Executive Semantic Processing Is Underpinned by
a Large-scale Neural Network: Revealing the
Contribution of Left Prefrontal, Posterior
Temporal, and Parietal Cortex to
Controlled Retrieval and
Selection Using TMS
Carin Whitney1, Marie Kirk1, Jamie OʼSullivan1,
Matthew A. Lambon Ralph2, and Elizabeth Jefferies1
Abstract
■ To understand the meanings of words and objects, we need
to have knowledge about these items themselves plus executive
mechanisms that compute and manipulate semantic information
in a task-appropriate way. The neural basis for semantic control
remains controversial. Neuroimaging studies have focused on
the role of the left inferior frontal gyrus (LIFG), whereas neuro-
psychological research suggests that damage to a widely distrib-
uted network elicits impairments of semantic control. There is
also debate about the relationship between semantic and execu-
tive control more widely. We used TMS in healthy human volun-
teers to create “virtual lesions” in structures typically damaged in
patients with semantic control deficits: LIFG, left posterior mid-
dle temporal gyrus (pMTG), and intraparietal sulcus (IPS). The
influence of TMS on tasks varying in semantic and nonsemantic
control demands was examined for each region within this
hypothesized network to gain insights into (i) their functional
specialization (i.e., involvement in semantic representation, con-
trolled retrieval, or selection) and (ii) their domain dependence
(i.e., semantic or cognitive control). The results revealed that
LIFG and pMTG jointly support both the controlled retrieval
and selection of semantic knowledge. IPS specifically participates
in semantic selection and responds to manipulations of non-
semantic control demands. These observations are consistent
with a large-scale semantic control network, as predicted by le-
sion data, that draws on semantic-specific (LIFG and pMTG) and
domain-independent executive components (IPS). ■
INTRODUCTION
Semantic cognition refers to the ability to assign and use
the meanings of words, sounds, objects, and faces to
interact with the environment. This capacity relies on both
stored semantic knowledge (semantic representations)
and executive control mechanisms that shape semantic
activation in line with current goals and constraints (se-
mantic control). We know a vast amount about any given
concept—yet only particular aspects of our knowledge will
be relevant in a given situation. For example, we know
many different things about bananas, including that they
are peeled before being eaten and that they can make
you slip when dropped on the ground. To understand
the relationship between “banana” and “slip,” it is necessary
to focus on a relatively obscure aspect of meaning (i.e., that
a banana has a slimy texture) as opposed to more domi-
nant aspects that are thought to be retrieved automati-
1University of York, 2University of Manchester
cally (Corbett, Jefferies, & Lambon Ralph, 2009; Badre
& Wagner, 2007; Jefferies, Baker, Doran, & Lambon
Ralph, 2007; Jefferies & Lambon Ralph, 2006; Badre,
Poldrack, Pare-Blagoev, Insler, & Wagner, 2005; Wagner,
Pare-Blagoev, Clark, & Poldrack, 2001; Thompson-Schill,
DʼEsposito, Aguirre, & Farah, 1997). Semantic control pro-
cesses therefore are a principle component of semantic
cognition and interact with stored semantic knowledge
during meaning retrieval.
Neuroimaging and neuropsychological research inves-
tigating the brain mechanisms underpinning semantic con-
trol have highlighted the importance of the left inferior
frontal gyrus (LIFG). LIFG activation typically increases
when weak or unusual relationships need to be identified
in an association task, subordinate meanings of an ambig-
uous word need to be accessed, or the number of response
options is increased, strengthening competition among
potential target items in the semantic network (e.g., Badre
et al., 2005; Noppeney, Phillips, & Price, 2004; Wagner
et al., 2001; Thompson-Schill et al., 1997). Moreover, LIFG
© 2011 Massachusetts Institute of Technology
Published under a Creative Commons Attribution 3.0 Unported (CC-BY 3.0) license
Journal of Cognitive Neuroscience 24:1, pp. 133–147
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
4
/
1
2
4
1
/
3
1
3
/
1
1
9
3
4
3
3
/
4
1
2
7
9
8
o
0
c
2
n
6
_
9
a
/
_
j
0
o
0
c
1
n
2
3
_
a
p
_
d
0
0
b
1
y
2
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
f
t
.
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
lesions in patients with stroke aphasia (SA) produce im-
pairments on similar tasks, establishing a causal relation-
ship between LIFG and semantic control deficits (Noonan,
Jefferies, Corbett, & Lambon Ralph, 2010; Corbett, Jefferies,
Ehsan, & Lambon Ralph, 2009; Corbett, Jefferies, & Lambon
Ralph, 2009; Novick, Kan, Trueswell, & Thompson-Schill,
2009; Soni et al., 2009; Jefferies, Patterson, & Lambon Ralph,
2008). These studies show that individuals with SA have
difficulty selecting appropriate concepts in the face of po-
tent distracters—for example, they have difficulty retrieving
the subordinate meanings of ambiguous words and strug-
gle to reject highly associated distractor words in synonym
judgment. Moreover, they profit from cues provided to re-
duce the requirement for internally generated semantic
control (e.g., /t/ to cue “tiger” during picture naming), dem-
onstrating that semantic knowledge itself is preserved in
the face of impaired semantic control.
Contradictory conclusions, however, have been drawn
about the broader semantic control network, extending
beyond LIFG. One source of controversy follows from the
fact that semantic control deficits are associated with le-
sions to both LIFG and/or temporo-parietal cortex (Novick
et al., 2009; Jefferies & Lambon Ralph, 2006). Although
this suggests a large-scale distributed network, the patientsʼ
lesions are typically extensive, encompassing potentially
separable sites—for example, posterior middle temporal
gyrus (pMTG; BA 21/BA 37) and parietal areas (e.g., intra-
parietal sulcus [IPS] and BA 39/BA 40; Noonan et al., 2010).
Moreover, individuals rarely have specific behavioral impair-
ments but are likely to have damage to more than one
cognitive function. Performance is impaired on a variety
of standard assessments probing attentional/executive skills
outside the verbal domain (e.g., deficits occurred on the
WCST and the Brixton Spatial Rule Attainment task; Novick
et al., 2009; Jefferies & Lambon Ralph, 2006), suggest-
ing that some of the regions affected in SA might serve a
domain-independent control function.
Neuroimaging studies offer higher spatial resolution, but
the interpretation of brain activation remains ambiguous.
For example, in most fMRI investigations, high semantic
control demands are confounded with the number of po-
tential target concepts; thus, these conditions might gen-
erate greater activation in the semantic store (cf. Snijders
et al., 2009; Gennari, MacDonald, Postle, & Seidenberg,
2007; Noppeney et al., 2004). In line with this, pMTG has
been described as a store for semantic knowledge that re-
ceives modulatory signals from prefrontal cortex during
the process of meaning retrieval (Binder, Desai, Graves,
& Conant, 2009; Hickok & Poeppel, 2004; Indefrey &
Levelt, 2004; Gold & Buckner, 2002). This interpretation
is in stark contrast to the neuropsychological profile of
semantically impaired patients with SA: Irrespective of
whether they have left prefrontal or posterior damage
(encompassing pMTG plus other posterior temporal and
inferior parietal areas), SA patients are able to retrieve con-
ceptual knowledge when the control demands of semantic
tasks are reduced, suggesting that this region does not
constitute a key semantic store (e.g., Jefferies & Lambon
Ralph, 2006).
Although pMTG is specifically implicated in semantic
processing, the left IPS, in contrast, does not appear to
be specific for semantic operations, as neural activity in
this area is modulated by a variety of cognitive tasks that
probe executive or attention processes, including spatial
orientation, tone discrimination, finger movement sequenc-
ing, and categorization of faces, as well as tasks using seman-
tic stimuli (Hedden & Gabrieli, 2010; Ciaramelli, Grady,
& Moscovitch, 2008; Collette, Hogge, Salmon, & Van der
Linden, 2006; Duncan, 2006; Wager, Jonides, & Reading,
2004). Because of this domain independence, IPS has
been implicated in the “multiple-demand” (MD) network
alongside medial and dorsal prefrontal structures (Duncan,
2006, 2010; Cristescu, Devlin, & Nobre, 2006; Duncan &
Owen, 2000; Owen, Schneider, & Duncan, 2000). Some of
the impairments seen after temporo-parietal infarcts in SA
patients might, thus, be a consequence of damage to a MD
region (i.e., IPS) rather than lesions to a semantic-specific
control area. We propose that semantic control draws on
both areas that are selectively engaged during tasks that
require manipulation of conceptual knowledge (i.e., LIFG
and pMTG) plus regions that serve a more general purpose
(i.e., the allocation of attention; IPS). However, patient
studies cannot investigate the separate roles of pMTG and
IPS in these aspects of semantic cognition and attention,
because SA patients with posterior lesions typically have
damage to both of these structures.
The aim of this study was to explore the contribution
of LIFG, pMTG, and IPS to control processes focusing
on (i) semantic knowledge and (ii) perceptual decisions
with low conceptual content (i.e., “nonsemantic” control).
