Fusiform Gyrus Face Selectivity Relates to Individual
Differences in Facial Recognition Ability
Nicholas Furl, Lúcia Garrido, Raymond J. Dolan, Jon Driver,
and Bradley Duchaine
Abstract
■ Regions of the occipital and temporal lobes, including a re-
gion in the fusiform gyrus (FG), have been proposed to con-
stitute a “core” visual representation system for faces, in part
because they show face selectivity and face repetition suppres-
sion. But recent fMRI studies of developmental prosopagnosics
(DPs) raise questions about whether these measures relate to
face processing skills. Although DPs manifest deficient face
processing, most studies to date have not shown unequivocal
reductions of functional responses in the proposed core re-
gions. We scanned 15 DPs and 15 non-DP control participants
with fMRI while employing factor analysis to derive behavioral
components related to face identification or other processes.
Repetition suppression specific to facial identities in FG or to
expression in FG and STS did not show compelling relation-
ships with face identification ability. However, we identified
robust relationships between face selectivity and face identifi-
cation ability in FG across our sample for several convergent
measures, including voxel-wise statistical parametric mapping,
peak face selectivity in individually defined “fusiform face areas”
(FFAs), and anatomical extents (cluster sizes) of those FFAs.
None of these measures showed associations with behavioral
expression or object recognition ability. As a group, DPs had
reduced face-selective responses in bilateral FFA when com-
pared with non-DPs. Individual DPs were also more likely than
non-DPs to lack expected face-selective activity in core regions.
These findings associate individual differences in face process-
ing ability with selectivity in core face processing regions.
This confirms that face selectivity can provide a valid marker
for neural mechanisms that contribute to face identification
ability. ■
INTRODUCTION
Developmental prosopagnosia (DP) is a condition in which
individuals (DPs) with otherwise normal vision, intelli-
gence, and social functioning show face recognition im-
pairments in the absence of evident brain injury. Although
numerous behavioral studies have enhanced understanding
of the cognitive bases of DP (Bentin, Degutis, DʼEsposito, &
Robertson, 2007; Humphreys, Avidan, & Behrmann, 2007;
Duchaine & Nakayama, 2006b; Behrmann, Avidan, Marotta,
& Kimchi, 2005), progress on the neural basis of DP is more
limited to date, with the few existing studies of this reviewed
below.
An influential account of the functional neuroanatomy
of face perception (Haxby, Hoffman, & Gobbini, 2000)
describes a so-called core system in the occipital and tem-
poral lobes. These core regions are considered responsible
for visual recognition of facial attributes (Haxby et al.,
2000) and are distinguished from a more “extended sys-
tem,” which further processes information resulting from
visual recognition. The proposed “core” regions have
been associated with face selectivity: they typically respond
University College London
more strongly to faces than to nonface objects (cf. Yovel &
Kanwisher, 2005). Neural responses in core regions can
also attenuate in response to repeated facial information
(i.e., repetition suppression). In normal observers, iden-
tity repetition typically suppresses regions in the fusiform
gyrus (FG) while repeating more changeable attributes,
such as expression, typically leads to BOLD suppression
in other regions such as STS (e.g., see Fox, Moon, Iaria,
& Barton, 2009; Rotshtein, Henson, Treves, Driver, &
Dolan, 2005; Andrews & Ewbank, 2004; Winston, Henson,
Fine-Goulden, & Dolan, 2004).
Although these core face-related brain regions suggest
possible loci for neural deficits in DPs, there is scant evi-
dence to date showing functional deficits in these re-
gions. DP case studies using fMRI have mainly reported
relatively normal face-selective responses in FG (e.g., see
Minnebusch, Suchan, Köster, & Daum, 2009; Thomas
et al., 2009; Behrmann, Avidan, Gao, & Black, 2007; Williams,
Berberovic, & Mattingley, 2007; Avidan, Hasson, Malach,
& Behrmann, 2005; Hasson, Avidan, Deouell, Bentin, &
Malach, 2003). Despite these numerous “null” (i.e., ap-
parently normal) findings, some atypical findings in in-
dividual cases have been identified, including some DPs
with no face selectivity (Minnebusch et al., 2009; Bentin
et al., 2007; Hadjikhani & de Gelder, 2002), weakened
© 2011 Massachusetts Institute of Technology
Journal of Cognitive Neuroscience 23:7, pp. 1723–1740
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
–
p
r
d
t
i
2
c
3
l
7
e
–
1
p
7
d
2
f
3
/
1
2
9
3
4
/
1
7
4
/
3
1
7
7
o
2
c
3
n
/
1
2
0
7
1
7
0
5
4
2
9
1
7
5
4
/
5
j
o
p
c
d
n
.
b
y
2
0
g
1
u
0
e
.
s
t
2
o
1
n
5
4
0
5
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
/
f
t
.
.
o
n
1
8
M
a
y
2
0
2
1
face selectivity (Avidan & Behrmann, 2009), or expression-
dependent FG face selectivity (van de Stock, van de Riet,
Righart, & de Gelder, 2009). One study (using an unusually
large sample of DPs) found that voice recognition deficits
in DPs were associated with reduced face selectivity in
core regions (von Kriegstein et al., 2008). Studies examining
repetition suppression have reported apparently normal
suppression to repeated (familiar) faces in DPs (Avidan &
Behrmann, 2009; Williams et al., 2007; Avidan et al., 2005).
Thus, functional imaging of DP has provided inconsis-
tent or incomplete evidence for substantially altered neural
function in the “core” regions that have been associated
with face processing in studies of people with typical face
skills. But such evidence might still be reconciled with
a role for these core regions in DP if, for example, face-
related effects were reduced or variable in DPs rather than
being completely abolished. It remains possible that the
magnitude of functional abnormalities might relate sys-
tematically to the extent of reduced face skills. Indeed
one open possibility, which has not been rigorously inves-
tigated to date, is that DPs might constitute the lower tail
of a fairly continuous distribution of face identification
ability. If so, one would not predict discrete, all-or-nothing
functional deficits that should be easily observed in every
individual DP. Instead, one would predict functional neuro-
anatomic correlates of performance might be best de-
tected by testing for continuous relationships between
fMRI measures and a wide range of behavioral face iden-
tification ability across both DPs and individuals exhibiting
normal variation in face skills.
Here, we aimed to implement such an “individual differ-
ences” approach by testing not only a sample large enough
to allow conventional group contrasts (DPs vs. non-DPs)
but also a more systematic analysis of neuroimaging data
in relation to the subject-by-subject scores on a battery of
tests. We recently applied this approach successfully in a
purely structural MRI analysis of 17 DPs and 18 non-DP
comparison participants (Garrido et al., 2009). We tested
these participants with a battery of behavioral tasks assess-
ing judgments of facial identity, facial expression, and non-
face object recognition. Using factor analysis, we showed
that we could derive summary scores for orthogonal com-
ponents of behavioral performance from this battery. Em-
pirically, this yielded separate face identity, face expression,
and nonface object behavioral components. In our MRI
morphometric analysis, these factor scores showed linear
relationships with gray matter volume in temporal cortex
across the entire sample (Garrido et al., 2009).
Using the same factor analytic approach to individual dif-
ferences in behavioral performance, we now extend our
analysis of this sample by exploring the relationship of face
identification performance deficits to the aforementioned
fMRI measures of functional face selectivity and repetition
suppression. We hypothesized that the use of these mea-
sures may reveal hitherto undiscovered relationships be-
tween face identification ability and fMRI signal in the core
face-related brain regions, such as FG.
METHODS
Participants
Our sample initially included 20 right-handed DPs who
contacted the Web site www.faceblind.org and reported
severe face recognition difficulties in everyday life. See
Garrido et al. (2009) for more information about this partici-
pant sample. We confirmed that these participants merited
classification as DPs by using the Cambridge Face Memory
Test (CFMT in its original form; Duchaine & Nakayama,
2006a) and the Famous Faces Test (FFT; Duchaine &
Nakayama, 2005). These tests have been validated on DPs
in previous studies that also provided the mean CFMT value
(mean ± SD = 57.90 ± 7.91) from 50 non-DP controls
(Duchaine & Nakayama, 2006a) and the mean FFT value
(0.89 ± 0.09) from 22 non-DP controls (Garrido, Duchaine,
& Nakayama, 2008). We observed that all of the DPs in the
present study scored more than two standard deviations
below those non-DP means (on both tests). All non-DP
participants (see below) in the present study scored better
than this criterion for the CFMT and all but two for the FFT.
All the DPs showed impaired performance compared with
the non-DP averages using a modified t test (Crawford &
Howell, 1998) for single cases. Three DPs were excluded
from the study because of neurological or visual abnormality
(epilepsy, brain injury, strabismus). The remaining 17 DPs
(11 women) were compared with a sample of 18 non-DP
participants (11 women), who were matched for age and
IQ. Scores on the CFMT and FFT tests in the present study
are shown in Figure 1 for the DPs and the non-DP group.
The ages of DP and non-DP groups did not significantly dif-
fer (mean ± SD: DPs = 30.9 ± 7.5, non-DP = 28.9 ± 5.7),
t(33) = 0.89, p = .38, nor did IQ (DPs = 123.9 ± 7.8,
non-DP = 118.9 ± 8.8), t(33) = 1.7, p = .1 (note that IQ
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
–
p
r
d
t
i
2
c
3
l
7
e
–
1
p
7
d
2
f
3
/
1
2
9
3
4
/
1
7
4
/
3
1
7
7
o
2
c
3
n
/
1
2
0
7
1
7
0
5
4
2
9
1
7
5
4
/
5
j
o
p
c
d
n
.
b
y
2
0
g
1
u
0
e
.
s
t
2
o
1
n
5
4
0
5
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
f
/
.
.
t
.
o
n
1
8
M
a
y
2
0
2
1
Figure 1. Face identification ability in non-DPs and DPs. Performance
of all study participants on the original Cambridge Face Memory Test
(CFMT) is indicated on the x-axis, and Famous Faces Test (FFT) is
indicated on the y-axis. Each point represents one participant. We defined
some participants as DPs on the basis of performance significantly
lower than published non-DP means on both of these two tests.
1724
Journal of Cognitive Neuroscience
Volume 23, Number 7
measurements were unavailable for two DPs) as measured
by Wechsler Abbreviated Scale of Intelligence (PsychCorp,
Harcourt Assessment Inc., San Antonio, TX). A neuroradiol-
ogist inspected the structural T1- and T2-weighted MRI
scans for all these participants and found no evident neuro-
logical abnormity in any. All 35 participants showed normal
or corrected-to-normal visual acuity as tested with Test
Chart 2000 (Thompson Software Solutions, Hatfield, UK).
Of these participants, 15 DPs and 15 non-DP controls elected
to return for the fMRI experiment. Informed consent was ob-
tained in accordance with procedures approved by The Joint
Ethics Committee of The National Hospital for Neurology
and Neurosurgery and The Institute of Neurology, London.
cal Neuroscience, Psychology Section, Karolinska Institute).
For each of four male identities (Figure 2A), we chose photo-
graphs acquired on two different days which depicted three
viewpoints (left or right three-quarters plus frontal) and
four emotional expressions (happy, fearful, neutral, and
angry). We converted all photographs to grayscale, cropped
the faces to occlude hair and clothing, normalized the im-
ages to have luminance distributed with equal mean and
range, and then placed them on a gray background. We also
digitized photographs of four model classic cars in left or
right three-quarters plus frontal views (Figure 2A). These
photographs were resized to approximate the size of the
face stimuli, and then the same image preparations were
implemented as described for the face stimuli.
