Kohl, S.H., Melies, P., Uttecht, J., Lührs, M., Bell, L., Mehler, D.M.A., Soekadar, S.R., Viswanathan, S., &
Konrad, K. (2023). Successful Modulation of Temporoparietal Junction Activity and Stimulus-Driven
Attention by fNIRS-based Neurofeedback – a Randomized Controlled Proof-of-Concept Study. Imaging
Neuroscience, Advance Publication. https://doi.org/10.1162/imag_a_00014
Successful Modulation of Temporoparietal Junction Activity and
Stimulus-Driven Attention by fNIRS-based Neurofeedback – a
Randomized Controlled Proof-of-Concept Study
Simon H. Kohl1,2*, Pia Melies2, Johannes Uttecht2, Michael Lührs3,4, Laura Bell2,5, David M. A.
Mehler6,7, Surjo R. Soekadar8, Shivakumar Viswanathan9, Kerstin Konrad1,2
1JARA-Institute Molecular Neuroscience and Neuroimaging (INM-11), Forschungszentrum Jülich, 52425 Jülich,
Germany
2Child Neuropsychology Section, Department of Child and Adolescent Psychiatry, Psychosomatics and Psychotherapy,
Faculty of Medicine, RWTH Aachen University, 52074 Aachen, Germany
3Brain Innovation B.V., Research Department, 6229 EV Maastricht, the Netherlands
4Faculty of Psychology and Neuroscience, Department of Cognitive Neuroscience, Maastricht University, 6200 EV
Maastricht, the Netherlands
5Audiovisual Media Center, Medical Faculty, RWTH Aachen University, 52074 Aachen, Germany
6Department of Psychiatry, Psychotherapy and Psychosomatics, Faculty of Medicine, RWTH Aachen University, 52074
Aachen, Germany
7Institute for Translational Psychiatry, University of Münster, 48149 Münster, Germany
8Clinical Neurotechnology Laboratory, Dept. of Psychiatry and Neurosciences, Charité Campus Mitte (CCM), Charité –
Universitätsmedizin Berlin, Charitéplatz 1, 10117 Berlin, Germany
9Institute of Neuroscience and Medicine – Cognitive Neuroscience (INM-3), Forschungszentrum Jülich, 52425 Jülich,
Germany
*Corresponding author: Simon H. Kohl, Forschungszentrum Jülich, Wilhelm-Johnen-Strasse, 52425 Jülich, Germany;
si.kohl@fz-juelich.de, simon.h.kohl@gmail.com ; ORCID: 0000-0003-0949-6754
© 2023 Massachusetts Institute of Technology. Published under a Creative Commons Attribution 4.0 International
(CC BY 4.0) license. 1
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023
Graphical abstract
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023
2
Abstract
The right temporoparietal junction (rTPJ) is a core hub in neural networks associated with reorienting
of attention and social cognition. However, it remains unknown whether participants can learn to
actively modulate their rTPJ activity via neurofeedback. Here, we explored the feasibility of
functional near-infrared spectroscopy (fNIRS)-based neurofeedback in modulating rTPJ activity and
its effect on rTPJ functions such as reorienting of attention and visual perspective taking. In a
bidirectional regulation control group design, 50 healthy participants were either reinforced to up- or
downregulate rTPJ activation over four days of training.
Both groups showed an increase in rTPJ activity right from the beginning of the trainingbut only the
upregulation group maintained this effect, while the downregulation group showed a decline from the
initial rTPJ activation. This suggests a learning effect in the downregulation exclusively, making it
challenging to draw definitive conclusions about the effectiveness of rTPJ upregulation training.
However, we observed group-specific effects on the behavioral level. We found a significant group x
time interaction effect in the performance of the reorienting of attention task and group-specific
changes, with decreased reaction times (RTs) in the upregulation group and increased RTs in the
downregulation group across all conditions after the neurofeedback training. Those with low baseline
performance showed greater improvements. In the perspective-taking task, however, only time
effects were observed that were non-group-specific.These findings demonstrate that fNIRS-based
neurofeedback is a feasible method to modulate rTPJ functions with preliminary evidence of
neurophysiologically specific effects, thus paving the way for future applications of non-invasive
rTPJ modulation in neuropsychiatric disorders.
Keywords: Neuromodulation, Neurofeedback, Functional Near-Infrared Spectroscopy (fNIRS),
Temporoparietal Junction, Attention, Social Cognition
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023
3
Highlights
the right temporoparietal junction (rTPJ) as a core hub for attentive and socio-cognitive
functions is a promising target for neuromodulatory interventions
first single-blinded, randomized controlled study demonstrates feasibility and
effectiveness of the fNIRS-based neurofeedback training of the rTPJ in healthy adults
subjects are able to regulate the rTPJ with different learning characteristics
first evidence of a neurophysiologically specific effect on stimulus-driven attention
findings have important implications for clinical translation of neurofeedback
interventions targeting the rTPJ
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023
4
1 Introduction
The right temporoparietal junction (TPJ) is considered a central hub of the human brain being
involved in diverse mental functions. Theoretical models stress its involvement in stimulus-driven
attention and social cognition and discuss its essential role in detecting violations of expectations,
contextual updating, mental state shifting, and sense of agency (Corbetta et al., 2008; Decety &
Lamm, 2007; Geng & Vossel, 2013; Krall et al., 2015; van Overwalle, 2009). Due to its diverse
anatomical and functional connections, the TPJ is also considered an important brain region for
communication with neighboring, partially overlapping networks, forming a potential hub where
multiple networks converge and interact (Carter & Huettel, 2013; Mars et al., 2012).
Neuromodulation of such high degree network hubs or control points may result in greater changes in
neural networks and associated behaviors and cognitive functions than neuromodulation of low
degree nodes. Therefore, they are considered hot spots for targeted brain-based interventions
(Murphy & Bassett, 2017).
Furthermore, targeting such high degree hubs using non-invasive neuromodulation, such as
neurofeedback, is interesting from a translational perspective. Testing the causal role of the hub in the
network by neuromodulation followed by observation of its behavioral/functional consequences may
inform therapeutic interventions for brain disorders associated with this hub, e.g., autism spectrum
disorder (ASD), depression and schizophrenia (Kana et al., 2015; Penner et al., 2018). In turn, testing
these potential interventions will increase our understanding of this neural network hub and its role in
the respective disorder.
Previous neuromodulation studies mostly relied on neurostimulation techniques such as
transcranial magnetic stimulation (TMS) or transcranial direct current stimulation (tDCS) to disrupt
or enhance TPJ functions while neurofeedback was utilized to a lesser extent.
TMS studies have demonstrated a decrease in spatial attention performance when disrupting
activation in the right TPJ (rTPJ; Krall et al., 2016; Mengotti et al., 2022). Conversely, a study
conducted by Roy et al. (2015) used tDCS to enhance activation in the right posterior parietal cortex,
which includes parts of the rTPJ, resulting in improved attention re-orienting following stimulation.
Regarding socio-cognitive abilities such as visual perspective taking (vPT) and imitation control, the
evidence for potential enhancement through tDCS is promising but mixed (Santiesteban et al., 2012,
2015; Nobusako et al., 2017; Yang et al., 2020).However, tDCS studies have reported no significant
enhancing effects on other complex socio-cognitive abilities, including theory of mind (ToM;
5
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023
Santiesteban et al., 2015), empathy, emotion recognition, and joint attention (Pereira et al., 2021). In
fact, inhibitory tDCS for ToM and empathy (Mai et al., 2016), as well as inhibitory TMS for ToM
(Krall et al., 2016), have shown disruptive effects.
Together, these studies provide first evidence that neuromodulation of the rTPJ can be used to
improve reorienting of attention and certain facets of socio-cognitive abilities, such as vPT.
Therefore, the rTPJ may also be a promising target for neurofeedback interventions, offering
potentially new treatment options for neuropsychiatric disorders characterized by deficient TPJ
functions such as ASD (Esse Wilson et al., 2018; Salehinejad et al., 2021)
Neurofeedback based on functional near-infrared spectroscopy (fNIRS) is similar to
neurostimulation a causative neuromodulation technique for modulating activation of circumscribed
neocortical brain regions, although likely with less specific and more global effects on brain
networks than neurostimulation. By providing real-time feedback of hemodynamic correlates of
neural activity (e.g., changes in oxyhemoglobin), participants can learn to regulate the brain activity
of specific target regions. In particular, fNIRS-based neurofeedback offers several advantages when
it comes to clinical translation. It is an easy-to-use, non-invasive, and endogenous form of
neuromodulation, which allows long-term learning through the reinforcement of neural activity and
cognitive strategies with therapeutic potential. Moreover, it is safe and well tolerated, and is therefore
associated with fewer ethical concerns than other neuromodulation techniques (Kohl et al., 2020;
Soekadar et al., 2021). Across different studies, preliminary but compelling evidence suggests that
the activation of a neural network, including the TPJ, can be successfully modulated by
neurofeedback based on functional magnetic resonance imaging (fMRI; Harmelech et al., 2015;
Emmert et al., 2016; Direito et al., 2019, 2021; Pamplona et al., 2020). However, behavioral effects
and specificity of findings are less clear, and no study has yet targeted rTPJ activity using fNIRS-
based neurofeedback.
In the current study, we aimed to fill this gap and investigated the feasibility and effectiveness
of fNIRS-based neurofeedback training employing social/monetary reward (Mathiak et al., 2015) to
control rTPJ activity in healthy participants. We conducted a randomized, controlled proof-of-
concept study employing a bidirectional-regulation control group design, which allows for the
detection of neurophysiologically specific effects (Sorger et al., 2019). More specifically, we aimed
to explore three aspects: (1) Can participants learn to increase/decrease the activity of the rTPJ using
fNIRS-based neurofeedback and how is their learning behavior over the course of the training
characterized? (2) Are there any specific behavioral effects in stimulus-driven attention and vPT
6
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023
following the neurofeedback training? (3) What are potential predictors of behavioral improvements?
We hypothesized that healthy adult participants could gain control over activation of the rTPJ with
fNIRS-based neurofeedback and that successful upregulation would be accompanied by improved
performance in a reorienting of attention task and a vPT task. In contrast, we assumed that
downregulation would be associated with either decreased performance or no change in performance.
Based on previous findings on specific traits associated with rTPJ function, e.g., empathy and autistic
traits (Donaldson et al., 2018; Kana et al., 2014; Yang et al., 2020), we tested predictors of behavioral
change on an exploratory level. Due to the scarcity of neurofeedback studies targeting rTPJ
activation, we identified a rather broad set of potential predictors without stating directed hypothesis
for each of them (see methods section).
2 Methods
2.1 Participants
Fifty right-handed healthy participants (age 18-30 years) were recruited via flyer and social media
announcements. Participants were screened during a telephone interview prior to participating in the
study and were excluded if they had a history of psychiatric or neurological diseases, drug or alcohol
abuse, or if they were undergoing current psychopharmacological or psychotherapeutic treatments.
Participants were informed about the study procedure and signed an informed consent document. At
the end of the study, they received a financial compensation of at least 60€ for attending all sessions,
along with an additional monetary reward depending on the success of the neurofeedback training.
The study protocol was approved by the local ethics committee (EK 148/18) and conducted in
accordance with the Declaration of Helsinki (World Medical Association, 2013), with the exception
that it was not pre-registered on a publicly accessible database.
The participants were randomly allocated to the study groups, which were balanced out in terms of
gender and the order of task assessments. Twenty subjects were allocated to the downregulation
group and ten more (30 participants) to the upregulation group in order to provide higher statistical
power for later subgroup analyses in this group.
After the first eleven participants, we noticed an error in the online preprocessing script
(motion correction algorithm), which led to small deviations of the feedback displayed during the
neurofeedback training. We simulated the feedback signal of these participants using the corrected
script and calculated the accordance with the original feedback signal. Five participants (3 in the
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023
7
upregulation group, 2 in the downregulation group) showed an accordance below 90% and were
therefore excluded from further analysis.
No a priori power analysis was conducted. However, according to a sensitivity analysis, a
mixed analysis of variance (ANOVA) including 45 participants was sufficiently powered (80%) to
detect a group x time interaction effect of at least f = 0.43 (assuming no violation of sphericity and a
correlation among repeated measure of 0.8) or 0.77 for an independent t-test.
2.2 Study design
We applied a single-blinded, randomized controlled between-subject design. Participants were
blinded to group assignments, but experimenters were not. We followed the recently published best
practices for fNIRS publications (Yücel et al., 2021) and the consensus on the reporting and
experimental design of clinical and cognitive-behavioral neurofeedback studies (CRED-nf checklist
(Ros et al., 2020;see Supplementary Material 2)).
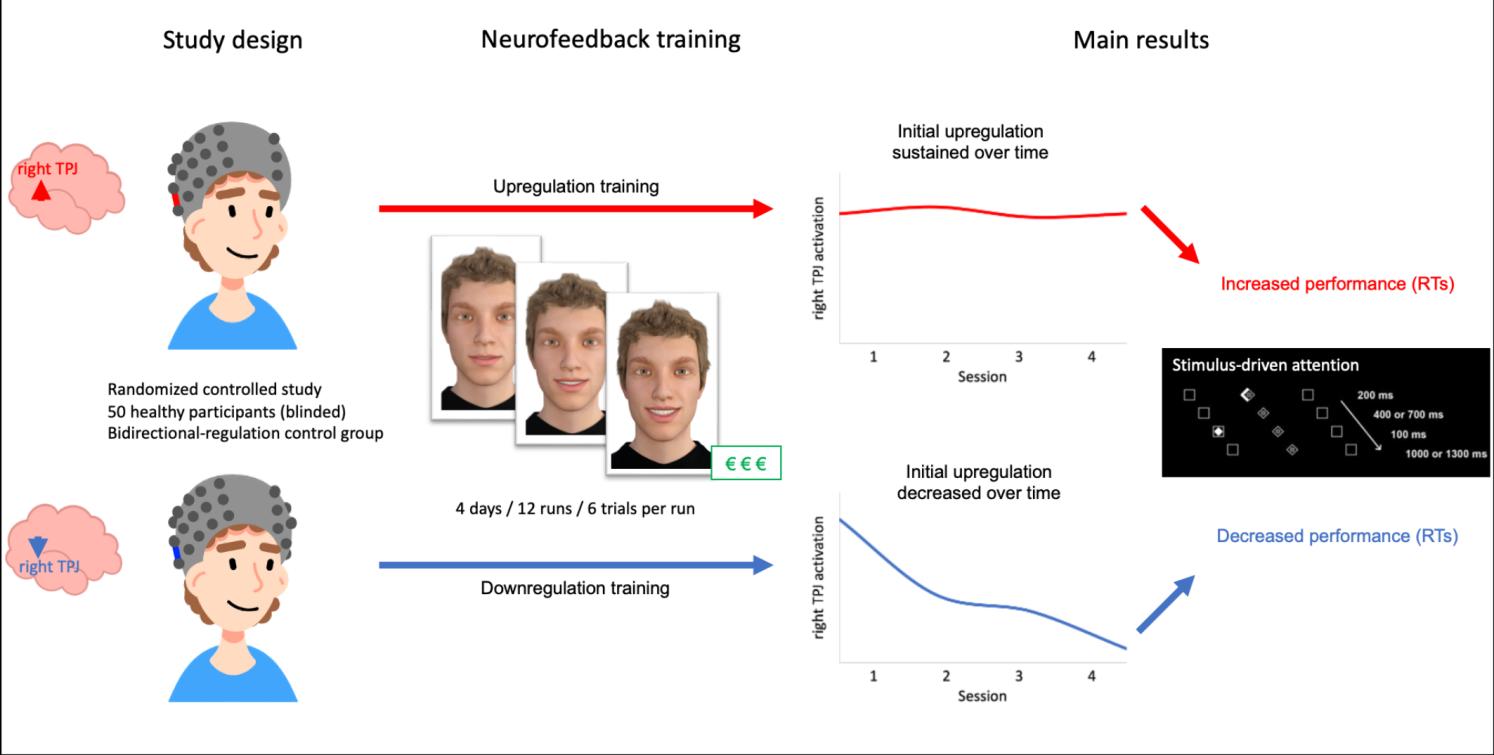
Figure 1. Study design
Procedure
All participants took part in four appointments, including a pre- and post-assessment session with one
additional short neurofeedback training session (day 1 and day 4) as well as two longer
neurofeedback training sessions (day 2 and day 3; see Figure 1). The four appointments were
scheduled within one week (M = 6.71 ± 2.23 days) and the pre- and post-assessment sessions at the
same time of day. The procedure on each day was as follows:
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023
8
Day 1: To evaluate self-efficacy as a potential mechanism of neurofeedback effects and address
group differences, participants completed the German version of the general self-efficacy scale
(Schwarzer and Jerusalem, 1995). Additionally, a questionnaire was administered to assess the
participants’ expectations and motivation towards the neurofeedback training, offering further
controls for non-specific psychological mechanisms. After a short practice session, the pre-
assessment of the (1) reorienting of attention task and (2) perspective-taking task took place. The
order of these two tasks was counterbalanced across participants within both groups. Participants
subsequently received specific instructions about the neurofeedback training and underwent two runs
of neurofeedback training.
