Initial experiences with Direct Imaging of Neuronal Activity (DIANA)
in humans
Shota Hodonoa,*, Reuben Rideauxb,c,*, Timo van Kerkoerled, Martijn A. Cloosa,e
aCentre for Advanced Imaging, The University of Queensland, Brisbane, Australia
bQueensland Brain Institute, The University of Queensland, Brisbane, Australia
cSchool of Psychology, The University of Sydney, Camperdown, Australia
dCognitive Neuroimaging Unit, CEA, INSERM, Université Paris- Saclay, NeuroSpin Center, Gif/Yvette, France
eARC Training Centre for Innovation in Biomedical Imaging Technology (CIBIT), The University of Queensland, Brisbane, Australia
*S.H. and R.R. contributed equally to this work
Corresponding Author: Shota Hodono (s.hodono@uq.edu.au)
ABSTRACT
Functional MRI (fMRI) has been widely used to study activity patterns in the human brain. It infers neuronal activity
from the associated hemodynamic response, which fundamentally limits its spatiotemporal specificity. In mice, the
Direct Imaging of Neuronal Activity (DIANA) method revealed MRI signals that correlated with extracellular electric
activity, showing high spatiotemporal specificity. In this work, we attempted DIANA in humans. Five experimental
paradigms were tested, exploring different stimulus types (flickering noise patterns, and naturalistic images), stimulus
durations (50– 200 ms), and imaging resolution (2 × 2 × 5 mm3 and 1 × 1 × 5 mm3). Regions of interest (ROI) were
derived from Blood Oxygen Level Dependent (BOLD) fMRI acquisitions (both EPI and FLASH based) and T1- weighted
anatomical scans. In Paradigm I (n = 1), using flickering noise patterns, signals were detected that resembled possible
functional activity from a small ROI. However, changes in stimulus duration did not lead to corresponding signal
changes (Paradigm II; n = 1). Therefore, care should be taken not to mistake artifacts for neuronal activity. In Paradigm
III (n = 3), when averaged across multiple subjects, a ~200 ms long 0.02% signal increase was observed ~100 ms after
the stimulus onset (10x smaller than the expected signal). However, white matter control ROIs showed similarly large
signal fluctuations. In Paradigm IV (n = 3), naturalistic image stimuli were used, but did not reveal signs of a potential
functional signal. To reduce partial voluming effects and improve ROI definition, in Paradigm V (n = 3), we acquired
data with higher resolution (1 × 1 × 5 mm3) using naturalistic images. However, no sign of activation was found. It is
important to note that repetitive experiments with short interstimulus intervals were found to be strenuous for the
subjects, which likely impacted data quality. To obtain better data, improvements in sequence and stimulus designs
are needed to maximize the DIANA signal and minimize confounds. However, without a clear understanding of DIANA’s
biophysical underpinnings it is difficult to do so. Therefore, it may be more effective to first investigate DIANA signals
with simultaneously recorded electrophysiological signals in more controlled settings, e.g., in anesthetized mice.
Keywords: functional MRI, neuronal activity, DIANA, physiological noise, artifacts
1.
INTRODUCTION
The development of functional magnetic resonance
imaging (fMRI) in the 1990s revolutionized neuroscience,
offering a way to non- invasively map human brain func-
tion ( Bandettini, 2007; Logothetis, 2008; Ogawa et al.,
1990). Yet, fMRI’s dependence on changes in the Blood
Oxygen Level Dependent (BOLD) signal as a surrogate
for neuronal activity limits its spatiotemporal specificity
Received: 14 July 2023 Accepted: 14 August 2023 Available Online: 18 August 2023
Imaging Neuroscience, Volume 1, 2023
https://doi.org/10.1162/imag_a_00013
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00013/2157533/imag_a_00013.pdf by guest on 07 September 2023
© 2023 Massachusetts Institute of Technology. Published under a Creative Commons Attribution 4.0 International (CC BY 4.0) license. Research ArticleS. Hodono, R. Rideaux, T. van Kerkoerle et al.
Imaging Neuroscience, Volume 1, 2023
( Kim & Ugurbil, 2003; Logothetis et al., 2001; Polimeni
et al., 2010; Turner, 2002; Zhao et al., 2004). In particular,
Gradient Recalled Echo (GRE)- BOLD signals are biased
toward the larger drainage veins near the pial surface, far
from the site of neuronal activity ( Turner, 2002). Combin-
ing Ultra- High field MRI with advanced techniques such
as Spin- Echo (SE)- BOLD ( Han et al., 2021; Koopmans &
Yacoub, 2019; Zhao et al., 2004) or Vascular Space Occu-
pancy (VASO) ( Huber et al., 2017; Yu et al., 2019) can shift
BOLD sensitivity towards the capillary network, closer to
the area of neuronal activity, but capillaries can also
respond to the activity of neurons that are relatively far
away ( Chen et al., 2011).
Optical measurements of neuronally driven hemody-
namics suggest the spatial specificity of fMRI has not yet
reached the physiological limit ( Drew et al., 2011;
Hillman, 2014; Sirotin et al., 2009); however, fMRI’s big-
gest limitation may be its temporal specificity. In small
animals, it has been demonstrated that BOLD fMRI can
reveal some aspects of information flow within the cor-
tex ( Jung et al., 2021; Silva & Koretsky, 2002; Yu et al.,
2014). In 2002, Silva and Koretsky implemented very
high temporal resolution fMRI by swapping the phase
encoding and measurement loops, and reported that
BOLD fMRI can differentiate the response onset time
within the cortex. Yu et al. (2014) implemented line scan-
ning fMRI which also demonstrated that fMRI response
onset coincides with neuronal inputs. In 2021, Jung et al.
revealed information flow in the somatosensory network
by analyzing BOLD response onset time. Although sur-
prisingly responsive ( Hodono, Polimeni, Reutens, et al.,
2022; Lewis et al., 2016), the hemodynamic response
function is sluggish compared to the rapid fluctuations in
activity observed at the level of neurons ( Friston et al.,
1994). Invasive techniques in animal models have already
indicated that many processes in the brain are so fast
and confined to such small areas that they would not be
accessible with BOLD fMRI ( Siegel et al., 2015; Steinmetz
et al., 2019). Animal studies contribute useful knowl-
edge; however, in the pursuit of understanding the
human brain, non- invasive neuroimaging techniques are
essential. To this end, a non- invasive method that can
provide measurements of neuronal activity with high
spatiotemporal resolution would be a valuable tool.
Over the years, various approaches have been pro-
posed to enable more direct observations of neuronal acti-
vation through MRI ( Bihan et al., 2006; Patz et al., 2019;
Roth, 2023; Stanley & Raz, 2018; Yu et al., 2014). However,
each of these methods has key limitations. Diffusion-
weighted fMRI ( Bihan et al., 2006) is easily overshadowed
by BOLD effects ( Hodono, Polimeni, & Cloos, 2022; Miller
et al., 2007), and specific absorption rate (SAR) and
peripheral nerve stimulation considerations make mea-
surements with sub- second temporal resolution difficult.
Functional MR spectroscopy ( Stanley & Raz, 2018)
observes activation- related variations in metabolite con-
centrations. This is also subject to experimental consider-
ations such as SAR and signal- to- noise ratio (SNR), which
limit its temporal specificity to ~1 s. Elastography- based
functional MRI ( Patz et al., 2019) provides access to high-
frequency neuronal activity, but its model- based recon-
struction, relying on spatial derivatives of the signal, makes
it difficult to obtain a high degree of spatial specificity.
Recently, Toi et al. (2022) further extended the limits of
fMRI spatiotemporal specificity by introducing a new MRI
method that aims to enable Direct Imaging of Neuronal
Activity (DIANA). In their work, they employed Silva and
Koretsky’s acquisition strategy ( Silva & Koretsky, 2002),
and showed signal changes that closely followed electro-
physiological recordings and captured the thalamocorti-
cal pathway.
