Functional Specificity and Sex Differences in the
Neural Circuits Supporting the Inhibition of
Automatic Imitation
Kohinoor M. Darda*, Emily E. Butler*, and Richard Ramsey
Abstract
■ Humans show an involuntary tendency to copy other people’s
actions. Although automatic imitation builds rapport and affilia-
tion between individuals, we do not copy actions indiscriminately.
Instead, copying behaviors are guided by a selection mechanism,
which inhibits some actions and prioritizes others. To date, the
neural underpinnings of the inhibition of automatic imitation
and differences between the sexes in imitation control are not
well understood. Previous studies involved small sample sizes
and low statistical power, which produced mixed findings regard-
ing the involvement of domain-general and domain-specific neu-
ral architectures. Here, we used data from Experiment 1 (N = 28)
to perform a power analysis to determine the sample size re-
quired for Experiment 2 (N = 50; 80% power). Using indepen-
dent functional localizers and an analysis pipeline that bolsters
sensitivity, during imitation control we show clear engagement
of the multiple-demand network (domain-general), but no sensi-
tivity in the theory-of-mind network (domain-specific). Weaker
effects were observed with regard to sex differences, suggesting
that there are more similarities than differences between the
sexes in terms of the neural systems engaged during imitation
control. In summary, neurocognitive models of imitation require
revision to reflect that the inhibition of imitation relies to a greater
extent on a domain-general selection system rather than a
domain-specific system that supports social cognition. ■
INTRODUCTION
Human social interactions are guided by nonverbal cues,
such as copying behaviors. In the last two decades, much
research has investigated the involuntary tendency to
copy other’s actions—a phenomenon known as auto-
matic imitation (Heyes, 2011). Automatic imitation is
thought to be beneficial in social situations because it
develops affiliative attitudes, better cooperation, and feel-
ings of closeness between interacting partners (Chartrand
& Lakin, 2013). Prior neuroscience research has shown
that imitation is supported by the mirror neuron system,
a neural network engaged in perceiving and performing
actions (Iacoboni, 2009; Rizzolatti & Craighero, 2004;
Iacoboni et al., 1999). Imitation, however, is unlikely to
rely on a single cognitive or brain system (Southgate &
Hamilton, 2008). For example, in many circumstances,
imitation is maladaptive and requires inhibition (Cross &
Iacoboni, 2014; Cross, Torrisi, Losin, & Iacoboni, 2013;
van Schie, van Waterschoot, & Bekkering, 2008; Newman-
Norlund, van Schie, van Zuijlen, & Bekkering, 2007). In
such situations, a selection mechanism is required to
suppress the tendency to imitate and prioritize alternative
actions (Brass, Ruby, & Spengler, 2009). To date, studies
investigating the neural mechanisms of imitation control
Bangor University
*Contributed equally to this work.
have been limited by small sample sizes and low statisti-
cal power, which has produced mixed findings (Table 1).
Furthermore, no neuroscience research has investigated
how individual differences such as sex modulate imita-
tion control, even though behavioral research has shown
that imitative tendencies vary as a function of sex (Butler,
Ward, & Ramsey, 2015; Sonnby-Borgström, Jönsson, &
Svensson, 2008; Dimberg & Lundquist, 1990). Across
two fMRI experiments, which had higher statistical power
and functional sensitivity than prior studies, we investi-
gated the extent to which imitation inhibition relies on a
domain-specific or domain-general neural network, which
varies its response as a function of sex.
Much like cognitive science in general (Kanwisher,
2010; Hirschfeld & Gelman, 1994), inhibitory control
research has focused on a neat division between domain-
general and domain-specific mental operations. Domain-
general inhibitory systems, which operate across multiple
tasks, have been identified in dorsal frontoparietal cortices
(Aron, Robbins, & Poldrack, 2014; Hazeltine, Poldrack, &
Gabrieli, 2007; Nee, Wager, & Jonides, 2007; Wager et al.,
2005; Bunge, Hazeltine, Scanlon, Rosen, & Gabrieli, 2002).
This brain circuit has been labeled the multiple demand
(MD) network because of its engagement in a diversity
of mental operations (Duncan, 2010). By contrast, evi-
dence from fMRI, neurostimulation, and neuropsycholog-
ical patient studies has suggested that a domain-specific
© 2018 by the Massachusetts Institute of Technology. Published under a
Creative Commons Attribution 4.0 International (CC BY 4.0) license.
Journal of Cognitive Neuroscience 30:6, pp. 914–933
doi:10.1162/jocn_a_01261
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
0
6
9
1
4
1
7
8
7
5
2
9
/
j
o
c
n
_
a
_
0
1
2
6
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
circuit in an anterior portion of medial prefrontal cortex
(mPFC) and right temporo-parietal junction (rTPJ) oper-
ates during the inhibition of imitation (Bardi, Gheza, &
Brass, 2017; Santiesteban, Banissy, Catmur, & Bird, 2012,
2015; Sowden & Catmur, 2015; Hogeveen et al., 2014;
Klapper, Ramsey, Wigboldus, & Cross, 2014; Wang, Ramsey,
& Hamilton, 2011; Spengler, von Cramon, & Brass, 2009,
2010; Brass, Derrfuss, Matthes-von Cramon, & von Cramon,
2003; Brass, Zysset, & von Cramon, 2001). Beyond the con-
trol of imitation, mPFC and rTPJ have been consistently im-
plicated in a variety of social cognition functions, which
require distinguishing between self and other, as well as
reasoning about other people’s mental states (theory of
mind [ToM]; Van Overwalle, 2009; Amodio & Frith, 2006;
Saxe & Kanwisher, 2003; Frith & Frith, 1999). These results
led to theorizing that a key neural circuit for social cogni-
tion also regulates imitative tendencies (Brass et al., 2009).
Although theories of imitation control have been devel-
oped that are based on functioning of the ToM network,
evidence from fMRI studies that used an RT measure of
imitation inhibition have not provided consistent support
for the involvement of a domain-specific neural network
(Table 1). The RT measure of imitation involves making
finger movements while simultaneously watching compat-
ible or incompatible finger movements (Brass, Bekkering,
Wohlschlager, & Prinz, 2000; Stürmer et al., 2000). The dif-
ference between RTs in these two conditions (i.e., the gen-
eral compatibility effect) has been argued to index imitative
control, as greater cognitive resources are required to inhibit
movements that are incompatible to one’s own responses
(Heyes, 2011; Brass & Heyes, 2005). Approximately half of
the fMRI studies using this paradigm failed to find
engagement of rTPJ and anterior mPFC. In addition, a
number of studies showed engagement of regions asso-
ciated with the MD network, including dorsal fronto-
parietal cortex, supplementary motor area (SMA) and
anterior insula (Marsh, Bird, & Catmur, 2016; Cross &
Iacoboni, 2013; Mengotti, Corradi-Dell’Acqua, & Rumiati,
2012; Crescentini, Mengotti, Grecucci, & Rumiati, 2011; Bien,
Roebroeck, Goebel, & Sack, 2009). Moreover, the most com-
mon measure of imitation interference is confounded by
spatial compatibility or the tendency to respond faster to a
stimulus when it is on the same side of space as the re-
sponse (e.g., Simon, 1969). To measure imitation interfer-
ence independent of spatial compatibility effects, spatial
and imitative processes need to be dissociated (Gowen,
Bolton, & Poliakoff, 2016; Marsh et al., 2016; Boyer, Longo,
& Bertenthal, 2012; Cooper, Catmur, & Heyes, 2012; Catmur
& Heyes, 2011; Wiggett, Hudson, Tipper, & Downing, 2011;
Bertenthal, Longo, & Kosobud, 2006). Therefore, the extent
to which imitation inhibition relies on domain-specific and
domain-general architectures remains unclear. Indeed, no
Table 1. fMRI Studies Investigating Imitation Control Using Modified Versions of the Imitation Inhibition Task
Sample
(Male:Female)
Dissociation of Imitative
and Spatial Processes
Analysis
ToM
ROI
Whole-brain
mPFC
rTPJ
MD
Brain Networks
Brass et al., 2001
Brass et al., 2005
Brass et al., 2009
Spengler et al., 2009
Bien et al., 2009
Crescentini et al., 2011
10 (4:6)
20 (8:12)
20a
18 (9:9)
15 (5:10)
19 (9:10)
Cross & Iacoboni, 2013
24 (12:12)
Mengotti et al., 2012
22 (10:12)
Cross et al., 2013
Klapper et al., 2014
Marsh et al., 2016
Wang et al., 2011
25 (5:15)
19 (2:17)
24 (7:17)
20 (5:15)
ü
ü
ü
ü
ü
ü
ü
ü
ü
ü
ü
ü
ü
ü
ü
ü
ü
ü
üb
ü
ü
ü
ü
ü
ü
ü
ü
ü
ü
ü
ü
ü
ü
ü
ü
ü
ü
ü
ü
ü
Evidence that the engagement of mPFC and rTPJ is inconsistent across fMRI studies that investigated imitation control using modified versions of the
imitation inhibition task. For all studies, engagement of mPFC or rTPJ is reported only for contrasts that test for inhibiting the urge to automatically
imitate. Engagement of the MD network is reported only for whole-brain analyses. Except for Wang et al. (2011), which used hand movements, all
other tasks used modified versions of the imitation inhibition tasks involving finger movements (Brass et al., 2000). For a more detailed version of
this table, see Supplementary Table S6.
aNumber of male and female individuals not mentioned.
bmPFC showed engagement only at p < .005, uncorrected.
Darda, Butler, and Ramsey
915
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
6
9
1
4
1
7
8
7
5
2
9
/
j
o
c
n
_
a
_
0
1
2
6
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
research to date has dissociated spatial from imitative
processes and used a functional ROI (fROI) approach
(Fedorenko, Duncan, & Kanwisher, 2013; Kanwisher,
2010). Using a fROI approach enables investigation of
how functionally defined brain circuits, such as the MD
and ToM networks, operate during the control of imitation.
A further area of imitation research that has received
little attention is the extent to which imitative control varies
across individuals, especially between the sexes. It has
been argued that imitation is modulated by stable indi-
vidual differences, such as empathy (Chartrand & Lakin,
2013) and sex (Butler et al., 2015; Sonnby-Borgström et al.,
2008). Although it has been suggested that women excel
across a range of social processes compared with men
(Baron-Cohen, 2002), only a limited number of studies have
investigated sex differences in social cognition, and the re-
sults are often mixed, do not replicate, or are specific to very
select contexts or samples (Hyde, 2014; Miller & Halpern,
2014). Furthermore, studies of sex differences in social cog-
nition have mainly focused on emotional expression per-
ception and mental state reasoning with little emphasis
placed on imitation (Krach et al., 2009; Russell, Tchanturia,
Rahman, & Schmidt, 2007; Rahman, Wilson, & Abrahams,
2004; Campbell et al., 2002; Thayer & Johnsen, 2000).
A recent study that used an RT measure of imitation
inhibition (Brass et al., 2000) showed that female indi-
viduals showed a greater level of interference than male
individuals (Butler et al., 2015). It is possible that this
sex difference in imitation control may be mediated by
empathy—female individuals have been shown to be more
empathetic compared with male individuals (Christov-
Moore et al., 2014; Baron-Cohen & Wheelwright, 2004).
However, even though empathy has been associated with
different types of imitation paradigms (Müller, Leeuwen,
Baaren, Bekkering, & Dijksterhuis, 2013; Sonnby-Borgström,
Jönsson, & Svensson, 2003; Sonnby-Borgström, 2002;
Chartrand & Bargh, 1999), the evidence to date suggests
that there is no link between imitation, as measured by
RTs, and empathy (Genschow et al., 2017; Butler et al.,
2015). In addition, in the study by Butler and colleagues
(2015), it is unclear whether sex modulates the tendency
to automatically imitate or the tendency to automatically
respond in the same spatial location to the observed
action. The former indicates a sex difference that is spe-
cifically tied to imitation control, whereas the latter might
indicate a sex difference in processes associated with re-
solving spatial conflict. More recent work also showed a
greater interference effect for female individuals com-
pared with male individuals (Genschow et al., 2017), as
well as greater error rates for predominantly female sam-
ples than male samples (Cracco et al., 2018). The imitation
task used by Genschow and colleagues (2017) was con-
trolled for left–right spatial compatibility by presenting
the stimulus hand orthogonal to the response. Even
though this shows that the sex difference remains when
spatial compatibility is reduced, it does not rule out the
possibility of orthogonal spatial compatibility ( Weeks &
Proctor, 1990). More generally, sex differences have been
found on a wide range of inhibitory control tasks, including
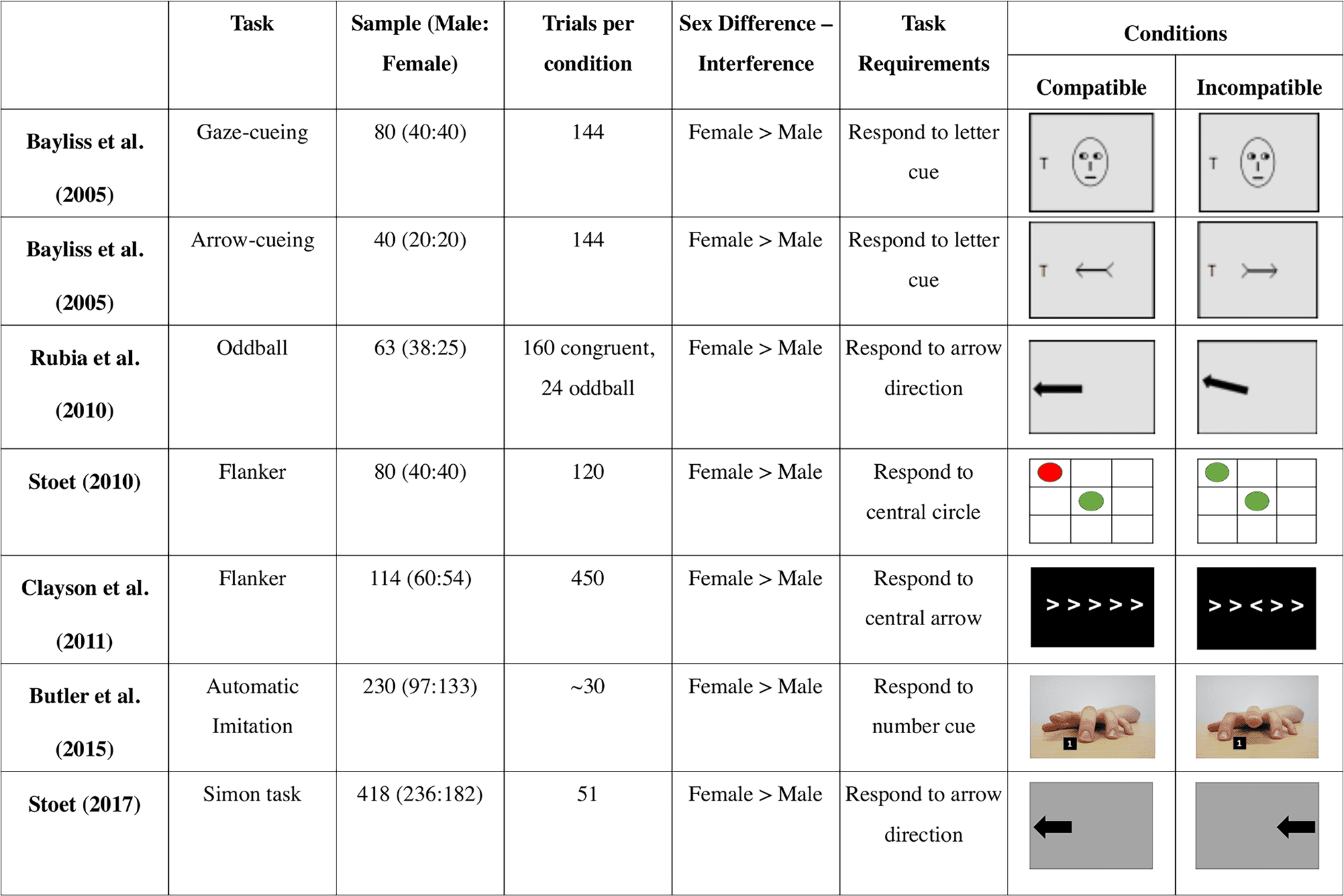
flanker, gaze cueing, arrow cueing, oddball, and Simon
tasks, wherein female individuals have been shown to re-
quire more cognitive resources than male individuals to
inhibit automatic response tendencies (Figure 1; Stoet,
2010, 2017; Clayson, Clawson, & Larson, 2011; Rubia, Hyde,
Halari, Giampietro, & Smith, 2010; Bayliss, di Pellegrino, &
Tipper, 2005). It is possible, therefore, that a domain-
general system may underpin the sex differences observed
across these tasks, including during imitation control, but
no research to date has directly investigated this proposal.
