitment and cognitive aging: Two hemispheres
are better than one especially as you age. Psychological
Science, 10, 494–500.
Rubia, K., Russell, T., Overmeyer, S., Brammer, M. J., Bullmore,
E. T., Sharma, T., et al. (2001). Mapping motor inhibition:
Conjunctive brain activations across different versions of
go/no-go and stop tasks. Neuroimage, 13, 250–261.
Rypma, B., & DʼEsposito, M. (2000). Isolating the neural
mechanisms of age-related changes in human working
memory. Nature Neuroscience, 3, 509–515.
Rypma, B., Eldreth, D. A., & Rebbechi, D. (2007). Age-related
differences in activation–performance relations in delayed-
response tasks: A multiple component analysis. Cortex,
43, 65–76.
Sakai, K., & Passingham, R. E. (2006). Prefrontal set activity
predicts rule-specific neural processing during subsequent
cognitive performance. Journal of Neuroscience, 26,
1211–1218.
Sakai, K., Ramnani, N., & Passingham, R. E. (2002). Learning
of sequences of finger movements and timing: Frontal
lobe and action-oriented representation. Journal of
Neurophysiology, 88, 2035–2046.
Salthouse, T. A. (1996). The processing-speed theory of adult
age differences in cognition. Psychological Review, 103,
403–428.
Shallice, T. (2004). The fractionation of supervisory control.
In M. S. Gazzaniga (Ed.), The cognitive neurosciences
(pp. 943–955). Cambridge, MA: MIT Press.
Smith, E. E., Geva, A., Jonides, J., Miller, A., Reuter-Lorenz, P.,
& Koeppe, R. A. (2001). The neural basis of task-switching
in working memory: Effects of performance and aging.
Proceedings of the National Academy of Sciences, U.S.A.,
98, 2095–2100.
Stroop, J. R. (1935). Studies of interference in serial verbal
reactions. Journal of Experimental Psychology, 18,
643–662.
Stuss, D. T., & Alexander, M. P. (2007). Is there a dysexecutive
syndrome? Philosophical Transactions of the Royal
Society of London, Series B, Biological Sciences, 362,
901–915.
Stuss, D. T., Shallice, T., Alexander, M. P., & Picton, T. W.
(1995). A multidisciplinary approach to anterior attentional
functions. Annals of the New York Academy of Sciences,
769, 191–211.
Sweeney, J. A., Rosano, C., Berman, R. A., & Luna, B. (2001).
Inhibitory control of attention declines more than working
memory during normal aging. Neurobiology of Aging,
22, 39–47.
Vallesi, A., Hasher, L., & Stuss, D. T. (in press). Age-related
changes in transfer costs: Evidence from go/nogo tasks.
Psychology and Aging.
Vallesi, A., McIntosh, A. R., Alexander, M., & Stuss, D. T. (2009).
fMRI evidence of a functional network setting the criteria
for withholding a response. Neuroimage, 45, 537–548.
Vallesi, A., McIntosh, A. R., Shallice, T., & Stuss, D. T. (2009).
When time shapes behaviour: fMRI evidence of brain
correlates of strategic preparation over time. Journal of
Cognitive Neuroscience, 21, 1116–1126.
Vallesi, A., & Stuss, D. T. (2010). Excessive sub-threshold
motor preparation for non-target stimuli in normal aging.
Neuroimage, 50, 1251–1257.
Vallesi, A., Stuss, D. T., McIntosh, A. R., & Picton, T. W. (2009).
Age-related differences in processing irrelevant information:
Evidence from event-related potentials. Neuropsychologia,
47, 577–586.
Velanova, K., Lustig, C., Jacoby, L. L., & Buckner, R. L. (2007).
Evidence for frontally mediated controlled processing
differences in older adults. Cerebral Cortex, 17, 1033–1046.
Verhaeghen, P., Cerella, J., & Basak, C. (2006). Aging, task
complexity, and efficiency modes: The influence of working
memory involvement on age differences in response times
for verbal and visuospatial tasks. Neuropsychology,
Development, and Cognition: Section B, Aging,
Neuropsychology and Cognition, 13, 254–280.
Vincent, J. L., Kahn, I., Snyder, A. Z., Raichle, M. E., & Buckner, R. L.
(2008). Evidence for a frontoparietal control system
revealed by intrinsic functional connectivity. Journal
of Neurophysiology, 100, 3328–3342.
Viskontas, I. V., Morrison, R. G., Holyoak, K. J., Hummel, J. E.,
& Knowlton, B. J. (2004). Relational integration, inhibition,
and analogical reasoning in older adults. Psychology and
Aging, 19, 581–591.
Willis, S. L., & Nesselroade, C. S. (1990). Long-term effects
of fluid ability training in old–old age. Developmental
Psychology, 26, 905–910.
Zarahn, E., Rakitin, B., Abela, D., Flynn, J., & Stern, Y. (2007).
Age-related changes in brain activation during a delayed
item recognition task. Neurobiology of Aging, 28, 784–798.
Zysset, S., Schroeter, M. L., Neumann, J., & Yves von Cramon, D.
(2007). Stroop interference, hemodynamic response and
aging: An event-related fMRI study. Neurobiology of Aging,
28, 937–946.
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
–
p
r
d
t
i
2
c
3
l
4
e
–
8
p
0
d
1
f
1
/
9
2
4
3
0
/
9
4
7
/
0
8
o
0
c
1
n
/
1
2
0
7
1
7
0
5
2
2
1
1
3
4
9
/
0
j
o
p
c
d
n
.
b
2
y
0
g
1
u
0
e
.
s
t
2
o
1
n
4
9
0
0
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
.
.
t
.
/
f
o
n
1
8
M
a
y
2
0
2
1
Vallesi, McIntosh, and Stuss
815