METHODS
Nonparametric test for connectivity detection in
multivariate autoregressive networks and
application to multiunit activity data
M. Gilson
1
, A. Tauste Campo
1,2
, X. Chen
3
3
, A. Thiele
, and G. Deco
1,4
1Computational Neuroscience Group, Departament de Tecnologies de la Informació i les Comunicacions, Universitat
Pompeu Fabra, Barcelona, Spain
2Epilepsy Monitoring Unit, Department of Neurology, Hospital del Mar Medical Research Institute, Barcelona, Spain
Institute of Neuroscience, Newcastle University, Newcastle upon Tyne, United Kingdom
4Institució Catalana de Recerca i Estudis Avançats, Barcelona, Spain
3
a n o p e n a c c e s s
j o u r n a l
Keywords: Network connectivity detection, Nonparametric significance method, Multivariate
autoregressive process, Granger causality, Multiunit activity
ABSTRACT
Directed connectivity inference has become a cornerstone in neuroscience to analyze
multivariate data from neuroimaging and electrophysiological techniques. Here we propose
a nonparametric significance method to test the nonzero values of multivariate autoregressive
model to infer interactions in recurrent networks. We use random permutations or circular
shifts of the original time series to generate the null-hypothesis distributions. The underlying
network model is the same as used in multivariate Granger causality, but our test relies on
the autoregressive coefficients instead of error residuals. By means of numerical simulation
over multiple network configurations, we show that this method achieves a good control
of false positives (type 1 error) and detects existing pairwise connections more accurately
than using the standard parametric test for the ratio of error residuals. In practice, our
method aims to detect temporal interactions in real neuronal networks with nodes possibly
exhibiting redundant activity. As a proof of concept, we apply our method to multiunit
activity (MUA) recorded from Utah electrode arrays in a monkey and examine detected
interactions between 25 channels. We show that during stimulus presentation our method
detects a large number of interactions that cannot be solely explained by the increase in the
MUA level.
INTRODUCTION
In recent years, there has been a growing interest in developing multivariate techniques to
infer causal relations among time series. The initial formulation of the problem goes back
to the seminal work by Granger in the 1960s (Granger, 1969) motivated by the analysis of
the pairwise influence between economic time series.
In this work, Granger decomposes
the cross-spectrum of two autoregressive time series into two directional components that
account for the potential causal influences between each other. A general solution of the
problem in multivariate scenarios was developed a decade later by the introduction of mul-
tivariate autoregressive (MVAR) processes, which allow the estimation of causal relationships
between nodes in networks with linear feedback based on their observed activity (Amemiya,
1974; Geweke, 1982, 1984; Lütkepohl, 2005). The MVAR was further combined with spec-
tral analysis to develop the directed transfer entropy function (Kaminski & Blinowska,
1991; Kami ´nski, Ding, Truccolo, & Bressler, 2001), which has been employed to analyze
Citation: Gilson, M., Tauste Campo, A.,
Chen, X., Thiele, A., & Deco, G.
(2017). Nonparametric test for
connectivity detection in multivariate
autoregressive networks and
application to multiunit activity data.
Network Neuroscience, 1(4), 357–380.
https://doi.org/10.1162/netn_a_00019
DOI:
https://doi.org/10.1162/netn_a_00019
Supporting Information:
http://www.scipy.org
http://dally.nimh.nih.gov/index.html
Received: 3 January 2017
Accepted: 1 June 2017
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Authors:
Adria Tauste Campo
adria.tauste@upf.edu
Matthieu Gilson
matthieu.gilson@upf.edu
Handling Editor:
Pedro Valdes-Sosa
Copyright: © 2017
Massachusetts Institute of Technology
Published under a Creative Commons
Attribution 4.0 International
(CC BY 4.0) license
The MIT Press
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
1
4
3
5
7
1
0
9
1
8
7
2
n
e
n
_
a
_
0
0
0
1
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Nonparametric MVAR connectivity detection for MUA data
Noise-diffusion network:
Stochastic model used in statistics to
capture linear interactions among
multiple time series.
Granger causality analysis:
Estimation of directional interactions
between time series from the
residuals of two MVAR linear
regressions.
Error residuals:
Unexplained variability of the node
activity in a multivariate process
(e.g., MVAR) fitted to observed
time series.
Parametric and nonparametric
significance testing:
Comparison of estimated value with
a known (parametric) or surrogate
(nonparametric) reference probability
distribution.
connectivity patterns in neurobiological systems (Babiloni et al., 2005; Wilke, Ding, & He,
2008). Granger causality analysis is nowadays often used to evaluate the influence of a group
of variables onto another, which corresponds to the influence of a subgroup of nodes onto
another one in networks. In particular, it is also applied to detect individual connections be-
tween pairs of nodes (each subgroup being a single node), which sets the context of the present
paper.
In neuroscience, this inference problem has been transposed to analyze interactions be-
tween neuronal populations from spiking activity or neuroimaging measurements such as
fMRI, EEG, and MEG (Lusch, Maia, & Kutz, 2016; Messé, Rudrauf, Benali, & Marrelec, 2014;
Michalareas, Schoffelen, Paterson, & Gross, 2013; Rogers, Katwal, Morgan, Asplund, & Gore,
2010; Seth, Barrett, & Barnett, 2015; Storkey et al., 2007). Two types of estimation proce-
dures may be distinguished: measures relying on an underlying interaction model such as
Granger causality analysis (M. Ding, Chen, & Bressler, 2006) and dynamic causal modeling
(DCM) (Friston, Harrison, & Penny, 2003) on the one hand; and model-free measures such
as transfer entropy (Schreiber, 2000) and directed information (Massey, 1990), which make
minimal model assumptions, on the other hand. Although model-free approaches have
proven useful
(So, Koralek, Ganguly,
Gastpar, & Carmena, 2012; Tauste Campo et al., 2015), certain assumptions are required when
estimating interactions at the neuronal population level, in which broader spatial and tem-
poral scales contribute to shaping the signals. Motivated by data-driven practical problems,
methodological refinements of Granger causality analysis (or MVAR-based methods) have
considered additive noise (Vinck et al., 2015) or measurement noise via state-space models
(Barnett & Seth, 2015; Friston et al., 2014). However, in the majority of cases, the ratio behind
the detection test concerns submodel error residuals, which might become too similar when
connections are placed in a highly redundant network, thus increasing the missed detection
rate (Stramaglia, Cortes, & Marinazzo, 2014).
to describe neural propagation at spike-train level
To detect directed connections in the general context of large networks, we propose to test
the significance of the MVAR coefficients using a nonparametric procedure. As a generative
model, the MVAR process is canonically related to Granger causality analysis: the linear re-
gression in the upper right inset of Fig. 1A provides both coefficients and residuals, the latter
being viewed as the remaining uncertainty in the prediction of the target time series by its
source(s). By comparing the residuals of two linear regressions—one involving a supposed
driver node and one without it—in a log ratio, traditional tests for Granger causality estimate
the effective interaction of one node onto another (Barrett & Barnett, 2013). Since these log
ratios asymptotically converge to known distributions, parametric statistical tests have been
developed to assess the significance of these interactions (Barnett & Seth, 2014). Instead, our
proposed method evaluates the significance of the MVAR coefficients to infer the existence
of network connections. To achieve this, we propose a nonparametric significance test in the
regression coefficients space. Previous literature on nonparametric testing for Granger causal-
ity has resorted to surrogate data generated by trial shuffling (Dhamala, Rangarajan, & Ding,
2008; Nedungadi, Rangarajan, Jain, & Ding, 2009), bootstrap procedures (Diks & DeGoede,
2001), or phase randomization in frequency-domain measures (L. Ding, Worrell, Lagerlund, &
He, 2007; Li et al., 2016). Here we focus on within-trial surrogate tests for time-domain co-
efficients as done in previous studies (Faes, Marinazzo, Montalto, & Nollo, 2014; Schreiber &
Schmitz, 1996; Winkler, Ridgway, Webster, Smith, & Nichols, 2014).
Our approach is motivated by the growing of multichannel recording techniques in neu-
roscience, which require tailored multivariate analysis. In the context of recurrent networks,
Network Neuroscience
358
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
1
4
3
5
7
1
0
9
1
8
7
2
n
e
n
_
a
_
0
0
0
1
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Nonparametric MVAR connectivity detection for MUA data
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
1
4
3
5
7
1
0
9
1
8
7
2
n
e
n
_
a
_
0
0
0
1
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 1. Network model and connectivity estimation. (A) For a given directed connectivity A
and input covariances Σ (left), the network activity (middle) is simulated using Equation 1. From the
observed time series, the existing interactions in the original connectivity can be estimated (right):
Granger causality analysis uses the residuals of linear regressions ((cid:2) in the upper right equation; see
Methods for details about the residuals used in the log ratio), whereas MVAR corresponds to the
and (cid:2)Q1
coefficients. Note that MVAR can be obtained using the empirical covariance matrices (cid:2)Q0
;
see in Equation 7 for τ = 0 and 1. (B) The left panel compares the estimated values with the original
values for all connections in the network. The upper thread displays the distributions of estimated
values for existing and nonexisting connections in the original network. Using a sliding threshold
(vertical dashed gray line) on the estimated values, one can calculate the ROC curve (right). The
lower thread compares the estimated value for a single connection with a null distribution. From
this, the choice of whether the connection exists is made for each individual connection.
