Enhanced Alpha-oscillations in Visual Cortex during
Anticipation of Self-generated Visual Stimulation
Max-Philipp Stenner1,2, Markus Bauer1, Patrick Haggard1,
Hans-Jochen Heinze2, and Ray Dolan1
Abstract
■ The perceived intensity of sensory stimuli is reduced when
these stimuli are caused by the observerʼs actions. This phe-
nomenon is traditionally explained by forward models of sen-
sory action–outcome, which arise from motor processing.
Although these forward models critically predict anticipatory
modulation of sensory neural processing, neurophysiological
evidence for anticipatory modulation is sparse and has not been
linked to perceptual data showing sensory attenuation. By
combining a psychophysical task involving contrast discrimina-
tion with source-level time–frequency analysis of MEG data, we
demonstrate that the amplitude of alpha-oscillations in visual
cortex is enhanced before the onset of a visual stimulus when
the identity and onset of the stimulus are controlled by partic-
ipantsʼ motor actions. Critically, this prestimulus enhancement
of alpha-amplitude is paralleled by psychophysical judgments of
a reduced contrast for this stimulus. We suggest that alpha-
oscillations in visual cortex preceding self-generated visual
stimulation are a likely neurophysiological signature of motor-
induced sensory anticipation and mediate sensory attenuation.
We discuss our results in relation to proposals that attribute
generic inhibitory functions to alpha-oscillations in prioritizing
and gating sensory information via top–down control. ■
INTRODUCTION
Immediate consequences of voluntary actions are subject
to sensory attenuation. A prominent example is the per-
ceived intensity of self-applied tactile stimuli, which is
reduced when compared with identical, externally gener-
ated stimuli (Bays, Wolpert, & Flanagan, 2005). Similarly,
a reduction of perceived loudness and visual contrast have
been demonstrated for self-generated auditory (Weiss,
Herwig, & Schütz-Bosbach, 2011) and visual (Cardoso-
Leite, Mamassian, Schütz-Bosbach, & Waszak, 2010) stim-
uli, respectively.
Across modalities, sensory attenuation has been con-
sidered within theoretical frameworks that link motor
processing to perception of sensory action consequences
(Waszak, Cardoso-Leite, & Hughes, 2012; Schütz-Bosbach
& Prinz, 2007; Wolpert, Ghahramani, & Jordan, 1995).
Here, sensory attenuation is often explained by consis-
tency between forward models of anticipated action
effects (corollary discharge; Sperry, 1950) and actual sen-
sory feedback. Neural processes that functionally cor-
respond to corollary discharge have been reported based
on invasive recordings in animals (Poulet & Hedwig,
2007; Sommer & Wurtz, 2006). Equivalent evidence in
humans is sparse, predominantly indirect (i.e., speaking
in favor of motor prediction pathways upstream of sensory
areas), and rarely linked to sensory attenuation (Ostendorf,
Kilias, & Ploner, 2012; Kühn, Seurinck, Fias, & Waszak,
1University College London, 2University of Magdeburg
2010; Voss, Ingram, Haggard, & Wolpert, 2009; Haggard
& Whitford, 2004).
Because of its presumed motor origin, corollary dis-
charge is regarded as physiologically distinct from sen-
sory predictions that are purely based on external
events. Because corollary discharge is thought to reflect
motor forward modeling, it implies anticipated control
over a stimulus. Corollary discharge signals should there-
fore vary with the extent to which an action is expected
to control a subsequent stimulus and should not simply
reflect predictability of this stimulus from other sensory
events. However, in a recent review, Hughes, Desantis,
and Waszak (2012) concluded that the majority of studies
on sensory attenuation potentially confound stimulus
control and stimulus predictability. Because stimulus pre-
dictability is well known to modulate sensory processing
(e.g., Alink, Schwiedrzik, Kohler, Singer, & Muckli, 2010),
this confound can lead to misinterpretations. Other stud-
ies of sensory attenuation are based on a task design that
introduces another potential confounding factor. These
studies typically compare stimuli that are contingent on
an action with identical stimuli that are not preceded by a
motor response (e.g., Weiss et al., 2011; see Hughes
et al., 2012, for a review). Such a comparison introduces
a difference in motor output in addition to differences in
control, which complicates interpretation of any observed
effects as specific to control.
Here, we used a task that varied the extent to which an
action controlled a subsequent stimulus while keeping
both stimulus predictability and motor output comparable
© 2014 Massachusettes Institute of Technology Published under a
Creative Commons Attribution 3.0 Unported (CC BY 3.0) license
Journal of Cognitive Neuroscience 26:11, pp. 2540–2551
doi:10.1162/jocn_a_00658
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
6
/
1
2
1
6
/
2
1
5
1
4
/
0
2
1
5
9
4
4
0
7
/
8
1
6
4
7
8
o
2
c
0
n
7
_
2
a
/
_
j
0
o
0
c
6
n
5
8
_
a
p
_
d
0
0
b
6
y
5
g
8
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
.
/
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
across conditions. We combined MEG and a visual con-
trast discrimination task to test for an anticipatory, that
is, prestimulus, modulation of sensory processing in
relation to perceptual evidence of sensory attenuation.
The majority of previous MEG/EEG studies on sensory
attenuation in humans have restricted their analyses to
specific stimulus-evoked responses (Gentsch & Schütz-
Bosbach, 2011; Hughes & Waszak, 2011; Hesse, Nishitani,
Fink, Jousmäki, & Hari, 2010; Baess, Widmann, Roye, Schröger,
& Jacobsen, 2009; Martikainen, Kaneko, & Hari, 2005).
Here, we focus on neural oscillations in a prestimulus time
interval, motivated by the finding that induced responses
convey a corollary discharge signal in speech (Chen et al.,
2011). Accumulating evidence points to alpha-oscillations
in visual cortex as a representation of pulsed inhibition
by feedback projections that prioritize the processing of
task-relevant (Jensen & Mazaheri, 2010) and salient stimuli
( Jensen, Bonnefond, & VanRullen, 2012). Closely related
functions have been proposed for sensory attenuation,
namely the prevention of self-induced desensitization
and the prioritizing of externally caused stimuli (Lally &
Frendo, 2011; Poulet & Hedwig, 2007). On this basis, we
hypothesized that anticipation of a visual stimulus that is
controlled by oneʼs action would be reflected in an increase
in alpha-amplitude in visual cortex and predict sensory
attenuation at the perceptual level.
METHODS
Participants
Ten healthy volunteers participated in the study (mean
age = 24.8 years, SD = 4.7 years, six women). All par-
ticipants were recruited via an on-line database. They
gave written informed consent before participation with
the right to exit the study at any time. The study was
approved by the local ethics committee (University College
London, UK). Participants received £10 per hour as
reimbursement.
Task and Experimental Procedure
The task was designed to vary participantsʼ control over
the onset and orientation of a visual stimulus while mini-
mizing differences in motor output and stimulus pre-
dictability across conditions. There were three blocked
conditions: a “motor/control” condition, in which par-
ticipants controlled the onset and orientation of a static
Gabor patch on a computer screen with their button
presses; a “motor/no control” condition, in which par-
ticipantsʼ button presses had no influence on the onset
and orientation of this Gabor patch; and a “no motor”
condition, in which the Gabor patch was presented with-
out a preceding button press. The “motor/no control”
condition served to avoid a potential confound of control
with motor output present in many previous studies. In
addition, cues that contained information about the orien-
tation and timing of the Gabor patch in this condition
controlled for stimulus predictability. The “no motor”
condition was included for comparability with previous
studies in which conditions without motor output served
as a baseline (typically labeled “effect only” conditions in
these studies).
All conditions included a contrast discrimination task
to test for psychophysical evidence of sensory attenua-
tion. For this discrimination task, each Gabor patch was
followed by a second Gabor patch, which was either of
higher, lower, or identical contrast relative to the first.
Participants had to indicate whether the first or the sec-
ond of the two stimuli was the one of higher contrast by
pressing one of two buttons. Sensory attenuation is
known to decrease over the course of a few hundred milli-
seconds (Aliu, Houde, & Nagarajan, 2009; Bays et al.,
2005). Accordingly, we expected sensory attenuation to
affect the first of the two Gabor patches, presented about
50 msec after the button press, more strongly than the
second, which followed the first after a SOA between
1000 and 1250 msec. Therefore, we could predict that
sensory attenuation would become manifest as a stronger
bias to report the first stimulus as the low-contrast one,
similar to previous studies that reported shifts in the point
of subjective equality (PSE) as a measure of sensory atten-
uation (Desantis, Weiss, Schütz-Bosbach, & Waszak, 2012;
Weiss et al., 2011; Haggard & Whitford, 2004).
