Inferring Causality from Noninvasive Brain Stimulation in
Cognitive Neuroscience
Til Ole Bergmann1 and Gesa Hartwigsen2
Abstract
■ Noninvasive brain stimulation (NIBS) techniques, such as
transcranial magnetic stimulation or transcranial direct and al-
ternating current stimulation, are advocated as measures to en-
able causal inference in cognitive neuroscience experiments.
Transcending the limitations of purely correlative neuroimaging
measures and experimental sensory stimulation, they allow to
experimentally manipulate brain activity and study its conse-
quences for perception, cognition, and eventually, behavior.
Although this is true in principle, particular caution is advised
when interpreting brain stimulation experiments in a causal
manner. Research hypotheses are often oversimplified, disre-
garding the underlying (implicitly assumed) complex chain of
causation, namely, that the stimulation technique has to gener-
ate an electric field in the brain tissue, which then evokes or
modulates neuronal activity both locally in the target region
and in connected remote sites of the network, which in conse-
quence affects the cognitive function of interest and eventually
results in a change of the behavioral measure. Importantly, ev-
ery link in this causal chain of effects can be confounded by
several factors that have to be experimentally eliminated or con-
trolled to attribute the observed results to their assumed cause.
This is complicated by the fact that many of the mediating and
confounding variables are not directly observable and dose–
response relationships are often nonlinear. We will walk the
reader through the chain of causation for a generic cognitive
neuroscience NIBS study, discuss possible confounds, and ad-
vise appropriate control conditions. If crucial assumptions are
explicitly tested (where possible) and confounds are experi-
mentally well controlled, NIBS can indeed reveal cause–effect
relationships in cognitive neuroscience studies. ■
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
3
2
1
9
5
1
8
6
2
5
1
7
/
j
o
c
n
_
a
_
0
1
5
9
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
INTRODUCTION
Noninvasive brain stimulation (NIBS) techniques, such as
TMS or transcranial direct and alternating current stimu-
lation (TDCS/TACS), allow to experimentally manipulate
neuronal activity in the healthy human brain in a tempo-
rally and spatially specific manner, thereby overcoming
the merely correlative nature of electrophysiological
and neuroimaging techniques (Bergmann, Karabanov,
Hartwigsen, Thielscher, & Siebner, 2016). Their ability
to bypass sensory input channels and directly affect brain
activity makes them unparalleled tools for studying
cause–effect relationships between neuronal activity and
cognitive function. Shortly after its invention (Barker,
Jalinous, & Freeston, 1985), TMS was already demonstrated
to be capable of suppressing visual perception (Amassian
et al., 1989), and by now, the “disruptive” or “interfering”
effects of TMS have a long-standing tradition in cognitive
neuroscience, following the so-called virtual lesion ap-
proach (Pascual-Leone, Walsh, & Rothwell, 2000; Walsh &
“This Special Focus Review derives from a symposium at the
2019 annual meeting of the Cognitive Neuroscience Society, or-
ganized by Romy Lorenz.”
1Leibniz Institute for Resilience Research, Mainz, Germany,
2Max Planck Institute for Human Cognitive and Brain Sciences,
Leipzig, Germany
Cowey, 2000). Later, TDCS (Nitsche & Paulus, 2000) and
TACS (Antal et al., 2008) were discovered as methods for
subthreshold modulation of neuronal activity and thus of
cognitive function ( Vosskuhl, Strüber, & Herrmann,
2018; Yavari, Jamil, Mosayebi Samani, Vidor, & Nitsche,
2018). Although cognitive neuroscience studies using these
NIBS techniques often claim to test the “causal relevance”
of a specific brain region or neuronal activity pattern for a
specific cognitive function or behavior, the underlying
cause–effect relationships are rarely made explicit.
However, to justify such causal inference, the theoretically
assumed chain of causation, leading from the applied stim-
ulation to the observed behavioral change, has to hold for
a concrete empirical experiment. Crucially, every single
cause–effect link in this causal chain can be interrupted
or confounded by several factors, which are best eliminat-
ed or controlled experimentally to attribute the observed
results to their assumed cause. We will start by briefly out-
lining the core element of this paper: a simplified five-step
chain of causation for cognitive neuroscience NIBS studies
and its principal confounders. We will then introduce gen-
eral experimental approaches using NIBS and discuss the
concept of causal inference for the case of experimental
NIBS studies in cognitive neuroscience before we walk
the reader step-by-step through the five-step chain of cau-
sation. Afterward, we will discuss potential confounders in
© 2020 by the Massachusetts Institute of Technology. Published under
a Creative Commons Attribution 4.0 International (CC BY 4.0) license.
Journal of Cognitive Neuroscience 33:2, pp. 195–225
https://doi.org/10.1162/jocn_a_01591
more detail and review the available experimental control
conditions to counteract them before we conclude by pro-
viding 12 general recommendations for designing valid
NIBS studies.
Please note that, in the context of this paper, causal in-
ference simply means “inferring causality” or “inferring
that one variable is the cause of another” (Scheines,
2005), an inference that may either be based on the con-
trolled randomized experiment or, under certain condi-
tions, on observational data alone, when using the causal
inference framework developed by Judea Pearl (Pearl &
Mackenzie, 2018; Pearl, 2010) and others. A more general
introduction into the latter and its implications for neuro-
imaging studies is outside the scope of this paper. While
adopting elements from this particular causal inference
framework, this review remains largely focused on the clas-
sical experimentalist’s framework of inferring causation via
experimental manipulation. We primarily aim to raise
awareness for the underlying (often implicitly assumed)
chain of causation in NIBS studies, their potential con-
founds, and respective experimental control measures,
encouraging the conduction of well-planned and well-
controlled NIBS experiments that actually justify causal
inference, that is, the conclusion of cause–effect relation-
ships between neuronal activity and cognitive function.
The Chain of Causation in NIBS Studies
A chain of causation (or causal chain) refers to an unin-
terrupted concatenation of cause–effect pairs, leading
from an initial cause of interest via a number of mediating
variables to an eventual effect of interest. Given that most
effects have many causes and themselves cause many ef-
fects, such a chain represents only one specific path
through an entire causal diagram. A causal diagram can
be formalized as variables connected with arrows that in-
dicate causation (A ➔ B) instead of mere association (A,
B), with the left and right variables representing cause
and effect for a particular cause–effect pair (Pearl &
Mackenzie, 2018). Importantly, there is no ultimate cause
or effect, and the partial chain to be considered depends
entirely on the research question at hand. Once the hy-
pothesis has defined cause and effect of interest, any in-
termediate element within a causal path connecting them
is referred to as “mediator.” In contrast, elements that are
associated with the cause of interest (i.e., causing it or
merely covarying with it but not caused by it) and that
influence the effect of interest are considered con-
founders, because their influence on the effect of interest
is mixed with that of the cause of interest, preventing the
straightforward attribution of causal influence to the lat-
ter and therefore the identification of an unambiguous
causal path through the causal diagram.
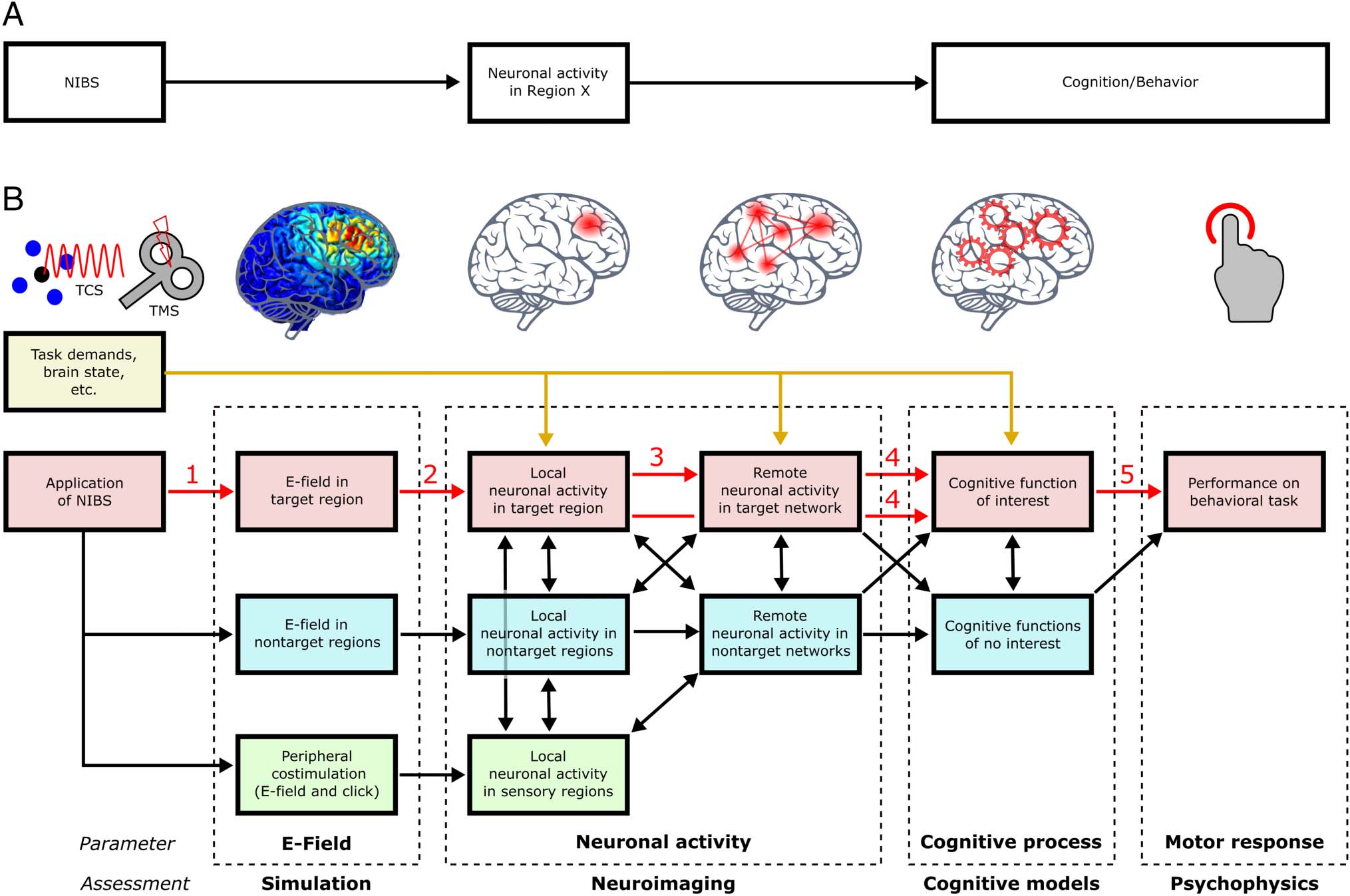
The somewhat naive level at which hypotheses are often
phrased in NIBS studies is depicted in Figure 1A. The stim-
ulation is expected to affect a single circumscribed brain
region, which has an effect on behavior, because the brain
region is causally relevant for producing that behavior. In
Figure 1B, we provide a more elaborated causal diagram,
but for the sake of comprehensibility, it is still simplified
and reduced to the key variables (some of them summa-
rizing multiple smaller ones). The red elements connected
by red arrows indicate the core chain of causation leading
from application of NIBS to an observable behavioral ef-
fect. The yellow box and arrows indicate the causal route
by which task demands and the current brain state drive
local and network activity and thereby the respective cog-
nitive function and behavioral responses. This causal path
is not only a source of confounding but is relevant for task
performance in the absence of NIBS and the main drive for
the cognitive function of interest. In fact, NIBS-induced
brain activity per se is insufficient to cause more complex
cognitive functions and only modulates ongoing task-
related neuronal activity. The black arrows indicate addi-
tional causal relationships that result from or eventually
affect elements of the main causal chain of interest and may
thereby produce confounding via central (blue boxes) and
peripheral (green boxes) off-target stimulation. We segre-
gate the core chain of causation from NIBS application to
behavior into five cause–effect pairs as described below.
Arrow 1: The application of NIBS produces an electric
field (E-field) in the brain tissue, either via transcranial
electro-magneto-electric induction (TMS) or the direct
administration of weak transcranial currents (transcranial
current stimulation [TCS]).
Arrow 2: The E-field then interacts with the neurons’
membrane potential to immediately (online) evoke neuro-
nal firing (TMS) or modulate the threshold for spontaneous
firing (TCS) locally in the targeted brain region, activating
specific intracortical circuit motifs and associated neuro-
nal signatures ( Womelsdorf, Valiante, Sahin, Miller, &
Tiesinga, 2014). In the case of offline TMS/TCS protocols,
it additionally triggers processes of synaptic plasticity.
Arrow 3: If driving output neurons of the targeted net-
work node to suprathreshold levels, local neuronal activ-
ity transsynaptically spreads to other connected brain
regions of the targeted network via intercortical axonal
projections, activating large-scale and remote circuit mo-
tifs as well as remote changes in synaptic strength.
Arrow 4: The immediate (online) or subsequent (off-
line) effects on local and large-scale circuit motifs can dis-
turb or facilitate the specific task-relevant neuronal
computations mediating the cognitive function of inter-
est, which is reflected in either respective changes of
the outcome or the completion time of these processes.
Arrow 5: With regard to the motor responses exerted
in the context of a specific behavioral task, these altered
cognitive processes eventually result in changed error
rates or RTs.
Confounding in NIBS Studies
Most cognitive neuroscience studies investigate the neuro-
nal implementation of a cognitive function and therefore
196
Journal of Cognitive Neuroscience
Volume 33, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
3
2
1
9
5
1
8
6
2
5
1
7
/
j
o
c
n
_
a
_
0
1
5
9
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
3
2
1
9
5
1
8
6
2
5
1
7
/
j
o
c
n
_
a
_
0
1
5
9
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 1. Causal diagram for NIBS studies in cognitive neuroscience. (A) Naive chain of causation stated for many NIBS studies: Stimulation is
expected to affect a single brain region, which has an effect on behavior, because the brain region is causally relevant for producing that behavior. (B)
A more elaborated causal diagram, with red arrows indicating the core chain of causation; yellow arrows indicating the impact of task demands and
brain state; and black arrows indicating additional causal relationships that may produce confounding. Application of NIBS produces an E-field in the
brain tissue (1), which evokes or modulates local neuronal activity in the target region (2), which then spreads via synaptic connections to other brain
regions within the same target network (3) and affects the task-relevant cognitive processes of interest mediated by the local target region or the
target network (4), which eventually results in a motor response as part of this task (5). However, NIBS also produces E-fields and thereby neuronal
activity in nontarget regions, spreading in nontarget networks and affecting cognitive processes of interest and of no interest (blue boxes), and
thereby behavioral outcome. NIBS also creates an E-field in the periphery, causing afferent input and neuronal activity in sensory regions (green
boxes), affecting both target and nontarget networks as well as cognitive processing. Importantly, task demands and the current brain state are the
main drive of local and network neuronal activity and thus engage the cognitive function itself, whereas NIBS-related brain activity is merely
modulating task-related neuronal activity and cognition.
hypothesize that neuronal activity in a certain brain region is
causal to that cognitive function of interest. NIBS is merely
considered a means to manipulate the cause “neuronal
activity” (via an E-field), whereas behavioral measures are
used as an observable proxy to assess the hidden effect
“cognitive function.” Every single link in this causal path
can be confounded by several (known or unknown) factors
(yellow, blue, and green boxes in Figure 1B), which are
best experimentally eliminated or controlled for to rule
out alternative explanations for the observed data and to
draw strong conclusions regarding the hypothesized
cause–effect relationship. This is complicated by the facts
that (i) many of the mediating and confounding variables
are not directly observable and have to be approximated
by simulation (E-field), neuroimaging (neuronal activity),
or modeling (cognitive function) and (ii) few of the rele-
vant cause–effect pairs express linear dose–response
relationships. Depending on the expertise and educational
background of the researcher, some of these links are typ-
ically less thoroughly elaborated than others (e.g., the kind
of neuronal activity induced by the E-field or the precise
behavioral changes to be expected from changes in cogni-
tive function), crucial assumptions about mediators re-
main untested (e.g., whether the TMS-induced E-field
actually produced the neuronal activity that was aimed
for), and potential confounds are uncontrolled (e.g., the
sensory input because of peripheral costimulation).
GENERAL APPROACHES FOR NIBS IN
COGNITIVE NEUROSCIENCE
Although NIBS is often depicted as a means by which we
can simply “switch off” or “knock out” entire brain regions
or realistically mimic endogenous oscillatory activity, the
Bergmann and Hartwigsen
197
reality is more complicated, and the mechanism of action
and applicability of a specific technique need to be consid-
ered before choosing it to manipulate neuronal activity in a
specific study. Although often lumped together as “NIBS,”
TMS and TCS rely on different neurophysiological princi-
ples of action. Sharing the principal mediating mechanism
of an E-field being imposed on the brain tissue, the result-
ing neuronal effects differ markedly. The fast-changing,
high-amplitude E-field gradients (< 300 μs) caused by
TMS are sufficient to fully depolarize the membrane poten-
tial of cortical neurons, causing the immediate emergence
of action potentials (APs; suprathreshold stimulation). In
contrast, the much weaker constant or alternating E-fields
caused by TCS are assumed to merely shift the neurons’
membrane potential slightly toward depolarization or hy-
perpolarization, thus modulating the likelihood of APs to
emerge spontaneously (subthreshold stimulation). This
fundamental difference has important consequences for
the kind of experimental approaches suitable for TMS
and TCS, respectively. Transcranial ultrasound stimulation
is yet another promising NIBS technique for neuromodu-
lation, which has recently received growing attention be-
cause of its capability of stimulating very circumscribed
volumes deep in the brain while sparing the overlying
tissue, and can be expected to amend the NIBS toolbox
for human applications in the near future (Fomenko,
Neudorfer, Dallapiazza, Kalia, & Lozano, 2018). Transcranial
ultrasound stimulation is not based on the induction of
E-fields but presumably involves the mechanic impact of
focused sound pressure waves on the neurons’ membrane
and/or ion channels ( Jerusalem et al., 2019) and will not
be discussed within the scope of this paper.
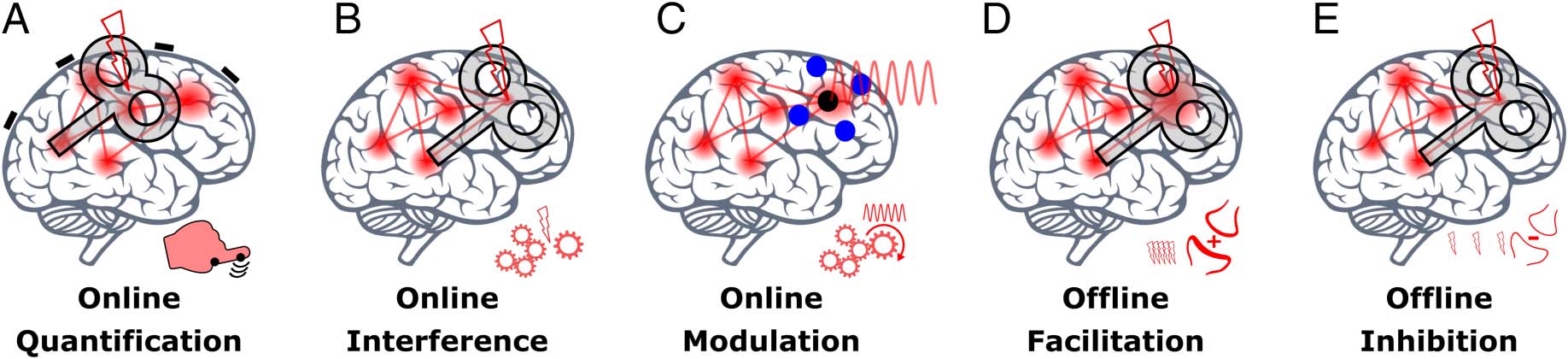
minutes to hours (Figure 2; for a more detailed discus-
sion of NIBS approaches, see Bergmann et al., 2016).
Online approaches, assessing the immediate neural re-
sponse to stimulation, can be used to (i) quantify prop-
erties such as cortical excitability or connectivity and their
modulation by brain state (including task engagement;
Figure 2A), (ii) interfere with ongoing task-related or
spontaneous neuronal activity and thereby with cogni-
tion (Figure 2B), or (iii) modulate, more gently, the level
(“gating”) and timing (“entrainment”) of neuronal activity
and thereby cognitive function (Figure 2C). In contrast,
offline approaches can be utilized to either (iv) inhibit
or (v) facilitate neuronal excitability for an extended pe-
riod via mechanisms of synaptic plasticity, assessing its
subsequent effects on neuronal activity and cognition
(Figure 2D and E). Although relying on different neuro-
physiological mechanisms (which we will discuss in a
later section), both online interference and offline inhibi-
tion approaches have been referred to as “virtual lesion”
and are most frequently used in cognitive neuroscience
to test whether and when a cortical region is “causally
relevant” for a cognitive function. Online modulation
additionally allows to investigate which neuronal patterns,
for example, oscillatory frequencies, are mediating or
supporting a specific cognitive function. Whereas TMS
works for all these approaches, the much weaker TCS
cannot be used for quantification (as it does not trigger
APs) and hardly for interference. In contrast, it is optimally
suited to modulate without disruption and can produce
lasting offline effects. We will see that net increases and
decreases in cortical excitability (online as well as offline)
do not simply translate into respective behavioral improve-
ments and impairments.
Experimental Approaches Using NIBS
Depending on their specific stimulation parameters, both
TMS and TCS are able to exert not only immediate
(online) effects during stimulation but also subsequent
(offline) effects that outlast the stimulation itself for
CAUSAL INFERENCE FROM NEUROIMAGING
VS. NIBS IN COGNITIVE NEUROSCIENCE
Causal inference refers to the process of inferring cause–
effect relationships based on the observed changes of an
Figure 2. Schematic representation of noninvasive brain stimulation approaches. (A) Online quantification: a stimulation strong enough to cause a direct
output of the targeted region/network (with TMS, not TCS) that allows to quantify cortical excitability via motor-evoked potentials or phosphene reports.
(B) Online interference: a disruption of ongoing task-related or spontaneous brain activity (with TMS, rather not TCS) that disturbs a cognitive function.
(C) Online modulation: a moderate modulation of the level (“gating” via low-intensity TMS or TDCS) or timing (“entrainment” via TACS or rhythmic TMS) of
neuronal activity that interacts with ongoing task-related or spontaneous neuronal activity without disrupting it. (D) Offline facilitation: an increase in cortical
excitability (triggered by repetitive TMS [rTMS] rTMS or prolonged TCS) presumably mediated via long-term potentiation of the stimulated synapses.
(E) Offline inhibition: a decrease in cortical excitability (triggered by rTMS or prolonged TCS) presumably mediated via LTD of the stimulated synapses.
198
Journal of Cognitive Neuroscience
Volume 33, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
2
1
9
5
1
8
6
2
5
1
7
/
j
o
c
n
_
a
_
0
1
5
9
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
effect, after changes of its hypothesized cause. In contrast
to inferring a mere association of the two variables by
means of conditional probabilities, that is, P(effect|
cause), causal inference assigns a direction to their rela-
tionship, assuming that active manipulation of the cause
(experimentally or counterfactually), with everything else
held constant (“ceteris paribus”), produces the effect, but
not vice versa. This asymmetric relationship has been
formalized via the do operator, where P(effect|cause) =
P (effect|do(cause)), whereas P (cause|effect) ≠ P(cause|
do(effect)) = P (cause) (Pearl & Mackenzie, 2018). A sim-
ple example in the context of NIBS would be that TMS
(with sufficient intensity) of the primary motor cortex
(M1) hand area causes a contralateral finger movement
∼20 msec later or at least increases its likelihood (P(con-
traction|do(TMS)), but a spontaneous finger movement
does not affect the likelihood of TMS to occur (P[TMS|
do(contraction)] = P(TMS)). In fact, this asymmetry
holds for every single cause–effect pair in the causal
chain mediating the effect from TMS pulse to finger
movement (i.e., TMS pulse ➔ E-field ➔ APs in cortical
neurons in M1 Layer 2/3 or premotor cortex ➔ APs in cor-
ticospinal output neurons in M1 Layer 5 ➔ APs in spinal
motoneurons ➔ muscular APs ➔ muscle contraction ➔
finger movement). For behavioral task performance in
cognitive neuroscience studies, the causal chain is typi-
cally even more complex, and for the causal diagram in
Figure 1B, the numerous single steps have been consol-
idated into a few categories for the sake of simplification.
