Magnetoencephalographic Activity Related to Conscious
Perception Is Stable within Individuals across Years
but Not between Individuals
Kristian Sandberg1,2,5, Gareth Robert Barnes2, Geraint Rees2,3,
and Morten Overgaard1,4
D
o
w
n
l
o
a
d
e
d
f
r
o
m
Abstract
■ Studies indicate that conscious perception is related to
changes in neural activity within a time window that varies be-
tween 130 and 320 msec after stimulus presentation, yet it is
not known whether such neural correlates of conscious percep-
tion are stable across time. Here, we examined the generalization
across time within individuals and across different individuals.
We trained classification algorithms to decode conscious percep-
tion from neural activity recorded during binocular rivalry using
magnetoencephalography (MEG). The classifiers were then used
to predict the perception of the same participants during differ-
ent recording sessions either days or years later as well as
between different participants. No drop in decoding accuracy
was observed when decoding across years compared with days,
whereas a large drop in decoding accuracy was found for
between-participant decoding. Furthermore, underlying percept-
specific MEG signals remained stable in terms of latency, am-
plitude, and sources within participants across years, whereas
differences were found in all of these domains between individ-
uals. Our findings demonstrate that the neural correlates of con-
scious perception are stable across years for adults, but differ
across individuals. Moreover, the study validates decoding based
on MEG data as a method for further studies of correlations be-
tween individual differences in perceptual contents and between-
participant decoding accuracies. ■
INTRODUCTION
The last two decades have seen an upsurge of empirical
research into the neural correlates of conscious percep-
tion, yet it remains unknown whether candidate neural
signatures of conscious perception determined by such
earlier work are stable across time. Here, we addressed
this question by measuring and comparing magneto-
encephalography (MEG) signals associated with conscious
perception that were acquired from the same individuals
days or years apart.
We focused on one very consistent neural correlate
of conscious perception, the so-called visual awareness
negativity (VAN; Koivisto & Revonsuo, 2003). VAN refers
to awareness-specific event-related EEG activity occurring
within a time window of around 130–320 msec after stimu-
lus presentation and has been observed in more than
30 independent EEG studies (Koivisto & Revonsuo, 2010),
and at least three MEG experiments have reported activ-
ity corresponding to the VAN (Sandberg et al., 2013;
Liu, Paradis, Yahia-Cherif, & Tallon-Baudry, 2012; Vanni,
Revonsuo, Saarinen, & Hari, 1996). The VAN consists of
1Aarhus University Hospital, 2University College London, 3Well-
come Trust Centre for Neuroimaging, Institute of Neurology,
London, UK, 4Aalborg University, 5Hammel Neurorehabilitation
and Research Center
an early and a late part (Koivisto & Revonsuo, 2010), and
in some experiments, these two parts of the VAN are
observed as separate ERP/event-related field (ERF) com-
ponents (e.g., Sandberg et al., 2013; Fahrenfort, Scholte, &
Lamme, 2007).
Although there are differences in the interpretation of
the VAN and its individual parts/components, studies with
very different theoretical backgrounds consistently report
VAN-like findings. Koivisto and Revonsuo (2010), for in-
stance, interpret the VAN as reflecting recurrent process-
ing in sensory areas associated with conscious experience
in the theory of Lamme (2010), and research by Lammeʼs
own group report activity around the first VAN component
at 110–210 msec as the main correlate of visual aware-
ness in masking tasks (Van Loon, Scholte, van Gaal,
van der Hoort, & Lamme, 2012; Fahrenfort et al., 2007).
In contrast, Dehaene and others report that signals around
the second VAN component, peaking around 270 msec,
correlate with subjective, graded ratings of visibility,
although they consider later, bimodal responses as the cor-
relates of conscious report (Dehaene, Changeux, Naccache,
Sackur, & Sergent, 2006; Sergent, Baillet, & Dehaene,
2005; Sergent & Dehaene, 2004; Dehaene, Kerszberg, &
Changeux, 1998). In this study, we remain agnostic regard-
ing the theoretical interpretation of the VAN, but simply
use it as the object of our analysis as one of the components
that correlate with the content of consciousness.
© 2014 Massachusetts Institute of Technology Published under a
Creative Commons Attribution 3.0 Unported (CC-BY 3.0) license
Journal of Cognitive Neuroscience 26:4, pp. 840–853
doi:10.1162/jocn_a_00525
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
6
/
4
2
6
8
/
4
4
0
/
1
8
9
4
4
0
6
/
6
1
2
7
3
8
o
1
c
0
n
1
_
4
a
/
_
j
0
o
0
c
5
n
2
5
_
a
p
_
d
0
0
b
5
y
2
g
5
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
.
/
t
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Several studies have examined the stability of different
aspects of the EEG signal within individuals across time.
Many such studies have focused on visual-evoked poten-
tials ( VEPs), typically evoked by a reversing checkerboard
stimulus, which elicit two early components, an N75
(at around 75 msec) and a P100 (at around 100 msec).
For latency, moderate-to-strong correlations of 0.3–0.8
is often found across 0.2–13 months, although for the
P100 correlations as high as .93 have been observed
(Sarnthein, Andersson, Zimmermann, & Zumsteg, 2009;
Oken, Chiappa, & Gill, 1987; Hall, Rappaport, Hopkins,
& Griffin, 1973). Also for amplitude, defined as the
N75-P100 difference, moderate correlations of .4–.7 have
been reported (Sarnthein et al., 2009; Schellberg, Gasser,
& Köhler, 1987). One study reported that interindividual
VEP differences were generally larger than intraindi-
vidual differences (Sarnthein et al., 2009), although for
children aged 10–13 years, intraindividual differences
across 10 months appear to be of the same magnitude
as interindividual differences (Schellberg et al., 1987).
Other studies have examined error-related negativity, a
component found around 50 msec after the individual
makes an error, and found a moderate-to-strong ampli-
tude correlation across 2–6 weeks and a moderate cor-
relation across 1.5–2.5 years (Weinberg & Hajcak, 2011).
Finally, studies have reported relatively high stability of
the EEG power spectra related to resting as well as related
to performing a working memory task across up to
40 months (Näpflin, Wildi, & Sarnthein, 2007, 2008).
Although these previous studies indicate that some
degree of interindividual difference is to be expected
and possibly that smaller intraindividual difference is to
be expected, it is difficult to draw parallels between
studies because of the difference in the examined com-
ponents. Most of the reported studies were concerned
with very early components, which are generally not
considered to be modulated by conscious content. Fur-
thermore, this study examines not only the components
related to a certain visual stimulation but also endogenous,
consciousness-specific modulation of these components,
thus making it difficult to make predictions regarding the
stability of activity in the VAN time window across years
based on previous studies.