Stimuli for fMRI Study
The stimuli were digitized photographs taken from the
KDEF database (Lundqvist, Flykt, & Öhman, 1998; The
Karolinska Directed Emotional Faces, Department of Clini-
fMRI Experiment: Design and Procedures
Throughout the fMRI experiment, participants viewed
15.2-sec blocks, followed by 4 sec of fixation (Figure 2B).
Figure 2. Stimuli and design.
(A) Examples of the car
stimuli plus facial identities
and expressions. (B) The time
course of an example block
(IdEd condition shown). (C)
Participants viewed six types
of blocks. Cs = the same
car appeared eight times;
Cd = four different cars
each appeared twice in
random order; IdEd =
four identities, and four
expressions each appeared
twice; IsEd = the same
identity appeared throughout,
but four expressions each
appeared twice; IdEs = four
identities each appeared
twice, but the expression
was the same throughout;
IsEs = all faces showed the
same identity and expression
throughout (but still across
different viewpoints, see
main text).
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
–
p
r
d
t
i
2
c
3
l
7
e
–
1
p
7
d
2
f
3
/
1
2
9
3
4
/
1
7
4
/
3
1
7
7
o
2
c
3
n
/
1
2
0
7
1
7
0
5
4
2
9
1
7
5
4
/
5
j
o
p
c
d
n
.
b
y
2
0
g
1
u
0
e
.
s
t
2
o
1
n
5
4
0
5
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
.
.
t
/
f
o
n
1
8
M
a
y
2
0
2
1
Furl et al.
1725
Each block comprised eight successive stimulus presenta-
tions, which were 1700 msec each and were preceded by
a 200-msec fixation cross. Participants fixated on a cross
at the center of the screen (and image) and pressed a
key when this cross appeared black on a randomly se-
lected one third of image presentations (otherwise white).
Throughout a block, participants viewed images of either
all faces or all cars. Each image depicted one of the three
views, selected pseudorandomly subject to the constraint
that the same view was never immediately repeated (Fig-
ure 2B). There were two types of car blocks: In cars-same
blocks (Cs), participants viewed eight images of a particular
one of the four cars. In cars-different blocks (Cd), partici-
pants saw all four cars, each presented twice in random
order. There were four types of face blocks (Figure 2C),
which conformed to the four cells of a 2 × 2 factorial design
(Winston et al., 2004). The first factor was Identity repeti-
tion: identities could be the same (Is) or different (Id).
The second factor was Expression repetition: expressions
could be same (Es) or different (Ed). For identity-different/
expression-different blocks (IdEd), each of the four iden-
tities and each of the four expressions appeared twice
in random order. For identity-same/expression-different
blocks (IsEd), all eight images depicted the same identity,
with each emotion expressed twice throughout the block.
For identity-different/expression-same blocks (IdEs), we
presented the four identities twice, always expressing the
same emotion. For identity-same/expression-same blocks
(IsEs), every image depicted one of the four identities,
always expressing one of the four emotions (Figure 2B).
Throughout the experiment, each of these six block types
was presented 24 times.
fMRI Data Acquisition
We used a Siemens Trio 3T system (Siemens, Erlangen,
Germany) to acquire T2*-weighted echo-planar functional
brain volumes. For each participant, we acquired 430 vol-
umes in each of three scanning sessions. A volume com-
prised 32 transverse slices aligned parallel to the inferior
aspect of the temporal lobes. Images were acquired at a
volume repetition time (TR) of 2176 msec with an in-plane
resolution of 3 × 3 mm, 2 mm slice thickness, and 1 mm
slice gap, with echo time = 30 msec and a flip angle of
90°. We discarded the two volumes commencing each ses-
sion to avoid magnetic equilibrium contamination. In
another scanning session (Garrido et al., 2009), T1-weighted
MDEFT and T2-weighted structural scans were acquired.
Factor Analysis of Behavioral Test Battery
We administered a test battery comprising 11 behavioral
measures to the 35 participants (17 DPs and 18 non-DPs;
see Table 1 and Supplementary Table 1). Each task required
recognition of facial identity, facial emotional expression,
or nonface objects. These data, for all measures and all
participants (i.e., across the entire sample, including both
DPs and non-DP participants, in accord with our “individ-
ual differences” approach), were submitted to a standard
Table 1. Varimax-rotated Principal Component Weights for Behavioral Tests
Test
Old/new face identity (A0)
CFMT (original version)
FFT
Sequential identity matching
Cambridge Face Perception
Old/new horses (A0)
Old/new cars (A0)
Cambridge hair memory
Facial expression films
Eyes test
Sequential expression matching
1
0.87a
0.86
0.80
0.76
0.43
0.04
0.23
0.16
0.13
0.13
0.39
Factor
2
0.11
0.38
0.34
0.16
0.63
0.85b
0.84
0.61
0.12
0.21
−0.15
3
0.15
0.17
0.25
0.13
0.00
0.16
−0.06
0.17
0.87c
0.80
0.49
CFMT = Cambridge face memory test; FFT = Famous faces test.
aBold items in this column are identity recognition tests.
bBold items in this column are object recognition tests.
cBold items in this column are expression recognition tests.
Reference
Duchaine and Nakayama (2005)
Duchaine and Nakayama (2006a, 2006b)
Duchaine and Nakayama (2005)
Garrido et al. (2009)
Duchaine, Germine, and Nakayama (2007), Duchaine,
Yovel, and Nakayama (2007)
Duchaine and Nakayama (2005)
Duchaine and Nakayama (2005)
Garrido et al. (2009)
Garrido et al. (2009)
Baron-Cohen, Wheelwright, Hill, Raste, and Plumb (2001)
Garrido et al. (2009)
1726
Journal of Cognitive Neuroscience
Volume 23, Number 7
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
–
p
r
d
t
i
2
c
3
l
7
e
–
1
p
7
d
2
f
3
/
1
2
9
3
4
/
1
7
4
/
3
1
7
7
o
2
c
3
n
/
1
2
0
7
1
7
0
5
4
2
9
1
7
5
4
/
5
j
o
p
c
d
n
.
b
y
2
0
g
1
u
0
e
.
s
t
2
o
1
n
5
4
0
5
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
/
.
.
.
f
t
o
n
1
8
M
a
y
2
0
2
1
PCA of the correlation matrix with varimax rotation of the
ensuing eigenvector components, as implemented by the
SPSS software package for Windows (Rel. 11.0.1. 2001;
SPSS Inc., Chicago, IL). We evaluated the first three fac-
tors (eigenvalues > 1.0) and computed all participantsʼ
factor scores on these three factors. These factor scores
were then used as independent variables in regressions in-
cluding the fMRI data for all participants.
fMRI Preprocessing and SPM First-level Analysis
We preprocessed and analyzed the fMRI data using SPM5
(Wellcome Trust Centre for Neuroimaging, London; http://
www.fil.ion.ucl.ac.uk/spm/) in conjunction with in-house
MATLAB programs (The Mathworks, Natick, MA). Func-
tional scans were realigned, normalized to the standard
Montreal Neurological Institute (MNI) echo-planar image
template, and then smoothed to an 8-mm FWHM Gaussian
kernel in accord with the standard SPM approach.
At the first level of individual subjects, we computed
mass univariate time-series models for each participant
using proportional scaling, AR(1) autocorrelation mod-
eling, a 1/128-Hz high-pass filter, and regressor convo-
lution with the SPM canonical hemodynamic response
model. Covariates of no interest in the design matrix
included the six head-motion parameters computed dur-
ing realignment. Six experimental regressors modeled
the onset of each image within the two car and four face
block types. We statistically controlled for any correla-
tions between low-level image similarity and block type
by adding additional parametric regressors, which mod-
eled the pixel-wise correlation of each presented image
and its predecessor (Winston et al., 2004). The first im-
age in each block (which had no predecessor) was as-
signed the average similarity for the rest of the block
for this aspect of the analysis. The mean pixel-wise corre-
lations in each condition were as follows: IdEd = 0.1633,
IsEd = 0.2511, IdEs = 0.1743, IsEs = 0.2831, Cd = 0.1639,
Cs = 0.2059.
Analysis of the group data then proceeded in two parallel
ways. For SPM group analysis, inferences about brain ac-
tivations were made on the basis of whole-brain statistical
parametric maps, using the mass univariate approach and
family-wise error (FWE) correcting for multiple corrections
at the cluster level using the SPM5 software. This standard
SPM approach was thus conducted in a voxel-wise manner.
For the ROI group analysis, ROIs were identified in indi-
vidual participants, with visual confirmation of their ana-
tomical location. Then parameter estimates from the peak
of each individually defined ROI were extracted for the vari-
ous conditions in the main experiment for further analy-
sis in MATLAB (see below). Thus, the ROI analyses differ
from the group SPM analysis in considering functionally
defined individual ROIs that can fall at somewhat different
voxels in different participants, unlike the voxel-wise SPM
group analysis. We also analyzed the cluster sizes of partic-
ular ROIs (see below).
SPM Group Analysis
For first-level, fixed-effects analysis, we computed our
contrasts of interest for each participant (e.g., face > car se-
lectivity, identity repetition suppression and expression
repetition suppression). For our SPM group analysis, whole-
brain images of these contrasts were tested statistically
using “second level” analyses, which treated participants
as a random effect. To test face selectivity and repetition
suppression within groups, contrast images were subjected
to one-tailed, one-sample t tests. To test whether non-DPs
controls differed from DPs, contrast estimates were sub-
jected to two-sample t tests. Finally, these contrasts were
tested using linear regressions with the behavioral factor
scores. These regressions were carried out by computing
the best-fitting line to the data from all participants, when
considering the data from all participants (i.e., from both
groups), in accord with our “individual differences” ap-
proach (for the same approach to structural MR data, see
also Garrido et al., 2009).
We first identified clusters of contiguously significant vox-
els at an uncorrected threshold ( p < .005, as also used for
display purposes in the figures). We then tested these clus-
ters for cluster-level FWE correction p < .05 using Gaussian
random field theory. We corrected using the whole-brain
volume or used conventional small-volume corrections
only for regions about which we had a priori hypotheses.
To use small volumes conservatively, we required cluster-
level FWE-corrected significance on the basis of both (a) a
10-mm radius sphere surrounding coordinates found in at
least one previous fMRI study reporting face-selective or
face repetition suppression effects (Fox et al., 2009; Hein
& Knight, 2008; Rotshtein et al., 2005; Andrews & Ewbank,
2004; Winston et al., 2004; Allison, Puce, & McCarthy, 2000)
and (b) single hemisphere anatomic masks of the posterior
fusiform cortex or superior or middle temporal gyrus (for
posterior and mid-STS), as derived from the Harvard-Oxford
probabilistic atlas available with FSL 4.1 (FMRIB, Oxford, UK;
http://www.fmrib.ox.ac.uk/fsl) and thresholded at 25% of
maximum intensity. Exactly the same masks were also used
for small volume correction in the structural study of Garrido
et al. (2009). For completeness, we additionally tested small
volume corrections in the vicinity of the precuneus and pos-
terior cingulate on the basis of a location recently implicated
in DP (Avidan & Behrmann, 2009), but no significant results
were obtained using small volumes for these structures. We
also allowed small-volume correction of amygdala for con-
trasts related to existing hypotheses about expression pro-
cessing. These were implemented on the basis of 10-mm
radius spheres surrounding MNI coordinates reported in
previous studies of facial expression processing (Das et al.,
2005; Vuilleumier, Armony, Driver, & Dolan, 2003).