We used three constant stimulus levels, each presented
in one third of trials in each of the three conditions. The
contrast of the first Gabor was held constant at 0.25
Michelson contrast to keep it predictable, and only the
contrast of the second Gabor was varied from trial to trial.
Contrast levels for trials in which the second Gabor was of
higher or lower contrast than the first were individually
determined by a staircase procedure before the main
task.1 Participants were informed that the two Gabor
patches presented per trial were always of different con-
trasts. Responses in the contrast discrimination task were
given by pressing one of two buttons, operated with the
left middle and index fingers. There was no time limit
for these responses. Gabor patches (spatial frequency =
0.8 cycles/degree, phase = +90°, size (SD) = 1.25°) were
presented in the lower left visual field (at −3.0°/−3.0°) on
a gray background for 250 msec. This eccentric stimulus
presentation was chosen for two reasons: First, MEG is
less sensitive to signals evoked by foveally presented
visual stimuli versus stimuli above or below fixation
(due to the orientation and functional architecture of
the calcarine sulcus). And second, stimulus presentation
in the left visual hemifield helped to spatially dissociate
motor-related signals (in sensorimotor areas of the left
hemisphere; relevant responses were given with the
right hand) and consequence-related signals (in visual
areas of the right hemisphere). The paradigm, including
all stimuli, was programmed using Presentation software
(Neurobehavioral Systems, www.neurobs.com).
Stenner et al.
2541
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
6
/
1
2
1
6
/
2
1
5
1
4
/
0
2
1
5
9
4
4
0
7
/
8
1
6
4
7
8
o
2
c
0
n
7
_
2
a
/
_
j
0
o
0
c
6
n
5
8
_
a
p
_
d
0
0
b
6
y
5
g
8
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
/
.
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
“Motor/Control” Condition
In the “motor/control” condition (Figure 1A, top) partic-
ipants pressed a button to trigger the sequential presen-
tation of the two Gabor patches on the screen. The onset
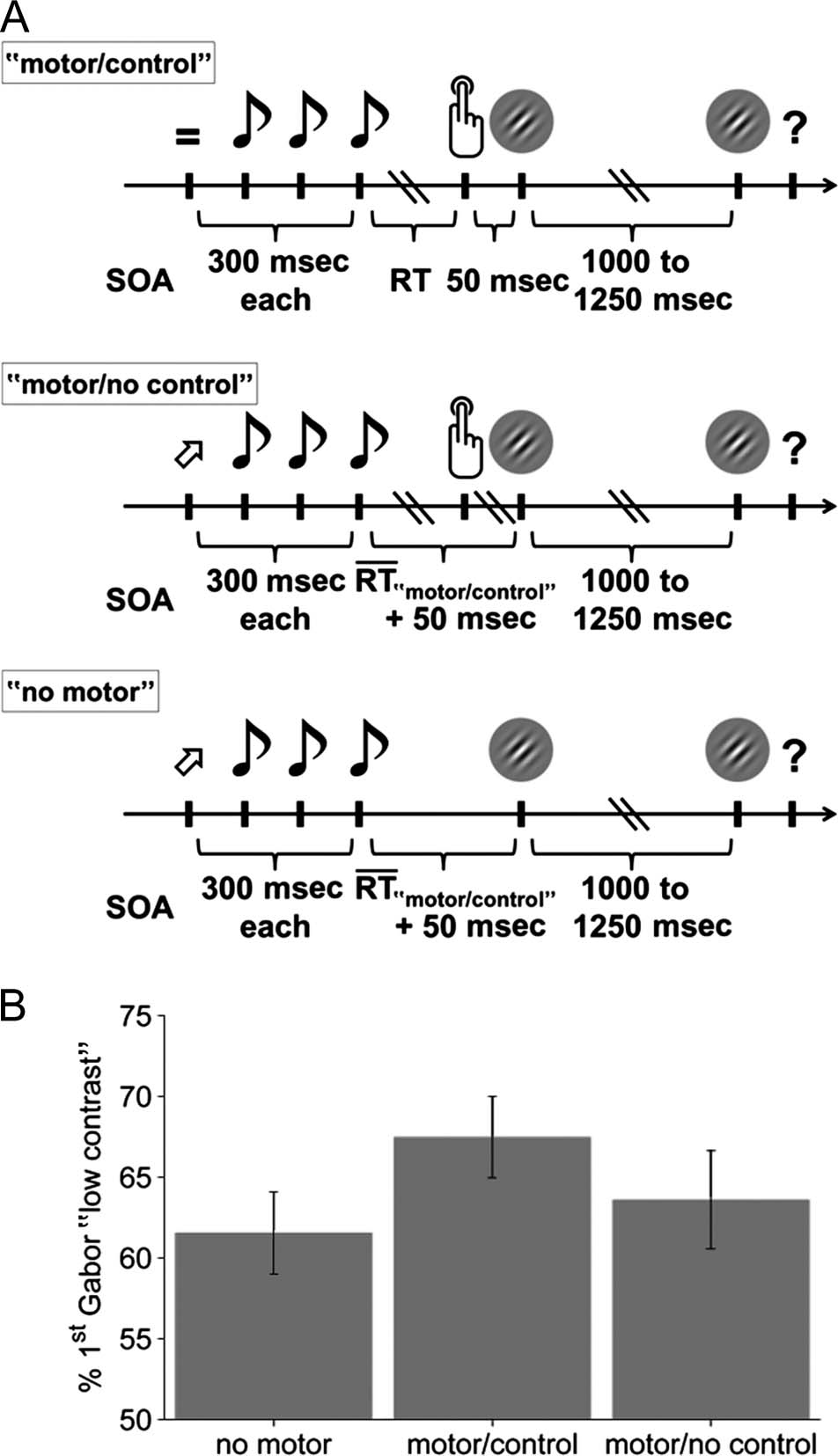
Figure 1. Task and behavioral results. (A) Timing of events in “motor/
control” (top), “motor/no control” (middle), and “no motor” (bottom)
conditions. Each trial started with the presentation of a visual cue,
which signaled the button/orientation mapping in the “motor/control”
condition and the orientation of upcoming Gabor patches in “motor/
no control” and “no motor” conditions. This visual cue was followed by
an auditory cue, which signaled the start of the response window in
“motor/control” and “motor/no control” conditions and predicted the
onset time of the first Gabor in “motor/no control” and “no motor”
conditions. Two Gabor patches were presented sequentially in each
trial. Participants had to indicate which of the two was of higher
contrast at the end of each trial (first or second Gabor patch). The
crucial experimental manipulation was a variation of the extent to which
participantsʼ button presses determined the onset and orientation of
the first Gabor patch. Note that trial events and average timing of these
events was balanced across “motor/control” and “motor/no control”
conditions. (B) Percentage of trials in which the first Gabor patch
was reported as the lower-contrast stimulus in the three conditions
(mean ± SEM ).
time and orientation of the first Gabor and the orienta-
tion of the second Gabor were fully determined by this
button press. Participants were required to perform these
button presses as follows.
On each trial, participants chose between two alter-
native buttons, operated by their right index and middle
fingers. They were instructed to choose the two buttons
approximately equally often across each block (of 26
trials). They received feedback on the relative fre-
quencies of their choices at the end of each block. On
each trial, this choice determined the orientation of the
two subsequently presented Gabor patches, which could
be 45° clockwise or 45° counterclockwise. The mapping
of the two buttons onto the two orientation varied from
trial to trial, so that, for example, pressing the index finger
button led to Gabor patches oriented 45° clockwise in
some trials and to stimuli of the opposite orientation in
others. On any given trial, the current valid mapping was
signaled by a visual cue from the beginning of each trial
onwards (presented centrally, visual angle 1°, white). This
cue could either be an equal sign, meaning that the index
finger button would lead to Gabor patches oriented
45° counterclockwise and the middle finger button to
Gabor patches oriented 45° clockwise, or a bidirectional
arrow, which signaled the opposite mapping. Our motiva-
tion to avoid a constant mapping was to keep consistency
of the mapping across trials similarly low as in the “motor/
no control” condition, where this mapping was incidental
(see “motor/no control” condition below). Note that, be-
cause of the visual cue, participants could fully predict
the orientation of Gabor patches from their button choice
on each trial. Participants were familiarized with both
mappings and the meaning of the visual cues in a practice
session before the main experiment.2
In addition to controlling stimulus orientation, partici-
pants also determined the onset time of the first Gabor,
which was programmed to be 50 msec after the button
press (measured time between button press and Gabor
onset: 59 ± 5 msec [mean ± SD]; this slight deviation
from the programmed value results from the jitter of re-
sponse timing with respect to vertical refreshes of the
screen [at 60 Hz]). To be able to achieve, on average,
comparable timings of trial events in the “motor/control”
condition as in the “motor/no control” condition (where
Gabor onset was independent of RT, see below), par-
ticipants had to press the button that triggered Gabor
presentation within a short time window. This time win-
dow was signaled by an auditory cue, which consisted of
three consecutive sine wave tones (700 Hz, 700 Hz, and
1000 Hz; duration = 75 msec; SOA = 300 msec, bin-
aurally presented over headphones; the first tone was
played 300 msec after the onset of the visual cue). Par-
ticipants had to press a button within 500 msec after
the end of the third tone of this auditory cue in order
for the Gabor patches to be presented. Earlier or later
button presses produced an error sound and no Gabor
presentation.