To test the stability of the VAN over extended pe-
riods of time, we used intermittent binocular rivalry (BR;
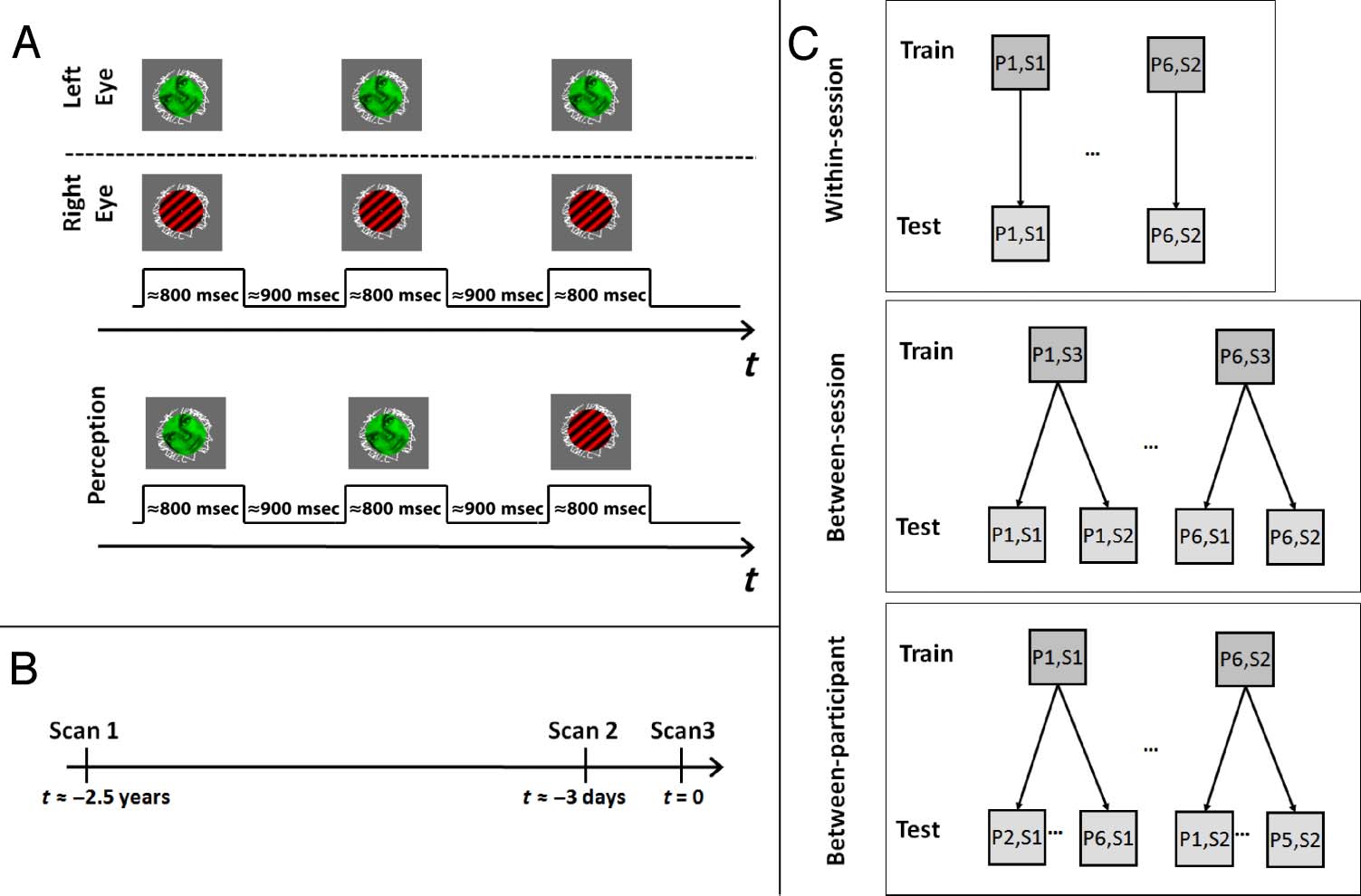
Figure 1A). During BR, two images are presented dich-
optically, and perception alternates spontaneously between
the two images. When BR stimuli are presented inter-
mittently, that is, for short durations of less than a second
separated by a short break with no visual stimulation, the
Figure 1. Experimental
design. (A) Rivaling stimuli
(face/grating) were presented in
trials of ∼800 msec separated
by blank periods of ∼900 msec.
Stimuli were dichoptically
presented to the eyes of the
participant and rotated in
opposite directions at a rate
of 0.7 rotations per second.
Participants reported which
of the two images they
perceived with a button press.
(B) Time line of recording
sessions. Session 1 was
performed around 2.5 years
before Session 2, which was
performed 2–5 days before
Session 3. (C) Classification
procedure. SVMs were trained
to distinguish MEG activity
related to conscious face and
grating perception for each
participant. The SVMs were
then used to decode the
perception of (1) the same
participant (P) on different
trials from the same recording
session (S; top), (2) trials from
the same participant but from
a different recording session
(middle), and (3) trials from
recording sessions from
each of the other participants
(bottom).
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
6
/
4
2
6
8
/
4
4
0
/
1
8
9
4
4
0
6
/
6
1
2
7
3
8
o
1
c
0
n
1
_
4
a
/
_
j
0
o
0
c
5
n
2
5
_
a
p
_
d
0
0
b
5
y
2
g
5
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
f
/
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Sandberg et al.
841
participant typically reports only one percept per trial,
thus allowing examination of percept-specific signals that
are time-locked to stimulus onset.
We examined whether multivariate support vector
machines (SVM) could decode the percept that a partici-
pant reported on any given trial when trained and tested
on different data sets, that is, when the training data
came from the same participant, but was gathered days
or even years apart from the testing data, or when the
training and testing data came from different individuals
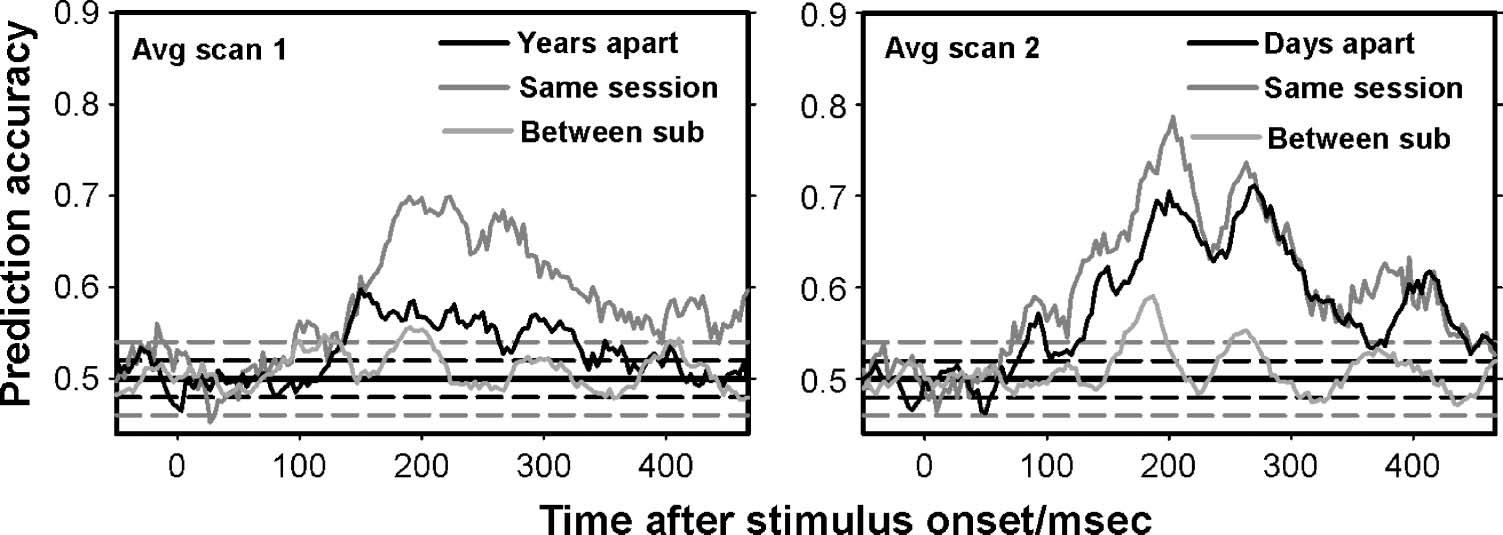
(Figure 1B, C). The degree to which prediction accuracy
did (or did not) fall when generalizing across data sets
in this way served as an index of the stability of the
multivariate signal that correlated with conscious percep-
tion at baseline, either over time within the same indi-
vidual or across individuals at the same time point. We
further explicitly examined the latency and amplitude
of ERF components during the VAN as well as the acti-
vated sources to interpret changes in decoding accuracy
more fully.
The MEG of eight participants was recorded in Session 1,
and the analyses reported here were based on an in-depth
examination of four participants who were available
2.5 years later. The full analysis of all eight participants of
recording Session 1 is reported elsewhere (Sandberg et al.,
2013; see Methods for details). We emphasize that the
drop in between-participant prediction accuracy reported
in this study of four participants is highly similar to the
drop previously reported for the full group of eight par-
ticipants, thus indicating that the current four participants
are representative of the group. We further emphasize
that significant differences observed with a relatively
small sample of participants reflect that the effect size is
larger (and thus more relevant) than if a larger sample
of participants were needed to obtain the same level of
significance (Friston, 2012).
METHODS
As mentioned above, comprehensive analyses of the data
from Session 1 are reported elsewhere (Sandberg et al.,
2013). The goal of this previous study was to examine
the MEG correlates of the content of consciousness during
intermittent BR. The main analysis was decoding using an
SVM classifier, and decoding was based on field strength
amplitude (using various filters) as well as power estimates
across a wide range of power spectra, and it was estab-
lished that field strength amplitude was the most infor-
mative signal. Furthermore, the potential advantage of
feeding the SVM with data across multiple time points
was examined, yet no such advantage was found. Analyses
were performed at the sensor level as well as on data re-
constructed at a number of sources distributed across
cortex achieving similar results in the two conditions. Ad-
ditionally, successful decoding across participants was
achieved, yet a relatively large drop in decoding accuracy
was observed.