To resolve the mystery of the human mind, non-
invasive techniques that provide high spatiotemporal
specificity would be instrumental. DIANA may be able to
fulfill this role if it can be translated from animals to human
experiments ( Kerkoerle & Cloos, 2022). Here, we describe
our initial experience attempting to observe neuronal
activation in humans using DIANA.
2. METHOD
2.1. Simulations
The DIANA method is based on a Spoiled Gradient
Recalled Echo (SPGRE) sequence. Depending on the
exact SPGRE sequence parameters, it can take many
repetitions for the magnetization to stabilize. Full Bloch
simulations were performed using different T1 values to
estimate the number of dummy pulses needed to reach
the steady state, assuming sequence parameters from
the original DIANA paper (TR = 5 ms, flip angle (FA) = 4˚
( Toi et al., 2022)). The MATLAB (MathWorks, USA) code
used in these simulations can be found at https://
bitbucket . org / shotahodono / diana _ spgre _ sim.
2.2. DIANA sequence implementation and setup
Our implementation of the DIANA sequence was based
on a product SPGRE sequence using 50˚ quadratic phase
increments. As originally proposed by Silva and Koretsky
(2002), the DIANA sequence swaps the phase and
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00013/2157533/imag_a_00013.pdf by guest on 07 September 2023
2
S. Hodono, R. Rideaux, T. van Kerkoerle et al.
Imaging Neuroscience, Volume 1, 2023
temporal
resolutions
measurement loops. Combined with a synchronized
repetitive functional paradigm, it becomes possible to
obtain extremely high
(e.g.,
TR = 5 ms), especially when used to image a single slice.
Under these conditions, each trial samples the same line
in k- space M times once for each image in the final time
series. Trials are then repeated N times (number of phase
encoding lines), each adding one line to the time series.
Thus, collectively it takes at least M × N × TR to collect
one fully sampled dataset, hereafter referred to as “run.”
To ensure a stable baseline signal, an option was
added to enable sufficient dummy pulses to reach the
steady state, and trigger signals were added to synchro-
nize the acquisition and functional paradigm. All DIANA
experiments were performed with 2 × 2 mm2 or 1 × 1 mm2
in- plane resolution and a 5 mm slice thickness (TR = 5 ms,
TE = 2.4 ms, and FA = 4˚) at 7 Tesla (Siemens Magnetom,
Germany) using a 32- channel head coil (Nova Medical,
USA). The exact sequence implementation including gra-
dient amplitudes can be found in Supplementary Figure 1.
2.3. Phantom experiments
Phantom experiments were performed to evaluate the
stability of the MRI signal. After allowing fluid motion to
settle (~30 min), a custom phantom containing 50 mL
centrifuge tubes with different concentrations of manga-
nese chloride was imaged using both the SPGRE and
DIANA sequences. The protocol was chosen such that
both measurements produced an equal number of read-
outs. The SPGRE collected 1024 sequential images in 2
runs, with the first 21 measurements (~2000 TR) removed
to ensure that the SPGRE signal reached the steady
state. Two scans of 4 runs of DIANA measurements were
collected with 700 ms trials, using 2000 dummy TRs to
stabilize the signal at the start.
2.4. DIANA paradigms
Four different paradigms were tested, each in a different
scan session (summarized in Table 1). Data for paradigms
Table 1. Paradigm configurations and numbers of subjects.
I– V were collected using a single oblique axial slice cen-
tered on the calcarine sulcus (Fig. 1).
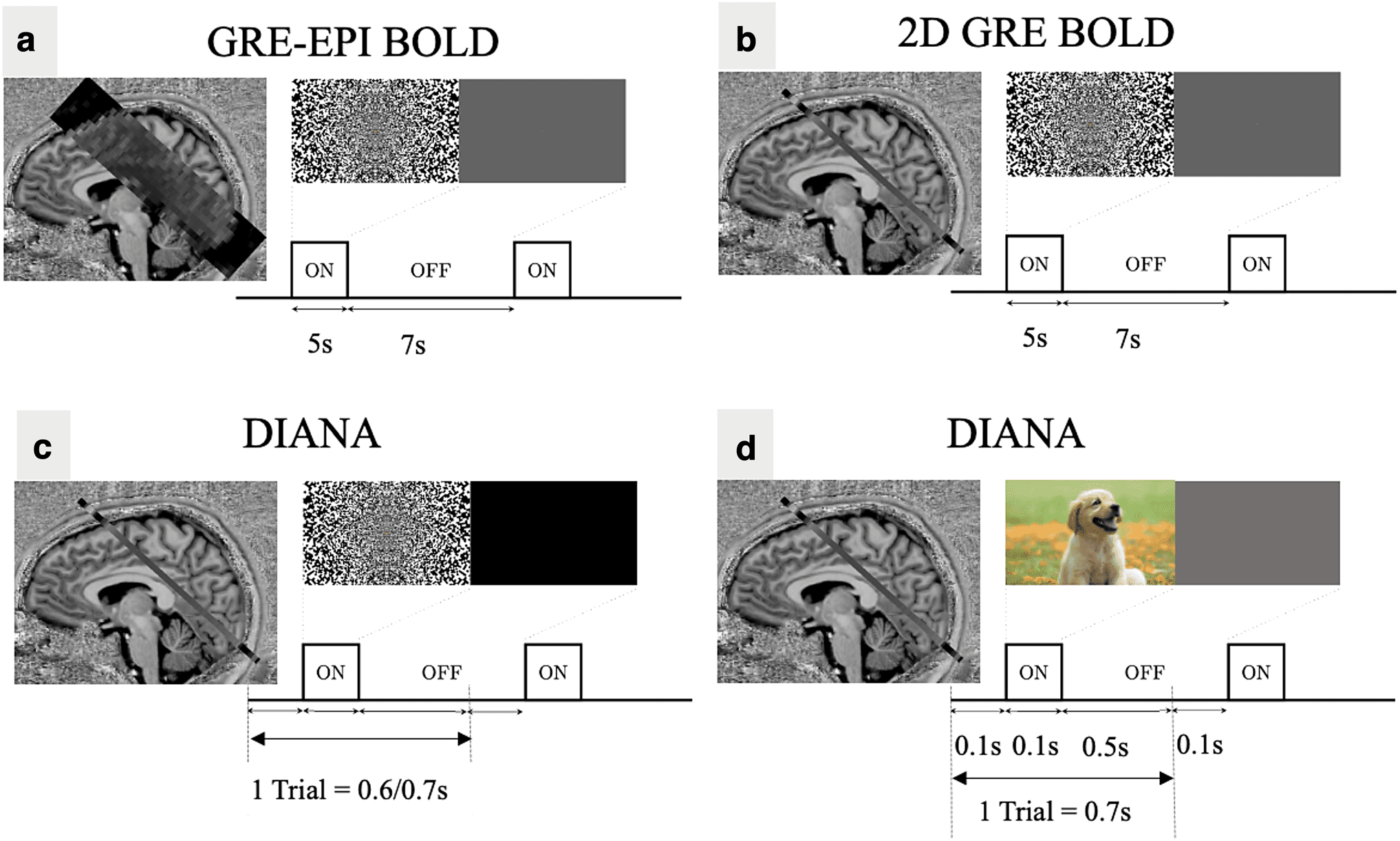
Each trial in the DIANA paradigm consisted of a 50–
200 ms visual stimulus with a 500– 600 ms interstimulus
interval (ISI) (Table 1). Visual stimuli were either pseudo-
randomly configured noise patterns or naturalistic images
that changed configuration on each trial. During ISIs, a
blank (black/gray) screen was presented to minimize
visual stimulation, e.g., caused by blinking. Lights in the
scanner room were dimmed to reduce contrast between
ambient light and closed eyes. In total, 3 scans of 10 runs
were collected (1056 trials, 50 min of DIANA scan time per
subject) for Paradigm I, 3 scans of 10 runs for Paradigm II,
3 scans of 11 runs per subject for Paradigm III, and 4
scans of 11 runs per subject for Paradigms IV and V.
