Flexible Coding of Task Rules in Frontoparietal Cortex:
An Adaptive System for Flexible Cognitive Control
Alexandra Woolgar, Soheil Afshar, Mark A. Williams, and Anina N. Rich
Abstract
■ How do our brains achieve the cognitive control that is re-
quired for flexible behavior? Several models of cognitive control
propose a role for frontoparietal cortex in the structure and
representation of task sets or rules. For behavior to be flexible,
however, the system must also rapidly reorganize as mental fo-
cus changes. Here we used multivoxel pattern analysis of fMRI
data to demonstrate adaptive reorganization of frontoparietal
activity patterns following a change in the complexity of the
task rules. When task rules were relatively simple, frontoparie-
tal cortex did not hold detectable information about these
rules. In contrast, when the rules were more complex, fronto-
parietal cortex showed clear and decodable rule discrimination.
Our data demonstrate that frontoparietal activity adjusts to task
complexity, with better discrimination of rules that are behav-
iorally more confusable. The change in coding was specific to
the rule element of the task and was not mirrored in more spe-
cialized cortex (early visual cortex) where coding was indepen-
dent of difficulty. In line with an adaptive view of frontoparietal
function, the data suggest a system that rapidly reconfigures in
accordance with the difficulty of a behavioral task. This system
may provide a neural basis for the flexible control of human
behavior. ■
INTRODUCTION
Human behavior is remarkably diverse and flexible. How
does the brain achieve the flexible cognitive control that
is required? Several theories implicate pFC, damage to
which results in characteristically inflexible and persever-
ative behavior (Luria, 1966). In particular, pFC is thought
to have a key role in structuring and representing task sets
or rules (e.g., Duncan, 2013; Miller & Cohen, 2001). Sev-
eral theories of prefrontal organization predict the in-
volvement of different prefrontal regions, depending on
the characteristics of the rule being used (e.g., O’Reilly,
2010; Badre & D’Esposito, 2009; Botvinick, 2008; Christoff
& Keramatian, 2007; Koechlin & Summerfield, 2007;
Bunge & Zelazo, 2006; Koechlin & Jubault, 2006). For ex-
ample, one prominent suggestion is that the pFC follows
a rostrocaudal gradient with different regions recruited
according to the control demands of the task (Badre &
D’Esposito, 2007; Koechlin & Summerfield, 2007). In
contrast, the adaptive coding hypothesis (Duncan,
2001) proposes that single prefrontal neurons dynam-
ically adjust their responses to code the task distinctions
that are currently relevant for behavior. The suggestion is
that a specific network of prefrontal and parietal regions,
active in many different tasks, supports flexible behavior
by rapidly reconfiguring to represent the information that
is currently relevant as mental focus changes (Duncan &
Miller, 2013; Duncan, 2001, 2010). Similar to other models
of prefrontal function (Miller & Cohen, 2001; Dehaene,
Macquarie University, Sydney, Australia
Kerszberg, & Changeux, 1998; Norman & Shallice,
1980), cognitive control is then exerted through influence
over more specialized processing areas, for example,
by biasing processing toward task-relevant informa-
tion (Duncan, 2006; Duncan & Miller, 2002; Desimone
& Duncan, 1995).
Recruitment of different brain regions could follow a
posterior–anterior pFC axis along which control signals
of progressively greater temporal abstraction are orga-
nized (Koechlin & Summerfield, 2007; Koechlin, Ody,
& Kouneiher, 2003). According to this proposal, rules
governing habitual sensorimotor associations are pro-
cessed in the premotor cortex, whereas contextual rules
(e.g., “apply Rule 1 for red and Rule 2 for blue”) addition-
ally recruit posterior lateral pFC. Engagement of the
more anterior region of the inferior frontal sulcus (IFS)
requires episodic control (i.e., control by past events,
for example, a cue given at the start of a block of trials).
A related organizational scheme was proposed by Badre
and D’Esposito (2007, 2009) and Badre (2008), who sug-
gested a caudal-rostral gradient mapping onto increasing
layers of conditional dependency. For example, the rule
“press 1 for stimulus A, and 2 for stimulus B” has one layer,
whereas the rule “if the cue is red, press 1 for stimulus A,
and 2 for stimulus B, but if the cue is blue then press 2
for stimulus A, and 1 for stimulus B” has two layers, one
governing the mapping between stimulus and response
and a second governing which mapping to use. Process-
ing of a single layer rule is proposed to be confined to the
caudal part of pFC (dorsal premotor cortex), processing
© 2015 Massachusetts Institute of Technology. Published under a
Creative Commons Attribution 3.0 Unported (CC BY 3.0) license
Journal of Cognitive Neuroscience 27:10, pp. 1895–1911
doi:10.1162/jocn_a_00827
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
7
/
1
2
0
7
/
1
1
8
0
9
/
5
1
1
8
9
9
4
5
9
/
6
1
2
6
7
8
o
3
c
2
n
4
_
3
a
/
_
j
0
o
0
c
8
n
2
7
_
a
p
_
d
0
0
b
8
y
2
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
/
t
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
of two layers additionally recruits anterior dorsal premo-
tor cortex and so on, with additional layers recruiting pro-
gressively more rostral regions.
These theories account for our ability to flexibly solve a
wide range of different tasks by positing the involvement
of different brain regions in different tasks, while the
response of each brain region is specialized for a partic-
ular information type. In contrast, the adaptive coding
hypothesis (Duncan, 2001) suggests that individual neu-
rons, capable of supporting behavior in a wide range of
different tasks, adjust their response according to task
demands. Brain regions showing adaptive coding would
therefore be expected to be involved in a wide range of
tasks, to code a wide range of task features as necessary
for behavior, and to adjust their coding profiles according
to the demands of the current task (Duncan, 2010).
Adaptive coding can be seen in the firing rates of single
units recorded in nonhuman primates. The activity of sin-
gle cells in lateral frontal cortex can discriminate a wide
range of task features including task rules, cues, stimuli,
responses, rewards, and combinations of these features
(for a review, see Duncan, 2001). Moreover, the activity
profile of many of these cells adjusts to make different
task-relevant distinctions during single trials as attentional
focus is established (Kadohisa et al., 2013), as dictated by
the current phase of the task (Stokes et al., 2013; Sigala,
Kusunoki, Nimmo-Smith, Gaffan, & Duncan, 2008; Rao,
Rainer, & Miller, 1997) and after retraining (Cromer,
Roy, & Miller, 2010; Freedman, Riesenhuber, Poggio, &
Miller, 2001). A similar generality of response is seen for cer-
tain cells in lateral parietal cortex, which discriminate a range
of task features including aspects of cues, stimuli, rules, and
responses (e.g., Ibos, Duhamel, & Ben Hamed, 2013; Gail
& Andersen, 2006; Stoet & Snyder, 2004; Andersen, Essick,
& Siegel, 1985) and adjust their responses after retraining
(Freedman & Assad, 2006).
In the human brain, candidate regions for adaptive
coding are a set of regions in frontal and parietal cortices
that are active for a wide range of task demands (Duncan,
2001, 2006). These encompass the IFS, anterior insula/
frontal operculum (AI/FO), dorsal anterior cingulate/
pre-supplementary motor area (ACC/pre-SMA), and intra-
parietal sulcus (IPS). In functional imaging, a characteris-
tic pattern of activity in these regions is found for a range
of different cognitive demands, including response conflict,
task switching, perceptual difficulty, working memory, ep-
isodic memory, and semantic memory (e.g., Niendam
et al., 2012; Stiers, Mennes, & Sunaert, 2010; Dosenbach
et al., 2006; Naghavi & Nyberg, 2005; Owen, McMillan,
Laird, & Bullmore, 2005; Nyberg et al., 2003; Duncan &
Owen, 2000). The same result is seen across tasks at the
level of single subjects (Fedorenko, Duncan, & Kanwisher,
2013). Evidence from neuropsychology suggests that these
regions may be critical for fluid intelligence, the aspect of
general intelligence that predicts performance on a wide
range of tasks, with the extent of behavioral deficit linearly
predicted by the extent to which these regions are dam-
aged ( Woolgar et al., 2010) and compensatory activity
seen throughout the system after frontal lobe damage
(Woolgar, Bor, & Duncan, 2013).
Given the generality of their response, we refer to
these regions as “multiple-demand” or MD regions (Duncan,
2006, 2010). They have elsewhere been referred to as the
“cognitive control network” (Cole & Schneider, 2007),
“task positive” network (Fox et al., 2005), “task activation
ensemble” (Seeley et al., 2007), or “frontoparietal control
system” ( Vincent, Kahn, Snyder, Raichle, & Buckner,
2008). They are suggested to form a possible neural basis
for a “global workspace” (Dehaene & Changeux, 2011;
Dehaene & Naccache, 2001) and are widely implicated
in neural models of executive function and cognitive con-
trol (e.g., Cole & Schneider, 2007; Corbetta & Shulman,
2002; Miller & Cohen, 2001).
The adaptive coding hypothesis predicts that the MD
regions dynamically adjust the information they discrim-
inate when task demands change. Information coding
has traditionally been difficult to examine in humans,
but multivoxel pattern analysis (MVPA) of fMRI data pre-
sents an opportunity to test this prediction. MVPA tests
for consistency in the fine-grained patterns of BOLD re-
sponse: Information coding is inferred where patterns can
consistently discriminate between different task events
(Haynes & Rees, 2005). In line with an adaptive view of
MD function, multivoxel patterns of activity in these re-
gions discriminate a range of task-relevant information in-
cluding stimuli, task rules, and participant responses (e.g.,
Harel, Kravitz, & Baker, 2014; Woolgar, Thompson, Bor, &
Duncan, 2011; Stiers et al., 2010; Bode & Haynes, 2009; Li,
Ostwald, Giese, & Kourtzi, 2007).
In addition to coding a range of qualitatively different
task features, regions showing adaptive coding should
adjust the strength of coding of single task features as
task demands change ( Woolgar, Hampshire, Thompson,
& Duncan, 2011). Moreover, this adjustment should be in
a direction that is adaptive for behavior. This presents a
counterintuitive prediction: that information that is more
difficult to discriminate on a behavioral level will be bet-
ter distinguished at the level of multivoxel codes. Consis-
tent with this, we recently demonstrated that MD regions
code visual stimuli more strongly under conditions of
high, compared to low, perceptual difficulty ( Woolgar,
Williams, & Rich, 2015; Woolgar, Hampshire, et al.,
2011), suggesting that MD representation of perceptual
information adapts to cope with increased perceptual
difficulty.
Beyond perceptual characteristics, a mechanism for
cognitive control requires representation of task rule in-
formation, governing the appropriate behavioral response
for different stimuli. The MD regions are already strongly
implicated in the representation of task rules in a variety
of experimental contexts ( Waskom, Kumaran, Gordon,
Rissman, & Wagner, 2014; Soon, Namburi, & Chee, 2013;
Zhang, Kriegeskorte, Carlin, & Rowe, 2013; Momennejad,
2012; Reverberi, Gorgen, & Haynes, 2012a, 2012b; Cole,
1896
Journal of Cognitive Neuroscience
Volume 27, Number 10
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
7
/
1
2
0
7
/
1
1
8
0
9
/
5
1
1
8
9
9
4
5
9
/
6
1
2
6
7
8
o
3
c
2
n
4
_
3
a
/
_
j
0
o
0
c
8
n
2
7
_
a
p
_
d
0
0
b
8
y
2
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
/
t
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Etzel, Zacks, Schneider, & Braver, 2011; Woolgar,
Hampshire, et al., 2011; Woolgar, Thompson, et al., 2011;
Bode & Haynes, 2009; Haynes et al., 2007), including, for
example, rules pertaining to perceptual and conceptual as-
pects of visual stimuli (Harel et al., 2014). However, to our
knowledge, the question of whether MD rule representa-
tion is adaptive, namely, stronger when the rule aspect of
the task is more challenging, has not been examined.