It has been argued that causal relationships can provi-
sionally be inferred from observational data alone within
the causal inference framework when using the do-
calculus to decide which confounding variables should
be statistically adjusted for and which should rather not
to avoid the introduction of spurious effects (Pearl &
Mackenzie, 2018; Pearl, 2010). This approach has gained
increasing interest in the functional neuroimaging com-
munity as well, where its applicability is fiercely debated
(Reid et al., 2019; Mehler & Kording, 2018; Grosse-
Wentrup, Janzing, Siegel, & Schölkopf, 2016; Weichwald
et al., 2015; Ramsey et al., 2010). In contrast, the classical
experiment solves this caveat elegantly also with respect
to unknown confounders via the randomized allocation
of levels of the independent variable (IV) to observational
units across multiple experimental repetitions (i.e., do
[cause]), while observing the resulting changes in the de-
pendent variable (DV) as a function of the IV level (i.e.,
P(effect|do(cause))). Although manipulation of the IV
and resulting changes of the DV relate to cause and effect
for the case of a randomized controlled trial with a single
cause–effect pair, it is more ambiguous for more complex
chains of causation with multiple cause–effect pairs lined
up (like in Figure 1B), where only the first cause (here,
application of stimulation) relates to the experimental
manipulation of an IV and only the last effect (here, be-
havior) is assessed as DV, whereas all intermediate steps
are hidden variables that can neither be manipulated
nor directly be observed. We will thus use the terms
“cause” and “effect” within the causal inference frame-
work, but “IV” and “DV” when taking the experimentalist’s
perspective.
Mapping Correlational Relationships with
Noninvasive Neuroimaging
In neuroimaging studies, the cognitive function of inter-
est is isolated experimentally as the only difference be-
tween respective task conditions (levels of the IV ),
whereas the associated neural activity is assessed (as
DV) and contrasted between conditions. By experimental
variation of the IV in randomized controlled trials, clever
design of task conditions, and careful experimental con-
trol of possible confounders, such experiments, unlike
mere observational studies, allow to attribute the ob-
served DV change to variations of the IV. Nonetheless,
the causal direction between cognitive function and brain
activity is not easily derived from such experiments.
Commonly, this allocation of IV and DV does not imply
that a certain mental state is expected to cause the re-
spective brain state (P[“brain state”|do(“mental state”)])
but rather the opposite (P[“mental state”|do(“brain
state”)]) or, at least, that the noncausal relation of super-
venience is assumed (Dijkstra & de Bruin, 2016). While
trying to avoid taking any particular philosophical posi-
tion with respect to the mind–body problem (for an in-
troduction, see Chambliss, 2018; Nagel, 1993), we will,
for the ease of argument, assume here that a cognitive
function causally depends on a specific neuronal sub-
strate (i.e., the structure) and temporospatial patterns
of neuronal activity it produces (P[“cognitive function”|
do(“brain activity”)]). This should not be misunderstood
as a dualist view on mental causation, because “cognitive
function” does not refer to a conscious, phenomenal ex-
perience or mental state but rather pragmatically to the
mechanisms of information processing and the computa-
tions that eventually give rise to a certain behavior, in the
following generously spanning anything from perception,
via higher-order cognitive processes, to motor function.
From that perspective, a cognitive neuroimaging experi-
ment thus engages the participant in a task requiring for
its completion the recruitment of certain neuronal net-
works and mechanisms implementing the respective tar-
get cognitive function, while measuring brain activity as a
function of task condition (for a detailed discussion, see
Dijkstra & de Bruin, 2016). However, because not all as-
sociated brain activities may be causally contributing to
the cognitive function engaged by the task, the measured
neuronal activity cannot qualify unambiguously as its
cause but merely as its neuronal correlate.
Mapping Causal Relationships with NIBS
This ambiguity of causal direction can be resolved when
using NIBS to experimentally manipulate brain activity
Bergmann and Hartwigsen
199
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
2
1
9
5
1
8
6
2
5
1
7
/
j
o
c
n
_
a
_
0
1
5
9
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
independent of task engagement instead of merely ob-
serving it via neuroimaging, while measuring behavioral
task performance as a proxy for the integrity of a cogni-
tive function (Sack, 2006; for a detailed discussion, see
Dijkstra & de Bruin, 2016). Therefore, NIBS-related changes
in behavior can principally be interpreted as causal effects
of the experimentally induced change in brain activity
(P[“cognitive function”|do(“brain activity”)]). There are
few examples when NIBS alone is sufficient to produce
motor behavior or a perceptual phenomenon, such as the
well-established induction of muscle responses as quanti-
fied by the motor-evoked potentials (MEPs) or phosphenes
(illusory perceptions of light) after TMS of the primary
motor and visual cortex, respectively (Kammer, 1998;
Mills, Boniface, & Schubert, 1992). For these cases, the
online quantification approach (Figure 2A) can be em-
ployed to demonstrate a clear causal relationship between
neuronal activity in the respective brain structure (e.g.,
TMS-induced firing of corticospinal motor neurons in M1)
and behavioral outcome (e.g., contraction of a contralateral
hand muscle), and even dose–response relationships can
be identified (e.g., the sigmoidal function relating increas-
ing TMS intensity to increasing MEP amplitude). However,
with NIBS in humans, most other brain targets do not
result in overt outputs. Instead, behavioral tasks are re-
quired to engage the cognitive function of interest and
its neuronal correlate, while using NIBS to manipulate
the activity or excitability of a target brain region before
(offline; Figure 2D and E) or during (online; Figure 2B
and C) task performance to reveal its causal contribution
to this cognitive function. This brings us back to the heart
of this paper: the chain of causation that is tacitly hypoth-
esized for NIBS studies in cognitive neuroscience and its
many possible confounders that complicate the causal
interpretation of NIBS results (Figure 1B). In the next
sections, we will walk the reader step-by-step through
the causal path and discuss for each step under which
conditions a cause–effect relationship can be assumed.
Afterward, we will describe how these causal links can
be confounded by variables that systematically covary
with the cause and constitute an alternative cause for its
effect. Only if an uninterrupted chain of causation can be
established without confounding causes for the individual
links, the conclusion can be drawn that “neuronal activity
in region X is causing behavior Y.”
FROM NIBS APPLICATION TO E-FIELDS
(ARROW 1)
The first cause–effect pair (Arrow 1) is often implicitly as-
sumed without further discussion, namely, that the ap-
plied NIBS technique produces an E-field of desired
intensity, extent, and direction in the target brain region,
without affecting nontarget brain regions and without the
target brain region being inadvertently affected by other
factors associated with the stimulation. This first crucial
step is far from trivial, and simply holding a TMS coil or
attaching a TCS electrode over the assumed target region
is not sufficient for many reasons.
Identifying the Target
Before we can attempt to stimulate a specific target site of
interest, we need to determine its location. Depending on
the spatial specificity of the NIBS method, these targets are
easily underspecified or overspecified. For TMS, effective
current densities are restricted to less than a cubic centi-
meter (Brasil-Neto, McShane, Fuhr, Hallett, & Cohen,
1992), and targets such as the “posterior parietal cortex”
or “dorsolateral pFC” are very unspecific, given that (i)
the functional organization of most brain areas is topo-
graphically more fine-grained and (ii) only a portion of that
anatomical structure will receive effective stimulation. For
TCS, in contrast, even entire brain regions can hardly be
stimulated in isolation. Irrespective of the spatial specific-
ity of the NIBS technique, the target site can principally be
determined based on (i) its function, (ii) its neuroanatom-
ical location, or (iii) even its location relative to the skull
alone. A functional TMS localizer can be used to determine
a motor or phosphene hot spot based on MEPs and phos-
phenes, respectively. This approach is highly specific as it
allows fine-tuned coil positioning based on the immediate
feedback from the output variables, ensuring that the
intended neuron population is effectively stimulated.
Unfortunately, this method is only available for very few
targets (i.e., motor and visual cortex; see Bergmann
et al., 2016). The second-best option is a localizer via
fMRI, for example, to determine the FEF from a covert spa-
tial attention task (Marshall, O’Shea, Jensen, & Bergmann,
2015) or the extrastriate body area from contrasting body
parts versus other objects (Zimmermann, Verhagen, de
Lange, & Toni, 2016). While providing no information on
coil orientation, the target voxel can be determined based
on individual statistical maps. For functions tightly linked
to an identifiable anatomical location (e.g., motor hand
knob), an individual structural MRI scan alone may be used
to identify the target coordinates, yet allowing consider-
able uncertainty within that area. When ignoring interindi-
vidual variability in a structure–function relationship,
standard coordinates from the literature can be utilized
after transforming them to native space with the help
of an individual structural MRI (Duecker et al., 2014).
When disregarding individual brain anatomy altogether,
the 10–20 EEG electrode system can be used to roughly
estimate the location of specific brain regions (e.g., F3 for
the left dorsolateral pFC or P4 for the right posterior
parietal cortex). Systematic comparisons revealed that,
with decreasing individualization across the above-
described methods, the number of participants required
to observe a significant effect increases dramatically (Sack
et al., 2009; Sparing, Buelte, Meister, Pauš, & Fink, 2008).
Importantly, all approaches besides the TMS localizer
200
Journal of Cognitive Neuroscience
Volume 33, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
2
1
9
5
1
8
6
2
5
1
7
/
j
o
c
n
_
a
_
0
1
5
9
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
and the 10–20 system require an MR-informed frameless
stereotactic neuronavigation setup to position the TMS
coil over the target site, which is considered state-of-
the-art to maintain coil position within and across exper-
imental sessions.
Reaching the Target
Once we know where to stimulate, how can we ensure
that the desired E-field is expressed in the target site?
For TMS, a high-voltage current pulse (< 300 μs), running
through an insulated coil held tangential to the scalp,
produces a magnetic field that painlessly penetrates the
skull and in turn produces an electric current in the un-
derlying brain tissue. Importantly, the magnetic field is
not attenuated by the intermediate bone, but the induced
E-field in the brain simply decays exponentially with dis-
tance from the TMS coil (Thielscher & Kammer, 2004).
This highlights the role of the scalp–cortex distance,
which is known to vary across both brain regions and in-
dividuals and can partially be accounted for by adjusting
stimulation intensity to the actual scalp–cortex distance
(Stokes et al., 2005). In addition, the local E-field distri-
bution depends on the anatomical distribution of brain
tissues with different conductivities (gray matter, white
matter, corticospinal fluid) and the individual gyrification
of the underlying cortex (Opitz, Windhoff, Heidemann,
Turner, & Thielscher, 2011; Thielscher, Opitz, & Windhoff,
2011). The E-field induced by TCS, in contrast, has to pass
through the bone, which is a major barrier of low conduc-
tivity, causing large portions of the stimulation current
to be shunted via the scalp and to enter the skull via
openings, such as eyeballs, ear canals, or small foramen
for the cranial nerves. Within the brain, the E-field distri-
bution depends again on the distribution of brain tissues,
but unlike for TMS, the TCS-related E-field extends across
a much larger brain volume (depending on the specific
electrode type and montage; Opitz, Paulus, Will, Antunes,
& Thielscher, 2015; Datta et al., 2009). Importantly, E-field
simulations based on anatomically precise individual
head models revealed that the location of the maximum
E-field varies across brain regions and individuals and
is not simply located directly underlying the TMS coil
(Weise, Numssen, Thielscher, Hartwigsen, & Knösche, 2020;
Gomez-Tames, Hamasaka, Laakso, Hirata, & Ugawa, 2018)
or TCS electrode (Opitz et al., 2015; Saturnino, Antunes, &
Thielscher, 2015). To establish that an effective stimula-
tion intensity is achieved at the target coordinate, indi-
vidualized E-field modeling is advisable for both TCS
(Alekseichuk, Falchier, et al., 2019; Kasten, Duecker,
Maack, Meiser, & Herrmann, 2019) and TMS (Weise et al.,
2020; Bungert, Antunes, Espenhahn, & Thielscher, 2017).
Yet, although spatial E-field parameters are reliably simulated,
its absolute intensity ( V/m) at the target coordinate unfor-
tunately is more uncertain (Saturnino, Thielscher, Madsen,
Knösche, & Weise, 2019).
FROM E-FIELDS TO LOCAL NEURONAL
EFFECTS (ARROW 2)
The second cause–effect pair (Arrow 2) refers to the im-
pact of the E-field on local neuronal activity. For the sake
of simplicity and to prevent overloading the causal dia-
gram in Figure 1B, local neuronal activity here also refers
to effects secondary to the initial neuronal response,
such as the activation of local circuit motifs, shifts in neu-
ronal excitability, entrainment of local neuronal oscilla-
tors, and local synaptic plasticity. The key question is
thus not only whether the applied E-field directly excites
local neuronal elements in the target brain region but
also whether it generates the specific neuronal effects re-
quired for the chosen experimental approach (Figure 2).
Although there is some principal understanding of the
neuronal effects of TMS and TCS, combining noninvasive
electrophysiological and neuroimaging techniques with
NIBS can help to verify for a specific experiment that
the desired neuronal effects were successfully induced.
Inducing Immediate (Online) Effects in Local
Neuronal Activity
Membrane polarization is presumably the main mecha-
nism of action for both TMS and TDC, although addi-
tional mechanisms have been discussed (Peterchev
et al., 2012). For TMS, the E-field dynamics are sufficiently
fast and strong to depolarize the neuronal membrane to
suprathreshold levels, presumably at the level of axons or
axon terminals (Aberra, Wang, Grill, & Peterchev, 2020),
to a degree that APs emerge and spread along the mem-
brane. These APs then transsynaptically affect connected
neurons, causing excitatory and inhibitory postsynaptic
potentials, via glutamate and GABA-A/GABA-B receptors,
depending on the initially depolarized neuron type.
Spatial and temporal integration of postsynaptic poten-
tials then causes excitatory and inhibitory postsynaptic
neurons to fire. Although the intracortical circuitry re-
sponding to TMS has been studied in great detail using
paired-pulse protocols and pharmacological interven-
tions in the primary motor cortex (Di Lazzaro &
Ziemann, 2013; Di Lazzaro, Ziemann, & Lemon, 2008)
and some insights have been generated by work in
rodents or nonhuman primates (Romero, Davare,
Armendariz, & Janssen, 2019; Li et al., 2017; Mueller
et al., 2014), the specific circuit motifs activated in most
human cortical regions can only be speculated about. In
any case, TMS evokes highly synchronized neuronal re-
sponses of entire intracortical circuits, not only because
the E-field initially depolarizes a large number of different
neurons but also because the activation spreads among
them. Consequently, there will be both excitation and
inhibition within a neuronal population or brain region
after TMS, and the net effect on its excitability or informa-
tion processing capabilities is complex (as will be dis-
cussed for Arrow 4).
Bergmann and Hartwigsen
201
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
2
1
9
5
1
8
6
2
5
1
7
/
j
o
c
n
_
a
_
0
1
5
9
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
For TCS, the E-field is much weaker and assumed to
merely shift the membrane potential slightly toward de-
polarization or hyperpolarization, changing neuronal ex-
citability on a subthreshold scale (Liu et al., 2018; Stagg &
Nitsche, 2011), either constantly (TDCS) or rhythmically
(TACS). The E-fields induced by standard stimulation in-
tensities (1–2 mA) in humans are much lower than those
in mice or monkeys (Alekseichuk, Mantell, Shirinpour, &
Opitz, 2019), and the effectiveness of TCS in humans is
thus highly debated (Filmer, Mattingley, & Dux, 2020;
Liu et al., 2018; Vöröslakos et al., 2018), although
TCS-induced E-fields as small as 0.2–1 V/m have already
proven effective in causing tiny shifts in spontaneous
neuronal firing rates (Krause, Vieira, Csorba, Pilly, &
Pack, 2019; Liu et al., 2018; Reato, Rahman, Bikson, &
Parra, 2010). Although small, the TCS-induced E-field is
broad, and the effects may accumulate across large neu-
ron populations. Again, entire circuits will be stimulated
both directly by the E-field and indirectly via synaptic con-
nections, but the response will be less synchronized and
more strongly dependent on ongoing brain activity com-
pared to TMS. Importantly, the impact of the E-field on
different neuronal structures at the cellular level depends
on both their shape and their orientation with the brain
and is thus highly complex, with the net effect on a given
neuron depending on the integration of diverse depolar-
ization and hyperpolarization of its parts (Rahman et al.,
2013). Even more so, the net effect on an entire brain region
arises from the integration of the individual neurons’ excit-
ability changes, highlighting the impossibility of a simple
relationship between TCS polarity and the resulting net
excitability change of the target brain region.
Quantifying Excitability and Connectivity
The quantification of motor or visual cortical excitability
via TMS-induced MEPs or phosphenes comes with the in-
herent proof of suprathreshold stimulation of target neu-
rons (Figure 2A). There are many studies that elegantly
employ MEP or phosphene measurement to demon-
strate the modulation of motor or visual cortical excitabil-
ity under various task conditions (Lepage, Saint-Amour,
& Théoret, 2008; Sparing et al., 2002). Using dual-coil
TMS, effective connectivity with those brain regions can
be assessed in a task-dependent fashion (Murakami,
Restle, & Ziemann, 2012; Davare, Lemon, & Olivier,
2008). However, these studies typically use NIBS in a cor-
relative manner, treating target brain region excitability as
a DV, not an IV, and do not probe the causal impact of
brain activity on cognition.
Interfering with Spontaneous or Task-related
Neuronal Activity
Many classic cognitive neuroscience TMS studies aim to
interfere with neuronal activity during task processing
(online) in a specific brain region to demonstrate its
causal relevance for a cognitive function (Figure 2B).
Unfortunately, we lack a precise understanding of most
neuronal activity patterns implementing a specific com-
putation and thus do not know exactly what to interfere
with. Accordingly, TMS for interference often uses either
high-intensity single-pulse TMS when aiming for a good
temporal resolution and thus short period of interference
(Amassian et al., 1998) or short TMS bursts at a high fre-
quency (mostly 10–20 Hz) covering several hundred mil-
liseconds to ensure sufficiently long disruption of
neuronal processing (Capotosto, Babiloni, Romani, &
Corbetta, 2012; Taylor, Nobre, & Rushworth, 2007).
Although the TMS-evoked neuronal activity in local
circuits is complex, it can safely be assumed that TMS
(i) excites random neural elements (those optimally
located relative to the E-field), including those not acti-
vated by the task; (ii) results in subsequent suppression
of neuronal activity, also in neurons activated by the task,
for ∼50–150 msec after initial excitation (Li et al., 2017;
Moliadze, Zhao, Eysel, & Funke, 2003), potentially
because of GABA-B-receptor-mediated inhibition, paral-
leling the motor cortical phenomena of the cortical silent
period (Stetkarova & Kofler, 2013; Chen, Lozano, &
Ashby, 1999) and long-interval intracortical inhibition
(McDonnell, Orekhov, & Ziemann, 2006; Valls-Solé,
Pascual-Leone, Wassermann, & Hallett, 1992); and (iii)
causes highly synchronized neuronal activity in the target
region based on the time-locked excitation–inhibition
pattern artificially evoked in a comparably large neuron
population (Romero et al., 2019). We will discuss possi-
ble implications of these neuronal effects for the neuro-
nal computations mediating cognition for Arrow 4.
Although stimulation intensity and frequency are likely
key parameters for determining a successful interference
protocol, there has been no systematic comparison of
stimulation intensities, frequencies, and train durations
regarding their principal suitability for interference
protocols.
Modulating (“Gating”) Neuronal Excitability
Online TDCS is often supposed to modulate the excitabil-
ity of a certain brain region during a task with the ratio-
nale to facilitate task-relevant neuronal processing
(Figure 2C). Animal work has indeed shown a polarity-
dependent modulation of spontaneous neuronal spiking
(Fröhlich & McCormick, 2010; Bindman, Lippold, &
Redfearn, 1964). However, given the complexity of neu-
ronal excitability changes, in humans, anodal and cathodal
TDCS can generally not be equated with excitability in-
crease and decrease outside the primary motor cortex
(M1). This issue is somewhat resolved for transcranial
random noise stimulation, composed of various (particu-
larly high, > 140 Hz) TACS frequencies (Terney, Chaieb,
Moliadze, Antal, & Paulus, 2008). Even for a fixed polarity,
no simple dose–response curve can be observed for TCS
202
Journal of Cognitive Neuroscience
Volume 33, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
3
2
1
9
5
1
8
6
2
5
1
7
/
j
o
c
n
_
a
_
0
1
5
9
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
(Esmaeilpour et al., 2018), and these nonlinear effects
complicate the determination of appropriate stimulation
dosages. Irrespective of these challenges, a noteworthy
approach is to induce excitability changes during a learn-
ing task to gate learning-induced synaptic plasticity, which
can result in long-lasting effects, not resulting from
stimulation- but learning-related plasticity (O’Shea et al.,
2017; Snowball et al., 2013; Vollmann et al., 2013). This
approach effectively increases the low anatomical preci-
sion of the TCS by the task-related activation of highly
specific circuits.
Entraining Neuronal Activity (and Oscillations)
Rhythmic TMS or TACS at a certain frequency is used to
entrain neuronal activity with the aim to synchronize and
enhance endogenous brain oscillations (Figure 2C) and test
their causal role for cognition (Vosskuhl et al., 2018; Antal
& Herrmann, 2016; Herrmann, Rach, Neuling, & Strüber,
2013; Thut, Schyns, & Gross, 2011; Thut, Veniero, et al.,
2011). However, the underlying neurophysiological
assumptions are often not made explicit. Neuronal oscil-
lations in EEG/magnetoencephalography (MEG) reflect
the summed potentials/fields from large synchronized
neuron populations with parallelly oriented dendritic
trees (Cohen, 2017). Their amplitude increases when
the postsynaptic activity of more neurons becomes syn-
chronized, and the entrainment of a neuronal oscillation
by rhythmic NIBS typically refers to the synchronization
of spontaneously but yet independently oscillating
neurons (Thut, Schyns, et al., 2011). However, it is also
possible that random neuronal activity is entrained in-
stead of an already ongoing endogenous oscillation
(Herring, Esterer, Marshall, Jensen, & Bergmann, 2019).
Entrainment may also work differently for TMS and TCS.
Whereas rhythmic (suprathreshold) TMS may directly
evoke waves of synchronized excitation and inhibition,
potentially phase-resetting existing oscillatory activity
(Herring, Thut, Jensen, & Bergmann, 2015), TACS merely
shifts the membrane potential forth and back, biasing
spontaneous neuronal firing. The weaker impact of
TACS thus likely requires it to be more well targeted,
for example, adjusted to the individual frequency of
the target oscillation ( Vosskuhl et al., 2018; Thut et al.,
2017). Unfortunately, direct proof of neuronal entrain-
ment during rhythmic NIBS is currently difficult to
impossible because of the strong NIBS-related artifacts
(Gebodh et al., 2019; Rogasch et al., 2017; Noury,
Hipp, & Siegel, 2016; Herring et al., 2015; Ilmoniemi &
Kičić, 2010). However, there have been a few successful
attempts using more indirect measures of neuronal
entrainment (Herring et al., 2019; Hanslmayr, Matuschek,
& Fellner, 2014; Helfrich, Schneider, et al., 2014; Thut,
Veniero, et al., 2011). Note that any lasting increase in oscil-
latory power after TACS reflects synaptic aftereffects (off-
line) in the oscillation-generating circuits, not ongoing
entrainment per se (Vossen, Gross, & Thut, 2015; Zaehle,
Rach, & Herrmann, 2010).
Inducing Aftereffects (Offline) in Local Neuronal
Excitability Based on Synaptic Plasticity
Repetitive TMS (rTMS) or prolonged TDCS can produce
transient changes in neuronal excitability, mediated by
synaptic plasticity and outlasting the stimulation protocol
itself by minutes to hours (Figure 2D and E). Several NIBS
protocols have been developed for M1, producing bidirec-
tional changes in corticospinal excitability as indexed by
MEP amplitude, primarily depending on the frequency
or pattern of rTMS or the polarity of TDCS (Ziemann
et al., 2008). In principle, classic high-frequency (∼5 Hz)
versus low-frequency (∼1 Hz) rTMS results in long-term
potentiation (LTP)-like and long-term depression (LTD)-
like facilitation and inhibition of corticospinal excitability,
respectively (Fitzgerald, Fountain, & Daskalakis, 2006),
whereas for theta burst stimulation, the specific timing
of TMS trains and pauses determines the direction of ef-
fects (Huang, Edwards, Rounis, Bhatia, & Rothwell,
2005). Likewise, TDCS in classic M1-contralateral forehead
montage produces lasting increases and decreases in cor-
ticospinal excitability, respectively, depending on whether
anode or cathode overlays M1 (Nitsche & Paulus, 2000).