Three human adult males and two females (23– 40 yo)
participated in the experiments, having provided written
informed consent. All participants had either normal or
corrected- to- normal vision and were screened for MRI
contraindications prior to scanning. The study was
approved by the local human research ethics committee
in accordance with national guidelines.
2.5.
In vivo slice placement
The DIANA imaging slice was identified using a BOLD-
based GRE- EPI functional localizer (9 slices, TR = 1 s,
TE = 20 ms, FA = 60˚, 2 × 2 × 5 mm, 360 volumes). A visual
paradigm was used (5 s on, 7 s off; Fig. 1a) and quickly
analyzed using the Fourier- transform (along the measure-
ment dimension) to identify voxels that match the expected
spectral function based on the convolution of the canoni-
cal hemodynamic response function with the functional
paradigm ( Hodono, Polimeni, Reutens, et al., 2022).
2.6. Functional localization and ROI selection
In Paradigms III and IV, the target slice was also imaged
using a single slice SPGRE sequence (TR = 31 ms,
TE = 20 ms, FA = 10˚, 2 × 2 × 5 mm, GRAPPA = 3, 360
volumes) to obtain a BOLD- based map of functional
Type
Stimulus
duration [ms]
ISI [ms]
Number
of subjects
Runs × scans
Resolution
Paradigm I
Paradigm II
Paradigm III
Paradigm IV
Paradigm V
Noise
Noise
Noise
Naturalistic images
Naturalistic images
50
200
100
100
100
1
1
3
3
3
11 × 3
10 × 3
11 × 3
11 × 4
11 × 4
2 × 2 × 5 mm3
2 × 2 × 5 mm3
2 × 2 × 5 mm3
2 × 2 × 5 mm3
1 × 1 × 5 mm3
550
500
600
600
600
3
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00013/2157533/imag_a_00013.pdf by guest on 07 September 2023
S. Hodono, R. Rideaux, T. van Kerkoerle et al.
Imaging Neuroscience, Volume 1, 2023
Fig. 1. Slice placement and paradigm designs for in vivo experiments. (a) GRE- EPI BOLD for slice placement in
Paradigms I, II, and III. (b) GRE BOLD for functional localization in Paradigm III. (c) DIANA acquisition in Paradigms I, II,
and III. (d) DIANA acquisition in Paradigm IV.
activity without geometric distortion (5 s on, 7 s off;
Fig. 1b). In Paradigm V, GRE BOLD was modified to
1 × 1 × 5 mm, keeping all other parameters the same. The
z- score maps were obtained through general linear mod-
eling analysis with FSL (https://fsl . fmrib . ox . ac . uk / fsl/).
Voxels with z- scores above 5 (in the general vicinity of V1)
were used to identify BOLD- based regions of interest
(ROI) where a DIANA signal may be expected.
Given the relatively low spatial resolution used in this
study (2 × 2 × 5 mm), reasonable voxel- wise coincidence
between BOLD and DIANA signals may be expected,
even though the GRE BOLD signal is weighted towards
the drainage veins at the surface ( Polimeni et al., 2010;
Turner, 2002). Nevertheless, we also collected a T1-
weighted image of the target slice using a 2D adaptation
of the MP2RAGE sequence ( Marques et al., 2010). These
images were used to manually draw anatomically informed
ROI using ITK- SNAP ( Yushkevich et al., 2006). Two control
ROIs were drawn in white matter and gray matter areas
where no activation is expected. We matched the number
of voxels in the gray matter control ROI to the manually
drawn anatomical ROI to equate the statistical power.
2.7. DIANA acquisition
In Paradigms I to IV, DIANA acquisition was performed
with the following parameters: voxel size = 2 × 2 × 5 mm3,
matrix size = 96 × 96, no parallel imaging, TR = 5 ms,
TE = 2.4 ms, FA = 4˚, and readout bandwidth = 650 Hz/
pixel. In Paradigm V, DIANA was acquired with a higher
in- plane resolution, 1 × 1 mm2 with 5 mm slice thickness,
matrix size = 192 × 192, TE = 2.3 ms, TR = 5 ms, FA = 4˚,
and readout bandwidth = 810 Hz/pixel. A GRAPPA factor
of 2 was used in Paradigm V so that the scan time per run
remained the same, with the aim of maintaining a similar
level of motion sensitivity. The sequence diagrams,
including the actual gradient amplitudes, are shown in
Supplementary Figure 1.
2.8.
Image reconstruction & data analysis
All DIANA data were reconstructed offline with a 16- bit
dynamic range (MATLAB, MathWorks, USA). After the
reconstruction, 2D in- plane motion correction was per-
formed. First, a reference image was computed by aver-
aging one of the runs. Then, individual images were
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00013/2157533/imag_a_00013.pdf by guest on 07 September 2023
4
S. Hodono, R. Rideaux, T. van Kerkoerle et al.
Imaging Neuroscience, Volume 1, 2023
coregistered to the reference image using 2D rigid
transformation (https://bitbucket . org / shotahodono / diana
_ coregi). The raw signal measured in each voxel for every
trial was first converted to a percent signal change as a
function of time, then linearly detrended and smoothed
with a gaussian kernel (width = 3 time points). Percent
signal change was then averaged across the ROI. Mean
percent signal change and 95% confidence intervals
were computed across the runs. Group means and the
confidence intervals were computed over all runs from all
subjects. The mean temporal SNR (tSNR) value in each
ROI was calculated for all paradigms (Supplementary
Table 1). The tSNR was calculated for each voxel based
on the mean signal (120 time points for Paradigm I and
140 time points for Paradigms II, III, IV, and V) divided by
the standard deviation.
3. RESULTS AND DISCUSSION
3.1. Simulations
~67 s to complete a single DIANA experiment with a
96 × 96 matrix and 140 time points (N = 96, M = 140), it is
considerably more efficient to complete multiple DIANA
measurements per scan. In our experiments, we ran 11
runs per scan (~12.5 min). Note that if a small number of
N and M are used (e.g., when using a shorter ISI), the
center of k- space can be reached before the magnetiza-
tion settles into the steady state.