On the basis of the results of this previous study, de-
coding using field strength data at single time points was
chosen as the main method of analysis in this study. This
study expands on the previous by examining the stability
of consciousness-specific modulation of neural activ-
ity across time for the first time and by performing an
in-depth analysis of the cause of the drop in between-
participant decoding. To accomplish this, the approach
was supplemented by three new analyses. Specifically,
we estimated the power and latency of the components
previously found to predict conscious content, and we
used multivariate Bayesian (MVB) modeling to estimate
the source composition of these components. All these
methods are explained in-depth below.
Participants
Eight healthy adults (six women) with normal or corrected-
to-normal vision gave written informed consent to partici-
pate in the experiment and performed recording Session 1.
Four of these (three women) were available 2.5 years
later for recording Sessions 2 and 3. Data for these four
participants were analyzed here. The ages of the partici-
pants was between 26 and 32 years (mean = 29 years,
SD = 2.5 years). The experiments were approved by the
University College London Research Ethics Committee.
Apparatus and MEG Recording
Stimuli were generated using the MATLAB toolbox
Cogent (www.vislab.ucl.ac.uk/cogent.php). They were
projected onto a 19-in. screen (resolution = 1024 ×
768 pixels; refresh rate = 60 Hz) using a JVC D-ILA,
DLA-SX21 projector. Participants viewed the stimuli
through a mirror stereoscope positioned at approximately
50 cm from the screen. MEG data were recorded in a
magnetically shielded room with a 275-channel CTF
Omega whole-head gradiometer system (VSM MedTech,
Coquitlam, BC, Canada) with a 600-Hz sampling rate.
Head localizer coils were attached to the nasion and
1 cm anterior of the left and right outer canthus to
monitor head movement during the recording sessions
and to establish head position for source analysis.
Stimuli
A red Gabor patch (contrast = 100%, spatial frequency =
3 cycles/degree, SD of the Gaussian envelope = 10 pixels)
was presented to the right eye of the participants, and
a green, female face was presented to the left eye (Fig-
ure 1A). To avoid piecemeal rivalry, the stimuli rotated
at a rate of 0.7 rotations per second in opposite directions,
and to ensure that stimuli were perceived in overlapping
areas of the visual field, each stimulus was presented
within an annulus (inner/outer r = 1.3/1.6 degrees of
visual angle) consisting of randomly oriented lines. In the
center of the circle was a small circular fixation dot.
842
Journal of Cognitive Neuroscience
Volume 26, Number 4
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
6
/
4
2
6
8
/
4
4
0
/
1
8
9
4
4
0
6
/
6
1
2
7
3
8
o
1
c
0
n
1
_
4
a
/
_
j
0
o
0
c
5
n
2
5
_
a
p
_
d
0
0
b
5
y
2
g
5
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
.
t
f
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Procedure
The MEG of the participants was recorded on three sepa-
rate occasions. Recording 1 was at around t = −2.5 years,
Recording 2 was at around t = −3 days, and Recording 3
was at t = 0. For all recordings, the following procedure
was used.
First, the stereoscope was calibrated by adjusting the
mirrors until the fixation circles around the stimuli fused.
Second, to minimize perceptual bias (Carter & Cavanagh,
2007), the relative luminance of the images was adjusted
for each participant until each image was reported
equally often (±5%) during a 1-min long continuous pre-
sentation. During both these calibrations runs and the
actual experiment participants reported their perception
using three buttons each corresponding to either face,
grating, or mixed perception. Participants swapped the
hand used to report between blocks. This was done to
prevent the classification algorithm from associating a
perceptual state with neural activity related to a specific
motor response.
Each participant completed six to nine runs of 12 blocks
of 20 trials, that is, 1440–2160 trials were completed per
participant. On each trial, the stimuli were displayed for
approximately 800 msec (this stimulation period was cali-
brated individually for each participant so that the percept
had time to form and did not switch during the stimula-
tion period—for all participants, the stimulus duration
was between 750 and 850 msec). Each trial was separated
by a uniform gray screen appearing for around 900 msec.
Between blocks, participants were given a short break of
8 sec. After each run, participants signaled when they
were ready to continue.
Preprocessing
Using SPM8 (www.fil.ion.ucl.ac.uk/spm/), data were
downsampled to 300 Hz and high-pass filtered at 1 Hz.
Behavioral reports of perceptual state were used to
divide stimulation intervals into face, grating, or mixed
epochs starting 200 msec before stimulus onset and ending
600 msec after. Trials containing artifacts were rejected at
a threshold of 3 pT.
Source Space Activity Reconstruction
To minimize confounds caused by differences in the head
position of the participant in the MEG system between
recording sessions, some analyses were performed on
data reconstructed in source space. The source reconstruc-
tion took into account the head position of the partici-
pants, which was measured at the beginning and end
of each run using localizer coils as described above. The
mean difference in position between start and end head
position was 3.4 mm (SD = 2.1). We identified the sources
that were most active 0–400 msec after stimulus onset
using the multiple sparse priors (MSP) algorithm (Friston,
Harrison, et al., 2008). MSP operates by finding the mini-
mum number of patches on a canonical cortical mesh that
explain the largest amount of variance in the MEG data;
this tradeoff between complexity and accuracy is optimized
through maximization of model evidence. The MSP per-
formed a group-level reconstruction based on template
structural MR scans using all trials (labeled identically)
from all 12 recording sessions (4 participants × 3 sessions/
participant). The 35 most active sources were identified
(see Table 1). SPM was then used to reconstruct the activ-
ity of the correctly (face/grating) labeled trials across the
35 sources for each recording, and these data sets were
used in the analyses below. Note that the use of generic
anatomy (head model and cortical mesh) means that dif-
ferences in source reconstruction will be due primarily to
differences in sensor level data (rather than differences
in cortical folding over individuals). For this reason, the
use of generic models will result in more similar source
estimates over subjects than the use of models based on
individual MRIs, and larger differences between individuals
than within individuals (between sessions) cannot be an
artifact of the source estimation process.
Multivariate Prediction Analysis
Multivariate pattern classification of the evoked responses
was performed using the linear SVM of the MATLAB Bio-
informatics Toolbox (Mathworks, Natick, MA). The SVM
attempted to decode the trial type (face or grating) inde-
pendently for each time point along the epoch. Classifica-
tion was based on 2–10 Hz filtered data, as the components
of interest in the 130–320 msec time windows are <10 Hz.
Application of a 1–20 Hz filter or 1-Hz high-pass filter
only does not change MEG results qualitatively (Sandberg
et al., 2013).
We attempted to decode conscious perception both
within participants across time and between participants.
For all analyses, 100 randomly selected trials each kind
(face>
7.6 and p < .0001 in all cases].