Stimulation intensity and duration are crucial determi-
nants for offline effects with both TMS (Ziemann et al.,
2008) and TCS (Nitsche et al., 2008). However, Bonaiuto
and Bestmann (2015) emphasized that the “sliding-scale
rationale” (assuming the magnitude of cortical excitability
increases to scale with stimulation intensity) is incorrect,
as nonlinearity has been clearly demonstrated even for
M1 (Batsikadze, Moliadze, Paulus, Kuo, & Nitsche, 2013;
Moliadze, Atalay, Antal, & Paulus, 2012). For a detailed dis-
cussion of the neurophysiological mechanisms mediating
NIBS-induced LTP/LTD-like plasticity, the reader is re-
ferred to previous reviews (Hoogendam, Ramakers, & Di
Lazzaro, 2010; Ziemann et al., 2008). Importantly, these af-
tereffects show large intraindividual and interindividual
variability (as discussed below), often emerge with a cer-
tain delay (Huang et al., 2005), and wash out after an un-
known duration, typically 30–60 min (Ziemann et al.,
2008). Although the effectiveness of NIBS can immediately
be assessed via MEP amplitudes for M1, such a manipula-
tion check for other brain regions requires neuroimaging
techniques (see below). Although common practice, we
cannot assume every NIBS protocol to easily translate from
motor to nonmotor regions, and without a manipulation
check, we can only hope for the desired excitability effects
to occur in the target region.
Mapping NIBS-related Neuronal Effects
with Neuroimaging
Both online and offline NIBS effects can be assessed in
humans with noninvasive neuroimaging techniques
Bergmann and Hartwigsen
203
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
3
2
1
9
5
1
8
6
2
5
1
7
/
j
o
c
n
_
a
_
0
1
5
9
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
(e.g., fMRI, EEG, or MEG). For a detailed discussion of the
challenges associated with the combination of NIBS and neu-
roimaging, see previous review papers (Bergmann et al.,
2016; Bestmann & Feredoes, 2013; Siebner, Bergmann,
et al., 2009). Neuroimaging is crucial to provide proof of
target engagement, that is, to verify the assumption that
the applied NIBS protocol has effectively induced the in-
tended neuronal activity in the target region. The high
spatial resolution of BOLD fMRI helps to detect net
changes in spontaneous or task-related neuronal activity
via resting-state or task fMRI, but for demonstrating
entrainment or interference effects, the superior tempo-
ral resolution of EEG or MEG is usually required.
Neuroimaging also allows to screen for unintended coac-
tivation of nontarget brain regions, which may otherwise
cause confounding and prevent the unambiguous identi-
fication of structure–function relationships (cf. Arrows 3
and 4). In the absence of behavioral effects, neuronal
activity may be the only readout available to investigate
network effects such as compensation (cf. Arrow 3),
whereas in the presence of behavioral effects, brain–
behavior correlations may further corroborate the causal
link between NIBS-induced neuronal and behavioral
effects in terms of dose–response relationships.
FROM LOCAL TO NETWORK EFFECTS
(ARROW 3)
The third cause–effect pair (Arrow 3) refers to the impact of
NIBS-induced local neuronal activity on other connected
nodes of the target network. This spread of activation
may be desired or considered a potential confound, but
it is in any case an inherent feature of the brain, not a
shortcoming of the method. Network effects always need
to be considered when attributing changes in cognitive
function to NIBS-induced changes in the targeted brain
region.
Remote Effects of NIBS
The most direct evidence of transsynaptic spread is the
MEP after TMS of M1, which relies on several synaptic
connections from the initially excited neural elements
in M1 via corticospinal output neurons and spinal moto-
neurons to the muscle. Yet, cortico-cortical spread has
been demonstrated by combined NIBS–fMRI studies,
which revealed strong remote effects of TMS in networks
for various motor and cognitive functions (Bergmann
et al., 2016; Bestmann & Feredoes, 2013). Concurrent
TMS–fMRI studies, applying TMS to M1, FEFs, or intrapar-
ietal sulcus, found strong (dose- and state-dependent)
effects in remote but anatomically connected cortical
and subcortical areas (Ruff et al., 2008; Bestmann,
Baudewig, Siebner, Rothwell, & Frahm, 2003, 2005), even
for subthreshold intensities (Bestmann et al., 2003).
Likewise, concurrent (Antal, Polania, Schmidt-Samoa,
Dechent, & Paulus, 2011) and consecutive (Polania,
Paulus, & Nitsche, 2012) TCS–fMRI studies reported
widespread BOLD effects (Turi, Paulus, & Antal, 2012).
Yet, for TCS, direct unfocal stimulation effects (i.e., direct
effects of the widespread E-field on neuronal activity out-
side the target region) are difficult to disentangle from
actual network effects (i.e., spread of local E-field-
induced changes in neuronal activity to remote regions
via long-range axonal projections and synaptic connec-
tions). In addition, concurrent TMS–EEG studies report
TMS-evoked potentials spreading within the targeted net-
work (Harquel et al., 2016; Massimini et al., 2005), and
dual-coil TMS studies typically build on this feature when
testing effective connectivity between two brain regions
(Silvanto, Lavie, & Walsh, 2005; Ferbert et al., 1992).
Neuroimaging can be used to read out both immediate
(online) effects as well as subsequent (offline) effects me-
diated via synaptic plasticity (Bergmann et al., 2016). For
aftereffects on remote neuronal activity, the question re-
mains, however, whether they are caused by local synap-
tic plasticity in the target site, subsequently affecting
remote activity via changes in functional connectivity or
via synaptic plasticity in the remote site itself induced by
spread of activity during the stimulation.
Consequences for Network Activity
In any case, both online and offline effects on remote
nodes can be functionally relevant. For instance, in a
consecutive TMS–fMRI study, the rTMS-induced increase
in the inhibitory influence of the stimulated area on a
remote node predicted the individual TMS-induced
response delay in a language task (Hartwigsen et al.,
2017). Inhibitory stimulation effects are thus not restricted
to the stimulated area but can affect large parts of the
network, also modulating the functional interaction of
its elements. Such remote network effects remain hidden
in purely behavioral studies if single-site TMS or TCS is used
and are usually ignored when drawing conclusions about
the causal relevance of the stimulated area for a given
task. Yet, network effects are potential confounders, es-
pecially when relying on plasticity-inducing offline proto-
cols that leave the brain time for adaptive plasticity in
response to the intervention and rapid short-term reorga-
nization of the network. Note that remote effects can be
inhibitory, facilitatory, or both in different parts of the
network, and the direction of network effects is difficult
to predict a priori. NIBS-induced inhibition of a key target
node sometimes decreases task-related activity in larger
parts of the network (Hartwigsen et al., 2017; O’Shea,
Johansen-Berg, Trief, Göbel, & Rushworth, 2007), which
in turn disinhibits and increases activity in other network
nodes, thereby compensating for the disruption and pre-
venting behavioral effects. Such compensatory upregula-
tion can occur in contralateral homologous regions
(O’Shea et al., 2007), ipsilateral network nodes (Hallam,
Whitney, Hymers, Gouws, & Jefferies, 2016), and neigh-
boring regions relevant for other cognitive functions,
204
Journal of Cognitive Neuroscience
Volume 33, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
3
2
1
9
5
1
8
6
2
5
1
7
/
j
o
c
n
_
a
_
0
1
5
9
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
including domain-general areas (Hartwigsen et al., 2017).
This short-term reorganization in response to focal dis-
ruption stresses the strong potential for flexible redistri-
bution of resources and the high degree of degeneracy
in the brain (Price & Friston, 2002; Edelman & Gally,
2001). Combining NIBS with neuroimaging provides a
means of mapping both local and remote network effects
at the systems level and relating these effects to changes
in behavior.
FROM NEURONAL (NETWORK) ACTIVITY TO
COGNITIVE EFFECTS (ARROW 4)
The fourth cause–effect pair (Arrow 4) refers to the tran-
sition from neuronal network activity to a cognitive ef-
fect. The latter is not directly observable but must be
operationalized as a specific task to be assessed via be-
havioral performance. Importantly, there is no one-to-
one mapping of brain activity to cognitive functions, as
the same region is likely involved in multiple functions
and the same cognitive function relies on the interaction
of multiple regions. Moreover, NIBS protocols can influ-
ence the interaction between network activity and cogni-
tive function but rarely produce a direct behavioral
output. The main question of this section is thus whether
the desired modulation of local and network activity af-
fects the target cognitive function of interest it is assumed
to mediate. Both online and offline NIBS approaches can
either facilitate or inhibit a cognitive function (Figure 1B),
and although a causal discovery per se (e.g., cortical
area X but not Y is causally relevant for a cognitive
process A but not B) can be made independently from
the estimation of direction, size, or specific function of
the cause–effect relationship, the latter is crucial for
understanding the neuronal mechanism underlying a
cognitive function and for developing theory-based applica-
tions. It can thus be considered a key challenge to pre-
dict a priori the direction and size of the induced effects
for a given NIBS protocol, cognitive function, and exper-
imental setting.
Impairing vs. Improving Cognitive Functions
with NIBS
Many NIBS studies in cognitive neuroscience rely on the
“virtual lesion” approach to map causal relationships be-
tween neuronal activity in a given brain region and a cog-
nitive function of interest, assuming that disturbing or
inhibiting task-related neuronal activity by an online or
offline NIBS protocol will result in impairment of the in-
vestigated cognitive function. The online interference ap-
proach (Figure 2B) aims at transiently disturbing a
cognitive function with TMS during task execution,
whereas the offline inhibition approach (Figure 2E) relies
on a decrease in cortical excitability during the task, me-
diated by the preceding weakening of synapses after an
inhibitory offline NIBS protocol. In both cases, decreases
in task performance are expected. Yet, the term “virtual
lesion” is misleading, because TMS does not simply switch
off a brain region, and offline and online approaches
rely on different neuronal mechanisms (cf. Arrow 2).
Moreover, “virtual lesions” can only explain perfor-
mance impairments, and improvements in response to
“inhibitory” protocols are often referred to as “paradox-
ical facilitation” ( Walsh & Cowey, 2000; Kapur, 1996).
The alternative rationale for demonstrating causal rele-
vance of a brain region or neuronal activity pattern for a
given cognitive function is to facilitate it during task ex-
ecution and show positive consequences for perfor-
mance. Again, this is typically tried either with online
modulation (Figure 2C), entraining task-relevant oscilla-
tions via rhythmic TMS or TACS or increasing immediate
cortical excitability via TDCS during a task, or by offline
facilitation (Figure 2D). The latter is supposed to induce
a lasting increase of spontaneous neuronal activity dur-
ing a subsequent task via the strengthening of synapses
with facilitatory offline NIBS. Because it is generally easier
to disturb than to improve an insufficiently understood
process, the “facilitatory” approach is used less frequently.
NIBS-induced facilitation of behavior is nonetheless
tempting, as it opens interesting avenues for therapeutic
applications or neuroenhancement. Importantly, the
choice of NIBS protocols for a specific study is often built
on oversimplified assumptions, which partially explains
the many null findings and controversial results. Below,
we will discuss the possible mechanisms of action trans-
lating neuronal into cognitive effects and highlight some
of the modulating factors.
Impairing a Cognitive Function by Online TMS
The most effective approach for this aim may be online
interference via TMS (Figure 2B), which presumably
builds on three neuronal effects (cf. Arrow 2). First, the
initial excitation of random neural elements causes neu-
ronal noise in the stimulated circuits (Ruzzoli, Marzi, &
Miniussi, 2010; Siebner, Hartwigsen, Kassuba, &
Rothwell, 2009). Noise pervades all levels of information
processing in the nervous system, from receptor signal
transduction to behavioral responses (Faisal, Selen, &
Wolpert, 2008). The artificial induction of noise may im-
pair or delay task-relevant neuronal computations be-
cause neural activity needs to be sampled longer to
discriminate signal and noise. Second, the initial excita-
tion is inevitably followed by GABA-B-ergic feedback inhi-
bition, suppressing neuronal activity for ∼50–150 msec
after TMS (Inghilleri, Berardelli, Cruccu, & Manfredi,
1993; Haug, Schönle, Knobloch, & Köhne, 1992), inter-
rupting and delaying neuronal processing or even caus-
ing signal loss during crucial processing steps. This
effect may come closest to the “virtual lesion” idea of
silencing neuronal activity. Third, the evoked excitation–
inhibition sequence artificially synchronizes larger neu-
ron populations, thereby lowering the number of
Bergmann and Hartwigsen
205
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
3
2
1
9
5
1
8
6
2
5
1
7
/
j
o
c
n
_
a
_
0
1
5
9
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
possible neuronal activity patterns in the network. This
loss of entropy (Shannon & Weaver, 1949) in local neu-
ronal activity reduces the information representation ca-
pacity of the synchronized network (Hanslmayr, Staudigl,
& Fellner, 2012; Schneidman et al., 2011; Tononi, 2008),
leading to a degradation of task-relevant information and
a disruption of neuronal computations. This may result in
prolonged processing time (because of the need for
compensatory iterations or the recruitment of additional
processing resources) or even an incorrect outcome of
the computation. Importantly, online disruption does
not leave the targeted network time for functional reor-
ganization (see below), potentially leading to stronger
stimulation effects and simplifying the conclusions in
comparison to offline approaches.
Improving a Cognitive Function via Entrainment by
Online TACS or TMS
To actively improve task-related function, specific as-
sumptions are needed regarding the neuronal mecha-
nisms of action. Comparably simple targets are neuronal
oscillations, supposedly underlying a variety of cognitive
functions (Buzsáki & Draguhn, 2004). To entrain a neuro-
nal oscillation by means of phasic online modulation via
rhythmic TMS or TACS (Figure 2C), not only the brain
region or network but also the oscillatory frequency
needs to be made explicit. When able to transcranially en-
train (and augment) a neuronal oscillation and thereby
improve the cognitive function, this provides proof of
its causal relevance ( Vosskuhl et al., 2018). However,
whether an increase in synchronization is beneficial de-
pends on the very mechanism by which the oscillation
mediates the relevant neuronal computations. Beyond
mere excitability fluctuations (Bergmann, Lieb, Zrenner,
& Ziemann, 2019; Bergmann et al., 2012; Schroeder &
Lakatos, 2009), oscillations supposedly enable more com-
plex processes, such as interarea (Fries, 2015) and cross-
frequency communication via phase–phase or phase–
amplitude coupling (Jensen & Colgin, 2007), phase coding
( Jensen, Gips, Bergmann, & Bonnefond, 2014; Lisman &
Jensen, 2013), and potentially phase-dependent plasticity
(Bergmann & Born, 2018). These complex processes are
more difficult to optimize, although (multifocal) TACS
has successfully been used to produce behaviorally rele-
vant interarea synchronization (Reinhart & Nguyen,
2019) and working memory enhancement (Alekseichuk,
Turi, Amador de Lara, Antal, & Paulus, 2016). However,
an increase in synchronization, which may unintentionally
also recruit task-irrelevant neurons, or an entrainment-
induced phase shift of the endogenous oscillation may al-
so be detrimental to task-relevant neuronal computations.
Interestingly, the same TMS protocols (e.g., four to five
pulse bursts of 10- or 20-Hz TMS) are often used for both
behavioral interference (Hartwigsen, Price, et al., 2010)
and neuronal entrainment, for example, of alpha or beta
oscillatory activity (Romei et al., 2016; Thut, Veniero,
et al., 2011), and whether a TMS burst impairs or im-
proves a cognitive function may thus depend on whether
or not rhythmically synchronized brain activity in the tar-
get network is beneficial for the task.
Bidirectionally Modulating (Gating) a Cognitive
Function with Online or Offline NIBS
In contrast to disrupting or actively driving the neuronal
processes mediating a cognitive function, tonic online
modulation via TDCS (Figure 2C) is assumedly able to bi-
directionally modulate (decrease or increase) task-related
neuronal activity, depending on stimulation polarity
(among other factors). As discussed for Arrow 2, there
is no straightforward relationship between TDCS polarity,
intensity, and net excitability changes. There is also no
simple mapping from cortical excitability to cognitive
performance for any given NIBS protocol. It is well con-
ceivable that a net excitability increase in the circuits me-
diating task-relevant computations augments task-related
signal and thus boosts signal-to-noise ratio (SNR).
Alternatively, increased excitability may also augment
spontaneous but task-irrelevant activity, thereby increas-
ing noise and lowering the SNR. The same ambiguity ap-
plies also for excitability decreases. Importantly, although
additional noise may degrade task-relevant SNR for a
well-tuned neuronal representation (e.g., when distin-
guishing similar stimuli in a discrimination task) and thus
impair task performance, noise may be beneficial for
other tasks (e.g., by lifting a weak perceptual stimulus
above threshold in a detection paradigm). The actual
effects of online NIBS depend on the complex interaction
of spontaneous and task-induced brain states, specific
task demands, participant-specific characteristics, and
stimulation parameters (Fertonani & Miniussi, 2017).
Similar considerations also apply to the rationale of in-
hibition or facilitation via offline TMS or TCS protocols
(Figure 2D and E), although the bidirectional modulation
of cortical excitability after offline NIBS is based on differ-
ent neuronal mechanisms (cf. Arrow 2). The LTP-like
strengthening or LTD-like weakening of synapses in the
target network results only indirectly in subsequent in-
creases or decreases in neuronal excitability and respec-
tive changes in spontaneous and task-related neuronal
activity. It should also be noted that evidence for a bidir-
ectionality of offline effects is based almost entirely on
the respective modulation of MEP amplitudes after M1
stimulation and may not easily generalize to other mon-
tages and cortical areas (Parkin, Bhandari, Glen, & Walsh,
2019). Importantly, as for the effects of online modula-
tion, excitability changes are not homogenously distrib-
uted within the targeted brain circuits, because only a
random selection of functionally heterogeneous synapses
is affected. Despite a possible net facilitation or inhibition
of excitability, random changes in either direction are
most likely to produce noise in the neuronal activity pat-
terns generated by these circuits, with the above-
206
Journal of Cognitive Neuroscience
Volume 33, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
3
2
1
9
5
1
8
6
2
5
1
7
/
j
o
c
n
_
a
_
0
1
5
9
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
discussed positive or negative consequences for task per-
formance. These considerations are highly relevant for
training studies and therapeutic applications of NIBS,
which often assume an offline NIBS-induced facilitation
to improve behavior in a subsequently performed task
(Luber & Lisanby, 2014).
The Paradox of Paradoxical Facilitation
“Paradoxical facilitation” usually refers to an unexpected
positive effect of an “interfering” or “inhibitory” NIBS
protocol on a cognitive function. As emphasized above,
noise is a central concept to explain the cognitive effects
of NIBS protocols (Ruzzoli et al., 2010; Siebner,
Hartwigsen, et al., 2009). Taking stochastic resonance
into account, adding noise to a nonlinear system like
the human brain may produce opposite effects. Whereas
an appropriate amount of noise can add to the weak
neuronal signal of a subthreshold stimulus, elevate it
above threshold, and result in behavioral facilitation
(Schwarzkopf, Silvanto, & Rees, 2011; Miniussi, Ruzzoli,
& Walsh, 2010), exceeding noise levels may rather mask
the task-relevant neuronal signal. Importantly, the NIBS-
induced activity or neural noise is not totally random
(Ruzzoli et al., 2010) and also not independent of the
task-induced neural activity or brain state. Thus, depend-
ing on the activated neuron population, the induced ac-
tivity can even be considered both as noise and as part
of the signal (Miniussi et al., 2010). If the induced neuronal
noise is synchronized with the ongoing relevant activity
(Ermentrout, Galán, & Urban, 2008), it may augment
the signal (Miniussi, Harris, & Ruzzoli, 2013). In other
words, behavioral facilitation may result from an opti-
mum level of noise in the system. Although originally de-
scribed for online TMS studies (Abrahamyan, Clifford,
Arabzadeh, & Harris, 2011), these principles also hold
for offline TMS and TCS protocols (cf. Fertonani &
Miniussi, 2017). Indeed, offline NIBS applied before task
processing may transiently prime activity in the stimulated
area to a level that facilitates subsequent task perfor-
mance, although homeostatic metaplasticity may lead to
opposite effects (see below). Besides positive conse-
quences of noise, paradoxical facilitation may also arise
from NIBS-induced inhibition of task-irrelevant areas that
compete for resources (“addition-by-subtraction” [Luber
& Lisanby, 2014]) or the disruption of distracting stimulus
elements, facilitating task-relevant processing ( Walsh,
Ellison, Battelli, & Cowey, 1998). Finally, stimulation-
induced disinhibition of distant connected areas may fa-
cilitate cognitive processing (Sandrini, Umiltà, & Rusconi,
2011).
Unfortunately, given the complex interaction between
task, brain state, stimulation protocol, and intensity, in
most cognitive neuroscience experiments, the exact cir-
cumstances under which a given NIBS protocol results,
on average, in behavioral impairment or facilitation re-
main unknown, and the above explanations are mainly
used in a post hoc fashion. It is even conceivable that,
in some cases, for example, as a consequence of in-
creased neuronal noise, the true direction of the NIBS ef-
fect on cognitive performance varies across participants
or even within participants across trials. In such a case,
the mere increase in variance (beyond measurement
noise) after NIBS may be considered evidence for a
cause–effect relationship, even when lacking a clear di-
rection. However, such a relationship would be less infor-
mative regarding the neuronal mechanisms underlying
the cognitive function and more difficult to exploit for
therapeutic applications.
Various NIBS-induced neuronal effects can affect task-
relevant neuronal computations, although little is known
about the factors making a specific neuronal process sus-
ceptible to or robust against this influence. NIBS effects
are often small, and the brain is capable to compensate
for weak disturbances, likely contributing to the null ef-
fects observed in many NIBS studies.
Multi-site Approaches to Study
Network Interactions
Because all cognitive functions rely on distributed pro-
cesses organized in large-scale neural networks, there is
increasing interest in disrupting several network nodes
for a given function in a simultaneous or subsequent
fashion to study stimulation-induced network effects on
cognitive functions. Multifocal TMS can provide insights
into functional network interactions and elucidate their
compensatory potential. Functional interactions can
be studied either online by simultaneously targeting
more than one area (“multi-site” approach) or by com-
bining offline and online TMS over different regions
(“condition-and-perturb” approach). Multi-site TMS ap-
proaches are particularly suited to map the immediate
consequences of the disruption of several brain regions
because the acute TMS-induced interference during
task performance leaves the system no time to develop
adaptive plasticity. This allows to test whether the inter-
ference effect over one area may be increased by the si-
multaneous disruption of other key regions, ipsilateral
(Ellison & Cowey, 2009) or contralateral (Hartwigsen,
Baumgaertner, et al., 2010) to the stimulation site. In a
complementary fashion, the condition-and-perturb ap-
proach can be used to study rapid network redistribution
and compensation (Hartwigsen, 2018). It combines the
plastic aftereffects of offline modulation with the imme-
diate perturbation effects of online interference, follow-
ing the rationale that offline conditioning of one area
may sensitize another network node to the disruptive ef-
fect of online interference (e.g., Hartwigsen et al., 2012).
In some cases, offline conditioning of a single target area
does not affect task-related behavior, whereas additional
online disruption of a second area effectively impairs task
performance, unmasking the disruptive effect of the off-
line protocol (Hartwigsen et al., 2015; Sack et al., 2009). A
Bergmann and Hartwigsen
207
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
3
2
1
9
5
1
8
6
2
5
1
7
/
j
o
c
n
_
a
_
0
1
5
9
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
likely explanation is that both areas contribute to the
function of interest and that offline conditioning of one
area can be compensated by a stronger contribution of
the other node, changing their functional weights within
the network. The additional online perturbation in-
creases the overall “lesion load” and thus results in cog-
nitive disruption. When combined with neuroimaging,
condition-and-perturb approaches can also be used to
study rapid reorganization at the network level. O’Shea
et al. (2007) demonstrated that offline TMS over the left
premotor cortex decreased task-related activity in the
stimulated area during an action selection task and in-
duced compensatory upregulation in other areas of the
motor network, including the contralateral homologous
region. Targeting the “reorganized” homologous premo-
tor cortex with subsequent online TMS impaired task per-
formance, demonstrating the functional relevance of the
observed compensatory upregulation. This shows how
the combination of neuroimaging and multifocal TMS
can provide insight into the compensatory dynamics of
task-specific neural networks.