Days 2 and 3: On days 2 and 3, participants underwent neurofeedback sessions with four runs each.
To assess pre-post changes in mood states and resting state brain activity, we assessed the German
short version of the Profile of Mood States (McNair et al., 1981) and recorded a 10 min resting-state
fNIRS measurement (to be reported elsewhere) before the training started on day 2 and after the
training was completed on day 3. Between days 2 and 3, participants completed standardized
questionnaires to account for variations in socio-cognitive traits among groups and predict
neurofeedback effects. These traits, including autistic traits, empathy, cognitive styles, as well as
ADHD symptoms have the potential to impact rTPJ functioning (Barman et al., 2015; Donaldson et
al., 2018; Kana et al., 2014). All traits were assessed dimensionally using the German version of the
Social Responsiveness Scale (Bölte, 2012), the Adult ADHD Self-Report Scale v1.1 (Kessler et al.,
2005), the German version of the Interpersonal Reactivity Index (IRI;Davis, 1983), the autism-
spectrum quotient (Baron-Cohen et al., 2001), the systemizing quotient (Baron-Cohen et al., 2003),
and the empathy quotient (Baron-Cohen & Wheelwright, 2004).
Day 4: Participants underwent a short neurofeedback training session of two runs, followed by the
post-assessment of the reorienting of attention and perspective-taking task. At the end of the session,
participants filled in the general self-efficacy scale again as well as a debriefing questionnaire to
further assess feasibility and unspecific mechanisms. This questionnaire included items assessing
participants’ evaluation of the neurofeedback training, for example “I believe the training helped to
improve my attention”, “I enjoyed the training”, “The experimenter was trustworthy”, etc.
Furthermore, they were asked to guess the group condition they had been randomly assigned to.
The fNIRS system was set up at the beginning of each day, except for day 1, which began
with comprehensive instructions and practice runs. For all tasks, stimuli were presented on a 24-inch
LCD screen (1920 x 1080 pixels) using the Psychtoolbox on Matlab 2017a (The Mathworks Inc,
9
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023
Natick, MA) and being run on a Windows PC. Participants viewed the screen at a distance of
approximately 50cm. Responses were acquired using a standard keyboard.
Neurofeedback training
Participants were blinded to their group assignment and were told that, depending on their group
assignments, the goal of the training was to increase or decrease activation of a specific brain region.
Irrespective of group assignment, participants in both groups received the same instructions.
All participants received standardized information and instructions about the neurofeedback
training (see Supplementary Material 1) based on Greer et al. (2014). They were instructed not to use
any respiratory or motor strategies but to remain still, breathe regularly, and only rely on mental
strategies to regulate their brain activity. The training took place on all four days and comprised 12
runs in total: two runs on days 1 and 4 (~12 minute/day), and four runs on days 2 and 3 (~25
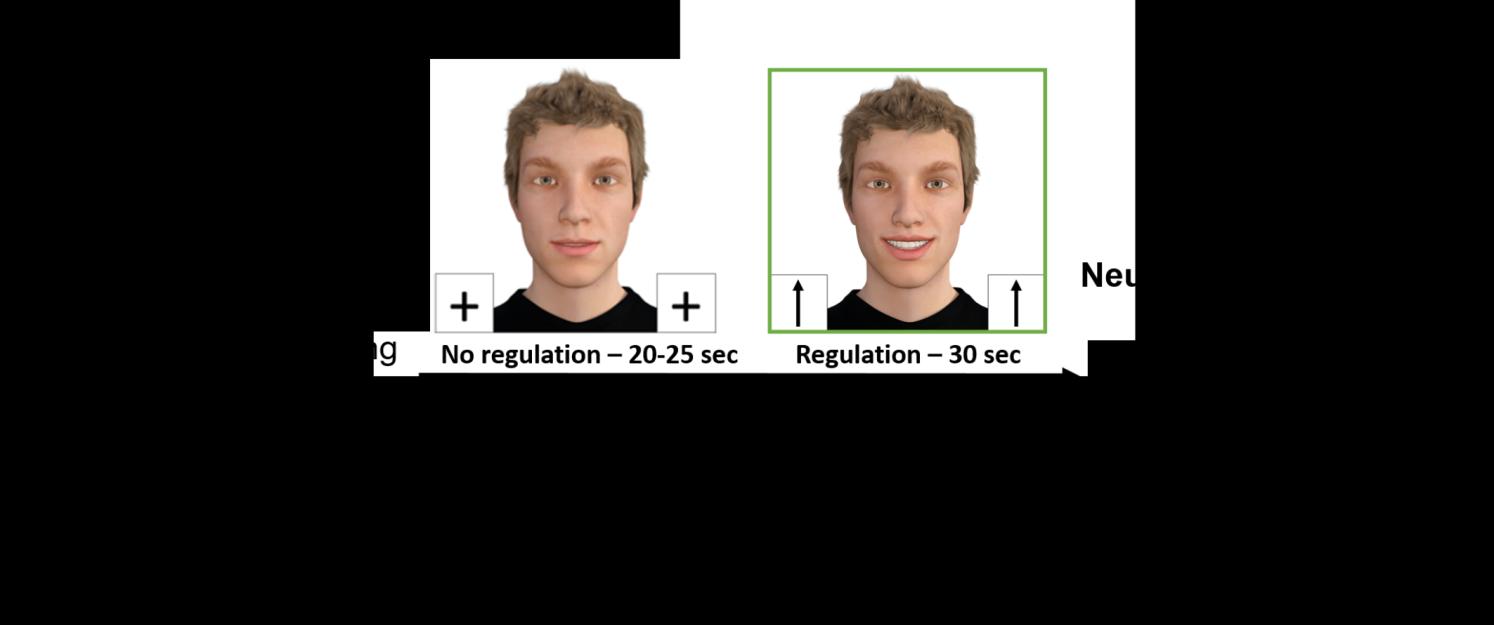
minutes/day). Each neurofeedback run consisted of six blocks. Each block started with a 25s/30s no-
regulation condition followed by a 30s regulation condition, and the block ended with a 2s reward
presentation (see Figure 1). We varied the durations of the no-regulation condition to avoid
synchronization with physiological confounds, such as breathing patterns and Mayer waves, during
the task and to increase design efficiency (Kinoshita et al., 2016; Yücel et al., 2021). On each block,
the face of a human avatar was continuously displayed on the screen. We used DAZ Studio 4.9 (DAZ
Productions, Inc., USA) to create a modified version of the stimuli validated by Hartz et al. (2021).
Eleven pictures of the avatar with different levels of smiling were created for the visualization of the
feedback signal.
During the no-regulation condition, participants were instructed to passively look at the
avatar, which maintained a neutral facial expression. During the regulation condition, real-time
feedback of rTPJ activity was presented visually on a screen using a smiling avatar (social reward).
Participants were instructed to regulate and make the avatar smile, which was modulated in real time
by their rTPJ activation.
To foster motivation, participants received a monetary reward for successful regulation. In
each regulation trial, the participant received 0.01€ per second exceeding a certain individual
threshold (see real-time fNIRS data processing (online analysis)). Whenever participants exceeded
this reward threshold, a green frame appeared around the feedback display, indicating that their
regulation was earning an incentive. The total amount earned on each trial was presented on the
screen at the end of the trial. This threshold was adapted according to individual regulation
performance (see 0. for a detailed description).
10
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023
In neurofeedback training, providing explicit mental strategies is not necessary but initially
seems to facilitate learning (Scharnowski et al., 2015). Therefore, we provided some example
strategies that could be helpful to regulate rTPJ activity (e.g., strategies related to ToM, empathy,
thinking, imagination of positive events, counting, etc.; see Supplementary Material 1). However,
participants were encouraged to find their own individual successful strategy by trial and error. After
each neurofeedback run, we asked participants to verbally report which strategies they used and how
successful they rated this strategy (Likert scale ranging from 1 to 5). After each session, we also
assessed participants’ motivation to continue participating in the training and their beliefs about
being able to control their brain activity.
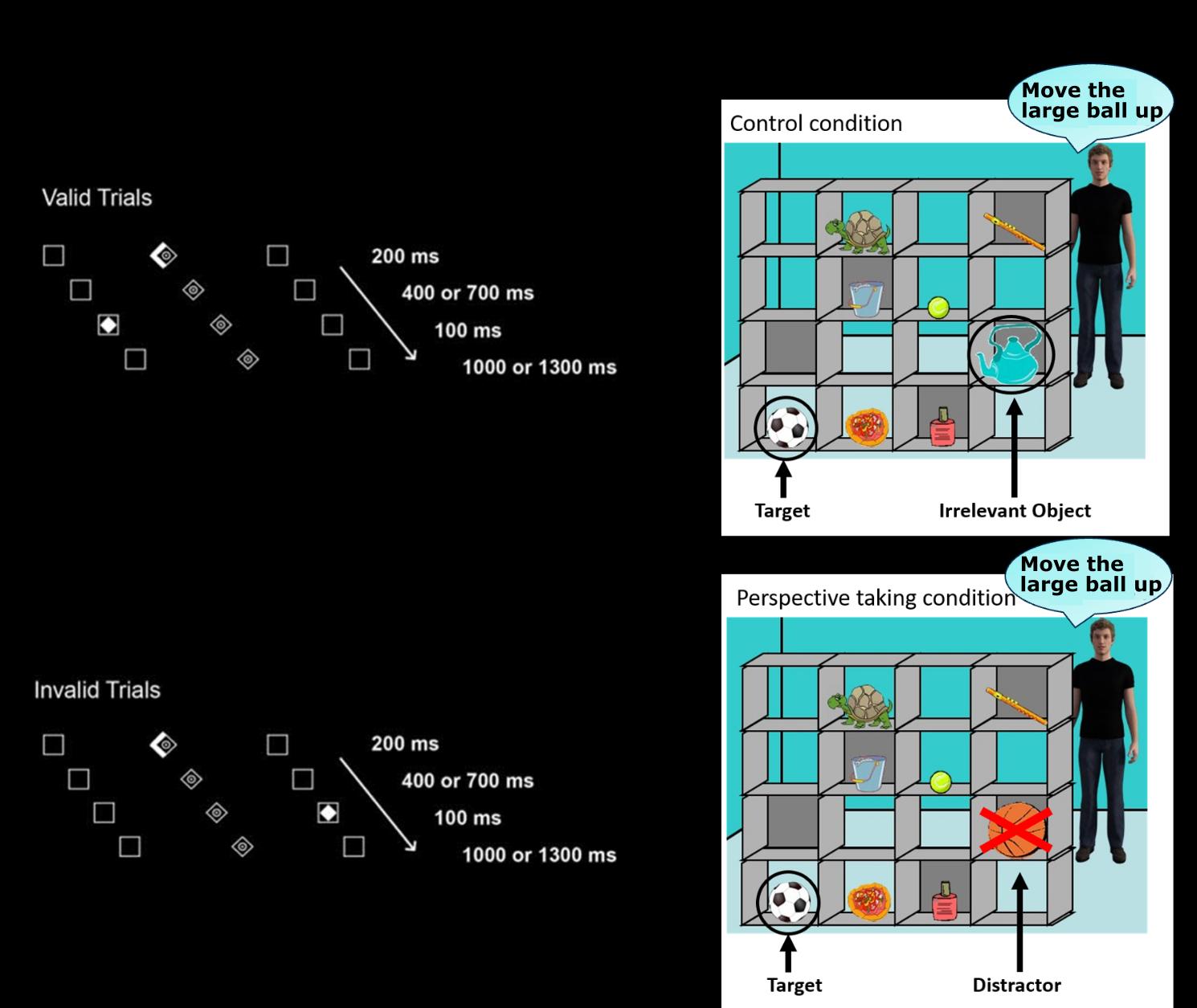
Reorienting of attention task
Reorienting of attention is defined as the capacity to alter the focus of attention to unexpected,
external stimuli while expecting another task/situation. We assessed the reorienting of attention using
a modified version of the Posner paradigm (Krall et al., 2016; Posner, 1980; Vossel et al., 2009). In
this task (see Figure 2), a central diamond (fixation point) was displayed between two horizontally
arranged boxes. For each trial, a central cue was presented for 200ms indicating whether a target
would appear on the right or the left side of the screen (brightening of the diamond to the right or left,
respectively). After a variable cue-target interval of 400ms or 700ms, the target (white diamond)
appeared for 100ms with a certain probability at the cued (valid cueing) or at the non-cued location
(invalid cueing) and the participant had to indicate on which side it appeared by pressing a button
using his/her right hand. The target-cue stimulus onset asynchrony was either 1000ms or 1300ms. All
stimuli were presented on a black background. Since fNIRS was assessed during the task, the trials
were presented in a blocked design. The task consisted of a total of twelve blocks, with six invalid
blocks and six valid blocks. Each invalid block comprised twelve valid and eight invalid trials and
each valid block comprised 20 valid trials only. Hence, the overall distribution of valid trials (192 of
240) and invalid trials (48 of 240) was 80% vs. 20%. The blocks were presented in a randomized
order to mitigate anticipatory effects. Participants were told that the cue was not always informative,
but they were not informed about the different blocks beforehand. The task blocks with a 40s
duration were separated by 20s or 25s rest periods in which the same visual stimuli, but no cues or
targets, were presented.
Visual perspective-taking (vPT) task
vPT refers to the ability to infer spatial relationships between objects from different viewing angles.
We assessed vPT with the widely used Director paradigm according to Dumontheil et al. (2010) and
11
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023
Symeonidou et al. (2016), as this task has been successfully used to assess the effects of tDCS
stimulation of the TPJ (Santiesteban et al., 2012, 2015). In this task, participants saw a visual scene
with a 4 x 4 set of shelves containing eight different objects (see Figure 2) and were instructed to take
the perspective of a “director” standing behind the shelves and giving them auditory instructions to
move certain objects on the shelves by clicking a mouse on the respective target object. Importantly,
some of the objects were occluded from the view of the director, which participants had to take into
account in order to respond correctly in the perspective taking (PT) condition. This can be seen in
Figure 2 where the “director” refers to the football instead of the large basketball (distractor), which
is occluded from his view. In the control condition (non-perspective-taking (NPT) condition), the
distractor is replaced by an irrelevant object. For a more detailed description of this task, see
Dumontheil et al. (2010) and Symeonidou et al. (2016). Each block consisted of four trials. The PT
and NPT blocks were presented in a pseudo-randomized order in such a way that no more than two
blocks of the same condition were presented consecutively. The task blocks (24s) were separated by
a rest period with a duration of 20s or 25s. RTs were recorded from the onset of the auditory
instruction to the participant’s mouse click response.
