Eight Problems for the Mirror Neuron Theory
of Action Understanding in Monkeys
and Humans
Gregory Hickok
D
o
w
n
l
o
a
d
e
d
Abstract
& The discovery of mirror neurons in macaque frontal cortex
has sparked a resurgence of interest in motor/embodied theo-
ries of cognition. This critical review examines the evidence
in support of one of these theories, namely, that mirror neu-
rons provide the basis of action understanding. It is argued
that there is no evidence from monkey data that directly tests
this theory, and evidence from humans makes a strong case
against the position. &
INTRODUCTION
. . . we understand action because the motor
representation of that action is activated in
our brain. (Rizzolatti, Fogassi, & Gallese, 2001,
p. 661)
The [motor] theory is so simple and so easy to
present that every one is glad to believe it. The
only question that any one cares to raise is how
much of it will the known facts permit one to
accept. ( Walter B. Pillsbury, 1911, p. 84)
Motor theories of cognition have a long history in psy-
chology (Scheerer, 1984), dating back at least to Berkeley’s
(1709) motor interpretation of depth perception, and
have been proposed as explanations for a wide range of
mental processes. For example, in the early part of the
20th century, Margaret Floy Washburn proposed a mo-
tor theory of mental imagery (Washburn, 1914, 1916),
and John B. Watson explained thought as nothing more
than speech-related sensory–motor processes: ‘‘accord-
ing to my view, thought processes are really motor hab-
its in the larynx’’ ( Watson, 1913, p. 174). As early as 1910,
in the Presidential Address at the American Psychological
Association meeting in Minneapolis, Walter B. Pillsbury
summarized the prevalence of motor theories in the his-
tory of psychology succinctly, ‘‘. . . there is nothing in
the mind that has not been explained in terms of move-
ment’’ (Pillsbury, 1911, p. 84). He also highlighted the
widespread popularity of motor theories in his own time,
commenting that, ‘‘A reader of some of the texts lately
University of California, Irvine
published would be inclined to believe that there was
nothing in consciousness but movement, and that the
presence of sense organs, or of sensory and associatory
tracts in the cortex was at the least a mistake on the part
of the Creator’’ (Pillsbury, 1911, p. 83).
The mirror neuron theory of action understanding
(Rizzolatti & Craighero, 2004; Rizzolatti et al. 2001;
Gallese, Fadiga, Fogassi, & Rizzolatti, 1996; di Pellegrino,
Fadiga, Fogassi, Gallese, & Rizzolatti, 1992) is the latest
in this long line of motor theories—the motor theory
of speech perception (Liberman, Cooper, Shankweiler,
& Studdert-Kennedy, 1967) being a prominent mid-
century representative—and as with motor theories of
the past, seems to have a firm grasp on the field. In
fact, judging from the frequency of appearance of mirror
neuron-related publications in prominent journals, and
the range of abilities and disorders to which the theory
has been extended (e.g., speech perception, music per-
ception, empathy, altruism, emotion, theory of mind,
imitation, autism spectrum disorder, among others),
the comments of Pillsbury, appropriately updated, are
equally applicable today as they were a century ago.
Pillsbury’s goal in his address was ‘‘to attempt a critical
if sympathetic survey of the different formulations of the
theory and to compare it with the facts’’ (p. 84). My goal
here with respect to mirror neuron theory is the same.
Mirror neurons are an interesting class of cells that de-
serve to be thoroughly investigated and their function
fully understood. My view is that the intense focus on
one interpretation of mirror neuron function, that of
action understanding, has impeded progress on mirror
neuron research. Although the action understanding hy-
pothesis is interesting and worthy of investigation, I will
argue that it fails dramatically on empirical examination
(Negri et al., 2007; Mahon & Caramazza, 2005). I will
D 2008 Massachusetts Institute of Technology
Journal of Cognitive Neuroscience 21:7, pp. 1229–1243
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
–
p
r
d
t
i
2
c
1
l
7
e
–
1
p
2
d
2
f
9
/
1
2
9
1
3
/
7
7
7
/
8
1
9
2
o
2
c
9
n
/
1
2
0
7
0
6
9
0
1
2
9
1
0
1
8
/
9
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
1
8
0
9
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
f
.
/
t
.
.
o
n
1
8
M
a
y
2
0
2
1
start by providing a brief review of the properties of mir-
ror neurons, followed by a discussion of eight problems
for the mirror neuron theory of action understanding.
MIRROR NEURONS: THE DATA
Mirror neurons, which famously respond both when the
monkey makes active movements and when it observes
the experimenter making meaningful movements, were
discovered in frontal area F5 of the macaque monkey
(Macaca nemestrina) (Gallese et al., 1996; di Pellegrino
et al., 1992). Studies of F5 before the discovery of mirror
neurons revealed that most cells in that region respond
during the execution of motor acts such as grasping, hold-
ing, and tearing, and a fraction of these also respond to
passive somatosensory ((cid:1)40%) or visual ((cid:1)17%) stimu-
lation in the absence of action (Rizzolatti et al., 1988).
Accordingly, the region’s function was interpreted as
supporting a motor ‘‘vocabulary where proximal and
distal movement necessary for reaching, grasping, hold-
ing and bringing food to the mouth are represented’’
(Rizzolatti et al., 1988, p. 506). In this context, responses
to visual objects or somatosensory stimulation were in-
terpreted not as the neural basis of object or tactile
understanding, but as a mechanism for sensory stimula-
tion to access various motor acts (Rizzolatti et al., 1988).
interpretation
Since the discovery of mirror neurons,
of nonmirror neurons in F5 has not changed among
most F5 experts. For example, with respect to ‘‘canonical’’
(i.e., nonmirror) object-responsive neurons in F5, Nelissen,
Luppino, Vanduffel, Rizzolatti, and Orban (2005) state,
‘‘These neurons are known to play an important role
in the visuomotor transformation for grasping, but they
do not appear to have any role in objects’ identification’’
(p. 334).1
According to the most detailed early study (Gallese
et al., 1996), mirror neurons constituted 17% of sampled
cells in the portion of F5 that was examined, and exhibit
the following properties. The cells were activated when
the monkey observed hand and/or mouth movements
that were directed toward objects (‘‘goal-directed’’ ac-
tions). Roughly half (55%) were selective for one type
of action, with grasping the most frequently represented
movement across the population of cells (75% of cells).
The majority of cells were either strictly or broadly con-
gruent with their action execution response properties.
The cells did not respond to visually presented objects or
food items, faces, non-goal-directed body movements,
goal-directed actions made using tools (although see
Ferrari, Rozzi, & Fogassi, 2005), mimicking of grasping in
the absence of an object (pantomime), or gestures having
emotional meaning. The cells do not exhibit movement
preparation activity: They discharge when the monkey
observes an action, stop firing when the action termi-
nates, and remain quiet even if the object is moved to-
ward the monkey, firing again only when the monkey
initiates its own action. This is an important fact as this
property distinguishes mirror neurons from well-known
‘‘set-related’’ neurons in nearby monkey area 6 that dis-
charge before movement onset (Wise & Mauritz, 1985;
Weinrich, Wise, & Mauritz, 1984). As important controls
for the possibility that ‘‘mirror activity’’ reflected some
form of covert movement, Gallese et al. (1996) recorded
from the hand area of primary motor cortex (F1 or M1),
and recorded EMG activity from several hand and mouth
muscles during action observation. No M1 cells fired, and
no EMG activity was elicited in response to action obser-
vation. On the basis of this evidence, mirror neurons were
hypothesized to support ‘‘action understanding.’’
Since these early studies, mirror neurons have also
been found in monkey parietal cortex (Gallese, Fogassi,
Fadiga, & Rizzolati, 2002), and problematically (see be-
low), in M1 (Tkach, Reimer, & Hatsopoulos, 2007).
MIRROR NEURONS: THE THEORY
Unlike the majority of the (nonmirror) neurons in macaque
area F5, which are argued to support a ‘‘motor vocabu-
lary’’ (Rizzolatti et al., 1988), mirror neurons are claimed
to support ‘‘action understanding’’ (Rizzolatti & Craighero,
2004; Rizzolatti et al., 2001; Gallese et al., 1996; di
Pellegrino et al., 1992). ‘‘Action understanding’’ is de-
fined somewhat differently in various papers. Gallese
et al. (1996) define it as ‘‘the capacity to recognize that
an individual is performing an action, to differentiate this
action from others analogous to it, and to use this infor-
mation in order to act appropriately’’ (p. 606). Rizzolatti
et al. (2001) propose that action understanding is ‘‘the
capacity to achieve the internal description of an action
and to use it to organize appropriate future behaviour’’
(p. 661). Rizzolatti and Craighero (2004) claim, ‘‘Each
time an individual sees an action done by another indi-
vidual, neurons that represent that action are activated
in the observer’s premotor cortex. This automatically
induced, motor representation of the observed action
corresponds to that which is spontaneously generated
during active action and whose outcome is known to the
acting individual. Thus, the mirror system transforms vi-
sual information into knowledge’’ (p. 172). Nelissen et al.
(2005) state that ‘‘A mere visual representation [of an
action], without involvement of the motor system, pro-
vides a description of the visible aspects of the move-
ment of the agent, but does not give information critical
for understanding action semantics, i.e., what the action
is about, what its goal is, and how it is related to other
actions’’ (p. 332). The notion ‘‘action understanding’’ has
been generalized in humans to include speech percep-
tion (Wilson, Saygin, Sereno, & Iacoboni, 2004; Rizzolatti
& Arbib, 1998; Gallese et al., 1996).
It is not obvious from the definitions quoted above
what ‘‘action understanding’’ means. For example, with
respect to the first definition, upon seeing an individual
producing meaningless, non-goal-directed actions (e.g.,
flailing the arms, which should yield no mirror neuron
1230
Journal of Cognitive Neuroscience
Volume 21, Number 7
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
–
p
r
d
t
i
2
c
1
l
7
e
–
1
p
2
d
2
f
9
/
1
2
9
1
3
/
7
7
7
/
8
1
9
2
o
2
c
9
n
/
1
2
0
7
0
6
9
0
1
2
9
1
0
1
8
/
9
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
1
8
0
9
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
t
.
f
/
.
o
n
1
8
M
a
y
2
0
2
1
activity), one could presumably ‘‘recognize’’ that actions
are being performed, ‘‘differentiate’’ such actions from
other actions (e.g., swinging the arms rhythmically), and
‘‘act appropriately’’ in response (walk away or call 911),
all without ‘‘understanding’’ the meaning of the actions
in the goal-directed sense. The nature of the ‘‘internal de-
scription’’ in the second definition is itself undefined and
therefore adds little clarity to the nature of action under-
standing. In the third definition, the idea that under-
standing is achieved by knowing the ‘‘outcome’’ is also
somewhat vague because ‘‘outcome’’ is not defined. The
fourth definition also includes concepts that are under-
specified: What is the action of grasping a peanut ‘‘about’’?
What is the ‘‘goal’’ of such an action? And on what level
of analysis is ‘‘relation’’ between actions defined?