Cognitive Models and
Computational Neurostimulation
To map stimulation-induced changes on a cognitive func-
tion of interest, a valid cognitive model is mandatory that
can be translated into a task. For instance, dual-route
models have been used to explain stimulus–response
compatibility effects in conflict tasks (Ridderinkhof,
2002a). Such models assume parallel routes for
decision-making that can be dissociated behaviorally
(i.e., a direct, stimulus-based activation route and a con-
trolled, deliberate response activation route) and con-
verge at the level of response activation processes. The
dynamics of these processes can be captured with distri-
butional analyses that map interference effects during
decision-making (see next section). Thereby, cognitive
models help to decompose abstract and complex con-
structs into several subcomponents that can be opera-
tionalized by specific tasks. NIBS can probe the
dynamics between these subcomponents and the func-
tional relevance of different brain regions for these
processes. More recently, cognitive models have been
complemented by computational neurostimulation ap-
proaches, which simulate emergent network dynamics
and compare them with real data (Bonaiuto & Bestmann,
2015). Computational models may, for example, more
accurately reflect choice dynamics by modeling influ-
ences of previous trials to capture choice repetition biases
(Bonaiuto, de Berker, & Bestmann, 2016; Hämmerer,
Bonaiuto, Klein-Flügge, Bikson, & Bestmann, 2016). As
humans tend to repeat recent choices in real-life situa-
tions, modeling the choice history provides valid estima-
tions of decision-making outside the laboratory (Bonaiuto
et al., 2016). Such models generate predictions about the
effects of NIBS-induced modulations of network dynamics
on behavioral measures. They rely on the assumption
that perturbation of the dynamics of a biophysical net-
work model via membrane depolarization affects cogni-
tive function (e.g., value-based decision-making) in a
predictable way, by modulating the susceptibility of net-
work dynamics to background noise. Thereby, they allow
for predicting large-scale network effects of neurostimula-
tion that can be experimentally validated (Bonaiuto et al.,
2016; Hämmerer et al., 2016).
FROM COGNITIVE FUNCTION TO
BEHAVIORAL RESPONSE (ARROW 5)
The fifth cause–effect pair (Arrow 5) refers to the impact
of a cognitive function (or its modulation) on the perfor-
mance in a specific behavioral task. The cognitive func-
tion of interest needs to be operationalized by a
specific task to measure how it is affected by a given
NIBS protocol, and this task has to be sufficiently difficult
to be sensitive enough for the (usually very small) cogni-
tive changes induced by NIBS. Usually, the interaction of
several cognitive functions is necessary for task comple-
tion, and a control task is needed that differs selectively
in the target cognitive component to contrast out the in-
fluence of other cognitive processes and establish task
specificity.
The Benefit of Behavioral Modeling Approaches
To bridge the gap between cognitive model and behav-
ioral outcome measures, distributional analyses can
provide insight into different response strategies
(Ridderinkhof, 2002a). One advantage relative to com-
posite measures like mean response speed or task accu-
racy is that they take the whole response distribution
into account (i.e., both correct and incorrect responses)
and are more sensitive to experimental dynamics and
individual differences in response strategies (van den
Wildenberg et al., 2010). They help to overcome the poor
statistical sensitivity of composite measures ( Voss,
Nagler, & Lerche, 2013). Distributional analyses map re-
sponse strategies, such as the speed–accuracy trade-off,
by disentangling whether NIBS-induced interference in-
creases errors for fast responses, indicating a potential
emphasis on speed, or rather decreases the overall un-
certainty in task processing, resulting in increased errors
for slower responses. Such analyses further help to distin-
guish subprocesses of cognitive theories. For instance,
within the framework of dual-process models, they
have been used to dissociate the role of direct response
activation (based on a target stimulus) and selective
suppression of activation based on precues in conflict
tasks (Ridderinkhof, 2002b). These processing dynamics
are usually lost when relying on composite scores.
Distributional analyses have demonstrated TDCS-
induced modulations of different error types underlying
impulsive responses (Spieser, van den Wildenberg,
208
Journal of Cognitive Neuroscience
Volume 33, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
3
2
1
9
5
1
8
6
2
5
1
7
/
j
o
c
n
_
a
_
0
1
5
9
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Hasbroucq, Ridderinkhof, & Burle, 2015), as well as TMS-
induced changes in response strategies during action re-
programming (Hartwigsen & Siebner, 2015; Hartwigsen
et al., 2012) and conflict paradigms (van Campen,
Kunert, van den Wildenberg, & Ridderinkhof, 2018).
Other behavioral modeling approaches rely on sequential
sampling models such as the drift diffusion model
(Ratcliff, Smith, Brown, & McKoon, 2016; Ratcliff, 1978),
which assumedly reflect the underlying processes contrib-
uting to a particular response distribution and also capture
decision biases. Such models are particularly sen-
sitive toward slight adaptations of response strategies that
may be overlooked when relying on composite mea-
sures (Ratcliff & McKoon, 2008; Voss & Voss, 2007), espe-
cially in NIBS studies suffering from relatively small effect
sizes (see also Hartwigsen et al., 2015). Aside from binary
choice tasks, sequential sampling models have been
adapted for more complex multichoice decisions (Kohl,
Spieser, Forster, Bestmann, & Yarrow, 2019) that might
better match real-life decisions.
Composite Measures to Quantify NIBS Effects
Yet, most NIBS studies rely on composite measures de-
rived from the individual mean response speed or accu-
racy, which are usually analyzed with ANOVAs or t tests.
However, NIBS studies can benefit from mixed models,
allowing to model nonlinear individual characteristics
and providing more flexibility when handling missing
data (Kaarre et al., 2018; Payne & Tainturier, 2018). Such
approaches are especially useful for longitudinal NIBS de-
signs where missing data for single time points might
otherwise lead to participant exclusion. Sometimes, mo-
tor responses can also be assessed by electrophysiologi-
cal means, for example, for MEPs recorded from orofacial
muscles during stimulation of motor and premotor areas
in speech production tasks (Möttönen, van de Ven, &
Watkins, 2014; Murakami, Restle, & Ziemann, 2011).
For other tasks, psychometric functions (Zazio,
Bortoletto, Ruzzoli, Miniussi, & Veniero, 2019; Cattaneo
et al., 2011) or response biases (Riddle, Hwang, Cellier,
Dhanani, & D’Esposito, 2019; Smalle, Rogers, &
Möttönen, 2015) may be the measure of choice. No mat-
ter which behavioral measure is used, NIBS protocols are
likely to first affect task efficiency, leading to increased
(or decreased) response latencies, because response
speed is a more sensitive performance metric than task
accuracy (Bonaiuto et al., 2016). However, sometimes,
task accuracy is affected without any influences on task
efficiency (e.g., Ward et al., 2010; Amassian et al.,
1998). As noted above, speed and accuracy reflect differ-
ent cognitive strategies that can be disentangled with
cognitive models. With respect to the potential mecha-
nisms related to TMS-induced interference effects on ei-
ther of the two processes, the severity of the interference
effect likely depends on the perceptional threshold. For
instance, strong interference may suppress a visual
stimulus below the perception threshold (Amassian
et al., 1998), resulting in decreased task accuracy that
cannot be compensated by increased response speed,
and weak visual stimuli closer to the perception thresh-
old might be affected first. In contrast, slight modulations
of task-related activity may selectively delay response
speed, with increases in latencies preventing effects on
task accuracy. The timing of the pulses and the stimula-
tion frequency likely play a crucial role for the outcome
of a stimulation protocol. For instance, the absence of
any impairments in task accuracy in the presence of
strong delays in response speed during a visual discrim-
ination task was explained with the employed 10-Hz
rTMS protocol, arguing that TMS might disrupt process-
ing for a brief period within each 100-msec interpulse in-
terval but never completely interferes with the relevant
information for discrimination (Ellison & Cowey, 2009).
Rather, 10-Hz rTMS may merely delay processing by the
summed periods of disruption. A similar reasoning likely
explains why a 500-msec stimulation period of 10-Hz
rTMS does not result in a 500-msec increase in response
speed ( Walsh & Cowey, 2000). Yet, to the best of our
knowledge, no study has systematically varied the stimu-
lation frequency to investigate whether higher frequencies
might affect both discrimination speed and accuracy. Most
studies assume that TMS might affect both task speed and
accuracy, but it remains unclear whether the modulation
of either parameter relies on different neuroal mechanisms.
Cognitive and computational models may help to specify
the expected effects on both outcome parameters a
priori, as the conceptualization of different subprocesses
of a cognitive function helps to dissociate the expected
outcomes.
CONFOUNDING FACTORS CHALLENGING
THE ASSUMED CHAIN OF CAUSATION
As outlined in Figure 1B, several factors may confound
the hypothesized chain of causation (red boxes), by af-
fecting both sides of the investigated core cause–effect
pair, that is, the structure–function (or brain–behavior)
relationship, namely, the targeted neuronal activity (sup-
posedly caused by NIBS) and the cognitive function of
interest (supposedly causing behavioral task perfor-
mance). Some factors are associated with the application
of NIBS, such as the unintended costimulation of nontar-
get brain regions, either directly (blue boxes) or via pe-
ripheral sensory pathways (green boxes). Other factors
(yellow box) do not originate from NIBS application
but rather from the experimental setup (e.g., task de-
mands), the participants’ predisposition (e.g., current
brain state, cognitive abilities, beliefs, and expectations),
or an interaction of both (e.g., learning effects). All these
factors either directly influence brain activity and cogni-
tive function or via a modulation of their response to
NIBS. Showing considerable intraindividual and/or inter-
individual variability, unsystematic variance in these
Bergmann and Hartwigsen
209
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
3
2
1
9
5
1
8
6
2
5
1
7
/
j
o
c
n
_
a
_
0
1
5
9
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
factors can compromise the overall effectiveness of a
NIBS protocol to modulate the target cognitive function
(producing false negatives), whereas systematic variance
across experimental conditions can introduce systematic
confounding (false positives) for both within- and
between-participants designs. To prevent confounding,
these factors either need to be eliminated, kept constant
across experimental conditions, or explicitly included as
experimental control condition (see next section).
Costimulation of Nontarget Regions and Networks
The application of TMS and TCS can have side effects
creating relevant confounding of Arrow 2, namely, an
effective E-field in nontarget brain regions and the
costimulation of peripheral neuronal structures. If the
stimulation intensity is sufficiently large, the TMS-
induced E-field will reach effective levels also in adjacent
nontarget locations (blue boxes), especially in more
superficially located ones. It is thus unlikely to exclu-
sively stimulate a coordinate deep in the sulcus, limiting
focal stimulation to the gyral crowns (Siebner, 2020;
Thielscher et al., 2011). For TCS, the widespread E-field
results in even more extended off-target stimulation at
similar or higher intensities compared to the target.
Although the E-field is more focal for multielectrode
montages with small electrodes as compared to classical
two-electrode montages with large electrodes (Datta
et al., 2009), the effective E-field will not be confined
by the borders of the targeted brain area. In addition,
both TMS and TCS also induce effective E-fields in the
cranial periphery (green boxes), with magnitudes in the
skin being inevitably larger than those in the brain
(Asamoah, Khatoun, & McLaughlin, 2019), exciting
efferent fibers of the facial nerve innervating the facial
muscles (Chen, Chauvette, Skorheim, Timofeev, &
Bazhenov, 2012), or afferent fibers of the trigeminal
nerve innervating the scalp, face, and meninges
(Siebner, Auer, Roeck, & Conrad, 1999; Schmid, Møller,
& Schmid, 1995). In particular, the TCS-induced E-field,
shunted via highly conductive skin tissue, also extends to
peripheral neuronal structures in the retina (Lorenz et al.,
2019; Schutter, 2016) and the vestibular system (Kwan,
Forbes, Mitchell, Blouin, & Cullen, 2019). In addition to
the E-field itself, physical side effects of the stimulation,
such as the “click” sound and mechanical vibration gen-
erated by the discharging TMS coil, also affect mechano-
receptors in the skin and reach the inner ear via both
airborne sound waves and bone conduction. Activation
of peripheral sensory structures then causes unintended
activation of primary and secondary sensory brain regions
and, eventually, also higher-order areas. Thus, both path-
ways (blue and green) can eventually activate remote re-
gions and affect nontarget cognitive functions, for
example, via the modulation of attentional orienting or
distraction of working memory content (Duecker &
Sack, 2015). Therefore, both local and remote network
effects can confound the effect of NIBS-induced target ac-
tivity on the target cognitive function. For example, when
targeting higher cortical association areas like the inferior
parietal cortex, which integrates information from several
modalities (Seghier, 2013), its NIBS-related activation
could result from transcranial or sensory stimulation.
Complex domain-specific cognitive functions usually en-
gage several domain-general processes like attention,
working memory, or executive functions, and sensory dis-
ruption of these processes (or other cognitive functions
of no interest) may severely affect task processing and
modulate response speed or accuracy.
Participants’ Beliefs and Expectations
Participants in NIBS studies have certain expectations and
beliefs about the impact of neurostimulation and poten-
tial side effects. These expectations and beliefs can influ-
ence their performance in a way that may be congruent
or incongruent with the experiment’s underlying hypoth-
esis. Participants may also develop expectations and actual
knowledge with regard to the temporal structure of the
experiment and the current stimulation condition (effec-
tive or sham), such that they may anticipate and prepare
for stimulation trials in online NIBS studies or behave dif-
ferently after an offline NIBS protocol.
Task Demands, Learning Effects, and
Cognitive Abilities
Many cognitive functions vary considerably over time
(e.g., as a consequence of learning or fatigue) and across
participants. Differences in baseline performance levels
because of different cognitive abilities (low vs. high per-
formers) contribute to the large interindividual variability
observed in NIBS studies of cognition. In visual priming
studies, single-pulse TMS facilitated behavioral responses
in low performers but delayed response speed in high
performers, and these effects interacted with task diffi-
culty, indicating a complex interaction between stimula-
tion, brain state, and task-induced state (e.g., Silvanto,
Bona, Marelli, & Cattaneo, 2018; Schwarzkopf et al.,
2011). Such interactions complicate the conclusions
drawn from a given NIBS protocol. Moreover, learning
or generalization effects influence performance, especially
if the same task is measured repeatedly under different
stimulation conditions, as usually done for within-
participant designs. Potential influences of these effects
need to be controlled by counterbalancing the order of
tasks and stimulation conditions across participants. In
some learning paradigms, mirrored or alternative se-
quences or stimulus lists can be used for a second ses-
sion. In case of implicit tasks or learning paradigms
that cannot be repeated, between-participant designs
need to be employed. Other unspecific effects such as
time of day and hormonal cycle may also influence cog-
nitive function. Some of these factors can (and should)
210
Journal of Cognitive Neuroscience
Volume 33, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
3
2
1
9
5
1
8
6
2
5
1
7
/
j
o
c
n
_
a
_
0
1
5
9
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
be kept constant across experimental conditions, for in-
stance, by performing repeated measures in the same
participant at the same time of day.
The Current Brain State
The magnitude and direction of NIBS effects may also
vary because of differences in the current brain state at
the time of stimulation, both within and between partic-
ipants (Miniussi et al., 2013). The concept of “state de-
pendency” has been first introduced in the visual
system (Silvanto, Muggleton, & Walsh, 2008; Silvanto,
Muggleton, Cowey, & Walsh, 2007), and state-dependent
effects have been described across a variety of (cognitive)
domains (Silvanto & Cattaneo, 2017). An impressive ex-
ample for state dependency is the quantification of corti-
cospinal excitability via the MEP acquired at rest and
under precontraction. When compared to a relaxed tar-
get muscle, slight precontraction leads to a considerable
increase in the MEP size (Siebner, Hartwigsen, et al.,
2009). State dependency has also been demonstrated in
remote network nodes by simultaneous TMS–fMRI when
comparing the effects of different TMS intensities on neu-
ral activity at rest and during a grip task (e.g., Bestmann
et al., 2008). The key assumption is that the brain state
affects the distribution of excitability in the stimulated
population of neurons, which in turn affects their respon-
siveness to stimulation. A modulation of MEP amplitude
has also been shown as a function of the current ampli-
tude and phase of neuronal oscillations during sleep
(Bergmann et al., 2012) and wakefulness (Bergmann
et al., 2019; Thies, Zrenner, Ziemann, & Bergmann,
2018). Such oscillatory modulations of cortical excitability
led to the idea of brain-state-dependent brain stimula-
tion, allowing to confine stimulation to a certain target
state (Bergmann, 2018). Brain state dynamics may also
change the NIBS interference effect, such that intensities,
which normally impair perception, suddenly have a facil-
itatory effect (Silvanto et al., 2018; Silvanto & Cattaneo,
2017). For instance, when applied immediately before
or early during a task, TMS resulted in a priming effect
by increasing activity in the target area to a level optimal
for task performance (e.g., Klaus & Hartwigsen, 2019;
Töpper, Mottaghy, Brügmann, Noth, & Huber, 1998).
In contrast, TMS impaired performance when given dur-
ing the same task ( Wassermann et al., 1999; Flitman
et al., 1998). The current brain state may also interact
with the polarity in TDCS studies, which is particularly
crucial for learning studies that engage different training
phases (Dockery, Hueckel-Weng, Birbaumer, & Plewnia,
2009). Although initially introduced for online TMS, state
dependency has also been suggested to influence offline
NIBS protocols (Nguyen, Deng, & Reinhart, 2018;
Miniussi et al., 2013). The facilitatory aftereffect of
10-Hz TACS on subsequent EEG alpha band power, for
example, is only evident when TACS is applied with eyes
open but not eyes closed and thus during different alpha
oscillation amplitudes (Neuling, Rach, & Herrmann,
2013). In addition, classical offline NIBS protocols show
large intraindividual and interindividual variability in their
aftereffects, depending on several factors that vary across
sessions within an individual (e.g., the current brain state,
history of synaptic activity, hormonal levels, circadian
rhythms) or across individuals (e.g., sex, age, individual
depth and orientation of target region, genetics;
Ridding & Ziemann, 2010), sometimes leading to null re-
sults (Beaulieu, Flamand, Massé-Alarie, & Schneider,
2017; Heidegger et al., 2017). The individual variability
in response to offline TMS protocols in the motor system
may be influenced by the specific interneuron networks
recruited, and corticospinal excitability measures may help
to dissociate responders and nonresponders (Hamada,
Murase, Hasan, Balaratnam, & Rothwell, 2013). However,
it is unclear how this translates to cognitive functions,
because excitability of M1 is not related to the responsive-
ness of other areas, and interindividual differences in
cognitive abilities and functional organization further
contribute to large interindividual variability in response
to NIBS protocols when targeting cognitive functions.
Metaplasticity
Offline stimulation protocols may further be affected by
metaplasticity, such that synaptic plasticity itself may vary
depending on the history of a neuron’s postsynaptic ac-
tivity (Abraham & Bear, 1996). Metaplasticity contributes
to network function and behavior and may be homeostatic
or nonhomeostatic. Homeostatic metaplasticity has
been demonstrated in human M1 for several combina-
tions of plasticity-inducing NIBS protocols (Karabanov
et al., 2015; Müller-Dahlhaus & Ziemann, 2015). For
instance, application of the same priming and test theta-
burst protocols may reverse the effect of the test pro-
tocol, whereas priming with the opposite protocol
was shown to increase the effect of a test protocol
(Mastroeni et al., 2013; Murakami, Müller-Dahlhaus, Lu,
& Ziemann, 2012). Notably, effects of metaplasticity in-
duced by two consecutive NIBS protocols strongly de-
pend (among other factors like stimulation intensity)
on the timing between priming and test protocol
(Müller-Dahlhaus & Ziemann, 2015), as the subsequent
combination of two similar theta-burst protocols may also
result in a nonhomeostatic additive effect on corticosp-
inal excitability (e.g., Goldsworthy, Müller-Dahlhaus,
Ridding, & Ziemann, 2015). At the behavioral level, it
was shown that the capacity of the motor cortex to un-
dergo LTP-like plasticity in response to paired associative
stimulation was abolished immediately after motor train-
ing (Stefan et al., 2006). These findings are particularly
relevant for cognitive neuroscience studies because many
training interventions are combined with NIBS protocols
to augment the effect of the behavioral intervention,
and the principle of homeostatic metaplasticity may
Bergmann and Hartwigsen
211
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
3
2
1
9
5
1
8
6
2
5
1
7
/
j
o
c
n
_
a
_
0
1
5
9
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
generalize across cortical areas (e.g., Bocci et al., 2014;
Gatica Tossi, Stude, Schwenkreis, Tegenthoff, & Dinse,
2013). Yet, the optimal timing between task and stimu-
lation remains unknown for most interventions, and po-
tential effects of metaplasticity are usually ignored. The
above-discussed results suggest that metaplasticity may
augment, diminish, or even reverse the expected direc-
tion of a given NIBS protocol. Although it remains large-
ly unclear how this affects cognitive processes, such
effects may contribute to unexpected results or null
findings of NIBS studies in the field of cognitive
neuroscience.
CONTROL CONDITIONS—HOW TO DESIGN A
VALID NIBS STUDY
Having outlined the numerous factors potentially inter-
rupting or confounding the chain of causation for a typ-
ical cognitive neuroscience NIBS study, we now discuss
the most important experimental control conditions.
Control conditions are used to control influences that
cannot be completely eliminated in NIBS experiments,
such as the unintentional costimulation of nontarget
brain regions or networks and peripheral sensory struc-
tures, the influence of the participants’ expectations
and beliefs regarding the NIBS procedure, and the inev-
itable dependence of task performance on additional
nontarget cognitive functions. Importantly, the control
conditions included in an experiment determine the con-
clusions that can be drawn from the results. The typical
rationale of NIBS experiments is that, if a given NIBS pro-
tocol, applied at a certain time point relative to a task,
affects behavioral performance, this is proof of the causal
relevance of the targeted area and/or neuronal activity
pattern for the cognitive function operationalized by
the task. Depending on the hypothesis, several explicit
(or implicit) assumptions are made regarding the speci-
ficity of the investigated brain–behavior relationship with
respect to the targeted anatomical location, temporal
window, and/or oscillatory frequency, as well as the af-
fected aspects of the task. For feasibility reasons, it is
rarely possible to realize all control conditions in a single
study, particularly not in a fully crossed factorial design.
However, any specificity claim needs to be explicitly
tested with an appropriate control condition.
Stimulation-free Condition
Earlier studies often included trials without stimulation
(i.e., “no TMS”) as a control condition, sometimes ran-
domly interleaved with effective stimulation to control
for carry-over and practice effects (Sandrini et al.,
2011). However, this does not control for any unspecific
side effects, and the difference between TMS and no TMS
will be obvious for the participant. To avoid carry-over ef-
fects between trials or conditions, the duration of
intertrial intervals and/or TMS trains may be adopted or
trials of similar conditions may be grouped in short
blocks.
Sham TMS
Sham TMS is meant to mimic sensory costimulation and
serve as a placebo condition to control for participants’
beliefs and expectations. Different sham TMS approaches
have been adopted, including physical separation of the
coil from the scalp with a spacer, placing an additional
coil with 90° tilt on top of the active coil and selectively
discharging the former as a sham condition, slightly tilt-
ing a coil on the scalp to avoid stimulation of the under-
lying brain region, employing a commercially available
sham coil, or developing other “realistic” sham condi-
tions. Moving the coil away from the scalp preserves
the airborne sound induced by coil charging but induces
little or no bone conduction and completely lacks so-
matosensory costimulation (ter Braack, de Vos, & van
Putten, 2015). Likewise, an additional coil on top or tilt-
ing the coil such that only the edge touches the head
produces roughly comparable auditory inputs compared
to active TMS, yet almost no somatosensory inputs be-
cause of a lack of peripheral nerve stimulation. The same
holds true for conventional sham coils that reduce the
effective stimulation intensity by shielding or opposed
current flow, which may prevent effective stimulation of
the target area but also of peripheral structures.
Consequently, participant blinding is hard to achieve with
these approaches. This is particularly problematic for
within-participant designs often used in cognitive neuro-
science studies. To overcome these limitations, “realistic”
sham conditions aim to also mimic the somatosensory
side effects. State-of-the-art realistic sham conditions thus
combine individually adjusted auditory noise masking to
attenuate the click sound for both active and sham con-
ditions (Duecker & Sack, 2015), foam padding beneath
the coil to attenuate vibration, a TMS coil discharging at
an ineffective distance to control for residual auditory in-
put, and individually adjusted electric stimulation of the
skin via surface electrodes beneath the TMS coil to mimic
peripheral nerve stimulation (Conde et al., 2019). Yet, even
if electric stimulation intensity is individually adjusted,
experienced participants likely notice the difference
between realistic sham and effective stimulation, as the
skin sensation of the electric stimulation will be different
(e.g., Mennemeier et al., 2009; Rossi et al., 2007). This is
particularly problematic when online TMS is applied at
higher intensities to areas where the stimulation may
yield unpleasant side effects (such as cranial/facial muscle
twitches). These side effects can substantially confound
the obtained results, as shown by a negative association
between working memory performance and individual
ratings of the unpleasantness of TMS (Abler et al.,
2005). An active control site is thus always preferable.
However, the choice of an active control site can be
212
Journal of Cognitive Neuroscience
Volume 33, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
3
2
1
9
5
1
8
6
2
5
1
7
/
j
o
c
n
_
a
_
0
1
5
9
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
tricky, especially if complex cognitive functions are stud-
ied that are widely distributed across the brain, preclud-
ing the use of most well-matched regions as control sites.
