Functional Context Affects Scene Processing
Elissa M. Aminoff1
and Michael J. Tarr2
Abstract
■ Rapid visual perception is often viewed as a bottom–up pro-
cess. Category-preferred neural regions are often characterized
as automatic, default processing mechanisms for visual inputs of
their categorical preference. To explore the sensitivity of such
regions to top–down information, we examined three scene-
preferring brain regions, the occipital place area (OPA), the para-
hippocampal place area (PPA), and the retrosplenial complex
(RSC), and tested whether the processing of outdoor scenes is
influenced by the functional contexts in which they are seen.
Context was manipulated by presenting real-world landscape
images as if being viewed through a window or within a picture
frame—manipulations that do not affect scene content but do
affect one’s functional knowledge regarding the scene. This
manipulation influences neural scene processing (as measured by
fMRI): The OPA and the PPA exhibited greater neural activity when
participants viewed images as if through a window as compared
with within a picture frame, whereas the RSC did not show this dif-
ference. In a separate behavioral experiment, functional context
affected scene memory in predictable directions (boundary exten-
sion). Our interpretation is that the window context denotes three-
dimensionality, therefore rendering the perceptual experience of
viewing landscapes as more realistic. Conversely, the frame context
denotes a 2-D image. As such, more spatially biased scene represen-
tations in the OPA and the PPA are influenced by differences in top–
down, perceptual expectations generated from context. In contrast,
more semantically biased scene representations in the RSC are
likely to be less affected by top–down signals that carry information
about the physical layout of a scene. ■
INTRODUCTION
Although rapid visual perception is often considered as a
primarily bottom–up process, it is well established that
the processing of visual input involves both bottom–up
and top–down mechanisms (Kay & Yeatman, 2017; Fang,
Boyaci, Kersten, & Murray, 2008; Lamme & Roelfsema,
2000; Felleman & Van Essen, 1991). For example, the
responses of the scene-selective network of category-
preferred brain regions are affected by top–down informa-
tion regarding learned contextual associations (Bar &
Aminoff, 2003). This network of regions, the parahip-
pocampal place area (PPA)/lingual region (Epstein &
Kanwisher, 1998), the retrosplenial complex (RSC;
Maguire, 2001), and the occipital place area (OPA; also
known as the transverse occipital sulcus; Dilks, Julian,
Paunov, & Kanwisher, 2013), appears to represent a
wide variety of scene characteristics (reviewed in Epstein
& Baker, 2019). The list of scene-relevant properties
includes spatial layout, three-dimensionality, landmark
processing, navigability, environment orientation and
retinotopic bias, scene boundaries, scene categories, ob-
jects within a scene, and the contextual associative nature
of the scene (Lescroart & Gallant, 2019; Lowe, Rajsic,
Gallivan, Ferber, & Cant, 2017; Baldassano, Fei-Fei, &
Beck, 2016; Çukur, Huth, Nishimoto, & Gallant, 2016;
1Fordham University, 2Carnegie Mellon University
© 2021 Massachusetts Institute of Technology
Julian, Ryan, Hamilton, & Epstein, 2016; Aminoff & Tarr,
2015; Marchette, Vass, Ryan, & Epstein, 2015; Park, Konkle,
& Oliva, 2015; Silson, Chan, Reynolds, Kravitz, & Baker, 2015;
Troiani, Stigliani, Smith, & Epstein, 2014; Harel, Kravitz, &
Baker, 2013; Auger, Mullally, & Maguire, 2012; Nasr &
Tootell, 2012; Henderson, Zhu, & Larson, 2011; Kravitz,
Peng, & Baker, 2011; Park, Brady, Greene, & Oliva, 2011;
Bar, Aminoff, & Schacter, 2008; Janzen & van Turennout,
2004; Levy, Hasson, Avidan, Hendler, & Malach, 2001).
One of the significant open questions regarding the
representation of scene properties is how they come to
be encoded; that is, to what extent are the associated
neural responses driven by visual properties within scenes
as opposed to nonperceptual high-level scene properties,
such as learned functional properties1 and semantics? We
address this question by exploring whether prior experi-
ence and expectations modulate scene-selective neural
activity.
We used fMRI to measure neural responses while partic-
ipants viewed the otherwise identical outdoor scenes in
two different contexts: in a window frame (“WIN” condi-
tion) or in a picture frame (“PIC” condition; Figure 1).
We hypothesize that viewing scene images surrounded
by a window invokes a more naturalistic context that is
closer to the perceptual experience of real-world scene
processing. More specifically, a window connotes that
the scene is 3-D, navigable, and extends beyond the
boundaries presented. In contrast, we hypothesize that
viewing scene images surrounded by a picture frame
Journal of Cognitive Neuroscience 33:5, pp. 933–945
https://doi.org/10.1162/jocn_a_01694
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
3
5
9
3
3
1
9
5
9
4
5
5
/
j
o
c
n
_
a
_
0
1
6
9
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
3
5
9
3
3
1
9
5
9
4
5
5
/
j
o
c
n
_
a
_
0
1
6
9
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 1. Sample stimuli showing the same scenes in both the picture frame (PIC) and the window frame ( WIN) conditions. See Methods for more
information.
invokes a less realistic context in which the scene is viewed
as a 2-D picture without extension beyond the frame; as a
consequence, inferential scene properties such as spatial
affordances are likely to be limited. Based on these
assumptions, we predict that the perception of a scene
image will vary based on the context in which the image
is situated. Under the assumption that the network of
scene-preferred brain regions (PPA, RSC, and OPA) sub-
serves different computational functions, we also predict
that these regions will respond differently from one another
across the manipulation of scene context. Alternatively, if
scene preference is purely a function of scene content,
one should predict no differences in responses across
these regions.
To further explore the effect of functional context, we ex-
amined how the picture frame versus window frame manip-
ulation affects boundary extension—a well-documented
distortion of scene memory (Intraub, 2010, 2014; Intraub
& Richardson, 1989). Boundary extension has been dis-
cussed as a memory distortion directly related to scene
representation—a phenomenon that is intertwined with
the spatial affordances arising from the process of scene
perception applied to picture viewing (Intraub, 2010,
2020). When we experience a real-world scene via either
direct viewing or a picture, we are not just perceiving the
scene as a finite entity but as a percept that continues
beyond the edges of our perception. Thus, if we manipulate
the functional context of scenes by presenting them explic-
itly in picture frames, we are limiting the spatial context
necessary for scene understanding and boundary extension
should be reduced. As such, we predicted greater boundary
extension for window-framed scenes as compared with
picture-framed scenes.
More broadly, the manipulation of functional context
addresses the question of whether scene-preferred brain
regions process category-relevant inputs in a primarily
bottom–up manner or whether they are sensitive to top–
down influences. At the same time, the pattern of neural
modulation across different scene-preferred brain regions
adds to our understanding of the different functional roles
for each.
METHODS
fMRI Experiment
Participants
Eighteen individuals participated in this experiment; 17 were
included in the analysis (mean age = 23.6 years, range =
18–30 years; 8 women, 9 men; 1 left-handed). One participant
was removed from the analysis because of extremely poor
performance, indicative of falling asleep (missing 22% of
the repeated trials in a trivial 1-back task). All participants
had normal or corrected-to-normal vision and were not
taking any psychoactive medication. Written informed
consent was obtained from all participants before testing
in accordance with the procedures approved by the insti-
tutional review board of Carnegie Mellon University.
Participants were financially compensated for their time.
Stimuli
The main experiment included 120 outdoor scenes, in-
cluding both manmade outdoor scenes such as a garden
patio, as well as natural landscapes such as a mountain
range. A majority of the stimuli were found and obtained
through Google Image Search. There were two versions
of each scene: one within the context of a window frame
and the other within the context of a picture frame (see
Figure 1).
A pool of 13 window frames and 13 picture frames was
used across the 120 scenes. Each scene presented within
the frame subtended 5.5° of visual angle, and the average
extent of the frames was 9° with 0.68° ( WIN) and 0.61°
(PIC) standard deviations across the different frame exem-
plars. The frames were set against a gray rectangular back-
ground that subtended 10° of visual angle; the remainder
of the screen background was black.
In a post hoc analysis, the brightness, contrast, and spa-
tial frequency were measured for all stimulus images.
Images in the PIC and WIN conditions were found to be
matched across contrast and spatial frequency. However,
there was a difference in brightness with PIC images
brighter on average than WIN images.
934
Journal of Cognitive Neuroscience
Volume 33, Number 5
Stimuli in the localizer experiment included 60 scenes
(outdoor and indoor, nonoverlapping with the stimuli
used in the main experiment), 60 weak contextual objects
(Bar & Aminoff, 2003), and 60 phase-scrambled scenes.
Phase-scrambled scenes were generated by running a
Fourier transform of each scene image, scrambling the
phases, and then performing an inverse Fourier transform
back into the pixel space. All stimuli were presented at a
5.5° visual angle against a gray background.
Procedure
During fMRI scanning, images were presented to the partic-
ipants via 24-in. MR compatible LCD display (BOLDScreen,
Cambridge Research Systems LTD.) located at the head
of the bore and reflected through a head coil mirror to
the participant. There were two functional runs in the
WIN/PIC experiment. Functional scans used a blocked
design alternating WIN blocks and PIC blocks with fixation
in between. The order of the blocks was balanced both
across and within participants. Each functional scan began
and ended with 12 sec of a white fixation cross (“+”) pre-
sented against a black background. Images were presented
for 750 msec, with a 250-msec ISI. Each block contained 10
unique images and two repeated images, for a total block
duration of 12 sec. Each run consisted of six blocks per con-
dition. There were 10 sec of fixation between task blocks.
