FOCUS FEATURE:
Connectivity, Cognition, and Consciousness
Effects of classic psychedelic drugs on turbulent
signatures in brain dynamics
Josephine Cruzat1,2,3
, Yonatan Sanz Perl2, Anira Escrichs2, Jakub Vohryzek2,3,
Christopher Timmermann4, Leor Roseman4, Andrea I. Luppi5,6,7,8, Agustin Ibañez1,9,10,
David Nutt4, Robin Carhart-Harris4,11, Enzo Tagliazucchi1,12, Gustavo Deco2,13,14,15,
and Morten L. Kringelbach3,16,17
1Latin American Brain Health Institute (BrainLat), Universidad Adolfo Ibañez, Santiago, Chile
2Computational Neuroscience Group, Center for Brain and Cognition, Department of Information and Communication
Technologies, Universitat Pompeu Fabra, Barcelona, Catalonia, Spain
3Centre for Eudaimonia and Human Flourishing, Linacre College, University of Oxford, Oxford, United Kingdom
4Centre for Psychedelic Research, Division of Psychiatry, Department of Brain Sciences, Imperial College London,
London, United Kingdom
5Division of Anaesthesia, School of Clinical Medicine, University of Cambridge, Cambridge, United Kingdom
6Department of Clinical Neurosciences, University of Cambridge, Cambridge, United Kingdom
7Leverhulme Centre for the Future of Intelligence, University of Cambridge, Cambridge, United Kingdom
8The Alan Turing Institute, London, United Kingdom
9Cognitive Neuroscience Center (CNC), Universidad de San Andrés, and CONICET, Buenos Aires, Argentina
10Global Brain Health Institute (GBHI), University of California San Francisco (UCSF), San Francisco, CA, USA,
and Trinity College Dublin (TCD), Dublin, Ireland
11Psychedelics Division–Neuroscape, Department of Neurology, University of California San Francisco, San Francisco, CA, USA
12Physics Department, University of Buenos Aires, and Buenos Aires Physics Institute, Buenos Aires, Argentina
13Institució Catalana de la Recerca i Estudis Avancats (ICREA), Barcelona, Spain
14Department of Neuropsychology, Max Planck Institute for Human Cognitive and Brain Sciences, Leipzig, Germany
15School of Psychological Sciences, Turner Institute for Brain and Mental Health, Monash University, Melbourne, VIC, Australia
16Department of Psychiatry, University of Oxford, Oxford, United Kingdom
17Center for Music in the Brain, Department of Clinical Medicine, Aarhus University, Denmark
Keywords: Psychedelics, Psilocybin, LSD, Turbulence
ABSTRACT
Psychedelic drugs show promise as safe and effective treatments for neuropsychiatric disorders,
yet their mechanisms of action are not fully understood. A fundamental hypothesis is that
psychedelics work by dose-dependently changing the functional hierarchy of brain dynamics,
but it is unclear whether different psychedelics act similarly. Here, we investigated the changes in
the brain’s functional hierarchy associated with two different psychedelics (LSD and psilocybin).
Using a novel turbulence framework, we were able to determine the vorticity, that is, the local
level of synchronization, that allowed us to extend the standard global time-based measure of
metastability to become a local-based measure of both space and time. This framework produced
detailed signatures of turbulence-based hierarchical change for each psychedelic drug, revealing
consistent and discriminate effects on a higher level network, that is, the default mode network.
Overall, our findings directly support a prior hypothesis that psychedelics modulate (i.e.,
“compress”) the functional hierarchy and provide a quantification of these changes for two
different psychedelics. Implications for therapeutic applications of psychedelics are discussed.
AUTHOR SUMMARY
Significant progress has been made in understanding the effects of psychedelics on brain
function. One of the main hypotheses is that psychedelics work by changing the functional
a n o p e n a c c e s s
j o u r n a l
Citation: Cruzat, J., Perl, Y. S.,
Escrichs, A., Vohryzek, J.,
Timmermann, C., Roseman, L., Luppi,
A. I., Ibañez, A., Nutt, D., Carhart-Harris,
R., Tagliazucchi, E., Deco, G., &
Kringelbach, M. L. (2022). Effects of
classic psychedelic drugs on turbulent
signatures in brain dynamics. Network
Neuroscience, 6(4), 1104–1124. https://
doi.org/10.1162/netn_a_00250
DOI:
https://doi.org/10.1162/netn_a_00250
Supporting Information:
https://doi.org/10.1162/netn_a_00250
Received: 25 November 2021
Accepted: 6 April 2022
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Authors:
Josephine Cruzat
josecruzat@gmail.com
Morten L. Kringelbach
morten.kringelbach@psych.ox.ac.uk
Handling Editor:
Olaf Sporns
Copyright: © 2022
Massachusetts Institute of Technology
Published under a Creative Commons
Attribution 4.0 International
(CC BY 4.0) license
The MIT Press
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
6
4
1
1
0
4
2
0
5
6
2
8
8
n
e
n
_
a
_
0
0
2
5
0
p
d
.
/
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Turbulent signatures in brain dynamics under psychedelics
hierarchy of brain dynamics in a dose-dependent manner, modulating the encoding of the
precision of priors, beliefs, or assumptions in the brain. We used a novel turbulence framework
to investigate the changes in the brain’s functional hierarchy associated with two different
psychedelics (LSD and psilocybin). This framework produced detailed signatures of
turbulence-based hierarchical change for each psychedelic drug, revealing consistent and
discriminate effects on a higher level network, that is, the default mode network.
INTRODUCTION
In recent years, psychedelic (mind-manifesting) drug research has made a strong comeback
(Nichols, 2016; Pollan, 2019), promising to deliver effective and safe treatments for neuropsychi-
atric disorders such as treatment-resistant depression (Carhart-Harris et al., 2021; Carhart-Harris,
Bolstridge, et al., 2016) and addiction (Bogenschutz et al., 2015; Johnson, Garcia-Romeu, &
Griffiths, 2017). Research so far indicates that the potential benefits far outweigh the risks
(Johnson, Griffiths, Hendricks, & Henningfield, 2018; Johnson, Hendricks, Barrett, & Griffiths,
2019). Yet, a deeper knowledge of how psychedelics function in the human brain is needed to
ensure best use of these compounds.
Here, we use a novel framework to directly characterize the effects of psychedelics on tur-
bulent signatures in brain dynamics and gain insights into one of the leading hypotheses of
how psychedelics function, namely that they work by relaxing hierarchical processing in
the brain. This has been expressed in the REBUS (RElaxed Beliefs Under pSychedelics) and
the anarchic brain hypothesis, which integrates Friston’s free-energy principle (Friston,
2010) with Carhart-Harris’s entropic brain hypothesis (Carhart-Harris, 2018). The authors
hypothesize that psychedelics bring about a relaxation of the precision of high-level priors
or “beliefs” (REBUS), allowing (anarchic) bottom-up information flow.
At the heart of this hypothesis is the idea that psychedelics modulate the encoding of the
precision of priors, beliefs, or assumptions in the brain. This effect could manifest in various
ways, including an alteration (i.e., a reduction) in the power of canonical brain rhythms, and
the integrity of large-scale networks. There is compelling evidence that the action of classic
psychedelics begins with agonism of 5-HT2A receptors (Nichols, 2016). A key functional
effect of 5-HT2A receptor agonism via psychedelics is an increase in the sensitivity of excit-
atory neurons expressing the receptor (e.g., deep-layer pyramidal neurons) resulting in a spike-
wave-decoupling and dysregulation of spontaneous population-level activity. Mapping these
effects to subjective phenomena is a work in progress; however, it is tempting to speculate that
subjective phenomena like ego dissolution (Milliere, 2017; Nour, Evans, Nutt, & Carhart-
Harris, 2016), unitive experiences (Griffiths et al., 2016; Roseman, Nutt, & Carhart-Harris,
2018), and a sense of the ineffable (Pollan, 2019) relate to the relaxation (or “breakdown”)
of particularly core or high-level priors or beliefs, encoded via the functioning of high-level
systems in the brain.
The present paper uses a novel turbulence framework to determine the functional hierarchy
of any brain state, by calculating measures of information processing in the human brain
inspired by turbulence theory (Kolmogorov, 1941b; Kuramoto, 1984) but applied to neurosci-
ence (Sheremet, Qin, Kennedy, Zhou, & Maurer, 2019). Long-standing research in fluid
dynamics has shown that turbulence facilitates the rapid transfer of energy through fluids
(Frisch, 1995; Kolmogorov, 1941b). In terms of brain dynamics, the turbulent core is deter-
mined by the local synchronization between brain areas that, in turn, are linked to the
Turbulence:
In coupled oscillators, turbulence is
characterized by the high variability
across space and time of the local
Kuramoto parameter capturing the
local synchronization, so in this
sense it is a spatiotemporal extension
of the level of metastability.
Functional hierarchy:
General and intrinsic organizing
principle of brain function that arises
because of the progressive
integration of information.
Brain state:
Continuously evolving dynamics of
self-organized, condition-dependent,
system-wide patterns of synchronous
neural activity.
Network Neuroscience
1105
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
6
4
1
1
0
4
2
0
5
6
2
8
8
n
e
n
_
a
_
0
0
2
5
0
p
d
.
t
/
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Turbulent signatures in brain dynamics under psychedelics
Information transfer:
A measure that characterizes how
the information propagates across
space at a particular scale.
Information cascade flow:
A measure that estimates the stream
of information between a given scale
and a subsequent lower scale in
consecutive time steps.
Information cascade:
A measure that captures the entire
efficient information-processing
behavior across scales.
rotational vortices found in fluid dynamics. As such, turbulence has been demonstrated in
large-scale neuroimaging data from healthy participants scanned with functional magnetic res-
onance imaging (fMRI) (Deco & Kringelbach, 2020) and ensures efficient information transfer
(Deco, Kemp, & Kringelbach, 2021; Deco & Kringelbach, 2020). The size of the turbulent
vortices is what defines the different information-processing scales, and it has also been shown
how rare long-range connections enhance information processing throughout the brain (Deco,
Perl, et al., 2021; Escrichs et al., 2021).
In order to determine the psychedelic-specific changes to the functional hierarchy, our
framework employed two complementary model-free and model-based approaches inspired
by the turbulence theory of physics (Frisch, 1995; Kolmogorov, 1941b; Kuramoto, 1984).
Using the model-free approach, we measured information transmission across space and time-
scales using four novel and distinct measures recently introduced by Deco and Kringelbach
(2020): turbulence, information transfer, information cascade flow, and information cascade.
The model-based approach used a whole-brain model to mechanistically explain the statisti-
cal dependencies of the neuronal dynamics observed in the empirical neuroimaging data
(Deco, Cabral, et al., 2017; Deco & Kringelbach, 2020; Deco, Kringelbach, Jirsa, & Ritter,
2017). The model allowed us to study simulated perturbation-elicited changes in global and
local brain activity to assess the system’s susceptibility and information-encoding capability,
offering new insights into the brain’s reactivity to external perturbations, such as perturbations
induced pharmacologically or by electrical stimulations.
We were able to demonstrate that this framework differentiates between the different psy-
chedelics in terms of providing a specific signature of the way they perturb the hierarchy of
brain dynamics. Overall, this provides novel information on how different psychedelics work.
RESULTS
We used a novel turbulence framework (Deco & Kringelbach, 2020) to investigate changes in
brain hierarchy induced by different psychedelic drugs measured with fMRI in healthy volun-
teers from two different datasets. Figure 1 provides a schematized version of the general frame-
work used to determine the functional hierarchy in brain dynamics.
Participants received intravenous injections of lysergic acid diethylamide (LSD) (N = 15) or
psilocybin (N = 9), and placebo in two separate studies (Carhart-Harris, Muthukumaraswamy,
et al., 2016; Carhart-Harris et al., 2012). Experimental designs differed but all were placebo
controlled (saline injection), with participants blinded to condition. Here we focused on the
effects of psychedelics on the brain’s turbulent dynamics based on the recent demonstration
that the brain exhibits a turbulent dynamic intrinsic backbone that facilitates large-scale net-
work communication (Deco & Kringelbach, 2020).