In the current study, we focused on the predictions of
the adaptive coding hypothesis. We asked whether MD
regions would change their coding of task rules when
rule complexity changed. To address this, we compared
multivoxel discrimination of two relatively simple stimulus–
response mapping rules previously found to be coded in
these regions (Woolgar, Thompson, et al., 2011) with dis-
crimination of two new stimulus–response mapping rules
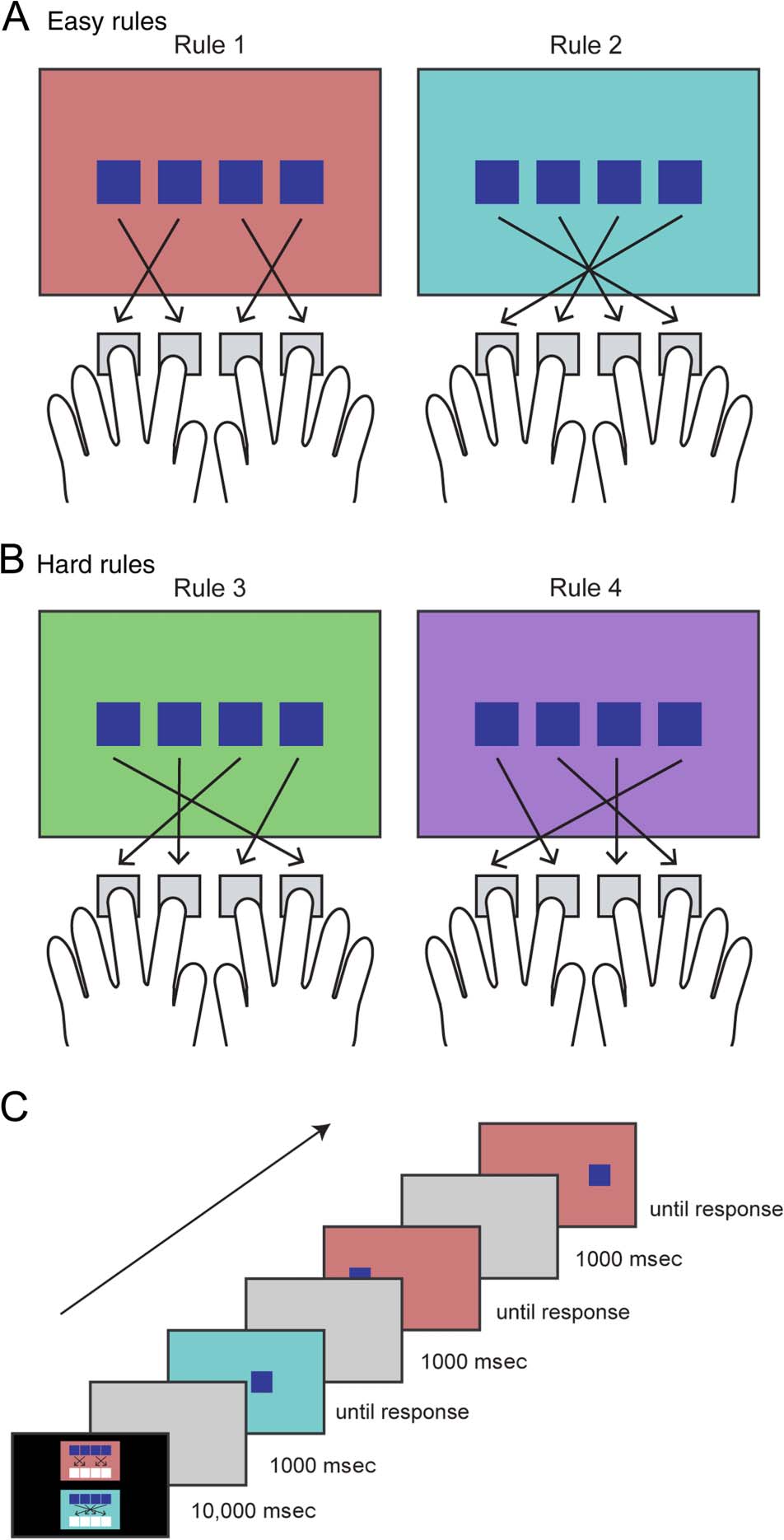
that were more complex and highly confusable (Figure 1).
Each rule defined the mapping between four stimulus po-
sitions and four button press responses, and on each trial
the current rule to use was cued by the background color
of the screen.
The adaptive coding hypothesis (Duncan, 2001) pre-
dicts stronger coding of the more challenging rules rela-
tive to the simple ones as the system responds to the
change in difficulty of the task by augmenting the repre-
sentation of the most challenging task feature.
At the opposite extreme, hierarchical organization ac-
counts predict that, because our rules depend on pro-
cessing information immediately available when the
stimulus is presented (Koechlin & Summerfield, 2007)
and consist of only two layers (Badre & D’Esposito,
2007), processing should be confined to the anterior dor-
sal premotor cortex and not recruit, for example, the IFS
MD region. However, given previous observations of ac-
tivation for (Crittenden & Duncan, 2014) and discrimina-
tion of ( Woolgar, Hampshire, et al., 2011; Woolgar,
Thompson, et al., 2011) rules of this type throughout
the MD system, such an outcome seems unlikely.
It is also possible that the MD system always codes
rules equally, regardless of complexity. This might be
predicted given a specific role for the MD regions in
maintaining task rules (Miller & Cohen, 2001), commen-
surate with previous empirical studies that found the rep-
resentation of rules in frontoparietal cortex to be stronger
than representation of other task features (e.g., Riggall &
Postle, 2012; Woolgar, Thompson, et al., 2011; Sigala
et al., 2008). Because even our “easy” rules were cogni-
tively challenging and have previously been found to be
represented in the MD system ( Woolgar, Thompson,
et al., 2011), with the strength of representation robust
to changes in perceptual difficulty ( Woolgar, Hampshire,
et al., 2011), it was possible that both these and the new
set of hard rules would be represented equally. Further-
more, because our easy and hard rules do not differ on the
number of layers of conditional dependency (Badre &
D’Esposito, 2009) nor require integration of information
over different timescales (Koechlin & Summerfield,
Figure 1. Participants learned four stimulus–response mappings
governing which of four response keys they should press for each of the
four possible stimulus positions. Rules 1 and 2 were simple (“easy”, A),
whereas Rules 3 and 4 were more complex (“hard”, B). The easy rules
comprised one consistent position–response transformations and were
internally symmetrical, whereas the hard two rules comprised four
unique position–response transformations and no internal symmetry.
The background color of the screen indicated which rule to use on each
trial (red: Rule 1, blue: Rule 2, green: Rule 3, purple: Rule 4). (C)
Illustration of three trials at the start of a block of easy rules. The correct
responses would be buttons 2, 2, and 3.
2007), the accounts of Badre and D’Esposito (2009) and
Koechlin and Summerfield (2007) would not predict dif-
ferential coding of the hard and easy rules.
Finally, if rule decoding depends on the similarity of
stimulus–response transformations within each rule, we
might see more coding of the easy relative to the hard
rules. For example, Rules 1 and 2 can be thought of as
Woolgar et al.
1897
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
7
/
1
2
0
7
/
1
1
8
0
9
/
5
1
1
8
9
9
4
5
9
/
6
1
2
6
7
8
o
3
c
2
n
4
_
3
a
/
_
j
0
o
0
c
8
n
2
7
_
a
p
_
d
0
0
b
8
y
2
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
f
t
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
containing only one stimulus–response transformation
operation each (Duncan, 1977; Rule 1: alternate finger
within hemisphere, Rule 2: reflection across the midline;
Figure 1), whereas Rules 3 and 4 both require the use of
four distinct stimulus–response transformations (mirror
across the midline, direct mapping, alternate finger of al-
ternate hand, and alternate finger within hemisphere).
Because our rule decoding analysis identifies commonal-
ities in multivoxel patterns for one rule as opposed to an-
other, representation of the rules at this level should
drive better classification for the easy rules where the
transformations are more similar within rule and more
distinct between rules.
To preempt our results, the data revealed greater MD
discrimination of the two complex rules relative to the
simpler rules. That is, multivoxel discrimination in the
MD system was greater for rules that behaviorally were
more difficult to distinguish. In contrast, visual cortex en-
coded the two (visually cued) rules equally. The results
suggest an adaptive frontoparietal system that reconfi-
gures in response to changing task difficulty, providing
a possible mechanism for cognitive control of flexible
behavior.
METHODS
Participants
Twenty-two participants (14 women, mean age =
24.9 years, SD = 4.51) took part in this study. All partici-
pants were right-handed and had normal or corrected-to-
normal vision. For multivariate analyses, three participants
were rejected because of poor behavioral performance
(see Behavioral Thresholding section) leaving 19 partici-
pants (12 women, mean age = 23.9 years, SD = 4.25).
All participants gave written informed consent and were
reimbursed for their time. The study was approved by
the Macquarie University Human Research Ethics
Committee.
Task Design
Participants were scanned while completing a visual
stimulus–response task that we used previously to sepa-
rate multivoxel coding of visual stimuli, task rules, and
button-press responses ( Woolgar, Hampshire, et al.,
2011; Woolgar, Thompson, et al., 2011). On each trial,
the stimulus was a blue square measuring 2° × 2° presented
on a projector and viewed through a head-coil-mounted
mirror in the scanner. It could appear in one of four posi-
tions, arranged in a horizontal row in the center of the
screen, and separated by 3° middle to middle. Participants
responded by pressing one of four response keys using
the index and middle fingers from each hand. Participants
learnt four incompatible stimulus–response mappings
(“rules”) between the four stimulus positions and the four
response keys (Figure 1). Two of the rules were relatively
simple (“easy”), whereas the other two rules were more
complex (“hard”). The easy rules each comprised one
“consistent” position–response transformation (Rule 1: al-
ternate finger within hemisphere, Rule 2: reflection across
the midline; Duncan, 1977, 1978), and were internally sym-
metrical. The two hard rules were “inconsistent” (Duncan,
1977), each comprising four different position–response
transformations (mirror across the midline, direct map-
ping, alternate finger of alternate hand, and alternate finger
within hemisphere) with no internal symmetry, and were
mirror images of one another. On each trial, the current
rule to use was indicated by the background color of the
screen (red: Rule 1, blue: Rule 2, green: Rule 3, purple:
Rule 4; color rule mappings fixed across participants).
Colors were presented at 50% saturation and 70% bright-
ness. The colors for the two easy rules and the two hard
rules were each separated by 180° of hue.
Training
Participants learned and practiced each rule outside the
scanner. Training order was counterbalanced over partic-
ipants so that half the participants learned the hard rules
first whereas the other half learned the easy rules first,
and within this, the order in which the two easy and
two hard rules were learned was also counterbalanced
across participants. For participants that learned the easy
rules first, they first learned (12 trials) and practiced (12 tri-
als) one of the easy rules and then learned (12 trials) and
practiced (12 trials) the other easy rule. During the
12 learning trials, participants were shown each stimulus
together with a graphical depiction of the correct button
to press. For this, trial order was fixed such that the blue
square appeared in each of the horizontal positions in se-
quence from left to right, repeated three times. During
the 12 practice trials, the stimuli were presented in ran-
dom order, and the correct button to press was not
shown. Participants then practiced a mixed block (36 tri-
als) in which the four stimulus positions and two easy
rules were presented in random order, with the back-
ground color on each trial indicating the rule to be ap-
plied. The same procedure was then used for the hard
rules. Participants were instructed to respond as quickly
as possible without making any mistakes. During prac-
tice, participants were given feedback on each trial
(“CORRECT,” for 500 msec; or “INCORRECT! The correct
answer was x” where x indicates the number of the cor-
rect response button, for 1500 msec). If the participant’s
response was incorrect, they were shown the stimulus
from the previous trial again, together with a graphical
depiction of correct button to press, and were required
to press the correct response key. At the end of each
block, the participant’s accuracy and mean RT were dis-
played for 4 sec. If participants scored below 60%
(chance = 25%) on any rule in any practice block, they
were required to repeat that block until their perfor-
mance reached 60%. The same training scheme was used
1898
Journal of Cognitive Neuroscience
Volume 27, Number 10
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
7
/
1
2
0
7
/
1
1
8
0
9
/
5
1
1
8
9
9
4
5
9
/
6
1
2
6
7
8
o
3
c
2
n
4
_
3
a
/
_
j
0
o
0
c
8
n
2
7
_
a
p
_
d
0
0
b
8
y
2
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
.