Across two fMRI experiments, the current study inves-
tigated functional specificity and sex differences in imita-
tion control. Several aspects of the experimental design
provide grounds to extend current understanding in
meaningful and concrete ways. First, this is the first study
to use independent functional localizers to identify MD
and ToM networks in single subjects and directly test
the involvement of these networks in imitation control.
By doing so, we can directly test hypotheses regarding
the role of functionally defined neural circuits (i.e., MD
and ToM networks) and therefore minimize the reliance
on reverse inference to infer cognitive function based on
anatomical localization (Poldrack, 2006). Second, we
used data from Experiment 1 to perform a power analysis
to determine the sample size required to achieve a de-
sired level of power in Experiment 2. Given the inconsis-
tent findings in prior studies, which had relatively small
sample sizes, this multiexperiment approach made sure
that our key experiment had over 80% power to detect
expected effect sizes. Third, to avoid spatial compatibility
confounds, in Experiment 2, we used a modified version
of the imitation inhibition paradigm that allowed for an
independent measure of spatial and imitative compatibil-
ity (Catmur & Heyes, 2011). If the inhibition of automatic
imitation relies on a domain-specific neural architecture
that is associated with social cognition, as proposed by
Brass and colleagues (2009), mPFC and rTPJ would be
engaged in imitative control. In contrast, engagement
of the MD network would suggest that domain-general
processes subserve imitation control. Furthermore, the
sex difference found previously (Butler et al., 2015) may
be supported by differences in ToM or MD networks.
METHODS
Overview of the Experimental Approach
Experiment 1 used a group-level whole-brain analysis,
which provided the basis for power analyses that set up
Experiment 2 as the critical experiment with high sta-
tistical power (80%). In Experiment 2, to increase sensi-
tivity and functional resolution, we used independent
localizers to identify key functional circuits (i.e., MD
and ToM networks), and analyses were performed in
single subjects to precisely quantify the consistency of
916
Journal of Cognitive Neuroscience
Volume 30, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
6
9
1
4
1
7
8
7
5
2
9
/
j
o
c
n
_
a
_
0
1
2
6
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
6
9
1
4
1
7
8
7
5
2
9
/
j
o
c
n
_
a
_
0
1
2
6
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 1. Sex differences in inhibitory control tasks. Female individuals experience greater interference than male individuals in multiple inhibitory
control tasks. Images are produced based on figures and description in each experiment apart from Butler et al. (2015), which are the actual images used.
Also, in Rubia et al. (2010), the sex difference showed increased interference by the oddball trials rather than the incongruent trials, and this is what is
represented by the images. Finally, in Butler et al. (2015), participants completed 60 trials that were 30 ± 2 trials per compatible and incompatible condition.
network engagement across individuals (Nieto-Castañón
& Fedorenko, 2012; Kanwisher, 2010). Group-level anal-
yses require responses across individuals to overlap in in-
dividual voxels. In contrast, the fROI approach allows
identification of corresponding functional regions with-
out the requirement of exact voxel overlap across individ-
uals. Therefore, the same voxels need not be active
across individuals, as long as voxels within a functionally
defined ROI are consistently active across individuals.
Consequently, group-level analyses may underestimate
functional specificity, whereas fROI analyses can show in-
creased sensitivity (Nieto-Castañón & Fedorenko, 2012).
In addition, because of a constrained search volume,
fROI analyses typically have higher statistical power than
whole-brain analyses (Fedorenko, Hsieh, Nieto-Castañón,
Whitfield-Gabrieli, & Kanwisher, 2010; Saxe, Brett, &
Kanwisher, 2006).
Experiment 1
Participants
Twenty-eight participants (Mage = 23.96, SDage = 5.52;
14 women) participated for monetary compensation of
£15. Participants gave informed consent in line with the
guidelines set by the Research Ethics and Governance
Committee of the School of Psychology at Bangor Uni-
versity, were right-handed, had normal or corrected-to-
normal vision, and reported no history of neurological
damage.
Design and Procedure
All participants performed the imitation task inside the
scanner. The participants also did four additional tasks
in the same scanning session as part of another experi-
ment. The scanning session started with the imitation
task, followed by a run of a face perception task, a flanker
task (Erikson & Erikson, 1974), another run of the face
perception task, a dynamic face localizer (Pitcher, Dilks,
Saxe, Triantafyllou, & Kanwisher, 2011), and a ToM loca-
lizer (Dodell-Feder, Koster-Hale, Bedny, & Saxe, 2011).
The order of the tasks was counterbalanced across partic-
ipants such that, of the 28 participants, 14 participants
did the imitation task first, and 14 participants did the
flanker task first, with the order of the other tasks remain-
ing the same.
Darda, Butler, and Ramsey
917
The imitation inhibition task. The imitation task was
based on a stimulus–response compatibility paradigm
developed by Brass et al. (2000) consisting of observation
and execution of finger-lifting movements during fMRI scan-
ning. Before the task, participants were instructed to hold
down the “blue” and “yellow” buttons on the response
box with their index and middle fingers of the right hand,
respectively, A number cue (either “1” or “2”) was presented
to participants, and they were asked to lift their index finger
on presentation of the number “1” and the middle finger for
the number “2.” Simultaneously, they also viewed an image
of an index or middle finger lift of a left hand viewed from the
third-person perspective, such that the fingers extended to-
ward the participants. Thus, there were four trial types in an
event-related design that led to two conditions—participants
performing the same (congruent) or different (incongruent)
finger movement to the observed hand image.
Each trial started with a fixation cross (500 msec) followed
by a neutral hand (for a random ISI of 500, 700, or
1000 msec) and a hand image with an index/middle finger
lift, which stayed onscreen for 2000 msec, irrespective of
when the participant made the response. Sequencing the
hand images in such a way led to the appearance of appar-
ent motion of the finger. After 2000 msec, the next trial
started immediately with a fixation cross (500 msec). To sep-
arately model the influence of individual events in an event-
related design, the four trial types were pseudorandomized,
such that each trial type was preceded by each other trial
type and by itself an equal number of times ( Wager &
Nichols, 2003; Josephs & Henson, 1999). There were 17
trials in each block. The first trial was used to set up the
randomization sequence but excluded from the analysis as
it was not preceded by any other trial. The remaining 16 tri-
als within a block were analyzed and consisted of eight trials
per condition. Each run consisted of five blocks separated
by a 3-, 4-, or 5-sec fixation cross. All participants completed
one run of the imitation task. Thus, there were 80 trials of
interest (40 congruent and 40 incongruent).
Behavioral Data Analysis
RT on the imitation inhibition task was measured as the
time from number cue onset to when participants made
a response. To ensure participants were engaging cor-
rectly with the task, participants who had less than 80%
accuracy were removed. In addition, RTs more than 3 SDs
away from the mean were excluded from the analyses.
Furthermore, trials on which participants made an “error”
were excluded from the analyses. Errors included an in-
correct response, no response, a response after 2000 msec,
and pressing an invalid key. The general compatibility effect
was calculated as the RT difference between incompatible
and compatible trials. A one-sample t test was performed
to verify the presence of a general compatibility effect. A
one-tailed independent sample t test was performed to
determine if the compatibility effect was greater for female
than male individuals. Mean differences, 95% confidence
intervals (CI), and Cohen’s d (Cohen, 1992) are reported
for all effects of interest. For the one-sample t test, Cohen’s
dz was calculated as mean difference divided by the stan-
dard deviation of the sample (Lakens, 2013). The 95% CI is
reported for the lower bound for a one-tailed t test. For
the independent samples t test, Cohen’s d was calculated
as mean difference between the two groups divided by
the pooled standard deviation (Cohen, 1992).
fMRI Data Analysis
Data acquisition. Participants were placed supine in a
3-T Philips MRI scanner using a SENSE 32-channel phased
array coil. They were requested to avoid head motion
during the scanning session and were presented stimuli
on a computer screen placed behind the scanner made
visible by a mirror attached to the head coil. Responses
on the task were recorded with the help of a button box
that recorded RTs. Thirty-five axial slices were acquired in
an ascending order using a T2*-weighted EPI sequence.
The reference slice for slice time correction was the slice
acquired in the middle of the sequence (Slice 17). Param-
eters are as follows: voxel size = 3 × 3 × 4 mm, repetition
time = 2000 msec, echo time = 30 msec, flip angle = 90°,
slice thickness = 4 mm, slice gap = 0.8 mm, field of view =
230 × 230 × 167 mm3. One hundred seventy-four volumes
were collected for the imitation task.
Four dummy scans collected at the beginning of each
run of the task were not included in any analyses. A high-
resolution T1-weighted anatomical image was also col-
lected with the following parameters: repetition time =
12 msec, echo time = 3.5 msec, flip angle = 8°, number
of axial slices = 170, voxel size = 1 mm3, field of view =
250 × 250 × 170 mm3.
Data preprocessing and general linear model. Func-
tional images were preprocessed in SPM-8. Data were
realigned, unwarped, and corrected for slice timing. Data
were normalized to the Montreal Neurological Institute
(MNI) template with a resolution of 3 mm3, and images
were spatially smoothed (8 mm).
For the imitation task, a design matrix was fit for each
participant with three regressors: one each for the cor-
rect trials of the two conditions, and one for the “new”
trials (i.e., the first trial of each block). The new trials
were not used in any further analyses. Stimulus onsets
were time-locked to the presentation of the number
cue with a duration of 0 sec and convolved with the stan-
dard hemodynamic response function.
Whole-brain analyses. Contrast images (incompatible >
compatible) were calculated at the single-subject level for
the imitation inhibition task to identify regions of the brain
showing a compatibility effect. Group-level contrast images
were created from these single-subject contrast images to
identify regions that were consistently engaged for the
compatibility effect across the sample using one-sample
918
Journal of Cognitive Neuroscience
Volume 30, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
0
6
9
1
4
1
7
8
7
5
2
9
/
j
o
c
n
_
a
_
0
1
2
6
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
t tests. To identify a neural signature of the sex difference
in imitation inhibition, a Sex × Compatibility ANOVA was
computed (female [incompatible > compatible] > male
[incompatible > compatible]) as female individuals have
been shown to have a higher compatibility effect than
male individuals in the imitation task (Butler et al., 2015).
For all analyses, contrast images were taken to the group
level and thresholded using a voxel-level threshold of
p < .001 and a voxel extent of 10 voxels. Correction for
multiple comparisons was performed at the cluster level
(Friston, Worsley, Frackowiak, Mazziotta, & Evans, 1994),
with clusters that survive correction for multiple correc-
tions using a family-wise error correction ( p < .05; shown
in bold font in Table 2A and B; see Results). This restricts
the likelihood of false positives (Eklund et al., 2016). Clus-
ters of activity were identified with the SPM Anatomy
toolbox (Eickhoff et al., 2005).
Experiment 2
Participants
Fifty-five participants (Mage = 22.04, SDage = 3.70; 27 women)
were recruited from the Bangor community and were either
reimbursed with £15 or three course credits for their partic-
ipation. Informed consent was obtained in line with the
guidelines set by the Research Ethics and Governance Com-
mittee of the School of Psychology at Bangor University. All
participants were right-handed, did not have dyslexia or
dyspraxia, were not on any medication, did not report
neurological damage, and had normal or corrected-to-
normal vision. The sample size was determined by a
power analysis based on Experiment 1 data (see Results).
Design and Procedure
Each participant performed three tasks inside the scanner—
the automatic imitation task, a ToM network localizer
task, and an MD network localizer task. The order of the
tasks was as follows: two runs of the MD network localizer
task were interspersed between three runs of the imitation
task to offset boredom. This was followed by two runs of
the ToM network localizer task. The ToM task was always
presented at the end to reduce the likelihood that belief
reasoning during the ToM task would influence perfor-
mance in the imitation task. The order was the same for
all participants. Participants also completed a 50-item Inter-
national Personality Item Pool questionnaire (Donnellan,
Oswald, Baird, & Lucas, 2006; Goldberg, 1992; unrelated
to the current study) and a stimulus rating form where they
were asked to rate the hand stimulus from the imitation
task as either male, female, or neutral. The entire session
lasted approximately 1.5 hr, with 60 min inside the scanner.
All stimulus presentation was coded in MATLAB 2015b and
presented with PsychToolBox 3.0.6.
The imitation inhibition task. The automatic imitation
task was similar to the one used in Experiment 1, but
with two changes. First, we used a different hand stimu-
lus, which was rated as sex-neutral by observers. The sex
of the hand was an important consideration to minimize
the possibility of an own-sex bias while exploring sex dif-
ferences in imitation inhibition. As such, we conducted
pilot work that asked observers to evaluate a range of
hand stimuli in terms of masculinity and femininity, and
we selected the most sex-neutral stimulus (see Supple-
mentary Information, Development of Stimuli). We only
used one hand stimulus to simplify the design space.