Participants performed a 1-back task where they pressed
a button if the picture immediately repeated, two per
block. Each run presented all 120 stimuli, 60 presented in
the WIN condition, and 60 presented in the PIC condition.
The second run presented all 120 stimuli again, but with the
presentation condition (PIC or WIN) swapped. The condi-
tion in which a stimulus was presented first was balanced
across participants.
Most participants had two functional localizer runs (two
participants had only one run because of time constraints)
to functionally define scene-preferred regions.2 Localizer
runs consisted of three conditions: scenes, objects, and
phase-scrambled scenes. These runs began and ended
with 12 sec of a black fixation cross (“+”) presented
against a gray background. Each run had four blocks per
condition. Images were presented for 800 msec, with
200-msec ISI, with the exception that the first stimulus
in each block other than the first block was presented
for 2800 msec. Each block contained 12 unique images
with two repeated images, for a total block duration of
14 sec for the first block and 16 sec thereafter because
of the longer presentation of the first stimulus. There were
10 sec of fixation between task blocks. Participants per-
formed a 1-back task where they pressed a button if the
picture immediately repeated, two per block. The localizer
runs occurred after the WIN/PIC functional runs.
fMRI Data Acquisition
fMRI data were collected on a 3T Siemens Verio MR scan-
ner at the Scientific Imaging and Brain Research Center at
Carnegie Mellon University using a 32-channel head coil.
Functional images were acquired using a T2*-weighted
echo-planar imaging multiband pulse sequence (69 slices
aligned to the AC/PC, in-plane resolution 2 mm × 2 mm,
2 mm slice thickness, no gap, repetition time [TR] =
2000 msec, echo time [TE] = 30 msec, flip angle = 79°,
multiband acceleration factor = 3, field of view =
192 mm, phase encoding direction A >> P, ascending ac-
quisition). Number of acquisitions per run was 139 for the
WIN/PIC runs and 162 for the scene localizer. High-
resolution anatomical scans were acquired for each partic-
ipant using a T1-weighted MPRAGE sequence (1 mm ×
1 mm × 1 mm, 176 sagittal slices, TR = 2.3 sec, TE =
1.97 msec, flip angle = 9°, GRAPPA = 2, field of view =
256). A field-map scan was also acquired to correct for
distortion effects using the same slice prescription as the
EPI scans (69 slices aligned to the AC/PC, in-plane resolu-
tion 2 mm × 2 mm, 2 mm slice thickness, no gap, TR =
724 msec, TE1 = 5 msec, TE2 = 7.46 msec, flip angle =
70°, field of view = 192 mm, phase encoding direction
A >> P, interleaved acquisition).
fMRI Data Analysis
All fMRI data were analyzed using SPM12 (www.fil.ion.ucl
.ac.uk/spm/software/spm12/ ). All data were preprocessed
to correct for motion and to unwarp for geometric distor-
tions using the field-map scan acquired. Data were
smoothed using an isotropic Gaussian kernel (FWHM =
4 mm). Only data used for the group average activation
maps were normalized to the Montreal Neurological
Institute template. Otherwise, data used were in native
space (i.e., all ROI analyses). The data were analyzed as a
block design using a general linear model and canonical
hemodynamic response function. A high-pass filter using
128 sec was implemented. The six motion parameter esti-
mates that output from realignment were used as addi-
tional nuisance regressors. An autoregressive model of
order 1, AR(1), was used to account for the temporal cor-
relations of the residuals. For the whole-brain analysis in
the group average, the contrasts were passed to a second-
level random-effects analysis that consisted of testing the
contrast against zero using a voxel-wise single-sample t test.
All group-averaged activity maps are examined through a
whole-brain analysis using a false discovery rate correction
of q = .05. For visualization purposes, these average maps
were rendered onto a 3-D inflated brain using CARET (Van
Essen et al., 2001).
All ROIs analyzed were defined and extracted at the
individual level using the MarsBaR toolbox (marsbar
.sourceforge.net/index.html) or in-house MATLAB (The
MathWorks) scripts and analyzed in native space. Scene-
preferred regions (PPA, RSC, and OPA) were functionally
defined using the contrast of scenes greater than the com-
bined conditions of objects and phase-scrambled scenes
from the localizer runs. Typically, a threshold of family-
wise error, p < .001, was used to define the set of voxels.
Aminoff and Tarr
935
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
5
9
3
3
1
9
5
9
4
5
5
/
j
o
c
n
_
a
_
0
1
6
9
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
In a post hoc analysis, the effect of stimulus brightness
was evaluated. To test whether stimulus brightness con-
tributed to any of our observed effects, we measured the
mean brightness across all images within a block (as pre-
sented to the individual participant during the fMRI run).
Blocks within the same frame condition (PIC, WIN) were
separated into the brighter blocks (n = 6) and the darker
blocks (n = 6), thereby yielding four conditions: PIC
Bright, PIC Dark, WIN Bright, and WIN Dark. Conditions
were compared to determine whether the differences in
the WIN and PIC conditions could be accounted for by
image brightness.
Behavioral Experiment
Participants
Thirty-seven individuals participated in the behavioral
experiment examining boundary extension. Data from
36 individuals were included in the analysis, one partici-
pant was removed because of a technical error related to
which buttons were pressed. The participants were under-
graduates at Fordham University who were either paid for
their participation or received course credit (mean age =
20.0 years, SD = 1.36 years, range = 18–22 years; 28 women,
7 men; 4 left-handed). Written informed consent was ob-
tained from all participants before testing in accordance
with the procedures approved by the institutional review
board of Fordham University.
Stimuli
The stimuli for this experiment were 200 unique scenes,
which included the 120 scenes used in the fMRI experi-
ment as well as an additional 80 outdoor scenes added
to increase the total number of trials. As in the fMRI exper-
iment, there were two formats for each scene: one within
the context of a window frame ( WIN) and the other in the
context of a picture frame (PIC). The same pool of window
frames and picture frames from the fMRI experiment was
applied to the 80 new pictures. Pictures were divided into
two groups of 100 scenes, Group A and Group B. Images
were presented to the participants on a 27-in. iMac using
Psychtoolbox (Brainard, 1997) and MATLAB.
Procedure
Participants were instructed to memorize all of the scenes
presented in the experiment. In the study phase, a single
scene image was presented on each trial, and participants
judged whether there was water in the picture. Each trial
was composed of a white fixation cross presented against
a gray background for 250 msec, a scene presented for
250 msec, and a repeat of the fixation cross for 250 msec.
Following the second fixation cross, participants viewed a
response screen showing: “(b) Water (n) No Water.”
Participants had up to 2500 msec to respond with the
appropriate key press (b or n). Immediately after the par-
ticipant responded, the next trial started.
Trials were broken into blocks of 25 trials, between which
participants were offered a break. Each block consisted of
pictures from a single condition, either PIC or WIN.
Condition order alternated, starting with the WIN condi-
tion. Group A stimuli were presented in the WIN condition,
and Group B stimuli were presented in the PIC condition.
After 200 trials—a total of eight blocks, four from each
condition—participants’ memory for the scenes was tested.
In the test phase, a fixation cross was presented for
250 msec, followed by a picture of a scene shown during
the study phase, except without a frame. Participants
judged whether the scene was identical to the version they
had seen at study (absent the frame), was zoomed in (i.e.,
closer) relative to the version they had seen at study, or
was zoomed out (i.e., wider) relative to the version they
had seen at study. Participants responded on a 5-point
scale: very close, close, same, wide, and very wide. The
response screen was self-paced. After participants judged
the amount of “zoom,” they rated their confidence on a
3-point scale: sure, pretty sure, or don’t remember pic-
ture. This screen was self-paced as well. Trials were broken
into blocks of 25 trials, and as before, each block consisted
of pictures from a single condition, either PIC or WIN. All
scenes presented in the test phase were actually shown
with the “same” boundaries as presented in the study
phase—that is, with no zoom in or out. Thus, the correct
answer was always “same.”
After the 200 test trials, participants were presented
with another 200 study and 200 test trials using the same
200 scenes, but appearing in the opposite condition at
study as compared with the first study/test session. Here,
Group A stimuli appeared in the PIC condition, and
Group B stimuli appeared in the WIN condition. The con-
dition order again alternated across blocks, but here, start-
ing with the PIC condition. Although presentation order
was randomized for both sessions, a technical bug resulted
in the stimuli and order of conditions not being balanced
across conditions. See Results for detailed analysis demon-
strating that this error did not affect the results.
Responses at test were converted to an integer score
from −2 to +2 (corresponding to very close, close, same,
wide, and very wide), where positive values denote when
participants perceived the scene at test to be “wider” than
they remembered seeing it at study (i.e., boundary con-
traction), zero represents no change from study to test,
and negative values denote when participants perceived
the scene at test to be “closer” than they remembered see-
ing it at study (i.e., boundary extension). Scores were
summed across all test trials separately for the WIN and
PIC conditions. Responses with RTs exceeding 3 SDs from
the participant’s mean were considered outliers and
removed from the analysis. A t test ( WIN/PIC) was per-
formed on these summed scores. A second analysis was
run based on the confidence of the participant. If the par-
ticipant responded “Don’t remember picture,” that trial
936
Journal of Cognitive Neuroscience
Volume 33, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
5
9
3
3
1
9
5
9
4
5
5
/
j
o
c
n
_
a
_
0
1
6
9
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
was removed from the analysis to ensure any effects
arose from the frame context manipulation and not a
failure of memory.