Within the model-free framework, we studied changes in information transmission flow
across spatial and temporal scales using four distinct measures: turbulence, information transfer,
information cascade flow, and information cascade. These analyses were performed based on
the Kuramoto order parameter describing the local degree of synchronization of a brain region,
n, as a function of space, (cid:1)x, and time t, at a given scale λ. The scale of the local synchronization
is defined by the parameter λ, which determines the spatial distance where the synchronization
is assessed. We explored scales ranging from 0.01 (∼100 mm) to 0.21 (∼5 mm), in steps of
0.03, where high values of λ denote short distances in the brain and vice versa.
Compared with placebo, psychedelics induce significant increases in turbulence (Figure 2A). In
the LSD condition, these increases were found across all spatial scales but were more pronounced
Network Neuroscience
1106
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
6
4
1
1
0
4
2
0
5
6
2
8
8
n
e
n
_
a
_
0
0
2
5
0
p
d
/
t
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Turbulent signatures in brain dynamics under psychedelics
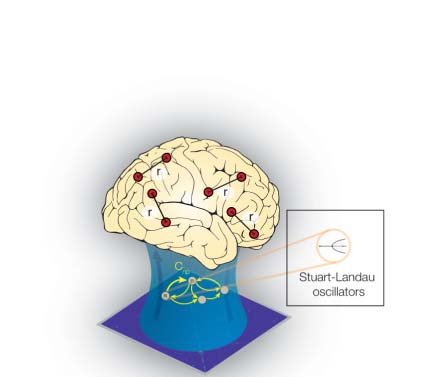
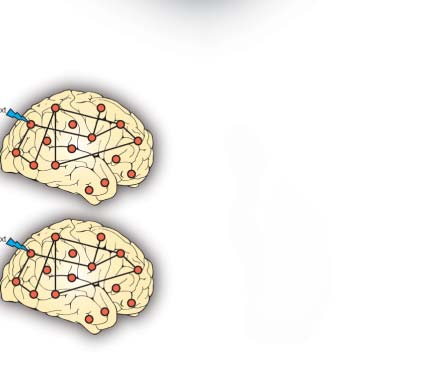
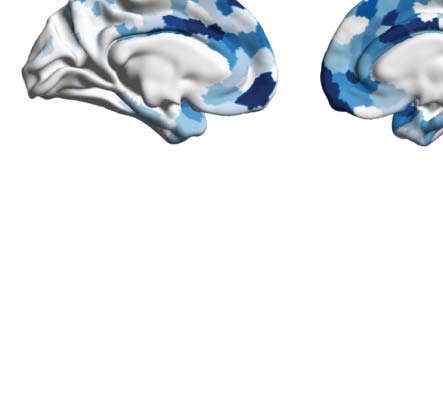
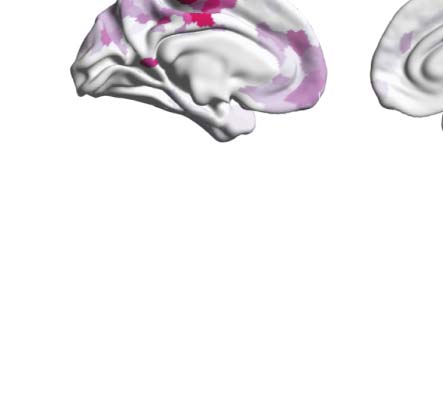
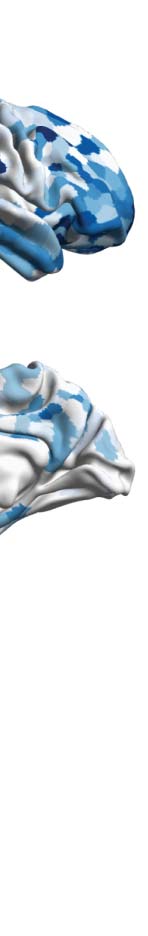
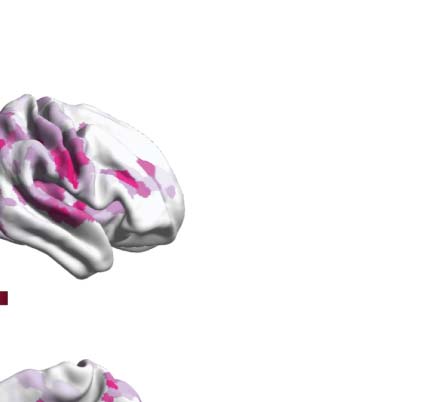
Figure 1. Determining the functional hierarchy in brain dynamics using a turbulence framework. (A) The turbulence framework uses model-
free and model-based measures. The left panels show a schematic of how measures of turbulence, information transfer, information cascade
flow, and information cascade are obtained from estimating the local level of synchronization characterized by the Kuramoto local order
parameter (R; see the Methods section) at different scales. The right panel shows a Hopf whole-brain model that describes the dynamics of
each brain area through a Stuart Landau nonlinear oscillator. The system of local oscillators is connected through the anatomical connectivity
to simulate the global dynamics induced by each psychedelic. Whole-brain modeling allows us to obtain measures that arise from the per-
turbative approach. We simulated external perturbations and evaluated the model’s response for each condition’s brain state by quantifying
the susceptibility and information capacity measures. (B) The brain is hierarchically organized such that information flows from lower sensory
regions to higher association regions (Smallwood et al., 2021). The top left panel sketches how information comes into primary visual ( V1) and
somatosensory (S1) regions and flows through the distinct visual (orange) and somatomotor (yellow) networks before reaching the default mode
network (DMN, light blue). The top right panel renders the seven Yeo resting-state networks on the human brain, which forms the basis for the
hierarchical measure obtained by using the turbulence framework on the two different psychedelics (LSD and psilocybin). The brain plots
show the resulting node-level turbulence for each psychedelic drug. The spider plots at the bottom show how this gives rise to different sig-
natures of the degree of participation of the Yeo resting-state networks for each psychedelic.
at the longer ones. In contrast, the effects in psilocybin were observed only at longer distances in
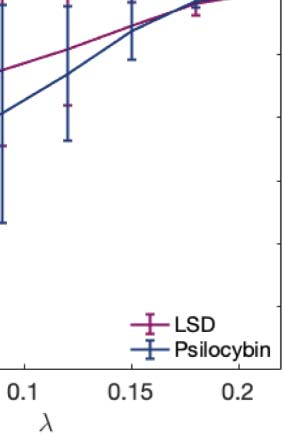
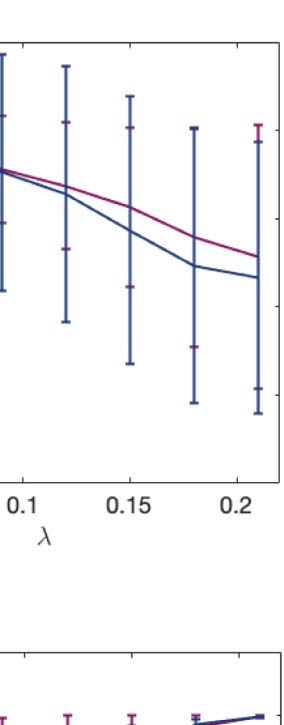
the brain (λ < 0.06). Complementarily, we calculated a linear fit to the mean turbulence at each
λ and captured the slopes of each condition representing its level of turbulence at each spatial
scale. Figure 2B resumes the measures of information transmission through time and space
calculated by changes in turbulence on each different scale. Both psychedelics monotonically
decrease the slope at longer spatial scales.
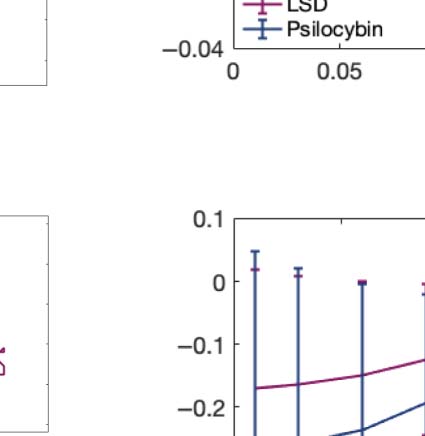
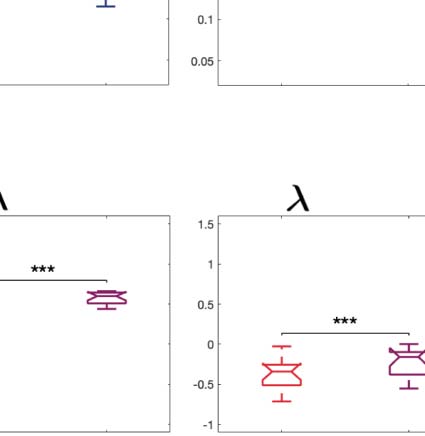
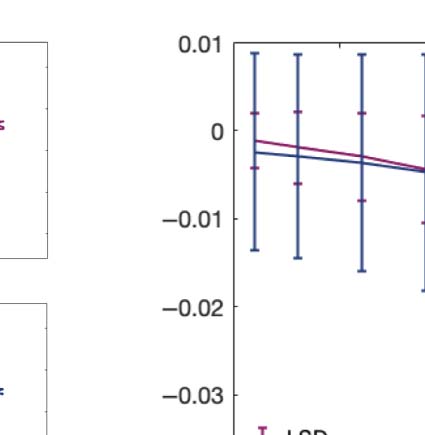
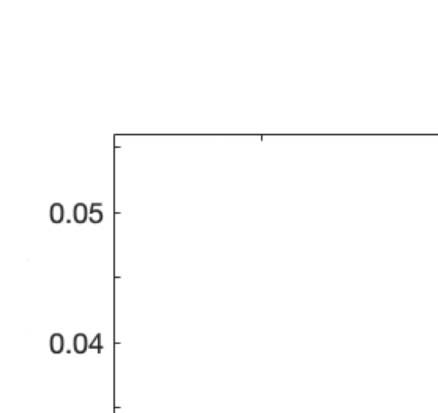
We computed the information transfer measure, which denotes how information travels
across space at a given spatial scale. We found that psychedelics significantly increase infor-
mation transfer across all spatial scales in the brain, favoring information transmission.
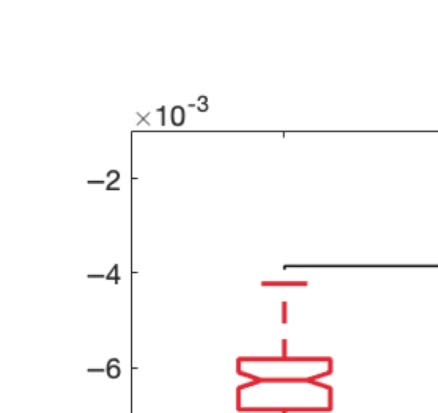
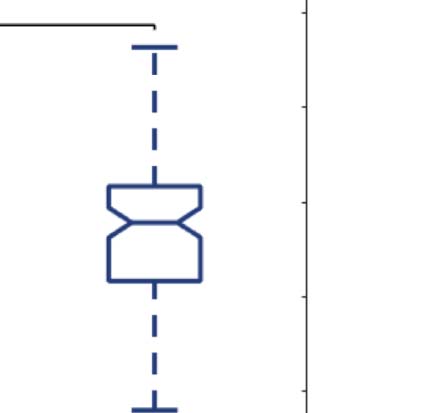
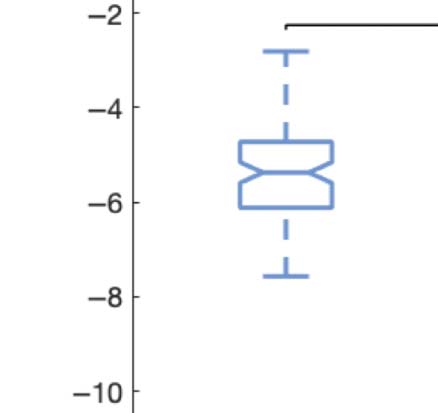
Figure 2C shows boxplots of the statistically significant differences between conditions that
passed the permutation-based paired t test with p < 0.05. We performed the exact computa-
tions presented in Figure 2B, but now for the information cascade measurement. Figure 2D
summarizes the behavior of this measure at different spatial scales. LSD and psilocybin present
a similar slope-scale relationship at shorter spatial scales (larger lambdas), while their differ at
the shorter ones suggesting that psilocybin is more sensitive to differences in the degree of
information transmission across scales.
