Dopamine and Consolidation of Episodic Memory:
Timing Is Everything
John Grogan1, Rafal Bogacz2, Demitra Tsivos3, Alan Whone1,3, and Elizabeth Coulthard1,3
Abstract
■ Memory consolidation underpins adaptive behavior and do-
paminergic networks may be critical for prolonged, selective in-
formation storage. To understand the time course of the
dopaminergic contribution to memory consolidation in hu-
mans, here we investigate the effect of dopaminergic medica-
tion on recall and recognition in the short and longer term in
Parkinson disease (PD). Fifteen people with PD were each
tested on or off dopaminergic medication during learning/early
consolidation (Day 1) and/or late consolidation (Day 2). Fif-
teen age-matched healthy participants were tested only once.
On Day 1 participants learned new information, and early ep-
isodic memory was tested after 30 min. Then on Day 2, recall
and recognition were retested after a 24-hr delay. Participants
on medication on Day 1 recalled less information at 30 min
and 24 hr. In contrast, patients on medication on Day 2 (8–
24 hr after learning) recalled more information at 24 hr than
those off medication. Although recognition sensitivity was un-
affected by medication, response bias was dependent on dopa-
minergic state: Medication during learning induced a more
liberal bias 24 hr later, whereas patients off medication during
learning were more conservative responders 24 hr later. We
use computational modeling to propose possible mechanisms
for this change in response bias. In summary, dopaminergic
medication in PD patients during learning impairs early consol-
idation of episodic memory and makes delayed responses
more liberal, but enhances late memory consolidation presum-
ably through a dopamine-dependent consolidation pathway
that may be active during sleep. ■
INTRODUCTION
Traditionally, memory impairment in neurodegenerative
disease has often been considered to be explained by
loss of cholinergic neurons (e.g., Bartus, Dean, Beer, &
Lippa, 1982). The role of dopamine in memory is more
controversial as are the nature and degree of memory
impairments in patients with dopaminergic loss such as
those with Parkinson disease (PD; Foerde, Braun, &
Shohamy, 2013; Foerde & Shohamy, 2011; Shohamy,
Myers, Geghman, Sage, & Gluck, 2006; Cools, Barker,
Sahakian, & Robbins, 2001). Conventional accounts of
memory posit three main stages: encoding (initial learn-
ing of the information), consolidation (maintenance of
the stored memory over a period of minutes, hours, days,
or years), and retrieval (accessing this memory). Dopami-
nergic effects have been reported across these stages.
Dopamine antagonist infusion during or immediately
after encoding worsens delayed recall in animals (Bethus,
Tse, & Morris, 2010; O’Carroll, Martin, Sandin, Frenguelli,
& Morris, 2006), suggesting a benefit of dopaminergic ac-
tivity on encoding and perhaps early consolidation. This is
also supported by a recent observation that optogenetic
stimulation of dopaminergic neurons during learning en-
hances memory retention (McNamara, Tejero-Cantero,
1University of Bristol, 2University of Oxford, 3North Bristol NHS
Trust
Trouche, Campo-Urriza, & Dupret, 2014). Importantly this
recent study found that stimulating the dopaminergic pro-
jections from the ventral tegmental area (VTA) to the hip-
pocampal CA1 subfield during learning did not speed up
learning but did increase memory at a 1-hr delayed test, as
well as increasing the reactivation of memory traces during
sleep after learning. The authors suggested that dopami-
nergic input to the CA1 increases the reactivation of newly
formed neuronal assemblies to allow for consolidation of
the memories they encode.
Dopamine appears to improve memory consolidation
in animals, but only at certain time points after initial
learning. A dopamine D1/D5 agonist infused into the
CA1 in the hippocampus of rats before inhibitory avoid-
ance learning had no effect when tested 24 hr later, nor
when it was infused 9 hr after learning (Bernabeu et al.,
1997). However, when it was infused 3 or 6 hr after learn-
ing, it improved (increased) the step-down latencies of
the rats significantly. A dopamine D1/D5 antagonist had
the opposite effect, decreasing the latency only when in-
fused 3 or 6 hr after learning. Adenylyl cyclase activator
infusion 3 or 6 hr after learning also increased memory,
and a PKA inhibitor decreased it, suggesting that this
dopamine-dependent consolidation effect is mediated
by the cAMP/PKA pathway in the hippocampus. Further in-
sights into the molecular mechanisms come from Rossato
and colleagues, who demonstrated that NMDAR antagonist
© 2015 Massachusetts Institute of Technology. Published under a
Creative Commons Attribution 3.0 Unported (CC By 3.0) license.
Journal of Cognitive Neuroscience 27:10, pp. 2035–2050
doi:10.1162/jocn_a_00840
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
7
/
1
2
0
7
/
2
1
0
0
3
/
5
2
1
0
9
3
4
5
9
/
4
1
5
2
7
8
o
3
c
3
n
4
_
1
a
/
_
j
0
o
0
c
8
n
4
0
_
a
p
_
d
0
0
b
8
y
4
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
t
.
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
infusion to the VTA 12 hr after learning impaired memory
14 days later, and this was reversed by dopamine agonist
infusion to the CA1 (Rossato, Bevilaqua, Izquierdo, Medina,
& Cammarota, 2009). The authors propose that NMDAR
activation in the VTA 12 hr after learning excites those cells
and causes greater dopaminergic activity projected to the
hippocampus, which causes dopamine-dependent
consolidation.
Very recent work demonstrated that a D2 inverse ago-
nist (similar to an antagonist, but it decreases the recep-
tor activity below baseline activity) injected postlearning
decreased novelty preference 24 hr later (França et al.,
2015). Importantly, a putative mechanism for such an ef-
fect stems from the findings that the decrease in memory
was accompanied by a decrease in CaMKII from 3 to
12 hr after learning, Zif-268 6 hr after learning, and brain-
derived neurotrophic factor (BDNF) 12 hr after learning,
along with decreased REM sleep. These proteins are all
activated at different times after learning, are involved
in plasticity and consolidation, and are affected by dopa-
mine just after the time of learning, as well as at their
time of activation (Rossato et al., 2009). This suggests
that dopamine affects plasticity protein levels, possibly
via REM sleep. Perhaps not surprisingly given the known
dose dependence of dopamine responses and complex-
ity of dopaminergic networks, somewhat conflicting
results have emerged when different experimental para-
digms are used (e.g., Furini, Myskiw, Schmidt, Marcondes,
& Izquierdo, 2014; Péczely et al., 2014; Rossato et al., 2009,
2013). However, the overall picture is that dopamine-
dependent networks have a time-critical role in animal
memory consolidation that may to some extent depend
on a period of sleep.
One theory of consolidation that incorporates a role of
dopamine is the synaptic tagging and capture theory
(Clopath, 2012; Redondo & Morris, 2011; Clopath, Ziegler,
Vasilaki, Büsing, & Gerstner, 2008; Sajikumar & Frey,
2004; Frey & Morris, 1997, 1998). This theory states that
an input to a synapse can cause early synaptic plasticity
and can also “tag” the synapse so that when plasticity-
related products are synthesized they can be captured
by the tagged synapse and used to stabilize the early plas-
ticity changes to allow consolidation to take place. In the
computational model of this theory (Clopath et al., 2008),
the threshold for production of the plasticity-related
products is set by tonic dopamine levels. As this theory
states that the tags only last a few hours, which could
account for some of the findings mentioned above, it
cannot easily account for the consolidation effects of
dopamine at later time points found in other studies,
without extensions made to the model.
Although these models of memory consolidation have
been developed based on animal experimentation, the
focus of our work is the contribution of dopamine to hu-
man memory. The global application of dopaminergic
drugs in humans limits the conclusions that can be drawn
and poses a challenge when designing studies in people.
However, it is critical to establish how dopamine influ-
ences human memory, particularly given that memory
loss is such a prominent problem affecting our increas-
ingly elderly population.
Older adults given levodopa, a dopamine precursor,
before learning showed no benefits on memory after
2 hr, but did show dose-dependent improvement in scene
recognition 6 hr after learning (Chowdhury, Guitart-Masip,
Bunzeck, Dolan, & Düzel, 2012). Such an inverted U-shaped
correlation between dose and performance perhaps im-
plies that low concentrations were insufficient to have an
impact, whereas high concentrations “overdosed” the
brain (Cools, 2006; Cools et al., 2001). The delayed benefi-
cial effects of levodopa suggest dopamine might be impor-
tant for human late memory consolidation, but effects of
dopamine over longer timescales in humans have not been
investigated.
Genetic studies suggest that increased levels of dopa-
minergic activity improve memory (Wittmann, Tan, Lisman,
Dolan, & Düzel, 2013; De Frias et al., 2004). However, work
in humans has not normally fully dissociated encoding,
consolidation, and retrieval, and results have conflicted
with some studies finding that dopamine replacement
medications given before learning improve memory
(Chowdhury et al., 2012; Coulthard et al., 2012) whereas
another found that they impaired encoding (Macdonald
et al., 2013). Thus, it is not clear exactly how dopamine con-
tributes to the earliest stages of memory processing and
whether human dopaminergic contributions to memory
mirror those observed in animals.