Group Analysis of Individual ROIs
Whereas SPM group analysis provided inference at every
voxel-wise location in the brain in the form of a statistical
Furl et al.
1727
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
3
l
7
e
-
1
p
7
d
2
f
3
/
1
2
9
3
4
/
1
7
4
/
3
1
7
7
o
2
c
3
n
/
1
2
0
7
1
7
0
5
4
2
9
1
7
5
4
/
5
j
o
p
c
d
n
.
b
y
2
0
g
1
u
0
e
.
s
t
2
o
1
n
5
4
0
5
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
t
f
.
.
/
o
n
1
8
M
a
y
2
0
2
1
parametric map, ROI group analyses provided inference
about activity in localized regions as defined in individual
participants (which might thus correspond to somewhat
different voxels, albeit from similar brain structures, in
different participants). The anatomical location of each
ROI was confirmed visually with reference to canonical
and subject-specific T1-weighted scans in MNI space. Here,
we used “FG” to refer in general to neuroanatomic loca-
tions within the FG, and we reserved the term “fusiform
face area” (FFA) to refer more specifically to just the func-
tionally defined (individually face-selective) ROI located
within FG. We identified bilateral FFAs and occipital face
areas (OFAs), plus an individual ROI in the right posterior
STS, using the contrast all faces > all cars thresholded at
p < 1 × 10−4 uncorrected. Voxels in these regions fell
within the identifiable boundary of just one ROI. For partic-
ipants who lacked a face-selective area with this criterion,
we explored liberal uncorrected significance thresholds (as
liberal as p < .05) to confirm that obvious face-selective
areas were not visibly discernable; no areas were found
for these cases even with the more lax criteria.
We computed the extent of the ROI cluster sizes for non-
DP participants versus DPs. When participants lacked an
ROI, the cluster size for that ROI was designated as zero
for this analysis. We also sought a measure of individual
ROI response independent of cluster size. For this pur-
pose, we extracted beta parameters from first-level fixed-
effects models at the peak voxel in each ROI (peak for
the functionally defining contrast, e.g., all faces > all cars).
Participants who lacked an identifiable face-selective re-
gion had no peak voxel and so were excluded from the
analysis of individual peaks. The ROI peaks and the cluster
sizes were each submitted to two-sample t tests, com-
paring non-DP participants and DPs. Importantly, we also
tested for any relationships these two measures (for ROI
peaks and cluster sizes) might have with the behavioral
factor scores using linear regression across the entire sam-
ple, in accord with our “individual differences” approach.
Please note that all our ROIs were defined independently
of their relation with the behavioral scores and so did not
bias any such relationship.
RESULTS
Factor Scores for Identity-related
Recognition Ability
Our factor analysis identified three orthogonal factors that
were readily interpretable as related to an individualʼs abil-
ity to recognize facial identities, nonface objects, and facial
expressions respectively. These results were previously
published in Garrido et al. (2009) and so are only briefly
summarized in Supplementary Results and Supplementary
Table 1. We were especially interested in scores on the
first behavioral factor, which provide a summary measure
of facial identification ability that is necessarily orthogonal
to the object- and expression-related factors. We ran linear
regression analyses comparing the scores derived from
these factors with fMRI responses in the whole sample
(both non-DPs and DPs considered together, in accord
with our “individual-differences” approach, as described
later). For completeness, in the Supplementary Results,
we considered any relations of the fMRI data to the be-
havioral factors within one or other group (DP or non-
DP) considered alone, although that approach inevitably
has less power.
Face Selectivity: SPM Group Analysis
We began by searching every voxel in the whole brain for
face selectivity (i.e., we tested the contrast: all faces > all
cars) at p < .05 FWE-corrected cluster level. We report
whole-brain corrected effects or small volume corrections
where applicable. All participants were included in the
SPM group analyses, regardless of whether they also ex-
pressed individually defined ROIs.
For non-DP participants, we observed face-selective
peaks in regions of bilateral FG, bilateral posterior STS,
right mid-STS, anterior temporal cortex, and bilateral amyg-
dala, with significant effects also spreading into bilateral
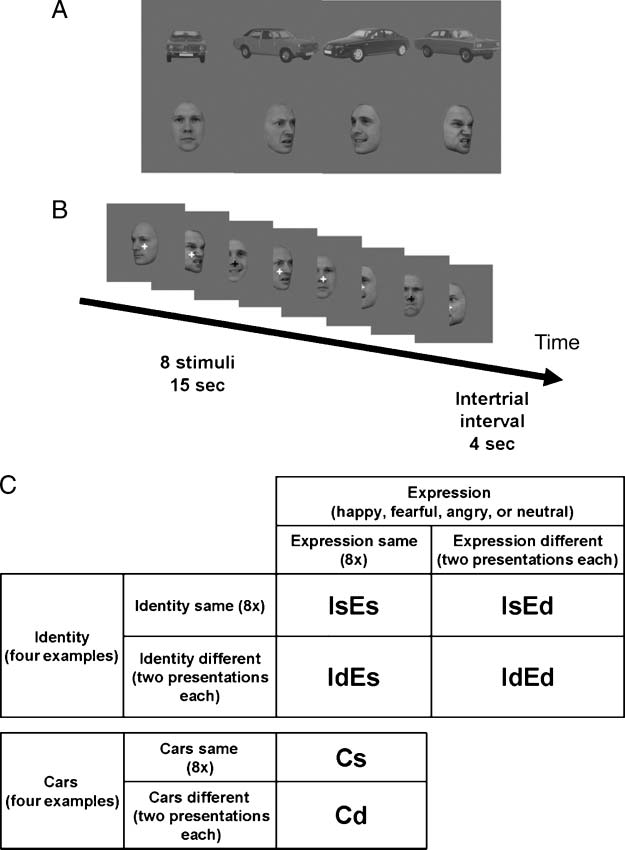
medial-temporal cortex (Figure 3 and Table 2). When we
computed the same contrast for the DP participants (Fig-
ure 3 and Table 2), substantially fewer voxels were visible:
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
3
l
7
e
-
1
p
7
d
2
f
3
/
1
2
9
3
4
/
1
7
4
/
3
1
7
7
o
2
c
3
n
/
1
2
0
7
1
7
0
5
4
2
9
1
7
5
4
/
5
j
o
p
c
d
n
.
b
y
2
0
g
1
u
0
e
.
s
t
2
o
1
n
5
4
0
5
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
f
.
t
.
/
o
n
1
8
M
a
y
2
0
2
1
Figure 3. SPM random-effects analysis of face selectivity. Regions in
stereotactic space showing significant differences between all faces
and all cars in non-DP controls (top) and DPs (bottom) include right
and left FG and right posterior and mid-STS. The right hemisphere
is shown on the right-hand side of the transverse images. All clusters
p(FWE) < .05 at the cluster level, whole brain corrected, except left
FG (small volume corrected).
1728
Journal of Cognitive Neuroscience
Volume 23, Number 7
Table 2. Linear Relationships between Face Selectivity and Behavioral Factor Scores
Peak MNI (x y z)
z Score
Cluster Size ( Voxels)
Non-DPs: All Faces > All Cars
Right STSa
Bilateral amygdalae, medial-temporal cortices and hippocampia
Right FGa
Left STSa
Left FGb
DPs: All Faces > All Cars
Right amygdalaa
Right STS, middle temporal gyrus and FGa
Positive Linear Associations with Identification Factor (1)
Left temporal polea
Right FGb
Left FGb
Positive Linear Associations with Expression-related Factor (3)
Right visual cortexa
Right amygdala/uncusb
p(FWE) < .05 cluster-level corrected.
aWhole brain corrected.
bSmall volume corrected.
54 −42 8
20 −4 −18
46 −48 −24
−56 −46 12
46 −50 −28
18 −6 −20
54 −44 14
−34 18 −28
46 −62 −18
−48 −58 −28
12 −88 20
22 0 −32
5.87
5.72
5.07
4.72
3.99
4.96
4.53
5.14
3.55
4.26
4.74
4.53
2464
6821
298
1692
213
418
1575
299
98
193
237
1575
1,993 voxels in DP clusters, but 11,488 voxels in non-DP
clusters. For DPs, one cluster peaked in right amygdala
and spread into right medial-temporal cortex, whereas an-
other peaked in right posterior STS and spread inferiorly
into the right FG. In DPs, anterior temporal cortex and
left FG showed no significantly face-selective voxels. De-
spite the visible differences in the significant statistical
parametric maps for the two groups considered separately,
when we contrasted non-DPs and DPs directly using a
one-tailed two-sample t test, no regions were significant
at p(FWE) < .05, using either whole brain or small volume
correction.
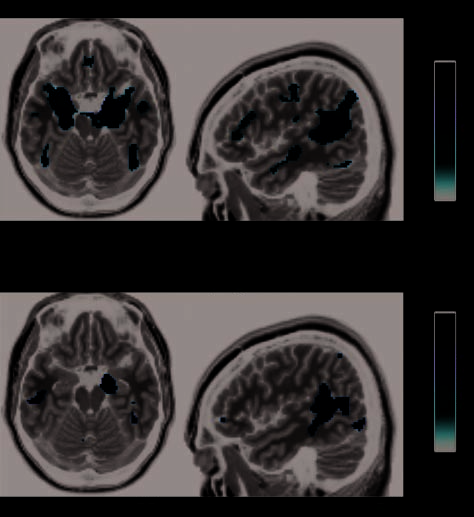
We nevertheless observed significant effects of face rec-
ognition ability when using linear regression to test for
quantitative relationships between face selectivity in fMRI
and the identity-related factor scores as measured behav-
iorally (Figure 4 and Table 2) across the entire sample in
accord with our “individual differences” approach. No-
tably, we found such effects in bilateral FG, within the
distribution of face-selective effects observed in non-DP
participants. A region near the left temporal pole was also
significant at p(FWE) < .05 cluster level using whole-brain
correction (Figure 4 and Table 2). In all three regions (right
and left FG and left temporal pole), better face identifi-
cation factor scores were associated with increased face
selectivity.
We also tested for significant linear relationships with
the second and third behavioral factors. As the second
and third factors were derived to be orthogonal to the
first, they reflect noncorrelated abilities in our participants.
We observed no significant interactions between the re-
gressor for first factor and those of the other two factors.
Moreover, the regressor for the second (object-related)
factor showed no significant linear relationship. For the
third (expression-related) factor, across the whole sample,
we observed correlations with face selectivity in a region
bordering the right posterior amygdala and uncus (small
volume corrected for the right amygdala) and a region
(whole brain corrected) of early visual cortex (Figure 4
and Table 2).
In sum, we found robust correlations between face iden-
tification ability across our entire sample and face selectivity
in FG and anterior temporal cortex; but we found no com-
parable relationship when considering the other factor
scores. The lack of a relationship with FG for the third factor
seems unlikely to reflect a simple lack of statistical power,
as linear relationships were detected outside of FG and tem-
poral pole (in visual cortex and amygdala) for that factor.
Furl et al.
1729
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
3
l
7
e
-
1
p
7
d
2
f
3
/
1
2
9
3
4
/
1
7
4
/
3
1
7
7
o
2
c
3
n
/
1
2
0
7
1
7
0
5
4
2
9
1
7
5
4
/
5
j
o
p
c
d
n
.
b
y
2
0
g
1
u
0
e
.
s
t
2
o
1
n
5
4
0
5
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
t
/
f
.
.
