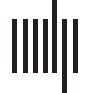FOCUS FEATURE:
New Trends in Connectomics
ADHD and attentional control: Impaired
segregation of task positive and task
negative brain networks
1
Brian D. Mills
1
, Oscar Miranda-Dominguez
, Sarah L. Karalunas
1
Julia Painter
1
, Kathryn L. Mills
, Joel T. Nigg
3,1
3,1
, Eric Earl
1
1
, Michaela Cordova
1,2,3
,
, and Damien A. Fair
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
0
2
0
2
2
0
0
1
0
9
2
1
5
7
n
e
n
_
a
_
0
0
0
3
4
p
d
.
t
/
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
a n o p e n a c c e s s
j o u r n a l
1Department of Behavioral Neuroscience, Oregon Health and Science University, Portland, OR, USA
2Advanced Imaging Research Center, Oregon Health and Science University, Portland, OR, USA
3Department of Psychiatry, Oregon Health and Science University, Portland, OR, USA
Keywords: ADHD, Functional connectivity MRI, Attentional control, Default mode network, Task
positive network, Negative connectivity
ABSTRACT
In children with attention deficit hyperactivity disorder (ADHD) difficulty maintaining task
focus may relate to the coordinated, negatively correlated activity between brain networks
that support the initiation and maintenance of task sets (task positive networks) and networks
that mediate internally directed processes (i.e., the default mode network). Here, resting-state
functional connectivity MRI between these networks was examined in ADHD, across
development, and in relation to attention. Children with ADHD had reduced negative
connectivity between task positive and task negative networks (p = 0.002). Connectivity
continues to become more negative between these networks throughout development
(7–15 years of age) in children with ADHD (p = 0.005). Regardless of group status, females
had increased negative connectivity (p = 0.003). In regards to attentional performance, the
ADHD group had poorer signal detection (d’) on the continuous performance task (CPT)
(p < 0.0001), more so on easy than difficult d’ trials (p < 0.0001). The reduced negative
connectivity in children with ADHD also relates to their attention, where increased negative
connectivity is related to better performance on the d’ measure of the CPT (p = 0.008).
These results highlight and further strengthen prior reports underscoring the role of
segregated system integrity in ADHD.
AUTHOR SUMMARY
Maintaining task focus has been thought to relate to the coordinated activity between brain
networks that support the initiation and maintenance of task sets (task positive networks) and
networks that mediate internally directed processes (i.e., the default mode network). Here
we find that segregation between these functional networks is impaired in children with
ADHD, shows developmental lag, and is related to attentional impairments as measured
by the continuous performance task. These results highlight and further strengthen prior
reports underscoring the role of segregated system integrity in ADHD and its relationship to
impairments in attention.
Citation: Mills, B. D., Miranda-
Dominguez, O., Mills, K., Earl, E.,
Cordova, M., Painter, J., . . . Fair, D. A.
(2018). ADHD and attentional control:
Impaired segregation of task positive
and task negative brain networks.
Network Neuroscience, 2(2), 200–217.
https://doi.org/10.1162/netn_a_00034
DOI:
https://doi.org/10.1162/netn_a_00034
Supporting Information:
https://doi.org/10.1162/netn_a_00034
Received: 1 May 2017
Accepted: 30 October 2017
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Author:
Damien A. Fair
faird@ohsu.edu
Handling Editor:
Lucina Q. Uddin
Copyright: © 2018
Massachusetts Institute of Technology
Published under a Creative Commons
Attribution 4.0 International
(CC BY 4.0) license
The MIT Press
ADHD and attentional control: Impaired segregation of brain networks
Resting-state functional connectivity:
An fMRI method for mapping
correlated patterns in spontaneous
blood oxygen level dependent signal
across the brain during wakeful rest.
Default mode network:
Also referred to as the task negative
network. The default mode is a
collection of brain regions whose
activities are highly correlated during
rest; during goal-directed tasks the
network reduces its baseline level of
activity.
Task positive network:
A collection of brain networks that
during goal-directed tasks typically
increase in baseline level of activity.
INTRODUCTION
Attention deficit hyperactivity disorder (ADHD) is a major public health concern that affects
an estimated 5% of children in the United States (Boyle et al., 2011; Faraone et al., 2015).
Children with ADHD have impairments in attention that are manifested in real-world negative
outcomes both in and out of the classroom (Biederman et al., 2004; Diamantopoulou, Rydell,
Thorell, & Bohlin, 2007; Miller & Hinshaw, 2010). Several lines of evidence suggest that
aspects of resting-state functional brain connectivity MRI (rs-fcMRI) are atypical in ADHD
(Castellanos & Aoki, 2016; Costa Dias et al., 2013; Fair, Nigg, et al., 2012; Konrad & Eickhoff,
2010; Krain & Castellanos, 2006).
In particular, prior theories have suggested that the ab-
sence of a negative correlation between task positive and task negative systems in the brain
might relate to attentional issues in participants in ADHD (Castellanos et al., 2008; Hoekzema
et al., 2014; Sonuga-Barke & Castellanos, 2007; Sripada, Kessler, & Angstadt, 2014; Sun et al.,
2012). These brain systems are believed to be important for external and internal information
processing, respectively (Dosenbach et al., 2007; Fox et al., 2005; J. D. Power, Fair, Schlaggar,
& Petersen, 2010). The negative correlations between them have been shown to be reliable and
robust across processing techniques (Fox, Zhang, Snyder, & Raichle, 2009; Hutchison et al.,
2012; C. Keller et al., 2013; Kelly et al., 2008; Yan, Craddock, Zuo, Zang, & Milham, 2013), are
related to modulations in local field potential power (Hutchison, Hashemi, Gati, Menon, &
Everling, 2015; C. Keller et al., 2013; Popa, Popescu, & Pare, 2009), and may reflect the
ongoing balance of specialization (and integration) between the distinct functional networks
they are composed of (Fox et al., 2005; Hutchison et al., 2015).
In the current report, we characterize connectivity between internally oriented systems
(i.e., the default mode network, DMN), and task positive, externally oriented networks, which
include regions in the cingulo-opercular, salience,
fronto-parietal, dorsal attention, and
ventral attention networks (Dosenbach et al., 2007; Fox et al., 2005; J. D. Power et al., 2010).
The DMN is a large and robustly replicable network that reduces its baseline level of activity
during a task (Broyd et al., 2009; Buckner, Andrews-Hanna, & Schacter, 2008). In contrast,
task positive networks increase their level of activity while engaging in tasks. These regions
have been implicated in response selection, planning of willful acts, and preparing to respond
to environmental events (Fransson, 2005, 2006). The task positive system is composed of
several separate subnetworks including the fronto-parietal, cingulo-opercular, salience, dorsal
attention, and ventral attention networks (Dosenbach et al., 2007; Fox et al., 2005; J. D. Power
et al., 2010). There is a striking degree of negative correlation between the DMN and task
positive networks that suggests a reciprocal relationship and a potential antagonism in terms
of the psychological functions these networks serve (Cocchi, Zalesky, Fornito, & Mattingley,
In fact, when individuals transition from
2013; Fox et al., 2005; Fransson, 2005, 2006).
internally focused cognition to goal-directed, attentionally demanding behaviors, the DMN
becomes more deactivated (Buckner et al., 2008).
The dynamics between these two networks not only may be important for characterizing
ADHD, but may be a critical component of typical brain organization. Interestingly, in typ-
ically developing children negative correlations between the DMN and regions in the task
positive network matures from childhood to early adulthood (7–21 years), and connections
that show age-related effects also show larger ADHD group differences (Sripada, Kessler, &
Angstadt, 2014). This suggests that ADHD may be characterized by dysmaturation between
connections that continue to mature during development (Sripada, Kessler, & Angstadt, 2014).
However, direct relationships between this system and age in ADHD has not been explored
Network Neuroscience
201
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
0
2
0
2
2
0
0
1
0
9
2
1
5
7
n
e
n
_
a
_
0
0
0
3
4
p
d
t
/
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
ADHD and attentional control: Impaired segregation of brain networks
Continuous performance task (CPT):
A behavioral measure of attentional
vigilance, which requires an
individual to respond to rare target
events presented amid frequent
nontarget events.
and it remains unclear whether the system changes in any group during early childhood and
adolescence (7–15 years).
Although ADHD has been generally characterized by underconnectivity between a subset
of connections between the DMN and task positive networks in ADHD (Castellanos et al.,
2008;Castellanos & Aoki, 2016;Hoekzema et al., 2014;Sripada, Kessler, Fang, et al., 2014;Sun
et al., 2012), findings are often connection specific, and the evidence remains inconclusive.
Deficits in this system are thought to play a key role in ADHD-related dysfunction but are
also potentially related to typical attentional processing. Kessler and colleagues showed that
maturation of this system may be related to attentional performance in healthy controls (Kessler,
Angstadt, & Sripada, 2016), but the role this system plays during earlier development and its
direct relationship to failures in various aspects of attention in ADHD have not been tested.
Of course, attention is a multifaceted construct with the specific subcomponents varying
somewhat by model (Fan & Posner, 2004; Posner & DiGirolamo, 1999). In the current report
we focus on the relationship of negative connectivity between task positive and task negative
networks and vigilance (which is related to the arousal function of attention). Vigilance is
defined as the ability to maintain attentional focus to rare events (Riccio & Reynolds, 2006;
Tucha et al., 2009). Vigilance is classically indexed by the continuous performance task (CPT),
which requires an individual to respond to rare target events presented amid frequent nontarget
events (Huang-Pollock, Karalunas, Tam, & Moore, 2012; Tucha et al., 2009). In many versions
of the task used in ADHD populations, the difficulty of discriminating target and nontarget
events is relatively easy (e.g., respond to the letter X but to no other letters), but it is also pos-
sible to manipulate the discrimination difficulty by including stimuli that are either more or
less distinct from each other (Cornblatt, Risch, Faris, Friedman, & Erlenmeyer-Kimling, 1988;
Huang-Pollock et al., 2012). There is also evidence that task difficulty may play a role with
several studies finding larger ADHD performance differences on easy relative to difficult dis-
criminations, interpreted as children with ADHD being less able to endogenously direct the
focus of their attention and thus more reliant on task demands to exogenously engage attention
(Friedman-Hill et al., 2010; Lenartowicz et al., 2014). Vigilance impairments are a prominent
deficit in children with ADHD (Huang-Pollock et al., 2012; Tucha et al., 2009); however, the
neural mechanisms of these attentional impairments in ADHD remain poorly understood.