To resolve some of the ambiguities arising from previous
research, we utilized TMS, which induces a focal and tran-
sient disruption of neural processing when applied re-
petitively (i.e., a “virtual lesion”; Pascual-Leone, Walsh, &
Rothwell, 2000; Walsh & Cowey, 2000; Walsh & Rushworth,
1999; Pascual-Leone et al., 1998). Although this technique
has been successfully used to complement neuroimag-
ing and neuropsychological studies of semantic process-
ing (Lambon Ralph, Pobric, & Jefferies, 2009; Devlin &
Watkins, 2007; Pobric, Jefferies, & Lambon Ralph, 2007;
Devlin, Matthews, & Rushworth, 2003; Wassermann et al.,
1999), there are next-to-no TMS studies specifically fo-
cused on semantic control. The TMS method enabled us
to explore the individual contribution of relatively small
cortical fields, which cannot be easily separated in studies
of neuropsychological cases (i.e., sites such as pMTG and
inferior parietal lobe). Furthermore, TMS is an interfer-
ence technique and can establish whether stimulated re-
gions play an essential role in particular functions, unlike
fMRI. For example, pMTG shows activation during high-
control semantic tasks in fMRI, but this activation may
not be necessary for semantic control—instead, it may re-
flect the retrieval of a greater number of concepts in these
conditions. However, if TMS to pMTG disrupts semantic
134
Journal of Cognitive Neuroscience
Volume 24, Number 1
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
4
/
1
2
4
1
/
3
1
3
/
1
1
9
3
4
3
3
/
4
1
2
7
9
8
o
0
c
2
n
6
_
9
a
/
_
j
0
o
0
c
1
n
2
3
_
a
p
_
d
0
0
b
1
y
2
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
f
.
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
control, this would be powerful evidence that this region
does play a critical role.
To investigate the specific function of each of the target
areas, we employed two manipulations of semantic con-
trol. There were two tasks with high semantic control de-
mands, which were compared with a condition involving
minimal control requirements (see Figure 1). Both control
manipulations have been shown to produce greater activa-
tion in LIFG, plus pMTG and/or IPS (Badre et al., 2005;
Thompson-Schill et al., 1997; Wagner et al., 2001), making
these tasks ideal to assess the extent of the semantic con-
trol network outside LIFG. In the first of the high-control
tasks, participants had to choose a target concept that was
weakly related to the cue word, as opposed to strongly re-
lated (e.g., salt–grain vs. salt–pepper). In these situations,
additional executive resources are required to retrieve the
target concept from memory (i.e., “controlled semantic
retrieval”), because the cue will not effectively activate
the target via spreading activation in the semantic network
(Wagner et al., 2001; Masson, 1991; Collins & Loftus, 1975).
In the other task, participants were asked to attend to
specific, typically less salient features of word meaning
(e.g., color: salt–dove; both concepts are white) while ignor-
ing strong but task-irrelevant semantic associations at the
same time (e.g., pepper was also presented as a distractor;
see Figure 1). This semantic control process differed from
controlled semantic retrieval in two ways: First, participants
had to select a particular semantic feature, which was task rel-
evant over competing prepotent but irrelevant information
(i.e., “semantic selection”; Badre et al., 2005; Thompson-
Schill et al., 1997). Second, the task required a strategic
top–down approach for meaning recovery, evoked by spe-
cific task instructions (i.e., “associate: color!”). Theories that
differentiate between strategic (top–down) and stimulus-
driven (bottom–up) forms of attention have linked IPS
to conditions where prior information biases task perfor-
mance, for example, when cues indicate the position of
the target on the screen versus no cues (Ciaramelli et al.,
2008). In the feature selection task, the instruction biased
participantsʼ attention toward a specific semantic dimen-
sion and away from strong associations, which acted as dis-
tractors. Hence, we predicted that TMS over IPS would
disrupt the feature selection task, but not the retrieval of
weak associations, for which no biases/cues were neces-
sary. In contrast, TMS to pMTG was expected to disrupt
both tasks tapping semantic control, because fMRI has
revealed activation increases during both conditions (i.e.,
during attention to specific features and for weakly related
cue–target pairs; Wagner et al., 2001; Thompson-Schill et al.,
1997).
To explore the roles of LIFG, pMTG, and IPS in con-
trol functions beyond the semantic domain, a perceptual
matching task was designed using compound letters (e.g.,
an A made of small Bs), with two different levels of execu-
tive/attentive demand (Navon, 1977). Task demands were
higher when participants had to match a cue letter to the
local elements of a compound letter as opposed to its over-
all shape (see Figure 1). This process required inhibition
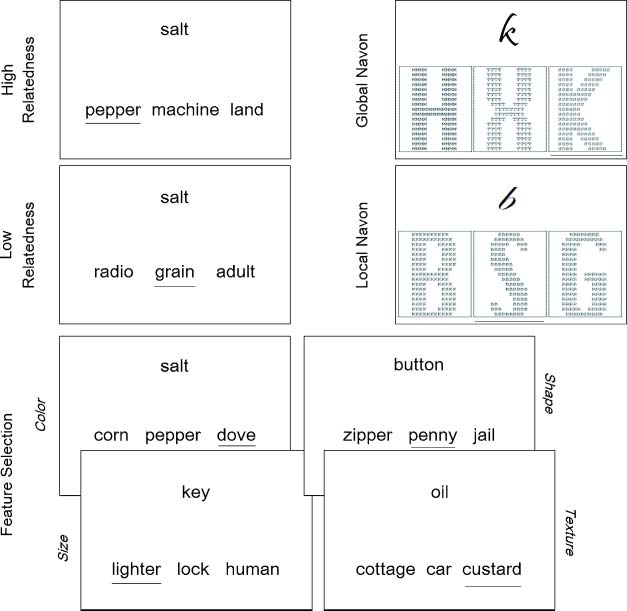
Figure 1. Example trials for
the semantic tasks and the
Navon tasks. Participants had
to select the target word that
was either strongly related to
the cue shown above (high
relatedness), weakly related
(low relatedness), or unrelated
but similar to the cue in
one of the following semantic
dimensions: color, shape, size,
or texture (feature selection).
In the Navon tasks, participants
had to choose the target
compound letter that
resembled the cue letter
either in its global shape
(global Navon) or in its local,
smaller elements (local Navon).
Target items are underlined,
and compound letters are
enlarged for illustration
purposes.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
4
/
1
2
4
1
/
3
1
3
/
1
1
9
3
4
3
3
/
4
1
2
7
9
8
o
0
c
2
n
6
_
9
a
/
_
j
0
o
0
c
1
n
2
3
_
a
p
_
d
0
0
b
1
y
2
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
/
.
f
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Whitney et al.
135
of the visually dominant global shape of the compound
letter plus top–down attention directed toward selective,
subordinate features of the stimulus. As such, the condi-
tion was comparable to the semantic feature selection task
with the exception that participants had to orient toward
spatial/perceptual properties as opposed to semantic fea-
tures of the stimulus (e.g., color). fMRI and TMS studies
have shown parietal cortex to be crucial for the Navon task
(Hedden & Gabrieli, 2010; Billington, Baron-Cohen, &
Bor, 2008; Mevorach, Shalev, Allen, & Humphreys, 2008;
Mevorach, Humphreys, & Shalev, 2005), which is consis-
tent with the view that this region serves an MD function
not limited to semantic processes. The Navon task was,
therefore, used to probe the domain independence of po-
tential semantic control regions. We predicted that TMS
over IPS would impair performance on the Navon task—
because of its function in the MD network mediating top–
down control—but that no TMS effects would emerge
after stimulation of LIFG nor pMTG because of the low
semantic content of the Navon stimuli.
METHODS
Participants
Sixteen right-handed native English speakers from the
University of York participated in this study (eight women;
mean age = 22.25 years, SD = 3.55). All subjects passed
TMS and MRI safety screening (Wassermann, 1998), were
free of medication, and did not have any personal or family
history of neurological or psychiatric illness. Participants
had normal or corrected-to-normal vision and gave in-
formed consent before the beginning of the study. A re-
imbursement of £40 was paid for participation. The study
was approved by the local ethics committee.
Tasks
Three semantic judgment tasks requiring different levels
of executive semantic demand were employed (i.e., judg-
ments involving high relatedness, low relatedness, and
feature selection; see Figure 1). In each task, a cue word
appeared above a row of three potential target words. Par-
ticipants were asked to decide which target was related to
the cue by pressing one of three buttons with their right
hand, corresponding to the position of the response item
(left, middle, and right).
In the high relatedness task, the target was strongly re-
lated to the cue and appeared with two unrelated distractor
items (salt–pepper, machine, land). Semantic control de-
mands were minimal because target retrieval benefitted
from automatic spreading activation (Masson, 1991; Neely,
1990; Collins & Loftus, 1975). In the low relatedness condi-
tion, cue–target associations were weak (salt–grain, radio,
adult) and consequently target retrieval required addi-
tional executive resources that helped to direct the search
and recovery of the relevant item, that is, “controlled se-
mantic retrieval” (Badre et al., 2005; Wagner et al., 2001).
In the feature selection task, the target shared a particular
semantic dimension (color, size, shape, or texture) with
the cue (e.g., color: salt–dove, corn, pepper). The target ap-
peared together with a strong semantic associate and an
unrelated distractor. Target retrieval required the explicit
selection of the appropriate semantic feature (e.g., white)
and the suppression of the dominant but irrelevant asso-
ciate (pepper; “semantic selection”; Badre et al., 2005;
Thompson-Schill et al., 1997). Participantsʼ attention was
biased toward a particular semantic feature before stim-
ulus presentation (via an instruction slide, for example,
“associate: color!”). Therefore, the feature selection task
and the low relatedness task tapped two different forms of
semantic control.
Nonsemantic control tasks were constructed from the
global–local Navon letter-matching task (Navon, 1977).
We produced easy and difficult versions of this task to
establish whether rTMS effects over LIFG/pMTG/IPS re-
mained specific to the semantic domain when control de-
mands were increased. In both Navon conditions, a cue
letter appeared above three larger compound letters,
which were composed of smaller letters (e.g., a large letter
S made of small Bs; see Figure 1). In the easy condition,
participants were asked to decide which compound letter
matched the cue in global shape, irrespective of the letters
that appeared as smaller elements inside the compound.