.
o
n
1
8
M
a
y
2
0
2
1
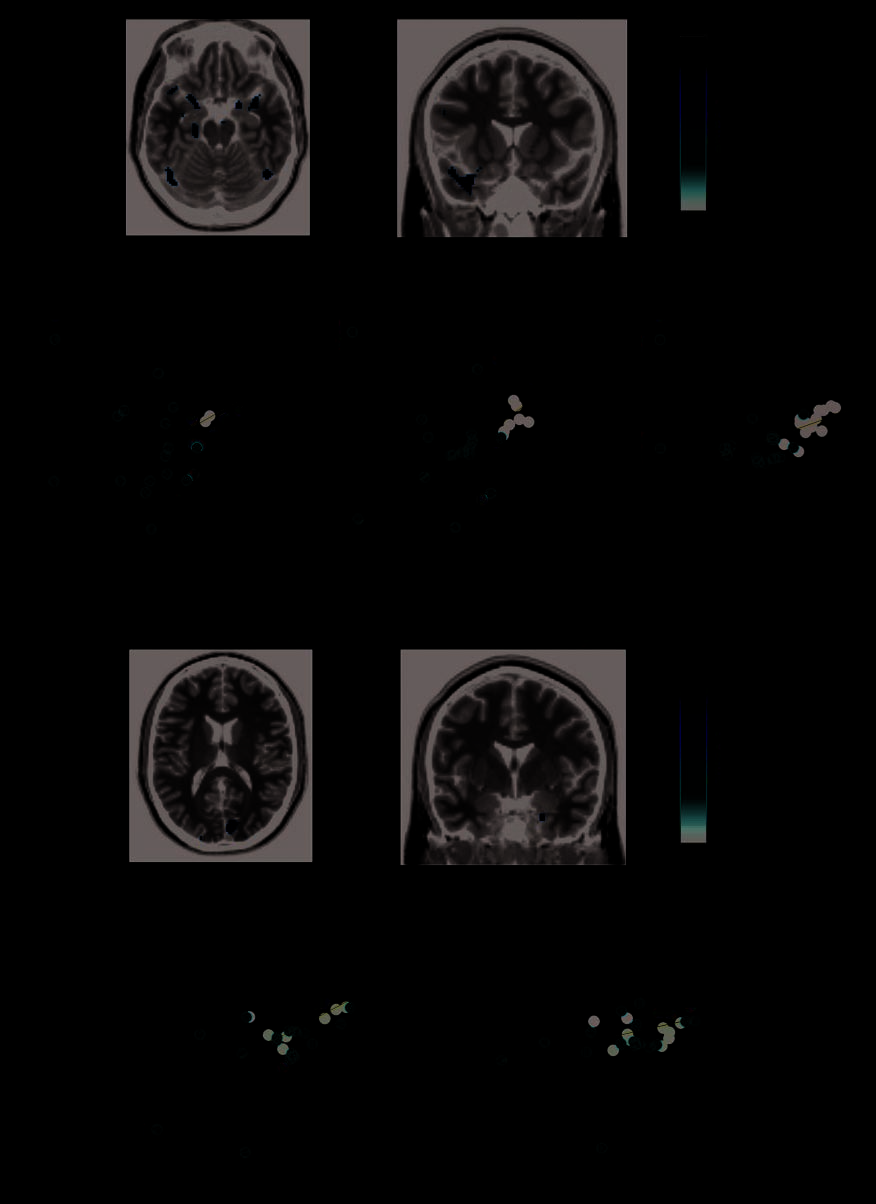
Identification of Individual ROIs
We also assessed face-selective ROIs for every individual
participant. As in previous studies (Minnebusch et al.,
2009; Bentin et al., 2007; Williams et al., 2007; Avidan
et al., 2005; Hasson et al., 2003; Hadjikhani & de Gelder,
2002), most DPs evinced right FFAs. There were three ex-
ceptions (Figure 5 and Supplementary Table 3): one DP
lacked any reliable right ventral temporal face selectivity
(hence obtained an ROI cluster size of zero for this). Two
additional DPs showed right temporal lobe face selectivity
that was diffuse and not divided into distinct regions (even
at more conservative thresholds), so we eliminated those
two DPs from further ROI group analysis. The absence of
left FFA was more common. Five DPs and two non-DPs
lacked left FFAs. Eight DPs and three non-DPs lacked any
reliable face-selective activation within the right STS. One
of the aforementioned DPs with diffuse right temporal lobe
face selectivity had no separable region in right posterior
STS and so was excluded from ROI group analysis of right
posterior STS. We note that posterior STS face selectivity
was not likely to be missed in these DPs because of our
choice of threshold. Indeed, most DPs lacking STS selec-
tivity nevertheless showed some abnormal face selectivity
in the form of a nearby cluster located near V5, which was
visibly outside the STS. As we have no unequivocal evi-
dence suggesting this region might be homologous to pos-
terior STS as observed in non-DP participants, we did not
classify it as STS. The presence or absence of clearly defined
OFA was not associated with DP. Five DPs and six non-DPs
lacked right OFA, whereas nine DPs and seven non-DPs
lacked left OFA.
We note that the operational definition of an ROI de-
pended on the choice of significance threshold. To avoid
false positives, we could have relied on conservative thresh-
olds that were multiple comparison corrected at the first
level (e.g., see Avidan & Behrmann, 2009) or required re-
sponses to faces to be numerically at least twice that of re-
sponses to nonface objects (cf. Yovel & Kanwisher, 2005).
However, these more conservative criteria inevitably in-
crease the tendency to “miss” effects that might be apparent
at less stringent uncorrected thresholds. Accordingly, for
purely descriptive purposes, we also explored more liberal
uncorrected significance thresholds such as p < .01 or p <
.05 to examine if any trend for a particular ROI to emerge
would then become apparent in an individual who did not
show an ROI at our preselected more stringent threshold.
However, at such lowered thresholds, many participants
showed first-level results contaminated by uniformly distrib-
uted background noise permeating the whole brain, from
which it proved problematic to discern any weakly signifi-
cant ROIs. We therefore optimized our operational def-
inition by selecting an uncorrected significance level that
offered unambiguously interpretable ROIs (for at least some
of the core areas) in every participant, relative to the pre-
sence of background noise and obvious false-positives. We
note also that although our ROI selection procedure neces-
sarily had led to the exclusion of some participants from the
ROI group analysis (Figure 5 and Supplementary Table 3),
no participants were excluded from the SPM group analysis.
We thus sought to avoid any potential weaknesses associ-
ated with either individual ROI or group SPM approaches
alone by demonstrating key findings that were convergent
across both approaches.
To summarize, all participants were included in the SPM
group analysis. All participants were also included in the
ROI group analysis of cluster sizes, except for the two par-
ticipants with undifferentiated face selectivity (see above).
For the analysis of individual ROI peaks, participants were
excluded from analysis only if they failed to clearly manifest
the relevant ROI. The excluded participants are shown in
black in the matrix shown in Figure 5 and listed as “no
ROI” in Supplementary Table 3.
Face Selectivity and Cluster Size: Group Analysis
of Individual ROIs
The matrix in Figure 5 shows the peak difference between
faces and cars for each ROI in each participant. Rows are
sorted within group by the identity-related behavioral fac-
tor. Rows filled in black depict the participants mentioned
above who did not show any identifiable ROI and were
therefore excluded from further analyses of ROI peaks.
From this matrix, a positive association is evident between
the identity-related factor scores and face selectivity at the
ROI peaks. These visual observations were confirmed sta-
tistically as follows.
Using one-tailed, two-sample t tests, we found significant
differences in peak face selectivity (faces > cars) between
non-DPs and DPs for the FFA in the right hemisphere,
t(25) = 2.62, p = .007, and left hemisphere, t(21) = 2.29,
p = .016 (Figure 7). In agreement with the SPM group
analysis (which had been in voxel-wise stereotactic space,
Figure 4. Behavioral factor scores in relation to brain face selectivity: SPM random-effects analysis. (A) Linear relationships with identity-related
behavioral factor scores, across the entire sample (including both DP and non-DP participants) emerge in the right and left FG (both p(FWE) <
.05, cluster level, small volume corrected) and anterior temporal cortex ( p(FWE) < .05, cluster level, whole brain corrected). The right hemisphere
is shown on the right-hand side. (B) The contrast (difference in beta parameters) for all faces > all cars (computed for each participant at the
fixed-effects level and grand mean scaled to zero) is plotted as a function of the identity-related factor scores, with each point corresponding to
one individual participant. The regression lines (fit to all participants) are shown in blue. (C) Right visual cortex ( p(FWE) < .05, cluster level,
whole brain corrected) and amygdala ( p(FWE) < .05, cluster level, small volume corrected) regions showing relationships with expression-related
behavioral factor scores. The right hemisphere is shown on the right-hand side. (D) The contrast all faces > all cars from the peak voxels in
right visual cortex and amygdala plotted as a function of expression-related factor score. The regression lines (fit to the whole sample) are
shown in blue. Note all participants were included in SPM analyses.
1730
Journal of Cognitive Neuroscience
Volume 23, Number 7
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
–
p
r
d
t
i
2
c
3
l
7
e
–
1
p
7
d
2
f
3
/
1
2
9
3
4
/
1
7
4
/
3
1
7
7
o
2
c
3
n
/
1
2
0
7
1
7
0
5
4
2
9
1
7
5
4
/
5
j
o
p
c
d
n
.
b
y
2
0
g
1
u
0
e
.
s
t
2
o
1
n
5
4
0
5
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
f
.
t
.
.
/
o
n
1
8
M
a
y
2
0
2
1
rather than for individually defined ROIs as now), we ob-
served a significantly positive linear relationship between
the peak face selectivity and the identification factor scores
for FFA (Figure 5) on the right, β = 0.41, t(25) = 2.89,
p = .004, and left, β = 0.28, t(21) = 2.36, p = .015. These
significant effects were complemented by significant cor-
relations with behavioral Factor 1 (when partialling out Fac-
tors 2 and 3) in right FFA (ρ = 0.52, p = .004) and left FFA
(ρ = 0.48, p = .015). We also analyzed whether the func-
tional size (volume) of face-selective ROIs (measured as
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
–
p
r
d
t
i
2
c
3
l
7
e
–
1
p
7
d
2
f
3
/
1
2
9
3
4
/
1
7
4
/
3
1
7
7
o
2
c
3
n
/
1
2
0
7
1
7
0
5
4
2
9
1
7
5
4
/
5
j
o
p
c
d
n
.
b
y
2
0
g
1
u
0
e
.
s
t
2
o
1
n
5
4
0
5
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
/
f
.
.
.
t
o
n
1
8
M
a
y
2
0
2
1
Furl et al.
1731
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
–
p
r
d
t
i
2
c
3
l
7
e
–
1
p
7
d
2
f
3
/
1
2
9
3
4
/
1
7
4
/
3
1
7
7
o
2
c
3
n
/
1
2
0
7
1
7
0
5
4
2
9
1
7
5
4
/
5
j
o
p
c
d
n
.
b
y
2
0
g
1
u
0
e
.
s
t
2
o
1
n
5
4
0
5
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
t
f
.