3.2. Phantom experiments
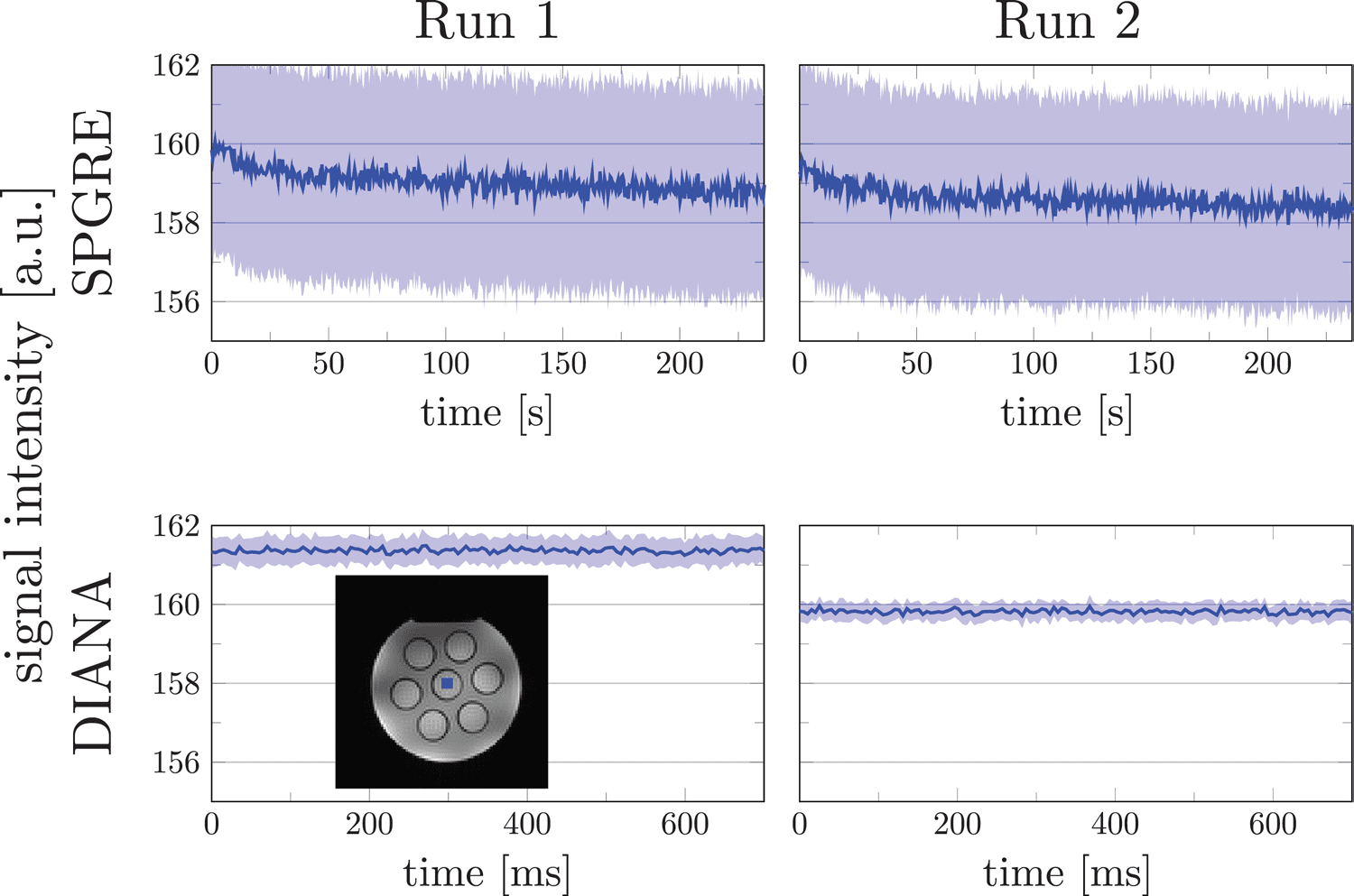
The mean signal in each image is dominated by the cen-
ter for k- space. The standard SPGRE sequence passes
through the center of k- space once every N × TR, such
that subsequent measurements directly reflect scanner
drift (Fig. 2). Interestingly, although simulations suggested
that 2000 dummy TR were adequate to reach the steady
state, some samples still showed signs of a residual tran-
sient. Therefore, in the following DIANA analyses, we dis-
carded the first run from each scan, in addition to the
initial 2000 dummy TRs.
Bloch simulations indicated that 1000 to 1500 dummy
pulses were needed to reach the steady state for all tis-
sues (Supplementary Fig. 2). With a 5 ms TR, this trans-
lates to 10 s of dummy pulses. Given that it requires
The DIANA sequence rapidly passes through the cen-
ter of k- space for all time points in one trial, but then will
not revisit the k- space center until N × M × TR later. Con-
sequently, signal drift now presents itself as a step
Fig. 2. Phantom measurements showing the effect of scanner drift on traditional SPGRE and DIANA sequences. The top
panels show two sequentially collected traditional SPGRE measurements, indicating a gradual drift in SPGRE. The bottom
panels show two sequentially collected DIANA measurements, indicating a step change. Both measurements comprise
an equal number of readouts. Blue shaded areas indicate 95% confidence intervals across voxels for SPGRE and across
runs and voxels for DIANA.
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00013/2157533/imag_a_00013.pdf by guest on 07 September 2023
5
S. Hodono, R. Rideaux, T. van Kerkoerle et al.
Imaging Neuroscience, Volume 1, 2023
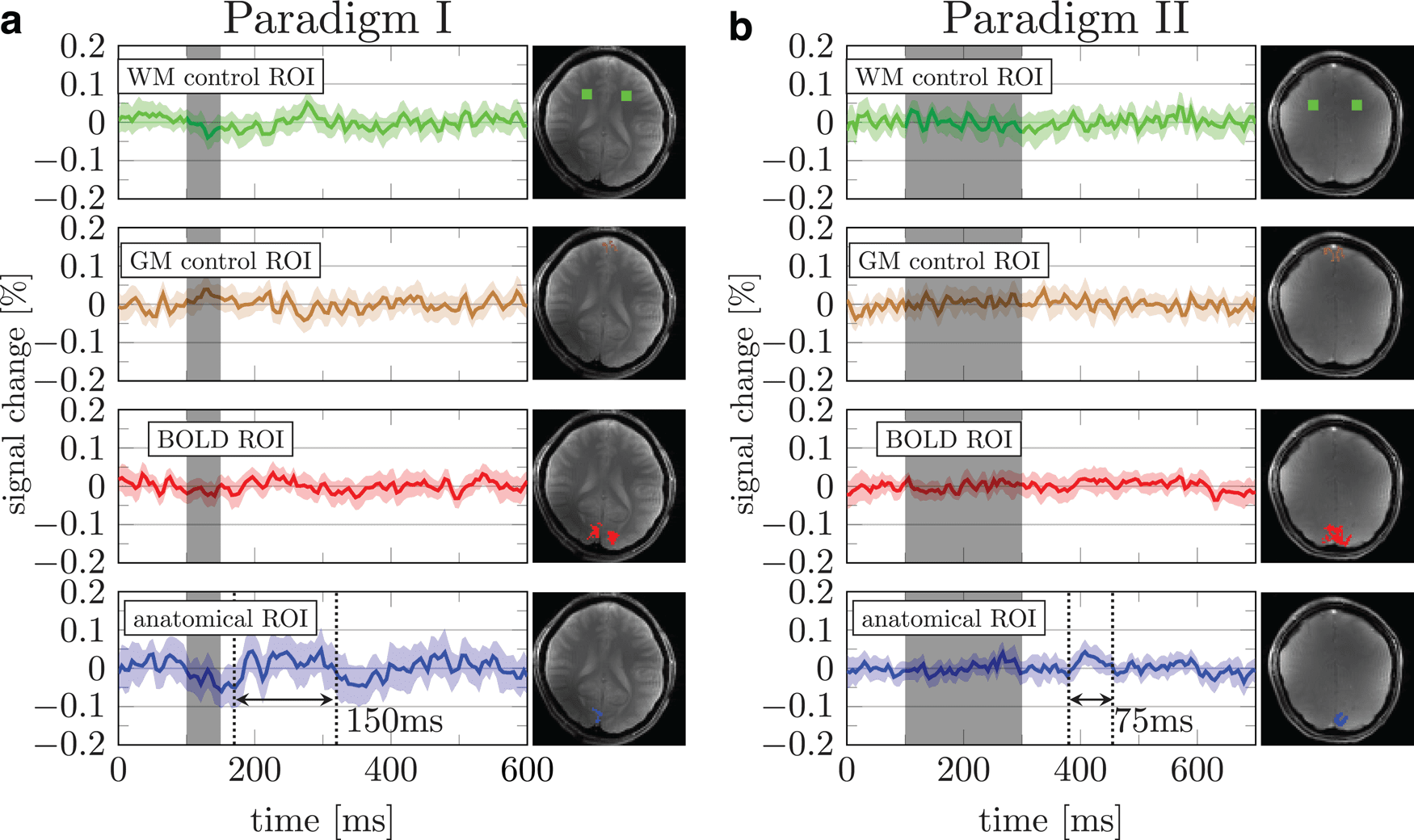
Fig. 3. Mean response over runs using paradigms I & II. (a & b) Green, brown, red, and blue data represent signal
changes from white matter control, gray matter control, BOLD based, and manually drawn anatomical ROIs, respectively.
Shaded areas indicate 95% confidence interval across runs from the subject.
function every N × M × TR (Fig. 2). Importantly, because
each trial is individually normalized based on its mean
signal before averaging, these drifts are effectively elimi-
nated from the DIANA analysis. Confidence intervals for
SPGRE acquisition were calculated across voxels within
ROI. For the DIANA acquisition, the mean signal over the
ROI was first calculated and confidence intervals were
calculated across runs. The different procedures resulted
in wider confidence intervals in SPGRE acquisition.