False alarm (or false positive):
Wrong rejection of the null
hypothesis in a detection test (here
detecting a connection that is absent
in the original connectivity).
which are ubiquitous in neuroscience, we provide numerical evidence that these tests can
achieve a good control of the false-alarm rate and might improve the miss rate by properly
adapting the null distribution to each connection. The focus of the present analysis is on the
case where we observe more time samples (a few thousands per node) than the network size
(about a hundred nodes), a usual ground for electrophysiological data. Within this regime,
we test the robustness of the detection method for a broad range of network parameters and
various nontrivial topologies inspired by neuronal networks.
Network Neuroscience
359
Nonparametric MVAR connectivity detection for MUA data
Multivariate autoregressive (MVAR)
process:
Dynamic network model where
nodes linearly interact between each
other while receiving input noise; in
discrete time, this definition is
mathematically equivalent to the
MVAR process.
Ordinary least square (OLS) linear
regression:
Optimization technique for the
MVAR process to estimate the linear
interaction strengths.
METHODS: MULTIVARIATE AUTOREGRESSIVE MODEL AND
CONNECTIVITY ESTIMATION
The activity in the MVAR process—also known as noise-diffusion discrete-time network—is
described by the following equation:
xt = Axt−1 + ζt ,
(1)
where the connection matrix A describes the interactions between coordinates of the vector
xt = (xt
i ) with time t being an integer and node index 1 ≤ i ≤ N. Here we constrain our study
to the case where ζt
is Gaussian (possibly cross-correlated noise), whose realizations are time
independent for successive time steps. Without loss of generality, we assume that all variables
ζt
are zero mean. We only consider MVAR processes of order 1 in a first place, but will extend
the work to the case of order 2 in a later section.
Granger Causality Analysis
Granger causality analysis is usually presented using time series, and the estimation of nonzero
coefficients in A from observed activity over a period 1 ≤ t ≤ T relies on the linear regression
of the activity xt
i of a given node i at time t by the past activity of a subset S of network nodes:
i = ∑
xt
j∈S
aijxt−1
j + (cid:2)t
(2)
for 2 ≤ t ≤ T. When T is large and S contains all nodes, the estimated coefficients aij converge
toward Aij. With abuse of notation, we define the residual (cid:2) as the standard deviation of the
(cid:2)t
for the ordinary least square (OLS) regression in Equation 2, standard deviation of error
residuals
(cid:3)
(cid:2)
x2≤t≤T
i
|x1≤t≤T−1
i
(cid:5)
(cid:4)
=
((cid:2)t)2 ,
∑
t
(3)
with a notation similar to conditional probabilities; the superscript t indicates the considered
time range and the subscripts indicate the nodes involved. To detect the existence of connec-
tion j → i in a network, two types of Granger causality analysis exist: “unconditional” and
“conditional” (Geweke, 1982, 1984); they consider the comparisons of the following residuals:
GRu(xj → xi) = ln
GRc(xj → xi) = ln
(cid:3)
(cid:3)
(cid:3)
(cid:2)
(cid:2)
(cid:2)
⎡
⎣
⎡
⎣
x2≤t≤T
i
x2≤t≤T
i
x2≤t≤T
i
(cid:3)
(cid:2)
|x1≤t≤T−1
1,··· ,j−1,j+1,···,N
|x1≤t≤T−1
1,··· ,N
(cid:4)
x2≤t≤T
i
⎤
(cid:4)
⎦ .
⎤
⎦ ;
(cid:4)
(cid:4)
(4)
i
|x1≤t≤T−1
|x1≤t≤T−1
i,j
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
1
4
3
5
7
1
0
9
1
8
7
2
n
e
n
_
a
_
0
0
0
1
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
For both GRu and GRc, which have a univariate target node xi, the usual parametric test
for significance relies on the F statistic, which performs better for a small number of samples
(Barnett & Seth, 2014). The null hypothesis of no interaction for GRu(xj → xi) corresponds
to m = T, p = 1, nx = 1, and ny = 2 using the notation in Barnett and Seth (2014):
(cid:3)
(cid:2)
x2≤t≤T
i
(cid:4)
|x1≤t≤T−1
(cid:3)
i
(cid:2)
x2≤t≤T
i
(cid:3)
x2≤t≤T
− (cid:2)
i
(cid:4)
|x1≤t≤T−1
i,j
(cid:4)
|x1≤t≤T−1
i,j
= [exp(GRuij) − 1] >
φ(α, 1, T − 3)
T − 3
(5)
Network Neuroscience
360
Nonparametric MVAR connectivity detection for MUA data
with α the desired sensitivity and φ the inverse survival function of the F distribution
(Scientific Python Library, n.d.). The equivalent for GRc corresponds to ny = N, yielding
(cid:3)
(cid:2)
x2≤t≤T
i
(cid:4)
|x1≤t≤T−1
1,··· ,N
(cid:3)
(cid:2)
x2≤t≤T
i
(cid:3)
x2≤t≤T
− (cid:2)
i
|x1≤t≤T−1
1,··· ,j−1,j+1,··· ,N
(cid:4)
|x1≤t≤T−1
(cid:4)
1,··· ,j−1,j+1,··· ,N
>
φ(α, 1, T − N − 1)
T − N − 1
.
(6)
Circular shift of time series:
The new start is set to a given
position and remaining early entries
are sequentially moved after the end
of the original series.
We also use nonparametric tests for GRc by performing a circular shift (see details in the
section below, Generation of Surrogate Time Series) either on the target node for each connec-
tion (Faes et al., 2014) or independently on the time series of all nodes (in order to save time
in estimating the full network’s connectivity by shuffling somehow all targets simultaneously).
Both cases provide a null distribution for the log ratio, with which the actual estimated log
ratio can be compared.
Multivariate Autoregressive Estimation
To detect the existence of connections Aij > 0, another possibility is to estimate the coefficients
themselves, which can be done using the covariances of the observed activity variables xt
(Lütkepohl, 2005):
1
(cid:2)Qτ
ij =
T − τmax − 1 ∑
1≤t≤T−τmax
(xt+τ
i − ¯xi)(xt
j − ¯xj) ,
(7)
where T denotes the number of successive samples indexed by t, τ ∈ {0, 1} is the time shift
(here τmax = 1), and the observed mean activity for each node is ¯xi = 1
i . The Yule-
Walker equation gives a consistency equation for the theoretical covariance matrices (Qτ
) in
terms of the connectivity A in the dynamics described by Equation 1:
T ∑t xt
Q1 = A Q0 .
(8)
The estimation of network connections relies on evaluating A from Equation 8 for the
empirical covariance matrices defined as Equation 7 and calculated for a given time series:
a = (cid:2)Q1( (cid:2)Q0)−1 .
(9)
1,··· ,N |x1≤t≤T−1
x2≤t≤T
Note that this OLS estimate corresponds to the linear regression related to (cid:2)
and also to the linear model with maximum likelihood under the assumption that the observed
process is Gaussian.
1,··· ,N
(cid:3)
(cid:4)
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
1
4
3
5
7
1
0
9
1
8
7
2
n
e
n
_
a
_
0
0
0
1
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
MVAR of Order 2
Equation 1 can be extended to the case where the activity vector xt
previous time steps:
is determined by the two
xt = A1xt−1 + A2xt−2 + ζt .
(10)
For the second order, we use τmax = 2 in Equation 7 and the estimation of A1
Yule-Walker equation is given by (Lütkepohl, 2005, p. 86)
and A2
via the
˜a = ˜Q1 ( ˜Q0)−1,
(11)
361
Network Neuroscience
Nonparametric MVAR connectivity detection for MUA data
with the block matrices
˜a =
˜Q0 =
˜Q1 =
(cid:10)
(cid:12)
(cid:10)
(cid:11)
a1
a2
(cid:2)Q0
( (cid:2)Q1)†
(cid:2)Q1
(cid:2)Q2
,
(cid:2)Q1
(cid:2)Q0
(cid:11)
.
(cid:13)
,
(12)
Random permutation of time series:
Shuffling of the time labels in time
series to destroy their temporal
arrangement when generating
surrogates for nonparametric test.
The coefficients of A1
inversion involving the covariances, as with the first-order case in Equation 9.
can thus be estimated using a matrix multiplication and an
and A2
Generation of Surrogate Time Series
In this paper, we consider circular shifts (CS), random permutations (RP), and phase random-
ization (PR) to shuffle the time points of the observed time series. From the original xt
i with
1 ≤ t ≤ T,
CS draws a random integer t0 ∈ {1, · · · , T} and returns (xt0
RP draws a random permutation σ of {1, · · · , T} such that each integer appears once
(and only once) and returns xσ(t)
PR calculates the discrete Fourier transform F (xt
of the T coefficients of F (xt
performs the inverse transform.
i , then multiplies each
randomly chosen in [0, 2π], and
i ) by exp(2πıφt) with φt
i ) of the original xt
i , · · · , xt0−1
i , · · · , xT
i , x1
);
;
i
i
Importantly, these operations are applied to each time series independently of the others.
In addition, we consider the replacement of all time series in the network by T zero-mean
and normally distributed variables with a standard deviation equal to the average standard
deviation of xt
i along the time axis, over all nodes. We refer to these surrogates as STD.