The most reasonable interpretation (in my mind) is
that what is being ‘‘understood’’ by mirror neurons while
observing peanut-grasping is something closer to the
‘‘grasping-with-the-hand.’’ However, Nelissen
concept,
et al. suggest that mirror neurons are coding more than
the ‘‘essence of grasping’’ (p. 334) which they believe is
coded in a more anterior region of F5 (Nelissen et al.,
2005). In short, the concept of action understanding
has been evolving, but at its core is the idea that self-
generated actions have an inherent semantics and that
observing the same action in others affords access to
this action semantics.
The existence of mirror neurons has been inferred to
exist in humans, beginning with the earliest mirror neu-
ron reports (Gallese et al., 1996; di Pellegrino et al., 1992).
These early claims (Gallese et al., 1996) were based on
(i) the fact that pantomime recognition deficits exist in
aphasia (Gainotti & Lemmo, 1976), (ii) a PET study in
humans showing activation in Broca’s region during ac-
tion observation (Rizzolatti et al., 1996), and (iii) a trans-
cranial magnetic stimulation (TMS) study that showed
enhanced distal muscle motor-evoked potentials (MEPs)
during action observation (Fadiga, Fogassi, Pavesi, &
Rizzolatti, 1995). However, the empirical basis for the gen-
eralization of the mirror neurons to humans was dubious
from the start based on the very data that was claimed to
support it: (i0) Mirror neurons do not respond to panto-
mimed actions and so pantomime recognition should not
rely on the mirror system. Further, pantomime recogni-
tion deficits were not associated with frontal lesions, but
rather were predominantly associated with posterior le-
sions (Heilman, Rothi, & Valenstein, 1982). (ii0) The PET
study showing Broca’s region activation during action ob-
servation failed to show overlapping activation during
grasping production (Rizzolatti et al., 1996), in contrast
to the central mirror neuron observation. And (iii0) the
TMS finding of peripheral motor activation during action
observation directly contradicted the early demonstration
in monkeys that M1 and the peripheral motor system did
not exhibit mirror properties (Gallese et al., 1996).
Mirror neuron findings were also quickly generalized
to speech (Gallese et al., 1996; di Pellegrino et al., 1992)
on the basis of analogy to the motor theory of speech
perception (Liberman et al., 1967). But despite its mirror
neuron-led resurgence in popularity among nonspeech
scientists, the motor theory of speech perception ‘‘has
few proponents within the field of speech perception’’
(Galantucci, Fowler, & Turvey, 2006, p. 361). Thus, the
theoretical grounding of mirror neuron theory in the
speech domain was not particularly strong.
Mirror neurons have also been generalized to explain
imitation (Rizzolatti & Craighero, 2004). This function of
mirror neurons, however, has been restricted to humans
because macaques (at least adult macaques; Ferrari
et al., 2006) do not imitate (Visalberghi & Fragaszy,
2001). This means that mirror neuron function, as it is
studied in macaque monkeys, cannot be the basis of imi-
tation. Rizzolatti and Craighero (2004) emphasize that
‘‘the primary function of mirror neurons cannot be ac-
tion imitation’’ (p. 172). Any evidence regarding the
neural basis of imitation in humans, therefore, cannot
be empirically linked to mirror neurons.
Although the ‘‘mirror system’’ has been used as the
basis for understanding a range of behaviors, we will
focus our attention on the core function supposed to
hold across species, namely, action understanding. If the
mirror neuron theory fails to stand up empirically with
respect to its core claim, as I will argue, then linkage
between mirror neurons and the many systems and dis-
orders linked to their function is highly dubious.
The perception of a graspable object is sufficient to
trigger the activation of cells in motor area F5 (Rizzolatti
et al., 1988). Most mirror neuron theorists do not endow
cells that respond to the perception of objects with an
object semantics. Instead, they propose that F5 contains
a motor vocabulary, and that sensory (object) responses
in F5 cells reflect a means for grasping-related sensory
information to access that vocabulary.2 When consider-
ing mirror neuron function, it is helpful to adopt this
view of F5 function as the null hypothesis, namely, that
F5 is fundamentally a motor area that is capable of sup-
porting sensory–motor associations. In order to make
a serious case for mirror neurons as the basis of action
understanding, one has to show that they are qualita-
tively different from other sensory–motor cells in F5,
specifically, that they are coding more than just a sensory–
motor association (they have a semantics that other
sensory cells in F5 do not). In what follows, I will detail
eight problems that undermine the claim that mirror
neurons go beyond other sensory–motor cells in F5 and
support action understanding.
1. There Is No Evidence in Monkeys That Mirror
Neurons Support Action Understanding
The mirror neuron theory of action understanding pre-
dicts that disruption of motor areas in F5 should pro-
duce deficits in action perception. Although functional
Hickok
1231
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
–
p
r
d
t
i
2
c
1
l
7
e
–
1
p
2
d
2
f
9
/
1
2
9
1
3
/
7
7
7
/
8
1
9
2
o
2
c
9
n
/
1
2
0
7
0
6
9
0
1
2
9
1
0
1
8
/
9
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
1
8
0
9
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
f
.
/
t
.
.
o
n
1
8
M
a
y
2
0
2
1
disruption of macaque area F5 has been shown to dis-
rupt grasping behavior (Fogassi et al., 2001), the pre-
dicted corresponding decrement in action perception
has never been reported. Rizzolatti and Craighero (2004)
argue that such studies are not feasible. This is because
(i) the mirror system is bilateral and involves parietal
structures, (ii) there are other mechanisms that mediate
action recognition, and (iii) if one lesioned the entire
mirror neuron system, more general cognitive deficits
would result, making interpretation difficult (Rizzolatti
& Craighero, 2004). However, if the claim is that motor
systems underlie action understanding, and if it is pos-
sible to impair motor behavior by disruption of motor
systems in F5 (Fogassi et al., 2001), then it should follow
that action understanding will be commensurately im-
paired. If, on the other hand, motor behavior and action
understanding dissociate in macaque following F5 dis-
ruption, this would constitute evidence against a critical
role for motor systems (and area F5) in action under-
standing, independently of whether the mirror system
extends beyond F5 or not.
In place of the standard lesion method, three studies
are held up as evidence that mirror neurons in monkeys
support action understanding. One involves the dem-
onstration that some mirror neurons (15%) respond to
action-associated sounds presented in isolation (cracking
peanut shell, ripping paper) (Kohler et al., 2002). The
logic here is that ‘‘If mirror neurons mediate action un-
derstanding, their activity should reflect the meaning of
the observed action, not its visual features’’ (Rizzolatti &
Craighero, 2004, p. 173). According to this logic, Kohler
et al.’s findings indicate that 85% of mirror neurons do
not mediate action understanding because their activity
does not reflect the meaning of the perceived action.3
This still leaves a population of 15% of mirror neurons—
the audiovisual type—that may code the meaning of ac-
tions. Does the existence of these audiovisual mirror neu-
rons prove that they are coding meaning? No. A more
straightforward interpretation of this result is that sounds
just as
can be associated with actions in F5 neurons,
objects can be associated with actions in F5 neurons
(Rizzolatti et al., 1988). Framed in terms of a priming ex-
planation, we might argue that the animal has associated
the action of breaking a peanut with the sound of break-
ing a peanut, and when hearing only the sound, the activa-
tion spreads to F5; a form of partial cue retrieval. Right or
wrong, the point is that we do not need to endow these
cells with semantic properties to explain the finding.
The second experiment showed that although mirror
neurons do not respond to pantomimed actions (ac-
tions without the object present), they do respond if an
action is directed toward an object that is hidden behind
a screen such that the monkey knows it is there (Umilta`
et al., 2001). In this scenario, more than half of the mir-
ror neurons that were tested also responded in the hid-
den condition. The logic here is the same, that it is not
the physical features of the action that drives the re-
sponse, but rather the knowledge of the ‘‘meaning’’ of
the action. Again, following the logic, the results of the
study indicate that half of all mirror neurons are not cod-
ing action meaning, and again there is a simpler explana-
tion. The monkey can represent the object in working
involves
memory which, according to popular views,
the same systems that represent the object when it is
physically present (Postle, 2006; Pasternak & Greenlee,
2005; Ruchkin, Grafman, Cameron, & Berndt, 2003;
Fuster, 1995). This information can then be used in the
normal manner as if the object was visible.
Rizzolatti and Craighero (2004) claim that these studies
show that ‘‘the activity of mirror neurons correlates with
action understanding’’ (p. 174). However, action under-
standing was never actually measured, and there is a
simpler explanation of both results, one that fits well with
the hypothesized function of the nonmirror neurons in
F5, namely, that perceptual information—including ob-
jects, tactile stimulation, sounds, and actions—can be
associatively linked to and can prime a ‘‘motor vocabu-
lary’’ in F5 (Rizzolatti et al., 1988).
The third study by Fogassi et al. (2005) uses a differ-
ent approach to argue for abstract, action understanding
properties of mirror neurons. These authors present
very interesting data from the inferior parietal lobule
(IPL) of monkeys, which also contains mirror neurons,
as noted above. Monkeys were trained either to grasp
a piece of food and put it in his (the monkey’s) mouth,
or to pick up an object and put it in a container. In
some conditions, the container was next to the mon-
key’s mouth such that the mechanics of the movement
were very similar between grasping-to-eat and grasping-
to-place. In addition, a condition was also implemented
in which the monkey grasped and placed a piece of
food in the container to control for differences between
food items and objects, both visually and tactilely. In all
variants of the experiment, the authors report that some
IPL cells preferentially responded to the goal of the ac-
tion: grasping-to-eat versus grasping-to-place. Again, this
was true even when the placing-action terminated in
close proximity to the mouth and involved grasping
a piece of food. Some of these cells also responded
selectively and congruently during the observation of
grasping-to-eat and grasping-to-place. Thus, both in per-
ception and action, there are IPL cells that seem to be
selective for the specific goal of an action rather than
the sensory or motor features of an action—a very in-
triguing result. Fogassi et al. discuss their motor findings
in the context of ‘‘intentional chains’’ in which differ-
ent motor acts forming the entire action are linked in
such a way that each act is facilitated in a predictive and
goal-oriented fashion by the previous ones. They give
an example of IPL neurons observed in another unpub-
lished study that respond to flexion of the forearm, have
tactile receptive fields around the mouth, and respond
during grasping actions of the mouth and suggest that
‘‘these neurons appear to facilitate the mouth opening
1232
Journal of Cognitive Neuroscience
Volume 21, Number 7
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
–
p
r
d
t
i
2
c
1
l
7
e
–
1
p
2
d
2
f
9
/
1
2
9
1
3
/
7
7
7
/
8
1
9
2
o
2
c
9
n
/
1
2
0
7
0
6
9
0
1
2
9
1
0
1
8
/
9
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
1
8
0
9
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
t
.
f
/
.