RESULTS
fMRI Experiment
We hypothesized that the PIC versus the WIN context
manipulation would give rise to different top–down driven
inferences—reflected in responses in scene-preferred brain
regions—about the nature of the viewed scene. Neural re-
sponses were measured using fMRI in a block design, and
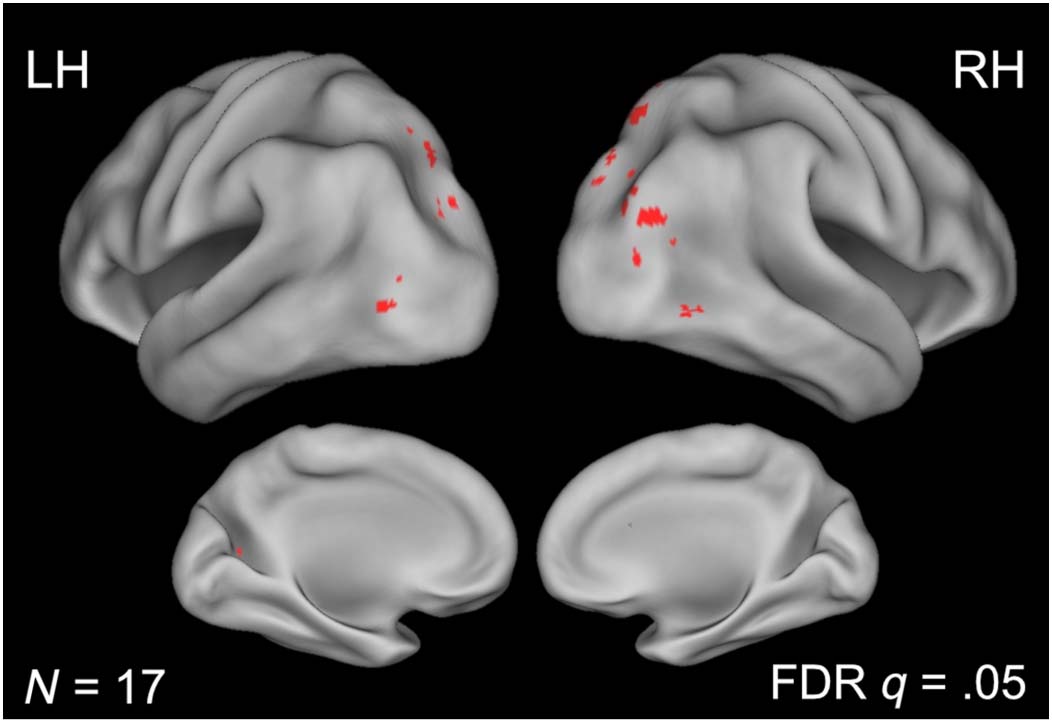
we performed a whole-brain analysis comparing the BOLD
activity elicited by WIN versus PIC blocks. This comparison
revealed no voxel responses with larger magnitudes for the
PIC as compared with the WIN condition (false discovery
rate threshold at q = .05). In contrast, there were many
voxel responses of larger magnitude for the WIN as com-
pared with the PIC condition. These voxels were located
within the dorsal visual stream, within the occipital cortex,
and within the parietal cortex, close to the inferior portion
(Figure 2).
We next examined how our context manipulation af-
fects different scene-preferred brain regions (Figure 3).
An independent functional localizer was used to de-
fine ROIs commonly observed to be selective for scene
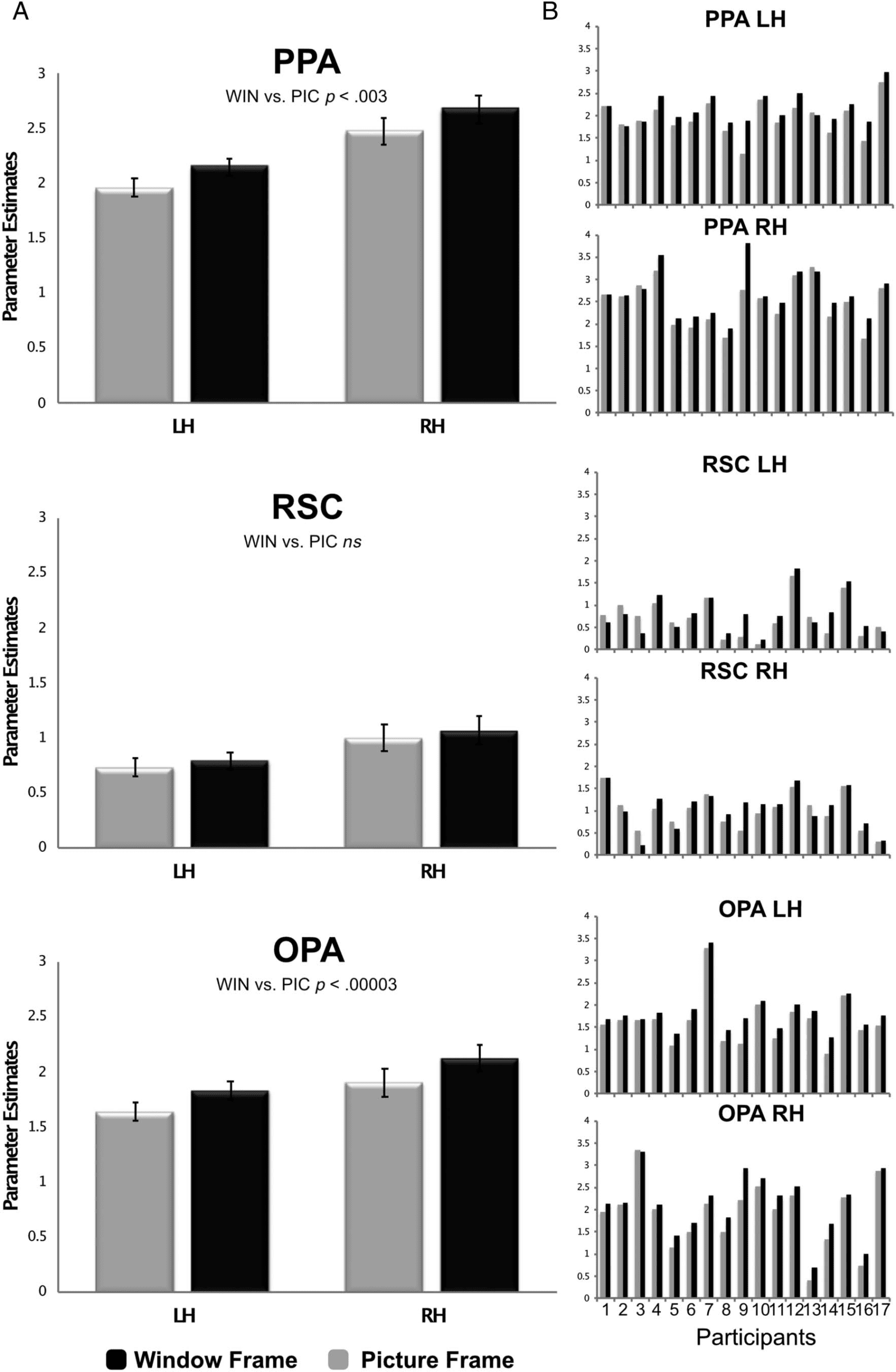
processing—PPA, RSC, and OPA. An ANOVA with ROI ×
Hemisphere × Condition as factors revealed a significant
main effect of Condition, with WIN eliciting more activity
than PIC, F(1, 16) = 11.83, p < .003, ηp
2 = .425. There was
also a main effect of ROI, F(2, 32) = 85.02, p < 1.57 ×
10−13, ηp
2 = .842, with the PPA showing the highest mag-
nitude response (2.3 parameter estimate) as compared
with either the OPA (1.9 parameter estimate, p < .001 in
planned comparisons) or the RSC (0.89 parameter esti-
mate, p < .0001); the OPA response was also significantly
higher than the RSC response ( p < .0001). The effect of
Hemisphere was significant, with the right hemisphere
eliciting more activity than the left hemisphere, F(1, 16) =
19.07, p < .0005, ηp
2 = .544. There was also a significant
interaction between ROI × Condition, F(2, 32) = 10.95, p <
.0003, ηp
2 = .407. Pairwise ROI × Condition comparisons
revealed that this interaction was driven by significant differ-
ences between both the PPA and OPA as compared with the
RSC: PPA versus RSC, F(1, 16) = 21.26, p < .0003, ηp
2 = .571;
OPA versus RSC, F(1, 16) = 15.09, p < .001, ηp
2 = .485.
There was no significant effect when comparing the PPA
to the OPA, F(1, 16) = 0.080, p > .78, ηp
2 = .005. No other
interactions were significant.
To explore the effect of the context manipulation within
each specific scene-preferred region, we ran separate
ANOVAs for each ROI (Hemisphere × Condition). In the
PPA, there was a significant main effect of Condition, F(1,
16) = 12.45, p < .003, ηp
2 = .438, with WIN eliciting signif-
icantly more activity than PIC. There was also a significant
difference in Hemisphere, F(1, 16) = 17.72, p < .001, ηp
2 =
.526, with the right hemisphere showing more activity
than the left hemisphere. The interaction was not signifi-
cant ( p > .9). In the OPA, there was a significant main
effect of Condition, F(1, 16) = 33.71, p < .00003, ηp
2 =
.678, with WIN eliciting significantly more activity than
PIC. Neither the main effect of Hemisphere nor the
Hemisphere × Condition interaction were significant
( ps > .15). In the RSC, there was no significant main effect
of Condition ( p > .24) nor any interaction between
Hemisphere × Condition. However, there was a main
effect of Hemisphere, with the right-hemisphere response
being greater than the left-hemisphere response, F(1, 16) =
11.27, p < .004, ηp
2 = .413.