We then characterized the transmission of information across spatial and temporal scales
using the information cascade flow measurement described as the correlation of the signal at a
given scale λ, with the signal at a lower scale λ − Δλ, in consecutive time steps, t + Δt. As
shown in Figure 3A, we found that LSD significantly increased information cascade flow at all
Network Neuroscience
1107
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
6
4
1
1
0
4
2
0
5
6
2
8
8
n
e
n
_
a
_
0
0
2
5
0
p
d
/
t
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Turbulent signatures in brain dynamics under psychedelics
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
6
4
1
1
0
4
2
0
5
6
2
8
8
n
e
n
_
a
_
0
0
2
5
0
p
d
/
t
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
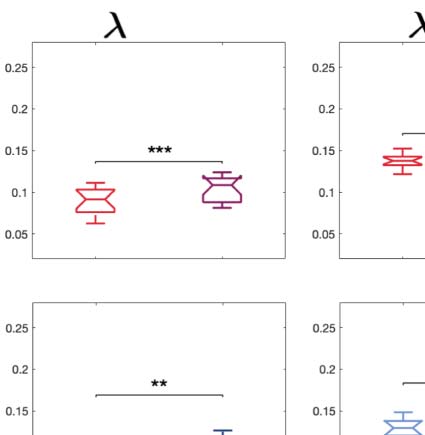
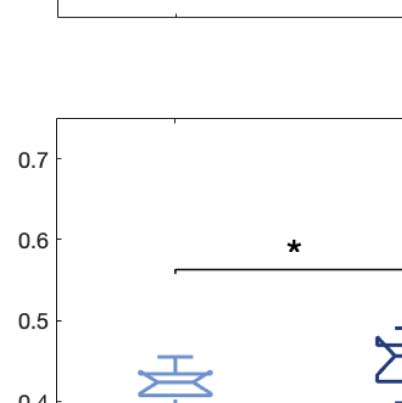
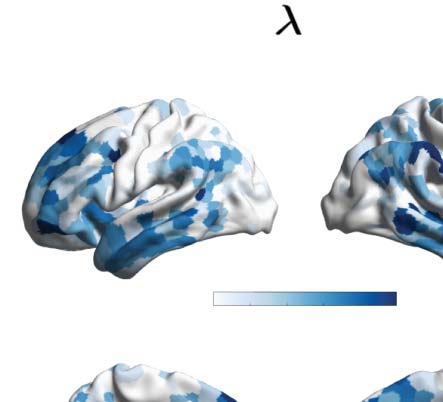
Figure 2. Turbulence and information transfer changes across scales under the psychedelic state. Within the model-free framework, we
investigated the degree to which psychedelics modulate information processing at the whole-brain level, characterized by turbulence and
information transfer measurements. (A) Turbulence estimates the level of local synchronization at different spatial scales by measuring the
spatiotemporal variability of the local Kuramoto order parameter. We calculated the turbulence for spatial scales (λ) ranging from 0.01 to
0.21 in steps of 0.03. Both psychedelics, LSD (above) and psilocybin (below), increase the level of turbulence compared with placebo,
particularly at larger spatial scales (smaller λ). The effect size between statistically significant results was assessed by Cohen’s d. D values
for LSD, respectively: d = 0.9441 (λ = 0.01), d = 0.7013 (λ = 0.09), d = 0.4237 (λ = 0.18). D values for psilocybin, respectively: d =
0.8034 (λ = 0.01), d = 0.5230 (λ = 0.09), d = 0.4451 (λ = 0.18). (B) Complementarily, we computed a linear fitting of the mean level of
turbulence at each spatial scale. The panel shows the slope obtained as a function of λ, and the vertical lines represent the standard deviation
across subjects. The two slopes show a similar pattern, although they slightly differ by showing at higher spatial scales (lower λ). (C) The
spatial information transfer measurement informs how the information travels across space at a specific scale (λ). This measure is defined as
the slope of a linear fitting in log-log scale of the time correlation between the Kuramoto local order parameter of two brain areas at the same
scale as a function of its Euclidean distance (r) within the inertial subrange. Psychedelics significantly increase information transfer at all
spatial scales. The effect size between statistically significant results is clear. D values for LSD, respectively: d = 1.0332 (λ = 0.01), d = 1.0281
(λ = 0.09), d = 0.8809 (λ = 0.18). D values for psilocybin, respectively: d = 0.7920 (λ = 0.01), d = 1.2115 (λ = 0.09), d = 0.9206 (λ = 0.18). (D)
Following the same procedure as in panel B, we computed the linear fit of the information transfer measure. As can be seen, psilocybin differs
from the LSD slope patterns, showing a much steeper slope-scale, denoting higher sensitivity to differences in the degree of transmission
of information across scales. The vertical lines represent the standard deviation across subjects. * Indicates p < 0.05, ** indicates p < 0.01,
and *** indicates p < 0.001.
λ scales compared with placebo, while psilocybin also did so at higher spatial scales, that is,
lower λ denoting long distances in the brain. Finally, we found evidence of significantly
increased information cascade under the psychedelic state compared with placebo (Figure 3B).
This measure characterizes the global degree of information transmission across scales and is
obtained by averaging the information cascade flow across all λ scales.
The increases in turbulence, information transfer, and information cascade observed under
LSD did not correlate with the subjective experience reports. For psilocybin, we observed that
only the increases in information cascade correlated with ratings on ego-dissolution (see the
Supporting Information, Figures S1–S3).
Network Neuroscience
1108
Turbulent signatures in brain dynamics under psychedelics
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
6
4
1
1
0
4
2
0
5
6
2
8
8
n
e
n
_
a
_
0
0
2
5
0
p
d
/
t
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Information flow across scales and information cascade changes under the psychedelic state. (A) The information flow across scales
Figure 3.
outlines the degree of predictability of the synchronization level at a certain scale (λ) given by the precedent scale (λ − Δλ), where Δλ is the
discretization of the scale. LSD significantly increases information flow at all spatial scales, while psilocybin does so only at larger spatial
scales (λ < 0.09). (B) Similarly, the information cascade flow, defined as the average information flow across scales, significantly increases
under the psychedelic state, for both LSD and psilocybin, compared with placebo. * Indicates p < 0.05, ** indicates p < 0.01, and *** indicates
p < 0.001. D values for LSD: d = 1.0437; and psilocybin: d = 0.6566.
Furthermore, we calculated the node variability of the local synchronization defined as the
standard deviation across time of the local Kuramoto order parameter for each condition. We
calculated the similarity of the node-level turbulence between each psychedelic compound
and the counterpart placebo using the Kolmogorov–Smirnov distance (KSD) between them at
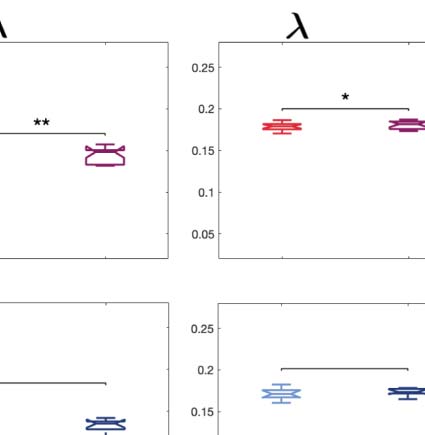
each spatial scale. Figure 4A shows that the difference between conditions is more prominent
at lower λ scales for both psychedelic drugs. For each condition, we then computed the abso-
lute difference of the node-level turbulence between the psychedelic state and the placebo at
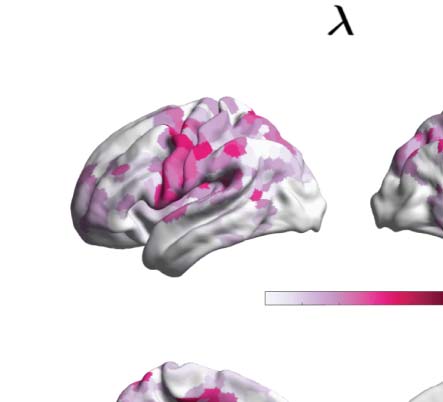
λ = 0.12. Figure 4B renders this absolute difference onto the brain’s cortex. Subsequently, we
picked the nodes for each comparison of the upper 15% quantile, identified the resting-state
network to which they belong, and counted the number of nodes per network. This strategy
allowed us to describe the precise signature of turbulence-based hierarchical change for each
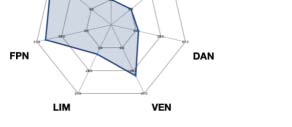
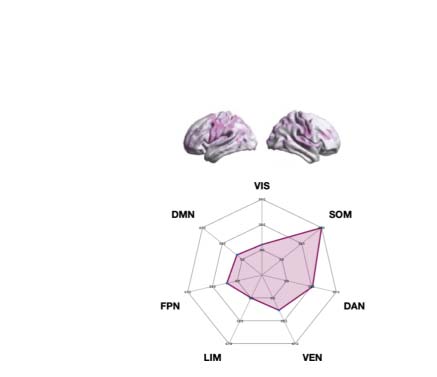
psychedelic drug, revealing how each one impacts the dynamics of the networks. As can be
seen in Figure 4C, differences between LSD and placebo were mainly found in the somato-
motor (SOM) and dorsal attention networks (DAN), and between psilocybin and placebo in
the default mode network (DMN), frontoparietal (FPN), and ventral network ( VEN).
We applied a whole-brain computational modeling approach based on the sensitivity of
these models to react to external in silico perturbations and their capability to offer insights
into the mechanisms underlying the global complexity and dynamical stability of brain activity
(Deco et al., 2018; Jobst et al., 2021) (see the Methods section). For each of the four brain
states (LSD vs. placebo, and psilocybin vs. placebo), we constructed a whole-brain dynamical
model based on the normal form of a supercritical Hopf bifurcation (Deco, Kringelbach, et al.,
2017). The model was coupled to the structural connectivity obtained through diffusion tensor
Network Neuroscience
1109
Turbulent signatures in brain dynamics under psychedelics
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
6
4
1
1
0
4
2
0
5
6
2
8
8
n
e
n
_
a
_
0
0
2
5
0
p
d
t
/
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
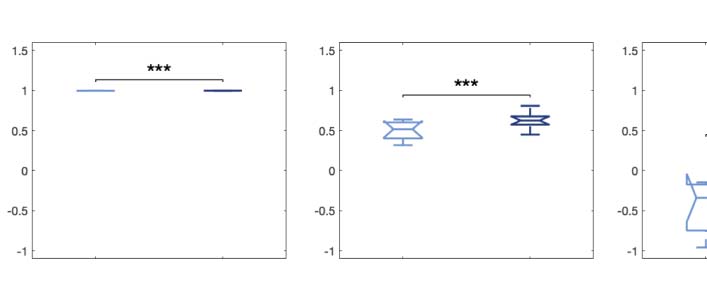
Figure 4. Node-level turbulence. We computed the node variability of the local synchronization defined as the standard deviation across
time of the local Kuramoto order parameter. (A) The panels plot the Kolmogorov–Smirnov distance (KSD) quantifying the difference between
the distributions of the node-level turbulence for each psychedelic and placebo, at each spatial scale. The smaller the KSD values, the more
similar the distributions; therefore, the greatest differences were found at larger spatial scales (lower λ values). (B) The brain schemes show the
absolute difference of the node-level turbulence for λ = 0.12 as an example (shown in yellow in panel A). (C) We then picked the upper 15%
quantile of the absolute difference of the node-level turbulence between conditions, indexed the resting-state network to which they belong,
according to Yeo’s resting-state network, and estimated the number of nodes per network. The spiderweb charts outline the number of nodes in
the higher 15% quantile of the absolute difference for each comparison and network ( VIS: visual; SOM: somatomotor; DAN: dorsal attention
network; VEN: ventral network; LIM: limbic; FPN: frontoparietal network; DMN: default mode network). The networks that showed the most
significant differences between LSD and placebo are the SOM and DAN; and between psilocybin and placebo, the FPN, DMN, and VEN. The
results generalize to the other spatial scales.
imaging and used the exponential distance rule of anatomical connections as a cost-of-wiring
principle (Ercsey-Ravasz et al., 2013; Markov et al., 2013; Markov et al., 2014). Each model
was fitted to optimally reproduce the empirical spatiotemporal dependencies of each brain
state characterized using the functional connectivity (FC) measure as a function of the global
coupling parameter G. For each G and condition, we performed 100 simulations and calcu-
lated the fitting performance as the Euclidean distance between the empirical and the
simulated FC. The minimum distance between the empirical and the simulated FC defines
the optimal operating point of the model. Then, to evaluate how each model-based brain state
reacts to external stimuli, we systematically applied in silico perturbations and quantified the
Network Neuroscience
1110
Turbulent signatures in brain dynamics under psychedelics
functional consequences of these perturbations using the susceptibility and information capa-
bility measures. Perturbations were implemented by randomly changing the bifurcation
parameter of each brain area an in the model within the range [−0.02:0]. The susceptibility
measure reflects the sensitivity of the whole-brain model to react to external perturbations.