We designed a paradigm to differentiate the effects
of dopaminergic activity on encoding, consolidation and
retrieval in PD patients. We aimed to see whether exoge-
nous dopamine present during encoding/early consolida-
tion or late consolidation would enhance 24-hr delayed
recall and recognition memory in people with PD. Impor-
tantly, we were able to distinguish between the stages of
memory by probing (i) initial learning, (ii) early consolida-
tion and retrieval (at 30 min), and (iii) late consolidation
and retrieval at 24 hr (after overnight consolidation) sepa-
rately. By withdrawing patients from their dopaminergic
medications before learning on Day 1 and/or testing on
Day 2, we could see the effects of dopaminergic activity
on each component process within the memory system.
METHODS
Participants
Fifteen patients with PD and 15 age-matched healthy par-
ticipants were tested (see Table 1 for details). Patients
were identified through movement disorder and general
neurology clinics in North Bristol NHS Trust, and all had a
clinical diagnosis of PD and no other neurological diagno-
ses. The PD patients were all on dopaminergic medica-
tion (levodopa and/or dopamine agonists) and were
not taking cholinesterase inhibitors, monoamine oxidase
2036
Journal of Cognitive Neuroscience
Volume 27, Number 10
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
7
/
1
2
0
7
/
2
1
0
0
3
/
5
2
1
0
9
3
4
5
9
/
4
1
5
2
7
8
o
3
c
3
n
4
_
1
a
/
_
j
0
o
0
c
8
n
4
0
_
a
p
_
d
0
0
b
8
y
4
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
.
f
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Table 1. Demographics and Questionnaires Scores for Patients and Healthy Participants
Group
Number
Age
Years since diagnosis
Number on levodopa/
dopamine agonists/both
Levodopa dose equivalency
(mg/day)
MMSE
UPDRS on meds
UPDRS off meds
DASS
(Depression/
Anxiety/
Stress)
BIS
REI (Rationality/
Experientiality)
LARS
Number of Missing
Patients/HPP
PD Patients
Healthy
Participants
One-way ANOVA: Patients vs. Healthy
Participants (df, F, p)
15
15
71.53 (2.40)
71.00 (2.63)
(1, 28), 0.076, .785
5.20 (1.38)
9/1/5
603.00 (71.64)
D
o
w
n
l
o
a
d
e
d
f
r
o
m
27.00 (0.31)
27.90 (0.34)
(1, 28), 5.000, .033
19.90 (3.17)
24.90 (3.86)
1/0
22.50 (2.57)
10.30 (3.17)
(1, 27), 10.943, .003
1/0
0/4
6.53 (0.84)
2.94 (0.82)
(1, 27), 16.079, <.001
8.18 (1.28)
3.176 (0.83)
(1, 27), 14.312, .001
6.41 (0.96)
5.06 (0.98)
(1, 27), 2.640, .116
63.40 (1.68)
62.10 (1.37)
(1, 27), 0.736, .398
3.33 (0.15)
3.67 (0.21)
(1, 24), 1.828, .189
2.89 (0.12)
2.91 (0.15)
(1, 24), 0.10, .920
−19 (1.56)
−26 (1.32)
(1, 28), 13.094, .001
Means with SEM in parentheses unless stated otherwise. MMSE = Mini Mental State Exam; UPDRS = Unified Parkinson Disease Rating Scale; DASS =
Depression, Anxiety and Stress Scale; BIS = Barratt Impulsivity Scale; REI = Rational–Experiential Inventory; LARS = Lille Apathy Rating Scale.
inhibitors, or anti-psychotic medications or treated with
deep brain stimulation or other functional neurosurgery.
Aside from Parkinsonian medications, two patients were tak-
ing medications for cholesterol (simvastatin), and one of
these was also on amlodipine and clopidogrel, which are an-
tihypertensive and antiplatelet medications. One was taking
alfuzosin for enlarged prostate, one an SSRI (sertraline) and
calcium supplements, one quinine sulfate (for nocturnal
leg cramps), and one latanoprast and dorzolamide for glau-
coma and salazopyrin for arthritis. Levodopa dose equiva-
lency was calculated to get a measure of total daily
dopaminergic medication taken (Tomlinson et al., 2010).
Healthy participants were either the spouses of the PD
patients who accompanied them to the session or were
recruited from the BRACE Centre’s Healthy Volunteer da-
tabase. Healthy participants taking any dopaminergic or
noradrenergic medications were excluded from the
study. None had any neurological diagnoses or reported
memory problems. Of the healthy participants, one was
taking simvastatin and aspirin (cholesterol and antiplate-
let medication), one an ACE inhibitor (ramipril), and one
fluoxetine (SSRI).
Ethics approval was granted by the North Bristol NHS
Trust Research Ethics Committee, and all participants
gave written consent in accordance with the Declaration
of Helsinki.
Task
The Hopkins Verbal Learning Task-Revised (HVLT-R) is
an episodic memory test with immediate and 30-min de-
layed recall and recognition components (www4.parinc.
com/Products/Product.aspx?ProductID=HVLT-R). The
experimenter reads aloud 12 words at a rate of one per
second, after which the participant recalls as many words
as they can in any order. These words are drawn from
three semantic categories (four words from each) such
as mammals, fuels, and tools. The list is read out twice
more in the same order, each time followed by immedi-
ate recall to give a total of three immediate recall trials.
After a 30-min delay, there is another recall trial, along
with a recognition trial where the experimenter reads
aloud the 12 target words and 12 new distractor words.
Six of these distractors are from the same three semantic
categories as the target words (two from each category),
and six are not. The words are presented in a randomized
order, and no feedback is given. Participants respond
“yes” if they think the word was on the learning list
and “no” if they think it is a new word. In an addition
to the standard HVLT, at 24 hr another recall test was
performed along with a further recognition test, which
was the same as the 30-min test only with new distractor
words. A different version of the task was given in each of
the four conditions (Versions 1, 3, 4, 5) in a randomized
Grogan et al.
2037
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
0
7
/
2
1
0
0
3
/
5
2
1
0
9
3
4
5
9
/
4
1
5
2
7
8
o
3
c
3
n
4
_
1
a
/
_
j
0
o
0
c
8
n
4
0
_
a
p
_
d
0
0
b
8
y
4
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
f
t
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
order between participants. Each word appeared in only
one version of the task. As patients completed the HVLT
four times, the delayed tests could not be kept a surprise,
so all patients were told at the beginning that they would
be tested on the list both later on in the current session
and in the session the next day.
Procedure
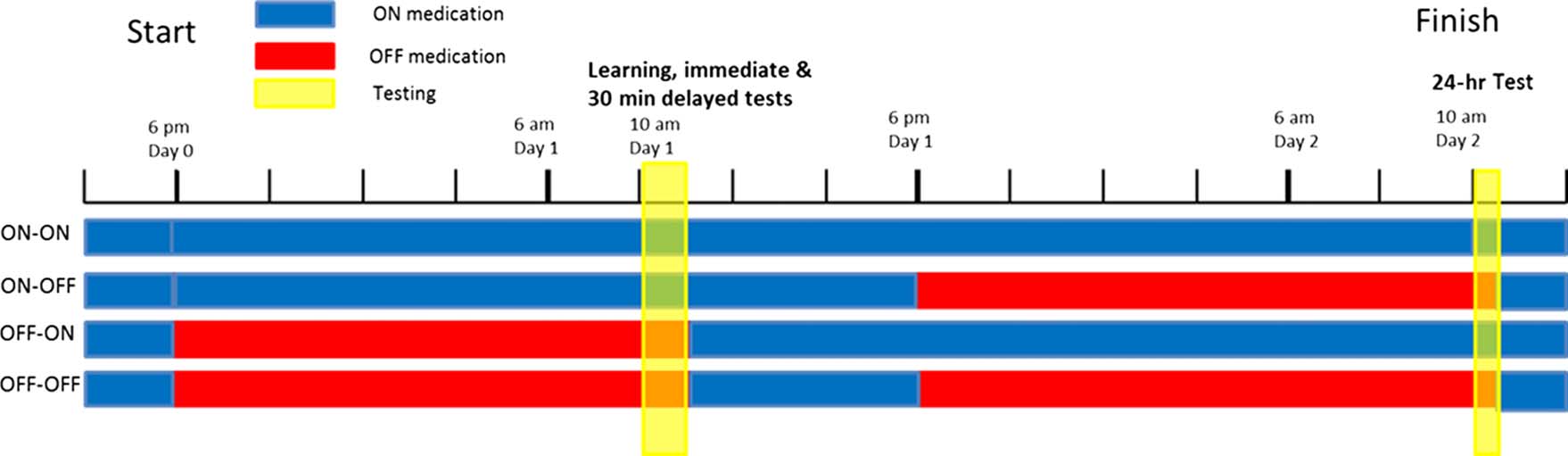
Testing took place over two consecutive days (see Figure 1
for details). On Day 1 participants completed the immedi-
ate recall and 30-min delayed recall and recognition tests,
along with paper-based questionnaires: Barratt Impul-
sivity Scale (Patton, Stanford, & Barratt, 1995), Rational–
Experiential Inventory (REI; Pacini & Epstein, 1999),
Depression Anxiety Stress Scale (DASS; Lovibond &
Lovibond, 1995), Mini Mental State Exam (MMSE; Folstein,
Folstein, & McHugh, 1975), and Lille Apathy Rating Scale
(LARS; Zahodne et al., 2009; Sockeel et al., 2006). The
PD patients also completed a standard motor symptom
rating (Unified Parkinson Disease Rating Scale-III; Goetz
et al., 2008). These assessments were completed during
the 30-min delay.