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023
12
Figure 2. Illustration of experimental tasks used for pre-post measurements. The reorienting of
attention task (left; adopted from Krall et al., 2016) and the visual perspective-taking task (right;
adopted from Symeonidou et al., 2016) were used to measure the effects of the neurofeedback
training. [Note that in the perspective-taking task (right), the speech bubbles are only shown for
illustration. Auditory instructions were provided to the participants by the director (see text).]
2.3 fNIRS acquisition
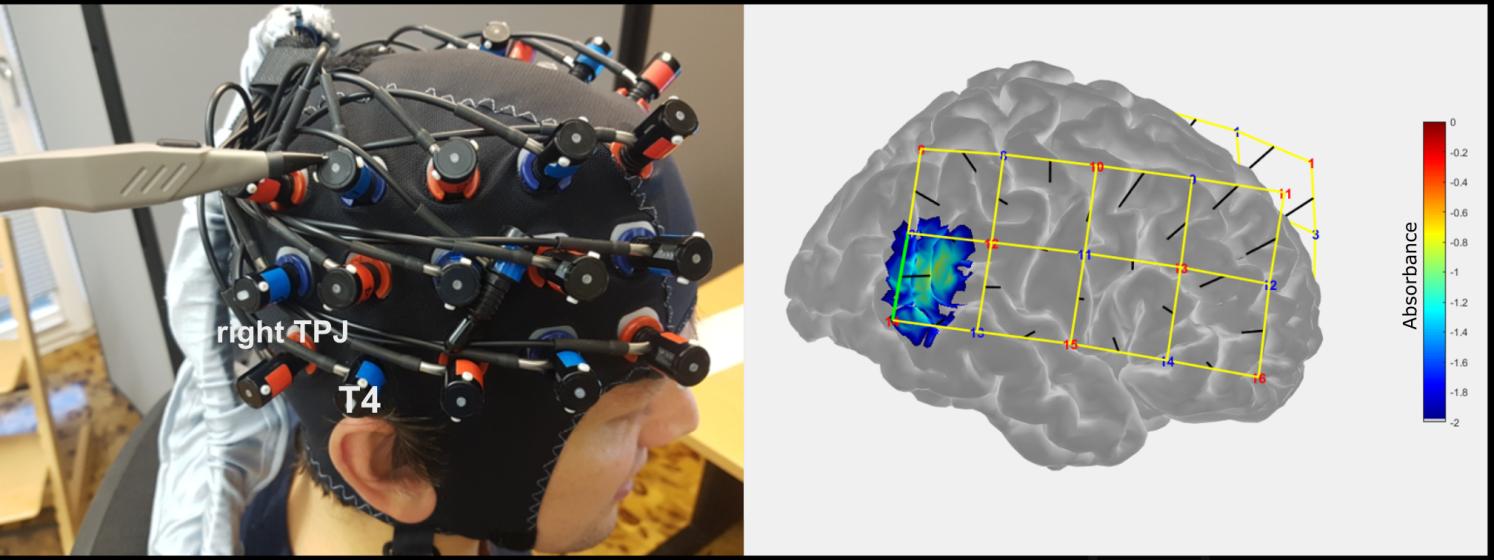
We used the ETG-4000 continuous wave system (Hitachi Medical Corporation, Tokyo, Japan) to
measure changes in oxy-(HbO) and deoxyhemoglobin (HbR) concentrations at a rate of 10Hz with
two wavelengths (695nm and 830nm). Two 3 × 5 probe sets (2 × 22 measurement channels) were
placed bitemporally on the participant’s head to cover temporal and frontal brain regions and were
attached using electroencephalography (EEG) caps (Easycap GmbH, Herrsching, Germany). The
interoptode distance was 3cm. The probe sets were placed on the participants’ heads in such a way
that the second most posterior optode of the lowest row was placed over T3/T4 of the EEG 10-20
13
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023
system (Jasper, 1958) and the most anterior optode of the lowest row was placed approximately over
the eyebrow (see Figure 3). If necessary, hair was moved away from optode holders in order to
increase the quality of the signal. Furthermore, we instructed participants each day to stay relaxed,
breathe regularly, and keep the movement of their heads to a minimum.
To select the best channel for the feedback processing that covers the rTPJ, prior to the
current study, we conducted digitizer measurements using a Patriot 3D Digitizer (Polhemus,
Colchester, Vermont) in a separate sample of five pilot participants wearing an fNIRS optode
arrangement available from a previous study. In all five subjects, the same channel corresponded to
anterior parts of the rTPJ (see Figure 3). To confirm the anatomical specificity of this channel, we
also conducted digitizer measurements in all participants of the current study after each experimental
session. Anatomical locations of the optodes in relation to standard head landmarks (nasion, inion,
Cz, and preauricular points) were assessed. Cortical sensitivities of all channels were estimated
through Monte Carlo photon migration simulations (1,000,000 photons) using AtlasViewer
implemented in Homer v2.8 (Aasted et al., 2015; Huppert et al., 2009). Montreal Neurological
Institute (MNI) coordinates for each subject and session were extracted and averaged for each
participant. In total, 5% of data (i.e., 10 of the 200 samples obtained from 50 participants and 4
sessions) were excluded from this analysis due to errors during the digitizer measurements, which
resulted in implausible estimations of MNI coordinates. The average MNI coordinate of the feedback
channel (x = 56 ± 6.4, y = -49 ± 4.6, z = 18 ± 6.9) corresponded to anterior parts of the rTPJ,
previously reported in a meta-analysis for reorienting of attention and theory of mind contrasts (Krall
et al., 2015).
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023
14
Figure 3. fNIRS optode arrangement and sensitivity profile for the feedback channel. The
feedback channel corresponds to anterior parts of the rTPJ (MNI: x = 56 ± 6.4, y = -49 ± 4.6, z = 18
± 6.9).
2.4 Real-time fNIRS data processing (online analysis)
Participants received feedback in real time about the instantaneous HbO activity at one single
channel placed over the rTPJ (see section on fNIRS acquisition). The procedure to convert the HbO
activity into feedback (updated every 100ms) involved several steps as described below (see Figure
4).
Figure 4. Real-time fNIRS data processing. Correlation-based signal improvement (CBSI)
algorithm (Cui et al., 2010).
The raw signal was first preprocessed by the ETG-4000 using a high-pass filter of 0.01Hz and
a low-pass filter of 1Hz and a moving average of 5s. This preprocessed HbO signal was then sent in
real time to an external computer where it was further processed using a customized Matlab script.
15
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023
Motion artifacts in the signal were then removed using the correlation-based signal
improvement (CBSI) algorithm (Cui et al., 2010). This algorithm calculates the corrected signal as a
linear combination of HbO and HbR scaled by their standard deviations, based on the assumption
that HbO and HbR are highly negatively correlated. Furthermore, the algorithm assumes that the
signal has been offset-corrected to have zero mean.
Using the CBSI algorithm, the corrected HbO signal at time t (denoted as Xcorr (t)) was
obtained using the following expression:
where X(t) and Y(t) are the measured values of the HbO and HbR values, respectively, at time t (after
offset correction), and α is the ratio of the noise amplitude in the HbO and HbR signals. To estimate
the noise amplitude ratio α and perform the offset correction, we used the HbO and HbR signals from
the last 30s of the no-regulation period (i.e. from the period [–30s, 0s] relative to the start of the
regulation period at 0s) as follows:
This preprocessed and motion-corrected signal Xcorr (t) was then normalized relative to the
HbO signal from the last five seconds of the no-regulation period (baseline) using the following
formula:
The feedback signal was further smoothed using linear interpolation over 1s. The final step
was to convert the feedback value into visual feedback. This was implemented by mapping the
feedback value onto a scale that ranged from 0 to 10, based on the following expression:
To receive positive feedback on downregulation, this value was multiplied by –1 for the
downregulation group. In this expression, the maximum feedback value (level 10) was defined as a
percentage (k) of a threshold value (T) that was determined for each participant based on their rTPJ
activation during the reorienting of attention and perspective-taking tasks at pre-assessment. Similar
to the calculation of the feedback signal during the neurofeedback task, the rTPJ activation during
valid/invalid and perspective-taking/non-perspective-taking blocks was estimated and averaged over
16
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023
blocks. The mean between the contrasts of invalid vs valid (reorienting of attention task) and
perspective taking vs non-perspective taking (vPT task) was calculated using the following formula:
If this value was negative, only the positive contrast was used. If both contrasts were
negative, the initial threshold was set to the default value T = 1. To scale the feedback signal, we
defined level 10 of the feedback value as k = 0.25 (i.e., 25% of the threshold value T). The feedback
value at each time was used to update the visual display in two ways. Each level of this scale from 0
to 10 was associated with eleven images of the avatar smiling to different extents with 10 being the
largest smile. Therefore, the feedback value was used to update the image of the avatar displayed on
the screen. Additionally, whenever the participant exceeded a feedback level of 5, a green frame
appeared around the feedback display indicating a monetary reward (1 cent/sec).
To maintain the difficulty of the task across runs, the value of k was incremented by 0.25 for
the next run if the participant exceeded level 5 for 75% of the time on each run.
2.5 Data processing and analysis
Statistical methods and software
The additional fNIRS offline analyses were carried out using Homer v2.8 (Huppert et al.,
2009)(Huppert et al., 2009) and in-house Matlab scripts (Matlab 2018b; The Mathworks Inc, Natick,
MA). Statistical analyses were performed using R (R Core Team, 2021). To assess the effects of the
neurofeedback training, we calculated linear mixed models using the R packages lme4 (Bates et al.,
2015) and lmerTest (Kuznetsova et al., 2017). The models were fitted using REML. In the case of
non-normal residuals, robust nonparametric analysis of longitudinal data in factorial designs was
carried out using the nparLD package (Noguchi et al., 2012), and ANOVA-type statistics (ATS) were
reported. In addition, we calculated paired and Welch’s unequal variances t-test and Mann-
Whitney/Wilcoxon tests for comparisons of mean values. Spearman’s rank correlation was used to
assess relationships between neurofeedback regulation success, behavioral effects, and psychosocial
factors. To explore predictors of behavioral improvements, we calculated stepwise multiple
regression models and applied an Akaike information criterion (AIC) stepwise model selection
algorithm (Akaike, 1974) to select the best models. Data are presented as means ± standard deviation
(SD) unless indicated otherwise. For all analyses, a p-value below 0.05 was considered significant.
Bonferroni correction was applied for the correlational analyses. We calculated Cohen’s d for mean
comparisons or the correlation coefficient after non-parametric tests and partial eta-squared (p²) for
linear mixed models.
17
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023
Neurofeedback regulation success
Further preprocessing
To analyze neurofeedback regulation success, we analyzed the time series of the feedback signal
based on the online analysis adding further steps for artifact removal. First, we detected and
automatically removed noisy channels by calculating coefficients of variation (CoV) and excluding
channels with a CoV > 10% in HbO or HbR or channels with a variation difference between the
chromophores of over 5%. In addition, channels in which we identified a flat line of at least 1s were
removed (Bell et al., 2020). If the channel covering the rTPJ (COI) was detected as a noisy channel,
we visually inspected the raw and preprocessed time series of the respective channel, and reincluded
the channel if the high CoV was driven by spikes or drifts that could be removed by our
preprocessing pipeline. The removed values were replaced by the average activation of the six
neighboring trials. Second, outliers were removed if they exceeded 3SDs from the mean on-trial level
and replaced by the last observation.
Additional robustness checks
Since short channel measurements were unfortunately not available for our system, we carried out an
additional stepwise offline analysis approach to further test the robustness of the observed effects and
to rule out the possibility that the neurofeedback signal change was driven by systemic physiological
signals. For the offline robustness checks we used raw fNIRS signals of the same data sets as for the
online analysis and carried out the same preprocessing and analysis steps as in the online analysis
(bad channel removal, outlier detection, interpolation, bandpass filter, 5s moving average filter, and
CBSI). In addition, we applied a more stringent bandpass filter (0.01-0.09 Hz), which is
recommended by Pinti et al. (2019) and should remove most of the systemic physiological signals
(first robustness check). In the second, more conservative robustness check, we applied the common
average reference (CAR) using the average time series of the 22 channels placed over the left
hemisphere and subtracting it from the feedback channel time series. The CAR is considered to be a
viable approach when short channel measurements are not available (Yücel et al., 2021), albeit a
suboptimal one, since there is a risk of overcorrecting the signal or inducing additional effects
depending on network activity during the task (Hudak et al., 2018; Klein et al., 2022; Kohl et al.,
2020).
For all three analysis approaches, we calculated the median and standard deviation of the
feedback signal time series for each of the 30s trials, normalized to the last five seconds of the no-
18
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023
regulation period (see formula for the calculation of the feedback signal). Mean values were
calculated for each neurofeedback run and used for further group-level analyses.
Neurofeedback success measures
The main goal of the study was to test for successful control of rTPJ activation. However, there is no
consensus on how to define neurofeedback regulation success (Kohl et al., 2020; Paret et al., 2019),
and there is evidence of insufficient reporting quality in the field of fNIRS-based neurofeedback
(Kohl et al., 2020), making it difficult to assess the effectiveness of a newly developed
neurofeedback training protocol. Therefore, here we report several different neurofeedback success
measures on the group and on the individual level, each of which has implications for the conclusion
of the successful control of rTPJ activation (see Table 1). Besides measure of signal amplitude, we
also include measures of signal variability in our analysis as they might be indicative of learning as
well (see Kohl et al., 2020).
First, we tested whether a participant is able to (1) activate the target region in the desired
direction and maintain it over the course of the training compared to a within-baseline condition
(neurofeedback performance as compared to baseline – maintenance of activation). On the group
level, we used one-sample t-tests to test for a regulation effect against baseline for both groups
separately. For the analysis on the individual level, we followed an exploratory approach according
to Haugg et al. (2021) and Auer et al. (2015) to classify successful participants who maintained up-
or downregulation throughout the training, independent of a learning effect. A run was classified as
successful if the medians of the feedback signal over trials were positive in the upregulation group
and negative in the downregulation group. Participants who demonstrated more than 50% successful
neurofeedback runs were then classified as “successful”, and participants below 50% as
“unsuccessful”. The numbers of successful participants are reported. In addition, we report on the
numbers of successful runs per participant (see Table S1 and S2).
These measures only provide necessary evidence for the control of baseline brain activation,
but they alone are insufficient to draw conclusions about the successful regulation of brain activation
through neurofeedback. This limitation arises because the activation of the target region could be
influenced by factors inherent to the experimental paradigm, such as stimulation from the
experimental stimuli, or the use of mental strategies, rather than solely attributable to the effects of
neurofeedback.
Stronger evidence for control would be if a participant showed a voluntary change in (2)
amplitudes and (3) variability of the feedback signal over time compared to a within-baseline
19
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023
condition (neurofeedback improvement or learning). On the group level, we tested for a time effect of
rTPJ regulation using linear mixed models or non-parametric ANOVAs for both groups separately.
On the individual level, the neurofeedback improvement of each participant was calculated based on
the slope of the linear regression over all neurofeedback runs. Here, a participant was classified as
“successful” if he or she showed a slope larger than 0 in the upregulation group and smaller than 0 in
the downregulation group. In addition, since learning does not necessarily follow a linear trajectory,
we compared the regulation success of the last session with the first session. On the group level, we
used the paired t-test for both groups separately to compare rTPJ activation in the last session
compared to the first session. On the individual level, we classified a participant with a
positive/negative value as “successful” and vice versa.
Lastly, we tested for a specific effect of regulation (specific evidence for control) by
comparing measures 1-3 with the between-group control condition. To do so, we calculated linear
mixed models or non-parametric ANOVAs and tested for a significant group effect (1) and a
significant group × time interaction (2-3).