Sham TCS
In contrast to most TMS studies, TCS studies usually in-
clude sham conditions but no active control sites (i.e.,
control montages). Sham TCS is commonly realized by
ramping up the current to target intensity for, for exam-
ple, 10–30 sec and immediately ramping it down again
without any further active stimulation, thus producing
some cutaneous (tingling, itching, burning) sensations
in the beginning, when they are also strongest for real
TDCS before they habituate, supposedly making real
and sham TDCS indistinguishable (Gandiga, Hummel,
& Cohen, 2006). However, recent evidence shows that,
even if low stimulation intensities around 1 mA are used,
participant blinding is compromised (Greinacher, Buhôt,
Möller, & Learmonth, 2019; Turi et al., 2019). A recently
introduced sham TCS protocol combines a multielec-
trode montage with controlled shunting of currents via
a model-based quantification of transcutaneous and
transcranial effects, ensuring constant scalp sensations
across the whole stimulation procedure and similar sen-
sations relative to effective TCS (Neri et al., 2020). This
protocol was suggested to be superior in participant
blinding relative to conventional bifocal ramp-up, ramp-
down sham protocols and may provide a realistic sham
condition for TCS. As for TMS, carefully matched active
control montages are the gold standard also for TCS stud-
ies, and an additional sham session with the same mon-
tage should serve only as low-level baseline. In TDCS
studies, polarity specificity can be tested additionally by
applying both anodal and cathodal TDCS with respect
to the target area of interest.
Control via Ineffective Stimulation Protocols
For some NIBS protocols, it is possible to create an inef-
fective stimulation protocol, which can be applied to the
same region without causing the crucial neuronal effects.
For example, a change in intertrain duration made the in-
termediate theta burst stimulation protocol ineffective
(Huang et al., 2005), and replacing theta bursts (triplets)
with single pulses also produced no aftereffects ( Volman,
Roelofs, Koch, Verhagen, & Toni, 2011), since 200 pulses
at 5 Hz have likely no lasting effect (Peinemann et al.,
2004). The paired associative stimulation protocol
(Stefan, Kunesch, Cohen, Benecke, & Classen, 2000) be-
comes ineffective when using, imperceptibly for the par-
ticipant, a random selection of per se ineffective ISIs
(Bergmann et al., 2008). In general, a dose reduction
can make an ineffective protocol, for example, by reduc-
ing stimulation intensity, number of pulses, or total dura-
tion of the application. In addition, a change in TMS coil
orientation (usually from orthogonal to parallel with
respect to the target gyrus) can be sufficient to abolish
a specific effect, not only for MEPs evoked in M1 (Mills
et al., 1992) but possibly also for other brain regions
(Thielscher et al., 2011; Thut, Veniero, et al., 2011).
Except for a simple dose reduction, inevitably changing
the amount of sensory costimulation and, possibly, the
participants’ beliefs regarding the effectiveness of the
stimulation, these controls can be considered high-level
control conditions matching the effective experimental
protocol relatively well, although they come with the risk
of residual transcranial effects on the brain.
Control Tasks (Task Specificity)
Task specificity implies that, if a cortical area X is relevant
for a cognitive process A but not B, then a given NIBS
protocol over area X should selectively modulate task A
but not B (Miniussi et al., 2013). This requires a control
task that differs from the task of interest selectively in the
cognitive function of interest but is matched with respect
to task difficulty, low-level sensory input, and supporting
cognitive functions (e.g., perceptual, attentional, working
memory, executive, or motor demands). Without
evidence for task specificity, a NIBS protocol may simply
interfere with any of those cognitive functions instead,
potentially resulting in the same effect on behavioral per-
formance. Importantly, some tasks (e.g., visual attention
or orienting tasks) may also be more strongly affected by
sensory costimulation.
Control Regions (Anatomical Specificity)
Anatomical specificity means that a NIBS protocol only
affects task A when applied to area X but not area Y, be-
cause only the former causally contributes to the cogni-
tive function of interest. An active TMS control site or
TCS electrode montage is thus needed to demonstrate
that the observed effects actually depend on the stimula-
tion of a specific brain region and not only of the brain
per se or its sensory input structures. Numerous studies
have used the vertex as an active control site, with
the rationale that auditory and somatosensory inputs
should be roughly similar to that of other target sites,
but the brain tissue is located deeper under the scalp
and thought to mainly contain sensorimotor representa-
tions of the lower body, thus not influencing cognitive task
performance ( Jung, Bungert, Bowtell, & Jackson, 2016;
Duecker, de Graaf, Jacobs, & Sack, 2013). However, de-
pending on the target area of interest, differences in lat-
eralization and unpleasantness are a potential issue. For
instance, lateralized sham TMS was shown to pull covert
spatial attention toward the corresponding side of space,
thereby facilitating target detection in this hemifield
(Duecker & Sack, 2013), which can hardly be achieved
with vertex stimulation. TMS of pFC and (anterior) tem-
poral cortex can be particularly unpleasant because of
costimulation of the facial nerves and muscles, and vertex
Bergmann and Hartwigsen
213
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
3
2
1
9
5
1
8
6
2
5
1
7
/
j
o
c
n
_
a
_
0
1
5
9
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
stimulation might not be an adequate control site for these
areas. Moreover, using simultaneous TMS–fMRI, it was dem-
onstrated that suprathreshold low-frequency rTMS ap-
plied over the vertex induced widespread deactivations in
the default mode network, although the BOLD signal in
the stimulated area was not affected ( Jung et al., 2016).
Although the origin of these remote effects remains un-
clear, the implication is that the vertex may not be a suit-
able control site for tasks that involve or interact with the
default mode network. Given the complex interaction of
the task positive network and the default mode network,
this would preclude the use of vertex stimulation for
most cognitive functions, especially those directly associated
with default mode engagement such as semantic process-
ing, social cognition, autobiographical memory, self-
related thinking, and consciousness (e.g., Binder, Desai,
Graves, & Conant, 2009; Buckner, Andrews-Hanna, &
Schacter, 2008). For some areas, homologous regions
may be adequate active control sites, especially if later-
alization of a particular cognitive function is of interest.
Yet, TMS may also affect contralateral areas via transcallo-
sal connections (cf. Arrow 3), especially if high intensities
or frequencies are used. For some cognitive processes,
lateralization is less clear, and the homologous region
might contribute to a given task. The choice of an active
control site that is matched for the most important dimen-
sions can be guided by a recently introduced atlas for TMS
studies (Meteyard & Holmes, 2018). Also for TCS, control
montages, well matched for peripheral costimulation
effects, should be mandatory, allowing at least some
degree of anatomical specificity to be claimed. For
offline NIBS protocols, different stimulation sites should
be targeted in different sessions several days apart to
prevent any carry-over effects of the stimulation.
Control Time Points (Temporal Specificity)
Depending on the research question and the applied
NIBS protocol, temporal specificity can be crucial.
Indeed, chronometric approaches have substantially ad-
vanced the current knowledge on the time course of dif-
ferent cognitive processes such as visual perception
(Amassian et al., 1989), visual orientation and awareness
(de Graaf, Duecker, Fernholz, & Sack, 2015; Jacobs,
Goebel, & Sack, 2012), motion-driven attention
(Alexander, Laycock, Crewther, & Crewther, 2018),
sound localization (At, Spierer, & Clarke, 2011), working
memory (Mottaghy, Gangitano, Krause, & Pascual-Leone,
2003), or language (Schuhmann, Schiller, Goebel, &
Sack, 2012). Most chronometric TMS studies that system-
atically vary the time point of stimulation argue that no
control site is needed because specificity is shown by
the difference between time points. However, this as-
sumption is invalid, because online TMS given immedi-
ately before or with stimulus onset acts as an alerting
signal and causes unspecific intersensory facilitation ef-
fects on response speed (Duecker et al., 2013; Terao
et al., 1997). If TMS occurs after the stimulus in some
trials, response speed can be delayed because of expec-
tancy violations causing the participant to “wait” for the
TMS pulse (de Graaf, Jacobs, Roebroeck, & Sack, 2009).
Again, temporal controls cannot replace an active control
site, as they are designed to reveal different specificities.
Control Frequencies (Frequency Specificity)
Frequency specificity is ignored in most NIBS studies.
However, for some research questions, this may be cru-
cial, especially if conclusions are drawn about entrain-
ment or plastic aftereffects of rhythmic NIBS protocols.
For instance, in a hypothetical experiment, the conclusion
that beta-TACS over the pFC affects working memory
would only be valid if one could show these effects to
be frequency specific (including control frequencies), in
addition to the anatomical specificity (including a control
montage and sham stimulation as baseline) and task
specificity (including a well-matched control task).
Indeed, frequency specificity is often considered in cog-
nitive neuroscience studies employing TACS, but many
studies still simply compare a frequency of interest
against sham stimulation, which does not justify any
conclusion regarding the relevance of the stimulation
frequency. Some studies have explicitly employed
symmetrical control frequencies below and above the tar-
get frequency (Herring et al., 2019; Romei et al., 2016) to
prevent confounds with stimulation duration or number
of cycles. Yet, no clear procedure has been established to
define the number of control frequencies or the distance
from the frequency of interest (Herrmann et al., 2013).
Obviously, the control frequencies should not be en-
gaged in the task of interest. Individual adjustment of
the stimulation frequency may be favorable, at least in
the alpha band, although it is still unclear whether stim-
ulation is more effective when it matches the eigenfre-
quency of the brain (Reato, Rahman, Bikson, & Parra,
2013) or is slightly different ( Vossen et al., 2015;
Helfrich, Knepper, et al., 2014). Notably, TACS can in-
duce rhythmic retinal phosphenes and cutaneous sensa-
tions at most frequencies, although with different
thresholds (Kanai, Chaieb, Antal, Walsh, & Paulus,
2008), highlighting again that frequency controls alone
are not sufficient and control montages need to be
employed.
Further Considerations
To guarantee accurate coil or electrode placement and
maintenance across the experiment and avoid confounds
induced by movement of the stimulation device, the use
of a stereotactic neuronavigation system and individual
T1-weighted images is highly recommended. In general,
TCS studies benefit from optimized montages or compu-
tationally optimized multichannel arrangements that may
help to focalize the stimulated area and minimize
214
Journal of Cognitive Neuroscience
Volume 33, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
3
2
1
9
5
1
8
6
2
5
1
7
/
j
o
c
n
_
a
_
0
1
5
9
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
unwanted peripheral costimulation (e.g., Khatoun et al.,
2018). E-field modeling should be employed to estimate
the focality of electrode montages or coil placement,
minimize the impact of noncortical stimulation, and op-
timize stimulation efficiency. As many other studies in
the field of cognitive neuroscience, NIBS studies often
suffer from relatively small and homogeneous samples
(i.e., mainly healthy young student volunteers, with sam-
ple sizes < 30 participants), which are not representative
of the general population. A priori calculations of statisti-
cal power may guide sample size selection, and we prin-
cipally advocate larger sample sizes, but for most NIBS
studies, the effect size is unknown beforehand. Whenever
possible, NIBS experiments should be conducted as a
within-participant design to reduce interindividual vari-
ability and confounds based on imperfect randomization
of group membership. If control conditions are realized
in a between-participant design (e.g., for a post hoc control
experiment) the control condition needs to have the same
sample size as the experimental condition to rule out
statistical power as a confound.
Twelve General Recommendations for Designing
Valid NIBS Studies
As a summary, we make the following 12 general recom-
mendations to be considered when designing a NIBS
study in cognitive neuroscience (the applicability of these
recommendations may vary depending on the specifics
of your research question as well as the technical/logistic
feasibility in your laboratory):
1. Know your target! Ensure you have identified the
stimulation target with a spatial precision appropri-
ate for your NIBS technique of choice. Consider an
fMRI-based localizer for target identification if
possible.
2. Simulate the E-field! Use individual E-field modeling
based on realistic MR-based head models to corrob-
orate that the E-field is maximal at the target site
(sensitivity) and as limited to it as possible
(specificity).
3. Adjust stimulation intensity! Stimulation intensity
should be adjusted individually, taking coil–cortex
distance into account, even if no ideal reference val-
ue exists yet (% RMT is established). The estimation
of induced E-field strength is not (yet) state-of-the-
art but recommended.
4. Use neuronavigation! Individual MR-based neurona-
vigation ensures precise TMS coil/TCS electrode
placement and maintenance within and across ses-
sions and is a basic prerequisite for high-quality
TMS and high-definition TCS studies.
5. Combine NIBS with neuroimaging! Use neuroimag-
ing (e.g., fMRI, EEG/MEG) to establish proof or tar-
get engagement, that is, demonstrate that the
desired neuronal effects have been induced in the
6.
7.
8.
9.
target region/network and that no unintended coac-
tivations of other regions/networks can confound
the results. This also allows to relate NIBS-induced
neuronal and behavioral effects.
Include a control site! An active control site (TMS) or
montage (TCS), well matched for sensations and an-
noyance, is a strong control for sensory costimula-
tion confounds and serves to establish anatomical
specificity. An additional (realistic) sham condition
as baseline is optimal.
Include a control task! An appropriate control task,
which does not involve the cognitive function of in-
terest but is matched for task difficulty, is a strong
control for possible confounding via coaffected sup-
porting cognitive functions (e.g., attention) and
serves to establish task specificity.
Include control time windows (if applicable)! If the
timing of the target neuronal process matters (e.g.,
when doing mental chronometry), stimulation at
different time points/windows is necessary to estab-
lish temporal specificity.
Include control frequencies (if applicable)! If you
want to demonstrate entrainment or similar
frequency-specific effects, neighboring control fre-
quencies (ideally both lower and higher) are manda-
tory. An arrhythmic control stimulation can control
for the number of stimulation pulses/cycles and the
sheer presence of rhythmicity but cannot establish
frequency specificity.
10. Reduce variability wherever possible! Within-
participant designs reduce interindividual variability,
but between-participant designs are necessary
when repeated stimulation or task performance is
problematic for reasons of blinding or learning ef-
fects. Internal and external contextual factors (e.g.,
time of day, arousal) should be kept as comparable
as possible between conditions.
11. Prevent carry-over and order effects! For offline
NIBS protocols, control conditions need to be con-
ducted in separate sessions several days apart and
counterbalanced to ensure that previously induced
synaptic plasticity cannot systematically interact
with the current protocol.
12. Choose your DV wisely! Thoroughly consider the
outcome measure best reflecting the expected
change in cognitive function. Titrate task difficulty
individually to a level where performance becomes
sensitive to even small disturbances of underlying
computation.
Conclusion
We have outlined major challenges and potential pitfalls
for experimentally testing and interpreting the chain of
causation for NIBS studies in cognitive neuroscience.
We hope to raise awareness for the potential confounds
Bergmann and Hartwigsen
215
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
2
1
9
5
1
8
6
2
5
1
7
/
j
o
c
n
_
a
_
0
1
5
9
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
and provide a guide for designing valid NIBS experi-
ments. On the basis of the above-discussed studies, a
promising avenue for the future will be the multimethod
combination of NIBS with computational modeling and
neuroimaging to map stimulation-induced changes at
the neuronal and network level and link these changes
with cognitive and behavioral effects. With respect to
computational modeling approaches, particular chal-
lenges include the modulation of the dynamics of long-
term plastic aftereffects of different NIBS protocols and
the transfer from relatively easy decision-making processes
to more complex cognitive functions (e.g., language,
social cognition, or problem solving). Recent advances in
behavioral modeling may further help to bridge the gap
between cognitive theories and behavioral outcome
measures of NIBS experiments, and the inclusion of more
natural tasks or stimuli will increase the ecological validity.
To deepen our understanding of the modulatory effects of
NIBS protocols, future studies should relate the simulated
strength of the induced E-field in the target area to
neuroimaging-based assessments of target engagement
and behavioral outcome measures. As NIBS studies are
complex and time consuming and the modulatory effects
are often small and variable across participants, a particular
challenge for the future is the inclusion of larger sample
sizes to guarantee sufficient experimental power. Here,
multicenter approaches may identify valid paradigms for
NIBS studies that may provide further insight into causal
structure–function relationships.
Acknowledgments
T.O.B. was supported by the Boehringer Ingelheim Foundation.
G.H. was supported by the Max Planck Society.
Reprint requests should be sent to Til Ole Bergmann, Leibniz
Institute for Resilience Research, Wallstraße 7-9, 55122 Mainz,
Germany, or via e-mail: til-ole.bergmann@lir-mainz.de.
REFERENCES
Aberra, A. S., Wang, B., Grill, W. M., & Peterchev, A. V. (2020).
Simulation of transcranial magnetic stimulation in head
model with morphologically-realistic cortical neurons. Brain
Stimulation, 13, 175–189. DOI: https://doi.org/10.1016/j.brs
.2019.10.002, PMID: 31611014
Abler, B., Walter, H., Wunderlich, A., Grothe, J., Schönfeldt-
Lecuona, C., Spitzer, M., et al. (2005). Side effects of
transcranial magnetic stimulation biased task performance
in a cognitive neuroscience study. Brain Topography, 17,
193–196. DOI: https://doi.org/10.1007/s10548-005-6028-y,
PMID: 16110769
Abraham, W. C., & Bear, M. F. (1996). Metaplasticity: The plasticity
of synaptic plasticity. Trends in Neurosciences, 19, 126–130.
DOI: https://doi.org/10.1016/S0166-2236(96)80018-X
Abrahamyan, A., Clifford, C. W. G., Arabzadeh, E., & Harris, J. A.
(2011). Improving visual sensitivity with subthreshold
transcranial magnetic stimulation. Journal of Neuroscience,
31, 3290–3294. DOI: https://doi.org/10.1523/JNEUROSCI.6256
-10.2011, PMID: 21368040, PMCID: PMC6623934
Alekseichuk, I., Falchier, A. Y., Linn, G., Xu, T., Milham, M. P.,
Schroeder, C. E., et al. (2019). Electric field dynamics in the
brain during multi-electrode transcranial electric stimulation.
Nature Communications, 10, 2573. DOI: https://doi.org
/10.1038/s41467-019-10581-7, PMID: 31189931, PMCID:
PMC6561925
Alekseichuk, I., Mantell, K., Shirinpour, S., & Opitz, A. (2019).
Comparative modeling of transcranial magnetic and electric
stimulation in mouse, monkey, and human. Neuroimage,
194, 136–148. DOI: https://doi.org/10.1016/j.neuroimage
.2019.03.044, PMID: 30910725, PMCID: PMC6536349
Alekseichuk, I., Turi, Z., Amador de Lara, G., Antal, A., & Paulus,
W. (2016). Spatial working memory in humans depends on
theta and high gamma synchronization in the prefrontal
cortex. Current Biology, 26, 1513–1521. DOI: https://doi.org
/10.1016/j.cub.2016.04.035, PMID: 27238283
Alexander, B., Laycock, R., Crewther, D. P., & Crewther, S. G.
(2018). An fMRI-neuronavigated chronometric TMS
investigation of V5 and intraparietal cortex in motion driven
attention. Frontiers in Human Neuroscience, 11, 638. DOI:
https://doi.org/10.3389/fnhum.2017.00638, PMID: 29354043,
PMCID: PMC5758491
Amassian, V. E., Cracco, R. Q., Maccabee, P. J., Cracco, J. B.,
Rudell, A., & Eberle, L. (1989). Suppression of visual perception
by magnetic coil stimulation of human occipital cortex.
Electroencephalography and Clinical Neurophysiology, 74,
458–462. DOI: https://doi.org/10.1016/0168-5597(89)90036-1
Amassian, V. E., Cracco, R. Q., Maccabee, P. J., Cracco, J. B.,
Rudell, A. P., & Eberle, L. (1998). Transcranial magnetic
stimulation in study of the visual pathway. Journal of Clinical
Neurophysiology, 15, 288–304. DOI: https://doi.org/10.1097
/00004691-199807000-00002, PMID: 9736464
Antal, A., Boros, K., Poreisz, C., Chaieb, L., Terney, D., & Paulus,
W. (2008). Comparatively weak after-effects of transcranial
alternating current stimulation (tACS) on cortical excitability
in humans. Brain Stimulation, 1, 97–105. DOI: https://doi
.org/10.1016/j.brs.2007.10.001, PMID: 20633376
Antal, A., & Herrmann, C. S. (2016). Transcranial alternating
current and random noise stimulation: Possible mechanisms.
Neural Plasticity, 2016, 3616807. DOI: https://doi.org/10
.1155/2016/3616807, PMID: 27242932, PMCID: PMC4868897
Antal, A., Polania, R., Schmidt-Samoa, C., Dechent, P., & Paulus, W.
(2011). Transcranial direct current stimulation over the primary
motor cortex during fMRI. Neuroimage, 55, 590–596. DOI:
https://doi.org/10.1016/j.neuroimage.2010.11.085, PMID:
21211569
Asamoah, B., Khatoun, A., & McLaughlin, M. (2019). tACS motor
system effects can be caused by transcutaneous stimulation
of peripheral nerves. Nature Communications, 10, 266.
DOI: https://doi.org/10.1038/s41467-018-08183-w, PMID:
30655523, PMCID: PMC6336776
At, A., Spierer, L., & Clarke, S. (2011). The role of the right
parietal cortex in sound localization: A chronometric single
pulse transcranial magnetic stimulation study. Neuropsychologia,
49, 2794–2797. DOI: https://doi.org/10.1016/j.neuropsychologia
.2011.05.024, PMID: 21679720
Barker, A. T., Jalinous, R., & Freeston, I. L. (1985). Non-invasive
magnetic stimulation of human motor cortex. Lancet, 325,
1106–1107. DOI: https://doi.org/10.1016/S0140-6736(85)
92413-4
Batsikadze, G., Moliadze, V., Paulus, W., Kuo, M.-F., & Nitsche,
M. A. (2013). Partially non-linear stimulation intensity-
dependent effects of direct current stimulation on motor
cortex excitability in humans. Journal of Physiology, 591,
1987–2000. DOI: https://doi.org/10.1113/jphysiol.2012
.249730, PMID: 23339180, PMCID: PMC3624864
Beaulieu, L.-D., Flamand, V. H., Massé-Alarie, H., & Schneider,
C. (2017). Reliability and minimal detectable change of
216
Journal of Cognitive Neuroscience
Volume 33, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
2
1
9
5
1
8
6
2
5
1
7
/
j
o
c
n
_
a
_
0
1
5
9
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
transcranial magnetic stimulation outcomes in healthy adults:
A systematic review. Brain Stimulation, 10, 196–213. DOI:
https://doi.org/10.1016/j.brs.2016.12.008, PMID: 28031148
Bergmann, T. O. (2018). Brain state-dependent brain stimulation.
Frontiers in Psychology, 9, 2108. DOI: https://doi.org/10.3389
/fpsyg.2018.02108, PMID: 30443236, PMCID: PMC6221926
Bergmann, T. O., & Born, J. (2018). Phase–amplitude coupling:
A general mechanism for memory processing and synaptic
plasticity? Neuron, 97, 10–13. DOI: https://doi.org/10.1016
/j.neuron.2017.12.023, PMID: 29301097
Bergmann, T. O., Karabanov, A., Hartwigsen, G., Thielscher, A.,
& Siebner, H. R. (2016). Combining non-invasive transcranial
brain stimulation with neuroimaging and electrophysiology:
Current approaches and future perspectives. Neuroimage,
140, 4–19. DOI: https://doi.org/10.1016/j.neuroimage.2016
.02.012, PMID: 26883069
Bergmann, T. O., Lieb, A., Zrenner, C., & Ziemann, U. (2019).
Pulsed facilitation of corticospinal excitability by the
sensorimotor μ-alpha rhythm. Journal of Neuroscience, 39,
10034–10043. DOI: https://doi.org/10.1523/JNEUROSCI
.1730-19.2019, PMID: 31685655, PMCID: PMC6978939
Bergmann, T. O., Mölle, M., Marshall, L., Kaya-Yildiz, L., Born, J.,
& Siebner, H. R. (2008). A local signature of LTP- and LTD-
like plasticity in human NREM sleep. European Journal of
Neuroscience, 27, 2241–2249. DOI: https://doi.org/10.1111
/j.1460-9568.2008.06178.x, PMID: 18445215
Bergmann, T. O., Mölle, M., Schmidt, M. A., Lindner, C.,
Marshall, L., Born, J., et al. (2012). EEG-guided transcranial
magnetic stimulation reveals rapid shifts in motor cortical
excitability during the human sleep slow oscillation. Journal
of Neuroscience, 32, 243–253. DOI: https://doi.org/10.1523
/JNEUROSCI.4792-11.2012, PMID: 22219286, PMCID:
PMC6621327
Bestmann, S., Baudewig, J., Siebner, H. R., Rothwell, J. C., &
Frahm, J. (2003). Subthreshold high-frequency TMS of
human primary motor cortex modulates interconnected
frontal motor areas as detected by interleaved fMRI–TMS.