.
o
n
1
8
M
a
y
2
0
2
1
when an object is touched or grasped’’ (p. 665). Re-
garding the action perception response properties of
the IPL neurons in their study, Fogassi et al. conclude,
‘‘that IPL mirror neurons, in addition to recognizing the
goal of the observed motor act, discriminate identical
motor acts according to the action in which these acts
are embedded. Because the discriminated motor act is
part of a chain leading to the final goal of the action, this
neuronal property allows the monkey to predict the goal
of the observed action and, thus, to ‘read’ the intention
of the acting individual’’ (p. 666).
According to Fogassi et al., IPL mirror neurons code
action goals and can ‘‘read the intention’’ of the acting
individual. Perhaps Fogassi et al.’s notion of predictive
coding and their example of the IPL neuron with re-
ceptive fields on the face can provide a simpler expla-
nation. Suppose the abstract goal of an action and/or
its meaning is coded outside of the motor system. And
suppose that Fogassi et al. are correct in that a com-
plex motor act leads to some form of predictive coding
(anticipatory opening of the mouth, salivation, perhaps
even forward modeling of the expected somatosensory
consequences of the action). The predictive coding in
the motor system is now going to be different for the
grasping-to-eat versus grasping-to-place actions. For eat-
ing, there may be anticipatory opening of the mouth, sal-
ivation, and perhaps anticipatory activity associated with
the expected somatosensory consequences of the action.
For placing, there will be no mouth-related coding, but
there may be other kinds of coding such as expectations
about the size, shape, or feel of the container, or the
sound that will result if the object is placed in it. If cells
in the IPL differ in their sensitivity to feedback from
these different systems, then it may look like the cells
are coding goals, when in fact they are just getting
differential feedback input from the forward models.
Observing an action may activate this system with similar
electrophysiological consequences, not because it is read-
ing the intention of the actor, but simply because the
sensory event is associated with particular motor acts.
2. Action Understanding Can Be Achieved via
Nonmirror Neuron Mechanisms
Rizzolatti and Craighero (2004) noted that the mirror
neuron system may not be the only mechanism that
can support action understanding. Rizzolatti et al. (2001)
also emphasize that ‘‘these [mirror neuron] findings do
not exclude the possibility that other areas are involved
in the description of biological movement and the un-
derstanding of action’’ (p. 662). The existence of other
mechanisms for action understanding is a problem for
the mirror neuron theory of action understanding be-
cause, it places action understanding on par with ‘‘object
understanding.’’ Object responses in F5 are not generally
interpreted as the neural basis for object understand-
ing (Rizzolatti et al., 1988), presumably because other
neural systems in the ventral visual stream support ob-
ject recognition/understanding. Object information, pro-
cessed for ‘‘meaning’’ in the temporal lobe, can gain
access to motor programs as appropriate for behaviors
such as grasping, thus explaining the object response
properties of F5 cells, even though the meaning of the
objects is not coded in these motor areas (Nelissen
et al., 2005). If there is a neural network outside of the
mirror system that can support action understanding, as
Rizzolatti and colleagues suggest, then we can propose
an identical form of interaction. Actions are processed
for ‘‘meaning’’ in this other system, which via the same
associative mechanisms can gain access to motor pro-
grams in F5, thus producing ‘‘mirror’’ responses, anal-
ogous to object responses.
A candidate region for an action understanding alter-
native to mirror neurons is the superior temporal sulcus
(STS). Cells in portions of the macaque STS respond to
a wide range of actions in a manner that appears more
sophisticated than that found in mirror neurons. STS
neurons respond to actions such as walking toward or
away, head turning, movement into or out of view, arm
movements, and hand–object interaction where there
is selectivity for specific actions including reaching, re-
trieving, manipulating, picking, tearing, presenting to
the monkey, and holding (Perrett, Mistlin, Harries, &
Chitty, 1990; Perrett et al., 1985). These cells do not have
motor properties in that they do not appear to fire
during action execution (although this has not been
investigated thoroughly). Interestingly, the region of in-
ferior parietal cortex that contains mirror neurons (PF),
and which projects to F5, receives input from the STS
(Rizzolatti & Craighero, 2004). This would seem to be an
ideal circuit for representing actions (STS) and coordi-
nating their interaction (PF) with the motor system (F5).
3. M1 Contains Mirror Neurons
It was recently observed that mirror neurons exist in pri-
mary motor cortex of macaque monkeys (Tkach et al.,
2007). Although this is consistent with the MEP work in
humans (Fadiga et al., 1995), it undermines an impor-
tant control observation in the original mirror neuron
reports. Recall that the lack of mirror neurons in M1 was
taken as evidence against the possibility that the mon-
keys were covertly generating movement responses dur-
ing the perception of actions. In other words, it ruled
out the possibility that ‘‘mirror’’ responses were merely
some kind of unimplemented motor command, and
opened the door to a more interesting, higher-level
function. Now with the demonstration of ‘‘mirror’’ re-
sponses in low-level motor circuitry (M1 in macaque, and
distal muscles in humans, as demonstrated with TMS), it
is entirely possible that ‘‘mirror’’ responses are nothing
more than the facilitation of the motor system via learned
associations. Tkach et al. (2007) suggest a similar inter-
pretation of their data, namely, ‘‘that the neural activity
Hickok
1233
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
–
p
r
d
t
i
2
c
1
l
7
e
–
1
p
2
d
2
f
9
/
1
2
9
1
3
/
7
7
7
/
8
1
9
2
o
2
c
9
n
/
1
2
0
7
0
6
9
0
1
2
9
1
0
1
8
/
9
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
1
8
0
9
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
t
/
f
.
.
.
o
n
1
8
M
a
y
2
0
2
1
during observation is attributable to the covert generation
of a motor command and that [the reason] we observe
congruent neural activity during observation [is] because
the visual goal, and thus the motor command generated,
is the same as during active movement’’ (p. 13247).
4. The Relation between Macaque Mirror Neurons
and the ‘‘Mirror System’’ in Humans Is Either
Nonparallel or Undetermined
As noted above, mirror neuron function has been gen-
eralized to a wide range of human behaviors. Indeed,
much of the excitement over mirror neurons is directly
related to their potential to explain complex human
capacities and disorders. A statement by Oberman et al.
(2005) illustrates both the extent of the generaliza-
tion and the excitement: ‘‘Mirror neurons are primarily
thought to be involved in perception and comprehen-
sion of motor actions, but they may also play a critical
role in higher order cognitive processes such as imi-
tation, theory of mind, language, and empathy, all of
which are known to be impaired in individuals with
autism spectrum disorders’’ (pp. 190–191, citation num-
bers omitted).
The problem with statements such as this, and many
like it, is that the species that has been shown to possess
mirror neurons does not, to our knowledge, possess
any of these higher-order cognitive processes, and the
species that possesses the higher-order cognitive pro-
cesses has not been shown conclusively to possess mir-
ror neurons (Chong, Cunnington, Williams, Kanwisher,
& Mattingley, 2008; Dinstein, 2008; Dinstein, Thomas,
Behrmann, & Heeger, 2008; Dinstein, Hasson, Rubin, &
Heeger, 2007). To be sure, there have been a host of
studies aimed at investigating the ‘‘mirror system’’ in hu-
mans, but much of this work has investigated behaviors
that mirror neurons could not possibly support given
their response properties in monkeys,4 and therefore,
the connection between these behaviors and mirror
neurons is tenuously based on a chain of assumptions:
Mirror neurons exist in humans (there are individual
cells that respond both during action execution and ac-
tion perception), mirror neurons have evolved to sup-
port functions in humans that they do not support in
monkeys, this evolution has conserved the functional
properties found in monkeys, and mirror neurons are
responsible for the behavior in question. There is noth-
ing wrong with using animal models to generate testable
hypotheses in humans—indeed, this is a productive
and important research strategy. The problem in the
case of mirror neurons is that the system has been gen-
eralized to humans without systematic validation, and
with the wholesale adoption of the mirror neuron doc-
trine concerning action understanding. When a human
study starts with the assumption that mirror neurons
support action understanding (see above quote from
Oberman et al.), and that a homologous and functionally
enriched system exists in humans, it is then an easy and
prima facie logical inference that the human mirror sys-
tem can support higher-order functions such as lan-
guage and empathy. However, this inference falls apart
if any of the assumptions about mirror neurons are in-
correct. Thus, my caution here is not that we cannot or
even should not use mirror neurons to guide human
research, but that we have to first validate our assump-
tions before making inferences regarding human behav-
iors, especially those that do not exist in monkeys.
Let me illustrate the problem with an abstract argu-
ment. Suppose that Rizzolatti and colleagues are correct,
namely, that mirror neurons in monkeys are the basis for
action understanding, but not imitation (because adult
macaques don’t imitate). In humans, the mirror system
behaves differently than in monkeys such that it appears
to support imitation (Rizzolatti & Craighero, 2004): It
activates during the perception and execution of even
meaningless movements (Iacoboni et al., 1999). This ob-
servation has led some mirror neuron theorists to argue
that the mirror system in humans has evolved to sup-
port not only action understanding (based on inferences
from monkey data), but also imitation (based on human
data) (Rizzolatti & Craighero, 2004). The assumption
made by these authors is that in the evolution of this
system, old properties of mirror neurons are fully con-
served. But what if the mirror system evolved in humans
such that it now supports imitation but no longer sup-
ports action understanding? Perhaps humans evolved a
more sophisticated semantic system, distinct from the
motor system, that freed the mirror system to support
imitation. Possibilities such as this are not considered in
mainstream mirror neuron theorizing. Instead, monkey
data and theories are typically imported to human work
without empirical validation of the assumptions.
Here is a concrete example of how monkey data are
assumed to hold, problematically, in human work. In the
context of studying the human mirror system, a number
of functional imaging experiments have investigated the
perception of meaningless gestures, pantomimed ges-
tures, and imitation (Koski, Iacoboni, Dubeau, Woods,
& Mazziotta, 2003; Koski et al., 2002; Iacoboni et al.,
1999; Grezes, Costes, & Decety, 1998; Decety et al., 1997).
These studies, which often implicate portions of the
inferior frontal gyrus (IFG) and the inferior precentral
gyrus, are cited as evidence for the existence of a hu-
man mirror system that has evolved to support imitation
(Rizzolatti & Craighero, 2004). However, this is not the
only interpretation. There are at least three logical pos-
sibilities. (i) Mirror neurons do not exist in humans,
and the activation in these studies results from the func-
tion of some other system. (ii) Mirror neurons exist in
humans exactly as they do in monkeys (with the same
properties), and the activations in these studies result
from the function of some other system. (iii) Mirror neu-
rons exist in humans, but have evolved such that they
now support pantomime recognition and imitation. The
1234
Journal of Cognitive Neuroscience
Volume 21, Number 7
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
–
p
r
d
t
i
2
c
1
l
7
e
–
1
p
2
d
2
f
9
/
1
2
9
1
3
/
7
7
7
/
8
1
9
2
o
2
c
9
n
/
1
2
0
7
0
6
9
0
1
2
9
1
0
1
8
/
9
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
1
8
0
9
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
f
.