Experimental Setup and Processing of Electrode Measurements to Extract MUAe Activity
All procedures were carried out in accordance with the European Communities Council
Directive RL 2010/63/EC, the U.S. National Institutes of Health Guidelines for the Care and
Use of Animals for Experimental Procedures, and the U.K. Animals Scientific Procedures Act.
Two male macaque monkeys (5–14 years of age) were used in the experiment; only the data
for the first one are used here. A surgical operation was performed under sterile conditions,
in which a custom-made head post (Peek, Tecapeek) was embedded into a dental acrylic
head stage. Details of surgical procedures and postoperative care have been published pre-
viously (Thiele, Delicato, Roberts, & Gieselmann, 2006). During the surgery microelectrode
chronic Utah arrays (5 × 5 grids), attached to a CerePort base (Blackrock Microsystems) were
implanted into V1. Electrodes were 1 mm in length in line with procedures described in
Supèr and Roelfsema (2005). Stimulus presentation was controlled using Cortex software (Lab-
oratory of Neuropsychology, NIMH, http://dally.nimh.nih.gov/index.html) on a computer with
an Intel Core i3-540 processor. Stimuli were displayed at a viewing distance of 0.54 m, on a
25
Sony Trinitron CRT monitor with a resolution of 1280 by 1024 pixels, yielding a resolution
of 31.5 pixels / degree of visual angle (dva). The monitor refresh rate was 85 Hz for monkey
1, and 75 Hz for monkey 2. A gamma correction was used to linearize the monitor output,
and the gratings had 50% contrast. Monkeys performed a passive viewing task where they
fixated centrally while stationary sinusoidal grating of either horizontal or vertical orientation
and 2 cycle per degree spatial frequency were presented in a location that covered all receptive
(cid:5)(cid:5)
Network Neuroscience
362
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
1
4
3
5
7
1
0
9
1
8
7
2
n
e
n
_
a
_
0
0
0
1
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Nonparametric MVAR connectivity detection for MUA data
fields recorded from the 25 electrode tips. Stimuli were presented 500 ms after fixation onset
for 150 ms. Raw data were acquired at a sampling frequency of 32556 Hz using a 64-channel
Digital Lynx 16SX Data Acquisition System (Neuralynx, Inc.). Following each recording ses-
sion, the raw data were processed offline using commercial software (Neuralynx, Inc.). Signals
were extracted using Cheetah 5 Data Acquisition Software, with bandpass filtering set to allow
for spike extraction (600-9000 Hz) and saved at 16-bit resolution.
In the present study, we focus on the period starting 200 ms before and finishing 200 ms after
the stimulus onset, for 4 conditions (vertical gradings with pre/post cue in the receptor/opposite
field) that will not be compared in details. The electrode recordings is firstly down-sampled
from 32,556 Hz to 1,000 Hz. A high-pass filter above 400 Hz is then applied—third-order
Butterworth filter at 0.8 of the Nyquist frequency (Scientific Python Library, n.d.)—followed
by a smoothing of 4 ms to extract the envelope of the resulting signal, thus retaining the 250
time points of 1,000-ms period surrounding the stimulus onset.
BENCHMARK OF DETECTION PERFORMANCE FOR SYNTHETIC DATA
The workflow of the benchmark for the estimation procedure is schematically represented in
Figure 1A. We first consider an MVAR process defined by Equation 1 with given connectiv-
ity matrix A and input covariances (obtained by mixing independent Gaussian processes) to
generate the activity of the network. From the observed activity over a period of duration T,
we estimate the coefficients matrix A using the covariances as described in Equation 9. We
also perform the linear regressions of each node activity over the past activity of given subsets
of nodes corresponding to the unconditional (GRu) and conditional (GRc) Granger causality
analysis, from which we calculate the ratios of residuals in Equation 4. Actually, these esti-
mates correspond to the same OLS regression (top right in Figure 1A) and the difference resides
in the spaces where they lie: coefficients versus residuals.
For each method, the prediction power can be measured by the relationship between the
estimated values and the original connectivity values, as illustrated in Figure 1B (left). To dis-
criminate between existing and absent connections, one can apply a common threshold for
all connections (top thread); by sliding this threshold, we obtain the ROC (receiver-operating
characteristic) curve with the rates of false alarms on the x-axis and of true positives on the
y-axis. The area under the curve indicates in a single value how well the ranking of estimates
performs for the detection of connections with respect to the original connectivity. Alterna-
tively, an individual test can be made for each connection with respect to the network, for
example by comparing the estimated value to a null distribution (bottom thread). We then
obtain the false-positive and true-positive rates by pooling the results over all connections.
Coefficients from Linear Regression Potentially Predict Better Existing Connections Than Residuals
We start with the comparison between the predictability of coefficients and residuals for MV,
GRu, and GRc for all connections in a given network. To do so, we simulate 500 randomly
connected networks, which are simulated with different sizes (N = 50 to 150 nodes), density,
and connectivity weights (uniformly drawn in a randomly chosen range [wmin, wmax]). Here
inputs are not correlated; the ζi are independent across node indices in Equation 1. For each
network configuration, we evaluate the accuracy for connection detection via the area under
the ROC curve (see the upper thread in Figure 1B). Figure 2A displays this ROC-based accuracy
as a function of the number of observed time samples (x-axis) represented by violin plots for
ROC curve:
Curve describing the performance of
a binary detector when moving its
discrimination threshold.
Network Neuroscience
363
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
1
4
3
5
7
1
0
9
1
8
7
2
n
e
n
_
a
_
0
0
0
1
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Nonparametric MVAR connectivity detection for MUA data
A
C
residuals GRu
residuals GRc
coefficients MV
Effect of number of samples
B
Match estimate – original A
Effect of network parameters
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
1
4
3
5
7
1
0
9
1
8
7
2
n
e
n
_
a
_
0
0
0
1
9
p
d
.
t
Figure 2. ROC-based prediction power. (A) Area under ROC for estimated A obtained from log
ratios of residuals obtained from Granger causality analysis (unconditional for GRu and conditional
for GRc) and MVAR. The x-axis indicates three sample size T for the observed network activity.
The violin plots correspond to 500 simulated networks of various sizes and connectivity strengths
(the horizontal black bar indicates the median). (B) Match of the ranking between GRu, GRc, and
MVAR estimates and the original connectivity weights A, as measured by the Spearman correlation
coefficient. The plotted values correspond to the 500 networks in A and the x-axis indicates the
sample size T. (C) Effect of network parameters on ROC-based performance. Influence of network
size N, connectivity density, sum of recurrent connectivity strengths, minimum weight wmin in A,
mean noise on the diagonal of Σ, and mean off-diagonal noise in Σ on the ROC-based accuracy
in Figure 2A. In each plot, the network configurations have been grouped in quartiles according to
the parameter plotted on the x-axis, and the corresponding group mean and standard deviations are
indicated; the curves are displaced horizontally to improve legibility.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
500 randomly connected networks. When considering many samples (104
), all methods per-
form well. However, for smaller sample sets, the MVAR method exhibits superior performance
than GRu: as measured by the Mann-Whittney test, p < 10−45
for
the three values of observed samples, respectively.
, and p < 10−5
, p < 10−19
Network Neuroscience
364
Nonparametric MVAR connectivity detection for MUA data
Although error residual ratios are in a different space from the true weights in A, one expects
some degree of correlation between them, such that Granger causality analysis effectively de-
tects connections. In Figure 2B, both GRu and GRc estimates have a ranking similar to the
original A weights (as measured by the Spearman correlation) for T = 10, 000 observed time
samples, but this weakens dramatically for T ≤ 3, 000. In contrast, the ranking for estimated
MVAR coefficients reflects much better the original A for T ≤ 3, 000. In the studied networks,
GRu performs slightly better than GRc. As analyzed in previous studies, this can be a conse-
quence of the balance between redundant and synergistic activity exhibited by the simulated
network nodes (Stramaglia et al., 2014). To shed light into the effect of the network structure,
we next examine how the ROC-based performance in Figure 2A depends on the controlled
network parameters. The four panels in Figure 2C display the trends of the values for the
500 networks as a function of the network size N, the network density, the minimum weight
in the original network (wmin mentioned above), and the mean sum of incoming weights per
node. For illustration purpose, the 500 networks are grouped in quartiles for each parameter.
Not surprisingly, the estimation accuracy of all methods decreases as a function of the network
size N and density, and increases as a function of the minimum connectivity weight and the
mean incoming weight per node. More interestingly, in challenging configurations with small
weights, MVAR consistently shows a superior performance by a larger gap compared with
Granger causality analyses. These findings support the use of coefficients to robustly detect
connections in recurrently connected networks. Note that GRu performs on average slightly
better than GRc here: The discrepancy decreases as a function of the network density, which
may follow from lower redundancy in the recurrent network (Stramaglia et al., 2014).
A Robust Nonparametric Significance Test for MVAR
We have so far examined the performance of different estimation methods based on the area
under the ROC curve, which corresponds to a single threshold for all connections in a net-
work and combines the information about false alarms and true detection over the whole range
of estimated values. However, in the context of real data, the decision for the existence of a
connection typically relies on comparing the value of the connection estimate with a given sta-
tistical threshold. For GRu and GRc, such parametric tests have been developed, for example,
based on the F statistics (Barnett & Seth, 2014). Equivalently, it is sufficient to know how the
values of the estimates for absent connections are distributed, in order to select a desired rate
of false alarms (type 1 error). In this section, we develop a significance test for the estimated
MVAR coefficients by providing a null-hypothesis distribution for absent connections.