advantage of source-based versus sensor-based decoding
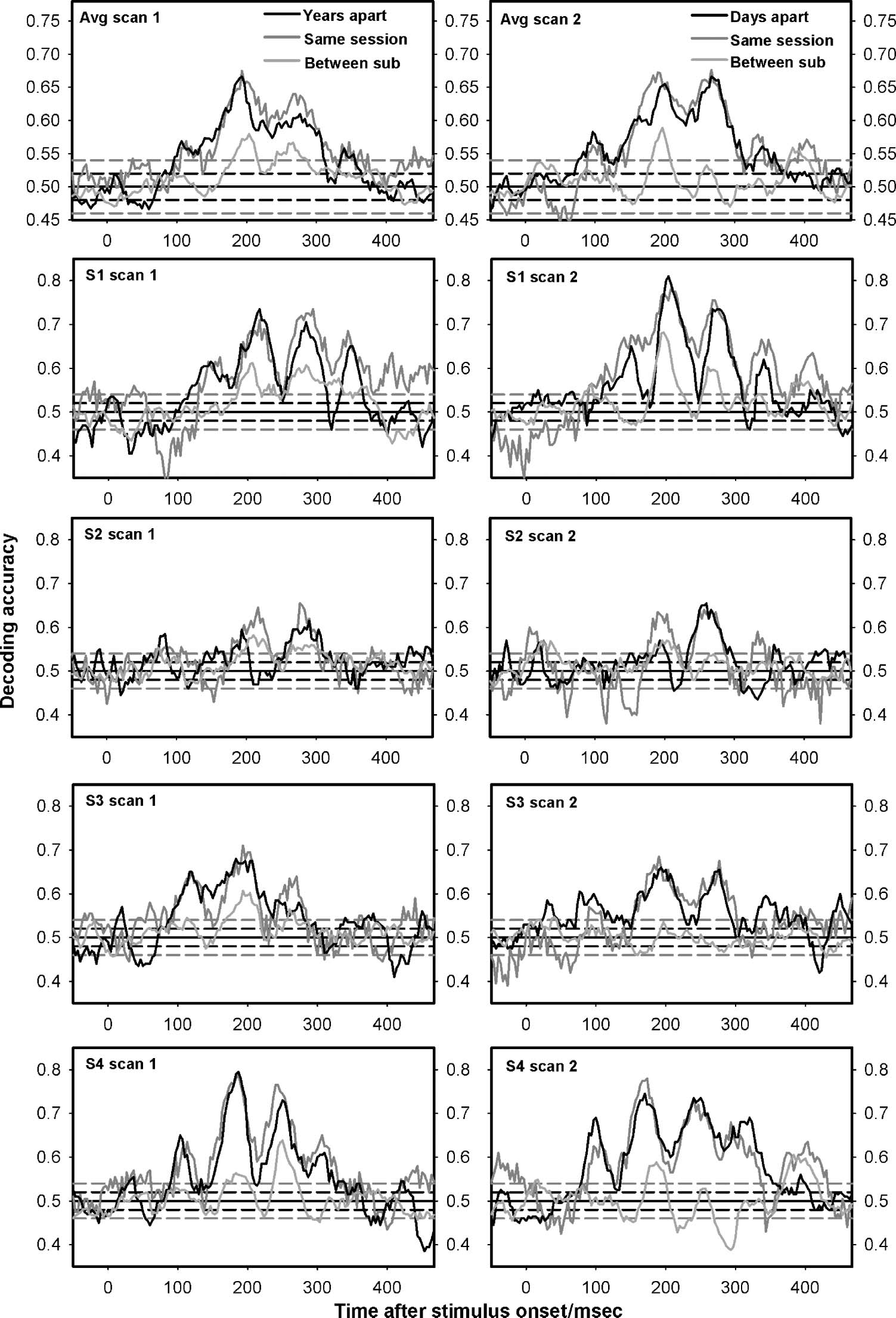
is presented in Figure 5. In sensor space, a large drop in
decoding accuracy was observed across years. As no such
drop was not observed in source space, the sensor space
difference should not be taken as a sign of difference in
neural activity, but rather a trivial difference in head posi-
tion demonstrating the necessity of source space activity
reconstruction.
As mentioned above, all decoding analyses were per-
formed using data reconstructed at the source level. The
The VAN is most often found around two ERF compo-
nents around the 130–320 msec time range (Koivisto &
Sandberg et al.
847
Figure 5. Average decoding
accuracy across subjects when
classifiers were trained and
tested on sensors data instead
of source data. SVMs were
trained to decode perception.
Decoding accuracy is plotted for
conditions where the SVM
training and testing data came
from the same participants and
the same recording session
(dark gray lines), for conditions
where training and testing
data came from the same
participant but were gathered
days (right, black line) or years (left, black line) apart, and finally for conditions where training and testing data came from different participants
(light gray lines). Horizontal, solid black lines represent chance. Horizontal, dotted black and gray lines represent the 95% binomial confidence