3.3. Paradigms I & II
The DIANA signal obtained with Paradigm I showed
some promise in an anatomically defined V1 ROI (Fig. 3a,
blue). A 0.05% increase in signal, approximately half that
observed in mice ( Toi et al., 2022), followed the stimulus
onset by ~75 ms. This signal increase persisted for
~150 ms. The tSNR per voxel in the ROI was >1000, and
the tSNR in the ROI is expected to be increased by the
square root of the number of voxels (Supplementary
Table 1). Therefore, the 0.05% signal change may be
detectable, albeit close to the detection limit.
Unlike the mice experiments of Toi et al., no electro-
physiological recordings were available for comparison.
Therefore, we devised a different test in which we
increased the stimulus duration from 50 to 200 ms in Par-
adigm II. While some localized areas showed a peak in
the time averaged signal (0.05%) after the stimulus
(Fig. 3b, blue), the duration of the putative signal peak
was reduced. If the change in activity was related to the
stimulus, one would expect to observe a prolonged peak
exceeding the duration of that observed in Paradigm I
(150 ms) ( Mirpour & Esteky, 2009). It should also be noted
that the onset of the putative signal peak in Paradigm II is
inconsistent with the temporal dynamics of the visual
processing cascade. That is, the peak occurs ~300 ms
after the stimulus onset, whereas responses in the early
visual cortex are evoked between 50 and 100 ms after
the stimulus onset ( Ringach et al., 2003). It may be that
the specific anatomical ROI shown in Paradigms I and II
included systaltic artifacts from large vessels (Supple-
mentary Figs. 3– 5).
3.4. Paradigm III
The results of Paradigms I and II suggest spurious results
may be found, possibly due to limited signal averaging
power and poorly defined ROI. Thus, we next sought to
collect more data across different participants. Further, in
the following set of experiments, we also collected
distortion matched SPGRE- BOLD and T1- weighted ana-
tomical data to improve the ROI definition.
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00013/2157533/imag_a_00013.pdf by guest on 07 September 2023
6
S. Hodono, R. Rideaux, T. van Kerkoerle et al.
Imaging Neuroscience, Volume 1, 2023
Fig. 4. Exemplary functional and anatomical localization data from an oblique axial experiment. The left panel shows
the activation estimate based on the Fourier analysis of GRE- EPI BOLD images. The middle panel shows a z- score map
obtained using single slice SPGRE- BOLD. The right panel shows the T1- weighted anatomical matching the target slice.
The bottom panel shows the z- score (>5) derived activation superimposed on the 2D adaptation of the MP2RAGE.
All participants showed clear stimulus- related activa-
tion across the visual cortex in the EPI- based functional
localizer (Fig. 4, left). In all cases, the target slice also
showed clear activation in the single slice distortion- free
2D GRE data (Fig. 4, middle), matching the expected
anatomy.
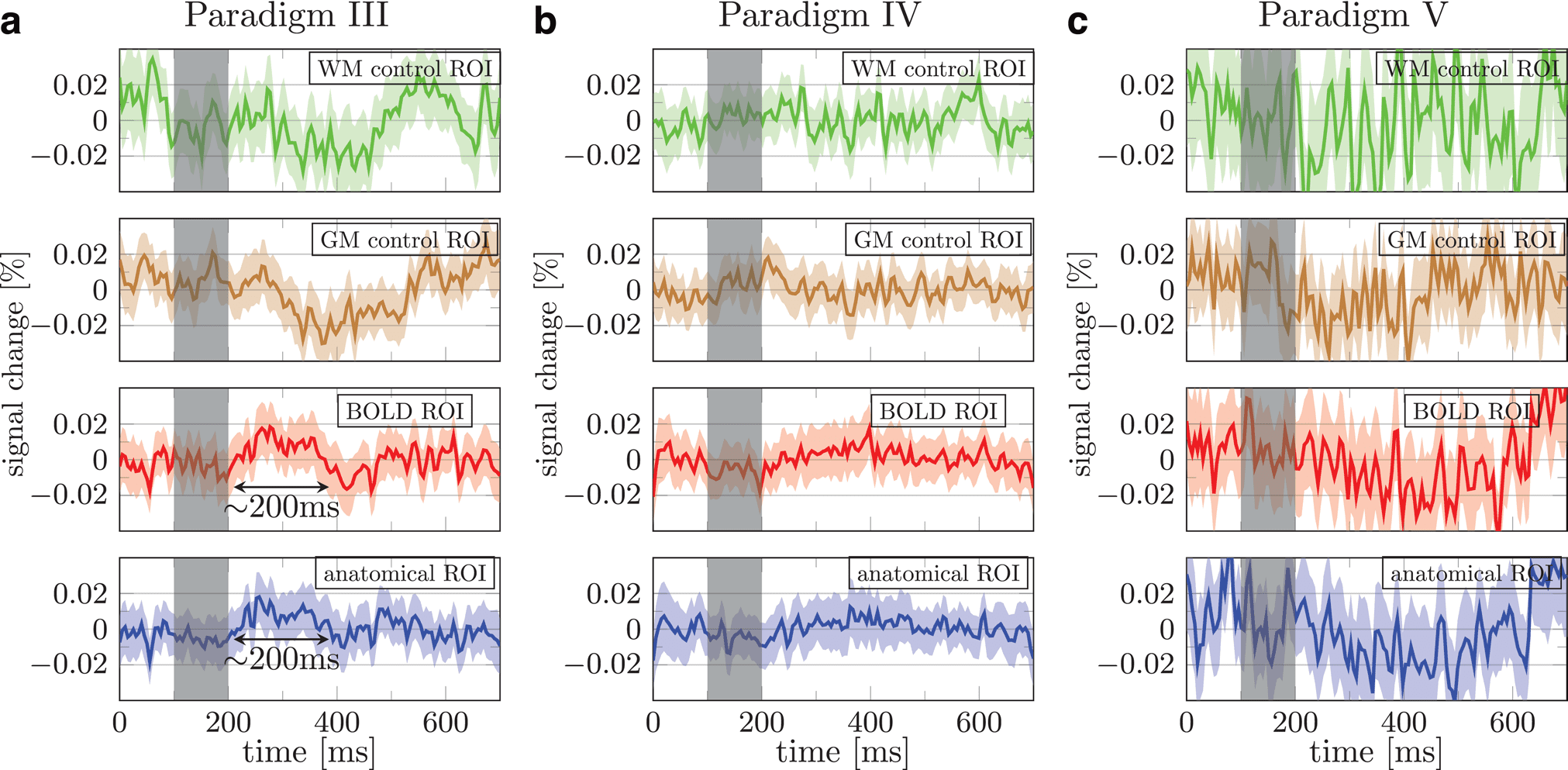
When averaged across BOLD or anatomical ROI, trial
averaged DIANA data showed no signs of activation in
individual subjects (Supplementary Fig. 6, the tSNR per
voxel and number of voxels in the ROIs are seen in Sup-
plementary Table 1). When averaged across all three sub-
jects, data from BOLD and anatomical ROIs showed a
~200 ms long 0.02% signal increase starting ~100 ms
after the stimulus onset (Fig. 5a). However, the observed
signal percent change was an order of magnitude smaller
than what was reported in mice ( Toi et al., 2022). More-
over, data from control ROI also showed signal changes
exceeding 0.02%. This suggests that the signal change
observed in V1 may not reflect neuronal activity.
Volunteers reported that the noise- like checkerboard
stimuli with ISI ~600 ms were intense, bordering uncom-
fortable, motivating them to blink in anticipation. In gen-
eral, blinking may be a confound in these visual paradigms
using rapid stimulus presentation. A typical eye blink
lasts ~100 ms, creating a visual contrast change in simi-
lar duration to the target stimulus. Although blinking
could be detected using an eye tracker, data scrubbing
may be difficult because DIANA inherently averages trials
during image reconstruction.
3.5. Paradigm IV
Based on our experiences in Paradigms I– III, we sur-
mised that tailored naturalistic images, which are more
engaging and comfortable to view, may be a superior
visual stimulus for the experiment.
