Neural Dynamics of Conflict Control in Working Memory
Khoi D. Vo1, Audrey Siqi-Liu1, Alondra Chaire2, Sophia Li1,
Elise Demeter3, Tobias Egner1, and Marty G. Woldorff1
Abstract
■ Attention and working memory ( WM) have classically been
considered as two separate cognitive functions, but more recent
theories have conceptualized them as operating on shared repre-
sentations and being distinguished primarily by whether attention
is directed internally ( WM) or externally (attention, traditionally
defined). Supporting this idea, a recent behavioral study docu-
mented a “WM Stroop effect,” showing that maintaining a color
word in WM impacts perceptual color-naming performance to
the same degree as presenting the color word externally in the
classic Stroop task. Here, we employed ERPs to examine the neu-
ral processes underlying this WM Stroop task compared to those
in the classic Stroop and in a WM-control task. Based on the
assumption that holding a color word in WM would (pre-)activate
the same color representation as by externally presenting that
color word, we hypothesized that the neural cascade of conflict–
control processes would occur more rapidly in the WM Stroop
than in the classic Stroop task. Our behavioral results replicated
equivalent interference behavioral effects for the WM and classic
Stroop tasks. Importantly, however, the ERP signatures of conflict
detection and resolution displayed substantially shorter latencies
in the WM Stroop task. Moreover, delay-period conflict in the WM
Stroop task, but not in the WM control task, impacted the ERP
and performance measures for the WM probe stimuli. Together,
these findings provide new insights into how the brain processes
conflict between internal representations and external stimuli,
and they support the view of shared representations between
internally held WM content and attentional processing of external
stimuli. ■
INTRODUCTION
We often perform tasks that require an externally directed
focus of attention (such as driving a car or cooking a
meal), while also keeping in mind internal information
(working memory [WM]) that may or may not be related
to the task at hand (e.g., remembering directions vs.
remembering to return a missed call from your in-laws).
What we maintain in WM can affect our attention and thus
substantially impact the way we interact with stimuli in
our environment. Likewise, our interaction with the exter-
nal world can also influence the information we are
endeavoring to maintain in WM. Although the interplay
between attention and WM is crucial to successfully navi-
gating everyday life, the nature of their relationship is not
well understood.
Attention and WM have classically been considered two
separate cognitive functions: Attention refers to the prior-
itized processing of a subset of information in the
environment (Treisman, 1960; Broadbent, 1958), whereas
WM has traditionally been thought of as a dedicated cog-
nitive system that acts as a buffer to temporarily process
and maintain information internally when such informa-
tion is no longer present in the environment (Baddeley,
1992; Baddeley & Hitch, 1974). Recent work, however,
1Duke University, Durham, NC, 2German Centre for Neuro-
degenerative Diseases, Magdeburg, Germany, 3University of
North Carolina at Charlotte
© 2021 Massachusetts Institute of Technology
has indicated that attention and WM are not as separable
as previously believed. Specifically, WM could be con-
ceived as an internally directed form of attention, with a
common set of representational substrates and attentional
control mechanisms that either activate representations
endogenously or facilitate their activation by external
stimuli by attending to them (Kiyonaga & Egner, 2013;
Gazzaley & Nobre, 2012; Postle, 2006; Awh & Jonides,
2001).
A compelling set of behavioral findings to support this
notion derives from a modified color-word Stroop task
devised by Kiyonaga and Egner (2014). These authors
showed that the classic Stroop effect (MacLeod, 1991;
Stroop, 1935)—slower and more error-prone responses
when participants have to name the font color of a
semantically incongruent word (e.g., the word BLUE in
red ink) compared to a congruent one (e.g., the word
BLUE in blue ink)—could be instantiated when partici-
pants were instead asked to hold a color word (e.g.,
“blue”) in mind during a simple perceptual color catego-
rization task (classifying colored swatches as being blue,
green, red, or yellow). This “WM Stroop effect” was also
subject to modulation by the proportion of incongruent
trials just like the classic Stroop effect (e.g., Bugg &
Crump, 2012), with smaller Stroop effects under condi-
tions of more frequent incongruent trials (Kiyonaga &
Egner, 2014). These findings thus indicated that internally
activated representations can interfere with externally
Journal of Cognitive Neuroscience 33:10, pp. 2079–2092
https://doi.org/10.1162/jocn_a_01744
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
1
0
2
0
7
9
1
9
6
2
1
3
5
/
/
j
o
c
n
_
a
_
0
1
7
4
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
attended information in a similar way as external distrac-
ters, with similar strategic effects on conflict-resolution
processes, suggesting that WM shares representation
and control processes with external attention.
However, because of the purely behavioral nature of
these findings, it is not known to what degree the under-
lying neural control mechanisms may differ between the
circumstances of an internal versus external source of
processing conflict. In particular, although many prior
studies have investigated the neural mechanisms of de-
tecting and resolving conflict in information processing
that stems from external distractors, such as in the classic
Stroop task (Appelbaum, Meyerhoff, & Woldorff, 2009;
Egner & Hirsch, 2005; Kerns et al., 2004; MacDonald,
Cohen, Stenger, & Carter, 2000), it is not known how
conflict stemming from an internal WM representation
might be detected and resolved. The core assumption
of shared representations between WM and attended ex-
ternal stimuli results in a strong prediction regarding the
temporal cascade of neural conflict–control processes.
Namely, a color word held in WM would result in a
“pre-activation” of the corresponding representation,
and thus the conflict generation (and resolution) in the
WM Stroop task would be expected to occur more rapidly
than in the classic Stroop task, where color and color-
word information are both presented externally and si-
multaneously. Given this timing-related prediction, a
particularly sensitive way to approach this question would
be to leverage the high temporal resolution of EEG.
Accordingly, we addressed this question here by com-
bining an adapted and extended version of the WM
Stroop task (Kiyonaga & Egner, 2014) with EEG, along
with an additional, specifically directed, control task.
Participants performed three variants of the Stroop task
(see Methods section). Of the three Stroop variant tasks,
two were delayed-match-to-sample WM tasks that inte-
grated the color discrimination into the delay period,
with the third being the classic Stroop task. The key com-
monality across these tasks was that subjects were re-
quired to perform a color discrimination on an external
stimulus in which the processing of the task-relevant
stimulus feature (a color swatch) could be interfered with
by other conflicting task-irrelevant information (i.e., the
color word). Between the two WM tasks, the key differ-
ence was whether the source of this conflict (i.e., how
task-irrelevant information was introduced) was from a
clash with another external stimulus feature (classic
Stroop task) or a clash with information being maintained
in WM ( WM Stroop task). The key difference between the
two WM tasks was whether or not the item being held in
WM overlapped in domain with the task demands during
the delay-period Stroop task. The standard WM Stroop
task (keeping color words in WM) involved such an over-
lap (color representations), whereas the newly devised
WM Stroop control task did not (participants were instead
asked to remember a number word rather than a color
word). Contrasts between these three tasks allowed us
to delineate the neural dynamics—in terms of amplitudes
and latencies of ERP measures of brain activity—of pro-
cessing conflict stemming from a clash between internal
information and externally presented stimuli compared to
conflict between two features of an externally presented
stimulus, while controlling for WM load.
METHODS
Participants
Thirty-three healthy volunteers provided written in-
formed consent to participate in our study in accordance
with the Duke University institutional review board.
Three participants were excluded because of an excessive
level of noise in the EEG data (n = 2) or poor behavioral
performance (n = 1), yielding 30 participants for the final
analyses (13 men; ages 18–30 years). All participants
passed a screening before the study that ensured they
were fluent in English, did not have any color-vision de-
ficiency, and were not taking any attention-enhancing
drugs. Participants were compensated $15 per hour for
their participation.
Tasks
Participants performed three variants of the Stroop task,
as introduced above and described in detail below.
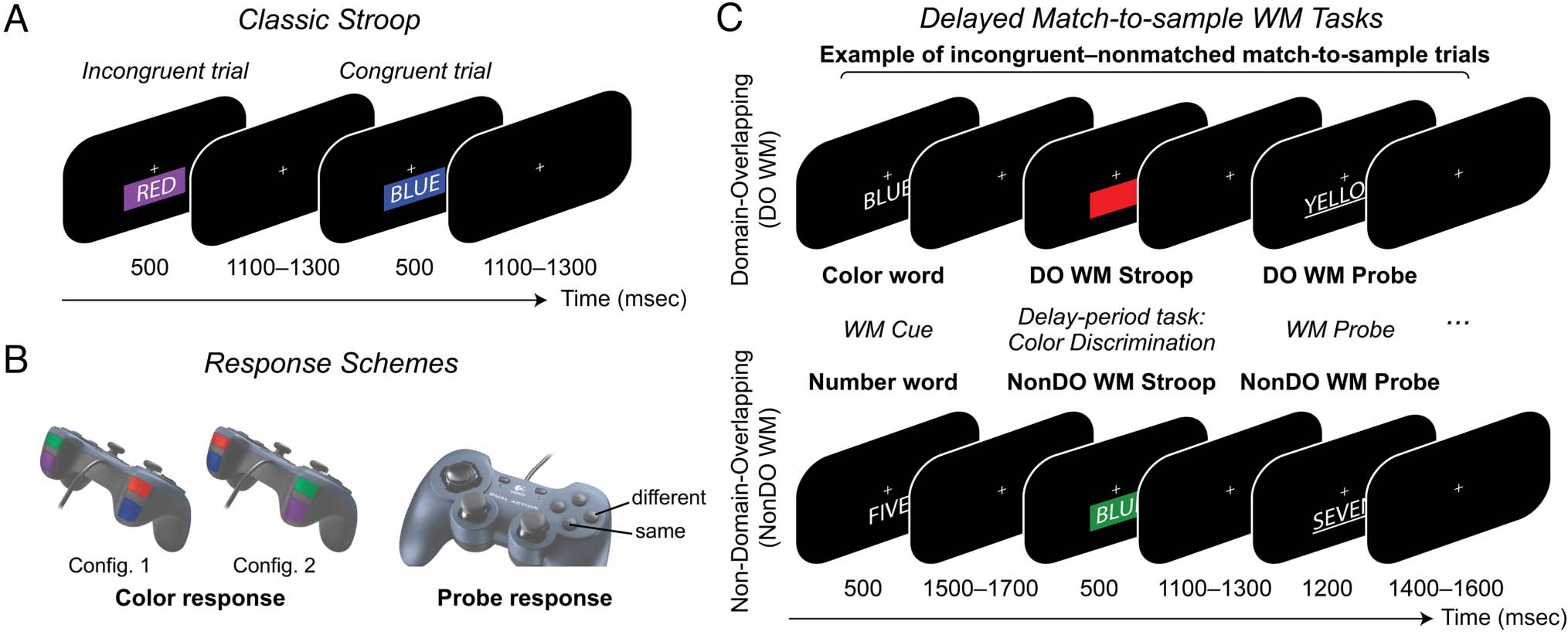
Classic Stroop Task
The prototypical Stroop task (Stroop, 1935) consists of a
series of color words presented in colored font where
participants are tasked to report the font color while
ignoring the semantic meaning of the word. However, in
order to approximately equate visual stimuli across tasks in
this study, our classic Stroop task (Figure 1A) consisted of
a series of color words (red, blue, green, purple) written in
white font (red, green, blue values: 200, 200, 200) and
superimposed on rectangular color swatches (red = 255,
0, 0; blue = 0, 0, 255; green = 34, 139, 34; purple = 153,
50, 204). Using one of four designated response keys on a
gamepad mapped to the possible colors, participants were
asked to report the color of the rectangular swatches and
to ignore the accompanying color word (Figure 1A).