Cognitive control demands were expected to be minimal
in these trials because the global shape is visually dominant
over local features (Navon, 1977). Moreover, neither dis-
tractor was related to the cue, that is, neither global shape
nor local letter features matched the cue in this condition
(see Figure 1). In contrast, the more difficult local Navon
task required participants to match the cue letter to the
local elements of one of the compounds, hence, to dis-
regard the dominant, global shape of the stimuli. Cognitive
control demands were further increased by presenting a
compound letter whose global shape was identical to the
cue, thus generating a strong task-irrelevant competitor
(see Figure 1). The local Navon task was therefore com-
parable to the feature selection condition, because both
required top–down control processes to direct attention
away from dominant targets and toward selective, sub-
ordinate attributes of the stimulus.
Design and Procedure
A within-subject factorial design was used, with stimula-
tion SITE (LIFG, pMTG, IPS), TMS (stimulation vs. no stimu-
lation), and TASK (three semantic tasks, two Navon tasks)
as within-subject factors. Each site was stimulated on a dif-
ferent day, with test sessions separated by at least 1 week.
The sequence of stimulation site was counterbalanced
across sessions. Furthermore, each session included re-
cordings of task performance immediately after TMS and
without any TMS intervention (“baseline” performance)
to identify the influence of TMS on cognitive behavior
136
Journal of Cognitive Neuroscience
Volume 24, Number 1
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
4
/
1
2
4
1
/
3
1
3
/
1
1
9
3
4
3
3
/
4
1
2
7
9
8
o
0
c
2
n
6
_
9
a
/
_
j
0
o
0
c
1
n
2
3
_
a
p
_
d
0
0
b
1
y
2
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
.
/
t
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
(TMS effect). Baseline performance was measured either
before TMS intervention or 30 min after TMS offset by
which time no TMS effect remains (Pobric, Jefferies, &
Lambon Ralph, 2010; Lambon Ralph et al., 2009; Pobric,
Lambon Ralph, & Jefferies, 2009; Pobric et al., 2007). The
order of baseline assessment was counterbalanced across
sessions.
The six experimental runs—that is, baseline and post-
TMS performance for each stimulation site (LIFG, pMTG,
IPS)—lasted about 6 min each (M = 5.93 min, SD = 0.49)
and included 30 trials per condition. Two miniblocks of
15 consecutive trials were created for each condition and
presented in a pseudorandomized order to control for
effects that relate to the fading of the TMS effect over time.
At the beginning of each block, an instruction slide was
shown, followed by a fixation cross for 500 msec in the
center of the screen. This was replaced by the first experi-
mental trial, which displayed the cue above three response
options for a maximum of 5 sec (Figure 1). As soon as
a response was made, the fixation cross appeared again,
followed by the next trial. A computer running E-prime
(Psychology Software Tools) was used to present the
stimuli and record the responses.
Stimuli
Each of the semantic conditions consisted of 180 cue–
target–distractor trials. The trials were arranged into six
matched sets of 30 trials each, used for each experimen-
tal run, and then split into miniblocks of 15 trials, which
were equated for word length, frequency, and cue–target
association strength. Stimuli were based on Badre et al.
(2005) but restricted to nouns only, and some trials were
amended to make them suitable for U.K. participants.
Words in all three conditions were matched in length in
letters (low relatedness: M = 5.1, SD = 1.4; high related-
ness: M = 5.2, SD = 1.8; feature selection: M = 5.5, SD =
1.8) and frequency (Kucera & Francis, 1967; low related-
ness: M = 54.3, SD = 105.1; high relatedness: M = 48.1,
SD = 90.4; feature selection: M = 44.5, SD = 103.6).
The high and low relatedness tasks were arranged such
that the same cue word was matched with a high or low
semantic associate, using several sets of association norms
(Moss & Older, 1996; Postman & Keppel, 1970). Associa-
tion strength was defined as the proportion of subjects
that named the target in response to the cue in free asso-
ciation. Each cue word was also paired with two unrelated
distractor items, for which no entry in the association norms
was found (e.g., high: salt–pepper, machine, land; low:
salt–grain, radio, adult). The mean association strength for
high and low related cue–target pairs differed significantly
(paired t test: high = 0.24, SD = 0.18; low = 0.04, SD =
0.10; t = 15.00; p < .001). For the feature selection task,
cue words were paired with an unrelated target word that
shared a particular semantic dimension with the cue (i.e.,
color, shape, size, or texture), a strong semantic associate
(mean association strength = 0.22, SD = 0.76) and a new,
unrelated distractor noun (e.g., color: salt–dove, pepper,
cone). Sixteen percent of the cues and strong semantic
associates were taken from the cue–target pairs in the high
relatedness condition and paired with a new target and
a new unrelated noun. Twelve percent of the trials in the
feature selection task needed to be repeated but never
within the same experimental session.
For the nonsemantic control conditions, 180 trials of
the global and local version of the Navon task were con-
structed and, again, broken down into sets of 15 trials.
Navon stimuli were taken from Hills and Lewis (2007).
These depicted 21 upper-case letters (excluding M, N, Q,
V, W) composed of smaller capital letters with a differ-
ent identity (e.g., an A made out of small Bs). The local
elements (width × length: 7 × 7 pixels) were arranged
densely in the shape of the larger compound letter (69 ×
166 pixels), with no gap in between. There were between
3 and 10 different versions of each of the 21 upper-case
letters (made up of different local letters), yielding a total
of 125 unique compound letters. The cues in the Navon
task were 21 lower-case letters that matched the local
elements or global shape of the target compound letter.
To increase the number of trials and to delay response
times, varying script fonts were used (Blackadder, Curlz
MT, Bradleyhand, Edwardian Script, and Pristina), yielding
74 individual cue letters. No cue letters were repeated in
a single experimental run.
TMS Protocol
A standard off-line “virtual lesion” rTMS protocol was
used, which was compatible with established TMS safety
guidelines (Rossi, Hallett, Rossini, & Pascual-Leone, 2009;
Wassermann, 1998). In the absence of any behavioral task,
repetitive trains of TMS were delivered at 1 Hz to the tar-
get brain area for 10 min. This repetitive stimulation results
in a temporary and reversible disruption of neural process-
ing in the underlying tissue, which interferes with tasks that
rely on the stimulated area (Lambon Ralph et al., 2009;
Pobric et al., 2007, 2009; Pascual-Leone et al., 1998). The
resulting behavioral deficits are typically reflected in a
delay in response times rather than a decline in accuracy
(Devlin et al., 2003; Pascual-Leone et al., 2000; Walsh &
Cowey, 2000).
A 50-mm figure-of-eight coil, attached to a Magstim
Rapid2 stimulator, was used for the repetitive delivery of
magnetic pulses. The center of the coil was aligned to
the point that marked the stimulation site on a tight-fitting
elastic cap, worn by the participant. The coil was held firmly
against the scalp throughout stimulation. Stimulation in-
tensity was determined before each rTMS administration
as 120% of active motor threshold (MT). MT was identified
as the lowest intensity that produced a visible muscle
twitch in the tense right hand when intensity was gradually
decreased during single-pulse stimulation of left motor
cortex. Intensity threshold was set to a maximum of 65% of
stimulator output (mean intensity = 62.40%, SD = 3.20).
Whitney et al.
137
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
1
2
4
1
/
3
1
3
/
1
1
9
3
4
3
3
/
4
1
2
7
9
8
o
0
c
2
n
6
_
9
a
/
_
j
0
o
0
c
1
n
2
3
_
a
p
_
d
0
0
b
1
y
2
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
.
f
/
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Coil orientation was manipulated to minimize partici-
pantsʼ discomfort during rTMS (particularly over LIFG),
as previous research found behavioral effects were insen-
sitive to the orientation of the coil (Niyazov, Butler, Kadah,
Epstein, & Hu, 2005). Also, six participants received a
slightly lower intensity for rTMS over this site, ranging from
109% to 116% of individual MT (M = 113%). Despite these
adaptations, LIFG stimulation yielded the strongest perfor-
mance deficits, which were comparable in size to the in-
terference observed in previous rTMS studies that used
the same stimulation protocol and similar semantic tasks
(e.g., Lambon Ralph et al., 2009; Pobric et al., 2007). More-
over, differences in sensory experiences across stimulation
sites (e.g., in general discomfort, noise, or muscle twitches,
which were highest during LIFG stimulation) cannot ac-
count for the TMS effects because (i) performance was
always measured in the absence of any ongoing brain
stimulation and (ii) various control tasks were used to de-
tect any task-independent consequences of TMS (i.e., the
high relatedness and Navon tasks, which were meant to
have no effect after LIFG stimulation).
Localization of Stimulation Sites
Structural T1-weighted MRI scans of each participant were
used to guide coil positioning using the Ascension Minibird
magnetic tracking device and MRIreg software. Five ana-
tomical landmarks (tip and bridge of the nose, left and right
tragus and vertex) were identified to coregister the partici-
pantʼs head, stabilized on a chin rest, with the MRI image
on the screen.
Our stimulation sites were derived from peak activa-
tions identified in fMRI studies that employed the same
stimulus set and/or the same tasks (i.e., low and high relat-
edness, feature selection; Badre et al., 2005; Wagner et al.,
2001; Thompson-Schill et al., 1997). One of these studies
reported activation in all three target sites during increased
executive semantic demands (Badre et al., 2005). Coordi-
nates were transformed into individual subject space using
the transformation matrix from the “segment” function
implemented in SPM5, after the origin of each individual
image was realigned to the anterior commissure. Visual
inspection ensured that coordinates referred to the tar-
get areas by making reference to anatomical landmarks
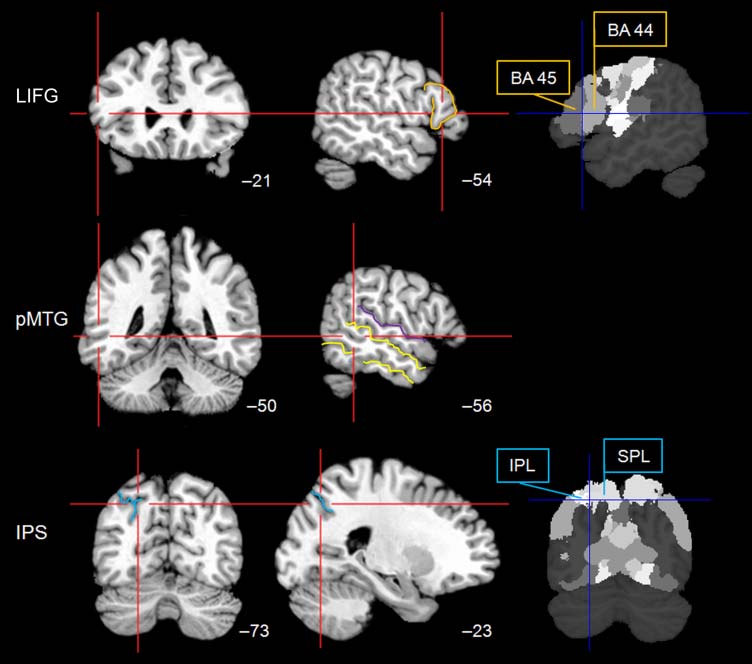
(Figure 2).