/
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
for participants that learned the hard rules first. Training
took approximately 20 min.
Acquisition
fMRI scans were acquired using a Siemens (Erlangen,
Germany) 3-T Verio scanner with 32-channel head coil,
at the Macquarie Medical Imaging facility in Macquarie
University Hospital, Sydney, Australia. We used a sequential
descending T2*-weighted EPI acquisition sequence with
the following parameters: repetition time = 2000 msec;
echo time = 30 msec; 34 oblique axial slices of 3.0 mm slice
thickness with 0.7 mm interslice gap; in-plane resolution =
3.0 × 3.0 mm; field of view = 210 mm; flip angle = 78°. We
also acquired T1-weighted MPRAGE structural images for
all participants (resolution 1.0 × 1.0 × 1.0 mm).
Stimuli were presented using Matlab with Psychophysics
Toolbox-3 (Brainard, 1997) and were back-projected onto
a screen viewed through a head-coil mounted mirror in
the scanner. Participants performed alternating 2-min
blocks of hard and easy rules, separated by 22 sec. Partic-
ipants performed two EPI acquisition runs each consisting
of eight blocks of trials and lasting 19 min 12 sec.
Block order was counterbalanced within participants
across runs, and run order was counterbalanced between
participants. At the start of each block, a graphical depic-
tion of the two rules to be used in the current block was
displayed for 10 sec, after which the screen returned to
gray for 1000 msec before the first stimulus was displayed
(Figure 1C). Within each block, the eight stimuli (4 posi-
tions * 2 background colors) were presented in random
order. Stimuli remained visible for 4000 msec or until the
participant responded. There was an intertrial interval of
1000 msec between response and display of the next
stimulus, during which time the screen was gray. Block
length was fixed at 2 min, in which time participants com-
pleted a varying number of trials (mean ± SD total num-
ber of trials over experiment, 857.18 ± 89.74). At the end
of each block, participants were shown a blank screen for
1000 msec, the message “End of Block” for 1000 msec,
followed by a 500-msec blank screen after which feed-
back (% correct and average RT) was displayed for
4000 msec. After this, there was a further 5000 msec
break in which the screen was blank before the next
block began.
Analysis
Univariate and multivariate analyses techniques were
used to examine complementary aspects of the data. Uni-
variate analyses examined overall differences in BOLD
response in hard and easy blocks. MVPA was used to ex-
amine whether multivoxel patterns of activation could re-
liably discriminate task rules, stimulus positions, and
responses in the hard and easy conditions separately.
Of central interest was whether MD coding of rule was
stronger in the hard condition than in the easy condition,
as predicted by the adaptive coding hypothesis (Duncan,
2001). Analyses therefore focused on prefrontal and pari-
etal MD ROIs with an additional ROI in the visual cortex
included for comparison. Exploratory whole brain (uni-
variate) or searchlight (multivariate) analyses were also
included to check the specificity of the ROI effects.
Preprocessing
Preprocessing was carried out using SPM5 ( Wellcome
Department of Imaging Neuroscience, London, UK;
www.fil.ion.ucl.ac.uk). EPI images were spatially re-
aligned to the first image and slice-time-corrected with
the first slice as the reference. The structural image was
coregistered to the mean EPI image and normalized
(simultaneous gray/white matter segment and normalize)
to derive normalization parameters for each participant.
For multivariate analyses, the EPI images were addition-
ally smoothed with a 4-mm FWHM Gaussian kernel. For
univariate analyses, the EPIs were warped into template
space using the derived normalization parameters and
smoothed with an 8-mm kernel. The time course of each
voxel was high-pass filtered with a cut off of 128 sec.
ROIs
MD ROIs were defined as described previously (Woolgar
et al., 2015; Woolgar, Hampshire, et al., 2011; Woolgar,
Thompson, et al., 2011; Cusack, Mitchell, & Duncan,
2010) using data from a review of activity associated with
diverse cognitive demands (Duncan & Owen, 2000).
There were seven MD ROIs (see Figure 3): left and right
IFS (center of mass, +/−38, 26, 24; volume, 17 cm3), left
and right AI/FO (+/−35, 19, 3; 3 cm3), left and right
IPS (+/−35, −58, 41; 7 cm3), and bilateral ACC/pre-
SMA (0, 23, 39; 21 cm3). Left and right visual cortex ROIs,
Brodmann’s area (BA) 17/18 (−13, −81, 3; 16, −79, 3;
54 cm3) were defined from the Brodmann’s template of
MRIcro (Rorden & Brett, 2000). Coordinates are given in
MNI152 space (McConnell Brain Imaging Centre, Montreal
Neurological Institute, Montreal, QC, Canada).
Behavioral Thresholding
For our main neural analyses we excluded any block of
trials where participants scored below 70% correct.1 This
only occurred on hard blocks. To balance the number of
blocks contributing to easy and hard rule discrimination,
we also rejected the corresponding easy block (e.g., if a
participant scored <70% on the third hard block, both
the third hard and the third easy block were omitted
from analysis). Three participants were excluded entirely
as they had <2 hard blocks remaining at this threshold.
Neural analyses were based on an average of 5.95 (SD =
2.22) easy and per
participant.
Woolgar et al.
1899
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
> .067).
To check the specificity of this finding, we repeated the
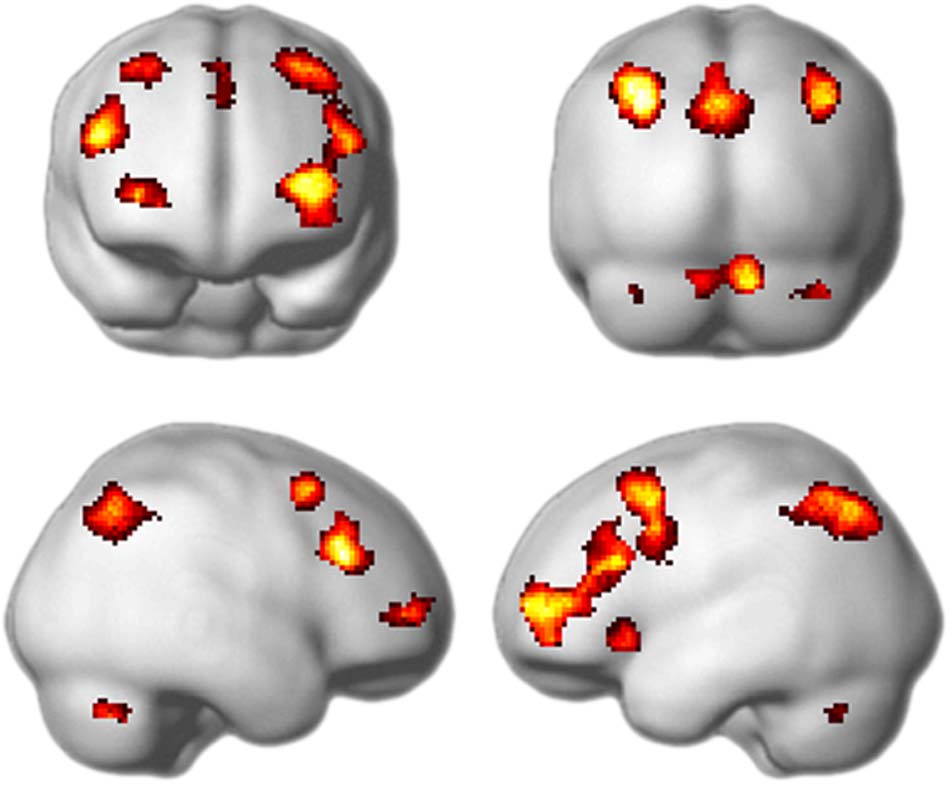
analysis using a mass-univariate approach. As can be seen
in Figure 2 and Table 2, the result was a pattern of frontal
and parietal activation dominated by the characteristic MD
Woolgar et al.
1901
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
7
/
1
2
0
7
/
1
1
8
0
9
/
5
1
1
8
9
9
4
5
9
/
6
1
2
6
7
8
o
3
c
2
n
4
_
3
a
/
_
j
0
o
0
c
8
n
2
7
_
a
p
_
d
0
0
b
8
y
2
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
t
f
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
.004, dz = 0.69) whereas the easy rules could not be dis-
criminated in any MD region (IPS: t(18) = 0.20, p = .16,
dz = 0.05, all other classifications not numerically greater
than 50%). The small ventral prefrontal region around
the AI/FO did not show coding of rule in either con-
dition (classification not numerically greater than 50%).
We performed an additional ANOVA to check for hemi-
spheric differences. This ANOVA had factors Difficulty
(easy, hard), Hemisphere (left, right), and Region (IPS,
IFS, AI/FO; data from bilateral ACC/pre-SMA ROI omit-
ted). There was no Hemisphere × Difficulty interaction
or three-way interaction and no main effect of Hemi-
sphere ( ps > .72). There was a significant Hemisphere ×
Region interaction (F(2, 36) = 4.38, p = .20, η2
p = 0.20),
which indicated that the hemisphere effect was not the
same across regions. To explore this, we performed
post hoc ANOVAs (factors: Difficulty and Hemisphere) in
each region separately. In the IPS region, there was a sig-
nificant main effect of Hemisphere (F(1, 18) = 4.59, p =
.046, η2
p = 0.20), indicating that there was more coding
overall in the left compared with right IPS region, but again
this did not interact with Difficulty (F(1, 18) = 0.022, p =
.88, η2
p < 0.01). There was no main effect of Hemisphere
or Hemisphere × Difficulty interaction in the IFS or AI/FO
ANOVAs ( ps > .35). There was no evidence that the in-
crease in rule coding for hard rules varied between the
two hemispheres.