Neuroimage, 20, 1685–1696. DOI: https://doi.org/10.1016
/j.neuroimage.2003.07.028, PMID: 14642478
Bestmann, S., Baudewig, J., Siebner, H. R., Rothwell, J. C., &
Frahm, J. (2005). BOLD MRI responses to repetitive TMS over
human dorsal premotor cortex. Neuroimage, 28, 22–29.
DOI: https://doi.org/10.1016/j.neuroimage.2005.05.027,
PMID: 16002305
Bestmann, S., & Feredoes, E. (2013). Combined neurostimulation
and neuroimaging in cognitive neuroscience: Past, present,
and future. Annals of the New York Academy of Sciences, 1296,
11–30. DOI: https://doi.org/10.1111/nyas.12110, PMID:
23631540, PMCID: PMC3760762
Bestmann, S., Swayne, O., Blankenburg, F., Ruff, C. C., Haggard,
P., Weiskopf, N., et al. (2008). Dorsal premotor cortex exerts
state-dependent causal influences on activity in contralateral
primary motor and dorsal premotor cortex. Cerebral Cortex,
18, 1281–1291. DOI: https://doi.org/10.1093/cercor/bhm159,
PMID: 17965128, PMCID: PMC2600427
Binder, J. R., Desai, R. H., Graves, W. W., & Conant, L. L. (2009).
Where is the semantic system? A critical review and meta-
analysis of 120 functional neuroimaging studies. Cerebral
Cortex, 19, 2767–2796. DOI: https://doi.org/10.1093/cercor
/bhp055, PMID: 19329570, PMCID: PMC2774390
Bindman, L. J., Lippold, O. C. J., & Redfearn, J. W. T. (1964). The
action of brief polarizing currents on the cerebral cortex of
the rat (1) during current flow and (2) in the production of
long-lasting after-effects. Journal of Physiology, 172, 369–382.
DOI: https://doi.org/10.1113/jphysiol.1964.sp007425, PMID:
14199369, PMCID: PMC1368854
Bocci, T., Caleo, M., Tognazzi, S., Francini, N., Briscese, L.,
Maffei, L., et al. (2014). Evidence for metaplasticity in the
human visual cortex. Journal of Neural Transmission, 121,
221–231. DOI: https://doi.org/10.1007/s00702-013-1104-z,
PMID: 24162796
Bonaiuto, J. J., & Bestmann, S. (2015). Understanding the
nonlinear physiological and behavioral effects of tDCS
through computational neurostimulation. Progress in Brain
Research, 222, 75–103. DOI: https://doi.org/10.1016/bs.pbr
.2015.06.013, PMID: 26541377
Bonaiuto, J. J., de Berker, A., & Bestmann, S. (2016). Response
repetition biases in human perceptual decisions are explained
by activity decay in competitive attractor models. eLife, 5,
e20047. DOI: https://doi.org/10.7554/eLife.20047, https://doi
.org/10.7554/eLife.20047.025
Brasil-Neto, J. P., McShane, L. M., Fuhr, P., Hallett, M., & Cohen,
L. G. (1992). Topographic mapping of the human motor
cortex with magnetic stimulation: Factors affecting accuracy
and reproducibility. Electroencephalography and Clinical
Neurophysiology, 85, 9–16. DOI: https://doi.org/10.1016
/0168-5597(92)90095-S
Buckner, R. L., Andrews-Hanna, J. R., & Schacter, D. L. (2008).
The brain’s default network: Anatomy, function, and
relevance to disease. Annals of the New York Academy of
Sciences, 1124, 1–38. DOI: https://doi.org/10.1196/annals
.1440.011, PMID: 18400922
Bungert, A., Antunes, A., Espenhahn, S., & Thielscher, A.
(2017). Where does TMS stimulate the motor cortex?
Combining electrophysiological measurements and realistic
field estimates to reveal the affected cortex position. Cerebral
Cortex, 27, 5083–5094. DOI: https://doi.org/10.1093/cercor
/bhw292, PMID: 27664963
Buzsáki, G., & Draguhn, A. (2004). Neuronal oscillations in
cortical networks. Science, 304, 1926–1929. DOI: https://doi
.org/10.1126/science.1099745, PMID: 15218136
Capotosto, P., Babiloni, C., Romani, G. L., & Corbetta, M. (2012).
Differential contribution of right and left parietal cortex to
the control of spatial attention: A simultaneous EEG–rTMS
study. Cerebral Cortex, 22, 446–454. DOI: https://doi.org
/10.1093/cercor/bhr127, PMID: 21666126, PMCID:
PMC3256411
Cattaneo, L., Barchiesi, G., Tabarelli, D., Arfeller, C., Sato, M., &
Glenberg, A. M. (2011). One’s motor performance
predictably modulates the understanding of others’ actions
through adaptation of premotor visuo-motor neurons. Social
Cognitive and Affective Neuroscience, 6, 301–310. DOI:
https://doi.org/10.1093/scan/nsq099, PMID: 21186167,
PMCID: PMC3110437
Chambliss, B. (2018). The mind–body problem. Wiley
Interdisciplinary Reviews: Cognitive Science, 9, e1463.
DOI: https://doi.org/10.1002/wcs.1463, PMID: 29727520
Chen, J.-Y., Chauvette, S., Skorheim, S., Timofeev, I., &
Bazhenov, M. (2012). Interneuron-mediated inhibition
synchronizes neuronal activity during slow oscillation.
Journal of Physiology, 590, 3987–4010. DOI: https://doi.org
/10.1113/jphysiol.2012.227462, PMID: 22641778, PMCID:
PMC3476644
Chen, R., Lozano, A. M., & Ashby, P. (1999). Mechanism of the
silent period following transcranial magnetic stimulation:
Evidence from epidural recordings. Experimental Brain
Research, 128, 539–542. DOI: https://doi.org/10.1007
/s002210050878, PMID: 10541749
Cohen, M. X. (2017). Where does EEG come from and what
does it mean? Trends in Neurosciences, 40, 208–218. DOI:
https://doi.org/10.1016/j.tins.2017.02.004, PMID: 28314445
Conde, V., Tomasevic, L., Akopian, I., Stanek, K., Saturnino, G. B.,
Thielscher, A., et al. (2019). The non-transcranial TMS-
evoked potential is an inherent source of ambiguity in TMS–
EEG studies. Neuroimage, 185, 300–312. DOI: https://doi
.org/10.1016/j.neuroimage.2018.10.052, PMID: 30347282
Bergmann and Hartwigsen
217
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
2
1
9
5
1
8
6
2
5
1
7
/
j
o
c
n
_
a
_
0
1
5
9
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Datta, A., Bansal, V., Diaz, J., Patel, J., Reato, D., & Bikson, M.
(2009). Gyri-precise head model of transcranial direct current
stimulation: Improved spatial focality using a ring electrode
versus conventional rectangular pad. Brain Stimulation, 2,
201–207. DOI: https://doi.org/10.1016/j.brs.2009.03.005,
PMID: 20648973, PMCID: PMC2790295
Davare, M., Lemon, R., & Olivier, E. (2008). Selective
modulation of interactions between ventral premotor cortex
and primary motor cortex during precision grasping in
humans. Journal of Physiology, 586, 2735–2742. DOI:
https://doi.org/10.1113/jphysiol.2008.152603, PMID:
18403420, PMCID: PMC2536583
de Graaf, T. A., Duecker, F., Fernholz, M. H. P., & Sack, A. T.
(2015). Spatially specific vs. unspecific disruption of visual
orientation perception using chronometric pre-stimulus TMS.
Frontiers in Behavioral Neuroscience, 9, 5. DOI: https://
doi.org/10.3389/fnbeh.2015.00005, PMID: PMC4311643
de Graaf, T. A., Jacobs, C., Roebroeck, A., & Sack, A. T. (2009).
fMRI effective connectivity and TMS chronometry:
Complementary accounts of causality in the visuospatial
judgment network. PLoS One, 4, e8307. DOI: https://doi.org
/10.1371/journal.pone.0008307, PMID: 20011541, PMCID:
PMC2789405
Dijkstra, N., & de Bruin, L. (2016). Cognitive neuroscience and
causal inference: Implications for psychiatry. Frontiers in
Psychiatry, 7, 129. DOI: https://doi.org/10.3389/fpsyt.2016
.00129, PMID: 27486408, PMCID: PMC4949233
Di Lazzaro, V., & Ziemann, U. (2013). The contribution of
transcranial magnetic stimulation in the functional evaluation
of microcircuits in human motor cortex. Frontiers in Neural
Circuits, 7, 18. DOI: https://doi.org/10.3389/fncir.2013
.00018, PMID: 23407686, PMCID: PMC3570771
Di Lazzaro, V., Ziemann, U., & Lemon, R. N. (2008). State of
the art: Physiology of transcranial motor cortex stimulation.
Brain Stimulation, 1, 345–362. DOI: https://doi.org/10.1016
/j.brs.2008.07.004, PMID: 20633393
Dockery, C. A., Hueckel-Weng, R., Birbaumer, N., & Plewnia, C.
(2009). Enhancement of planning ability by transcranial direct
current stimulation. Journal of Neuroscience, 29, 7271–7277.
DOI: https://doi.org/10.1523/JNEUROSCI.0065-09.2009, PMID:
19494149, PMCID: PMC6666475
Duecker, F., de Graaf, T. A., Jacobs, C., & Sack, A. T. (2013).
Time- and task-dependent non-neural effects of real and
sham TMS. PLoS One, 8, e73813. DOI: https://doi.org/10
.1371/journal.pone.0073813, PMID: 24040080, PMCID:
PMC3763998
Duecker, F., Frost, M. A., de Graaf, T. A., Graewe, B., Jacobs, C.,
Goebel, R., et al. (2014). The cortex-based alignment approach
to TMS coil positioning. Journal of Cognitive Neuroscience,
26, 2321–2329. DOI: https://doi.org/10.1162/jocn_a_00635,
PMID: 24702449
Ermentrout, G. B., Galán, R. F., & Urban, N. N. (2008). Reliability,
synchrony and noise. Trends in Neurosciences, 31, 428–434.
DOI: https://doi.org/10.1016/j.tins.2008.06.002, PMID:
18603311, PMCID: PMC2574942
Esmaeilpour, Z., Marangolo, P., Hampstead, B. M., Bestmann, S.,
Galletta, E., Knotkova, H., et al. (2018). Incomplete evidence
that increasing current intensity of tDCS boosts outcomes.
Brain Stimulation, 11, 310–321. DOI: https://doi.org/10.1016
/j.brs.2017.12.002, PMID: 29258808, PMCID: PMC7050474
Faisal, A. A., Selen, L. P. J., & Wolpert, D. M. (2008). Noise in the
nervous system. Nature Reviews Neuroscience, 9, 292–303.
DOI: https://doi.org/10.1038/nrn2258, PMID: 18319728,
PMCID: PMC2631351
Ferbert, A., Priori, A., Rothwell, J. C., Day, B. L., Colebatch, J. G.,
& Marsden, C. D. (1992). Interhemispheric inhibition of the
human motor cortex. Journal of Physiology, 453, 525–546.
DOI: https://doi.org/10.1113/jphysiol.1992.sp019243, PMID:
1464843, PMCID: PMC1175572
Fertonani, A., & Miniussi, C. (2017). Transcranial electrical
stimulation: What we know and do not know about
mechanisms. Neuroscientist, 23, 109–123. DOI: https://doi
.org/10.1177/1073858416631966, PMID: 26873962, PMCID:
PMC5405830
Filmer, H. L., Mattingley, J. B., & Dux, P. E. (2020). Modulating
brain activity and behaviour with tDCS: Rumours of its death
have been greatly exaggerated. Cortex, 123, 141–151. DOI:
https://doi.org/10.1016/j.cortex.2019.10.006, PMID: 31783223
Fitzgerald, P. B., Fountain, S., & Daskalakis, Z. J. (2006). A
comprehensive review of the effects of rTMS on motor
cortical excitability and inhibition. Clinical Neurophysiology,
117, 2584–2596. DOI: https://doi.org/10.1016/j.clinph.2006
.06.712, PMID: 16890483
Flitman, S. S., Grafman, J., Wassermann, E. M., Cooper, V.,
O’Grady, J., Pascual-Leone, A., et al. (1998). Linguistic processing
during repetitive transcranial magnetic stimulation. Neurology,
50, 175–181. DOI: https://doi.org/10.1212/WNL.50.1.175,
PMID: 9443476
Fomenko, A., Neudorfer, C., Dallapiazza, R. F., Kalia, S. K.,
& Lozano, A. M. (2018). Low-intensity ultrasound
neuromodulation: An overview of mechanisms and emerging
human applications. Brain Stimulation, 11, 1209–1217.
DOI: https://doi.org/10.1016/j.brs.2018.08.013, PMID:
30166265
Fries, P. (2015). Rhythms for cognition: Communication
through coherence. Neuron, 88, 220–235. DOI: https://
doi.org/10.1016/j.neuron.2015.09.034, PMID: 26447583,
PMCID: PMC4605134
Fröhlich, F., & McCormick, D. A. (2010). Endogenous electric
fields may guide neocortical network activity. Neuron, 67,
129–143. DOI: https://doi.org/10.1016/j.neuron.2010.06.005,
PMID: 20624597, PMCID: PMC3139922
Duecker, F., & Sack, A. T. (2013). Pre-stimulus sham TMS
Gandiga, P. C., Hummel, F. C., & Cohen, L. G. (2006).
facilitates target detection. PLoS One, 8, e57765. DOI: https://
doi.org/10.1371/journal.pone.0057765, PMID: 23469232,
PMCID: PMC3587629
Duecker, F., & Sack, A. T. (2015). Rethinking the role of sham
TMS. Frontiers in Psychology, 6, 210. DOI: https://doi.org
/10.3389/fpsyg.2015.00210, PMID: 25767458, PMCID:
PMC4341423
Edelman, G. M., & Gally, J. A. (2001). Degeneracy and complexity
in biological systems. Proceedings of the National Academy
of Sciences, U.S.A., 98, 13763–13768. DOI: https://doi.org
/10.1073/pnas.231499798, PMID: 11698650, PMCID:
PMC61115
Ellison, A., & Cowey, A. (2009). Differential and co-involvement
of areas of the temporal and parietal streams in visual tasks.
Neuropsychologia, 47, 1609–1614. DOI: https://doi.org/10
.1016/j.neuropsychologia.2008.12.013, PMID: 19133279
Transcranial DC stimulation (tDCS): A tool for double-blind
sham-controlled clinical studies in brain stimulation. Clinical
Neurophysiology, 117, 845–850. DOI: https://doi.org/10
.1016/j.clinph.2005.12.003, PMID: 16427357
Gatica Tossi, M. A., Stude, P., Schwenkreis, P., Tegenthoff, M., &
Dinse, H. R. (2013). Behavioural and neurophysiological
markers reveal differential sensitivity to homeostatic
interactions between centrally and peripherally applied
passive stimulation. European Journal of Neuroscience,
38, 2893–2901. DOI: https://doi.org/10.1111/ejn.12293,
PMID: 23834757
Gebodh, N., Esmaeilpour, Z., Adair, D., Chelette, K.,
Dmochowski, J., Woods, A. J., et al. (2019). Inherent
physiological artifacts in EEG during tDCS. Neuroimage,
185, 408–424. DOI: https://doi.org/10.1016/j.neuroimage
.2018.10.025, PMID: 30321643, PMCID: PMC6289749
218
Journal of Cognitive Neuroscience
Volume 33, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
2
1
9
5
1
8
6
2
5
1
7
/
j
o
c
n
_
a
_
0
1
5
9
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Goldsworthy, M. R., Müller-Dahlhaus, F., Ridding, M. C., &
Ziemann, U. (2015). Resistant against de-depression: LTD-
like plasticity in the human motor cortex induced by spaced
cTBS. Cerebral Cortex, 25, 1724–1734. DOI: https://doi.org
/10.1093/cercor/bht353, PMID: 24488942
Gomez-Tames, J., Hamasaka, A., Laakso, I., Hirata, A., & Ugawa, Y.
(2018). Atlas of optimal coil orientation and position for TMS:
A computational study. Brain Stimulation, 11, 839–848. DOI:
https://doi.org/10.1016/j.brs.2018.04.011, PMID: 29699821
Greinacher, R., Buhôt, L., Möller, L., & Learmonth, G. (2019).
The time course of ineffective sham-blinding during low-
intensity (1 mA) transcranial direct current stimulation.
European Journal of Neuroscience, 50, 3380–3388. DOI:
https://doi.org/10.1111/ejn.14497, PMID: 31228880, PMCID:
PMC6899874
Grosse-Wentrup, M., Janzing, D., Siegel, M., & Schölkopf, B.
(2016). Identification of causal relations in neuroimaging data
with latent confounders: An instrumental variable approach.
Neuroimage, 125, 825–833. DOI: https://doi.org/10.1016
/j.neuroimage.2015.10.062, PMID: 26518633
Hallam, G. P., Whitney, C., Hymers, M., Gouws, A. D., &
Jefferies, E. (2016). Charting the effects of TMS with fMRI:
Modulation of cortical recruitment within the distributed
network supporting semantic control. Neuropsychologia,
93, 40–52. DOI: https://doi.org/10.1016/j.neuropsychologia
.2016.09.012, PMID: 27650816, PMCID: PMC5155664
Hamada, M., Murase, N., Hasan, A., Balaratnam, M., & Rothwell,
J. C. (2013). The role of interneuron networks in driving
human motor cortical plasticity. Cerebral Cortex, 23,
1593–1605. DOI: https://doi.org/10.1093/cercor/bhs147,
PMID: 22661405
Hämmerer, D., Bonaiuto, J., Klein-Flügge, M., Bikson, M., &
Bestmann, S. (2016). Selective alteration of human value
decisions with medial frontal tDCS is predicted by changes
in attractor dynamics. Scientific Reports, 6, 25160. DOI:
https://doi.org/10.1038/srep25160, PMID: 27146700, PMCID:
PMC4857193
Hanslmayr, S., Matuschek, J., & Fellner, M.-C. (2014).
Entrainment of prefrontal beta oscillations induces an
endogenous echo and impairs memory formation. Current
Biology, 24, 904–909. DOI: https://doi.org/10.1016/j.cub
.2014.03.007, PMID: 24684933
Hanslmayr, S., Staudigl, T., & Fellner, M.-C. (2012). Oscillatory
power decreases and long-term memory: The information via
desynchronization hypothesis. Frontiers in Human
Neuroscience, 6, 74. DOI: https://doi.org/10.3389/fnhum
.2012.00074, PMID: 22514527, PMCID: PMC3322486
Harquel, S., Bacle, T., Beynel, L., Marendaz, C., Chauvin, A., &
David, O. (2016). Mapping dynamical properties of cortical
microcircuits using robotized TMS and EEG: Towards
functional cytoarchitectonics. Neuroimage, 135, 115–124.
DOI: https://doi.org/10.1016/j.neuroimage.2016.05.009,
PMID: 27153976
Hartwigsen, G. (2018). Flexible redistribution in cognitive
networks. Trends in Cognitive Sciences, 22, 687–698. DOI:
https://doi.org/10.1016/j.tics.2018.05.008, PMID: 29914734
Hartwigsen, G., Baumgaertner, A., Price, C. J., Koehnke, M.,
Ulmer, S., & Siebner, H. R. (2010). Phonological decisions
require both the left and right supramarginal gyri.
Proceedings of the National Academy of Sciences, U.S.A.,
107, 16494–16499. DOI: https://doi.org/10.1073/pnas
.1008121107, PMID: 20807747, PMCID: PMC2944751
Hartwigsen, G., Bergmann, T. O., Herz, D. M., Angstmann, S.,
Karabanov, A., Raffin, E., et al. (2015). Modeling the effects of
noninvasive transcranial brain stimulation at the biophysical,
network, and cognitive level. Progress in Brain Research,
222, 261–287. DOI: https://doi.org/10.1016/bs.pbr.2015
.06.014, PMID: 26541384
Hartwigsen, G., Bestmann, S., Ward, N. S., Woerbel, S., Mastroeni,
C., Granert, O., et al. (2012). Left dorsal premotor cortex
and supramarginal gyrus complement each other during
rapid action reprogramming. Journal of Neuroscience, 32,
16162–16171. DOI: https://doi.org/10.1523/JNEUROSCI.1010
-12.2012, PMID: 23152600, PMCID: PMC3558742
Hartwigsen, G., Bzdok, D., Klein, M., Wawrzyniak, M., Stockert, A.,
Wrede, K., et al. (2017). Rapid short-term reorganization in the
language network. eLife, 6, e25964. DOI: https://doi.org/10
.7554/eLife.25964, https://doi.org/10.7554/eLife.25964.017
Hartwigsen, G., Price, C. J., Baumgaertner, A., Geiss, G.,
Koehnke, M., Ulmer, S., et al. (2010). The right posterior
inferior frontal gyrus contributes to phonological word
decisions in the healthy brain: Evidence from dual-site TMS.
Neuropsychologia, 48, 3155–3163. DOI: https://doi.org/10
.1016/j.neuropsychologia.2010.06.032, PMID: 20600177,
PMCID: PMC3223523
Hartwigsen, G., & Siebner, H. R. (2015). Joint contribution of
left dorsal premotor cortex and supramarginal gyrus to rapid
action reprogramming. Brain Stimulation, 8, 945–952. DOI:
https://doi.org/10.1016/j.brs.2015.04.011, PMID: 26028563
Haug, B. A., Schönle, P. W., Knobloch, C., & Köhne, M.
(1992). Silent period measurement revives as a valuable
diagnostic tool with transcranial magnetic stimulation.
Electroencephalography and Clinical Neurophysiology, 85,
158–160. DOI: https://doi.org/10.1016/0168-5597(92)90081-L
Heidegger, T., Hansen-Goos, O., Batlaeva, O., Annak, O.,
Ziemann, U., & Lötsch, J. (2017). A data-driven approach to
responder subgroup identification after paired continuous
theta burst stimulation. Frontiers in Human Neuroscience,
11, 382. DOI: https://doi.org/10.3389/fnhum.2017.00382,
PMID: 28824394, PMCID: PMC5543102
Helfrich, R. F., Knepper, H., Nolte, G., Strüber, D., Rach, S.,
Herrmann, C. S., et al. (2014). Selective modulation of
interhemispheric functional connectivity by HD-tACS shapes
perception. PLoS Biology, 12, e1002031. DOI: https://doi.org
/10.1371/journal.pbio.1002031, PMID: 25549264, PMCID:
PMC4280108
Helfrich, R. F., Schneider, T. R., Rach, S., Trautmann-Lengsfeld,
S. A., Engel, A. K., & Herrmann, C. S. (2014). Entrainment
of brain oscillations by transcranial alternating current
stimulation. Current Biology, 24, 333–339. DOI: https://
doi.org/10.1016/j.cub.2013.12.041, PMID: 24461998
Herring, J. D., Esterer, S., Marshall, T. R., Jensen, O., &
Bergmann, T. O. (2019). Low-frequency alternating current
stimulation rhythmically suppresses gamma-band oscillations
and impairs perceptual performance. Neuroimage, 184,
440–449. DOI: https://doi.org/10.1016/j.neuroimage.2018
.09.047, PMID: 30243972
Herring, J. D., Thut, G., Jensen, O., & Bergmann, T. O. (2015).
Attention modulates TMS-locked alpha oscillations in the
visual cortex. Journal of Neuroscience, 35, 14435–14447.
DOI: https://doi.org/10.1523/JNEUROSCI.1833-15.2015,
PMID: 26511236, PMCID: PMC4623224
Herrmann, C. S., Rach, S., Neuling, T., & Strüber, D. (2013).
Transcranial alternating current stimulation: A review of the
underlying mechanisms and modulation of cognitive processes.
Frontiers in Human Neuroscience, 7, 279. DOI: https://doi
.org/10.3389/fnhum.2013.00279, PMID: 23785325, PMCID:
PMC3682121
Hoogendam, J. M., Ramakers, G. M. J., & Di Lazzaro, V. (2010).
Physiology of repetitive transcranial magnetic stimulation of
the human brain. Brain Stimulation, 3, 95–118. DOI: https://
doi.org/10.1016/j.brs.2009.10.005, PMID: 20633438
Huang, Y.-Z., Edwards, M. J., Rounis, E., Bhatia, K. P., &
Rothwell, J. C. (2005). Theta burst stimulation of the human
motor cortex. Neuron, 45, 201–206. DOI: https://doi.org
/10.1016/j.neuron.2004.12.033, PMID: 15664172
Bergmann and Hartwigsen
219
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
2
1
9
5
1
8
6
2
5
1
7
/
j
o
c
n
_
a
_
0
1
5
9
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Ilmoniemi, R. J., & Kičić, D. (2010). Methodology for combined
TMS and EEG. Brain Topography, 22, 233–248. DOI: https://
doi.org/10.1007/s10548-009-0123-4, PMID: 20012350, PMCID:
PMC2800178
Inghilleri, M., Berardelli, A., Cruccu, G., & Manfredi, M. (1993).