Activation peaks within LIFG, observed during previous
fMRI studies of semantic control, were typically large and
widely distributed, comprising both anterior and posterior
segments of this structure. We used the Montreal Neuro-
logical Institute (MNI) coordinates for LIFG (−54 21 12)
from Badre et al. (2005). This area (BA 44/45) in the pars
triangularis (cf. Keller, Crow, Foundas, Amunts, & Roberts,
2009) has been found to be sensitive to several executive
Figure 2. Stimulation sites.
rTMS was delivered to the
pars triangularis of LIFG,
pMTG, and IPS. Images on
the right include probability
maps, which were available
for target regions in BA 44
and BA 45 and the superior
(SPL) and inferior parietal
lobe (IPL). Stimulation sites
are displayed on axial and
saggital slices in MNI space,
with reference to y and x
coordinates, respectively.
orange = pars triangularis,
yellow = inferior and superior
temporal sulcus, purple =
Sylvian fissure, blue = IPS.
138
Journal of Cognitive Neuroscience
Volume 24, Number 1
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
1
2
4
1
/
3
1
3
/
1
1
9
3
4
3
3
/
4
1
2
7
9
8
o
0
c
2
n
6
_
9
a
/
_
j
0
o
0
c
1
n
2
3
_
a
p
_
d
0
0
b
1
y
2
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
/
t
.
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
semantic manipulations including low vs. high related-
ness, feature selection (as opposed to decisions based
on low relatedness), and a task that manipulated target
congruency during feature selection. Moreover, circum-
scribed lesions to this area resulted in poor performance
during executively demanding semantic tasks in aphasic
patients (Novick et al., 2009).
The location for left pMTG stimulation (−56 −50 3) lay
between the superior and inferior temporal sulcus and
was slightly anterior to an imaginary line perpendicular
to the most posterior horizontal segment of the Sylvian
fissure (cf. Gennari et al., 2007; Figure 2). This site, in
BA 21, was identified from the average MNI coordinates
of two studies (Badre et al., 2005; Wagner et al., 2001),
which both reported increased pMTG peak activity in
response to verbal low vs. high relatedness judgments
and when the number of response options was large as
opposed to small. This area is frequently affected in pa-
tients with semantic control deficits following temporo-
parietal infarcts (Noonan et al., 2010).
MNI coordinates for left parietal lobe (−23 −73 48) were
mean values based on Thompson-Schill and colleaguesʼ
study (1997) and referred to an area close to the posterior
bank of the IPS (BA 7; Figure 2). Enhanced parietal activa-
tion was observed during feature selection as opposed to
high relatedness and when the response set was increased
(see also Badre et al., 2005). Furthermore, damage to
the inferior parietal lobule, reaching up to IPS, has been
observed in some patients with SA (e.g., Noonan et al.,
2010; Soni et al., 2009).
Data Analysis
The primary performance measure was RT because RT is
sensitive to rTMS effects even in the absence of any decline
in accuracy (cf. Lambon Ralph et al., 2009; Pobric et al.,
2007; Devlin et al., 2003). RT data were screened for errors
and outliers (±2 SD). We then employed two complemen-
tary analyses. In the first, we used ANOVAs to compare the
impact of TMS across pairs of brain regions (i.e., LIFG
vs. pMTG, LIFG vs. IPS, pMTG vs. IPS) for the semantic
and Navon tasks separately. This is useful because a
three-way interaction would confirm, in line with our pre-
dictions, that the impact of TMS was task- and site-specific,
hence, that brain areas were functionally dissociable within
the control network being tested. Second, we used t tests
to test specific hypotheses regarding the specificity of the
TMS effects, establishing (i) which task(s) were significantly
impaired and (ii) at what site(s). For this analysis, differ-
ence scores were calculated from post-TMS and baseline
sessions for each subject in each condition at each site
(i.e., the TMS effect). These planned t test comparisons
determined whether rTMS-induced effects were present
(two-tailed one-sample t test) and whether these effects
were site-specific and task-specific (two-tailed paired
t tests). Error rates were analyzed using the same model.
RESULTS
RT
ANOVAs
We first considered data from the semantic conditions
(high relatedness, low relatedness, feature selection) to ex-
amine which areas worked together to underpin semantic
control. There was a significant three-way interaction be-
tween Site (n = 2), Task (n = 3), and TMS (baseline vs.
post-TMS) when IPS was compared with LIFG and when
IPS was compared with pMTG (see Table 1 and Figure 3).
However, there were no interactions with Site when LIFG
and pMTG were compared, suggesting that these regions
are functionally dissociable from IPS but similar to each
other in terms of their contribution to semantic control.
The second ANOVA compared the stimulation sites in
a pairwise fashion for the two Navon tasks to establish
whether any of the brain regions contributed to nonseman-
tic forms of control. Again, the results showed a three-way
interaction between Site (n = 2), Task (n = 2), and TMS
(baseline vs. post-TMS) for IPS versus LIFG and IPS versus
pMTG, but not when LIFG and pMTG were compared (see
Table 1 and Figure 3). These results suggest that IPS also
dissociates from LIFG and pMTG in terms of nonsemantic
functions.
TMS Effects across Tasks
We computed the size of the TMS effect for each task at
each stimulation site (i.e., RT for post-TMS minus baseline
performance; see Figure 4), allowing us to draw specific
inferences about the functional role played by each area.
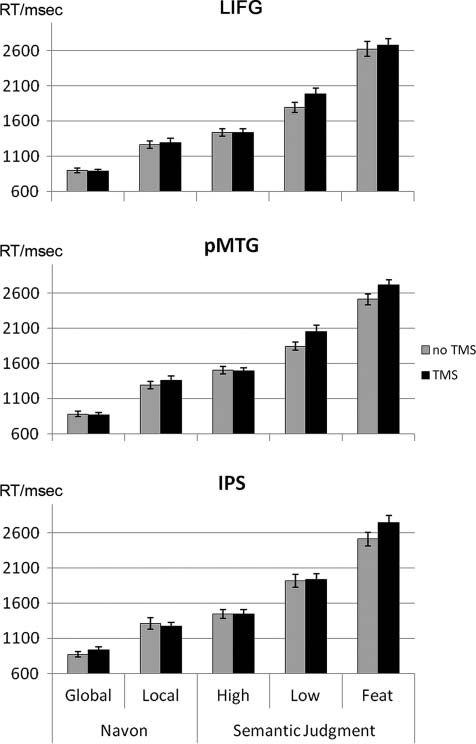
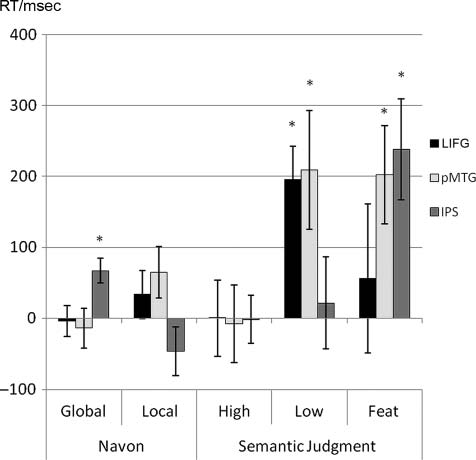
LIFG
rTMS over LIFG slowed performance during the low related-
ness task (one-sample t(15) = 4.24, p = .001) but not the
high relatedness task (one-sample t(15) < 1). Surprisingly,
there was no significant TMS effect for the feature selection
task (one-sample t(15) < 1). Also, none of the Navon tasks
was impaired after TMS (one-sample t(15) < 1). The direct
comparison between the effects for the high versus low re-
latedness condition was significant (paired t(15) = 3.35, p =
.004), supporting a role of LIFG in controlled semantic re-
trieval. Comparison between the two semantic tasks with
high-control demands however revealed that these tasks
were not dissociable (low relatedness vs. feature selection:
paired t(15) = 1.13, p = .28).
pMTG
Stimulation of pMTG disrupted both semantic tasks with
high-control demands (low relatedness: one-sample t(15) =
2.51, p < .05; feature selection: one-sample t(15) = 2.92,
p = .01). There was no TMS effect for the semantic task
with low-control demands (high relatedness: one-sample
t(15) < 1). Again, both Navon tasks remained unaffected
Whitney et al.
139
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
1
2
4
1
/
3
1
3
/
1
1
9
3
4
3
3
/
4
1
2
7
9
8
o
0
c
2
n
6
_
9
a
/
_
j
0
o
0
c
1
n
2
3
_
a
p
_
d
0
0
b
1
y
2
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
f
/
.