/
.
t
o
n
1
8
M
a
y
2
0
2
1
third interpretation is typically adopted, whereas the
other possibilities are not even considered. Why? In
monkey mirror neuron research, other possibilities
were considered. Gallese et al. (1996) considered both
the possibility that mirror neurons reflected ‘‘set-related’’
responses and the possibility that mirror neurons were
reflecting a nonimplemented motor plan (see above).
Because these possibilities were ruled out empirically in
monkeys, it is assumed (probably implicitly) that there
is no need to rule them out in humans. But this is faulty
logic. If mirror neurons exist in humans as is claimed, the
system is demonstrably different from that in the mon-
key. One therefore cannot assume that monkey data will
hold in the human system. The alternative possibilities
have to be ruled out empirically again. Consider in this
respect a highly cited study of imitation in the human
mirror system (Iacoboni et al., 1999), which found equiv-
alent activation during the passive perception of an ac-
tion (a moving hand), a static hand, and a rectangle with
a spatial cue (to which subjects were previously trained
to make a hand movement). The authors explain the
activation to the latter nonaction stimulus is this way:
‘‘During all scans the participants knew that the task was
either to move a finger or to refrain from moving it.
Thus mental
imagery of their finger (or of the finger
movement) should have been present even during sim-
ple observation’’ (Iacoboni et al., 1999, pp. 2526–2527).
This would suggest that it is not action perception that
is driving these ‘‘mirror activations,’’ but simply the in-
ternal activation of a motor act. Indeed, there is evidence
that human area 44, a presumed component of the hu-
man mirror system, is involved in movement preparation
(Krams, Rushworth, Deiber, Frackowiak, & Passingham,
1998).
There are also a number of studies that have investi-
gated the human mirror system using behaviors that do
hold of mirror neurons, namely, object-directed actions.
A recent meta-analyses of fMRI studies of the human
‘‘mirror system’’ (Morin & Grezes, 2008) has suggested
that human BA 44 is not the homologue of the macaque
mirror neuron F5 region, as this region is insensitive to
the presence or absence of target objects in action per-
ception. That is, BA 44 does not distinguish between
object-directed actions and actions that are non-object-
directed. Instead, Morin and Grezes (2008) point out
that a more posterior region, ventral premotor cortex
(BA 6), is activated significantly more often during the
perception of object-directed action than actions without
object goals. Accordingly, these authors propose ventral
BA 6 as the human homologue of the mirror system.
Although Morin and Grezes’ hypothesis is quite reason-
able and is based on direct parallels with the macaque
mirror neuron system, it remains to be experimentally
verified. For example, a nontrivial fraction (36%) of the
studies reviewed by Morin and Grezes reported that per-
ception of non-object-directed actions activated ventral
BA 6. What drove these activations? Also, it will be impor-
tant to confirm that this area has overlapping sensory–
motor response properties. Surprisingly, many investiga-
tions of the mirror system fail to confirm this fundamental
property of mirror neurons—another example of unver-
ified generalization from monkey to human work. In-
deed, Morin and Grezes’ review, which aimed explicitly
to identify the human homologue of the macaque mir-
ror neuron system, focused exclusively on perceptual
responses. It will be important to determine whether
the response properties of this ventral BA 6 region can
be linked directly to action processing, or whether it
might be performing a more general function, on which
action processes rely. For example, recent fMRI and le-
sion evidence has implicated this region in predicting
sequences of abstract nonbiological stimuli, suggesting
a more general functional role involving sequence pro-
cessing (Schubotz, Sakreida, Tittgemeyer, & von Cramon,
2004; Schubotz & von Cramon, 2004).
Other evidence often cited as support for the exis-
tence of a human ‘‘mirror system’’ is the demonstration
that viewing actions can result in peripheral motor po-
tentiation. TMS of motor cortex produces MEPs in distal
muscles. The amplitude of MEPs in distal muscles is en-
hanced during action observation (Fadiga et al., 1995),
which has been interpreted as evidence for a human
mirror system. Although this work shows clearly that
associations between observed actions and motor exe-
cution systems exist, it does not indicate that these as-
sociations are mediated by anything like macaque mirror
neurons. For example, TMS/MEP data cannot rule out
the possibility that the link between action observation
and action execution could be mediated by a nonmotor
conceptual representation.
The relation between the macaque mirror neuron sys-
tem and the hypothesized human homologue remains
to be elucidated (for recent discussion and evidence on
this debate, see Chong et al., 2008; Dinstein et al., 2007,
2008). This in itself, however, is not an argument against
the mirror neuron theory of action understanding. By
the same token, even if a network with mirror-neuron-
like properties can be fully outlined in humans, this in
itself would not be an argument for the mirror neuron
theory of action understanding. Such an argument re-
quires a different sort of evidence; this is the topic of the
next section.
5. Action Understanding in Humans Dissociates
from Neurophysiological Indices of the Human
‘‘Mirror System’’
There are examples in the human ‘‘mirror system’’ lit-
erature of dissociations between action understanding
and ‘‘mirror system’’ function. One study (Buccino et al.,
2004) examined functional activations during the per-
ception of biting actions or communicative gestures per-
formed by a human, a monkey, or a dog. Independent of
the species performing the action, viewing biting actions
Hickok
1235
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
–
p
r
d
t
i
2
c
1
l
7
e
–
1
p
2
d
2
f
9
/
1
2
9
1
3
/
7
7
7
/
8
1
9
2
o
2
c
9
n
/
1
2
0
7
0
6
9
0
1
2
9
1
0
1
8
/
9
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
1
8
0
9
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
t
.
/
.
.
f
o
n
1
8
M
a
y
2
0
2
1
activated regions thought to be part of the human mir-
ror system, the left IFG and the precentral gyrus (among
other areas). Viewing communicative gestures elicited
activation of these frontal mirror systems for actions
performed by a human (lip-reading) and a monkey (lip
smacking), but not a dog (barking). On the assumption
that the study participants ‘‘understood’’ all three com-
municative actions, it is interesting that only the human
and monkey actions resulted in ‘‘mirror system’’ activa-
tion. (At least subjects very likely understood that the
lip-reading action was associated with speech and the
barking action was associated with barking. The monkey
action arguably contained less semantic information.)
This result clearly shows that actions can be understood
without the mirror system, or more to the point, that
mirror system activity is not particularly correlated with
action understanding. Of interest is that the STS was ac-
tivated across all conditions.
Another demonstration of the dissociability of the
mirror system from action understanding comes from
a TMS/MEP study (Catmur, Walsh, & Heyes, 2007). The
authors used TMS to induce MEPs in the abductor mus-
cles of the hand. When subjects watched a video of a
hand with index finger abduction, the MEPs were greater
in the subjects own index finger, whereas when the
video showed movement of a hand with little finger ab-
duction, MEPs were greater in the little finger of the
observer. This is the standard ‘‘mirror’’ MEP effect. The
investigators then trained subjects to move their fingers
in a manner incongruent with the hand in the video:
Move little finger when index finger movement is shown
and vise versa. After training, MEPs were greater in the
little finger when index finger movement was observed,
and vise versa. ‘‘Mirror’’ effects can be trained simply by
sensory–motor association. The important implication of
this result is that study participants who exhibited in-
congruent MEP responses presumably did not mistake
the perception of index finger movement for little finger
movement and vise versa. This indicates that a prominent
indicator of human ‘‘mirror system’’ activity (Fadiga et al.,
1995) dissociates from action understanding.
It should not be surprising that measures of human
‘‘mirror system’’ function dissociate from action under-
standing, as we are fully capable of understanding ac-
tions we have never produced. For example, musically
untrained people can recognize, say, saxophone playing
even if they have never touched the instrument, just as
one can recognize actions of non-conspecifics (barking,
flying). Similarly, it would be surprising, maladaptive even,
if all observed actions resulted in the activation of the
exact same motor program in the observer. Indeed, most
sports would be impossible to play, as the observation of
an object-directed action (throwing a ball) would result in
the activation of the same action in the observer when
a very different action is required (catching or blocking).
The same is arguably true in many daily activities. The
results of Catmur et al. show that presumed mirror system
activity is not mirroring anything, but rather reflects
adaptive task-dependent sensory–motor associations.
6. Action Understanding and Action
Production Dissociate
As noted above, there is no evidence that deactivation of
the monkey mirror system disrupts action understand-
ing. The issue has been taken up in human research,
however, where there are now several published studies
that investigate action recognition. This work is suitable
for testing several predictions of the (human extrapo-
lated) mirror neuron theory of action understanding.
One such prediction is that action understanding and ac-
tion production should be strongly correlated. Although
it has been found that these two abilities can be corre-
lated in group studies, there is strong evidence that they
also are quite dissociable.
Several recent studies have investigated the issue. One
assessed a sample of 21 patients with limb apraxia and
found a strong correlation between gesture production
(imitation of meaningful gestures) and gesture recogni-
tion (determining which of two sequentially presented
gestures match a gesture name) (Buxbaum, Kyle, &
Menon, 2005). However, because the production mea-
sure has a perceptual component, a deficit affecting only
perception could lead to correlated deficits on the
recognition and production tasks. Further, the recogni-
tion task involved some form of working memory:
Subjects had to remember the gesture name (an audi-
torily and visually presented verb, e.g., ‘‘hammering’’),
and two gestures that were presented sequentially with
a 2-sec interstimulus interval. If working memory for
gestures recruits some form of motor-related rehearsal
component, as is the case for speech (Baddeley, 1992),
then both tasks shared a production component, which
may also have contributed to the correlation.
Another study (Pazzaglia, Smania, Corato, & Aglioti,
2008) also tested a sample of 21 patients with limb
apraxia and found a correlation (r (cid:3) .5) between a
gesture discrimination task (judging whether or not an
action is performed correctly) and a gesture production
task (asking subjects ‘‘to perform seven complex actions
that required the use of real objects,’’ p. 3031). How-
ever, a cluster analysis showed that while 14 of the 21
patients with limb apraxia had ‘‘a severe gesture rec-
ognition deficit,’’ 7 patients ‘‘presented with no deficit’’
(p. 3034), indicating that the two abilities are dissociable.
A third study (Tessari, Canessa, Ukmar, & Rumiati,
2007) of unselected left hemisphere damage patients
(n = 22) reported a weaker correlation between gesture
imitation and action (pantomime) recognition (r = .32;
again, not surprising because imitation involves a recog-
nition component), but no correlation between action
recognition and real object use (r = (cid:4).13), which
arguably provides a better assessment of ‘‘mirror sys-
tem’’ function. Importantly, double dissociations were
1236
Journal of Cognitive Neuroscience
Volume 21, Number 7
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
–
p
r
d
t
i
2
c
1
l
7
e
–
1
p
2
d
2
f
9
/
1
2
9
1
3
/
7
7
7
/
8
1
9
2
o
2
c
9
n
/
1
2
0
7
0
6
9
0
1
2
9
1
0
1
8
/
9
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
1
8
0
9
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
/
f
.