Previous work from our group has demonstrated that this
alternative presentation of Stroop stimuli produces similar
behavioral conflict effects as the standard Stroop stimulus
(Appelbaum, Boehler, Davis, Won, & Woldorff, 2014;
Appelbaum, Boehler, Won, Davis, & Woldorff, 2012;
Appelbaum et al., 2009). In this task, participants were
given up to 1000 msec to respond. Each color-word swatch
stimulus was presented on a black background and sepa-
rated in time by a jittered intertrial interval of 1100–
1300 msec. Equal proportions of congruent trials (semantic
meaning and color swatch in agreement) and incongruent
trials (semantic meaning and color swatch differing) were
2080
Journal of Cognitive Neuroscience
Volume 33, Number 10
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
1
0
2
0
7
9
1
9
6
2
1
3
5
/
/
j
o
c
n
_
a
_
0
1
7
4
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
1
0
2
0
7
9
1
9
6
2
1
3
5
/
/
j
o
c
n
_
a
_
0
1
7
4
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 1. Experimental paradigm with sample trial sequences. Participants completed each task in four independent blocks of 60 trials (240 trials per
task), where the common task requirement was to perform a color discrimination on an external stimulus (color patch). (A) Our version of a classic
Stroop task where the stimuli have been altered in order to control for visual presentation across the three tasks. (B) Two response schemes for color
responses (counterbalanced across participants) and one response scheme for probe responses (constant across participants). (C) Two delayed
match-to-sample WM tasks where participants were told that the item held in WM ( WM Cue) will be tested ( WM probe) after they performed a color
discrimination task during the delay period.
presented in randomized order. This task was used to
assess typical conflict–control processes outside of the
WM domain, where conflict originates from clashing
representations triggered by two features of an external
stimulus, that is, the color of the swatch and the semantic
meaning of the printed color word.
Domain-overlapping WM Stroop Task
This delayed match-to-sample task (Figure 1C, top) was
patterned after the WM-variant Stroop task from
Kiyonaga and Egner (2014). Each trial started with a color
word WM cue (the words RED, BLUE, GREEN, or
PURPLE, printed in white on a black background), which
participants were asked to remember for a subsequent
WM test at the end of the trial. Following the WM cue,
participants were shown a rectangular color swatch
(red, blue, green, or purple), with the task to report
the color of the swatch using the same button mapping
as the classic Stroop task. Because the internally attended
stimuli (i.e., the color word memoranda) and perceptual
stimuli during the delay-period task had semantic/
feature-domain overlap (i.e., color), we refer to it as the
domain-overlapping WM Stroop task (DO WM Stroop).
After the delay period, another color word, now under-
lined to clarify that it was the WM probe, was shown
and participants indicated whether this probe matched
the WM cue they had kept in memory. This DO WM task,
in particular its delay-period segment, was used to assess
conflict–control processes when conflict was caused by
incompatibility between an internally represented WM
item (the color word) and an external stimulus feature
(the color of the swatch).
Participants had up to 1000 msec to respond during
the DO WM Stroop task and up to 1200 msec to respond
to the DO WM probe. A jittered interstimulus interval
(ISI) of 1500–1700 msec separated the WM cue and the
color swatch, and a jittered ISI of 1100–1300 msec sepa-
rated the color swatch and the WM probe. A jittered in-
tertrial interval of 1400–1600 msec separated each
compound-event trial. Equal proportions of congruent
trials (WM cue meaning and color swatch in agreement)
and incongruent trials ( WM cue meaning and color
swatch differing), as well as equal proportions of trials of
matching and nonmatching WM probes, were presented
in randomized order.
Non-DO WM Stroop Task
We included an additional dual-task paradigm (Figure 1C,
bottom) as a comparison to the DO WM task that was not
present in the original Kiyonaga and Egner (2014) study.
This dual task was also a delayed match-to-sample WM
task, but here the memoranda did not overlap semanti-
cally with the relevant external stimulus feature during
the delay-period task, thus providing a WM-load control
condition with respect to the DO WM task. Specifically,
the WM cue item and probe in this condition were num-
ber words (four, five, six, seven) instead of color words,
with the delay-period task using the same structure and
stimuli as our classic Stroop task (color word superim-
posed on a rectangular color swatch). Because the WM
item and delay-period Stroop task did not have any
feature-domain overlap, we refer to this Stroop as the
non-DO WM Stroop task (NonDO WM Stroop). Thus,
both the NonDO WM Stroop and the classic Stroop
Vo et al.
2081
required control over conflict triggered by clashing external
stimulus features, rather than triggered by an internal WM
representation clashing with an external stimulus as in the
DO WM Stroop task. In addition, the fact that participants
had to also keep in mind a WM item in this condition
allowed us to compare the interplay between external
processing and WM maintenance as a function of whether
there was or was not domain/semantic overlap between
the external item and the internally held one (i.e., com-
paring the responses to the NonDO WM probe and the
DO WM probe).
The trial organization, timing, task requirements, and
response mappings of the NonDO WM task were the same
as the DO WM task. Likewise, equal proportions of congru-
ent (color word meaning and color swatch in agreement)
and incongruent (color word meaning and color swatch
differing) Stroop stimuli, as well as equal proportions of
matching and nonmatching WM probes, were presented
in randomized order. For the purpose of this report, WM
probes for the DO and NonDO WM tasks were analyzed
based on whether the preceding delay-period color dis-
crimination task was a congruent or incongruent trial,
but not as a function of whether they were matching or
nonmatching with the memoranda because this contrast
was too general and did not specifically highlight the im-
pact of delay-period task conflicts on WM maintenance.
Procedure
All tasks were programmed using the Presentation software
package (Neurobehavioral Systems, Inc.) and presented on
a 24-in. LCD monitor. Data were collected in a dimly lit,
electrically shielded room where participants were seated
in a comfortable chair positioned ∼60 cm from the screen.
To control for eye movement, participants were asked to
continuously fixate on a crosshair placed in the center of
the screen throughout the session. All stimuli were pre-
sented just below this fixation crosshair.
Each participant completed 240 trials per task (classic
Stroop, DO WM Stroop, NonDO WM Stroop) for a total
of 720 trials. Each task was divided into four blocks of
60 trials. Participants completed two blocks for each task
(two DO WM blocks, two NonDO WM blocks, two classic
Stroop blocks, randomized) before taking a break (dura-
tion ∼2–5 min, as needed). After the break, participants
completed the remaining six task blocks, which were again
randomized. This pseudorandomized block ordering was
to ensure that participants were exposed to all three tasks
in a fair manner and that no task was inadvertently ran-
domized to mostly occur during the beginning or the
end of a testing session. Lastly, for every block of 60 trials,
participants were able to take a short break at the midway
point or after the block if needed.
Response Mapping and Practice
Behavioral responses for this study were collected from
both hands using a Logitech Dual Action gamepad
(Figure 1C). To respond during the color discrimination
stage in each task, participants had to push one of the four
bumper buttons on the top of the gamepad using their
index and middle fingers. Each button was associated with
one color (e.g., red-left index, blue-left middle, green-right
index, purple-right middle). Two sets of button-color
mappings (Figure 1B) were counterbalanced across partic-
ipants. To respond to the WM probe, participants used
their right thumb to press button 2 when they judged the
WM probe and memoranda to be the same, and button 3
when they thought they were different. Before starting
the study, participants were given time to practice using
the gamepad, completing eight practice color discrimina-
tion trials to learn the button-color mapping and eight prac-
tice WM probe trials to learn the match and nonmatch
button mapping. Participants also practiced the two WM
tasks in full for eight trials in order to familiarize themselves
with the organization and timing of these delayed match-to-
sample trials (e.g., encode WM cue, perform color discrim-
ination, and respond to WM probe). Participants were asked
to repeat the practices until comfortable with the tasks and
familiar with the button mappings to meet the required cri-
teria for moving forward with the study (i.e., high accuracy
[> 90%] and below-threshold response times (RTs) [color
discrimination: < 1000 msec, WM probe: < 1200 msec]).
The practice trial data were not analyzed further.
EEG Data Acquisition and Preprocessing
EEG data (500-Hz sample rate per channel; online low-pass
anti-aliasing filter with a half-amplitude cutoff equivalent
of 130 Hz) were recorded using a 64-channel, custom-
designed, equidistant, extended-coverage cap ( Woldorff
et al., 2002) with active electrodes (ActiCAP) and a Brain
Products actiCHamp amplifier (Brain Products GmbH).
Data were recorded referenced to the right mastoid and
rereferenced off-line to the algebraic average of the left
and right mastoids. Channel impedances were kept be-
low 15 kOhms.
Off-line data analysis was performed using EEGLAB
(Delorme & Makeig, 2004) and FieldTrip (Oostenveld,
Fries, Maris, & Schoffelen, 2011) software packages.
Data were filtered off-line with a 0.05–30 Hz bandpass,
causal, finite impulse response filter and then down-
sampled to 250 Hz. Noisy channels were replaced using
an interpolation of surrounding channels before running
an independent component analysis, where the number
of interpolated channels were accounted for because of
independence considerations (principal components =
63 minus the number of interpolated channels).