Silent period evoked by transcranial stimulation of the
human cortex and cervicomedullary junction. Journal of
Physiology, 466, 521–534.
Jacobs, C., Goebel, R., & Sack, A. T. (2012). Visual awareness
suppression by pre-stimulus brain stimulation; a neural
effect. Neuroimage, 59, 616–624. DOI: https://doi.org
/10.1016/j.neuroimage.2011.07.090, PMID: 21840406
Jensen, O., & Colgin, L. L. (2007). Cross-frequency coupling
between neuronal oscillations. Trends in Cognitive Sciences,
11, 267–269. DOI: https://doi.org/10.1016/j.tics.2007.05.003,
PMID: 17548233
Jensen, O., Gips, B., Bergmann, T. O., & Bonnefond, M. (2014).
Temporal coding organized by coupled alpha and gamma
oscillations prioritize visual processing. Trends in
Neurosciences, 37, 357–369. DOI: https://doi.org/10.1016
/j.tins.2014.04.001, PMID: 24836381
Jerusalem, A., Al-Rekabi, Z., Chen, H., Ercole, A., Malboubi, M.,
Tamayo-Elizalde, M., et al. (2019). Electrophysiological–
mechanical coupling in the neuronal membrane and its role
in ultrasound neuromodulation and general anaesthesia.
Acta Biomaterialia, 97, 116–140. DOI: https://doi.org/10
.1016/j.actbio.2019.07.041, PMID: 31357005
Jung, J., Bungert, A., Bowtell, R., & Jackson, S. R. (2016). Vertex
stimulation as a control site for transcranial magnetic
stimulation: A concurrent TMS/fMRI study. Brain Stimulation,
9, 58–64. DOI: https://doi.org/10.1016/j.brs.2015.09.008,
PMID: 26508284, PMCID: PMC4720218
Kaarre, O., Äikiä, M., Kallioniemi, E., Könönen, M., Kekkonen,
V., Heikkinen, N., et al. (2018). Association of the N100 TMS-
evoked potential with attentional processes: A motor cortex
TMS–EEG study. Brain and Cognition, 122, 9–16. DOI:
https://doi.org/10.1016/j.bandc.2018.01.004, PMID: 29407789
Kammer, T. (1998). Phosphenes and transient scotomas
induced by magnetic stimulation of the occipital lobe: Their
topographic relationship. Neuropsychologia, 37, 191–198.
DOI: https://doi.org/10.1016/S0028-3932(98)00093-1
Kanai, R., Chaieb, L., Antal, A., Walsh, V., & Paulus, W. (2008).
Frequency-dependent electrical stimulation of the visual
cortex. Current Biology, 18, 1839–1843. DOI: https://doi.org
/10.1016/j.cub.2008.10.027, PMID: 19026538
Kapur, N. (1996). Paradoxical functional facilitation in brain–
behaviour research: A critical review. Brain, 119, 1775–1790.
DOI: https://doi.org/10.1093/brain/119.5.1775, PMID:
8931597
Karabanov, A., Ziemann, U., Hamada, M., George, M. S.,
Quartarone, A., Classen, J., et al. (2015). Consensus paper:
Probing homeostatic plasticity of human cortex with non-
invasive transcranial brain stimulation. Brain Stimulation, 8,
442–454. DOI: https://doi.org/10.1016/j.brs.2015.01.404,
https://doi.org/10.1016/j.brs.2015.06.017, https://doi.org/10
.1016/j.brs.2015.06.016, PMID: 26050599
Kasten, F. H., Duecker, K., Maack, M. C., Meiser, A., &
Herrmann, C. S. (2019). Integrating electric field modeling
and neuroimaging to explain inter-individual variability of tACS
effects. Nature Communications, 10, 5427. DOI: https://doi
.org/10.1038/s41467-019-13417-6, PMID: 31780668, PMCID:
PMC6882891
Khatoun, A., Breukers, J., Op de Beeck, S., Nica, I. G., Aerts,
J.-M., Seynaeve, L., et al. (2018). Using high-amplitude and
focused transcranial alternating current stimulation to entrain
physiological tremor. Scientific Reports, 8, 4927. DOI: https://
doi.org/10.1038/s41598-018-23290-w, https://doi.org/10.1038
/s41598-018-26013-3, PMID: 29795166, PMCID: PMC5964328
Klaus, J., & Hartwigsen, G. (2019). Dissociating semantic and
phonological contributions of the left inferior frontal gyrus to
language production. Human Brain Mapping, 40, 3279–3287.
DOI: https://doi.org/10.1002/hbm.24597, PMID: 30969004,
PMCID: PMC6916743
Kohl, C., Spieser, L., Forster, B., Bestmann, S., & Yarrow, K.
(2019). The neurodynamic decision variable in human
multi-alternative perceptual choice. Journal of Cognitive
Neuroscience, 31, 262–277. DOI: https://doi.org/10.1162
/jocn_a_01347, PMID: 30277429
Krause, M. R., Vieira, P. G., Csorba, B. A., Pilly, P. K., & Pack, C. C.
(2019). Transcranial alternating current stimulation entrains
single-neuron activity in the primate brain. Proceedings of the
National Academy of Sciences, U.S.A., 116, 5747–5755. DOI:
https://doi.org/10.1073/pnas.1815958116, PMID: 30833389,
PMCID: PMC6431188
Kwan, A., Forbes, P. A., Mitchell, D. E., Blouin, J.-S., & Cullen,
K. E. (2019). Neural substrates, dynamics and thresholds
of galvanic vestibular stimulation in the behaving primate.
Nature Communications, 10, 1904. DOI: https://doi.org
/10.1038/s41467-019-09738-1, PMID: 31015434, PMCID:
PMC6478681
Lepage, J.-F., Saint-Amour, D., & Théoret, H. (2008). EEG and
neuronavigated single-pulse TMS in the study of the
observation/execution matching system: Are both techniques
measuring the same process? Journal of Neuroscience
Methods, 175, 17–24. DOI: https://doi.org/10.1016/j.jneumeth
.2008.07.021, PMID: 18762214
Li, B., Virtanen, J. P., Oeltermann, A., Schwarz, C., Giese, M. A.,
Ziemann, U., et al. (2017). Lifting the veil on the dynamics of
neuronal activities evoked by transcranial magnetic stimulation.
eLife, 6, e30552. DOI: https://doi.org/10.7554/eLife.30552.022,
https://doi.org/10.7554/eLife.30552, PMID: 29165241, PMCID:
PMC5722613
Lisman, J. E., & Jensen, O. (2013). The theta–gamma neural code.
Neuron, 77, 1002–1016. DOI: https://doi.org/10.1016/j.neuron
.2013.03.007, PMID: 23522038, PMCID: PMC3648857
Liu, A., Vöröslakos, M., Kronberg, G., Henin, S., Krause, M. R.,
Huang, Y., et al. (2018). Immediate neurophysiological effects
of transcranial electrical stimulation. Nature Communications,
9, 5092. DOI: https://doi.org/10.1038/s41467-018-07233-7,
PMID: 30504921, PMCID: PMC6269428
Lorenz, R., Simmons, L. E., Monti, R. P., Arthur, J. L., Limal, S.,
Laakso, I., et al. (2019). Efficiently searching through large tACS
parameter spaces using closed-loop Bayesian optimization.
Brain Stimulation, 12, 1484–1489. DOI: https://doi.org/10.1016
/j.brs.2019.07.003, PMID: 31289013, PMCID: PMC6879005
Luber, B., & Lisanby, S. H. (2014). Enhancement of human
cognitive performance using transcranial magnetic stimulation
(TMS). Neuroimage, 85, 961–970. DOI: https://doi.org/10
.1016/j.neuroimage.2013.06.007, PMID: 23770409, PMCID:
PMC4083569
Marshall, T. R., O’Shea, J., Jensen, O., & Bergmann, T. O. (2015).
Frontal eye fields control attentional modulation of alpha and
gamma oscillations in contralateral occipitoparietal cortex.
Journal of Neuroscience, 35, 1638–1647. DOI: https://doi.org
/10.1523/JNEUROSCI.3116-14.2015, PMID: 25632139, PMCID:
PMC4308606
Massimini, M., Ferrarelli, F., Huber, R., Esser, S. K., Singh, H., &
Tononi, G. (2005). Breakdown of cortical effective connectivity
during sleep. Science, 309, 2228–2232. DOI: https://doi.org/10
.1126/science.1117256, PMID: 16195466
Mastroeni, C., Bergmann, T. O., Rizzo, V., Ritter, C., Klein, C.,
Pohlmann, I., et al. (2013). Brain-derived neurotrophic
factor—A major player in stimulation-induced homeostatic
metaplasticity of human motor cortex? PLoS One, 8, e57957.
DOI: https://doi.org/10.1371/journal.pone.0057957, PMID:
23469118, PMCID: PMC3585283
220
Journal of Cognitive Neuroscience
Volume 33, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
2
1
9
5
1
8
6
2
5
1
7
/
j
o
c
n
_
a
_
0
1
5
9
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
McDonnell, M. N., Orekhov, Y., & Ziemann, U. (2006). The role of
GABAB receptors in intracortical inhibition in the human motor
cortex. Experimental Brain Research, 173, 86–93. DOI:
https://doi.org/10.1007/s00221-006-0365-2, PMID: 16489434
Mehler, D. M. A., & Kording, K. P. (2018). The lure of causal
statements: Rampant mis-inference of causality in estimated
connectivity. arXiv:1812.03363.
Mennemeier, M. S., Triggs, W. J., Chelette, K. C., Woods, A. J.,
Kimbrell, T. A., & Dornhoffer, J. L. (2009). Sham transcranial
magnetic stimulation using electrical stimulation of the scalp.
Brain Stimulation, 2, 168–173. DOI: https://doi.org/10.1016
/j.brs.2009.02.002, PMID: 20160893, PMCID: PMC2774907
Meteyard, L., & Holmes, N. P. (2018). TMS SMART—Scalp
mapping of annoyance ratings and twitches caused by
transcranial magnetic stimulation. Journal of Neuroscience
Methods, 299, 34–44. DOI: https://doi.org/10.1016/j.jneumeth
.2018.02.008, PMID: 29471064
Mills, K. R., Boniface, S. J., & Schubert, M. (1992). Magnetic
brain stimulation with a double coil: The importance of
coil orientation. Electroencephalography and Clinical
Neurophysiology, 85, 17–21. DOI: https://doi.org/10.1016
/0168-5597(92)90096-T
Miniussi, C., Harris, J. A., & Ruzzoli, M. (2013). Modelling
non-invasive brain stimulation in cognitive neuroscience.
Neuroscience & Biobehavioral Reviews, 37, 1702–1712.
DOI: https://doi.org/10.1016/j.neubiorev.2013.06.014, PMID:
23827785
Miniussi, C., Ruzzoli, M., & Walsh, V. (2010). The mechanism of
transcranial magnetic stimulation in cognition. Cortex, 46,
128–130. DOI: https://doi.org/10.1016/j.cortex.2009.03.004,
PMID: 19356747
Moliadze, V., Atalay, D., Antal, A., & Paulus, W. (2012). Close to
threshold transcranial electrical stimulation preferentially
activates inhibitory networks before switching to excitation
with higher intensities. Brain Stimulation, 5, 505–511. DOI:
https://doi.org/10.1016/j.brs.2011.11.004, PMID: 22445135
Moliadze, V., Zhao, Y., Eysel, U., & Funke, K. (2003). Effect of
transcranial magnetic stimulation on single-unit activity in
the cat primary visual cortex. Journal of Physiology, 553,
665–679. DOI: https://doi.org/10.1113/jphysiol.2003.050153,
PMID: 12963791, PMCID: PMC2343567
Mottaghy, F. M., Gangitano, M., Krause, B. J., & Pascual-Leone,
A. (2003). Chronometry of parietal and prefrontal activations
in verbal working memory revealed by transcranial magnetic
stimulation. Neuroimage, 18, 565–575. DOI: https://doi.org
/10.1016/S1053-8119(03)00010-7
Möttönen, R., van de Ven, G. M., & Watkins, K. E. (2014).
Attention fine-tunes auditory–motor processing of speech
sounds. Journal of Neuroscience, 34, 4064–4069. DOI:
https://doi.org/10.1523/JNEUROSCI.2214-13.2014, PMID:
24623783, PMCID: PMC3951700
Mueller, J. K., Grigsby, E. M., Prevosto, V., Petraglia, F. W., III,
Rao, H., Deng, Z.-D., et al. (2014). Simultaneous transcranial
magnetic stimulation and single-neuron recording in alert
non-human primates. Nature Neuroscience, 17, 1130–1136.
DOI: https://doi.org/10.1038/nn.3751, PMID: 24974797,
PMCID: PMC4115015
Müller-Dahlhaus, F., & Ziemann, U. (2015). Metaplasticity in
human cortex. Neuroscientist, 21, 185–202. DOI: https://
doi.org/10.1177/1073858414526645, PMID: 24620008
Murakami, T., Müller-Dahlhaus, F., Lu, M.-K., & Ziemann, U.
(2012). Homeostatic metaplasticity of corticospinal excitatory
and intracortical inhibitory neural circuits in human motor
cortex. Journal of Physiology, 590, 5765–5781. DOI: https://
doi.org/10.1113/jphysiol.2012.238519, PMID: 22930265,
PMCID: PMC3528990
Murakami, T., Restle, J., & Ziemann, U. (2011). Observation–
execution matching and action inhibition in human primary
motor cortex during viewing of speech-related lip movements
or listening to speech. Neuropsychologia, 49, 2045–2054.
DOI: https://doi.org/10.1016/j.neuropsychologia.2011.03.034,
PMID: 21458473
Murakami, T., Restle, J., & Ziemann, U. (2012). Effective
connectivity hierarchically links temporoparietal and frontal
areas of the auditory dorsal stream with the motor cortex lip
area during speech perception. Brain and Language, 122,
135–141. DOI: https://doi.org/10.1016/j.bandl.2011.09.005,
PMID: 22030113
Nagel, T. (1993). What is the mind–body problem? Ciba
Foundation Symposium, 174, 1–7.. DOI: https://doi.org
/10.1002/9780470514412.ch1, PMID: 8319503
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
2
1
9
5
1
8
6
2
5
1
7
/
j
o
c
n
_
a
_
0
1
5
9
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Neri, F., Mencarelli, L., Menardi, A., Giovannelli, F., Rossi, S.,
Sprugnoli, G., et al. (2020). A novel tDCS sham approach
based on model-driven controlled shunting. Brain Stimulation,
13, 507–516. DOI: https://doi.org/10.1016/j.brs.2019.11.004,
PMID: 31926812
Neuling, T., Rach, S., & Herrmann, C. S. (2013). Orchestrating
neuronal networks: Sustained after-effects of transcranial
alternating current stimulation depend upon brain states.
Frontiers in Human Neuroscience, 7, 161. DOI: https://doi
.org/10.3389/fnhum.2013.00161, PMID: 23641206, PMCID:
PMC3639376
Nguyen, J., Deng, Y., & Reinhart, R. M. G. (2018). Brain-state
determines learning improvements after transcranial
alternating-current stimulation to frontal cortex. Brain
Stimulation, 11, 723–736. DOI: https://doi.org/10.1016/j.brs
.2018.02.008, PMID: 29482970, PMCID: PMC6019559
Nitsche, M. A., Cohen, L. G., Wassermann, E. M., Priori, A., Lang, N.,
Antal, A., et al. (2008). Transcranial direct current stimulation:
State of the art 2008. Brain Stimulation, 1, 206–223. DOI:
https://doi.org/10.1016/j.brs.2008.06.004, PMID: 20633386
Nitsche, M. A., & Paulus, W. (2000). Excitability changes
induced in the human motor cortex by weak transcranial
direct current stimulation. Journal of Physiology, 527,
633–639. DOI: https://doi.org/10.1111/j.1469-7793.2000.t01
-1-00633.x, PMID: 10990547, PMCID: PMC2270099
Noury, N., Hipp, J. F., & Siegel, M. (2016). Physiological
processes non-linearly affect electrophysiological recordings
during transcranial electric stimulation. Neuroimage, 140,
99–109. DOI: https://doi.org/10.1016/j.neuroimage.2016
.03.065, PMID: 27039705
Opitz, A., Paulus, W., Will, S., Antunes, A., & Thielscher, A.
(2015). Determinants of the electric field during transcranial
direct current stimulation. Neuroimage, 109, 140–150. DOI:
https://doi.org/10.1016/j.neuroimage.2015.01.033, PMID:
25613437
Opitz, A., Windhoff, M., Heidemann, R. M., Turner, R., &
Thielscher, A. (2011). How the brain tissue shapes the
electric field induced by transcranial magnetic stimulation.
Neuroimage, 58, 849–859. DOI: https://doi.org/10.1016
/j.neuroimage.2011.06.069, PMID: 21749927
O’Shea, J., Johansen-Berg, H., Trief, D., Göbel, S., & Rushworth,
M. F. S. (2007). Functionally specific reorganization in human
premotor cortex. Neuron, 54, 479–490. DOI: https://doi.org
/10.1016/j.neuron.2007.04.021, PMID: 17481399
O’Shea, J., Revol, P., Cousijn, H., Near, J., Petitet, P., Jacquin-
Courtois, S., et al. (2017). Induced sensorimotor cortex
plasticity remediates chronic treatment-resistant visual neglect.
eLife, 6, e26602. DOI: https://doi.org/10.7554/eLife.26602,
https://doi.org/10.7554/eLife.26602.022
Parkin, B. L., Bhandari, M., Glen, J. C., & Walsh, V. (2019). The
physiological effects of transcranial electrical stimulation
do not apply to parameters commonly used in studies of
cognitive neuromodulation. Neuropsychologia, 128, 332–339.
DOI: https://doi.org/10.1016/j.neuropsychologia.2018.03.030,
PMID: 29630916
Bergmann and Hartwigsen
221
Pascual-Leone, A., Walsh, V., & Rothwell, J. (2000). Transcranial
magnetic stimulation in cognitive neuroscience—Virtual
lesion, chronometry, and functional connectivity. Current
Opinion in Neurobiology, 10, 232–237. DOI: https://doi.org
/10.1016/S0959-4388(00)00081-7
Payne, J. S., & Tainturier, M.-J. (2018). tDCS facilitation of picture
naming: Item-specific, task general, or neither? Frontiers in
Neuroscience, 12, 549. DOI: https://doi.org/10.3389/fnins
.2018.00549, PMID: 30147643, PMCID: PMC6095956
Pearl, J. (2010). An introduction to causal inference. International
Journal of Biostatistics, 6, 7. DOI: https://doi.org/10.2202/1557
-4679.1203, PMID: 20305706, PMCID: PMC2836213
Pearl, J., & Mackenzie, D. (2018). The book of why: The new
science of cause and effect. New York: Basic Books.
Peinemann, A., Reimer, B., Löer, C., Quartarone, A., Munchau, A.,
Conrad, B., et al. (2004). Long-lasting increase in
corticospinal excitability after 1800 pulses of subthreshold
5 Hz repetitive TMS to the primary motor cortex. Clinical
Neurophysiology, 115, 1519–1526. DOI: https://doi.org/
10.1016/j.clinph.2004.02.005, PMID: 15203053
Peterchev, A. V., Wagner, T. A., Miranda, P. C., Nitsche, M. A.,
Paulus, W., Lisanby, S. H., et al. (2012). Fundamentals of
transcranial electric and magnetic stimulation dose: Definition,
selection, and reporting practices. Brain Stimulation, 5,
435–453. DOI: https://doi.org/10.1016/j.brs.2011.10.001,
PMID: 22305345, PMCID: PMC3346863
Polania, R., Paulus, W., & Nitsche, M. A. (2012). Modulating
cortico-striatal and thalamo-cortical functional connectivity
with transcranial direct current stimulation. Human Brain
Mapping, 33, 2499–2508. DOI: https://doi.org/10.1002/hbm
.21380, PMID: 21922602, PMCID: PMC6870027
Price, C. J., & Friston, K. J. (2002). Degeneracy and cognitive
anatomy. Trends in Cognitive Sciences, 6, 416–421. DOI:
https://doi.org/10.1016/S1364-6613(02)01976-9
Rahman, A., Reato, D., Arlotti, M., Gasca, F., Datta, A., Parra, L. C.,
et al.. (2013). Cellular effects of acute direct current stimulation:
Somatic and synaptic terminal effects. Journal of Physiology,
591, 2563–2578. DOI: https://doi.org/10.1113/jphysiol.2012
.247171, PMID: 23478132, PMCID: PMC3678043
Ramsey, J. D., Hanson, S. J., Hanson, C., Halchenko, Y. O.,
Poldrack, R. A., & Glymour, C. (2010). Six problems for causal
inference from fMRI. Neuroimage, 49, 1545–1558. DOI:
https://doi.org/10.1016/j.neuroimage.2009.08.065, PMID:
19747552
Ratcliff, R. (1978). A theory of memory retrieval. Psychological
Review, 85, 59–108. DOI: https://doi.org/10.1037/0033-295X
.85.2.59
Ratcliff, R., & McKoon, G. (2008). The diffusion decision model:
Theory and data for two-choice decision tasks. Neural
Computation, 20, 873–922. DOI: https://doi.org/10.1162/neco
.2008.12-06-420, PMID: 18085991, PMCID: PMC2474742
Ratcliff, R., Smith, P. L., Brown, S. D., & McKoon, G. (2016).
Diffusion decision model: Current issues and history. Trends
in Cognitive Sciences, 20, 260–281. DOI: https://doi.org
/10.1016/j.tics.2016.01.007, PMID: 26952739, PMCID:
PMC4928591
Reato, D., Rahman, A., Bikson, M., & Parra, L. C. (2010). Low-
intensity electrical stimulation affects network dynamics by
modulating population rate and spike timing. Journal of
Neuroscience, 30, 15067–15079. DOI: https://doi.org/10
.1523/JNEUROSCI.2059-10.2010, PMID: 21068312, PMCID:
PMC3500391
Reato, D., Rahman, A., Bikson, M., & Parra, L. C. (2013). Effects
of weak transcranial alternating current stimulation on brain
activity—A review of known mechanisms from animal studies.
Frontiers in Human Neuroscience, 7, 687. DOI: https://doi
.org/10.3389/fnhum.2013.00687, PMID: 24167483, PMCID:
PMC3805939
Reid, A. T., Headley, D. B., Mill, R. D., Sanchez-Romero, R.,
Uddin, L. Q., Marinazzo, D., et al. (2019). Advancing functional
connectivity research from association to causation. Nature
Neuroscience, 22, 1751–1760. DOI: https://doi.org/10.1038
/s41593-019-0510-4, PMID: 31611705, PMCID: PMC7289187
Reinhart, R. M. G., & Nguyen, J. A. (2019). Working memory
revived in older adults by synchronizing rhythmic brain
circuits. Nature Neuroscience, 22, 820–827. DOI: https://doi
.org/10.1038/s41593-019-0371-x, PMID: 30962628, PMCID:
PMC6486414
Ridderinkhof, K. R. (2002a). Activation and suppression in
conflict tasks: Empirical clarification through distributional
analyses. In W. Prinz & B. Hommel (Eds.), Attention and
performance XIX: Common mechanisms in perception and
action (pp. 494–519). Oxford: Oxford University Press.
Ridderinkhof, K. R. (2002b). Micro- and macro-adjustments
of task set: Activation and suppression in conflict tasks.
Psychological Research, 66, 312–323. DOI: https://doi.org
/10.1007/s00426-002-0104-7, PMID: 12466928
Ridding, M. C., & Ziemann, U. (2010). Determinants of the
induction of cortical plasticity by non-invasive brain
stimulation in healthy subjects. Journal of Physiology,
588, 2291–2304. DOI: https://doi.org/10.1113/jphysiol
.2010.190314, PMID: 20478978, PMCID: PMC2915507
Riddle, J., Hwang, K., Cellier, D., Dhanani, S., & D’Esposito, M.
(2019). Causal evidence for the role of neuronal oscillations
in top–down and bottom–up attention. Journal of Cognitive
Neuroscience, 31, 768–779. DOI: https://doi.org/10.1162
/jocn_a_01376, PMID: 30726180, PMCID: PMC6701188
Rogasch, N. C., Sullivan, C., Thomson, R. H., Rose, N. S., Bailey,
N. W., Fitzgerald, P. B., et al. (2017). Analysing concurrent
transcranial magnetic stimulation and electroencephalographic
data: A review and introduction to the open-source TESA
software. Neuroimage, 147, 934–951. DOI: https://doi.org
/10.1016/j.neuroimage.2016.10.031, PMID: 27771347
Romei, V., Bauer, M., Brooks, J. L., Economides, M., Penny, W.,
Thut, G., et al. (2016). Causal evidence that intrinsic beta-
frequency is relevant for enhanced signal propagation in the
motor system as shown through rhythmic TMS. Neuroimage,
126, 120–130. DOI: https://doi.org/10.1016/j.neuroimage.2015
.11.020, PMID: 26584867, PMCID: PMC4739512
Romero, M. C., Davare, M., Armendariz, M., & Janssen, P. (2019).
