Linking the Brain with Behavior: The Neural Dynamics
of Success and Failure in Goal-directed Behavior
Amanda K. Robinson1,2
, Anina N. Rich1, and Alexandra Woolgar1,3
Abstract
■ The human brain is extremely flexible and capable of rapidly
selecting relevant information in accordance with task goals.
Regions of frontoparietal cortex flexibly represent relevant task
information such as task rules and stimulus features when par-
ticipants perform tasks successfully, but less is known about
how information processing breaks down when participants
make mistakes. This is important for understanding whether
and when information coding recorded with neuroimaging is
directly meaningful for behavior. Here, we used magnetoen-
cephalography to assess the temporal dynamics of information
processing and linked neural responses with goal-directed
behavior by analyzing how they changed on behavioral error.
Participants performed a difficult stimulus–response task using
two stimulus–response mapping rules. We used time-resolved
multivariate pattern analysis to characterize the progression of
information coding from perceptual information about the
stimulus, cue and rule coding, and finally, motor response.
Response-aligned analyses revealed a ramping up of perceptual
information before a correct response, suggestive of internal
evidence accumulation. Strikingly, when participants made a
stimulus-related error, and not when they made other types
of errors, patterns of activity initially reflected the stimulus
presented, but later reversed, and accumulated toward a repre-
sentation of the “incorrect” stimulus. This suggests that the
patterns recorded at later time points reflect an internally gen-
erated stimulus representation that was used to make the
(incorrect) decision. These results illustrate the orderly and
overlapping temporal dynamics of information coding in per-
ceptual decision-making and show a clear link between neural
patterns in the late stages of processing and behavior. ■
INTRODUCTION
A primary function of the human brain is to flexibly
respond to relevant perceptual information in accordance
with current context and task goals. The sound of a phone
ringing, for example, should prompt a different response if
it is your phone than if it belongs to someone else. This set
of complex processes, termed cognitive control, involves
interpreting incoming information given the current con-
text to determine an appropriate action (Posner & Presti,
1987; Posner & Snyder, 1975). Cognitive control involves
dynamic information exchange at different levels of pro-
cessing, from perceptual information processing to
decision-making and response selection. Understanding
how these different processes unfold could provide a great
deal of insight into how the brain achieves goal-directed
behavior.
A large body of neuroimaging research implicates fron-
toparietal brain regions in goal-directed behavior, which
form a distributed network responsible for cognitive
control (Duncan, 2010). This multiple-demand (MD)
network (Duncan, 2010), elsewhere referred to as the
cognitive control network (Cole & Schneider, 2007),
frontoparietal control system ( Vincent, Kahn, Snyder,
Raichle, & Buckner, 2008), or task-positive network (Fox
1Macquarie University, 2The University of Sydney, 3University of
Cambridge
et al., 2005), appears to flexibly represent different types
of information depending on task context. For example,
activity in these regions encodes task rules (e.g., Crittenden,
Mitchell, & Duncan, 2016; Woolgar, Afshar, Williams &
Rich, 2015; Woolgar, Hampshire, Thompson, & Duncan,
2011) and auditory, visual, and tactile stimulus features
(Long & Kuhl, 2018; Bracci, Daniels, & Op de Beeck, 2017;
Jackson, Rich, Williams, & Woolgar, 2017; Woolgar & Zopf,
2017; for a review, see Woolgar, Jackson, & Duncan, 2016).
These regions particularly encode task elements that are
demanding (Woolgar, Afshar, et al., 2015; Woolgar et al.,
2011) or at the focus of attention (Jackson & Woolgar, 2018;
Jackson et al., 2017; Woolgar, Williams, et al., 2015). Activity in
some of these regions has also been causally implicated in
selectively facilitating coding of task-relevant information
(Jackson, Feredoes, Rich, Lindner, & Woolgar, 2021). This
lends support to the possibility that flexible responses within
the MD regions play a causal role in goal-directed behavior
(e.g., Duncan, Assem, & Shashidhara, 2020; Woolgar,
Dermody, Afshar, Williams, & Rich, 2019; Woolgar, Duncan,
Manes, & Fedorenko, 2018; Woolgar et al., 2010).
A characteristic feature of cognitive control is that it
dynamically changes in response to task-relevant informa-
tion. Research using fMRI has yielded insight into the
brain networks involved in goal-directed behavior, but
the slow nature of the blood-oxygen-level response has
limited the exploration of the corresponding dynamics.
© 2022 Massachusetts Institute of Technology. Published under
a Creative Commons Attribution 4.0 International (CC BY 4.0) license.
Journal of Cognitive Neuroscience 34:4, pp. 639–654
https://doi.org/10.1162/jocn_a_01818
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
4
4
6
3
9
1
9
9
6
6
7
8
/
j
o
c
n
_
a
_
0
1
8
1
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Time-resolved neuroimaging methods such as magneto-
encephalography (MEG) and EEG have been more fruitful
in understanding how cognitive control unfolds over time
(for a review, see Gratton, Cooper, Fabiani, Carter, &
Karayanidis, 2017). For example, conflict-related process-
ing involving incongruent task features elicits a larger
evoked response than a congruent condition approxi-
mately 200–400 msec after stimulus onset, which has been
linked to activity within the anterior cingulate (Folstein &
Van Petten, 2008), and task-switching involves a larger
parietal positivity around 300 msec relative to task repeats
(Karayanidis et al., 2010). The newer method of multivar-
iate pattern analysis (MVPA) in conjunction with MEG/EEG
allows further insight into processing dynamics underly-
ing cognitive control. MVPA uses pattern classification
approaches applied to neuroimaging data to show what
information is being coded within the brain (Haxby,
2012; Haxby et al., 2001). Time-resolved MVPA has been
used to characterize how information coding changes over
time (Hebart & Baker, 2018; Carlson, Hogendoorn, Kanai,
Mesik, & Turret, 2011). For example, Hebart, Bankson,
Harel, Baker, and Cichy (2018) had participants perform
different tasks on visual object stimuli while measuring
MEG and showed that task-relevant object features were
enhanced at late stages of processing, more than 500 msec
after the stimulus was presented. Other work has shown
clear progression of task-relevant information during com-
plex tasks, with different dynamics for features such as
stimulus, task, and response (Kikumoto & Mayr, 2020;
Hubbard, Kikumoto, & Mayr, 2019; Wen, Duncan, &
Mitchell, 2019). This line of research has also highlighted
the importance of combining relevant task information for
successful behavior (Kikumoto & Mayr, 2020). These
MVPA studies provided great insight into the neural
dynamics of goal-directed behavior, but all used designs
where the task cue was presented before the target, allow-
ing participants to prepare for the task in advance. In addi-
tion, these studies focused on stimulus-aligned neural
responses. It seems likely that tracking the dynamic cod-
ing of relevant task features relative to both stimulus onset
and response, using a task that induces more flexible
behavior, might elucidate stronger links between dynamic
neural responses and goal-directed behavior.
Decades of neuroimaging research have focused on the
neural correlates of behavior, but assessing whether
particular patterns of brain activity are necessary for
behavior has presented a challenge. In MVPA, a classifier
algorithm is trained to distinguish between conditions
using patterns of neural data from multiple trials of each
condition. If a classifier can predict the conditions of
new neural data better than chance, this demonstrates that
the patterns of activity in the data must contain, or repre-
sent, information about the different conditions. How-
ever, the conclusion that decodable patterns represent
information has been questioned on theoretical grounds
(de Wit, Alexander, Ekroll, & Wagemans, 2016): Just
because information is decodable using machine learning
does not necessarily mean it is used by the brain to gener-
ate behavior. This awareness has led researchers to push
for more explicit links between MVPA patterns and behav-
ior, for example, comparing details of patterns with RTs or
accumulation rates in models of behavior (Grootswagers,
Cichy, & Carlson, 2018; Ritchie & Carlson, 2016; Ritchie,
Tovar, & Carlson, 2015).
Exploring how information coding changes when par-
ticipants make errors is another way to establish how
behaviorally meaningful patterns of activation are. For
example, Williams, Dang, and Kanwisher (2007) demon-
strated that multivariate fMRI patterns in lateral occipital
cortex, but not those in early visual regions, reduced to
chance when participants made errors on a shape discrim-
ination task, indicating that patterns in early visual cortex
were not directly read out in behavior. In another study,
participants performed a scene classification task
( Walther, Caddigan, Fei-Fei, & Beck, 2009), and classifier
prediction error patterns correlated with the types of
errors in behavior within high-level object and scene-
specific brain regions, but not within early visual cortex.
Using MEG, we have recently shown that this logic can
even be used to predict behavioral errors before they
occur (Karimi-Rouzbahani, Woolgar, & Rich, 2021).
A stronger requirement for a behaviorally meaningful
pattern of activity is that it should not only change on error
but also change to something that predicts the particular
error to be made (Woolgar et al., 2019). We tested this in
fMRI and found that patterns of activation in frontoparie-
tal cortex indeed reversed on error, such that patterns of
activation on error trials represented information that was
not presented to the participant, in a manner that was
diagnostic of the particular behavioral error they made
( Woolgar et al., 2019). In that study, participants per-
formed a difficult response-mapping task. When partici-
pants made a rule error, MD patterns of activity reflected
the incorrect rule, and when participants made other
errors, MD patterns of activity reflected the incorrect
stimulus ( Woolgar et al., 2019). Within visual cortex, in
contrast, there was no evidence of relevant information
(correct or incorrect) during errors. Thus, some multivar-
iate patterns appear to be more directly relatable to
behavior that others, and there is a tight link between
frontoparietal activity patterns and behavioral outcome.
In the current study, we used MEG and MVPA to exam-
ine the dynamics of this effect, asking whether information
coding through the course of a trial was equally associated
with behavior. We aimed to (1) characterize the neural
dynamics of multiple types of task-relevant information
and (2) examine their relationship to behavior over time.
Participants performed a difficult response-mapping task
that required different responses to a target stimulus
depending on the current rule. To determine what aspects
of this representation could be directly linked to behavior,
we examined information coding on incorrect trials:
when the wrong rule was applied or when there were
errors in perception. We found a clear progression in
640
Journal of Cognitive Neuroscience
Volume 34, Number 4
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
4
4
6
3
9
1
9
9
6
6
7
8
/
j
o
c
n
_
a
_
0
1
8
1
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
onset of information coding, such that stimulus features
are evident shortly after stimulus onset, followed by
abstract rule coding and then the response, with the
information about each task feature accumulating up to
the time of response. When participants made stimulus
errors, stimulus information was initially coded veridically
but later accumulated in the opposite direction, toward a
representation of the incorrect stimulus. By contrast,
stimulus information was encoded correctly when partici-
pants made rule errors. The data reveal the dynamics with
which information coding in the brain can be tightly
linked to participant behavior.
METHODS
All code and materials are available on the Open Science
Project at https://osf.io/2nwhr/.
Participants
Participants were 22 healthy adults (14 women, eight
men; age range: 18–38 years) with normal or corrected-
to-normal vision recruited from Macquarie University.
This study was approved by the Macquarie University
ethics committee, and informed consent was obtained
from all participants. Participants took part in two ses-
sions: a 1-hr behavioral session and a 2-hr MEG session,
on separate days. They were compensated $15 for the behavioral session and $40 for the MEG session. For
two participants, initial photodiode inaccuracies meant
that the timing for two and five trials, respectively, was
not adequately marked, so these trials were excluded
from analyses. Data from an additional two participants
(two men) were collected and excluded: Both participants
had very few stimulus position errors during the MEG ses-
sion (<10), and for one of the participants, there was a
recording error such that MEG data were recorded for
only 680 of 800 trials.