.
t
o
n
1
8
M
a
y
2
0
2
1
evident across patients in the latter relation: Case 23 per-
formed at 20% accuracy on action recognition, but 100%
on object use, whereas Case 15 performed at 100% accu-
racy on object recognition and 57% on object use. Other
cases showed similar dissociations.
Similar findings of group-level gesture perception–
production correlation, but case-level dissociations were
obtained by Negri et al. (2007). This study tested an un-
selected group of 37 patients with unilateral brain le-
sions on several tasks including pantomime recognition,
pantomime imitation, object use, and object recogni-
tion. Significant correlations were found between ob-
ject use and pantomime recognition (r = .58), object
use and object recognition (r = .37), pantomime imita-
tion and recognition (r = .59), and pantomime imitation
and object use (r = .79). However, despite these group
trends, subsets of patients demonstrated dissociations
between each of the correlated pairs of tests including
double dissociations between object use and panto-
mime recognition, and object use and object recogni-
tion. The authors of this study conclude that ‘‘. . .(a) The
ability to use objects is not necessary in order to be able
to recognize object-associated pantomimes; (b) the abil-
ity to imitate pantomimes is not necessary in order to be
able to recognize object-associated pantomimes; and (c)
the ability to use objects is not necessary in order to be
able to recognize objects’’ (p. 806).
Sign language provides additional evidence for the dis-
sociation between action production and action under-
standing. For example, Case ‘‘Gail D.’’ presented with
very severe deficits in sign language production asso-
ciated with a large left frontal lobe lesion, yet her com-
prehension of sign language was well-preserved (Poizner,
Klima, & Bellugi, 1987).
In summary, although gesture production and gesture
recognition can be correlated in groups of both apraxic
and unselected patients with focal brain lesions, these
abilities double dissociate, contrary to the prediction of
the mirror neuron theory of action understanding.
7. Damage to the Inferior Frontal Gyrus Is Not
Correlated with Action Understanding Deficits
If the human homologue of F5 is BA 44/6, then damage
to this region should result in action understanding defi-
cits. Available evidence does not support this predic-
tion. For example, based on earlier research, Heilman
et al. (1982) have argued that lesions to the parietal lobe
are associated with both production and comprehension
deficits, whereas frontal lesions produce only production
deficits. A mirror neuron proponent may counter that
the parietal lobe also contains mirror neurons, and thus,
the association between parietal
lobe damage and ac-
tion understanding deficits could be viewed as consistent
with ‘‘mirror system’’ claims. Following this line of ar-
gument, one would have to conclude that portions of
the mirror system that are more closely aligned with the
motor system, BA 44/6, do not support action under-
standing. This is clearly contrary to the central claim of
Rizzolatti et al. that it is motor representations that un-
derlie action understanding.
More recent studies using modern lesion analysis meth-
ods have provided mixed results regarding the anatomi-
cal correlate of action understanding deficits. One such
study (Buxbaum et al., 2005) confirmed earlier observa-
tions showing an association between deficits in object-
related gesture recognition and lesions to the inferior
lobe, whereas another study (Saygin, Wilson,
parietal
Dronkers, & Bates, 2004) reported that action compre-
hension is associated with lesions to BA 44/6/4. However,
this latter study examined a sample of aphasic patients
which may have biased their findings compared to studies
that use unselected patients or patients selected on the
basis of gesture-related deficits. Further, Saygin et al. did
not use dynamic actions for their stimuli, but rather static
pictures of pantomimed actions (the subject then pointed
to the pictured object that best fit the action). The relation
between action understanding in dynamic actions and
static actions is unknown, so interpretation of this study
is further compromised. However, it is relevant that de-
ficits in the understanding of linguistically specified ac-
tions (written phrases such as, ‘‘She is sweeping the. . .’’
followed by the same picture choices used in the ‘‘action’’
condition) dissociated behaviorally from understanding
of pictured actions, and were not associated with lesions
to BA 44/6/4, but with portions of the superior temporal
gyrus, insula, and inferior parietal lobe. One can conclude
from the behavioral and neural dissociation between pic-
tured actions and linguistically specific actions that what is
being mapped in this study, and associated with BA 44/6/4
in the picture condition, is not ‘‘action semantics,’’ as ac-
cess to this information is available via other routes. Thus,
this study provides evidence against the view that the
meaning of actions is encoded in motor representations
in motor cortex.
Another recent study (Pazzaglia et al., 2008) appears
to provide compelling evidence for an association be-
tween IFG damage and deficits in action understanding.
Lesions in patients with limb apraxia and gesture discrim-
ination deficits were compared with lesions in patients
with limb apraxia but without gesture discrimination defi-
cits. Subtraction of the lesions in these two groups of
patients identified the left IFG as being associated with
the limb apraxia plus gesture discrimination deficits. A
voxel-based lesion–symptom mapping analysis showed
the same result. However, an examination of the relation
between the amount of damaged tissue in the IFG and
gesture discrimination scores in the group of patients
who had gesture discrimination deficits showed no rela-
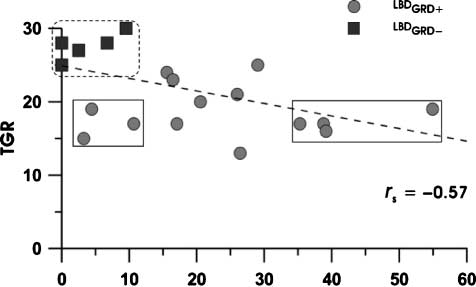
tion (Figure 1, circles). For example, the four patients
with the most IFG involvement (Figure 1, right solid rect-
angle) had gesture discrimination scores that are in-
distinguishable from the three patients with the least
IFG involvement (left solid rectangle), and the latter are
Hickok
1237
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
–
p
r
d
t
i
2
c
1
l
7
e
–
1
p
2
d
2
f
9
/
1
2
9
1
3
/
7
7
7
/
8
1
9
2
o
2
c
9
n
/
1
2
0
7
0
6
9
0
1
2
9
1
0
1
8
/
9
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
1
8
0
9
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
t
.
.
f
.
/
o
n
1
8
M
a
y
2
0
2
1
action judgments, vPMc stimulation yielded longer reac-
tion times than EBA stimulation, and the reverse held for
form judgments, longer reaction times for EBA stimula-
tion than vPMc stimulation. Stimulation had no effect on
accuracy. In the other study (Urgesi, Calvo-Merino, et al.,
2007), which seemed to involve more difficult stimuli
and only asked subjects to judge body configuration, an
effect of accuracy was observed with vPMc stimulation
associated with more errors on the configuration match-
ing task than with EBA stimulation. Oddly, there were
no reaction time effects.
Thus, two studies show that interference stimulation to
vPMc negatively affects performance on a body configu-
ration delayed matched-to-sample task. Again, because
these studies did not assess action understanding, they
cannot speak to the question of whether the mirror sys-
tem supports action understanding. However, they do
suggest that processing of body configurations at least in
the delayed match-to-sample task involves vPMc to some
extent. Given that the tasks involved working memory,
it seems possible that this region may support some sort
of working memory for body configurations. This is con-
sistent with many claims regarding the sensory–motor
nature of working memory systems (Pa, Wilson, Pickell,
Bellugi, & Hickok, in press; Buchsbaum & D’Esposito,
2008; Postle, 2006; Hickok, Buchsbaum, Humphries, &
Muftuler, 2003; Ruchkin et al., 2003; Wilson, 2001).
More work is needed to characterize the neural basis
of ‘‘action understanding.’’ Available evidence, however,
leads us to conclude that the IFG does not play a cen-
tral role.
8. Generalization of the Mirror System to Speech
Recognition Fails on Empirical Grounds
Mirror neuron function has been generalized to speech
perception from the earliest reports (Rizzolatti & Arbib,
1998; Gallese et al., 1996). Basing their speculation on
the motor theory of speech perception (Liberman &
Mattingly, 1985; Liberman et al., 1967), Rizzolatti et al.
suggested that mirror neurons may underlie the per-
ception of speech gestures. The motor theory of speech
perception had been all but abandoned among the
majority of speech scientists when mirror neurons were
discovered, but has enjoyed a healthy revival since. How-
ever, there is exceptionally strong evidence against the
motor theory of speech perception, and consequently,
the mirror neuron generalization of action understand-
ing to the speech domain.
A motor theory of speech perception makes a very
clear and strong prediction. Damage to the motor speech
areas should produce deficits in speech recognition. How-
ever, damage to motor speech areas, evidenced in many
cases by large left frontal lesions and severe speech pro-
duction deficits, do not typically lead to speech recog-
nition deficits. Paul Broca’s original case, Leborne,
is
representative of this pattern in that the patient could
Figure 1. Scatterplot showing the relation between gesture
recognition and the amount of lesioned tissue in the IFG of left
hemisphere damaged patients with apraxia. Dark square points are
patients without gesture recognition deficits; lighter circle points
are patients with gesture recognition deficits. Solid rectangles are
aligned on the y-axis and show that patients at the extremes of the
distribution of IFG tissue damage have indistinguishable scores
on gesture recognition. Dotted rectangle outlines patients without
gesture recognition deficits for comparison. Figure modified from
Pazzaglia et al. (2008). TGR = Transitive Gesture Recognition score.
themselves well within the distribution of patients with-
out gesture discrimination deficits (left dashed rectan-
gle) in terms of the amount of IFG involvement. Clearly,
IFG involvement is not predicting gesture discrimina-
tion performance. It is unclear why the lesion subtrac-
tion and voxel-based mapping analyses identified the IFG
in this study, but the fact that these analyses were cal-
culated using a measure that was not corrected for re-
sponse bias may be a factor (the task was implemented
using a signal detection paradigm, but percent correct
rather than the bias-corrected d0 statistic was used for
lesion analyses).
Two recent rTMS experiments have studied the ef-
fects of functional disruption of ventral premotor cortex
(vPMc) on visual discrimination of action-related pictures
(Urgesi, Calvo-Merino, Haggard, & Aglioti, 2007; Urgesi,
Candidi, Ionta, & Aglioti, 2007). In both of these stud-
ies, subjects were asked to make two-choice, match-to-
sample judgments: A picture of a body configuration was
presented (the sample) followed by a mask (500 msec),
and then a picture of two body configurations; the sub-
ject was asked to indicate which of the two matched the
sample. First, it is important to note that neither of these
studies actually tested action understanding. That is, dis-
crimination performance did not depend on understand-
ing the meaning of the actions, but could be performed
based on configural information alone. One study (Urgesi,
Candidi, et al., 2007) compared the effects of interference
stimulation of vPMc with interference stimulation of a
ventral temporal–occipital location (the extrastriate body
area, EBA) during action discrimination (which action
matches the sample?) versus form discrimination (which
actor matches the sample, independent of action?). For
1238
Journal of Cognitive Neuroscience
Volume 21, Number 7
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
–
p
r
d
t
i
2
c
1
l
7
e
–
1
p
2
d
2
f
9
/
1
2
9
1
3
/
7
7
7
/
8
1
9
2
o
2
c
9
n
/
1
2
0
7
0
6
9
0
1
2
9
1
0
1
8
/
9
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
1
8
0
9
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
t
.