Table 1. Neurofeedback success measures
Necessary evidence for control:
Neurofeedback performance as
compared to baseline
Stronger evidence for control:
Neurofeedback improvement or
learning
Individual level
>50% successful runs
(positive/negative median)
Group level
(de)activation over all runs (t-test)
Difference between last and first
session or slope ><0
change from first to last session (t-
test), slope (mixed model)
Specific evidence for control:
Significant group effects
N>1000 ms, as
well as incorrect key presses were excluded from the analysis. Harmonic means of valid and invalid
trials of the invalid blocks were calculated and analyzed. The harmonic mean, as recommended for
RT analysis by Ratcliff (1993),is a more unbiased estimator of the central tendency of RTs than the
arithmetic mean, which also reduces the effects of outliers while remaining high power. In addition,
RTs for invalid trials were subtracted from valid trials to estimate the costs of shifting attention from
the cued position to a non-cued target (reorienting effect). Two participants, one from each group,
had to be excluded due to a technical error or not understanding task instructions. For the vPT task,
20
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023
harmonic means of the RTs and mean accuracies were analyzed. One participant had to be excluded
from this analysis due to a technical error. For both tasks, linear mixed models or non-parametric
ANOVAs were calculated with the task condition and measurement time as within-subject factors
and the group as the between-subject factor. According to our hypotheses, we expected to see a
significant group × time interaction as well as a significant within-group time effect in the
upregulation group for both tasks.
To further confirm the specificity of the behavioral effects and control for unspecific
contributions of psychosocial factors, we calculated four sets of correlational analyses:
1-2) Observed behavioral effects in the attention task/vPT task were correlated with three different
neurofeedback success measures for both groups separately as well as across group resulting in 27
tests (3 conditions x 3 success measures x 3 groups) for each task.
3-4) Changes in RTs of the attention task/changes in accuracies of the vPT task across conditions
were correlated with eleven results of questionnaires assessing psychosocial factors (e.g.,
expectations toward the training, subjective evaluation of the training, etc.) resulting in 33 tests (11
questionnaire results x 3 groups) for each task.
We applied the Bonferroni correction separately for the four different sets of analyses, each
involving a distinct number of tests (i.e., 27, 27, 33, and 33).
Predicting behavioral improvements
As neurofeedback represents a potentially useful tool for application in clinical populations exploring
how subclinical symptoms, personality traits, and baseline task performance are related to specific
behavioral neurofeedback, effects in healthy samples can inform clinical translation. In terms of TPJ
functioning, these include ASD symptoms (Kana et al., 2014, 2015, 2016) and measures of empathy
as well as baseline cognitive and socio-cognitive performance data.
We only found a specific effect in the reorienting of attention task and no specific effect in the vPT
task. Therefore, we conducted an analysis for the effects in the reorienting of attention task using
absolute RTs across conditions as a dependent variable of a multiple regression model. We used the
results of questionnaires assessing autism-related traits and empathy (AQ, EQ, SQ, SRS, IRI) as well
as baseline task performance (RTs across conditions of the reorienting of attention task and
accuracies in PT trials of the vPT task) as predictor variables (7 in total). To avoid overfitting,
stepwise multiple regression models were calculated and the AIC stepwise model selection algorithm
(Akaike, 1974) was used to select the best model.
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023
21
Mental strategies underlying neurofeedback regulation
Based on a content analysis of the strategy reports, we identified 20 different categories of strategies
that participants employed to regulate their brain activity. Figure 8 in the results section shows the
different categories and their distribution. We classified the reported strategies into the different
categories, calculated how many strategies were used by each subject and how many participants
reported to have used a particular strategy. The mean number of strategies used, and the frequencies
of the different strategies were compared between groups.
3 Results
3.1 Baseline characteristics
There were no baseline differences between the two groups, i.e., neither in the questionnaire data nor
in the reaction times and accuracies in the (1) reorienting of attention task and (2) vPT task (all p >
0.05; see Table 2, Figure 6, and Table S1-2 for more detailed baseline characteristics and
questionnaire results). In addition, the thresholds for the feedback signal as determined by rTPJ
activation during the pre-assessments did not significantly differ between the groups (upregulation
group = 2.19 ± 1.45, range 0.45 – 6.2, downregulation group = 2.76 ± 1.84, Range 0.03 – 6.92).
These results demonstrate that our randomization procedure was successful. Seven participants (5 in
the upregulation and 2 in the downregulation group) showed negative contrasts for both tasks.
Therefore, their initial threshold was set to a default value of 1.
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023
22
Upregulation
Downregulation
(M ± SD)
(M ± SD)
27 (13 female)
18 (9 female)
p-value
N
Age (years)
pre RTs attention task – invalid
pre RTs attention task – valid
24.22 ± 3.03
497 ± 69 ms
452 ± 61 ms
pre accuracies attention task – invalid
0.98 ± 0.02
pre accuracies attention task – valid
0.99 ± 0.02
24.22 ± 2.71
508 ± 90 ms
468 ± 94 ms
0.98 ± 0.05
0.99 ± 0.03
pre RTs vPT task – PT
3667 ± 327 ms
3630 ± 302 ms
pre RTs vPT task – NPT
3611 ± 286 ms
3620 ± 272 ms
pre accuracies vPT task – PT
pre accuracies vPT task – NPT
pre rTPJ thresholds
AQ total
EQ total
SQ total
IRI total1
SRS: total
POMS: depression/anxiety
POMS: vigor
POMS: fatigue
POMS: hostility
0.946 ± 0.071
0.981 ± 0.035
2.19 ± 1.45
15.30 ± 6.14
45.19 ± 9.76
0.926 ± 0.091
0.971 ± 0.044
2.76 ± 1.84
13.94 ± 4.49
45.17 ± 8.28
29.96 ± 10.35
32.11 ± 13.75
56.78 ± 11.78
39.26 ± 19.36
0.40 ± 0.52
3.43 ± 1.01
1.61 ± 0.99
0.54 ± 0.93
52.39 ± 8.83
37.94 ± 12.94
0.42 ± 0.58
3.40 ± 0.90
1.53 ± 1.12
0.52 ± 0.77
General self-efficacy
30.59 ± 2.50
31.83 ± 3.57
Expectations
Motivation
2.42 ± 0.56
3.70 ± 0.40
2.67 ± 0.59
3.63 ± 0.50
Table 2. Baseline characteristics and questionnaire results
0.935
0.816
0.703
0.169
0.112
0.701
0.966
0.634
0.230
0.270
0.399
0.995
0.577
0.161
0.862
0.716
0.920
0.814
0.737
0.211
0.123
0.695
1 according to Cliffordson (2001) and Paulus (2012)Cliffordson (2001) and Paulus (2012); AQ, Autism Spectrum Quotient;
ASRS, Adult ADHD Self-Report Scale; EQ, Empathy Quotient; IRI, Interpersonal Reactivity Index; POMS, Profile of Mood
States; PT, perspective taking; NPT, non-perspective taking; SQ, Systemizing Quotient; SRS, Social Responsiveness Scale;
vPT, visual perspective taking.
3.2 Regulation behavior and rewards
Both groups were able to regulate the signal in the desired direction and remained above their
individual threshold. On average, the upregulation group remained above the threshold in each trial
for a longer period (M = 16±3.25 of 30s) than the downregulation group (M = 9.41±2.9 of 30s), but
23
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023
only the downregulation group improved over time (see Supplementary Material 3.1 and Figure S1
for detailed results). As a result, the upregulation group also received significantly more monetary
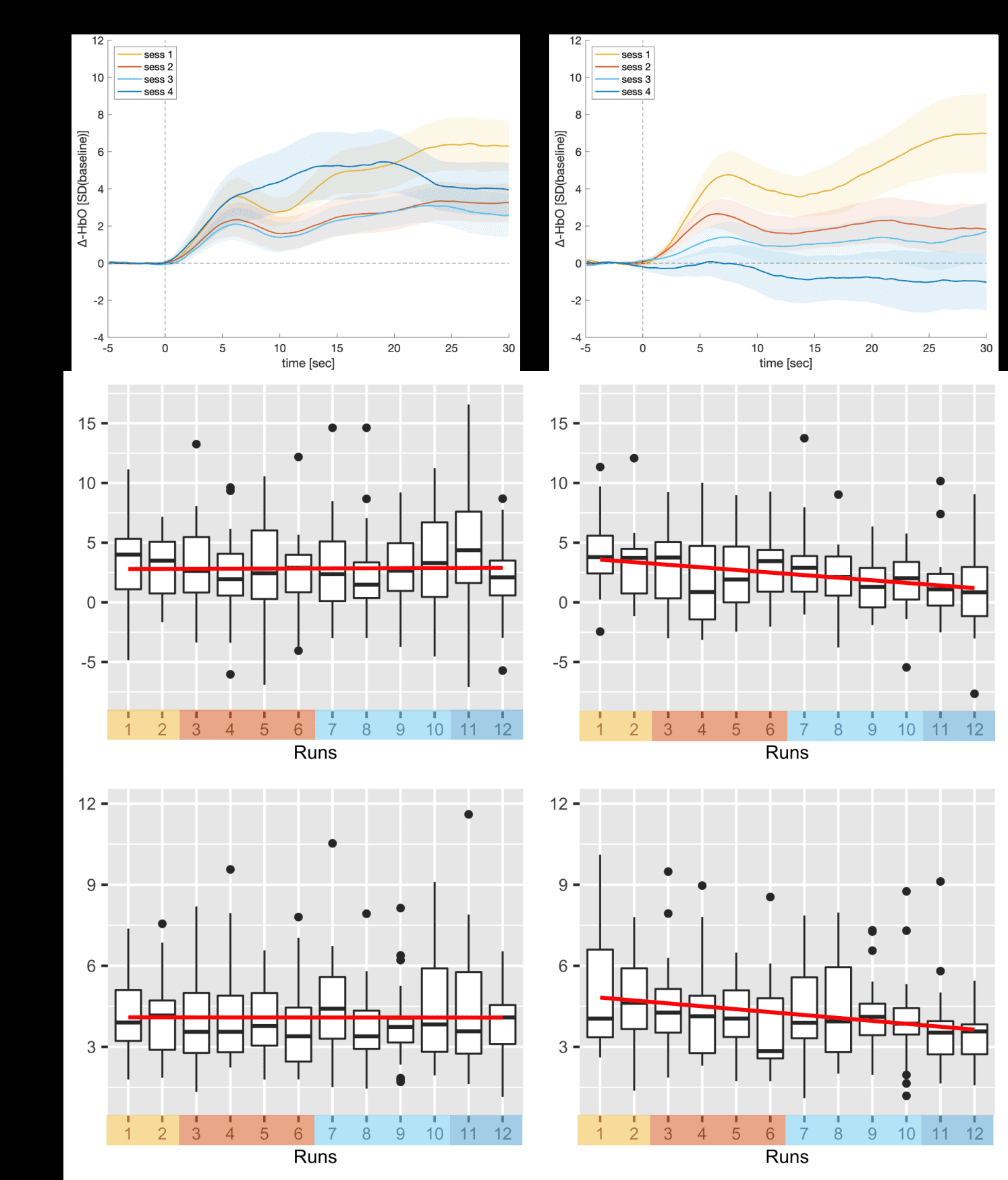
rewards than the downregulation group (M = 12.80±2.33€ vs M = 7.86€±2.16€; t(38,4) = 7.29, p < 0.001, d = 2.35). 3.3 Neurofeedback regulation success Table 3 shows the results for the different neurofeedback success measures and Figure 5 shows grand averages of HbO changes of the feedback signal for all four neurofeedback training days (sessions) and box plots of average feedback performance based on the online analysis for all twelve neurofeedback runs. Tables S5-6 show the individual results of neurofeedback regulation success. Table 3. Neurofeedback regulation success Upregulation group NF performance - compared to baseline NF improvement (slope) NF improvement (last vs first) Downregulation group Online analysis (amplitudes) Online analysis (variability) M±SD 2.84±2.2 0.01±0.34 0.02±3.7 n 26/27 14/27 15/27 p-value < 0.001* M±SD N/A n N/A 0.84 0.97 0.00±0.17 -0.06±1.69 17/27 17/27 p-value N/A 0.595 0.86 M±SD 2.39±1.89 n 4/18 p-value 1 M±SD N/A n N/A p-value N/A NF performance - compared to baseline 0.143 15/18 NF improvement (slope) NF improvement (last vs first) 0.004* 15/18 Neurofeedback regulation success according to different success measures for both groups. The p-values reflect the results of the group analysis as described in 2.5 “Data processing and analysis”; NF, neurofeedback. Regulation success (amplitudes) -0.11±0.17 -1.37±1.76 -0.22±0.35 -2.36±3.6 15/18 15/18 0.09 0.01* In the upregulation group, we observed high rTPJ activation that was sustained over the course of the training. In contrast, the downregulation group unexpectedly showed the same effect (activation instead of deactivation), which, however, disappeared over the course of the training. One-sample t- tests revealed a significant main effect of regulation over all runs in the upregulation group (M = 2.84±2.2, t(26) = 6.72, p < 0.001, d = 1.32) and the downregulation group (M = 2.39±1.89, t(17) = 5.6, p < 0.001, d = 1.36), meaning that on average, rTPJ activity also increased in the downregulation group. Paired-sample t-tests, however, only revealed a significant decrease between the last and the first session in the downregulation group (Mdiff = -2.36±3.6, t(17) = 2.79, p = 0.01, d = 0.68), but no significant increase in the upregulation group (Mdiff = 0.02±3.7, p > 0.98, d = -0.01). The non-
parametric ANOVA only revealed a non-significant time trend in the downregulation group (FATS
24
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023
(5.85, ∞) = 1.86, p = 0.09) and no effect in the upregulation group. No specific group effect or
significant group × time interaction was found.
The analysis on the individual level revealed that in the upregulation group, 96.30% of the
participants (26 of 27) were successfully upregulating rTPJ activity (>50% successful runs; M = 9.63
± 2.27), 51.85% (14 of 27) showed an improvement of regulation performance over runs as indicated
by a positive slope, and 55.56% (15 of 27) showed a higher regulation performance in the last session
compared to the first session. In the downregulation group, only 22.22 % (4 of 18) were successfully
downregulating rTPJ activity (>50% successful runs; M = 3 ± 2.66), 83.33% (15 of 18) showed an
improvement of regulation performance over runs as indicated by a negative slope and a higher
regulation performance in the last compared to the first session.
Regulation success (variability)
For the variability of the neurofeedback performance over time, similar results compared to the main
analysis of regulation success (analysis based on signal amplitudes) were observed. Paired-sample t-
tests also revealed a difference between the last and the first session in the downregulation group
(Mdiff = -1.37±1.76, t(17) = 3.29, p = 0.004, d = 0.8) but not in the upregulation group (Mdiff = –
0.06±1.69, p = 0.86, d = 0.03). The non-parametric ANOVA only revealed a non-significant time
trend in the downregulation group (FATS (7.27, ∞) = 1.55, p = 0.143). No specific group effect or
significant group × time interaction was found.
The individual analysis revealed that in the upregulation group, 62.96% of the participants
(17 of 27) showed decreasing standard deviations over runs, as indicated by a negative slope of the
regression and lower values in the last session compared to the first session. On the other hand, in the
downregulation group, 15 out of 18 participants (83.33%) showed decreasing standard deviations
over runs, as indicated by a negative slope of the regression and lower values in the last session
compared to the first session.
Robustness checks
Robustness check 1 successfully confirmed the results of the online analysis. However, none of the
effects survived the more conservative robustness check 2 (see Supplementary Material 3.3 for
detailed results).
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023
25
Figure 5. Neurofeedback regulation performance. The first row shows the grand averages of the
changes in HbO of the feedback channel for the four neurofeedback training days (sessions). The
second row shows box plots of the average feedback performance as assessed by the standardized
median change of rTPJ activation averaged over participants for all twelve neurofeedback runs
(sessions color-coded) based on the online analysis. The third row shows box plots of the standard
26
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023
deviations of feedback performance as assessed by the standardized median change of right TPJ
activation averaged over participants for each run. The regression lines of the linear models are
depicted in red. Paired-sample t-tests comparing the last session (run 11 and 12) with the first session
(run 1 and 2) revealed significant effects and the ANOVA over all neurofeedback runs only revealed
non-significant time trends in the downregulation group only. No time effect was observed in the
upregulation group (see main text).