Design and Procedure
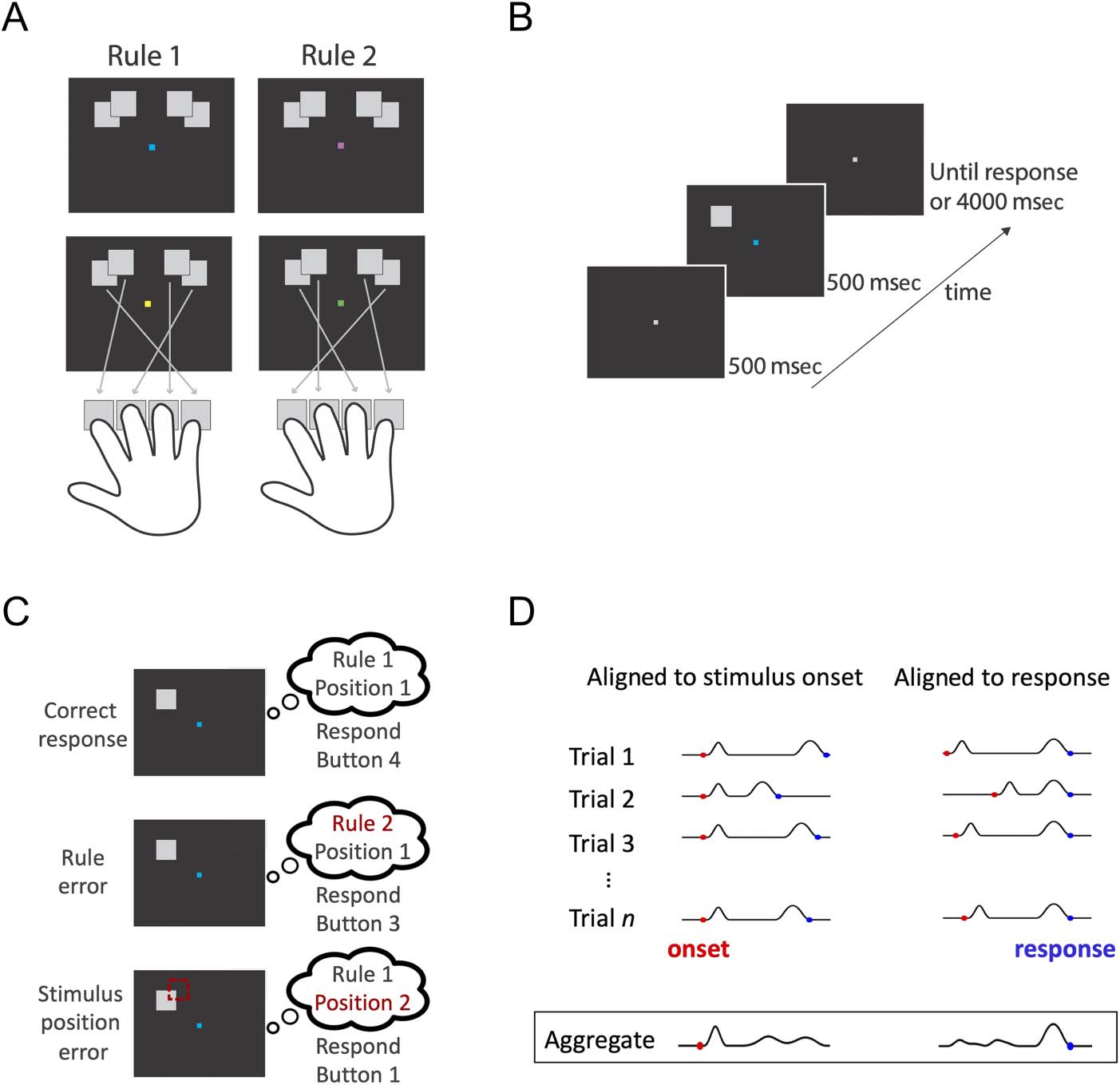
Participants learned to apply two difficult response-
mapping rules regarding the position of a target stimulus.
The target was a gray square approximately 2° × 2° of
visual angle that appeared in one of four positions. All
positions were equidistant from fixation at an eccentricity
of 4° of visual angle. Within the left and right visual fields,
the two possible target locations overlapped by 60% hori-
zontally and 65% vertically to create a high degree of posi-
tion uncertainty (Figure 1A). Participants had to respond
to the position of the stimulus using two possible
response-mapping rules (Figure 1A). The two rules each
comprised four unique position transformations and were
mirror images of each other. The color of a central fixation
square acted as a cue for the rule. There were two cues per
rule, to dissociate neural responses to cues from the
neural responses to rules (e.g., blue and yellow = Rule
1, pink and green = Rule 2; counterbalanced across partic-
ipants). Participants responded by pressing one of four
response keys with their right hand. The stimuli were pre-
sented using PsychToolbox in MATLAB (The MathWorks).
The stimulus–response rule mappings were designed to
distinguish correct responses and specific types of errors
(Figure 1C). An error was considered a “rule error” when
the button press response reflected the combination of
the correct stimulus position with the wrong rule. In con-
trast, a “stimulus error” was defined as a button press
response consistent with the combination of the adjacent
perceptually confusable position with the correct rule. For
example, under Rule 1, if the stimulus appeared on the far
left, the correct response would be Button 4, a rule error
(i.e., using Rule 2 applied to the correct position) would
lead to a Button 3 response, a stimulus error (i.e., using
Rule 1 correctly but confusing the stimulus with the
other left position) would lead to a Button 1 response,
and confusing both the rule and the stimulus led to a
Button 2 response.
Training Session
Participants learned each rule in a separate session outside
the MEG. They were trained to perform the task using
increasingly difficult blocks of trials (see below). Feedback
was given on every trial. For every incorrect response, par-
ticipants were shown the correct response.
Initially, stimuli were presented in nonoverlapping
positions (i.e., further apart than the final paradigm) so
there was no position uncertainty. Stimuli were pre-
sented on the screen until a response was made (i.e.,
not time limited). In the first block, participants learned
the first rule (Rule 1 or Rule 2, counterbalanced across
participants). Each stimulus position was shown with
its associated response four times (16 trials), and partic-
ipants had to press the appropriate button for each stim-
ulus. In each trial, cue color was chosen randomly from
the two possible cues for that rule. The second block
followed the same protocol, but for the other rule (Rule
1/Rule 2). In the third block, participants had to perform
the task by implementing both rules, but still with well-
separated stimuli. In the fourth block, the stimuli were
presented in their final, overlapping experimental posi-
tions. Finally, in the fifth block, the stimuli were presented
with the same procedure as the final experimental para-
digm: The stimuli were overlapping and were presented
for only 500 msec. Blocks 3–5 contained 32 trials each,
consisting of two repeats of each cue and stimulus posi-
tion, randomly ordered. In all blocks, participants had to
perform at 60% accuracy or above to progress to the next
block type. Blocks were repeated if they did not reach this
threshold. On average, participants completed 8.61 train-
ing blocks (SD = 2.46). Block 3 was most often repeated
(M = 3.09 repeats).
Robinson, Rich, and Woolgar
641
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
4
6
3
9
1
9
9
6
6
7
8
/
j
o
c
n
_
a
_
0
1
8
1
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 1. Experimental design.
(A) Response mapping rules.
Participants had to indicate the
position of a target stimulus that
appeared in one of four
possible locations. There were
two cues per rule, designated by
blue/yellow and green/pink
squares at fixation. The button
press associated with each
position is indicated by the
specific rule. (B) Trial timeline.
After a fixation screen, the
target stimulus and colored
fixation cue appeared
simultaneously, and participants
had to apply the correct
response-mapping rule using a
button press. (C) Behavioral
response types. In this example,
the stimulus was in Position 1
and the rule was Rule 1 (blue
cue), so the correct response
was Button 4. A rule error
occurred if the rule was
mistaken for Rule 2, leading
to a response of Button 3.
A stimulus position error
occurred if the position was
mistaken to be Position 2,
leading to a response of
Button 1. (D) Depiction of MEG
data collation: data aligned to
stimulus onset (left) or RT
(right). The temporal dynamics
of stimulus-related and decision-related neural responses vary across trials, with different processes aligned with onset and response. Aligning
the MEG data to stimulus onset versus response highlights different neural stages on the aggregate of all trials, although the content of each
trial is identical.
Experimental Session
In the second session, participants performed the task
while their neural activity was recorded using MEG. At
the start of each block, participants were shown a graphi-
cal depiction of the rules for at least 2 sec. When they were
ready, they pressed a button to begin the block. In each
trial, participants were shown a gray fixation marker for
500 msec, and then, the square target stimulus and col-
ored rule cue were presented for 500 msec (Figure 1B).
The participants were instructed to respond as quickly as
possible without sacrificing accuracy. After they
responded, there was an intertrial interval of 1000 msec
before the next trial started. There were 10 blocks of 80
trials, each containing five trials per stimulus and cue color
combination. Within each block, the order of the trials was
randomized. Instead of feedback on every trial, like in
training, participants were given feedback about their
mean accuracy and RTs at the end of each block.
system containing 160 gradiometers (Kado et al., 1999).
The participant’s head was fitted with a cap containing five
marker coils. The head shape and position of the marker
coils was marked using a Polhemus digitization system.
Once inside the MEG, the position of the marker coils
was measured to ensure the MEG sensors had good
coverage over the participant’s head. Marker position
measurements were repeated halfway through the exper-
iment and at the end of the session. Raw MEG data were
collected at 1000 Hz with online 0.03-Hz high-pass and
200-Hz low-pass filters.
Stimuli were projected onto the ceiling of the magneti-
cally shielded room. Stimulus timing was measured using a
photodiode placed on the projection mirror and marked
in an additional channel in the MEG recording. Partici-
pants indicated their response using a four-button fiber
optic response pad (Current Designs). Response tim-
ing was marked in the MEG recording using a parallel
port trigger.
MEG Acquisition
MEG Data Analysis
MEG data were collected at Macquarie University in the
KIT-MQ MEG facility with a whole-head supine Yokogawa
MEG data were analyzed using multivariate decoding,
which is very sensitive to reliable effects in the data and
642
Journal of Cognitive Neuroscience
Volume 34, Number 4
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
4
6
3
9
1
9
9
6
6
7
8
/
j
o
c
n
_
a
_
0
1
8
1
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
resistant to artifacts such as eye blinks that are not consis-
tent across time and condition (Carlson, Grootswagers, &
Robinson, 2020; Grootswagers, Wardle, & Carlson, 2017).
Because of the robustness of decoding to such artifacts,
data were minimally preprocessed using EEGLAB
(Delorme & Makeig, 2004). Data were filtered using a
Hamming window finite impulse response filter (default
EEGLAB filter pop_eegfiltnew) with high pass of 0.1 Hz
and low pass of 100 Hz and then downsampled to
200 Hz before epoching. For separate analyses, trials were
epoched relative to stimulus onset, marked by the photo-
diode, and to the button press response, marked by the
parallel port trigger.
Data were analyzed using time-resolved classification
methods (e.g., Carlson et al., 2020; Robinson, Grootswagers,
& Carlson, 2019; Grootswagers et al., 2017) and imple-
mented using the CoSMoMVPA toolbox (Oosterhof,
Connolly, & Haxby, 2016). For each time point, data were
pooled across all 160 MEG sensors, and we tested the ability
of a linear discriminant analysis classifier to discriminate
between the patterns of neural responses associated with
the different conditions. Trials were divided according to
their associated behavioral responses: correct trials, rule
errors, and stimulus position errors (Figure 1C). The classi-
fiers were always trained on correct trials. To ensure that
there were equal numbers of trials for each condition,
correct trials were subsampled to be equal for each position
and rule combination for each block per participant. To
ensure adequate trial numbers for each of the analyses,
blocks with fewer than two trials per Rule × Position
combination were excluded; this amounted to nine
excluded blocks in total across eight participants, with the
remaining 14 participants having all blocks included. The
total number of selected trials per participant was M =
437.09 (minimum = 280, maximum = 600).