/
.
o
n
1
8
M
a
y
2
0
2
1
Figure 5. Face selectivity in individually defined ROIs. (A) Data are shown for ROIs defined by the contrast all faces > all cars. Within each
group (DPs and non-DP controls), rows are sorted by each participantʼs identity-related factor score. The first column represents group (black =
DPs; white = non-DP controls); the second is the identity-specific performance score on behavioral Factor 1 (scaled to the range 0 to 1). The
remaining columns show face selectivity (beta parameters) for contrast all faces > all cars (scaled to between 0 and 1). A row is filled in black
when an ROI could not be identified for that individual participant. (B) The contrast (difference between beta weights) all faces > all cars was
computed at the fixed effects level for right (left) and left (center) individually defined FFA and then plotted here as a function of identity-related
factor score, with each point representing one participant. The regression lines (fit to the entire sample) are shown in blue. Participants lacking
ROIs (filled in black in Figure 5A) were not included in this analysis. The rightmost graph shows the cluster sizes (in numbers of voxels) of the
right FFA, plotted against the identity-related factor score. For this analysis, all participants were included except two (see Results for details).
number of contiguous face-selective voxels) was linearly
associated with the face identification factor score. In
right FFA only (Figure 5), we found a significant regression
using the identity-related behavioral Factor 1 scores, β =
129.63, t(24) = 2.51, p = .010. Moreover, we observed sig-
nificantly larger clusters for right FFA in non-DP controls
than DPs, t(26) = 1.89, p = .003. The correlation between
cluster size in right FFA and the behavioral identification
Factor 1 scores (partialling out Factors 2 and 3) was also
significant ρ = 0.46, p = .01. None of our face selectivity
ROI measures (peak or cluster size) showed effects in right
posterior STS or the OFAs.
In all our ROI regressions, we included the scores for
Factors 2 and 3 (object- and expression-related factors) as
regressors. These regressors were not significant, although
we observed robust positive slopes and correlations for
our (orthogonally derived) identification Factor 1 in bilat-
eral FFA. Notably, this pattern of results was convergent
across all our analyses of right FG or FFA activity (SPM group
analysis and ROI group analysis of peaks and cluster sizes)
and likewise for the temporal pole. Indeed, the numeric
difference in slope between the first factor and the others
was visibly striking in right FFA for both peaks and cluster
sizes (Supplementary Figure 1). Although these differences
1732
Journal of Cognitive Neuroscience
Volume 23, Number 7
appeared large and replicated across several convergent
analyses, we did not have enough statistical power to
detect interactions between one factor versus the other
in relation to the fMRI data (Supplementary Table 4). Our
results overall seemed consistent with the conclusion that
there exists a positive linear relationship between our
fMRI face selectivity measures and behavioral Factor 1,
but we could offer no evidence favoring any such rela-
tionships for Factors 2 or 3 other than the relationship of
Factor 3 to the amygdala and visual cortex in the SPM
analysis.
However, the FG region was absent when we tested the
identity suppression contrast. Also unlike non-DPs, there
was no significant expression suppression in right mid-
STS. Despite these apparent differences and the overall
weaker effects observed in DPs, we did not find any signif-
icant effects when testing group differences or regression
models using behavioral factor scores for the repetition
suppression effects. No interaction was found between
identity and expression repetition in non-DPs or DPs.
The regions discussed above showed no repetition sup-
pression to cars, neither in non-DP nor DPs.
D
o
w
n
l
o
a
d
e
d
SPM Group Analysis of Repetition Suppression
We evaluated identity and expression repetition suppres-
sion for each voxel in the brain (SPM group analysis) and
also for individually defined ROIs. For both types of analy-
sis, we tested identity suppression by computing the one-
tailed main effect of identity repetition (identity different >
identity same). This comparison contrasts the rows in Fig-
ure 2C: (IdEd + IdEs) > (IsEd + IsEs); see also Winston
et al. (2004). Similarly, expression suppression was tested
by computing the one-tailed main effect of expression re-
petition (expression different > expressions same), which
entails contrasting the columns of Figure 2C: (IdEd +
IsEd) > (IdEs + IsEs). We also tested for the two-tailed in-
teraction of identity and expression suppression (IdEs >
IsEd) versus (IdEd > IsEs), which would reveal regions for
which one type of suppression was modulated by whether
the other category was same or different. On the basis of
previous literature (Fox et al., 2009; Rotshtein et al., 2005;
Yovel & Kanwisher, 2005; Andrews & Ewbank, 2004),
we had expected that FG may show identity suppression
whereas right (posterior and/or mid) STS would show ex-
pression suppression and that there would be no inter-
action effect.
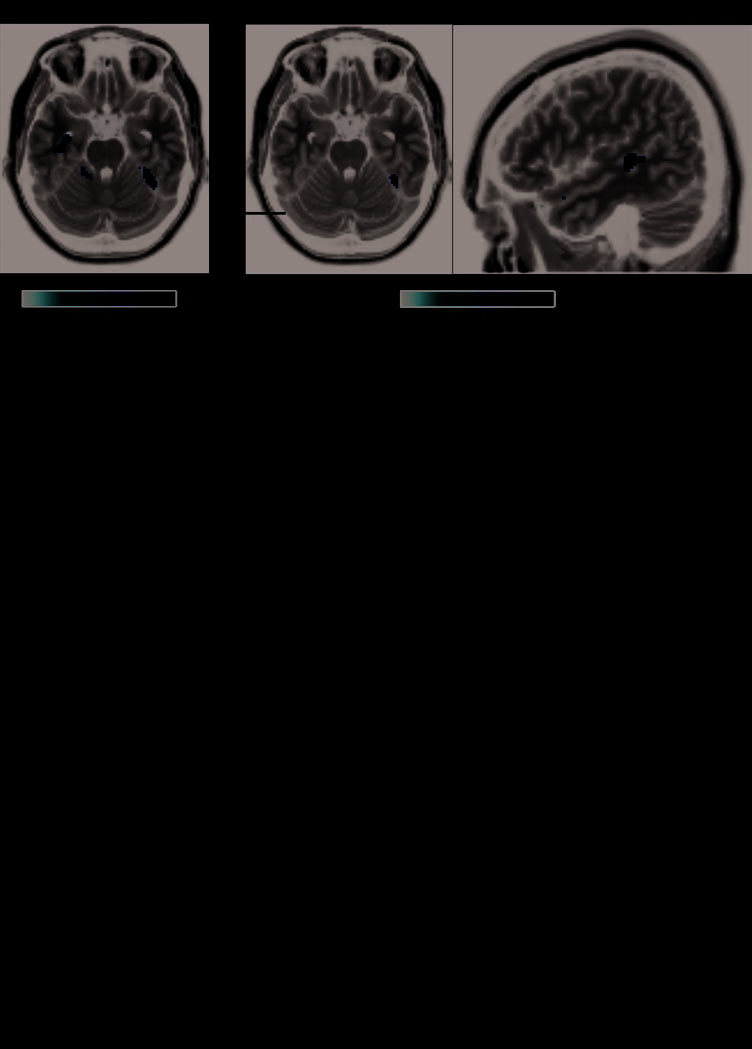
We began by using SPM group analysis to test whether
data from our non-DP participants replicates previous find-
ings (Fox et al., 2009; Winston et al., 2004). Both identity
and expression suppression contrasts yielded a right me-
dial FG cluster (Figure 6), sharing the same peak voxel
(MNI: 38 −44 −24). This region overlaps with the face-
selective activation seen in non-DPs (cf. Figure 3) but peaks
anterior and medial to the peak showing correlations be-
tween face selectivity and identity-related behavioral factor
scores (cf. Figure 4). The expression suppression contrast
also showed a region (Figure 6) in right mid-STS (MNI: 52
−30 4), anterior to the peak right posterior STS face selec-
tivity found in non-DPs (cf. Figure 4A). This mid-STS lo-
cation replicates that observed in previous studies (Fox
et al., 2009; Winston et al., 2004). Some studies have also
reported identity suppression in the posterior right STS
(Fox et al., 2009; Winston et al., 2004), but we did not ob-
serve this.
We tested whether this pattern of repetition suppres-
sion was present in DP participants. The SPM group results
showed expression suppression in the right FG region.
Repetition Suppression in Individually
Defined ROIs
We next analyzed the peak voxel in individually defined
face-selective ROIs for repetition suppression effects (Fig-
ures 7 and 8 and Supplementary Table 2), in case they might
provide a more sensitive assessment for any group differ-
ences. In agreement with the SPM group analysis, non-DP
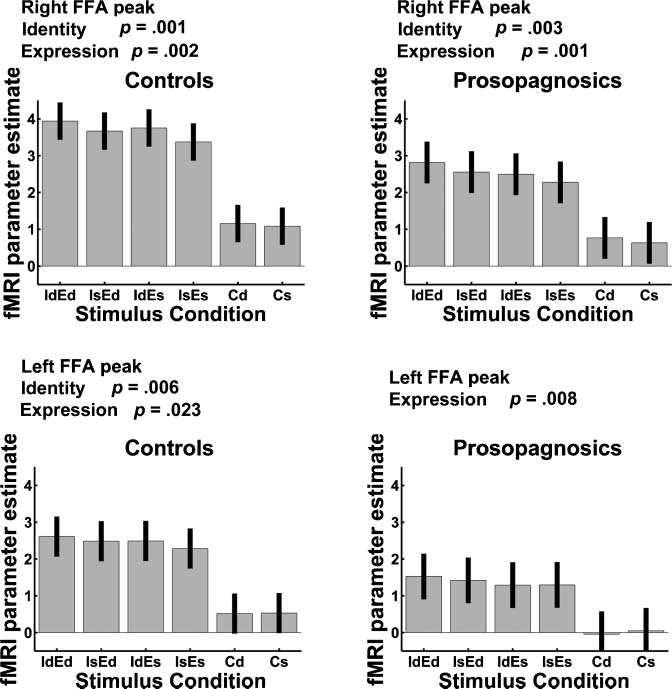
participants showed significant effects in their right FFA
ROIs (Figure 7) for both identity, t(14) = 4.37, p < .001,
and expression repetition suppression, t(14) = 3.59, p =
.002. We also found significant effects in left FFA for iden-
tity, t(12) = 3.01, p = .006, and expression repetition sup-
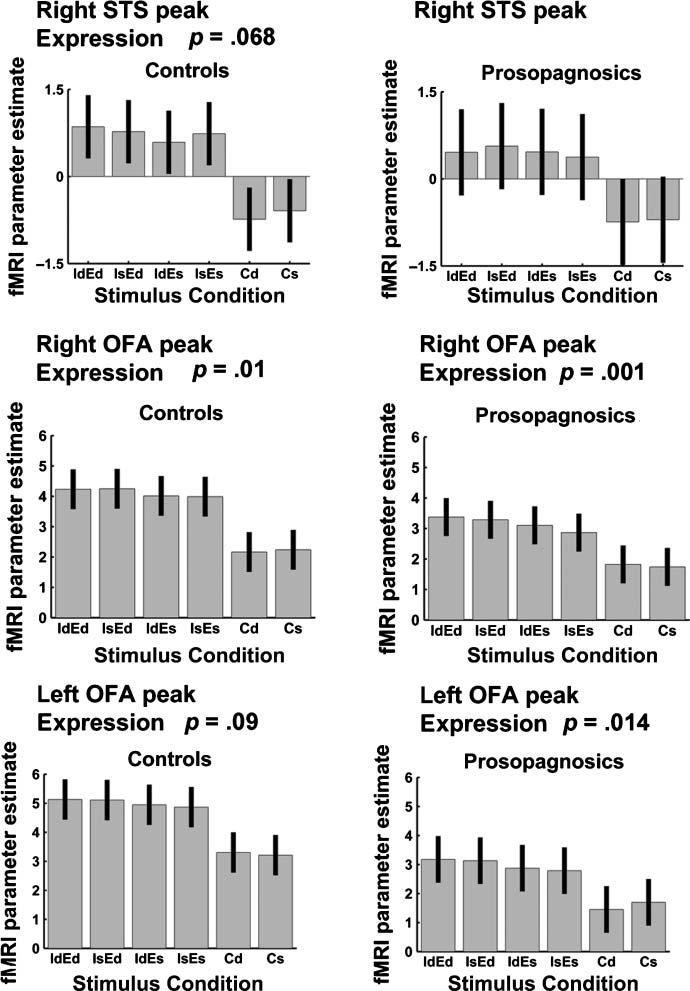
pression, t(12) = 2.19, p = .023. There was a nearly
significant trend toward expression repetition suppression
in right posterior STS, t(12) = 1.60, p = .068, but no identity
suppression. We note that the SPM group analysis had local-
ized expression repetition suppression effects to a mid-STS
region anterior to the face-selective individual peaks in
posterior STS (Figure 6). Therefore, the ROI group analysis
(now using those latter peaks) may have reduced power for
detecting these mid-STS expression effects, which appeared
(according to SPM) located anterior to the ROI peaks.