Neural effects of transcranial magnetic stimulation at the
single-cell level. Nature Communications, 10, 2642. DOI:
https://doi.org/10.1038/s41467-019-10638-7, PMID: 31201331,
PMCID: PMC6572776
Rossi, S., Ferro, M., Cincotta, M., Ulivelli, M., Bartalini, S.,
Miniussi, C., et al. (2007). A real electro-magnetic placebo
(REMP) device for sham transcranial magnetic stimulation
(TMS). Clinical Neurophysiology, 118, 709–716. DOI:
https://doi.org/10.1016/j.clinph.2006.11.005, PMID: 17188568
Ruff, C. C., Bestmann, S., Blankenburg, F., Bjoertomt, O.,
Josephs, O., Weiskopf, N., et al. (2008). Distinct causal
influences of parietal versus frontal areas on human visual
cortex: Evidence from concurrent TMS–fMRI. Cerebral
Cortex, 18, 817–827. DOI: https://doi.org/10.1093/cercor
/bhm128, PMID: 17652468, PMCID: PMC2601025
Ruzzoli, M., Marzi, C. A., & Miniussi, C. (2010). The neural
mechanisms of the effects of transcranial magnetic stimulation
on perception. Journal of Neurophysiology, 103, 2982–2989.
DOI: https://doi.org/10.1152/jn.01096.2009, PMID: 20457853
Sack, A. T. (2006). Transcranial magnetic stimulation, causal
structure–function mapping and networks of functional
relevance. Current Opinion in Neurobiology, 16, 593–599.
DOI: https://doi.org/10.1016/j.conb.2006.06.016, PMID:
16949276
Sack, A. T., Cohen Kadosh, R., Schuhmann, T., Moerel, M.,
Walsh, V., & Goebel, R. (2009). Optimizing functional
222
Journal of Cognitive Neuroscience
Volume 33, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
2
1
9
5
1
8
6
2
5
1
7
/
j
o
c
n
_
a
_
0
1
5
9
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
accuracy of TMS in cognitive studies: A comparison of methods.
Journal of Cognitive Neuroscience, 21, 207–221. DOI: https://
doi.org/10.1162/jocn.2009.21126, PMID: 18823235
Sandrini, M., Umiltà, C., & Rusconi, E. (2011). The use of
transcranial magnetic stimulation in cognitive neuroscience:
A new synthesis of methodological issues. Neuroscience &
Biobehavioral Reviews, 35, 516–536. DOI: https://doi.org
/10.1016/j.neubiorev.2010.06.005, PMID: 20599555
Saturnino, G. B., Antunes, A., & Thielscher, A. (2015). On the
importance of electrode parameters for shaping electric field
patterns generated by tDCS. Neuroimage, 120, 25–35. DOI:
https://doi.org/10.1016/j.neuroimage.2015.06.067, PMID:
26142274
Saturnino, G. B., Thielscher, A., Madsen, K. H., Knösche, T. R.,
& Weise, K. (2019). A principled approach to conductivity
uncertainty analysis in electric field calculations. Neuroimage,
188, 821–834. DOI: https://doi.org/10.1016/j.neuroimage.2018
.12.053, PMID: 30594684
Scheines, R. (2005). The similarity of causal inference in
experimental and non-experimental studies. Philosophy of
Science, 72, 927–940. DOI: https://doi.org/10.1086/508950
Schmid, U. D., Møller, A. R., & Schmid, J. (1995). Transcranial
magnetic stimulation of the trigeminal nerve: Intraoperative
study on stimulation characteristics in man. Muscle & Nerve,
18, 487–494. DOI: https://doi.org/10.1002/mus.880180503,
PMID: 7739635
Schneidman, E., Puchalla, J. L., Segev, R., Harris, R. A., Bialek,
W., & Berry, M. J., II (2011). Synergy from silence in a
combinatorial neural code. Journal of Neuroscience, 31,
15732–15741. DOI: https://doi.org/10.1523/JNEUROSCI
.0301-09.2011, PMID: 22049416, PMCID: PMC3446770
Schroeder, C. E., & Lakatos, P. (2009). Low-frequency neuronal
oscillations as instruments of sensory selection. Trends in
Neurosciences, 32, 9–18. DOI: https://doi.org/10.1016/j.tins
.2008.09.012, PMID: 19012975, PMCID: PMC2990947
Schuhmann, T., Schiller, N. O., Goebel, R., & Sack, A. T. (2012).
Speaking of which: Dissecting the neurocognitive network of
language production in picture naming. Cerebral Cortex, 22,
701–709. DOI: https://doi.org/10.1093/cercor/bhr155, PMID:
21685399
Schutter, D. J. L. G. (2016). Cutaneous retinal activation and
neural entrainment in transcranial alternating current
stimulation: A systematic review. Neuroimage, 140, 83–88.
DOI: https://doi.org/10.1016/j.neuroimage.2015.09.067,
PMID: 26453929
Schwarzkopf, D. S., Silvanto, J., & Rees, G. (2011). Stochastic
resonance effects reveal the neural mechanisms of
transcranial magnetic stimulation. Journal of Neuroscience,
31, 3143–3147. DOI: https://doi.org/10.1523/JNEUROSCI
.4863-10.2011, PMID: 21368025, PMCID: PMC3059801
Seghier, M. L. (2013). The angular gyrus: Multiple functions and
multiple subdivisions. Neuroscientist, 19, 43–61. DOI: https://
doi.org/10.1177/1073858412440596, PMID: 22547530, PMCID:
PMC4107834
Shannon, C. E., & Weaver, W. (1949). The mathematical theory
of communication. Urbana, IL: University of Illinois Press.
Siebner, H. R. (2020). Does TMS of the precentral motor hand
knob primarily stimulate the dorsal premotor cortex or the
primary motor hand area? Brain Stimulation, 13, 517–518.
DOI: https://doi.org/10.1016/j.brs.2019.12.015, PMID:
31924573
Siebner, H. R., Auer, C., Roeck, R., & Conrad, B. (1999). Trigeminal
sensory input elicited by electric or magnetic stimulation
interferes with the central motor drive to the intrinsic hand
muscles. Clinical Neurophysiology, 110, 1090–1099. DOI:
https://doi.org/10.1016/S1388-2457(98)00053-4.
Siebner, H. R., Bergmann, T. O., Bestmann, S., Massimini, M.,
Johansen-Berg, H., Mochizuki, H., et al. (2009). Consensus
paper: Combining transcranial stimulation with neuroimaging.
Brain Stimulation, 2, 58–80. DOI: https://doi.org/10.1016
/j.brs.2008.11.002, PMID: 20633405
Siebner, H. R., Hartwigsen, G., Kassuba, T., & Rothwell, J. C.
(2009). How does transcranial magnetic stimulation modify
neuronal activity in the brain? Implications for studies of
cognition. Cortex, 45, 1035–1042. DOI: https://doi.org/10
.1016/j.cortex.2009.02.007, PMID: 19371866, PMCID:
PMC2997692.
Silvanto, J., Bona, S., Marelli, M., & Cattaneo, Z. (2018). On the
mechanisms of transcranial magnetic stimulation (TMS):
How brain state and baseline performance level determine
behavioral effects of TMS. Frontiers in Psychology, 9, 741.
DOI: https://doi.org/10.3389/fpsyg.2018.00741, PMID:
29867693, PMCID: PMC5966578
Silvanto, J., & Cattaneo, Z. (2017). Common framework for
“virtual lesion” and state-dependent TMS: The facilitatory/
suppressive range model of online TMS effects on behavior.
Brain and Cognition, 119, 32–38. DOI: https://doi.org
/10.1016/j.bandc.2017.09.007, PMID: 28963993, PMCID:
PMC5652969
Silvanto, J., Lavie, N., & Walsh, V. (2005). Double dissociation of
V1 and V5/MT activity in visual awareness. Cerebral Cortex,
15, 1736–1741. DOI: https://doi.org/10.1093/cercor/bhi050,
PMID: 15703247
Silvanto, J., Muggleton, N., & Walsh, V. (2008). State-dependency
in brain stimulation studies of perception and cognition.
Trends in Cognitive Sciences, 12, 447–454. DOI: https://doi
.org/10.1016/j.tics.2008.09.004, PMID: 18951833
Silvanto, J., Muggleton, N. G., Cowey, A., & Walsh, V. (2007).
Neural adaptation reveals state-dependent effects of
transcranial magnetic stimulation. European Journal of
Neuroscience, 25, 1874–1881. DOI: https://doi.org/10.1111
/j.1460-9568.2007.05440.x, PMID: 17408427
Smalle, E. H. M., Rogers, J., & Möttönen, R. (2015). Dissociating
contributions of the motor cortex to speech perception and
response bias by using transcranial magnetic stimulation.
Cerebral Cortex, 25, 3690–3698. DOI: https://doi.org
/10.1093/cercor/bhu218, PMID: 25274987, PMCID:
PMC4585509
Snowball, A., Tachtsidis, I., Popescu, T., Thompson, J., Delazer,
M., Zamarian, L., et al. (2013). Long-term enhancement of
brain function and cognition using cognitive training and
brain stimulation. Current Biology, 23, 987–992. DOI: https://
doi.org/10.1016/j.cub.2013.04.045, PMID: 23684971, PMCID:
PMC3675670
Sparing, R., Buelte, D., Meister, I. G., Pauš, T., & Fink, G. R.
(2008). Transcranial magnetic stimulation and the challenge
of coil placement: A comparison of conventional and
stereotaxic neuronavigational strategies. Human Brain
Mapping, 29, 82–96. DOI: https://doi.org/10.1002/hbm
.20360, PMID: 17318831, PMCID: PMC6871049
Sparing, R., Mottaghy, F. M., Ganis, G., Thompson, W. L.,
Töpper, R., Kosslyn, S. M., et al. (2002). Visual cortex
excitability increases during visual mental imagery—A TMS
study in healthy human subjects. Brain Research, 938, 92–97.
DOI: https://doi.org/10.1016/S0006-8993(02)02478-2
Spieser, L., van den Wildenberg, W., Hasbroucq, T.,
Ridderinkhof, K. R., & Burle, B. (2015). Controlling your
impulses: Electrical stimulation of the human supplementary
motor complex prevents impulsive errors. Journal of
Neuroscience, 35, 3010–3015. DOI: https://doi.org/10.1523
/JNEUROSCI.1642-14.2015, PMID: 25698738, PMCID:
PMC6605584
Stagg, C. J., & Nitsche, M. A. (2011). Physiological basis of
transcranial direct current stimulation. Neuroscientist, 17,
37–53. DOI: https://doi.org/10.1177/1073858410386614,
PMID: 21343407
Bergmann and Hartwigsen
223
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
2
1
9
5
1
8
6
2
5
1
7
/
j
o
c
n
_
a
_
0
1
5
9
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Stefan, K., Kunesch, E., Cohen, L. G., Benecke, R., & Classen, J.
(2000). Induction of plasticity in the human motor cortex by
paired associative stimulation. Brain, 123, 572–584. DOI:
https://doi.org/10.1093/brain/123.3.572, PMID: 10686179.
Stefan, K., Wycislo, M., Gentner, R., Schramm, A., Naumann, M.,
Reiners, K., et al. (2006). Temporary occlusion of associative
motor cortical plasticity by prior dynamic motor training.
Cerebral Cortex, 16, 376–385. DOI: https://doi.org/10.1093
/cercor/bhi116, PMID: 15930370
Stetkarova, I., & Kofler, M. (2013). Differential effect of
baclofen on cortical and spinal inhibitory circuits. Clinical
Neurophysiology, 124, 339–345. DOI: https://doi.org/10
.1016/j.clinph.2012.07.005, PMID: 22877625
Stokes, M. G., Chambers, C. D., Gould, I. C., Henderson, T. R.,
Janko, N. E., Allen, N. B., et al. (2005). Simple metric for
scaling motor threshold based on scalp–cortex distance:
Application to studies using transcranial magnetic stimulation.
Journal of Neurophysiology, 94, 4520–4527. DOI: https://doi
.org/10.1152/jn.00067.2005, PMID: 16135552
Taylor, P. C. J., Nobre, A. C., & Rushworth, M. F. S. (2007). FEF
TMS affects visual cortical activity. Cerebral Cortex, 17, 391–399.
DOI: https://doi.org/10.1093/cercor/bhj156, PMID: 16525126
Terao, Y., Ugawa, Y., Suzuki, M., Sakai, K., Hanajima, R., Gemba-
Shimizu, K., et al. (1997). Shortening of simple reaction time
by peripheral electrical and submotor-threshold magnetic
cortical stimulation. Experimental Brain Research, 115,
541–545. DOI: https://doi.org/10.1007/PL00005724, PMID:
9262209
ter Braack, E. M., de Vos, C. C., & van Putten, M. J. A. M. (2015).
Masking the auditory evoked potential in TMS–EEG: A
comparison of various methods. Brain Topography, 28,
520–528. DOI: https://doi.org/10.1007/s10548-013-0312-z,
PMID: 23996091
Terney, D., Chaieb, L., Moliadze, V., Antal, A., & Paulus, W.
(2008). Increasing human brain excitability by transcranial
high-frequency random noise stimulation. Journal of
Neuroscience, 28, 14147–14155. DOI: https://doi.org/10
.1523/JNEUROSCI.4248-08.2008, PMID: 19109497, PMCID:
PMC6671476
Thielscher, A., & Kammer, T. (2004). Electric field properties of
two commercial figure-8 coils in TMS: Calculation of focality
and efficiency. Clinical Neurophysiology, 115, 1697–1708.
DOI: https://doi.org/10.1016/j.clinph.2004.02.019, PMID:
15203072
Thielscher, A., Opitz, A., & Windhoff, M. (2011). Impact of the
gyral geometry on the electric field induced by transcranial
magnetic stimulation. Neuroimage, 54, 234–243. DOI: https://
doi.org/10.1016/j.neuroimage.2010.07.061, PMID: 20682353
Thies, M., Zrenner, C., Ziemann, U., & Bergmann, T. O. (2018).
Sensorimotor mu-alpha power is positively related to
corticospinal excitability. Brain Stimulation, 11, 1119–1122.
DOI: https://doi.org/10.1016/j.brs.2018.06.006, PMID:
29945791
Thut, G., Bergmann, T. O., Fröhlich, F., Soekadar, S. R., Brittain,
J.-S., Valero-Cabré, A., et al. (2017). Guiding transcranial brain
stimulation by EEG/MEG to interact with ongoing brain
activity and associated functions: A position paper. Clinical
Neurophysiology, 128, 843–857. DOI: https://doi.org/10
.1016/j.clinph.2016.10.128, https://doi.org/10.1016/j.clinph
.2017.01.003, PMID: 28233641, PMCID: PMC5385293
Thut, G., Schyns, P. G., & Gross, J. (2011). Entrainment of
perceptually relevant brain oscillations by non-invasive
rhythmic stimulation of the human brain. Frontiers in
Psychology, 2, 170. DOI: https://doi.org/10.3389/fpsyg
.2011.00170, PMID: 21811485, PMCID: PMC3142861
Thut, G., Veniero, D., Romei, V., Miniussi, C., Schyns, P., &
Gross, J. (2011). Rhythmic TMS causes local entrainment of
natural oscillatory signatures. Current Biology, 21, 1176–1185.
DOI: https://doi.org/10.1016/j.cub.2011.05.049, PMID:
21723129, PMCID: PMC3176892
Tononi, G. (2008). Consciousness as integrated information: A
provisional manifesto. Biological Bulletin, 215, 216–242.
DOI: https://doi.org/10.2307/25470707, PMID: 19098144
Töpper, R., Mottaghy, F. M., Brügmann, M., Noth, J., & Huber,
W. (1998). Facilitation of picture naming by focal transcranial
magnetic stimulation of Wernicke’s area. Experimental
Brain Research, 121, 371–378. DOI: https://doi.org/10.1007
/s002210050471, PMID: 9746143
Turi, Z., Csifcsák, G., Boayue, N. M., Aslaksen, P., Antal, A.,
Paulus, W., et al. (2019). Blinding is compromised for
transcranial direct current stimulation at 1 mA for 20 min in
young healthy adults. European Journal of Neuroscience,
50, 3261–3268. DOI: https://doi.org/10.1111/ejn.14403,
PMID: 30888090
Turi, Z., Paulus, W., & Antal, A. (2012). Functional neuroimaging
and transcranial electrical stimulation. Clinical EEG and
Neuroscience, 43, 200–208. DOI: https://doi.org/10.1177
/1550059412444978, PMID: 22956648
Valls-Solé, J., Pascual-Leone, A., Wassermann, E. M., & Hallett,
M. (1992). Human motor evoked responses to paired
transcranial magnetic stimuli. Electroencephalography and
Clinical Neurophysiology, 85, 355–364. DOI: https://doi.org
/10.1016/0168-5597(92)90048-G
van Campen, A. D., Kunert, R., van den Wildenberg, W. P. M., &
Ridderinkhof, K. R. (2018). Repetitive transcranial magnetic
stimulation over inferior frontal cortex impairs the suppression
(but not expression) of action impulses during action conflict.
Psychophysiology, 55, e13003. DOI: https://doi.org/10.1111
/psyp.13003, PMID: 28929571
van den Wildenberg, W. P. M., Wylie, S. A., Forstmann, B. U.,
Burle, B., Hasbroucq, T., & Ridderinkhof, K. R. (2010). To head
or to heed? Beyond the surface of selective action inhibition:
A review. Frontiers in Human Neuroscience, 4, 222. DOI:
https://doi.org/10.3389/fnhum.2010.00222, PMID: 21179583,
PMCID: PMC3004391
Vollmann, H., Conde, V., Sewerin, S., Taubert, M., Sehm, B.,
Witte, O. W., et al. (2013). Anodal transcranial direct current
stimulation (tDCS) over supplementary motor area (SMA)
but not pre-SMA promotes short-term visuomotor learning.
Brain Stimulation, 6, 101–107. DOI: https://doi.org/10.1016
/j.brs.2012.03.018, PMID: 22659022
Volman, I., Roelofs, K., Koch, S., Verhagen, L., & Toni, I. (2011).
Anterior prefrontal cortex inhibition impairs control over
social emotional actions. Current Biology, 21, 1766–1770.
DOI: https://doi.org/10.1016/j.cub.2011.08.050, PMID:
22000109
Vöröslakos, M., Takeuchi, Y., Brinyiczki, K., Zombori, T., Oliva, A.,
Fernández-Ruiz, A., et al. (2018). Direct effects of transcranial
electric stimulation on brain circuits in rats and humans.
Nature Communications, 9, 483. DOI: https://doi.org/10
.1038/s41467-018-02928-3, PMID: 29396478, PMCID:
PMC5797140
Voss, A., Nagler, M., & Lerche, V. (2013). Diffusion models
in experimental psychology: A practical introduction.
Experimental Psychology, 60, 385–402. DOI: https://doi
.org/10.1027/1618-3169/a000218, PMID: 23895923
Voss, A., & Voss, J. (2007). Fast-dm: A free program for efficient
diffusion model analysis. Behavior Research Methods, 39,
767–775. DOI: https://doi.org/10.3758/BF03192967, PMID:
18183889
Vossen, A., Gross, J., & Thut, G. (2015). Alpha power increase
after transcranial alternating current stimulation at alpha
frequency (alpha-tACS) reflects plastic changes rather than
entrainment. Brain Stimulation, 8, 499–508. DOI: https://
doi.org/10.1016/j.brs.2014.12.004, PMID: 25648377, PMCID:
PMC4464304
224
Journal of Cognitive Neuroscience
Volume 33, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
2
1
9
5
1
8
6
2
5
1
7
/
j
o
c
n
_
a
_
0
1
5
9
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Vosskuhl, J., Strüber, D., & Herrmann, C. S. (2018). Non-
Weise, K., Numssen, O., Thielscher, A., Hartwigsen, G., &
invasive brain stimulation: A paradigm shift in understanding
brain oscillations. Frontiers in Human Neuroscience, 12,
211. DOI: https://doi.org/10.3389/fnhum.2018.00211, PMID:
29887799, PMCID: PMC5980979
Walsh, V., & Cowey, A. (2000). Transcranial magnetic stimulation
and cognitive neuroscience. Nature Reviews Neuroscience,
1,73–80. DOI: https://doi.org/10.1038/35036239, PMID:
11252771
Walsh, V., Ellison, A., Battelli, L., & Cowey, A. (1998). Task-specific
impairments and enhancements induced by magnetic
stimulation of human visual area V5. Proceedings of the
Royal Society of London, Series B, Biological Sciences, 265,
537–543. DOI: https://doi.org/10.1098/rspb.1998.0328,
PMID: 9569672, PMCID: PMC1688918
Ward, N. S., Bestmann, S., Hartwigsen, G., Weiss, M. M.,
Christensen, L. O. D., Frackowiak, R. S. J., et al. (2010).
Low-frequency transcranial magnetic stimulation over left
dorsal premotor cortex improves the dynamic control of
visuospatially cued actions. Journal of Neuroscience, 30,
9216–9223. DOI: https://doi.org/10.1523/JNEUROSCI.4499
-09.2010, PMID: 20610756, PMCID: PMC3717513
Wassermann, E. M., Blaxton, T. A., Hoffman, E. A., Berry, C. D.,
Oletsky, H., Pascual-Leone, A., et al. (1999). Repetitive
transcranial magnetic stimulation of the dominant hemisphere
can disrupt visual naming in temporal lobe epilepsy patients.
Neuropsychologia, 37, 537–544. DOI: https://doi.org/10.1016
/S0028-3932(98)00102-X
Weichwald, S., Meyer, T., Özdenizci, O., Schölkopf, B., Ball, T.,
& Grosse-Wentrup, M. (2015). Causal interpretation rules for
encoding and decoding models in neuroimaging. Neuroimage,
110, 48–59. DOI: https://doi.org/10.1016/j.neuroimage.2015
.01.036, PMID: 25623501
Knösche, T. R. (2020). A novel approach to localize cortical
TMS effects. Neuroimage, 209, 116486. DOI: https://doi.org
/10.1016/j.neuroimage.2019.116486, PMID: 31877374
Womelsdorf, T., Valiante, T. A., Sahin, N. T., Miller, K. J., &
Tiesinga, P. (2014). Dynamic circuit motifs underlying
rhythmic gain control, gating and integration. Nature
Neuroscience, 17, 1031–1039. DOI: https://doi.org/10.1038
/nn.3764, PMID: 25065440
Yavari, F., Jamil, A., Mosayebi Samani, M., Vidor, L. P., & Nitsche,
M. A. (2018). Basic and functional effects of transcranial
electrical stimulation (tES)—An introduction. Neuroscience
& Biobehavioral Reviews, 85, 81–92. DOI: https://doi.org
/10.1016/j.neubiorev.2017.06.015, PMID: 28688701
Zaehle, T., Rach, S., & Herrmann, C. S. (2010). Transcranial
alternating current stimulation enhances individual alpha
activity in human EEG. PLoS One, 5, e13766. DOI: https://
doi.org/10.1371/journal.pone.0013766, PMID: 21072168,
PMCID: PMC2967471
Zazio, A., Bortoletto, M., Ruzzoli, M., Miniussi, C., & Veniero, D.
(2019). Perceptual and physiological consequences of dark
adaptation: A TMS–EEG study. Brain Topography, 32,
773–782. DOI: https://doi.org/10.1007/s10548-019-00715-x,
PMID: 31076949
Ziemann, U., Paulus, W., Nitsche, M. A., Pascual-Leone, A., Byblow,
W. D., Berardelli, A., et al. (2008). Consensus: Motor cortex
plasticity protocols. Brain Stimulation, 1, 164–182. DOI:
https://doi.org/10.1016/j.brs.2008.06.006, PMID: 20633383
Zimmermann, M., Verhagen, L., de Lange, F. P., & Toni, I. (2016).
The extrastriate body area computes desired goal states during
action planning. eNeuro, 3, ENEURO.0020-16.2016. DOI:
https://doi.org/10.1523/ENEURO.0020-16.2016, PMID:
27066535, PMCID: PMC4821904
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
2
1
9
5
1
8
6
2
5
1
7
/
j
o
c
n
_
a
_
0
1
5
9
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Bergmann and Hartwigsen
225