On Day 2 participants completed the 24-hr delayed re-
call and recognition tasks and a sleep questionnaire
about their sleep the previous night (St. Mary’s Hospital
Sleep Questionnaire; Leigh, Bird, Hindmarch, Constable,
& Wright, 1988; Ellis et al., 1981)).
PD patients were on or off their medications on each
of the 2 days, giving four conditions for each subject
(Day1/Day2 = on/on, on/off, off/on, off/off ). When the
patients were in the “off” condition, they did not take
any dopaminergic medications for a minimum of 15 hr
before testing (usually none after 6 pm for testing at
10 am the next day). After the session had finished, they
took a dose and then carried on with their usual dose
schedule. For example, in the “off–off” condition (bot-
tom bar in Figure 1), patients were on medication until
6 pm Day 0, then off medication until after testing on
Day 1 (∼11 am), then on medication until 6 pm Day 1, then
off medication until after testing on Day 2 (∼10:30 am),
after which they resumed their normal medication sched-
ule. All patients were on their medication for at least a
few hours after learning on Day 1 (see Figure 1). This
meant that in Day 1 off conditions they took a dose after
the session finished and continued their normal dosage
until the evening where if they were to be off medication
on Day 2 they stopped their doses. This was to minimize
discomfort to the patients and to reduce the chance of
neuroleptic malignant syndrome, which can occur when
dopaminergic medications are stopped (Keyser &
Rodnitzky, 1991).
The order of the four conditions was counterbalanced
across participants. Healthy participants were tested
only once.
Data Analysis
Data were analyzed using within-subject repeated-
measures ANOVAs to test the effects of Day 1 and
Day 2 medication state on each separate measure in
the patient group. Between-subject ANOVAs were used
to compare patients and healthy participants. Raw de-
layed recall scores were measured at 30 min and 24 hr,
as well as the percentage retention, which was the de-
layed recall score divided by the highest of the second
and third immediate recall trial scores, multiplied by
100. This was calculated for the 30-min and 24-hr delayed
recall scores. The change between the two delayed scores
was also expressed as a percentage (24 hr/30 min × 100).
For the recognition tests, the raw measures were num-
ber of hits, misses, false alarms, and correct rejections.
Signal detection methods (Stanislaw & Todorov, 1999;
Macmillan, 1993; Green & Swets, 1966) were used to es-
0 measure of sensitivity between the targets
timate the d
and distractors,
d0 ¼ Z p hits
ð
ð
Þ
Þ−Z p false alarms
ð
ð
Þ
Þ
(1)
where Z is the inverse cumulative density of the standard
normal distribution and the hits and false alarms are
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
0
7
/
2
1
0
0
3
/
5
2
1
0
9
3
4
5
9
/
4
1
5
2
7
8
o
3
c
3
n
4
_
1
a
/
_
j
0
o
0
c
8
n
4
0
_
a
p
_
d
0
0
b
8
y
4
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
f
.
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 1. Timeline of the testing procedure from 6 pm Day 0 to 12 pm Day 2. Blue denotes when patients were on their dopaminergic medication,
red when they were withdrawn from it, and yellow the time of testing sessions. To have patients off their medication during the testing at 10 am,
patients had to be withdrawn from their dose the night before because of the long washout times of the medications.
2038
Journal of Cognitive Neuroscience
Volume 27, Number 10
expressed as probabilities, and response bias (a measure
of where the threshold for a “yes” response is set),
The change in weights after presentation of a novel
stimulus is calculated as
ð
c ¼ − Z p hits
ð
Þ
Þ þ Z p false alarms
ð
ð
2
Þ
Þ
(2)
Δwj ¼
−η
ð
Na 1−a
(cid:3)
(cid:2)
Þ h xj−a
(4)
Negative response biases indicate that less evidence is
needed for a “yes” response, which corresponds to
more “yes” responses—a liberal response bias. Positive
response biases mean more evidence is needed for a
“yes,” which leads to more “no” responses—a con-
servative response bias. Nonparametric measures of sen-
sitivity and response bias were also calculated (Stanislaw
& Todorov, 1999) but are not reported here as they
gave the same results as the more common parametric
measures.
The response bias score takes into account the propor-
tion of hits and false alarms. We use this measure rather
than discussing the raw hits and false alarms scores be-
cause there was no clear pattern of results emerging from
them.
All SEM bars for figures have been corrected for
between-subject variance using the Cousineau–Morey
method (O’Brien & Cousineau, 2014; Morey, 2008;
Cousineau, 2005), which removes the variance from
between-subject differences and only shows the variance
due to within-subject differences in a similar manner to
the way a repeated-measures ANOVA removes between-
subject variance.
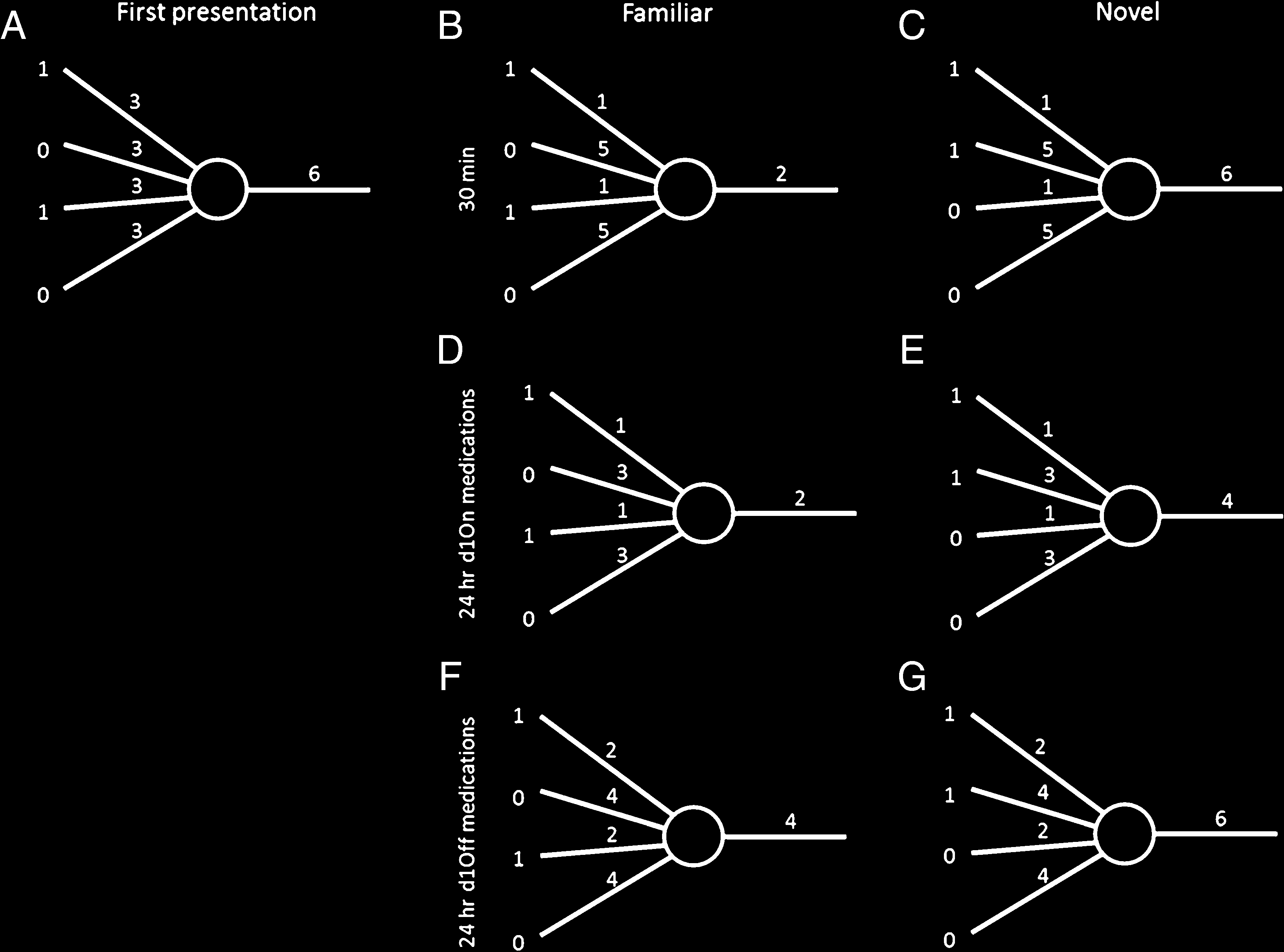
Methods of Simulation
To illustrate what changes in synaptic connections could
underlie the observed effects of dopaminergic medica-
tion on recognition memory (i.e., higher tendency to
classify both previously seen and unseen stimuli as famil-
iar when on medication during encoding; see Figure 4B),
we used a simple anti-Hebbian model of familiarity
discrimination (Bogacz & Brown, 2003a). The intuitive
description of model is provided in the Results section,
whereas here we describe details of the model (it is
recommended to read the Results section before these
details).