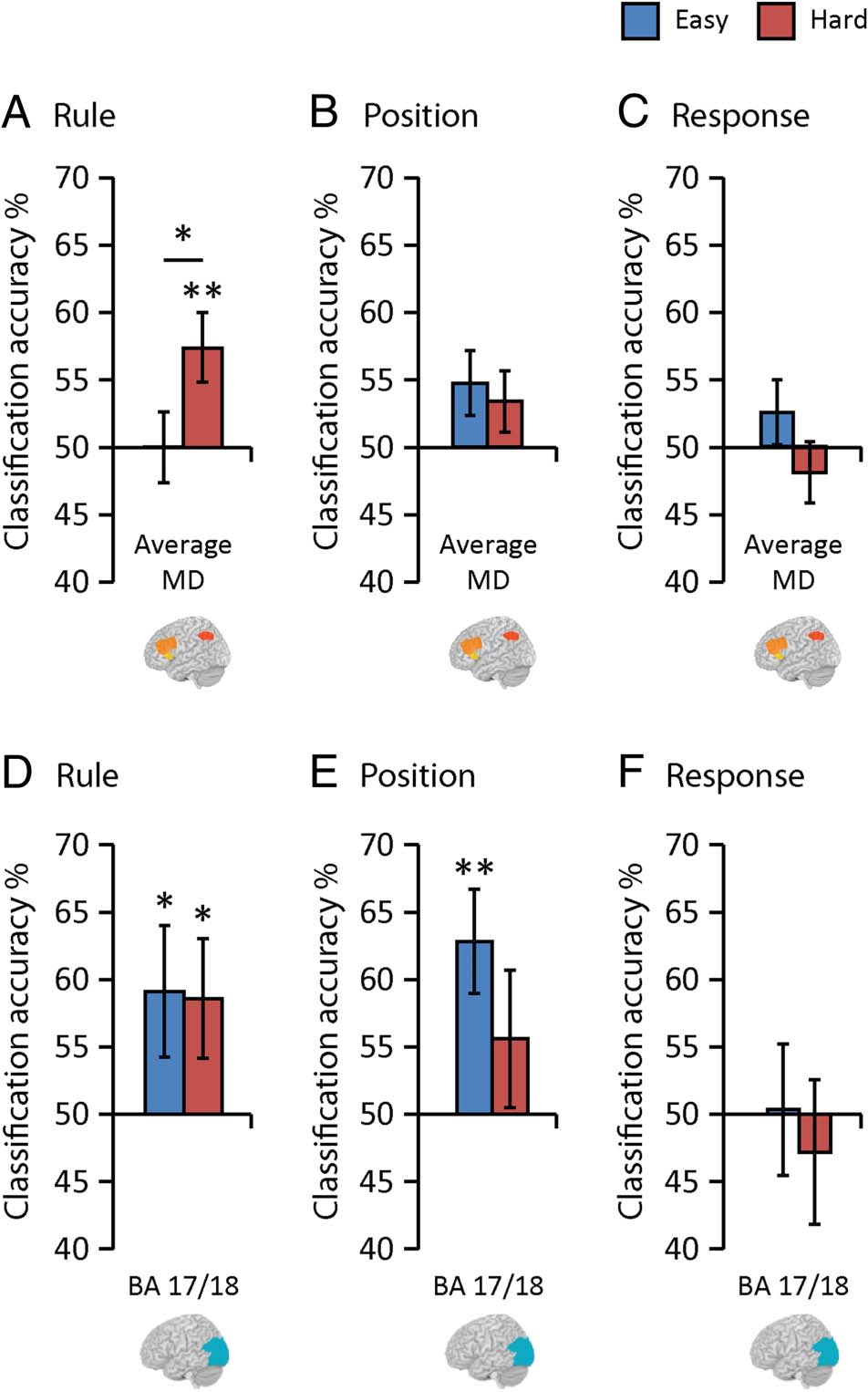
Next, we asked whether the increase in MD coding un-
der conditions of increased rule complexity was specific
to the representation of task rules. One possibility was a
general increase in coding of all task features, as might be
predicted by a general increase in attention or effort in
hard blocks. However, there was no main effect of Diffi-
culty in the ANOVA of position coding (F(1, 18) = 0.12,
p = .73, η2
p = 0.01; Figure 4B) or response coding (F(1,
18) = 1.17, p = .29, η2
p = 0.06; Figure 4C), nor any in-
teraction with Region ( ps > .66). Moreover, in the MD
system considered as a whole, coding of position and re-
sponse information was not significantly different from
chance in either the hard or the easy condition ( ps >
.07). In the context of challenging stimulus–response
mapping rules, multivoxel codes in the MD system were
dominated by rule coding.
Next, we checked the specificity of our result to the
MD system. For this, we examined rule coding in the vi-
sual cortex (BA 17/18), using an ANOVA with factors Dif-
ficulty (easy, hard) and Hemisphere (left, right). This
visual region held information about the (visually cued)
task rules, but, in contrast to the MD system, there was
no difference in rule coding between the easy and hard
conditions (no main effect of Difficulty: F(1, 18) = 0.01,
p = .93, η2
p < 0.01, Figure 4D). Rule discrimination was
above chance (50%) in both the easy (t(18) = 2.24, p =
.019, dz = 0.51) and hard (t(18) = 2.30, p = .017, dz =
0.53) conditions. This ROI also showed above chance
discrimination of position information in the easy condi-
tion (t(18) = 3.34, p = .002, dz = 0.77), but not in the
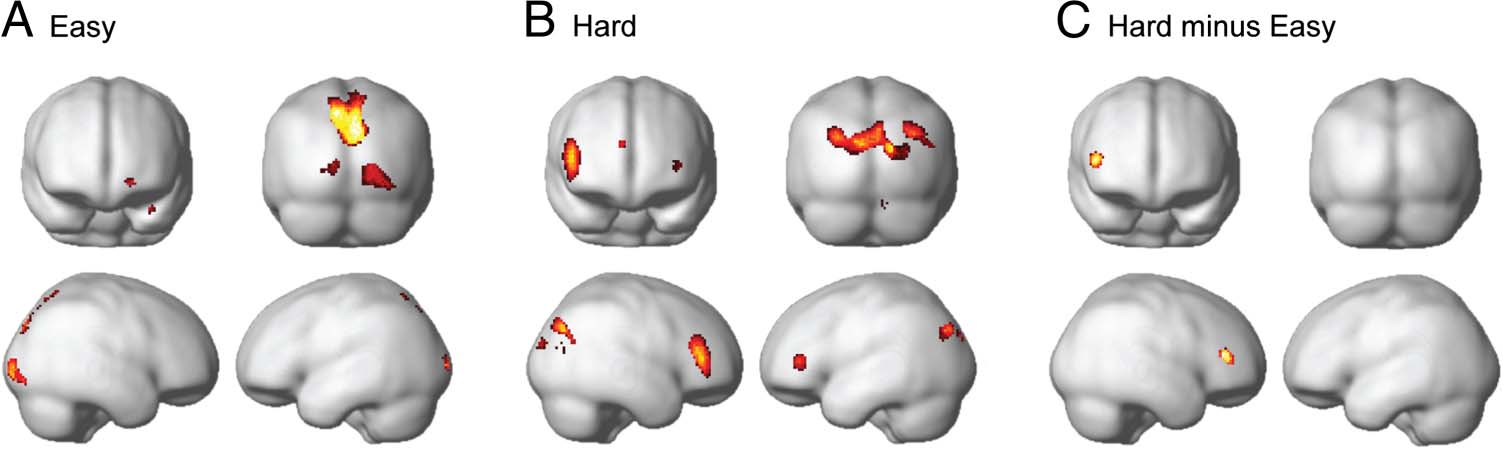
Figure 2. Regions showing increased activation for hard relative to easy
rules (whole-brain univariate analysis). Data are thresholded at p < .05
with FDR correction for multiple comparisons and an extent threshold
of 20 voxels. Coordinates of peak activation are given in Table 2.
pattern with additional restricted regions in the anterior
frontal lobe, FEFs, precuneus, and cerebellum.
Multivoxel Results
We examined multivoxel coding of rules, stimulus posi-
tion, and button press responses when participants per-
formed the stimulus–response mapping task using easy
and hard rules. Of central interest was whether coding
of task-relevant information in the MD system would ad-
just to the increased difficulty of the hard compared to
the easy rules. First, based on the adaptive coding hy-
pothesis (Duncan, 2001) and our previous results with
manipulations of perceptual difficulty ( Woolgar et al.,
2015; Woolgar, Hampshire, et al., 2011), we predicted
that when the two rules were more confusable, the MD
system would adjust to show a counter intuitive increase
in discrimination between the two rules. The decoding
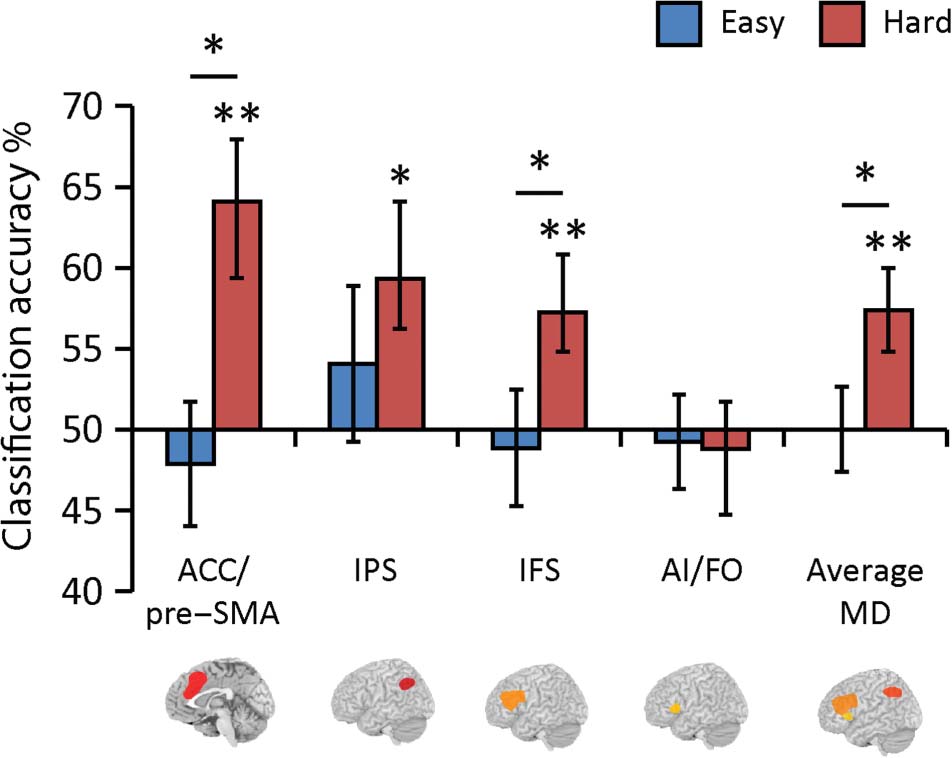
results were in line with this prediction (Figure 3). An
ANOVA with factors Difficulty (high, low) and Region
(ACC/pre-SMA, IPS, IFS, and AI/FO, collapsed across hemi-
sphere where appropriate) revealed a main effect of Diffi-
culty (F(1, 18) = 3.360, p = .042, η2
p = 0.16), indicating
that in the MD system as a whole, there was significantly
more coding of rule information for hard compared with
easy rules. This is striking, because it indicates increased
discrimination between rules that behaviorally are more
confusable. There was no Difficulty × Region interaction
(F(2.039, 36.7) = 2.310, p = .11, η2
p = 0.11).
When we compared rule classification accuracy to
chance (50%) in each region separately, we found that
the hard rules were coded in three of the four MD ROIs
(ACC/pre-SMA: t(18) = 2.79, p = .004, dz = 0.64; IFS:
t(18) = 3.03, p = .004, dz = 0.70; IPS: t(18) = 2.98, p =
1902
Journal of Cognitive Neuroscience
Volume 27, Number 10
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
0
7
/
1
1
8
0
9
/
5
1
1
8
9
9
4
5
9
/
6
1
2
6
7
8
o
3
c
2
n
4
_
3
a
/
_
j
0
o
0
c
8
n
2
7
_
a
p
_
d
0
0
b
8
y
2
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
f
/
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
hard condition (t(18) = 1.01, p = .14, dz = 0.23), al-
though the difference between conditions was again
not significant (main effect of difficulty, F(1, 18) = 1.33,
p = .26, η2
p = 0.07, Figure 4E). There was no coding of
response information in this ROI ( ps > .47, Figure 4F),
and no main effect of, or interaction with, Hemisphere in
any ANOVA (all ps > .33). The adaptive response to the
change in rule difficulty seen in the MD system was not
seen in this more specialized cortex.
Finally, to ensure that we had not overlooked any ad-
ditional brain regions showing rule coding we re-ran the
rule decoding analysis on a more exploratory basis using
a roaming searchlight (Kriegeskorte et al., 2006). Because
this analysis uses data that overlaps with the ROI analy-
ses, converging results would not be surprising but
would indicate that discrimination was possible based
on local patterns of activation on a substantially smaller spa-
tial scale (0.5 cm3 spherical searchlight vs. e.g., 17 cm3 IFS
ROI). The advantage of this analysis is in identifying any ad-
ditional regions that would have been missed by the ROI
approach. To be as inclusive as possible, we used a lenient
threshold of p < .001 uncorrected with extent threshold 20
voxels. The results are shown in Figure 5 and Table 3. For
the easy rules, coding was seen in left and right visual cor-
tex, left middle temporal cortex, left orbital frontal cortex,
and the precuneus (Figure 5A). For the hard rules, coding
was seen in left and right lateral frontal cortex, right ante-
rior cingulate, bilateral occipital cortex, right precuneus,
left caudate nucleus, and right cerebellum (Figure 5B).