This report studies a longitudinal sample of 432 subjects and 604 scans that were strictly
controlled for movement during the MRI. We investigate connectivity patterns between task
negative and task positive networks and their relationship with attentional processes utilizing
an equivalent-pairs CPT, a widely used and validated measurement sensitive to task vigilance
that also allows us to examine the relationship between brain networks and vigilance under
varying discrimination difficulty (Cornblatt et al., 1988; Huang-Pollock et al., 2012). We test
whether negative correlations between the DMN and task positive networks are reduced in
children with ADHD, whether these negative correlations strengthen with development, and
whether these connectivity patterns are related to attention measured by the CPT.
METHODS AND MATERIALS
Participants
This study included 502 participants with 816 MRI scans. Of these, a total of 432 children
with 604 scans met all our motion censoring criteria (see below) and were included in the final
analysis. The control group consisted of 176 children with a total of 231 total scans; 51 subjects
had two scans and two subjects had three scans. The ADHD group had 256 subjects, 373 total
Network Neuroscience
202
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
0
2
0
2
2
0
0
1
0
9
2
1
5
7
n
e
n
_
a
_
0
0
0
3
4
p
d
.
/
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
ADHD and attentional control: Impaired segregation of brain networks
scans; 85 subjects had two scans and 15 subjects had three scans. Children were between
7 and 11 years at baseline, and the lag design of approximately one year spans 7–15 years
(average lag 1.51 years control, 1.62 years ADHD). The average age of the control group was
10.67 years (SD = 1.53) and included 89 males. The average age of the ADHD group was
11.05 (SD = 1.58) and consisted of 181 males. Sex and age were included as covariates and
follow-up analyses matched the number of male and female scans included in each group.
See the Supplementary Information (Mills et al., 2018) for recruitment and diagnostics.
Human Connectome Project MRI Dataset
An independent dataset was used in order to define the connections that are most anti-
correlated between the task positive and task negative networks. For this analysis we used
data from the Human Connectome Project (HCP) consortium “500 Subject” release and
included 61 healthy adult control subjects (22–35 years of age, 26 males), which were se-
lected based on their optimal data quality and low motion (see Supplementary Information,
Mills et al., 2018). These data were obtained from the publically available HCP database
(https://db.humanconnectome.org).
Behavioral Testing: Continuous Performance Task
At each scan date, a version of the identical pairs continuous performance task (IP-CPT) was
used to examine vigilance and sustained attention (Cornblatt et al., 1988; Halperin, Sharma,
Greenblatt, & Schwartz, 1991). An IP-CPT was used because it is less vulnerable to the ceiling
effects that interfere with calculation of signal detection parameters in a less difficult CPT.
The version used here was modeled on tasks used successfully in other studies of ADHD
(Curko Kera, Marks, Berwid, Snatra, & Halperin, 2004). In the task, children viewed a series of
four-digit numbers displayed one at a time in pseudorandom order to ensure unpredictability
while achieving the required ratio of trial types. A total of 11 different four-digit numbers were
used. When two identical numbers appeared back-to-back, the child pushed the response
button. We used a 200 ms display followed by a 1,500 ms dark screen, for a total time per trial
of 1,700 ms. Target frequency was 20%. Another 20% of trials were “catch” trials in which
the back-to-back numbers differed by only one digit, creating a difficult discrimination, and
60% of trials were “stim” or “nontargets” in which subsequent numbers differed by multiple
digits, making them comparatively easy discriminations. With a total of 300 stimuli, the task
required about 10 min to complete. For both the easy (“stim”) and difficult (“catch”) trials, d’,
an index of perceptual sensitivity (i.e., the ability to discriminate between target and noise;
Stanislaw & Todorov, 1999), was calculated as our main index of vigilance (Cornblatt et al.,
1988; Halperin et al., 1991). A higher d’ score indicates better performance and a greater
sensitivity in distinguishing the target from the nontargets. D’ is typically interpreted as an
index of vigilance (Sergeant, Oosterlaan, & van der Meere, 1999) and is commonly impaired
in studies of ADHD (Huang-Pollock et al., 2012; Tucha et al., 2009; Willcutt, Doyle, Nigg,
Faraone, & Pennington, 2005). See the Supplementary Information (Mills et al., 2018) for
more detail.
Functional Connectivity MRI Processing
Children were scanned on a Siemens Tim Trio 3.0 Tesla MRI scanner, consisting of 15 min of
resting-state BOLD data acquired in three sequential 5-min runs. Strict motion censoring pro-
cedures were applied to resting-state functional maps, volumes with a framewise displacement
(FD; Fair, Nigg, et al., 2012; J. Power, Barnes, Snyder, Schlaggar, & Petersen, 2012) exceeding
0.2 mm were excluded, and only subjects with greater than 4 min of remaining motion-free
Network Neuroscience
203
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
0
2
0
2
2
0
0
1
0
9
2
1
5
7
n
e
n
_
a
_
0
0
0
3
4
p
d
t
.
/
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
ADHD and attentional control: Impaired segregation of brain networks
data were included in this analysis. For further motion quality control we removed an addi-
tional 30 scans whose average default to task positive connectivity was related to mean FD,
thus ensuring that data quality passed criteria as recently described by Siegel et al. (2016, see
Supplementary Information, Mills et al., 2018). Results are also presented without this sec-
ondary motion control (see Supplementary Information, Mills et al., 2018). In order to match
the amount of data included on all scans, connectivity matrices were calculated with 4 min
of randomly selected clean data for all scans. No differences in remaining FD were found
between groups (p = 0.48). For data analysis, we selected 333 regions of interest (ROIs) with
predefined network affiliation based on prior work (Gordon et al., 2014). These ROIs are in
good agreement with known functional brain systems and have been assigned functional iden-
tities and network definitions. For all participants, the resting blood oxygen level–dependent
time series for each region of interest was cross-correlated (Pearson correlations) between the
time series of all ROI pairs, yielding a correlation value that was then Fisher r to z transformed.
The final result was a 333 × 333 size correlation matrix for each subject. MRI scan parameters,
motion censoring, motion quality control, and preprocessing are described in full detail in the
Supplementary Information (Mills et al., 2018).
Network Definitions and Statistical Methods
Not all connections between task positive and task negative networks confer negative cor-
relations (Cocchi et al., 2013). Therefore, an independent dataset of 61 adults was used to
define the most highly negative connections between task negative (i.e., DMN) and task pos-
itive networks (i.e., fronto-parietal, cingulo-opercular, salience, dorsal attention, and ventral
attention networks; Gordon et al., 2014). The average connectivity between networks was then
computed and connections that were most highly anticorrelated were selected. For the main
analyses connections with an R < −0.35 were used (250 negative connections of 5,043 total
connections between default and task positive networks, corresponding to a connection den-
sity of 5%). See Figure 1 for these regions, their importance to the network mask, and their
network assignments. Less stringent thresholds and network masks corresponding to the DMN
to all negative connections, unrestricted to the task positive network, were also analyzed.
For the main analyses, connectivity between each of the 250 task positive and task neg-
ative connections was averaged for each scan. In each case, connectivity was then assessed
for main effects of group status, sex, age, and their interactions, controlling for subject head
motion (mean FD). If interaction terms are insignificant, only main effects will be modeled for
subsequent analyses, unless otherwise stated. Statistics were computed using a linear mixed-
effects model (R version 3.2.1; lme4 package).
Individual parameter effects were estimated
with the R package, lmerTest. Linear mixed effects modeling allows for an estimation of the
fixed effects while incorporating the longitudinal nature of the data by including within-subject
variation as nested random effects.
The R package MuMIn was used to evaluate the best fitting model and Akaike information
criterion (AIC) was evaluated for model selection and to guard against overfitting. Statistical
effects were evaluated with three models; Model 1 considered only main effects, Model 2
included a group by age interaction, and Model 3 was a full omnibus model including a three-
way interaction between age, group, and sex (see Supplementary Table 3 for formulas; Mills
et al., 2018). Model 1 had the lowest AIC and was used to test main effects and connection-
level effects; Model 3 was used to evaluate whether there were any interaction effects; and
Model 2 further tested significant interactions from Model 3, including the interaction between
age and group status on connectivity.
Network Neuroscience
204
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
0
2
0
2
2
0
0
1
0
9
2
1
5
7
n
e
n
_
a
_
0
0
0
3
4
p
d
/
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
ADHD and attentional control: Impaired segregation of brain networks
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
Figure 1. Regions with significant negative correlations between the (A) task negative and (B)
task positive networks. For the main analyses, 250 negative connections between systems were
analyzed (i.e., r < −0.35, 5% density). Regions are colored based on the importance of a given
region to the network mask, where brighter colors represent regions with more negative connections
between task positive and task negative systems. Unfilled regions do not have a connection within
this network mask threshold. Borders of the regions are colored based on network assignment
defined by Gordon et al. (2014). For illustrative purposes, (C) represents BOLD time series data
from a randomly selected control subject depicting the negatively correlated relationship between
a region of the task positive and task negative network.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
0
2
0
2
2
0
0
1
0
9
2
1
5
7
n
e
n
_
a
_
0
0
0
3
4
p
d
/
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Connection-level analyses separately modeled each of the 250 connections between
networks and identified connections that were significantly ( p < 0.05, false discovery rate
[FDR] corrected; Benjamini & Hochberg, 1995) related to a parameter of interest (using
Model 1). Results from these analyses are shown in Supplementary Table 4 (Mills et al., 2018).
Given that even trend-level results likely influence the conglomerate measures of connectiv-
ity, brain images shown in Figures 3 and 4 were constructed as follows. Significant t scores
( p < 0.05, uncorrected) for each connection were summed for each parameter effect of in-
terest in a given direction (i.e., ADHD < control). This summed t score for each region then
visualized on the cortical surface. This analysis depicts a region’s influence on each effect of
interest.
In order to ensure our results were not driven by an imbalance in gender distribution be-
tween groups (ADHD = 241 male scans, control = 123 male scans, 1.96:1 ADHD:control ra-
tio), we performed confirmatory analyses that matched the gender distribution between groups.
For these analyses 123 ADHD male scans and 100 ADHD female scans were randomly se-
lected (118 ADHD male scans and 8 ADHD female scans were randomly removed) in order
to match the control distribution. Statistical models were refit with this matched subset and
parameter effects were recomputed. This process was repeated 10,000 times, each time ran-
domly selecting a subset of ADHD subjects, and the distribution of p values were averaged to
Network Neuroscience
205
ADHD and attentional control: Impaired segregation of brain networks
obtain a gender-balanced effect for each effect of interest (age, group, gender, age by group
interaction, and CPT by behavior).