The model is a simple layer network with N input
neurons and novelty neurons in separate layers. For
the version we used, we simplified the model to only
have one novelty neuron and N = 100 input neurons.
The model assumes the novelty neuron receives con-
nections from all input neurons. The weights of the con-
nections from input neuron j to the novelty neuron are
denoted as wj. The activity of each input neuron is de-
noted by xj, and it is 1 if the neuron is active and 0 oth-
erwise. The activity of the novelty neuron (h) is thus
calculated,
h ¼
XN
j¼1
wjxj
(3)
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
0
7
/
2
1
0
0
3
/
5
2
1
0
9
3
4
5
9
/
4
1
5
2
7
8
o
3
c
3
n
4
_
1
a
/
_
j
0
o
0
c
8
n
4
0
_
a
p
_
d
0
0
b
8
y
4
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
f
/
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
where η is the learning rate and a is the sparseness pa-
rameter controlling the fraction of input neurons active
in the patterns. In particular for each input pattern (x),
the number of active neurons is determined by the
sparseness parameter a such that
XN
j¼1
xj ¼ aN
(5)
In all our simulations, parameters were fixed at a = 0.2 and
η = 0.2. After learning a series of randomly generated pat-
terns, we present the now-familiar patterns and some ran-
domly generated novel patterns. For each presented
pattern, the neuron’s activity is compared against a thresh-
old T: if h < T it is classified as familiar, and if h > T as novel.
We further denote the thresholds on 2 days of testing
by T1 and T2, respectively. In all our simulations, T1 = 1.
To simulate the effects of high and low dopaminergic
state, we proposed two versions of the model. In the first
version (the decay model), dopaminergic medications
would affect the decay of the weights. Dopamine has
been implicated in consolidation in animals (Bethus
et al., 2010; Rossato et al., 2009; Bernabeu et al., 1997),
and models have been proposed in which dopamine sets
the threshold for consolidation of weight changes at syn-
apses (Clopath et al., 2008; Frey & Morris, 1997). We
model this with two coefficients that control how much
of the weight change is expressed:
(cid:4)
if Δwij > 0; Δwij ¼ Δwijαþ
if Δwij < 0; Δwij ¼ Δwijα−
(6)
There is a separate coefficient for the decreases in
weights caused by coactivation of the input and novelty
neurons and the increase in weights caused by the acti-
vation of the input neurons alone. This change was only
applied after the 30-min test as consolidation takes time
to have an effect. This model has four parameters that
have been optimized: α+ and α− for PD patients on
and off medication on Day 1, respectively (α+,On, α−,Off,
α+,Off, α−,Off). In this model, the thresholds are assumed
to be the same on both days T2 = T1 = 1.
In the second version of the model (the threshold
model), the dopamine state during learning does not affect
the decay of the weight changes but instead the threshold
T2 for the 24-hr test. We assumed that T2 depends upon
Day 1 dopamine state; thus, the model has four free param-
eters, two of which correspond to the value of T2 when the
dopamine was on and off during Day 1 (T2,On, T2,Off). This
is akin to a consolidation or decay of the threshold inputs
dependent on dopamine 30–120 min after learning. Two
other free parameters correspond to α+ and α−, but these
were the same for Day 1 on and off conditions.
Grogan et al.
2039
We simulated the models and then compared the rec-
0 and response bias measures from
ognition accuracy d
the model with the behavioral data (Figure 4) using root
mean square deviation (RMSD),
RMSD ¼
s
ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
Xn
Þ2
ð
yi−di
n
i¼1
(7)
where yi is the behavioral data, di is the simulated data
for measure i, and n is the number of measures (n =
0 and response bias on and off med-
8; 30 min and 24 hr d
ications on Day 1).
We used 12 familiar patterns, each presented three
times in the same order (as in the HVLT-R), and for the
recognition tests all 12 familiar patterns along with
12 novel patterns were presented (different novel patterns
were used for each test). We repeated this 1000 times
with newly generated patterns and random starting
weights each time.
We used Matlab’s fminsearch function to find free pa-
rameters of each model that minimize this RMSD. The ini-
tial values of the free parameters (i.e., the starting point for
the search) were randomly generated from uniform distri-
butions between 0 and 2. For each model, the whole opti-
mization was repeated 300 times with different randomly
generated initial sets of parameters to avoid local minima.
RESULTS
No Effects of Dopaminergic Medication on
Immediate Recall
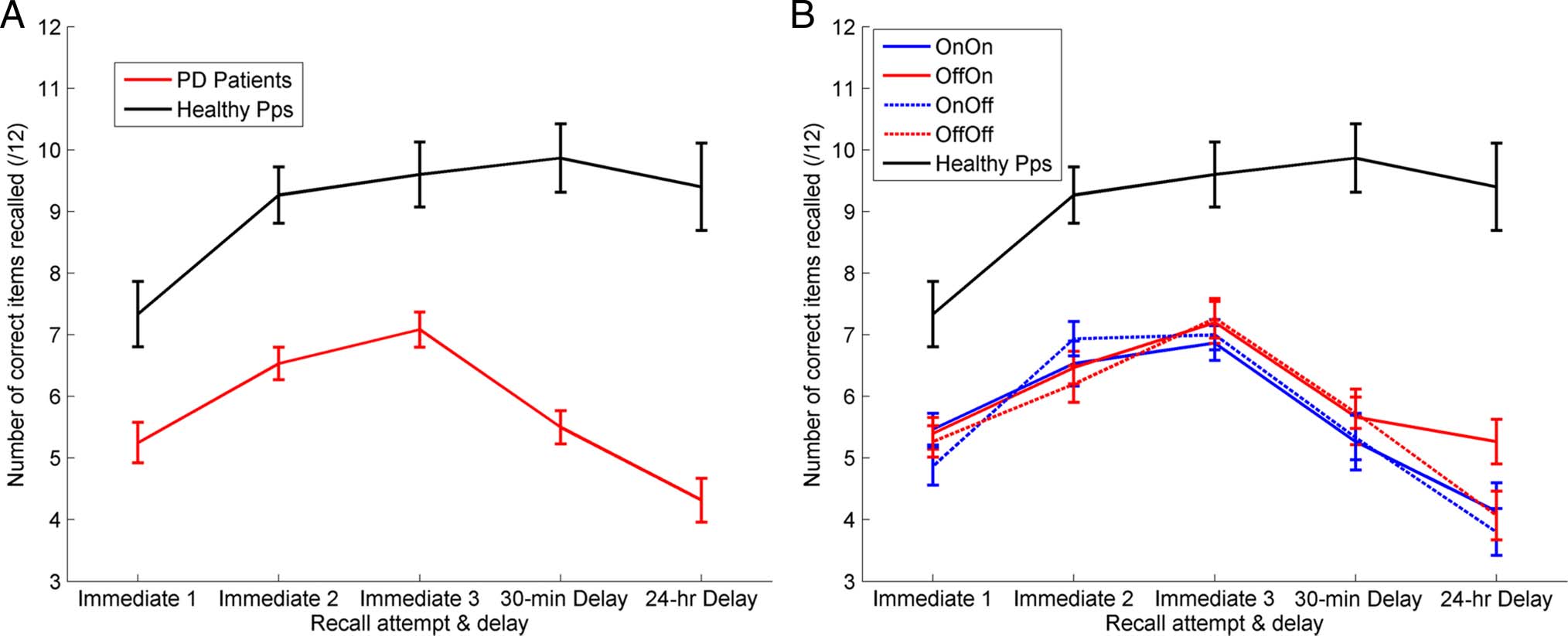
Patients and healthy participants show an increase in the
number of words recalled over the three immediate re-
call trials (Figure 2A), although healthy participants recall
significantly more words at each trial than patients (one-
way multivariate ANOVAs, p < .001). Healthy participants
only show a change in number of correctly recalled
words between the first and second recalls (one-way
repeated-measures ANOVA with Bonferroni-corrected pair-
wise comparisons, p = .005), with no further increases or
decreases ( p = 1). A two-way repeated-measures ANOVA
(Day 1 medication state × Time) on the PD patients’
immediate recall trials revealed a significant effect of
Time (F(2, 28) = 24.870, p < .001) but no significant
effect of Day 1 medication state (F(1, 14) = .008, p =
.931; see Figure 2B). PD patients take longer to learn
information and retain less information than healthy par-
ticipants, with their final immediate recall score about the
same as the healthy participants’ first, and show no do-
pamine medication effects.