Independent components were extracted using the ex-
tended infomax algorithm in EEGLAB. Components that
reflected eye blinks and horizontal eye movement were
removed. This meant one or two components were re-
moved for each participant, with the exception of four
participants who had three components removed. All
time-locked epochs were extracted from −1000 to 2500 msec
2082
Journal of Cognitive Neuroscience
Volume 33, Number 10
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
1
0
2
0
7
9
1
9
6
2
1
3
5
/
/
j
o
c
n
_
a
_
0
1
7
4
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Table 1. Number of Epochs Per Condition: Mean (SD)
DO WM
NonDO
WM
Classic
Stroop
Color discrimination (Stroop)
Congruent
Incongruent
103 (10)
103 (12)
114 (4)
100 (12)
102 (11)
112 (5)
WM probe
Prior Stroop Congruent
101 (10)
100 (13)
Prior Stroop Incongruent
99 (12)
99 (11)
–
–
and were baseline corrected from −200 to 0 msec, relative
to stimulus onset. Any artifacts remaining in the epochs
were detected using a threshold ranging between ±90 mV
over the time period between −500 and 1500 msec. Epochs
that were detected to have artifacts in no more than five
channels were kept and corrected by interpolating the noisy
channels. Remaining epochs with uncorrectable artifacts
were excluded from further analyses. For ERP plotting
and statistical analyses, another low-pass filter (20 Hz,
second-order Butterworth) and baseline correction from
−200 to 0 msec was applied to the averaged ERP data.
Only epochs related to the color discrimination or to the
WM probe were analyzed (Table 1). All ROIs used to ana-
lyze the ERP components described below were defined
based on where these components were greatest collapsed
across all relevant conditions.
ERP Components of Interest
In order to assess how conflict processing may differ as
a function of whether the conflict arises from a clash
between external stimulus features or from a clash
between an external stimulus feature and a representation
held in WM, we focused on two hallmark conflict-related
ERP components. These ERP components are elicited
time-locked to the color discrimination stimulus in each
task (i.e., DO WM Stroop, NonDO WM Stroop, and classic
Stroop) and are best visualized as the difference between
incongruent and congruent trials. Here, for each task, we
show both the raw waves as a function of congruency, as
well as the difference waves (incongruency minus
congruent).
Negative Incongruency Effect
The first conflict-related processing component is an early,
centrally distributed, negative-polarity wave peaking
around 300 to 500 msec, in which activity during incon-
gruent trials are more negative compared to activity dur-
ing congruent trials (herein referred to as the negative
incongruency component or Ninc). The Ninc is observed
in many conflict tasks with similar spatial distribution but
with varying latencies depending on the nature of the con-
flict and task parameters thought to affect the timing of
such conflict processing (Donohue, Appelbaum, McKay,
& Woldorff, 2016; Appelbaum et al., 2014; Donohue,
Liotti, Perez, & Woldorff, 2012; Coderre, Conklin, & van
Heuven, 2011). In general, the Ninc is thought to capture
the generation of conflict (e.g., detection) as it unfolds
during information processing before a behavioral re-
sponse. In a classic Stroop task, this negative incongruency
usually peaks around 450 msec poststimulus onset and is
often referred to as the N450 ( West & Alain, 1999, 2000).
However, because the timing of this incongruency effect
might vary across conditions in our study ( West & Alain,
2000), we will refer to such conflict-related negativity as
a Ninc to avoid specific latency assumptions (Donohue
et al., 2016). Specifically, we expected that conflict
generation/detection would occur earlier during the DO
WM Stroop compared to the other two tasks because of
the cognitive pre-instantiation of color processing from
having to maintain a color word in WM. We also expected
a later onset latency for the Ninc during the delay-period
NonDO WM Stroop as compared to the classic Stroop,
based on the assumption that the (unrelated) WM load
of the NonDO Stroop task would delay stimulus process-
ing (de Fockert, Rees, Frith, & Lavie, 2001). For this study,
we focused our analysis of the Ninc at a cluster of centro-
parietal channels (Cz, CPz, C1p, C2p) typically activated
by this neural response, and that also had the greatest
level of such activity across participants here, irrespective
of task.
Late Positive Complex
The second conflict-related processing component is the
posteriorly distributed late positive complex (LPC), also
known as the conflict slow potential. LPC activity is more
positive for incongruent relative to congruent trials, typi-
cally occurring immediately after the Ninc within the neural
cascade of conflict–control processes during Stroop tasks,
with an onset latency of ∼500 msec and lasting up to 900-
to 1000-msec poststimulus onset (e.g., Donohue et al.,
2016; Larson, Clayson, & Clawson, 2014; Appelbaum
et al., 2009). The LPC has been implicated as an index of
conflict resolution and adaptation processes (Larson et al.,
2014; McKay, van den Berg, & Woldorff, 2017), that is, the
up-regulation of controlled attention in response to
conflict, as demonstrated in studies of the congruency
sequence effect (or “conflict adaptation”), where interfer-
ence effects are reduced following an incongruent as com-
pared to a congruent trial (Donohue et al., 2012; Larson,
Kaufman, & Perlstein, 2009; Egner, 2007; Gratton, Coles, &
Donchin, 1992). In other words, larger LPC during high
conflict, incongruent trials (vs. low conflict, congruent
trials) reflects this up-regulation, which would ramify in
enhanced selective attention following such an event.
We leveraged the LPC to better understand the similarities
Vo et al.
2083
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
1
0
2
0
7
9
1
9
6
2
1
3
5
/
/
j
o
c
n
_
a
_
0
1
7
4
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
and differences in this later stage of conflict processing
during the DO WM Stroop as compared to the other two
tasks. Specifically, based on the same logic as our Ninc pre-
dictions, we expected an earlier LPC during the DO WM
Stroop as compared to the other two tasks and a later
LPC during the NonDO WM Stroop as compared to the
classic Stroop. The ROI for analyzing the LPC was composed
of channels (CPz, Pz, POz, P1, P2, PO1, PO2), again chosen
to be consistent with previous studies and because these
sites showed the strongest LPC activity here across the dif-
ferent conflict tasks.
P300 to the WM Probe
Besides the conflict-related processing components during
the Stroop tasks, we also examined the classic P300 compo-
nent elicited by the WM probe in the two delayed match-to-
sample tasks. The positive-polarity P300, which peaks within
a typical time window between 250 and 500 msec, is a widely
studied ERP component that has often been implicated
for its role as an index of cognitive control during atten-
tion and memory-related operations (Polich, 2007, 2011).
Our specific interest here was to examine how the
cognitive-control processes that occurred during the delay
period affected the processing of the memoranda during
WM retrieval. More explicitly, we expected greater P300
amplitude when the item held in WM had been better
maintained because of less interference from the delay-
period Stroop task (i.e., WM probes following congruent
vs. incongruent Stroop trials; Polich, 2007). The cluster
of interest for the P300 analysis focused on the classic
parietal channels typically used for such activity (Cp1, P1,
P2, Pz).
Statistical Analysis of ERPs
Statistical significance tests for the ERPs of interest were
conducted using permutation statistics for all experimental
contrasts (Maris & Oostenveld, 2007). In these permutation
tests, a null distribution was derived by assigning labels for
the specific conditions (color discrimination: congruent
and incongruent) randomly to the data extracted for each
task from the predefined ROIs. A difference wave (i.e., in-
congruent minus congruent) based on the new data labels
was calculated, and each time point in this difference wave
was tested against a null hypothesis that the difference
should be zero. This permutation procedure was repeated
10,000 times to create a bootstrap distribution of the t sta-
tistic of the null hypothesis for each time point. The t values
obtained from the real difference wave data (i.e., differ-
ence waves calculated using the original, true data labels)
were compared to the bootstrapped t distributions and
were considered significant if they fell within the top or
bottom 2.5% of the null distribution on a timepoint-by-
timepoint basis (corresponding to a two-tailed test at p <
.05). An additional cluster-based correction (maximum
cluster-level statistic) was applied to the final results over
prespecified broad time periods (based on prior reports
for the associated ERP) for each ERP of interest. The
Stroop-stimulus Ninc effects were examined from 200 to
600 msec, the Stroop-stimulus LPC effects from 400 up
to 1600 msec, and the WM Probe P300 effects from 200 to
700 msec. The significant maximal cluster that survived the
correction provided critical information about the onset, off-
set, and duration of a given ERP of interest that was signifi-
cantly different from the null hypothesis based on a
corrected critical alpha value of .05. Thus, reported p values
for all ERP analyses are cluster-based corrected p values.
In addition to deriving the onsets and durations of all ERPs
of interest using permutation statistics, we also conducted
repeated-measures analyses of variance (rmANOVAs) to test
for latency and amplitude differences across tasks and
congruency. To test for amplitude differences, averaged
voltages were extracted from across task-specific time win-
dows that contained the peak amplitudes (100-msec win-
dows for Ninc and 200-msec windows for LPC) and a 3 ×
2 rmANOVA (Task × Congruency) was conducted. To test
for latency differences, an additional factor of Time was in-
cluded, where ERPs extracted from five consecutive time
windows (100-msec windows for Ninc [0–500 msec] and
200-msec windows for LPC [200–1200 msec]) were ana-
lyzed in consideration of task and congruency. All analyses
were corrected for sphericity violations when necessary
using the Greenhouse–Geisser correction.