Even at our lenient threshold, the only cortical region to
show stronger coding of hard compared with easy rules
was a region in right lateral pFC at the location of the IFS
Table 2. Peak Activation in the Whole-brain Univariate Analyses of Hard Minus Easy Rules
Coordinates
Lobe
Frontal
Cluster
Hemisphere
IFS
Left
AI/FO
Right
Left
Right
ACC/pre-SMA
Bilateral
Anterior frontal
FEFs
IPS
Parietal
Precuneus
Cerebellum
Cerebellum
Left
Right
Left
Right
Left
Right
Bilateral
Bilateral
Left
Right
x
−34
−32
−54
52
−34
30
−8
6
8
−28
38
−24
28
−32
−34
−46
38
38
38
−6
8
−8
−34
30
y
4
22
22
28
20
24
22
22
18
60
58
14
14
−68
−58
−44
−60
−64
−46
−68
−84
−80
−64
−64
z
36
22
28
32
−6
2
44
44
52
8
6
48
50
40
40
44
42
52
34
36
−30
−30
−36
−36
BA
44
45
44
45
47
47
32
32
8
10
10
8
8
7
7
40
39
7
40
7
–
–
–
–
t Score
5.27
4.72
4.09
6.87
5.22
4.55
4.98
4.17
4.04
5.24
4.50
4.07
5.04
6.27
6.06
3.15
5.04
4.14
3.74
5.90
6.44
4.07
3.61
4.23
Results are thresholded at p < .05 with voxelwise FDR correction and an extent threshold of 20 voxels. Large continuous clusters are summarized by
representative peaks.
Woolgar et al.
1903
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
0
7
/
1
1
8
0
9
/
5
1
1
8
9
9
4
5
9
/
6
1
2
6
7
8
o
3
c
2
n
4
_
3
a
/
_
j
0
o
0
c
8
n
2
7
_
a
p
_
d
0
0
b
8
y
2
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
f
t
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
ety of fixed experimental contexts (Harel et al., 2014;
Waskom et al., 2014; Soon et al., 2013; Zhang et al.,
2013; Momennejad, 2012; Reverberi et al., 2012a, 2012b;
Cole et al., 2011; Woolgar, Hampshire, et al., 2011; Woolgar,
Thompson, et al., 2011; Bode & Haynes, 2009; Haynes et al.,
2007). In addition, univariate studies have implicated the
lateral pFC, ACC/pre-SMA, and IPS in the learning, selection,
retrieval, maintenance, and implementation of task rules
(e.g., Crittenden & Duncan, 2014; Wendelken, Munakata,
Baym, Souza, & Bunge, 2012; Dumontheil, Thompson, &
Duncan, 2011; Donohue, Wendelken, & Bunge, 2008;
Crone, Wendelken, Donohue, & Bunge, 2006; Dosenbach
et al., 2006; Passingham, Toni, & Rushworth, 2000; Brass
& von Cramon, 2004; Bunge, 2004), and work in non-
human primates strongly suggests a role for prefrontal
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
0
7
/
1
1
8
0
9
/
5
1
1
8
9
9
4
5
9
/
6
1
2
6
7
8
o
3
c
2
n
4
_
3
a
/
_
j
0
o
0
c
8
n
2
7
_
a
p
_
d
0
0
b
8
y
2
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
f
t
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 4. Multivoxel coding of rule, position, and response information
in the MD system (A, B, and C) and visual cortex (D, E, and F) in easy
(blue bars) and hard (red bars) rule conditions. Error bars indicate SEM.
Significance marking between bars indicate increased coding for hard
relative to easy rules; significance markings for individual bars indicate
coding significantly above chance (50%). *p < .05, **p < .01. Data in A
are repeated from Figure 3 for ease of comparison. The change in
information coding between hard and easy conditions was unique to the
coding of rule and to the MD system.
Figure 3. Rule coding in the MD system in easy and hard rule
conditions. Bars represent classification accuracy for the multivoxel
discrimination of easy rules (Rule 1 vs. Rule 2, blue bars) and hard rules
(Rule 3 vs. Rule 4, red bars). Error bars indicate SEM. Significance
marking between bars indicate increased coding for hard relative to
easy rules; significance markings for individual bars indicate coding
significantly above chance (50%). *p < .05, **p < .01.
ROI (Figure 5C). However, this analysis also revealed a clus-
ter of subcortical voxels in the dorsal lateral region of the
left caudate nucleus where coding was stronger for hard
relative to easy rules. There were no significant voxels for
the inverse comparison of easy minus hard rules. If a more
conservative threshold of p < .05 with FDR correction was
applied, a cluster of voxels in the precuneus for rule cod-
ing in the easy condition were the only voxels to survive
correction.
DISCUSSION
For cognitive control to be flexible to the ever-changing
demands of the world around us, we need a neural sys-
tem that can both represent the rules of our current task
and dynamically adjust as mental focus changes. Here we
tested candidate regions in frontal and parietal cortex,
defined on the basis of being responsive to a wide range
of demands, for flexible representation of task rules in a
stimulus–response mapping task. In line with the adap-
tive coding hypothesis (Duncan, 2001), the MD system
adjusted to yield a strong representation of task rules
when the rules were complex and behaviorally confusable,
but showed little or no representation when the rules
were simpler. This adjustment was only found for the rule
element of the task and was not mirrored in more special-
ized (visual) cortex, where the information was repre-
sented regardless of rule complexity.
Our results suggest an adaptive frontoparietal coding
system that adjusts to the difficulty of the task at hand.
Previous work has demonstrated robust frontopari-
etal multivoxel discrimination between rules in a vari-
1904
Journal of Cognitive Neuroscience
Volume 27, Number 10
Figure 5. Rule coding assessed
with a roaming searchlight.
Whole-brain maps show voxels
where patterns of activation in
the local neighborhood (5 mm
sphere) discriminated between
the two easy rules (A) or the
two hard rules (B). C shows
regions where rule coding was
stronger for hard rules than for
easy rules. Results are
thresholded at p < .001
(uncorrected, equivalent to t =
3.61), with an extent threshold of 20 voxels. There were no regions where coding was significantly stronger for easy compared to hard rules.
The threshold for p < .05 with FDR correction for easy rules would have been t = 4.40, at which threshold the only significant cluster consisted
of 579 voxels around the precuneus. No voxels survived FDR correction for the other comparisons. Coordinates of peak coding are given in
Table 3.
and parietal cells in rule representation (e.g., Stokes et al.,
2013; Buschman, Denovellis, Diogo, Bullock, & Miller,
2012; Sigala et al., 2008; Wallis & Miller, 2003; Wallis,
Anderson, & Miller, 2001; White & Wise, 1999; Hoshi,
Shima, & Tanji, 1998). However, the adaptive coding hy-
pothesis predicts not only that rules can be represented
in this system but also that rule representation will adapt
according to task demands. Here we present new data
showing that rule coding in the MD regions is sensitive to
task difficulty.
Previous evidence for adaptive coding has been drawn
primarily from the nonhuman primate literature. For ex-
ample, Rao et al. (1997) demonstrated that single lateral
prefrontal neurons can discriminate target identity and
location at different times within single trials of a task, ac-
cording to what information is needed in the current task
phase. Over a longer timescale, Freedman and colleagues
showed that monkey frontal (Freedman et al., 2001) and
parietal (Freedman & Assad, 2006) neurons adapt to
code new task-relevant distinctions among the same
stimuli when the animal is trained to use a new decision
boundary (see also Cromer et al., 2010; Roy, Riesenhuber,
Poggio, & Miller, 2010). More recent work testing pat-
terns of activation across multiple units has also shown
Table 3. Coordinates of Peak Decoding in the Searchlight Analyses of Rule Coding
Cluster
Hemisphere
x
y
z
BA
t Score Cluster Extent
Coordinates
Condition
Easy
Lobe
Frontal
Parietal
Hard
Frontal
Lateral frontal cortex (IFS)
Left
47/45 −34
Orbital frontal cortex
Left
11 −18
50 −6
Precuneus
Bilateral
7 −4 −70
52
Occipital
Primary visual cortex
Right
Temporal
Anterior fusiform gyrus
Left
Left
17
18 −100 −4
17 −10 −102
4
36 −34
−2 −28
Anterior cingulate
Parietal
Precuneus
Right
Right
Right
45
32
30
50
8
14 −48
Occipital
Extrastriate cortex
Bilateral
19/18 −28 −78
Subcortical Cerebellum
Caudate nucleus
Right
Right
Left
38
40
52
6
8
24
18
28
38
19
36 −80
12 −68 −24
–
–
−16
8
38
14
26
12
20
4.49
5.55
4.48
4.10
3.91
5.04
5.38
3.85
3.87
4.85
4.32
4.13
4.11
4.35
3.98
36
1503
213
47
35
97
424
30
60
1340
215
138
58
103
98
Hard minus easy
Frontal
Lateral frontal cortex (IFS) Right
Subcortical Caudate nucleus
Left
45
46
–
−16
The table shows peak decoding of rule for easy and hard rules separately and peaks where coding was significantly stronger for hard relative to easy
rules. There were no significant voxels showing stronger coding of easy rules than hard rules. Results are thresholded at p < .001 (uncorrected) with
an extent threshold of 20 voxels.
Woolgar et al.
1905
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
0
7
/
1
1
8
0
9
/
5
1
1
8
9
9
4
5
9
/
6
1
2
6
7
8
o
3
c
2
n
4
_
3
a
/
_
j
0
o
0
c
8
n
2
7
_
a
p
_
d
0
0
b
8
y
2
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
f
/
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
that population tuning profiles in the monkey pFC adjust
to accommodate different task contexts (Stokes et al.,
2013). In human functional imaging, interrogation of sin-
gle neurons is not possible, but a similar result was found
using multivoxel pattern analysis when participants at-
tended to different aspects of moving dot figures (Li
et al., 2007). Moreover, we previously demonstrated that
the MD system adjusts to give a stronger discrimination
of perceptual information under conditions of high, com-
pared to low, perceptual difficulty (Woolgar et al., 2015;
Woolgar, Hampshire, et al., 2011). Here we extend this
work to a non-perceptual domain, demonstrating that
the coding of task rules is sensitive to rule complexity,
with a stronger representation of rules that are behavior-
ally more confusable.
Adaptive rule representation provides a potential
mechanism by which cognitive control can adjust as task
demands change. Converging evidence comes from the
observation that MD coding of task rules is diminished
after a week of practice ( Woolgar, Hampshire, et al.,
2011). The current data show that rule coding adapts in
response to externally imposed changes in difficulty on a
much shorter timescale (between 2 min blocks), making
a clearer case for this system as supporting cognitive con-
trol on the rapid basis required for behavior. Consistent
with this interpretation, Waskom et al. (2014) showed
that frontoparietal coding of attentional context in the
IFS, IPS, and occipital temporal cortex was greatest for
trials immediately following a switch in context relative
to subsequent nonswitch trials.
The present results cannot easily be accounted for by
other major theories of prefrontal function that empha-
size fixed functional specialization of prefrontal regions.