Temporal Dynamics of Stimulus, Cue, Rule, and
Response Coding
We performed pattern classification analyses to determine
the time points at which stimulus position, cue, rule, and
response representations emerge in the brain. First, we
decoded stimulus position by comparing neural represen-
tations of the inner two stimulus positions (Positions 2
and 3) with those of the outer two stimulus positions
(Positions 1 and 4). Separating position in this manner
meant that motor responses and rule were balanced
across the two position conditions and could not drive
the classification results, ensuring we are detecting infor-
mation related to stimulus position.
Next, we assessed the time course of rule coding by
training a classifier to distinguish between Rule 1 and
Rule 2. In having two color cues per rule, this analysis
focused on rule coding over and above the physical prop-
erties of the cues (Rule 1 [blue and yellow cues] vs. Rule
2 [pink and green cues]).
We can also, however, decode cue coding separately
from rule coding. To assess how cue decoding differed
from rule decoding, we decoded between the two cues
per rule (i.e., blue vs. yellow color cue for Rule 1 and pink
vs. green for Rule 2). Cue coding was quantified as the
mean of the two pairwise analyses.
As a final analysis, we decoded motor response by
comparing the inner two button presses with the outer
two button presses. This comparison ensured that stim-
ulus position and rule were balanced within each class,
so that the classifier would be driven by the motor
response alone.
For each decoding analysis, classification analyses were
performed using a leave-one-block-out cross-validation
approach. This resulted in 10-fold cross-validation for
participants with no excluded blocks (n = 14). The
remaining participants used ninefold (n = 7) and eight-
fold cross-validation (n = 1). For all decoding analyses,
chance performance was 50%.
Error Representations
The next set of analyses focused on decoding neural
activity when participants made errors, to explore the
relationship between patterns of activity and behavior. To
investigate the representation of rule and stimulus errors,
we trained the classifier on the correct trials and tested on
the error trials. This allowed us to decode what information
was present in the patterns of response across sensors
when participants made different kinds of mistakes.
Specifically, the analyses assessed whether the error pat-
terns resembled the “correct” stimulus and rule patterns
(above-chance decoding) or the neural patterns associated
with the “incorrect” stimulus and rule (below-chance
decoding). Note that, in this approach, below-chance clas-
sification is meaningful: It indicates the representation of
the pattern that is instantiated when the other (incorrect
in this case) rule or stimulus position is encoded.
We performed error decoding for stimulus position and
rule information. In a comparable procedure to the cor-
rect trial analysis, we used leave-one-block-out cross-
decoding analyses. In each fold, the classifier was trained
on “correct” trials from all but one block and tested on all
“error” trials across the whole session. This ensured the
same training data (and thus decoding models) were
used as in the correct trial analyses but allowed well-
characterized results for the relatively small number of
error trials. Participants made an average of 5.71% rule
errors and 9.82% stimulus position errors (Figure 2A).
Table 1 shows the mean number of trials used for stimulus
and rule decoding per trial response type (correct, stimu-
lus error, rule error).
Exploratory Searchlight Analysis
To illustrate the spatial extent of the MEG signal containing
relevant task-related information, we applied searchlight
Robinson, Rich, and Woolgar
643
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
4
6
3
9
1
9
9
6
6
7
8
/
j
o
c
n
_
a
_
0
1
8
1
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
4
6
3
9
1
9
9
6
6
7
8
/
j
o
c
n
_
a
_
0
1
8
1
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
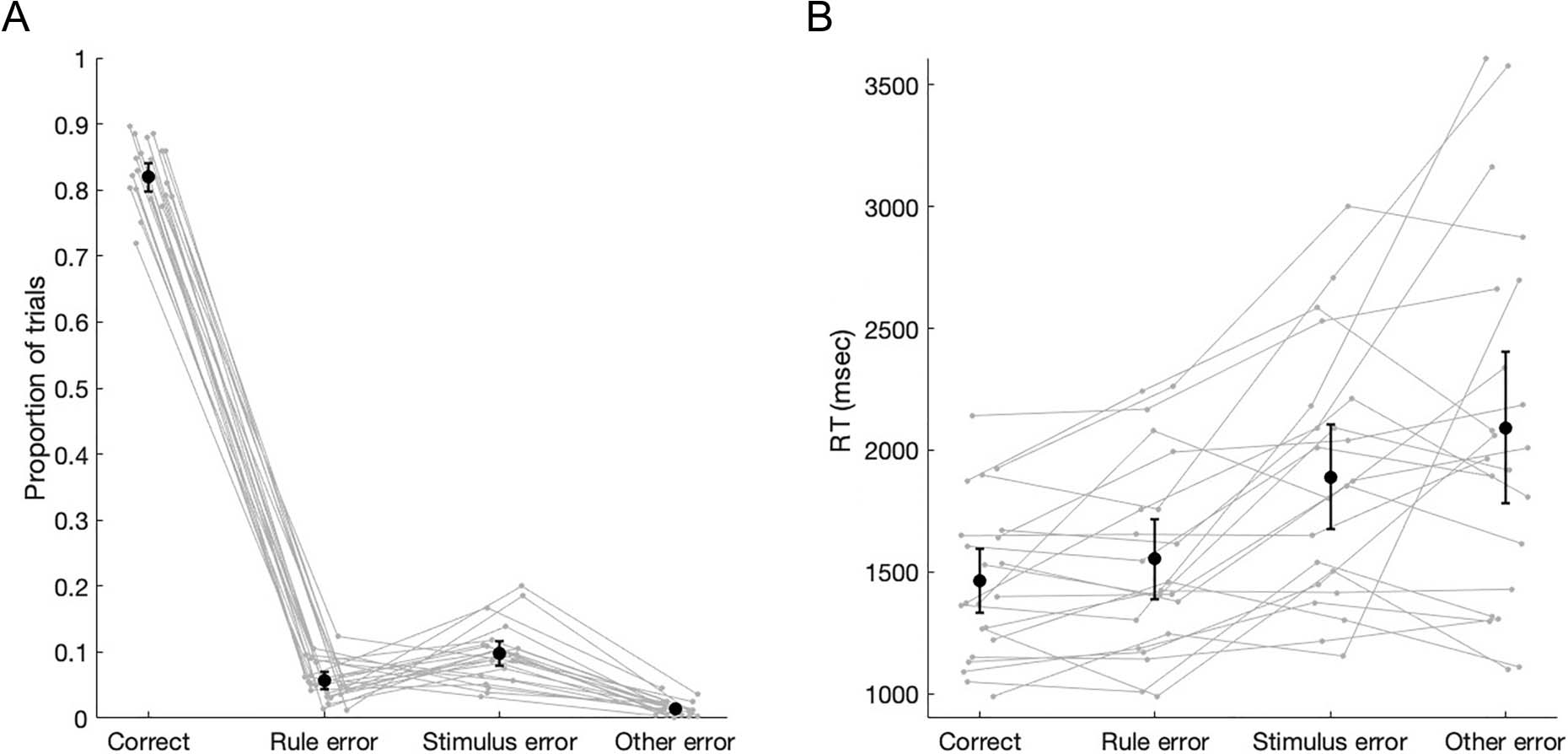
Figure 2. Behavioral results in the MEG session (n = 22). (A) Proportion of trials and (B) median RT for correct, rule errors, stimulus errors, and
other errors. Gray lines denote individual participants, and black markers denote group means. Error bars are 95% confidence intervals across
participants.
decoding. For each sensor in turn, we defined a search-
light consisting of that sensor and its immediate neighbors
(mean = 5.6 sensors per searchlight). We then ran the
same decoding schemes as on the whole head (above)
in each of these searchlights. Decoding accuracy for each
searchlight was plotted on the central sensor, resulting in a
head map of decoding accuracies, showing the regions
containing task-related information at a given time. For
each task feature and condition (e.g., Rule 1 vs. Rule 2
on stimulus-aligned correct trials), searchlight results were
plotted using 20-msec time windows of interest centered
around representative time points: 150 and 1000 msec for
the onset-aligned analyses, and −600 and −200 msec for
the response-aligned analyses.
Statistical Testing
To assess performance of the classifier, we used Bayesian
statistics to determine the evidence that decoding perfor-
mance was different from chance (Teichmann, Moerel,
Baker, & Grootswagers, 2021; Dienes, 2011, 2016; Rouder,
Speckman, Sun, Morey, & Iverson, 2009; Wagenmakers,
2007; Jeffreys, 1961). A Bayes factor (BF) is the probability
of the data under the alternative hypothesis relative to the
null hypothesis. In all analyses, the alternative hypotheses
of above- and below-chance (50%) decoding were tested
using the BayesFactor package in R (Morey et al., 2018).
BFs were calculated using a JZS prior, centered around
chance decoding of 50% (Rouder et al., 2009) with a
default scale factor of 0.707, meaning that for the alterna-
tive hypotheses of above- and below-chance decoding,
we expected to see 50% of parameter values falling
within −0.707 and 0.707 SDs from chance ( Wetzels &
Wagenmakers, 2012; Rouder et al., 2009; Zellner & Siow,
1980; Jeffreys, 1961). A null interval was specified as a
range of effect sizes between −0.5 and 0.5.
In accordance with the BF literature, we did not make a
correction for multiple comparisons for the large number
of time points tested (Teichmann et al., 2021; Świątkowski
& Carrier, 2020; Dienes, 2011, 2016). BFs assess the
strength of evidence for the null hypothesis versus the
alternative hypothesis; here, they are used to directly test
Table 1. Mean Number of Trials Used for Stimulus and Rule Decoding Analyses per Participant
Inner positions
Outer positions
Rule 1
Rule 2
Correct
218.55 (46.00)
218.55 (46.00)
218.55 (46.00)
218.55 (46.00)
Stimulus Error
30.00 (29.35)
48.50 (26.05)
38.14 (16.09)
40.36 (21.44)
Rule Error
17.55 (9.14)
28.14 (17.94)
22.18 (13.29)
23.50 (12.19)
Group means (standard deviation) for trial numbers are presented according to the two different stimulus classes (inner or outer positions; regard-
less of rule) and two rules (regardless of stimulus position). Trials are split by behavior: correct response, stimulus error, and rule error. Decoding
models were always trained using correct trials.
644
Journal of Cognitive Neuroscience
Volume 34, Number 4
the evidence for above-chance (or below-chance) decod-
ing and the evidence that decoding is equivalent to
chance. Thus, we assess the magnitude of evidence in
either direction, rather than a probability of the observed
data occurring by chance as in traditional null hypothesis
testing. BF > 3 is typically considered evidence for the
alternative hypothesis; and BF < 1/3, as evidence in favor
of the null hypothesis ( Wetzels et al., 2011; Jeffreys, 1961).
When the magnitude of BFs is interpreted at face value,
rather than applying a threshold for “significance,” there
is no need to correct for the number of tests (time points;
Teichmann et al., 2021). Additional tests provide addi-
tional evidence and can be interpreted as such without
correcting for multiple comparisons (Dienes, 2011,
2016). Accordingly, we do not interpret high BFs at iso-
lated time points as evidence for decoding; rather, we
assess the pattern of evidence through time in support
of the null or alternative hypotheses. Single time points
are not considered to provide substantial evidence if
neighboring time points support the opposite hypothesis.
We will refer to periods with sustained evidence for the
alternative hypothesis as times when information could
be decoded, indicating information was represented in
the brain.
RESULTS
Behavioral Results
All participants performed above 60% on the final block of
the response-mapping task in the training session and
therefore participated in the experimental MEG session.
In the MEG session, participants performed well above
chance (M = 81.92%) but still made both rule errors
(M = 5.71%) and stimulus position errors (M = 9.82%;
Figure 2A). There were very few other errors (M =
1.43%) or trials with no response (M = 1.13%). RTs were
slower for stimulus error trials than rule error and correct
trials (Figure 2B).