It is defined by the difference between the perturbed and unperturbed mean of the modulus
of the local order parameter across time (eRλs
(cid:1)x; t
Þ, respectively), averaged over all
ð
brain nodes. Complementarily, the information capability denotes the capacity of the brain to
encode external perturbations in brain dynamics and is defined as the standard deviation
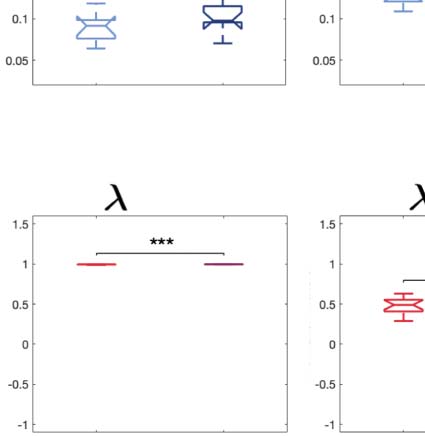
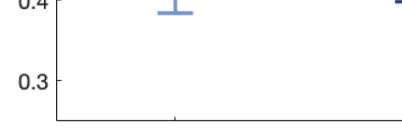
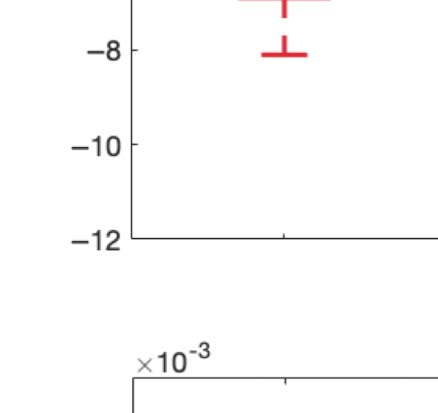
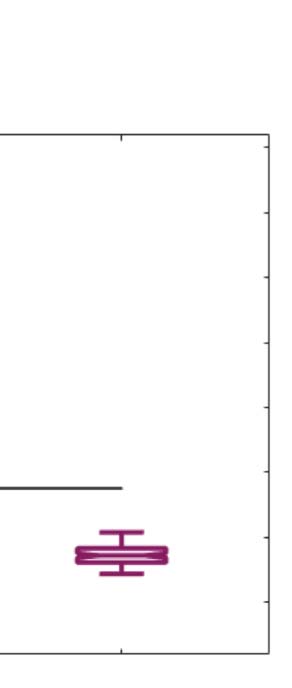
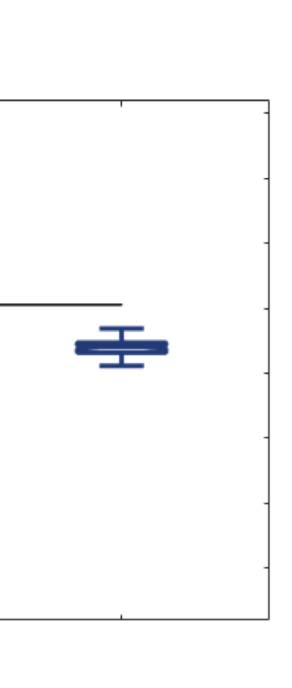
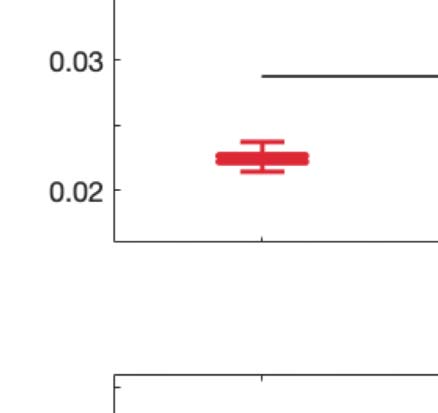
across trials of the susceptibility measure. Figure 5A shows that LSD and psilocybin signifi-
cantly decrease susceptibility. Complementarily, the information-encoding capacity signifi-
cantly increases following the psychedelic drug administration, compared with placebo
(p < 0.001, two-sided Wilcoxon rank sum test). The results for both measures were estimated
at λ = 0.18.
Þ and Rλs
(cid:1)x; t
ð
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
6
4
1
1
0
4
2
0
5
6
2
8
8
n
e
n
_
a
_
0
0
2
5
0
p
d
.
/
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
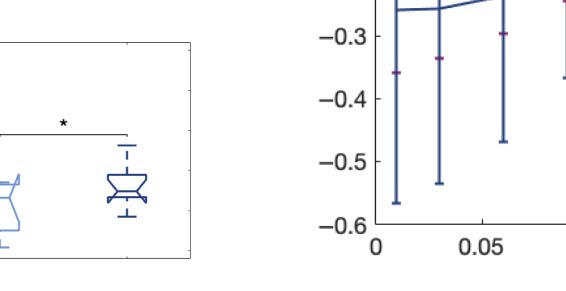
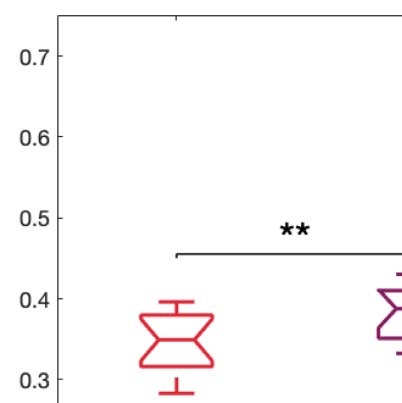
Figure 5. Model-based susceptibility and information capacity changes under the psychedelic state. We used a model-based approach to
assess the effects of external perturbations on brain dynamics. (A) We quantified the susceptibility measure, which estimates the responsiveness
of the in silico whole-brain dynamics to external perturbations. LSD and psilocybin significantly decrease the susceptibility level compared
with placebo. D values for LSD and psilocybin, respectively: d = 1.2522, d = 0.8336. (B) The information capacity measurement denotes the
encoding capability of the whole-brain model. In other words, this appraisal captures how the simulated external perturbations are encoded in
brain dynamics. Both psychedelic drugs significantly increase the information-encoding capacity. D values for LSD and psilocybin, respec-
tively: d = 1.400, d = 2.5615. P values were assessed using the Wilcoxon rank sum test and corrected for multiple comparisons. * Indicates
p < 0.05, ** indicates p < 0.01, and *** indicates p < 0.001.
Network Neuroscience
1111
Turbulent signatures in brain dynamics under psychedelics
DISCUSSION
Here we used two independent fMRI datasets of healthy participants receiving moderate to high
doses of LSD or psilocybin in placebo-controlled designs to determine the psychedelic-specific
modulation of the brain’s dynamic functional hierarchy and assess its impact on information
processing. Our framework comprised model-free and model-based approaches inspired by
signatures of turbulence in complex systems. Using this approach, we aimed to test a leading
hypothesis of the brain action of psychedelics, that is, that they relax the properties of canonical
systems in the brain—encoding internal models or assumptions. Results revealed generally
consistent increases in turbulent signatures across the psychedelics.
Serotonin 2A receptor agonism is a defining pharmacological property of the “classic”
psychedelics—and LSD and psilocybin are perhaps the most familiar examples. At the popu-
lation and systems level, psychedelics have been shown to increase the entropy of spontane-
ous brain activity (Carhart-Harris et al., 2014; Herzog et al., 2020), broaden the repertoire of
connectivity states (Atasoy, Vohryzek, Deco, Carhart-Harris, & Kringelbach, 2018; Tagliazucchi,
Carhart-Harris, Leech, Nutt, & Chialvo, 2014; Varley, Carhart-Harris, Roseman, Menon, &
Stamatakis, 2020), and enhance global connectivity between high-level networks and the rest
of the brain (Carhart-Harris, Muthukumaraswamy, et al., 2016; Tagliazucchi et al., 2016). The
merit of turbulence-related metrics, however, is the information they confer about information
transfer in the brain and their relevance to complexity science—which offers a rich and devel-
oping language for understanding the properties of complex systems.
Using a model-free approach, we first explored the global information transmission char-
acteristics. This approach is grounded in the innovative and sophisticated framework for ana-
lyzing whole-brain dynamics, recently developed by Deco and Kringelbach (2020), through
which they revealed that the healthy human brain shows turbulent dynamics (Frisch, 1995;
Kolmogorov, 1941b; Kuramoto, 1984). Summarizing the results, compared with placebo, both
psychedelics promoted greater information transmission in the brain, in both the spatial and
the temporal domains. Specifically, significant increases were observed in turbulence, infor-
mation transfer across scales, information cascade flow, and information cascade. The present
framework could equally well be described in terms of the local Kuramoto order parameter
and its use for characterizing information transmission, that is, as a generalization of the con-
cept of metastability in neuroscience, pioneered by Shanahan and Kuramoto (Kuramoto,
1984; Wildie & Shanahan, 2012). Nevertheless, such a description does not capture the
generality of the principles governing brain dynamics and most, if not all, physical systems.
The turbulence framework offers a characterization of the dynamics underlying different
brain states and provides a principled, mechanistic way to describe information transmission
across spacetime.
Thus, irrespective of the type of psychedelic, the psychedelic state appears to show a clear
trend towards a greater transmission of information—characterized by increases in turbulence
and information transfer values—through long-range spatial scales, that is, smaller lambdas.
Long-range cortical connections have long been thought to be crucial for the emergence of
consciousness. In fact, it has been shown that long-range connections are associated with
enhanced cognitive processing in healthy participants (Deco, Perl, et al., 2021), whereas a
decrease in global information processing and large-scale functional connectivity appears to
be a robust signature of reduced conscious levels (Bourdillon et al., 2020; King et al., 2013;
Vatansever et al., 2020). These findings suggest that the recruitment of long-distance connec-
tions plays a fundamental role in the flow of information through the cortex, allowing
communication between different brain areas and ultimately supporting the emergence of
Metastability:
Fundamental concept describing
complex systems’ behavior in terms
of the variability in the global state of
synchronization as a function
of time.
Network Neuroscience
1112
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
6
4
1
1
0
4
2
0
5
6
2
8
8
n
e
n
_
a
_
0
0
2
5
0
p
d
/
.
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Turbulent signatures in brain dynamics under psychedelics
conscious awareness. It is tempting to speculate that increased long-range information transfer
under psychedelics relates to their purported ability to facilitate psychological insight (Yaden
& Griffiths, 2020)—consistent with the etymology of the term “psychedelic”—that is,
“mind-revealing” (Hartogsohn, 2018; Nichols, 2016; Osmond, 1957).
We explored node-level changes in turbulence to identify the primary brain areas driving
the turbulent dynamic core and thus involved in changing the whole-brain dynamics under-
lying each psychedelic state. Noteworthily, we found that brain regions belonging to the soma-
tomotor and dorsal attention network showed the most critical differences in the LSD-induced
brain state; and for psilocybin the default mode, frontoparietal and ventral networks showed
the greatest differences. These results highlight the specific functional changes to the func-
tional hierarchy, revealing that psilocybin directly impacts high-level networks, whereas
LSD has stronger effects on primary visual-sensory areas. Given that the level of turbulence
is increased for both LSD and psilocybin at the different spatial scales, we speculate that
changes in the setting such as music may increase turbulence and therefore increase the
observed effect. However, this is out of the scope of the present paper and deserves thorough
investigation.