2542
Journal of Cognitive Neuroscience
Volume 26, Number 11
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
6
/
1
2
1
6
/
2
1
5
1
4
/
0
2
1
5
9
4
4
0
7
/
8
1
6
4
7
8
o
2
c
0
n
7
_
2
a
/
_
j
0
o
0
c
6
n
5
8
_
a
p
_
d
0
0
b
6
y
5
g
8
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
f
t
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Lastly, to emphasize the dependency of stimulus pre-
sentation on button presses, we introduced infrequent
no-go trials, which were signaled by a color change of
the fixation cross from white to red at the beginning of
the trial (replacing the visual cue present in all other
trials). No Gabor and no error sound were presented
upon successfully withholding any right-hand button
press in these trials. There were two no-go trials in each
block (of 26 trials).
In summary, participants could control and predict the
presentation, timing, and orientation of Gabor patches by
their button presses in the “motor/control” condition.
“Motor/No Control” Condition
Similar to the “motor/control” condition, the “motor/no
control” condition (Figure 1A, middle) required partici-
pants to press one of the two right-hand buttons follow-
ing the auditory cue. However, RT and choice of button
had no influence on the timing and orientation of the
first Gabor patch, as its orientation was predetermined
pseudorandomly and its onset was time-locked to the
auditory cue.
Nevertheless, both Gabor orientation and onset were
predictable from the visual and auditory cues in this
condition: Orientation was cued from the beginning
of each trial onwards by arrows pointing “northwest”
(for Gabor orientation of 45° counterclockwise) or
“northeast” (for Gabor orientation of 45° clockwise)
(presented centrally, visual angle 1°, white).3 Gabor onset
followed the last tone of the auditory cue after the mean RT
in the preceding “motor/control” block plus 50 msec (the
programmed action–outcome delay in “motor/control”
blocks).
Participants were explicitly instructed to choose button
presses independently of orientation cues and succeeded
in doing so (button choices corresponded to the cued
direction in 49.8% of trials across participants, t(9) = .4,
p = .68 [dependent samples t test on the number of trials
in which button choice corresponded to the cued vs. the
opposite orientation]). Similar to the “motor/control”
condition, participants were also instructed to press the
left and right buttons approximately equally often across
each block. They received feedback on the relative fre-
quencies of these button choices at the end of each
block.
As in the “motor/control” condition, “motor/no control”
blocks also contained infrequent no-go trials, in which
Gabor patches were presented irrespective of whether
participants withheld their response or not to further
reduced the action–stimulus contingency.
Thus, our design ensured that “motor/control” and
“motor/no control” conditions were balanced for motor
output, visual input, and, at least to some extent, pre-
dictability of stimulus onset and orientation. They differed
primarily in participantsʼ control over stimulus onset and
orientation.
“No Motor” Condition
For comparability with previous studies and as a localizer
condition for our MEG analysis, we included “no motor”
blocks (Figure 1A, bottom). These were identical to
“motor/no control” blocks, including the information
carried by the visual and auditory cues with respect to
Gabor orientation and onset, with the exception that
participants were instructed to omit right-hand responses
altogether throughout the entire blocks.
Experimental Procedure
All three parts of the experiment (the staircase, the prac-
tice, and the main task) were completed twice by each
participant, on two consecutive days, to increase power
of the behavioral data. MEG data were acquired on the
second day only. In the main task, blocks of the “motor/
control” and “motor/no control” conditions alternated
(starting with a “motor/control” block). There was one
“no motor” block every four blocks. In total, participants
completed 30 blocks on the first day (12 “motor/control,”
12 “motor/no control,” 6 “no motor”) and 20 blocks on the
second (8 “motor/control,” 8 “motor/no control,” 4 “no motor”).
Each block consisted of 26 trials and lasted approximately
2–3 min.
MEG Recording and Analysis
MEG data were recorded continuously from 274 axial
gradiometers (one SQUID was defunct) and 35 reference
channels of a CTF Omega system, Port Coquitlam, BC,
Canada at a sampling frequency of 1200 Hz. Head posi-
tion was measured via three coils at the nasion and the
tragi. Analysis used FieldTrip (http://fieldtrip.fcdonders.nl/;
Oostenveld, Fries, Maris, & Schoffelen, 2011) and SPM8
(www.fil.ion.ucl.ac.uk/spm; Litvak et al., 2011). Procedures
for preprocessing and artifact treatment were similar to
previous work (Bauer, Kennett, & Driver, 2012; Bauer,
Kluge, et al., 2012). In short, MEG data were epoched
and inspected visually for amplitude jumps, muscle arti-
facts, and eye blink artifacts using standard options in
FieldTrip (“ft_rejectvisual”) based on a threshold for the
maximum amplitude variance across channels in each trial.
Remaining artifacts arising from eye movements were
removed using PCA. Artifact rejection was done blind to
condition. Line noise was removed from 5-sec periods
around each trial using a narrow-band notch filter.
For sensor-level analysis, the artifact-free data were inter-
polated to a common sensor array template across partici-
pants to correct for interindividual variations in head
positions. This spatial interpolation method, based on a
minimum-norm projection of the data as an intermediate
step, is a standard function in FieldTrip (“ft_megrealign”)
described in detail in previous work (Bauer, Kennett,
et al., 2012). Planar gradients were then calculated using
a nearest-neighbor method (Bastiaansen & Knösche,
Stenner et al.
2543
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
6
/
1
2
1
6
/
2
1
5
1
4
/
0
2
1
5
9
4
4
0
7
/
8
1
6
4
7
8
o
2
c
0
n
7
_
2
a
/
_
j
0
o
0
c
6
n
5
8
_
a
p
_
d
0
0
b
6
y
5
g
8
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
/
t
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
2000) to better estimate the spatial distribution of cortical
generators of the MEG field at the sensor level (Bauer,
Kennett, et al., 2012).
Source reconstruction was based on individual T1
weighted MRI warped to the standard MNI brain. The
first steps of forward modeling, including coregistration
of MEG data and individual MRI, segmentation, calcu-
lation of a cortical source model with 8196 grid points
(down-sampled to 823 grid points using reducepath.m
in MatLab after computation of the lead fields), and com-
putation of a single-shell volume conduction model, were
performed using SPM8 (Litvak et al., 2011). These source-
and volume-conduction models were entered into the
lead field computation in FieldTrip. We used linearly con-
strained minimum variance beamforming to project the
sensor data onto the cortical grid, based on the covari-
ance matrix of the low-pass filtered data (<40 Hz), as
described in detail previous work (Bauer, Kluge, et al.,
2012). These filters were identical across conditions. For
filter computation, orientation was fixed to the direction
of spatial component with largest projected power
separately for each grid point (a standard option Field-
Trip). The regularization parameter λ set 0 (the
default).
Spectral analysis of time courses at sensor level
and source level channel and were
performed between 2.5 40 Hz steps Hz. A
Fourier-transformed Hanning taper 400-msec window
length multiplied Fourier-transformed data
segments sampled every 100 msec 500 msec
before after stimulus onset. For sensor-level data,
vertical horizontal components planar gradient
were combined spectral analysis. Power cal-
culated as square magnitude complex-
valued Fourier spectra. low-frequency analyses, data
were analyzed separately before removal of
phase-locked (“evoked”) responses from spectrum
(we show data phase-locked compo-
nents throughout Results section). Phase-locked re-
sponses removed complex-valued Fourier
spectra computation power by subtracting the
average spectra trials single-trial
data individual, described work
(Bauer, al., 2012).