Our approach relies on the fact that covariances reflect the underlying connectivity. We thus
construct the null distribution for estimates by performing a random permutation for each of
the observed time samples, which “destroys” the covariance structure apart from the variances
on the diagonal of (cid:2)Q0
as illustrated in Figure 3A; other methods will be tested in a later section.
From the resulting covariance matrices, we evaluate a surrogate connectivity matrix. The core
result underlying our surrogate approach is shown in Figure 3B. The distribution of surrogate
estimates (thick black line) is compared against the distribution of existing (red) and absent
(blue) connections in a simulated random network model. The surrogate distribution in black
provides a good approximation for the distribution of estimates for nonexisting connections in
blue.
We consider two options—corresponding to the two threads in Figure 1B—to test the exis-
tence of a connection from an MVAR estimate while keeping the false-alarm rate under control.
Network Neuroscience
365
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
1
4
3
5
7
1
0
9
1
8
7
2
n
e
n
_
a
_
0
0
0
1
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Nonparametric MVAR connectivity detection for MUA data
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
1
4
3
5
7
1
0
9
1
8
7
2
n
e
n
_
a
_
0
0
0
1
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. Nonparametric test to assess significance for MVAR coefficients based on random per-
mutations. (A) Schematic illustration of random permutation (numbers indicate time) applied in-
dependently to all observed time series (green curves) to generate the surrogate covariances (right
panels).
(B) Pooled distributions of estimated weights for the existing (in red) and absent (blue)
connections. The thick black curve indicates the distribution of connections over 100 surrogates,
which closely matches the blue distribution. (C) For a given connection, we compare two methods:
for “local” in red, the null distribution corresponds to the matrix elements for the same connection
in 200 surrogates; for “global” in black, the null distribution is the pooled distribution for all N2
elements of the 200 surrogate matrices (same as in B). The dark red and gray dashed lines indicate
the detection thresholds corresponding to the 4% tail for those two options. (D) The performance
of the two nonparametric methods for the thresholds described in B and C is displayed on the ROC
curve for a desired false-alarm rate ranging from 1% to 5%. Triangles indicate the global test and
circles the local test. (E) Comparison of the desired (% of the tail of null distribution) and actual rate
of false alarms for the local and global tests when varying the the number S of surrogates (see figure
in legend). Error bars indicate one standard deviation for 500 random networks; importantly, inputs
for these networks have cross-correlation, unlike Figure 2. (F) Influence of the strength of original
weight on the detection performance for the 500 random networks and a desired false-alarm rate
set to 2% in E. In both panels, lighter colors indicate smaller numbers of surrogates S, in red for the
local test and gray for the global test (see legends).
Network Neuroscience
366
Nonparametric MVAR connectivity detection for MUA data
The global test relies on the null distribution corresponding to the black histogram in
Figure 3C, which is obtained by grouping together all SN2
matrix elements of all matrices
for S = 200 surrogates. From that surrogate distribution, we perform a detection test by
setting a threshold corresponding to a percentage of the right tail equal to the desired
false-alarm rate (here 2%), as illustrated by the vertical gray dashed line.
Instead, the local test uses for each connection the surrogate distribution of S values,
corresponding to the same matrix element in each of the S surrogates. From that distri-
bution in red in Figure 3C, the detection threshold is defined similarly (vertical dark red
dashed line).
The rationale behind these two choices lies in the trade-off between taking into account
spatial heterogeneity in the network and gaining larger sample size, as illustrated by the dis-
tinct thresholds in Figure 3C. Note also that the F statistical test for Granger causality analysis
corresponds to a global threshold on the log ratio values. When varying the desired false-alarm
rate, the two tests perform well, as illustrated in Figure 3D by their location close to the ROC
curve (circles and triangles for local and global, respectively).
To quantify the small variability observed in Figure 3D over the randomness of network
configurations, we simulate 500 randomly connected networks with the same parameters as
in Figure 2, except for the size 50 ≤ N ≤ 90 and the presence of input cross-correlations.
Note that, from Figure 2, the chosen size N corresponds to a situation where Granger causality
analysis performs relatively well as compared with MVAR. The control of the false-alarm rate
is displayed in Figure 3E for both local and global tests with various numbers S of surrogates.
The control of false-alarm rates is close to perfect across various values for all S and both
tests (local and global), demonstrating the robustness of the proposed method for randomly
connected networks. Next, we fix the desired false-alarm rate to 2% and evaluate the miss
rate (true negatives) of both methods depending on the actual weight strength: In Figure 3F,
connections are grouped in terciles for each network configuration.
Interestingly, the local
test improves with the number S of surrogates (right panel), whereas the global test exhibits a
constant performance for all S (left panel). Note that the advantage of the local test over the
global test particularly concerns connections with small weights, which are difficult to detect,
in line with Figure 2C (see influence of the minimum original weight).
Because GRu does not take all network nodes into account, the presence of spatially corre-
lated noise (indicated by the purple dashed arrows in Figure 1A) dramatically affects the false-
alarm rate when using the parametric significance F test (Barnett & Seth, 2014), as shown in
Figure 4A by the dark blue dashed curve. This is solved by the “complete” linear regression in
GRc, achieving a quasi perfect control irrespective of the input correlation level for both the
parametric and two nonparametric tests (cyan, green, and blue-green dashed curves, respec-
tively), as our nonparametric tests do (red and gray; recall also Figure 3E). We consider two
nonparametric tests for GRc: “T” stands for target, where the null distribution of a connec-
tion is obtained by shuffling only the target, and “F” stands for full, where we shuffle all time
series simultaneously as in our coefficient-based tests. Both perform equally in terms of false
alarms.
The main result of the paper is described in Figure 4B, where the dashed line corresponds to
the miss rate for parametric GRc: Our nonparametric method exhibits a better than miss rate—
that is, decrease—for both local and global tests (in red and gray, respectively) on average over
the same 500 random networks as in Figure 4A. For S ≥ 200, the local test even becomes bet-
ter in all cases. Note that the small miss-rate improvements of about 7% actually correspond
Network Neuroscience
367
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
1
4
3
5
7
1
0
9
1
8
7
2
n
e
n
_
a
_
0
0
0
1
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Nonparametric MVAR connectivity detection for MUA data
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
1
4
3
5
7
1
0
9
1
8
7
2
n
e
n
_
a
_
0
0
0
1
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. Comparison of our coefficient-based method with Granger causality analysis. (A) Com-
parison of the parametric tests for GRu (blue curve) and GRc (cyan) with the nonparametric methods
for GRc (green for GRcF and blue-green for GRcT; see the text for details) and MVAR (red for local
test and gray for global). The x-axis indicates the strength of input correlations (i.e., pink noise) in
the simulated network. The desired false-alarm rate is set to 2% as in Figure 3F and the number of
observed time samples is T = 3, 000. Error bars indicate one standard deviation over the 500 ran-
dom networks as in Figure 3E. (B) Comparison of the miss rate improvement (decrease) with respect
to parametric GRc for the 500 networks in A as a function of the number S of surrogates (x-axis).
Red indicates the local test, gray the global test, green the full-network nonparametric GRc, and
blue-green the target-only nonparametric GRc. (C) Details of the performance of the five methods
in B as a function of the mean incoming weight per node (left) and the network density (right). The
plots for the miss rate are similar to those for the ROC-based prediction power in Figure 2C. (D)
Comparison of the computational cost for the surrogate-based method and parametric tests as a
function of the number T of observed samples (left) and network size (right). Only GRcF is shown,
as GRcT takes a much longer time in the unoptimized version that we use.
to more than 50 existing connections per network here. In contrast, both nonparametric tests
for GRc perform worse than the parametric test here, with the target-shuffling surrogate con-
verging faster to the nonparametric GRc. Figure 4C displays the trends of the performance of
all five tests in Figure 4B as a function of two network properties: the mean incoming weight
Network Neuroscience
368
Nonparametric MVAR connectivity detection for MUA data
per node (left) and the density (right). The main result here is that the local test performs better
especially in difficult configurations with small weights and dense connectivity.
From Figures 3F and 4B–C, we conclude that the local test is preferable to the global test
provided S ≥ 200 surrogates are generated. However, the computational cost increases lin-
early with S, as illustrated in Figure 4D by the red curves. Note that the parametric GRc (in
cyan) takes the same time to calculate as S = 50 surrogates. However, our nonparametric
method scales better than parametric GRc when the network size increases. As a compari-
son, the full-network nonparametric test for GRc takes a longer time to compute, but further
optimization of the calculations could be made that were not incorporated here.
Comparison of Generation Methods for Surrogates for Nonparametric MVAR
The fact that the OLS MVAR estimates can be obtained via the two covariance matrices (with
and without time shift; see Figure 1A) hints at possible methods to generate surrogate by de-
stroying the structure in these covariances. Methods to generate surrogate time series have
been widely used in the past: circular shifts of the time series (Faes et al., 2014), random
permutation (Winkler et al., 2014), and phase randomization (Schreiber & Schmitz, 1996) to
generate a null distribution for the ratios in Equation 4; they are referred to here as CS, RP, and
PR, respectively. We thus consider these three methods (cf. box in Figure 5), as well as sur-
rogate time series that only preserve the mean standard deviation averaged over the network
(STD), so as to test to what extent it is important to preserve the spatial heterogeneity of the
nodes’ activity. See Methods for details about the calculations.