Temporal Dynamics of Goal-directed Behavior
First, we investigated neural coding during correct trials by
decoding different task-related information from the MEG
signal when each trial was aligned to stimulus onset (anal-
ogous to classic event-related analyses). We then realigned
the MEG signal of each trial to the response and per-
formed the same decoding analyses (see Figure 1D for a
depiction of realignment). This gives us a unique insight
into the time course of the processing stages during
goal-directed behavior.
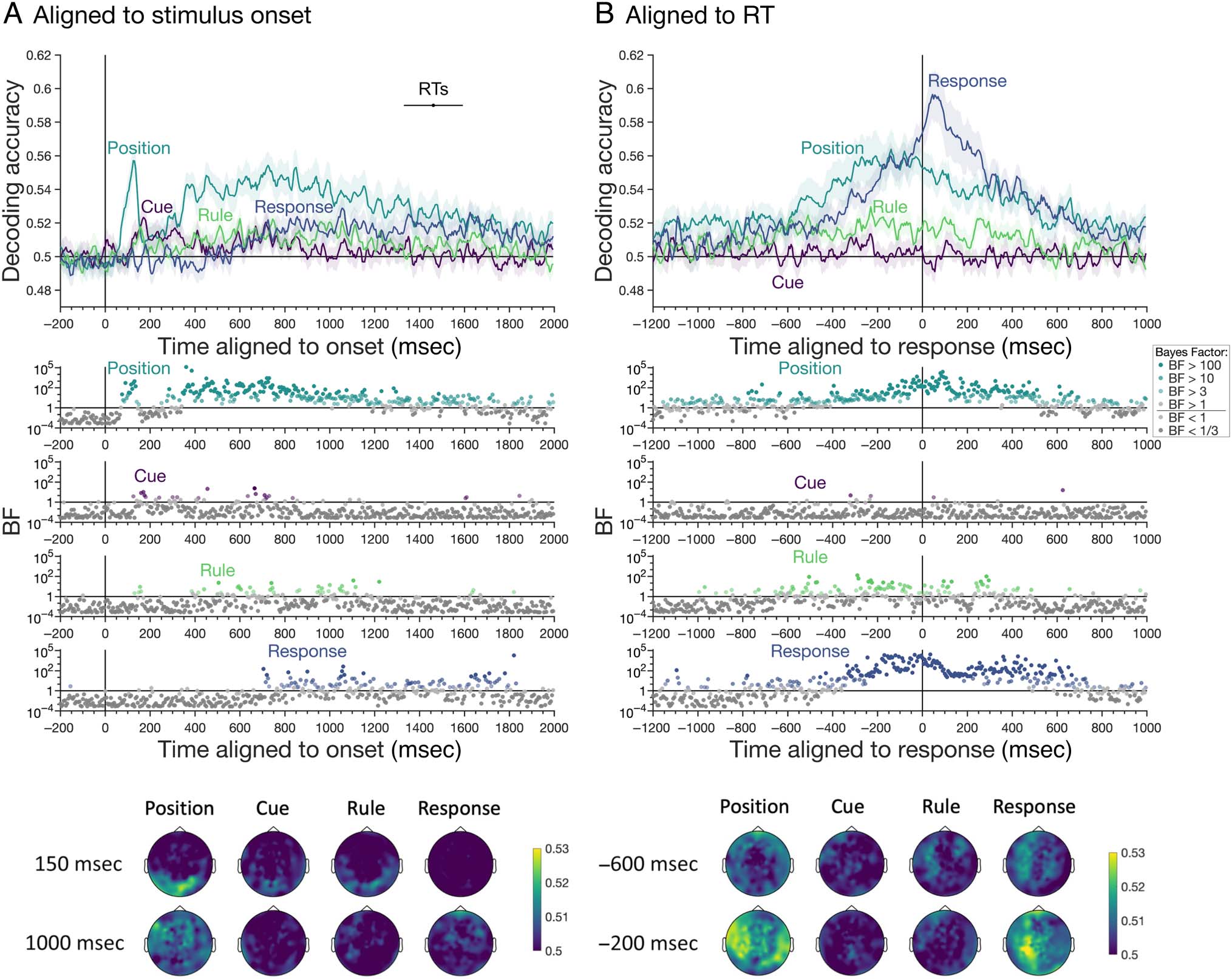
Time-resolved decoding performed relative to stimulus
onset revealed a progression of relevant information over
time (Figure 3A). Stimulus position information was repre-
sented in the neural signal from approximately 75 msec
after the stimulus appeared, with a double-peak response.
Cue information could be decoded from 170 msec, and
the dynamics were similar for the blue versus yellow
(53.34% at 170 msec) and green versus pink (51.86% at
170 msec) cue color decoding (mean presented in
Figure 3A), indicating the decoding reflects general cue
information rather than being specific to one set of cues.
The timing of stimulus and cue information was thus
consistent with early visual stages of retinotopic position
(Battistoni, Kaiser, Hickey, & Peelen, 2020; Carlson et al.,
2011) and color processing (Teichmann, Grootswagers,
Carlson, & Rich, 2019), respectively.
Rule information was briefly evident at about the same
time as the cues (around 150 msec), but also prominent
from approximately 400 to 1200 msec, likely coinciding
with higher-level cognitive stages of processing. Rule
information was quite low in general. One possibility is
that there was a carryover effect such that the rule type
on a given trial subsequently affected rule coding on the
next trial. In an additional exploratory analysis, we
assessed rule decoding separately for rule switch and rule
repeat trials by training Rule 1 versus Rule 2 on correct tri-
als (as in the original analysis) and testing on repeat and
switch trials separately. We found that rule decoding
exhibited similar dynamics regardless of the rule on the
previous trial, with slightly higher decoding on switch tri-
als (see OSF repository). Thus, inertia from the previous
trial did not appear to cause a switch cost in the represen-
tation of rule information in the brain.
Finally, we assessed the temporal dynamics of response
information. The button press response was represented
from about 715 msec, corresponding with RTs on some of
the faster trials. Together, decoding accuracy peaked at
125 msec for stimulus position, 175 msec for cue color,
600 msec for rule, and 1055 msec for response button,
showing a clear progression in information processing
through time.
Searchlight decoding results showed that position, cue,
and rule information at 150 msec localized to posterior
regions of the brain (Figure 3A, bottom). At a later period,
1000 msec, position and response information were more
frontal and diffuse across the MEG sensors. Rule informa-
tion was much lower in general but again showed a disso-
ciation for the early versus late time points, with frontal
and central topography at the later period.
As is typical in difficult tasks, there was a wide variation
in RTs across trials and participants, indicating that the
dynamics of high-level task-related processes such as
decision-making vary trial to trial with respect to stimulus
onset. Time-resolved decoding relies on processes occur-
ring at the same time across trials, so this temporal jitter
can mask results ( Vidaurre, Myers, Stokes, Nobre, &
Woolrich, 2019). To capture processes that are more
closely aligned with the response, we next realigned the
MEG data to the RT (Figure 1D) and performed the same
decoding analyses. The temporal dynamics of relevant
task-related information were markedly different com-
pared with onset-aligned results (Figure 3B). Notably,
cue information could no longer be reliably detected,
Robinson, Rich, and Woolgar
645
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
4
6
3
9
1
9
9
6
6
7
8
/
j
o
c
n
_
a
_
0
1
8
1
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
4
6
3
9
1
9
9
6
6
7
8
/
j
o
c
n
_
a
_
0
1
8
1
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. The temporal dynamics of correct stimulus position, cue color, rule, and response information coding. (A) Decoding analyses conducted
relative to stimulus onset. (B) Decoding analyses conducted relative to RT. Shaded areas show standard error across participants (n = 22). Decoding
accuracy is smoothed with a 20-msec window for visualization. Line at the top of Plot A marks the mean and 95% confidence interval of median
RTs per participant. BFs for above-chance decoding are displayed below the x axes for every time point using a log scale and color coded according
to the evidence for above-chance decoding (see inset). Head maps depict sensor searchlight decoding results for each task-related feature at
representative periods. The color bar indicates decoding accuracy per sensor searchlight, calculated as the mean of a 20-msec time window centered
on the time of interest.
presumably because cue color representations were
transient and tightly stimulus locked because, by design,
the cue distinctions were irrelevant as soon as rule infor-
mation could be extracted from them. In contrast, decod-
ing of stimulus position coding was evident more than
1000 msec before the response, and rule coding was
evident more than 600 msec before the response, with evi-
dence for stimulus processing earlier than rule processing.
Motor response coding was sporadically present from
more than 1000 msec before the response but was sus-
tained from around 485 msec before when the response
was made. Response coding peaked after the response
was given, potentially reflecting the contribution of
somatosensory feedback from the different button
presses. Response coding 200 msec before the response
was associated with highest decoding over central-left
sensors, which would be consistent with motor and
somatosensory cortex activity associated with a right-hand
response. Interestingly, the representation of stimulus
position and response information appeared to ramp up
before the response, plausibly reflecting the accumulation
of evidence leading to a decision.
Error Representations
We were particularly interested in understanding whether
and how the task-related information we can decode with
MVPA is related to participant performance. Specifically,
we investigated how information coding changes when
an error is made. Recall that our design explicitly allows
us to identify the likely source of the error based on the
behavioral response (Figure 1C). We assessed stimulus
646
Journal of Cognitive Neuroscience
Volume 34, Number 4
and rule information in the neural signal when participants
made stimulus errors versus rule errors. On the basis of
our previous work with fMRI ( Woolgar et al., 2019), we
hypothesized that the brain would represent the incorrect
stimulus before a stimulus error and the incorrect rule
before a rule error. Classifiers were trained to classify stim-
ulus position and rule using correct trials and tested on
incorrect trials. Therefore, for each time point on each
error trial, the analysis reveals whether activation patterns
were more similar to the usual patterns for the presented
rule and stimulus (correct rule and stimulus) or the alter-
nate one (incorrect) corresponding to the participant’s
decision (as shown by the behavior response).