The participants indeed reported that the naturalistic
images were more comfortable, making it easier to remain
attentive, allowing us to obtain 4 scans per subject (44
runs per subject). Nevertheless, the responses averaged
over three subjects show no convincing evidence of a
DIANA signal (Fig. 5b; data for individual subjects are
provided in Supplementary Figure 7, the tSNR per voxel
and number of voxels in the ROIs are seen in Supplemen-
tary Table 1).
We were unable to reproduce the positive signal
change observed in Paradigm III when averaged across
three subjects. It is possible that the natural stimuli pro-
duced less activation than the noise pattern used in Par-
adigm III. However, considering the scale of the signal, it
is also possible that the increased activation found in
Paradigm III was an artifact or spurious.
3.6. Paradigm V
In Paradigms I– IV, we employed in- plane resolution of
2 × 2 mm2. Despite care taken when we drew the ana-
tomical ROIs, the relatively large voxel size is prone to
partial volume effects. In Paradigm V, we used a finer
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00013/2157533/imag_a_00013.pdf by guest on 07 September 2023
7
S. Hodono, R. Rideaux, T. van Kerkoerle et al.
Imaging Neuroscience, Volume 1, 2023
Fig. 5. Mean signal percent change averaged across all three subjects in paradigms III (a), IV (b), and V (c). The
green, brown, red, and blue data show the trial averaged signal obtained using the white matter control, gray matter
control, BOLD based, and anatomical ROIs, respectively. Shaded areas show 95% confidence interval across runs
from three subjects (90 runs for (a) and 120 runs for (b) and (c)). Individual subjects’ results and the ROIs are provided in
Supplementary Figures 6– 8.
in- plane resolution, 1 × 1 mm2, to reduce partial volume
effects and improve the ROI (Supplementary Fig. 8).
Although the tSNR per voxel was smaller than in Para-
digms I– IV, each ROI contained a larger number of vox-
els, partially compensating for the reduced tSNR and per
voxel (Supplementary Table 1). Nevertheless, no sign of
neuronal activity was found in individual subjects (Sup-
plementary Fig. 8) or averaged across 3 subjects (Fig. 5).
3.7. Recommendations for future experiments
If DIANA is possible with MRI, using the method to study
human brains will require establishing an effective para-
digm. In addition to visual stimuli, we also used auditory
stimuli, which were more comfortable for the subject and
could benefit from the high temporal specificity of the
auditory system ( Gazzaniga, 2000; Zanker & Harris,
2002), but ultimately failed to produce a reliable DIANA
signal (data not shown). However, in the auditory experi-
ment, the BOLD response was also notably weaker, pre-
sumably due to the auditory noise produced by the
scanner itself, which may be even more problematic
when using short ISIs. Finger tapping may be an effective
alternative. However, care should be taken not to inad-
vertently induce head motion.
The expected percent change in DIANA experiments is
very small (<0.5%). For individual subject analysis with rea-
sonable scan times, such small signal changes are at the
edge of the detectability. Care should be taken not to mis-
take small artifacts for DIANA signals (Supplementary
Note). In animal imaging, the validity of such candidate sig-
nals can be tested by comparison against simultaneously
recorded intracranial electrophysiological signals. However,
such recordings are generally not feasible in humans. As an
alternative, candidate signals can be tested by changing
the experimental paradigm. In Paradigms I and II, we
changed the stimulus duration. It is expected that longer
stimulus block alters the response dynamics accordingly if
it is neuronal. Alternatively, one could also add additional
stimuli within a single trial, with the expectation that this will
produce an additional signal in the trial averaged response.
In the original DIANA experiment, anesthetized mice
were used. Therefore, motion was not a major issue. How-
ever, in humans, motion is likely to occur even when scan-
ning compliant participants. Although in- plane motion can
be corrected to some extent, the single slice imaging makes
retrospective through- plane motion correction impossible.
Real time motion correction ( Lee et al., 1996; Maclaren
et al., 2013; White et al., 2010) could help, but adding nav-
igators may change the repetition time. An external tracking
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00013/2157533/imag_a_00013.pdf by guest on 07 September 2023
8
S. Hodono, R. Rideaux, T. van Kerkoerle et al.
Imaging Neuroscience, Volume 1, 2023
device may be a solution ( Zaitsev et al., 2006). In addition,
physiological noise contribution is also concerned (Supple-
mentary Note); as shown in Supplementary Figure 3, there
is an inflow effect. By introducing a saturation module,
such artifacts may be mitigated. However, separate satura-
tion modules lengthen the TR significantly. A clever pulse
design may help reduce motion artifacts while also sup-
pressing potential inflow effects ( Hodono et al., 2023).
One other challenge is spatial resolution. In Paradigms
I– IV, we employed 2 × 2 × 5 mm3 resolution to have suffi-
cient SNR to detect the order of 0.1% signal change and to
mitigate through- plane motion. However, such relatively
large voxels may contain some amount of white matter or
cerebral spinal fluid (CSF). Combined with minute motions
and brain pulsations, such partial voluming effect can
decrease the effective tSNR. However, high resolution
acquisition will reduce the absolute SNR and require more
phase encoding lines, which increases motion sensitivity. To
avoid increased motion sensitivity, we employed an accel-
eration factor of 2, to obtain 1 × 1 × 5 mm3 resolution with-
out acquiring additional phase encoding lines (Paradigm V).
Even finer resolution might be feasible with higher accelera-
tion factors. However, the SNR will drop further and g-factor
noise amplification will increase (Supplementary Figure 4),
which could impede the observation of DIANA responses
within individual subjects and reasonable scan times.
Despite these challenges, even finer resolution might
be required to resolve spatially confined neuronal signals.
It can be argued that the detection of fast and transient
neuronal signals necessitates not only a high temporal
resolution, but also a high spatial resolution. In electro-
physiology, it is commonly found that signals with high
temporal frequency are coherent across smaller volumes
of tissue than signals with low temporal frequencies
( Bullock, 1997), and therefore also require electrodes
with smaller surface areas to measure reliably ( Sindhu
et al., 2023; Worrell et al., 2008). In the extreme case,
reliably recording single units, which are signals with a
main frequency component around 1 kHz, requires an
electrode with a diameter similar to that of the soma of a
single neuron ( Starr et al., 1973; Viswam et al., 2019). As
a historical note, single unit electrophysiology, and
thereby systems neuroscience ( Hubel, 1982), can be said
to only really have taken off with the development of a
method to make electrodes that were both rigid and thin
enough to allow contact sites of a few micrometers
( Hubel, 1915). It could be that measuring fast transients
with MRI requires a similar methodological advance.
Interestingly, a recent preprint highlights difficulties
reproducing the original work in mice at very high spatio-
temporal resolution at 15 Tesla ( Choi et al., 2023), raising
questions about the reliability and even validity of the
DIANA method. An important hurdle to resolve this
debate is that our current understanding of DIANA’s bio-
physical underpinning is still very limited. Without more
detailed knowledge of the biophysical underpinning of
the DIANA signal, it will be difficult to make an informed
decision regarding the sequence parameterization and
experimental paradigm needed to reproduce and opti-
mize DIANA, first of all in rodents and then possibly in
human imaging. To bridge this knowledge gap and in the
search for non- hemodynamic fMRI signals more broadly,
we believe that it may be necessary to run dedicated
studies with setups that are both highly controlled and
biologically representative ( Morrison et al., 2023).
DATA AND CODE AVAILABILITY
The data and code used for analysis are publicly avail-
able at https://osf . io / x3yab/.
AUTHOR CONTRIBUTIONS
All authors contributed to the designed experiments,
data interpretation, and writing of the paper. S.H., R.R.,
and M.A.C. acquired and analyzed the data. M.A.C.
supervised the work.
DECLARATION OF COMPETING INTEREST
None.
ACKNOWLEDGMENTS
This work was supported by the young investigator award
from the French National Research Agency (ANR- 19-
CE37- 003- 01), the Australian government through the
Australian Research Council (ARC) Future fellowship
grant FT200100329, ARC Discovery Early Career
Researcher Award (DE210100790), and ARC Centre
grant (IC170100035). The authors acknowledge the facil-
ities of the National Imaging Facility at the Centre for
Advanced Imaging. They thank Chia- Yin Wu for help
during the experiments.