RESULTS
Color Discrimination (Stroop) Behavioral Results
To examine how color discrimination performance was
impacted by conflicting information that was either exter-
nally presented or held internally in WM, we conducted a
3 (Task: DO WM Stroop, NonDO WM Stroop, classic
Stroop) × 2 (Congruency: congruent vs. incongruent)
rmANOVA on RT (Figure 2A) and mean accuracy
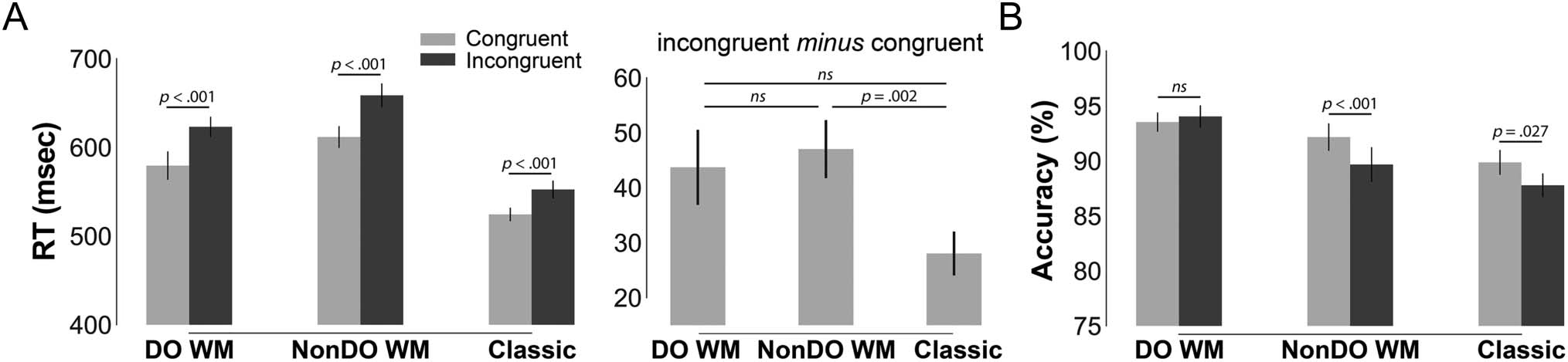
(Figure 2B). Participants were slower to respond during
incongruent Stroop trials than during congruent ones,
demonstrating a main effect of Congruency, F(1, 29) =
147.2, p < .001, η2 = .84. There was also a main effect
of Task on RT, F(2, 29) = 104.9, p < .001, η2 = .78, with
responses being quickest during the classic Stroop and
slowest during the NonDO WM Stroop ( pbonf’s < .001).
These main effects were qualified by a marginally signifi-
cant Task × Congruency interaction effect after correcting
for violations of sphericity, F(1.5, 29) = 3.5, p = .052, η2 =
.12. While congruency effects were comparable between
the DO WM Stroop and classic Stroop ( pbonf = .29), as well
as between the DO WM and NonDO WM Stroop tasks
( pbonf = 1), this effect was significantly greater in the
NonDO WM Stroop compared to classic Stroop ( pbonf =
.002). Given that the stimuli and task demand for the
color discrimination in the NonDO WM Stroop task were
identical to the classic Stroop task, this finding indicates
that the additional WM load in the NonDO WM Stroop
2084
Journal of Cognitive Neuroscience
Volume 33, Number 10
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
1
0
2
0
7
9
1
9
6
2
1
3
5
/
/
j
o
c
n
_
a
_
0
1
7
4
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
1
0
2
0
7
9
1
9
6
2
1
3
5
/
/
j
o
c
n
_
a
_
0
1
7
4
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
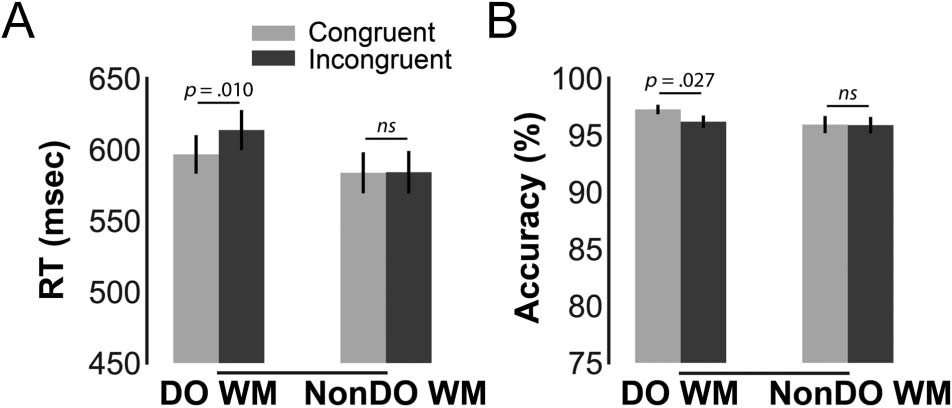
Figure 2. Behavioral results (mean ± standard errors) for the color discrimination (Stroop) responses across the three tasks. (A) Across all tasks,
participants were slower to respond during incongruent trials than congruent trials and slower to respond during the WM tasks than during the
standalone classic Stroop (left). However, this incongruency effect (incongruent minus congruent) was only significantly different between the
NonDO WM and classic tasks (middle). (B) Aside from the DO WM task, participants were more accurate in their responses during congruent trials
relative to incongruent trials.
led to greater RT Stroop interference in that task as com-
pared to its classic Stroop counterpart.
With respect to accuracy, we observed the classic main
effect of Congruency, F(1, 29) = 6.8, p = .015, η2 = .19,
because of a greater number of errors on incongruent
than congruent trials, and a main effect of Task, F(2, 29) =
20.8, p < .001, η2 = .42; pairwise comparisons indicated
that overall accuracy was lowest during the classic Stroop
and highest during the DO WM Stroop ( pbonf’s < .05).
Th ese ma in e ffe ct s w er e qua li f ie d by a T as k ×
Congruency interaction, F(1.4, 29) = 4.2, p = .034, η2 =
.126. This interaction was because of the fact that congru-
ency effects on accuracy were observed in the classic
Stroop (congruent: 89.9 ± 6.1%; incongruent: 87.8 ±
5.9%; t(29) = −2.3, p = .027, d = −0.42) and the NonDO
WM Stroop (congruent: 92.2 ± 6.8%; incongruent: 89.7 ±
8.6%; t(29) = −3.9, p < .001, d = −0.71), but not in the
DO WM Stroop (congruent: 93.5 ± 4.8%; incongruent:
94.0 ± 5.5%; t(29) = 0.55, p = .588, d = 0.10).
In summary, we replicated prior behavioral results
from Kiyonaga and Egner (2014), namely, that the classic
Stroop RT interference effect was emulated during the
DO WM Stroop task. Although participants were generally
slower to respond during the delay-period Stroop tasks
compared to the classic Stroop task (Figure 2A left), the
relative Stroop interference effect was only significantly
different between the NonDO WM Stroop and the classic
Stroop (Figure 2A middle). Controlling for the effect of
WM load, there were comparable Stroop interference
effects between the two delay-period Stroop tasks.
Neural Conflict-detection Processes during
Delay-period Color Discrimination
Time-locked to the onset of the color discrimination
(Stroop) stimulus, a Ninc (negative-polarity incongruency
effect) was observed in all three tasks. The individual
traces for congruent and incongruent trials and the cor-
responding difference waves are depicted in Figure 3,
along with the corresponding topographic maps and
onset/offset latencies defined by nonparametric permuta-
tion testing (see Methods section). The Ninc during the
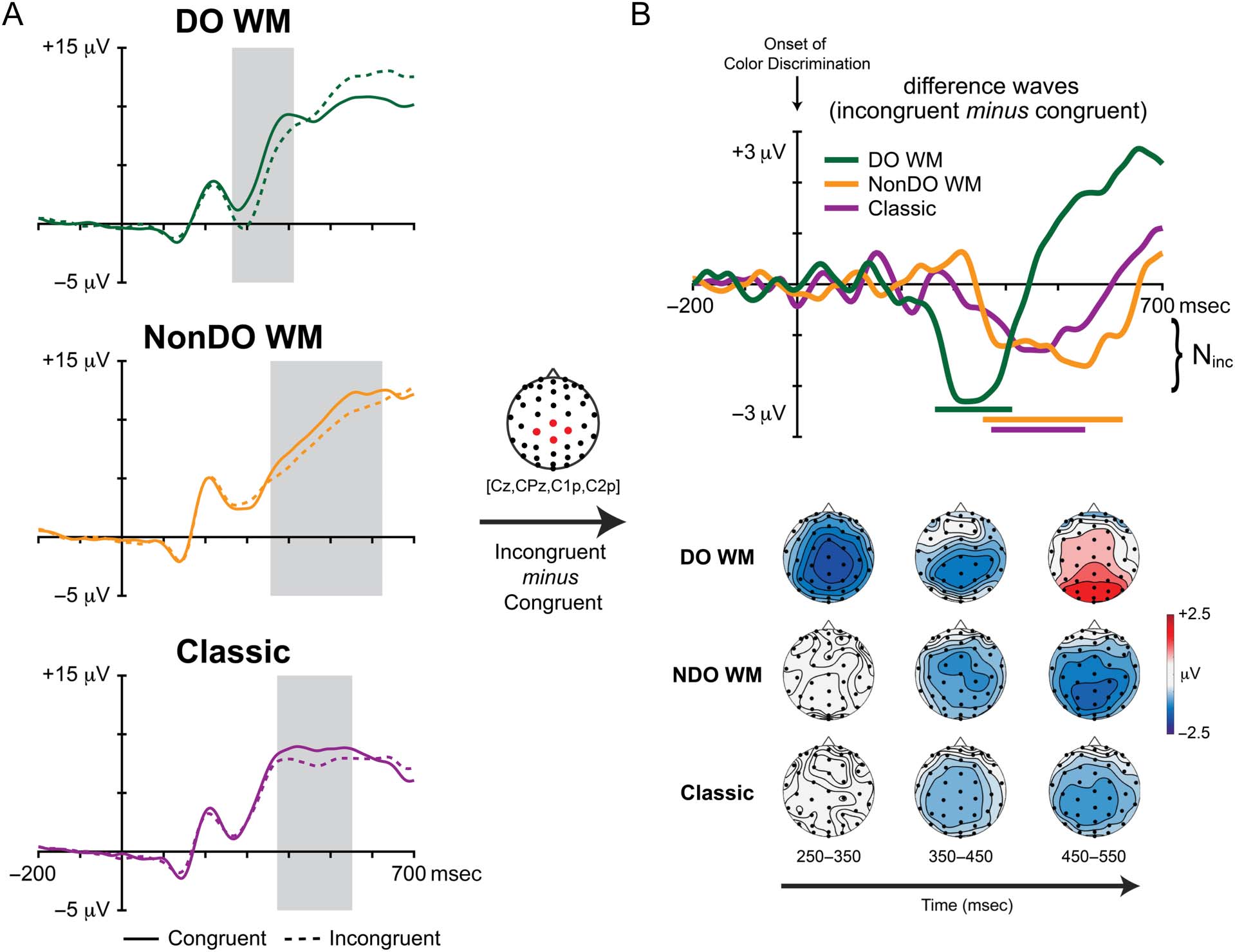
NonDO WM Stroop (onset: 356 msec; offset: 624 msec)
resembled a prototypical Stroop N450 effect and was
visually comparable with the Ninc from the classic Stroop
(onset: 372 msec; offset: 552 msec). However, as hypoth-
esized, the Ninc during the DO WM Stroop had a substan-
tially earlier latency (onset: 264 msec; offset: 412 msec) as
compared to the other two Stroop tasks.