Interestingly, we also detected (unpredicted) expression
repetition suppression in the individual right OFA ROIs (Fig-
ure 8), t(8) = 2.93, p < .010, with some concordant non-
significant tendency in the same direction for the left OFA,
t(7) = 1.52, p = .09.
Finally we tested whether DPs could replicate the repeti-
tion suppression pattern observed in non-DP controls. The
pattern of results was similar to those of non-DPs in show-
ing right FFA suppression for repeated identities, t(11) =
3.47, p = .003, and expressions, t(11) = 3.91, p = .001. The
left FFA in DPs showed significant expression repetition
suppression, t(9) = 2.99, p = .008, but no effect of identity
repetition. As with the SPM group analysis, DPs showed
no effects or trends in right posterior STS. Similarly to
non-DPs, DPs showed significant expression repetition
suppression in right OFA, t(9) = 4.83, p < .001, and left
OFA, t(5) = 3.04, p < .014. The ROI group analysis of rep-
etition suppression thus did not provide a different out-
come. In line with the SPM group results, we found the
overall pattern of repetition effects to be weaker in DPs
compared with non-DPs, yet but without any overall signif-
icant group differences or behavioral correlations in ROIs,
Furl et al.
1733
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
3
l
7
e
-
1
p
7
d
2
f
3
/
1
2
9
3
4
/
1
7
4
/
3
1
7
7
o
2
c
3
n
/
1
2
0
7
1
7
0
5
4
2
9
1
7
5
4
/
5
j
o
p
c
d
n
.
b
y
2
0
g
1
u
0
e
.
s
t
2
o
1
n
5
4
0
5
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
/
.
f
.
t
.
o
n
1
8
M
a
y
2
0
2
1
Figure 6. Identity and
expression repetition
suppression: SPM random-
effects analysis. (A) Regions in
non-DP control participants
showing significant identity
repetition suppression in the
right FG (left) and expression
repetition suppression in
right FG (middle) and right
mid-STS (right). The right
side of each transverse image
represents the right hemisphere.
(B) Pattern of mean “adjusted
responses” (condition-wise
beta weights, adjusted for
other regressors in general
linear model) in non-DP
control participants from
the peak right FG and right
mid-STS voxels found in
both identity and expression
repetition suppression contrasts.
(C) For comparison, we
show the pattern of adjusted
responses in DPs using the
peak right FG and mid-STS
voxels shown in (B). All
participants were included
in SPM analyses.
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
3
l
7
e
-
1
p
7
d
2
f
3
/
1
2
9
3
4
/
1
7
4
/
3
1
7
7
o
2
c
3
n
/
1
2
0
7
1
7
0
5
4
2
9
1
7
5
4
/
5
j
o
p
c
d
n
.
b
y
2
0
g
1
u
0
e
.
s
t
2
o
1
n
5
4
0
5
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
t
f
.
.
/
.
o
n
1
8
M
a
y
2
0
2
1
for either identity or expression repetition suppression.
Also in agreement with the SPM group analysis, we de-
tected no interactions between identity and expression
suppression in any ROI, either for non-DPs or for DPs.
Car repetition did not show any effects in any ROI, either
for non-DPs or for DPs.
In summary, our measures of identity- and expression-
specific repetition suppression replicated the expected
findings in non-DP participants. FG showed suppression
to repeated expressions and identities, whereas right mid-
STS showed suppression to expressions. However, this did
not differ significantly between groups (although repetition
suppression tended to be weaker in DPs) nor did it relate
significantly to the behavioral factor scores.
This contrasts with the robust statistical relationships
we had observed between face selectivity per se (faces >
cars) in FG (or individually defined FFA) with the be-
havioral Factor 1 of identity-related recognition ability.
That effect was observed for multiple measures of face
selectivity, including face selectivity at individual voxels
in SPM group analysis, at the peak face selectivity of ROIs
functionally defined in individuals, and the anatomic extent
1734
Journal of Cognitive Neuroscience
Volume 23, Number 7
of the face selectivity in these ROIs. Our results thus pro-
vide new evidence that the neural mechanisms expressing
face selectivity do relate to individual differences in face
identification performance, although repetition suppres-
sion effects apparently may not.
DISCUSSION
We explored the relationships between behavioral face
recognition ability and two fMRI measures of neural pro-
cessing for faces (face selectivity and repetition suppres-
sion), exploiting the wide range of face identification
performance among 15 DPs and 15 matched non-DP par-
ticipants. As a group, DPs showed reduced face-selective
responses in the bilateral FFA and smaller face-selective
clusters in right FFA. We employed a factor analytic ap-
proach to our behavioral battery, as recently reported in
our related structural MR study (Garrido et al., 2009). For
this approach, we derived behavioral scores from standard
factor analysis of all participantsʼ performance on the test
battery. This yielded three orthogonal components of be-
havioral variability related to recognition for facial identity,
nonface objects, and facial expressions.
Our analyses demonstrate that the identity-related be-
havioral factor was linearly related (when considering every
individual in our sample, in accord with our “individual
differences” approach) to fMRI face selectivity in the left
anterior temporal lobe and right FG. This effect was ob-
served for FG face selectivity as identified in SPM group
analysis and for the peaks and anatomic extent of individu-
ally defined FFA ROIs. These results demonstrate that bet-
ter performance in facial identity processing tasks was
positively associated with fMRI face selectivity in FG bilat-
erally and left anterior temporal lobe, with peak face selec-
tivity in the right and left individually defined FFA, and with
the size of the right FFA. The significant brain–behavior
relations we observed applied only to the behavioral com-
ponent relating to face identification performance (but see
Supplementary Table 4), even when controlling for the
other two factors (related to performance for facial ex-
pressions or nonface objects). The second and the third
factors were not related to any of our three face selectivity
fMRI measures: SPM voxels, ROI peaks, or ROI cluster size.
The absence of face-selective ROIs, including for right
posterior STS, was also associated with DP-related deficits.
Thus, we provide abundant new evidence linking face
identification ability to individual differences in function
for core face-selective areas (FG, posterior STS) in relation
to fMRI face selectivity (assessed here by the face > car
contrast). Although we found several forms of repetition
suppression in these and related regions, the repetition
Figure 7. Identity and
expression repetition
suppression in individually
defined FFA. Average peak
parameter estimate (beta
weight) with 95% confidence
intervals in right and left
for the six conditions. Also
shown are p values less than
.05 for identity repetition
suppression (denoted
“identity”) and expression
repetition suppression
(denoted “expression”).
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
–
p
r
d
t
i
2
c
3
l
7
e
–
1
p
7
d
2
f
3
/
1
2
9
3
4
/
1
7
4
/
3
1
7
7
o
2
c
3
n
/
1
2
0
7
1
7
0
5
4
2
9
1
7
5
4
/
5
j
o
p
c
d
n
.
b
y
2
0
g
1
u
0
e
.
s
t
2
o
1
n
5
4
0
5
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
/
t
.
.
.
f
o
n
1
8
M
a
y
2
0
2
1
Furl et al.
1735
Figure 8. ROI identity
and expression repetition
suppression in posterior
STS and OFA. Average peak
parameter estimate (beta
weight) with 95% confidence
intervals in right and left
OFA plus right posterior STS
for the six conditions. Also
shown are significant and
near-significant p values
for identity repetition
suppression (denoted
“identity”) and expression
repetition suppression
(denoted “expression”).
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
–
p
r
d
t
i
2
c
3
l
7
e
–
1
p
7
d
2
f
3
/
1
2
9
3
4
/
1
7
4
/
3
1
7
7
o
2
c
3
n
/
1
2
0
7
1
7
0
5
4
2
9
1
7
5
4
/
5
j
o
p
c
d
n
.
b
y
2
0
g
1
u
0
e
.
s
t
2
o
1
n
5
4
0
5
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
/
.
f
t
.
phenomena were not strongly linked to individual differ-
ences in behavior, unlike the face selectivity per se.
Face Selectivity and Successful Recognition
of Identities
The FG and the FFA have received considerable research
attention. Its robust face selectivity across many normal
observers suggests that it may provide an important mech-
anism for face representation. More controversially, it has
been claimed that FG and/or FFA may provide specialized
mechanisms for individuating identities as opposed to rec-
ognition of other physical attributes of faces (Rotshtein
et al., 2005; Haxby et al., 2000). Such claims have led to
the prediction that behavioral success in differentiating
identities (and individual differences in this ability) may
be correlated with FG face selectivity. Our new data in re-
lation to individual differences in face identification skill
(across non-DP and DP participants considered as one
sample of variation in such skill) provide new evidence
consistent with this view.
Our approach allowed us to detect quantitative rela-
tionships between fMRI measures and the magnitude of
behavioral skill. A quantitative (rather than all-or-none) re-
lationship between the extent of face identification ability
and face selectivity may, in part, help explain the hetero-
geneous results seen in the previously published DP case
studies, typically examining a smaller number of participants
o
n
1
8
M
a
y
2
0
2
1
1736
Journal of Cognitive Neuroscience
Volume 23, Number 7
(Minnebusch et al., 2009; Bentin et al., 2007; Williams et al.,
2007; Avidan et al., 2005; Hasson et al., 2003; Hadjikhani
& de Gelder, 2002). In our study, nearly all DPs (12 of
15 DPs) evinced normal-appearing right FFAs when we
inspected cases individually. We observed more quantita-
tive relationships here, which may be consistent with DP
individuals forming the tail end of a potentially more con-
tinuous distribution of facial recognition ability. Such con-
tinuous variability might also potentially explain why the
SPM group analysis did not reveal significant effects in FG
for categorical comparisons between group means. Sig-
nificant FG findings were instead most apparent for regres-
sions aimed at detecting continuous linear relationships
rather than discrete differences. Although our ROI group
analysis of peak selectivity and cluster size was sensitive
to some discrete differences, our data suggest that many
DPs may be part of the wider spectrum of face identifica-
tion ability, although this “continuum” versus “dichotomy”
issue requires further research with even larger samples.
Beyond FG/FFA, our data implicate additional regions
in face identification ability. Right posterior STS face se-
lectivity was absent or abnormal in many DPs (Figure 5),
although we did not find statistically robust regression ef-
fects for the face identification factor in this region (but
note that there were fewer participants in the analysis
for STS ROIs). Intriguingly, right STS was also implicated
in our previous study of the structural correlates of DP-
related face impairments, using the same participant popu-
lation (Garrido et al., 2009). In that study, a mid-STS region
(somewhat anterior to the more posterior STS face selec-
tivity we observed here) showed decreased gray matter
volume in our DP population compared with the same
matched non-DP control sample.
It is notable also that the SPM group analysis revealed
a sizable correlation of fMRI face selectivity with behavioral
face identification near the left temporal pole (Table 2).