Presentation order effects were explored by comparing
Runs 1 and 2—where the same scene images appeared in
different contexts. An ANOVA for each ROI was run with
Hemisphere × Condition × Run as factors. Suggesting that
order made no difference in neural responses, the main
effect of Run was insignificant for each ROI ( p > .18, ηp
2 <
.11), as was the interaction between Condition × Run ( p >
.14, ηp
2 < .14). The interaction of Hemisphere × Run was
not significant in the RSC ( p > .68, ηp
2 < .01), was margin-
ally significant for the PPA ( p < .07, ηp
2 < .19), and was
Figure 2. Whole-brain analysis
examining activity elicited for
scenes in window frames ( WIN)
as compared with the activity
for scenes in picture frames
(PIC).
Aminoff and Tarr
937
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
5
9
3
3
1
9
5
9
4
5
5
/
j
o
c
n
_
a
_
0
1
6
9
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
5
9
3
3
1
9
5
9
4
5
5
/
j
o
c
n
_
a
_
0
1
6
9
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. ROI analyses for both the group average (A) and individual participants (B). WIN condition = black; PIC condition = gray.
938
Journal of Cognitive Neuroscience
Volume 33, Number 5
significant in the OPA ( p < .02, ηp
2 < .31). The overall pat-
tern does show greater activity in Run 1 as compared with
Run 2, which is consistent with adaptation to the stimuli,
regardless of condition. However, we found this effect to
be modulated by hemisphere. In the PPA, the effect of ad-
aptation was marginally greater in the left hemisphere than
in the right hemisphere (Run 1 minus Run 2: left hemi-
sphere 0.14, right hemisphere 0.05). In the OPA, adaptation
was again observed in the left hemisphere (0.11); however,
in the right hemisphere, there was slightly greater activity in
Run 2 compared with Run 1, yielding the significant interac-
tion (right hemisphere −0.02). The three-way interaction
of Hemisphere × Condition × Run was not significant
2 < .0; RSC, p < .34, ηp
(PPA, p < .94, ηp
2 < .06; OPA, p <
.07, ηp
2 < .2).
A significant Hemisphere effect was found in a number
of our analyses. However, our main manipulation of inter-
est ( WIN vs. PIC) did not interact with Hemisphere.
However, our results do reflect a preference for scene pro-
cessing in the right hemisphere—an effect that is difficult
to compare to prior findings in that many studies examin-
ing scene selectivity collapse across hemispheres without
statistical support. As such, the pervasiveness of this hemi-
spheric effect is unknown. We suggest several reasons for
observing a hemispheric difference in our study. First, the
left hemisphere may preferentially process high spatial fre-
quencies, whereas the right hemisphere may preferen-
tially process low spatial frequencies (for a review, see
Kauffmann, Ramanoël, & Peyrin, 2014). Low spatial fre-
quencies have a unique role in the rapid processing of
contextual and scene information (Greene & Oliva,
2009; Bar, 2004; Oliva & Torralba, 2001). Second, the right
hemisphere may be biased toward perceptual properties
of a scene, whereas the left hemisphere may be biased to-
ward conceptual information (Stevens, Kahn, Wig, &
Schacter, 2012; van der Ham, van Zandvoort, Frijns,
Kappelle, & Postma, 2011). However, this difference
would not seem to be able to account for why, in our
study, scene processing recruits the right hemisphere
preferentially, in that performing the 1-back task would
seem to recruit both perceptual and conceptual informa-
tion and that both levels of description are relevant to
judging whether one image matches another.
A post hoc analysis was run to test whether differences
in brightness accounted for the observed effects. When
overall image brightness was considered as a separate fac-
tor, we failed to find any significant effect of brightness
(PIC Bright = PIC Dark, WIN Bright = WIN Dark, ps >
.25). Moreover, in 13 of the 17 participants, we were able
to equate brightness across the PIC and WIN conditions,
allowing us to directly compare the PIC and WIN condi-
tions with equal average brightness for the images across
the two conditions. Despite equivalent average bright-
ness, we again found the predicted significant effect of
context: left-hemisphere PPA, t(12) = 2.40, p < .033;
left-hemisphere OPA, t(12) = 3.54, p < .004; right-
hemisphere PPA, t(12) = 2.69, p < .02; right-hemisphere
OPA, t(12) = 4.17, p < .001; left- and right-hemisphere
RSC, ns). As such, we conclude that differences in low-
level properties do not underlie our contextual interpreta-
tion of the observed differences between conditions.
Behavioral Experiment
Our neuroimaging results suggest that window frames
render scene images more “scene-like”—that is, perceived
as more realistic. But what does “more realistic” entail?
Viewing a scene in a window frame versus a picture frame
affects the functional context and thus the associated spa-
tial affordances. More specifically, a scene in a picture
frame is understood in the functional context of “what is
in the picture is what is important,” whereas a scene in a
window is understood to be only a part of the overall
scene. For example, when we view only part of a real-world
scene (e.g., the position of a bed in a bedroom), we know
to turn our head to perceive and interpret additional fea-
tures of the scene (e.g., the location of the closet). Under
this view, we predict that differences found in the neural
representations of the WIN and PIC scene conditions
should also manifest in behavioral measures of scene per-
ception because of these differences in functional context.
In particular, boundary extension is a phenomenon where
observers remember scenes with wider boundaries (i.e.,
more zoomed out) than what was originally experienced
(Intraub, 2014; Intraub & Richardson, 1989). The bound-
ary extension phenomenon is held to be specific to scene
memory (for an alternative account, see Bainbridge &
Baker, 2020). Moreover, there is evidence that boundary
extension manipulations also recruit the PPA (Chadwick,
Mullally, & Maguire, 2013; Park, Intraub, Yi, Widders, &
Chun, 2007). As such, we do see consistency across
boundary extension studies and our fMRI experiment in
that PPA appears to correlate with BE and the observed sig-
nificant recruitment of the PPA for our frame manipula-
tion. Here, on the basis of the assumed differences
between the window and picture frame contexts, we hy-
pothesized a larger boundary extension effect for scenes
presented in windows than for scenes presented in picture
frames. This context manipulation—the same as used in
our fMRI experiment—was included during the study
phase of this experiment. During the subsequent test
phase, the same scenes were presented without any
frame, and participants’ memory was probed via reports
as to whether each scene was identical (minus the frame)
to its presentation at study, zoomed in (i.e., closer), or
zoomed out (i.e., wider).
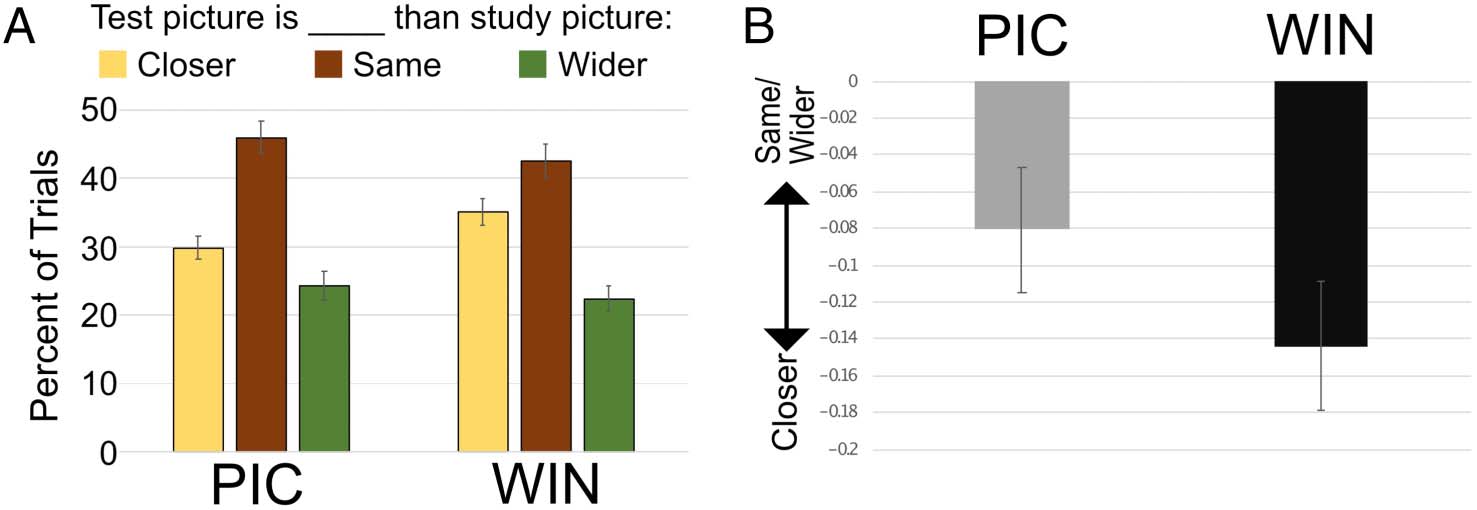
Across both study contexts, participants remembered
the scene at test as being closer than what was actually pre-
sented at study (i.e., boundary extension; 32% of the trials)
more often than the scene at test being farther than at
study (i.e., boundary contraction; 23% of the trials)—a sig-
nificant difference, t(35) = 3.3, p < .002. Relevant to our
hypothesis, participants more often remembered that
scenes in the WIN condition were closer at test relative
Aminoff and Tarr
939
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
5
9
3
3
1
9
5
9
4
5
5
/
j
o
c
n
_
a
_
0
1
6
9
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. Boundary extension
results. (A) Percentage of trials
at test the participants thought
the test image was closer, the
same as, or wider than the
study image. (B) The average
converted bias scores where
negative denotes that responses
were biased to remember the
test image as closer than what
was actually presented at study.
to scenes in the PIC condition (35% vs. 30% of test trials;
Figure 4). To measure this bias in scene memory, we com-
puted an average based on the integer values assigned to
each response (see Methods): The bias score for the WIN
condition was −0.14, whereas the bias score for the PIC
condition was −0.08 (Figure 4). This difference in memory
bias indicates that participants were more likely to remem-
ber the WIN scenes as wider compared with the PIC
scenes, t(35) = 2.85, p < .007. We also examined the bias
removing any trials in which the participants responded
“Don’t remember picture” in their confidence judgment.