/
.
f
.
o
n
1
8
M
a
y
2
0
2
1
produce little more than the syllable ‘‘tan’’ yet ‘‘under-
stood almost all that was said to him’’ (p. 63) (Broca,
1861/1960). Much subsequent work has confirmed the
pattern at least at the single word level (Goodglass,
Kaplan, & Barresi, 2001; Goodglass, 1993).5 For exam-
ple, a recent study reported that Broca’s aphasics (n =
9) were indistinguishable from control subjects on an
auditory word comprehension test involving 236 items
(Moineau, Dronkers, & Bates, 2005). Lesions associated
with Broca’s aphasia tend to be relatively large, involving
most of the lateral frontal lobe, motor cortex, and ante-
rior insula but often also extending posteriorly to include
the parietal
lobe (Dronkers, Redfern, & Knight, 2000;
Damasio, 1991, 1992); thus the entire left hemisphere
‘‘mirror system’’ can be affected in Broca’s aphasia. A
motor theory of speech recognition has no explanation
for the existence of a syndrome such as Broca’s aphasia.
Although proponents of motor theories of speech rec-
ognition typically completely ignore the speech com-
prehension abilities of Broca’s aphasics—for instance, a
recent review of the motor theory of speech perception
failed to even mention the syndrome (Galantucci et al.,
2006)—it is more often noted by motor theorists that
Broca’s aphasics can be impaired on syllable discrimi-
nation tasks, i.e., the ability to judge whether pairs of
nonsense syllables are the same (/ba/–/ba/) or different
(/ba/–/da/) (Blumstein, 1995). Although this would ap-
pear to provide evidence favoring a motor theory of
speech recognition, it does not. Performance on such
tasks doubly dissociates from measures of auditory com-
prehension. For example, Miceli, Gainotti, Caltagirone,
and Masullo (1980) report 19 patients who were im-
paired relative to controls on a syllable discrimination
task, yet performed at 100% accuracy in matching a spo-
ken word (e.g., ‘‘bear’’) to a picture presented along
with three foil pictures, one phonemically related (e.g.,
PEAR), one semantically related (e.g., MOOSE), and one
unrelated (e.g., GRAPES). Another nine patients in that
study showed the reverse pattern of performance; they
were impaired relative to controls on the auditory com-
prehension task, but performed normally on the syllable
discrimination task. Patients with nonfluent speech pro-
duction deficits, such as Broca’s aphasics, are typically
the most impaired on syllable discrimination tasks (Basso,
Casati, & Vignolo, 1977) (Figure 2). This indicates that
syllable discrimination tasks are tapping some ability, or
abilities (e.g., working memory, executive, or attentional
processes), that is/are not necessary for normal, ecologi-
cally valid speech recognition, and therefore, are not valid
measures for assessing speech recognition (see Hickok
& Poeppel, 2000, 2004, 2007, for review and extensive
discussion).
Another syndrome that clearly demonstrates the disso-
ciability of motor speech functions and speech understand-
ing is mixed transcortical aphasia, sometimes referred to
as ‘‘isolation of the speech zone’’ (Bogousslavsky, Regli,
& Assal, 1988; Geschwind, Quadfasel, & Segarra, 1968).
This syndrome is characterized by a severe deficit in the
comprehension of speech, despite the well-preserved
ability to repeat heard speech, sometimes compulsively.
The syndrome is associated with damage to left fron-
tal and posterior parietal regions but with sparing of
peri-sylvian speech-related areas such as Broca’s area,
superior temporal gyrus, and the tissue in between.
This pattern of damage leaves sensory–motor functions
of speech intact (explaining repetition ability), while
apparently disrupting systems involved in mapping
speech onto conceptual–semantic representations. This
is the reverse dissociation compared to Broca’s apha-
sia, and indicates that preservation of motor speech
functions is neither necessary nor sufficient for speech
understanding.
In short, data from lesion studies of speech processing
unequivocally demonstrate that the motor theory/mirror
neuron theory of speech perception is incorrect in any
strong form. This is not to say that sensory–motor circuits
cannot contribute to speech recognition. Top–down pro-
cesses initiated in any frontal circuit (not just motor) may
be able to influence speech recognition to some extent
via sensory–motor circuits. This may be particularly useful
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
–
p
r
d
t
i
2
c
1
l
7
e
–
1
p
2
d
2
f
9
/
1
2
9
1
3
/
7
7
7
/
8
1
9
2
o
2
c
9
n
/
1
2
0
7
0
6
9
0
1
2
9
1
0
1
8
/
9
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
1
8
0
9
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
f
.
.
/
.
t
Figure 2. Reconstructed
brain lesions in two patients
from Caplan, Gow, and
Makris (1995). Case R. Wi. is
a Broca’s aphasic with good
auditory comprehension (by
definition), whereas Case M. L.
is a Wernicke’s aphasic with
relatively poor comprehension
(by definition). On a syllable
discrimination task, R. Wi.
performed at 72% correct
(A0 = 0.72), whereas M. L.
performed much better at
90% correct (A0 = 0.90).
Figure modified from Caplan
et al. (1995).
o
n
1
8
M
a
y
2
0
2
1
Hickok
1239
under noisy listening conditions (Moineau et al., 2005).
However, this influence is modulatory, not primary.
ing is dependent on the motor system. This is a desirable
result and deserves empirical evaluation.
WHAT ROLE DOES THE ‘‘MIRROR SYSTEM’’
PLAY IN ACTION UNDERSTANDING?
The evidence reviewed above shows that action under-
standing can doubly dissociate from ‘‘mirror system’’ func-
tion, both in the domain of manual gesture and speech.
Clearly then, the ‘‘mirror system’’ is not the basis for ac-
tion understanding.
What does the mirror system reflect? There are two
possibilities, as suggested by Mahon and Caramazza (2008)
in their critical analysis of ‘‘embodied cognition’’ hy-
potheses, generally. One is that it reflects pure Pavlovian
association. Pair a tone with a puff of air to the eye and
pretty soon the tone alone will elicit a blink response.
This motor response to a sensory event does not indi-
cate that the blink response is coding the meaning of
the tone. Perhaps the activity of mirror neurons simply
reflects sensory–motor pairings. The fact that ‘‘mirror
system’’ activity can be dynamically remapped with train-
ing (Catmur et al., 2007) lends some support for this
idea. Mahon and Caramazza suggest another possibil-
ity. Concepts, including action-related concepts, involve
an abstract level of representation that is distinct from
sensory–motor systems. These abstract representations
are sufficient for recognition, but can be associated with
related sensory–motor information that ‘‘colors concep-
tual processing, enriches it, and provides it with a rela-
tional context’’ (p. 68). Thus, one can have a perfectly
viable concept of ‘‘saxophone playing’’ without ever hav-
ing touched such an instrument, and without the con-
cept being tied to a specific sensory–motor situation
(e.g., the concept may apply equally well to playing
an alto or tenor sax, a toy instrument, or to mimicking
saxophone playing on some other object). However,
knowledge of how to grasp a saxophone, finger the
keys, and position one’s mouth on the mouthpiece, can,
according to Mahon and Caramazza, augment the ab-
stract concept by providing a specific sensory–motor
association. This information might even lead to a dif-
ferent ‘‘understanding’’ of a saxophone-playing action;
for example, in a situation where the player is holding
the instrument improperly, the observer with sensory–
motor experience with a saxophone might recognize
that the player is not an expert, whereas someone with-
out such experience may not be able to access this
‘‘enriched’’ knowledge. Or in other situations, sensory–
motor knowledge may allow the observer to generate
predictions about subsequent actions that could in-
fluence sensory systems in a top–down fashion and
facilitate subsequent perceptual recognition. The view
promoted by Mahon and Caramazza admits that motor
knowledge can influence or augment action ‘‘under-
standing’’ to some degree, but without committing to
the empirically untenable position that action understand-
Although it seems entirely possible that motor expe-
rience can augment conceptual understanding in some
situations, in others, mirror-like activity appears to re-
flect sensory–motor associations that are devoid of mea-
ningful conceptual content. ‘‘Mirror system’’ activity that
has been observed during the imitation of meaningless
gestures (Iacoboni et al., 1999) is one such situation. The
demonstration that ‘‘mirror activity’’ associated with view-
ing actions can be remapped such that it becomes
associated with a completely different action (Catmur
et al., 2007) is another. Thus, perhaps both of the pos-
sibilities raised by Mahon and Caramazza apply to the
‘‘mirror system.’’
CONCLUSION
Mirror neurons are a fascinating class of cells that de-
serve to be thoroughly investigated in the monkey,
and explored systematically for possible homologues in
humans. The early hypothesis that these cells under-
lie action understanding is likewise an interesting and
prima facie reasonable idea. However, despite its wide-
spread acceptance, the proposal has never been ade-
quately tested in monkeys, and in humans there is strong
empirical evidence, in the form of physiological and neuro-
psychological (double) dissociations, against the claim.
Why does the hypothesis remain prominent, indeed
all but accepted as fact, despite solid evidence to the
contrary? I suggest that Pillsbury was right. Motor theo-
ries are simple and easy to understand: ‘‘we understand
action because the motor representation of that action
is activated in our brain’’ (Rizzolatti et al., 2001, p. 661).
We see someone pouring liquid from a bottle into a
glass; this activates a motor representation associated
with our own liquid-pouring experiences, and voila`, we
have understanding. But scratch the surface of action
understanding and it is immediately clear that the prob-
lem is not that simple (Pinker, 1989, 2007). For example,
the motor act of pouring liquid from a bottle into a glass
could be understood as pouring,
filling, emptying,
tipping, rotating, inverting, spilling (if the liquid missed
its mark), defying/ignoring/rebelling (if the pourer was
instructed not to pour), and so on. A motor representa-
tion cannot distinguish between the range of possible
meanings associated with such an action. A mirror neu-
ron theorist might protest that it is the goal or inten-
tion that is coded by mirror neurons, not the specific
actions (Fogassi et al., 2005). But a goal, say to fill a glass
with water, can be accomplished with any number of
individual actions or sequence of actions: pouring from
a pitcher, turning a spigot, dipping the glass in a lake,
setting the glass in the rain, positioning an array of leaves
to collect and funnel dew into the glass, digging a well
and pumping water into the glass, or even commanding
someone else to do any of these! Given the range of
1240
Journal of Cognitive Neuroscience
Volume 21, Number 7
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
–
p
r
d
t
i
2
c
1
l
7
e
–
1
p
2
d
2
f
9
/
1
2
9
1
3
/
7
7
7
/
8
1
9
2
o
2
c
9
n
/
1
2
0
7
0
6
9
0
1
2
9
1
0
1
8
/
9
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
1
8
0
9
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
/
.
t
.