RESULTS
Negative Correlations
An independent dataset of 61 adults was used to define the connections between task neg-
ative (default) and task positive networks that were most highly anticorrelated. Connections
that survived a threshold of R < −0.35 (corresponding to the 5% strongest negative connec-
tions) were included in the network mask used for the main analyses. To ensure these effects
were not threshold dependent, supplementary analyses tested anticorrelated network masks of
R < −0.3 and R < −0.375 (see Supplementary Table 2, Mills et al., 2018). For the main anal-
yses, we found 250 negative connections from the default mode; the majority were from the
default to the cingulo-opercular networks (170 connections), followed by the dorsal attention
network (76 connections), and a small number of connections with the fronto-parietal network
(four connections, from two regions). No negative connections were found at this threshold
between the default and ventral attention or salience networks. See Supplementary Table 1
(Mills et al., 2018) for the number of regions included in each network and the number of con-
nections between each network. Follow-up analyses show a similar distribution of negative
connections throughout the task positive network when default connections were unrestricted
to task positive regions (see Supplementary Table 1, Mills et al., 2018).
A major aim of this study was to examine whether negative connectivity between task
positive and task negative networks is distinct in children with ADHD, between sexes, and
whether these connectivity patterns continue to mature across development. See Supplemen-
tary Table 3 (Mills et al., 2018) for the linear mixed-effects models to test these effects and their
interactions. Model 1 (main effects only) was the best fitting model (d f = 7, AIC = −1, 011)
followed by Model 2, which included all two-way interactions, (d f = 8, AIC = −1, 003),
followed by Model 3 which also included a three-way interaction between group, age, and
sex (d f = 11, AIC = −979).
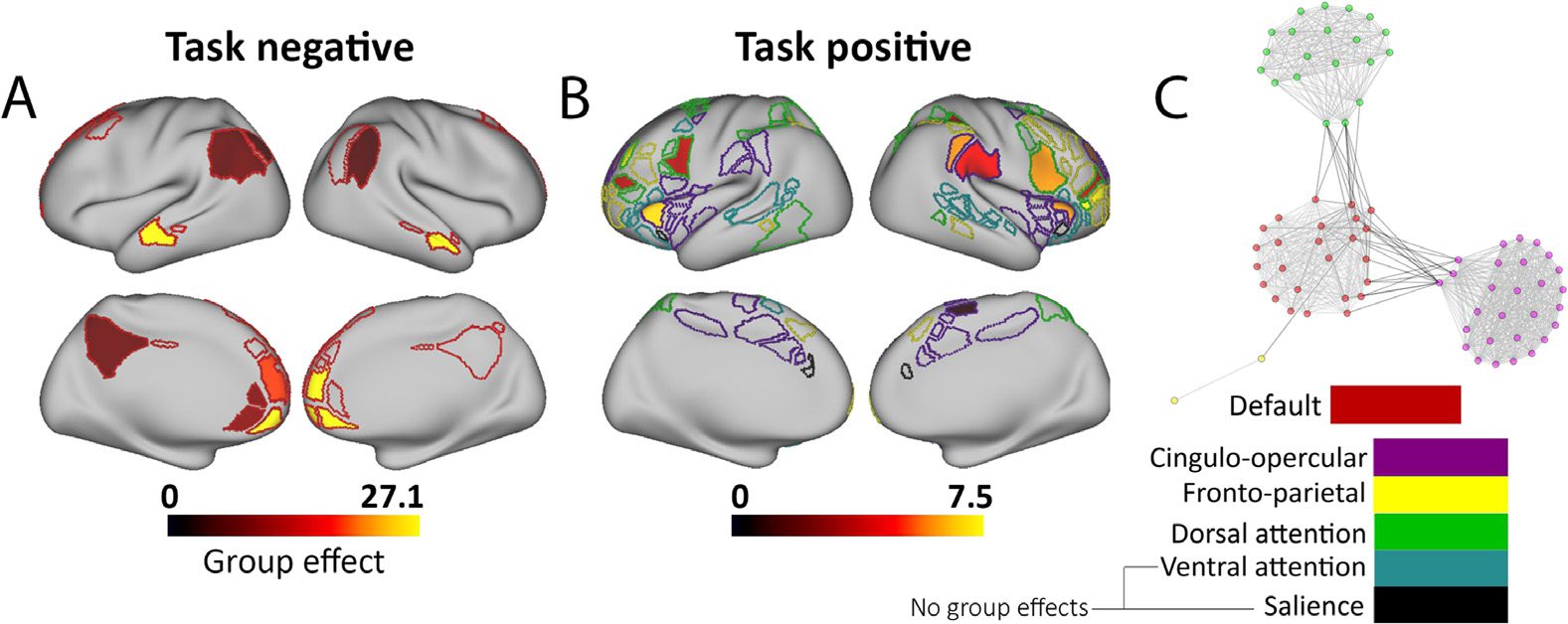
Negative connectivity is greater in controls than in children with ADHD
We found that ADHD subjects had reduced negative connectivity than control subjects. This
was consistent across all three statistical models (Model 1; main effect of group status, t = 3.03,
p = 0.002, TD M = −0.291, SD = 0.098, ADHD M = −0.264, SD = 0.109; see Figure 2, and
Supplementary Table 3, Mills et al., 2018, for each statistical model). The main effect of group
status was also robust to connection density when defining the most negative connections
between task negative and positive networks (R < −0.3, −0.35, −0.375; see Supplementary
Table 2, Mills et al., 2018), and the main effect of ADHD was preserved in analyses focusing
on these alternative network masks, including those from the default network to the rest of the
brain (see Supplementary Table 2, Mills et al., 2018). An additional analysis added CPT (d’
easy) as a covariate in the full model (Model 3). Although less significant than without including
this measure in the model, group (p = 0.040) and sex (p = 0.044) effects remained significant.
This suggests that these differences in connectivity are not purely due to vigilance deficits,
but are also related to other aspects of ADHD and sex.
In order to examine which connections were likely to drive effects in the main analyses,
the individual connections with the group effects (p < 0.05, uncorrected) are visualized in
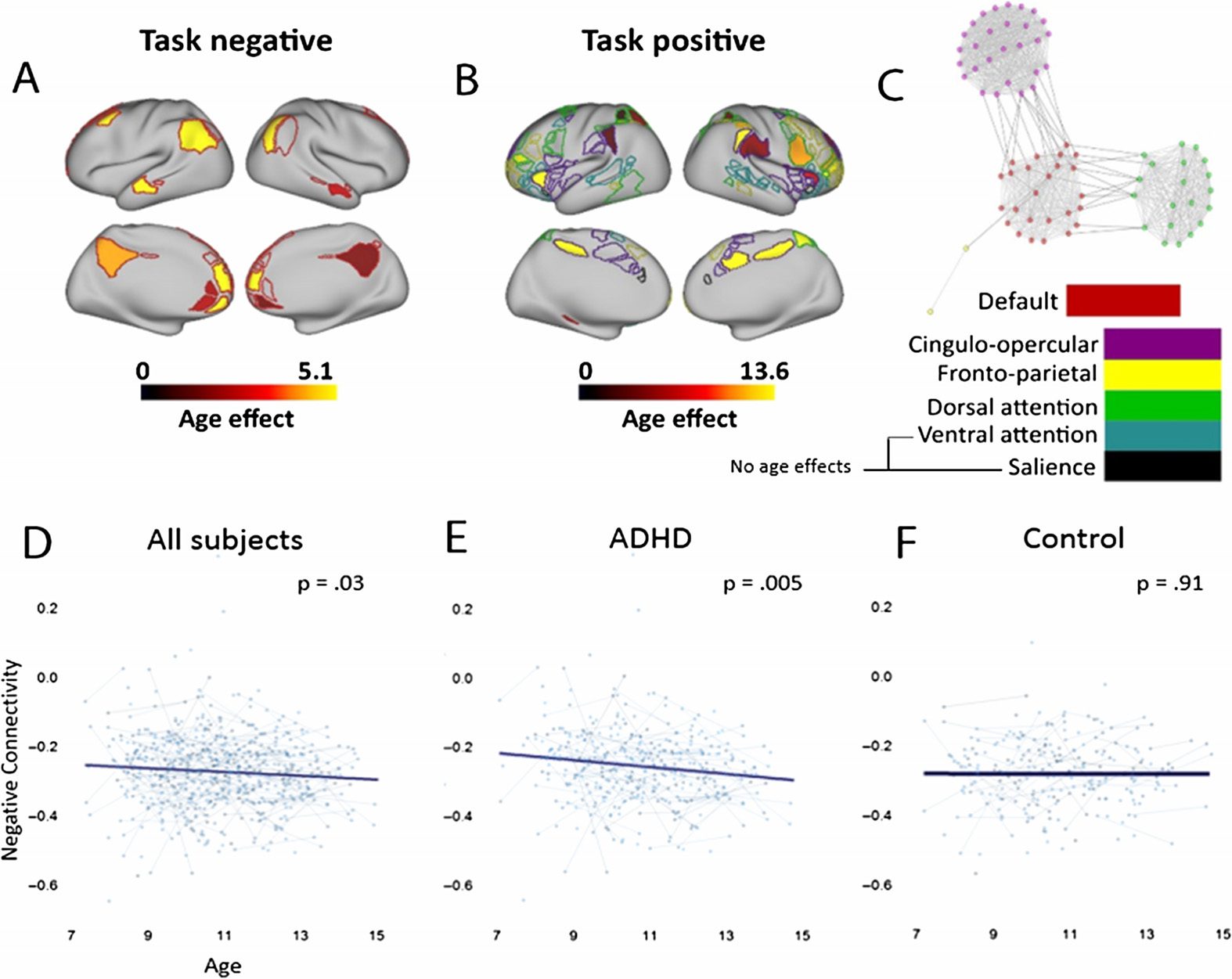
Figure 3, and each connection that shows a group effect that passes an FDR-corrected p < 0.05
is listed in Supplementary Table 4 (Mills et al., 2018).
Network Neuroscience
206
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
0
2
0
2
2
0
0
1
0
9
2
1
5
7
n
e
n
_
a
_
0
0
0
3
4
p
d
.
t
/
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
ADHD and attentional control: Impaired segregation of brain networks
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
Figure 2. Relationships between average negative connectivity, group status, gender, and motion.
(A) ADHD subjects have weaker negative connectivity between the task negative and task positive
networks. (B) Across groups, females have increased negative connectivity. (C) The ADHD group
difference is similar in males and females (i.e, no group by sex interaction). Given the longitudi-
nal nature of the data, main effects and interactions were tested in a linear mixed-effects model.