The control of false alarms for local tests in Figure 5A and B is better for CS and RP, whereas
the detection of true connections is similar for the four methods over 500 random networks
of size N = 70. However, CS fails to detect self-connections (Figure 5C). The reason is that,
because CS surrogates preserve the autocovariances in the time-shifted covariance, they fail to
build a proper null distribution for self-connections. The influence of the number of samples
used in the estimation is similar for all methods, as illustrated in Figure 5D. The comparison
with STD (purple), which averages the covariance statistics over the whole network, suggests
that the local test makes a good use of the heterogeneous information across nodes. As a
conclusion, we retain RP as the best option.
Influence of Network Topology
In this part, we test and compare the robustness of global and local surrogate-based detec-
tion tests to specific connections and topological configurations. Here, T = 3, 000 observed
samples and we compare the local and global tests with S = 400 surrogates for 500 networks
of each type. In all cases, the simulated networks have the same size N = 70, but they vary
in connectivity density, distribution of recurrent weights and level of input cross-correlation.
We compare the miss rate for unidirectional, reciprocal, and self-connections in the random
networks examined until now (and a desired 2% of false alarms). Figure 6A shows that the miss
rate is similar in unidirectional and reciprocal connections with the local test, which performs
slightly better than the global test (as in Figure 3F).
Modular and hierarchical network
topology:
Nonrandom connectivity giving
heterogeneous statistics (e.g., number
of connections) across nodes.
Now we consider more elaborate network topologies than the random connectivity (Erdös-
Rényi) considered so far, namely modular and hierarchical networks. In Figure 6B, we simulate
500 modular networks with two groups (green and blue) linked by hubs (red, about 5 to 15% of
the nodes). Interestingly, intragroup and hub-group connections have a similar miss rate with
Network Neuroscience
369
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
1
4
3
5
7
1
0
9
1
8
7
2
n
e
n
_
a
_
0
0
0
1
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Nonparametric MVAR connectivity detection for MUA data
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
1
4
3
5
7
1
0
9
1
8
7
2
n
e
n
_
a
_
0
0
0
1
9
p
d
.
t
Figure 5. Comparison of the local test for four surrogate generation methods. Surrogates are
generated by independently performing for each original time series (1) random permutations, (2)
circular shifts, (3) phase randomization, and (4) replacing the original by a new random time se-
ries with the mean standard deviation averaged over all of the original time series in the network.
(A) Comparison of the control of the false-alarm rate for various thresholds on the tail of the distri-
butions of 400 surrogates (% indicated on the x-axis). The error bars correspond to the variability
over 500 random networks similar to Figure 3 with T = 3, 000 observed time samples and the local
test. (B) Influence of the number S of surrogates on the detection performance for the four surrogate
methods (x-axis). The violin plots indicate the distribution of false-alarm rates (left panel) and miss
rates (right) for the 500 networks in A with a desired false-alarm rate set to 2% (dashed line in the
left panel). Lighter to darker colors correspond to 50, 100, 200, and 400 surrogates, respectively.
(D) Influence of the number T of
(C) Same as the miss rate in B, but only for self-connections.
observed time samples on the miss rate for S = 400 surrogates.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
regard to using local and global surrogates. In Figure 6C, we simulate hierarchical networks
of three layers, for which connections either link the center and an intermediate node, or link
an intermediate node and a leaf, or are self-connections. This network type is much sparser
than the two types in A and B, yielding a quasi perfect detection performance for all types of
connections (miss rate < 0.1 in Figure 6C). In all cases, the local test performs better than the
global test. However, the control of false-alarm rate is similar for both tests with all topologies,
as can be seen in Figure 6D.
Network Neuroscience
370
Nonparametric MVAR connectivity detection for MUA data
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
1
4
3
5
7
1
0
9
1
8
7
2
n
e
n
_
a
_
0
0
0
1
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 6. Robustness to nontrivial network topology.
(A) Detection performance for unidirec-
tional, reciprocal, and self-connections in 500 of the randomly connected networks used so far in
(B) Detection performance for modular topology schematically represented in the left:
Figure 3.
each of the 500 networks composed of two groups connected by hubs. The connections are sepa-
rated depending on whether the connect group nodes or hubs, as indicated by the diagram on the
left. (C) Similar to B with a hierarchical topology, where connections are grouped in three subsets:
from center to intermediate nodes; from intermediate nodes to leaves; self-connections. Note that
the density for all 500 networks is very low. (D) Control of false-alarm rate for the local (left) and
global (right) significance tests and the three network topologies; the plot is similar to Figure 3E.
(E) Control of false-alarm rate and miss rate for networks with both excitatory and inhibitory
connections.
Network Neuroscience
371
Nonparametric MVAR connectivity detection for MUA data
Finally, we consider a network with both excitatory and inhibitory connections (with an
inhibitory ratio equal to 5 to 50% of all) and perform the test by defining a threshold on both
tails of the null distributions. As can be seen in Figure 6E, the positive/negative nature of the
connection weights affects neither the false-alarm nor the miss rate. However, the performance
is poorer than with excitatory connections only.
We conclude that, in those networks with spatial heterogeneity as with randomly connected
networks, the local test with an individual null distribution per connection performs better than
the global test. Recall that an improvement of the miss rate by 1% in a network with a density
of 20% actually corresponds to N20.2/100 (cid:7) 10 existing connections here, so the plotted
improvements concern about 50 connections.
Applicability to Second-Order MVAR Process
As explained in Methods, an MVAR process whose state depends on the two previous time
steps can be estimated with the covariances with time shifts τ = 0, 1, and 2; see Equations 11
and 12 for details. Here we simply focus on random connectivity for the two corresponding
matrices A1
and A2
, with size N that is randomly drawn between 30 and 80; we construct
and A2
A1
such that a connection j → i cannot be in both matrices, but at most in one. The
existing connections are detected with the nonparametric local test relying on RP surrogates for
each matrix separately, as a proof of concept. The control of false alarms in Figure 7A and the
overall detection performance in Figure 7B suggest that our surrogate method can be extended
satisfactorily to higher-order MVAR processes. Note that the improvement by generating more
A
False-alarm control
B
Miss rate
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
1
4
3
5
7
1
0
9
1
8
7
2
n
e
n
_
a
_
0
0
0
1
9
p
d
.
t
C
Detection of A1 and A2
D
Influence of network size
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 7. Connectivity detection for second-order MVAR process. (A) Control of false-alarm rate
for the local test with S = 100, 200, and 400 in both connectivity matrices A1
, corresponding
to each time step. Error bars correspond to the variability over 500 network configurations with
random connectivity and T = 3, 000 observed samples. (B) Influence of the number T of observed
samples (x-axis) on the miss rate for the 500 networks in A. Lighter to darker red indicates the
number of surrogates S. (C) Details of the detection performance for A1
separately, as well
as connections in either A1
. The number of observed samples is indicated on the x-axis as in
B. (D) Influence of the network size N on the detection performance in C for the local and global
tests with S = 400 surrogates.
and A2
and A2
or A2
Network Neuroscience
372
Nonparametric MVAR connectivity detection for MUA data
surrogates is rather weak here.
A1
Importantly, there is no difference between the detection in
, as demonstrated in Figure 7C. Last, the network size worsens the miss rate in
and A2
Multiunit activity:
High-frequency signal from
simultaneous extracellular recordings
(e.g., electrode array), presumably
related to the spiking neuronal
activity.
Figure 7D, which affects more dramatically the global test as compared with the local test.
APPLICATION TO EXPERIMENTAL DATA
Multiunit Activity Data Obtained from Utah Electrode Array in Monkey
Now we consider data recorded from a monkey performing a visual task, where the stimulus
corresponds to vertical gratings covering all recorded V1 receptive fields from the Utah arrays
(see Methods for details). We aim to provide a proof of concept for the connectivity analysis
for this type of data, so as to complement the more classical analysis based on the activity
of individual channels; therefore, we do not focus on comparing the four stimulus conditions
with each other.
The multiunit activity envelope (MUAe) is obtained as described in Methods. In Figure 8A,
the resulting MUAe is represented for two out of the 26 channels (red and purple) for two trials
in the top and middle panels, 400 ms before and 600 ms after the stimulus onset. The typical
analysis of MUAe activity consists of averaging over 200 trials, which exhibits a peak imme-
diately after the stimulus for the two channels in the bottom panel. Among the 26 channels,
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
1
4
3
5
7
1
0
9
1
8
7
2
n
e
n
_
a
_
0
0
0
1
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 8. Application to multiunit activity (MUAe) data. (A) Example of two trials (top and middle
panels) of multiunit activity envelope (MUAe) for two channels of recordings using Utah electrode
array in the primary visual cortex of a monkey (in arbitrary units; see text for further details). The
bottom panel represents the average over 200 trials (with standard-error mean for the thickness
of the curve). The stimulus is presented at time point 100 (actually 400 ms, since the smoothing
corresponds to a smoothing window of 4 ms). (B) Example of cross-covariance between the two
channels in A averaged over 1, 20, and 200 trials.