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Table 1. F and p Values for the ANOVA for RT
Site
TMS
Task
Site × TMS
Site × Task
TMS × Task
Site × TMS × Task
Semantic Conditions Only (High Relatedness, Low Relatedness, Feature Selection)
df
1, 15
LIFG, IPS
<1
p
.89
1, 15
7.31
.02
2, 30
1, 15
371.14
<1
<.001
.97
pMTG, IPS
<1
13.88
486.93
<1
p
.74
.002
<.001
.42
LIFG, pMTG
<1
12.61
420.01
<1
p
.58
.003
<.001
.48
Nonsemantic Conditions Only (Global and Local Navon Task)
df
1, 15
1, 15
LIFG, IPS
<1
<1
p
.74
pMTG, IPS
<1
p
.94
.46
1.23
.29
1, 15
99.32
<.001
1, 15
<1
.89
108.07
<1
<.001
.67
LIFG, pMTG
<1
<1
121.45
<1
p
.78
.39
<.001
.69
2, 30
<1
.64
<1
.63
1.84
1.76
1, 15
<1
.97
2.27
.15
4.44
.052
2, 30
3.27
.069a
4.99
.014
4.55
.03a
1, 15
4.34
.055
<1
.53
4.78
.045
2, 30
3.44
.045
3.26
.05
<1
.44
1, 15
10.01
.006
17.84
.001
1.91
.19
Pairs of brain regions that were compared are listed in the first column. The three-way interaction omnibus ANOVA, including all sites and
conditions, was also significant (F(8, 120) =2.36; p (Huynh–Feldt).
by TMS (> .09). The size of the TMS effects did not differ
between the two high-control semantic conditions (paired
t(15) < 1), but both effects were larger compared with
trials requiring minimal semantic control, indicating the
importance of pMTG for various types of semantic control
(low vs. high relatedness: paired t(15) = 2.41, p = .03;
feature selection vs. high relatedness: paired t(15) = 3.41,
p = .004).
IPS
rTMS over IPS interfered with the feature selection task
(one-sample t(15) = 3.35, p < .005) and the global Navon
task (one-sample t(15) = 3.86, p < .005). The feature selec-
tion task was more impaired than the other two semantic
conditions (feature selection vs. high relatedness: paired
t(15) = 2.99, p = .01; feature selection vs. low relatedness:
paired t(15) = 2.24, p = .04), consistent with the specific
function of IPS in top–down mediated selection. The global
Navon task was also more affected than the local version
(paired t(15) = 3.28, p = .005).
TMS Effects across Sites
The TMS effects for LIFG and pMTG in the low relatedness
task were larger than the effect of TMS over IPS, which is
in line with our prediction that IPS does not support con-
trolled semantic retrieval (LIFG vs. IPS: paired t(15) = 2.55,
p = .02; pMTG vs. IPS: paired t(15) = 2.00, p = .06). There
was no difference in the effect of TMS across LIFG and
pMTG (paired t(15) < 1), suggesting that both of these
sites play a critical role in the controlled retrieval of seman-
tic information (unlike IPS). In contrast, the TMS effect for
the feature selection task did not differ across the three
sites, implying that all targeted brain areas contribute to
semantic selection (IPS vs. LIFG: paired t(15) = 1.41, p =
.18; pMTG vs. LIFG: paired t(15) = 1.15, p = .27; IPS vs.
MTG: paired t(15) < 1; see Figure 4). Finally, the TMS ef-
fect for the global Navon task was larger for IPS compared
with LIFG or pMTG (IPS vs. LIFG: paired t(15) = 2.59, p =
.02; IPS vs. pMTG: paired t(15) = 3.05, p = .008). This
suggests, together with the findings from the ANOVA, that
IPS contributes to nonsemantic decisions as well as to
aspects of semantic control, unlike LIFG or pMTG.
In summary, the results of the ANOVAs showed that
LIFG and pMTG performed similar functions, while IPS
dissociated from both regions during semantic and non-
semantic control tasks (see Figures 3 and 4). Moreover,
in line with our predictions, planned comparisons revealed
that LIFG and pMTG exclusively mediated semantic control
functions, while IPS contributed to control processes in
both domains (semantic and nonsemantic; Figure 4). Dif-
ferences also emerged regarding the type of semantic con-
trol that was supported by LIFG and pMTG as opposed to
140
Journal of Cognitive Neuroscience
Volume 24, Number 1
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
1
2
4
1
/
3
1
3
/
1
1
9
3
4
3
3
/
4
1
2
7
9
8
o
0
c
2
n
6
_
9
a
/
_
j
0
o
0
c
1
n
2
3
_
a
p
_
d
0
0
b
1
y
2
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
/
f
t
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
highly competitive but irrelevant response (e.g., when “c”
was the cue and a large “C” was the distractor). If the auto-
matic association between globally identical cue and target
letters was reduced by the use of different fonts, strong
inhibition processes may have no longer been required,
which might explain the lack of a TMS effect for the local
Navon task.
Error Rates
ANOVAs
Table 2 lists the error rates that were entered into the
ANOVAs. These analyses revealed that no effects were
driven by Site in any of the six ANOVAs, which compared
pairs of brain regions separately for semantic and Navon
tasks (as in the RT analysis; Table 3). Instead, TMS effects
were dependent on the tasks in the semantic conditions
(i.e., the TMS × Task interaction was significant in all three
ANOVAs comparing pairs of brain regions), while TMS
had no influence on performance of the Navon tasks (i.e.,
no main effects or interactions with TMS were observed;
F < 1).
Planned Comparisons
Planned comparisons on the difference scores between
post-TMS and baseline performance were used to test
specific predictions about the impact of TMS on task per-
formance, separately for each target area. Two effects
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
1
2
4
1
/
3
1
3
/
1
1
9
3
4
3
3
/
4
1
2
7
9
8
o
0
c
2
n
6
_
9
a
/
_
j
0
o
0
c
1
n
2
3
_
a
p
_
d
0
0
b
1
y
2
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
f
/
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 4. TMS effect. Difference scores between post-TMS and
baseline performance (TMS–no TMS) for reaction time. Positive
values indicate a decline in performance after brain stimulation,
whereas negative values indicate improvement. High = high
relatedness; Low = low relatedness; Feat = feature selection.
*p < .05. Error bars denote SEM.
Whitney et al.
141
Figure 3. Reaction times. Performance at baseline (no TMS) and
post-TMS following stimulation of LIFG, pMTG, and IPS. High = high
relatedness; Low = low relatedness; Feat = feature selection. Error
bars denote SEM.
IPS: IPS was the only region that did not engage in con-
trolled semantic retrieval but contributed to feature selec-
tion only.
Unexpectedly, TMS over IPS disrupted the easier global
Navon task, although fMRI studies have shown increased
brain activity in the left IPS when participants attended to
local and less-salient dimensions of Navon letters (corre-
sponding to the Local Navon task; Mevorach et al., 2008).
One possibility is that the intended automatic mapping
of cue and target letter during the global Navon task was
hindered by our use of different fonts, which meant that
sometimes the shape of the cue deviated significantly from
the global shape of the target compound and could be
ambiguous (e.g., a lower-case “e” in font “Curlz” could be
mistaken for a “c”). Therefore, the global Navon task might
have required a stronger level of cognitive control than
expected. Simultaneously, these manipulations had the
opposite effect on the control requirements of the local
Navon task. Here, a globally related letter was used as
one of the distractor items, which was meant to act as a
Table 2. Error Rates
High Relatedness
Low Relatedness
Feature Selection
Global Navon
Local Navon
LIFG
No TMS
TMS
Difference
pMTG
No TMS
TMS
Difference
IPS
No TMS
TMS
Difference
2.00 (2.42)
3.13 (2.99)
1.13 (3.74)
2.94 (2.59)
3.25 (3.66)
0.31 (3.94)
2.81 (3.97)
5.38 (4.21)
2.57 (6.03)
5.75 (4.91)
11.25 (6.44)
5.50 (8.63)
7.44 (6.31)
11.25 (5.69)
3.81 (7.47)
9.06 (8.81)
9.06 (4.93)
0.00 (6.68)
17.31 (10.01)
12.25 (8.51)
−5.06 (11.32)
2.63 (3.07)
1.81 (2.97)
4.75 (12.31)
4.38 (3.50)
−0.82 (4.82)
−0.37 (12.96)
15.06 (7.86)
13.13 (8.37)
−1.93 (8.23)
20.94 (10.85)
15.81 (15.01)
−5.13 (10.76)
1.19 (2.04)
1.50 (1.55)
0.31 (2.82)
3.44 (5.25)
2.81 (2.48)
−0.63 (4.47)
1.81 (2.46)
1.94 (2.62)
0.13 (3.52)
7.38 (13.16)
4.00 (5.27)
−3.38 (13.68)
Percentage of errors committed during performance at baseline (no TMS) and after TMS (TMS) to LIFG, pMTG, and IPS for each task separately.
Difference scores (TMS–no TMS) reflect the TMS effect, with positive values indicating a decline in performance after brain stimulation whereas
negative values signal improvement. Standard deviation is given in parentheses.
Table 3. F and p Values for the ANOVA for Error Rate
Site
TMS
Task
Site × TMS
Site × Task
TMS × Task
Site × TMS × Task
Semantic Conditions Only (High Relatedness, Low Relatedness, Feature Selection)
df
LIFG, IPS
p
pMTG, IPS
p
1, 15
1, 15
2.89
.11
3.12
.10
<1
.86
<1
.94
LIFG, pMTG
<1
<1
p
.70
.44
2, 30
35.49
<.001a
27.75
<.001
41.34
<.001a
1, 15
1.05
.32
<1
<1
.34
.89
Nonsemantic Conditions Only (Global and Local Navon Task)
df
1, 15
1, 15
LIFG, IPS
<1
<1
p
pMTG, IPS
p
LIFG, pMTG
p
.76
2.44
.14
1.49
.24
.36
1.82
.20
<1
6.66
.02
6.31
.02
4.65
.72
.048
<1
<1
<1
.67
.46
.82
2, 30
<1
.44
2.65
.09
<1
.68
1, 15
<1
.56
1.27
.28
<1
.77
2, 30
6.29
.014a
4.42
.036a
7.58
.008a
1, 15
<1
.51
1.30
.27
<1
.89
2, 30
2.07
.16a
44.74
.10
1.01
.38
1, 15
<1
<1
<1
.51
.57
.68
Pairs of brain regions that were compared are listed in the first column.
aSphericity-corrected (Huynh–Feldt).