Although using one sex-neutral hand stimulus provided
greater experimental control, it may have harmed our abil-
ity to study or elicit sex differences. Future work could
probe this further by varying the sex of the stimulus and/or
by using more sex-typical stimuli.
The second change that we made was to calculate an
imitative compatibility effect independent of spatial com-
patibility (Catmur & Heyes, 2011). To do so, participants
viewed an image of an index or middle finger lift of either
a right or left hand but always responded with their right
hand. Using right- and left-hand images produced eight
trial types and four main conditions of interest (see
Figure 2A). For example, when cued to lift their index
finger while observing a left-hand index finger lift, the
observed movement is both imitatively compatible (same
finger), as well as spatially compatible (same side of space
to the executed movement). In contrast, when observing
a right-hand index finger lift, the participant’s response is
imitatively compatible (same finger), but it is not on the
same side of space (they are spatially incompatible). Thus,
participants performed the same (imitatively compatible)
or different (imitative incompatible) finger movement on
the same (spatially compatible) or different (spatially in-
compatible) side of space to the observed finger move-
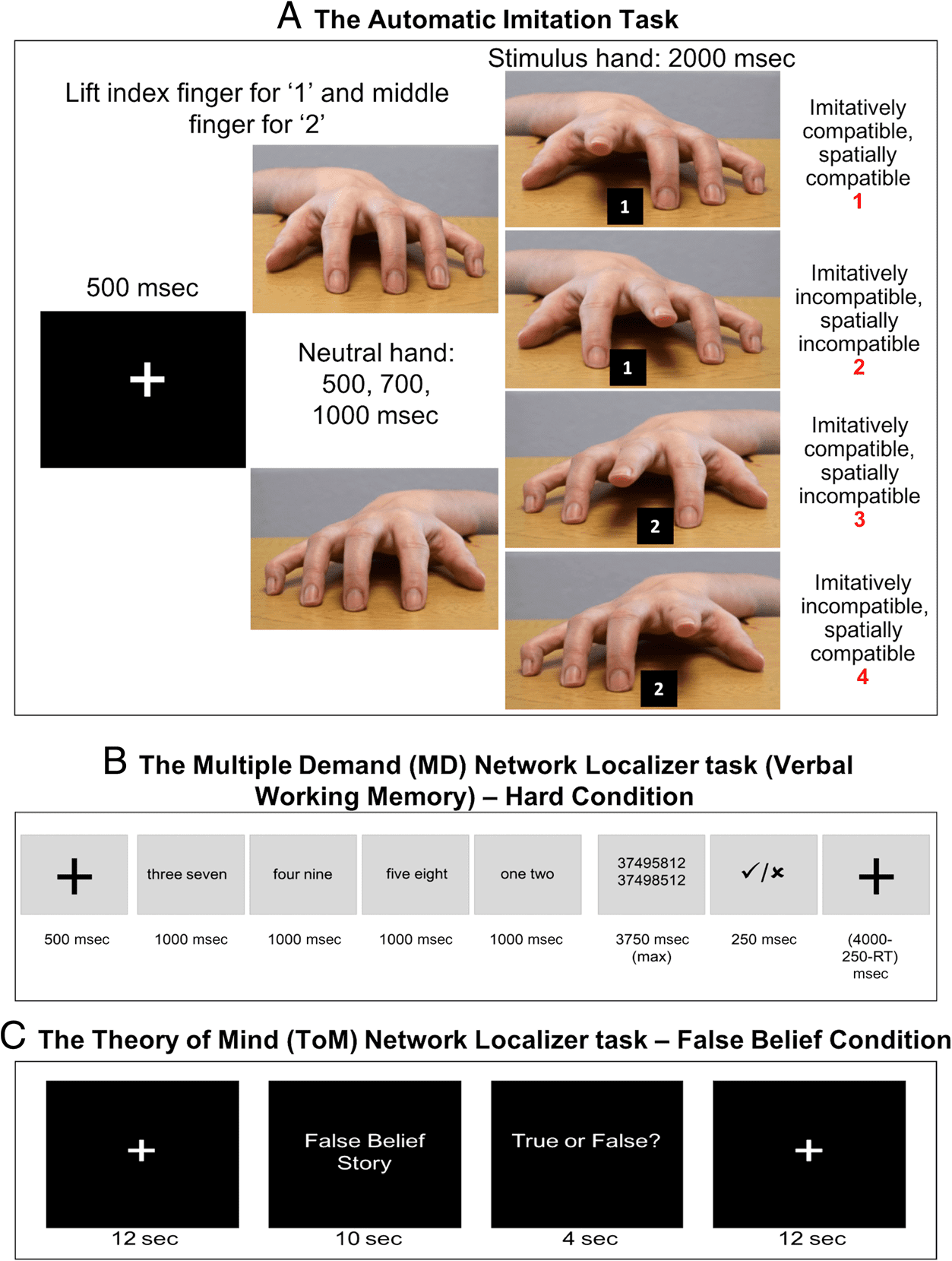
ment, giving rise to the following four conditions:
1.
2.
3.
4.
Imitatively and spatially compatible
Imitatively and spatially incompatible
Imitatively compatible and spatially incompatible
Imitatively incompatible and spatially compatible
Sequencing information and pseudorandomization was
the same as Experiment 1. There were 65 trials in each
block. The first trial was used to set up the randomization
sequence but excluded from the analysis as it was not
preceded by any other trial. The remaining 64 trials were
analyzed, consisting of 16 trials per condition. Each run
consisted of two blocks separated by a 3-sec fixation
cross. All participants completed three runs of the imita-
tion task. In total, there were 384 trials of interest, 96 per
condition. Experiment 2, therefore, had more than twice
the number of trials per condition than Experiment 1.
Localizer tasks
The MD network localizer. To identify regions of the
MD network, a verbal working memory ( WM) task was
used (Fedorenko, Behr, & Kanswisher, 2011). Participants
Darda, Butler, and Ramsey
919
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
6
9
1
4
1
7
8
7
5
2
9
/
j
o
c
n
_
a
_
0
1
2
6
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Table 2. General Compatibility Effect and Sex × Compatibility Interaction for the Imitation Inhibition Task (Experiment 1)
MNI Coordinates
Region
Cluster Size
p FWE Corr
t
x
y
(A) General Compatibility Effect (Incompatible > Compatible)
L IPL extending into SPL and
superior frontal gyrus
986
<.001
L cerebellum
150
.001
R cerebellum
198
<.001
R PrecG extending across superior
183
<.001
frontal gyrus and MFG
R postcentral gyrus extending into
481
<.001
SPL and IPL
R posterior middle temporal gyrus
L insula
R posterior medial frontal cortex
L posterior medial frontal cortex
R pallidum extending into thalamus
L paracentral lobule
R middle cingulate cortex
41
24
20
55
11
11
20
.179
.458
.564
.083
.834
.834
.564
8.40
6.50
6.38
5.79
4.95
4.72
5.71
5.12
4.32
5.12
5.04
4.39
5.18
4.55
4.50
4.61
4.13
3.63
4.67
4.79
4.40
3.82
4.14
3.80
3.89
3.85
3.78
3.67
z
43
70
70
−41
−35
−44
−44
−32
−23
70
58
61
73
67
37
1
−8
13
−2
73
52
52
−2
7
79
43
49
52
70
46
52
−38
−29
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
6
9
1
4
1
7
8
7
5
2
9
/
j
o
c
n
_
a
_
0
1
2
6
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
−39
−36
−27
−21
−30
−9
21
45
39
27
42
39
33
42
48
66
57
60
−36
3
−3
−6
21
15
−40
−37
−7
−55
−55
−70
−58
−46
−55
−1
2
−10
−40
−40
−34
−46
−40
−43
17
−4
−1
11
−7
−6
−12
−19
9
6
6
14
8
17
−37
−19
−31
−55
−55
(B) Sex × Compatibility [Female (Incompatible > Compatible) > Male (Incompatible > Compatible)]
−21
L SPL extending into postcentral gyrus
.011
4.98
93
L cerebellum
16
.679
4.80
4.60
4.34
4.04
−30
−24
−24
−21
Regions surviving a voxel-level threshold of p < .001, and 10 voxels are reported for the (A) general compatibility effect and (B) Sex × Compatibility
interaction for the imitation inhibition task. Subclusters at least 8 mm from the main peak are listed. Bold font indicates clusters that survive correction
for multiple corrections using a family-wise error (FWE) correction ( p < .05). MNI = Montreal Neurological Institute; SPL = superior parietal lobule;
IPL = inferior parietal lobule; PrecG = precentral gyrus; MFG = middle frontal gyrus; L = left hemisphere; R = right hemisphere.
920
Journal of Cognitive Neuroscience
Volume 30, Number 6
Figure 2. Stimuli for the
imitation inhibition and
functional localizer tasks.
Stimuli and trial design for the
imitation inhibition task (A),
the Multiple Demand (MD)
network localizer task (B), and
the Theory-of-Mind network
localizer task (C). For the
automatic imitation task, spatial
compatibility was calculated as
spatial incompatible (2+3)
minus (1+4), and imitative
compatibility was calculated
as imitatively incompatible
(2+4) minus imitative
compatible (1+3) trials.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
6
9
1
4
1
7
8
7
5
2
9
/
j
o
c
n
_
a
_
0
1
2
6
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
were asked to remember the sequence in which either
four (easy condition) or eight (hard condition) digit se-
quences were presented on screen (see Figure 2B). After
each trial, participants had to choose between two digit
sequences presented numerically, one of which matched
the sequence in which the digits were presented as words.
Feedback was provided as to whether they answered cor-
rectly or incorrectly. The hard > easy contrast has been
found to robustly activate regions of the MD network
(Fedorenko et al., 2011, 2013). Each run consisted of 10 ex-
perimental blocks (each 34 sec long) and 6 fixation blocks
(each 16 sec long). The total run lasted for 436 sec. Each
participant completed two runs of the WM task.
The ToM localizer. To localize brain regions involved in
mental state reasoning, we used a paradigm developed
by Dodell-Feder and colleagues (2011; saxelab.mit.edu/
superloc.php). This localizer task (see Figure 2C) includes
20 stories, each describing a false representation. Ten
stories included out-of-date beliefs (the false belief condi-
tion), and the other 10 included out-of-date physical repre-
sentations (photographs/maps; the false photograph
condition). The false belief > false photograph contrast
has been shown in prior work to robustly activate regions
involved in mentalizing (Dufour et al., 2013). All trials con-
sisted of a story (10 sec), followed by a true or false question
(4 sec). Each story was separated by a 12-sec rest period.
The order of the stories and conditions was the same for
all participants. Each participant completed two runs of this
task, with five trials per condition presented in each run.
Behavioral Data Analysis
RT and accuracy were recorded in the same way as Exper-
iment 1. Compatibility effects were calculated as follows:
spatial compatibility = spatially incongruent trials − spa-
tially congruent trials; imitative compatibility = imitatively
incongruent trials − imitatively congruent trials. Behavioral
data were analyzed in the same fashion as Experiment 1,
only separately for imitative and spatial compatibility
effects. The main aim of the experiment was to test for
the presence of imitative and spatial compatibility effects,
Darda, Butler, and Ramsey
921
as well as for differences between the sexes (female >
male). Hence, we used a one-sample t test to verify the
presence of spatial and imitative compatibility effects and
a one-tailed independent samples t test to test whether
female individuals showed a higher spatial/imitative com-
patibility effect than male individuals.
the MD localizer). The onset and duration of each con-
dition was specified and convolved with the standard
hemodynamic response function. Contrast images were
then calculated for each individual subject to identify
regions that responded to cognitive demand (hard >
easy) and mentalizing (belief > photo).
fMRI Data Analysis
Data acquisition. Data acquisition procedures were
the same as Experiment 1. There were 249 volumes col-
lected for the imitation task, 219 for the MD network
localizer, and 136 for the ToM localizer for each run.
Data preprocessing and general linear model. All MRI
data were preprocessed in SPM-8. Data were realigned,
unwarped, and corrected for slice timing. Data were nor-
malized to the MNI template with a resolution of 3 mm3.
Normalizing to a common space instead of the individ-
ual’s native anatomical space allows for comparisons with
previous studies (relying on the common space) and is
preferred when definition of fROIs is based on group-
constrained functional data (Nieto-Castañón & Fedorenko,
2012). Images were spatially smoothed (8 mm).
For the imitation task, a design matrix was fit for each
participant with five regressors: one each for the correct
trials of the four conditions and one for “new” trials (i.e.,
the first trial of each block). Stimulus onsets were time-
locked to the presentation of the number cue with a
duration of 0 sec and convolved with the standard hemo-
dynamic response function. Contrast images were calcu-
lated for each individual participant to identify regions of
the brain showing a spatial (spatially incompatible > spa-
tially compatible) or imitative (imitatively incompatible >
imitatively compatible) compatibility effect.
For the localizer tasks, the design matrix consisted of
regressors for each experimental condition (“Belief” and
“Photo” for the ToM localizer and “Hard” and “Easy” for
Definition of group-constrained subject-specific analyses.
For the group-constrained subject-specific (GSS) analyses,
the spm_ss toolbox was used, which runs in SPM using
MATLAB (web.mit.edu/evelina9/www/funcloc.html). The
GSS approach developed by Fedorenko et al. (2010) and
Julian, Fedorenko, Webster, and Kanwisher (2012) was
used to define fROIs for each participant. These fROIs
were defined using (1) each individual’s activation map
for the localizer tasks and (2) group-constraints or masks.
These masks refer to a set of “parcels,” which demarcate
areas in the brain where prior work has been shown to
exhibit activity for the localizer contrasts.