(D–E). Head motion, measured by mean remaining frame displacement (FD), is not related to
average negative connectivity in either (D) control subjects (r = 0.08, p = 0.19) or (E) ADHD
(r = 0.06, p = 0.22).
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
0
2
0
2
2
0
0
1
0
9
2
1
5
7
n
e
n
_
a
_
0
0
0
3
4
p
d
/
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
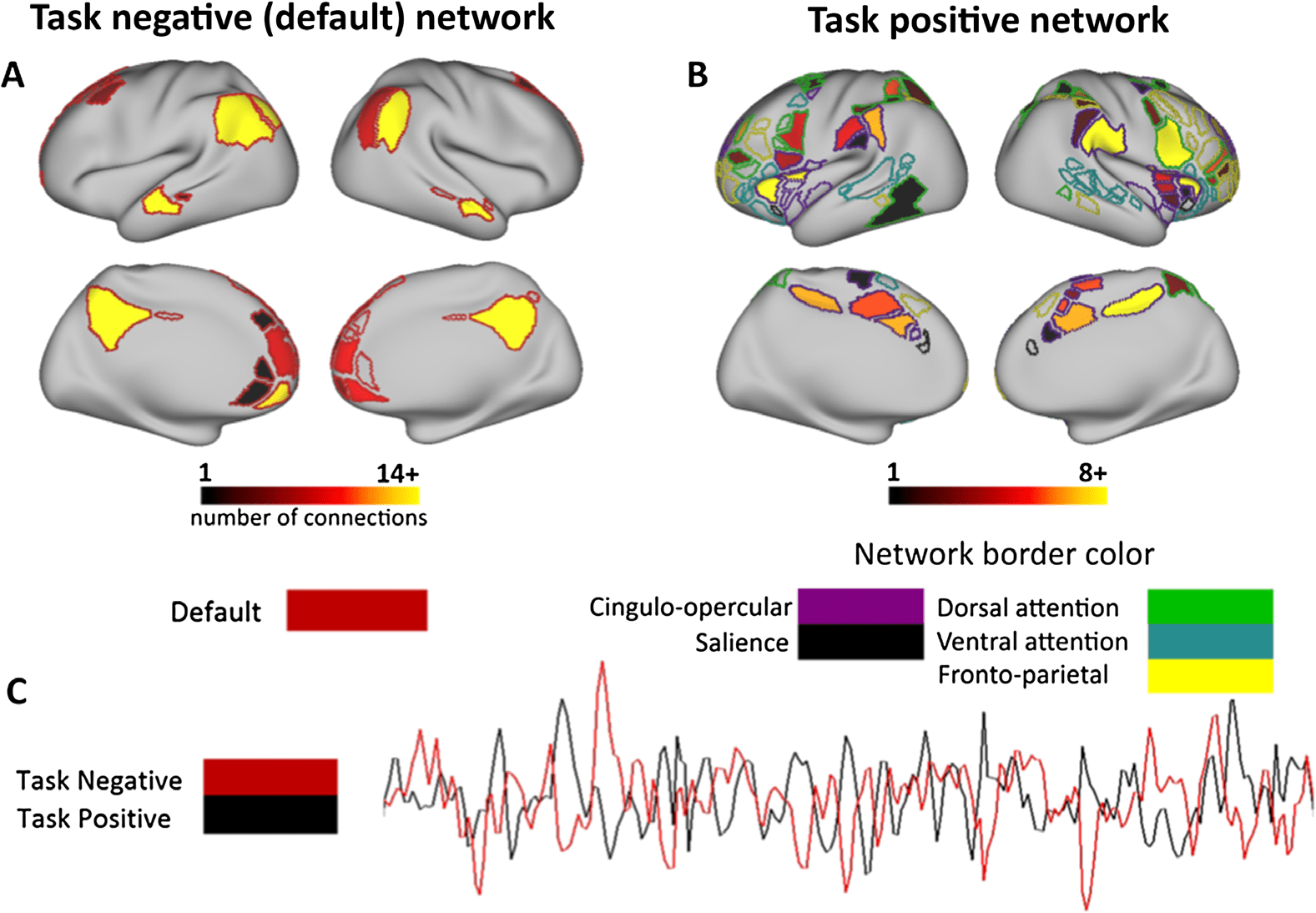
Figure 3. ADHD effects visualized on the brain. ADHD subjects have reduced negative connec-
tivity between many connections of the task negative and task positive networks. The group effect
(summed t scores for all group effects in a given region) are shown, where brighter colors indicate a
region where connections are weaker in ADHD subjects than controls. Connections are visualized
between the (A) task negative and (B) task positive network. (C) A statistical distance metric visual-
izes the number and origin of differences in negative connectivity between groups. Nodes that are
closer to the center are connected by solid black lines and denote a significant group effect from
a given task positive network to the task negative network (default mode in the center). Note that
many of the group effects that show weaker negative connectivity are between the default network
and the cingulo-opercular and dorsal attention network.
Network Neuroscience
207
ADHD and attentional control: Impaired segregation of brain networks
Negative connectivity is greater in females than in males
Across groups, girls had greater negative connectivity than boys (t = −2.93, p = 0.003; see
Figure 2B). When examining only control subjects, this sex effect nearly reached statistical
significance (p = 0.056). We found no group by sex interaction (p > 0.5; see Figure 2C). This
suggests that the ADHD effect is similar for males and females and that the increased negative
connectivity observed in females is independent of diagnosis.
Negative connectivity is related to age in children with ADHD
In order to assess whether this system continues to mature through development, a main effect
of age shows that negative connectivity continues to increase with age (main effect of age
Model 1, t = −2.249, p = 0.025). However, we also found a trend towards an age by group
interaction (Model 3, t = −1.91, p = 0.057; Model 2, t = −1.836, p = 0.06), where children
with ADHD showed a stronger age by connectivity relationship (age parameter effect with
only ADHD subjects, t = −2.76, p = 0.005) than controls (only control subjects, t = 1.01,
p = 0.91). This suggests that the effect of age may be driven by children with ADHD (see
Figures 4D, 4E, and 4F). No other interaction term reached significance.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
0
2
0
2
2
0
0
1
0
9
2
1
5
7
n
e
n
_
a
_
0
0
0
3
4
p
d
t
/
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. Connectivity between the task positive and task negative network becomes increasingly
negative with age. The relative age effects (summed t scores for all age effects in a given region)
are shown for regions within the (A) task negative and (B) task positive network. Brighter colors
indicate regions with more age effects, where negative connectivity increases with age. (C) Solid
black lines denote a significant age effect from a given region within the task positive network, to
a region within the task negative network. (D) Averaged negative connectivity between networks
decreases with age, across both groups. This effect is driven by a significant relationship between
age and connectivity in (E) ADHD rather than (F) control subjects.
Network Neuroscience
208
ADHD and attentional control: Impaired segregation of brain networks
Overall, these findings suggest that negative connectivity between task positive and task
negative networks is decreased in children with ADHD, that negative connectivity is greater
in females regardless of group status, and that negative connectivity continues to increase in
children with ADHD.
Effect of Head Motion on Connectivity and Behavior
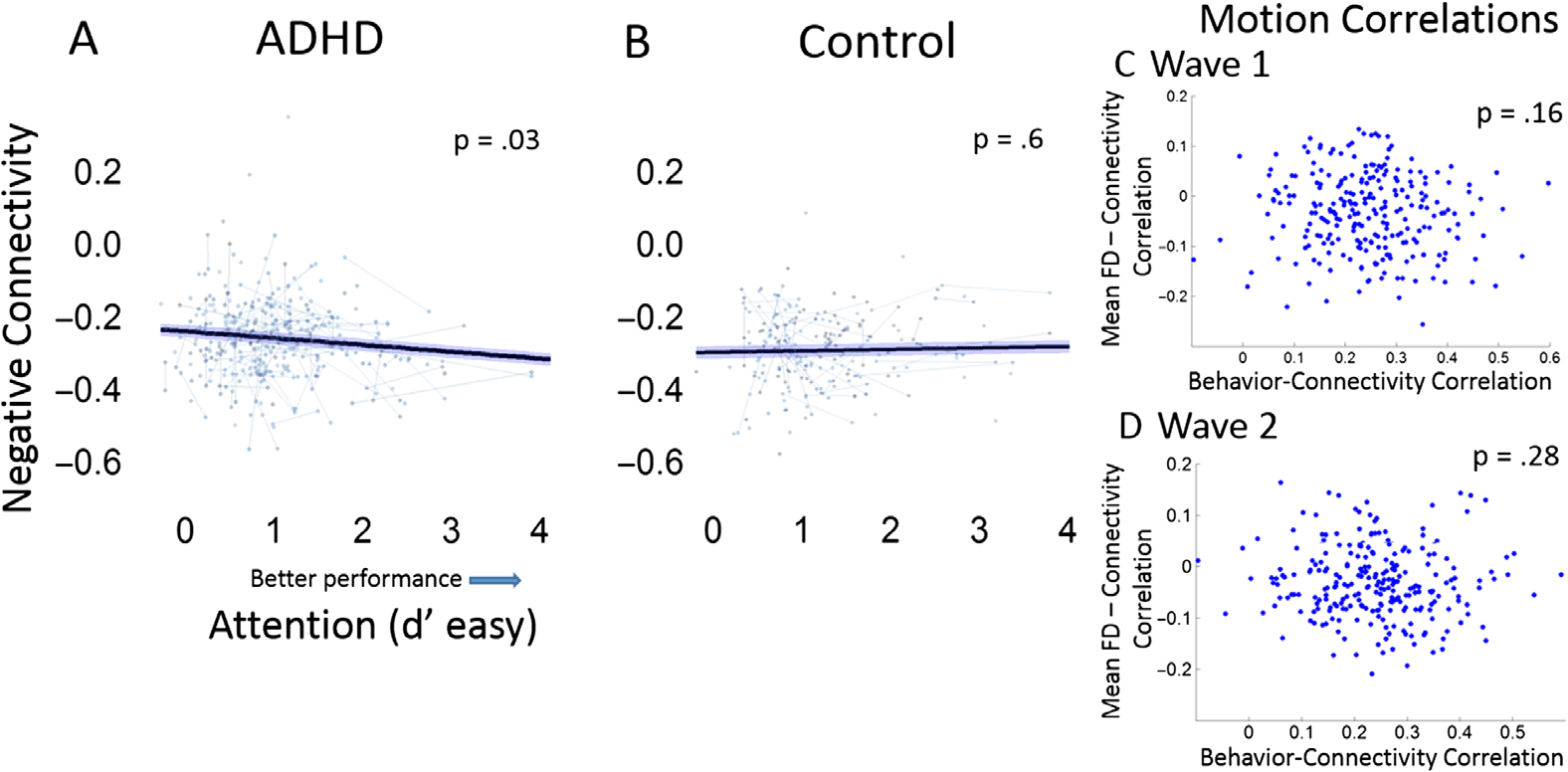
Importantly, head motion, a major confound in neuroimaging studies, was not related to the
average connectivity metric or average connectivity by behavior findings (see Figures 2D and 2E
and Figures 5C and 5D). Age was also unrelated to mean remaining FD (R = −0.042, p = 0.3).