For example, although our study was not designed to
be a direct test of these frameworks, because each rule
required contextual control by a cue presented concur-
rent with the stimulus (Koechlin & Summerfield, 2007;
Koechlin et al., 2003) and each had two layers of abstrac-
tion (Badre & D’Esposito, 2007, 2009), the accounts of
Badre and D’Esposito (Badre & D’Esposito, 2007, 2009;
Badre, 2008) and Koechlin and Summerfield (2007)
would not predict the involvement of regions anterior
to posterior pFC. Yet, significant rule coding was seen
here in our IFS ROI, as in our previous studies ( Woolgar,
Hampshire, et al., 2011; Woolgar, Thompson, et al.,
2011), and the searchlight analysis suggested this may
have been driven by the anterior portion of the IFS in
both hemispheres. In this regard, our results parallel a
recent univariate study specifically designed to address
this question that found activation throughout the MD
system, but no frontal lobe hierarchy, for rules similar
to ours (Crittenden & Duncan, 2014). Moreover, for
our rules the required stimulus–response transformation
was always conditional on a cue determining which set of
stimulus–response transformations to use, so our easy
and hard rules did not differ on the number of layers
of conditional dependency (Badre & D’Esposito, 2007,
2009), and the cue indicating which rule to use was al-
ways presented concurrently with the stimulus to be dis-
criminated, so our easy and hard rules did not differ in
the need to integrate information over different time-
scales (Koechlin & Summerfield, 2007; Koechlin et al.,
2003). Given this, it is not clear that these theories would
predict our observed difference in rule coding for any
given prefrontal area.
What aspect of each rule is encoded in the MD system?
Although it does not rule out concurrent representa-
tion at other levels, all our analyses examined multivoxel
representation at a specific level of encoding: that at which
patterns of activation were consistent across the stimulus–
response pairs of one rule and different from the set of
pairs in the other rule. This makes the results presented
here all the more surprising. If we consider each rule to
be a set of stimulus–response transformations (Duncan,
1977; Rabbitt & Vyas, 1973), the coding requirements of
the easy and hard rules are clearly distinct. For each of
the two easy rules, there is a single transformation to be
used across all stimulus positions (Rule 1: alternate finger
within hemisphere, Rule 2: reflection across the midline;
Figure 1A; Duncan, 1977, 1978). This means that, across
the different stimulus instances, a code representing the
transformation would be consistent within rule and differ-
ent between the rules, making the two easy rules decod-
able. In contrast, for the two hard rules there are four
separate stimulus–response transformations to encode,
and the same transformations are used in both rules
(Figure 1B). Across different stimuli, the code within rule
would be inconsistent, whereas across rules the codes
consist of the same transformations, making them more
confusable and the two hard rules less decodable. There-
fore, if these rules were encoded purely at the level of
transformations, we would see stronger decoding of the
easy rules (internally consistent, distinct across rules) com-
pared to the hard rules (internally inconsistent, sim-
ilar across rules), not the opposite. Alternatively, if each
stimulus–response pair were encoded separately, we
would see equal classification for the two rules: because
all rules consist of four unique stimulus–response pairs,
the two easy and the two hard rules differ in all compo-
nents. Instead, because our analyses required the classifier
to generalize over specific stimulus–response combina-
tions, the observed pattern of results is only possible if
the multivoxel patterns reflect representation of the set
of stimulus–response transformations in each rule. Further
work is needed to understand precisely what combination
of factors makes the hard rules more difficult to imple-
ment than the easy ones, but our results imply that these
rules must at some level be coded as whole sets of trans-
formations, with better discrimination between the more
challenging wholes although they consist of more sim-
ilar parts.
Could behavioral differences have contributed to the
difference in neural coding between our conditions? Trial
by trial differences in RT were accounted for at the first
1906
Journal of Cognitive Neuroscience
Volume 27, Number 10
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
0
7
/
1
1
8
0
9
/
5
1
1
8
9
9
4
5
9
/
6
1
2
6
7
8
o
3
c
2
n
4
_
3
a
/
_
j
0
o
0
c
8
n
2
7
_
a
p
_
d
0
0
b
8
y
2
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
f
/
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
level of analysis by using a variable epoch model (Grinband
et al., 2008; Henson, 2007), in which each trial was mod-
eled as a box car from stimulus onset until response. This
model explicitly takes into account the time on task on
each trial, which should mitigate the contribution of RT
differences to multivoxel patterns (Woolgar et al., 2014).
Additionally, the RT differences between the two rules
was not significantly different between the two difficulty
conditions, and the direction of the numerical difference
(a bigger difference between the two easy rules than be-
tween the two hard rules) should have acted against our
effect, driving more coding in the easy condition (Todd
et al., 2013). Participants were also faster and more accu-
rate on the two easy compared to the two hard rules,
meaning that there were more correct trials contributing
to the analysis of easy rule coding. In terms of amount of
data, this should again act against our effect, driving, if
anything, stronger decoding in the easy condition. A final
possibility is that the increased effort in the hard condi-
tion might simply lead to more coding in general in the
hard condition. However, the increase in coding was spe-
cific to the task feature being manipulated (increased
coding of rule whereas coding of position and response
was unchanged) and specific to the MD system (no
change in coding in the visual ROI), ruling out this alter-
native explanation.
In our design, each rule was consistently cued by a sin-
gle background color, meaning that multivoxel discrimi-
nation between rules could be based on differential
coding of the two visual colors rather than a representa-
tion of the set of stimulus–response mappings them-
selves. We have addressed this question elsewhere
(Woolgar, Thompson, et al., 2011) and found the contri-
bution of the background color to be minimal in the MD
system. In our previous study, discrimination between
two background colors cueing the same rule was weak,
and rule information was shown to be extracted away
from (and generalize over) cue color. Moreover, it seems
unlikely that the visual properties of the different colors
could drive the pattern of results seen here in the MD
system, whereby rule coding was stronger for the more
complex rules. This does, however, offer a simple expla-
nation for the observation that rules could be distin-
guished in the pattern of activation in the occipital
cortex (BA 17/18), because the strength of this classifica-
tion did not vary with rule difficulty.
The MD representation of the easy rules used here was
noticeably weaker than the representation of the same
stimulus–response mapping rules in a previous experi-
ment ( Woolgar, Thompson, et al., 2011), raising the in-
triguing possibility that the wider experimental context
can influence which aspects of the task are represented
in the MD regions. Although one possible interpretation
of the data presented here is simply that the MD regions
only code rules of the “hard” type, integration of the cur-
rent data set with the literature and our previous work
makes this interpretation unlikely. Here, the inclusion
of the more difficult rules in the current study changed
the context in which the easy rules were performed.
We speculate that, in the context of the (even more) dif-
ficult set of rules, processing resources were differentially
recruited, resulting in relatively weak MD coding of the
easy rules. There are a few caveats, however, which need
to be considered. First, in the previous study, each rule
was cued by two different screen background colors,
whereas here we used only one color per rule. This
may, for example, have encouraged participants to gener-
ate a more abstract representation of each rule in the pre-
vious study. However, strong rule coding was also seen in
a subsequent study using very similar rules ( Woolgar,
Hampshire, et al., 2011), in which only one color cued
each rule. Second, in the current study, participants were
somewhat slower and less accurate on the easy rules
compared to the participants performing the same rules
in the previous study. It is difficult to relate differences in
information coding to behavioral success across studies,
but this hints at an interesting avenue for future research
linking coding in the MD regions to successful task per-
formance. Finally, because there were more conditions,
there was substantially less fMRI data contributing to
the estimated BOLD response for each rule in the current
study than in our previous experiment. Further work is
needed to understand the details of this possible redistri-
bution of processing resources with wider task context
and its implications for performance.
We were motivated to examine information coding in a
specific and well-defined network of brain regions
thought to be candidate regions for adaptive coding,
therefore our main analyses focused on the MD regions.
However, our exploratory searchlight analysis revealed
two unexpected findings. First, significant discrimination
between the two easy rules was seen for a large cluster of
voxels in the dorsal precuneus, which survived voxelwise
FDR correction for multiple comparisons. This anatomi-
cal region is commonly considered to be a major compo-
nent of the default mode network (e.g., Fransson &
Marrelec, 2008; Cavanna, 2007), and indeed, our cluster
of significant rule decoding was almost entirely within the
canonical definition of the default mode network given
by Power et al. (2011). This was surprising given that ac-
tivity in the default mode network is usually associated
with the task negative or resting state (e.g., Fox et al.,
2005; Raichle et al., 2001; Shulman et al., 1997), and with
internal modes of cognition such as introspection, epi-
sodic memory, thinking about the future, and first-person
perspective taking (e.g., Buckner & Carroll, 2007) rather
than active maintenance or processing of externally driven
task-relevant information. However, recent work has sug-
gested that the functional connectivity of the precuneus
may be state dependent, showing functional connectivity
to the default mode network at rest but functional con-
nectivity with a frontoparietal network overlapping with
the MD network during active task performance (Utevsky,
Smith, & Huettel, 2014). Here we show multivoxel
Woolgar et al.
1907
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
0
7
/
1
1
8
0
9
/
5
1
1
8
9
9
4
5
9
/
6
1
2
6
7
8
o
3
c
2
n
4
_
3
a
/
_
j
0
o
0
c
8
n
2
7
_
a
p
_
d
0
0
b
8
y
2
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
.
/
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
discrimination of task-relevant rule information in this re-
gion under conditions of active task performance. One in-
teresting possibility is that, in the context of a more
challenging task that occupies the MD network, this re-
gion may be recruited to represent the relatively simple
task information. However, note that in our data there
was no evidence that rule coding in this region was stron-
ger for easy compared to hard rules. Additionally, our
whole-brain univariate analysis found the opposite result
for activation: Along with the MD system, this region was
more active for hard than for easy rules. Further work is
needed to understand what this region contributes to
active tasks of different difficulties.
An additional region identified by our searchlight anal-
ysis, which did not survive correction for multiple com-
parisons, was a dorsal region of the left caudate
nucleus, which showed stronger rule coding for hard rel-
ative to easy rules. Although rule coding in this region
would be consistent with a role in learning, potentiating,
and switching between stimulus–response associations
(e.g., Helie, Ell, & Ashby, 2015; Packard & Knowlton,
2002; Wise, Murray, & Gerfen, 1996) and dorsolateral re-
gion of the caudate nucleus does receive anatomical pro-
jections from dorsolateral prefrontal and posterior
parietal cortex (Alexander, DeLong, & Strick, 1986, see
also e.g., Postuma & Dagher, 2006; Lehericy et al.,
2004; Strafella, Paus, Barrett, & Dagher, 2001), we wish
to replicate this finding before placing much emphasis
on it.
The adaptive coding hypothesis is sometimes mis-
represented as suggesting that the pFC is undifferenti-
ated or there is no specificity of function between different
prefrontal regions. In fact, the proposal is that adaptive
coding is a property of a specific set of prefrontal regions,
which are observed to be active in many different tasks
(Duncan, 2010, 2013). It allows, for example, the possibil-
ity that a distinct set of cognitive functions is associated
with the most anterior parts of the pFC (e.g., Roca et al.,
2011). In our data, we found that the MD regions showed
an adaptive representation of task rules, and our explor-
atory searchlight analysis did not suggest the involvement
of any additional prefrontal regions. However, future work
may benefit from inclusion of a priori prefrontal control
regions to provide a more powerful test of the proposed
regional specificity.
For flexible human behavior we need a cognitive con-
trol system that can both represent the rules of our task
and dynamically adjust as the demands of our task
change. According to the adaptive coding hypothesis
(Duncan, 2001, 2010), neurons in specific frontal and pa-
rietal regions dynamically adjust their responses to code
the information that is currently needed for behavior.
Here we demonstrate adaptive coding of rule in the hu-
man MD system. Multivoxel representation of stimulus–
response mapping rules was sensitive to current task dif-
ficulty, the adjustment occurred dynamically, and the di-
rection of the adjustment was such that MD coding of
rule was stronger for rules that were more complex and
behaviorally more confusable. The results suggest a fronto-
parietal system that rapidly and adaptively reorganizes in
response to changing demands, providing a possible
mechanism for the cognitive control of flexible human
behavior.
Acknowledgments
This work was funded under the Australian Research Council
(ARC)’s Discovery Projects funding scheme (DP12102835).