To substantiate the latency differences across tasks, we
conducted a 3 × 5 rmANOVA on the congruency differ-
ence waves (i.e., incongruent minus congruent), with the
factors of Task (DO WM Stroop, NonDO WM Stroop,
classic Stroop) and Time Window (0–100 msec, 100–200,
200–300, 300–400, 400–500). The time window of 0–
500 msec was chosen for these sequential time windows
to sufficiently capture the temporal characteristics of the
Ninc across all the task conditions. There was no main
effect of Task, F(2, 58) = 0.85, p = .43, η2 = .03, but there
was a significant main effect of Time, F(2.96, 85.83) =
17.65, p < .001, η2 = .378, which was expected because
of differences in averaged voltage signal within each time
bin driven by the rise and fall of the Ninc. Importantly,
there was an interaction effect of Task × Time, F(4.7,
136.4) = 11.74, p < .001, η2 = .29, indicating that conflict
detection processes, as proxied by the Ninc, occurred at
different latencies across the tasks.
To probe this interaction further, we conducted three
follow-up 2 × 5 rmANOVAs comparing different pairs of
tasks (DO vs. NonDO, DO vs. classic, NonDO vs. classic)
along the same time factor. There was a significant Task ×
Time interaction when comparing between the DO WM
Stroop and NonDO WM Stroop tasks, F(2.9, 85.2) =
17.5, p < .001, η2 = .15, and between the DO WM
Stroop and classic Stroop tasks, F(3, 87) = 20.42, p <
.001, η2 = .41. However, this interaction was not signifi-
cant for the comparison between the NonDO WM
Stroop and classic Stroop tasks, F(2.6, 76.4) = 0.90, p =
.44, η2 = .03. These results indicate that when the conflict
during the color categorization task was because of a clash
Vo et al.
2085
between an internally held color word and the externally
perceived color swatch (in the DO WM Stroop), conflict
processing occurred substantially earlier than when con-
flict was caused from two clashing external stimulus fea-
tures. In contrast, the conflict-detection onset latencies
were comparable between the NonDO WM Stroop and
classic Stroop. Notably, this meant that the additional
WM load manipulation during the NonDO WM Stroop
did not significantly impact the onset of conflict detection
processes as compared to its standalone counterpart, the
classic Stroop.
Neural Conflict Resolution Processes during
Delay-period Color Discrimination
LPC activity was also analyzed time-locked to the onset of
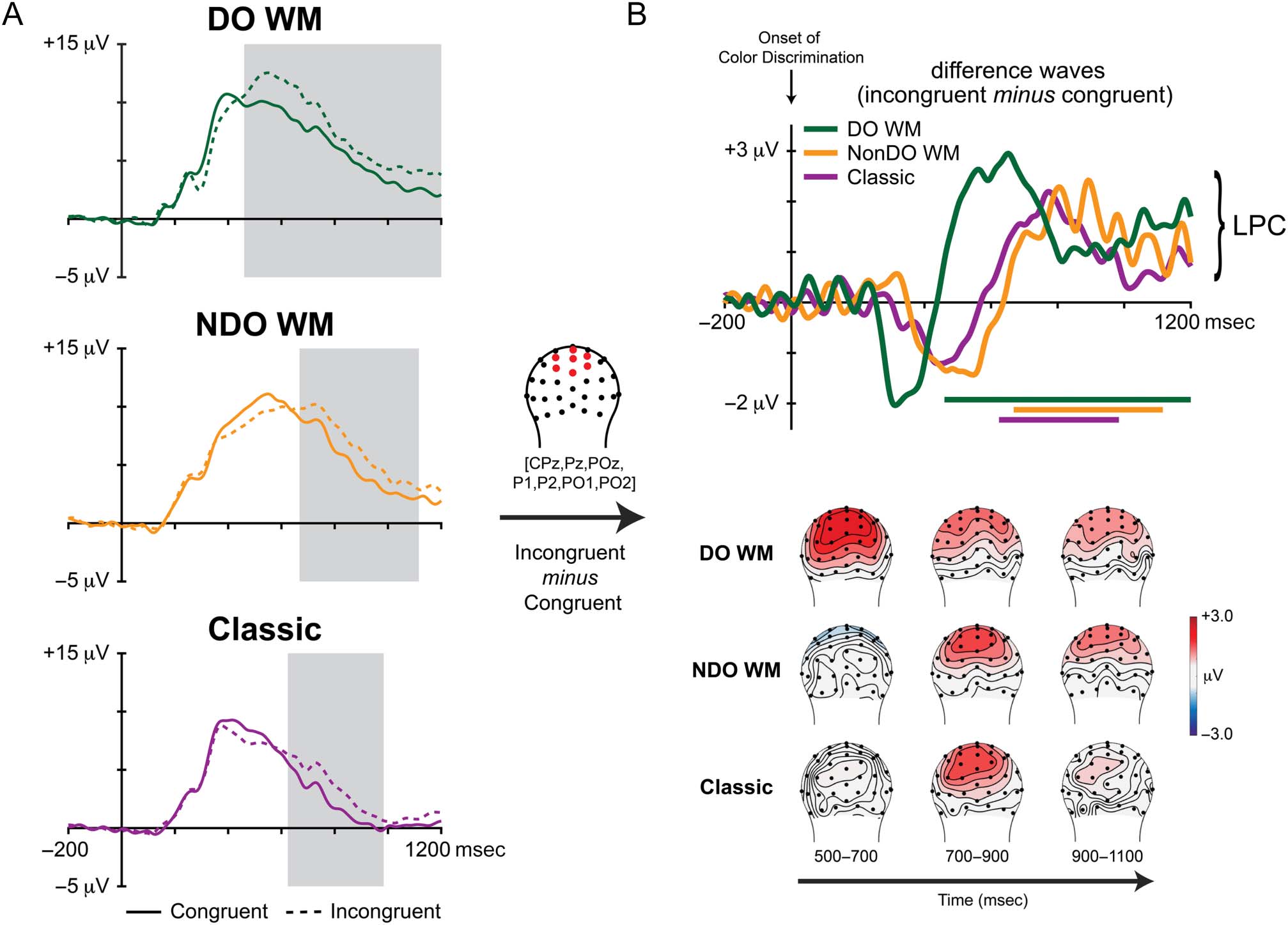
the color discrimination Stroop stimulus (Figure 4). LPC
activity was detected in all three tasks, with the earliest
latency occurring during the DO WM Stroop (onset:
460 msec; offset: 1200 msec) as compared to both the
NonDO WM Stroop (onset: 668 msec; offset: 1116 msec)
and the classic Stroop (onset: 624 msec; offset: 984 msec).
Note that although the LPC was significant for the longest
period of time in the DO WM condition, all three LPC com-
ponents resolved well after average RT to the Stroop stim-
uli. Thus, it is unlikely that any differences in LPC duration
would have affected the observed RTs.
Similar to the analyses for the Ninc, the latency differ-
ences across tasks for the LPC were substantiated
through a 3 × 4 rmANOVA on the difference waves, with
the factors of Task (DO WM Stroop, NonDO WM Stroop,
classic Stroop) and Time Window (200–400 msec, 400–
600, 600–800, 800–1000). There was no main effect of
Task, F(2, 58) = 2.54, p = .087, η2 = .081, but there
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
1
0
2
0
7
9
1
9
6
2
1
3
5
/
/
j
o
c
n
_
a
_
0
1
7
4
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. ERP activity for conflict detection (Ninc) during color discrimination. Plotted ERP waveforms are averaged from a cluster of four
centro-parietal channels (Cz, CPz, C1p, C2p). (A) Raw ERP waveforms for congruent and incongruent trials, along with associated windows of
significant differences between congruent and incongruent traces, as identified by permutation statistics (pcorrected < .05). Cluster correction was
done between 200 and 600 msec. (B) Difference waves (incongruent minus congruent, top) were calculated for each condition and plotted with the
same cluster-corrected windows of significance along with topographic maps of these differences waves (bottom). Conflict detection, as measured by
the negative incongruency component (Ninc), occurred much earlier for color discrimination during the DO task, but the degree, or strength, of
conflict detection did not differ across tasks. Because of the latency shift during the DO task, part of the LPC was also captured in the superior-scalp
topographic map associated with this task, although this wave tends to be largest over more posterior sites (see Figure 4).
2086
Journal of Cognitive Neuroscience
Volume 33, Number 10
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
1
0
2
0
7
9
1
9
6
2
1
3
5
/
/
j
o
c
n
_
a
_
0
1
7
4
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. ERP activity for conflict resolution (LPC) during color discrimination. Plotted ERP waveforms are averaged from a cluster of seven posterior
channels (CPz, Pz, POz, P1, P2, PO1, and PO2). (A) Raw ERP waveforms for congruent and incongruent trials for each task with associated windows
of significant differences between the trial conditions ( pcorrected < .05). Cluster correction for this set of permutation statistics was done between
400 and 1600 msec. (B) Difference waves (incongruent minus congruent) with the same windows of cluster-corrected significance (top) and
associated topographic maps for each task (bottom). Conflict resolution following color discrimination, as measured by the late positivity complex
(LPC), occurred earliest during the DO WM Stroop and slowest during the NonDO WM Stroop. The degree, or intensity, of this resolution process
was greater during the DO WM task only.
was an expected main effect of Time, F(2.04, 59.12) =
45.95, p < .001, η2 = .61. Most importantly, we observed
a significant interaction of Task × Time, F(4.45, 129.1) =
22.51, p < .001, η2 = .44, indicating that LPC onset laten-
cies differed across the tasks. We conducted follow-up
tests to further probe the Task × Time interaction, com-
paring all pairings of tasks along the same time window
factor. All three follow-ups yielded significant interaction
effects between task and time, namely, that conflict res-
olution processes started earlier for both DO WM Stroop,
F(3, 87) = 20.41, p < .001, η2 = .41, and classic Stroop,
F(3, 87) = 5.45, p = .002, η2 = .16, as compared to the
NonDO WM Stroop; and earliest during DO WM Stroop
when comparing between the DO and NonDO WM
delay-period Stroops, F(3, 87) = 35.18, p < .001,η2 =
.55. Extra WM load during the NonDO WM Stroop delayed
the onset of LPC activity during this task as compared to
the classic Stroop, unlike the Ninc where this load manip-
ulation had no effect.