Results are also robust to connection density of the network mask (see Supplementary Informa-
tion for r < −0.3 and r < −0.375; Mills et al., 2018) and are maintained without the inclusion
of mean FD as a model covariate (data not shown); these relationships are also maintained
after the reinclusion of the 30 scans filtered based on the secondary motion-censoring criteria
(see Supplementary Figure 1, Mills et al., 2018).
Gender-Balanced Analyses
Finally, in order to address the gender imbalance in our sample (male subjects ~2:1 ADHD:
control), a permutation approach was taken to randomly select subjects from the ADHD group
to match the gender distribution in the control sample. P values for each parameter effect were
then averaged across 10,000 permutations to assess each effect’s significance. When model-
ing the main effects (Model 1), group status (p = 0.0081) and gender (p = 0.0083) remained
significantly related to negative connectivity. After balancing the distribution of genders be-
tween groups, the main effect of age is no longer evident across groups (p = 0.187), but may
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
0
2
0
2
2
0
0
1
0
9
2
1
5
7
n
e
n
_
a
_
0
0
0
3
4
p
d
.
/
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 5. Attention as measured by the CPT (d’ easy low-discrimination trials) is related to nega-
tive connectivity between task positive and negative networks in subjects with ADHD. (A) In ADHD
subjects, greater negative connectivity between networks is related to better attentional performance
on the CPT (t = −2.12, p = 0.03). (B) No relationship is found in TD children (t = 0.61, p = 0.60).
Each dot represents a scan and gray lines connect longitudinal scans for each subject. 95% confi-
dence intervals around the slope are shown. Importantly, the behavior connectivity relationship is
not related to in scanner head motion at either behavioral and MRI scan (C) Wave 1 (p = 0.16) or
(D) Wave 2 (p = 0.28).
Network Neuroscience
209
ADHD and attentional control: Impaired segregation of brain networks
be driven by a trend-level effect in ADHD subjects (group by age interaction effect, Model 2,
p = 0.087).
Connection-Level Analyses
Each of the analyses above focused on main effects on an averaged metric of negative con-
nectivity between task positive and task negative networks. In order to examine the particular
connections driving this effect, we next assessed the individual connections driving group, sex,
and age effects. A list of connections between these networks (250-connection network mask),
which showed significant main effects of group, age, and sex, are listed in Supplementary
Table 4 (Mills et al., 2018). Significance was calculated based on a p < 0.05 (FDR corrected)
using Model 1. Of note is that an abundance of connections influencing the group and age ef-
fects are found between the default mode and dorsal attention and cingulo-opercular networks,
and that only connections between default and the cingulo-opercular drive gender effects.
Behavioral Results
As expected, we found that children with ADHD had worse performance on the d’ measure
of the CPT, an index assessing attentional vigilance (group effect controlling for age, sex, and
task difficulty, p < 0.0001). We also found that performance on the CPT improved with age
(p < 0.0001). Importantly, we found a group by task difficulty interaction (interaction effect
controlling for age and sex (p < 0.0001), where ADHD children had worse performance than
controls on easy discrimination trials (TD M = 2.76, SD = 0.84; ADHD M = 2.21, SD = 0.93)
than on difficult trails (TD M = 1.34, SD = 0.79; ADHD M = 1.01, SD = 0.69). Although
children with ADHD were impaired on both easy and difficult discriminations, the larger differ-
ence on easy trials suggests that children with ADHD have more attentional difficulties when
task demands are low. No other interactions were significant, and gender was unrelated to
performance on the CPT (p = 0.61).
Brain Behavior Relationships
In a repeated measures analyses including all subjects and both d’ measures of task difficulty
(i.e., easy and difficult trials), more negative connectivity was related to better attentional per-
formance (p = 0.0064). This relationship was similar for easy and difficult trials (interaction
effect between connectivity and d’ difficulty, p = 0.162), suggesting that connectivity was re-
lated to both difficult (B = −1.33, t = −2.11, p = 0.038) and easy (B = −2.81, t = −3.362,
p = 0.008) d’ measures (all analyses controlled for mean FD). This relationship was not moder-
ated by age or sex. However, a trend was observed where ADHD subjects showed a stronger
relationship between average connectivity and the d’ (easy) than controls (group by behavior
interaction p = 0.072). Post hoc analyses in the control group show no relationship between
connectivity and d’ easy (p = 0.603) but do show a significant relationship when only an-
alyzing ADHD subjects (p = 0.033). Difficult d’ conditions are trend level in the ADHD
group (p = 0.103) and nonsignificant in control subjects (p = 0.342). Taken together this
suggests that the relationship between easy discriminability trials on the CPT and average con-
nectivity is driven by subjects with ADHD. These findings were also unrelated to motion (see
Figures 5C and 5D), and the CPT-connectivity relationship is conserved with the inclusion of
the 30 scans filtered based on the secondary motion-censoring criteria (see Supplementary
Information, Mills et al., 2018). In the gender-balanced analyses, we found a significant re-
lationship between CPT and negative connectivity in the easy (p = 0.045) but not difficult
trials (p = 0.13). In the gender-balanced analysis, the CPT-connectivity relationship was only
Network Neuroscience
210
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
0
2
0
2
2
0
0
1
0
9
2
1
5
7
n
e
n
_
a
_
0
0
0
3
4
p
d
t
/
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
ADHD and attentional control: Impaired segregation of brain networks
somewhat moderated by group status (p = 0.11), potentially because of a decrease in statistical
power.
DISCUSSION
Negatively correlated activity between task positive and task negative brain networks has pre-
viously been proposed to support attentional processes (Castellanos & Aoki, 2016; Weissman,
Warner, & Woldorff, 2004). This study extends an emerging body of literature suggesting that
negative functional connectivity between this system is a locus of dysfunction in ADHD and
shows developmental lag in ADHD. It also demonstrates that these networks are negatively
connected in females regardless of group status and that dysfunction of this system may be
related to attentional impairments in childhood ADHD.
There Are Several Benefits of the Current Study Relative to the State of the Literature
Reduced negative connectivity between negative and task positive networks in ADHD was a
robust finding across statistical models, with and without matching gender distributions in our
sample, and was significant across a range of connection densities. Although other studies
have found reduced negative connectivity between particular task positive to task negative
connections (Castellanos et al., 2008; Castellanos & Aoki, 2016; Hoekzema et al., 2014;
Sripada, Kessler, Fang, et al., 2014; Sun et al., 2012), this study benefited from an independent
and well-curated dataset that defined the set of the most highly negative connections between
these well-defined networks. This network mask was then used, at various thresholds, to com-
pute an average connectivity metric for each scan. This allowed for a direct and straightforward
assessment of the effect of age, sex, their interactions, and relationships to behavior. This was
all done without suffering from issues dealing with multiple comparisons and easing inter-
pretability of these effects. That being said, the work also highlights particular connections
and networks that may be most perturbed within this averaged connectivity metric. These in-
clude a number of connections showing reduced negative connectivity in ADHD between the
default, cingulo-opercular, and dorsal attention networks. It should be noted, however, that the
particular connection and network effects may be biased towards the number of ROIs within
each network, and these effects may shift based on connection density used in anticorrelated
network mask and may change based on parcellation scheme. Nonetheless, the general trends
found here are unlikely to be affected by such factors.
Another strength of this study is the large longitudinal sample collected at a single site. To
the best of our knowledge, previous literature examining similar group effects have generally
consisted of around 20 subjects per group (Castellanos et al., 2008; Hoekzema et al., 2014;
Sun et al., 2012), with the exception of one large multisite study (Sripada, Kessler, & Angstadt,
2014). Although multisite studies can be powerful in their own right, many potential confounds
can arise when combining subjects across studies and across sites, including differences in MRI
acquisition protocols and data quality (Abraham et al., 2017; Friedman, Hastie, & Tibshirani,
2008), differences in participant instructions (e.g., eyes open vs. eyes closed; Yan et al., 2013),
as well as differences in recruitment strategy and methods for diagnoses (Abraham et al., 2017).
Even if differences between sites are known, they can be difficult to accurately model, and
can make it more challenging to isolate the true relationships of the variable of interest. Our
study avoids these issues because it is large and homogenous in all of its procedures, and
ridged diagnoses. This does not mean, of course, that other multisite studies are not useful or
helpful, or that they don’t have their own advantages, but having a large homogenous sample
for scientific purposes is a strong positive of the current study. Another strength of this study is
Network Neuroscience
211
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
0
2
0
2
2
0
0
1
0
9
2
1
5
7
n
e
n
_
a
_
0
0
0
3
4
p
d
t
.
/
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
ADHD and attentional control: Impaired segregation of brain networks
in its use of the human connectome processing pipeline, which has many advantages including
more accurate spatial localization and improved signal to noise (Smith et al., 2013; Van Essen
et al., 2012). The current study also uses the most up-to-date and stringent motion correction
criteria, an important confound particularly in developmental studies (J. Power et al., 2012;
Yerys et al., 2009). These safeguards helped ensure that both group differences as well as
imaging-behavior relationships (Siegel et al., 2016) were unrelated to in-scanner head motion.
Negative Connectivity Continues to Develop in Children With ADHD
As stated previously, children with ADHD showed a significant relationship between aver-
age negative connectivity and age, and no such relationship was found in typical children
during this age bracket (7–15 years). These findings might suggest a few things. The first is
that in typical children, this system is developed by early childhood and will not continue
to mature into the early teen years. While there are several pieces of literature that would
suggest the continued maturation of this system (J. Keller et al., 2015; Kelly et al., 2008; Luna,
Marek, Larsen, Tervo-Clemmens, & Chahal, 2015; Marek et al., 2015; Sato et al., 2016; Spreng,
Stevens, Viviano, & Schacter, 2016; Sripada, Kessler, & Angstadt, 2014; Sun et al., 2012), strict
experimental controls on this large of a functional dataset in ADHD is the first of its kind, and
thus might run counter to prior reports. Moreover, we are referring here to findings related to
average connectivity between these systems, and individual connections may show develop-
mental maturation between these ages in typical controls. It remains possible that individual
connections increase in negative connectivity throughout typical development, but that these
are subtler changes, or more variable across connections. This would result in differences that
would be averaged out in our single metric of negative connectivity and appear as a lack of
development of this system in typical controls.