(C) Autocovariance profiles of MUAe signals
for all 25 channels and time shifts up to 12 ms averaged over 200 trials plotted with a log y-axis:
comparison of signals before (gray) and after (red) the stimulus presentation. (D) MVAR estimates of
the connectivity between the 25 channels for the MUAe activity 200 ms before and after stimulus
presentation (i.e., 50 time points each), averaged over 200 trials. The scaling has been optimized
to enhance the legibility of off-diagonal elements. (E) Comparison of the cumulative distribution
of connectivity weights (off-diagonal elements in D) for the four conditions. Gray and red indicate
before and after the stimulus, respectively.
Network Neuroscience
373
Nonparametric MVAR connectivity detection for MUA data
about a third show a large increase in activity after the stimulus onset as compared with before
(namely, a poststimulus mean activity larger by more than three standard deviations compared
with the prestimulus activity); almost all channels show a moderate increase of one standard
deviation. One channel is discarded for a much larger activity (by 5 times) than all others.
To further investigate the temporal information conveyed by MUAe jointly for pairs of chan-
nels, we calculate the pairwise covariances between them, after centering the MUAe activity
individually for each trial. Figure 8B shows the stabilization of the cross-covariance between
the two channels in Figure 8A from a single trial to averages over 20 and 200 trials. Note
the asymmetry with respect to time difference: This information is extracted by the network
model to estimate the interactions between the neuronal populations recorded by the chan-
nels. Then we verify that the model can be applied to these data, by examining the MUAe
autocovariances in Figure 8C, which exhibit a profile corresponding to an exponential decay
up to two time shifts (i.e., 8 ms for the downsampling every 4 ms), that is, a straight line in the
log plot. This suits an autoregressive model with large positive values on the diagonal of the
connectivity matrix A.
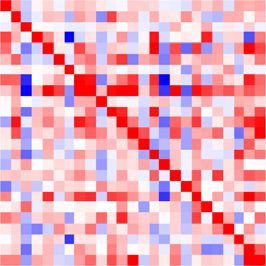
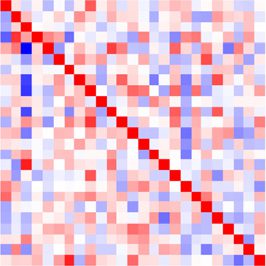
Both connectivity matrices for the 25 channels estimated using the MVAR method before
and after the stimulus are illustrated in Figure 8D for condition 1: we find larger off-diagonal
values for the period after the stimulus than before. This is actually true for all conditions,
as indicated by the more spread distributions in red as compared with gray in Figure 8E. The
channels appear to be coordinated at the considered time scale of 4 ms, and their collective
interaction scheme is affected by the stimulus presentation.
Significance Test for Real Data: Interactions Related to Stimulus Presentation
We then use the local and global tests based on 1,000 surrogates (with random permutation)
to retain only significant interactions from the estimates in Figure 8D: this leaves a few inter-
actions for the prestimulus period in Figure 9A (left panel), 8 out of 650, which is of the order
of the desired false-alarm rate set to 1% (namely, the extreme 0.5% of each tail). In contrast,
many more poststimulus interactions survive the significance tests in the right panel: almost all
these interactions are unidirectional. The counterpart for circular shift for Figure 9B involves 24
interactions in common with Figure 9A. On average over the four conditions, 22 poststimulus
interactions are common between the two shuffling methods, to be compared with 7 for the
prestimulus period (both with a standard deviation of 4); this corresponds to 3.5% of all possi-
ble interactions. Almost all detected interactions are unidirectional, as illustrated in Figure 9C
for both local and global tests for the poststimulus period. Varying the threshold on the tail of
the null distributions, we see that the number of detected interactions is close to the desired
false-alarm rate for the prestimulus period in Figure 9D (dark red and black curves, respec-
tively). In contrast, poststimulus interactions are many more for both local and global tests (light
red and gray). The global test detects fewer interactions than the local test, indicating the ne-
cessity to take into account the disparities across channels. Around 57% of poststimulus inter-
actions detected by the global test (largest values in absolute value) are found by the local test.
Finally, we check the relationship between the strengths of significant interactions—in ab-
solute value—and the increase of average MUAe observed in Figure 8A (lower panel).
In
Figure 9E, the plotted dots correspond to the pre-post change in the sum of incoming
(left panel) and outgoing (right panel) significant interactions for each node. The summed
interaction values positively correlate with the MUAe difference (post minus pre) only for the
incoming interactions: p = 0.03 with a coefficient of 0.21. In contrast, outgoing interactions
Network Neuroscience
374
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
1
4
3
5
7
1
0
9
1
8
7
2
n
e
n
_
a
_
0
0
0
1
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Nonparametric MVAR connectivity detection for MUA data
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
1
4
3
5
7
1
0
9
1
8
7
2
n
e
n
_
a
_
0
0
0
1
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 9. Detection of significant interactions. (A) Examples of significant interactions with the RP
surrogate method; left and right panels correspond to pre- and poststimulus periods, respectively.
The p value corresponds to the upper and lower 0.5% tails of 1,000 surrogates (local test). Many
more interactions are found for post- than prestimulus period. (B) Same as A for the CS surrogate
method and poststimulus period.
(C) Comparison of number of asymmetric interactions versus
symmetric interactions for the local (red) and global (gray) tests over the four conditions. (D) Ratios
of detected interactions for pre- and poststimulus periods with the desired false-alarm rate equal to
1% to 5% corresponding to both local and global tests. (E) Change between pre- and poststimulus
periods of the sum of incoming (left) and outgoing (right) significant weights—in absolute value—
plotted against the change in the change in mean MUAe. Each dot represents a channel, and all
four conditions are grouped here. (F) Similar plot to D for parametric GRc. (G) Similar plot to E
with sums of GRc values for each node over its incoming (dots) and outgoing (crosses) connections.
Only significant interactions passing the parametric test with p value of 0.01 in F are retained here.
Network Neuroscience
375
Nonparametric MVAR connectivity detection for MUA data
exhibit a nonsignificant negative correlation (p (cid:8) 0.1). This suggests a stimulus-driven gating
of the effective gain for incoming anatomical connections to recorded cell populations. The
application of our method thus unravels stimulus-driven directed interactions and cannot be
merely explained by an increase of single-channel MUAe activity.
In comparison, similar detection with parametric GRc testing gives more interactions for
the poststimulus period in Figure 9F (bright green), twice as much as MVAR for a p value of
0.01 (corresponding to 1% in Figure 9D). Moreover, the distributions of MVAR coefficients in
Figure 9E and the corresponding ones for GRc estimated values have similar KS distance when
comparing—for each condition—the pre- and post-periods (mean of 0.17 with a standard de-
viation of 0.01 over the four conditions). This means that GRc values collectively discriminate
between the two periods as well as the estimated MVAR coefficients. However, the pre-post
changes in incoming and outgoing GRc values strongly correlate with the change in mean
MUAe (p < 10−20
for both incoming and outgoing connections); note that the estimated con-
nectivity is not symmetric, though. This contrasts with the two plots in Figure 9E and suggests
that the two methods may capture distinct effects at work in the network of neuronal popu-
lations. Note that the nonparametric method for GRc did not work for the experimental data
here, failing to detect more interactions than the expected false-alarm rate.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
1
4
3
5
7
1
0
9
1
8
7
2
n
e
n
_
a
_
0
0
0
1
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
DISCUSSION
Nonparametric MVAR-Based Detection of Linear Feedback in Recurrent Networks
This paper proposed a nonparametric method to detect pairwise feedback connections in bi-
ological networks with possibly strong and/or dense recurrent feedback. We examined the
benefit of detecting directional connections in MVAR-like models estimated using the OLS
autoregressive coefficients instead of the error residual ratios (Figure 1). To our understanding,
the good performance of the presented method has three reasons. First, the ROC-based pre-
diction power in Figure 2, which relies on the estimated ranking of connections in the network
(i.e., from small to strong weights), is more robust for the regression coefficients than residual
log ratios for recurrent networks with relatively large density (0.1 − 0.3%); these networks over-
all imply many redundancy and convergence patterns of connections (M. Ding et al., 2006;
Stramaglia et al., 2014). Second, practical coefficient-based connectivity detection performed
using nonparametric significance tests based on time series randomization (red distribution in
Figure 4B) yields better results than conditional Granger causality ratio test in the time domain,
either with the standard parametric F test (Barnett & Seth, 2014, dashed line) or with nonpara-
metric testing (green and blue-green distributions). Finally, the use of connection-specific sig-
nificance testing achieves higher accuracy than a network-pooled alternative, especially when
asymptotic assumptions do not hold (e.g., small number of time samples), as illustrated by the
local nonparametric test in Figures 3F and 4B (red versus dark gray). Together, our results high-
light the need for testing strategies that capture the heterogeneity of sufficiently large networks
to detect individual connections. Further note that assessing the connectivity via the regression
coefficients space brings an additional advantage for network studies: The estimated connec-
tion weights can be interpreted and compared across the whole network, for example using
graph theory (Sporns, 2013).