WM Probe Behavioral Results
To evaluate how WM maintenance was affected when at-
tentional resources were diverted to the color discrimina-
tion task during the delay period, performance measures
of RTs and mean accuracy for WM probe responses were
analyzed for the DO and NonDO Stroop tasks. For these
analyses, only responses following correct color discrim-
ination responses were analyzed to ensure that conflict
had been successfully resolved during the delay period.
First, we conducted a 2 × 2 rmANOVA ([Task: DO WM
probe vs. NonDO WM probe] × [Prior color discrimina-
tion (Stroop) Congruency: congruent vs. incongruent])
on the RTs for probe responses (Figure 5A). Main effects
of task and prior congruency indicated that responses
during DO WM probe were slower than that of NonDO
WM probe, F(1, 29) = 14.04, p < .001, η2 = .326, as well
as during WM probes that followed an incongruent
delay-period Stroop stimulus compared to those follow-
ing a congruent one, F(1, 29) = 5.76, p = .023, η2 = .166.
Vo et al.
2087
and delay-period perceptual task overlapped domain-
wise. Although there was an equivalent WM load main-
tained during the NonDO WM task, WM maintenance
was not affected by conflict during the delay period when
there was no domain overlap.
Neural Effect of Delay-period Stroop Conflict on
Subsequent WM Probe P300
To understand how conflict during the delay period influ-
enced retrieval during the WM probe, we analyzed EEG
data time-locked to the DO and NonDO WM probes
based on whether the associated preceding delay-period
Stroop trial was congruent versus incongruent. To exam-
ine whether there were differences in evoked P300 activity
across the four individual contrasts (Figure 6A), we con-
ducted a 2 × 2 × 5 rmANOVA with factors of Task (DO
and NonDO) and Prior Stroop Congruency (congruent
and incongruent) on averaged ERP data within consecu-
tive 100-msec time windows between 200 and 700 msec
(time window: 200–300 msec, 300–400, 400–500, 500–
600, 600–700). This time window was chosen to capture
the entire duration in which the P300 occurred in the
probe response. The difference waves (congruent minus
incongruent) are displayed in Figure 6B.
These analyses showed that there was a main effect of
Task, with P300 amplitude being greater in response to the
DO WM probes compared to the NonDO WM probes, F(1,
29) = 6.77, p = .014, η2 = .19. We also observed a main
effect of Prior Congruency, as probes that followed con-
gruent delay-period Stroop trials had greater P300 ampli-
tude than probes that followed incongruent Stroop trials,
F(1, 29) = 9.55, p = .004, η2 = .25. There was also an ex-
pected main effect of Time Window, F(2.32, 67.33) =
23.86, p < .001, η2 = .45, reflecting the rise, peak, and fall
in P300 amplitude over time. These main effects were qual-
ified by a significant interaction between Task and Time
Window, F(2.13, 67.76) = 6.77, p = .002, η2 = .19, and,
most importantly, by a significant interaction between
Task and Prior Congruency, F(1, 29) = 10.92, p = .003,
Figure 5. Behavioral results (mean ± standard errors) for WM probe
responses during the DO and NonDO WM tasks. (A) Participants were
slower to respond during probe trials following delay periods with
incongruent color discrimination than with congruent, but only during
the DO WM task. (B) Likewise, participants were more accurate in their
WM probe responses following congruent color discrimination than
following incongruent, but, again, only for the DO WM task.
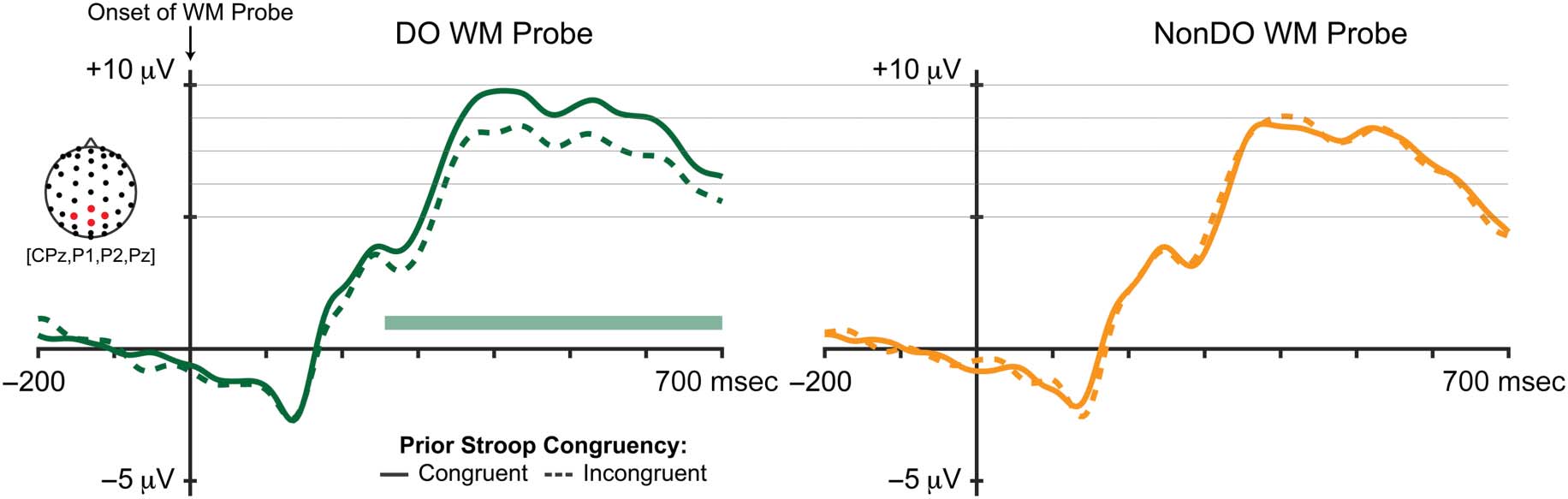
Importantly, these main effects were qualified by an inter-
action between Task and Congruency, F(1, 29) = 4.93, p =
.034, η2 = .145: The congruency effect on probe RT was
greater for DO WM probes, t(29) = 2.738, p = .010, d =
0.500, than for NonDO WM probes, t(29) = −0.049, p =
.961, d = −0.009. In other words, conflict during the
delay-period Stroop resulted in a delay in retrieving the
memorandum and matching it against the WM probe
(cf. Kiyonaga & Egner, 2014), but only when that conflict
involved a clash between the external stimulus and the
WM representation in the same featural domain (i.e., do-
main overlapping).
Running the same 2 × 2 rmANOVA on the accuracy of
the valid probe responses (Figure 5B) did not yield any
significant main or interaction effects of the aforemen-
tioned factors (all ps > .05). However, there was a simple
main effect of Congruency on the amount of errors made
during DO WM probes, with greater errors following
incongruent versus congruent delay-period Stroop trials
(F = 5.39, p = .027).
In summary, conflict during the delay-period Stroop
resulted in poorer WM maintenance, making retrieval
harder during WM probe, but only when the memoranda
Figure 6. Raw ERP waveforms during WM probe. Modulation by prior Stroop congruency was only significant during the DO WM task (period of
significance marked by a green bar, pcorr < .05). P300 activity was averaged across four centro-posterior channels (CPz, P1, P2, and Pz). Guide lines
(light gray) are included to allow for easier comparisons between tasks.
2088
Journal of Cognitive Neuroscience
Volume 33, Number 10
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
1
0
2
0
7
9
1
9
6
2
1
3
5
/
/
j
o
c
n
_
a
_
0
1
7
4
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
η2 = .27. This was because of the fact that the significant
effect of prior congruency on probe P300 was driven by
large differences for the DO WM probes ( p < .001), which
were not observed in response NonDO WM probes ( p =
.89). There were no other interaction effects. In summary,
P300 amplitude was greater during WM probes following
congruent (relative to incongruent) Stroop trials, but only
when the memoranda overlapped representationally with
the delay-period task demands. There was no such facilita-
tion for NonDO WM probes following congruent versus
incongruent Stroop trials.
DISCUSSION
In this study, we investigated how conflict stemming
from either clashing external stimulus features or be-
tween an external stimulus and an item held in WM
may differ in terms of the underlying neural cascade of
conflict processes. Our behavioral results replicated
those of Kiyonaga and Egner (2014), showing that re-
sponses during a delay-period color discrimination task
are substantially slowed when an incongruent color word
is being maintained in WM, and this Stroop effect during
the WM-variant Stroop (DO WM Stroop) was comparable
to that during the classic Stroop. However, the Stroop
interference during the WM-control task (NonDO WM
Stroop), a new control condition included in this study,
was more pronounced compared to its standalone classic
Stroop counterpart. This is presumably because of in-
creased cognitive load from having to maintain a numer-
ical representation in WM while performing the color
discrimination. Most importantly, the current demon-
strated that behaviorally equivalent effects between the
classic Stroop and its WM variant were underpinned by
a divergent neural cascade of conflict–control processes,
namely, that the onset latencies for conflict detection
(Ninc) and conflict resolution (LPC) processes occurred
substantially earlier when the distractor (i.e., color word)
was held in WM before the color discrimination task
(DO WM Stroop) as compared to when the distractor
was visually perceived at the same time during the color
discrimination task (classic Stroop, as well as NonDO
WM Stroop). In addition, WM maintenance during the
dual-task paradigms was affected only when the delay-
period task demands overlapped domain-wise (i.e., se-
mantically) with the memoranda (DO WM task) but
not when there was no overlap (NonDO WM task).
We further discuss the implications of these results
below.