Furthermore, we used a model-based approach to investigate how whole-brain dynamics
underlying each brain state impact the brain’s capacity to encode external perturbations. To do
so, we built a whole-brain model based on the normal form of a supercritical Hopf bifurcation
(Deco, Kringelbach, et al., 2017), simulating the empirical fMRI statistical dependencies. We
applied in silico external perturbations to evaluate how each model-based brain state reacts to
(simulated) external perturbations. We found that the brain is less sensitive to external pertur-
bations under psychedelics as shown by the susceptibility measure. Conversely, consistent
increases in the brain’s information-encoding capacity were observed across all psychedelics,
suggesting that the brain becomes more selective and enhances specificity when dealing with
information processing.
It is important to note that the susceptibility measure is not a measure of complexity but
rather a measure of the ability of a system to be perturbed, and it is also not a measure of
the ensuing complexity of the response (as in TMS-PCI studies). In contrast, the information
capacity measures the ability of the system to encode external inputs, and as such is more
closely related to complexity measures such as Lempel-Ziv-Welch (LZW), automatic com-
plexity evaluator (ACE), and synchrony coalition entropy (SCE) (used and defined by Schartner
et al., 2015).
First proposed by Massimini et al. (2005), the perturbative approach has been increasingly
applied to investigate brain function because of its potential to relate local neural activity
changes to global brain dynamics and reveal the underlying detailed causal mechanisms. This
approach is a suitable complement to observational approaches that are typically descriptive
and correlative and that thus do not offer insights into the underlying mechanisms. Data-
constrained whole-brain models are key to studying perturbation-induced changes in neural
activity, as they allow parameter optimization and targeted perturbation to be systematically
explored in different brain regions. Perturbations in dynamical models of whole-brain activity
have been shown to dissociate different brain states (Deco et al., 2018; Jobst et al., 2021; Sanz
Perl et al., 2021), providing a robust metric and computational tool capable of characterizing
and unraveling brain states. While a previous study compared perturbation-induced changes
in brain dynamics in the LSD state versus placebo using a similar model-based approach
( Jobst et al., 2021), it did not focus on the information-processing characteristics of brain
states. Still, as in the present study, this previous one did show greater variability in the
Network Neuroscience
1113
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
6
4
1
1
0
4
2
0
5
6
2
8
8
n
e
n
_
a
_
0
0
2
5
0
p
d
/
t
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Turbulent signatures in brain dynamics under psychedelics
perturbational responses in the LSD condition compared with placebo (Jobst et al., 2021).
This variability refers to what we call here information-encoding capability, a measure that
captures how the external perturbations are encoded in whole-brain dynamics. The previous
study also reported that brain dynamics under LSD took longer to recover to baseline activity
after perturbation.
Psychedelic drugs appear to broaden the brain’s dynamical repertoire (Atasoy et al., 2017;
Lord et al., 2019; Tagliazucchi et al., 2014) and enhance global functional connectivity by
shifting the brain’s global working point towards a more globally connected profile (Preller
et al., 2018; Tagliazucchi et al., 2016). The present study’s findings are consistent with this
characterization and advance on it by suggesting that psychedelics also promote a greater
spread of neural activity and an associated increase in information transfer and “mixing”
across domains and scales throughout the brain.
Taken together, our findings show that psychedelics exhibit a particular pattern of (generally
increased) turbulent dynamics that may relate to their characteristic effects on conscious expe-
rience. While helping to enrich our understanding of the brain basis of the psychedelic state,
these findings also deepen our understanding of the functional relevance of turbulence-related
metrics in relation to brain function more broadly.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
6
4
1
1
0
4
2
0
5
6
2
8
8
n
e
n
_
a
_
0
0
2
5
0
p
d
t
/
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
METHODS
To quantify the functional hierarchy under the acute effects of different psychedelics (LSD and
psilocybin), we applied a novel turbulence framework to two psychedelic datasets in human
participants. We used the power of whole-brain model-free and model-based approaches to
characterize the hierarchy for the different compounds. For the model-free approach, we
estimated the levels of turbulence, information transfer, information cascade flow, and infor-
mation cascade. For the model-based approach, we estimated the susceptibility and
information-encoding capability measures. Below we briefly summarize the participants, study
settings, data acquisition, and preprocessing protocols.
Ethics Statement
The National Research Ethics Service Committee (NRES) London–West London approved both
studies, which were conducted following the revised Declaration of Helsinki (2000), the Inter-
national Committee on Harmonization Good Clinical Practice guidelines, and the National
Health Service Research Governance Framework. Imperial College London sponsored the
research carried under a Home Office license for research with schedule 1 drugs. All partic-
ipants provided written informed consent prior to participation.
Participants
LSD. A complete description of the LSD study protocol can be found in the original paper
(Carhart-Harris, Muthukumaraswamy, et al., 2016). Twenty healthy subjects (four females,
mean age = 30.9 ± 7.8 years) recruited via word of mouth participated after study guidance
and physical and mental health screening. The screening for physical health included an elec-
trocardiogram, routine blood tests, and urine tests for recent drug use and pregnancy. All par-
ticipants provided full disclosure of their drug use history in a psychiatric interview. Critical
exclusion criteria included the following: <21 years of age, pregnancy, personal history of
diagnosed psychiatric illness, immediate family a psychotic disorder, an absence
of previous experience with classic psychedelic drug (e.g., LSD, mescaline, psilocybin>40 units consumed per week), or a med-
ically significant condition rendering the volunteer inadequate for the study. Participants
attended two scanning sessions on different days at 8:00 (LSD and placebo) at least two
weeks apart in balanced order, within-participants design. LSD (75 μg in 10 ml saline) or
placebo (10 ml saline) was delivered as bolus injections over 2 min while participants were
instructed to close their eyes and relax. Following an acclimatization period of 60 min inside
a mock MRI scanner, three fMRI scans were conducted in the following order: eyes-closed
resting state, rest while listening to music, and another eyes-closed resting-state session. Par-
ticipants rated the Visual Analog Scale ( VAS) via button press and a digital display screen
presented after each scan. We report results for the pre- and post-music resting-state scans
concatenated in time.
Psilocybin. A complete description of the psilocybin study protocol can be found in the orig-
inal paper (Carhart-Harris et al., 2012). In brief, 15 participants were included in the study
following rigorous exclusion criteria: no younger than 21 years of age, pregnancy, history of
psychiatric disorders, cardiovascular disease, substance dependence, claustrophobia, blood or
needle phobia, or adverse response to hallucinogens. Furthermore, participants were excluded
if their mean framewise displacement exceeded 0.4 mm. All participants underwent two
12-min eyes-closed resting-state fMRI scans over separate sessions, at least seven days apart.
In each session, subjects were injected intravenously with either psilocybin (2 mg dissolved in
10 mL of saline, 60-s injection) or a placebo (10 mL of saline, 60-s injection) in a counter-
balanced design. The injections were given manually by a medical doctor within the scanning
suite. The infusions began precisely 6 min after the start of the 12-min scans and lasted 60 s.
The subjective effects of psilocybin were felt almost immediately after injection and sustained
for the remainder of the scanning session and were rated via button press using the VAS scale.
All participants had previous experience with a hallucinogenic drug but not within six weeks
of the study.
MRI Data Acquisition
Imaging was performed on a 3T GE HDx system. High-resolution anatomical images
LSD.
were acquired with 3D fast spoiled gradient echo scans in an axial orientation, with a field
of view = 256 × 256 × 192 and matrix = 256 × 256 × 129 to yield 1-mm isotropic voxel
resolution. TR/TE = 7.9/3.0 ms; inversion time = 450 ms; flip angle = 20. BOLD-weighted fMRI
data were acquired using a gradient-echo planar imaging sequence, TR/TE = 2,000/35 ms,
field of view = 220 mm, 64 × 64 acquisition matrix, parallel acceleration factor = 2, 90 flip
angles. Thirty-five oblique axial slices were acquired interleaved, each 3.4 mm thick with zero
slice gap (3.4-mm isotropic voxels). The precise length of each of the BOLD scans was 7 min
20 s. One subject aborted the experiment because of anxiety, and four others we excluded for
excessive head motion in the scanner (defined as >15% of volumes with mean framewise
displacement > 0.5 (Carhart-Harris, Muthukumaraswamy, et al., 2016), leaving 15 subjects
with 434 TRs each for analysis.
Imaging acquisitions were identical to the LSD experiment with the following
Psilocybin.
exceptions: BOLD-weighted fMRI data were acquired at TR/TE = 3,000/35 ms, field of view =
192 mm. Thirty-three oblique axial slices were acquired interleaved, each 3 mm thick with
zero slice gap (3 × 3 × 3 mm voxels). Six participants were excluded for excessive motion,
leaving nine participants with 97 TRs for analysis.
Network Neuroscience
1115
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
6
4
1
1
0
4
2
0
5
6
2
8
8
n
e
n
_
a
_
0
0
2
5
0
p
d
.
t
/
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Turbulent signatures in brain dynamics under psychedelics
Resting-State Preprocessing
We used the CONN toolbox, version 17f (CONN; https://www.nitrc.org/projects/conn;
Whitfield-Gabrieli & Nieto-Castanon, 2012) based on Statistical Parametric Mapping 12
(https://www.fil.ion.ucl.ac.uk/spm), implemented in MATLAB to preprocess and denoise the LSD
and psilocybin fMRI data. For each condition (placebo, LSD; placebo, psilocybin), we
applied a standard pipeline including the following steps: removal of the first three volumes
to eliminate saturation effects and achieve steady-state magnetization; functional realign-
ment to correct for movement; slice-timing correction to account for variations in time of
acquisition among slices; identification of outlier scans for subsequent scrubbing through
the quality assurance/artifact rejection software art (https://www.nitrc.org/projects/artifact
_detect), using the default CONN settings of five global signal Z-values and 0.9 mm for
the identification of outlier volumes; normalization to Montreal Neurological Institute
(MNI152) standard space with 2-mm isotropic resampling resolution, using the segmented
gray matter image from each volunteer’s high-resolution T1-weighted image, together with
an a priori gray matter template; spatial smoothing with a Gaussian kernel of 6 mm full
width at half-maximum. For denoising details, see Luppi et al. (2021), Luppi et al. (2019),
and Luppi et al. (2020).
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
6
4
1
1
0
4
2
0
5
6
2
8
8
n
e
n
_
a
_
0
0
2
5
0
p
d
/
.
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Tractography Analysis
Following Deco and Kringelbach (2020), we used the freely available Human Connectome
Project (HCP) database comprising diffusion spectrum and T2-weighted neuroimaging data
from 32 healthy subjects. A comprehensive account of the acquisition parameters can be
found on the HCP website (Schaefer et al., 2018). The free Lead-DBS software package
(https://www.lead-dbs.org/) provides the preprocessing described in detail in Horn, Neumann,
Degen, Schneider, and Kühn (2017). Concisely, the data were processed using a q-sampling
imaging algorithm implemented in DSI Studio (https://dsi-studio.labsolver.org). Segmentation
of the T2-weighted anatomical images produced a white matter mask and coregistering the
images to the b0 image of the diffusion data using SPM12. For each HCP participant,
200,000 fibers were sampled within the white matter mask. Fibers were transformed into
MNI space using Lead-DBS (Horn & Blankenburg, 2016). Lastly, we adopted the standardized
methods in Lead-DBS to obtain the structural connectomes from the Schaefer 1000 parcella-
tion (Schaefer et al., 2018).
Parcellation Scheme
We used the Schaefer atlas with 1,000 parcels (Schaefer et al., 2018) to set brain nodes and
derive the corresponding time series. Also, we computed the Euclidean distances between the
centers of gravity of the parcels in Schaefer 1000 parcellation in MNI space.
Model-Free Framework
Turbulence. The amplitude turbulence, Rλ x(cid:1); t
local order parameter for a given brain node as a function of time:
ð
Þ, is defined as the modulus of the Kuramoto
ð
Rλ x(cid:1); t
(cid:2)
(cid:2)
(cid:2)
Þ ¼ k
(cid:2)
Z ∞
−∞
dx(cid:1)0Gλ x(cid:1) − x(cid:1)0
ð
Þeiφ x(cid:1)0;t
ð
Þ
(cid:2)
(cid:2)
(cid:2)
(cid:2);
(1)
where ‖
e−λj(cid:1)xj , φ x(cid:1); t
‖ is the modulus of the complex number, Gλ is the local weighting kernel Gλ x(cid:1)ð Þ ¼
Þ are the phases of the spatiotemporal data, k is the normalization factor
ð
Network Neuroscience
1116
Turbulent signatures in brain dynamics under psychedelics
ð
Þ
(cid:3)
R ∞
−∞ dx(cid:1)0Gλ x(cid:1) − x(cid:1)0
(cid:4)−1, and λ defines the spatial scaling. Hence, Rλ represents the local levels
of synchronization at a given scale, λ, as function of space, x(cid:1), and time, t. Inspired by
the rotational vortices found in fluid dynamics, the turbulence measure characterizes the brain
vortex space, Rλ, over time. Note that t can take values from 1 to 434 for the LSD condition,
and from 1 to 97 for the psilocybin condition; and x(cid:1) can take values from 0.2165 to
172.9464.