Under this context, in regards to the significant age relationship in children with ADHD,
our findings might suggest the underdevelopment of this system relative to controls, and thus,
their negative connectivity continues to mature during 7–15 years of age. Consistent with some
prior reports (El-Sayed, Larsson, Persson, Santosh, & Rydelius, 2003; Shaw et al., 2007; Sripada,
Kessler, & Angstadt, 2014), this would hint at a maturational lag. The findings and this inter-
pretation are also consistent with others who have found that individual negative connections
that show ADHD effects are also the connections that show continued development with age
(Sripada, Kessler, & Angstadt, 2014). As larger datasets are acquired and children are tracked,
future research should directly assess developmental lag in ADHD across the life span.
Females Have Increased Negative Connectivity Compared With Males
Interestingly, across groups, females had increased negative connectivity between the DMN
and task positive networks. Boys are over twice as likely to be diagnosed with ADHD as girls
(Boyle et al., 2011), and this led to an imbalance in the proportion of males in the ADHD group
compared with the control group (~2:1). Importantly, our results were robust to permutation
tests randomly matching groups by gender distribution. It has been reported that females have
greater connectivity within the DMN (Allen et al., 2011; Filippi et al., 2013; Tomasi & Volkow,
2012); however, studies of attention networks are mixed (Allen et al., 2011; Bluhm et al., 2008;
Filippi et al., 2013; Scheinost et al., 2015). To the best of our knowledge, this work is the first to
show that negative connectivity from the DMN to task positive system may also be an impor-
tant locus of sex-dependent differences in brain connectivity during early childhood. Although
females with ADHD had greater negative connectivity compared with males with ADHD, the
relative decrease compared with controls was the same for both genders. That being said,
Network Neuroscience
212
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
0
2
0
2
2
0
0
1
0
9
2
1
5
7
n
e
n
_
a
_
0
0
0
3
4
p
d
/
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
ADHD and attentional control: Impaired segregation of brain networks
ADHD females had a similar degree of negative connectivity as control males. It is challenging
to speculate whether the similar blunting of this network signifies a shared mechanism for
ADHD across genders, or whether the relatively intact network structure in ADHD females
suggests a distinct etiology and presentation of ADHD. That being said, decreased baseline
connectivity between these networks in males could represent a vulnerability for males to
develop attentional difficulties, and potentially contribute to the 2:1 male bias in ADHD.
These preliminary speculations highlight an avenue for future research to explore.
Attentional Performance Is Decreased in ADHD and Is Related to Negative Connectivity
As expected, we found that children with ADHD show impairments on attentional vigilance as
measured by the CPT. These deficits were greater in easy discrimination trials, compared with
more difficult discrimination trials. This is in line with previous research showing that attention
performance may improve in ADHD with increasing cognitive load (Forster & Lavie, 2016;
Friedman-Hill et al., 2010; Lenartowicz et al., 2014) and suggests that children with ADHD
may show deficits in attentional filtering particularly when task demands are low, although
evidence is mixed (Friedman-Hill et al., 2010; Huang-Pollock et al., 2005).
Importantly, reduced negative connectivity was related to worse vigilance, specifically in
children with ADHD. This suggests that impaired development of this system may contribute
to the observed problems in attention. No relationships were found in typical children, which
could be due to a number of reasons. One possible reason is that it is unlikely that this system
is the only driver of performance on the CPT; rather, when this system is highly atypical it can
disrupt performance on this task. It is also possible that given a more difficult task, correla-
tions between these resting-state networks and attention may emerge, even within the typical
variation seen in controls. Importantly, not all connections between the default mode and task
positive networks confer negative correlations. Here we select only the most consistently neg-
ative connections defined in an independent dataset. The selection of these connections may
be somewhat restrictive and could potentially shift based on transitory brain states that are not
captured during a rest (Cocchi et al., 2013). Therefore, these relationships could potentially
change if sampled throughout all connections between these networks rather than only from
the most consistently anticorrelated connections between task positive and negative networks.
Last, we note that ADHD is a vastly heterogeneous disorder encompassing different cognitive
(Fair, Bathula, Nikolas, & Nigg, 2012; Karalunas et al., 2014) as well as neural phenotypes
(Costa Dias et al., 2013; Gates, Molenaar, Iyer, Nigg, & Fair, 2014). Just as importantly, the
control population is also heterogeneous in a similar fashion (Fair, Bathula et al., 2012). It is
possible that performance on the CPT and its relationship to negative connectivity might vary
relative to various cognitive profiles, which might be skewed within the control and ADHD
samples. Further work will be needed to clarify such possibilities, and to refine whether this
network is implicated in deficits in a diverse set of attentional mechanisms, such as alerting,
orienting, or top-down attention (Posner & Petersen, 1990), or whether it selectively supports
only one domain of attention.
CONCLUSIONS
In summary, in a large cohort of children with ADHD with strict motion correction criteria,
negative connectivity between task positive and task negative networks is reduced in ADHD,
continues to develop with age, and is related to impairments in attentional vigilance. These
findings highlight a potential neurophenotype for at least a subset of children with ADHD and
serves as a potential marker of attentional impairments in this population.
Network Neuroscience
213
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
0
2
0
2
2
0
0
1
0
9
2
1
5
7
n
e
n
_
a
_
0
0
0
3
4
p
d
t
/
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
ADHD and attentional control: Impaired segregation of brain networks
AUTHOR CONTRIBUTIONS
Brian D. Mills: Conceptualization; Data curation; Formal analysis; Investigation; Methodol-
ogy; Project administration; Validation; Visualization; Writing – original draft; Writing – review
& editing. Oscar Miranda-Dominguez: Data curation; Methodology; Software. Kathryn Mills:
Methodology; Writing – review & editing. Eric Earl: Data curation; Writing – review & editing.
Michaela Cordova: Data curation; Investigation.
Julia Painter: Data curation; Investigation.
Sarah L. Karalunas: Conceptualization; Methodology; Writing – review & editing. Joel T. Nigg:
Funding acquisition; Investigation; Methodology; Project administration; Resources; Writing –
review & editing. Damien A. Fair: Conceptualization; Funding acquisition; Investigation;
Methodology; Project administration; Resources; Software; Supervision; Writing – review &
editing.
FUNDING INFORMATION
Funding for this project comes from the grants MH086654 (Nigg, Fair), MH096773 (Fair),
MH099064 (Nigg), and MH091238 (Fair). Data were also provided (in part) by the Human
Connectome Project, WU-Minn Consortium (principal investigators: David Van Essen and
Kamil Ugurbil; 1U54MH091657) funded by the 16 NIH Institutes and Centers that support
the NIH Blueprint for Neuroscience Research; OHSU Fellowship for Diversity and Inclusion
in Research Program (Oscar Miranda-Dominguez); and by the McDonnell Center for Systems
Neuroscience at Washington University.
REFERENCES
Abraham, A., Milham, M. P., Di Martino, A., Craddock, R. C.,
Samaras, D., Thirion, B., & Varoquaux, G. (2017). Deriving
reproducible biomarkers from multi-site resting-state data: An
autism-based example. NeuroImage, 147, 736–745. https://doi.
org/10.1016/j.neuroimage.2016.10.045
Allen, E. A., Erhardt, E. B., Damaraju, E., Gruner, W., Segall,
J. M., Silva, R. F., . . . Calhoun, V. D. (2011). A baseline for the
multivariate comparison of resting-state networks. Frontiers in
Systems Neuroscience, 5, 2. https://doi.org/10.3389/fnsys.2011.
00002
Benjamini, Y., & Hochberg, Y.
(1995). Controlling the false
discovery rate: A practical and powerful approach to multi-
ple testing. Journal of the Royal Statistical Society: B, 57(1),
289–300.
Biederman, J., Monuteaux, M. C., Doyle, A. E., Seidman, L. J.,
Wilens, T. E., Ferrero, F., . . . Faraone, S. V.
Impact of
executive function deficits and attention-deficit/hyperactivity
disorder (ADHD) on academic outcomes in children. Journal of
Consulting and Clinical Psychology, 72(5), 757–766. https://doi.
org/10.1037/0022-006X.72.5.757
(2004).
Bluhm, R. L., Osuch, E. A., Lanius, R. A., Boksman, K., Neufeld,
(2008). Default mode
R. W., Theberge,
J., Williamson, P.
network connectivity: Effects of age, sex, and analytic ap-
proach. Neuroreport, 19(8), 887–891. https://doi.org/10.1097/
WNR.0b013e328300ebbf
Boyle, C. A., Boulet, S., Schieve, L. A., Cohen, R. A., Blumberg,
S. J., Yeargin-Allsopp, M., . . . Kogan, M. D. (2011). Trends in the
prevalence of developmental disabilities in US children, 1997–
2008. Pediatrics, 127(6), 1034–1042. https://doi.org/10.1542/
peds.2010-2989
Broyd, S. J., Demanuele, C., Debener, S., Helps, S. K., James, C. J.,
& Sonuga-Barke, E. J. S. (2009). Default-mode brain dysfunction
in mental disorders: A systematic review. Neuroscience and
Biobehavioral Reviews, 33(3), 279–296. https://doi.org/10.1016/
j.neubiorev.2008.09.002
Buckner, R. L., Andrews-Hanna, J. R., & Schacter, D. L.
(2008).
The brain’s default network: Anatomy, function, and relevance
to disease. Annals of the New York Academy of Sciences, 1124,
1–38. https://doi.org/10.1196/annals.1440.011
Castellanos, F. X., & Aoki, Y. (2016). Intrinsic functional connec-
tivity in attention-deficit/hyperactivity disorder: A science in de-
velopment. Biological Psychiatry: Cognitive Neuroscience and
Neuroimaging,1(3), 253--261. https://doi.org/10.1016/j.bpsc.2016.