Previous fMRI work suggests that (in non-DP participants)
anterior temporal cortex may subserve identity-specific
face perception (Rotshtein et al., 2005) and may represent
person-specific conceptual knowledge (Simmons & Martin,
2009). At least in the right hemisphere, anterior temporal
cortex further can signal information about individual iden-
tities (Kriegeskorte, Formisano, Sorger, & Goebel, 2007).
Moreover, for DPs, there are recent reports of some struc-
tural abnormalities in temporal pole (Behrmann et al.,
2007). Our previous structural study showed a similar tem-
poral pole effect in the same sample of DPs we report
here, although in the right hemisphere (Garrido et al.,
2009). This further accords with recent findings that the
white matter integrity of connections between regions in
the occipital and posterior temporal lobes and more anterior
temporal regions are reduced in some DPs, as suggested
recently using diffusion tensor tractography (Thomas et al.,
2009). Interestingly, a DP case study (von Kriegstein,
Kleinschmidt, & Giraud, 2006) seems to accord with such
an interregional perspective by showing not only reduced
face responses in the anterior temporal lobe but also re-
duced functional connectivity between right FG and left
anterior lobe compared with a non-DP control population.
Although these anterior temporal regions may be struc-
turally deficient in DPs, Behrmann et al. (2007) did not
find structural or functional deficits that overlapped with
core functional regions (such as posterior or mid-FG). In
contrast, our previous structural study with the same par-
ticipants as reported here (Garrido et al., 2009) revealed
reduced gray matter in DPs for a mid-FG region, which
was relatively near to our face-selective FG region here.
To provide an initial exploration into possible relations
between structural MR outcomes and functional MR out-
comes within the same sample, we used our previously
published structural measures (see Garrido et al., 2009)
to explore the idea that reduced functional face selectivity
(in fMRI) might relate to deficient underlying gray matter
in structural MR (Supplementary Results and Supplemen-
tary Table 4). However, this preliminary analysis found
no significant correlations between functional and struc-
tural measures for any of the ROIs showing face selectivity
reductions with worse face identification here. Future re-
search with larger samples will be needed to unravel the
structural-functional relation in the context of face iden-
tification skills. Moreover, this may require sophisticated
multivariate analysis approaches because it is possible that
structural variation in one (or several) particular brain re-
gion could lead to functional variation in remote but inter-
connected regions. Hence, those areas showing structural
variation may not have a one-to-one correspondence with
those showing functional variation.
Measuring Orthogonal Components of
Recognition Ability
We included several tasks (including the CFMT and the
FFT) that putatively measure face identification ability. Per-
formance on any one of these tasks, taken by itself, is pre-
sumably attributable in part to identity-related processing.
Yet any raw putative identity-related measure (taken by
itself ) may be potentially confounded by variability attrib-
utable to other visual processes also or to the participantʼs
attention during that task. The factor analysis method can
provide some greater assurance that the variability as-
sociated with the emergent identity-related performance
factor is related only to that portion of variability held in
common by a diverse range of face identification-related
tasks. Moreover, the factor analysis also ensures this vari-
ability will be orthogonal to that associated with other fac-
tors, as for the object and expression components here.
Despite the effectiveness of this factor analytic approach,
we believe that it could be further improved. A greater num-
ber and diversity of behavioral measures and participants
will presumably produce increasingly “pure,” interpretable,
and useful resultant factors. For example, our face identi-
fication factor here related to several memory tests (e.g.,
CFMT) and so might be rendered more identity specific by
including more perception-based tests. It is reassuring in
Furl et al.
1737
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
–
p
r
d
t
i
2
c
3
l
7
e
–
1
p
7
d
2
f
3
/
1
2
9
3
4
/
1
7
4
/
3
1
7
7
o
2
c
3
n
/
1
2
0
7
1
7
0
5
4
2
9
1
7
5
4
/
5
j
o
p
c
d
n
.
b
y
2
0
g
1
u
0
e
.
s
t
2
o
1
n
5
4
0
5
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
t
.
f
.
.
/
o
n
1
8
M
a
y
2
0
2
1
this respect that the identity-matching test loaded highly
on our Factor 1, despite minimal demands on long-term
memory. However, it is interesting that another perceptual
test, CFPT, did not load as highly. Indeed, the factor analy-
sis partitioned variability in CFPT performance not only into
that held in common with other facial identification tasks
but also (separately) into some CFPT variability held more
in common with object tasks. This finding underscores the
potential advantage of factor analysis over reporting rela-
tionships with raw measures alone, as the latter approach
could potentially mix variability related to different abilities
that jointly contribute to any single task.
We acknowledge also that our power for detecting rela-
tionships with object- and expression-related factor scores
here may have been more limited than for face identifica-
tion factor scores because of restricted ranges (compared
with the identity-related factor). We were specifically in-
terested here in detecting correlations related to identity
recognition, so we recruited individuals (DPs) deficient in
identity processing in addition to the non-DP participants.
The resulting sample may not include individuals varying
to the same degree on object or expression recognition.
This could be addressed in future work by specifically
sampling also (or instead) individuals with marked object
or expression deficits. Despite the potential range restric-
tion, we nevertheless already sampled sufficient variability
to detect some brain–behavior relationships not related to
facial identity. In particular, we note our finding that amyg-
dala face selectivity related to the expression factor score,
a finding consistent with numerous studies showing amyg-
dala involvement in facial expression and emotion repre-
sentation (e.g., Das et al., 2005; Vuilleumier et al., 2003).
Repetition Suppression
In addition to investigating face selectivity, we were also
interested in repetition suppression as a further measure
potentially relevant to facial identity representations. Our
results in non-DP participants are similar to previous re-
sults from other studies and appear largely consistent with
the model of Haxby et al. (2000). As expected, we found
that identity repetition leads to reduced responses in right
mid-FG; expression repetition leads to reduced fMRI re-
sponses in right STS, whereas no interaction effect was
found between the two types of repetition (see also Winston
et al., 2004). We further detected expression repetition
effects in right mid-FG (see also Fox et al., 2009). Our find-
ings of both identity and expression repetition suppres-
sion in FG may bear on whether anatomically separable or
functionally independent pathways subserve identity ver-
sus expression processing, potentially favoring the alterna-
tive proposal that form-based representations of identities
and expressions may share some common visual represen-
tations (Calder & Young, 2005).
These suppression effects were also expressed in DPs,
albeit more weakly. Thus, our results further replicated
other previous findings that did not show clear repetition
suppression anomalies in DPs (Avidan & Behrmann, 2009;
Avidan et al., 2005). The absence of a group difference for
repetition suppression and, in particular, of any correlation
for such suppression with the face identification behavioral
factor contrasts with the positive results obtained using face
selectivity fMRI measures. This difference between two
measures of face-related neural processing deserves further
exploration, as it raises interesting questions about the ex-
tent to which face selectivity and repetition suppression
may reflect the same or different processes. Although many
studies a priori assumed repetition suppression to be a
conclusive marker for neural coding of the repeated infor-
mation, in fact the mechanisms underlying repetition sup-
pression are not well understood (Grill-Spector, Henson,
& Martin, 2006), complicating direct interpretation of their
results. A recent study (Mur, Ruff, Bodurka, Bandettini, &
Kriegeskorte, 2010) suggests that identity suppression may
not always reflect neural coding of identity but could poten-
tially result from a variety of other causes including declin-
ing attention. Moreover, abnormal repetition suppression
can arise despite intact face selectivity, in some cases of ac-
quired prosopagnosia with lesions in the vicinity of OFA
(Steeves et al., 2009; Schiltz et al., 2006).
One testable possibility is that repetition suppression
reflects representations that support different behaviors
than face selectivity. For example, repetition suppression
may better relate to repetition-related behavioral effects
such as priming (which were not examined behaviorally
here), whereas face selectivity might support more explicit
recognition judgments, like those made here. Further stud-
ies could go on to test whether individual differences in
repetition suppression are associated with more implicit
measures of face processing. Given that repetition sup-
pression effects in fMRI experiments can be much smaller
than face selectivity effects, magneto-encephalography
might provide more comparably sized measures of face
selectivity and repetition suppression (for large repetition
suppression effects in magneto-encephalography, see Furl,
van Rijsbergen, Treves, Friston, & Dolan, 2007; Harris &
Nakayama, 2006). Repetition suppression measures could
also be compared against other representational markers,
such as multivariate pattern decoding, in the future (e.g.,
Kriegeskorte et al., 2007).
Finally, we found that identity repetition suppressed FG
activity, despite variations in expression, viewpoint, and
other low-level image features across the repeated identi-
ties here. Although this result contrasts with some previous
studies that did not report clear identity-specific repeti-
tion effects in FG (Davies-Thompson, Gouws, & Andrews,
2009; Pourtois, Schwartz, Seghier, Lazeyras, & Vuilleumier,
2005a, 2005b; Andrews & Ewbank, 2004), there may be
some limited viewpoint dependence (Ewbank & Andrews,
2008). Pourtois et al. (2005b) found limited generalizability
of identity-specific suppression between three-quarter view
and frontal faces (as we used) in FG, whereas Ewbank and
Andrews (2008) showed that generalization declined with
increasing change in viewpoint angle.
1738
Journal of Cognitive Neuroscience
Volume 23, Number 7
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
–
p
r
d
t
i
2
c
3
l
7
e
–
1
p
7
d
2
f
3
/
1
2
9
3
4
/
1
7
4
/
3
1
7
7
o
2
c
3
n
/
1
2
0
7
1
7
0
5
4
2
9
1
7
5
4
/
5
j
o
p
c
d
n
.
b
y
2
0
g
1
u
0
e
.
s
t
2
o
1
n
5
4
0
5
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
.
t
f
/
.
o
n
1
8
M
a
y
2
0
2
1
Conclusions
Face selectivity and repetition suppression in core face
processing regions have become increasingly accepted
as neural measures for face processing. Previous studies
of DP had typically not found definitive evidence for re-
duced face selectivity or repetition suppression in these
core areas. To address this issue, we scanned 15 DPs and
15 matched non-DP controls in conjunction with a factor
analytic method for deriving identity-related summary
behavioral scores from a battery of tests (as opposed to
expression or nonface object relates scores). Although
we found no compelling relationship between these
behavioral scores and repetition suppression, we found
clear evidence that fusiform face selectivity in fMRI re-
lates systematically to individual differences in behavioral
face identification ability. This was found across voxel-wise
SPM group analysis, for the peaks of individually defined
FFAs, and for the anatomic extent of the individual FFAs.
No such relationship was found between FG/FFA activity
or size with object or expression recognition ability. These
results indicate that fusiform face selectivity provides a
marker for neural mechanisms that determine variation
in face identification ability.
Acknowledgments
The authors are grateful to their colleagues at the University
College London, namely, Laura Germine and Raka Tavashmi for
assistance with behavioral measurement and/or fMRI data collec-
tion, Bogdan Draganski and Ferath Kherif for advice and support,
and John Stevens for clinical evaluation of the participants. This
work was supported by the Portuguese Foundation for Science
and Technology (grant no. SFRH/BD/22580/2005 to L. G.), the
Economic and Social Research Council (grant no. RES-061-23-
0400 to B. D.), and The Wellcome Trust (R. J. D. and J. D.).
J. D. is a Royal Society Research Professor. Editorial assistance
for the manuscript was provided by the National Institutes of
Health Fellows Editorial Board.
Reprint requests should be sent to Nicholas Furl, National Insti-
tutes of Health, Building 49, 1B80, 49 Convent Dr., Bethesda, MD,
20892, or via e-mail: furlno@mail.nih.gov.