The level of amplitude turbulence, Dλ, is defined as the standard deviation across time and
space of the modulus of local Kuramoto order parameter (Rλ).
(cid:5)
Dλ ¼ Rλ
2
(cid:6)
(cid:1)x;t
− Rλh
i2
(cid:1)x;t
;
(2)
where the brackets h
i(cid:1)x; t denote averages across time and space.
Information transfer. The spatial information transfer shows how the information travels across
space at a specific scale, λ. This measure is defined as the slope of a linear fitting in log-log
scale of the time correlation between the Kuramoto local order parameter of two brain areas,
at the same scale as a function of its Euclidean distance (r) within the inertial subrange (the
limited range where turbulence energy is transferred from larger to smaller scales without
loss). We used the linear fit only to quantify the level of decay of the correlation of the local
level of synchronization with distance. Please note that we compute the linear fit in a limited
log-log space and, as such, we do not have a distribution, and thus are not trying to fit a
power law.
log corrt Rλ (cid:1)xð Þ; Rλ (cid:1)x þ r
ð
ð
ð
Þ
Þ ¼ Aλ (cid:2) log rð Þ þ Bλ;
Þ
(3)
where corrt is the pairwise correlation across time, Aλ and Bλ are the fitting parameters for
each scale (λ), where r is the spatial distance in the brain. The negative slope (Aλ) stands for
the transfer in the spatial direction r of the information in terms of time correlation of the local
level of synchronization. In this regard, when the slope is steeper, the information travels
across shorter distances, while a flatter slope indicates that the information is transferred
across longer distances. Thus, the negative slope stands for the spatial information transfer.
Note that the parameter A depends on only λ. It depends neither on t as the correlation is
over time, nor on the brain areas, as the pairwise correlations are organized as a function of
the Euclidean distances.
Information cascade flow. The information cascade flow characterizes the stream of informa-
tion between a given scale (λ) and a subsequent lower scale (λ − Δλ, where Δλ is a scale step)
in consecutive time steps (t and t + Δt ). In this way, the information cascade flow covers the
information transfer across scales computed as the time correlation between the Kuramoto
local order parameter in two consecutive scales and times:
F λð Þ ¼ corrt Rλ (cid:1)x; t þ Δt
ð
h
ð
Þ; Rλ−Δλ (cid:1)x; t
ð
Þ
Þ
i(cid:1)x
;
(4)
where the brackets h
wise time correlations.
i(cid:1)x denote averages across time and space, and corrt refers to the pair-
Information cascade. Finally, the information cascade is defined by averaging the informa-
tion transfer across scales λ, capturing the entire information-processing behavior across
scales.
Network Neuroscience
1117
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
6
4
1
1
0
4
2
0
5
6
2
8
8
n
e
n
_
a
_
0
0
2
5
0
p
d
/
t
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Turbulent signatures in brain dynamics under psychedelics
Node variability of local synchronization. The node variability of the local synchronization is
defined as the standard deviation across time of the local Kuramoto order parameter, such
as the following:
(cid:5)
NLS n; λð
Þ ¼ Rλ nð Þ tð Þ2
(cid:6)
t
(cid:5)
(cid:6)
2
− Rλ nð Þ tð Þ
t
;
(5)
where n is a node, and the brackets h
it denote the average values across time points. Further,
we used the discrete version of the node-level Kuramoto order parameter in Equation 2
(because we are computing a node-level metric over a given parcellation), with modulus R,
indicating a spatial average of the complex phase factor of the local oscillators weighted by
the coupling calculated through
Rλ nð Þ tð Þ ¼
X
(cid:2)
(cid:2)
(cid:2)
(cid:2)
(cid:2)
”
#
Cλ npð
ÞP
qCλ npð
Þ
p
(cid:2)
(cid:2)
(cid:2)
;
(cid:2)
(cid:2)
eiφ
p tð Þ
(6)
‖ is the modulus of the complex number, φp(t) are the phases of the spatiotemporal
where ‖
data, Cλ(np) is the local weighting kernel between node n and p, and λ defines the spatial
scaling:
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
6
4
1
1
0
4
2
0
5
6
2
8
8
n
e
n
_
a
_
0
0
2
5
0
p
d
/
t
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Cnp ¼ e−λ r n;pð
ð
Þ;
Þ
(7)
where r(n, p) is the Euclidian distance between the brain areas n and p in MNI space.
Model-Based Framework
Whole-brain computational model. We built a whole-brain computational model where the
local dynamics of each brain area (node) are described by the normal form of a supercritical
Hopf bifurcation (also known as Stuart-Landau), which can describe the transition from
noise-induced oscillations to fully sustained oscillations (Deco, Cabral, et al., 2017). This
model is characterized by two parameters that rule the global dynamical behavior: the
multiplicative factor, G, denoting the global conductivity of the fibers, scaling the structural
connectivity between brain areas, which is assumed to be equal throughout the brain (Deco,
Cabral, et al., 2017; Deco, Tagliazucchi, Laufs, Sanjuán, & Kringelbach, 2017); and the local
bifurcation parameter (aj), which rules the dynamical behavior of each area between noise-
induced, leading the system to a stable fixed point (a < 0), a stable limit cycle producing
self-sustained oscillations (a > 0), or a critical behavior between both (a ∼ 0). We optimized
the model parameters to better fit the empirical functional connectivity as a function of dis-
tance, r, within the inertial subrange. The model considered 1,000 cortical brain areas from
the Schaefer atlas mentioned above. The underlying anatomical matrix Cnp was added to
link the brain structure and functional dynamics and was obtained by measuring the expo-
nential distance rule as defined in Equation 2. The spontaneous local dynamics of each
brain area were described by the normal form of a supercritical Hopf bifurcation, which
simulates the dynamics for each brain area from noisy to oscillatory dynamics as follows:
(cid:3)
¼ anxn − x 2
n
(cid:4)
xn − ωnyn þ νη
n tð Þ;
þ y 2
n
(cid:3)
¼ anyn − x 2
n
(cid:4)
þ y 2
n
yn þ ωnxn þ νη
n tð Þ;
dxn
dt
dyn
dt
(8)
(9)
1118
Network Neuroscience
Turbulent signatures in brain dynamics under psychedelics
where ηn(t) is additive Gaussian noise with standard deviation ν = 0.01. The frequency ωn of
each brain area was determined from the empirical fMRI data as the peak of the power
spectrum. This normal form has a supercritical bifurcation at an = 0, such that for an > 0
the system is in a stable limit cycle oscillation with frequency fn = ωn/2π, whereas for an < 0
the local dynamics are in a stable point (i.e., noisy state). Lastly, the whole-brain dynamics
were described by the following set of coupled equations:
(cid:3)
¼ anxn − x 2
n
(cid:4)
þ y 2
n
xn − ωnyn þ G
X
N
p¼1
(cid:7)
Cnp xp tð Þ − xn
(cid:8)
þ νη
n tð Þ;
(cid:3)
¼ anyn − x 2
n
(cid:4)
yn þ ωnxn þ G
þ y 2
n
X
N
p¼1
(cid:7)
Cnp yp tð Þ − yp
(cid:8)
þ νη
n tð Þ;
dxn
dt
dyn
dt
(10)
(11)
where the global coupling factor G, scaled equally for each brain area, denotes the input
received in a region n from every other region p.
Functional connectivity fitting. Kolmogorov’s structure-function (Frisch, 1995; Kolmogorov,
1941a, 1941b) of a variable u was applied to the BOLD signal of the data. This measure is
based on the functional correlations between each pair of brain areas with equal Euclidean
distance and was defined as the following:
(cid:5)
S rð Þ ¼ u x(cid:1) þ r
ð
ð
Þ − u x(cid:1)ð Þ
Þ2
(cid:6)
(cid:1)x;t
¼ 2 FC 0ð Þ − FC rð Þ
½
(cid:3);
(12)
where FC(r ) is the spatial correlations of two points separated by a Euclidean distance r, which
is given by
FC rð Þ ¼ u x(cid:1)þ r
h
ð
Þu x(cid:1)ð Þ
;
i(cid:1)x;t
(13)
i(cid:1)x; t refers to the average across the spatial location x of the brain
where the symbol h
areas and time, and assuming stationarity. Thus, the structure functions are characterizing
the evolution of the functional connectivity as function of the Euclidean distance between
equally distant nodes, which is different from the usual definition of FC that does not
include distance. The simulated and the empirical FC(r) were compared using the
Euclidian distance within the inertial range defined in Deco and Kringelbach (2020),
allowing for adequate model
fitting performance testing in describing the changes
observed in the resting-state FC.
Susceptibility. The susceptibility measure of the whole-brain model was defined as the brain’s
sensitivity to react to external perturbations. The Hopf model was perturbed for each G by
randomly changing the local bifurcation parameter, an, in the range [−0.02:0]. The sensitivity
of the perturbations on the spatiotemporal dynamics was calculated by measuring the modu-
lus of the local Kuramoto order parameter as the following:
(cid:11)
D
(cid:9)
D
χ ¼
eRλs x(cid:1); t
ð
Þ
E
t
h
− Rλs x(cid:1); t
ð
(cid:10)
E
(cid:12)
Þ
i
t
trials
;
(cid:1)x
(14)
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
6
4
1
1
0
4
2
0
5
6
2
8
8
n
e
n
_
a
_
0
0
2
5
0
p
d
/
t
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
where eRλs
h
it, h
(cid:1)x; t
ð
itrials, and h
Þ corresponds to the perturbed case, the Rλs
(cid:1)x; t
ð
Þ to the unperturbed case, and
i(cid:1)x to the average across time, trials, and space, respectively.
Network Neuroscience
1119
Turbulent signatures in brain dynamics under psychedelics
Information-encoding capability. The information-encoding capability captures how the exter-
nal perturbations are encoded in whole-brain dynamics. The information capability, I, was
defined as the standard deviation across trials of the difference between the perturbed
eRλs
(cid:1)x; t
Þ mean of the modulus of the local Kuramoto order parameter
ð
across time t, averaged across all brain areas n as the following:
Þ and unperturbed Rλs
(cid:1)x; t
ð
(cid:11)
(cid:11)
(cid:9)
D
I ¼
eRλs
ð
(cid:1)x; t
E
Þ
t
− Rλs
h
(cid:1)x; t
ð
Þ
i
t
(cid:12)
(cid:10)
2
(cid:12)
(cid:11)
D
(cid:9)
D
−
trials
(cid:1)x
eRλs
(cid:1)x; t
ð
E
Þ
t
− Rλs
h
ð
(cid:1)x; t
Þ
i
t
(cid:10)
E
2
(cid:12)
trials
;
(cid:1)x
(15)
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
6
4
1
1
0
4
2
0
5
6
2
8
8
n
e
n
_
a
_
0
0
2
5
0
p
d
.
t
/
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
where the brackets h
it, h
itrials, and h
i(cid:1)x denote the averages defined as above.
Statistical Analyses
Differences between conditions in turbulence, information transfer, flow, and capability were
statistically assessed using a permutation-based paired t test. This nonparametric test uses per-
mutations of group labels to estimate the null distribution computed independently for each
condition. We compared conditions employing a t test to each of the 1,000 permutations and
used a significance threshold of α = 0.05. Additionally, we used the false discovery rate at the
0.05 level of significance to correct multiple comparisons (Hochberg & Benjamini, 1990). We
applied the Wilcoxon rank sum method to compute the difference between conditions in sus-
ceptibility and information-encoding capacity measures. The similitude of the distributions of
node-level turbulence per spatial scale, between each psychedelic drug and its counterpart
placebo, was assessed using the Kolmogorov–Smirnov distance (KSD). The KSD quantifies the
maximal difference between the cumulative distribution functions of the two samples.