.
f
o
n
1
8
M
a
y
2
0
2
1
meanings associated with a specific action and the range
of actions that can achieve a specific goal, there must be
a clear distinction between goals and the motor routines
that are implemented in a given circumstance to achieve
those goals. If mirror neurons are reflecting goals and not
actions, then a statement about mirror neurons such as,
‘‘we understand action because the motor representation
of that action is activated in our brain’’ (Rizzolatti et al.,
2001, p. 661) is either false because mirror neurons do
not code actions, or it is false because motor representa-
tions are not the basis of action understanding.
Unfortunately, more than 10 years after their discov-
ery, little progress has been made in understanding the
function of mirror neurons. I submit that this is a direct
result of an overemphasis on the action understanding
theory, which has distracted the field away from investi-
gating other possible (and potentially equally important)
functions.
Acknowledgments
I thank Richard Ivry, Steven Pinker, David Poeppel, and Stephen
Wilson for invaluable comments on an earlier draft of this article.
This work was supported by NIH grant no. DC0361.
Reprint requests should be sent to Gregory Hickok, Department
of Cognitive Sciences, Center for Cognitive Neuroscience, Univer-
sity of California, Irvine, CA 92697, or via e-mail: greg.hickok@
uci.edu.
Notes
Some theorists have suggested that object recognition is
1.
dependent on action-related motor systems (Gallese & Lakoff,
2005). This theoretical position will not be discussed here, but
see Negri et al. (2007) and Mahon & Caramazza (2005) for a
critical evaluation.
2. As Richard Ivry has pointed out (personal communication),
the ‘‘sensory’’ activity in F5 may be explained rather straight-
forwardly in terms of motor priming. Suppose action concepts
are represented upstream to F5. In self-generated movement,
the links between an action concept and its associated motor
code in F5 become activated. During object or action observa-
tion, these links are automatically reactivated, primed, as a result
of their prior association. So on this view, ‘‘sensory’’ activity in
F5 cells need not even involve a mechanism to access a motor
vocabulary, but rather may be the motor reflection of that access
process.
3. A reviewer suggested that this argument is ‘‘totally non-
sense’’ and suggested instead that the finding may indicate ‘‘that
only 15% of mirror neurons code the meaning of the perceived
action also on the basis of its sound and not only on the basis of
its visible outcome.’’ Let me be clear: This is not my argument. It
is Rizzolatti and colleagues’. If the claim is that the 15% of mirror
neurons respond to action-related sounds because they are cod-
ing the abstract meaning of the action irrespective of the sensory
input, ‘‘audiovisual mirror neurons code abstract contents, the
meaning of actions’’ (Kohler et al., 2002, p. 846)—then it follows
that the remaining 85% are not coding the abstract meaning,
but rather something sensory specific.
4. For example, the strongest evidence to date for the exis-
tence of mirror neurons in humans comes from a study (Chong
et al., 2008) that used fMRI to assess adaptation (repetition
suppression) across gesture execution–observation tasks. This
study reported an adaptation effect in the right parietal lobe
using pantomimed gestures—a stimulus that does not activate
macaque mirror neurons. The location of the effect is also
puzzling in that it is inconsistent with human data from apraxia
which typically is associated with left hemisphere disease (see
below).
5. Broca’s patients often have some comprehension difficulty
at the sentence level. However, these deficits are primarily re-
stricted to sentences in which successful comprehension de-
pends on accurate syntactic analysis (e.g., ‘‘He showed her
the baby pictures’’ vs. ‘‘He showed her baby the pictures’’).
If lexical information provides clues to correct interpretation
(‘‘The apple that the boy ate was red’’), Broca’s aphasics usu-
ally perform well
in comprehension assessments, providing
further evidence for well-preserved word-level comprehension
(Caramazza & Zurif, 1976).
REFERENCES
Baddeley, A. D. (1992). Working memory. Science, 255,
556–559.
Basso, A., Casati, G., & Vignolo, L. A. (1977). Phonemic
identification defects in aphasia. Cortex, 13, 84–95.
Berkeley, G. (1709). An essay towards a new theory of
vision. Dublin: Pepyat.
Blumstein, S. (1995). The neurobiology of the sound
structure of language. In M. S. Gazzaniga (Ed.), The
cognitive neurosciences (pp. 913–929). Cambridge,
MA: MIT Press.
Bogousslavsky, J., Regli, F., & Assal, G. (1988). Acute
transcortical mixed aphasia. A carotid occlusion syndrome
with pial and watershed infarcts. Brain, 111, 631–641.
Broca, P. (1861/1960). Remarks on the seat of the faculty
of articulate language, followed by an observation of
aphemia. In G. von Bonin (Ed.), Some papers on the
cerebral cortex. Oxford: Blackwell Scientific Publications.
Buccino, G., Lui, F., Canessa, N., Patteri, I., Lagravinese, G.,
Benuzzi, F., et al. (2004). Neural circuits involved in the
recognition of actions performed by nonconspecifics:
An fMRI study. Journal of Cognitive Neuroscience, 16,
114–126.
Buchsbaum, B. R., & D’Esposito, M. (2008). The search for
the phonological store: From loop to convolution. Journal
of Cognitive Neuroscience, 20, 762–778.
Buxbaum, L. J., Kyle, K. M., & Menon, R. (2005). On beyond
mirror neurons: Internal representations subserving
imitation and recognition of skilled object-related actions
in humans. Brain Research, Cognitive Brain Research,
25, 226–239.
Caplan, D., Gow, D., & Makris, N. (1995). Analysis of lesions
by MRI in stroke patients with acoustic–phonetic processing
deficits. Neurology, 45, 293–298.
Caramazza, A., & Zurif, E. B. (1976). Dissociation of algorithmic
and heuristic processes in sentence comprehension:
Evidence from aphasia. Brain and Language, 3, 572–582.
Catmur, C., Walsh, V., & Heyes, C. (2007). Sensorimotor
learning configures the human mirror system. Current
Biology, 17, 1527–1531.
Chong, T. T., Cunnington, R., Williams, M. A., Kanwisher, N.,
& Mattingley, J. B. (2008). fMRI adaptation reveals mirror
neurons in human inferior parietal cortex. Current Biology,
18, 1576–1580.
Damasio, A. R. (1992). Aphasia. New England Journal of
Medicine, 326, 531–539.
Damasio, H. (1991). Neuroanatomical correlates of the
aphasias. In M. Sarno (Ed.), Acquired aphasia (2nd ed.,
pp. 45–71). San Diego, CA: Academic Press.
Hickok
1241
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
–
p
r
d
t
i
2
c
1
l
7
e
–
1
p
2
d
2
f
9
/
1
2
9
1
3
/
7
7
7
/
8
1
9
2
o
2
c
9
n
/
1
2
0
7
0
6
9
0
1
2
9
1
0
1
8
/
9
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
1
8
0
9
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
/
.
.
f
t
o
n
1
8
M
a
y
2
0
2
1
Decety, J., Grezes, J., Costes, N., Perani, D., Jeannerod, M.,
Procyk, E., et al. (1997). Brain activity during observation
of actions: Influence of action content and subject’s
strategy. Brain, 120, 1763–1777.
di Pellegrino, G., Fadiga, L., Fogassi, L., Gallese, V., &
Rizzolatti, G. (1992). Understanding motor events: A
neurophysiological study. Experimental Brain Research,
91, 176–180.
Dinstein, I. (2008). Human cortex: Reflections of mirror
neurons. Current Biology, 18, R956–R959.
Dinstein, I., Hasson, U., Rubin, N., & Heeger, D. J. (2007).
Brain areas selective for both observed and executed
movements. Journal of Neurophysiology, 98, 1415–1427.
Dinstein, I., Thomas, C., Behrmann, M., & Heeger, D. J. (2008).
A mirror up to nature. Current Biology, 18, R13–R18.
Dronkers, N. F., Redfern, B. B., & Knight, R. T. (2000). The
neural architecture of language disorders. In M. S. Gazzaniga
(Ed.), The new cognitive neurosciences (pp. 949–958).
Cambridge, MA: MIT Press.
Fadiga, L., Fogassi, L., Pavesi, G., & Rizzolatti, G. (1995).
Motor facilitation during action observation: A magnetic
stimulation study. Journal of Neurophysiology, 73,
2608–2611.
Ferrari, P. F., Rozzi, S., & Fogassi, L. (2005). Mirror neurons
responding to observation of actions made with tools in
monkey ventral premotor cortex. Journal of Cognitive
Neuroscience, 17, 212–226.
Ferrari, P. F., Visalberghi, E., Paukner, A., Fogassi, L.,
Ruggiero, A., & Suomi, S. J. (2006). Neonatal imitation
in rhesus macaques. PLoS Biology, 4, e302.
Fogassi, L., Ferrari, P. F., Gesierich, B., Rozzi, S., Chersi, F.,
& Rizzolatti, G. (2005). Parietal lobe: From action
organization to intention understanding. Science, 308,
662–667.
Fogassi, L., Gallese, V., Buccino, G., Craighero, L., Fadiga, L.,
& Rizzolatti, G. (2001). Cortical mechanism for the visual
guidance of hand grasping movements in the monkey:
A reversible inactivation study. Brain, 124, 571–586.
Fuster, J. M. (1995). Memory in the cerebral cortex.
Cambridge, MA: MIT Press.
Gainotti, G., & Lemmo, M. (1976). Comprehension of
symbolic gestures in aphasia. Brain and Language, 3,
451–460.
Galantucci, B., Fowler, C. A., & Turvey, M. T. (2006). The
motor theory of speech perception reviewed. Psychonomic
Bulletin & Review, 13, 361–377.
Gallese, V., Fadiga, L., Fogassi, L., & Rizzolatti, G. (1996).
Action recognition in the premotor cortex. Brain, 119,
593–609.
Gallese, V., Fogassi, L., Fadiga, L., & Rizzolati, G. (2002).
Action representation and the inferior parietal lobule.
In W. Prinz & B. Hommel (Eds.), Attention & performance:
XIX. Common mechanisms in perception and action
(pp. 247–266). Oxford: Oxford University Press.
Gallese, V., & Lakoff, G. (2005). The brain’s concepts: The
role of the sensory–motor system in conceptual knowledge.
Cognitive Neuropsychology, 22, 455–479.
Geschwind, N., Quadfasel, F. A., & Segarra, J. M. (1968).
Isolation of the speech area. Neuropsychologia, 6, 327–340.
Goodglass, H. (1993). Understanding aphasia. San Diego:
Academic Press.
Heilman, K. M., Rothi, L. J., & Valenstein, E. (1982). Two
forms of ideomotor apraxia. Neurology, 32, 342–346.
Hickok, G., Buchsbaum, B., Humphries, C., & Muftuler, T.
(2003). Auditory–motor interaction revealed by fMRI:
Speech, music, and working memory in area Spt. Journal
of Cognitive Neuroscience, 15, 673–682.
Hickok, G., & Poeppel, D. (2000). Towards a functional
neuroanatomy of speech perception. Trends in Cognitive
Sciences, 4, 131–138.