3.4 Primary behavioral outcomes
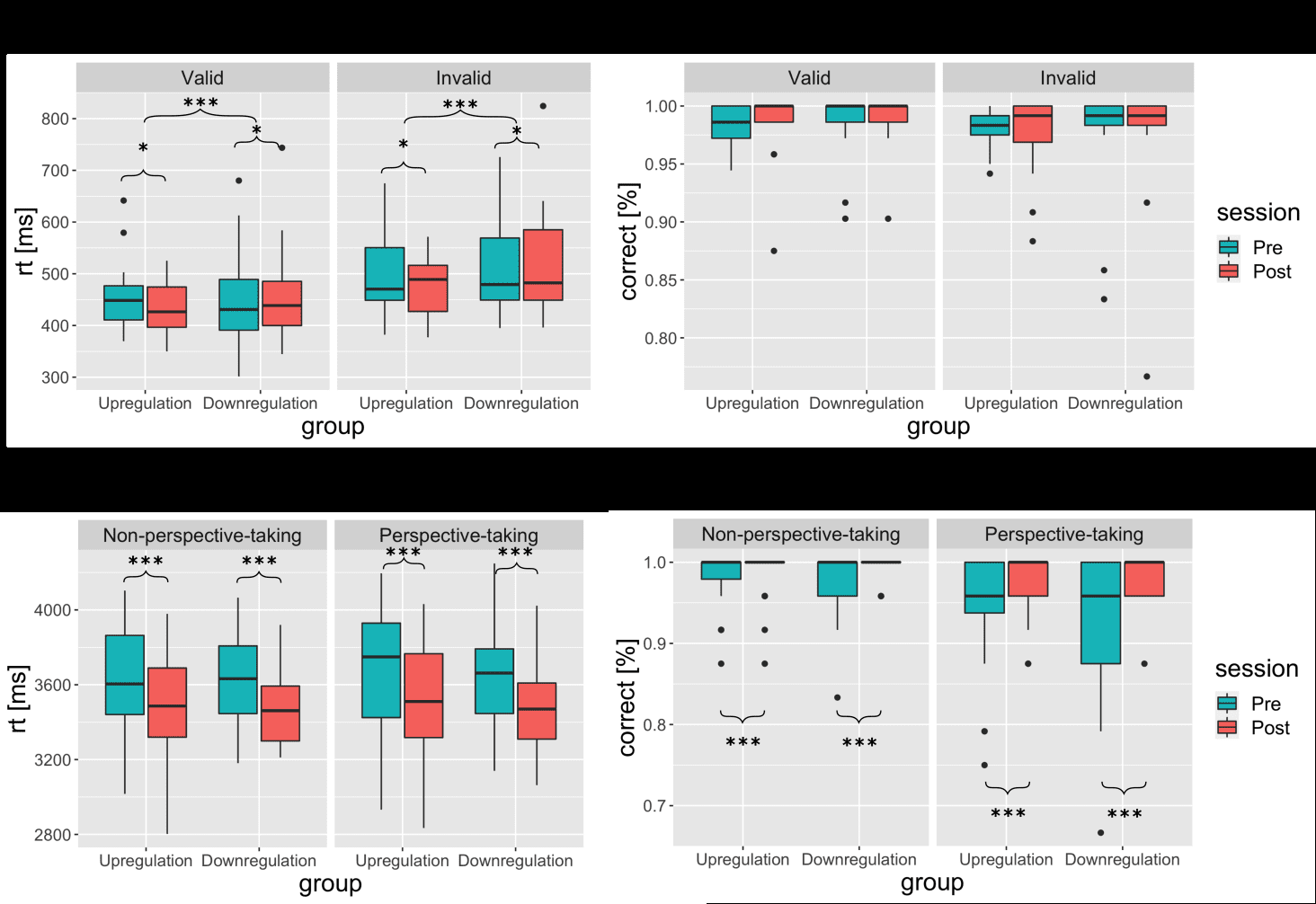
Figure 6. Primary behavioral outcomes. Results of the reorienting of attention task (upper panel) and
visual perspective-taking task (lower panel). For detailed descriptive statistics, see Table S2.
Boxplots show interquartile range ± 1.5 (whiskers). Asterisks denote the significance for the group ×
time interaction and within-group time effects across task conditions; *** p < 0.001; * p < 0.05.
Reorienting of attention task
As expected, we found a significant main effect of condition for RTs (F(1,123) = 111.21, p < 0.001,
p² = 0.47), and accuracy data (FATS (1, ∞) = 17.18, p < 0.001), reflecting a significant reorienting
27
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023
effect in both groups across time points (mean RTs valid = 456 ± 77ms, mean RTs invalid = 499 ±
80ms). The hypothesized three-way interaction of group × time × condition was not observed, i.e., no
significant time effects or group × time interaction effects were observed for the reorienting effect.
However, we found a significant group × time interaction (F(1,123) = 17.17, p < 0.001, p² = 0.12),
and a significant main effect of time in both groups (upregulation group: (FATS (1, ∞) = 6.20, p =
0.013), downregulation group: (FATS (1, ∞) = 4.42, p = 0.036), indicating a group-specific effect of
the training on RTs across conditions. The pre-post comparisons revealed that after the
neurofeedback training, reaction times across conditions decreased in the upregulation group (pre =
474 ± 68ms, post = 457 ± 57ms, d = 0.51) and increased in the downregulation group across
conditions (pre = 488 ± 93ms, post = 503 ± 108ms, d = -0.56). No other main effects or interactions
were found (see Figure 6).
If we included trials from the valid only blocks, results did not change, but the time effect in
the downregulation group (FATS (1, ∞) = 3.25, p = 0.071) failed to reach significance (see
Supplementary Material 4).
vPT task
Contrary to our hypothesis, the three-way interaction of group × time × condition was neither
observed for RTs nor for accuracies in the vPT task. RTs decreased in both groups (F(1,126) = 55.58,
p < 0.001, , p² = 0.31) irrespective of condition (pre = 3630 ± 270ms, post = 3500 ± 295ms, d =
0.83). No other main effects or interactions were significant. Accuracies increased in both groups
(pre = 95.8 ± 3.1%, post = 98.3 ± 6.5%, d = 0.58), as indicated by a significant time effect (FATS (1,
∞) = 11.91, p < 0.001) and a significant condition effect (FATS (1, ∞) = 10.75, p < 0.005), but no
interaction effect occurred.
However, a ceiling effect was observed in this task. The majority of the participants
responded with 100% accuracy in this task during the pre-assessment (29 in the NPT and 18 in the
PT condition) and during the post-assessment (34 in the NPT and 29 in the PT condition; see Figure
6)).
3.5 Mental strategies, secondary outcomes, and unspecific psychological effects
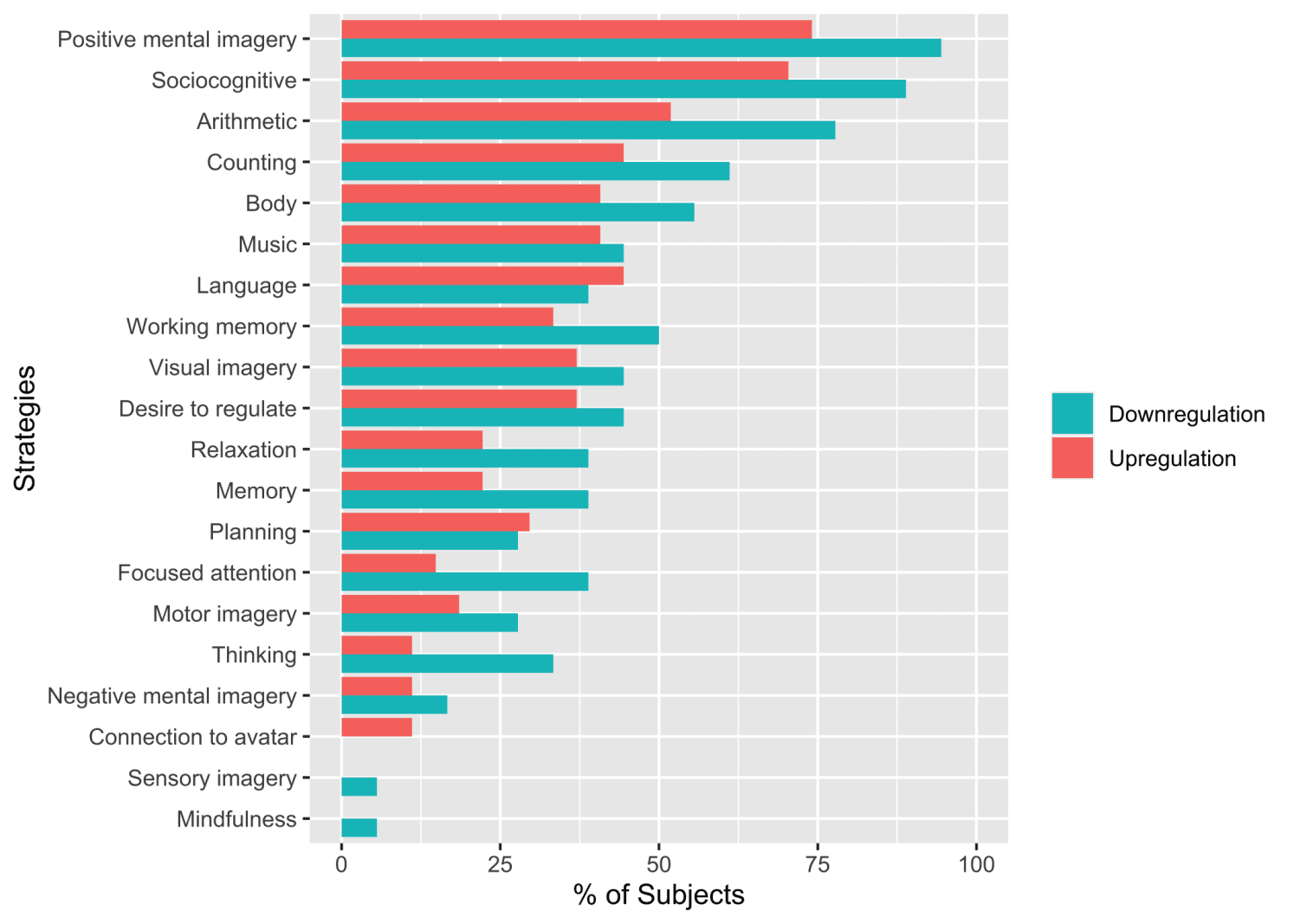
Mental strategies underlying neurofeedback regulation
The downregulation group used significantly more different strategies (M = 8.66±2.47) during the
neurofeedback training compared to the upregulation group (M = 6.26±3.24; t(42.11) = 2.82, p =
0.007, d = 0.87). Figure 7 shows the distribution of strategies as reported by the participants of both
28
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023
groups. Fisher’s exact Chi-square test revealed no significant association between the group and
reported strategies (p = 0.982), indicating that similar strategies were used for both upregulating and
downregulating TPJ activity. Table S8 shows the percentages of strategies relative to the total
number of strategies reported per group and their mean success rating. In total, most strategies were
reported to be more successful in the upregulation group (mean success rating: 3.35) than in the
downregulation group (mean success rating: 2.74), and socio-cognitive strategies and positive mental
imagery were reported most frequently in both groups (see Supplementary Material 5).
Figure 7. Strategies as reported by participants for each group.
Motivation, self-control beliefs, self-efficacy, and mood
Motivation to take further part in the neurofeedback training as assessed after each session
was high in both groups (Mdn = 9, on a 10-point rating scale), but decreased slightly in the
upregulation group over the course of the sessions. There was a significant time effect (FATS (2.34, ∞)
= 3.11, p = 0.04), which was driven by a simple main effect of time in the upregulation group (FATS
29
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023
(2.54, ∞) = 3.48, p = 0.02). No time effect was observed in the downregulation group. Post hoc
comparisons of the last session with the first session confirmed a slight, but significant, decrease of
motivation in the upregulation group (first session, Mdn = 9, last session, Mdn = 8, p = 0.02, r =
0.491) and no effect in the downregulation group (first session, Mdn = 9.5, last session, Mdn = 9.5, p
= 0.43, r = 0.124; see Table S4).
For the general self-efficacy scale, we found a significant time effect (F(1,43) = 4.93, p =
0.03, p² = 0.10). Although the group × time interaction failed to reach significance (F(1,43) = 2.39,
p = 0.129, p² = 0.05), this effect seemed to be driven by an increase in the upregulation group from
pre- (M = 30.59 ± 2.5) to post-assessments (M = 32.04 ± 3.16). This was indicated by a simple main
effect of time, which, however, failed to reach significance (F(1,52) = 3.48, p = 0.07, p² = 0.06). No
time effect was observed in the downregulation group (F(1,34) = 0.02, p = 0.89, p² = 0; see Table
S3).
Participants’ beliefs of how well they could control the neurofeedback signal was lower in the
downregulation group at the beginning of the training, but increased to the level of the upregulation
group towards the end of the training, as indicated by a significant time effect (F(3,128.3) = 3.36, p =
0.02, p² = 0.07), group effect (F(1,42.98) = 11.88, p = 0.001, p² = 0.26) as well as a significant
group time interaction (F(3,128.2) = 6.17, p < 0.001, p² = 0.12). A simple main effect of time was
only observed in the downregulation group (F(3,68) = 4.75, p = 0.005, p² = 0.15). Post hoc t-tests
indicated that there was a group difference in the first neurofeedback session (upregulation group =
7.19±1.42, downregulation group = 4.44±1.82, t(30.53) = 5.37, p < 0.001, d = 1.95) and the second
neurofeedback session (upregulation group = 6.92±1.5, downregulation group = 5.72±1.64, t(34.46)
= 2.48, p = 0.02, d = 0.87), which disappeared in the third session (upregulation group = 7.08±1.35,
downregulation group = 6.56±1.82) and the last session (upregulation group = 6.88±1.82,
downregulation group = 6.36±2.11) see Table S4). The neurofeedback training showed no significant
effect on mood states, as assessed with the POMS (see Table S3).
Expectations and evaluations of the training
No differences were found between the groups with respect to the expectation towards the
neurofeedback training and the subjective evaluation of the training (believed efficacy, joy, and
experimenter). The debriefing questionnaires revealed, however, that 71.11% of the participants
(80.77% in the upregulation and 55.56% in the downregulation group) guessed the group assignment
correctly, although most participants reported that they were not confident about their judgement.
30
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023
3.6 Correlations of behavioral outcomes with regulation performance and psychosocial factors
We found a significant negative correlation between changes in RTs in the valid trials of the
reorienting of attention task and neurofeedback performance, as assessed by the number of successful
runs (rho = -0.47, p = 0.045, Bonferroni corrected), indicating higher improvements of RTs in
participants with more successful runs in both groups. Subgroup analysis revealed no significant
effect after Bonferroni correction.
For the perspective-taking task, we found a significant correlation between neurofeedback
improvement (slopes) and improvements in the accuracies of NPT trials across groups (rho = -0.49, p
= 0.02), indicating greater performance improvements in participants who were more successful in
learning downregulation over the course of the training. This significant correlation was only
observed in the downregulation group (rho = -0.71, p = 0.039, Bonferroni corrected).
None of the psychosocial factors correlated significantly with behavioral outcomes after
Bonferroni correction. For more details including significant correlations on the uncorrected level,
see Supplementary Material 6.
3.7 Predicting behavioral improvements
For the neurophysiologically specific improvements observed in the attention task, we found that IRI
total scores and baseline performance in the attention task predicted changes in performance (see
Table 4). The subgroup models revealed that baseline attentional performance and EQ scores only
predicted behavioral improvements in the upregulation group, thus indicating greater improvements
in participants with lower baseline performance and higher EQ scores. In the downregulation group,
IRI scores and baseline vPT performance predicted decreased performance in the attention task after
the training.