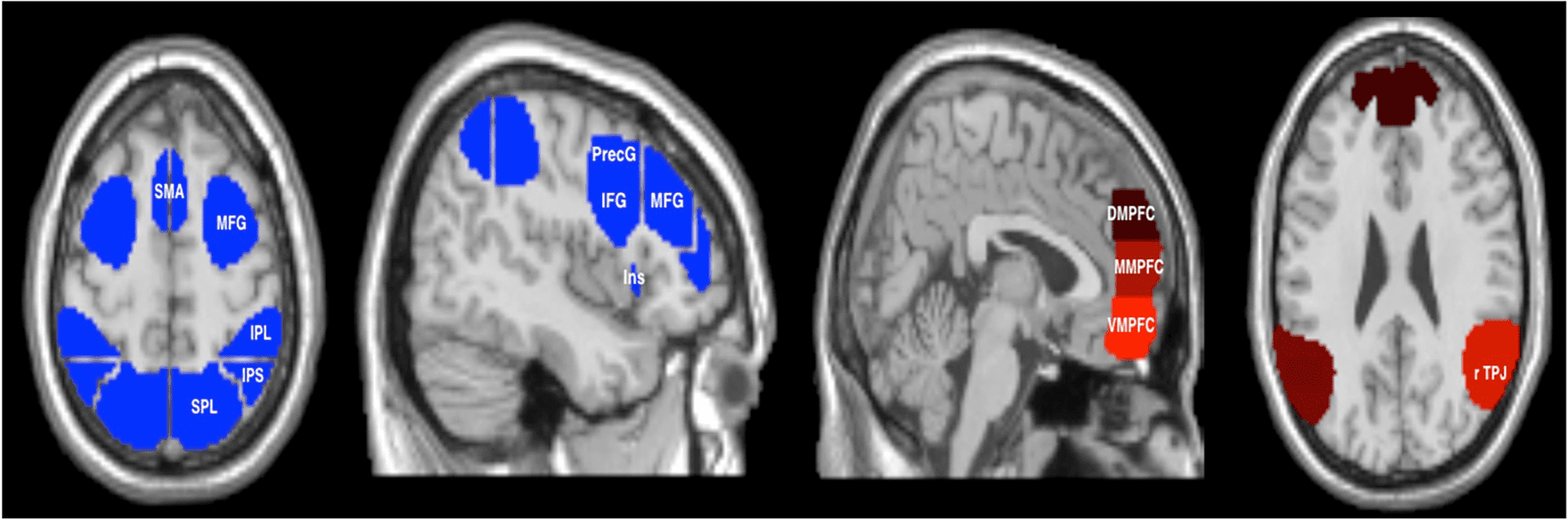
Two sets of fROIs were defined (Figure 3): MD net-
work fROIs that have been known to exhibit activity for
a variety of cognitive control tasks (Fedorenko et al.,
2013; Duncan, 2010) and ToM network fROIs that
support mentalizing and have been specifically impli-
cated for imitation inhibition (Brass et al., 2009; Saxe &
Kanwisher, 2003). For the ToM network, four parcels
were derived from a group-level map from 462 partici-
pants for the false belief > false photograph contrast
(Dufour et al., 2013). These regions included the dorsal,
middle, and ventral mPFC (DMPFC, MMPFC, and VMPFC,
respectively) and the rTPJ. For the MD network, we used
16 parcels derived from a set of functional parcels cre-
ated by Idan Blank based on a probabilistic overlap map
from 197 participants (available at https://evlab.mit.edu/
funcloc/download-parcels). These included areas in bilat-
eral superior and inferior parietal lobules (SPL and IPL,
respectively), inferior parietal sulcus (IPS), inferior and
middle frontal gyri (IFG, MFG), precentral gyrus (PrecG),
Figure 3. Graphical representation of the parcels used to define the MD and ToM network fROIs. The MD network consisted in 16 parcels, and the
ToM network included 4 parcels.
922
Journal of Cognitive Neuroscience
Volume 30, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
0
6
9
1
4
1
7
8
7
5
2
9
/
j
o
c
n
_
a
_
0
1
2
6
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
insula, and the SMA. These areas were chosen for two
reasons: (1) they were part of the MD network (Fedorenko
et al., 2013) and (2) they have been shown to respond in
prior work to the specific type of interference control of
relevance to the current study (Marsh et al., 2016; see also
Experiment 1).
For each individual, these masks were used to con-
strain the selection of subject-specific fROIs. For each
individual, for the ToM network mask, the belief > photo
contrast was used, and the top 10% of voxels (based on
t values) within each parcel were defined as that indi-
vidual’s fROI. Similarly, for the MD network mask, each
individual’s top 10% of voxels (based on t values) in
the hard > easy contrast were defined as that individual’s
fROI. Using the top 10% of voxels, rather than a fixed
threshold (e.g., all voxels with p < .001), ensures a con-
stant size of each fROI across individuals (Blank,
Kanwisher, & Fedorenko, 2014). We also ran the analyses
using a fixed threshold ( p < .001, uncorrected) and
found the same pattern of results (see Supplementary
Tables S1.1 and S1.2). All analyses reported below are
based on the top 10% of voxels that were generated from
the localizer data fROIs. Percent signal change values were
extracted from all fROIs. For the main analysis, all runs of
the localizer tasks were used to define fROIs in each indi-
vidual. Responses in these fROIs were estimated for spatial
and imitative compatibility effects.
In a supplementary analysis, responses to the localizer
contrasts were also estimated to ensure that all the fROIs
showed the expected response with respect to the locali-
zer contrasts. This supplementary analysis ensured that
the ToM network showed a robust belief > photo and
the MD network showed a robust hard > easy effect.
For these localizer analyses, an across-run cross-validation
approach was used (Nieto-Castañón & Fedorenko, 2012)
to ensure that data used for defining fROIs were indepen-
dent of data used for estimating response (Kriegeskorte,
Simmons, Bellgowan, & Baker, 2009).
As implemented in GSS, statistical tests were per-
formed on the percent signal change values using stan-
dard Student’s t tests. One-sample t tests were performed
to investigate the response of the MD and ToM network
fROIs to spatial and imitative compatibility effects. Based
on prior behavioral findings, which showed greater RT
interference for female than male individuals during imi-
tation inhibition (Butler et al., 2015), we expected to
observe sex differences in those regions that also show
simple compatibility effects. That is, we expected brain
regions that were generally involved in spatial and/or
imitative control to show sex differences. As such, we only
investigated sex difference in those fROIs that showed
spatial or imitative compatibility effects. To do so, one-
tailed independent samples t tests were performed that
tested for greater engagement for female than male indi-
viduals. False discovery rate (FDR) multiple-comparison
correction ( p < .05) was used to correct for the number
of fROIs in each functional network.
RESULTS
Experiment 1
Behavioral Results
A one-sample t test confirmed a general compatibility
effect (mean = 80.02, SE = 8.19), t(27) = 9.77, p ≤
.001, 95% CI (63.22, 96.82), Cohen’s dz = 1.85. A one-
tailed independent samples t test showed no differences
between male individuals (mean = 70.94, SE = 13.30)
and female individuals (mean = 89.10, SE = 9.43), t(26) =
1.114, p = .138, 95% mean difference = 18.16, CI (−9.64),
Cohen’s d = 0.42. All participants had >80% accuracy;
hence, all were included in the analysis. Trials on which
participants made an incorrect response (0.95%) did not
make a response or responded after 2000 msec (0.52%)
or pressed an invalid key or responded too fast (0.09%)
were excluded from the analyses.
fMRI Results
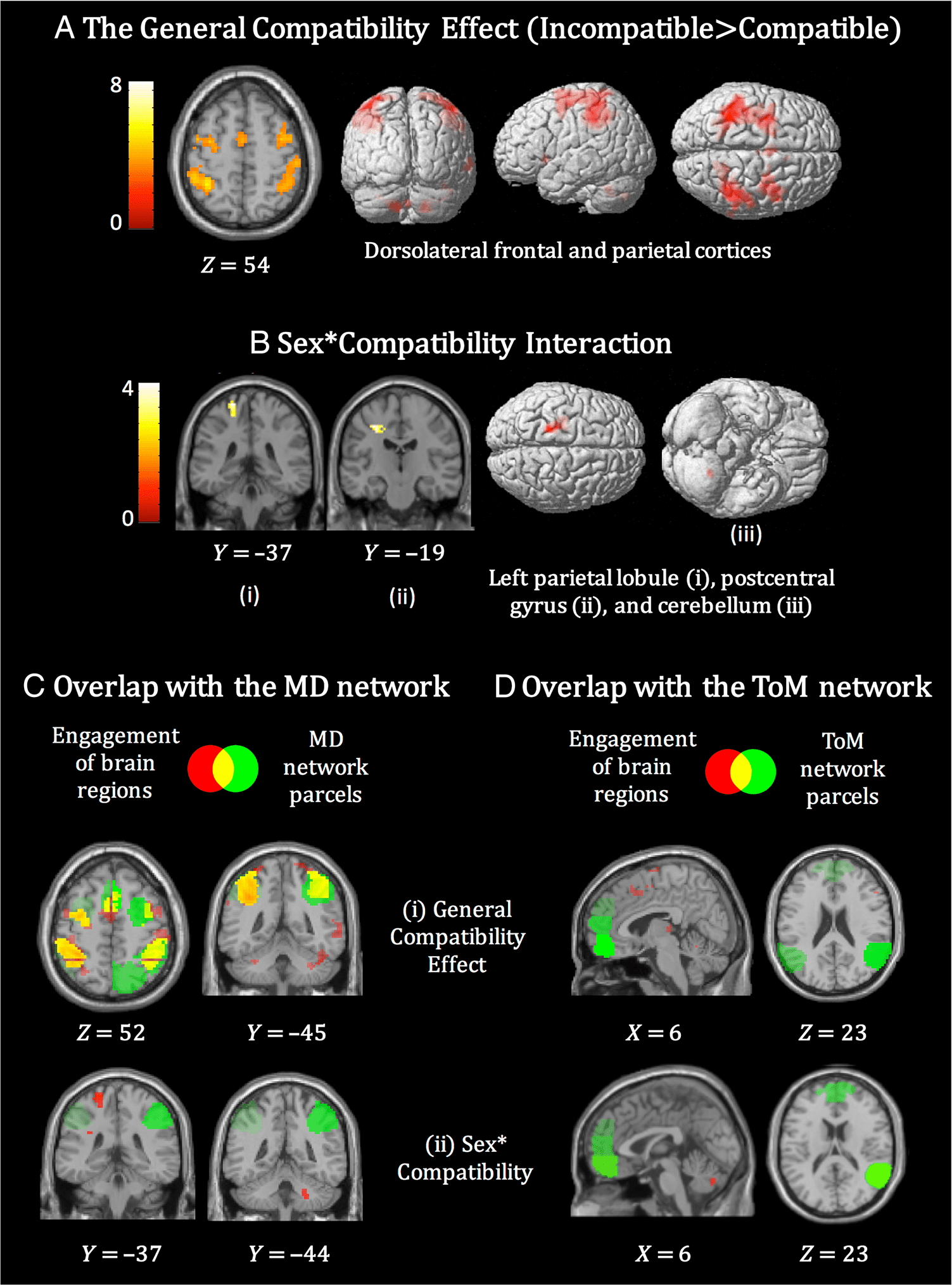
In a whole-brain analysis, compatibility effects (general
incompatible > general compatible) were observed in
dorsomedial frontal cortex and bilaterally in dorsolateral
frontal and parietal cortices (Figure 4A; Table 2A). A small
volume correction (SVC) using MD and ToM network
parcels was performed to restrict the search area to
ToM and MD networks. Using the MD network SVC, re-
sults showed widespread activation of frontal and parietal
regions, which survived correction for multiple compari-
sons (Figure 4A, Ci). In contrast, using the ToM network
SVC, no clusters survived correction for multiple compar-
ison, and only rTPJ showed a compatibility effect at more
lenient threshold ( p < .001, uncorrected; see Supple-
mentary Tables S2.1 and S2.2). Anterior mPFC did not
show the general compatibility effect even at this more
lenient threshold.
The Sex × Compatibility interaction revealed clusters
in left SPL extending into postcentral gyrus and a further
cluster in the cerebellum (Figure 4B; Table 2B). No clus-
ters emerged following an SVC analysis using the MD and
ToM network masks, which demonstrate that the clusters
emerging from the Sex × Compatibility interaction do not
overlap with the MD or ToM networks (see Supplementary
Tables S2.1 and S2.2; Figure 4Cii and Dii).
Power Analysis
We set up Experiment 1 to estimate the appropriate
sample size for our critical experiment (Experiment 2).
To this end, a power analysis was performed using the
fMRIpower software package (fMRIpower.org; Mumford
& Nichols, 2008). We performed the power analysis as
follows: First, a whole-brain map of the imitation task
general compatibility effect (incompatible > compati-
ble) from Experiment 1 was entered into fMRIpower.
Next, two ROIs were identified: the MD network
(Duncan, 2010) and the ToM network (Saxe & Kanwisher,
Darda, Butler, and Ramsey
923
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
0
6
9
1
4
1
7
8
7
5
2
9
/
j
o
c
n
_
a
_
0
1
2
6
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. General compatibility
effect and Sex × Compatibility
interaction in the imitation
inhibition task (Experiment 1).
(A) Results for the general
compatibility effect
(incompatible > compatible).
Clusters emerged in the
dorsal frontoparietal cortices.
(B) Results for the Sex ×
Compatibility interaction
(defined as [female
(compatibility effect) > male
(compatibility effect)]. Clusters
emerged in the left superior
parietal cortex extending into
the postcentral gyrus. The MD
network parcels were overlapped
with the general compatibility
effect (C(i)) and the Sex ×
Compatibility interaction
(C(ii)). An overlap was found
between MD parcels and
regions engaged by the general
compatibility effect. The ToM
network parcels were also
overlapped with the general
compatibility effect (D(i)) and
Sex × Compatibility interaction
(D(ii)). No overlapping regions
were found for the ToM
network and regions engaged
for the general compatibility
effect or for the Sex ×
Compatibility interaction. Voxel-
wise threshold used for all
images was p < .001, k = 10.
For a complete set of results,
see Table 2 and Supplementary
Tables S2.1 and S2.2.
2003). The MD and ToM network masks used were the
same as in Experiment 2 (see Methods). As recommended,
we corrected the alpha value by the number of ROIs (0.05/
2 = 0.025) before performing power analyses (Mumford,
2012).
Results from these power analyses showed that testing
50 participants in Experiment 2 would provide 80% power
to detect effects as large as (or larger than) the average
effect size that was observed across all nodes in the MD
network in Experiment 1 (Cohen’s d = 0.4, mean signal
change = 0.23, SD = 0.58). We did not have the same
level of power to detect smaller effects than these, such
as those observed in the ToM network in Experiment 1.
Indeed, the effects in the ToM network in Experiment 1
were so small that we would have needed an impractically
large sample size to achieve 80% power. As such, in
Experiment 2, we decided to test participants until we
had 50 usable data sets.