Statistical sensor- source-level power
was based on a nonparametric randomization test (Maris
& Oostenveld, 2007) implemented FieldTrip, which
corrects multiple comparisons channels> .2). Sensitivity (d
cantly between conditions (one-way repeated-measures
0 in
ANOVA, factor Condition: F(2, 18) = .74, p > .4; d
the three conditions, mean ± SD: 1.4 ± 0.58 [“no motor”],
1.34 ± 0.42 [“motor/control”], 1.47 ± 0.42 [“motor/no
control”]).
In summary, our behavioral results are consistent with
sensory attenuation of the first Gabor patch in the “motor/
control” conditions when compared with the “no motor”
condition as well as to the “motor/no control” conditions.
When participants controlled stimulus onset and orien-
tation, they were more likely to report the first Gabor
patch as the lower-contrast stimulus.
MEG
Here, we focused on alpha-power (7.5–12.5 Hz) in a time
window between 500 msec before and 500 msec after
Gabor onset. We focused on differences between the
“motor/control” condition and the “motor/no control”
condition, as these were balanced for motor output (as
well as stimulus predictability). We predicted enhanced
alpha-power over right occipital cortex in a prestimulus
time window in the “motor/control” condition as com-
pared with the “motor/no control.” Responses that were
phase-locked to stimulus onset (“evoked”) did not con-
tribute to the low-frequency spectra reported here as
they were removed for both sensor- and source-level
analyses (see Methods for details). Virtually the same
results were obtained when looking at the total power
without removal of transient phase-locked responses.
Sensor-level Analysis
Sensory attenuation is known to decrease over the course
of a few hundred milliseconds after a motor action (Bays
et al., 2005). Therefore, we first examined differential
effects of stimulus control on the power spectral densities
of the two stimuli (i.e., on the difference power(first Gabor) −
power(second Gabor)), effectively testing for a “control” ×
“Gabor” interaction. We used this approach to determine
channels that showed a significant differential modulation
by “control” for the two stimuli, equivalent to the effect of
stimulus control on our perceptual metric of sensory atten-
uation in the two alternative forced-choice task.
When testing for this interaction, correcting for multiple
comparisons across all channels and time bins between
500 msec before and after stimulus onset, we found a
significant positive cluster between 7.5 and 12.5 Hz during
the 300 msec before stimulus onset in right occipital and
parietal sensors (contralateral to the visual hemifield in
which the Gabor patches were presented; p = .044, cluster-
based correction for multiple comparisons across all
channels and across all time bins between −500 msec and
+500 msec; Figure 2A).5
This effect of stimulus control on the power difference
was predominantly due to higher alpha-band power
in the “motor/control” condition compared with the
“motor/no control” condition in the prestimulus period
corresponding to the first Gabor patch as opposed to
the second: Power between 7.5 and 12.5 Hz was sig-
nificantly higher in the “motor/control” condition than
in the “motor/no control” conditions in right occipital
and parietal sensors during the 300 msec before onset
of the first Gabor patch ( p = .034, cluster-based correc-
tion for multiple comparisons across all channels; power
spectrum in Figure 2B, top, averaged across the channels
highlighted in Figure 2A, top). The same contrast for the
prestimulus period of the second Gabor patch did not
yield any significant cluster (power spectrum in Figure 2B,
bottom, averaged across the channels highlighted in
Figure 2A, top). When comparing the “motor/control”
condition and the “no motor” condition, we found a
very similar cluster of right occipital sensors, which
showed significantly higher alpha-power (7.5–12.5 Hz)
up to 400 msec before the onset of the first Gabor patch
( p = .036, corrected for multiple comparisons across all
sensors). We found no significant difference between the
“motor/no control” condition and the “no motor” condition
Stenner et al.
2545
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
6
/
1
2
1
6
/
2
1
5
1
4
/
0
2
1
5
9
4
4
0
7
/
8
1
6
4
7
8
o
2
c
0
n
7
_
2
a
/
_
j
0
o
0
c
6
n
5
8
_
a
p
_
d
0
0
b
6
y
5
g
8
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
/
t
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
6
/
1
2
1
6
/
2
1
5
1
4
/
0
2
1
5
9
4
4
0
7
/
8
1
6
4
7
8
o
2
c
0
n
7
_
2
a
/
_
j
0
o
0
c
6
n
5
8
_
a
p
_
d
0
0
b
6
y
5
g
8
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
.
t
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
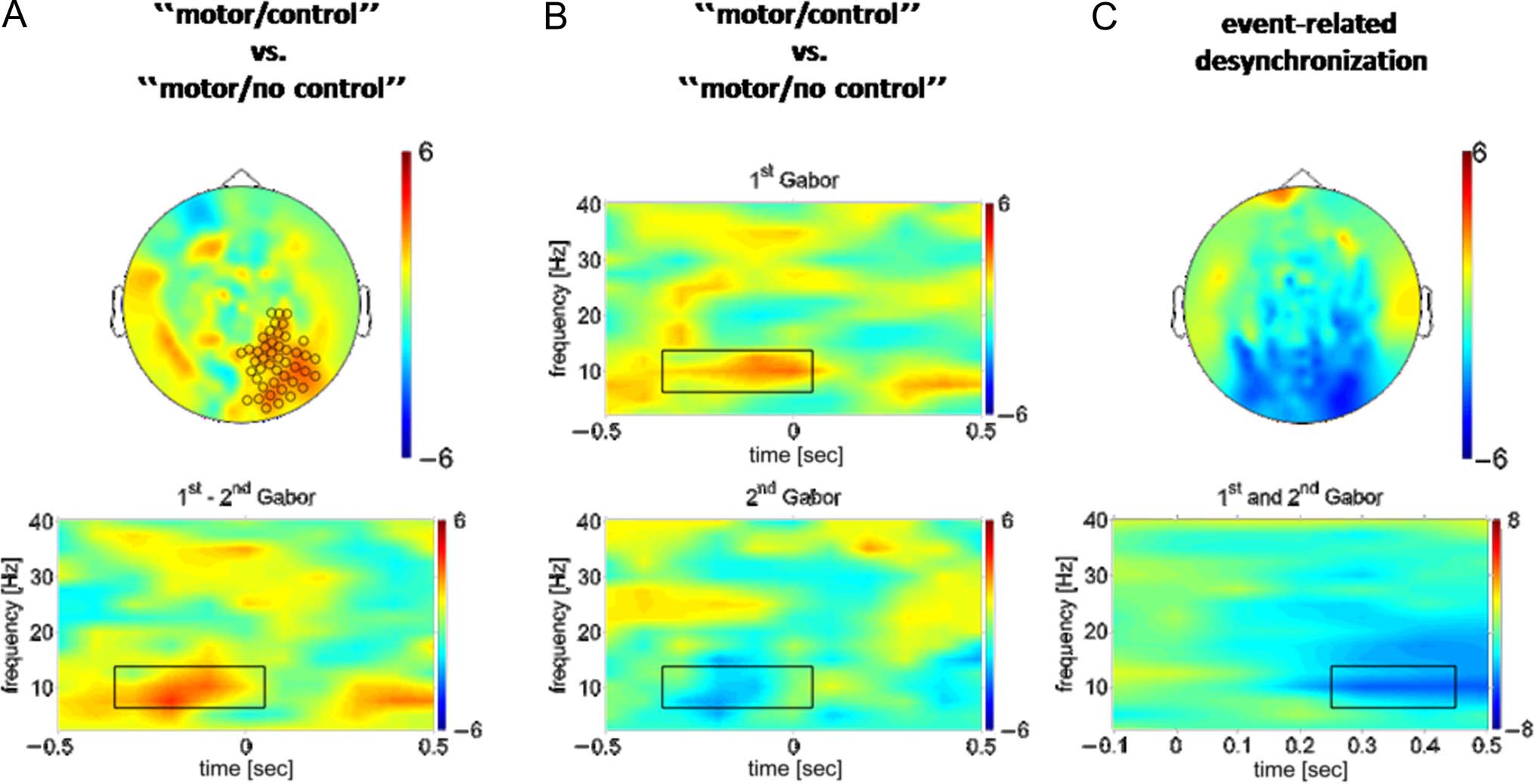
Figure 2. Effects of stimulus control and of Gabor patch presentation on peristimulus alpha-oscillations at the sensor level. Color codes for
t values in all panels. (A) Sensor-level planar gradient topography (top) and time–frequency representation (bottom) of the effect of stimulus
control (“motor/control” vs. “motor/no control”) on the difference in alpha-power (7.5–12.5 Hz) between the first and the second Gabor patch
(first − second). Markers in the sensor topography indicate channels that showed significantly higher alpha-power in the “motor/control” condition
than in the “motor/no control” condition within the prestimulus time–frequency window depicted in the bottom panel (box). Data were
averaged across these channels for the time–frequency representation. (B) Time–frequency representations of the effect of stimulus control
(“motor/control” vs. “motor/no control”) on the peristimulus low-frequency power spectrum, averaged across the channels highlighted in (A),
top, around the onset of the first (top) and second (bottom) Gabor patch. (C) Topography and time–frequency representation of the effect of
Gabor patch presentation on alpha-power (event-related alpha-desynchronization). The topography (top) is based on power averaged across the
window depicted in the time–frequency representation (bottom), which in turn shows t values for power averaged across significant channels.
in this time and frequency window (smallest cluster-level
p value: p = .28).