Table 4. Summary statistics of the stepwise multiple linear regression model predicting
behavioral improvements.
Model summary – both groups
R2
0.377
Coefficients
Step
Intercept
IRI total
pre RTs attention task
Adjusted R2
0.346
Residual SE
0.022
Beta
-0.102
0.001
0.17
SE
0.027
0.0004
0.047
F(2,39)
11.82
t-value
-3.836
2.143
3.651
p-value
<0.001***
p-value
<0.001***
0.038*
<0.001***
Model summary – Upregulation group
31
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023
R2
0.513
Coefficients
Step
Intercept
EQ
pre RTs attention task
Adjusted R2
0.471
Residual SE
0.026
Beta
0.169
-0.001
-0.271
SE
0.039
0.001
0.086
F(2,23)
12.12
t-value
4.308
-2.311
-3.170
p-value
<0.001***
p-value
<0.001***
0.03*
0.004**
Adjusted R2
0.38
Model summary – Downregulation group
R2
0.463
Coefficients
Step
Intercept
IRI total
pre accuracy vPT task
Note that we used absolute DRTs in the reorienting of attention task in the model including both groups but the real
DRTs in the subgroup models.
p-value
0.008**
0.017*
0.021*
t-value
-3.106
2.752
2.634
Beta
-0.403
0.003
0.286
SE
0.13
0.001
0.108
Residual SE
0.023
p-value
0.018*
F(2,13)
5.596
4 Discussion
This is the first study demonstrating the feasibility and effectiveness of neurofeedback training of the
rTPJ based on fNIRS. We demonstrated successful activation of the rTPJ compared to baseline
(necessary evidence for control) within the first training session (2 neurofeedback runs) in the
upregulation group. Only one of 27 participants in this group failed to activate (<50% successful
trials). However, we observed no significant effect of neurofeedback improvement; almost half of the
participants (13 of 27) failed to show a positive slope. Successful downregulation, on the other hand,
required at least four sessions (12 neurofeedback runs) or more. Most participants failed to
successfully downregulate, but a significant neurofeedback improvement effect was observed in this
group and only three of 18 participants failed to show such an effect. Surprisingly, participants in the
downregulation group were also activating their rTPJ at the beginning of the training but learnt to
downregulate or at least to not activate it anymore towards the end of the training. This can be
interpreted as strong evidence for control in the downregulation group.
While only unspecific improvements were observed for vPT, specific up/down-regulatory
effects on stimulus-driven attention were observed in the reorienting of attention task, providing
evidence for a neurophysiological specific effect of rTPJ regulation on stimulus-driven spatial
attention, although not specifically related to the reorienting process of attention (as indicated by a
reduced invalidity effect). Neurophysiological specificity was further confirmed by the fact that non-
specific psychological mechanisms and mental strategies did not differ between groups and therefore
cannot explain the group effect. The training was well received by the young and healthy participants
32
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023
with no dropouts as well as high levels of motivation and feelings of control reported throughout the
training.
4.1 Neurofeedback regulation success
While we demonstrate the feasibility and effectiveness of a neurofeedback training of the rTPJ, the
specific results of different neurofeedback success measures, in conjunction with the findings of the
behavioral effects, yield a complex picture. As both groups showed high activation of the rTPJ from
the beginning of the training and only the downregulation group showed a learning effect we cannot
derive definitive conclusion regarding the effectiveness of a neurofeedback upregulation training.
The initial high activation of the rTPJ might be explained by the contribution of general
neurofeedback regulation mechanism by a neurofeedback controller network. Such a separate
controller network involves neural populations of the TPJ (Emmert et al., 2016; Sitaram et al., 2017)
possibly related to the integration of visual feedback as well as other feedback-related processes such
as prediction processing (Bzdok et al., 2013). Therefore, there might have been an overlap of the
neural populations of the neurofeedback controller network with the neurofeedback target region,
meaning that the measured activity at rTPJ could have been a combination of the two. This potential
overlap complicates the interpretation of activity changes and the relevance of feedback. One may
speculate that during the regulation period, the controller network initially increased activity at rTPJ,
but extended learning led to changes in the network, potentially reducing its activity and leading to
complex effects on the measured upregulation and downregulation conditions. In such a scenario
decreased activity in the controller network counteracted a potential increase over time in the target
region, diminishing an observable learning effect. While the baseline period served as a control for
stimuli-evoked activity, it did not account for baseline activity specifically related to the controller
network, which was only engaged during the regulation period. However, we acknowledge the
speculative nature of this account, which can only be confirmed through fMRI studies employing
more fine-grained measures and estimations of the neurofeedback controller network.
Furthermore, the complexity of the social neurofeedback stimuli as well as the instructions
used in our design may explain the initial high activation of the rTPJ. The rTPJ involves parts of the
posterior STS, an area which has been attributed to the face processing network, and a subregion of
the STS closely located to anterior parts of the rTPJ which has been associated with biological
motion as well as emotional face processing (Beauchamp, 2015; Müller et al., 2018). Although we
used digitizer measurements to ensure the correct placements of the feedback channels over anterior
parts of the rTPJ, given the spatial resolution of fNIRS and the variability in optode placements we
33
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023
cannot exclude the possibility that the feedback channel captured the activation of this subregion of
the STS – at least in some of the participants. The activation of the feedback channel might therefore
have been partly induced by the feedback stimuli when the avatar started smiling or even by
participants paying more attention to the facial stimuli during the regulation condition.
Lastly, this effect might be explained by the fact that both groups received the same strategy
instructions and as a result relied heavily on socio-cognitive strategies associated with rTPJ
activation (Bzdok et al., 2013).
The absence of a learning effect in the upregulation group made it also difficult to detect, and
may explain, the absence of a significant group x time interaction effect in the current study (specific
evidence for control).
Nevertheless, the observed neurofeedback learning effect in the downregulation group, along
with the specific effects on the behavioral level, provides interesting and encouraging findings as
they indicate a neurophysiologically specific mechanism of rrTPJ regulation on stimulus-driven
attention. These results have important implications for future study designs and clinical translation
we discuss below (see section 4.2 and 4.5).
The first robustness check further confirmed the results of the online analysis, but the second,
more conservative robustness check did not. This should be interpreted with caution, since given the
limited spatial resolution and coverage in our study the CAR approach involves the risk of
overcorrecting the signal or inducing additional effects depending on network activity during the task
(Hudak et al., 2018; Klein et al., 2022; Kohl et al., 2020). In particular, the first approach (bandpass
filter of 0.01-0.09 Hz) is capable of removing most of the frequencies associated with systemic
physiology, including heart rate (~1Hz), breathing rate (~0.3Hz), and Mayer waves (~0.1Hz; Pinti et
al., 2019). Moreover, we took care to keep the contribution of systemic physiological changes in our
experimental paradigm at a low level by using variable stimulus onsets and instructing our young and
healthy participants to calm down before the experiment, breathe regularly, and avoid unnecessary
movements. Therefore, we can assume that it is very unlikely that the observed effects were driven
by systemic physiology, but instead by the real neural activation of the rTPJ.
4.2 Primary behavioral outcomes
The upregulation group showed increased performance and the downregulation group decreased
performance in the reorienting of attention task across conditions. Our single-blinded, bidirectional-
regulation control group design allowed us to properly control for neurofeedback non-specific or
general non-specific effects and demonstrate neurophysiological specificity of the observed
34
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023
behavioral effects. It is unlikely that differences in the employed strategies can explain the behavioral
effects, since although the downregulation group showed a higher variation of strategies, both groups
relied on the same strategies to regulate their brain activity. The absence of group-specific
correlations of regulation success with behavioral outcomes and the missing group-specific effect in
the vPT task, together with other non-significant correlations between behavioral improvements and
psychosocial factors, underline the role of other non-specific psychosocial mechanisms such as
reward, control beliefs, and expectations in explaining the behavioral effects. However, given the
strength of our study design, the observed dissociation in the reorienting of attention task provides
evidence for a specific neurophysiological effect of rTPJ regulation on stimulus-driven attention.
Contrary to our hypothesis, we did not find a specific effect on the reorienting of attention,
i.e., a specific improvement in invalid trials in the upregulation group. This is surprising given the
assumed specific role of the rTPJ in reorienting of attention (Krall et al., 2015), which has been
supported by previous neurostimulation studies (Krall et al., 2016; Roy et al., 2015). However,
neurofeedback is different from neurostimulation. Instead of passively receiving neurostimulation,
neurofeedback training requires the active participation of the participant and the skill of neural
regulation to be successfully learned, which involves the recruitment of additional neural networks
throughout the training (Emmert et al., 2016; Sitaram et al., 2017) that may have induced additional
behavioral effects. Possible downstream effects of rTPJ regulation on other brain activities may have
also resulted in additional behavioral effects (Kvamme et al., 2022). Furthermore, the specific role of
the rTPJ in early stimulus-driven attentional reorienting has been questioned by Geng and Vossel
(2013), who suggest a rather general role in post-perceptual contextual updating and adjustments of
top-down expectations.
We found increased RTs after downregulation training indicating decreased performance in
stimulus-driven attention, which is an interesting albeit preliminary finding. To date, only a few
studies have applied a bidirectional control group approach and demonstrated group-specific
changes, also including decreases in performance, for example sustained attention and response
inhibition in the study of Yamashita et al. (2017). Future studies should test the robustness of these
effects, assess potential long-term effects, and explore if decreased performance after downregulation
can also be observed in other cognitive domains. If so, the bidirectional control group approach
would be an interesting tool for cognitive neuroscience studies, since it is more efficient and the
demonstration of such a dissociation provides stronger (causal) evidence than just an upregulation
effect. However, caution is advised with respect to long-term effects and when such designs are
applied to clinical populations.
35
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023
Only unspecific improvements were found for vPT. The observed improvement might have
been the result of a retest effect or a ceiling effect, which was observed for most of the participants
and may have masked a group-specific effect in this task. Beneficial effects of rTPJ stimulation were
demonstrated by tDCS studies (Santiesteban et al., 2012, 2015), but the evidence is mixed (Nobusako
et al., 2017; Yang et al., 2020) with most of the studies applying a between-subject design lacking a
baseline control.
Moreover, accuracies were substantially lower than in our study, particularly after sham or
occipital stimulation, which left more room for improvement in these samples than in ours.
Therefore, investigating whether clinical or subclinical samples that are characterized by decreased
perspective-taking performance may benefit from neurofeedback of the rTPJ should be addressed in
future studies. Lastly, more difficult perspective-taking tasks need to be designed to avoid ceiling
effects in participants with high cognitive performance.
4.3 Secondary outcomes and non-specific mechanisms
Self-efficacy improved after the training, and, although we did not find a significant interaction
effect, this effect seemed to be slightly more pronounced in the upregulation group. A number of
neurofeedback studies have demonstrated improvements in domain-specific or general self-efficacy
in different clinical samples and have discussed improvements of self-efficacy as a psychological
mechanism mediating the effect of neurofeedback training (Hershaw et al., 2020; Ko & Park, 2018;
Markiewicz et al., 2021; Mehler et al., 2018; Schmidt & Martin, 2016, 2020). We were unable to find
significant correlations between changes in self-efficacy and behavioral improvements in cognitive
tasks. Self-efficacy might therefore be a psychological mechanism that mediates the effects on
symptom improvement in clinical samples, but this was not observed in the current sample and thus
cannot be responsible for the cognitive improvement observed in the reorienting of attention task in
young and healthy participants.
Regarding non-specific mechanisms, we were unable to find between-group differences in
expectation towards the neurofeedback training and with respect to the evaluation of the training.
Motivation slightly decreased in the upregulation group, although it remained at a high level. The
decrease in motivation might be explained by the lower level of difficulty of the upregulation training
compared to the downregulation training, which was experienced to be more challenging.
Participants in the downregulation group also showed lower control beliefs than the upregulation
group at the beginning of the training, but this difference disappeared over the course of the training
once participants in the downregulation group were regulating more successfully. At the end of the
36
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023
training, we found a significant group difference in the amount of the monetary rewards received,
which occurred due to the higher regulation success in the upregulation group. Unfortunately,
subjective reward experience was not assessed in this study. It is possible, however, that higher
reward experience in the upregulation group may have contributed to the differential behavioral
effects in the reorienting of attention task. Indeed, we found a small, albeit insignificant, correlation
of reward with improvements in this task. It is worth noting, however, that such a difference in
reward experience should have affected the perspective-taking task as well, and differences in mood
states, motivation, evaluation of the training, or control beliefs were not found.
In summary, these findings indicate a low influence of non-specific psychological
mechanisms such as reward, treatment expectations, motivation, and control beliefs (Ros et al., 2020)
and further support a neurophysiologically specific mechanism of rTPJ regulation on stimulus-driven
attention.
4.4 Predicting neurofeedback success
The finding that improvements in stimulus-driven attention were predicted by lower baseline
performance is promising for clinical translation. Clinical populations characterized by difficulties in
stimulus-driven attention, for example ASD (Kana et al., 2014; Landry & Parker, 2013), may benefit
more from the training than our healthy sample.
In particular, while these findings are promising from a clinical translation perspective, we
acknowledge that conclusions are limited to a healthy population. Nevertheless, these exploratory
findings allow us to hypothesize that measures of empathy, as well as the baseline task performance
of the outcome measures, have a predictive value for the behavioral effects of neurofeedback training
of the rTPJ. In this context, it is also noteworthy that comorbid impulsivity symptoms may moderate
the effects of a neurofeedback intervention in ASD and should therefore also be assessed in future
studies (Prillinger et al., 2022). Testing these hypotheses in confirmatory study designs including
clinical samples will allow to identify and select responders of a neurofeedback intervention, which is
important when it comes to the clinical translation of personalized TPJ neurofeedback protocols.
4.5 Limitations and future directions
This study has some limitations that are worth discussing. Since this was the first study
investigating the efficacy of fNIRS neurofeedback of the rTPJ, potential effect sizes were unknown
and therefore the sample size was not determined based on an a-priori statistical power analysis.
37
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023
While this may have resulted in insufficient statistical power to detect small effect sizes such as the
hypothesized group x time interaction effect in regulation performance, it is worth underlining the
large sample size in our study compared to the current state of the fNIRS neurofeedback field (Kohl
et al., 2020). Secondly, both groups showed high activation from the beginning of the training and no
learning effect in the upregulation group was observed, which may lead to the conclusion that mere
mental rehearsal and stimulation through social stimuli is sufficient, and that neurofeedback is not
necessary to regulate rTPJ activity.
In subsequent studies, it could prove advantageous to implement longer training regimes to
possibly foster a learning effect in the upregulation group as well. Additionally, employing more
neutral stimuli, like thermometer images, and refraining from suggesting example strategies, as well
as implementing controls for the activation of a coincident neurofeedback control network, could
assist in isolating a specific mechanism of rTPJ upregulation. Finally, the inclusion of extra control
groups, such as a mental rehearsal group or a sham feedback group, would aid in affirming a specific
mechanism as observed in the attention task.
Thirdly, this study did not involve short-distance measurement or other recommended
measures of systemic physiology (Yücel et al., 2021). With the increasing availability of state-of-the
art online artifact control measures and signal processing methods (Klein and Kranczioch, 2019;
Lühmann et al., 2020; Klein et al., 2022; Schroeder et al., 2023), as well as hardware featuring an
expanded channel count, spatial coverage and the inclusion of short-distance channels, future studies
will be able to better control for systemic physiology but also for signals from irrelevant brain
regions (spatial specificity). However, we took care to keep the contribution of systemic
physiological changes in our design to a minimum and assessed the robustness of the online analysis
through an additional offline analysis using more stringent preprocessing methods
Furthermore, care should be taken when setting and adapting feedback thresholds, particularly
when differences in target regulation difficulty can be expected. Feedback thresholds were based on
online assessments of rTPJ activity during the tasks before the training. We found a large variation in
the assessments (range: 0.03 – 6.92), which may have been the result of suboptimal online processing
methods and artifact control and may have made the training too easy or too difficult for some of the
participants. Future studies can use better online processing methods and thus exclude extreme values
that are likely the result of noisy measures. To avoid differences in rewards, the thresholds should be
adapted more carefully throughout the training and take into account differences in regulation
difficulty, as present in our bidirectional control group design, for example the downregulation group
should start with lower and smaller increases of feedback thresholds.