142
Journal Cognitive Neuroscience
Volume 24, Number 1
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
> words;
Heim et al., 2005; Fiebach et al., 2002). Activation in BA 45
also fluctuated with the degree of semantic processing
involved (e.g., semantic vs. phonological fluency or deci-
sions; Heim, Eickhoff, Ischebeck, et al., 2009; Amunts
et al., 2004). In contrast, changes in phonological stim-
ulus attributes or task demands altered activity in BA 44,
which is a brain region implicated in the dorsal language
pathway (“phonological route”; Heim, Eickhoff, & Amunts,
2009; Saur et al., 2008; Hickok & Poeppel, 2004). In line
with these data, a previous study has shown that TMS
over BA 44 impaired phonological judgments and spared
Whitney et al.
143
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
4
/
1
2
4
1
/
3
1
3
/
1
1
9
3
4
3
3
/
4
1
2
7
9
8
o
0
c
2
n
6
_
9
a
/
_
j
0
o
0
c
1
n
2
3
_
a
p
_
d
0
0
b
1
y
2
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
f
.
/
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
semantic decisions, while the reverse behavioral pattern
was observed after stimulation of BA 45/BA 47 (Gough,
Nobre, & Devlin, 2005). Our study extends these findings,
showing that activity in BA 45 is causally linked to the level
of semantic control needed to retrieve the target concept.
The Role of Left pMTG in Semantic Control
The performance deficits observed after left pMTG stimu-
lation were indistinguishable from the effects of TMS over
LIFG, suggesting that pMTG and LIFG work together as
part of a distributed executive semantic network support-
ing semantic selection and controlled retrieval. These re-
sults are consistent with neuropsychological studies of
patients with SA, who show similar performance deficits
on tasks that tap semantic control following either damage
to temporo-parietal areas, including pMTG, or in combina-
tion with LIFG (Corbett, Jefferies, & Lambon Ralph, 2011;
Noonan et al., 2010; Jefferies & Lambon Ralph, 2006).
Using voxel-based lesion symptom mapping, Schwartz and
colleagues (2009) demonstrated that naming deficits in
aphasia patients could not be attributed to damage in LIFG
or posterior temporal cortex once executive semantic con-
trol processes were controlled for. In contrast, the contribu-
tion of other “semantic” regions in more anterior temporal
lobe was not linked to semantic control. Both of these
lines of research suggest that left pMTG and LIFG perform
similar semantic control functions; however, up until now,
there was little direct evidence that selective disruption
of pMTG and LIFG can produce equivalent deficits in
semantic control.
The TMS results further resolve some of the ambiguity
from the neuroimaging literature regarding pMTG function-
ing, which has linked activation in this area to confound-
ing increases in representational processes during tasks
with high semantic control requirements (Bedny, McGill, &
Thompson-Schill, 2008; Badre et al., 2005; Gold & Buckner,
2002; Wagner et al., 2001). For example, in the feature selec-
tion task, strongly associated but task-irrelevant concepts
are activated alongside the target item (via automatic spread-
ing of activation in the semantic store). Because in our
study, deficits in semantic control were observed as a direct
consequence of temporary disruption to pMTG, it is no
longer plausible to suggest that brain activation in pMTG
during fMRI is a by-product rather than a causal consequence
of manipulations in semantic control processes. Further sup-
port is provided by severely aphasic patients with lesions to
pMTG who are asked to perform sentence–picture match-
ing tasks with different levels of difficulty. Comprehension
is best when the meaning of the sentence can be derived
from the high-frequency nouns alone, hence demonstrat-
ing spared semantic knowledge, compared with sentences
where understanding the more complex verb/verb ar-
gument structure is crucial (Dronkers, Wilkins, Van Valin,
Redfern, & Jaeger, 2004).
An alternative proposal is that pMTG acts as a semantic
store that encodes specific semantic attributes, associated
only with a subset of the stimuli tested in our experiment
(i.e., motion attributes; Wallentin et al., 2011; Dick,
Goldin-Meadow, Hasson, Skipper, & Small, 2009; Martin,
2007; Damasio, Grabowski, Tranel, Hichwa, & Damasio,
1996). It seems unlikely that this could explain why pMTG
stimulation specifically disrupted the control-demanding
semantic tasks and not the low-control condition, because
all three semantic tasks included stimuli from a wide range
of categories. Moreover, pMTG showed activation during
fMRI for the same stimuli, suggesting that even if pMTG
activation is modulated by semantic category/feature, this
site does not have a single role tightly restricted to a spe-
cific category. Further research is clearly needed to estab-
lish whether the same pMTG region responds to semantic
control demands and feature manipulations, such as ac-
tion judgments, and if so, why.
Apart from pMTG, other parts of temporal cortex have
been linked to storing semantic representation, including
more anterior and inferior temporal cortex (for reviews,
see Binder et al., 2009; Patterson, Nestor, & Rogers, 2007).
Bilateral atrophy focused on anterior inferior temporal
cortex results in a gradual degradation of semantic knowl-
edge, as seen in patients with semantic dementia (Binney,
Embleton, Jefferies, Parker, & Lambon Ralph, 2010; Hodges
& Patterson, 2007; Jefferies & Lambon Ralph, 2006; Mummery
et al., 2000; Hodges, Patterson, Oxbury, & Funnell, 1992).
The impairment in these patients is highly consistent across
tasks with varying control demands and depends on fac-
tors that describe the complexity of the semantic represen-
tation being retrieved, such as familiarity and typicality. In
contrast, in individuals with semantic control deficits, le-
sions affect left prefrontal, posterior temporal, and parietal
structures, while the anterior temporal lobe is spared, and
conceptual knowledge remains accessible once semantic
control requirements are reduced (e.g., Jefferies et al.,
2008; Jefferies & Lambon Ralph, 2006). This double dis-
sociation between executive and representational aspects
of semantic cognition in the left temporal cortex was con-
firmed in a recent fMRI investigation (Whitney, Jefferies, &
Kircher, 2011). Semantic selection requirements were linked
to LIFG, pMTG, and parietal cortex, while manipulations of
the number of meanings likely to be activated in a trial
loaded onto more anterior and inferior parts of left temporal
lobe (i.e., BA 20). Together, these observations imply that
semantic representation and control processes rely on dif-
ferent regions within left temporal cortex.
The Role of Left IPS in Semantic and Nonsemantic
Forms of Control
The function of left IPS in the control network was distinct
from that of LIFG and pMTG in two ways. First, IPS was
the only brain region that responded solely to manipu-
lations in semantic selection requirements as opposed to
controlled meaning retrieval. This response pattern was ex-
pected based on studies of attention that implicated IPS
in tasks that required top–down control, elicited by explicit
144
Journal of Cognitive Neuroscience
Volume 24, Number 1
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
4
/
1
2
4
1
/
3
1
3
/
1
1
9
3
4
3
3
/
4
1
2
7
9
8
o
0
c
2
n
6
_
9
a
/
_
j
0
o
0
c
1
n
2
3
_
a
p
_
d
0
0
b
1
y
2
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
/
.
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
cues meant to bias behavior even before stimulus presen-
tation (here: e.g., attend to color), as opposed to control
functions that are driven by the stimulus and are not cued
(Ciaramelli et al., 2008). Second, participation of IPS was
independent of whether semantic or nonsemantic stimuli
were used. rTMS affected response times in both the fea-
ture selection and the global Navon task, indicating that IPS
plays a wider role in cognition than either LIFG or pMTG.
Neuroimaging data reveals that IPS, unlike LIFG and
pMTG, is a brain region that forms part of a fronto-parietal
MD network, supporting tasks that require high executive
control, independently of stimulus modality (although se-
mantic control, as it is examined here, has not been explic-
itly tested; Duncan, 2010; Duncan & Owen, 2000; Owen
et al., 2000). Furthermore, semantically impaired patients
with lesions to temporo-parietal cortex, including IPS, suf-
fer from executive deficits that go far beyond the semantic
domain (Corbett, Jefferies, Ehsan, et al., 2009; Jefferies &
Lambon Ralph, 2006). The strongest support, however,
for a distinction between MD regions (in and around IPS)
and semantic-specific control areas (situated in inferior
frontal and posterior temporal cortex) comes from fMRI
research that has directly compared semantic and non-
semantic executive functions. Consistently, these results
point toward an engagement of the left IPS in any form
of control (Binney et al., 2010; Nagel, Schumacher, Goebel,
& DʼEsposito, 2008; Cristescu et al., 2006). In contrast,
activation in LIFG and pMTG is limited to tasks with high
semantic control demands. Using TMS, we were able to
verify these observations and clearly establish a dissocia-
tion between components in the semantic control network:
Although all three areas (LIFG, pMTG, IPS) were important
for semantic aspects of control, only IPS contributed to
executive functions beyond the semantic domain.
Acknowledgments
This work was supported by a Wellcome project grant (078734/
Z05/Z to E. J. and M. A. L. R.). We thank David Badre, Peter Hills,
and Michael Lewis for supplying their stimulus materials.
Reprint requests should be sent to Carin Whitney or Elizabeth
Jefferies, Department of Psychology, University of York, YO10 5DD,
York, United Kingdom, or via e-mail: c.whitney@psych.york.ac.uk;
ej514@york.ac.uk.