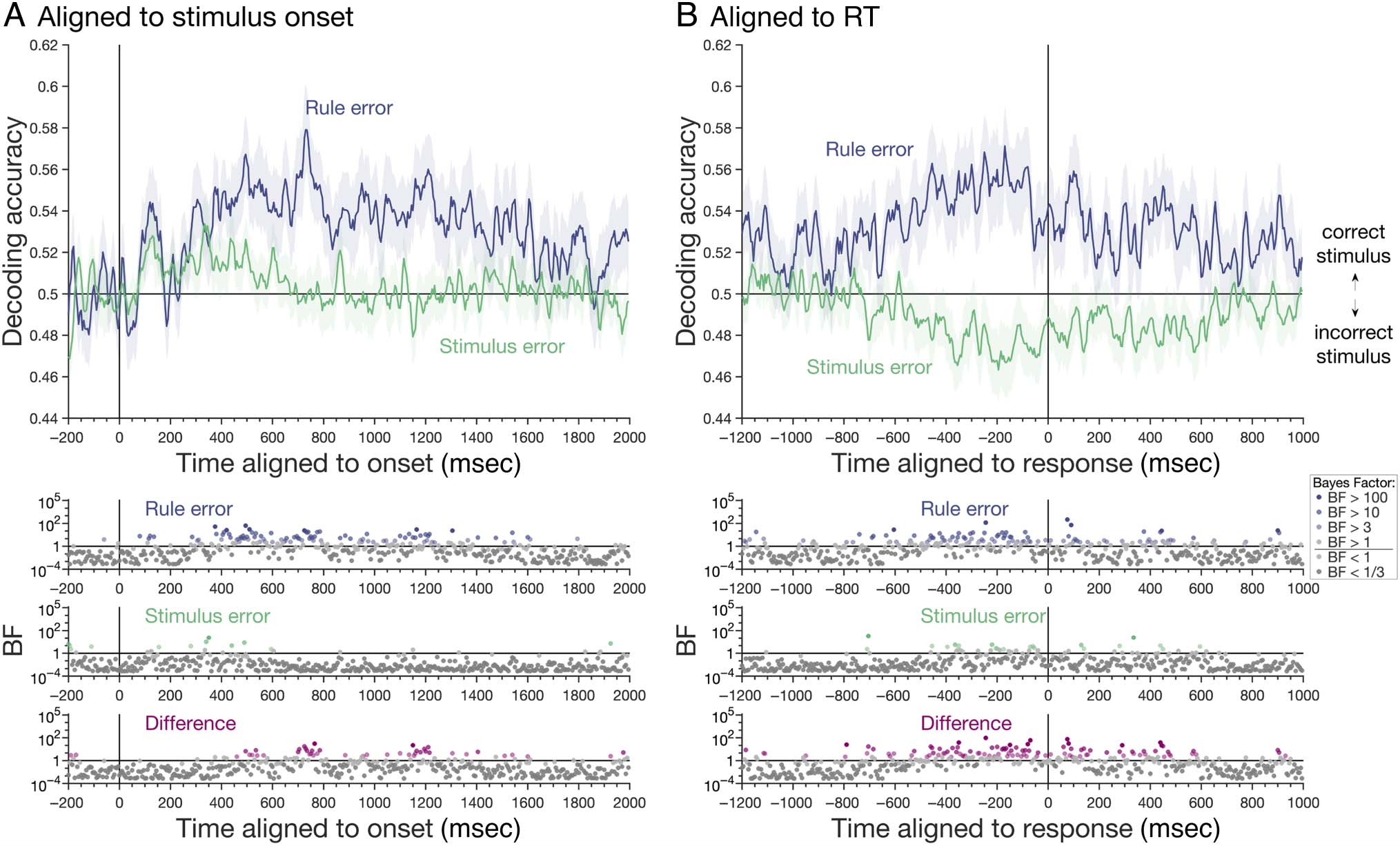
Stimulus Decoding—Aligned to Stimulus Onset
In this analysis, we looked at how stimulus position was
coded on error trials. We found that, when participants
made rule errors, in which behavior suggested that the
stimulus was encoded correctly but the incorrect rule
had been used, there was sustained stimulus decoding
with similar dynamics to that on correct trials (Figure 4A,
blue line). Stimulus coding on stimulus errors, however,
was present only transiently at 335 msec after which cod-
ing attenuated (Figure 4A, green line). After 495 msec,
there was substantial evidence that stimulus coding was
higher when participants made errors based on applying
the wrong rule than when the response suggested they
had misperceived the stimulus. This indicates that, when
participants made stimulus errors, correct stimulus infor-
mation was lost.
Stimulus Decoding—Aligned to RT
Next, we asked the same question but with the data rea-
ligned to the RT. On rule errors, there was a gradual ramp-
ing up of stimulus coding in the lead up to the response
(Figure 4B, blue line), as we had observed for correct
trials (Figure 3B). In contrast, on stimulus errors, activa-
tion patterns ramped toward the patterns encoding
the incorrect stimulus, as indexed by below-chance
decoding from approximately 355 msec before the
response (Figure 4B, green line). Given that the correct
stimulus had been encoded in the early part of these same
trials (Figure 4A), this suggests an evolution of information
coding toward the incorrect stimulus decision. Stimulus
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
4
6
3
9
1
9
9
6
6
7
8
/
j
o
c
n
_
a
_
0
1
8
1
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. Stimulus position decoding on rule and stimulus error trials. (A) Decoding analyses conducted relative to stimulus onset revealed initial
stimulus coding on both rule and stimulus error trials, with sustained stimulus coding on rule error trials (similar to correct trials; Figure 3), but no
stimulus coding at later time points on stimulus error trials. The interaction, shown by BF difference (pink), confirmed that at later time points, there
was more evidence for stimulus coding on rule errors than stimulus errors. (B) Decoding analyses conducted relative to RT revealed evidence for
correct stimulus coding on rule error trials and evidence for “incorrect” stimulus coding on stimulus error trials, evident as below-chance decoding
accuracy. Decoding accuracy is smoothed with a 20-msec window for visualization. BFs are shown on a log scale and color coded according to
amount of evidence.
Robinson, Rich, and Woolgar
647
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
4
6
3
9
1
9
9
6
6
7
8
/
j
o
c
n
_
a
_
0
1
8
1
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 5. Rule decoding on rule and stimulus error trials. (A) Decoding analyses conducted relative to stimulus onset. (B) Decoding analyses
conducted relative to RT. Decoding accuracy is smoothed with a 20-msec window for visualization. There was substantial evidence for the null (i.e.,
that rule could not be decoded) on both stimulus error and rule error trials, indicating the rule coding did not reverse for either type of error.
decoding accuracy on rule errors was higher than that
on stimulus errors for the bulk of the epoch, particularly
from about 795 msec before the response to 600 msec
after the response. Together, this finding shows that
stimulus coding in the latter part of the trial reflected
the decision ultimately made by participants, rather than
the stimulus presented.
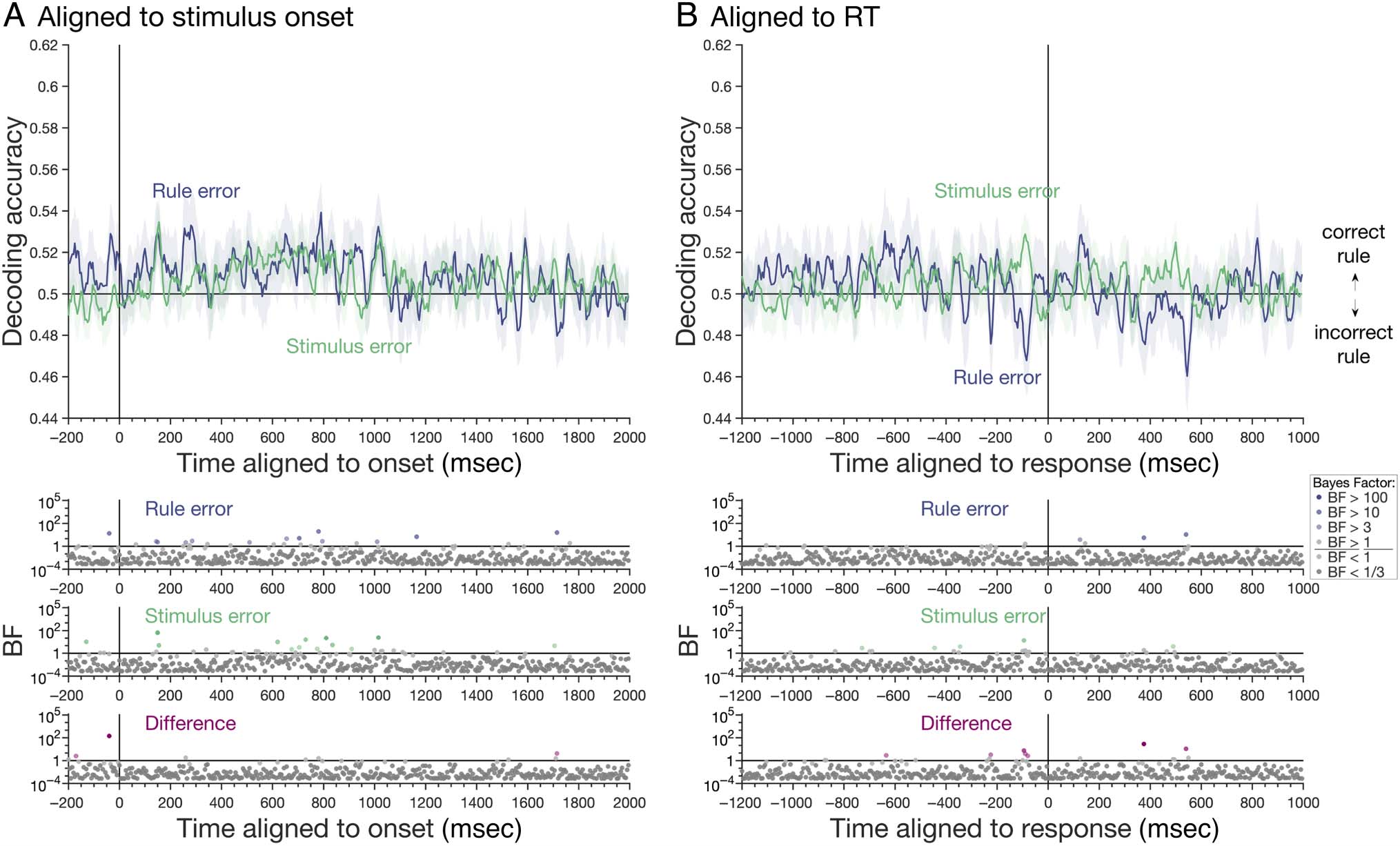
Rule Decoding
Next, we asked whether the representation of task rule
in the correct trials would also generalize to error trials.
However, there were only very brief periods of evidence
for rule information coding on rule errors and stimulus
errors, whether we aligned the MEG data to the stimulus
onset (Figure 5A) or response (Figure 5B). There was
also no difference in rule coding between error types.
For onset-aligned analyses, BFs indicated evidence for
above-chance decoding on rule and stimulus errors for
some time points, but it was not sustained. There were
also some brief periods of below-chance decoding on rule
errors for response-aligned analyses, which would indicate
coding of the incorrect rule, consistent with behavior, but
this did not reach our interpretation levels for BFs (two
consecutive time points, BF > 3). Overall, rule information
that had been (weakly) present on correct trials was largely
absent on both types of behavioral error. Moreover, the
“reversal” in coding—in this case, coding of the incorrect
rule—was not evident as it was for stimulus coding.
Taken together, the MEG decoding results show that,
on correct trials, all task-relevant aspects (stimulus posi-
tion, cue color, rule, and response) could be decoded.
The dynamics of coding varied such that analyses revealed
stimulus-locked coding of perceptual features (stimulus
position, cue) early in the time course, and analyses
aligned to the response revealed coding of relevant task
aspects (position, rule) and the resulting motor response
ramping up before the response being given. Strikingly,
the error decoding analysis showed that the increased
stimulus position coding in the latter part of the epoch
reflected the participants’ decision about the stimulus
more closely than the physical stimulus presented to
them, providing strong evidence for the connection
between specific neural responses decoded with MVPA
and behavior.
DISCUSSION
In this study, we used MVPA with MEG to characterize the
neural dynamics of stimulus, cue, rule, and response
648
Journal of Cognitive Neuroscience
Volume 34, Number 4
coding in a difficult response-mapping task as well as the
link between these codes and behavior. Our results
showed a clear and orderly progression of task-relevant
information coding after the stimulus was presented,
whereas analyses aligned to the RT revealed that informa-
tion coding for the stimulus and motoric response ramped
up over the ∼1 sec before the response was given, in a
manner reminiscent of evidence accumulation (e.g.,
Tagliabue et al., 2019; Pisauro, Fouragnan, Retzler, &
Philiastides, 2017). Strikingly, for trials on which partici-
pants made an error in the stimulus position, information
coding initially corresponded to physical stimulation, but
later accumulated in the opposite direction, so that activity
patterns at later time points resembled those encoding the
“incorrect” stimulus. This provides a crucial demonstra-
tion that patterns of neural activity recorded and classified
in this way can be predictive of behavior. These findings
give insight into the dynamics of processes underlying
cognitive control and provide a clear link between neural
responses and behavior.
The difficult response-mapping task implemented in
this study required complex processing for successful per-
formance. The task involved processing different types of
perceptual information (cue, stimulus), conversion of the
cue into the appropriate rule, application of the relevant
rule to the stimulus position, and selection of the correct
button-press response. Using MEG decoding with a
carefully balanced experimental design, we were able to
investigate the coding of each of these types of relevant
information over time and observe the succession of the
different task-related features. We summarize and con-
sider the findings below.