Again, we observed a difference in memory bias: The bias
score for the WIN condition was −0.15, whereas the bias
score for the PIC condition was −0.09, t(35) = 2.96, p <
.006. These results support our prediction that scenes in a
window frame context will elicit a greater boundary exten-
sion effect—consistent with the greater scene-selective
neural responses observed in our fMRI study.
Presentation order effects were explored by comparing
the two study/test sessions where the same scene images
appeared in counterbalanced contexts. The main effect
of Session was not significant, F(1, 35) = 1.159, p =
.289; ηp
2 = .032; the main effect of Condition was significant
(PIC or WIN), F(1, 35) = 8.808, p < .007, ηp
2 = .188; and
there was a significant interaction, F(1, 35) = 14.23, p <
.001, ηp
2 = .289. This interaction reflects similar boundary
extension across conditions in the first session ( WIN =
−.13, PIC = −.14), whereas in the second session, there
was stronger boundary extension for the WIN condition
( WIN = −.16, PIC = −.02). We believe that this session
interaction may be a consequence of a counterbalancing
error—an issue that we further address next.
As mentioned in Methods, a technical error meant that
the stimuli were not balanced across sessions or partici-
pants. Scenes were split into two static groups (A and B)
across all participants. Group A was always shown first in
the WIN condition, and Group B was always presented first
in the PIC condition. To examine whether this contributed
to the observed interactions, we performed an item anal-
ysis to investigate whether specific scenes consistently
elicited greater boundary extension regardless of condi-
tion. Or critically, whether the “same scene” elicits greater
boundary extension in the WIN condition as compared
with the PIC condition. In this item analysis, we replicated
the overall effect of boundary extension across all stimuli
and all conditions, mean = −.11, t(199) = −4.15, p <
.00005, as well as a greater boundary extension effect for
each scene in the WIN condition as compared with the PIC
condition (WIN = −.14, PIC = −.08), t(199) = 2.969, p <
.003. To rule out an effect driven by specific scenes, we com-
pared the boundary extension of Group B—presented in
the second session in the WIN condition—with Group A.
When collapsing across the PIC and WIN conditions, both
Groups A and B showed an overall boundary extension ef-
fect (A = −.08, B = −.15; no significant difference), t(99) =
1.438, p = .15, indicating that our observed context manip-
ulation effects were not the result of any imbalance in which
scenes appeared in which condition, but rather the result of
the manipulation itself. However, Group B did elicit greater
overall boundary extension (even in the PIC condition, al-
though, critically, still greater for the WIN condition), which
may have reduced the difference between PIC and WIN ob-
served in the first presentation, yielding the significant in-
teraction with session mentioned above. Overall, the item
analysis provides further evidence that functional context
affects how scenes are processed and perceived.
DISCUSSION
Rapid scene understanding is often construed as a feedfor-
ward process in which category-preferred neural substrates
are mandatorily recruited. At the same time, there is clear
evidence for high-level properties influencing scene per-
ception (Biederman, Mezzanotte, & Rabinowitz, 1982;
Biederman, 1981). We built on the idea of high-level knowl-
edge influencing scene processing by asking whether
the functional context in which a given scene is viewed
(as opposed to the scene content in and of itself ) affects
scene perception. To address this question, we examined
whether there is a difference in scene-selective neural
responses when viewing a scene through a window as
compared with in a picture frame. We found that two
scene-preferring regions of the brain, the OPA and the
PPA, respond differently when otherwise identical scenes
are viewed in these two contexts. Consistent with the
conception of these brain regions supporting real-world
scene understanding, the more ecologically valid context,
940
Journal of Cognitive Neuroscience
Volume 33, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
5
9
3
3
1
9
5
9
4
5
5
/
j
o
c
n
_
a
_
0
1
6
9
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
through a window, elicited stronger neural responses as
compared with the more artificial context, in a picture
frame. These results support the proposal that high-level,
top–down knowledge—even extraneous to the scene
content—influences scene processing. We posit that this
effect arises as a result of the window context triggering
a set of task-related expectations with respect to scenes
that modulate the manner in which the visual system pro-
cesses incoming scene information.
Why should the context specified by the frame affect
how we process scenes? In both conditions, each scene
is a 2-D picture that participants are viewing on a screen.
It seems highly unlikely that participants perceive the
window-framed picture as if it were a real scene being
viewed through a window (e.g., eventually seeing some-
thing move in the scenery). At the same time, statistical
inference plays an important role in perception, and a
variety of associations may automatically come into play
because they are coupled with specific features (i.e.,
window frames). In our present experiment, we are capi-
talizing on such statistical regularities—in this case, those
that give rise to specific functional contexts and spatial
affordances. For example, previous studies have demon-
strated differences in neural adaptation between the pro-
cessing of 2-D pictures and 3-D real-world objects (Snow
et al., 2011). However, Snow et al.’s (2011) study directly
compared physical stimuli and pictorial stimuli—as such,
there may be a variety of low- and mid-level visual cues,
along with high-level inferences, that differed between
their two presentation conditions. In contrast, the only
differences between our presentation conditions would
be carried by the frames rather than the images them-
selves (which were identical). Although it is possible—
particularly in light of the differences in processing seen
in Snow et al.’s study—that real-world stimuli would have
prompted different results, the differences we observe in
our presentation conditions must arise from either low-
level image differences in the frames or high-level infer-
ences about the frames that impact the processing of
the contained scenes. We have tried to rule out the former
and suggest that the latter is our preferred explanation. In
this light, we argue that further research with physical
stimuli may be needed to better characterize differences be-
tween perceiving 2-D and 3-D scenes (Snow et al., 2011,
used object, not scene, stimuli). We do note that one
way to address this issue is to examine whether our
presentation manipulation has a behavioral effect,
which would lend credence to the ecological validity
of the manipulation—a question we address in the next
section.
To better understand the functional impact of this neu-
ral processing difference, we examined how viewing
scenes in windows and picture frames affects scene mem-
ory. More specifically, we explored whether boundary ex-
tension, a memory phenomenon associated with scene
processing in which observers tend to remember scenes
as wider than as actually presented, would be modulated
by functional context. We predicted that boundary ex-
tension would be greater for those scenes presented
in window frames relative to those scenes presented
in picture frames because of the more ecologically valid
context afforded by windows. Our results were consistent
with this prediction, demonstrating stronger boundary
extension for scenes appearing in a window. Overall,
we find support for the view that the functional context
in which we view scenes can alter the perceived realism
and the spatial cognitive affordances of those scenes
(e.g., the multisource model; Intraub, 2010), thereby
influencing the manner in which they are perceptually
processed—an effect seen in both the magnitude of
scene-preferred neural responses and the level of distor-
tion of scene memories.
More broadly, scene-selective brain regions and mental
processes are not simply responding to inputs that fall
within their preferred domain. Instead, scene-preferred
responses reflect some interplay between bottom–up
and top–down information, including the associations/
expectations that observers have formed about visual
categories over their lifetimes. We posit that the responses
of other category-preferred regions similarly reflect both
feedforward and feedback processing (e.g., Hebart,
Bankson, Harel, Baker, & Cichy, 2018; Brandman &
Peelen, 2017; Vaziri-Pashkam & Xu, 2017; Çukur et al.,
2016; Kaiser, Oosterhof, & Peelen, 2016; Kok, Brouwer,
van Gerven, & de Lange, 2013; Yi & Chun, 2005).
We next turn to ask why the OPA and the PPA, but not
the RSC, are sensitive to functional context. How might we
account for higher neural responses for the window frame
context as compared with the picture frame context for
these two regions? Recent reports indicate that scene se-
lectivity within the OPA reflects the processing of spatial
properties. For example, the OPA was found to preferen-
tially process scene boundaries and geometry relative to
other properties such as landmarks ( Julian et al., 2016).
The OPA has also been found to process not just spatial
information per se but spatial information that carries as-
sociative content (i.e., explicit coding of spatial relations
within a scene and their relevance to a broader context;
Aminoff & Tarr, 2015). Under this view, spatial properties
such as boundaries not only help define a scene as a scene
but also provide task-relevant information as to how an ob-
server might navigate within their perceived environment.