Medication Impairs Early Consolidation but
Improves Late Consolidation
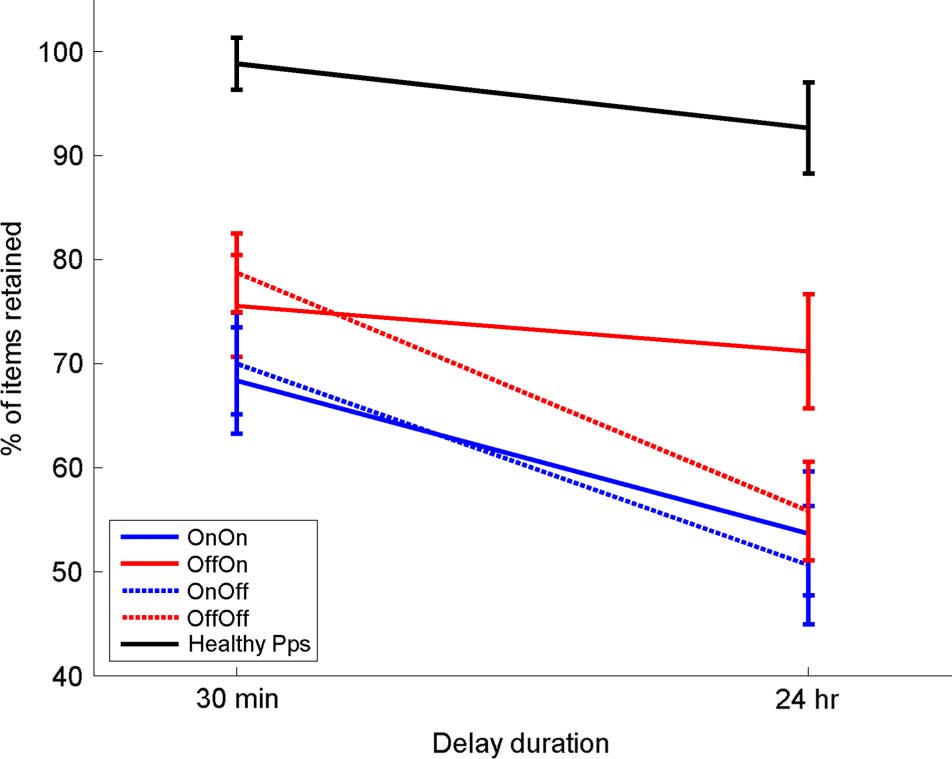
Healthy participants retain information across both 30-min
and 24-hr delays (Figure 2A). Patients, however, show a
decrease across both delays: 30-min delayed recall is
significantly lower than the third immediate recall (one-
way repeated-measures ANOVA with Bonferroni-corrected
pairwise comparisons; p = .001), and 24-hr recall score
is significantly lower than both the third immediate re-
call score and the 30-min delayed recall score ( p < .001
for both). On average, 1–1.5 words are lost across both
30-min and 24-hr delays in patients.
The recall retention scores in different medication condi-
tions are shown in Figure 3, and the patients were statisti-
cally analyzed with a three-way repeated-measures ANOVA
(Delay × Day 1 medication state × Day 2 medication
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
0
7
/
2
1
0
0
3
/
5
2
1
0
9
3
4
5
9
/
4
1
5
2
7
8
o
3
c
3
n
4
_
1
a
/
_
j
0
o
0
c
8
n
4
0
_
a
p
_
d
0
0
b
8
y
4
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
.
t
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 2. The mean number of correct words recalled at each recall test. (A) The red line is the patients’ raw recall scores averaged across all
conditions (SEM bars). Healthy participants showed significantly higher accuracy on all recall trials ( p < .001 for all) and no decrease over the delays
(F(1, 14) = 2.507, p = .136). PD patients learn more slowly and forget more over 30-min and 24 hr-delays. (B) Each PD condition separately, there
were no significant differences for the immediate recall trials.
2040
Journal of Cognitive Neuroscience
Volume 27, Number 10
ing that whereas the off–on condition may contribute
majorly to the effect, the on–on condition does also.
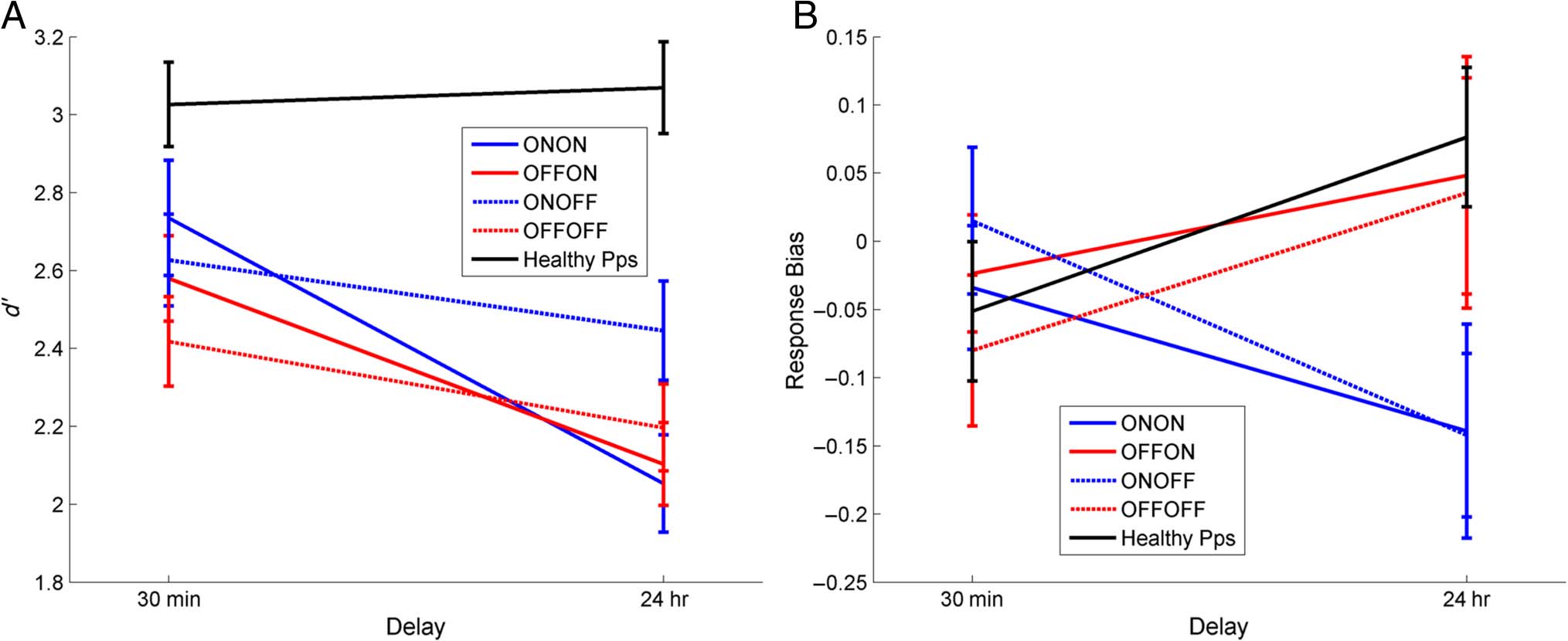
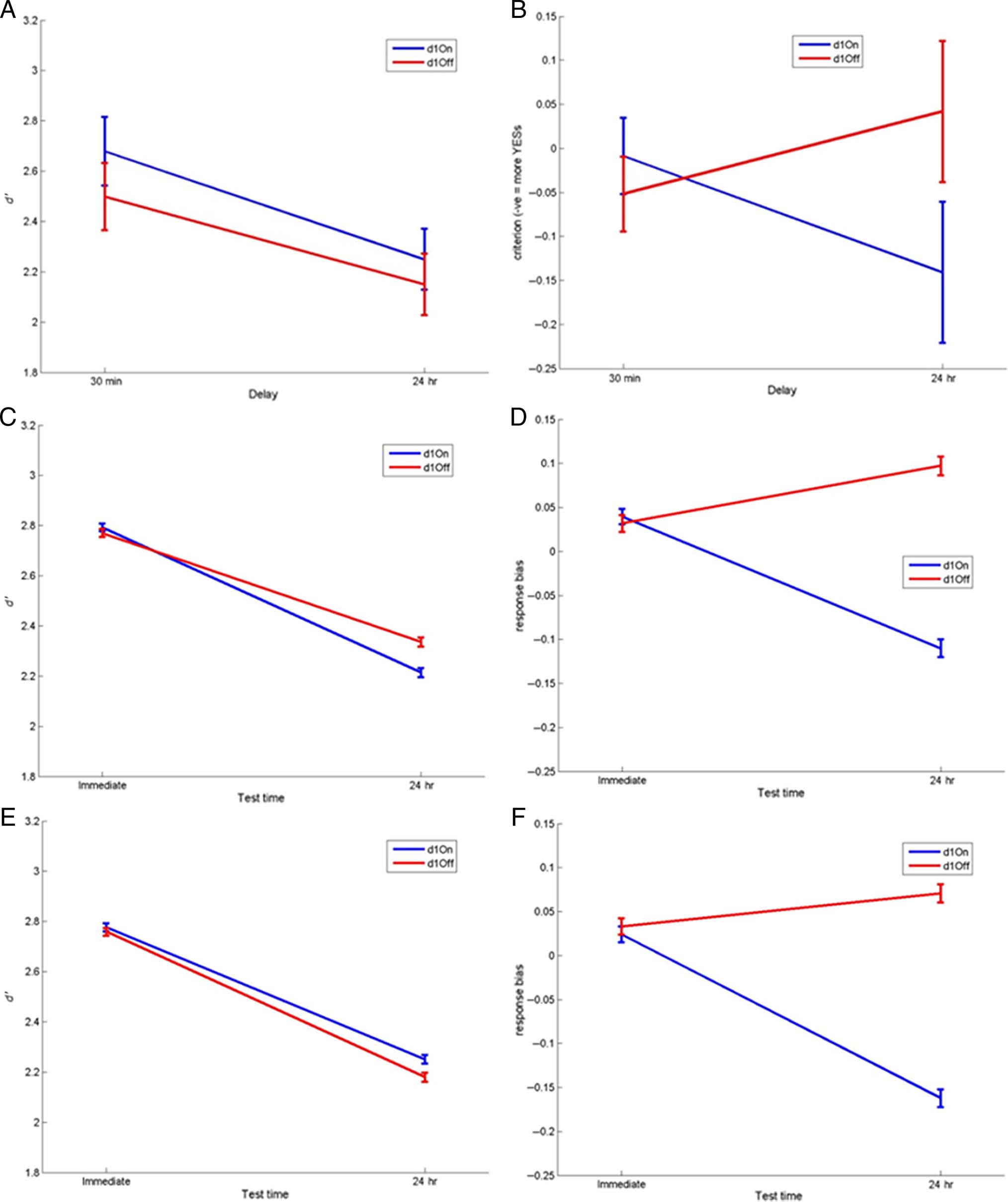
Response Bias, but Not Recognition Sensitivity, Is
Affected by Dopamine
As shown in Figure 4A, the patients had significantly
0) on the 24-hr recognition task than
lower sensitivity (d
on the 30-min test (effect of delay: F(1, 14) = 14.089,
p = .002), but there was no significant effect of medica-
0 scores at
tion state. Healthy participants have the same d
both time delays (paired t test: T = 0.363, p = .722),
which are much higher than the patients’ scores in both
cases.
Figure 4B shows that when patients were on medica-
tion on Day 1 they had a more liberal (negative) response
bias after 24 hr, but when patients were off medication
on Day 1 they had a more conservative (positive) bias
24 hr later, regardless of Day 2 medication (interaction
of time and Day 1 medication: F(1, 14) = 15.083, p =
.002). In other words, dopaminergic medication during
learning and early consolidation led to more “yes” re-
sponses 24 hr later, whereas a lack of dopaminergic med-
ication during learning led to more “no” responses 24 hr
later. In contrast, response bias of healthy participants
increased over 24 hr; they were more conservative at the
24-hr recognition task, responding “no” more often and
decreasing the number of hits and false alarms. This is
the same pattern as PD patients off medication on Day 1.
Modeling the Response Bias Effects
The response bias effects were the most unexpected re-
sults we found, and we were unable to come up with a
simple explanation, so we turned to computational
models of familiarity discrimination to see if an interac-
tion of delay and Day 1 medication state could be found
in the models.
We have used an abstract recognition model to at-
tempt to replicate our recognition memory results. The
anti-Hebbian model (Bogacz & Brown, 2002, 2003b;
Kohonen, Oja, & Rouhonen, 1974) was originally devel-
oped to capture the finding that perirhinal cortex neu-
rons that discriminate on the basis of familiarity show
high firing activity for novel, unfamiliar stimuli, but low
firing activity for familiar stimuli (Brown & Aggleton,
2001). For example, when presenting the same stimulus
twice, in the first time the neurons will fire wildly, and in
the second time the neurons will fire at a lower rate. This
differential activity allows discrimination between novel
and familiar stimuli.
In the model, when a novel stimulus is presented as a
pattern of activity to the novelty neuron, it elicits an out-
put activity (Figure 5A). Consequently, the synaptic
weights from the activated inputs are decreased, which
results in lower output activity for the same stimulus pre-