interval around chance (uncorrected) for between-participant and within-participant decoding, respectively. Note the large drop in classification
accuracy across years compared with when analyses were performed in source space (Figure 1). The topography thus differed across years,
whereas the sources did not, indicating that head position in relation to the sensors was a large artifact for sensor space decoding. For this
reason, analyses in the main text were performed on data reconstructed in source space.
Revonsuo, 2010). For face processing, these two compo-
nents are the M170 and the P2m (Sandberg et al., 2013).
The drop in decoding accuracy for between-participant
versus within-participant classification could be caused
by changes in the latency, amplitude, as well as the
sources of these two components. All these aspects are
examined in the following.
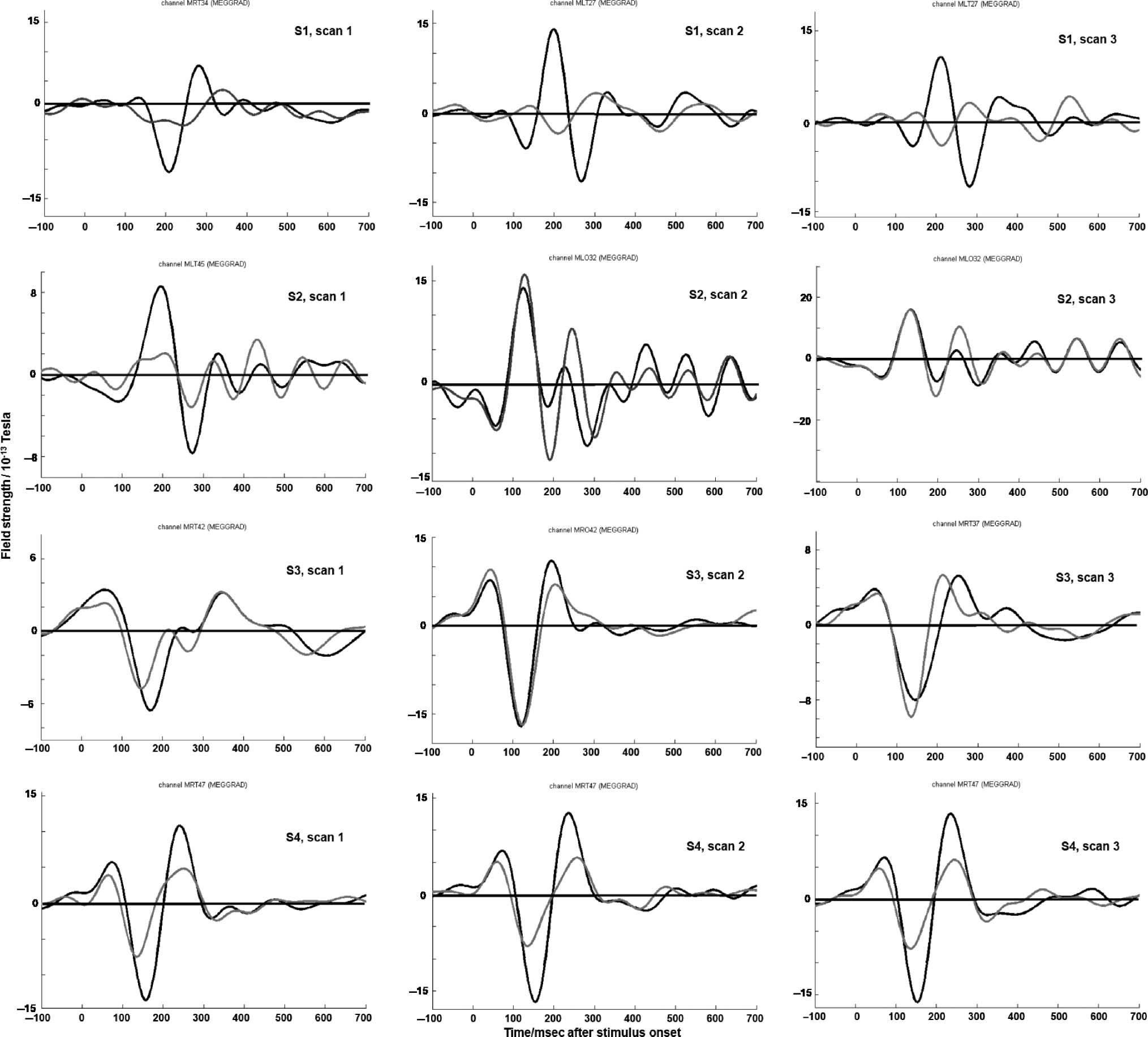
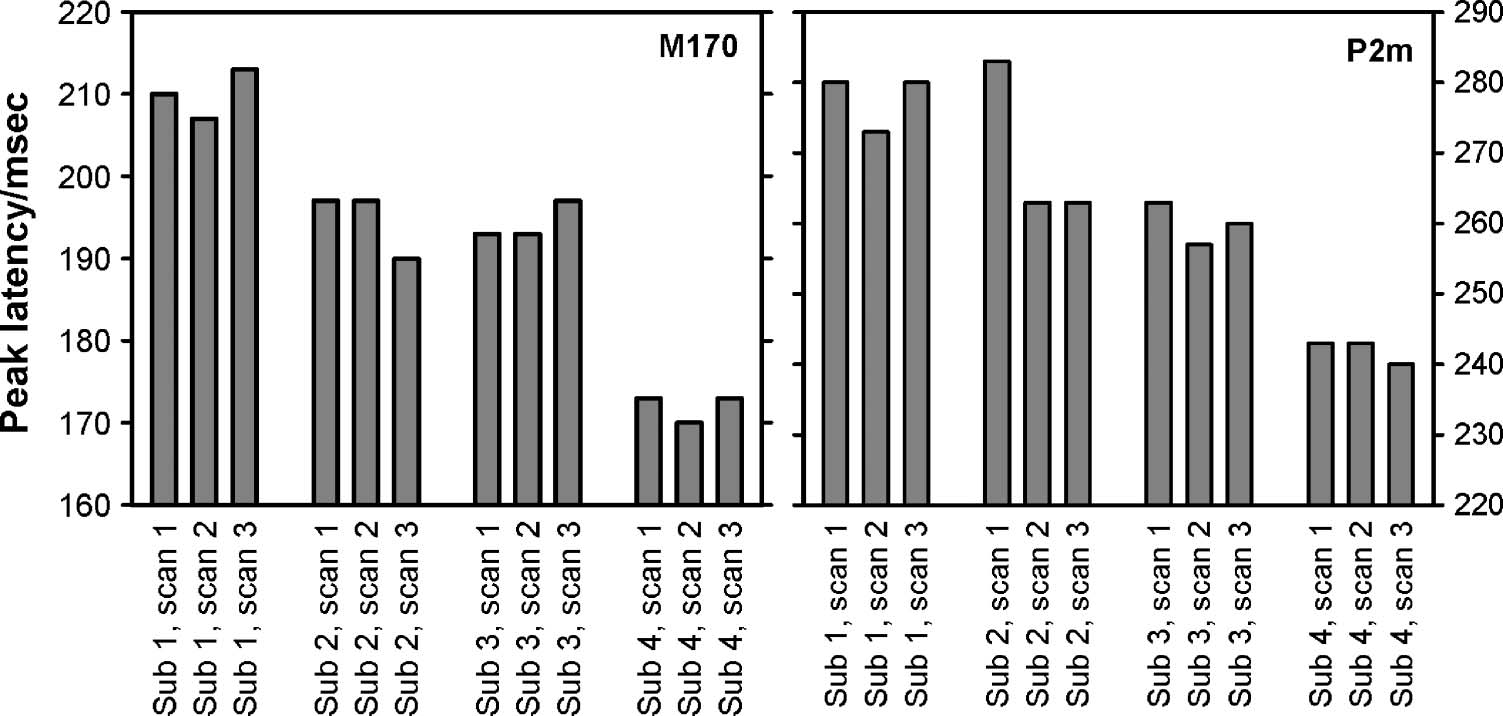
Variability in Latency
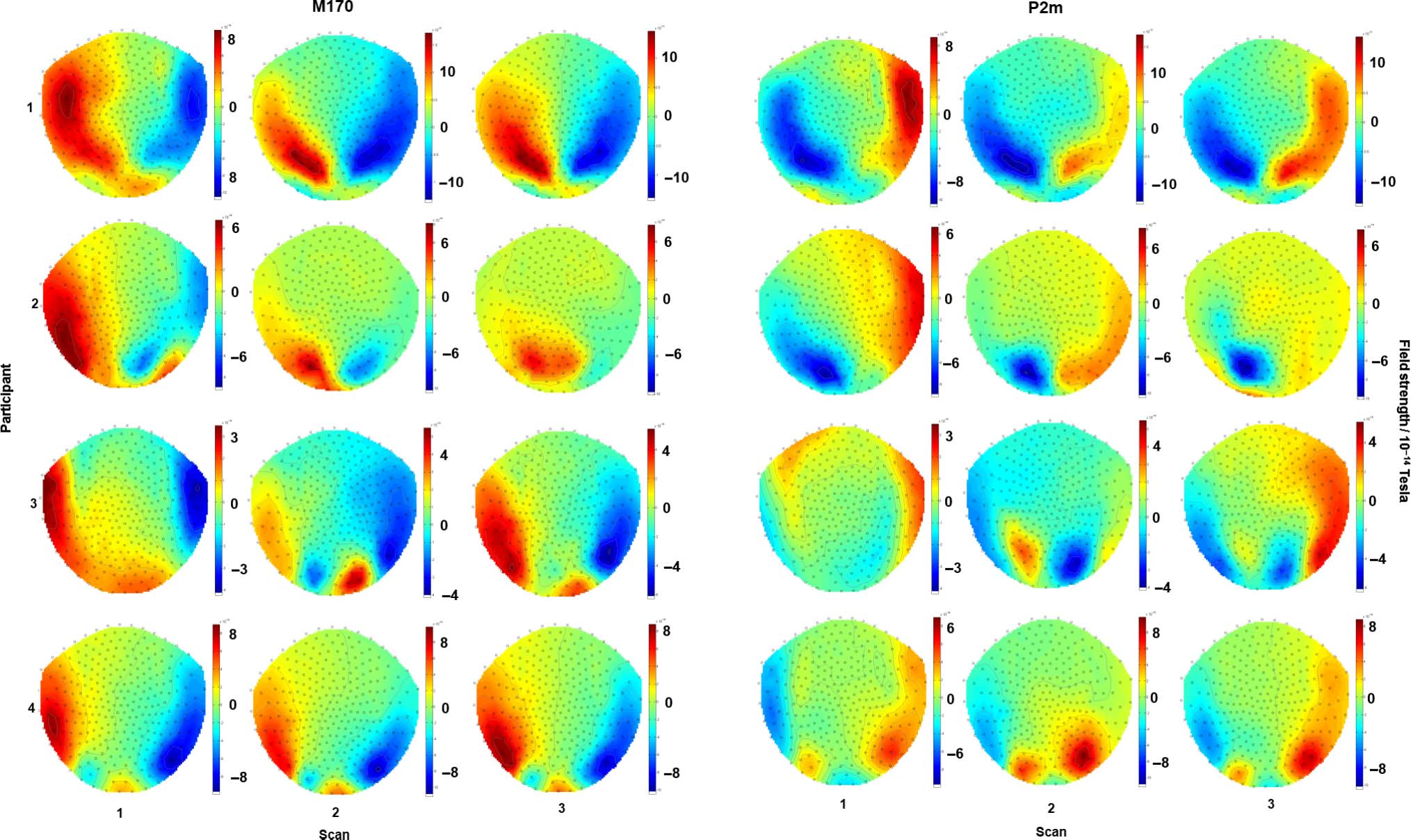
The latencies of the M170 and the P2m are plotted in
Figure 6 (the topographies around the peak are plotted
in Figure 2, and the percept-related ERFs are plotted in
Figure 3). Kruskal–Wallis tests revealed that the latency
of both components varied across participants [M170:
χ2(3) = 9.4, p = .025; P2m: χ2(3) = 8.9, p = .031],
whereas no such difference was found within participants
across time [M170: χ2(2) = 0.1, p = .97; P2m: χ2(2) =
1.0, p = .59].