Information Coding After Stimulus Onset:
Correct Trials
Our results demonstrated different dynamics for percep-
tual, rule-related, and motor processes for analyses aligned
to stimulus onset (Figure 3A). Stimulus position was
represented early in the time course (<100 msec after
stimulus onset), consistent with early retinotopic visual
processes (Im, Gururajan, Zhang, Chen, & He, 2007;
Di Russo et al., 2005). The cue was represented shortly
thereafter at a time that is general color
processing and in line previous work found color
decoding most evident from 135–155 after
image onset (Teichmann 2019, 2020). Rule informa-
tion, by contrast, around 600 and
maintained for longer than information, perhaps
reflecting the ongoing process of combining stimulus
and rule information to derive response. Motor
responses emerged last exhibited broad, shallow
peak, perhaps reflective wide range RTs the
task. Our data thus emphasize progression coding
for different types task-relevant over time,
despite relevant sensory (stimulus and
cue) being presented simultaneously.
These onset-aligned analyses are with
previous time-resolved multivariate showed
progression task-related after stimulus pre-
sentation (Kikumoto Mayr, 2020; Hubbard 2019;
Wen 2019; Hebart 2018). For example, Hebart
et al. (2018) showed was
evident MEG signal shortly task pre-
sented but ramped up again target object was
presented. In more complex design, al.
(2019) used cued task-switching EEG, which allowed
them look coding multiple aspects
over using oscillatory power neural signal. Like
our results, they pre-
ceded decoding pro-
longed throughout trial period. Relevant irrelevant
stimulus stimuli were
presented, response present later
in Here, we show similar cascade infor-
mation arises when simultaneously. results this work
show features the
brain temporal dynamics, there are
periods represented,
potentially giving correlates information
integration needed on these tasks.
The color observed indicative of
transient processing before was
selected, point distinctions (e.g., blue
vs. yellow, both indicate 1) become irrelevant.
In prolonged position
and (far exceeding presentation of
500 msec) likely reflects position information
being maintained, accumulated,>500 msec after stimulus onset)
would be theoretically consistent with processing within
higher-level frontoparietal regions. For example, previous
combined MEG–fMRI research has shown task-related
enhancement of relevant features occurred after 500 msec
following stimulus presentation, and task coding in poste-
rior parietal cortex and lateral pFC seemed to peak from
500 msec (Hebart et al., 2018). Moreover, attention
enhances task-relevant information in frontoparietal
regions from 500 msec (Moerel, Rich, & Woolgar, 2021).
On the other hand, task-relevant stimulus information also
seems to persist in occipital regions until these late time
points (Moerel et al., 2021; Hebart et al., 2018; Goddard,
Carlson, Dermody, & Woolgar, 2016). Our exploratory
sensor searchlight analyses suggested that early position,
Robinson, Rich, and Woolgar
649
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
4
4
6
3
9
1
9
9
6
6
7
8
/
j
o
c
n
_
a
_
0
1
8
1
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
cue, and rule information was evident in occipital sensors,
whereas later position information was more diffuse
across the brain, and later rule information was more
frontal. Future work could address the spatial nature of
these processes with more precision, perhaps using
computational methods to combine fMRI and MEG data
such as similarity-based fusion (Cichy, Pantazis, & Oliva,
2014, 2016).
Information Coding Before the Response:
Correct Trials
The analyses aligned to the RT provided rich additional
information about the temporal dynamics of stimulus,
cue, rule, and response coding (Figure 3B). We expected
that these response-aligned analyses would emphasize
higher-level decision-related processes required for
behavior, which might not be so salient in data aligned
to stimulus onset because of variability in their timing
( Vidaurre et al., 2019). Previous EEG work has shown neu-
ral signals ramp up during perceptual decision-making,
which has been described as evidence accumulation
(e.g., Tagliabue et al., 2019; Pisauro et al., 2017), but these
effects could be related to a general decision-making pro-
cess rather than involving information about the stimulus
of interest and could be confounded with preparatory
motor activity. Here, using decoding, we were able to
assess the dynamics of different types of task-related infor-
mation, separate from and in addition to response infor-
mation, that was represented in the brain before the
response. The results revealed an increase in stimulus
information from approximately 1000 msec before the
response that peaked around the RT, a pattern that was
noticeably absent in the onset-locked analyses. Response
coding, by contrast, showed a later, sharper ramping
in information that peaked just after the response was
made. The ramping of stimulus position and response
coding was, for the most part, when the stimulus and
cue were no longer visible; the stimulus and cue were only
presented for 500 msec, and the median RT was over
1400 msec, so on a typical trial, there was no stimulus pre-
sented in the 900 msec before the response. Therefore,
instead of perceptual accumulation, these pre-response
representations appear to be internally generated codes
that reflect the system moving toward different end states
as the person arrives at his or her decision.
The results revealed concurrent coding of position,
rule, and response information before the response,
which might reflect the need to combine position and rule
information to select the appropriate response. Kikumoto
and Mayr (2020) recently investigated the temporal
dynamics of action selection using EEG in a cued rule
selection task and provided evidence that conjunctions
between task-relevant features are necessary for action
selection. In addition to the succession of individual task
features, they found rule stimulus–response conjunctive
representations could be decoded using stimulus-aligned
EEG, and the strength of the conjunctive information was
associated with faster responses, providing a link with
behavior. Other work has used temporal decomposition
of EEG data and concluded that stimulus–response bind-
ings have different temporal profiles to stimulus informa-
tion, with gradual activation and decay over time (Takacs,
Mückschel, Roessner, & Beste, 2020). Our exploratory
searchlight decoding results showed that position and rule
information before the response was mostly lateral and dif-
fuse across the sensors but with similar spatial patterns
(although lower for rule decoding), potentially reflecting
the integration of task-relevant information within brain
regions. We did not explicitly set out to look at conjunctive
representations, but our results certainly fit with this
account. During goal-directed behavior, it seems that mul-
tiple task-relevant features are represented concurrently,
presumably reflecting the need for this information to
be maintained, and are then combined over time.
Our onset-aligned and response-aligned analyses
revealed complementary aspects of the data. The pattern
of results suggests that onset-aligned analyses may be
most sensitive to perceptual responses, whereas response-
aligned analyses may capture processes that are time-
varied relative to stimulus onset and more closely yoked
to the time of response, such as higher decision and motor
preparation processes. Specifically, we found that stimulus
position and cue color had sharp initial decoding when
aligned to stimulus onset, which was not visible after
realignment to response. However, neural representa-
tions of stimulus and response exhibited a ramping accu-
mulation before the button was pressed, which was not
visible in onset-aligned data. This highlights the utility of
including both approaches, perhaps particularly for diffi-
cult tasks with substantial RT variability, to yield additional
information about the dynamics underlying successful
task performance.
Information Coding Leading to Incorrect Behavior:
Error Trials
To test whether the neural coding of task-relevant infor-
mation detected with MVPA reflects activity necessary to
successfully perform a task, we examined how these codes
changed when participants made errors. We focused on
decoding stimulus and rule information during stimulus
errors and rule errors, situations in which the decision
made could be dissociated from the stimulus and rule
cue presented. Stimulus errors consisted of trials on which
participants correctly applied the rule but confused the
stimulus. Despite the behavioral evidence for correct rule
use on these trials, there was only some evidence of rule
coding, perhaps reflecting weak rule coding in general (on
correct trials) and the limited number of error trials. How-
ever, stimulus position coding on stimulus error trials
revealed a striking result: Initial stimulus coding showed
some fleeting evidence of the correct stimulus neural pat-
tern, but before the response, stimulus coding became
650
Journal of Cognitive Neuroscience
Volume 34, Number 4
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
4
4
6
3
9
1
9
9
6
6
7
8
/
j
o
c
n
_
a
_
0
1
8
1
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
consistent with the incorrect stimulus. Thus, onset-aligned
analyses and responses at early time points reflected
perception, whereas response-aligned analyses and
coding at later time points reflected behavior. Recall in
the paradigm that stimulus position was designed to be
confusable, and a stimulus position error, by definition,
means participants confused two (of four) stimulus posi-
tions. When the stimulus was presented, participants
would see it, which is consistent with brief veridical stim-
ulus position decoding, but the insufficient maintenance
of this information correlates with the behavioral perfor-
mance: Participants could not localize the stimulus pre-
cisely, which led to a decision in favor of the wrong
stimulus. It is this internal decision-related process that
seems to be reflected in below-chance (incorrect stim-
ulus) decoding before the response. Previous work has
shown that higher, but not early, perceptual regions
reflect behavior in terms of accuracy ( Walther et al.,
2009; Williams et al., 2007) and RT (Grootswagers et al.,
2018), although none of these studies revealed the code
reversal needed for a strong link with behavior. Here, we
used the temporal domain to show “what” was coded on
error trials at different stages of processing. There was a
dissociation between the coding of early perceptual
information and the stimulus decision used to generate
the behavioral response.
Rule errors consisted of trials on which participants
appeared (in their behavior) to apply the wrong rule to
the correct stimulus. Accordingly, for stimulus position
coding, the classifier trained on correct trials could suc-
cessfully classify the stimulus position after onset and
before the response on rule error trials. This indicates that
the stimulus coding reversal seen above was diagnostic of
the particular type of behavioral error, rather than reflec-
tive of errors in general, indicating a tight link with the
specific decision made and reflected in behavior.
For rule coding, we again found little evidence for rule
coding on rule error trials. A couple of time points just
before the response showed patterns of activity consistent
with the incorrect rule, as we had predicted for a full dou-
ble dissociation, but the effect was so transient that it is
difficult to interpret with confidence. This may reflect
the very small number of rule error trials and/or the rela-
tively weak coding of rule information in general in our
data (potentially attributable to more variability in timing
of this task aspect and/or relatively poor signal from frontal
regions that are further from the sensors in our supine
MEG system). This limitation means that, in contrast to
the stimulus information, we cannot conclude with confi-
dence whether or not the rule patterns we decoded were
closely linked to behavior.
Our research contributes to the growing literature
drawing links between neural responses and behavior
using MVPA. Using spatial and temporal neuroimaging,
classifier prediction errors and distances from the classifier
boundary have been shown to correlate with behavioral
error patterns and RTs (e.g., González-García, Formica,
Wisniewski, & Brass, 2021; Grootswagers et al., 2018;
Carlson, Ritchie, Kriegeskorte, Durvasula, & Ma, 2014;
Walther et al., 2009). Here, we argue that a tighter link
between brain and behavior can be found by testing
“what” is represented on error trials when an incorrect
decision is made. The results parallel fMRI work showing
frontoparietal MD regions represent the correct stimulus
but wrong rule during a rule error as well as the correct
rule but wrong stimulus during other errors ( Woolgar
et al., 2019). The current study extends this work by
elucidating the dynamics with which the incorrect repre-
sentations evolve over time, with early representations
reflecting the stimuli presented, but a late gradual accu-
mulation toward the opposite stimulus at time points just
before behavioral response. We also show here that there
is a dissociation in the perceptual coding (indexed by
onset-aligned analyses) and high-level decision coding
(indexed by response-aligned analyses) for error trials.
Specifically, on stimulus errors, after a transient representa-
tion of the veridical stimulus, activity accumulated toward
a pattern state reflecting the opposite and incorrect stim-
ulus, apparently reflecting the internal generation of
accumulation toward the wrong decision. This pattern
was specifically diagnostic of behavioral errors attributable
to stimulus misperception, as position information was
coded correctly on other types of behavioral errors.
The results of this study provide new insights into how
task-relevant information is processed in the human brain
to allow successful goal-directed behavior. There was a
clear progression of the onset of task-relevant information
in the brain, from stimulus position and cue, to rule and
then response information. Complimentary response-
aligned analyses, which highlight later high-level pro-
cesses aligned in time to behavior, additionally revealed
dynamics of information coding resembling an accumu-
lation of multiple types of task-relevant information.