A. W. and M. A. W. are recipients of ARC Fellowships (Discovery
Early Career Researcher Award, DECRA, DE120100898, Queen
Elizabeth II Research Fellowship DP0984919, respectively). We
thank Sehr Javed for help with behavioral piloting of the para-
digm and John Duncan for discussion of experimental design
and interpretation of results.
Reprint requests should be sent to Alexandra Woolgar, Percep-
tion in Action Research Centre and Department of Cognitive
Science, Macquarie University, North Ryde, Sydney, New South
Wales, Australia, 2109, or via e-mail: Alexandra.woolgar@mq.
edu.au.
Note
1. To ensure that the results presented here were not depen-
dent on this arbitrary performance threshold, we repeated our
main analyses using 60% and 80% cutoffs. The same pattern of
results was seen in all cases.
REFERENCES
Alexander, G. E., DeLong, M. R., & Strick, P. L. (1986). Parallel
organization of functionally segregated circuits linking basal
ganglia and cortex. Annual Review of Neuroscience, 9,
357–381.
Andersen, R. A., Essick, G. K., & Siegel, R. M. (1985). Encoding
of spatial location by posterior parietal neurons. Science, 230,
456–458.
Badre, D. (2008). Cognitive control, hierarchy, and the rostro–
caudal organization of the frontal lobes. Trends in Cognitive
Sciences, 12, 193–200.
Badre, D., & D’Esposito, M. (2007). Functional magnetic
resonance imaging evidence for a hierarchical organization of
the prefrontal cortex. Journal of Cognitive Neuroscience, 19,
2082–2099.
Badre, D., & D’Esposito, M. (2009). Is the rostro-caudal axis of
the frontal lobe hierarchical? Nature Reviews Neuroscience,
10, 659–669.
Bode, S., & Haynes, J. D. (2009). Decoding sequential stages of
task preparation in the human brain. Neuroimage, 45,
606–613.
Botvinick, M. M. (2008). Hierarchical models of behavior and
prefrontal function. Trends in Cognitive Sciences, 12, 201–208.
Brainard, D. H. (1997). The Psychophysics Toolbox. Spatial
Vision, 10, 433–436.
Brass, M., & von Cramon, D. Y. (2004). Decomposing
components of task preparation with functional magnetic
resonance imaging. Journal of Cognitive Neuroscience, 16,
609–620.
Brett, M., Anton, J.-L., Valabregue, R., & Poline, J.-B. (2002).
Region of interest analysis using an SPM toolbox [abstract].
8th International Conference on Functional Mapping of
the Human Brain. Sendai, Japan, Neuroimage, 16,
No. 2 (abstract 497).
1908
Journal of Cognitive Neuroscience
Volume 27, Number 10
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
0
7
/
1
1
8
0
9
/
5
1
1
8
9
9
4
5
9
/
6
1
2
6
7
8
o
3
c
2
n
4
_
3
a
/
_
j
0
o
0
c
8
n
2
7
_
a
p
_
d
0
0
b
8
y
2
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
f
t
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Buckner, R. L., & Carroll, D. C. (2007). Self-projection and the
brain. Trends in Cognitive Sciences, 11, 49–57.
Bunge, S. A. (2004). How we use rules to select actions: A
review of evidence from cognitive neuroscience. Cognitive,
Affective, & Behavioral Neuroscience, 4, 564–579.
Bunge, S. A., & Zelazo, P. D. (2006). A brain-based account of
the development of rule use in childhood. Current
Directions in Psychological Science, 15, 118–121.
Buschman, T. J., Denovellis, E. L., Diogo, C., Bullock, D.,
& Miller, E. K. (2012). Synchronous oscillatory neural
ensembles for rules in the prefrontal cortex. Neuron, 76,
838–846.
Cavanna, A. E. (2007). The precuneus and consciousness. CNS
Spectrums, 12, 545–552.
Chang, C.-C., & Lin, C.-J. (2011). LIBSVM: A library for support
vector machines. ACM Transactions on Intelligent Systems
and Technology, 2, 27.
Christoff, K., & Keramatian, K. (2007). Abstraction of mental
representations: Theoretical considerations and
neuroscientific evidence. In S. A. Bunge & J. D. Wallis (Eds.),
Perspectives on Rule-guided Behavior (pp. 107–126).
New York: Oxford University Press.
Cohen, J. (2013). Statistical power analysis for the behavioral
sciences. New York: Academic Press.
Cole, M. W., Etzel, J. A., Zacks, J. M., Schneider, W., & Braver,
T. S. (2011). Rapid transfer of abstract rules to novel contexts
in human lateral prefrontal cortex. Frontiers in Human
Neuroscience, 5, 142.
Cole, M. W., & Schneider, W. (2007). The cognitive control
network: Integrated cortical regions with dissociable
functions. Neuroimage, 37, 343–360.
Corbetta, M., & Shulman, G. L. (2002). Control of goal-directed
and stimulus-driven attention in the brain. Nature Reviews
Neuroscience, 3, 201–215.
Crittenden, B. M., & Duncan, J. (2014). Task difficulty
manipulation reveals multiple demand activity but no frontal
lobe hierarchy. Cerebral Cortex, 24, 532–540.
Cromer, J. A., Roy, J. E., & Miller, E. K. (2010). Representation of
multiple, independent categories in the primate prefrontal
cortex. Neuron, 66, 796–807.
Crone, E. A., Wendelken, C., Donohue, S. E., & Bunge, S. A.
(2006). Neural evidence for dissociable components of task-
switching. Cerebral Cortex, 16, 475–486.
Cusack, R., Mitchell, D. J., & Duncan, J. (2010). Discrete
object representation, attention switching, and task difficulty
in the parietal lobe. Journal of Cognitive Neuroscience, 22,
32–47.
Dehaene, S., & Changeux, J. P. (2011). Experimental and
theoretical approaches to conscious processing. Neuron, 70,
200–227.
Dehaene, S., Kerszberg, M., & Changeux, J. P. (1998). A
neuronal model of a global workspace in effortful cognitive
tasks. Proceedings of the National Academy of Sciences,
U.S.A., 95, 14529–14534.
Dehaene, S., & Naccache, L. (2001). Towards a cognitive
neuroscience of consciousness: Basic evidence and a
workspace framework. Cognition, 79, 1–37.
Desimone, R., & Duncan, J. (1995). Neural mechanisms of
selective visual attention. Annual Review of Neuroscience,
18, 193–222.
Donohue, S. E., Wendelken, C., & Bunge, S. A. (2008). Neural
correlates of preparation for action selection as a function of
specific task demands. Journal of Cognitive Neuroscience,
20, 694–706.
Dosenbach, N. U., Visscher, K. M., Palmer, E. D., Miezin,
F. M., Wenger, K. K., Kang, H. C., et al. (2006). A core
system for the implementation of task sets. Neuron, 50,
799–812.
Dumontheil, I., Thompson, R., & Duncan, J. (2011). Assembly
and use of new task rules in fronto-parietal cortex. Journal of
Cognitive Neuroscience, 23, 168–182.
Duncan, J. (1977). Response selection errors in spatial choice
reaction tasks. Quarterly Journal of Experimental
Psychology, 29, 415–423.
Duncan, J. (1978). Response selection in spatial choice reaction:
Further evidence against associative models. The Quarterly
Journal of Experimental Psychology, 30, 429–440.
Duncan, J. (2001). An adaptive coding model of neural function
in prefrontal cortex. Nature Reviews Neuroscience, 2,
820–829.
Duncan, J. (2006). EPS Mid-Career Award 2004: Brain
mechanisms of attention. Quarterly Journal of Experimental
Psychology (Hove), 59, 2–27.
Duncan, J. (2010). The multiple-demand (MD) system of the
primate brain: Mental programs for intelligent behaviour.
Trends in Cognitive Sciences, 14, 172–179.
Duncan, J. (2013). The structure of cognition: Attentional
episodes in mind and brain. Neuron, 80, 35–50.
Duncan, J., & Miller, E. K. (2002). Cognitive focus through
adaptive neural coding in the primate prefrontal cortex.
In D. T. Stuss & R. T. Knight (Eds.), Principals of frontal
lobe function (pp. 278–291). New York: Oxford University
Press.
Duncan, J., & Miller, E. K. (2013). Adaptive neural coding in
frontal and parietal cortex. In D. T. Stuss & R. T. Knight
(Eds.), Principles of frontal lobe function (2nd ed.,
pp. 292–301). Oxford, UK: Oxford University Press.
Duncan, J., & Owen, A. M. (2000). Common regions of the
human frontal lobe recruited by diverse cognitive demands.
Trends in Neurosciences, 23, 475–483.
Fedorenko, E., Duncan, J., & Kanwisher, N. (2013). Broad
domain generality in focal regions of frontal and parietal
cortex. Proceedings of the National Academy of Sciences,
110, 16616–16621.
Fox, M. D., Snyder, A. Z., Vincent, J. L., Corbetta, M., Van Essen,
D. C., & Raichle, M. E. (2005). The human brain is intrinsically
organized into dynamic, anticorrelated functional networks.
Proceedings of the National Academy of Sciences, U.S.A.,
102, 9673–9678.
Fransson, P., & Marrelec, G. (2008). The precuneus/posterior
cingulate cortex plays a pivotal role in the default mode
network: Evidence from a partial correlation network
analysis. Neuroimage, 42, 1178–1184.
Freedman, D. J., & Assad, J. A. (2006). Experience-dependent
representation of visual categories in parietal cortex. Nature,
443, 85–88.
Freedman, D. J., Riesenhuber, M., Poggio, T., & Miller, E. K.
(2001). Categorical representation of visual stimuli in the
primate prefrontal cortex. Science, 291, 312–316.
Gail, A., & Andersen, R. A. (2006). Neural dynamics in monkey
parietal reach region reflect context-specific sensorimotor
transformations. The Journal of Neuroscience, 26,
9376–9384.
Grinband, J., Wager, T. D., Lindquist, M., Ferrera, V. P., &
Hirsch, J. (2008). Detection of time-varying signals in event-
related fMRI designs. Neuroimage, 43, 509–520.
Harel, A., Kravitz, D. J., & Baker, C. I. (2014). Task context
impacts visual object processing differentially across the
cortex. Proceedings of the National Academy of Sciences,
111, E962–E971.
Haynes, J. D., & Rees, G. (2005). Predicting the orientation of
invisible stimuli from activity in human primary visual cortex.
Nature Neuroscience, 8, 686–691.
Haynes, J. D., Sakai, K., Rees, G., Gilbert, S., Frith, C., &
Passingham, R. E. (2007). Reading hidden intentions in the
human brain. Current Biology, 17, 323–328.
Woolgar et al.
1909
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
0
7
/
1
1
8
0
9
/
5
1
1
8
9
9
4
5
9
/
6
1
2
6
7
8
o
3
c
2
n
4
_
3
a
/
_
j
0
o
0
c
8
n
2
7
_
a
p
_
d
0
0
b
8
y
2
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
t
f
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Hebart, M. N., Görgen, K., & Haynes, J.-D. (2015). The
Decoding Toolbox (TDT): A versatile software package for
multivariate analyses of functional imaging data. Frontiers in
Neuroinformatics, 8, 8.
Helie, S., Ell, S. W., & Ashby, F. G. (2015). Learning robust
cortico-cortical associations with the basal ganglia: An
integrative review. Cortex, 64C, 123–135.
Henson, R. N. (2007). Efficient experimental design for fMRI.
In R. S. Frackowiak, J. T. Ashburner, S. J. Kiebel, T. E. Nichols,
& W. D. Penny (Eds.), Statistical parametric mapping.
The analysis of functional brain images (pp. 193–210).
London: Academic Press.
Hoshi, E., Shima, K., & Tanji, J. (1998). Task-dependent
selectivity of movement-related neuronal activity in the
primate prefrontal cortex. Journal of Neurophysiology, 80,
3392–3397.