sented again (Figure 5B; see Banks, Bashir, & Brown,
Grogan et al.
2041
Figure 3. The mean percentage retention over 30-min and 24-hr
delays. Healthy participants (black line) do not show a significant
decrease over 24 hr (F(1, 14) = 2.507, p = .136). For PD patients (red
and blue lines), there is a significant effect of delay (F(1, 14) = 49.885,
p < .0001) and a clear effect of Day 1 medication state (F(1, 14) = 7.329,
p = .017), with patients off medication on Day 1 during learning
(red lines) having higher retention of words at 30 min and 24 hr than
patients on medication on Day 1 (blue lines). It also shows a significant
interaction of Delay and Day 2 medication state (F(1, 14) = 11.4, p =
.005), with patients on medication on Day 2 (solid lines) having higher
retention on Day 2 than patients off dopamine on Day 2 (dashed lines).
All bars show SEM.
state). The recall scores after 24 hr were significantly lower
than scores after 30 min (effect of delay: F(1, 14) = 49.855,
p < .0001). Patients who were on dopaminergic medica-
tions on Day 1 recalled less at both 30-min and 24-hr recall,
which can be seen in Figure 3 where both blue lines are
below the red lines (effect of Day 1 medication state:
F(1, 14) = 7.320, p = .017). In stark contrast, patients on
dopaminergic medication on Day 2 retained significantly
more words between 30 min and 24 hr, which can be seen
in Figure 3 where both solid lines are flatter than the
dashed lines (interaction of delay and Day 2 medication:
F(1, 14) = 11.4, p = .005; note that Day 2 medication effect
could only be an interaction, as Day 2 medication could not
possibly affect the 30-min recall score on Day 1). Thus, do-
paminergic medication impairs delayed memory when
present during learning (without affecting immediate re-
call) but improves it when present overnight after learning
and during testing the next day.
Looking at Figure 3B, it seems that the Delay × Day 2
medication state effects may be driven mostly by the
off–on condition and that this may be significantly differ-
ent to the other three conditions by itself. However, the
three-way interaction from the ANVOA (Delay × Day 1
medication state × Day 2 medication state) did not return
a significant result (F(1, 14) = 1.274, p = .278), which
means that the significant differences are not due just to
the off–on condition. There is only a significant effect
when delay and Day 2 medication state are factors, mean-
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
0
7
/
2
1
0
0
3
/
5
2
1
0
9
3
4
5
9
/
4
1
5
2
7
8
o
3
c
3
n
4
_
1
a
/
_
j
0
o
0
c
8
n
4
0
_
a
p
_
d
0
0
b
8
y
4
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
t
f
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
0
7
/
2
1
0
0
3
/
5
2
1
0
9
3
4
5
9
/
4
1
5
2
7
8
o
3
c
3
n
4
_
1
a
/
_
j
0
o
0
c
8
n
4
0
_
a
p
_
d
0
0
b
8
y
4
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
f
/
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 4. Recognition memory. (A) d0 measure of sensitivity and (B) response bias for 30-min and 24-hr delayed tests (SEM bars). Healthy
0 across the delay whereas patients do (F(1, 14) = 14.089, p = .002). Response bias has a significant interaction of
participants show no decrease in d
Day 1 medication state and delay (F(1, 14) = 15.083, p = .002), with patients on dopamine on Day 1 (blue lines) showing a decrease in response bias
over 24 hr (more “yes” responses) and patients off dopamine during learning (red lines) and healthy participants showing an increase in response bias
(more “no” responses) irrespective of Day 2 medication (solid/dashed lines).
2012, for review of experimental evidence, suggesting
that such anti-Hebbian synaptic plasticity in the perirhinal
cortex underlies familiarity discrimination). Additionally,
during learning the weights from the inactivate inputs
are increased to balance the overall excitability for the
neuron. When a new novel pattern is presented, it is likely
to return a similar level of output activity as the original
novel pattern (here shown by the output number 6;
Figure 5C), which would not be the case without this in-
crease. The new novel pattern has some overlapping in-
put activity because of the finite number of inputs. The
output activity of the novelty neuron in the model is a
simple number, as this is a phenomenological model,
but could correspond to either the number of spikes
fired, the level of depolarization, or the probability of fir-
ing (if the model were to include more than one novelty
neuron).