Our approach for generating surrogate distributions can be encompassed in the family of
constrained randomization methods (Schreiber, 1998; Schreiber & Schmitz, 1996). Here we
have shown that random permutation provides a good estimation of all types of connections
(Figure 5) in false-alarm and miss rates. Comparatively, the circular shift method performs
Network Neuroscience
376
Nonparametric MVAR connectivity detection for MUA data
as well except for self-connections that are not detected at all; this also holds for nonran-
dom topologies (results not shown). Hence, preserving the autocovariance structure in the
generation of surrogates does not provide a substantial advantage here (Figure 5B–C). Both
methods show a good control for the false-alarm rate in comparison to phase randomization
(Schreiber & Schmitz, 1996) and a control Gaussian approximation over the whole network
(STD), which lead to an excess of about 1% of false alarms (i.e., ∼ 50 connections for a net-
work of 70 nodes). These results show the importance of choosing a surrogate method adapted
to the detection problem. For distinct dynamics governing the nodal activity such as non-
linearities, conclusions may differ and further research along these lines is necessary.
As mentioned earlier, the use of an individual null distribution for each connection (local
test) gives better results for the miss rate (by a few percentage) than lumping together all matrix
elements of all surrogates (global test), provided sufficiently many surrogates are generated.
For the size of networks considered here, computation time is not an issue (Figure 4D) and
our results support the choice of the local test over the global test to attain between accuracy
in the true-positive detection. This may be especially true for specific topologies or networks
with both excitatory and inhibitory connections; see Figure 6. In other words, the local test in-
corporates to a better extent the network heterogeneities in order to build the null distribution
for each connection. The present study was limited to OLS estimates for MVAR, but there exist
alternative estimators such as the locally weighted least square regression (Ruppert & Wand,
1994) that may perform better for particular network topologies. The extension of the pre-
sented surrogate techniques to the case where observations are sparser than connections—
implying that the covariance matrix is not invertible—is another interesting direction to explore
(Castelo & Roverato, 2006).
The problem of multiple comparison is intrinsic to brain connectivity detection as the num-
ber of testable connections across brain regions is massive (Rubinov & Sporns, 2010). In this
context, different approaches have been developed to control the family-wise error rate in the
weak sense. For instance, many studies on neuroimaging data (Genovese, Lazar, & Nichols,
2002; Nichols & Hayasaka, 2003) or electrophysiology (Lage-Castellanos, Martínez-Montes,
Hernández-Cabrera, & Galán, 2010) have resorted to procedures that control the false dis-
covery rate (FDR) (Benjamini & Hochberg, 1995), namely, the expected number of falsely de-
clared connections among the total number of detections. These methods make decisions on
single connections relying on the entire sequence of p values computed for each connection
and yield substantial statistical power gains over more conservative methods such as Šidák-
Bonferroni (Abdi, 2007). With the ever growing application of graph theory to brain connec-
tivity, new methods have been proposed that exploit the clustered structure of the declared
connections (Han, Yoo, Seo, Na, & Seong, 2013; Zalesky, Fornito, & Bullmore, 2010) to pro-
pose cluster-based statistical tests (Maris & Oostenveld, 2007) attaining similar performance
to FDR methods. The present work can therefore be understood as a primary step before per-
forming any or several multiple-correction procedures. By defining an accurate null model
of inexistent connections, p value estimates per connection are improved and cluster-based
surrogate distributions can be better approximated, which is expected to empower the overall
control of false positive rates in network connectivity analysis.
Applications to Real Electrophysiological and Neuroimaging Data
A motivation for our method is the detection of neuronal interactions between electrode
channels, which is often performed using spectral Granger causality analysis on local-field
potential (low-passed signal of electrode measurements) or electrocorticography measure-
Network Neuroscience
377
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
1
4
3
5
7
1
0
9
1
8
7
2
n
e
n
_
a
_
0
0
0
1
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Nonparametric MVAR connectivity detection for MUA data
ments. As an alternative, we have applied our connectivity detection method to MUAe recorded
from macaque area V1 in order to provide a proof of concept. Multichannel recording devices
have been developed in the past years to obtain this type of data (Fan et al., 2011; Roy & Wang,
2012) and a recent cognitive study has highlighted group properties of MUAe activity for a
similar experimental setup to the one used here (Engel et al., 2016). Looking at the variabil-
ity for individual trials in Figure 8A (upper and middle panels), it is rather surprising that the
MUAe conveys temporal information about joint activity for pairs of channels (Figure 8B),
which can be related to causal directed interactions. This means that the high trial-to-trial
variability is not an absolute limitation to temporal coordination, even though the latter only
becomes apparent over multiple trial repetitions, just as the poststimulus increase of MUAe
for single channels (lower panel in Figure 8A). The procedure detects a number of significant
directional interactions well above the false positive rate that were not merely explained by
the MUAe increase (Figures 9D and E). In comparison, the control for the prestimulus period
in the four different conditions detects just above the expected false-alarm rate. We find that
the parametric Granger causality test also detects many more interactions after the stimulus
than before; however, these interactions happen to link channels exhibiting the strongest in-
creases in MUAe activity. This presents the caveat of providing little information in addition
to the changes observed at the single-node level, when interpreting the obtained results. The
research of optimal preprocessing—in particular the filtering to obtain MUAe—to obtain a
robust detection of interactions is left for a later study. Likewise, such electrode recordings
are often analyzed with respect to specific frequency bands (e.g., alpha and gamma), but the
adaptation of our framework along the lines of previous work (Dhamala et al., 2008; L. Ding
et al., 2007) and comparison of detected interactions with established methods will be done
in future research.
More generally, our methodology requires adequate preprocessing of multivariate time
series—activity aggregation over hundreds of voxels for fMRI and 4-ms smoothing of MUAe
for electrode recordings—such that the autocovariance profiles match the exponential de-
cay of the dynamic network model with linear feedback, which underlies the connectivity
analysis. Although filtered and smoothed signals fall into the class of autoregressive moving-
average (ARMA) models, our approach is applicable provided the autocovariances exhibit a
profile resembling Figure 8C. We further expect nonparametric testing methods for be extend-
able to ARMA processes, complementing approaches developed by Barnett & Seth (2015) and
Friston et al. (2014). In theory, stationarity of the time series remains a critical issue as we need
sufficiently many observed samples to obtain precise covariances from which we estimate
the connectivity, but this may not be a strong limitation for MUAe in practice in view of the
poststimulus average response shown in Figure 8A.
ACKNOWLEDGMENTS
The authors are grateful to Robert Castelo and Inma Tur for constructive and inspirational
discussions.
AUTHOR CONTRIBUTIONS
Matthieu Gilson: Conceptualization (equal); Formal Analysis; Software; Writing – original
Draft (equal). Adriá Tauste Campo: Conceptualization (equal); Formal Analysis (equal);
Writing – original Draft (equal). Xing Chen:
Investigation. Alexander Thiele: Resources;
Writing – original Draft (supporting). Gustavo Deco: Resources; Writing – original Draft
(supporting).
Network Neuroscience
378
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
1
4
3
5
7
1
0
9
1
8
7
2
n
e
n
_
a
_
0
0
0
1
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Nonparametric MVAR connectivity detection for MUA data
FUNDING INFORMATION
MG acknowledges funding from the Marie Sklodowska-Curie Action (grant H2020-MSCA-
656547). MG and GD were supported by the Human Brain Project (grant FP7-FET-ICT-604102
and H2020-720270 HBP SGA1). GD and ATC were supported by the European Research
Council Advanced Grant DYSTRUCTURE (Grant 295129). AT was supported by the UK
Medical Research Council (grant MRC G0700976).
REFERENCES
Abdi, H.
(2007). The bonferonni and šidák corrections for multi-
ple comparisons. Encyclopedia of Measurement and Statistics,
3, 103–107.
Amemiya, T.
(1974). Multivariate regression and simultaneous
equation models when the dependent variables are truncated
Journal of the Econometric Society,
normal.
42(6), 999–1012.
Econometrica:
Babiloni, F., Cincotti, F., Babiloni, C., Carducci, F., Mattia, D.,
Astolfi, L., . . . He, B. (2005). Estimation of the cortical functional
connectivity with the multimodal integration of high-resolution
EEG and fMRI data by directed transfer function. Neuroimage,
24(1), 118–131.
Barnett, L., & Seth, A.
(2014). The MVGC multivariate Granger
causality toolbox: A new approach to Granger-causal inference.
Journal of Neuroscience Methods, 223, 50–68.
Barnett, L., & Seth, A.
(2015). Granger causality for state-space
models. Physical Review E, 91, 040101. https://doi.org/10.1103/
PhysRevE.91.040101
Barrett, A., & Barnett, L. (2013). Granger causality is designed to
measure effect, not mechanism. Frontiers in Neuroinformatics,
7, 6. https://doi.org/10.3389/fninf.2013.00006
Benjamini, Y., & Hochberg, Y. (1995). Controlling the false discov-
ery rate: A practical and powerful approach to multiple testing.
Journal of the Royal Statistical Society. Series B (Methodological),
289–300.
Castelo, R., & Roverato, A. (2006). A robust procedure for Gaussian
graphical model search from microarray data with p larger than
n. The Journal of Machine Learning Research, 7, 2621–2650.
Retrieved from http://dl.acm.org/citation.cfm?id=1248641
Cortex [Computer software].
Laboratory of Neuropsychology,
NIMH. http:dally.nimh.nih.gov/index.html
Dhamala, M., Rangarajan, G., & Ding, M.
(2008). Analyzing in-
formation flow in brain networks with nonparametric Granger
causality. NeuroImage, 41(2), 354–362.
Diks, C., & DeGoede, J.
(2001). A general nonparametric boot-
strap test for Granger causality.
In Bernd Krauskopf, Henk W.