SUPPLEMENTARY MATERIALS
Supplementary material for this article is available with
the online version here: https://doi . org / 10 . 1162 / imag _ a
_ 00013.
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00013/2157533/imag_a_00013.pdf by guest on 07 September 2023
9
S. Hodono, R. Rideaux, T. van Kerkoerle et al.
Imaging Neuroscience, Volume 1, 2023
REFERENCES
Bandettini, P. (2007). Functional MRI today. Int J
Psychophysiol, 63, 138–145. https://doi . org / 10 . 1016 / j
. ijpsycho . 2006 . 03 . 016
Bihan, D. L., Urayama, S., Aso, T., Hanakawa, T., &
Fukuyama, H. (2006). Direct and fast detection of
neuronal activation in the human brain with diffusion
MRI. Proc Natl Acad Sci U S A, 103, 8263–8268. https://
doi . org / 10 . 1073 / pnas . 0600644103
Bullock, T. H. (1997). Signals and signs in the nervous
system: The dynamic anatomy of electrical activity is
probably information- rich. Proc Natl Acad Sci U S A, 94,
1–6. https://doi . org / 10 . 1073 / pnas . 94 . 1 . 1
Chen, B. R., Bouchard, M. B., McCaslin, A. F. H., Burgess,
S. A., & Hillman, E. M. C. (2011). High- speed vascular
dynamics of the hemodynamic response. Neuroimage,
54, 1021–1030. https://doi . org / 10 . 1016 / j . neuroimage
. 2010 . 09 . 036
Choi, S.- H., Im, G. H., Choi, S., Yu, X., Bandettini, P. A.,
Menon, R. S., & Kim, S.- G. (2023). No replication
of direct neuronal activity- related (DIANA) fMRI in
anesthetized mice. bioRxiv, 2023.05.26.542419. https://
doi.org/10.1101/2023.05.26.542419
Drew, P. J., Shih, A. Y., & Kleinfeld, D. (2011). Fluctuating
and sensory- induced vasodynamics in rodent cortex
extend arteriole capacity. Proc Natl Acad Sci U S A, 108,
8473–8478. https://doi . org / 10 . 1073 / pnas . 1100428108
Friston, K. J., Jezzard, P., & Turner, R. (1994). Analysis of
functional MRI time‐series. Hum Brain Mapp, 1, 153–
171. https://doi . org / 10 . 1002 / hbm . 460010207
Gazzaniga, M. S. (2000). The new cognitive neurosciences.
MIT press. ISBN 9780262071956.
Han, S., Eun, S., Cho, H., Uludaǧ, K., & Kim, S.- G. (2021).
Improvement of sensitivity and specificity for laminar
BOLD fMRI with double spin- echo EPI in humans at 7
T. Neuroimage, 241, 118435. https://doi . org / 10 . 1016 / j
. neuroimage . 2021 . 118435
Hillman, E. M. C. (2014). Coupling mechanism and
significance of the BOLD signal: A status report.
Neuroscience, 37, 161–181. https://doi . org / 10 . 1146
/ annurev - neuro - 071013 - 014111
Hodono, S., Polimeni, J., & Cloos, M. (2022). Detection
of fast responses in diffusion fMRI of the human visual
cortex through reduced vascular contamination.
Proceedings of the Joint Annual Meeting of ISMRM-
ESMRMB, London, United Kingdom; 2115. https://
archive.ismrm.org/2022/2115.html
Hodono, S., Polimeni, J., Reutens, D., & Cloos, M. (2022).
Tracking rapid stimulus- driven BOLD oscillations in the
human primary motor cortex and somatosensory cortex.
Proceedings of the Joint Annual Meeting of ISMRM-
ESMRMB, London, United Kingdom; 0526. https://
archive.ismrm.org/2022/0526.html
Hodono, S., Wu, C., Dixon, C., MAillet, D., Jin, J., Polimeni,
J., & Cloos, M. (2023). Simultaneous saturation and
excitation (SatEx) pulse. Proceedings of the Annual
Meeting of ISMRM, Toronto, Canada; 4564.
Hubel, D. H. (1915). Tungsten microelectrode for recording
from single units. Science, 125, 549–550. https://doi . org
/ 10 . 1126 / science . 125 . 3247 . 549
Hubel, D. H. (1982). Evolution of ideas on the primary visual
cortex, 1955– 1978: A biased historical account. Biosci
Rep, 2, 435–469. https://doi . org / 10 . 1007 / bf01115245
Huber, L., Handwerker, D. A., Jangraw, D. C., Chen,
G., Hall, A., Stüber, C., Gonzalez- Castillo, J., Ivanov,
D., Marrett, S., Guidi, M., Goense, J., Poser, B. A., &
Bandettini, P. A. (2017). High- resolution CBV- fMRI allows
mapping of laminar activity and connectivity of cortical
input and output in human M1. Neuron, 96, 1253.e7–
1263.e7. https://doi . org / 10 . 1016 / j . neuron . 2017 . 11 . 005
Jung, W. B., Im, G. H., Jiang, H., & Kim, S.- G. (2021). Early
fMRI responses to somatosensory and optogenetic
stimulation reflect neural information flow. Proc Natl
Acad Sci U S A, 118, e2023265118. https://doi . org / 10
. 1073 / pnas . 2023265118
Kerkoerle, T. van, & Cloos, M. A. (2022). Creating a window
into the mind. Science, 378, 139–140. https://doi . org / 10
. 1126 / science . ade4938
Kim, S.- G., & Ugurbil, K. (2003). High- resolution functional
magnetic resonance imaging of the animal brain.
Methods, 30, 28–41. https://doi . org / 10 . 1016 / s1046
- 2023(03)00005 - 7
Koopmans, P. J., & Yacoub, E. (2019). Strategies and
prospects for cortical depth dependent T2 and T2*
weighted BOLD fMRI studies. Neuroimage, 197, 668–
676. https://doi . org / 10 . 1016 / j . neuroimage . 2019 . 03 . 024
Lee, C. C., Jack, C. R., Grimm, R. C., Rossman, P. J.,
Felmlee, J. P., Ehman, R. L., & Riederer, S. J. (1996).
Real‐time adaptive motion correction in functional MRI.
Magn Reson Med, 36, 436–444. https://doi . org / 10 . 1002
/ mrm . 1910360316
Lewis, L. D., Setsompop, K., Rosen, B. R., & Polimeni, J. R.
(2016). Fast fMRI can detect oscillatory neural activity in
humans. Proc Natl Acad Sci U S A, 113, E6679–E6685.
https://doi . org / 10 . 1073 / pnas . 1608117113
Logothetis, N. K. (2008). What we can do and what we
cannot do with fMRI. Nature, 453, 869–878. https://doi
. org / 10 . 1038 / nature06976
Logothetis, N. K., Pauls, J., Augath, M., Trinath, T., &
Oeltermann, A. (2001). Neurophysiological investigation
of the basis of the fMRI signal. Nature, 412, 150–157.
https://doi . org / 10 . 1038 / 35084005
Maclaren, J., Herbst, M., Speck, O., & Zaitsev, M. (2013).
Prospective motion correction in brain imaging: A review.