03.004
Castellanos, F. X., Margulies, D. S., Kelly, A. M. C., Uddin,
L. Q., Ghaffari, M., Kirsch, A., . . . Kelly, C. (2008). Cingulate-
precuneus interactions: A new locus of dysfunction in adult
attention-deficit/hyperactivity disorder. Biological Psychiatry,
63(3), 332–337. https://doi.org/10.1016/j.biopsych.2007.06.025
Cocchi, L., Zalesky, A., Fornito, A., & Mattingley, J. B. (2013).
Dynamic cooperation and competition between brain systems
during cognitive control. Trends in Cognitive Sciences, 17(10),
493–501. https://doi.org/10.1016/j.tics.2013.08.006
Cornblatt, B. A., Risch, N. J., Faris, G., Friedman, D., & Erlenmeyer-
Kimling, L. (1988). The continuous performance test, identical
pairs version (CPT-IP): I. New findings about sustained atten-
tion in normal families. Psychiatry Research, 26(2), 223–38.
Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/3237915
Iyer, S. P.,
Mills, K. L., Thurlow, B. L., . . . Fair, D. A. (2013). Reward
Costa Dias, T. G., Wilson, V. B., Bathula, D. R.,
Network Neuroscience
214
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
0
2
0
2
2
0
0
1
0
9
2
1
5
7
n
e
n
_
a
_
0
0
0
3
4
p
d
t
.
/
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
ADHD and attentional control: Impaired segregation of brain networks
circuit connectivity relates to delay discounting in children with
attention-deficit/hyperactivity disorder. European Neuropsycho-
pharmacology, 23(1), 33–45. https://doi.org/10.1016/j.euroneuro.
2012.10.015
Curko Kera, E. A., Marks, D. J., Berwid, O. G., Snatra, A., &
Halperin, J. M. (2004). Self-report and objective measures of
ADHD-related behaviors in parents of preschool children at risk
for ADHD. CNS Spectrums, 9(9), 639–647.
Diamantopoulou, S., Rydell, A.-M., Thorell, L. B., & Bohlin, G.
(2007). Impact of executive functioning and symptoms of at-
tention deficit hyperactivity disorder on children’s peer relations
and school performance. Developmental Neuropsychology, 32,
521–542. https://doi.org/10.1080/87565640701360981
Dosenbach, N., Fair, D. A., Miezin, F. M., Cohen, A. L.,
Wenger, K. K., Dosenbach, R. A. T., . . . Petersen, S. E.
(2007).
Distinct brain networks for adaptive and stable task control
in humans. Proceedings of
the National Academy of Sci-
ences, 104(26), 11073–11078. https://doi.org/10.1073/pnas.
0704320104
El-Sayed, E., Larsson, J., Persson, H., Santosh, P., & Rydelius, P.
(2003). “Maturational lag” hypothesis of attention deficit hyper-
activity disorder: An update. Acta Paediatrica, 92(7), 776–784.
https://doi.org/10.1080/08035250310002777
Fair, D. A., Bathula, D., Nikolas, M. A., & Nigg, J. T. (2012). Distinct
neuropsychological subgroups in typically developing youth in-
form heterogeneity in children with ADHD. Proceedings of the
National Academy of Sciences, 109(17), 6769–6774. https://doi.
org/10.1073/pnas.1115365109
Fair, D. A., Nigg, J. T., Iyer, S., Bathula, D., Mills, K. L., Dosenbach,
N. U. F., . . . Milham, M. P. (2012). Distinct neural signatures de-
tected for ADHD subtypes after controlling for micro-movements
in resting state functional connectivity MRI data. Frontiers in
Systems Neuroscience, 6(February), 80. https://doi.org/10.3389/
fnsys.2012.00080
Fan,
J., & Posner, M.
(2004). Human attentional networks.
Psychiatrische Prax, 31(Suppl. 2), S210–S214. Retrieved from
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&
db=PubMed&dopt=Citation&list_uids=15586312
Faraone, S. V., Asherson, P., Banaschewski, T., Biederman,
J.,
(2015).
Buitelaar, J. K., Ramos-Quiroga, J. A., . . . Franke, B.
Attention-deficit/hyperactivity disorder. Nature Reviews Disease
Primers, 1, 15020. https://doi.org/10.1038/nrdp.2015.20
Filippi, M., Valsasina, P., Misci, P., Falini, A., Comi, G., & Rocca,
M. A. (2013). The organization of intrinsic brain activity dif-
fers between genders: A resting-state fMRI study in a large co-
hort of young healthy subjects. Human Brain Mapping, 34(6),
1330–1343. https://doi.org/10.1002/hbm.21514
Forster, S., & Lavie, N.
(2016). Establishing the attention-
distractibility trait. Psychological Science, 27(2), 203–212. https://
doi.org/10.1177/0956797615617761
Fox, M. D., Snyder, A. Z., Vincent, J. L., Corbetta, M., Van Essen,
D. C., & Raichle, M. E. (2005). The human brain is intrinsically
organized into dynamic, anticorrelated functional networks.
Proceedings of the National Academy of Sciences, 102(27),
9673–9678. https://doi.org/10.1073/pnas.0504136102
works. Journal of Neurophysiology, 101(6), 3270–3283. https://
doi.org/10.1152/jn.90777.2008
Fransson, P.
(2005). Spontaneous low-frequency BOLD signal
fluctuations: An fMRI investigation of the resting-state default
mode of brain function hypothesis. Human Brain Mapping,
26(1), 15–29. Retrieved from http://www.ncbi.nlm.nih.gov/entrez/
query.fcgi?cmd=Retrieve&db=PubMed&dopt=Citation&list_
uids=15852468
Fransson, P. (2006). How default is the default mode of brain func-
tion? Further evidence from intrinsic BOLD signal fluctuations.
Neuropsychologia, 44(14), 2836–2845. https://doi.org/10.1016/
j.neuropsychologia.2006.06.017
Friedman, J., Hastie, T., & Tibshirani, R. (2008). Sparse inverse co-
variance estimation with the graphical lasso. Biostatistics, 9(3),
432–441. https://doi.org/10.1093/biostatistics/kxm045
Friedman-Hill, S. R., Wagman, M. R., Gex, S. E., Pine, D. S.,
(2010). What does dis-
Leibenluft, E., & Ungerleider, L. G.
tractibility in ADHD reveal about mechanisms for top-down
attentional control? Cognition, 115(1), 93–103. https://doi.org/
10.1016/j.cognition.2009.11.013
Gates, K. M., Molenaar, P. C. M., Iyer, S. P., Nigg, J. T., & Fair,
(2014). Organizing heterogeneous samples using com-
D. A.
munity detection of GIMME-derived resting state functional net-
works. PloS One, 9(3), e91322. https://doi.org/10.1371/journal.
pone.0091322
Gordon, E. M., Laumann, T. O., Adeyemo, B., Huckins, J. F., Kelley,
W. M., & Petersen, S. E. (2014). Generation and evaluation
of a cortical area parcellation from resting-state correlations.
Cerebral Cortex, 26(1), 288–303. https://doi.org/10.1093/cercor/
bhu239
Halperin,
J. M., Sharma, V., Greenblatt, E., & Schwartz, S. T.
(1991). Assessment of the continuous performance test: Relia-
bility and validity in a nonreferred sample. Psychological Assess-
ment, 3, 603–608.
Hoekzema, E., Carmona, S., Ramos-Quiroga,
J. A., Richarte
Fernández, V., Bosch, R., Soliva, J. C., . . . Vilarroya, O. (2014).
An independent components and functional connectivity analy-
sis of resting state fMRI data points to neural network dysregu-
lation in adult ADHD. Human Brain Mapping, 35(4), 1261–72.
https://doi.org/10.1002/hbm.22250
Huang-Pollock, C. L., Karalunas, S. L., Tam, H., & Moore, A. N.
(2012). Evaluating vigilance deficits in ADHD: A meta-analysis
Journal of Abnormal Psychology, 121(2),
of CPT performance.
360–371. https://doi.org/10.1037/a0027205
Huang-Pollock, C. L., Nigg, J. T., & Carr, T. H.
(2005). Deficient
attention is hard to find: Applying the perceptual load model
of selective attention to attention deficit hyperactivity disorder
subtypes. Journal of Child Psychology and Psychiatry and Allied
Disciplines, 46(11), 1211–1218. https://doi.org/10.1111/j.1469-
7610.2005.00410.x
Hutchison, R. M., Hashemi, N., Gati,
J. S., Menon, R. S., &
Everling, S.
(2015). Electrophysiological signatures of sponta-
neous BOLD fluctuations in macaque prefrontal cortex. Neuro-
Image, 113, 257–267. https://doi.org/10.1016/j.neuroimage.2015.
03.062
Fox, M. D., Zhang, D., Snyder, A. Z., & Raichle, M. E. (2009). The
global signal and observed anticorrelated resting state brain net-
Hutchison, R. M., Womelsdorf, T., Gati, J. S., Leung, L. S., Menon,
R. S., & Everling, S. (2012). Resting-state connectivity identifies
Network Neuroscience
215
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
0
2
0
2
2
0
0
1
0
9
2
1
5
7
n
e
n
_
a
_
0
0
0
3
4
p
d
/
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
ADHD and attentional control: Impaired segregation of brain networks
distinct
functional networks in macaque cingulate cortex.
Cerebral Cortex, 22(6), 1294–1308. https://doi.org/10.1093/
cercor/bhr181
Karalunas, S. L., Fair, D., Musser, E. D., Aykes, K., Iyer, S. P.,
& Nigg, J. T.
(2014). Subtyping attention-deficit/hyperactivity
disorder using temperament dimensions: Toward biologically
JAMA Psychiatry, 71(9), 1015–1024.
based nosologic criteria.
https://doi.org/10.1001/jamapsychiatry.2014.763
Keller, C., Bickel, S., Honey, C., Groppe, D., Entz, L., Craddock,
R. C., . . . Mehta, A. D.
(2013). Neurophysiological investiga-
tion of spontaneous correlated and anticorrelated fluctuations of
the BOLD signal. Journal of Neuroscience, 33(15), 6333–6342.
https://doi.org/10.1523/JNEUROSCI.4837-12.2013
Keller, J., Hedden, T., Thompson, T., Anteraper, S., Gabrieli, J.,
& Whitfield-Gabrieli, S.
(2015). Resting-state anticorrelations
between medial and lateral prefrontal cortex: Association with
working memory, aging, and individual differences. Cortex, 64,
271–280. https://doi.org/10.1016/j.cortex.2014.12.001
Kelly, A. M., Di Martino, A., Uddin, L. Q., Shehzad, Z., Gee,
D. G., Reiss, P. T., . . . Milham, M. P. (2008). Development of
anterior cingulate functional connectivity from late childhood
to early adulthood. Cerebral Cortex. Retrieved from http://www.
ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed
&dopt=Citation&list_uids=18653667
Kessler, D., Angstadt, M., & Sripada, C.
(2016). Growth charting
of brain connectivity networks and the identification of attention
impairment in youth. JAMA Psychiatry, 73(5), 481. https://doi.
org/10.1001/jamapsychiatry.2016.0088
Konrad, K., & Eickhoff, S. B. (2010). Is the ADHD brain wired dif-
ferently? A review on structural and functional connectivity in
attention deficit hyperactivity disorder. Human Brain Mapping,
31(6), 904–916. https://doi.org/10.1002/hbm.21058
Krain, A. L., & Castellanos, F. X. (2006). Brain development and
ADHD. Clinical Psychology Review, 26(4), 433–444. Retrieved
from http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=
Retrieve&db=PubMed&dopt=Citation&list_uids=16480802
Lenartowicz, A., Delorme, A., Walshaw, P. D., Cho, A. L.,
Bilder, R. M., McGough, J. J., . . . Loo, S. K.
(2014). Electro-
encephalography correlates of spatial working memory deficits
in attention-deficit/hyperactivity disorder: Vigilance, encoding,
and maintenance. Journal of Neuroscience, 34(4), 1171–1182.
https://doi.org/10.1523/JNEUROSCI.1765-13.2014
Luna, B., Marek, S., Larsen, B., Tervo-Clemmens, B., & Chahal, R.
(2015). An integrative model of the maturation of cognitive con-
trol. Annual Review of Neuroscience, 38(1), 151–170. https://doi.
org/10.1146/annurev-neuro-071714-034054
Marek, S., Hwang, K., Foran, W., Hallquist, M. N., Luna, B., &
Wheeler, M.
(2015). The contribution of network organization
and integration to the development of cognitive control. PLoS
Biology, 13(12), e1002328. https://doi.org/10.1371/journal.pbio.
1002328
Miller, M., & Hinshaw, S. P. (2010). Does childhood executive func-
tion predict adolescent functional outcomes in girls with ADHD?
Journal of Abnormal Child Psychology, 38(3), 315–326. https://
doi.org/10.1007/s10802-009-9369-2
Mills, B. D., Miranda-Dominguez, O., Mills, K., Earl, E., Cordova,
M., Painter, J., . . . Fair, D. A. (2018). Supplemental material for
“ADHD and attentional control:
Impaired segregation of task
positive and task negative brain networks.” Network Neuro-
science, 2(2), 200–217. https://doi.org/10.1162/netn_a_00034
Popa, D., Popescu, A. T., & Pare, D.
(2009). Contrasting activity
profile of two distributed cortical networks as a function of at-
tentional demands. Journal of Neuroscience, 29(4), 1191–1201.
https://doi.org/10.1523/JNEUROSCI.4867-08.2009
Posner, M. I., & DiGirolamo, G. J. (1999). Attention in cogni-
tive neuroscience: An overview. In M. S. Gazzaniga (Ed.), The
new cognitive neurosciences (2nd ed.). Cambridge, MA: MIT
Press.
Posner, M. I., & Petersen, S. E. (1990). The attention system of the
human brain. Annual Review of Neuroscience, 13, 25–42.
Power, J., Barnes, K., Snyder, A., Schlaggar, B., & Petersen, S. (2012).
Spurious but systematic correlations in functional connectivity
MRI networks arise from subject motion. NeuroImage, 59(3),
2142–2154. https://doi.org/10.1016/j.neuroimage.2011.10.018
J. D., Fair, D., Schlaggar, B. L., & Petersen, S. E.
(2010). The development of human functional brain networks.
Neuron, 67(5), 735–48. https://doi.org/10.1016/j.neuron.2010.
08.017
Power,
Riccio, C., & Reynolds, C.
(2006). Continuous performance tests
are sensitive to ADHD in adults but lack specificity. Annals of
the New York Academy of Sciences, 931(1), 113–139. https://
doi.org/10.1111/j.1749-6632.2001.tb05776.x
Sato, J. R., Salum, G. A., Gadelha, A., Crossley, N., Vieira, G.,
Manfro, G. G., . . . Bressan, R. A.
(2016). Default mode net-
work maturation and psychopathology in children and adoles-
cents. Journal of Child Psychology and Psychiatry, 57(1), 55–64.
https://doi.org/10.1111/jcpp.12444
Scheinost, D., Finn, E. S., Tokoglu, F., Shen, X., Papademetris, X.,
Hampson, M., & Constable, R. T.
(2015). Sex differences in
normal age trajectories of functional brain networks. Human
Brain Mapping, 36(4), 1524–1535. https://doi.org/10.1002/hbm.
22720
Sergeant,
J. J. (1999).
J., & van der Meere,
Information processing and energetic factors
in attention
deficit/hyperactivity disorder. In H. C. Quay & A. E. Hogan
(Eds.), Handbook of disruptive behavior disorders. New York:
Kluwer/Plenum Publishers.
J. A., Oosterlaan,
Shaw, P., Eckstrand, K., Sharp, W., Blumenthal, J., Lerch, J. P.,
Greenstein, D., . . . Rapoport, J. L. (2007). Attention-deficit/
hyperactivity disorder is characterized by a delay in corti-
cal maturation. Proceedings of the National Academy of Sci-
ences, 104(49), 19649–19654. https://doi.org/10.1073/pnas.
0707741104
Siegel, J. S., Mitra, A., Laumann, T. O., Seitzman, B. A., Raichle, M.,
Corbetta, M., & Snyder, A. Z.
(2016). Data quality influences
observed links between functional connectivity and behavior.
Cerebral Cortex. https://doi.org/10.1093/cercor/bhw253
Smith, S. M., Beckmann, C. F., Andersson, J., Auerbach, E. J.,
Bijsterbosch, J., Douaud, G., . . . Glasser, M. F. (2013). Resting-
state fMRI in the human connectome project. NeuroImage, 80,
144–168. https://doi.org/10.1016/j.neuroimage.2013.05.039
Sonuga-Barke, E. J. S., & Castellanos, F. X. (2007). Spontaneous
attentional fluctuations in impaired states and pathological
conditions: A neurobiological hypothesis. Neuroscience and
Network Neuroscience
216
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
0
2
0
2
2
0
0
1
0
9
2
1
5
7
n
e
n
_
a
_
0
0
0
3
4
p
d
/
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
ADHD and attentional control: Impaired segregation of brain networks
Biobehavioral Reviews, 31(7), 977–986. https://doi.org/10.1016/
j.neubiorev.2007.02.005
Spreng, R. N., Stevens, W. D., Viviano, J. D., & Schacter, D. L.
(2016). Attenuated anticorrelation between the default and dor-
sal attention networks with aging: Evidence from task and rest.
Neurobiology of Aging, 45, 149–160. https://doi.org/10.1016/j.
neurobiolaging.2016.05.020
Sripada, C., Kessler, D., & Angstadt, M. (2014). Lag in maturation
the brain’s intrinsic functional architecture in attention-
of
deficit/hyperactivity disorder. Proceedings of the National Acad-
emy of Sciences, 111(39), 14259–14264. https://doi.org/10.1073/
pnas.1407787111
Sripada, C., Kessler, D., Fang, Y., Welsh, R. C., Prem Kumar, K.,
& Angstadt, M.
(2014). Disrupted network architecture of the
resting brain in attention-deficit/hyperactivity disorder. Human
Brain Mapping, 35(9), 4693–4705. https://doi.org/10.1002/hbm.
22504
Stanislaw, H., & Todorov, N.
(1999). Calculation of signal
detection theory measures. Behavior Research Methods, Instru-
ments, and Computers, 31(1), 137–149. https://doi.org/10.3758/
BF03207704
Sun, L., Cao, Q., Long, X., Sui, M., Cao, X., Zhu, C., . . . Wang,
Y.
(2012). Abnormal functional connectivity between the an-
terior cingulate and the default mode network in drug-naïve
boys with attention deficit hyperactivity disorder. Psychiatry Re-
search, 201(2), 120–127. https://doi.org/10.1016/j.pscychresns.
2011.07.001
Tomasi, D., & Volkow, N. D. (2012). Gender differences in brain
functional connectivity density. Human Brain Mapping, 33(4),
849–860. https://doi.org/10.1002/hbm.21252
Tucha, L., Tucha, O., Walitza, S., Sontag, T. A., Laufkätter, R.,
Linder, M., & Lange, K. W. (2009). Vigilance and sustained
Journal of
attention in children and adults with ADHD.
Attention Disorders, 12(5), 410–421. https://doi.org/10.1177/
1087054708315065
Van Essen, D. C., Ugurbil, K., Auerbach, E., Barch, D., Behrens,
T. E., Bucholz, R., . . . Yacoub, E. (2012). The human connectome
project: A data acquisition perspective. NeuroImage, 62(4),
2222–2231. https://doi.org/10.1016/j.neuroimage.2012.02.018
(2004).
The neural mechanisms for minimizing cross-modal distraction.
Journal of Neuroscience, 24(48), 10941–10949. Retrieved from
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&
db=PubMed&dopt=Citation&list_uids=15574744
Weissman, D. H., Warner, L. M., & Woldorff, M. G.
(2005). Validity of
Willcutt, E. G., Doyle, A. E., Nigg, J. T., Faraone, S., & Pennington,
B. F.
the executive function theory of
attention-deficit/hyperactivity disorder: A meta-analytic review.
Biological Psychiatry, 57(11), 1336–1346. https://doi.org/10.1016/
j.biopsych.2005.02.006
Yan, C. G., Craddock, R. C., Zuo, X. N., Zang, Y. F., & Milham, M. P.
(2013). Standardizing the intrinsic brain: Towards robust mea-
surement of inter-individual variation in 1000 functional con-
nectomes. NeuroImage, 80, 246–262. https://doi.org/10.1016/
j.neuroimage.2013.04.081
Yerys, B. E., Jankowski, K. F., Shook, D., Rosenberger, L. R., Barnes,
K. A., Berl, M. M., . . . Gaillard, W. D. (2009). The fMRI success
rate of children and adolescents: Typical development, epilepsy,
attention deficit/hyperactivity disorder, and autism spectrum dis-
orders. Human Brain Mapping, 30(10), 3426–3435. https://doi.
org/10.1002/hbm.20767
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
0
2
0
2
2
0
0
1
0
9
2
1
5
7
n
e
n
_
a
_
0
0
0
3
4
p
d
.
/
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Network Neuroscience
217