Differences in the Timing of
Conflict–Control Processes
By and large, the neural cascades of conflict control pro-
cesses were very similar during Stroop performance
when the color word distractor was visually presented
at the same time as the color patch during the NonDO
WM Stroop and the classic Stroop. However, there were
two notable differences: (1) The durations of conflict de-
tection and resolutions processes, as indexed by the Ninc
and LPC, respectively, were extended during the NonDO
WM Stroop, and (2) although the onset latency of the con-
flict detection process (reflected by the Ninc) did not differ
between the NonDO and classic Stroop tasks, the onset
latency of the conflict resolution processes (LPC) was
slightly, but significantly, delayed in the NonDO WM
Stroop. We posit that this pattern of activity differences
is attributable to the additional WM load present in the
NonDO WM task, in the form of an additional feature di-
mension to attend to, and that may have resulted in some
delay and/or inefficiency in conflict processing during the
delay period in the NonDO Stroop task as compared to
the classic Stroop task. Notably, however, WM probe per-
formance and associated neural activity (P300) did not dif-
fer as a function of delay-period Stroop congruency
during the NonDO WM probe despite the increased cog-
nitive demands throughout the NonDO WM task.
Of greater interest was the comparison of the neural
cascades of conflict–control processes between the
WM-variant Stroop and the classic Stroop, to characterize
whether and how processing would differ between the
case of a clash between a WM representation (i.e., color
word) and an external item (i.e., color patch) and the
hallmark case of conflict between two clashing external
stimulus features in the classic Stroop paradigm. From
our results, it is clear that the neural cascades of conflict
processing between the WM-variant Stroop and the clas-
sic Stroop differed substantially, as reflected by much ear-
lier onset latencies for the Ninc (conflict detection) and
LPC (conflict resolution) neural processes during the
WM-variant Stroop. It is important to note, however, that
these earlier onset latencies for conflict detection in the
WM-variant Stroop did not always correspond with smaller
congruency effects behaviorally. Namely, the DO WM
Stroop was associated with smaller (nonsignificant)
accuracy congruency effects, but was not associated with
smaller RT congruency effects compared to the classic
Stroop. Rather, RT congruency effects were only signifi-
cantly larger in the NonDo WM Stroop compared to the
classic Stroop. This pattern of results suggests that, in the
current paradigm, RT congruency effects were more
strongly affected by WM load differences between tasks,
rather than the conflict-detection onset latency.
The most likely explanation for the substantially earlier
onset latency for the Ninc and subsequently the LPC during
WM-variant Stroop falls in line with current theories of WM
(D’Esposito & Postle, 2015), where maintaining a color
word in WM would be assumed to result in activating the
perceptual and semantic long-term memory representa-
tion of that color, and that representations activated by
the WM process are the same as those that are activated
by external, perceptual inputs. Since in the WM-variant
Stroop, those representations are already active when
the external stimulus is presented (i.e., at the onset of
Vo et al.
2089
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
1
0
2
0
7
9
1
9
6
2
1
3
5
/
/
j
o
c
n
_
a
_
0
1
7
4
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
the delay-period color swatch), conflict will occur more
rapidly—and thus be detected and resolved sooner—in
this condition compared to conditions where no such
color representation was pre-activated (NonDO WM
Stroop and classic Stroop).
Alternatively, it is also possible that Ninc and LPC activ-
ity could reflect different cognitive processes in the DO
WM Stroop than in the classic Stroop conditions, rather
than reflecting earlier onset of the same or similar con-
flict detection and resolution processes. That is, maybe
these earlier onsetting effects reflect WM maintenance
conflict in the face of interference from the Stroop color
patch, with that WM conflict occurring only in the DO
WM Stroop condition. Although we cannot completely
discount this possibility, we consider it to be highly un-
likely for several reasons. First, the RT congruency effects
were comparable between the DO WM Stroop and classic
Stroop tasks, suggesting similar levels of task interference
during the Stroop task completion. Second, the Ninc and
LPC exhibited similar topographies in all three of the
tasks, with both just occurring earlier in time in the DO
WM Stroop task, suggesting relatively similar underlying
neural processes. Thirdly, prior work from our group
(Appelbaum et al., 2009) investigating how varying the
temporal separation of relevant (e.g., color swatch) and
irrelevant (e.g., color word) Stroop stimulus features
(by up to ±200 msec), when there was no WM compo-
nent to the task at all, showed analogous acceleration of
the Sroop neural processing of conflict. More specifically,
when the irrelevant stimulus in that study came earlier
than the relevant one by either 100 or 200 msec (relative
to occurring simultaneously), both the Ninc and the LPC
occurred earlier in time, reflecting earlier incongruency
detection and processing, analogous to the current study.
Appelbaum et al.’s (2009) results thus show that latency
shifts in conflict–control processes can be observed even
when the irrelevant stimulus dimension does not need to
be stored in WM to be retrieved later on and when it was
not associated with an alternate response. Accordingly, it
seems fairly unlikely that the earlier Ninc and LPC effects
observed in the DO WM task in the current study reflect
activity associated with different cognitive processes re-
sulting from additional interference from the WM task,
and more likely that they reflect an acceleration of the
same or similar conflict–control processes observed in
the other conditions. Given that the maximum SOA
manipulation between Stroop components was only
200 msec in this prior study, however, the color word
was not considered to be held in WM but rather held
in sensory memory, which is still visual attention by
nature. As such, by having a color word held in WM in
the case of our WM-variant Stroop task, the considerably
earlier resultant instantiation of color representation in
WM would be expected to lead to a still greater Ninc onset
latency shift, as observed here (∼200 msec earlier).
qualitatively similar across tasks (albeit shifted in onset
latencies). At face value, this suggests that the underlying
conflict detection and resolution processes may also be
equivalent between these conditions, but just occurring
at shifted times. However, to draw stronger conclusions
regarding this equivalence would require additional re-
search. For instance, one could probe whether conflict
adaptation processes would generalize from the classic
Stroop task to the WM Stroop task and vice versa
(Egner, 2008).
Facilitation of P300 in Response to the WM Probe
The P300 activity in response to the WM probe was facili-
tated in the DO WM task following congruent compared to
incongruent Stroop trials. This pattern of P300 activity dif-
ferences during the processing of the DO WM probes as a
function of prior Stroop congruency mirrored behavioral
performance in the WM task, where responses to WM
probes were slower following incongruent Stroop trials
(which was also observed in Kiyonaga & Egner, 2014).
However, the analogous P300 activity in response to the
NonDO WM probes was not modulated by prior Stroop
congruency, and neither were RTs in that task. In fact,
following the incongruent delay-period stimuli, the P300
amplitudes were similar for all WM probes for the
NonDO WM task as well as for WM probes for the DO task.
We speculate that this pattern of neural activity could be
explained by an incidental beneficial effect on the me-
moranda on congruent trials in the DO WM Stroop.
Specifically, attending and responding to a congruent ex-
ternal stimulus may have enhanced the activation of the
(identical) internally maintained representation of that
stimulus (equivalent to attentional “refreshing” of the
memorandum; cf. Barrouillet & Camos, 2012). Such re-
freshing during the delay period would have strengthened
the WM representation, thereby leading to faster retrieval
and greater P300 responses in response to the WM probe
trials that followed congruent color discrimination trials in
the DO WM task as compared to the other probe trial con-
ditions. In contrast, in the NonDO WM task, the congruent
external stimulus was unrelated to the item being held in
WM, thereby not enabling such refreshing and, thus, not
learning to any effects on RT or the P300 to the WM probe
following congruent versus incongruent trials in that task.
Lastly, it isimportant tokeep inmind that these RT and P300
effects were in response to the WM probe, which occurred
substantially later than the Ninc and LPC incongruency-
reflecting components elicited by the Stroop stimulus.
Thus, it is unlikely that probe-related responses were a cause
of the differential timing of those earlier Stroop-stimulus
incongruency effects as a function of condition.
Conclusions
The topography and amplitudes of the conflict-related
ERP components investigated in this study were
In summary, this study enhanced our understanding of the
interplay between internally attended ( WM) information
2090
Journal of Cognitive Neuroscience
Volume 33, Number 10
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
1
0
2
0
7
9
1
9
6
2
1
3
5
/
/
j
o
c
n
_
a
_
0
1
7
4
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
and the processing of externally presented stimuli. We rep-
licated prior behavioral results from Kiyonaga and Egner
(2014)—showing that a mismatch between an activated
color word representation in WM and an externally
perceived color patch during a WM-variant Stroop task
can produce behavioral Stroop interference effects compa-
rable to that of a classic Stroop task. Importantly, however,
we demonstrate that the cascade of underlying neural pro-
cessing differs between these two interference effects. In
particular, we show that neural conflict detection and reso-
lution processes occur substantially earlier in the WM
Stroop than its classic Stroop counterpart. This finding
supports the fundamental assumption that WM representa-
tions overlap with representations activated by external
stimuli (Kiyonaga & Egner, 2013), such that a pre-activated
color representation in WM results in earlier conflict when
a mismatching external stimulus is presented than when
two incongruent color cues are presented externally. In
further support of this conclusion, we also showed that
WM probe performance and neural processing were mod-
ulated by delay-period conflict only when that conflict in-
volved the WM representation (in the DO WM Stroop) but
not when it was unrelated to the information being held in
WM (the NonDO Stroop). In summary, by revealing new
insights into the timing of conflict–control processes in-
volving WM versus externally activated representations,
the present findings support recent theories of shared rep-
resentations and control processes between WM and per-
ceptual attention.
Acknowledgments
This work was supported by National Institutes of Health grant
R01/R56-NS051048 to M. G. W.
Reprint requests should be sent to Marty G. Woldorff, Center
for Cognitive Neuroscience, Duke University, Box 90999,
Durham, NC 27708, or via e-mail: woldorff@duke.edu.
Funding Information
Marty G. Woldorff, National Institute of Neurological
Disorders and Stroke (https://dx.doi.org/10.13039
/100000065), grant number: R01/R56-NS051048.
Diversity in Citation Practices
A retrospective analysis of the citations in every article pub-
lished in this journal from 2010 to 2020 has revealed a per-
sistent pattern of gender imbalance: Although the
proportions of authorship teams (categorized by estimated
gender identification of first author/last author) publishing
in the Journal of Cognitive Neuroscience ( JoCN) during
this period were M(an)/M = .408, W(oman)/M = .335,
M/ W = .108, and W/ W = .149, the comparable proportions
for the articles that these authorship teams cited were
M/M = .579, W/M = .243, M/ W = .102, and W/ W =
.076 (Fulvio et al., JoCN, 33:1, pp. 3–7). Consequently,
JoCN encourages all authors to consider gender balance
explicitly when selecting which articles to cite and gives
them the opportunity to report their article’s gender cita-
tion balance.