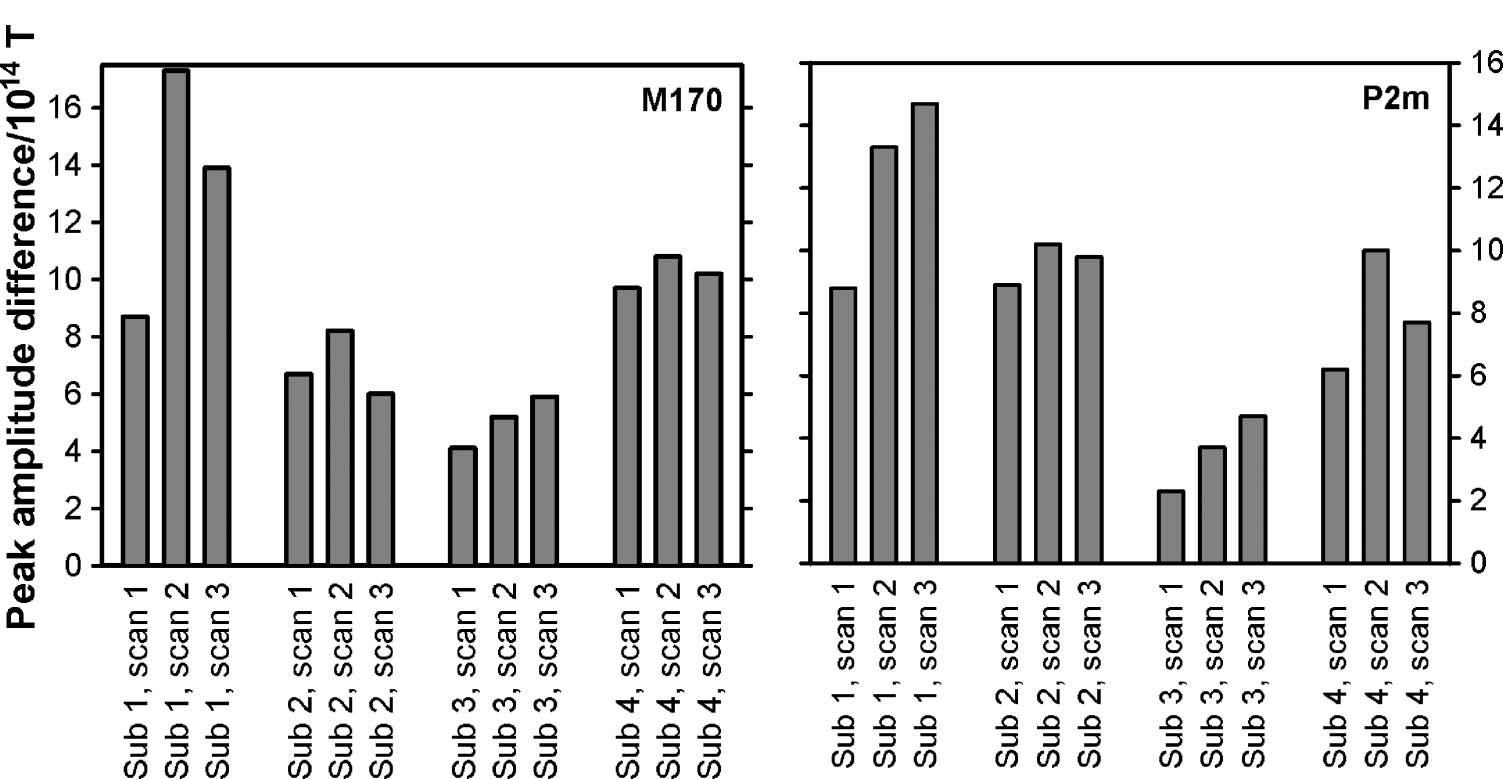
Variability in Amplitude
The maximum amplitude differences between face and
grating perception around the M170 and the P2m are
plotted in Figure 7. Kruskal–Wallis tests revealed that
the amplitude differences of both components varied
across participants [M170: χ2(3) = 9.5, p = .024; P2m:
χ2(3) = 7.8, p = .050], whereas no clear difference was
found within participants across time [M170: χ2(2) = 0.8,
p = .67; P2m: χ2(2) = 2.0, p = .37].
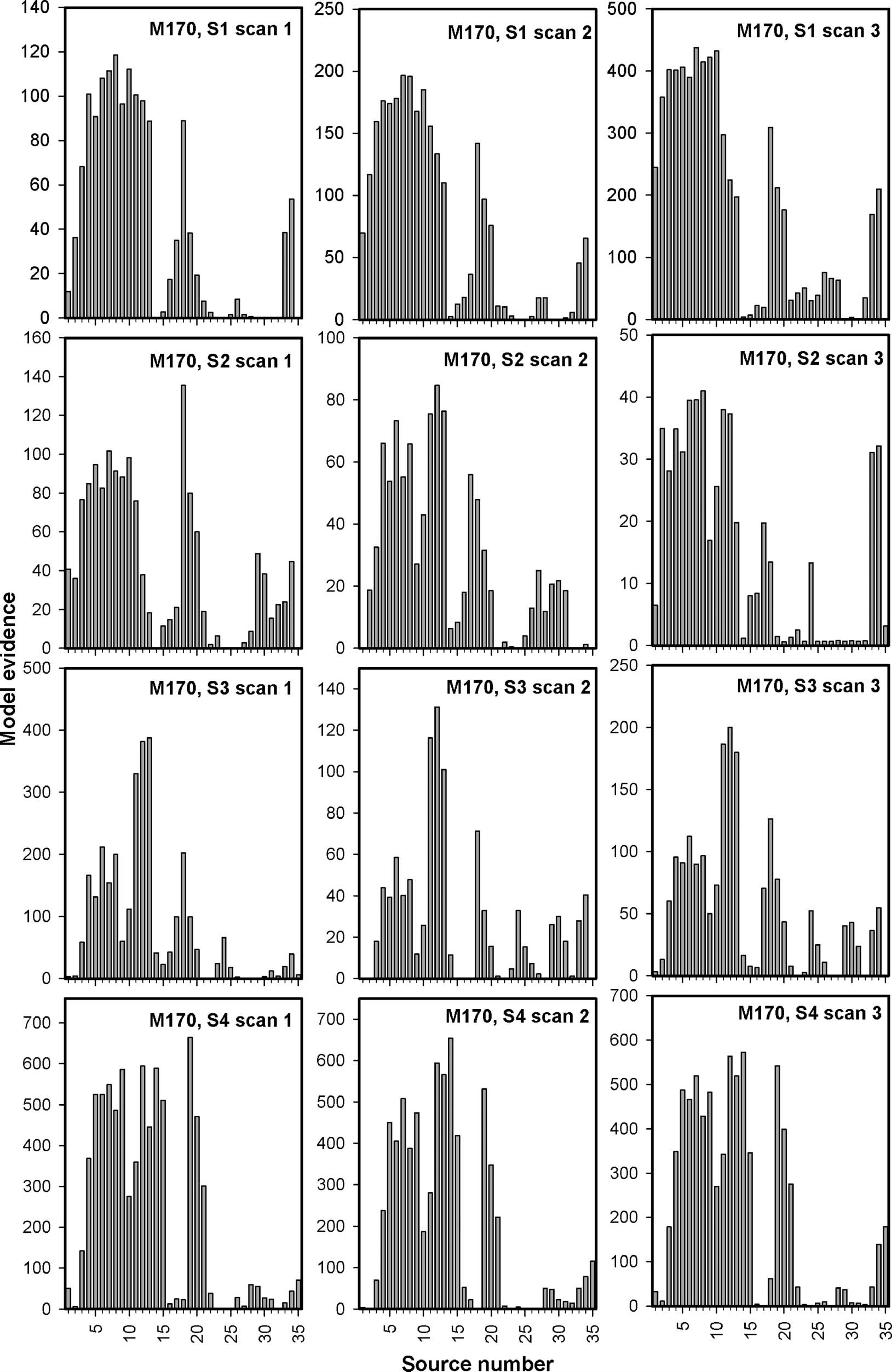
Variability of Sources
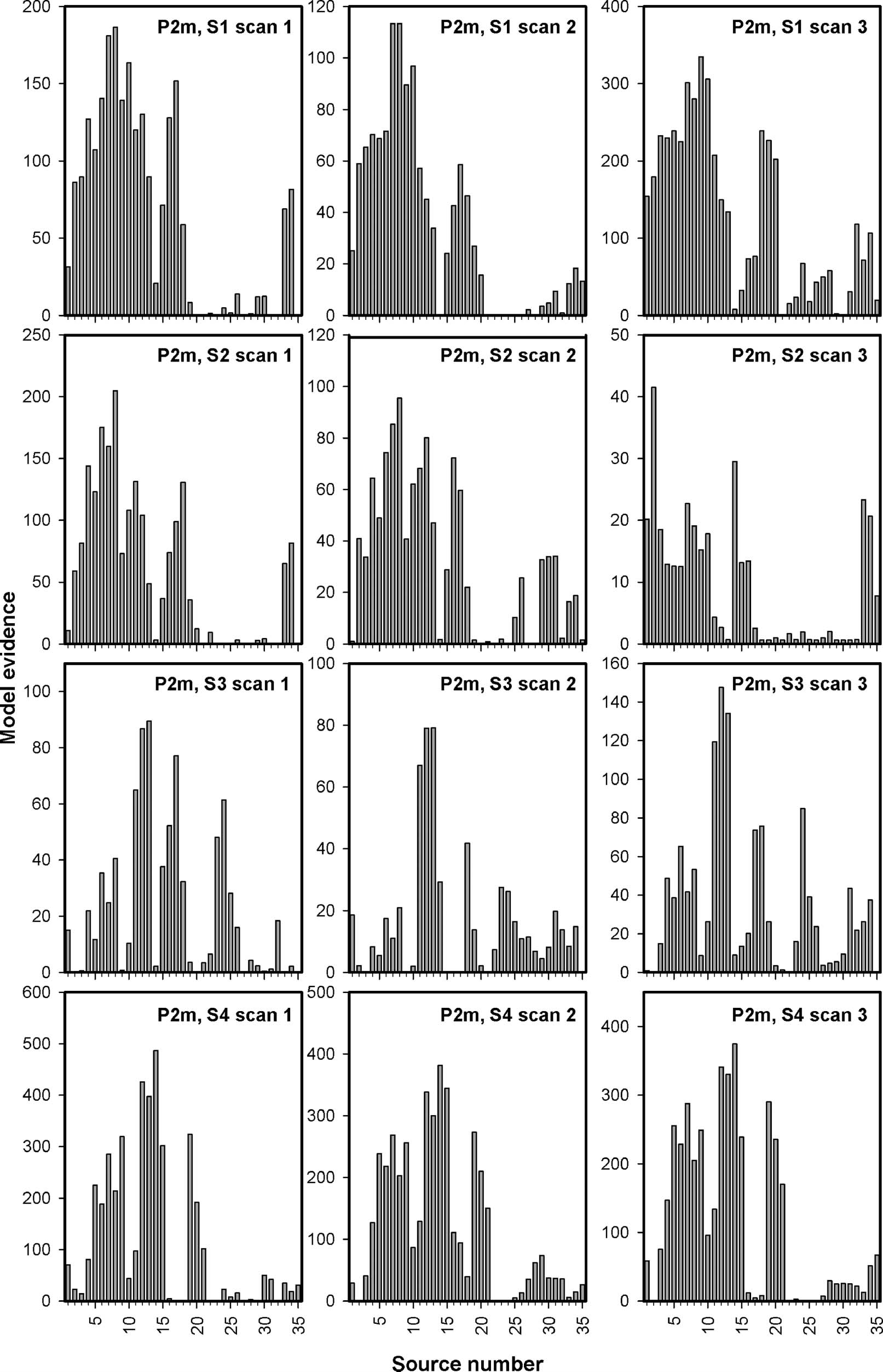
MVB models were made individually at the M170 and the
P2m for all 35 sources for all recordings. The model evi-
dence is plotted in Figure 8 for the M170 and in Figure 9
for the P2m. Conclusions should not be based on the
absolute values of the model evidence, but within a data
set, the model evidence provides an estimate of how
well the activity at a particular source predicts perception
Figure 6. Latency of M170
and P2m. The average peak
latencies for the M170 (left) and
the P2m (right) are plotted for
all data sets for all participants.