REFERENCES
Amunts, K., Schleicher, A., Burgel, U., Mohlberg, H., Uylings,
H. B., & Zilles, K. (1999). Brocaʼs region revisited:
Cytoarchitecture and intersubject variability. Journal
of Comparative Neurology, 412, 319–341.
Amunts, K., Weiss, P. H., Mohlberg, H., Pieperhoff, P., Eickhoff, S.,
Gurd, J. M., et al. (2004). Analysis of neural mechanisms
underlying verbal fluency in cytoarchitectonically defined
stereotaxic space—The roles of Brodmann areas 44 and 45.
Neuroimage, 22, 42–56.
Badre, D. (2008). Cognitive control, hierarchy, and the
rostro-caudal organization of the frontal lobes. Trends
in Cognitive Sciences, 12, 193–200.
Badre, D., & DʼEsposito, M. (2007). Functional magnetic
resonance imaging evidence for a hierarchical organization
of the prefrontal cortex. Journal of Cognitive Neuroscience,
19, 2082–2099.
Badre, D., Poldrack, R. A., Pare-Blagoev, E. J., Insler, R. Z.,
& Wagner, A. D. (2005). Dissociable controlled retrieval
and generalized selection mechanisms in ventrolateral
prefrontal cortex. Neuron, 47, 907–918.
Badre, D., & Wagner, A. D. (2007). Left ventrolateral
prefrontal cortex and the cognitive control of memory.
Neuropsychologia, 45, 2883–2901.
Bedny, M., McGill, M., & Thompson-Schill, S. L. (2008).
Semantic adaptation and competition during word
comprehension. Cerebral Cortex, 18, 2574–2585.
Billington, J., Baron-Cohen, S., & Bor, D. (2008). Systemizing
influences attentional processes during the Navon task:
An fMRI study. Neuropsychologia, 46, 511–520.
Binder, J. R., Desai, R. H., Graves, W. W., & Conant, L. L.
(2009). Where is the semantic system? A critical review
and meta-analysis of 120 functional neuroimaging studies.
Cerebral Cortex, 19, 2767–2796.
Binney, R. J., Embleton, K. V., Jefferies, E., Parker, G. J., &
Lambon Ralph, M. A. (2010). The ventral and inferolateral
aspects of the anterior temporal lobe are crucial in semantic
memory: Evidence from a novel direct comparison of
distortion-corrected fMRI, rTMS, and semantic dementia.
Cerebral Cortex, 20, 2728–2738.
Ciaramelli, E., Grady, C. L., & Moscovitch, M. (2008). Top–down
and bottom–up attention to memory: A hypothesis (AtoM)
on the role of the posterior parietal cortex in memory
retrieval. Neuropsychologia, 46, 1828–1851.
Collette, F., Hogge, M., Salmon, E., & Van der Linden, M.
(2006). Exploration of the neural substrates of executive
functioning by functional neuroimaging. Neuroscience,
139, 209–221.
Collins, A. M., & Loftus, E. F. (1975). Spreading activation theory
of semantic processing. Psychological Review, 82, 407–428.
Corbett, F., Jefferies, E., Ehsan, S., & Lambon Ralph, M. A.
(2009). Different impairments of semantic cognition in
semantic dementia and semantic aphasia: Evidence from
the non-verbal domain. Brain, 132, 2593–2608.
Corbett, F., Jefferies, E., & Lambon Ralph, M. A. (2009).
Exploring multimodal semantic control impairments in
semantic aphasia: Evidence from naturalistic object use.
Neuropsychologia, 47, 2721–2731.
Corbett, F., Jefferies, E., & Lambon Ralph, M. A. (2011).
Deregulated semantic cognition follows prefrontal and
temporo-parietal damage: Evidence from the impact
of task constraint on nonverbal object use. Journal of
Cognitive Neuroscience, 23, 1125–1135.
Cristescu, T. C., Devlin, J. T., & Nobre, A. C. (2006).
Orienting attention to semantic categories. Neuroimage,
33, 1178–1187.
Damasio, H., Grabowski, T. J., Tranel, D., Hichwa, R. D., &
Damasio, A. R. (1996). A neural basis for lexical retrieval.
Nature, 380, 499–505.
Devlin, J. T., Matthews, P. M., & Rushworth, M. F. (2003).
Semantic processing in the left inferior prefrontal cortex:
A combined functional magnetic resonance imaging
and transcranial magnetic stimulation study. Journal
of Cognitive Neuroscience, 15, 71–84.
Devlin, J. T., & Watkins, K. E. (2007). Stimulating language:
Insights from TMS. Brain, 130, 610–622.
Dick, A. S., Goldin-Meadow, S., Hasson, U., Skipper, J. I.,
& Small, S. L. (2009). Co-speech gestures influence
neural activity in brain regions associated with processing
semantic information. Human Brain Mapping, 30,
3509–3526.
Whitney et al.
145
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
4
/
1
2
4
1
/
3
1
3
/
1
1
9
3
4
3
3
/
4
1
2
7
9
8
o
0
c
2
n
6
_
9
a
/
_
j
0
o
0
c
1
n
2
3
_
a
p
_
d
0
0
b
1
y
2
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
.
f
/
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Dronkers, N. F., Wilkins, D. P., Van Valin, R. D., Jr., Redfern,
B. B., & Jaeger, J. J. (2004). Lesion analysis of the brain
areas involved in language comprehension. Cognition,
92, 145–177.
Duncan, J. (2006). EPS Mid-Career Award 2004: Brain
mechanisms of attention. Quarterly Journal of
Experimental Psychology: (Colchester), 59, 2–27.
Duncan, J. (2010). The multiple-demand (MD) system of the
primate brain: Mental programs for intelligent behavior.
Trends in Cognitive Sciences, 14, 172–179.
Duncan, J., & Owen, A. M. (2000). Common regions of
the human frontal lobe recruited by diverse cognitive
demands. Trends in Neurosciences, 23, 475–483.
Fiebach, C. J., Friederici, A. D., Muller, K., & von Cramon,
D. Y. (2002). fMRI evidence for dual routes to the mental
lexicon in visual word recognition. Journal of Cognitive
Neuroscience, 14, 11–23.
Gennari, S. P., MacDonald, M. C., Postle, B. R., & Seidenberg,
M. S. (2007). Context-dependent interpretation of words:
Evidence for interactive neural processes. Neuroimage,
35, 1278–1286.
Gold, B. T., & Buckner, R. L. (2002). Common prefrontal
regions coactivate with dissociable posterior regions
during controlled semantic and phonological tasks.
Neuron, 35, 803–812.
Gough, P. M., Nobre, A. C., & Devlin, J. T. (2005). Dissociating
linguistic processes in the left inferior frontal cortex with
transcranial magnetic stimulation. Journal of Neuroscience,
25, 8010–8016.
Hedden, T., & Gabrieli, J. D. (2010). Shared and selective neural
correlates of inhibition, facilitation, and shifting processes
during executive control. Neuroimage, 51, 421–431.
Heim, S., Alter, K., Ischebeck, A. K., Amunts, K., Eickhoff,
S. B., Mohlberg, H., et al. (2005). The role of the left
Brodmannʼs areas 44 and 45 in reading words and
pseudowords. Cognitive Brain Research, 25, 982–993.
Heim, S., Eickhoff, S. B., & Amunts, K. (2009). Different roles
of cytoarchitectonic BA 44 and BA 45 in phonological and
semantic verbal fluency as revealed by dynamic causal
modelling. Neuroimage, 48, 616–624.
Heim, S., Eickhoff, S. B., Ischebeck, A. K., Friederici, A. D.,
Stephan, K. E., & Amunts, K. (2009). Effective connectivity
of the left BA 44, BA 45, and inferior temporal gyrus during
lexical and phonological decisions identified with DCM.
Human Brain Mapping, 30, 392–402.
Hickok, G., & Poeppel, D. (2004). Dorsal and ventral streams:
A framework for understanding aspects of the functional
anatomy of language. Cognition, 92, 67–99.
Hills, P. J., & Lewis, M. B. (2007). Temporal limitation of
Navon effect on face recognition. Perceptual and Motor
Skills, 104, 501–509.
Hodges, J. R., & Patterson, K. (2007). Semantic dementia:
A unique clinicopathological syndrome. Lancet Neurology,
6, 1004–1014.
Hodges, J. R., Patterson, K., Oxbury, S., & Funnell, E. (1992).
Semantic dementia. Progressive fluent aphasia with temporal
lobe atrophy. Brain, 115, 1783–1806.
Indefrey, P., & Levelt, W. J. (2004). The spatial and temporal
signatures of word production components. Cognition,
92, 101–144.
Jefferies, E., Baker, S. S., Doran, M., & Lambon Ralph, M. A.
(2007). Refractory effects in stroke aphasia: A consequence
of poor semantic control. Neuropsychologia, 45, 1065–1079.
Jefferies, E., & Lambon Ralph, M. A. (2006). Semantic
impairment in stroke aphasia versus semantic dementia:
A case-series comparison. Brain, 129, 2132–2147.
Jefferies, E., Patterson, K., & Lambon Ralph, M. A. (2008).
Deficits of knowledge versus executive control in semantic
cognition: Insights from cued naming. Neuropsychologia,
46, 649–658.
Keller, S. S., Crow, T., Foundas, A., Amunts, K., & Roberts, N.
(2009). Brocaʼs area: Nomenclature, anatomy, typology
and asymmetry. Brain and Language, 109, 29–48.
Koechlin, E., Ody, C., & Kouneiher, F. (2003). The architecture
of cognitive control in the human prefrontal cortex.
Science, 302, 1181–1185.
Kucera, H., & Francis, W. N. (1967). Computational analysis of
present-day English. Providence, RI: Brown University Press.
Lambon Ralph, M. A., Pobric, G., & Jefferies, E. (2009).