Design differences between Experiments 1 and 2 are
worth considering when interpreting these power calcu-
lations because we may be underestimating the power of
our design in Experiment 2. The toolbox used to run
power calculations (fmripower.org) can only estimate
power for a future experiment with the same design as
the current data set (Mumford & Nichols, 2008). How-
ever, the designs of Experiments 1 and 2 differed in
924
Journal of Cognitive Neuroscience
Volume 30, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
6
9
1
4
1
7
8
7
5
2
9
/
j
o
c
n
_
a
_
0
1
2
6
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Table 3. Responses in Each MD Network fROI for Spatial and Imitative Compatibility
ROI
L_SPL
L_IPS
L_IPL
L_MFG
L_PrecG
L_IFG
L_Insula
L_SMA
R_SPL
R_IPS
R_IPL
R_MFG
R_PrecG
R_IFG
R_Insula
R_SMA
ROI Size
1173
287
641
536
338
181
197
294
1181
227
599
535
269
265
184
328
Intersubject
Overlap
Average ROI
Mask Size
( Voxels)
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
117
28
64
53
33
18
19
29
118
22
59
53
26
26
18
32
Spatial Compatibility
Imitative Compatibility
t
2.00
2.00
2.72
2.16
2.17
1.83
2.78
2.52
2.30
2.03
2.65
3.57
2.43
2.61
2.09
2.30
p
.026
.026
.005
.018
.018
.040
.004
.008
.013
.024
.005
<.001
.009
.006
.021
.022
p-FDR
.028
.028
.019
.028
.028
.037
.019
.020
.026
.028
.019
.006
.021
.019
.028
.028
t
1.13
1.96
2.05
0.53
0.91
1.53
0.52
0.39
1.56
2.30
2.50
1.55
1.59
2.53
1.24
1.06
p
.131
.028
.023
.301
.184
.066
.304
.349
.062
.013
.008
.064
.060
.007
.120
.148
p-FDR
.191
.089
.089
.324
.227
.118
.324
.349
.118
.069
.063
.118
.118
.063
.175
.198
For each individual, for the MD network mask, the hard > easy contrast was used, and the top 10% of voxels (based on t values) within each parcel
were defined as that individual’s fROI. Uncorrected p values as well as FDR-corrected p values are reported. Cells in bold are fROIs that survive
correction for multiple comparisons ( p < .05, FDR-corrected).
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
6
9
1
4
1
7
8
7
5
2
9
two ways. First, Experiment 1 measured a general com-
patibility effect, whereas in Experiment 2, we broke this
effect down into spatial and imitative compatibility ef-
fects. Second, Experiment 2 had more than double the
amount of trials per condition as Experiment 1. There-
fore, the primary contrast used to determine power
was not identical to the contrast used in Experiment 2,
but due to a greater number of trials per condition to
estimate the effects of interest, we may underestimate
power in Experiment 2. Given the lack of sex differences
in Experiment 1 in our ROIs, we did not have sufficient
power to convincingly investigate neural differences
between male and female individuals in Experiment 2.
However, given our a priori predictions regarding sex,
we continue to report sex difference analyses through-
out the article.
Experiment 2
Behavioral Results
The hand stimulus used in Experiment 2 for the imitation
inhibition task was perceived as “neutral” by most partic-
ipants (meanrating = 5.20, SDrating = 2.04; rated on a scale
of 1–9, where 1 = most masculine, 5 = neutral, and 9 =
most feminine). To ensure participants were engaging
/
j
o
c
n
_
a
_
0
1
2
6
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
correctly with the task, runs on which participants had less
than 80% accuracy (two runs of one participant) were re-
moved. In addition, RTs more than 3 SDs away from the
mean (two runs of one participant and one run of another
participant) were excluded from the analyses. Further-
more, trials on which participants made an incorrect re-
sponse (1.52%), did not make a response or responded
after 2000 msec (0.61%), or pressed an invalid key
(0.22%) were also excluded from the analyses. Figure 5
shows the imitative and spatial compatibility effects for
both the sexes. For RT data, see Supplementary Table S3.
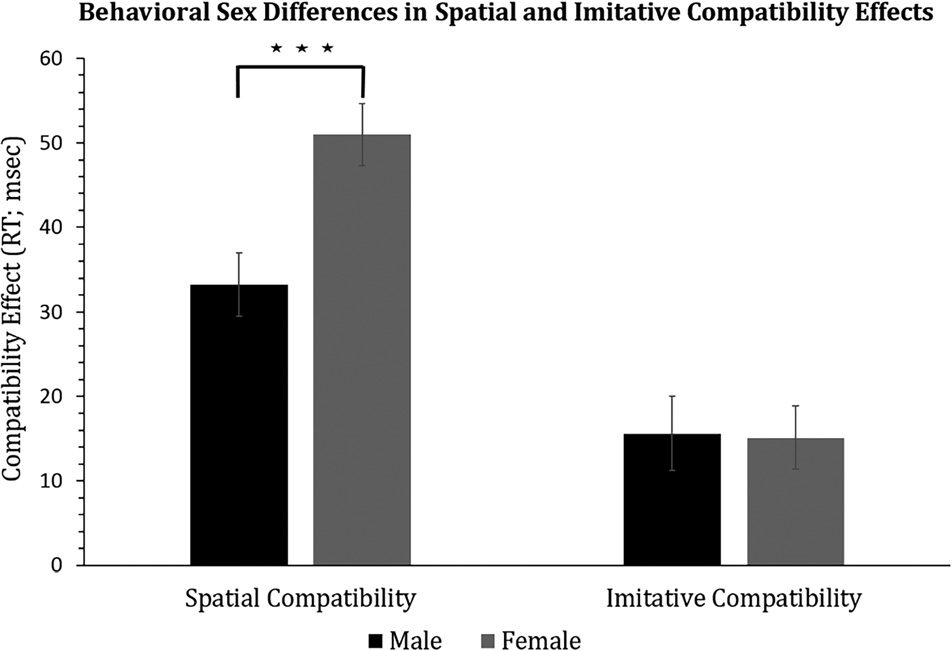
Spatial compatibility. A one-sample t test confirmed a
spatial compatibility effect (mean = 41.94, SE = 2.87),
t(54) = 14.618, p ≤ .001, 95% CI (36.19, 47.69), Cohen’s
dz = 1.97. A one-tailed independent samples t test
evidenced a greater spatial interference effect for female
individuals (mean = 50.98, SE = 3.67) as compared with
male individuals (mean = 33.20, SE = 3.75), t(53) =
−3.38, p < .001, mean difference = 17.76, 95% CI (8.91);
Cohen’s d = 0.91.
Imitative compatibility. A one-sample t test showed a
significant imitative compatibility effect (mean = 15.37,
SE = 2.86), t(54) = 5.37, p < .001, 95% CI (9.63, 21.11),
Cohen’s dz = 0.72. There was no significant difference
Darda, Butler, and Ramsey
925
and one participant’s data for one session of the imitation
task could not be used because the data file was corrupted.
Localizer Tasks
All fROIs showed the predicted responses to the localizer
contrasts (as estimated using data not used for defining
ROIs; see Methods). All the MD network fROIs showed a
robust hard > easy effect (ts > 9.13, ps < .0001), and
ToM network fROIs showed a robust belief > photo ef-
fect (ts > 5.70, ps < .0001). For responses for each indi-
vidual fROI separately, see Supplementary Tables S4.1
(MD) and S4.2 (ToM).
The Automatic Imitation Task
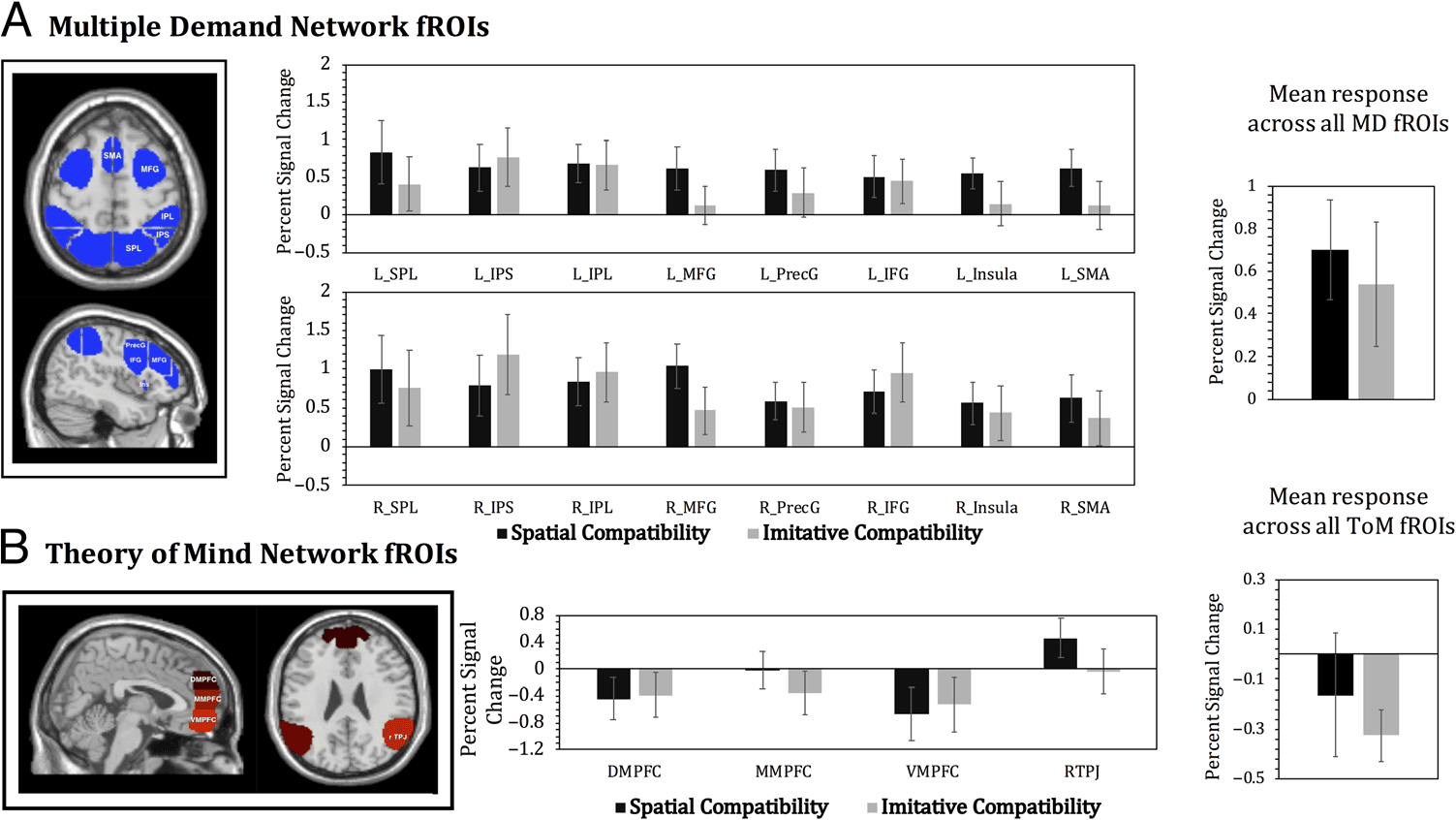
GSS analyses. Figure 6 shows the mean percent signal
change for each fROI in the MD and ToM networks for
spatial (spatial incompatible > spatial compatible) and
imitative compatibility (imitative incompatible > imita-
tive compatible) effects.
MD network fROIs
Spatial compatibility. All 16 fROIs of the MD network
showed a spatial compatibility effect (ts > 1.8, ps < .04;
Figure 6A, Table 3), which survived correction for multi-
ple comparisons ( p < .05, FDR-corrected). The mean
percent signal change across the MD network for spatial
compatibility was 0.70, SD = 1.66, Cohen’s d = 0.42. No
significant differences were found between male and
female individuals in percent signal change values in any
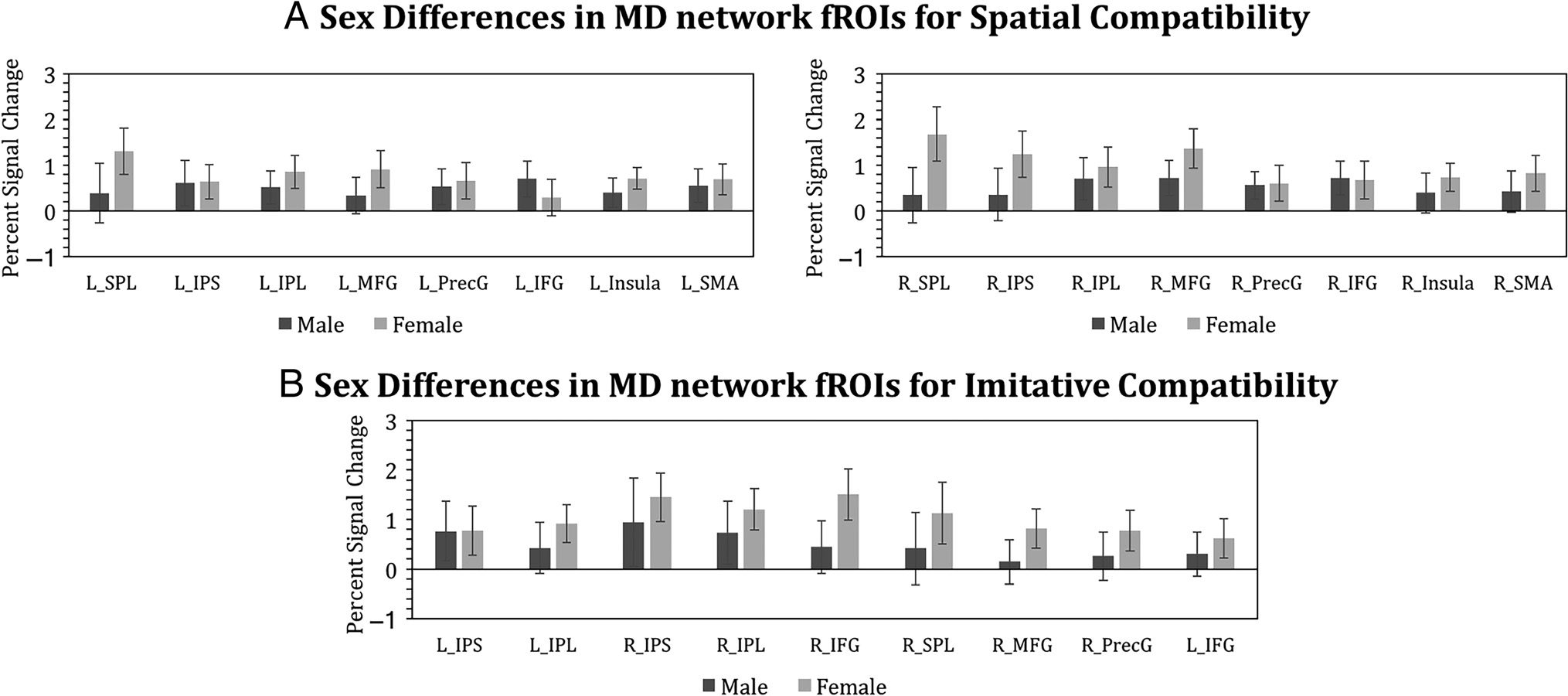
of the fROIs (ts < 1.6, ps > .1), except right SPL which
approached significance ( p = .062; Figure 7A).