The topography of this prestimulus cluster in the alpha-
band (Figure 2A, top) overlapped with the spatial dis-
tribution of the well-known stimulus-induced response
in the same frequency band (Figure 2C, t values, post-
stimulus [300–400 msec] vs. baseline [−200 msec] across
both Gabor patches for the “no motor” condition as a
localizer). Note that the reconstructed planar gradient
used here represents local cortical activity underneath
corresponding sensors (Bastiaansen & Knösche, 2000).
The prestimulus modulation of alpha-band power by
stimulus control thus involved similar cortical regions as
the early event-related alpha-desynchronization, which
is one of the strongest neural signatures of afferent
stimulation of sensory cortex (Pfurtscheller & Aranibar,
1977).
Source-level Analysis
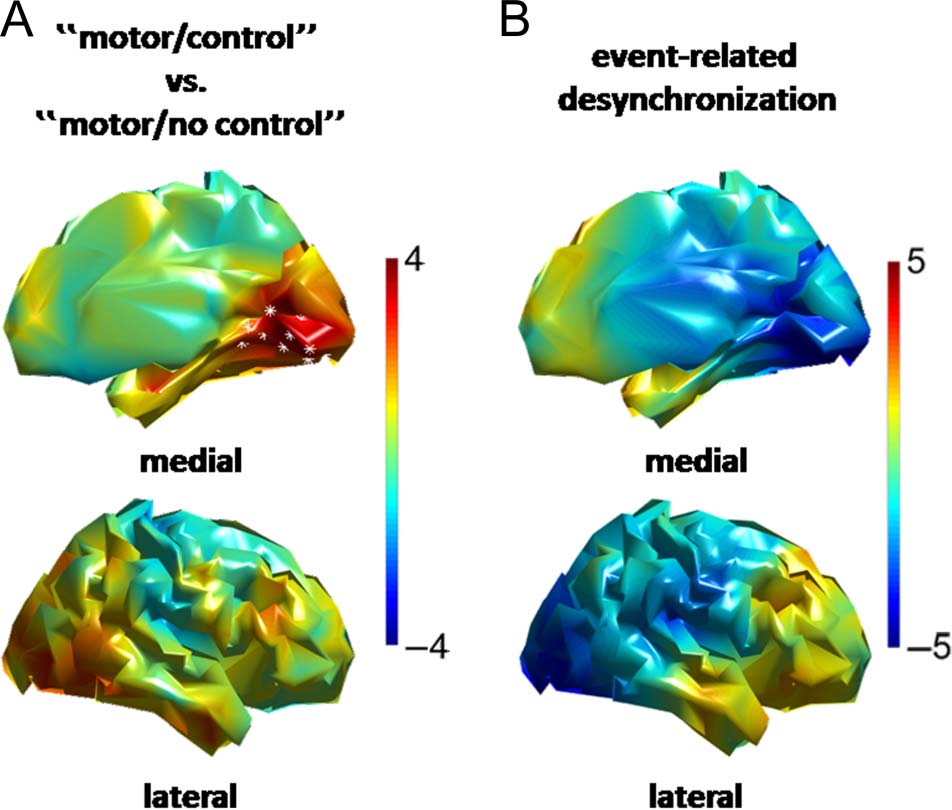
We next determined the cortical source of the effect of
stimulus control on prestimulus alpha-power and com-
pared it to the topography of the stimulus-induced de-
crease in alpha-band power. As expected, stimulus
control significantly modulated the source-level alpha-
power difference between the first and second Gabor
in a prestimulus time interval (around −200 msec). This
prestimulus effect of stimulus control survived cluster-
based whole-brain correction in a region including the
calcarine sulcus and ventral occipital cortex of the right
hemisphere (contralateral to the hemifield in which the
Gabor patches were presented; Figure 3A, top: medial
view, bottom: lateral view; p = .016, cluster-based cor-
rection for multiple comparisons across all cortical grid
points).
When comparing the source of the effect of stimulus
control on prestimulus alpha-power with the cortical
topography of the event-related alpha-desynchronization,
we found highly overlapping regions: Alpha-power was
significantly suppressed by stimulus presentation in a
region comprising the calcarine sulcus, parieto-occipital
sulcus, cuneus, ventral occipital cortex, and middle and
inferior occipital gyri of the right hemisphere (Figure 3B;
poststimulus [300–400 msec] vs. baseline [−200 msec]
across both Gabor patches for the “no motor” condition
as a localizer, p < .001, cluster-based correction for multi-
ple comparisons across all cortical grid points). Thus,
stimulus control modulated prestimulus alpha-power in
2546
Journal of Cognitive Neuroscience
Volume 26, Number 11
avoided this confound. Stimulus onset and orientation
were determined by the timing and the choice of pre-
ceding button presses in the “motor/control” condition
and independent of an irrelevant, but otherwise identical
motor response in the “motor/no control” condition.
Hughes et al. (2012) recently emphasized the impor-
tance of dissociating the relative contributions of four
components of causal control to sensory attenuation,
namely the contributions of stimulus onset predictability,
predictability of stimulus identity, control over stimulus
onset and control over stimulus identity. Here, we manip-
ulated causal control while keeping temporal and identity
predictability comparable across conditions. Because of
the cues at the beginning of each trial, the identity (orien-
tation) of the stimulus was equally predictable across
conditions. Stimulus onset was fully predictable by the
timing of button presses in “motor/control” condition
and became temporally predictable within the first few
trials of each “motor/no control” block because of the
fixed time lag between the auditory cue and stimulus
onset. Together with temporal control, we manipulated
the “outcome density” (Vallée-Tourangeau, Murphy, &
Baker, 2005), namely the probability of stimulus presenta-
tion in the absence of action (in no-go trials).
It is difficult to control for both stimulus predictability
and motor output at the same time. In particular, equal
subjective predictability of the onset of a stimulus in
“motor/control” and “motor/no control” conditions is
challenging when the temporally predictive cue and the
to-be-predicted stimulus are separated by a motor action
that is irrelevant to stimulus timing. Despite any potential
insufficiency of temporal cueing in “motor/no control”
conditions in our study, our main physiological finding
—enhanced anticipatory alpha-power in “motor/control”
conditions—is the opposite of what the previous litera-
ture predicts with respect to low-frequency oscillations
under conditions of increased temporal expectancy.
Specifically, in analogy to the well-established anticipa-
tory modulation of low-frequency oscillations in sensory
cortex during spatial attention (e.g., Thut, Nietzel, Brandt,
& Pascual-Leone, 2006), Rohenkohl and Nobre (2011)
reported stronger occipital alpha-desynchronization when
comparing conditions of high versus low temporal expec-
tancy of visual stimuli. An equivalent result—temporal
specificity of the typical prestimulus desynchronization of
low-frequency oscillations during attention—has been re-
ported in the somatosensory modality (Van Ede, de Lange,
Jensen, & Maris, 2011). If temporal predictability and,
thereby, temporal expectancy were indeed higher in the
“motor/control” condition than in the “motor/no control”
condition, we would have expected the opposite to what
we found, namely decreased anticipatory alpha-power.
Higher predictability and, as a result, greater expec-
tancy of the stimulus in “motor/control” versus “motor/
no control” conditions would therefore only strengthen
our physiological finding of enhanced anticipatory alpha-
power.
Stenner et al.