Moreover, when participants made behavioral errors,
the direction of accumulation was reversed. Under these
conditions, the trajectory of representation moved in the
opposite direction such that the neural pattern increas-
ingly represented the incorrect stimulus (which had not
been shown) in a manner diagnostic of the subsequent
behavioral choice. The data highlight the orderly but
overlapping dynamics with which several task elements
can be represented in brain activity. Our findings empha-
size a particular role for the trajectory of information
coding at later time points in determining behavioral suc-
cess or failure and also demonstrate the utility of aligning
neural data differently to examine high-level complex
cognitive processes.
Acknowledgments
We thank Christopher Whyte for assistance in data collection
and Dr. Tijl Grootswagers for helpful discussions. We acknowl-
edge the Sydney Informatics Hub and the University of
Sydney’s high-performance computing cluster Artemis for
Robinson, Rich, and Woolgar
651
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
4
4
6
3
9
1
9
9
6
6
7
8
/
j
o
c
n
_
a
_
0
1
8
1
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
providing high-performance computing resources that contrib-
uted to these research results.
Reprint requests should be sent to Amanda K. Robinson,
School of Psychology, The University of Sydney, Camperdown,
NSW 2006, Australia, or via e-mail: amanda.robinson@sydney
.edu.au.
Funding Information
This work was funded by Australian Research Council
D i s c o v e r y P r o j e c t ( h t t p s : / / d x . d o i . o r g / 1 0 . 1 3 0 3 9
/501100000923), grant number: 170101840, Australian
Research Council Future Fellowship (https://dx.doi.org
/10.13039/501100000923), grant number: FT170100105,
Medical Research Council (UK) intramural funding, grant
number: SUAG/052/G101400; and Australian Research
Council Discovery Early Career Researcher Award (https://
dx.doi.org/10.13039/501100000923), grant number:
DE200101159.
Diversity in Citation Practices
Retrospective analysis of the citations in every article pub-
lished in this journal from 2010 to 2021 reveals a persistent
pattern of gender imbalance: Although the proportions of
authorship teams (categorized by estimated gender iden-
tification of first author/last author) publishing in the
Journal of Cognitive Neuroscience ( JoCN) during this
period were M(an)/M = .407, W(oman)/M = .32, M/W =
.115, and W/W = .159, the comparable proportions for the
articles that these authorship teams cited were M/M =
.549, W/M = .257, M/W = .109, and W/W = .085 (Postle
and Fulvio, JoCN, 34:1, pp. 1–3). Consequently, JoCN
encourages all authors to consider gender balance explic-
itly when selecting which articles to cite and gives them
the opportunity to report their article’s gender citation
balance. The authors of this article report its proportions
of citations by gender category to be as follows: M/M =
.536, W/M = .145, M/ W = .130, and W/ W = .188.
REFERENCES
Battistoni, E., Kaiser, D., Hickey, C., & Peelen, M. V. (2020). The
time course of spatial attention during naturalistic visual
search. Cortex, 122, 225–234. https://doi.org/10.1016/j.cortex
.2018.11.018, PubMed: 30563703
Bracci, S., Daniels, N., & Op de Beeck, H. (2017). Task context
overrules object- and category-related representational content
in the human parietal cortex. Cerebral Cortex, 27, 310–321.
https://doi.org/10.1093/cercor/bhw419, PubMed: 28108492
Carlson, T. A., Grootswagers, T., & Robinson, A. K. (2020). An
introduction to time-resolved decoding analysis for M/EEG.
In M. G. Gazzaniga, G. R. Mangun & D. Poeppel (Eds.), The
cognitive neurosciences. Cambridge, MA: MIT Press.
Carlson, T. A., Hogendoorn, H., Kanai, R., Mesik, J., & Turret, J.
(2011). High temporal resolution decoding of object position
and category. Journal of Vision, 11, 1–17. https://doi.org/10
.1167/11.10.9.Introduction, PubMed: 21920851
Carlson, T. A., Ritchie, J. B., Kriegeskorte, N., Durvasula, S., &
Ma, J. (2014). Reaction time for object categorization is
predicted by representational distance. Journal of Cognitive
Neuroscience, 26, 132–142. https://doi.org/10.1162/jocn_a
_00476, PubMed: 24001004
Cichy, R. M., Pantazis, D., & Oliva, A. (2014). Resolving human
object recognition in space and time. Nature Neuroscience, 17,
455–462. https://doi.org/10.1038/nn.3635, PubMed: 24464044
Cichy, R. M., Pantazis, D., & Oliva, A. (2016). Similarity-based
fusion of MEG and fMRI reveals spatio-temporal dynamics
in human cortex during visual object recognition. Cerebral
Cortex, 26, 3563–3579. https://doi.org/10.1093/cercor/bhw135,
PubMed: 27235099
Cole, M. W., & Schneider, W. (2007). The cognitive control
network: Integrated cortical regions with dissociable
functions. Neuroimage, 37, 343–360. https://doi.org/10.1016
/j.neuroimage.2007.03.071, PubMed: 17553704
Crittenden, B. M., Mitchell, D. J., & Duncan, J. (2016). Task
encoding across the multiple demand cortex is consistent
with a frontoparietal and cingulo-opercular dual networks
distinction. Journal of Neuroscience, 36, 6147–6155. https://doi
.org/10.1523/JNEUROSCI.4590-15.2016, PubMed: 27277793
Delorme, A., & Makeig, S. (2004). EEGLAB: An open source
toolbox for analysis of single-trial EEG dynamics including
independent component analysis. Journal of Neuroscience
Methods, 134, 9–21. https://doi.org/10.1016/j.jneumeth.2003
.10.009, PubMed: 15102499
de Wit, L., Alexander, D., Ekroll, V., & Wagemans, J. (2016). Is
neuroimaging measuring information in the brain?
Psychonomic Bulletin & Review, 23, 1415–1428. https://doi
.org/10.3758/s13423-016-1002-0, PubMed: 26833316
Di Russo, F., Pitzalis, S., Spitoni, G., Aprile, T., Patria, F., Spinelli,
D., et al. (2005). Identification of the neural sources of the
pattern-reversal VEP. Neuroimage, 24, 874–886. https://doi
.org/10.1016/j.neuroimage.2004.09.029, PubMed: 15652322
Dienes, Z. (2011). Bayesian versus orthodox statistics: Which
side are you on? Perspectives on Psychological Science,
6, 274–290. https://doi.org/10.1177/1745691611406920,
PubMed: 26168518
Dienes, Z. (2016). How Bayes factors change scientific practice.
Journal of Mathematical Psychology, 72, 78–89. https://doi
.org/10.1016/j.jmp.2015.10.003
Duncan, J. (2010). The multiple-demand (MD) system of the
primate brain: Mental programs for intelligent behavior.
Trends in Cognitive Sciences, 14, 172–179. https://doi.org/10
.1016/j.tics.2010.01.004, PubMed: 20171926
Duncan, J., Assem, M., & Shashidhara, S. (2020). Integrated
intelligence from distributed brain activity. Trends in
Cognitive Sciences, 24, 838–852. https://doi.org/10.1016/j.tics
.2020.06.012, PubMed: 32771330
Folstein, J. R., & Van Petten, C. (2008). Influence of cognitive
control and mismatch on the N2 component of the ERP: A
review. Psychophysiology, 45, 152–170. https://doi.org/10
.1111/j.1469-8986.2007.00602.x, PubMed: 17850238
Fox, M. D., Snyder, A. Z., Vincent, J. L., Corbetta, M., Van Essen,
D. C., & Raichle, M. E. (2005). The human brain is intrinsically
organized into dynamic, anticorrelated functional networks.
Proceedings of the National Academy of Sciences, U.S.A.,
102, 9673–9678. https://doi.org/10.1073/pnas.0504136102,
PubMed: 15976020
Goddard, E., Carlson, T. A., Dermody, N., & Woolgar, A. (2016).
Representational dynamics of object recognition: Feedforward
and feedback information flows. Neuroimage, 128, 385–397.
https://doi.org/10.1016/j.neuroimage.2016.01.006, PubMed:
26806290
González-García, C., Formica, S., Wisniewski, D., & Brass, M.
(2021). Frontoparietal action-oriented codes support novel
instruction implementation. Neuroimage, 226, 117608.
https://doi.org/10.1016/j.neuroimage.2020.117608, PubMed:
33271270
652
Journal of Cognitive Neuroscience
Volume 34, Number 4
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
4
4
6
3
9
1
9
9
6
6
7
8
/
j
o
c
n
_
a
_
0
1
8
1
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Gratton, G., Cooper, P., Fabiani, M., Carter, C. S., & Karayanidis, F.
(2017). Dynamics of cognitive control: Theoretical bases,
paradigms, and a view for the future. Psychophysiology, 55,
e13016. https://doi.org/10.1111/psyp.13016
Grootswagers, T., Cichy, R. M., & Carlson, T. A. (2018). Finding
decodable information that can be read out in behavior.
Neuroimage, 179, 252–262. https://doi.org/10.1016/j
.neuroimage.2018.06.022, PubMed: 29886145
Grootswagers, T., Wardle, S. G., & Carlson, T. A. (2017).
Decoding dynamic brain patterns from evoked responses: A
tutorial on multivariate pattern analysis applied to time series
neuroimaging data. Journal of Cognitive Neuroscience, 29,
677–697. https://doi.org/10.1162/jocn_a_01068, PubMed:
27779910
Haxby, J. V. (2012). Multivariate pattern analysis of fMRI: The
early beginnings. Neuroimage, 62, 852–855. https://doi.org
/10.1016/j.neuroimage.2012.03.016, PubMed: 22425670
Haxby, J. V., Gobbini, M. I., Furey, M. L., Ishai, A., Schouten,
J. L., & Pietrini, P. (2001). Distributed and overlapping
representations of faces and objects in ventral temporal
cortex. Science, 293, 2425–2430. https://doi.org/10.1126
/science.1063736, PubMed: 11577229
Hebart, M. N., & Baker, C. I. (2018). Deconstructing
multivariate decoding for the study of brain function.
Neuroimage, 180, 4–18. https://doi.org/10.1016/j.neuroimage
.2017.08.005, PubMed: 28782682
Hebart, M. N., Bankson, B. B., Harel, A., Baker, C. I., & Cichy, R. M.
(2018). The representational dynamics of task and object
processing in humans. eLife, 7, 1–21. https://doi.org/10.7554
/eLife.32816, PubMed: 29384473
Hubbard, J., Kikumoto, A., & Mayr, U. (2019). EEG decoding
reveals the strength and temporal dynamics of goal-relevant
representations. Scientific Reports, 9, 9051. https://doi.org/10
.1038/s41598-019-45333-6, PubMed: 31227796
Im, C.-H., Gururajan, A., Zhang, N., Chen, W., & He, B. (2007).
Spatial resolution of EEG cortical source imaging revealed by
localization of retinotopic organization in human primary visual
cortex. Journal of Neuroscience Methods, 161, 142–154. https://
doi.org/10.1016/j.jneumeth.2006.10.008, PubMed: 17098289
Jackson, J. B., Feredoes, E., Rich, A. N., Lindner, M., & Woolgar,
A. (2021). Concurrent neuroimaging and neurostimulation
reveals a causal role for dlPFC in coding of task-relevant
information. Communications Biology, 4, 1–16. https://doi
.org/10.1038/s42003-021-02109-x, PubMed: 34002006
Jackson, J., Rich, A. N., Williams, M. A., & Woolgar, A. (2017).
Feature-selective attention in frontoparietal cortex:
Multivoxel codes adjust to prioritize task-relevant information.