Imitative compatibility. None of the 16 MD network
fROIs showed an imitative compatibility effect, which
Figure 5. Behavioral sex differences in imitative and spatial compatibility
effects. The spatial and imitative compatibility effects (RTs) in male
and female individuals displayed in milliseconds. Error bars denote
standard error of mean.
between male individuals (mean = 15.62, SE = 4.39) and
female individuals (mean = 15.11, SE = 3.73), t(53) = 0.09,
p = .465, mean difference = −0.51, 95% CI (−10.18),
Cohen’s d = 0.02.
fMRI Results
Five participants were excluded from the fMRI analyses
due to lower than 80% accuracy in two runs of the imita-
tion task and the MD network localizer task (n = 1) and
excessive head motion (n = 4; displacement > 4 mm) in all
runs of the imitation task and/or all runs of either of the
localizer tasks. Thus, the final sample consisted of 50 partic-
ipants (Mage = 22.26, SDage = 3.71; 24 female). From these
50 participants, two sessions of the imitation task were also
excluded for one participant due to excessive head motion
Figure 6. The parcels used to
define individual fROIs and the
responses to spatial and
imitative compatibility effects in
the MD (A) and ToM (B) network
fROIs are shown. Error bars
denote standard error of mean.
All MD network fROIs were
sensitive to spatial compatibility
effects (FDR-corrected, p < .05).
Bilateral IPL, bilateral IPS, and the
right IFG showed a significant
response for imitative
compatibility effects, but at an
uncorrected threshold of p <
.001. No ToM network fROIs
showed engagement for either
spatial or imitative compatibility
effects.
926
Journal of Cognitive Neuroscience
Volume 30, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
6
9
1
4
1
7
8
7
5
2
9
/
j
o
c
n
_
a
_
0
1
2
6
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
6
9
1
4
1
7
8
7
5
2
9
/
j
o
c
n
_
a
_
0
1
2
6
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 7. Responses to spatial (A) and imitative compatibility (B) effects separately for male and female participants in the MD network. Error bars
denote standard error of mean. None of the fROIs showed a sex difference either in imitative or spatial compatibility that survived correction for
multiple comparisons.
survived correction for multiple comparisons (all ps > .05,
FDR-corrected). Five MD network fROIs showed an imita-
tive compatibility effect at an uncorrected threshold (ts >
1.95, ps < .05). These fROIs include bilateral IPL, bilateral
IPS, and the right IFG (Figure 6A, Table 3). Four further
fROIs showed an imitative compatibility effect that ap-
proached significance, which included left IFG ( p = .07),
right SPL, right MFG, and right PrecG ( p = .06). The mean
percent signal change across the MD network for imitative
compatibility was 0.54, SD = 2.06, Cohen’s d = 0.26. There
was no significant difference between male and female in-
dividuals in any of these fROIs (ts < 1.5, ps > .08; see
Figure 7B).
ToM network fROIs. None of the ToM network fROIs
showed imitative (ts < 1.3, ps > .50) or spatial (ts <
1.6, ps > .06) compatibility effects, even at an uncor-
rected significance threshold (Figure 6B, Table 4). rTPJ
showed a spatial compatibility effect that approached
significance ( p = .065). The mean percent signal change
across the ToM network for spatial compatibility was
−0.16, SD = 1.88, Cohen’s d = −0.08, and the mean
percent signal change across the ToM network for imitative
compatibility was −0.32, SD = 2.02, Cohen’s d = −0.16.
Whole-brain analyses. For completeness and for use in
future meta-analyses, we also computed group-level whole-
brain analyses separately for general, spatial, and imitative
compatibility effects, as well as for Sex × Compatibility
interactions (see Supplementary Table S4).
Open Science
Data for Experiments 1 and 2 are freely available online
including behavioral and fROI data (osf.io/45x6z), as well
as whole-brain t maps (https://neurovault.org/collections/
3218).
Table 4. Responses in Each ToM Network fROI for Spatial and Imitative Compatibility
ROI
ROI Size
DMPFC
MMPFC
VMPFC
rTPJ
576
494
382
1018
Intersubject
Overlap
Average ROI
Mask Size
( Voxels)
1
1
1
1
57
49
38
101
Spatial Compatibility
Imitative Compatibility
t
−1.38
−0.043
−1.690
1.543
p
.913
.517
.951
.065
p-FDR
t
.951
.951
.951
.258
−1.167
−1.081
−1.286
−0.106
p
.876
.857
.898
.542
p-FDR
.898
.898
.898
.898
For each individual, for the ToM network mask, the Belief > Photo contrast was used, and the top 10% of voxels (based on t values) within each
parcel were defined as that individual’s fROI. Uncorrected p values as well as FDR-corrected p values are reported.
Darda, Butler, and Ramsey
927
DISCUSSION
The current study provides the most robust neuro-
imaging evidence to date for a lack of functional specific-
ity in the neural circuits supporting the inhibition of
automatic imitation. With higher statistical power and
functional sensitivity than prior studies, across two exper-
iments the results demonstrate that imitation inhibition
engages a domain-general neural network as opposed
to a brain network that supports social cognition. As
such, models of imitation control need updating to in-
clude an increased role for domain-general processes
and a reduced or altered role for domain-specific pro-
cesses. Furthermore, in terms of behavior, female individ-
uals showed a higher spatial but not imitative compatibility
effect than male individuals. However, there was no sex
difference in the neural mechanisms underlying spatial or
imitation control, which suggests that further exploration
of sex differences in inhibitory control is required.
Functional Specificity in Imitation Inhibition
Our findings show that brain regions that are engaged in
a verbal WM task, which are associated with the operation
of the MD network (Fedorenko et al., 2013; Duncan, 2010),
are also engaged during spatial and imitative conflict reso-
lution. These results support the involvement of a domain-
general cognitive and neural system during the control of
imitation. By contrast, brain regions that are engaged in a
belief reasoning task, which are associated with the opera-
tion of the ToM network ( Van Overwalle, 2009; Saxe &
Kanwisher, 2003; Frith & Frith, 1999), show no engage-
ment during the inhibition of imitation. As such, we pro-
vide no evidence for domain specificity in cognitive and
neural systems that control imitation.
Brass and colleagues (2009) proposed that, in the con-
text of imitation control, rTPJ is involved in self–other dis-
tinction, and mPFC enforces the self-generated action
over the observed action. Our findings are inconsistent
with the hypothesis that a specific neural system related
to social cognition is engaged in the inhibition of auto-
matic imitative tendencies. mPFC and rTPJ have both
been implicated in imitation inhibition by some studies
( Wang et al., 2011; Brass et al., 2009; Spengler et al.,
2009; Brass, Derrfuss, & von Cramon, 2005). In contrast,
other studies found engagement of mPFC only (Cross
et al., 2013; Brass et al., 2001) or of domain-general re-
gions rather than mPFC and rTPJ (Marsh et al., 2016;
Cross & Iacoboni, 2013; Crescentini et al., 2011; Bien
et al., 2009). In both experiments in the current study,
we had larger sample sizes than prior experiments, and
in Experiment 2, we had sufficient statistical power to
be confident in detecting effects as large as previously
observed in mPFC and rTPJ, should they exist. Taken
together with prior findings (Table 1), we suggest that,
during the inhibition of imitation, the consistency of mPFC
and rTPJ engagement across individuals is relatively low,
whereas the consistency of MD network engagement
across individuals is relatively high.
These results have potential implications for self–other
control theories of social cognition more generally. Mostly
based on imitation research, which previously suggested
that mPFC and rTPJ are engaged in imitation inhibition,
self–other control is thought to be a candidate mechanism
for a diverse set of social functions (de Guzman, Bird,
Banissy, & Catmur, 2016; Sowden & Shah, 2014; Brass
et al., 2009). For example, self–other control processes
have been linked to autism, empathy, and theory of mind
(de Guzman et al., 2016; Sowden & Shah, 2014; Spengler
et al., 2009). However, recent behavioral findings, which
used larger sample sizes than prior work and meta-analytical
approaches, do not support the view that the control of
imitation varies as a function of social disposition as in-
dexed by autistic-like traits and empathy (Cracco et al.,
2018; Genschow et al., 2017; Butler et al., 2015). In light
of these recent behavioral results, the lack of engagement
of mPFC and rTPJ in the current study raises an important
question about the reliance of imitation inhibition on a
self–other distinction. One possibility is that, instead of a
distinctly social mechanism (Bertenthal & Scheutz, 2013;
Boyer et al., 2012), inhibiting imitative tendencies may
involve the same cognitive processes that are used when
inhibiting other nonsocial external influences (Cooper
et al., 2012; Heyes, 2011).
Alternatively, the engagement of mPFC and rTPJ during
self–other control processes may be more complicated
than current models of social cognition suggest. Indeed,
a small number of neurostimulation studies have shown
that modulation to rTPJ can influence performance on
RT measures of imitation (Sowden & Catmur, 2015;
Hogeveen et al., 2014). In addition, mPFC and rTPJ have
been found to be involved in the modulation of auto-
matic imitation. For example, Klapper et al. (2014)
found a higher response in rTPJ when an interaction
partner looked human and was believed to be human
compared with when neither of these animacy cues
was present. Wang and colleagues (2011) demonstrated
that mPFC had a top–down influence on other brain cir-
cuits during social modulation of imitation via direct
gaze. These studies suggest that mPFC and/or rTPJ
may have a regulatory role, be sensitive to social con-
text, and be functionally connected to other regions
during the inhibition of automatic imitation. Indeed,
regions that do not show direct engagement in a cog-
nitive process of interest have been known to have a
regulatory influence on other regions that are directly
engaged (Burnett & Blakemore, 2009). In line with this
proposal, Cross and colleagues (2013) suggested that
imitation control involves top–down regulation between
a domain-general cognitive control network and a domain-
specific network relevant for imitation. More generally,
research from other domains of social cognition shows
growing evidence for higher complexity and functional
interplay within and between so-called domain-specific
928
Journal of Cognitive Neuroscience
Volume 30, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
0
6
9
1
4
1
7
8
7
5
2
9
/
j
o
c
n
_
a
_
0
1
2
6
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
and domain-general networks (Spunt & Adolphs, 2015;
Baetens, Ma, Steen, & Van Overwalle, 2014; Quadflieg
et al., 2011; Zaki, Hennigan, Weber, & Ochsner, 2010).
These studies suggest that models including neat divisions
between these networks may be an overly simplistic char-
acterization of mental function (Michael & D’Ausilio, 2015;
Barrett, 2012). Much like social cognition in general, there-
fore, imitation control may be best explained by interactions
between component functional circuits, which themselves
need not be domain-specific (Spunt & Adolphs, 2017). A
crucial direction for future research is testing for more
complex models of imitation, which may involve connec-
tivity in and between regions of the MD and ToM networks.
An important point to note, however, is that any con-
clusions made regarding possible domain specificity of
mPFC and rTPJ are based on the assumption that mPFC
and rTPJ are at least partly specialized for social cognition
(Brass et al., 2009). Recent evidence suggests that mPFC
and rTPJ may be functionally versatile in the sense that
they show general cognitive properties, which may not be
specific to social cognition (Dugué, Merriam, Heeger, &
Carrasco, 2017; Schurz, Tholen, Perner, Mars, Sallet, 2017;
Schuwerk, Schurz, Müller, Rupprecht, & Sommer, 2017;
de la Vega, Chang, Banich, Wager, & Yarkoni, 2016; Carter
& Huettel, 2013; Alexander & Brown, 2011; Yarkoni,
Poldrack, Nichols, Van Essen, & Wager, 2011). Thus, the
argument that the engagement of mPFC and rTPJ in
imitation inhibition may be specific to social cognition
might need further validation. In addition, social cognition
itself has been broken down in “bottom–up” and “top–
down” domains (Zaki & Ochsner, 2012). The bottom–up
domain refers to prereflective processes that are fast and
stimulus driven, whereas the top–down domain maps on
reflective, cognitively laborious, and flexible processes
(Bohl & van den Bos, 2012). When extended to imitation
control, prior research has consistently implicated regions
involved in top–down control for automatic imitation (Brass
et al., 2009). However, recent studies suggest that imitation
control (and social cognition more broadly) relies on inter-
actions between bottom–up and top–down processes
(Christov-Moore, Conway, & Iacoboni, 2017; Cross &
Iacoboni, 2014; Bohl & van den Bos, 2012). Thus, another
important avenue for future research would be to investi-
gate imitation control based on bottom–up and top–down
processes and their interactions, rather than considering
these processes as mutually exclusive.
Nonetheless, results from the current study remain
clear: The basic imitation inhibition mechanism engages
the MD network, which has been consistently associated
with domain-general processes (Duncan, 2010). Given
the mixed findings in prior imitation studies (Table 1)
as well as in psychology and neuroscience more generally
(Open Science Framework, 2015; Button et al., 2013),
future fMRI research may also consider reliability and
reproducibility as key concerns in imitation research and
consider the possible use of fROI approaches as a means
to quantify consistency across individuals.
Sex Differences in Imitation Inhibition
This study is the first to investigate sex differences in the
neural mechanisms that inhibit imitation. The behavioral
data demonstrated that female individuals show a greater
spatial but not imitative compatibility effect than male in-
dividuals. This result extends prior behavioral research
on sex differences, which did not separate spatial (or
orthogonal spatial) from imitative responses in imitation
control (Genschow et al., 2017; Butler et al., 2015). The
result is also consistent with reports in a wide range of non-
social inhibitory control tasks, which show similar sex dif-
ferences (Figure 2; Stoet, 2010, 2017; Clayson et al., 2011;
Rubia et al., 2010; Bayliss et al., 2005). All these tasks share
a common feature—they require the inhibition of a re-
sponse to a task-irrelevant spatial feature to enforce a task-
relevant response. Taken together, this pattern of results
suggests that response inhibition relating to spatial conflict
differs between the sexes, rather than a process that is tied
to the control of imitation. An alternative possibility is that
the difference between the sexes for spatial compatibility
is larger than for imitative compatibility, and we were
unable to detect the imitative effect behaviorally. Future
research will have to probe these possibilities further.
Given the proposed role of MD and ToM networks in
imitation control, we anticipated sex differences in one or
both of these networks. The neuroimaging data, how-
ever, demonstrated no sex differences in the ToM or
MD networks in either experiment. Furthermore, even
though regions outside our ROIs mediated the sex dif-
ference in Experiment 1, these regions were not consis-
tently engaged differently for male and female individuals
in Experiment 2. Thus, based on data across both exper-
iments, our best estimate is that univariate analyses,
which assess the magnitude of BOLD response, do not
show large effects of sex in MD or ToM neural networks.