2547
Figure 3. Main effects of stimulus control and of Gabor patch
presentation on alpha-oscillations at the source level. Color codes for
t values in all panels. (A) Right-hemisphere topography of the main
effect of stimulus control on the prestimulus (−200 msec) difference
in alpha-power (10 Hz) between the first and the second stimulus.
White asterisks indicate significant grid points. Top, medial view;
bottom, lateral view. (B) Right-hemisphere topography of the event-
related alpha-desynchronization (300–400 msec poststimulus vs.
baseline [−200 msec]). Top, medial view; bottom, lateral view.
regions that were subsequently engaged in stimulus-driven
processing as reflected by alpha-desynchronization.
DISCUSSION
We demonstrate that causal control over both onset and
identity of visual stimuli results in enhanced anticipa-
tory alpha-power in visual cortex, an enhancement we
link to a psychophysical measure of sensory attenua-
tion. We suggest that this prestimulus effect of causal
control on alpha-power reflects an inhibitory signal of
motor-induced anticipation in visual cortex that reduces
its excitability to afferent input, thereby leading to sensory
attenuation.
Specificity to Stimulus Control
Our study was motivated by the fact that a majority of
previous studies on sensory attenuation confound volun-
tary causal control over a sensory outcome with execu-
tion of a motor response and/or stimulus predictability
(Hughes et al., 2012). These studies compare stimuli that
are contingent on motor actions (typically labeled “motor-
to-effect” conditions) to stimuli that are not preceded by
motor responses (typically labeled “effect only” condi-
tions). By comparing stimuli that were either controlled
by a preceding action or independent of this action, we
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
1
2
1
6
/
2
1
5
1
4
/
0
2
1
5
9
4
4
0
7
/
8
1
6
4
7
8
o
2
c
0
n
7
_
2
a
/
_
j
0
o
0
c
6
n
5
8
_
a
p
_
d
0
0
b
6
y
5
g
8
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
t
f
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Bias as a Measure of Sensory Attenuation
We used bias in a contrast discrimination task as a mea-
sure of sensory attenuation, similar to many previous
studies of sensory attenuation, which report shifts in
the PSE in discrimination tasks (e.g., Desantis et al.,
2012; Weiss et al., 2011; Haggard & Whitford, 2004). A
change in bias (or PSE) due to sensory attenuation is in
line with theoretical predictions of an influential com-
putational model of motor control (Wolpert & Miall,
1996) that is often referred to explain sensory attenua-
tion. This model predicts that a close match between
predicted and observed action consequences results in
signal cancellation (Blakemore, Wolpert, & Frith, 1998).
Signal cancellation is distinct from a change of the sen-
sory gain function, which has been considered an alter-
native mechanism underlying sensory attenuation (Brown,
Adams, Parees, Edwards, & Friston, 2013). The effects of
signal cancellation and of a change in gain on the two main
signal detection theory measures, sensitivity and bias,
depend on the task design. In a detection paradigm, both
signal cancellation and gain reduction predict a decrease
in sensitivity. However, in discrimination paradigms, like
the one used here, mere cancellation of a sensory signal
leads to a perceptual bias without change in sensitivity.
In contrast, a change in sensory gain affects both signal
amplitude and variance (signal-to-noise ratio) and, thereby,
affects discriminability from the reference stimulus, that
is, discrimination sensitivity. Indeed, absence of a sensitivity
effect in our discrimination task speaks against any major
confounding effect of our manipulation of stimulus control
on attention. An attentional effect would typically alter the
signal-to-noise ratio and, thereby, the sensory gain function
(Hillyard, Vogel, & Luck, 1998).
In signal detection theory, bias has traditionally been
regarded as a measure of decision-related rather than
purely sensory processes. However, an interpretation of
bias as a purely decisional parameter, indexing only late
response stages, has been questioned on the grounds of
recent evidence that bias (“criterion”) can in fact reflect
the baseline activity of signal-selective units at an early,
sensory stage (Nobre, Summerfield, & Wyart, 2012).
Our manipulation of stimulus control leads to a change
of bias together with a physiologically plausible MEG
signal in the relevant sensory cortex. This co-occurrence
suggests that our behavioral result reflects a change in
sensory processing rather than a purely decisional effect.
Sensory Attenuation and Anticipatory
Alpha-oscillations
The original definition of sensory attenuation, based on
the observation that detection of somatosensory stimuli
is reduced when the stimulated limb is engaged in active
movement (Chapman, Bushnell, Miron, Duncan, & Lund,
1987), has recently been extended to include other
sensory modalities and abstract action–outcome associa-
tions in psychophysical and electrophysiological studies
(Gentsch & Schütz-Bosbach, 2011; Weiss et al., 2011;
Cardoso-Leite et al., 2010; Martikainen et al., 2005). The
lack of data on neurophysiological mechanisms, particu-
larly in humans, which may contribute to sensory atten-
uation, has meant that similarities in physiology and
function of these sensory phenomena, as implied by the
common term, have remained speculative.
A traditional theoretical framework used to explain
sensory attenuation across sensory modalities and motor
programs is based on motor-induced sensory predictions
(Waszak et al., 2012; Schütz-Bosbach & Prinz, 2007;
Wolpert et al., 1995). Although a number of studies have
examined the functional anatomy of putative efference
copy pathways upstream of primary sensory processing
by focusing on cortical motor areas (Voss et al., 2009;
Haggard & Whitford, 2004), the FEF (Ostendorf et al.,
2012), or on the thalamus (Ostendorf, Liebermann, &
Ploner, 2010; Bellebaum, Hoffmann, Koch, Schwarz,
& Daum, 2006; Bellebaum, Daum, Koch, Schwarz, &
Hoffmann, 2005), direct neural evidence of anticipatory
sensory activity as a result of such motor-induced pre-
dictions is sparse (Chen et al., 2011; Kühn et al., 2010)
and rarely studied in relation to sensory attenuation.
We note that Chen et al. recently reported enhanced
phase-locking in the gamma-frequency band between
Brocaʼs area and auditory cortex 50 msec before speech
onset using ECoG and a paradigm that compared vocal-
izing and listening. This enhanced gamma-phase syn-
chrony correlated interindividually with attenuation of the
auditory N100 in response to self-produced speech when
compared with replay (Chen et al., 2011).
Studying visual action consequences, we show a pre-
stimulus effect of voluntary causation on neural oscilla-
tions in sensory cortex that differs from that reported
by Chen et al. for speech. Of the two Gabor patches that
followed the motor response in our task, the first was
preceded by enhanced alpha-amplitude by up to 300 msec
when its onset and orientation were fully determined by
the action, whereas the amplitude of alpha-oscillations
was slightly (and not significantly) reduced during the pre-
stimulus interval of the second, temporally uncontrollable
and later stimulus. We thus observe an anticipatory effect in
sensory cortex that starts earlier than reported by Chen
et al., affects low-frequency oscillations rather than
gamma-frequencies, and depends on the temporal control
of the stimulus by the action and/or the time lag between
the two.
Occipital alpha-oscillations have been implicated in
top–down control of visual perception (Von Stein, Chiang,
& König, 2000) across a variety of cognitive tasks (Bauer,
Kluge, et al., 2012; Jensen & Mazaheri, 2010). An inhibitory
role of alpha-oscillations is widely accepted (Lange,
Oostenveld, & Fries, 2013; Klimesch, Sauseng, & Hanslmayr,
2007) and has given rise to proposals of generic functions
of sensory alpha-oscillations, namely gating ( Jensen &
Mazaheri, 2010) and prioritizing ( Jensen et al., 2012) of
2548
Journal of Cognitive Neuroscience
Volume 26, Number 11
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
1
2
1
6
/
2
1
5
1
4
/
0
2
1
5
9
4
4
0
7
/
8
1
6
4
7
8
o
2
c
0
n
7
_
2
a
/
_
j
0
o
0
c
6
n
5
8
_
a
p
_
d
0
0
b
6
y
5
g
8
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
/
.
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
sensory processing. Prestimulus occipital alpha-oscillations
have been causally linked to performance on visual detec-
tion tasks (Romei, Gross, & Thut, 2010; Van Dijk et al.,
2008; Hanslmayr et al., 2007), with impaired target detec-
tion following higher alpha-amplitudes. In light of this
inhibitory role of (prestimulus) alpha-oscillations in the
top–down control of visual perception, our finding of
alpha-amplitude enhancement before self-generated visual
stimuli is a plausible correlate of motor-induced sensory
anticipation such as corollary discharge. The fact that
alpha-amplitude enhancement precedes both stimulus
onset and motor execution does not speak against this
proposal. The origin of efference copies has been linked
to motor processing upstream of primary motor cortex in
humans (Voss et al., 2009; Haggard & Whitford, 2004), and
anticipatory sensory activity might conceivably precede
the execution of the motor command.