REFERENCES
Allison, T., Puce, A., & McCarthy, G. (2000). Social perception
from visual cues: Role of the STS region. Trends in
Cognitive Sciences, 4, 267–278.
Andrews, T. J., & Ewbank, M. P. (2004). Distinct representations
for facial identity and changeable aspects of faces in the
human temporal lobe. Neuroimage, 23, 905–913.
Avidan, G., & Behrmann, M. (2009). Functional MRI reveals
compromised neural integrity of the face processing network in
congenital prosopagnosia. Current Biology, 19, 1146–1150.
Avidan, G., Hasson, U., Malach, R., & Behrmann, M. (2005).
Detailed exploration of face-related processing in congenital
prosopagnosia: II. Functional neuroimaging findings.
Journal of Cognitive Neuroscience, 17, 1150–1167.
Baron-Cohen, S., Wheelwright, S., Hill, J., Raste, Y., & Plumb, I.
(2001). The “Reading the Mind in the Eyes” test revised
version: A study with normal adults, and adults with Asperger
syndrome or high-functioning autism. Journal of Child
Psychology and Psychiatry, 42, 241–251.
Behrmann, M., Avidan, G., Gao, F., & Black, S. (2007). Structural
imaging reveals anatomical alterations in inferotemporal
cortex in congenital prosopagnosia. Cerebral Cortex, 17,
2354–2363.
Behrmann, M., Avidan, G., Marotta, J. J., & Kimchi, R. (2005).
Detailed exploration of face-related processing in congenital
prosopagnosia: I. Behavioral findings. Journal of Cognitive
Neuroscience, 17, 1130–1149.
Bentin, S., Degutis, J. M., DʼEsposito, M., & Robertson, L. C.
(2007). Too many trees to see the forest: Performance,
event-related potential, and functional magnetic resonance
imaging manifestations of integrative congenital
prosopagnosia. Journal of Cognitive Neuroscience,
19, 132–146.
Calder, A. J., & Young, A. W. (2005). Understanding the
recognition of facial identity and expression. Nature
Reviews Neuroscience, 6, 641–651.
Crawford, J. R., & Howell, D. C. (1998). Comparing an
individualʼs test score against norms derived from small
samples. Clinical Neuropsychology, 12, 482–486.
Das, P., Kemp, A. H., Liddell, B. J., Brown, K. J., Olivieri, G.,
Peduto, A., et al. (2005). Pathways for fear perception:
Modulation of amygdala activity by thalamocortical
systems. Neuroimage, 26, 141–148.
Davies-Thompson, J., Gouws, A., & Andrews, T. J. (2009).
An image-dependent representation of familiar and
unfamiliar faces in the human ventral stream.
Neuropsychologia, 47, 1627–1635.
Duchaine, B., Germine, L., & Nakayama, K. (2007). Family
resemblance: Ten family members with prosopagnosia and
within-class object agnosia. Cognitive Neuropsychology,
24, 419–430.
Duchaine, B., & Nakayama, K. (2005). Dissociations of face
and object recognition in developmental prosopagnosia.
Journal of Cognitive Neuroscience, 17, 249–261.
Duchaine, B., & Nakayama, K. (2006a). The Cambridge Face
Memory Test: Results for neurologically intact individuals
and an investigation of its validity using inverted face
stimuli and prosopagnosic participants. Neuropsychologia,
44, 576–585.
Duchaine, B. C., & Nakayama, K. (2006b). Developmental
prosopagnosia: A window to content-specific face
processing. Current Opinion in Neurobiology, 16,
166–173.
Duchaine, B., Yovel, G., & Nakayama, K. (2007). No global
processing deficit in the Navon task in 14 developmental
prosopagnosics. Social Cognitive and Affective
Neuroscience, 2, 104–113.
Ewbank, M. P., & Andrews, T. J. (2008). Differential sensitivity
for viewpoint between familiar and unfamiliar faces in
human visual cortex. Neuroimage, 40, 1857–1870.
Fox, C. J., Moon, S. Y., Iaria, G., & Barton, J. J. (2009).
The correlates of subjective perception of identity and
expression in the face network: An fMRI adaptation
study. Neuroimage, 44, 569–580.
Furl, N., van Rijsbergen, N. J., Treves, A., Friston, K. J., & Dolan,
R. J. (2007). Experience-dependent coding of facial expression
in superior temporal sulcus. Proceedings of the National
Academy of Sciences, U.S.A., 104, 13485–13489.
Garrido, L., Duchaine, B., & Nakayama, K. (2008). Face
detection in normal and prosopagnosic individuals.
Journal of Neuroscience, 2, 219–240.
Garrido, L., Furl, N., Draganski, B., Weiskopf, N., Stevens, J.,
Tan, G., et al. (2009). Voxel-based morphometry reveals
reduced grey matter volume in the temporal cortex of
developmental prosopagnosics. Brain, 132, 3443–3455.
Furl et al.
1739
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
–
p
r
d
t
i
2
c
3
l
7
e
–
1
p
7
d
2
f
3
/
1
2
9
3
4
/
1
7
4
/
3
1
7
7
o
2
c
3
n
/
1
2
0
7
1
7
0
5
4
2
9
1
7
5
4
/
5
j
o
p
c
d
n
.
b
y
2
0
g
1
u
0
e
.
s
t
2
o
1
n
5
4
0
5
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
t
.
f
/
.
o
n
1
8
M
a
y
2
0
2
1
Grill-Spector, K., Henson, R., & Martin, A. (2006). Repetition
and the brain: Neural models of stimulus-specific effects.
Trends in Cognitive Sciences, 10, 14–23.
Hadjikhani, N., & de Gelder, B. (2002). Neural basis of
prosopagnosia: An fMRI study. Human Brain Mapping,
16, 176–182.
Harris, A., & Nakayama, K. (2006). Rapid face-selective
adaptation of an early extrastriate component in MEG.
Cerebral Cortex, 17, 63–70.
Hasson, U., Avidan, G., Deouell, L. Y., Bentin, S., & Malach, R.
(2003). Face-selective activation in a congenital prosopagnosic
subject. Journal of Cognitive Neuroscience, 15, 419–431.
Haxby, J. V., Hoffman, E. A., & Gobbini, M. I. (2000). The
distributed human neural system for face perception.
Trends in Cognitive Sciences, 4, 223–233.
Hein, G., & Knight, R. T. (2008). Superior temporal sulcus–Itʼs
my area: Or is it? Journal of Cognitive Neuroscience, 20,
2125–2136.
Humphreys, K., Avidan, G., & Behrmann, M. (2007). A detailed
investigation of facial expression processing in congenital
prosopagnosia as compared to acquired prosopagnosia.
Experimental Brain Research, 176, 356–373.
Kriegeskorte, N., Formisano, E., Sorger, B., & Goebel, R. (2007).
Individual faces elicit distinct response patterns in human
anterior temporal cortex. Proceedings of the National
Academy of Sciences, U.S.A., 104, 20600–20605.
Lundqvist, D., Flykt, A., & Öhman, A. (1998). The Karolinska
directed emotional faces—KDEF [CD-ROM]. Stockholm,
Sweden: Department of Clinical Neuroscience, Psychology
Section, Karolinska Institutet.
Minnebusch, D. A., Suchan, B., Köster, O., & Daum, I.
(2009). A bilateral occipitotemporal network mediates face
perception. Behavioural Brain Research, 198, 179–185.
Mur, M., Ruff, D. A., Bodurka, J., Bandettini, P. A., &
Kriegeskorte, N. (2010). Face-identity change activation
outside the face system: “Release from adaptation” may
not always indicate neuronal selectivity. Cerebral Cortex,
doi:10.1093/cercor/bhp272.
Pourtois, G., Schwartz, S., Seghier, M. L., Lazeyras, F., &
Vuilleumier, P. (2005a). Portraits or people? Distinct
representations of face identity in the human visual
cortex. Journal of Cognitive Neuroscience, 17, 1043–1057.
Pourtois, G., Schwartz, S., Seghier, M. L., Lazeyras, F., &
Vuilleumier, P. (2005b). View-independent coding of face
identity in frontal and temporal cortices is modulated by
familiarity: An event-related fMRI study. Neuroimage, 24,
1214–1224.
Rotshtein, P., Henson, R. N., Treves, A., Driver, J., & Dolan,
R. J. (2005). Morphing Marilyn into Maggie dissociates
physical and identity face representations in the brain.
Nature Neuroscience, 8, 107–113.
Schiltz, C., Sorger, B., Caldara, R., Ahmed, F., Mayer, E.,
Goebel, R., et al. (2006). Impaired face discrimination
in acquired prosopagnosia is associated with abnormal
response to individual faces in the right middle fusiform
gyrus. Cerebral Cortex, 16, 574–586.
Simmons, W. K., & Martin, A. (2009). The anterior temporal
lobes and the functional architecture of semantic memory.
Journal of the International Neuropsychological Society,
15, 645–649.
Steeves, J., Dricot, L., Goltz, H. C., Sorger, B., Peters, J.,
Milner, A. D., et al. (2009). Abnormal face identity coding
in the middle fusiform gyrus of two brain-damaged
prosopagnosic patients. Neuropsychologia, 47, 2584–2592.
Thomas, C., Avidan, G., Humphreys, K., Jung, K. J., Gao, F.,
& Behrmann, M. (2009). Reduced structural connectivity
in ventral visual cortex in congenital prosopagnosia.
Nature Neuroscience, 12, 19–31.
van de Stock, J., van de Riet, W. A., Righart, R., & de Gelder, B.
(2009). Neural correlates of perceiving emotional faces and
bodies in developmental prosopagnosia: An event-related
fMRI-study. PLoS ONE, 3, e3195.
von Kriegstein, K., Dogan, O., Grüter, M., Giraud, A. L., Kell,
C. A., Grüter, T., et al. (2008). Simulation of talking faces
in the human brain improves auditory speech recognition.
Proceedings of the National Academy of Sciences, U.S.A.,
105, 6747–6752.
von Kriegstein, K., Kleinschmidt, A., & Giraud, A. L. (2006). Voice
recognition and cross-modal responses to familiar speakersʼ
voices in prosopagnosia. Cerebral Cortex, 16, 1314–1322.
Vuilleumier, P., Armony, J. L., Driver, J., & Dolan, R. J. (2003).
Distinct spatial frequency sensitivities for processing faces and
emotional expressions. Nature Neuroscience, 6, 624–631.
Williams, M. A., Berberovic, N., & Mattingley, J. B. (2007).
Abnormal fMRI adaptation to unfamiliar faces in a case
of developmental prosopamnesia. Current Biology, 17,
1259–1264.
Winston, J. S., Henson, R. N., Fine-Goulden, M. R., & Dolan,
R. J. (2004). fMRI-adaptation reveals dissociable neural
representations of identity and expression in face
perception. Journal of Neurophysiology, 92, 1830–1839.
Yovel, G., & Kanwisher, N. (2005). The neural basis of the
behavioral face-inversion effect. Current Biology, 15,
2256–2262.
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
–
p
r
d
t
i
2
c
3
l
7
e
–
1
p
7
d
2
f
3
/
1
2
9
3
4
/
1
7
4
/
3
1
7
7
o
2
c
3
n
/
1
2
0
7
1
7
0
5
4
2
9
1
7
5
4
/
5
j
o
p
c
d
n
.
b
y
2
0
g
1
u
0
e
.
s
t
2
o
1
n
5
4
0
5
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
f
.
t
/
.
.
o
n
1
8
M
a
y
2
0
2
1
1740
Journal of Cognitive Neuroscience
Volume 23, Number 7