This being said, there does seem to be a trend for greater
engagement in the MD network for female individuals
compared with male individuals for both spatial and
imitative effects, but this does not survive our statistical
thresholding (Figure 7). As a consequence, we are cau-
tious to interpret this null result as we did not have the
same level of statistical power to detect sex differences
as we did to detect simple compatibility effects. Indeed,
it remains a possibility that small univariate effects exist
or that the sex difference is underpinned by more complex
neural organization. Future studies that use connectivity
measures (Sporns, Tononi, & Kötter, 2005) or multivoxel
pattern analysis (Kriegeskorte, Mur, & Bandettini, 2008;
Norman, Polyn, Detre, & Haxby, 2006) may show increased
sensitivity and be better able to capture the complexity of
neural organization that we are aiming to measure.
Limitations
The primary limitation of the current work is that we
studied a relatively simple model of brain organization
Darda, Butler, and Ramsey
929
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
0
6
9
1
4
1
7
8
7
5
2
9
/
j
o
c
n
_
a
_
0
1
2
6
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
based on univariate measures. Given the mixed evi-
dence from prior studies regarding imitation control
(Table 1), we felt it was an important step to first estab-
lish the extent to which general and specific systems
were engaged in a univariate manner. By doing so, we
aimed to build an appropriate foundation for future
work to build upon. Moreover, as we only identified
the MD and ToM networks, it is possible that neural re-
gions outside our key networks may play a role in imi-
tation inhibition or mediate the sex difference in spatial
response inhibition or imitation control. Even though
our whole-brain analyses showed no consistent effects
outside our fROIs, this only shows that there was no
univariate engagement of extended brain regions. We
thus acknowledge that we have tested a relatively sim-
ple model of brain organization that is likely to under-
estimate the complexity of neural processes associated
with social and cognitive mechanisms such as imitation
control. As mentioned before, future work may con-
sider interactions between general and specific sys-
tems and more complex, multivariate measures of
brain organization.
A second limitation regards the functional localization
approach used to identify the ToM and MD networks in
Experiment 2. The validity of the fROI approach is
based on assumptions about the functional processes
that are engaged by the localizers used to identify fROIs.
For example, different ToM localizers may engage partly
nonoverlapping aspects of the ToM network (Schaafsma,
Pfaff, Spunt, & Adolphs, 2015; Spunt & Adolphs, 2014).
Therefore, our conclusions about the role of ToM and
MD networks are limited to the type of localizer para-
digms that we used in the current study. Future research
that uses different functional partitions of these networks
would be instructive.
A final potential limitation is that the order of tasks in
Experiment 2 could have influenced our results. We
ordered the tasks such that the ToM localizer was always
performed at the end, but the MD task was interspersed
between imitations runs in order to offset boredom. We
arranged blocks in this manner because we were pri-
marily concerned that asking people to perform a belief
reasoning task would introduce a social bias to treat the
person (hand image) in an artificially more social/belief
reasoning manner during the imitation inhibition task.
We did not share the same level of concern that per-
forming a memory task, which we used to localize the
domain-general system, would introduce a memory or
“cognitive control” bias to the imitation inhibition task.
However, we cannot rule out the possibility in the
current experiment that the MD task influenced the
way the imitation task was performed. This being said,
we did get the same results in Experiment 1, when the
MD task was not performed before the imitation task.
As such, although possible, we find it unlikely that
task order had a meaningful impact on our results in
Experiment 2.
Acknowledgments
This work was funded by a grant from the Economic and Social
Research Council (Grant ES/ K001884/1 to R. R.). We thank
Ruud Hortensius for comments on a previous version of the
manuscript.
Reprint requests should be sent to Richard Ramsey, Depart-
ment of Psychology, Bangor University College of Health and
Behavioural Sciences, Adeilad Brigantia, Penrallt Road, Bangor,
Gwynedd, LL57 2AS, United Kingdom, or via e-mail: r.ramsey@
bangor.ac.uk.
REFERENCES
Alexander, W. H., & Brown, J. W. (2011). Medial prefrontal
cortex as an action-outcome predictor. Nature Neuroscience,
14, 1338–1344.
Amodio, D. M., & Frith, C. D. (2006). Meeting of minds: The
medial frontal cortex and social cognition. Nature Reviews
Neuroscience, 7, 268–277.
Aron, A. R., Robbins, T. W., & Poldrack, R. A. (2014). Inhibition
and the right inferior frontal cortex: One decade on. Trends
in Cognitive Sciences, 18, 177–185.
Baetens, K., Ma, N., Steen, J., & Van Overwalle, F. (2014).
Involvement of the mentalizing network in social and non-
social high construal. Social Cognitive Affective
Neuroscience, 9, 817–824.
Bardi, L., Gheza, D., & Brass, M. (2017). TPJ-M1 interaction
in the control of shared representations: New insights
from tDCS and TMS combined. Neuroimage, 146,
734–740.
Baron-Cohen, S. (2002). The extreme male brain theory of
autism. Trends in Cognitive Sciences, 6, 248–254.
Baron-Cohen, S., & Wheelwright, S. (2004). The empathy quotient:
An investigation of adults with Asperger syndrome or high
functioning autism, and normal sex differences. Journal of
Autism and Developmental Disorders, 34, 163–175.
Barrett, H. C. (2012). A hierarchical model of the evolution of
human brain specializations. Proceedings of the National
Academy of Sciences, U.S.A., 109, 10733–10740.
Bayliss, A. P., di Pellegrino, G., & Tipper, S. P. (2005). Sex
differences in eye gaze and symbolic cueing of attention.
Quarterly Journal of Experimental Psychology, 58A,
631–650.
Bertenthal, B. I., Longo, M. R., & Kosobud, A. (2006). Imitative
response tendencies following observation of intransitive
actions. Journal of Experimental Psychology: Human
Perception and Performance, 32, 210.
Bertenthal, B. I., & Scheutz, M. (2013). In praise of a model but
not its conclusions: Commentary on Cooper et al. (2012).
Cognitive Science, 37, 631–641.
Bien, N., Roebroeck, A., Goebel, R., & Sack, A. T. (2009).
The brain’s intention to imitate: The neurobiology of
intentional versus automatic imitation. Cerebral Cortex,
19, 2338–2351.
Blank, I., Kanwisher, N., & Fedorenko, E. (2014). A functional
dissociation between language and multiple-demand systems
revealed in patterns of BOLD signal fluctuations. Journal
of Neurophysiology, 112, 1105–1118.
Bohl, V., & van den Bos, W. (2012). Toward an integrative
account of social cognition: Marrying theory of mind and
interactionism to study the interplay of Type 1 and Type 2
processes. Frontiers in Human Neuroscience, 6, 274.
Boyer, T. W., Longo, M. R., & Bertenthal, B. I. (2012). Is
automatic imitation a specialized form of stimulus–response
compatibility? Dissociating imitative and spatial compatibilities.
Acta Psychologica, 139, 440–448.
930
Journal of Cognitive Neuroscience
Volume 30, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
0
6
9
1
4
1
7
8
7
5
2
9
/
j
o
c
n
_
a
_
0
1
2
6
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Brass, M., Bekkering, H., Wohlschlager, A., & Prinz, W. (2000).
Cracco, E., Bardi, L., Desmet, C., Rigoni, D., Radkova, I., Deschrijver,
Compatibility between observed and executed finger
movements: Comparing symbolic, spatial, and imitative cues.
Brain and Cognition, 44, 124–143.
Brass, M., Derrfuss, J., Matthes-von Cramon, G., & von Cramon,
D. Y. (2003). Imitative response tendencies in patients with
frontal brain lesions. Neuropsychology, 17, 265–271.
Brass, M., Derrfuss, J., & von Cramon, D. Y. (2005). The inhibition
of imitative and overlearned responses: A functional double
dissociation. Neuropsychologia, 43, 89–98.
Brass, M., & Heyes, C. (2005). Imitation: Is cognitive neuroscience
solving the correspondence problem? Trends in Cognitive
Sciences, 9, 489–495.
Brass, M., Ruby, P., & Spengler, S. (2009). Inhibition of imitative
behaviour and social cognition. Philosophical Transactions
of the Royal Society of London, Series B, Biological Sciences,
364, 2359–2367.
Brass, M., Zysset, S., & von Cramon, D. Y. (2001). The
inhibition of imitative response tendencies. Neuroimage, 14,
1416–1423.
Bunge, S. A., Hazeltine, E., Scanlon, M. D., Rosen, A. C., &
Gabrieli, J. D. E. (2002). Dissociable contributions of prefrontal
and parietal cortices to response selection. Neuroimage, 17,
1562–1571.
Burnett, S., & Blakemore, S. J. (2009). Functional
connectivity during a social emotion task in adolescents
and in adults. European Journal of Neuroscience, 29,
1294–1301.
Butler, E. E., Ward, R., & Ramsey, R. (2015). Investigating the
relationship between stable personality characteristics and
automatic imitation. PloS One, 10, e0129651.
Button, K. S., Ioannidis, J. P., Mokrysz, C., Nosek, B. A., Flint, J.,
Robinson, E. S., et al. (2013). Power failure: Why small sample
size undermines the reliability of neuroscience. Nature
Reviews Neuroscience, 14, 365–376.
Campbell, R., Elgar, K., Kuntsi, J., Akers, R., Terstegge, J.,
Coleman, M., et al. (2002). The classification of “fear” from
faces is associated with face recognition skill in females.
Neuropsychologia, 40, 575–584.
Carter, R. M., & Huettel, S. A. (2013). A nexus model of the
temporal–parietal junction. Trends in Cognitive Sciences, 17,
328–336.
Catmur, C., & Heyes, C. (2011). Time course analyses confirm
independence of imitative and spatial compatibility. Journal
of Experimental Psychology. Human Perception and
Performance, 37, 409–421.
Chartrand, T. L., & Bargh, J. A. (1999). The chameleon effect:
The perception-behavior link and social interaction. Journal
of Personality and Social Psychology, 76, 893–910.
Chartrand, T. L., & Lakin, J. L. (2013). The antecedents and
consequences of human behavioural mimicry. Annual Review
of Psychology, 64, 285–308.
Christov-Moore, L., Conway, P., & Iacoboni, M. (2017). Deontological
dilemma response tendencies and sensorimotor representations
of harm to others. Frontiers in Integrative Neuroscience,
11, 34.
Christov-Moore, L., Simpson, E. A., Coudé, G., Grigaityte, K.,
Iacoboni, M., & Ferrari, P. F. (2014). Empathy: Gender effects
in brain and behavior. Neuroscience & Biobehavioral Reviews,
46, 604–627.
Clayson, P. E., Clawson, A., & Larson, M. J. (2011). Sex
differences in electrophysiological indices of conflict
monitoring. Biological Psychology, 87, 282–289.
Cohen, J. (1992). A power primer. Quantitative Methods in
Psychology, 112, 155–159.
Cooper, R. P., Catmur, C., & Heyes, C. (2012). Are automatic
imitation and spatial compatibility mediated by different
processes? Cognitive Science, 37, 605–630.
E., et al. (2018). Automatic imitation: A meta-analysis.
Psychological Bulletin. doi:10.1037.bul0000143.
Crescentini, C., Mengotti, P., Grecucci, A., & Rumiati, R. I.
(2011). The effect of observed biological and non-biological
movements on action imitation: An fMRI study. Brain
Research, 1420, 80–92.
Cross, K. A., & Iacoboni, M. (2013). Optimised neural coding?
Control mechanisms in large cortical networks implemented
by connectivity changes. Human Brain Mapping, 34,
213–225.
Cross, K. A., & Iacoboni, M. (2014). To imitate or not: Avoiding
imitation involves preparatory inhibition of motor resonance.
Neuroimage, 91, 228–236.
Cross, K. A., Torrisi, S., Losin, E. A. R., & Iacoboni, M. (2013).
Controlling automatic imitative tendencies: Interactions
between mirror neuron and cognitive control systems.
Neuroimage, 83, 493–504.
de Guzman, M., Bird, G., Banissy, M. J., & Catmur, C. (2016).
Self–other control processes in social cognition: From
imitation to empathy. Philosophical Transactions of the
Royal Society of London, Series B, Biological Sciences, 371,
20150079.
de la Vega, A., Chang, L. J., Banich, M. T., Wager, T. D., &
Yarkoni, T. (2016). Large-scale meta-analysis of human medial
frontal cortex reveals tripartite functional organization. Journal
of Neuroscience, 36, 6553–6562.
Dimberg, U., & Lundquist, L. O. (1990). Gender differences in
facial reactions to facial expressions. Biological Psychology,
30, 151–159.
Dodell-Feder, D., Koster-Hale, J., Bedny, M., & Saxe, R. (2011).
fMRI item analysis in a theory of mind task. Neuroimage,
55, 705–712.
Donnellan, M. B., Oswald, F. L., Baird, B. M., & Lucas, R. E.
(2006). The mini-IPIP scales: Tiny-yet-effective measures of
the Big Five factors of personality. Psychological Assessment,
18, 192–203.
Dufour, N., Redcay, E., Young, L., Mavros, P. L., Moran, J. M.,
Triantafyllou, C., et al. (2013). Similar brain activation
during false belief tasks in a large sample of adults with
and without autism. PloS One, 8, e75468.
Dugué, L., Merriam, E. P., Heeger, D. J., & Carrasco, M. (2017).
Specific visual subregions of TPJ mediate reorienting of
spatial attention. Cerebral Cortex, 13, 1–16.
Duncan, J. (2010). The multiple-demand (MD) system of the
primate brain: Mental programs for intelligent behaviour.
Trends in Cognitive Sciences, 14, 172–179.
Eickhoff, S. B., Stephan, K. E., Mohlberg, H., Grefkes, C.,
Fink, G. R., Amunts, K., et al. (2005). A new SPM toolbox
for combining probabilistic cytoarchitectonic maps and
functional imaging data. Neuroimage, 25, 1325–1335.
Eklund, A., Nichols, T. E., & Knutsson, H. (2016). Cluster failure:
Why fMRI inferences for spatial extent have inflated false-positive
rates. Proceedings of the National Academy of Sciences,
U.S.A., 113, 7900–7905.
Erikson, B. A., & Erikson, C. W. (1974). Effects of noise letters
upon the identification of a target letter in a nonsearch task.
Perception and Psychophysics, 16, 143–149.
Fedorenko, E., Behr, M. K., & Kanswisher, N. (2011). Functional
specificity for high-level linguistic processing in the human
brain. Proceedings of the National Academy of Sciences,
U.S.A., 108, 16428–16433.