Notice the apparent variability
in latency across participants
and the apparent stability of
latency within participants.
848
Journal of Cognitive Neuroscience
Volume 26, Number 4
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
4
2
6
8
/
4
4
0
/
1
8
9
4
4
0
6
/
6
1
2
7
3
8
o
1
c
0
n
1
_
4
a
/
_
j
0
o
0
c
5
n
2
5
_
a
p
_
d
0
0
b
5
y
2
g
5
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
.
f
t
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 7. Amplitude
differences of the M170 and
P2m between face and grating
perception. The average peak
amplitude difference for the
M170 (left) and the P2m (right)
are plotted for all recording
sessions for all participants.
Notice that the amplitude
difference varies more between
participants than within
participants.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
4
2
6
8
/
4
4
0
/
1
8
9
4
4
0
6
/
6
1
2
7
3
8
o
1
c
0
n
1
_
4
a
/
_
j
0
o
0
c
5
n
2
5
_
a
p
_
d
0
0
b
5
y
2
g
5
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
/
.
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
compared with the other sources. For each data set, MVB
modeling was thus used to rank the importance of the
sources, and these rankings were compared within and
between participants using Kruskal–Wallis tests. Again,
large differences were found around both components
across participants [M170: χ2(3) = 14.0, p = .0029; P2m:
χ2(3) = 20.7, p = .0001], and no clear, significant dif-
ferences were found within participants across time [M170:
χ2(2) = 3.4, p = .18; P2m: χ2(2) = 2.3, p = .31].
DISCUSSION
Between-participant Differences but No
Across-time Differences
We found that conscious perception could be decoded
around the time window of the VAN (140–310 msec
after stimulus onset). Decoding accuracy decreased little
or not at all across years compared with across days
(<1%), implying that the VAN-related neural correlates
of conscious perception were very stable across time
within individuals. Follow-up analyses revealed that the
consciousness-specific modulation of the two main ERF
components of the experiment, the M170 and the P2m,
remained stable over time in terms of latency, amplitude,
and sources.
In contrast, decoding accuracy dropped drastically when
training and testing classifiers across individuals as was
previously reported in Sandberg et al. (2013). Note that
even with only four of the original eight participants,
statistical testing was consistently significant. In this study,
further analyses revealed significant differences in the con-
sciousness-specific modulation of the M170 and the P2m
in terms of latency, amplitude, and sources. Crucially, the
between-participant drop in decoding accuracy cannot
be caused by (a) technical artifacts related to classifying
across different recording sessions (as within-participant
decoding did not show this drop) and (b) artifacts of the
source activity reconstruction process (as differences were
found between individuals in sensor as well as source
space). It is also unlikely that the results are caused artifacts
related to (consistent) eye movements as these would
be localized to the inferior part of the frontal cortex and
previous analyses of Recording Session 1 (Sandberg et al.,
2013) demonstrated that peak classification accuracy can
be obtained when classification is based on extrastriate
visual cortex activity alone. Similarly, occipital and tem-
poral lobe sources were consistently found to be the most
predictive of conscious perception in that study (Sandberg
et al., 2013).
The results are also compatible with the previously men-
tioned study showing much higher intraindividual stability
of VEPs for simple visual stimuli across months compared
with interindividual stability (Sarnthein et al., 2009). We
note again, however, that here we did not simply dem-
onstrate that the ERPs were stable, but rather that the
consciousness-specific modulation of these components
(given equivalent physical stimulation) was comparable
within participants across years. In other words, the en-
dogenous suppression of the nonperceived image during
BR remains highly comparable across years in the normal,
healthy human brain.
At least two different interpretations appear valid for
the finding of interindividual differences in the neural cor-
relates of conscious contents, and the study thus opens
up the possibility for further studies into this domain.
The results could be taken to indicate that conscious con-
tents are represented differently across individuals (i.e., the
weighted contribution of cortical sources may differ be-
tween individuals having the same conscious experience),
or alternatively that different individuals have slightly dif-
ferent perceptual experiences of the same object (for in-
stance, it might be expected that individual differences
in the degree of endogenous suppression of irrelevant
information could lead to differences in perceived visual
clarity). These interpretations could be examined in further
Sandberg et al.
849
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
4
2
6
8
/
4
4
0
/
1
8
9
4
4
0
6
/
6
1
2
7
3
8
o
1
c
0
n
1
_
4
a
/
_
j
0
o
0
c
5
n
2
5
_
a
p
_
d
0
0
b
5
y
2
g
5
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
/
f
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 8. MVB model evidence across sources for the M170. MVB model evidence is plotted for the M170 for all sessions for all participants.
Notice the larger similarity in the pattern of source evidence across data sets from the same participant (horizontal direction) than between
participants (vertical direction).
850
Journal of Cognitive Neuroscience
Volume 26, Number 4
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
4
2
6
8
/
4
4
0
/
1
8
9
4
4
0
6
/
6
1
2
7
3
8
o
1
c
0
n
1
_
4
a
/
_
j
0
o
0
c
5
n
2
5
_
a
p
_
d
0
0
b
5
y
2
g
5
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
/
f
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 9. MVB model evidence across sources for the P2m. MVB model evidence is plotted for the P2m for all sessions for all participants. Notice
the larger similarity in the pattern of source evidence across data sets from the same participant (horizontal direction) than between participants
(vertical direction).
Sandberg et al.
851
studies by correlating individual differences in detailed
perceptual reports to interindividual decoding accuracy.
For both this study and previous studies of interindi-
vidual EEG/MEG differences, the anatomical structure of
the participants is a potential confound. In particular, indi-
vidual differences in anatomy (e.g., cortical folding) means
that the same cortical area may have different orientation
and distance from the sensors over subjects. It is thus
likely that at least part of the differences in the MEG signal
across individuals is because of this individual mapping
from sources to sensors. It is difficult to assess the impact
of this confound, and it is likely to have a greater effect on
the estimate of the source level amplitudes than latencies.
One way to partially address this potential issue, in future
studies, is to use combined MEG–EEG to sample from an
increased measurement subspace (Sharon, Hämäläinen,
Tootell, Halgren, & Belliveau, 2007).