Conceptual knowledge is underpinned by the temporal
pole bilaterally: Convergent evidence from rTMS.
Cerebral Cortex, 19, 832–838.
Martin, A. (2007). The representation of object concepts in
the brain. Annual Review of Psychology, 58, 25–45.
Masson, M. (1991). A distributed memory model of
context effects in word identification. In D. Besner &
G. W. Humphreys (Eds.), Basic processes in reading:
Visual word recognition (pp. 233–263). Hilldale, NJ:
Lawrence Erlbaum Associates.
Mevorach, C., Humphreys, G. W., & Shalev, L. (2005). Attending
to local form while ignoring global aspects depends on
handedness: Evidence from TMS. Nature Neuroscience,
8, 276–277.
Mevorach, C., Shalev, L., Allen, H. A., & Humphreys, G. W. (2008).
The left intraparietal sulcus modulates the selection of low
salient stimuli. Journal of Cognitive Neuroscience, 21, 303–315.
Moss, H., & Older, L. (1996). Birbeck word association
norms. Hove, UK: Psychology Press.
Mummery, C. J., Patterson, K., Price, C. J., Ashburner, J.,
Frackowiak, R. S., & Hodges, J. R. (2000). A voxel-based
morphometry study of semantic dementia: Relationship
between temporal lobe atrophy and semantic memory.
Annals of Neurology, 47, 36–45.
Nagel, I. E., Schumacher, E. H., Goebel, R., & DʼEsposito, M.
(2008). Functional MRI investigation of verbal selection
mechanisms in lateral prefrontal cortex. Neuroimage, 43,
801–807.
Navon, D. (1977). Forest before trees: The precedence of
global features in visual perception. Cognitive Psychology,
9, 353–383.
Neely, J. (1990). Semantic priming in visual word recognition:
A selective review of current theories and findings.
In D. Besner & G. W. Humphreys (Eds.), Basic processes
in reading: Visual word recognition (pp. 264–336).
Hillsdale, NJ: Lawrence Erlbaum Associates.
Niyazov, D. M., Butler, A. J., Kadah, Y. M., Epstein, C. M., &
Hu, X. P. (2005). Functional magnetic resonance imaging
and transcranial magnetic stimulation: Effects of motor
imagery, movement and coil orientation. Clinical
Neurophysiology, 116, 1601–1610.
Noonan, K. A., Jefferies, E., Corbett, F., & Lambon Ralph,
M. A. (2010). Elucidating the nature of deregulated
semantic cognition in semantic aphasia: Evidence for
the roles of prefrontal and temporo-parietal cortices.
Journal of Cognitive Neuroscience, 22, 1597–1613.
Noppeney, U., Phillips, J., & Price, C. (2004). The neural
areas that control the retrieval and selection of semantics.
Neuropsychologia, 42, 1269–1280.
Novick, J. M., Kan, I. P., Trueswell, J. C., & Thompson-Schill,
S. L. (2009). A case for conflict across multiple domains:
Memory and language impairments following damage to
ventrolateral prefrontal cortex. Cognitive Neuropsychology,
26, 527–567.
Owen, A. M., Schneider, W. X., & Duncan, J. (2000). Executive
control and the frontal lobe: Current issues. Experimental
Brain Research, 133, 1–2.
146
Journal of Cognitive Neuroscience
Volume 24, Number 1
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
4
/
1
2
4
1
/
3
1
3
/
1
1
9
3
4
3
3
/
4
1
2
7
9
8
o
0
c
2
n
6
_
9
a
/
_
j
0
o
0
c
1
n
2
3
_
a
p
_
d
0
0
b
1
y
2
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
/
f
.
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Pascual-Leone, A., Tormos, J. M., Keenan, J., Tarazona, F.,
Canete, C., & Catala, M. D. (1998). Study and modulation
of human cortical excitability with transcranial magnetic
stimulation. Journal of Clinical Neurophysiology, 15,
333–343.
Pascual-Leone, A., Walsh, V., & Rothwell, J. (2000). Transcranial
magnetic stimulation in cognitive neuroscience—Virtual
lesion, chronometry, and functional connectivity. Current
Opinion in Neurobiology, 10, 232–237.
Patterson, K., Nestor, P. J., & Rogers, T. T. (2007). Where
do you know what you know? The representation of
semantic knowledge in the human brain. Nature Reviews
Neuroscience, 8, 976–987.
Pobric, G., Jefferies, E., & Lambon Ralph, M. A. (2007). Anterior
temporal lobes mediate semantic representation: Mimicking
semantic dementia by using rTMS in normal participants.
Proceedings of the National Academy of Sciences, U.S.A.,
104, 20137–20141.
Pobric, G., Jefferies, E., & Lambon Ralph, M. A. (2010).
Amodal semantic representations depend on both anterior
temporal lobes: Evidence from repetitive transcranial
magnetic stimulation. Neuropsychologia, 48, 1336–1342.
Pobric, G., Lambon Ralph, M. A., & Jefferies, E. (2009). The
role of the anterior temporal lobes in the comprehension
of concrete and abstract words: rTMS evidence. Cortex,
45, 1104–1110.
Postman, L., & Keppel, G. (1970). Norms of word association.
New York: Academic Press.
Rossi, S., Hallett, M., Rossini, P. M., & Pascual-Leone, A. (2009).
Safety, ethical considerations, and application guidelines for
the use of transcranial magnetic stimulation in clinical practice
and research. Clinical Neurophysiology, 120, 2008–2039.
Saur, D., Kreher, B. W., Schnell, S., Kummerer, D., Kellmeyer,
P., Vry, M. S., et al. (2008). Ventral and dorsal pathways
for language. Proceedings of the National Academy of
Sciences, U.S.A., 105, 18035–18040.
Schwartz, M. F., Kimberg, D. Y., Walker, G. M., Faseyitan, O.,
Brecher, A., Dell, G. S., et al. (2009). Anterior temporal
involvement in semantic word retrieval: Voxel-based
lesion-symptom mapping evidence from aphasia. Brain,
132, 3411–3427.
Snijders, T. M., Vosse, T., Kempen, G., Van Berkum, J. J.,
Petersson, K. M., & Hagoort, P. (2009). Retrieval and
unification of syntactic structure in sentence comprehension:
An fMRI study using word-category ambiguity. Cerebral
Cortex, 19, 1493–1503.
Snyder, H. R., Feigenson, K., & Thompson-Schill, S. L. (2007).
Prefrontal cortical response to conflict during semantic and
phonological tasks. Journal of Cognitive Neuroscience,
19, 761–775.
Soni, M., Lambon Ralph, M. A., Noonan, K., Ehsan, S.,
Hodgson, C., & Woollams, A. M. (2009). “L” is for tiger:
Effects of phonological (mis)cueing on picture naming in
semantic aphasia. Journal of Neurolinguistics, 22, 538–547.
Thompson-Schill, S. L., DʼEsposito, M., Aguirre, G. K., &
Farah, M. J. (1997). Role of left inferior prefrontal cortex
in retrieval of semantic knowledge: A reevaluation.
Proceedings of the National Academy of Sciences,
U.S.A., 94, 14792–14797.
Thompson-Schill, S. L., Swick, D., Farah, M. J., DʼEsposito, M.,
Kan, I. P., & Knight, R. T. (1998). Verb generation in patients
with focal frontal lesions: A neuropsychological test of
neuroimaging findings. Proceedings of the National
Academy of Sciences, U.S.A., 95, 15855–15860.
Wager, T. D., Jonides, J., & Reading, S. (2004). Neuroimaging
studies of shifting attention: A meta-analysis. Neuroimage,
22, 1679–1693.
Wagner, A. D., Pare-Blagoev, E. J., Clark, J., & Poldrack, R. A.
(2001). Recovering meaning: Left prefrontal cortex guides
controlled semantic retrieval. Neuron, 31, 329–338.
Wallentin, M., Nielsen, A. H., Vuust, P., Dohn, A., Roepstorff, A.,
& Lund, T. E. (2011). BOLD response to motion verbs in
left posterior temporal gyrus during story comprehension.
Brain and Language, 119, 222–225.
Walsh, V., & Cowey, A. (2000). Transcranial magnetic stimulation
and cognitive neuroscience. Natures Review Neuroscience,
1, 73–79.
Walsh, V., & Rushworth, M. (1999). A primer of magnetic
stimulation as a tool for neuropsychology. Neuropsychologia,
37, 125–135.
Wassermann, E. M. (1998). Risk and safety of repetitive
transcranial magnetic stimulation: Report and suggested
guidelines from the International Workshop on the
Safety of Repetitive Transcranial Magnetic Stimulation,
June 5–7, 1996. Electroencephalography and Clinical
Neurophysiology, 108, 1–16.
Wassermann, E. M., Blaxton, T. A., Hoffman, E. A., Berry,
C. D., Oletsky, H., Pascual-Leone, A., et al. (1999).
Repetitive transcranial magnetic stimulation of the
dominant hemisphere can disrupt visual naming in
temporal lobe epilepsy patients. Neuropsychologia, 37,
537–544.
Whitney, C., Jefferies, E., & Kircher, T. (2011). Heterogeneity
of the left temporal lobe in semantic representation and
control: Priming multiple versus single meanings of
ambiguous words. Cerebral Cortex, 21, 831–844.
Zempleni, M. Z., Renken, R., Hoeks, J. C., Hoogduin, J. M.,
& Stowe, L. A. (2007). Semantic ambiguity processing
in sentence context: Evidence from event-related fMRI.
Neuroimage, 34, 1270–1279.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
4
/
1
2
4
1
/
3
1
3
/
1
1
9
3
4
3
3
/
4
1
2
7
9
8
o
0
c
2
n
6
_
9
a
/
_
j
0
o
0
c
1
n
2
3
_
a
p
_
d
0
0
b
1
y
2
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
.
t
f
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Whitney et al.
147