Fedorenko, E., Duncan, J., & Kanwisher, N. (2013). Broad domain
generality in focal regions of frontal and parietal cortex.
Proceedings of the National Academy of Sciences, U.S.A.,
110, 16616–16621.
Fedorenko, E., Hsieh, P. J., Nieto-Castañón, A., Whitfield-Gabrieli,
S., & Kanwisher, N. (2010). New method for fMRI investigations
Darda, Butler, and Ramsey
931
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
0
6
9
1
4
1
7
8
7
5
2
9
/
j
o
c
n
_
a
_
0
1
2
6
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
of language: Defining ROIs functionally in individual
subjects. Journal of Neurophysiology, 104, 1177–1194.
Friston, K. J., Worsley, K. J., Frackowiak, R. S. J., Mazziotta, J. C.,
& Evans, A. C. (1994). Assessing the significance of focal
activations using their spatial extent. Human Brain Mapping,
1, 210–220.
Frith, C. D., & Frith, U. (1999). Interacting minds—A biological
basis. Science, 286, 1692–1695.
Genschow, O., van Den Bossche, S., Cracco, E., Bardi, L., Rigoni,
D., & Brass, M. (2017). Mimicry and automatic imitation are
not correlated. PLoS One, 12, e0183784.
Goldberg, L. R. (1992). The development of markers for the
Big-Five factor structure. Psychological Assessment, 4, 26–42.
Gowen, E., Bolton, E., & Poliakoff, E. (2016). Believe it or not:
Moving non-biological stimuli believed to have human origin can
be represented as human movement. Cognition, 146, 431–438.
Hazeltine, E., Poldrack, R., & Gabrieli, J. D. E. (2007). Neural
activation during response competition. Journal of Cognitive
Neuroscience, 12, 118–129.
Heyes, C. (2011). Automatic imitation. Psychological Bulletin,
137, 463.
Hirschfeld, L. A., & Gelman, S. A. (1994). Mapping the mind:
Domain specificity in cognition and culture. Cambridge,
UK: Cambridge University Press.
Hogeveen, J., Obhi, S. S., Banissy, M. J., Santiesteban, I., Press,
C., Catmur, C., et al. (2014). Task-dependent and distinct
roles of the temporoparietal junction and inferior frontal
cortex in the control of imitation. Social Cognitive and
Affective Neuroscience, 10, 1003–1009.
Hyde, J. S. (2014). Gender similarities and differences. Annual
Review of Psychology, 65, 373–398.
Iacoboni, M. (2009). Neurobiology of imitation. Current
Opinion in Neurobiology, 19, 661–665.
Iacoboni, M., Woods, R. P., Brass, M., Bekkering, H., Mazziotta,
J. C., & Rizzolatti, G. (1999). Cortical mechanisms of human
imitation. Science, 286, 2526–2528.
Josephs, O., & Henson, R. N. (1999). Event-related functional
magnetic resonance imaging: Modelling, inference and
optimization. Philosophical Transactions of the Royal Society
of London, Series B, Biological Sciences, 354, 1215–1228.
Julian, J. B., Fedorenko, E., Webster, J., & Kanwisher, N. (2012).
An algorithmic method for functionally defining regions of
interest in the ventral visual pathway. Neuroimage, 60,
2357–2364.
Kanwisher, N. (2010). Functional specificity in the human brain:
A window into the functional architecture of the mind.
Proceedings of the National Academy of Sciences, U.S.A.,
107, 11163–11170.
Klapper, A., Ramsey, R., Wigboldus, D., & Cross, E. S. (2014).
The control of automatic imitation based on bottom–up
and top–down cues to animacy: Insights from brain and
behavior. Journal of Cognitive Neuroscience, 26, 2503–2513.
Krach, S., Blumel, I., Marjoram, D., Lataster, T., Krabbendam, L.,
Weber, J., et al. (2009). Are women better mindreaders?
Sex differences in neural correlates of mentalizing detected
with functional MRI. BMC Neuroscience, 10, 9.
Kriegeskorte, N., Mur, M., & Bandettini, P. (2008). Representational
similarity analysis—Connecting the branches of systems
neuroscience. Frontiers in Systems Neuroscience, 2, 4.
Kriegeskorte, N., Simmons, W. K., Bellgowan, P. S., & Baker,
C. I. (2009). Circular analysis in systems neuroscience: The
dangers of double dipping. Nature Neuroscience, 12, 535–540.
Lakens, D. (2013). Calculating and reporting effect sizes to
facilitate cumulative science: A practical primer for t-tests
and ANOVAs. Frontiers in Psychology, 4, 863.
Marsh, L. E., Bird, G., & Catmur, C. (2016). The imitation game:
Effects of social cues on ‘imitation’ are domain-general in
nature. Neuroimage, 139, 368–375.
Mengotti, P., Corradi-Dell’Acqua, C., & Rumiati, R. I. (2012).
Imitation components in the human brain: An fMRI study.
Neuroimage, 59, 1622–1630.
Michael, J., & D’Ausilio, A. (2015). Domain-specific and domain-
general processes in social perception—A complementary
approach. Consciousness and Cognition, 36, 434–437.
Miller, D. I., & Halpern, D. F. (2014). The new science of
cognitive sex differences. Trends in Cognitive Sciences, 18,
37–45.
Müller, B. C., Leeuwen, M. L., Baaren, R. B., Bekkering, H., &
Dijksterhuis, A. (2013). Empathy is a beautiful thing: Empathy
predicts imitation only for attractive others. Scandinavian
Journal of Psychology, 54, 401–406.
Mumford, J. A. (2012). A power calculation guide for fMRI
studies. Social Cognitive and Affective Neuroscience, 7,
738–742.
Mumford, J. A., & Nichols, T. E. (2008). Power calculation for
group fMRI studies accounting for arbitrary design and
temporal autocorrelation. Neuroimage, 39, 261–268.
Nee, D. E., Wager, T. D., & Jonides, J. (2007). Interference
resolution: Insights from a meta-analysis of neuroimaging
tasks. Cognitive, Affective & Behavioral Neuroscience, 7,
1–17.
Newman-Norlund, R. D., van Schie, H. T., van Zuijlen, A. M., &
Bekkering, H. (2007). The mirror neuron system is more
active during complementary compared with imitative action.
Nature Neuroscience, 10, 817–819.
Nieto-Castañón, A., & Fedorenko, E. (2012). Subject-specific
functional localisers increase sensitivity and functional
resolution of multi-subject analyses. Neuroimage, 63,
1646–1669.
Norman, K. A., Polyn, S. M., Detre, G. J., & Haxby, J. V. (2006).
Beyond mind-reading: Multi-voxel pattern analysis of fMRI
data. Trends in Cognitive Sciences, 10, 424–430.
Open Science Collaboration. (2015). Estimating the
reproducibility of psychological science. Science.
doi:10.1126/science.aac4716.
Pitcher, D., Dilks, D. D., Saxe, R. R., Triantafyllou, C., &
Kanwisher, N. (2011). Differential selectivity for dynamic
versus static information in face-selective cortical regions.
Neuroimage, 56, 2356–2363.
Poldrack, R. A. (2006). Can cognitive processes be inferred from
neuroimaging data? Trends in Cognitive Sciences, 10, 59–63.
Quadflieg, S., Flannigan, N., Waiter, G. D., Rossion, B., Wig,
G. S., Turk, D. J., et al. (2011). Stereotype-based modulation
of person perception. Neuroimage, 57, 549–557.
Rahman, Q., Wilson, G. D., & Abrahams, S. (2004). Sex, sexual
orientation, and identification of positive and negative facial
affect. Brain and Cognition, 54, 179–185.
Rizzolatti, G., & Craighero, L. (2004). The mirror-neuron system.
Annual Review of Neuroscience, 27, 169–192.
Rubia, K., Hyde, Z., Halari, R., Giampietro, V., & Smith, A.
(2010). Effects of age and sex on developmental neural
networks of visual-spatial attention allocation. Neuroimage,
51, 817–827.
Russell, T. A., Tchanturia, K., Rahman, Q., & Schmidt, U. (2007).
Sex differences in theory of mind: A male advantage on Happé’s
“cartoon” task. Cognition and Emotion, 21, 1554–1564.
Santiesteban, I., Banissy, M. J., Catmur, C., & Bird, G. (2012).
Enhancing social ability by stimulating right temporoparietal
junction. Current Biology, 22, 2274–2277.
Santiesteban, I., Banissy, M. J., Catmur, C., & Bird, G. (2015).
Functional lateralization of temporoparietal junction—Imitation
inhibition, visual perspective-taking and theory of mind.
European Journal of Neuroscience, 42, 2527–2533.
Saxe, R., Brett, M., & Kanwisher, N. (2006). Divide and
conquer: A defense of functional localisers. Neuroimage, 30,
1088–1096.
932
Journal of Cognitive Neuroscience
Volume 30, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
0
6
9
1
4
1
7
8
7
5
2
9
/
j
o
c
n
_
a
_
0
1
2
6
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Saxe, R., & Kanwisher, N. (2003). People thinking about
thinking people: The role of the temporo-parietal junction
in “theory of mind”. Neuroimage, 19, 1835–1842.
Schaafsma, S. M., Pfaff, D. W., Spunt, R. P., & Adolphs, R. (2015).
Deconstructing and reconstructing theory of mind. Trends
in Cognitive Sciences, 19, 65–72.
Spunt, R. P., & Adolphs, R. (2015). Folk explanations of behavior:
A specialized use of a domain-general mechanism. Psychological
Science, 26, 724–736.
Spunt, R. P., & Adolphs, R. (2017). A new look at domain
specificity: Insights from social neuroscience. Nature Reviews
Neuroscience, 18, 559.
Schurz, M., Tholen, M. G., Perner, J., Mars, R. B., & Sallet, J.
Stoet, G. (2010). Sex differences in the processing of flankers.
(2017). Specifying the brain anatomy underlying
temporo-parietal junction activations for theory of mind:
A review using probabilistic atlases from different imaging
modalities. Human Brain Mapping, 38, 4788–4805.
Schuwerk, T., Schurz, M., Müller, F., Rupprecht, R., & Sommer,
M. (2017). The rTPJ’s overarching cognitive function in
networks for attention and theory of mind. Social Cognitive
and Affective Neuroscience, 12, 157–168.
Simon, J. R. (1969). Reactions toward the source of stimulation.
Journal of Experimental Psychology, 81, 174.
Sonnby-Borgström, M. (2002). Automatic mimicry reactions as
related to differences in emotional empathy. Scandinavian
Journal of Psychology, 43, 433–443.
Sonnby-Borgström, M., Jönsson, P., & Svensson, O. (2003).
Emotional empathy as related to mimicry reactions at
different levels of information processing. Journal of
Nonverbal behavior, 27, 3–23.
Sonnby-Borgström, M. A., Jönsson, P., & Svensson, O. (2008).
Gender differences in facial imitation and verbally reported
emotional contagion from spontaneous to emotionally
regulated processing levels. Scandinavian Journal of
Psychology, 49, 111–122.
Southgate, V., & Hamilton, A. F. (2008). Unbroken mirrors:
Challenging a theory of autism. Trends in Cognitive Sciences,
12, 225–229.
Sowden, S., & Catmur, C. (2015). The role of the right
temporoparietal junction in the control of imitation. Cerebral
Cortex, 25, 1107–1113.
Sowden, S., & Shah, P. (2014). Self-other control: A candidate
mechanism for social cognitive function. Frontiers in
Human Neuroscience, 8, 789.
Spengler, S., von Cramon, D. Y., & Brass, M. (2009). Control of
shared representations relies on key processes in mental
state reasoning. Human Brain Mapping, 30, 3704–3718.
Spengler, S., von Cramon, D. Y., & Brass, M. (2010). Resisting
motor mimicry: Control of imitation involves processes
central to social cognition in patients with frontal and
temporo-parietal lesions. Social Neuroscience, 5, 401–416.
Sporns, O., Tononi, G., & Kötter, R. (2005). The human
connectome: A structural description of the human brain.
PLoS Computational Biology, 1, e42.
Spunt, R. P., & Adolphs, R. (2014). Validating the why/how
contrast for functional MRI studies of theory of mind.
Neuroimage, 99, 301–311.
The Quarterly Journal of Experimental Psychology, 63,
633–638.
Stoet, G. (2017). Sex differences in the Simon task help to
interpret sex differences in selective attention. Psychological
Research, 81, 571–581.
Thayer, J. F., & Johnsen, B. H. (2000). Sex differences in
judgement of facial affect: A multivariate analysis of
recognition errors. Scandinavian Journal of Psychology,
41, 243–246.
Van Overwalle, F. (2009). Social cognition and the brain:
A meta-analysis. Human Brain Mapping, 30, 829–858.
van Schie, H. T., van Waterschoot, B. M., & Bekkering, H.
(2008). Understanding action beyond imitation: Reversed
compatibility effects of action observation in imitation and
joint action. Journal of Experimental Psychology: Human
Perception and Performance, 34, 1493.
Wager, T. D., & Nichols, T. E. (2003). Optimization of experimental
design in fMRI: A general framework using a genetic
algorithm. Neuroimage, 18, 293–309.
Wager, T. D., Sylvester, C. Y. C., Lacey, S. C., Nee, D. E.,
Franklin, M., & Jonides, J. (2005). Common and unique
components of response inhibition revealed by fMRI.
Neuroimage, 27, 323–340.
Wang, Y., Ramsey, R., & Hamilton, A. F. (2011). The control of
mimicry by eye contact is mediated by medial prefrontal
cortex. Journal of Neuroscience, 31, 12001–12010.
Weeks, D. J., & Proctor, R. W. (1990). Salient-features coding in
the translation between orthogonal stimulus and response
dimensions. Journal of Experimental Psychology: General,
119, 355–366.
Wiggett, A. J., Hudson, M., Tipper, S. P., & Downing, P. E.
(2011). Learning associations between action and perception:
Effects of incompatible training on body part and spatial
priming. Brain and Cognition, 76, 87–96.
Yarkoni, T., Poldrack, R. A., Nichols, T. E., Van Essen, D. C., &
Wager, T. D. (2011). Large-scale automated synthesis of human
functional neuroimaging data. Nature Methods, 8, 665.
Zaki, J., Hennigan, K., Weber, J., & Ochsner, K. N. (2010). Social
cognitive conflict resolution: Contributions of domain
general and domain specific neural systems. Journal of
Neuroscience, 30, 8481–8488.
Zaki, J., & Ochsner, K. N. (2012). The neuroscience of
empathy: Progress, pitfalls and promise. Nature
Neuroscience, 15, 675.
Darda, Butler, and Ramsey
933
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
0
6
9
1
4
1
7
8
7
5
2
9
/
j
o
c
n
_
a
_
0
1
2
6
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3