Magn Reson Med, 69, 621–636. https://doi . org / 10 . 1002
/ mrm . 24314
Marques, J. P., Kober, T., Krueger, G., Zwaag, W., van der
Moortele, P.- F. V., & de Gruetter, R. (2010). MP2RAGE,
a self bias- field corrected sequence for improved
segmentation and T1- mapping at high field. Neuroimage,
49, 1271–1281. https://doi . org / 10 . 1016 / j . neuroimage
. 2009 . 10 . 002
Miller, K. L., Bulte, D. P., Devlin, H., Robson, M. D., Wise,
R. G., Woolrich, M. W., Jezzard, P., & Behrens, T. E. J.
(2007). Evidence for a vascular contribution to diffusion
FMRI at high b value. Proc Natl Acad Sci U S A, 104,
20967–20972. https://doi . org / 10 . 1073 / pnas . 0707257105
Mirpour, K., & Esteky, H. (2009). State- dependent effects of
stimulus presentation duration on the temporal dynamics
of neural responses in the inferotemporal cortex of
macaque monkeys. J Neurophysiol, 102, 1790–1800.
https://doi . org / 10 . 1152 / jn . 91197 . 2008
Morrison, S., Webber, E., Dixon, C., Maillet, D., Wolvetang,
E., & Cloos, M. (2023). Investigating the direct imaging
of neuronal activity (DIANA) contrast mechanism using
human brain organoids. Proceedings of the Annual
Meeting of ISMRM, Toronto, Canada; 0916.
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00013/2157533/imag_a_00013.pdf by guest on 07 September 2023
10
S. Hodono, R. Rideaux, T. van Kerkoerle et al.
Imaging Neuroscience, Volume 1, 2023
Ogawa, S., Lee, T., Nayak, A. S., & Glynn, P. (1990).
Oxygenation‐sensitive contrast in magnetic resonance
image of rodent brain at high magnetic fields. Magn
Reson Med, 14, 68–78. https://doi . org / 10 . 1002 / mrm
. 1910140108
Patz, S., Fovargue, D., Schregel, K., Nazari, N., Palotai,
M., Barbone, P. E., Fabry, B., Hammers, A., Holm, S.,
Kozerke, S., Nordsletten, D., & Sinkus, R. (2019). Imaging
localized neuronal activity at fast time scales through
biomechanics. Sci Adv, 5, eaav3816. https://doi . org / 10
. 1126 / sciadv . aav3816
Polimeni, J. R., Fischl, B., Greve, D. N., & Wald, L. L.
(2010). Laminar analysis of 7T BOLD using an imposed
spatial activation pattern in human V1. Neuroimage, 52,
1334–1346. https://doi . org / 10 . 1016 / j . neuroimage . 2010
. 05 . 005
Ringach, D. L., Hawken, M. J., & Shapley, R. (2003).
Dynamics of orientation tuning in macaque V1: The role
of global and tuned suppression. J Neurophysiol, 90,
342–352. https://doi . org / 10 . 1152 / jn . 01018 . 2002
Roth, B. J. (2023). Can MRI be used as a sensor to record
neural activity? Sensors (Basel), 23, 1337. https://doi . org
/ 10 . 3390 / s23031337
Siegel, M., Buschman, T. J., & Miller, E. K. (2015). Cortical
information flow during flexible sensorimotor decisions.
Science, 348, 1352–1355. https://doi . org / 10 . 1126
/ science . aab0551
Silva, A. C., & Koretsky, A. P. (2002). Laminar specificity
of functional MRI onset times during somatosensory
stimulation in rat. Proc Natl Acad Sci U S A, 99, 15182–
15187. https://doi . org / 10 . 1073 / pnas . 222561899
Sindhu, K. R., Ngo, D., Ombao, H., Olaya, J. E., Shrey,
D. W., & Lopour, B. A. (2023). A novel method for
dynamically altering the surface area of intracranial EEG
electrodes. J Neural Eng, 20, 026002. https://doi . org / 10
. 1088 / 1741 - 2552 / acb79f
Sirotin, Y. B., Hillman, E. M. C., Bordier, C., & Das, A.
(2009). Spatiotemporal precision and hemodynamic
mechanism of optical point spreads in alert primates.
Proc Natl Acad Sci U S A, 106, 18390–18395. https://doi
. org / 10 . 1073 / pnas . 0905509106
Stanley, J. A., & Raz, N. (2018). Functional magnetic
resonance spectroscopy: The “new” MRS for cognitive
neuroscience and psychiatry research. Front Psychiatry,
9, 76. https://doi . org / 10 . 3389 / fpsyt . 2018 . 00076
Starr, A., Wise, K. D., & Csongradi, J. (1973). An evaluation
of photoengraved microelectrodes for extracellular
single- unit recording. IEEE Trans Biomed Eng, 20,
291–293. https://doi . org / 10 . 1109 / tbme . 1973 . 324194
Steinmetz, N. A., Zatka- Haas, P., Carandini, M.,
& Harris, K. D. (2019). Distributed coding of choice,
action and engagement across the mouse brain.
Nature, 576, 266–273. https://doi . org / 10 . 1038 / s41586
- 019 - 1787 - x
Toi, P. T., Jang, H. J., Min, K., Kim, S.- P., Lee, S.- K., Lee,
J., Kwag, J., & Park, J.- Y. (2022). In vivo direct imaging
of neuronal activity at high temporospatial resolution.
Science, 378, 160–168. https://doi . org / 10 . 1126 / science
. abh4340
Turner, R. (2002). How much cortex can a vein drain?
Downstream dilution of activation- related cerebral blood
oxygenation changes. Neuroimage, 16, 1062–1067.
https://doi . org / 10 . 1006 / nimg . 2002 . 1082
Viswam, V., Obien, M. E. J., Franke, F., Frey, U., &
Hierlemann, A. (2019). Optimal electrode size for multi-
scale extracellular- potential recording from neuronal
assemblies. Front Neurosci, 13, 385. https://doi . org / 10
. 3389 / fnins . 2019 . 00385
White, N., Roddey, C., Shankaranarayanan, A., Han, E.,
Rettmann, D., Santos, J., Kuperman, J., & Dale, A.
(2010). PROMO: Real‐time prospective motion correction
in MRI using image‐based tracking. Magn Reson Med,
63, 91–105. https://doi . org / 10 . 1002 / mrm . 22176
Worrell, G. A., Gardner, A. B., Stead, S. M., Hu, S.,
Goerss, S., Cascino, G. J., Meyer, F. B., Marsh, R., &
Litt, B. (2008). High- frequency oscillations in human
temporal lobe: Simultaneous microwire and clinical
macroelectrode recordings. Brain, 131, 928–937. https://
doi . org / 10 . 1093 / brain / awn006
Yu, X., Qian, C., Chen, D., Dodd, S. J., & Koretsky, A. P.
(2014). Deciphering laminar- specific neural inputs with
line- scanning fMRI. Nat Methods, 11, 55–58. https://doi
. org / 10 . 1038 / nmeth . 2730
Yu, Y., Huber, L., Yang, J., Jangraw, D. C., Handwerker,
D. A., Molfese, P. J., Chen, G., Ejima, Y., Wu, J., &
Bandettini, P. A. (2019). Layer- specific activation of
sensory input and predictive feedback in the human
primary somatosensory cortex. Sci Adv, 5, eaav9053.
https://doi . org / 10 . 1126 / sciadv . aav9053
Yushkevich, P. A., Piven, J., Hazlett, H. C., Smith, R. G.,
Ho, S., Gee, J. C., & Gerig, G. (2006). User- guided 3D
active contour segmentation of anatomical structures:
Significantly improved efficiency and reliability.
Neuroimage, 31, 1116–1128. https://doi . org / 10 . 1016 / j
. neuroimage . 2006 . 01 . 015
Zaitsev, M., Dold, C., Sakas, G., Hennig, J., & Speck, O.
(2006). Magnetic resonance imaging of freely moving
objects: Prospective real- time motion correction using an
external optical motion tracking system. Neuroimage, 31,
1038–1050. https://doi . org / 10 . 1016 / j . neuroimage . 2006
. 01 . 039
Zanker, J. M., & Harris, J. P. (2002). On temporal hyperacuity
in the human visual system. Vision Res, 42, 2499–2508.
https://doi . org / 10 . 1016 / s0042 - 6989(02)00301 - 2
Zhao, F., Wang, P., & Kim, S. (2004). Cortical depth‐
dependent gradient‐echo and spin‐echo BOLD fMRI at
9.4T. Magn Reson Med, 51, 518–524. https://doi . org / 10
. 1002 / mrm . 10720
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00013/2157533/imag_a_00013.pdf by guest on 07 September 2023
11