Source Space versus Sensor Space
Parts of the results were based on decoding using classifi-
cation algorithms on source space data, and parts were
based on conventional statistics in sensor space. The com-
bination of both methods was used to obtain converging
evidence and reject some potential criticisms. The most
important reason for basing the analyses on source space
data was to avoid confounding the results by larger dif-
ferences in head position in relation to the sensors across
years than days. Indeed, our sensor space compared with
source space decoding revealed that this confound was
present. Within-participant topographical map differences
across years were thus entirely caused by the trivial finding
that participants positioned their head very similarly on
sessions close in time but not on sessions years apart. This
confound, however, could not impact upon the estima-
tion of the M170 and P2m component latencies, and for
this reason, this analysis was performed in sensor space
to include data that had not been modified by source space
reconstruction. A slight impact of head position might be
expected upon the difference in activity between the com-
ponent amplitude for face and grating perception at the
peak latencies as the signal-to-noise ratio is influenced by
the distance between the source and the sensor, which of
course varies with head position. To minimize the im-
pact of this confound, the difference was measured at
the sensor showing the largest difference, although this
might not be the same sensor for every data set.
Decoding versus Traditional ERF Measures
Decoding accuracy was used to obtain a meaningful ob-
jective measure of quantitative differences between data
sets—that is, a measure that would indicate whether the
information considered important to predict the conscious
experience of one individual at a given time is also the
information that predicts the conscious of that individual
at a different time or the conscious experience of a differ-
ent individual. By allowing the multivariate classifier to
select a weighted combination of sources, the choice of
relevant information was as close to optimal as possible
and could not be biased by subjective choices regarding
which sensors/sources and time points to compare.
Conclusion
Taken together, the experiment demonstrates that the
neural correlates of conscious perception generalize very
well across years within individuals, but not across indi-
viduals. The large stability of the latency, amplitude, and
the relative importance of a large number of sources
argue against spontaneous changes in how specific con-
scious contents are represented (on the general scale that
MEG examines). This study thus indicates that, once a
brain has found a way to process and consciously repre-
sent an object, it will continue to use this even across years
(presumably until some drastic event such as intensive
training or neural damage forces a change).
Acknowledgments
This work was supported by the Wellcome Trust (G. R., G. R. B.,
and K. S.) and the European Research Council (K. S. and M. O.).
Reprint requests should be sent to Dr. Kristian Sandberg, Cog-
nitive Neuroscience Research Unit, Aarhus University Hospital,
Noerrebrogade 44, Building 10G, 8000 Aarhus C, Denmark, or
via e-mail: krissand@rm.dk.
REFERENCES
Carter, O., & Cavanagh, P. (2007). Onset rivalry: Brief
presentation isolates an early independent phase of
perceptual competition. PloS One, 2, e343.
Dehaene, S., Changeux, J.-P., Naccache, L., Sackur, J.,
& Sergent, C. (2006). Conscious, preconscious, and
subliminal processing: A testable taxonomy. Trends
in Cognitive Sciences, 10, 204–211.
Dehaene, S., Kerszberg, M., & Changeux, J. P. (1998).
A neuronal model of a global workspace in effortful
cognitive tasks. Proceedings of the National Academy
of Sciences, U.S.A., 95, 14529–14534.
Fahrenfort, J. J., Scholte, H. S., & Lamme, V. A. F. (2007).
Masking disrupts reentrant processing in human visual
cortex. Journal of Cognitive Neuroscience, 19, 1488–1497.
Friston, K. J. (2012). Ten ironic rules for non-statistical
reviewers. Neuroimage, 61, 1300–1310.
Friston, K. J., Chu, C., Mourão-Miranda, J., Hulme, O.,
Rees, G., Penny, W., et al. (2008). Bayesian decoding
of brain images. Neuroimage, 39, 181–205.
Friston, K. J., Harrison, L., Daunizeau, J., Kiebel, S., Phillips, C.,
Trujillo-Barreto, N., et al. (2008). Multiple sparse priors for
the M/EEG inverse problem. Neuroimage, 39, 1104–1120.
Hall, R. A., Rappaport, M., Hopkins, H. K., & Griffin, R. B.
(1973). Peak identification in visual evoked potentials.
Psychophysiology, 10, 52–60.
Koivisto, M., & Revonsuo, A. (2003). An ERP study of change
detection, change blindness, and visual awareness.
Psychophysiology, 40, 423–429.
852
Journal of Cognitive Neuroscience
Volume 26, Number 4
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
4
2
6
8
/
4
4
0
/
1
8
9
4
4
0
6
/
6
1
2
7
3
8
o
1
c
0
n
1
_
4
a
/
_
j
0
o
0
c
5
n
2
5
_
a
p
_
d
0
0
b
5
y
2
g
5
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
/
f
.
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Koivisto, M., & Revonsuo, A. (2010). Event-related brain
potential correlates of visual awareness. Neuroscience
& Biobehavioral Reviews, 34, 922–934.
Kornmeier, J., & Bach, M. (2004). Early neural activity in
Necker-cube reversal: Evidence for low-level processing
of a Gestalt phenomenon. Psychophysiology, 41, 1–8.
Kornmeier, J., & Bach, M. (2005). The Necker cube−An
ambiguous figure disambiguated in early visual processing.
Vision Research, 45, 955–960.
Lamme, V. A. F. (2010). How neuroscience will change our
view on consciousness. Cognitive Neuroscience, 1, 204–220.
Leopold, D. A., Wilke, M., Maier, A., & Logothetis, N. K. (2002).
Stable perception of visually ambiguous patterns. Nature
Neuroscience, 5, 605–609.
Liu, Y., Paradis, A.-L., Yahia-Cherif, L., & Tallon-Baudry, C.
(2012). Activity in the lateral occipital cortex between 200
and 300 msec distinguishes between physically identical
seen and unseen stimuli. Frontiers in Human Neuroscience,
6, article 211.
Näpflin, M., Wildi, M., & Sarnthein, J. (2007). Test-retest
reliability of resting EEG spectra validates a statistical
signature of persons. Clinical Neurophysiology: Official
Journal of the International Federation of Clinical
Neurophysiology, 118, 2519–2524.
Näpflin, M., Wildi, M., & Sarnthein, J. (2008). Test-retest
reliability of EEG spectra during a working memory task.
Neuroimage, 43, 687–693.
Oken, B. S., Chiappa, K. H., & Gill, E. (1987). Normal
temporal variability of the P100. Electroencephalography
and Clinical Neurophysiology, 68, 153–156.
Sandberg, K., Bahrami, B., Kanai, R., Barnes, G. R.,
Overgaard, M., & Rees, G. (2013). Early visual responses
predict conscious face perception within and between
subjects during binocular rivalry. Journal of Cognitive
Neuroscience, 25, 969–985.
Sarnthein, J., Andersson, M., Zimmermann, M. B., & Zumsteg, D.
(2009). High test-retest reliability of checkerboard reversal
visual evoked potentials ( VEP) over 8 months. Clinical
Neurophysiology, 120, 1835–1840.
Schellberg, D., Gasser, T., & Köhler, W. (1987). The
intraindividual reproducibility of flash-evoked potentials
in a sample of children. International Journal of
Psychophysiology: Official Journal of the International
Organization of Psychophysiology, 5, 135–143.
Sergent, C., Baillet, S., & Dehaene, S. (2005). Timing of
the brain events underlying access to consciousness
during the attentional blink. Nature Neuroscience, 8,
1391–1400.
Sergent, C., & Dehaene, S. (2004). Neural processes underlying
conscious perception: Experimental findings and a global
neuronal workspace framework. Journal of Physiology,
Paris, 98, 374–384.
Sharon, D., Hämäläinen, M. S., Tootell, R. B. H., Halgren, E.,
& Belliveau, J. W. (2007). The advantage of combining MEG
and EEG: Comparison to fMRI in focally stimulated visual
cortex. Neuroimage, 36, 1225–1235.
Van Loon, A. M., Scholte, H. S., van Gaal, S., van der Hoort,
B. J. J., & Lamme, V. A. F. (2012). GABA(A) agonist reduces
visual awareness: A masking-EEG experiment. Journal of
Cognitive Neuroscience, 24, 965–974.
Vanni, S., Revonsuo, A., Saarinen, J., & Hari, R. (1996).
Visual awareness of objects correlates with activity of
right occipital cortex. NeuroReport, 8, 183–186.
Weinberg, A., & Hajcak, G. (2011). Longer term test-retest
reliability of error-related brain activity: Long-term reliability
of the ERN. Psychophysiology, 48, 1420–1425.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
4
2
6
8
/
4
4
0
/
1
8
9
4
4
0
6
/
6
1
2
7
3
8
o
1
c
0
n
1
_
4
a
/
_
j
0
o
0
c
5
n
2
5
_
a
p
_
d
0
0
b
5
y
2
g
5
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
f
/
.
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Sandberg et al.
853