Reinforcing this claim, the OPA has also been associated
with the position of the observer within an environment
(Sulpizio, Committeri, Lambrey, Berthoz, & Galati, 2013)
and with navigational affordances—information about
where one can and cannot move in a local environment
(Bonner & Epstein, 2017).
At an even finer grain, there is evidence that the OPA
is not a singular functional area but is actually composed
of at least two distinct functional regions: the OPA and
the caudal inferior parietal lobule (cIPL). Baldassano,
Esteva, Fei-Fei, and Beck (2016) argue that the OPA is tied
to perceptual systems, whereas the cIPL is tied to
Aminoff and Tarr
941
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
5
9
3
3
1
9
5
9
4
5
5
/
j
o
c
n
_
a
_
0
1
6
9
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
memory systems. Although our functional ROIs did not
distinguish between the OPA and cIPL, our whole-brain
analysis suggests that higher responses for the window
frame context were localized to more dorsal regions that
may include or overlap with the cIPL. We posit that the
activation observed in these regions may be related to ex-
pectations arising from top–down information derived
from memories of viewing scenes through windows.
Such expectations facilitate task-related scene processing
by biasing the observer to scene properties relevant to the
local environment, for example, navigational affordances
or scene boundaries. Supporting this view, in our behav-
ioral experiment, we observed a boundary extension
effect—remembering scene images with wider boundaries
than were originally presented—when scene images were
placed within a window frame. One possibility is that the
perception and representation of scenes with wider
boundaries may account for some of the differential activ-
ity we observe within the OPA.
As with the OPA, we observed that a second scene-
preferred region, the PPA, is also sensitive to functional
context. The PPA is sensitive to high-level associative
scene content (Marchette et al., 2015; Aminoff & Tarr,
2015; Mégevand et al., 2014; Aminoff, Kveraga, & Bar,
2013; Diana, Yonelinas, & Ranganath, 2012; Troiani
et al., 2014; Cant & Goodale, 2011; Peters, Daum,
Gizewski, Forsting, & Suchan, 2009; Rauchs et al., 2008).
We speculate that the larger neural responses observed
for the window frame context reflect stronger associa-
tions arising from the more realistic nature of the experi-
ence. That is, scenes viewed through windows are more
likely to be perceived as “real” scenes and therefore more
likely to prompt the kinds of associations one experi-
ences in day-to-day life. In contrast, scenes viewed within
picture frames are understood to be depictions of scenes
and less likely to be perceived as real. To the extent that
the PPA is involved in bringing associative content, in-
cluding associations, experiences, and expectations, to
bear in scene perception, the more likely it is that the
PPA will be engaged to a greater extent for the window
frame context.
One caution is that, in our whole-brain analysis, the PPA
did not demonstrate significant differential activity across
context conditions. One possibility is that this lack of an
effect may be a consequence of individual differences
as to where within the PPA any differential activity was elic-
ited. The PPA processes information differentially based
on type of information; spatial information is biased to
posterior regions, whereas nonspatial information is bi-
ased to anterior regions (Baldassano, Esteva, et al., 2016;
Aminoff & Tarr, 2015; Aminoff, Gronau, & Bar, 2007).
Across individuals, the difference between context condi-
tions may be driven more by differences in the perception
of the spatial properties of the scene and therefore recruit
more posterior regions of the PPA, whereas in other indi-
viduals, the difference may be driven more by functional
properties and semantics of the scene (e.g., viewing a
picture vs. being within the scene) and recruit more ante-
rior regions of the PPA.
Finally, another scene-preferring region, the RSC, did
not show any effects of our context manipulation. The
RSC is believed to process nonperceptual aspects of
scenes that are involved in defining higher-order properties
such as strong contextual objects (Aminoff & Tarr, 2015;
Bar & Aminoff, 2003); landmarks (e.g., Auger et al.,
2012); or abstract, content-related episodic and autobio-
graphical scene memories (Baldassano, Esteva, et al.,
2016; Aminoff, Schacter, & Bar, 2008; Addis, Wong, &
Schacter, 2007). Reinforcing the idea that the RSC is in-
volved in more abstract aspects of scene processing, RSC
responses to scenes are typically tolerant of shallow ma-
nipulations of the stimulus (Mao, Kandler, McNaughton,
& Bonin, 2017). Similarly, the RSC generalizes across mul-
tiple views (e.g., Park & Chun, 2009), including indoor and
outdoor views of specific places (Marchette et al., 2015).
Such findings suggest that the RSC processes scenes
abstracted away from their physical properties, that is, in
terms of scene content and how this content relates to
high-level properties of scenes encoded in memory.
Given that our context manipulation focused on task-
relevant inferences regarding scene structure, but not
scene content, the lack of an effect of functional context
in the RSC is consistent with this characterization. That
is, irrespective of how one might interact with a scene,
its high-level identity remains constant.
In summary, we demonstrate that top–down informa-
tion modulates both the way the OPA and the PPA process
and represent scenes and how observers remember
scenes. In contrast, the RSC appears to be independent
of this process, encoding a high-level representation of
scene content that is not influenced by presentation con-
text. Such results add to our understanding of the different
roles of the OPA, PPA, and RSC in scene processing. More
generally, our results demonstrate that responses in
category-preferred brain regions do not arise solely from
the processing of inputs within their preferential domains,
but rather integrate high-level knowledge into their pro-
cessing. Both feedforward and feedback pathways appear
to play an important role in categorical perception and, in
particular, in the specific neural substrates that support
scene understanding.
Acknowledgments
We thank Alyssa Shannon for her work in the boundary extension
experiment.
Reprint requests should be sent to Elissa M. Aminoff, Department
of Psychology, Fordham University, Dealy Hall 332, 441
E. Fordham Rd., Bronx, NY 10458, or via e-mail: eaminoff
@fordham.edu.
Author Contributions
Elissa M. Aminoff: Conceptualization; Data curation; Formal
analysis; Writing—Original draft; Writing—Review & editing.
942
Journal of Cognitive Neuroscience
Volume 33, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
5
9
3
3
1
9
5
9
4
5
5
/
j
o
c
n
_
a
_
0
1
6
9
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Michael J. Tarr: Conceptualization; Formal analysis; Writing
—Original draft; Writing—Review & editing.
Funding Information
Elissa M. Aminoff, National Science Foundation (http://dx
.doi.org/10.13039/100000001), grant number: 1439237.
Diversity in Citation Practices
A retrospective analysis of the citations in every article
published in this journal from 2010 to 2020 has revealed a
persistent pattern of gender imbalance: Although the pro-
portions of authorship teams (categorized by estimated
gender identification of first author/last author) publishing
in the Journal of Cognitive Neuroscience ( JoCN) during
this period were M(an)/M = .408, W(oman)/M = .335,
M/W = .108, and W/W = .149, the comparable proportions
for the articles that these authorship teams cited were
M/M = .579, W/M = .243, M/W = .102, and W/W = .076
(Fulvio et al., JoCN, 33:1, pp. 3–7). Consequently, JoCN
encourages all authors to consider gender balance explicitly
when selecting which articles to cite and gives them the
opportunity to report their article’s gender citation balance.
Notes
“Functional properties” denotes high-level knowledge of
1.
how a visual stimulus is used and how it interacts with the envi-
ronment (including other objects and people).
2. The participants of this study were also part of a study dis-
cussed in Yang, Tarr, Kass, and Aminoff (2019), and thus, the
localizer data used here is common with the localizer data
described in that paper.
REFERENCES
Addis, D. R., Wong, A. T., & Schacter, D. L. (2007). Remembering
the past and imagining the future: Common and distinct
neural substrates during event construction and elaboration.
Neuropsychologia, 45, 1363–1377. DOI: https://doi.org/10
.1016/j.neuropsychologia.2006.10.016, PMID: 17126370,
PMCID: PMC1894691
Aminoff, E. M., Gronau, N., & Bar, M. (2007). The parahippocampal
cortex mediates spatial and nonspatial associations. Cerebral
Cortex, 17, 1493–1503. DOI: https://doi.org/10.1093/cercor
/bhl078, PMID: 16990438
Aminoff, E. M., Kveraga, K., & Bar, M. (2013). The role of the
parahippocampal cortex in cognition. Trends in Cognitive
Sciences, 17, 379–390. DOI: https://doi.org/10.1016/j.tics
.2013.06.009, PMID: 23850264, PMCID: PMC3786097
Aminoff, E. M., Schacter, D. L., & Bar, M. (2008). The cortical
underpinnings of context-based memory distortion. Journal
of Cognitive Neuroscience, 20, 2226–2237. DOI: https://doi
.org/10.1162/jocn.2008.20156, PMID: 18457503, PMCID:
PMC3786095
Aminoff, E. M., & Tarr, M. J. (2015). Associative processing
is inherent in scene perception. PLoS One, 10, e0128840.
DOI: https://doi.org/10.1371/journal.pone.0128840, PMID:
26070142, PMCID: PMC4467091
Auger, S. D., Mullally, S. L., & Maguire, E. A. (2012). Retrosplenial
cortex codes for permanent landmarks. PLoS One, 7, e43620.
DOI: https://doi.org/10.1371/journal.pone.0043620, PMID:
22912894, PMCID: PMC3422332
Bainbridge, W. A., & Baker, C. I. (2020). Boundaries extend and
contract in scene memory depending on image properties.