We considered two ways of modeling the effect of
medication to which we refer as the decay and threshold
models. The decay model assumes that PD patients on
medication on Day 1 would show a decay in the in-
creased weights (Figure 5D), which does not affect the
output for familiar stimuli but decreases it for novel stim-
uli (Figure 5E). This would lead to more false alarms and
a lower response bias. PD patients off medication on
Day 1 were expected to have a decay in all weight changes
(Figure 5F), which heightens the activity from familiar
stimuli and overall keeps it the same for novel patterns
(Figure 5G), meaning more rejections of familiar stimuli
(i.e., a more conservative, positive response bias).
The threshold model assumes that the dopaminergic
medications on Day 1 affect the threshold for the novelty
decision on Day 2. In the threshold model, there is still a
decay of the weight changes (Equation 6), but this is
the same for the two simulated medication conditions.
The threshold can be thought of as either the spiking
threshold for the novelty neuron or for a “decision neu-
ron” that receives input from the novelty neuron. Either
way, dopamine-dependent consolidation mechanisms
could affect the threshold.
We simulated each model with a variety of randomly
generated parameters and minimized the RMSD between
the simulated data and the PD patients’ behavioral data.
The best fitting parameters (with the lowest RMSD) for
0 data
each model are shown along with the simulated d
in Figure 6. We compare the two models directly using
the RMSD as both models have the same number of pa-
rameters, meaning that measures like the Akaike Infor-
mation Criterion are not needed.
Both models can provide good fits to the patient data
0
(Figure 6A and B), reproducing the similar decrease in d
in both conditions (Figure 6C and E), but with Day 1 on
condition leading to a lower, more liberal response bias
and Day 1 off medication leading to a higher, more con-
servative response bias (Figure 6D and F). Although the
RMSD for the threshold model is slightly lower than for
the decay model, the behavior of participants does not
differ from the predictions of either model; thus, given
the uncertainty in exact values of experimental data, we
feel that it is not justified to categorically say that one of
the models fit better.
The best fitting parameters for the decay model were
α+,On = 0.1515, α−,On = 0.8787, α+,Off = 0.4079 and
α−,Off = 0.8702. This represents greater decay of positive
weight changes and slightly lesser decay of negative
weight changes for PD on medication on Day 1 when
2042
Journal of Cognitive Neuroscience
Volume 27, Number 10
compared to PD Day 1 off. This means that PD patients
on dopamine during learning may be better preserving
the weight decreases associated with the active inputs
from the patterns, but at the cost of poorer consolida-
tion of the weight increases for the inactive inputs.
The best fitting parameters for the threshold model
were T2,On = 1.1594, T2,Off = 1.1108, α+ = 0.3685, α− =
0.7562. This means that, even though this model is nomi-
nally the threshold model, there were only very slight dif-
ferences between the two conditions’ thresholds. The
slightly higher threshold when on medication on Day 1
means that 24 hr later patterns with higher (more novel)
activities are accepted as familiar, leading to a more liberal
response bias. Both conditions shared the same decay
parameters (α+ and α−), but these two were not the same,
meaning that both on and off conditions had a large de-
crease in inactive connection weight increases and a
smaller decrease in active connection weight decreases,
which would decrease the novelty neuron’s activity and
then the slight differences in thresholds would be enough
to separate the two response biases.
In summary, both models can account for the current
data, and we mention in the Discussion additional exper-
iments that could distinguish between the models.
Questionnaires
A one-way ANOVA was run on the questionnaire data (PD
vs. healthy participants), which revealed that PD patients
scored significantly lower on the MMSE (see Table 1 for
p values) and higher on the DASS and LARS than healthy
participants. The DASS breaks down into subscores for
depression, anxiety, and stress, and there were significant
differences only for the depression and anxiety sub-
scores, not for stress.
The groups did not differ significantly on Rational–
Experiential Inventory, Barratt Impulsivity Scale, or age.
This means that the PD patients were more apathetic, de-
pressed, and anxious and had poorer memory as com-
pared to the healthy participants, which may have
biased the between-group comparisons, but the critical
results from this study are within subjects.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
0
7
/
2
1
0
0
3
/
5
2
1
0
9
3
4
5
9
/
4
1
5
2
7
8
o
3
c
3
n
4
_
1
a
/
_
j
0
o
0
c
8
n
4
0
_
a
p
_
d
0
0
b
8
y
4
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
t
f
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 5. Anti-Hebbian model with one novelty neuron. Numbers to the left show the activity of inputs (for simplicity just four are shown), the
numbers over the four connections indicate their synaptic weights, and the number on the right is the level of neuron’s activity. (A) A novel input
pattern is presented to the neuron, and it outputs an activity of 6. Anti-Hebbian learning takes place decreasing the weights of the active connections
and increasing the inactive weights to balance overall excitation leading to the weights shown in B. (B) The same pattern (now familiar) is presented
again, this time eliciting a lower output activity. (C) Presenting a new novel pattern returns the same output as the original novel pattern because of
the increased weights of the inactive connections balancing the excitation. (D) Simulated PD patients on dopamine during learning show decays in
the weight increase for the inactive connections, but the activated connections remain decreased so the output for the familiar pattern remains the
same. (E) Presentation of the novel pattern from C would now elicit a lowered output, meaning it is more likely to be accepted as familiar. (F)
Simulated PD patients off their medication during learning show decays in both increased and decreased weights, leading to higher activity for the
familiar stimuli and (G) regular activity for novel patterns, corresponding to an increased response bias.
Grogan et al.
2043
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
0
7
/
2
1
0
0
3
/
5
2
1
0
9
3
4
5
9
/
4
1
5
2
7
8
o
3
c
3
n
4
_
1
a
/
_
j
0
o
0
c
8
n
4
0
_
a
p
_
d
0
0
b
8
y
4
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
.
t
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
0 and response bias results for the patients and decay and threshold models. The results of the model
Figure 6. The behavioral and simulated d
fittings; (A) Behavioral d0 and (B) response bias data for PD patients split by Day 1 medications state (Day 1 on vs. Day 1 off ). (C) The simulated d0 for
the best fitting decay model (blue lines are simulated PD Day 1 on medication patients, and red lines are Day 1 off medication). (D) The simulated
response biases for the best fitting decay model (RMSD = 0.0778), showing the same pattern as the behavioral data. The best fitting parameters were
0 for the best fitting threshold model
as follows: Day 1 on: α+ = .1515, α− = 0.8787; Day 1 off: α+ = 0.4079, α− = 0.8702. (E) The simulated d
(RMSD = 0.0620) and (D) the simulated response bias results. The best fitting parameters were as follows: α+ = 0.3685, α− = 0.7562, Day 1 on:
T2 = 1.1594; Day 1 off: T2 = 1.1108. The threshold model has a better fit to the data, based on the lower RMSD, which may be due to Day 1 on d0
being slightly higher than Day 1 off, as in the behavioral data.
2044
Journal of Cognitive Neuroscience
Volume 27, Number 10
From the St. Mary’s Hospital Sleep Questionnaire ad-
ministered on every Day 2 session, we had self-reported
measures of sleep latency, sleep quality, number of hours
of sleep during the night and day, and the total number
of hours. We compared each of these measures with a
two-way repeated-measures ANOVA (Day 1 medication
state × Day 2 medication state). There were no signifi-
cant effects of Day 1 or Day 2 medication states on any
of the sleep measures.
Repetition Effects
As the patients completed the HVLT four times, albeit
with different versions of the HVLT each time, it was pos-
sible that the practice improved their recall and recog-
nition scores so that they performed better during
their fourth condition than their first. We ran repeated-
measures ANOVAs on the 30-min and 24-hr delayed recall
and recognition tests as well as the learning score (third
immediate recall trial – first) to test for this. The ANOVAs
revealed that there were no significant effects of order on
30-min recognition (F(3, 42) = 0.035, p = .991), 24-hr
recognition (F(3, 42) = 2.671, p = .06), 30-min recall
(F(3, 42) = 0.887, p = .456), 24-hr recall (F(3, 42) =
2.516, p = .071), or learning score (F(3, 42) = 1.444,
p = .897). The two 24-hr scores show p values ap-
proaching significance, but this is not due to an increase
in accuracy with repetition but instead a “U”-shaped
curve with the final value similar to the first.