REFERENCES
Appelbaum, L. G., Boehler, C. N., Davis, L. A., Won, R. J., &
Woldorff, M. G. (2014). The dynamics of proactive and
reactive cognitive control processes in the human brain.
Journal of Cognitive Neuroscience, 26, 1021–1038. https://
doi.org/10.1162/jocn_a_00542, PubMed: 24345171
Appelbaum, L. G., Boehler, C. N., Won, R., Davis, L., & Woldorff,
M. G. (2012). Strategic allocation of attention reduces
temporally predictable stimulus conflict. Journal of Cognitive
Neuroscience, 24, 1834–1848. https://doi.org/10.1162/jocn_a
_00209, PubMed: 22360623
Appelbaum, L. G., Meyerhoff, K. L., & Woldorff, M. G. (2009).
Priming and backward influences in the human brain:
Processing interactions during the Stroop Interference Effect.
Cerebral Cortex, 19, 2508–2521. https://doi.org/10.1093
/cercor/bhp036, PubMed: 19321654
Awh, E., & Jonides, J. (2001). Overlapping mechanisms of
attention and spatial working memory. Trends in Cognitive
Sciences, 5, 119–126. https://doi.org/10.1016/S1364-6613(00)
01593-X, PubMed: 11239812
Baddeley, A. D. (1992). Working memory. Science, 255,
556–559. https://doi.org/10.1126/science.1736359, PubMed:
1736359
Baddeley, A. D., & Hitch, G. (1974). Working memory. In G. H.
Bower (Ed.), Psychology of learning and motivation (Vol. 8,
pp. 47–89). Academic Press. https://doi.org/10.1016/S0079
-7421(08)60452-1
Barrouillet, P., & Camos, V. (2012). As time goes by: Temporal
constraints in working memory. Current Directions in
Psychological Science, 21, 413–419. https://doi.org/10.1177
/0963721412459513
Broadbent, D. E. (1958). Perception and communication.
Elsevier. https://doi.org/10.1016/C2013-0-08164-9
Bugg, J., & Crump, M. (2012). In support of a distinction
between voluntary and stimulus-driven control: A review of
the literature on proportion congruent effects. Frontiers in
Psychology, 3, 367. https://doi.org/10.3389/fpsyg.2012.00367
Coderre, E., Conklin, K., & van Heuven, W. J. B. (2011).
Electrophysiological measures of conflict detection and
resolution in the Stroop task. Brain Research, 1413, 51–59.
https://doi.org/10.1016/j.brainres.2011.07.017, PubMed:
21840503
D’Esposito, M., & Postle, B. R. (2015). The cognitive
neuroscience of working memory. Annual Review of
Psychology, 66, 115–142. https://doi.org/10.1146/annurev
-psych-010814-015031, PubMed: 25251486
de Fockert, J. W., Rees, G., Frith, C. D., & Lavie, N. (2001). The
role of working memory in visual selective attention. Science,
291, 1803–1806. https://doi.org/10.1126/science.1056496,
PubMed: 11230699
Delorme, A., & Makeig, S. (2004). EEGLAB: An open source
toolbox for analysis of single-trial EEG dynamics including
independent component analysis. Journal of Neuroscience
Methods, 134, 9–21. https://doi.org/10.1016/j.jneumeth.2003
.10.009, PubMed: 15102499
Donohue, S. E., Appelbaum, L. G., McKay, C. C., & Woldorff,
M. G. (2016). The neural dynamics of stimulus and response
conflict processing as a function of response complexity and
task demands. Neuropsychologia, 84, 14–28. https://doi.org
/10.1016/j.neuropsychologia.2016.01.035, PubMed: 26827917
Vo et al.
2091
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
1
0
2
0
7
9
1
9
6
2
1
3
5
/
/
j
o
c
n
_
a
_
0
1
7
4
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Donohue, S. E., Liotti, M., Perez, R., & Woldorff, M. G. (2012). Is
conflict monitoring supramodal? Spatiotemporal dynamics
of cognitive control processes in an auditory Stroop task.
Cognitive, Affective, & Behavioral Neuroscience, 12, 1–15.
https://doi.org/10.3758/s13415-011-0060-z, PubMed: 21964643
Egner, T. (2007). Congruency sequence effects and cognitive
control. Cognitive, Affective & Behavioral Neuroscience, 7,
380–390. https://doi.org/10.3758/CABN.7.4.380
Egner, T. (2008). Multiple conflict-driven control mechanisms in
the human brain. Trends in Cognitive Sciences, 12, 374–380.
https://doi.org/10.1016/j.tics.2008.07.001, PubMed: 18760657
Egner, T., & Hirsch, J. (2005). Cognitive control mechanisms
resolve conflict through cortical amplification of task-relevant
information. Nature Neuroscience, 8, 1784–1790. https://doi
.org/10.1038/nn1594, PubMed: 16286928
Gazzaley, A., & Nobre, A. C. (2012). Top–down modulation:
Bridging selective attention and working memory. Trends in
Cognitive Sciences, 16, 129–135. https://doi.org/10.1016/j.tics
.2011.11.014, PubMed: 22209601
Gratton, G., Coles, M. G., & Donchin, E. (1992). Optimizing the
use of information: Strategic control of activation of responses.
Journal of Experimental Psychology. General, 121, 480–506.
https://doi.org/10.1037/0096-3445.121.4.480, PubMed:
1431740
Kerns, J. G., Cohen, J. D., MacDonald, A. W., Cho, R. Y., Stenger,
V. A., & Carter, C. S. (2004). Anterior cingulate conflict
monitoring and adjustments in control. Science, 303,
1023–1026. https://doi.org/10.1126/science.1089910,
PubMed: 14963333
Kiyonaga, A., & Egner, T. (2013). Working memory as internal
attention: Toward an integrative account of internal and
external selection processes. Psychonomic Bulletin &
Review, 20, 228–242. https://doi.org/10.3758/s13423-012-0359
-y, PubMed: 23233157
Kiyonaga, A., & Egner, T. (2014). The working memory Stroop
effect: When internal representations clash with external
stimuli. Psychological Science, 25, 1619–1629. https://doi.org
/10.1177/0956797614536739, PubMed: 24958685
Larson, M. J., Clayson, P. E., & Clawson, A. (2014). Making sense
of all the conflict: A theoretical review and critique of conflict-
related ERPs. International Journal of Psychophysiology,
93, 283–297. https://doi.org/10.1016/j.ijpsycho.2014.06.007,
PubMed: 24950132
Larson, M. J., Kaufman, D. A. S., & Perlstein, W. M. (2009).
Neural time course of conflict adaptation effects on the
Stroop task. Neuropsychologia, 47, 663–670. https://doi.org
/10.1016/j.neuropsychologia.2008.11.013, PubMed: 19071142
MacDonald, A. W., Cohen, J. D., Stenger, V. A., & Carter, C. S.
(2000). Dissociating the role of the dorsolateral prefrontal
and anterior cingulate cortex in cognitive control. Science,
288, 1835. https://doi.org/10.1126/science.288.5472.1835,
PubMed: 10846167
MacLeod, C. M. (1991). Half a century of research on the Stroop
effect: An integrative review. Psychological Bulletin, 109,
163–203. https://doi.org/10.1037/0033-2909.109.2.163, PubMed:
2034749
Maris, E., & Oostenveld, R. (2007). Nonparametric statistical
testing of EEG- and MEG-data. Journal of Neuroscience
Methods, 164, 177–190. https://doi.org/10.1016/j.jneumeth
.2007.03.024, PubMed: 17517438
McKay, C. C., van den Berg, B., & Woldorff, M. G. (2017). Neural
cascade of conflict processing: Not just time-on-task.
Neuropsychologia, 96, 184–191. https://doi.org/10.1016/j
.neuropsychologia.2016.12.022, PubMed: 28017818
Oostenveld, R., Fries, P., Maris, E., & Schoffelen, J.-M. (2011).
FieldTrip: Open source software for advanced analysis
of MEG, EEG, and invasive electrophysiological data.
Computational Intelligence and Neuroscience, 2011,
156869. https://doi.org/10.1155/2011/156869, PubMed:
21253357
Polich, J. (2007). Updating P300: An integrative theory of P3a
and P3b. Clinical Neurophysiology, 118, 2128–2148. https://
doi.org/10.1016/j.clinph.2007.04.019, PubMed: 17573239
Polich, J. (2011). Neuropsychology of P300. In E. S. Kappenman
& S. J. Luck (Eds.), The Oxford handbook of event-related
potential components. Oxford: Oxford University Press.
https://doi.org/10.1093/oxfordhb/9780195374148.013.0089
Postle, B. R. (2006). Working memory as an emergent property
of the mind and brain. Neuroscience, 139, 23–38. https://doi
.org/10.1016/j.neuroscience.2005.06.005, PubMed: 16324795
Stroop, J. R. (1935). Studies of interference in serial verbal
reactions. Journal of Experimental Psychology, 18, 643–662.
https://doi.org/10.1037/h0054651
Treisman, A. M. (1960). Contextual cues in selective listening.
Quarterly Journal of Experimental Psychology, 12, 242–248.
https://doi.org/10.1080/17470216008416732
West, R., & Alain, C. (1999). Event-related neural activity
associated with the Stroop task. Cognitive Brain Research,
8, 157–164. https://doi.org/10.1016/S0926-6410, PubMed:
10407204
West, R., & Alain, C. (2000). Effects of task context and
fluctuations of attention on neural activity supporting
performance of the Stroop task. Brain Research, 873,
102–111. https://doi.org/10.1016/S0006-8993(00)02530-0,
PubMed: 10915815
Woldorff, M. G., Liotti, M., Seabolt, M., Busse, L., Lancaster, J. L.,
& Fox, P. T. (2002). The temporal dynamics of the effects in
occipital cortex of visual-spatial selective attention. Brain
Research. Cognitive Brain Research, 15, 1–15. https://doi.org
/10.1016/S0926-6410(02)00212-4, PubMed: 12433379
2092
Journal of Cognitive Neuroscience
Volume 33, Number 10
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
1
0
2
0
7
9
1
9
6
2
1
3
5
/
/
j
o
c
n
_
a
_
0
1
7
4
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3