To the best of our knowledge, this is the first study that
establishes empirical links between a psychophysical
measure of sensory attenuation and physiological evi-
dence of anticipatory sensory modulation during action.
Our findings extend on previous studies, which have
shown changes in stimulus-evoked responses alone. We
highlight frequency-specific effects of voluntary causal
control, specifically on alpha-oscillations, a frequency
band mechanistically linked to top–down inhibitory con-
trol of perception. Thus, our results link sensory attenu-
ation, motor-induced predictions, and alpha-oscillations
in sensory cortex.
Acknowledgments
This work was supported by the Wellcome Trust (Ray Dolan
Senior Investigator Award 098362/Z/12/Z). M. P. S. is receiving
a scholarship from the German Research Foundation (Deutsche
Forschungsgemeinschaft DFG, STE 2091/1-1). The Wellcome
Trust Centre for Neuroimaging was supported by core funding
from the Wellcome Trust (091593/ Z/10/ Z). We thank David
Bradbury for his help with data acquisition.
Reprint requests should be sent to Max-Philipp Stenner, Wellcome
Trust Centre for Neuroimaging, 12 Queen Square, WC1N 3BG,
UK, or via e-mail: max-philipp.stenner@med.ovgu.de.
Notes
1. Here, we used a weighted up/down staircase procedure
(3 up, 1 down; Kaernbach, 1991) to adjust the difference in
contrast of two sequentially presented Gabor patches. In four
interleaved staircases, the contrast of the test stimulus approached
the pedestal contrast (0.25 Michelson contrast) exponentially
from above and below, separately for each of two Gabor patch
orientations (45° clockwise or counterclockwise). The order of
the test stimulus and the pedestal varied from trial to trial.
Participants had to indicate whether the first or the second Gabor
patch was the high-contrast stimulus. On a given trial, the contrast
of the test stimulus was computed by (0.25 ± 0.1 × 1.06exponent).
Starting from zero, the exponent was varied by a 3 up, 1 down
manipulation, that is, the exponent of the current staircase
was decreased by 1 after a correct response and increased by
3 after an incorrect response. The staircase procedure was
stopped after a total of 200 trials, and discrimination thresholds
were determined as the average of the contrast levels of test
stimuli at all reversals (from decreasing to increasing contrast
differences and vice versa). The trial structure during the stair-
case procedure was identical to the “no motor” condition in
the main experiment.
2. According to Cardoso-Leite et al. (2010), variations of action–
stimulus mapping with respect to previous learning can modulate
the degree of sensory attenuation. Following up on this obser-
vation, we trained participants predominantly on one of the
two mappings during practice (index finger, 45° counterclock-
wise; middle finger, 45° clockwise) and, to avoid extinction, pre-
sented the predominantly learnt mapping slightly more often
during the main task (in 62.5% of trials). However, since we found
no significant perceptual effect of mapping, we collapsed all
“motor/control” trials across both mappings (but note that we
report control analyses that treat the two mappings separately).
3. The information carried by the visual cues in “motor/
control” and “motor/no control” blocks was therefore different.
In “motor/control” blocks, cues signaled the button/orientation
mapping, whereas cues in “motor/no control” blocks predicted
stimulus orientation independently of the following button
choice. This difference in the information carried by the cues
reflects the idea that sensory predictions that arise from motor
actions are physiologically distinct from associative sensory
predictions that arise from external cues (Hughes et al., 2012;
Wolpert & Flanagan, 2001).
4. The button-Gabor orientation mapping had no significant
effect in the “motor/control” and the “motor/no control” con-
ditions as assessed by dependent samples t tests.
5. We observed highly overlapping clusters of right occipital
and parietal sensors that showed significantly enhanced pre-
stimulus (−300 to −100 msec) power between 7.5 and 12.5 Hz
when testing “motor/control” trials with the two button stimulus
orientation mappings separately against the “motor/no control”
condition ( p = .008 and p = .024; cluster-based correction for
multiple comparisons across all channels).
REFERENCES
Alink, A., Schwiedrzik, C. M., Kohler, A., Singer, W., &
Muckli, L. (2010). Stimulus predictability reduces responses
in primary visual cortex. The Journal of Neuroscience, 30,
2960–2966.
Aliu, S. O., Houde, J. F., & Nagarajan, S. S. (2009). Motor-
induced suppression of the auditory cortex. Journal of
Cognitive Neuroscience, 21, 791–802.
Baess, P., Widmann, A., Roye, A., Schröger, E., & Jacobsen, T.
(2009). Attenuated human auditory middle latency
response and evoked 40-Hz response to self-initiated
sounds. The European Journal of Neuroscience, 29,
1514–1521.
Bastiaansen, M. C., & Knösche, T. R. (2000). Tangential
derivative mapping of axial MEG applied to event-related
desynchronization research. Clinical Neurophysiology,
111, 1300–1305.
Bauer, M., Kennett, S., & Driver, J. (2012). Attentional selection
of location and modality in vision and touch modulates low-
frequency activity in associated sensory cortices. Journal of
Neurophysiology, 107, 2342–2351.
Bauer, M., Kluge, C., Bach, D., Bradbury, D., Heinze, H. J.,
Dolan, R. J., et al. (2012). Cholinergic enhancement of
visual attention and neural oscillations in the human brain.
Current Biology, 22, 397–402.
Bays, P. M., Wolpert, D. M., & Flanagan, J. R. (2005).
Perception of the consequences of self-action is temporally
tuned and event driven. Current Biology, 15, 1125–1128.
Stenner et al.
2549
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
1
2
1
6
/
2
1
5
1
4
/
0
2
1
5
9
4
4
0
7
/
8
1
6
4
7
8
o
2
c
0
n
7
_
2
a
/
_
j
0
o
0
c
6
n
5
8
_
a
p
_
d
0
0
b
6
y
5
g
8
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
/
.
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Bellebaum, C., Daum, I., Koch, B., Schwarz, M., & Hoffmann,
K.-P. (2005). The role of the human thalamus in processing
corollary discharge. Brain, 128, 1139–1154.
Bellebaum, C., Hoffmann, K.-P., Koch, B., Schwarz, M., & Daum, I.
(2006). Altered processing of corollary discharge in thalamic
lesion patients. The European Journal of Neuroscience, 24,
2375–2388.
Blakemore, S. J., Wolpert, D. M., & Frith, C. D. (1998). Central
cancellation of self-produced tickle sensation. Nature
Neuroscience, 1, 635–640.
Brown, H., Adams, R. A., Parees, I., Edwards, M., & Friston, K.
(2013). Active inference, sensory attenuation and illusions.
Cognitive Processing, 14, 411–427.
Cardoso-Leite, P., Mamassian, P., Schütz-Bosbach, S., & Waszak, F.
(2010). A new look at sensory attenuation. Action-effect
anticipation affects sensitivity, not response bias.
Psychological Science, 21, 1740–1745.
Chapman, C. E., Bushnell, M. C., Miron, D., Duncan, G. H., &
Lund, J. P. (1987). Sensory perception during movement
in man. Experimental Brain Research, 86, 516–524.
Chen, C.-M. A., Mathalon, D. H., Roach, B. J., Cavus, I.,
Spencer, D. D., & Ford, J. M. (2011). The corollary
discharge in humans is related to synchronous neural
oscillations. Journal of Cognitive Neuroscience, 23,
2892–2904.
Desantis, A., Weiss, C., Schütz-Bosbach, S., & Waszak, F. (2012).
Believing and perceiving: Authorship belief modulates
sensory attenuation. PLoS One, 7, e37959.
induces FFA and PPA activity. Frontiers in Human
Neuroscience, 4, 54.
Lally, N., & Frendo, B. (2011). Sensory cancellation of
self-movement facilitates visual motion detection. Journal
of Vision, 11, 1–9.
Lange, J., Oostenveld, R., & Fries, P. (2013). Reduced occipital
alpha power indexes enhanced excitability rather than
improved visual perception. The Journal of Neuroscience,
33, 3212–3220.
Litvak, V., Mattout, J., Kiebel, S., Phillips, C., Henson, R.,
Kilner, J., et al. (2011). EEG and MEG data analysis in
SPM8. Computational Intelligence and Neuroscience,
2011, 1–32.