ACKNOWLEDGMENTS
The contents of this publication are solely the responsibility of the authors and do not represent
the official views of the funding institutions.
SUPPORTING INFORMATION
Supporting information for this article is available at https://doi.org/10.1162/netn_a_00250.
AUTHOR CONTRIBUTIONS
Josephine Cruzat: Conceptualization; Formal analysis; Investigation; Methodology; Valida-
tion; Visualization; Writing – original draft; Writing – review & editing. Yonatan Sanz Perl:
Conceptualization; Formal analysis; Methodology; Supervision; Validation; Visualization.
Anira Escrichs: Software. Jakub Vohryzek: Validation. Christopher Timmermann: Validation.
Leor Roseman: Data curation; Validation. Andrea I. Luppi: Data curation. Agustin Ibañez:
Funding acquisition; Writing – review & editing. David Nutt: Data curation; Validation.
Robin Carhart-Harris: Data curation; Funding acquisition; Supervision; Validation; Writing –
review & editing. Enzo Tagliazucchi: Supervision; Validation; Writing – review & editing.
Investigation; Methodology; Resources; Supervision;
Gustavo Deco: Conceptualization;
Validation; Writing – review & editing. Morten L. Kringelbach: Conceptualization; Inves-
tigation; Methodology; Resources; Supervision; Validation; Visualization; Writing – review
& editing.
Network Neuroscience
1120
Turbulent signatures in brain dynamics under psychedelics
FUNDING INFORMATION
Yonatan Sanz Perl, EU H2020 FET Flagship programme, Award ID: HBP SGA3 945539.
Anira Escrichs, EU H2020 FET Flagship programme, Award ID: HBP SGA3 945539. Jakub
Vohryzek, EU H2020 FET Proactive project Neurotwin, Award ID: 101017716. Andrea I.
Luppi, Gates Cambridge scholarship. Agustin Ibañez, Takeda, CW2680521; CONICET;
ANID/ FONDECYT Regular (1210195 and 1210176); FONCYT-PICT 2017-1820;
ANID/FONDAP/15150012; Sistema General de Regalías (BPIN2018000100059), Universi-
dad del Valle (CI 5316); Programa Interdisciplinario de Investigación Experimental en Comu-
nicación y Cognición (PIIECC), Facultad de Humanidades, USACH; Alzheimer’s Association
GBHI ALZ UK-20-639295; and the Multi-Partner Consortium to Expand Dementia Research
in Latin America [ReDLat, supported by National Institutes of Health, National Institutes of
Aging (R01 AG057234), Alzheimer’s Association (SG-20-725707), Rainwater Charitable
Foundation–Tau Consortium, and Global Brain Health Institute)]. Enzo Tagliazucchi,
Mercator Fellowship granted by the German Research Foundation, ANPCyT, Award ID:
PICT-2019-02294 and Award ID: PICT-2018-03103. Gustavo Deco, Spanish Ministry of
Science, Innovation and Universities (MCIU), Award ID: PID2019-105772GB-I00 MCIU
AEI), State Research Agency (AEI), EU H2020 FET Flagship programme, Award ID: HBP
SGA3 945539, SGR Research Support Group support, Award ID: 2017 SGR 1545, funded
by the Catalan Agency for Management of University and Research Grants (AGAUR), EU
H2020 FET Proactive programme, Award ID: 101017716), EU H2020 MSCA-ITN Innovative
Training Networks: euSNN European School of Network Neuroscience, Award ID: 860563,
CECH The Emerging Human Brain Cluster, Award ID: 001-P-001682, within the framework
of the European Research Development Fund Operational Program of Catalonia 2014–2020,
Fundacio La Marato TV3, Brain-Connects: Brain Connectivity during Stroke Recovery and
Rehabilitation, Award ID: 201725.33, FLAG–ERA JTC 2017, Spanish Ministry of Science,
Innovation and Universities (MCIU), State Research Agency (AEI), Award ID: PCI2018-
092891. Morten L. Kringelbach, Danish National Research Foundation, Award ID:
DNRF117, Pettit and Carlsberg Foundations.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
6
4
1
1
0
4
2
0
5
6
2
8
8
n
e
n
_
a
_
0
0
2
5
0
p
d
.
/
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
REFERENCES
Atasoy, S., Roseman, L., Kaelen, M., Kringelbach, M. L., Deco, G., &
Carhart-Harris, R. L. (2017). Connectome-harmonic decomposi-
tion of human brain activity reveals dynamical repertoire
re-organization under LSD. Scientific Reports, 7(1), 17661.
https://doi.org/10.1038/s41598-017-17546-0, PubMed: 29247209
Atasoy, S., Vohryzek, J., Deco, G., Carhart-Harris, R. L., &
Kringelbach, M. L. (2018). Common neural signatures of
psychedelics: Frequency-specific energy changes and reper-
toire expansion revealed using connectome-harmonic decom-
position. Progress in Brain Research, 242, 97–120. https://doi
.org/10.1016/bs.pbr.2018.08.009, PubMed: 30471684
Bogenschutz, M. P., Forcehimes, A. A., Pommy, J. A., Wilcox, C. E.,
Barbosa, P., & Strassman, R. J. (2015). Psilocybin-assisted
treatment for alcohol dependence: A proof-of-concept study.
Journal of Psychopharmacology, 29(3), 289–299. https://doi.org
/10.1177/0269881114565144, PubMed: 25586396
Bourdillon, P., Hermann, B., Guénot, M., Bastuji, H., Isnard, J.,
King, J.-R., … Naccache, L. (2020). Brain-scale cortico-cortical
functional connectivity in the delta-theta band is a robust
signature of conscious states: An intracranial and scalp EEG
study. Scientific Reports, 10, 14037. https://doi.org/10.1038
/s41598-020-70447-7, PubMed: 32820188
Carhart-Harris, R. (2018). The entropic brain-revisited. Neurophar-
macology, 142, 167–178. https://doi.org/10.1016/j.neuropharm
.2018.03.010, PubMed: 29548884
Carhart-Harris, R., Giribaldi, B., Watts, R., Baker-Jones, M.,
Murphy-Beiner, A., Murphy, R., … Nutt, D. J. (2021). Trial of
psilocybin versus escitalopram for depression. New England
Journal of Medicine, 384, 1402–1411. https://doi.org/10.1056
/NEJMoa2032994, PubMed: 33852780
Carhart-Harris, R., Leech, R., Hellyer, P. J., Shanahan, M., Feilding,
A., Tagliazucchi, E., … Nutt, D. (2014). The entropic brain: A
theory of conscious states informed by neuroimaging research
with psychedelic drugs. Frontiers in Human Neuroscience, 8,
20. https://doi.org/10.3389/fnhum.2014.00020, PubMed:
24550805
Carhart-Harris, R., Muthukumaraswamy, S., Roseman, L., Kaelen,
M., Droog, W., Murphy, K., … Nutt, D. J. (2016). Neural
Network Neuroscience
1121
Turbulent signatures in brain dynamics under psychedelics
correlates of the LSD experience revealed by multimodal
neuroimaging. Proceedings of
the National Academy of
Sciences, 113(17), 4853–4858. https://doi.org/10.1073/pnas
.1518377113, PubMed: 27071089
Carhart-Harris, R. L., Bolstridge, M., Rucker, J., Day, C. M. J.,
Erritzoe, D., Kaelen, M., … Nutt, D. J. (2016). Psilocybin
with psychological support for treatment-resistant depression:
An open-label
feasibility study. The Lancet Psychiatry, 3(7),
619–627. https://doi.org/10.1016/S2215-0366(16)30065-7,
PubMed: 27210031
Carhart-Harris, R. L., Erritzoe, D., Williams, T., Stone, J. M., Reed,
L. J., Colasanti, A., … Murphy, K. (2012). Neural correlates of
the psychedelic state as determined by fMRI studies with psilo-
cybin. Proceedings of the National Academy of Sciences, 109,
2138–2143. https://doi.org/10.1073/pnas.1119598109,
PubMed: 22308440
Deco, G., Cabral, J., Saenger, V. M., Boly, M., Tagliazucchi, E.,
Laufs, H., … Kringelbach, M. L. (2018). Perturbation of
whole-brain dynamics in silico reveals mechanistic differences
between brain states. NeuroImage, 169, 46–56. https://doi.org
/10.1016/j.neuroimage.2017.12.009, PubMed: 29225066
Deco, G., Cabral, J., Woolrich, M. W., Stevner, A. B. A., van
Hartevelt, T. J., & Kringelbach, M. L. (2017). Single or multiple
frequency generators in on-going brain activity: A mechanistic
whole-brain model of empirical MEG data. NeuroImage, 152,
538–550. https://doi.org/10.1016/j.neuroimage.2017.03.023,
PubMed: 28315461
Deco, G., Kemp, M., & Kringelbach, M. L. (2021). Leonardo da
Vinci and the search for order in neuroscience. Current Biology,
31, R704–R709. https://doi.org/10.1016/j.cub.2021.03.098,
PubMed: 34102114
Deco, G., & Kringelbach, M. L. (2020). Turbulent-like dynamics in
the human brain. Cell Reports, 33, 108471. https://doi.org/10
.1016/j.celrep.2020.108471, PubMed: 33296654
Deco, G., Kringelbach, M. L.,
Jirsa, V. K., & Ritter, P.
(2017).
The dynamics of resting fluctuations in the brain: Metastability
and its dynamical cortical core. Scientific Reports, 7, 3095.
https://doi.org/10.1038/s41598-017-03073-5, PubMed:
28596608
Deco, G., Perl, Y. S., Vuust, P., Tagliazucchi, E., Kennedy, H., &
Kringelbach, M. L. (2021). Rare long-range cortical connections
enhance human information processing. Current Biology, 31(20),
4436–4448. https://doi.org/10.1016/j.cub.2021.07.064,
PubMed: 34437842
Deco, G., Tagliazucchi, E., Laufs, H., Sanjuán, A., & Kringelbach,
M. L. (2017). Novel intrinsic ignition method measuring
local-global integration characterizes wakefulness and deep
sleep. eNeuro, 4(5). https://doi.org/10.1523/ENEURO.0106-17
.2017, PubMed: 28966977
Ercsey-Ravasz, M., Markov, N. T., Lamy, C., Van Essen, D. C.,
Knoblauch, K., Toroczkai, Z., & Kennedy, H. (2013). A predictive
network model of cerebral cortical connectivity based on a
distance rule. Neuron, 80, 184–197. https://doi.org/10.1016/j
.neuron.2013.07.036, PubMed: 24094111
Escrichs, A., Perl, Y. S., Uribe, C., Camara, E., Türker, B., Pyatigorskaya,
N., … Deco, G. (2021). Unifying turbulent dynamics framework
distinguishes different brain states. bioRxiv. https://doi.org/10
.1101/2021.10.14.464380
Frisch, U. (1995). Turbulence: The legacy of A. N. Kolmogorov.
C a m b r i d g e U n i v e r s i t y P r e s s . h t t p s : / / d o i . o rg / 1 0 . 1 0 1 7
/CBO9781139170666
Friston, K. (2010). The free-energy principle: A unified brain theory?
Nature Reviews Neuroscience, 11, 127–138. https://doi.org/10
.1038/nrn2787, PubMed: 20068583
Griffiths, R. R., Johnson, M. W., Carducci, M. A., Umbricht, A.,
Richards, W. A., Richards, B. D., … Klinedinst, M. A. (2016).
Psilocybin produces substantial and sustained decreases in
depression and anxiety in patients with life-threatening cancer: A
randomized double-blind trial. Journal of Psychopharmacology,
30(12), 1181–1197. https://doi.org/10.1177/0269881116675513,
PubMed: 27909165
Hartogsohn, I. (2018). The meaning-enhancing properties of
psychedelics and their mediator role in psychedelic therapy,
spirituality, and creativity. Frontiers in Neuroscience, 12, 129.
https://doi.org/10.3389/fnins.2018.00129, PubMed: 29559884
Herzog, R., Mediano, P. A. M., Rosas, F. E., Carhart-Harris, R., Perl,
Y. S., Tagliazucchi, E., & Cofre, R. (2020). A mechanistic model
of the neural entropy increase elicited by psychedelic drugs.