38
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023
In addition, we had some issues with blinding the participants. Participants were informed
about the bidirectional control group approach and the debriefing revealed that some participants
associated “downregulation” with being more difficult or less successful in the training, while
“upregulation” was associated with the opposite. This might explain why more than 80% in the
upregulation group guessed the group assignment correctly, although none of the participants were
confident about their judgement. Notably, this lack of blinding did not seem to have an influence on
participants, as evidenced by the absence of group differences in motivation, control beliefs,
expectation towards the training, and evaluation of the training. Future neurofeedback experiments
employing a bidirectional control group approach should take care to avoid such associations when
designing the instructions for the participants. If ethically justifiable, participants should not be
informed about the existence of a control condition, or at least not be informed about a
downregulation condition, but rather be instructed that there are two groups in which different
patterns of brain activity are reinforced.
4.5 Conclusion
In summary, this is the first study that demonstrated the feasibility and effectiveness of fNIRS-based
neurofeedback training of the rTPJ. We present preliminary causal evidence that regulation of rTPJ
activity affects stimulus-driven attention. However, it remains unclear if fNIRS-based neurofeedback
can modulate social cognition. Future studies including longer training regimes and better controls
are required to corroborate these initial findings in larger samples using state-of-the-art fNIRS
methods. This study sets the ground for future investigations in clinical populations that are
characterized by the aberrant functioning of the rTPJ or difficulties in stimulus-driven spatial
attention.
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023
39
Data availability statement
Data from this study will be made available on the Open Science Framework (https://osf.io/gbn2r/)
after publication of the study, as far as data protection regulations permit.
CRediT authorship contribution statement
SHK: Conceptualization, Methodology, Software, Formal analysis, Investigation, Data curation,
Writing – original draft, Writing - Review & Editing, Visualization, Project administration
PM: Investigation, Data curation
JU: Software, Formal analysis, Investigation, Data curation, Visualization
ML: Methodology, Software, Writing – Review & Editing
LB: Formal analysis, Writing – Review & Editing, Visualization
DMAM: Writing – Review & Editing
SRS: Writing – Review & Editing
SV: Methodology, Writing – Review & Editing, Visualization
KK: Conceptualization, Resources, Writing – Review & Editing, Supervision, Funding acquisition
Declaration of competing interests
SHK was an employee of MEDIACC GmbH, Berlin, an independent clinical research organization.
SHK and DMAM received payments to consult with Mendi Innovations AB, Stockholm, Sweden.
LB receives commissions for fNIRS visualizations. ML is an employee of the research company
Brain Innovation B.V., Maastricht, the Netherlands. None of the above-mentioned companies were in
relationship with or support of this work. The remaining authors declare no conflicts of interest.
Acknowledgments
This work was supported by the German Research Foundation’s (DFG) International Research
Training Group “The Neuroscience of Modulating Aggression and Impulsivity in Psychopathology”
(IRTG-2150). SHK was supported by a fellowship from the Japan Society for the Promotion of
Science (JSPS). The purchase of the Hitachi fNIRS system for the University Hospital RWTH
Aachen (Germany) was supported by funding from the German Research Foundation (DFG; INST
948/18-1 FUGG), awarded to KK. KK is further supported by the German Research Foundation –
DFG: Project-ID 431549029 –SFB 1451. DMAM is supported by a Junior Principal Investigator
(JPI) fellowship funded by the Excellence Strategy of the Federal 621 Government and the Laender
(grant reference number: JPI074-21). SRS is supported by European Research Council (ERC) under
the project NGBMI (759370) and TIMS (101081905), the Federal Ministry of Research and
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023
40
Education (BMBF) under the projects SSMART (01DR21025A), NEO (13GW0483C), QHMI
(03ZU1110DD) and QSHIFT (01UX2211), as well as the Einstein Foundation Berlin (A-2019-558).
We would like to thank Dr. Iroise Dumontheil for providing the stimuli of the perspective-taking task
and Dr. Simone Vossel for providing the stimuli of the reorienting of attention task. We would also
like to thank Mert Asil Türeli for helping to program the experimental paradigms and Růžena
Ceralová for her contribution to data management.
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023
41
References
Aasted, C. M., Yücel, M. A., Cooper, R. J., Dubb, J., Tsuzuki, D., Becerra, L., Petkov, M. P.,
Borsook, D., Dan, I., & Boas, D. A. (2015). Anatomical guidance for functional near-infrared
spectroscopy: AtlasViewer tutorial. Neurophotonics, 2(2), 020801.
https://doi.org/10.1117/1.nph.2.2.020801
Akaike, H. (1974). A New Look at the Statistical Model Identification. IEEE Transactions on
Automatic Control, 19(6), 716–723. https://doi.org/10.1109/TAC.1974.1100705
Auer, T., Schweizer, R., & Frahm, J. (2015). Training efficiency and transfer success in an extended
real-time functional MRI neurofeedback training of the somatomotor cortex of healthy subjects.
Frontiers in Human Neuroscience, 9(OCT), 547. https://doi.org/10.3389/fnhum.2015.00547
Barman, A., Richter, S., Soch, J., Deibele, A., Richter, A., Assmann, A., Wüstenberg, T., Walter, H.,
Seidenbecher, C. I., & Schott, B. H. (2015). Gender-specific modulation of neural mechanisms
underlying social reward processing by Autism Quotient. Social Cognitive and Affective
Neuroscience, 10(11), 1537–1547. https://doi.org/10.1093/SCAN/NSV044
Baron-Cohen, S., Richler, J., Bisarya, D., Gurunathan, N., & Wheelwright, S. (2003). The
systemizing quotient: An investigation of adults with Asperger syndrome or high-functioning
autism, and normal sex differences. Philosophical Transactions of the Royal Society B:
Biological Sciences, 358(1430), 361–374. https://doi.org/10.1098/rstb.2002.1206
Baron-Cohen, S., & Wheelwright, S. (2004). The empathy quotient: An investigation of adults with
asperger syndrome or high functioning autism, and normal sex differences. In Journal of Autism
and Developmental Disorders (Vol. 34, Issue 2, pp. 163–175). Kluwer Academic Publishers-
Plenum Publishers. https://doi.org/10.1023/B:JADD.0000022607.19833.00
Baron-Cohen, S., Wheelwright, S., Skinner, R., Martin, J., & Clubley, E. (2001). The Autism-
Spectrum Quotient (AQ): Evidence from Asperger Syndrome/High-Functioning Autism, Males
and Females, Scientists and Mathematicians. Journal of Autism and Developmental Disorders,
31(1), 5–17. https://doi.org/10.1023/A:1005653411471
Bates, D., Mächler, M., Bolker, B. M., & Walker, S. C. (2015). Fitting linear mixed-effects models
using lme4. Journal of Statistical Software, 67(1), 1–48. https://doi.org/10.18637/jss.v067.i01
Beauchamp, M. S. (2015). The social mysteries of the superior temporal sulcus. In Trends in
Cognitive Sciences (Vol. 19, Issue 9, pp. 489–490). Elsevier Ltd.
https://doi.org/10.1016/j.tics.2015.07.002
Bell, L., Scharke, W., Reindl, V., Fels, J., Neuschaefer-Rube, C., & Konrad, K. (2020). Auditory and
visual response inhibition in children with bilateral hearing aids and children with adhd. Brain
Sciences, 10(5), 307. https://doi.org/10.3390/brainsci10050307
Bölte, S. (2012). Brief report: The Social Responsiveness Scale for adults (SRS-A): Initial results in a
German cohort. Journal of Autism and Developmental Disorders, 42(9), 1998–1999.
https://doi.org/10.1007/s10803-011-1424-5
Carter, R. M., & Huettel, S. A. (2013). A Nexus Model of the Temporal-Parietal Junction.
https://doi.org/10.1016/j.tics.2013.05.007
Cliffordson, C. (2001). Parent’s Judgments and Students’ Self-Judgments of Empathy: The Structure
of Empathy and Agreement of Judgment Based on the Interpersonal Reactivity Index (IRI)
Grade and grade assignment: Function and effects View project Self care and health among
older people in Norway View project. Article in European Journal of Psychological Assessment.
https://doi.org/10.1027/1015-5759.17.1.36
Corbetta, M., Patel, G., & Shulman, G. L. (2008). The reorienting system of the human brain: from
environment to theory of mind. Neuron, 58(3), 306–324.
https://doi.org/10.1016/j.neuron.2008.04.017
42
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023
Cui, X., Bray, S., & Reiss, A. L. (2010). Functional near infrared spectroscopy (NIRS) signal
improvement based on negative correlation between oxygenated and deoxygenated hemoglobin
dynamics. NeuroImage, 49(4), 3039–3046. https://doi.org/10.1016/j.neuroimage.2009.11.050
Davis, M. H. (1983). Measuring individual differences in empathy: Evidence for a multidimensional
approach. Journal of Personality and Social Psychology, 44(1), 113–126.
https://doi.org/10.1037/0022-3514.44.1.113
Decety, J., & Lamm, C. (2007). The Role of the Right Temporoparietal Junction in Social
Interaction: How Low-Level Computational Processes Contribute to Meta-Cognition. The
Neuroscientist, 13(6), 580–593. https://doi.org/10.1177/1073858407304654
Direito, B., Lima, J., Simões, M., Sayal, A., Sousa, T., Lührs, M., Ferreira, C., & Castelo-Branco, M.
(2019). Targeting dynamic facial processing mechanisms in superior temporal sulcus using a
novel fMRI neurofeedback target. Neuroscience, 406, 97–108.
https://doi.org/10.1016/j.neuroscience.2019.02.024
Direito, B., Mouga, S., Sayal, A., Simões, M., Quental, H., Bernardino, I., Playle, R., McNamara, R.,
Linden, D. E. J., Oliveira, G., & Castelo Branco, M. (2021). Training the social brain: Clinical
and neural effects of an 8-week real-time functional magnetic resonance imaging neurofeedback
Phase IIa Clinical Trial in Autism. Autism, 25(6), 1746–1760.
https://doi.org/10.1177/13623613211002052
Donaldson, P. H., Kirkovski, M., Rinehart, N. J., & Enticott, P. G. (2018). Autism-relevant traits
interact with temporoparietal junction stimulation effects on social cognition: a high-definition
transcranial direct current stimulation and electroencephalography study. European Journal of
Neuroscience, 47(6), 669–681. https://doi.org/10.1111/EJN.13675
Dumontheil, I., Küster, O., Apperly, I. A., & Blakemore, S. J. (2010). Taking perspective into
account in a communicative task. NeuroImage, 52(4), 1574–1583.
https://doi.org/10.1016/j.neuroimage.2010.05.056
Emmert, K., Kopel, R., Sulzer, J., Brühl, A. B., Berman, B. D., Linden, D. E. J., Horovitz, S. G.,
Breimhorst, M., Caria, A., Frank, S., Johnston, S., Long, Z., Paret, C., Robineau, F., Veit, R.,
Bartsch, A., Beckmann, C. F., van de Ville, D., & Haller, S. (2016). Meta-analysis of real-time
fMRI neurofeedback studies using individual participant data: How is brain regulation
mediated? NeuroImage, 124, 806–812. https://doi.org/10.1016/j.neuroimage.2015.09.042
Esse Wilson, J., Trumbo, M. C., Wilson, J. K., & Tesche, C. D. (2018). Transcranial direct current
stimulation (tDCS) over right temporoparietal junction (rTPJ) for social cognition and social
skills in adults with autism spectrum disorder (ASD). Journal of Neural Transmission 2018
125:12, 125(12), 1857–1866. https://doi.org/10.1007/S00702-018-1938-5
Geng, J. J., & Vossel, S. (2013). Re-evaluating the role of TPJ in attentional control: Contextual
updating? Neuroscience & Biobehavioral Reviews, 37(10), 2608–2620.
https://doi.org/10.1016/J.NEUBIOREV.2013.08.010
Greer, S. M., Trujillo, A. J., Glover, G. H., & Knutson, B. (2014). Control of nucleus accumbens
activity with neurofeedback. NeuroImage, 96, 237–244.
https://doi.org/10.1016/j.neuroimage.2014.03.073
Harmelech, T., Friedman, D., & Malach, R. (2015). Differential Magnetic Resonance Neurofeedback
Modulations across Extrinsic (Visual) and Intrinsic (Default-Mode) Nodes of the Human
Cortex. Journal of Neuroscience, 35(6), 2588–2595. https://doi.org/10.1523/jneurosci.3098-
14.2015
Hartz, A., Guth, B., Jording, M., Vogeley, K., & Schulte-Rüther, M. (2021). Temporal Behavioral
Parameters of On-Going Gaze Encounters in a Virtual Environment. Frontiers in Psychology,
12, 3056. https://doi.org/10.3389/FPSYG.2021.673982/BIBTEX
Haugg, A., Renz, F. M., Nicholson, A. A., Lor, C., Götzendorfer, S. J., Sladky, R., Skouras, S.,
McDonald, A., Craddock, C., Hellrung, L., Kirschner, M., Herdener, M., Koush, Y., Papoutsi,
43
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023
M., Keynan, J., Hendler, T., Cohen Kadosh, K., Zich, C., Kohl, S. H., … Steyrl, D. (2021).
Predictors of real-time fMRI neurofeedback performance and improvement – A machine
learning mega-analysis. NeuroImage, 237, 118207.
https://doi.org/10.1016/J.NEUROIMAGE.2021.118207
Hershaw, J. N., Hill-Pearson, C. A., Arango, J. I., Souvignier, A. R., & Pazdan, R. M. (2020). Semi-
Automated Neurofeedback Therapy for Persistent Postconcussive Symptoms in a Military
Clinical Setting: A Feasibility Study. Military Medicine, 185(3–4), e457–e465.
https://doi.org/10.1093/MILMED/USZ335
Hudak, J., Rosenbaum, D., Barth, B., Fallgatter, A. J., & Ehlis, A. C. (2018). Functionally
disconnected: A look at how study design influences neurofeedback data and mechanisms in
attention-deficit/hyperactivity disorder. PLoS One, 13(8), 1–15.
https://doi.org/10.1371/journal.pone.0200931
Huppert, T. J., Diamond, S. G., Franceschini, M. A., & Boas, D. A. (2009). HomER: A review of
time-series analysis methods for near-infrared spectroscopy of the brain. Applied Optics, 48(10),
D280–D298. https://doi.org/10.1364/AO.48.00D280
Jasper, H. H. (1958). The Ten-Twenty Electrode System of the International Federation.
Electroencephalography and Clinical Neurophysiology, 10, 371–375.
Kana, R. K., Libero, L. E., Hu, C. P., Deshpande, H. D., & Colburn, J. S. (2014). Functional Brain
Networks and White Matter Underlying Theory-of-Mind in Autism. Social Cognitive and
Affective Neuroscience, 9(1), 98. https://doi.org/10.1093/SCAN/NSS106
Kana, R. K., Maximo, J. O., Williams, D. L., Keller, T. A., Schipul, S. E., Cherkassky, V. L.,
Minshew, N. J., & Just, M. A. (2015). Aberrant functioning of the theory-of-mind network in
children and adolescents with autism. Molecular Autism, 6(1), 59.
https://doi.org/10.1186/s13229-015-0052-x
Kana, R. K., Patriquin, M. A., Black, B. S., Channell, M. M., & Wicker, B. (2016). Altered Medial
Frontal and Superior Temporal Response to Implicit Processing of Emotions in Autism. Autism
Research, 9(1), 55–66. https://doi.org/10.1002/aur.1496
Kessler, R. C., Adler, L., Ames, M., Demler, O., Faraone, S., Hiripi, E., Howes, M. J., Jin, R.,
Secnik, K., Spencer, T., Ustun, T. B., & Walters, E. E. (2005). The World Health Organization
adult ADHD self-report scale (ASRS): A short screening scale for use in the general population.