Current Biology, 30, 537–543. DOI: https://doi.org/10.1016
/j.cub.2019.12.004, PMID: 31983637, PMCID: PMC7187786
Baldassano, C., Esteva, A., Fei-Fei, L., & Beck, D. M. (2016). Two
distinct scene-processing networks connecting vision and
memory. eNeuro, 3, ENEURO.0178-16.2016. DOI: https://
doi.org/10.1523/ENEURO.0178-16.2016, PMID: 27822493,
PMCID: PMC5075944
Baldassano, C., Fei-Fei, L., & Beck, D. M. (2016). Pinpointing the
peripheral bias in neural scene-processing networks during
natural viewing. Journal of Vision, 16, 9. DOI: https://doi
.org/10.1167/16.2.9, PMID: 27187606
Bar, M. (2004). Visual objects in context. Nature Reviews
Neuroscience, 5, 617–629. DOI: https://doi.org/10.1038
/nrn1476, PMID: 15263892
Bar, M., & Aminoff, E. M. (2003). Cortical analysis of visual
context. Neuron, 38, 347–358. DOI: https://doi.org/10.1016
/S0896-6273(03)00167-3, PMID: 12718867
Bar, M., Aminoff, E., & Schacter, D. L. (2008). Scenes unseen:
The parahippocampal cortex intrinsically subserves contextual
associations, not scenes or places per se. Journal of
Neuroscience, 28, 8539–8544. DOI: https://doi.org/10.1523
/JNEUROSCI.0987-08.2008, PMID: 18716212, PMCID:
PMC2707255
Biederman, I. (1981). On the semantics of a glance at a scene. In
M. Kubovy & J. R. Pomerantz (Eds.), Perceptual organization
(pp. 213–253). Hillsdale, NJ: Erlbaum. DOI: https://doi.org
/10.4324/9781315512372-8
Biederman, I., Mezzanotte, R. J., & Rabinowitz, J. C. (1982).
Scene perception: Detecting and judging objects undergoing
relation violations. Cognitive Psychology, 14, 143–177. DOI:
https://doi.org/10.1016/0010-0285(82)90007-X, PMID:
7083801
Bonner, M. F., & Epstein, R. A. (2017). Coding of navigational
affordances in the human visual system. Proceedings of the
National Academy of Sciences, U.S.A., 114, 4793–4798. DOI:
https://doi.org/10.1073/pnas.1618228114, PMID: 28416669,
PMCID: PMC5422815
Brainard, D. H. (1997). The Psychophysics Toolbox. Spatial
Vision, 10, 433–436. DOI: https://doi.org/10.1163
/156856897X00357, PMID: 9176952
Brandman, T., & Peelen, M. V. (2017). Interaction between
scene and object processing revealed by human fMRI and
MEG decoding. Journal of Neuroscience, 37, 7700–7710.
DOI: https://doi.org/10.1523/JNEUROSCI.0582-17.2017,
PMID: 28687603, PMCID: PMC6596648
Cant, J. S., & Goodale, M. A. (2011). Scratching beneath the
surface: New insights into the functional properties of the
lateral occipital area and parahippocampal place area.
Journal of Neuroscience, 31, 8248–8258. DOI: https://doi
.org/10.1523/JNEUROSCI.6113-10.2011, PMID: 21632946,
PMCID: PMC6622867
Chadwick, M. J., Mullally, S. L., & Maguire, E. A. (2013). The
hippocampus extrapolates beyond the view in scenes: An
fMRI study of boundary extension. Cortex, 49, 2067–2079.
DOI: https://doi.org/10.1016/j.cortex.2012.11.010, PMID:
23276398, PMCID: PMC3764338
Çukur, T., Huth, A. G., Nishimoto, S., & Gallant, J. L. (2016).
Functional subdomains within scene-selective cortex:
Parahippocampal place area, retrosplenial complex, and
occipital place area. Journal of Neuroscience, 36,
10257–10273. DOI: https://doi.org/10.1523/JNEUROSCI
.4033-14.2016, PMID: 27707964, PMCID: PMC5050324
Aminoff and Tarr
943
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
5
9
3
3
1
9
5
9
4
5
5
/
j
o
c
n
_
a
_
0
1
6
9
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Diana, R. A., Yonelinas, A. P., & Ranganath, C. (2012). Adaptation
to cognitive context and item information in the medial
temporal lobes. Neuropsychologia, 50, 3062–3069. DOI:
https://doi.org/10.1016/j.neuropsychologia.2012.07.035,
PMID: 22846335, PMCID: PMC3483447
Dilks, D. D., Julian, J. B., Paunov, A. M., & Kanwisher, N. (2013).
The occipital place area is causally and selectively involved in
scene perception. Journal of Neuroscience, 33, 1331–1336.
DOI: https://doi.org/10.1523/JNEUROSCI.4081-12.2013,
PMID: 23345209, PMCID: PMC3711611
Epstein, R. A., & Baker, C. I. (2019). Scene perception in the
human brain. Annual Review of Vision Science, 5, 373–397.
DOI: https://doi.org/10.1146/annurev-vision-091718-014809,
PMID: 31226012, PMCID: PMC6989029
Epstein, R. A., & Kanwisher, N. (1998). A cortical representation
of the local visual environment. Nature, 392, 598–601. DOI:
https://doi.org/10.1038/33402, PMID: 9560155
Fang, F., Boyaci, H., Kersten, D., & Murray, S. O. (2008).
Attention-dependent representation of a size illusion in
human V1. Current Biology, 18, 1707–1712. DOI: https://doi
.org/10.1016/j.cub.2008.09.025, PMID: 18993076, PMCID:
PMC2638992
Felleman, D. J., & Van Essen, D. C. (1991). Distributed
hierarchical processing in the primate cerebral cortex.
Cerebral Cortex, 1, 1–47. DOI: https://doi.org/10.1093
/cercor/1.1.1, PMID: 1822724
Greene, M. R., & Oliva, A. (2009). Recognition of natural scenes
from global properties: Seeing the forest without representing
the trees. Cognitive Psychology, 58, 137–176. DOI: https://
doi.org/10.1016/j.cogpsych.2008.06.001, PMID: 18762289,
PMCID: PMC2759758
Harel, A., Kravitz, D. J., & Baker, C. I. (2013). Deconstructing
visual scenes in cortex: Gradients of object and spatial layout
information. Cerebral Cortex, 23, 947–957. DOI: https://
doi.org/10.1093/cercor/bhs091, PMID: 22473894, PMCID:
PMC3593580
Hebart, M. N., Bankson, B. B., Harel, A., Baker, C. I., & Cichy, R. M.
(2018). The representational dynamics of task and object
processing in humans. eLife, 7, e32816. DOI: https://doi.org
/10.7554/eLife.32816, PMID: 29384473, PMCID: PMC5811210
Henderson, J. M., Zhu, D. C., & Larson, C. L. (2011). Functions
of parahippocampal place area and retrosplenial cortex in
real-world scene analysis: An fMRI study. Visual Cognition,
19, 910–927. DOI: https://doi.org/10.1080/13506285.2011
.596852
Intraub, H. (2010). Rethinking scene perception: A multisource
model. In B. H. Ross (Ed.), The psychology of learning and
motivation (Vol. 52, pp. 231–264). Burlington, VT: Academic
Press. DOI: https://doi.org/10.1016/S0079-7421(10)52006-1
Intraub, H. (2014). Visual scene representation: A spatial-
cognitive perspective. In K. Kveraga & M. Bar (Eds.), Scene
vision: Making sense of what we see (pp. 5–26). Cambridge,
MA: MIT Press. DOI: https://doi.org/10.7551/mitpress
/9780262027854.003.0001
Intraub, H. (2020). Searching for boundary extension. Current
Biology, 30, R1463–R1464. DOI: https://doi.org/10.1016
/j.cub.2020.10.031, PMID: 33352122
Intraub, H., & Richardson, M. (1989). Wide-angle memories
of close-up scenes. Journal of Experimental Psychology:
Learning, Memory, and Cognition, 15, 179–187. DOI:
https://doi.org/10.1037/0278-7393.15.2.179
Janzen, G., & van Turennout, M. (2004). Selective neural
representation of objects relevant for navigation. Nature
Neurosciece, 7, 673–677. DOI: https://doi.org/10.1038
/nn1257, PMID: 15146191
Julian, J. B., Ryan, J., Hamilton, R. H., & Epstein, R. A. (2016).
The occipital place area is causally involved in representing
environmental boundaries during navigation. Current
Biology, 26, 1104–1109. DOI: https://doi.org/10.1016/j.cub
.2016.02.066, PMID: 27020742, PMCID: PMC5565511
Kaiser, D., Oosterhof, N. N., & Peelen, M. V. (2016). The neural
dynamics of attentional selection in natural scenes. Journal
of Neuroscience, 36, 10522–10528. DOI: https://doi.org/10
.1523/JNEUROSCI.1385-16.2016, PMID: 27733605, PMCID:
PMC6601932
Kauffmann, L., Ramanoël, S., & Peyrin, C. (2014). The neural
bases of spatial frequency processing during scene perception.
Frontiers in Integrative Neuroscience, 8, 37. DOI: https://doi
.org/10.3389/fnint.2014.00037, PMID: 24847226, PMCID:
PMC4019851
Kay, K. N., & Yeatman, J. D. (2017). Bottom–up and top–down
computations in word- and face-selective cortex. eLife, 6,
e22341. DOI: https://doi.org/10.7554/eLife.22341, PMID:
28226243, PMCID: PMC5358981
Kok, P., Brouwer, G. J., van Gerven, M. A. J., & de Lange, F. P.