Scientific Reports, 10, 17725. https://doi.org/10.1038/s41598
-020-74060-6, PubMed: 33082424
Hochberg, Y., & Benjamini, Y. (1990). More powerful procedures
for multiple significance testing. Statistics in Medicine, 9(7),
811–818. https://doi.org/10.1002/sim.4780090710, PubMed:
2218183
Horn, A., & Blankenburg, F. (2016). Toward a standardized structural–
functional group connectome in MNI space. NeuroImage, 124,
310–322. https://doi.org/10.1016/j.neuroimage.2015.08.048,
PubMed: 26327244
Horn, A., Neumann, W. J., Degen, K., Schneider, G. H., & Kühn,
A. A. (2017). Toward an electrophysiological “sweet spot” for
deep brain stimulation in the subthalamic nucleus. Human Brain
Mapping, 38, 3377–3390. https://doi.org/10.1002/hbm.23594,
PubMed: 28390148
Jobst, B. M., Atasoy, S., Ponce-Alvarez, A., Sanjuán, A., Roseman,
L., Kaelen, M., … Deco, G. (2021). Increased sensitivity to strong
perturbations in a whole-brain model of LSD. NeuroImage, 230,
117809. https://doi.org/10.1016/j.neuroimage.2021.117809,
PubMed: 33524579
Johnson, M. W., Garcia-Romeu, A., & Griffiths, R. R. (2017). Long-
term follow-up of psilocybin-facilitated smoking cessation.
American Journal of Drug and Alcohol Abuse, 43, 55–60.
https://doi.org/10.3109/00952990.2016.1170135, PubMed:
27441452
Johnson, M. W., Griffiths, R. R., Hendricks, P. S., & Henningfield,
J. E. (2018). The abuse potential of medical psilocybin according
to the 8 factors of the Controlled Substances Act. Neuropharma-
cology, 142, 143–166. https://doi.org/10.1016/j.neuropharm
.2018.05.012, PubMed: 29753748
Johnson, M. W., Hendricks, P. S., Barrett, F. S., & Griffiths, R. R.
(2019). Classic psychedelics: An integrative review of epidemi-
ology, therapeutics, mystical experience, and brain network
function. Pharmacology and Therapeutics, 197, 83–102.
https://doi.org/10.1016/j.pharmthera.2018.11.010, PubMed:
30521880
King, J.-R., Sitt, J. D., Faugeras, F., Rohaut, B., El Karoui, I., Cohen,
L., … Dehaene, S. (2013). Information sharing in the brain
Network Neuroscience
1122
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
6
4
1
1
0
4
2
0
5
6
2
8
8
n
e
n
_
a
_
0
0
2
5
0
p
d
.
/
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Turbulent signatures in brain dynamics under psychedelics
indexes consciousness in noncommunicative patients. Current
Biology, 23, 1914–1919. https://doi.org/10.1016/j.cub.2013.07
.075, PubMed: 24076243
Kolmogorov, A. N. (1941a). Dissipation of energy in locally isotro-
pic turbulence. Proceedings of the USSR Academy of Sciences
(Atmos. Ocean. Phys.), 32, 16–18.
Kolmogorov, A. N. (1941b). The local structure of turbulence in
incompressible viscous fluid for very large Reynolds numbers.
Proceedings of the USSR Academy of Sciences, 30, 301–305.
Kuramoto, Y. (1984). Chemical turbulence. In Chemical oscilla-
tions, waves, and turbulence (pp. 111–140). Berlin: Springer.
https://doi.org/10.1007/978-3-642-69689-3_7
Lord, L. D., Expert, P., Atasoy, S., Roseman, L., Rapuano, K.,
Lambiotte, R., … Cabral, J. (2019). Dynamical exploration of
the repertoire of brain networks at rest is modulated by psilocybin.
NeuroImage, 199, 127–142. https://doi.org/10.1016/j.neuroimage
.2019.05.060, PubMed: 31132450
Luppi, A. I., Carhart-Harris, R. L., Roseman, L., Pappas, I., Menon,
D. K., & Stamatakis, E. A. (2021). LSD alters dynamic integration
and segregation in the human brain. NeuroImage, 227, 117653.
https://doi.org/10.1016/j.neuroimage.2020.117653, PubMed:
33338615
Luppi, A. I., Craig, M. M., Pappas, I., Finoia, P., Williams, G. B.,
Allanson, J., … Menon, D. K. (2019). Consciousness-specific
dynamic interactions of brain integration and functional diver-
sity. Nature Communications, 10, 4616. https://doi.org/10.1038
/s41467-019-12658-9, PubMed: 31601811
Luppi, A. I., Vohryzek, J., Kringelbach, M. L., Mediano, P. A. M.,
Craig, M. M., Adapa, R., … Finoia, P. (2020). Connectome
harmonic decomposition of human brain dynamics reveals a
landscape of consciousness. bioRxiv. https://doi.org/10.1101
/2020.08.10.244459
Markov, N. T., Ercsey-Ravasz, M., Van Essen, D. C., Knoblauch, K.,
Toroczkai, Z., & Kennedy, H. (2013). Cortical high-density coun-
terstream architectures. Science, 342(6158), 1238406. https://doi
.org/10.1126/science.1238406, PubMed: 24179228
Markov, N. T., Vezoli, J., Chameau, P., Falchier, A., Quilodran, R.,
Huissoud, C., … Ullman, S. (2014). Anatomy of hierarchy:
Feedforward and feedback pathways in macaque visual cortex.
Journal of Comparative Neurology, 522, 225–259. https://doi.org
/10.1002/cne.23458, PubMed: 23983048
Massimini, M., Ferrarelli, F., Huber, R., Esser, S. K., Singh, H., &
Tononi, G. (2005). Breakdown of cortical effective connectivity
during sleep. Science, 309, 2228–2232. https://doi.org/10.1126
/science.1117256, PubMed: 16195466
Milliere, R. (2017). Looking for the self: Phenomenology,
neurophysiology and philosophical significance of drug-
induced ego dissolution. Frontiers in Human Neuroscience, 11,
245. https://doi.org/10.3389/fnhum.2017.00245, PubMed:
28588463
Nichols, D. E. (2016). Psychedelics. Pharmacological Reviews, 68,
264–355. https://doi.org/10.1124/pr.115.011478, PubMed:
26841800
Nour, M. M., Evans, L., Nutt, D., & Carhart-Harris, R. L. (2016).
Ego-d i s s o l u t i o n a n d p s y c h e d e l i c s : Va l i d a t i o n o f t h e
ego-dissolution inventory (EDI). Frontiers in Human Neurosci-
ence, 10, 269. https://doi.org/10.3389/fnhum.2016.00269,
PubMed: 27378878
Osmond, H. (1957). A review of the clinical effects of psychotomi-
metic agents. Annals of the New York Academy of Sciences,
66(3), 418–434. https://doi.org/10.1111/j.1749-6632.1957
.tb40738.x, PubMed: 13425232
Pollan, M. (2019). How to change your mind: What the new
science of psychedelics teaches us about consciousness, dying,
addiction, depression, and transcendence. Penguin Books.
Preller, K. H., Burt, J. B., Ji, J. L., Schleifer, C. H., Adkinson, B. D.,
Stämpfli, P., … Anticevic, A. (2018). Changes in global and
thalamic brain connectivity in LSD-induced altered states of con-
sciousness are attributable to the 5-HT2A receptor. eLife, 7,
e35082. https://doi.org/10.7554/eLife.35082, PubMed: 30355445
Roseman, L., Nutt, D. J., & Carhart-Harris, R. L. (2018). Quality of
acute psychedelic experience predicts therapeutic efficacy of
psilocybin for treatment-resistant depression. Frontiers in Phar-
macology, 8, 974. https://doi.org/10.3389/fphar.2017.00974,
PubMed: 29387009
Sanz Perl, Y., Pallavicini, C., Pérez Ipiña, I., Demertzi, A.,
Bonhomme, V., Martial, C., … Kringelbach, M. (2021). Perturba-
tions in dynamical models of whole-brain activity dissociate
between the level and stability of consciousness. PLoS Computa-
tional Biology, 17, e1009139. https://doi.org/10.1371/journal
.pcbi.1009139, PubMed: 34314430
Schaefer, A., Kong, R., Gordon, E. M., Laumann, T. O., Zuo, X.-N.,
Holmes, A. J., … Yeo, B. T. T. (2018). Local-global parcellation of
the human cerebral cortex from intrinsic functional connectivity
MRI. Cerebral Cortex, 28, 3095–3114. https://doi.org/10.1093
/cercor/bhx179, PubMed: 28981612
Schartner, M., Seth, A., Noirhomme, Q., Boly, M., Bruno, M.-A.,
Laureys, S., & Barrett, A. (2015). Complexity of multi-dimensional
spontaneous EEG decreases during propofol induced general
anaesthesia. PLoS ONE, 10(8), e0133532. https://doi.org/10.1371
/journal.pone.0133532, PubMed: 26252378
Sheremet, A., Qin, Y., Kennedy, J. P., Zhou, Y., & Maurer, A. P.
(2019). Wave turbulence and energy cascade in the hippocam-
pus. Frontiers in Systems Neuroscience, 12, 62. https://doi.org/10
.3389/fnsys.2018.00062, PubMed: 30662397
Smallwood, J., Bernhardt, B. C., Leech, R., Bzdok, D., Jefferies, E.,
& Margulies, D. S. (2021). The default mode network in cogni-
tion: A topographical perspective. Nature Reviews Neuroscience,
22(8), 503–513. https://doi.org/10.1038/s41583-021-00474-4,
PubMed: 34226715
Tagliazucchi, E., Carhart-Harris, R., Leech, R., Nutt, D., & Chialvo,
D. R. (2014). Enhanced repertoire of brain dynamical states
during the psychedelic experience. Human Brain Mapping, 35,
5442–5456. https://doi.org/10.1002/ hbm.22562, PubMed:
24989126
Taglia zucchi, E., Roseman, L ., Kaelen, M., O r ban, C.,
Muthukumaraswamy, S. D., Murphy, K., … Carhart-Harris, R.
(2016). Increased global functional connectivity correlates with
LSD-induced ego dissolution. Current Biology, 26(8), 1043–1050.
https://doi.org/10.1016/j.cub.2016.02.010, PubMed: 27085214
Varley, T. F., Carhart-Harris, R., Roseman, L., Menon, D. K., &
Stamatakis, E. A. (2020). Serotonergic psychedelics LSD & psilo-
cybin increase the fractal dimension of cortical brain activity in
spatial and temporal domains. NeuroImage, 220, 117049. https://
doi.org/10.1016/j.neuroimage.2020.117049, PubMed:
32619708
Network Neuroscience
1123
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
6
4
1
1
0
4
2
0
5
6
2
8
8
n
e
n
_
a
_
0
0
2
5
0
p
d
t
.
/
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Turbulent signatures in brain dynamics under psychedelics
Vatansever, D., Schröter, M., Adapa, R. M., Bullmore, E. T., Menon,
D. K., & Stamatakis, E. A. (2020). Reorganisation of brain hubs
across altered states of consciousness. Scientific Reports, 10,
3402. https://doi.org/10.1038/s41598-020-60258-1, PubMed:
32099008
Whitfield-Gabrieli, S., & Nieto-Castanon, A. (2012). CONN: A
functional connectivity toolbox for correlated and anticorrelated
brain networks. Brain Connectivity, 2, 125–141. https://doi.org
/10.1089/brain.2012.0073, PubMed: 22642651
Wildie, M., & Shanahan, M. (2012). Metastability and chimera
states in modular delay and pulse-coupled oscillator networks.
Chaos, 22(4), 043131. https://doi.org/10.1063/1.4766592,
PubMed: 23278066
Yaden, D. B., & Griffiths, R. R. (2020). The subjective effects of
psychedelics are necessary for their enduring therapeutic
effects. ACS Pharmacology and Translational Science, 4(2),
568–572. https://doi.org/10.1021/acsptsci.0c00194, PubMed:
33861219
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
6
4
1
1
0
4
2
0
5
6
2
8
8
n
e
n
_
a
_
0
0
2
5
0
p
d
.
/
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Network Neuroscience
1124