Psychological Medicine, 35(2), 245–256. https://doi.org/10.1017/S0033291704002892
Kinoshita, A., Takizawa, R., Yahata, N., Homae, F., Hashimoto, R., Sakakibara, E., Kawasaki, S.,
Nishimura, Y., Koike, S., & Kasai, K. (2016). Development of a neurofeedback protocol
targeting the frontal pole using near-infrared spectroscopy. Psychiatry and Clinical
Neurosciences, 70(11), 507–516. https://doi.org/10.1111/pcn.12427
Klein, F., Lührs, M., Benitez-Andonegui, A., Roehn, P., & Kranczioch, C. (2022). Performance
comparison of systemic activity correction in functional near-infrared spectroscopy for methods
with and without short distance channels. Https://Doi.Org/10.1117/1.NPh.10.1.013503, 10(1),
013503. https://doi.org/10.1117/1.NPH.10.1.013503
Ko, S., & Park, W. (2018). Effects of Quantitative Electroencephalography Based Neurofeedback
Training on Autonomous Regulations in Patients with Alcohol Use Disorder. Asian Nursing
Research, 12(2), 136–144. https://doi.org/10.1016/J.ANR.2018.05.003
Kohl, S. H., Mehler, D. M. A., Lührs, M., Thibault, R. T., Konrad, K., & Sorger, B. (2020). The
potential of functional near-infrared spectroscopy-based neurofeedback – a systematic review
and recommendations for best practice. Frontiers in Neuroscience, 14, 594.
https://doi.org/10.3389/FNINS.2020.00594
Krall, S. C., Rottschy, C., Oberwelland, E., Bzdok, D., Fox, P. T., Eickhoff, S. B., Fink, G. R., &
Konrad, K. (2015). The role of the right temporoparietal junction in attention and social
44
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023
interaction as revealed by ALE meta-analysis. Brain Structure and Function, 220(2), 587–604.
https://doi.org/10.1007/s00429-014-0803-z
Krall, S. C., Volz, L. J., Oberwelland, E., Grefkes, C., Fink, G. R., & Konrad, K. (2016). The right
temporoparietal junction in attention and social interaction: A transcranial magnetic stimulation
study. Human Brain Mapping, 37(2), 796–807. https://doi.org/10.1002/hbm.23068
Kuznetsova, A., Brockhoff, P. B., & Christensen, R. H. B. (2017). lmerTest Package: Tests in Linear
Mixed Effects Models . Journal of Statistical Software, 82(13), 1–26.
https://doi.org/10.18637/jss.v082.i13
Kvamme, T. L., Ros, T., & Overgaard, M. (2022). Can neurofeedback provide evidence of direct
brain-behavior causality? NeuroImage, 258, 119400.
https://doi.org/10.1016/J.NEUROIMAGE.2022.119400
Landry, O., & Parker, A. (2013). A meta-analysis of visual orienting in autism. Frontiers in Human
Neuroscience, 0, 833. https://doi.org/10.3389/FNHUM.2013.00833
Mai, X., Zhang, W., Hu, X., Zhen, Z., Xu, Z., Zhang, J., & Liu, C. (2016). Using tDCS to Explore
the Role of the Right Temporo-Parietal Junction in Theory of Mind and Cognitive Empathy.
Frontiers in Psychology, 0(MAR), 380. https://doi.org/10.3389/FPSYG.2016.00380
Markiewicz, R., Markiewicz-Gospodarek, A., Dobrowolska, B., & Łoza, B. (2021). Improving
Clinical, Cognitive, and Psychosocial Dysfunctions in Patients with Schizophrenia: A
Neurofeedback Randomized Control Trial. Neural Plasticity, 2021.
https://doi.org/10.1155/2021/4488664
Mars, R. B., Sallet, J., Schüffelgen, U., Jbabdi, S., Toni, I., & Rushworth, M. F. S. (2012).
Connectivity-based subdivisions of the human right “temporoparietal junction area”: Evidence
for different areas participating in different cortical networks. Cerebral Cortex, 22(8), 1894–
1903. https://doi.org/10.1093/cercor/bhr268
Mathiak, K. A., Alawi, E. M., Koush, Y., Dyck, M., Cordes, J. S., Gaber, T. J., Zepf, F. D.,
Palomero-Gallagher, N., Sarkheil, P., Bergert, S., Zvyagintsev, M., & Mathiak, K. (2015).
Social reward improves the voluntary control over localized brain activity in fMRI-based
neurofeedback training. Frontiers in Behavioral Neuroscience, 9, 136.
https://doi.org/10.3389/fnbeh.2015.00136
McNair, D., Lorr, M., & Droppleman, L. (1981). POMS Profile of Mood States. Ein Verfahren zur
Messung von Stimmungszuständen. Internationale Skalen Für Psychiatrie.
Mehler, D. M. A., Sokunbi, M. O., Habes, I., Barawi, K., Subramanian, L., Range, M., Evans, J.,
Hood, K., Lührs, M., Keedwell, P., Goebel, R., & Linden, D. E. J. (2018). Targeting the
affective brain—a randomized controlled trial of real-time fMRI neurofeedback in patients with
depression. Neuropsychopharmacology, November 2017, 1–8. https://doi.org/10.1038/s41386-
018-0126-5
Mengotti, P., Käsbauer, A.-S., Fink, G. R., & Vossel, S. (2022). Combined TMS-fMRI Reveals
Behavior-Dependent Network Effects of Right Temporoparietal Junction Neurostimulation in
an Attentional Belief Updating Task. Cerebral Cortex.
https://doi.org/10.1093/CERCOR/BHAB511
Müller, V. I., Höhner, Y., & Eickhoff, S. B. (2018). Influence of task instructions and stimuli on the
neural network of face processing: An ALE meta-analysis. Cortex, 103, 240–255.
https://doi.org/10.1016/j.cortex.2018.03.011
Murphy, A. C., & Bassett, D. S. (2017). A network neuroscience of neurofeedback for clinical
translation. Current Opinion in Biomedical Engineering, 1, 63–70.
https://doi.org/10.1016/j.cobme.2017.03.003
Nobusako, S., Nishi, Y., Nishi, Y., Shuto, T., Asano, D., Osumi, M., & Morioka, S. (2017).
Transcranial Direct Current Stimulation of the Temporoparietal Junction and Inferior Frontal
Cortex Improves Imitation-Inhibition and Perspective-Taking with no Effect on the Autism-
45
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023
Spectrum Quotient Score. Frontiers in Behavioral Neuroscience, 11, 84.
https://doi.org/10.3389/FNBEH.2017.00084
Noguchi, K., Gel, Y. R., Brunner, E., & Konietschke, F. (2012). nparLD : An R Software Package for
the Nonparametric Analysis of Longitudinal Data in Factorial Experiments. Journal of
Statistical Software, 50(12). https://doi.org/10.18637/jss.v050.i12
Pamplona, G. S. P., Heldner, J., Langner, R., Koush, Y., Michels, L., Ionta, S., Scharnowski, F., &
Salmon, C. E. G. (2020). Network-based fMRI-neurofeedback training of sustained attention.
NeuroImage, 221, 117194. https://doi.org/10.1016/J.NEUROIMAGE.2020.117194
Paret, C., Goldway, N., Zich, C., Keynan, J. N., Hendler, T., Linden, D., & Cohen Kadosh, K.
(2019). Current progress in real-time functional magnetic resonance-based neurofeedback:
Methodological challenges and achievements. NeuroImage, 202, 116107.
https://doi.org/10.1016/j.neuroimage.2019.116107
Paulus, C. (2012). Ist die Bildung eines Empathiescores in der deutschen Fassung des IRI sinnvoll?
https://doi.org/doi:10.22028/D291-23347
Penner, J., Osuch, E. A., Schaefer, B., Théberge, J., Neufeld, R. W. J., Menon, R. S., Rajakumar, N.,
& Williamson, P. C. (2018). Temporoparietal Junction Functional Connectivity in Early
Schizophrenia and Major Depressive Disorder. Chronic Stress, 2.
https://doi.org/10.1177/2470547018815232
Pereira, H. C., Sousa, D., Simões, M., Martins, R., Amaral, C., Lopes, V., Crisóstomo, J., & Castelo-
Branco, M. (2021). Effects of anodal multichannel transcranial direct current stimulation (tDCS)
on social-cognitive performance in healthy subjects: A randomized sham-controlled crossover
pilot study. In Progress in Brain Research (Vol. 264, pp. 259–286). Elsevier B.V.
https://doi.org/10.1016/bs.pbr.2021.04.004
Pinti, P., Scholkmann, F., Hamilton, A., Burgess, P., & Tachtsidis, I. (2019). Current Status and
Issues Regarding Pre-processing of fNIRS Neuroimaging Data: An Investigation of Diverse
Signal Filtering Methods Within a General Linear Model Framework. Frontiers in Human
Neuroscience, 12, 505. https://doi.org/10.3389/fnhum.2018.00505
Posner, M. I. (1980). Orienting of attention. The Quarterly Journal of Experimental Psychology,
32(1), 3–25. https://doi.org/10.1080/00335558008248231
R Core Team. (2021). R: A language and environment for statistical computing. R Foundation for
Statistical Computing. https://www.R-project.org/
Ratcliff, R. (1993). Methods for dealing with reaction time outliers. Psychological Bulletin, 114(3),
510–532. https://doi.org/10.1037/0033-2909.114.3.510
Ros, T., Enriquez-Geppert, S., Zotev, V., Young, K. D., Wood, G., Whitfield-Gabrieli, S., Wan, F.,
Vuilleumier, P., Vialatte, F., van de Ville, D., Todder, D., Surmeli, T., Sulzer, J. S., Strehl, U.,
Sterman, M. B., Steiner, N. J., Sorger, B., Soekadar, S. R., Sitaram, R., … Thibault, R. T.
(2020). Consensus on the reporting and experimental design of clinical and cognitive-
behavioural neurofeedback studies (CRED-nf checklist). Brain, 143(6), 1674–1685.
https://doi.org/10.1093/brain/awaa009
Roy, L. B., Sparing, R., Fink, G. R., & Hesse, M. D. (2015). Modulation of attention functions by
anodal tDCS on right PPC. Neuropsychologia, 74, 96–107.
https://doi.org/10.1016/j.neuropsychologia.2015.02.028
Salehinejad, M. A., Paknia, N., Hosseinpour, A. H., Yavari, F., Vicario, C. M., Nitsche, M. A., &
Nejati, V. (2021). Contribution of the right temporoparietal junction and ventromedial prefrontal
cortex to theory of mind in autism: A randomized, sham-controlled tDCS study. Autism
Research, 1–13. https://doi.org/10.1002/AUR.2538
Santiesteban, I., Banissy, M. J., Catmur, C., & Bird, G. (2012). Enhancing social ability by
stimulating right temporoparietal junction. Current Biology, 22(23), 2274–2277.
https://doi.org/10.1016/j.cub.2012.10.018
46
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023
Santiesteban, I., Banissy, M. J., Catmur, C., & Bird, G. (2015). Functional lateralization of
temporoparietal junction - imitation inhibition, visual perspective-taking and theory of mind.
The European Journal of Neuroscience, 42(8), 2527–2533. https://doi.org/10.1111/ejn.13036
Schmidt, J., & Martin, A. (2016). Neurofeedback Against Binge Eating: A Randomized Controlled
Trial in a Female Subclinical Threshold Sample. https://doi.org/10.1002/erv.2453
Schmidt, J., & Martin, A. (2020). The Influence of Physiological and Psychological Learning
Mechanisms in Neurofeedback vs. Mental Imagery Against Binge Eating. Applied
Psychophysiology Biofeedback, 45(4), 293–305. https://doi.org/10.1007/S10484-020-09486-
9/TABLES/4
Schönenberg, M., Weingärtner, A. L., Weimer, K., & Scheeff, J. (2021). Believing is achieving - On
the role of treatment expectation in neurofeedback applications. Progress in Neuro-
Psychopharmacology and Biological Psychiatry, 105.
https://doi.org/10.1016/j.pnpbp.2020.110129
Schwarzer, R., & Jerusalem, M. (1995). General Self-Efficacy Scale (GSE) (tabel) : In Measures in
health psychology: A user’s portfolio. Causal and control beliefs. Generalised Self-Efficacy
Scale1st Ed, J Weinman, S Wright, M Johnston. Nfer-Nelson, WindsorUK, 1995, 35–37.
Sitaram, R., Ros, T., Stoeckel, L., Haller, S., Scharnowski, F., Lewis-Peacock, J., Weiskopf, N.,
Blefari, M. L., Rana, M., Oblak, E., Birbaumer, N., & Sulzer, J. (2017). Closed-loop brain
training: the science of neurofeedback. Nature Reviews Neuroscience, 18(2), 86–100.
https://doi.org/10.1038/nrn.2016.164
Soekadar, S. R., Kohl, S. H., Mihara, M., & von Lühmann, A. (2021). Optical brain imaging and its
application to neurofeedback. NeuroImage: Clinical, 30.
https://doi.org/10.1016/j.nicl.2021.102577
Sorger, B., Scharnowski, F., Linden, D. E. J., Hampson, M., & Young, K. D. (2019). Control freaks:
Towards optimal selection of control conditions for fMRI neurofeedback studies. NeuroImage,
186, 256–265. https://doi.org/10.1016/j.neuroimage.2018.11.004
Symeonidou, I., Dumontheil, I., Chow, W.-Y., & Breheny, R. (2016). Development of online use of
theory of mind during adolescence: An eye-tracking study. Journal of Experimental Child
Psychology, 149, 81–97. https://doi.org/10.1016/j.jecp.2015.11.007
van Overwalle, F. (2009). Social cognition and the brain: A meta-analysis. Human Brain Mapping,
30(3), 829–858. https://doi.org/10.1002/HBM.20547
Vossel, S., Weidner, R., Thiel, C. M., & Fink, G. R. (2009). What is “Odd” in Posner’s Location-
cueing Paradigm? Neural Responses to Unexpected Location and Feature Changes Compared.
Journal of Cognitive Neuroscience, 21(1), 30–41. https://doi.org/10.1162/jocn.2009.21003
World Medical Association. (2013). World Medical Association Declaration of Helsinki: Ethical
Principles for Medical Research Involving Human Subjects. JAMA, 310(20), 2191–2194.
https://doi.org/10.1001/JAMA.2013.281053
Yamashita, A., Hayasaka, S., Kawato, M., & Imamizu, H. (2017). Connectivity Neurofeedback
Training Can Differentially Change Functional Connectivity and Cognitive Performance.
Cerebral Cortex, 27(10), 4960–4970. https://doi.org/10.1093/CERCOR/BHX177
Yang, L.-Z., Zhang, W., Wang, W., Yang, Z., Wang, H., Deng, Z.-D., Li, C., Qiu, B., Zhang, D.-R.,
Kadosh, R. C., Li, H., & Zhang, X. (2020). Neural and Psychological Predictors of Cognitive
Enhancement and Impairment from Neurostimulation. Advanced Science, 7(4), 1902863.
https://doi.org/10.1002/ADVS.201902863
Yücel, M. A., Lühmann, A. v., Scholkmann, F., Gervain, J., Dan, I., Ayaz, H., Boas, D., Cooper, R.
J., Culver, J., Elwell, C. E., Eggebrecht, A., Franceschini, M. A., Grova, C., Homae, F., Lesage,
F., Obrig, H., Tachtsidis, I., Tak, S., Tong, Y., … Wolf, M. (2021). Best practices for fNIRS
publications. Neurophotonics, 8(1), 012101. https://doi.org/10.1117/1.NPH.8.1.012101
47
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00014/2154906/imag_a_00014.pdf by guest on 07 September 2023