Hickok, G., & Poeppel, D. (2004). Dorsal and ventral streams:
A framework for understanding aspects of the functional
anatomy of language. Cognition, 92, 67–99.
Hickok, G., & Poeppel, D. (2007). The cortical organization
of speech processing. Nature Reviews Neuroscience, 8,
393–402.
Iacoboni, M., Woods, R. P., Brass, M., Bekkering, H., Mazziotta,
J. C., & Rizzolatti, G. (1999). Cortical mechanisms of
human imitation. Science, 286, 2526–2528.
Kohler, E., Keysers, C., Umilta, M. A., Fogassi, L., Gallese, V.,
& Rizzolatti, G. (2002). Hearing sounds, understanding
actions: Action representation in mirror neurons. Science,
297, 846–848.
Koski, L., Iacoboni, M., Dubeau, M. C., Woods, R. P., &
Mazziotta, J. C. (2003). Modulation of cortical activity
during different imitative behaviors. Journal of
Neurophysiology, 89, 460–471.
Koski, L., Wohlschlager, A., Bekkering, H., Woods, R. P.,
Dubeau, M. C., Mazziotta, J. C., et al. (2002). Modulation
of motor and premotor activity during imitation of
target-directed actions. Cerebral Cortex, 12, 847–855.
Krams, M., Rushworth, M. F., Deiber, M. P., Frackowiak, R. S.,
& Passingham, R. E. (1998). The preparation, execution
and suppression of copied movements in the human
brain. Experimental Brain Research, 120, 386–398.
Liberman, A. M., Cooper, F. S., Shankweiler, D. P., &
Studdert-Kennedy, M. (1967). Perception of the speech
code. Psychological Review, 74, 431–461.
Liberman, A. M., & Mattingly, I. G. (1985). The motor theory
of speech perception revised. Cognition, 21, 1–36.
Mahon, B. Z., & Caramazza, A. (2005). The orchestration of
the sensory–motor systems: Clues from neuropsychology.
Cognitive Neuropsychology, 22, 480–494.
Mahon, B. Z., & Caramazza, A. (2008). A critical look at the
embodied cognition hypothesis and a new proposal for
grounding conceptual content. Journal of Physiology
(Paris), 102, 59–70.
Miceli, G., Gainotti, G., Caltagirone, C., & Masullo, C. (1980).
Some aspects of phonological impairment in aphasia.
Brain and Language, 11, 159–169.
Moineau, S., Dronkers, N. F., & Bates, E. (2005). Exploring
the processing continuum of single-word comprehension
in aphasia. Journal of Speech, Language, and Hearing
Research, 48, 884–896.
Morin, O., & Grezes, J. (2008). What is ‘‘mirror’’ in the
premotor cortex? A review. Neurophysiologie Clinique,
38, 189–195.
Negri, G. A., Rumiati, R. I., Zadini, A., Ukmar, M., Mahon,
B. Z., & Caramazza, A. (2007). What is the role of
motor simulation in action and object recognition?
Evidence from apraxia. Cognitive Neuropsychology,
24, 795–816.
Goodglass, H., Kaplan, E., & Barresi, B. (2001). The assessment
of aphasia and related disorders (3rd ed.). Philadelphia:
Lippincott Williams & Wilkins.
Grezes, J., Costes, N., & Decety, J. (1998). Top–down effect
of strategy on the perception of human biological motion:
A PET investigation. Cognitive Neuropsychology, 15,
553–582.
Nelissen, K., Luppino, G., Vanduffel, W., Rizzolatti, G., &
Orban, G. A. (2005). Observing others: Multiple action
representation in the frontal lobe. Science, 310, 332–336.
Oberman, L. M., Hubbard, E. M., McCleery, J. P., Altschuler,
E. L., Ramachandran, V. S., & Pineda, J. A. (2005). EEG
evidence for mirror neuron dysfunction in autism spectrum
disorders. Cognitive Brain Research, 24, 190–198.
1242
Journal of Cognitive Neuroscience
Volume 21, Number 7
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
–
p
r
d
t
i
2
c
1
l
7
e
–
1
p
2
d
2
f
9
/
1
2
9
1
3
/
7
7
7
/
8
1
9
2
o
2
c
9
n
/
1
2
0
7
0
6
9
0
1
2
9
1
0
1
8
/
9
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
1
8
0
9
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
t
f
.
.
/
.
o
n
1
8
M
a
y
2
0
2
1
Pa, J., Wilson, S. M., Pickell, B., Bellugi, U., & Hickok, G.
(in press). Neural organization of linguistic short-term
memory is sensory modality-dependent: Evidence from
signed and spoken language. Journal of Cognitive
Neuroscience.
Pasternak, T., & Greenlee, M. W. (2005). Working memory
in primate sensory systems. Nature Reviews Neuroscience,
6, 97–107.
Pazzaglia, M., Smania, N., Corato, E., & Aglioti, S. M. (2008).
Neural underpinnings of gesture discrimination in patients
with limb apraxia. Journal of Neuroscience, 28, 3030–3041.
Perrett, D. I., Mistlin, A. J., Harries, M. H., & Chitty, A. J. (1990).
Understanding the visual appearance and consequence of
hand actions. In M. A. Goodale (Ed.), Vision and action:
The control of grasping (pp. 163–180). Norwood, NJ: Ablex.
Perrett, D. I., Smith, P. A. J., Mistlin, A. J., Chitty, A. J., Head,
A. S., Potter, D. D., et al. (1985). Visual analysis of body
movements by neurones in the temporal cortex of the
macaque monkey: A preliminary report. Behavioural
Brain Research, 16, 153–170.
Pillsbury, W. B. (1911). The place of movement in
consciousness. Psychological Review, 18, 83–99.
Pinker, S. (1989). Learnability and cognition: The acquisition
of argument structure. Cambridge, MA: MIT Press.
Pinker, S. (2007). The stuff of thought: Language as a
window into human nature. New York: Viking.
Poizner, H., Klima, E. S., & Bellugi, U. (1987). What the
hands reveal about the brain. Cambridge, MA: MIT Press.
Postle, B. R. (2006). Working memory as an emergent property
of the mind and brain. Neuroscience, 139, 23–38.
Rizzolatti, G., & Arbib, M. (1998). Language within our grasp.
Trends in Neurosciences, 21, 188–194.
Rizzolatti, G., Camarda, R., Fogassi, L., Gentilucci, M.,
Luppino, G., & Matelli, M. (1988). Functional organization
of inferior area 6 in the macaque monkey: II. Area F5 and
the control of distal movements. Experimental Brain
Research, 71, 491–507.
Rizzolatti, G., & Craighero, L. (2004). The mirror-neuron
system. Annual Review of Neuroscience, 27, 169–192.
Rizzolatti, G., Fadiga, L., Matelli, M., Bettinardi, V., Paulesu, E.,
Perani, D., et al. (1996). Localization of grasp representations
in humans by PET: 1. Observation versus execution.
Experimental Brain Research, 111, 246–252.
Rizzolatti, G., Fogassi, L., & Gallese, V. (2001). Neurophysiological
mechanisms underlying the understanding and imitation
of action. Nature Reviews Neuroscience, 2, 661–670.
Ruchkin, D. S., Grafman, J., Cameron, K., & Berndt, R. S.
(2003). Working memory retention systems: A state of
activated long-term memory. Behavioral and Brain
Sciences, 26, 709–777.
Saygin, A. P., Wilson, S. M., Dronkers, N. F., & Bates, E.
(2004). Action comprehension in aphasia: Linguistic and
non-linguistic deficits and their lesion correlates.
Neuropsychologia, 42, 1788–1804.
Scheerer, E. (1984). Motor theories of cognitive structure:
A historical review. In W. Prinz & A. F. Sanders (Eds.),
Cognition and motor processes (pp. 77–98). Berlin:
Springer-Verlag.
Schubotz, R. I., Sakreida, K., Tittgemeyer, M., & von Cramon,
D. Y. (2004). Motor areas beyond motor performance:
Deficits in serial prediction following ventrolateral
premotor lesions. Neuropsychology, 18, 638–645.
Schubotz, R. I., & von Cramon, D. Y. (2004). Sequences of
abstract nonbiological stimuli share ventral premotor
cortex with action observation and imagery. Journal of
Neuroscience, 24, 5467–5474.
Tessari, A., Canessa, N., Ukmar, M., & Rumiati, R. I. (2007).
Neuropsychological evidence for a strategic control of
multiple routes in imitation. Brain, 130, 1111–1126.
Tkach, D., Reimer, J., & Hatsopoulos, N. G. (2007). Congruent
activity during action and action observation in motor
cortex. Journal of Neuroscience, 27, 13241–13250.
Umilta`, M., Kohler, E., Gallese, V., Fogassi, L., Fadiga, L.,
Keysers, C., et al. (2001). I know what you are doing.
A neurophysiological study. Neuron, 31, 155–165.
Urgesi, C., Calvo-Merino, B., Haggard, P., & Aglioti, S. M.
(2007). Transcranial magnetic stimulation reveals two
cortical pathways for visual body processing. Journal
of Neuroscience, 27, 8023–8030.
Urgesi, C., Candidi, M., Ionta, S., & Aglioti, S. M. (2007).
Representation of body identity and body actions in
extrastriate body area and ventral premotor cortex.
Nature Neuroscience, 10, 30–31.
Visalberghi, E., & Fragaszy, D. (2001). Do monkeys ape?
Ten years after. In K. Dautenhahn & C. L. Nehaniv (Eds.),
Imitation in animals and artifacts. Cambridge, MA:
MIT Press.
Washburn, M. F. (1914). The function of incipient motor
processes. Psychological Review, 21, 376–390.
Washburn, M. F. (1916). Movement and mental imagery:
Outlines of a motor theory of the complexer mental
processes. Boston: Houghton Mifflin.
Watson, J. B. (1913). Psychology as the behaviorist views it.
Psychological Review, 20, 158–177.
Weinrich, M., Wise, S. P., & Mauritz, K. H. (1984). A
neurophysiological study of the premotor cortex in the
rhesus monkey. Brain, 107, 385–414.
Wilson, M. (2001). The case for sensorimotor coding in
working memory. Psychonomic Bulletin & Review, 8,
44–57.
Wilson, S. M., Saygin, A. P., Sereno, M. I., & Iacoboni, M.
(2004). Listening to speech activates motor areas involved
in speech production. Nature Neuroscience, 7, 701–702.
Wise, S. P., & Mauritz, K. H. (1985). Set-related neuronal
activity in the premotor cortex of rhesus monkeys:
Effects of changes in motor set. Proceedings of the
Royal Society of London, Series B, Biological Sciences,
223, 331–354.
Hickok
1243
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
–
p
r
d
t
i
2
c
1
l
7
e
–
1
p
2
d
2
f
9
/
1
2
9
1
3
/
7
7
7
/
8
1
9
2
o
2
c
9
n
/
1
2
0
7
0
6
9
0
1
2
9
1
0
1
8
/
9
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
1
8
0
9
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
t
/
.
f
.
.
o
n
1
8
M
a
y
2
0
2
1