Journal of Cognitive Neuroscience, 29, 310–321. https://doi
.org/10.1162/jocn_a_01039, PubMed: 27626230
Jackson, J. B., & Woolgar, A. (2018). Adaptive coding in the
human brain: Distinct object features are encoded by
overlapping voxels in frontoparietal cortex. Cortex, 108,
25–34. https://doi.org/10.1016/j.cortex.2018.07.006, PubMed:
30121000
Jeffreys, H. (1961). Theory of probability (3rd ed.). Oxford:
Oxford University Press.
Kado, H., Higuchi, M., Shimogawara, M., Haruta, Y., Adachi, Y.,
Kawai, J., et al. (1999). Magnetoencephalogram systems
developed at KIT. IEEE Transactions on Applied
Superconductivity, 9, 4057–4062. https://doi.org/10.1109/77
.783918
Karayanidis, F., Jamadar, S., Ruge, H., Phillips, N., Heathcote, A., &
Forstmann, B. U. (2010). Advance preparation in task-switching:
Converging evidence from behavioral, brain activation, and
model-based approaches. Frontiers in Psychology, 1, 25.
https://doi.org/10.3389/fpsyg.2010.00025, PubMed: 21833196
Karimi-Rouzbahani, H., Woolgar, A., & Rich, A. N. (2021). Neural
signatures of vigilance decrements predict behavioral errors
before they occur. eLife, 10, e60563. https://doi.org/10.7554
/eLife.60563, PubMed: 33830017
Kikumoto, A., & Mayr, U. (2020). Conjunctive representations
that integrate stimuli, responses, and rules are critical for
action selection. Proceedings of the National Academy of
Sciences, U.S.A., 117, 10603–10608. https://doi.org/10.1073
/pnas.1922166117, PubMed: 32341161
Long, N. M., & Kuhl, B. A. (2018). Bottom–up and top–down
factors differentially influence stimulus representations
across large-scale attentional networks. Journal of
Neuroscience, 38, 2495–2504. https://doi.org/10.1523
/JNEUROSCI.2724-17.2018, PubMed: 29437930
Moerel, D., Rich, A. N., & Woolgar, A. (2021). Selective attention
and decision-making have separable neural bases in space and
time [preprint]. BioRxiv, 2021.02.28.433294. https://doi.org
/10.1101/2021.02.28.433294
Morey, R. D., Rouder, J. N., Jamil, T., Urbanek, S., Forner, K., &
Ly, A. (2018). Package “BayesFactor”. https://cran.r-project
.org/web/packages/BayesFactor/BayesFactor.pdf
Oosterhof, N. N., Connolly, A. C., & Haxby, J. V. (2016).
CoSMoMVPA: Multi-modal multivariate pattern analysis of
neuroimaging data in MATLAB/GNU octave. Frontiers in
Neuroinformatics, 10, 27. https://doi.org/10.3389/fninf.2016
.00027, PubMed: 27499741
Pisauro, M. A., Fouragnan, E., Retzler, C., & Philiastides, M. G.
(2017). Neural correlates of evidence accumulation during
value-based decisions revealed via simultaneous EEG–fMRI.
Nature Communications, 8, 15808. https://doi.org/10.1038
/ncomms15808, PubMed: 28598432
Posner, M. I., & Presti, D. E. (1987). Selective attention and
cognitive control. Trends in Neurosciences, 10, 13–17.
https://doi.org/10.1016/0166-2236(87)90116-0
Posner, M. I., & Snyder, C. R. R. (1975). Attention and cognitive
control. In R. L. Solso (Ed.), Information processing and
cognition: The Loyola symposium. Erlbaum.
Ritchie, J. B., & Carlson, T. A. (2016). Neural decoding and
“inner” psychophysics: A distance-to-bound approach for
linking mind, brain, and behavior. Frontiers in Neuroscience,
10, 1–8. https://doi.org/10.3389/fnins.2016.00190, PubMed:
27199652
Ritchie, J. B., Tovar, D. A., & Carlson, T. A. (2015). Emerging
object representations in the visual system predict reaction
times for categorization. PLoS Computational Biology, 11,
e1004316. https://doi.org/10.1371/journal.pcbi.1004316,
PubMed: 26107634
Robinson, A. K., Grootswagers, T., & Carlson, T. A. (2019). The
influence of image masking on object representations during
rapid serial visual presentation. Neuroimage, 197, 224–231.
https://doi.org/10.1016/j.neuroimage.2019.04.050, PubMed:
31009746
Rouder, J. N., Speckman, P. L., Sun, D., Morey, R. D., & Iverson,
G. (2009). Bayesian t tests for accepting and rejecting the null
hypothesis. Psychonomic Bulletin & Review, 16, 225–237.
https://doi.org/10.3758/PBR.16.2.225, PubMed: 19293088
Świątkowski, W., & Carrier, A. (2020). There is nothing magical
about Bayesian statistics: An introduction to epistemic
probabilities in data analysis for psychology starters. Basic
and Applied Social Psychology, 42, 387–412. https://doi.org
/10.1080/01973533.2020.1792297
Tagliabue, C. F., Veniero, D., Benwell, C. S. Y., Cecere, R.,
Savazzi, S., & Thut, G. (2019). The EEG signature of sensory
evidence accumulation during decision formation closely
tracks subjective perceptual experience. Scientific Reports, 9,
4949. https://doi.org/10.1038/s41598-019-41024-4, PubMed:
30894558
Takacs, A., Mückschel, M., Roessner, V., & Beste, C. (2020).
Decoding stimulus–response representations and their
stability using EEG-based multivariate pattern analysis.
Robinson, Rich, and Woolgar
653
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
4
4
6
3
9
1
9
9
6
6
7
8
/
j
o
c
n
_
a
_
0
1
8
1
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Cerebral Cortex Communications, 1, tgaa016. https://doi.org
/10.1093/texcom/tgaa016, PubMed: 34296094
Teichmann, L., Grootswagers, T., Carlson, T. A., & Rich, A. N.
(2019). Seeing versus knowing: The temporal dynamics of
real and implied color processing in the human brain.
Neuroimage, 200, 373–381. https://doi.org/10.1016/j
.neuroimage.2019.06.062, PubMed: 31254648
Teichmann, L., Moerel, D., Baker, C., & Grootswagers, T.
(2021). An empirically-driven guide on using Bayes factors for
M/EEG decoding. BioRxiv, 2021.06.23.449663. https://doi.org
/10.1101/2021.06.23.449663
Teichmann, L., Quek, G. L., Robinson, A. K., Grootswagers, T.,
Carlson, T. A., & Rich, A. N. (2020). The influence of
object-color knowledge on emerging object representations
in the brain. Journal of Neuroscience, 40, 6779–6789.
https://doi.org/10.1523/JNEUROSCI.0158-20.2020, PubMed:
32703903
Vidaurre, D., Myers, N. E., Stokes, M., Nobre, A. C., & Woolrich,
M. W. (2019). Temporally unconstrained decoding reveals
consistent but time-varying stages of stimulus processing.
Cerebral Cortex, 29, 874–863. https://doi.org/10.1093/cercor
/bhy290, PubMed: 30535141
Vincent, J. L., Kahn, I., Snyder, A. Z., Raichle, M. E., & Buckner,
R. L. (2008). Evidence for a frontoparietal control system
revealed by intrinsic functional connectivity. Journal of
Neurophysiology, 100, 3328–3342. https://doi.org/10.1152/jn
.90355.2008, PubMed: 18799601
Wagenmakers, E. (2007). A practical solution to the pervasive
problems of p values. Psychonomic Bulletin and Review, 14,
779–804. https://doi.org/10.3758/BF03194105, PubMed:
18087943
Walther, D. B., Caddigan, E., Fei-Fei, L., & Beck, D. M. (2009).
Natural scene categories revealed in distributed patterns of
activity in the human brain. Journal of Neuroscience, 29,
10573–10581. https://doi.org/10.1523/JNEUROSCI.0559-09
.2009, PubMed: 19710310
Wen, T., Duncan, J., & Mitchell, D. J. (2019). The time-course of
component processes of selective attention. Neuroimage,
199, 396–407. https://doi.org/10.1016/j.neuroimage.2019.05
.067, PubMed: 31150787
Wetzels, R., Matzke, D., Lee, M. D., Rouder, J. N., Iverson, G. J., &
Wagenmakers, E. J. (2011). Statistical evidence in experimental
psychology: An empirical comparison using 855 t tests.
Perspectives on Psychological Science, 6, 291–298. https://doi
.org/10.1177/1745691611406923, PubMed: 26168519
Wetzels, R., & Wagenmakers, E.-J. (2012). A default Bayesian
hypothesis test for correlations and partial correlations.
Psychonomic Bulletin & Review, 19, 1057–1064. https://doi
.org/10.3758/s13423-012-0295-x, PubMed: 22798023
Williams, M. A., Dang, S., & Kanwisher, N. G. (2007). Only some
spatial patterns of fMRI response are read out in task
performance. Nature Neuroscience, 10, 685–686. https://doi
.org/10.1038/nn1900, PubMed: 17486103
Woolgar, A., Afshar, S., Williams, M. A., & Rich, A. N.
(2015). Flexible coding of task rules in frontoparietal
cortex: An adaptive system for flexible cognitive
control. Journal of Cognitive Neuroscience, 27,
1895–1911. https://doi.org/10.1162/jocn_a_00827,
PubMed: 26058604
Woolgar, A., Dermody, N., Afshar, S., Williams, M. A., & Rich, A. N.
(2019). Meaningful patterns of information in the brain
revealed through analysis of errors [preprint]. BioRxiv,
673681. https://doi.org/10.1101/673681
Woolgar, A., Duncan, J., Manes, F., & Fedorenko, E. (2018).
Fluid intelligence is supported by the multiple-demand
system not the language system. Nature Human Behavior,
2, 200–204. https://doi.org/10.1038/s41562-017-0282-3,
PubMed: 31620646
Woolgar, A., Hampshire, A., Thompson, R., & Duncan, J. (2011).
Adaptive coding of task-relevant information in human
frontoparietal cortex. Journal of Neuroscience, 31,
14592–14599. https://doi.org/10.1523/JNEUROSCI.2616-11
.2011, PubMed: 21994375
Woolgar, A., Jackson, J., & Duncan, J. (2016). Coding of visual,
auditory, rule, and response information in the brain: 10
Years of multivoxel pattern analysis. Journal of Cognitive
Neuroscience, 28, 1433–1454. https://doi.org/10.1162/jocn_a
_00981, PubMed: 27315269
Woolgar, A., Parr, A., Cusack, R., Thompson, R., Nimmo-Smith,
I., Torralva, T., et al. (2010). Fluid intelligence loss linked
to restricted regions of damage within frontal and parietal
cortex. Proceedings of the National Academy of Sciences,
U.S.A., 107, 14899–14902. https://doi.org/10.1073/pnas
.1007928107, PubMed: 20679241
Woolgar, A., Williams, M. A., & Rich, A. N. (2015). Attention
enhances multi-voxel representation of novel objects in
frontal, parietal and visual cortices. Neuroimage, 109,
429–437. https://doi.org/10.1016/j.neuroimage.2014.12.083,
PubMed: 25583612
Woolgar, A., & Zopf, R. (2017). Multi-sensory coding in the
multiple-demand regions: Vibrotactile task information is
coded in frontoparietal cortex. Journal of Neurophysiology,
118, 703–716. https://doi.org/10.1152/jn.00559.2016,
PubMed: 28404826
Zellner, A., & Siow, A. (1980). Posterior odds ratios for selected
regression hypotheses. Trabajos de Estadistica y de
Investigacion Operativa, 31, 585–603. https://doi.org/10
.1007/BF02888369
654
Journal of Cognitive Neuroscience
Volume 34, Number 4
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
4
4
6
3
9
1
9
9
6
6
7
8
/
j
o
c
n
_
a
_
0
1
8
1
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3