(2013). Prior expectations bias sensory representations in
visual cortex. Journal of Neuroscience, 33, 16275–16284.
DOI: https://doi.org/10.1523/JNEUROSCI.0742-13.2013,
PMID: 24107959, PMCID: PMC6618350
Kravitz, D. J., Peng, C. S., & Baker, C. I. (2011). Real-world scene
representations in high-level visual cortex: It’s the spaces
more than the places. Journal of Neuroscience, 31,
7322–7333. DOI: https://doi.org/10.1523/JNEUROSCI.4588
-10.2011, PMID: 21593316, PMCID: PMC3115537
Lamme, V. A., & Roelfsema, P. R. (2000). The distinct modes
of vision offered by feedforward and recurrent processing.
Trends in Neurosciences, 23, 571–579. DOI: https://doi
.org/10.1016/S0166-2236(00)01657-X
Lescroart, M. D., & Gallant, J. L. (2019). Human scene-selective
areas represent 3D configurations of surfaces. Neuron, 101,
178–192. DOI: https://doi.org/10.1016/j.neuron.2018.11.004,
PMID: 30497771
Levy, I., Hasson, U., Avidan, G., Hendler, T., & Malach, R.
(2001). Center-periphery organization of human object areas.
Nature Neuroscience, 4, 533–539. DOI: https://doi.org/10
.1038/87490, PMID: 11319563
Lowe, M. X., Rajsic, J., Gallivan, J. P., Ferber, S., & Cant, J. S.
(2017). Neural representation of geometry and surface
properties in object and scene perception. Neuroimage, 157,
586–597. DOI: https://doi.org/10.1016/j.neuroimage
.2017.06.043, PMID: 28647484
Maguire, E. A. (2001). The retrosplenial contribution to human
navigation: A review of lesion and neuroimaging findings.
Scandinavian Journal of Psychology, 42, 225–238. DOI:
https://doi.org/10.1111/1467-9450.00233, PMID: 11501737
Mao, D., Kandler, S., McNaughton, B. L., & Bonin, V. (2017).
Sparse orthogonal population representation of spatial
context in the retrosplenial cortex. Nature Communications,
8, 243. DOI: https://doi.org/10.1038/s41467-017-00180-9,
PMID: 28811461, PMCID: PMC5557927
Marchette, S. A., Vass, L. K., Ryan, J., & Epstein, R. A. (2015).
Outside looking in: Landmark generalization in the human
navigational system. Journal of Neuroscience, 35, 14896–14908.
DOI: https://doi.org/10.1523/JNEUROSCI.2270-15.2015, PMID:
26538658, PMCID: PMC4635136
Mégevand, P., Groppe, D. M., Goldfinger, M. S., Hwang, S. T.,
Kingsley, P. B., Davidesco, I., et al. (2014). Seeing scenes:
Topographic visual hallucinations evoked by direct electrical
stimulation of the parahippocampal place area. Journal of
Neuroscience, 34, 5399–5405. DOI: https://doi.org/10.1523
/JNEUROSCI.5202-13.2014, PMID: 24741031, PMCID:
PMC6608225
Nasr, S., & Tootell, R. B. H. (2012). A cardinal orientation bias in
scene-selective visual cortex. Journal of Neuroscience, 32,
14921–14926. DOI: https://doi.org/10.1523/JNEUROSCI
.2036-12.2012, PMID: 23100415, PMCID: PMC3495613
944
Journal of Cognitive Neuroscience
Volume 33, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
5
9
3
3
1
9
5
9
4
5
5
/
j
o
c
n
_
a
_
0
1
6
9
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Oliva, A., & Torralba, A. (2001). Modeling the shape of the
scene: A holistic representation of the spatial envelope.
International Journal of Computer Vision, 42, 145–175.
DOI: https://doi.org/10.1023/A:1011139631724
Park, S., Brady, T. F., Greene, M. R., & Oliva, A. (2011).
Disentangling scene content from spatial boundary:
Complementary roles for the parahippocampal place area
and lateral occipital complex in representing real-world
scenes. Journal of Neuroscience, 31, 1333–1340. DOI:
https://doi.org/10.1523/JNEUROSCI.3885-10.2011, PMID:
21273418, PMCID: PMC6623596
Park, S., & Chun, M. M. (2009). Different roles of the
parahippocampal place area (PPA) and retrosplenial cortex
(RSC) in panoramic scene perception. Neuroimage, 47,
1747–1756. DOI: https://doi.org/10.1016/j.neuroimage.2009
.04.058, PMID: 19398014, PMCID: PMC2753672
Park, S., Intraub, H., Yi, D.-J., Widders, D., & Chun, M. M.
(2007). Beyond the edges of a view: Boundary extension in
human scene-selective visual cortex. Neuron, 54, 335–342.
DOI: https://doi.org/10.1016/j.neuron.2007.04.006, PMID:
17442252
Park, S., Konkle, T., & Oliva, A. (2015). Parametric coding of
the size and clutter of natural scenes in the human brain.
Cerebral Cortex, 25, 1792–1805. DOI: https://doi.org/10.1093
/cercor/bht418, PMID: 24436318, PMCID: PMC4459284
Peters, J., Daum, I., Gizewski, E., Forsting, M., & Suchan, B.
(2009). Associations evoked during memory encoding recruit
the context-network. Hippocampus, 19, 141–151. DOI:
https://doi.org/10.1002/hipo.20490, PMID: 18777560
Rauchs, G., Orban, P., Balteau, E., Schmidt, C., Degueldre, C.,
Luxen, A., et al. (2008). Partially segregated neural networks
for spatial and contextual memory in virtual navigation.
Hippocampus, 18, 503–518. DOI: https://doi.org/10.1002
/hipo.20411, PMID: 18240326
Silson, E. H., Chan, A. W.-Y., Reynolds, R. C., Kravitz, D. J., &
Baker, C. I. (2015). A retinotopic basis for the division of
high-level scene processing between lateral and ventral
human occipitotemporal cortex. Journal of Neuroscience,
35, 11921–11935. DOI: https://doi.org/10.1523/JNEUROSCI
.0137-15.2015, PMID: 26311774, PMCID: PMC4549403
Snow, J. C., Pettypiece, C. E., McAdam, T. D., McLean, A. D.,
Stroman, P. W., Goodale, M. A., et al. (2011). Bringing the real
world into the fMRI scanner: Repetition effects for pictures
versus real objects. Scientific Reports, 1, 130. DOI: https://
doi.org/10.1038/srep00130, PMID: 22355647, PMCID:
PMC3216611
Stevens, W. D., Kahn, I., Wig, G. S., & Schacter, D. L. (2012).
Hemispheric asymmetry of visual scene processing in the
human brain: Evidence from repetition priming and intrinsic
activity. Cerebral Cortex, 22, 1935–1949. DOI: https://doi
.org/10.1093/cercor/bhr273, PMID: 21968568, PMCID:
PMC3388897
Sulpizio, V., Committeri, G., Lambrey, S., Berthoz, A., & Galati, G.
(2013). Selective role of lingual/parahippocampal gyrus and
retrosplenial complex in spatial memory across viewpoint
changes relative to the environmental reference frame.
Behavioral Brain Research, 242, 62–75. DOI: https://doi
.org/10.1016/j.bbr.2012.12.031, PMID: 23274842
Troiani, V., Stigliani, A., Smith, M. E., & Epstein, R. A. (2014).
Multiple object properties drive scene-selective regions.
Cerebral Cortex, 24, 883–897. DOI: https://doi.org/10.1093
/cercor/bhs364, PMID: 23211209, PMCID: PMC3948490
van der Ham, I. J. M., van Zandvoort, M. J. E., Frijns, C. J. M.,
Kappelle, L. J., & Postma, A. (2011). Hemispheric differences
in spatial relation processing in a scene perception task: A
neuropsychological study. Neuropsychologia, 49, 999–1005.
DOI: https://doi.org/10.1016/j.neuropsychologia.2011.02.024,
PMID: 21356223
Van Essen, D. C., Drury, H. A., Dickson, J., Harwell, J., Hanlon,
D., & Anderson, C. H. (2001). An integrated software suite for
surface-based analyses of cerebral cortex. Journal of the
American Medical Informatics Association, 8, 443–459.
DOI: https://doi.org/10.1136/jamia.2001.0080443, PMID:
11522765, PMCID: PMC131042
Vaziri-Pashkam, M., & Xu, Y. (2017). Goal-directed visual
processing differentially impacts human ventral and dorsal
visual representations. Journal of Neuroscience, 37, 8767–8782.
DOI: https://doi.org/10.1523/JNEUROSCI.3392-16.2017, PMID:
28821655, PMCID: PMC5588467
Yang, Y., Tarr, M. J., Kass, R. E., & Aminoff, E. M. (2019). Exploring
spatiotemporal neural dynamics of the human visual cortex.
Human Brain Mapping, 40, 4213–4238. DOI: https://doi.org
/10.1002/hbm.24697, PMID: 31231899, PMCID: PMC6865718
Yi, D.-J., & Chun, M. M. (2005). Attentional modulation of
learning-related repetition attenuation effects in human
parahippocampal cortex. Journal of Neuroscience, 25,
3593–3600. DOI: https://doi.org/10.1523/JNEUROSCI.4677
-04.2005, PMID: 15814790, PMCID: PMC6725381
Aminoff and Tarr
945
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
5
9
3
3
1
9
5
9
4
5
5
/
j
o
c
n
_
a
_
0
1
6
9
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3