Ibos, G., Duhamel, J.-R., & Ben Hamed, S. (2013). A functional
hierarchy within the parietofrontal network in stimulus
selection and attention control. The Journal of
Neuroscience, 33, 8359–8369.
Kadohisa, M., Petrov, P., Stokes, M., Sigala, N., Buckley, M.,
Gaffan, D., et al. (2013). Dynamic construction of a coherent
attentional state in a prefrontal cell population. Neuron,
80, 235–246.
Keppel, G. (1991). Design and analysis: A researcher’s
handbook. Englewood Cliffs, NJ: Prentice-Hall, Inc.
Koechlin, E., & Jubault, T. (2006). Broca’s area and the
hierarchical organization of human behavior. Neuron, 50,
963–974.
Koechlin, E., Ody, C., & Kouneiher, F. (2003). The architecture
of cognitive control in the human prefrontal cortex. Science,
302, 1181–1185.
during working memory, episodic memory, and semantic
memory. Neuropsychologia, 41, 371–377.
O’Reilly, R. C. (2010). The what and how of prefrontal cortical
organization. Trends in Neurosciences, 33, 355–361.
Owen, A. M., McMillan, K. M., Laird, A. R., & Bullmore, E.
(2005). N-back working memory paradigm: A meta-analysis of
normative functional neuroimaging studies. Human Brain
Mapping, 25, 46–59.
Packard, M. G., & Knowlton, B. J. (2002). Learning and memory
functions of the basal ganglia. Annual Review of
Neuroscience, 25, 563–593.
Passingham, R. E., Toni, I., & Rushworth, M. F. S. (2000).
Specialisation within the prefrontal cortex: The ventral
prefrontal cortex and associative learning. Experimental
Brain Research, 133, 103–113.
Postuma, R. B., & Dagher, A. (2006). Basal ganglia functional
connectivity based on a meta-analysis of 126 positron
emission tomography and functional magnetic resonance
imaging publications. Cerebral Cortex, 16, 1508–1521.
Power, J. D., Cohen, A. L., Nelson, S. M., Wig, G. S., Barnes,
K. A., Church, J. A., et al. (2011). Functional network
organization of the human brain. Neuron, 72, 665–678.
Rabbitt, P., & Vyas, S. (1973). What is repeated in the “repetition
effect”. In S. Korublum (Ed.), Attention and performance IV
(pp. 327–342). Waltham, MA: Academic Press.
Raichle, M. E., MacLeod, A. M., Snyder, A. Z., Powers, W. J.,
Gusnard, D. A., & Shulman, G. L. (2001). A default mode of
brain function. Proceedings of the National Academy of
Sciences, U.S.A., 98, 676–682.
Rao, S. C., Rainer, G., & Miller, E. K. (1997). Integration of what
and where in the primate prefrontal cortex. Science, 276,
821–824.
Koechlin, E., & Summerfield, C. (2007). An information
Reverberi, C., Gorgen, K., & Haynes, J. D. (2012a).
theoretical approach to prefrontal executive function. Trends
in Cognitive Sciences, 11, 229–235.
Kriegeskorte, N., Goebel, R., & Bandettini, P. (2006).
Information-based functional brain mapping. Proceedings
of the National Academy of Sciences, U.S.A., 103,
3863–3868.
Lehericy, S., Ducros, M., Van de Moortele, P. F., Francois, C.,
Thivard, L., Poupon, C., et al. (2004). Diffusion tensor fiber
tracking shows distinct corticostriatal circuits in humans.
Annals of Neurology, 55, 522–529.
Compositionality of rule representations in human prefrontal
cortex. Cerebral Cortex, 22, 1237–1246.
Reverberi, C., Gorgen, K., & Haynes, J. D. (2012b). Distributed
representations of rule identity and rule order in human
frontal cortex and striatum. Journal of Neuroscience, 32,
17420–17430.
Riggall, A. C., & Postle, B. R. (2012). The relationship between
working memory storage and elevated activity as measured
with functional magnetic resonance imaging. Journal of
Neuroscience, 32, 12990–12998.
Li, S., Ostwald, D., Giese, M., & Kourtzi, Z. (2007). Flexible
Roca, M., Torralva, T., Gleichgerrcht, E., Woolgar, A.,
coding for categorical decisions in the human brain. Journal
of Neuroscience, 27, 12321–12330.
Luria, A. R. (1966). Higher cortical functions in man. London:
Tavistock.
Miller, E. K., & Cohen, J. D. (2001). An integrative theory of
prefrontal cortex function. Annual Review of Neuroscience,
24, 167–202.
Momennejad, H. (2012). Human anterior prefrontal cortex
encodes the “what” and “when” of future intentions.
Neuroimage, 61, 139–148.
Naghavi, H. R., & Nyberg, L. (2005). Common fronto-parietal
activity in attention, memory, and consciousness: Shared
demands on integration? Consciousness and Cognition, 14,
390–425.
Niendam, T. A., Laird, A. R., Ray, K. L., Dean, Y. M., Glahn, D. C.,
& Carter, C. S. (2012). Meta-analytic evidence for a
superordinate cognitive control network subserving diverse
executive functions. Cognitive, Affective & Behavioral
Neuroscience, 12, 241–268.
Norman, D. A., & Shallice, T. (1980). Attention to action: Willed
and automatic control of behaviour. Chip report 99,
University of California, San Diego.
Nyberg, L., Marklund, P., Persson, J., Cabeza, R., Forkstam, C.,
Petersson, K. M., et al. (2003). Common prefrontal activations
Thompson, R., Duncan, J., et al. (2011). The role of area 10
(BA 10) in human multitasking and in social cognition: A
lesion study. Neuropsychologia, 49, 3525–3531.
Rorden, C., & Brett, M. (2000). Stereotaxic display of brain
lesions. Behavioral Neurology, 12, 191–200.
Roy, J. E., Riesenhuber, M., Poggio, T., & Miller, E. K. (2010).
Prefrontal cortex activity during flexible categorization.
Journal of Neuroscience, 30, 8519–8528.
Seeley, W. W., Menon, V., Schatzberg, A. F., Keller, J., Glover,
G. H., Kenna, H., et al. (2007). Dissociable intrinsic connectivity
networks for salience processing and executive control.
Journal of Neuroscience, 27, 2349–2356.
Shulman, G. L., Fiez, J. A., Corbetta, M., Buckner, R. L., Miezin,
F. M., Raichle, M. E., et al. (1997). Common blood flow
changes across visual tasks: II. Decreases in cerebral cortex.
Journal of Cognitive Neuroscience, 9, 648–663.
Sigala, N., Kusunoki, M., Nimmo-Smith, I., Gaffan, D., &
Duncan, J. (2008). Hierarchical coding for sequential
task events in the monkey prefrontal cortex. Proceedings
of the National Academy of Sciences, U.S.A., 105,
11969–11974.
Soon, C. S., Namburi, P., & Chee, M. W (2013). Preparatory
patterns of neural activity predict visual category search
speed. Neuroimage, 66, 215–222.
1910
Journal of Cognitive Neuroscience
Volume 27, Number 10
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
0
7
/
1
1
8
0
9
/
5
1
1
8
9
9
4
5
9
/
6
1
2
6
7
8
o
3
c
2
n
4
_
3
a
/
_
j
0
o
0
c
8
n
2
7
_
a
p
_
d
0
0
b
8
y
2
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
f
t
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Stiers, P., Mennes, M., & Sunaert, S. (2010). Distributed task
coding throughout the multiple demand network of the
human frontal-insular cortex. Neuroimage, 52, 252–262.
Stoet, G., & Snyder, L. H. (2004). Single neurons in posterior
parietal cortex of monkeys encode cognitive set. Neuron,
42, 1003–1012.
Stokes, M. G., Kusunoki, M., Sigala, N., Nili, H., Gaffan, D., &
Duncan, J. (2013). Dynamic coding for cognitive control in
prefrontal cortex. Neuron, 78, 364–375.
Strafella, A. P., Paus, T., Barrett, J., & Dagher, A. (2001).
Repetitive transcranial magnetic stimulation of the human
prefrontal cortex induces dopamine release in the caudate
nucleus. Journal of Neuroscience, 21, RC157.
Todd, M. T., Nystrom, L. E., & Cohen, J. D. (2013). Confounds
in multivariate pattern analysis: Theory and rule
representation case study. Neuroimage, 77, 157–165.
Utevsky, A. V., Smith, D. V., & Huettel, S. A. (2014). Precuneus
is a functional core of the default-mode network. The Journal
of Neuroscience, 34, 932–940.
Vincent, J. L., Kahn, I., Snyder, A. Z., Raichle, M. E., & Buckner,
R. L. (2008). Evidence for a frontoparietal control system
revealed by intrinsic functional connectivity. Journal of
Neurophysiology, 100, 3328–3342.
Wallis, J. D., Anderson, K. C., & Miller, E. K. (2001). Single
neurons in prefrontal cortex encode abstract rules. Nature,
411, 953–956.
Wallis, J. D., & Miller, E. K. (2003). From rule to response:
Neuronal processes in the premotor and prefrontal cortex.
Journal of Neurophysiology, 90, 1790–1806.
Waskom, M. L., Kumaran, D., Gordon, A. M., Rissman, J., &
Wagner, A. D. (2014). Frontoparietal representations of task
context support the flexible control of goal-directed
cognition. Journal of Neuroscience, 34, 10743–10755.
Wendelken, C., Munakata, Y., Baym, C., Souza, M., & Bunge,
S. A. (2012). Flexible rule use: Common neural substrates in
children and adults. Developmental Cognitive Neuroscience,
2, 329–339.
White, I. M., & Wise, S. P. (1999). Rule-dependent neuronal
activity in the prefrontal cortex. Experimental Brain
Research, 126, 315–335.
Wise, S. P., Murray, E. A., & Gerfen, C. R. (1996). The frontal
cortex-basal ganglia system in primates. Critical Reviews in
Neurobiology, 10, 317–356.
Woolgar, A., Bor, D., & Duncan, J. (2013). Global increase
in task-related fronto-parietal activity after focal frontal
lobe lesion. Journal of Cognitive Neuroscience, 25,
1542–1552.
Woolgar, A., Golland, P., & Bode, S. (2014). Coping with
confounds in multivoxel pattern analysis: What should we do
about reaction time differences? A comment on Todd,
Nystrom & Cohen 2013. Neuroimage, 98, 506–512.
Woolgar, A., Hampshire, A., Thompson, R., & Duncan, J. (2011).
Adaptive coding of task-relevant information in human
frontoparietal cortex. Journal of Neuroscience, 31,
14592–14599.
Woolgar, A., Parr, A., Cusack, R., Thompson, R., Nimmo-Smith,
I., Torralva, T., et al. (2010). Fluid intelligence loss linked to
restricted regions of damage within frontal and parietal
cortex. Proceedings of the National Academy of Sciences,
U.S.A., 107, 14899–14902.
Woolgar, A., Thompson, R., Bor, D., & Duncan, J. (2011). Multi-
voxel coding of stimuli, rules, and responses in human
frontoparietal cortex. Neuroimage, 56, 744–752.
Woolgar, A., Williams, M. A., & Rich, A. N. (2015). Attention
enhances multi-voxel representation of novel objects in frontal,
parietal and visual cortices. Neuroimage, 109, 429–437.
Zhang, J., Kriegeskorte, N., Carlin, J. D., & Rowe, J. B. (2013).
Choosing the rules: Distinct and overlapping frontoparietal
representations of task rules for perceptual decisions.
Journal of Neuroscience, 33, 11852–11862.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
0
7
/
1
1
8
0
9
/
5
1
1
8
9
9
4
5
9
/
6
1
2
6
7
8
o
3
c
2
n
4
_
3
a
/
_
j
0
o
0
c
8
n
2
7
_
a
p
_
d
0
0
b
8
y
2
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
t
.
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Woolgar et al.
1911