Maris, E., & Oostenveld, R. (2007). Nonparametric statistical
testing of EEG- and MEG-data. Journal of Neuroscience
Methods, 164, 177–190.
Martikainen, M. H., Kaneko, K., & Hari, R. (2005).
Suppressed responses to self-triggered sounds in the human
auditory cortex. Cerebral Cortex, 15, 299–302.
Nobre, A. C., Summerfield, C., & Wyart, V. (2012). Dissociable
prior influences of signal probability and relevance on visual
contrast sensitivity. Proceedings of the National Academy of
Sciences, U.S.A., 109, 6354.
Oostenveld, R., Fries, P., Maris, E., & Schoffelen, J.-M.
(2011). FieldTrip: Open source software for advanced
analysis of MEG, EEG, and invasive electrophysiological
data. Computational Intelligence and Neuroscience,
2011, 3593–3598.
Gentsch, A., & Schütz-Bosbach, S. (2011). I did it: Unconscious
Ostendorf, F., Kilias, J., & Ploner, C. J. (2012). Theta-burst
expectation of sensory consequences modulates the
experience of self-agency and its functional signature.
Journal of Cognitive Neuroscience, 23, 3817–3828.
Haggard, P., & Whitford, B. (2004). Supplementary motor area
provides an efferent signal for sensory suppression. Brain
Research. Cognitive Brain Research, 19, 52–58.
Hanslmayr, S., Aslan, A., Staudigl, T., Klimesch, W., Herrmann,
C. S., & Bäuml, K.-H. (2007). Prestimulus oscillations predict
visual perception performance between and within subjects.
Neuroimage, 37, 1465–1473.
Hesse, M. D., Nishitani, N., Fink, G. R., Jousmäki, V., & Hari, R.
(2010). Attenuation of somatosensory responses to self-
produced tactile stimulation. Cerebral Cortex, 20, 425–432.
Hillyard, S. A., Vogel, E. K., & Luck, S. J. (1998). Sensory gain
control (amplification) as a mechanism of selective attention:
Electrophysiological and neuroimaging evidence. Philosophical
Transactions of the Royal Society of London, Series B,
Biological Sciences, 353, 1257–1270.
Hughes, G., Desantis, A., & Waszak, F. (2012). Mechanisms of
intentional binding and sensory attenuation: The role of
temporal prediction, temporal control, identity prediction,
and motor prediction. Psychological Bulletin, 139, 133–151.
Hughes, G., & Waszak, F. (2011). ERP correlates of action effect
prediction and visual sensory attenuation in voluntary action.
Neuroimage, 56, 1632–1640.
Jensen, O., Bonnefond, M., & VanRullen, R. (2012). An
oscillatory mechanism for prioritizing salient unattended
stimuli. Trends in Cognitive Sciences, 16, 200–206.
Jensen, O., & Mazaheri, A. (2010). Shaping functional
architecture by oscillatory alpha activity: Gating by inhibition.
Frontiers in Human Neuroscience, 4, 186.
Kaernbach, C. (1991). Simple adaptive testing with the
weighted up-down method. Perception & Psychophysics,
49, 227–229.
Klimesch, W., Sauseng, P., & Hanslmayr, S. (2007). EEG alpha
oscillations: The inhibition-timing hypothesis. Brain
Research Reviews, 53, 63–88.
Kühn, S., Seurinck, R., Fias, W., & Waszak, F. (2010). The
internal anticipation of sensory action effects: When action
stimulation over human frontal cortex distorts perceptual
stability across eye movements. Cerebral Cortex, 22,
800–810.
Ostendorf, F., Liebermann, D., & Ploner, C. J. (2010). Human
thalamus contributes to perceptual stability across eye
movements. Proceedings of the National Academy of
Sciences, U.S.A., 107, 1229–1234.
Pfurtscheller, G., & Aranibar, A. (1977). Event-related cortical
desynchronization detected by power measurements of scalp
EEG. Electroencephalography and Clinical Neurophysiology,
42, 817–826.
Poulet, J. F. A., & Hedwig, B. (2007). New insights into corollary
discharges mediated by identified neural pathways. Trends in
Neurosciences, 30, 14–21.
Rohenkohl, G., & Nobre, A. C. (2011). Alpha oscillations related
to anticipatory attention follow temporal expectations.
The Journal of Neuroscience, 31, 14076–14084.
Romei, V., Gross, J., & Thut, G. (2010). On the role of
prestimulus alpha rhythms over occipito-parietal areas in
visual input regulation: Correlation or causation? The Journal
of Neuroscience, 30, 8692–8697.
Schütz-Bosbach, S., & Prinz, W. (2007). Perceptual resonance:
Action-induced modulation of perception. Trends in
Cognitive Sciences, 11, 349–355.
Sommer, M. A., & Wurtz, R. H. (2006). Influence of the
thalamus on spatial visual processing in frontal cortex.
Nature, 444, 374–377.
Sperry, R. W. (1950). Neural basis of the spontaneous
optokinetic response produced by visual inversion.
Journal of Comparative and Physiological Psychology,
43, 482–489.
Thut, G., Nietzel, A., Brandt, S. A., & Pascual-Leone, A. (2006).
Alpha-band electroencephalographic activity over occipital
cortex indexes visuospatial attention bias and predicts
visual target detection. The Journal of Neuroscience, 26,
9494–9502.
Vallée-Tourangeau, F., Murphy, R. A., & Baker, A. G. (2005).
Contiguity and the outcome density bias in action–outcome
contingency judgements. The Quarterly Journal of
2550
Journal of Cognitive Neuroscience
Volume 26, Number 11
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
1
2
1
6
/
2
1
5
1
4
/
0
2
1
5
9
4
4
0
7
/
8
1
6
4
7
8
o
2
c
0
n
7
_
2
a
/
_
j
0
o
0
c
6
n
5
8
_
a
p
_
d
0
0
b
6
y
5
g
8
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
t
f
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Experimental Psychology: Series B, Comparative and
Physiological Psychology, 58, 177–192.
Van Dijk, H., Schoffelen, J.-M., Oostenveld, R., & Jensen, O.
(2008). Prestimulus oscillatory activity in the alpha band
predicts visual discrimination ability. The Journal of
Neuroscience, 28, 1816–1823.
Van Dijk, H., van der Werf, J., Mazaheri, A., Medendorp, W. P., &
Jensen, O. (2010). Modulations in oscillatory activity with
amplitude asymmetry can produce cognitively relevant event-
related responses. Proceedings of the National Academy of
Sciences, U.S.A., 107, 900–905.
Van Ede, F., de Lange, F., Jensen, O., & Maris, E. (2011).
Orienting attention to an upcoming tactile event involves a
spatially and temporally specific modulation of sensorimotor
alpha- and beta-band oscillations. The Journal of Neuroscience,
31, 2016–2024.
Von Stein, A., Chiang, C., & König, P. (2000). Top–down
processing mediated by interareal synchronization.
Proceedings of the National Academy of Sciences, U.S.A., 97,
14748–14753.
Voss, M., Ingram, J. N., Haggard, P., & Wolpert, D. M. (2009).
Sensorimotor attenuation by central motor command signals
in the absence of movement. Nature Neuroscience, 9, 26–27.
Waszak, F., Cardoso-Leite, P., & Hughes, G. (2012). Action
effect anticipation: Neurophysiological basis and functional
consequences. Neuroscience and Biobehavioral Reviews,
36, 943–959.
Weiss, C., Herwig, A., & Schütz-Bosbach, S. (2011). The self in
action effects: Selective attenuation of self-generated sounds.
Cognition, 121, 207–218.
Wolpert, D. M., & Flanagan, J. R. (2001). Motor prediction.
Current Biology: CB, 11, R729–R732.
Wolpert, D. M., Ghahramani, Z., & Jordan, M. I. (1995). An internal
model for sensorimotor integration. Science, 269, 1880–1882.
Wolpert, D. M., & Miall, R. C. (1996). Forward models for
physiological motor control. Neural Networks, 9, 1265–1279.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
1
2
1
6
/
2
1
5
1
4
/
0
2
1
5
9
4
4
0
7
/
8
1
6
4
7
8
o
2
c
0
n
7
_
2
a
/
_
j
0
o
0
c
6
n
5
8
_
a
p
_
d
0
0
b
6
y
5
g
8
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
.
f
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Stenner et al.
2551