Broer, & Gert Vegter (Eds.), Global analysis of dynamical systems
(pp. 391–403). Bristol, UK: Institute of Physics Publishing.
Ding, L., Worrell, G., Lagerlund, T., & He, B. (2007). Ictal source
analysis: Localization and imaging of causal interactions in hu-
mans. NeuroImage, 34(2), 575–586.
Ding, M., Chen, Y., & Bressler, S. (2006). Granger causality: Basic
theory and application to neuroscience.
In B. Schelter, M.
Winterhalder, & J. Timmer (Eds.), Handbook of time series
analysis: Recent theoretical developments and applications.
arXiv preprint q-bio/0608035: Wiley-VCH Verlage.
Engel, T. A., Steinmetz, N. A., Gieselmann, M. A., Thiele, A.,
(2016). Selective modulation of cor-
Moore, T., & Boahen, K.
tical state during spatial attention. Science, 354, 1140–1144.
Faes, L., Marinazzo, D., Montalto, A., & Nollo, G.
(2014).
Lag-specific transfer entropy as a tool to assess cardiovascu-
lar and cardiorespiratory information transfer. IEEE Transactions
on Biomedical Engineering, 61, 2556–2568. https://doi.org/10.
1109/TBME.2014.2323131
Fan, D., Rich, D., Holtzman, T., Ruther, P., Dalley, J. W., Lopez, A.,
. . . Yin, H. H. (2011). A wireless multi-channel recording system
for freely behaving mice and rats. PLoS ONE, 6, e22033.
Friston, K., Bastos, A., Oswal, A., van Wijk, B., Richter, C., &
Litvak, V. (2014). Granger causality revisited. NeuroImage, 101,
796–808. https://doi.org/10.1016/j.neuroimage.2014.06.062
Friston, K., Harrison, L., & Penny, W.
(2003). Dynamic causal
modelling. NeuroImage, 19(4), 1273–1302.
Genovese, C., Lazar, N., & Nichols, T. (2002). Thresholding of sta-
tistical maps in functional neuroimaging using the false discovery
rate. NeuroImage, 15(4), 870–878.
Geweke, J. (1982). Measurement of linear dependence and feed-
back between multiple time series. Journal of the American Sta-
tistical Association, 77, 304–313.
Geweke, J. (1984). Measures of conditional linear dependence and
feedback between time series. Journal of the American Statistical
Association, 79, 907–915.
Granger, C. (1969). Investigating causal relations by econometric
models and cross-spectral methods. Econometrica: Journal of
the Econometric Society, 424–438.
Han, C., Yoo, S., Seo, S., Na, D., & Seong, J.-K.
(2013). Cluster-
based statistics for brain connectivity in correlation with behav-
ioral measures. PLoS ONE, 8(8), e72332.
Kaminski, M., & Blinowska, K.
(1991). A new method of the de-
scription of the information flow in the brain structures. Biological
Cybernetics, 65(3), 203–210.
Kami ´nski, M., Ding, M., Truccolo, W., & Bressler, S. (2001). Eval-
uating causal relations in neural systems: Granger causality, di-
rected transfer function and statistical assessment of significance.
Biological Cybernetics, 85(2), 145–157.
Lage-Castellanos, A., Martínez-Montes, E., Hernández-Cabrera, J.,
& Galán, L.
(2010). False discovery rate and permutation test:
An evaluation in ERP data analysis. Statistics in Medicine, 29(1),
63–74.
Li, Y., Ye, X., Liu, Q., Mao, J., Liang, P., Xu, J., & Zhang, P. (2016).
Localization of epileptogenic zone based on graph analysis of
stereo-EEG. Epilepsy Research, 128, 149–157.
Network Neuroscience
379
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
1
4
3
5
7
1
0
9
1
8
7
2
n
e
n
_
a
_
0
0
0
1
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Nonparametric MVAR connectivity detection for MUA data
Lusch, B., Maia, P., & Kutz, J. (2016). Inferring connectivity in net-
worked dynamical systems: Challenges using Granger causality.
Physical Review E, 94, 032220.
Lütkepohl, H.
(2005). New introduction to multiple time series
analysis. Berlin: Springer Science & Business Media.
Maris, E., & Oostenveld, R. (2007). Nonparametric statistical test-
Journal of Neuroscience Methods,
ing of EEG- and MEG-data.
164(1), 177-190.
Massey, J.
(1990). Causality, feedback and directed information.
Proceedings ISITA 1990 International Symposium, 303–305.
Messé, A., Rudrauf, D., Benali, H., & Marrelec, G. (2014). Relat-
ing structure and function in the human brain: Relative contri-
butions of anatomy, stationary dynamics, and non-stationarities.
PLoS Computational Biology, 10, e1003530.
Michalareas, G., Schoffelen, J., Paterson, G., & Gross, J. (2013). In-
vestigating causality between interacting brain areas with multi-
variate autoregressive models of MEG sensor data. Human Brain
Mapping, 34(4), 890–913. https://doi.org/10.1002/hbm.21482
Nedungadi, A., Rangarajan, G., Jain, N., & Ding, M. (2009). Ana-
lyzing multiple spike trains with nonparametric Granger causal-
ity. Journal of Computational Neuroscience, 27(1), 55–64.
Nichols, T., & Hayasaka, S. (2003). Controlling the familywise error
rate in functional neuroimaging: A comparative review. Statisti-
cal methods in medical research, 12(5), 419–446.
Rogers, B., Katwal, S., Morgan, V., Asplund, C., & Gore, J. (2010).
Functional MRI and multivariate autoregressive models. Mag-
netic Resonance Imaging, 28(8), 1058–1065.
Roy, S., & Wang, X.
recording in freely moving and vocalizing primates.
Neuroscience Methods, 203, 28–40.
(2012). Wireless multi-channel single unit
Journal of
Rubinov, M., & Sporns, O. (2010). Complex network measures of
brain connectivity: Uses and interpretations. NeuroImage, 52(3),
1059–1069.
Ruppert, D., & Wand, M.
(1994). Multivariate locally weighted
least squares regression. Annals of Statistics, 22, 1346-1370.
Schreiber, T. (1998). Constrained randomization of time series data.
Physical Review Letters, 80, 2105.
Schreiber, T. (2000). Measuring information transfer. Physical Re-
view Letters, 85, 461.
Schreiber, T., & Schmitz, A.
(1996).
Improved surrogate data for
nonlinearity tests. Physical Review Letters, 77, 635.
Scientific Python Library. (n.d.). http://www.scipy.org
Seth, A., Barrett, A., & Barnett, L. (2015). Granger causality analysis
Journal of Neuroscience,
in neuroscience and neuroimaging.
35, 3293–3297. https://doi.org/10.1523/JNEUROSCI.4399-14.
2015
So, K., Koralek, A., Ganguly, K., Gastpar, M., & Carmena, J. (2012).
Assessing functional connectivity of neural ensembles using
Journal of Neural Engineering, 9(2),
directed information.
026004.
Sporns, O.
(2013). Making sense of brain network data. Nature
Methods, 10, 491–493.
Storkey, A., Simonotto, E., Whalley, H., Lawrie, S., Murray, L., &
Mcgonigle, D. (2007). Learning structural equation models for
In B. Schölkopf, J. Platt, & T. Hoffman (Eds.), Advances
fMRI.
in neural information processing systems 19 (pp. 1329–1336).
Cambridge, MA: MIT Press.
Stramaglia, S., Cortes, J., & Marinazzo, D. (2014). Synergy and re-
dundancy in the Granger causal analysis of dynamical networks.
New Journal of Physics, 16, 105003.
Supèr, H., & Roelfsema, P. (2005). Chronic multiunit recordings in
behaving animals: Advantages and limitations. Progress in Brain
Research, 147, 263–282.
Tauste Campo, A., Martinez-Garcia, M., Nácher, V., Luna, R.,
Romo, R., & Deco, G.
(2015). Task-driven intra- and inter-
area communications in primate cerebral cortex. Proceedings
of the National Academy of Sciences, 112(15), 4761–4766.
Thiele, A., Delicato, L., Roberts, M., & Gieselmann, M.
(2006).
A novel electrode-pipette design for simultaneous recording
of extracellular spikes and iontophoretic drug application in
awake behaving monkeys. Journal of Neuroscience Methods,
158, 207–211.
Vinck, M., Huurdeman, L., Bosman, C., Fries, P., Battaglia, F.,
Pennartz, C., & Tiesinga, P. (2015). How to detect the Granger-
causal flow direction in the presence of additive noise? Neuro-
Image, 108, 301–318. http://doi.org/10.1016/j.neuroimage.
2014.12.017
Wilke, C., Ding, L., & He, B. (2008). Estimation of time-varying con-
nectivity patterns through the use of an adaptive directed transfer
IEEE Transactions on Biomedical Engineering, 55(11),
function.
2557–2564.
Winkler, A., Ridgway, G., Webster, M., Smith, S., & Nichols, T.
(2014). Permutation inference for the general linear model. Neu-
roImage, 92, 381-397. https://doi.org/10.1016/j.neuroimage.
2014.01.060
Zalesky, A., Fornito, A., & Bullmore, E.
(2010). Network-based
statistic: Identifying differences in brain networks. NeuroImage,
53(4), 1197–1207.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
1
4
3
5
7
1
0
9
1
8
7
2
n
e
n
_
a
_
0
0
0
1
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network Neuroscience
380