DISCUSSION
We investigated the effect of dopaminergic medication
on memory consolidation in PD patients tested on and
off dopaminergic medications during learning/early con-
solidation and late consolidation/recall. Compared to age-
matched healthy participants, PD patients retain less in-
formation over 30 min and 24 hr than healthy elderly par-
ticipants. Remarkably, for free recall, dopamine during
learning impaired recall at 30 min and 24 hr (but not im-
mediate memory), whereas dopamine between 8 and
24 hr after learning (including a period of sleep) enhanced
recall at 24 hr. Thus, we have demonstrated a benefit of
dopaminergic medication on long-term memory storage
over 24 hr in PD patients. Although dopaminergic medi-
cation state had no overall effect on recognition sensitiv-
ity, dopamine during learning led to a more liberal
response bias 24 hr after learning regardless of dopamine
state during recognition.
Dopaminergic Medication during Learning Impairs
Early Consolidation of Memory
PD patients on medication during learning retained less af-
ter 30-min and 24-hr delays, suggesting that dopaminergic
medication is interfering with early consolidation of infor-
mation within 30 min of encoding and that this deleterious
effect persists despite subsequent changes to dopamine
levels. The effects of dopaminergic medication are unlikely
to reflect a role in encoding as there were no effects of do-
pamine state on immediate memory, which would be ex-
pected if encoding were compromised by dopaminergic
drugs. Also, we used the retention scores for the ANOVAs
on the delayed memory tests, which normalize the number
of words recalled by the maximum number recalled in the
immediate recall trials. This should have removed any ef-
fects of worse encoding of words. The results are not con-
sistent with a Day 1 effect of dopaminergic medication
purely on retrieval processes; such an effect would be sim-
ilar across immediate recall and 30 min and would not affect
performance after a 24-hr delay. Thus, the critical time point
at which dopaminergic activity appears to impair both
30-min and 24-hr recall is early consolidation (within
30 min of encoding). Note that this memory impairment
on dopamine medication is seen despite improvement of
motoric symptoms (see Unified Parkinson’s Disease Rating
Scale in Table 1) and so does not represent a general delete-
rious effect of dopamine medication on the patients’ overall
function, rather dopaminergic medication appears to specifi-
cally interfere with early consolidation of new information.
The only theory that predicts poorer performance
when dopamine is restored in PD is the dopamine over-
dose hypothesis (Cools et al., 2001). This posits that the
brain regions that are relatively spared from the dopami-
nergic cell loss in PD are flooded with dopamine from the
medication, and this impairs normal function. Thus, a
possible explanation is that encoding/early consolidation
mechanisms in brain regions spared the dopaminergic
depletion in PD and are therefore harmed by excess do-
pamine replacement therapy. Although the VTA is usually
implicated in situations such as these because of its rela-
tively preserved dopaminergic projections in PD (when
compared to the substantia nigra; Wittmann et al.,
2013; Cools, 2006; Gasbarri, Sulli, & Packard, 1997; Agid
et al., 1989), in this case the VTA is thought to underpin
improved memory performance when on medication
overnight (see below). Although the VTA can underlie
both encoding/early consolidation and sleep consolida-
tion mechanisms, we would expect that dopamine would
exert either positive or negative effects on both, not the
different directions of effects seen here. Therefore, al-
though the dopamine overdose hypothesis may fit with
the finding of impaired learning when on dopaminergic
medication, the exact underlying neural substrates neces-
sary for this effect are unknown. Neuroimaging experi-
ments offer a way to investigate this by examining
blood flow changes in brain regions during learning
when on and off dopaminergic medication.
Dopaminergic Activity Improves Late
Consolidation of Memory
We found that the number of items retained between
30 min and 24 hr decreased in the patients. However,
Grogan et al.
2045
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
0
7
/
2
1
0
0
3
/
5
2
1
0
9
3
4
5
9
/
4
1
5
2
7
8
o
3
c
3
n
4
_
1
a
/
_
j
0
o
0
c
8
n
4
0
_
a
p
_
d
0
0
b
8
y
4
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
f
.
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
patients who were on medication between 8 and 24 hr
after learning showed a smaller decrease, irrespective of
Day 1 medication state. Thus, dopaminergic medication
during late consolidation and retrieval enhances memory
in PD patients. It appears more likely that dopamine is
exerting effects during consolidation rather than re-
trieval, as retrieval was also required on Day 1 tests, when
dopaminergic medication did not enhance performance.
Although this is the first demonstration in humans of an
effect of dopamine on memory consolidation over 24 hr,
the finding is consistent with animal work (Bethus et al.,
2010; O’Carroll et al., 2006).
When coming off medication, patients spend a mini-
mum of 15 hr without dopaminergic medications before
testing, meaning they are off medication overnight dur-
ing sleep. In addition, on Day 1 all patients are on their
medications from 11 am until 6 pm regardless of Day 1 or
two condition (see Figure 1), meaning that the only time
Day 2 off patients differed from Day 2 on patients was
from 6 pm on Day 1 until testing on Day 2 (around
10 am). This suggests that any dopaminergic medication
effects on 24-hr recall are occurring within this window
(8–24 hr after learning), which coincides with a period
of sleep.
It has been proposed that sleep plays a role in the con-
solidation of memories, and therefore, a lack of dopa-
mine during sleep could impair this process. PD
patients often complain of sleep disturbances (Dhawan,
Healy, Pal, & Chaudhuri, 2006), and dopamine has been
suggested to play a role in the sleep–wake cycle (Rye,
2004) possibly because of the increased phasic firing of
VTA neurons during REM sleep (Lima, 2013; Dahan
et al., 2007). Indeed, D2 antagonist infusion has been
found to decrease REM sleep as well as the levels of plas-
ticity related proteins and memory performance (França
et al., 2015), and the VTA has been shown to activate the
dopaminergic projections to the hippocampus and in-
crease BDNF levels there (Rossato et al., 2009). McNamara
et al. (2014) also found dopaminergic effects on sleep, but
during SWS. They found that optogenetic stimulation of
VTA neurons or their axons onto hippocampal CA1 cells
increased memory performance on a spatial learning task.
This coincided with SWS reactivation of the firing patterns
present during awake exploration. This effect was removed
when a D1/D5 antagonist was infused before the explora-
tion, suggesting that dopaminergic activity during learning
can affect later SWS consolidation via VTA–hippocampal
connections. It may be that a similar process occurs when
dopamine is present during sleep itself. So dopaminergic
consolidation effects have been found in both REM and
SWS; an interesting focus for future work would be unpick-
ing the relative contribution of these two sleep stages to
memory consolidation.
If dopaminergic activity is restored during postlearning
sleep, this would allow the VTA firing to have its full effect
and, because of the VTA-hippocampal projections, affect
consolidation via increased dopamine release onto the
hippocampus. Our self-reported sleep questionnaires
did not yield any significant effects of Day 2 medication
state on sleep (the night when consolidation takes place
after learning); thus, it seems that, although dopamine
may effect overnight consolidation, it does this without
affecting overall self-reported sleep measures such as du-
ration and quality.
An alternative although not mutually exclusive mecha-
nism of overnight dopamine could involve selection of
memories to be consolidated. Evidence for this comes
from a placebo-controlled dopamine agonist study in hu-
mans, which found that a dopamine D2 receptor agonist
given overnight after learning increased memory for pic-
tures associated with low value rewards to the level of ac-
curacy seen for pictures associated with high value
rewards (Feld, Besedovsky, Kaida, Münte, & Born,
2014). This suggests that the greater dopaminergic activ-
ity mimicked the dopamine-dependent consolidation
usually only seen with items associated with rewards
( Wittmann et al., 2005). In other words, the greater do-
paminergic activity selected the low-reward stimuli to be
consolidated in the same way that the high-reward stim-
uli were selected by the dopamine bursts associated with
the rewards. Although our words were not associated
with rewards, it may be that the increased dopaminergic
activity overnight is still selecting these words as though
they had been reward related and thus consolidating
them.
One possible molecular mechanism for the overnight
consolidation effect of dopamine is suggested by the
synaptic-tagging and capture model (Clopath, 2012; Clopath
et al., 2008; Frey & Morris, 1997). In this model, synap-
tic activity induces potentiation or depression of the
synapse, and in order for this to persist, the number
of “tagged” synapses must exceed the threshold that
is set by dopamine levels. This triggers protein synthesis
and causes consolidation of the synaptic weight change.
In this TagTriC model, dopamine lowers the threshold
for the synthesis of a consolidation protein. In the mod-
el, there is only a fairly short time window for this pro-
tein to be synthesized—before the number of tagged
synapses has decayed to below the dopamine threshold
(<4 hr). However, dopamine consolidation over these
longer timescales could operate similarly to that pro-
posed by the TagTriC model, perhaps through a protein
with much slower decay rate, genetic modi-
fications synapses, or downstream proteins do not
depend on number of tagged synapses (e.g., BDNF),
perhaps depending instead previous protein
levels change in weight which has already oc-
curred. Extending current models cover effects
seen at longer help explain our
findings.
One interesting thing is see PD medication
condition most similar healthy participants’ behav-
ior. We might expect on–on condition would
have best overall performance as this has
2046
Journal Cognitive Neuroscience
Volume 27, Number 10
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
>