Dopamine Alters the Fidelity of Working Memory
Representations according to
Attentional Demands
Sean James Fallon1, Nahid Zokaei1, Agnes Norbury2,
Sanjay G. Manohar1,3, and Masud Husain1,3
D
o
w
n
l
o
a
d
e
d
f
r
o
m
Abstract
■ Capacity limitations in working memory (WM) necessitate the
need to effectively control its contents. Here, we examined the
effect of cabergoline, a dopamine D2 receptor agonist, on WM
using a continuous report paradigm that allowed us to assess the
fidelity with which items are stored. We assessed recall perfor-
mance under three different gating conditions: remembering only
one item, being cued to remember one target among distractors,
and having to remember all items. Cabergoline had differential
effects on recall performance according to whether distractors
had to be ignored and whether mnemonic resources could be
deployed exclusively to the target. Compared with placebo, caber-
goline improved mnemonic performance when there were no dis-
tractors but significantly reduced performance when distractors
were presented in a precue condition. No significant difference
in performance was observed under cabergoline when all items
had to be remembered. By applying a stochastic model of response
selection, we established that the causes of drug-induced changes
in performance were due to changes in the precision with which
items were stored in WM. However, there was no change in the
extent to which distractors were mistaken for targets. Thus, D2
agonism causes changes in the fidelity of mnemonic representa-
tions without altering interference between memoranda. ■
INTRODUCTION
Working memory (WM), the ability to store and manipu-
late information in the short term, is a limited capacity
system that is essential to our daily lives (Baddeley,
2012; Oberauer & Hein, 2012). Most studies usually as-
sess WM using binary report measures and examine the
quantity of information that can be maintained, for
example, the number of items that can be recalled. Recent
methodological developments also allow us to measure
the fidelity or quality of information that can be retained
from the latent structure of responding. Although contro-
versial (Ester, Vogel, & Awh, 2012; Zhang & Luck, 2011), it
has been proposed that WM might be best understood as
a finite resource that can be distributed among retained
items, with decrements in recall precision as set size
increases (Ma, Husain, & Bays, 2014; Bays, Catalao, &
Husain, 2009; Bays & Husain, 2008; Alvarez & Cavanagh,
2004; Wilken & Ma, 2004).
An important inference that has emerged from the
use of recall precision measures is that the distribution
of resources can be flexibly altered depending on task
demands. For example, dynamic reallocation of WM re-
source has been observed in both precueing and retro-
cueing experiments: When participants selectively attend
to one item among several others, the precision with
1University of Oxford, 2University of Cambridge, 3John Radcliffe
Hospital, Oxford, UK
which they recall it is significantly enhanced (Zokaei,
Manohar, Husain, & Feredoes, 2014; Zokaei, Ning, Manohar,
Feredoes, & Husain, 2014; Pertzov, Bays, Joseph, & Husain,
2013; Gorgoraptis, Catalao, Bays, & Husain, 2011; Bays &
Husain, 2008). Indeed, precueing can modulate the effect
distractors have on recall such that performance for the
cued item becomes equivalent to the case where no dis-
tractors are present (Gorgoraptis et al., 2011), suggesting
that top–down mechanisms are highly effective in pre-
venting irrelevant items from gaining mnemonic resources.
The neurotransmitter dopamine is a strong candidate
for supporting such reallocation given its long-standing
association with WM maintenance (Rypma et al., 2015;
Eckart, Fuentemilla, Bauch, & Bunzeck, 2014; Fischer
et al., 2010; Vijayraghavan, Wang, Birnbaum, Williams, &
Arnsten, 2007; Floresco, Magyar, Ghods-Sharifi, Vexelman,
& Maric, 2006; Goldman-Rakic, 1995) and the prominent
role attributed to the D2 receptor in controlling the con-
tents of WM (Bloemendaal et al., 2015; Cools & D’Esposito,
2011; Hazy, Frank, & O’Reilly, 2007; Mehta, Manes, Magnolfi,
Sahakian, & Robbins, 2004). Dopaminergic processing in
the striatum may make an essential contribution to WM
by modulating cortical processing (Chatham & Badre,
2015). In attention, it has been demonstrated that the
BG can modulate activity in the sensory cortex by
modulating the connectivity through boosting activity of
task-relevant areas and decreasing activity in task-irrelevant
areas (van Schouwenburg, den Ouden, & Cools, 2013).
© 2017 Massachusetts Institute of Technology. Published under a
Creative Commons Attribution 3.0 Unported (CC BY 3.0) license.
Journal of Cognitive Neuroscience 29:4, pp. 728–738
doi:10.1162/jocn_a_01073
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
9
/
4
2
9
7
/
2
4
8
/
1
7
9
2
5
8
2
/
5
1
2
7
6
8
o
6
c
1
n
5
_
9
a
/
_
j
0
o
1
c
0
n
7
3
_
a
p
_
d
0
1
b
0
y
7
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
.
/
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
More specifically, with regard to WM, a division of labor has
been proposed between the D1-dominated go and the D2
no-go corticostriatal pathways. It has been hypothesized
that, whereas the go pathway allows entry of items into
WM, the no-go pathway prevents it (Hazy et al., 2007; Frank
& O’Reilly, 2006). Thus, D2 receptors might be involved in
controlling the contents of WM by preventing irrelevant in-
formation from gaining access to scarce mnemonic re-
sources. Specifically, under this model, postsynaptic
stimulation of D2 receptors should inhibit the no-go path-
way, inhibiting the gating of information and thus making
WM more open to external input.
However, the impact that these subcortical filtering
mechanisms have on the subsequent mental representa-
tions of memoranda remains to be determined. Previous
studies that have attempted to resolve this question have
employed binary report measures, for example, change
detection tasks requiring a same/different judgment at re-
trieval (Figure 1A). These methods effectively probe WM to
determine whether an item has been retained, whereas re-
cent techniques that measure precision of recall have sug-
gested an alternative view to this all-or-nothing, “quantal”
account (reviewed in Ma et al., 2014). Furthermore, studies
that have varied reward levels suggest that subtle differ-
ences can appear in the corruptibility of items in WM, an
effect that may be dopaminergic in origin (Chumbley,
Dolan, & Friston, 2008). This raises the possibility that D2
receptor stimulation may modulate distractor resistance at
the subitem (i.e., feature) level and affect the resolution
with which certain elements of those items are stored
(Figure 1B). Alternatively, dopamine may have a role in al-
tering interference between the items stored in memory,
for example, on misbinding feature combinations
belonging to different memoranda (Figure 1C). These
two different sources of errors in WM can be teased apart
using a probabilistic model of response selection (Bays
et al., 2009; see also Figure 3A).
Here, we apply such a model to help understand how
dopamine modulates information in WM. We examined
the effect of a D2 agonist, cabergoline, on recall precision
using a double-blind crossover, placebo-controlled design.
We assessed recall performance under three different
gating conditions: remembering only one item, being cued
to remember only one target among distractors, and re-
membering all items (Figure 2). Crucially, we measured
the quality of retained information by probing recall with
an analog, continuous report scale (Gorgoraptis et al.,
2011), rather than a binary one. This allowed us to both
examine raw performance differences induced by caber-
goline and also apply a probabilistic model to dissect out
how different types of errors are affected by drugs.
METHODS
Participants
Nineteen male participants were recruited to take part in
the study (one provided incomplete data because of a
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
9
/
4
2
9
7
/
2
4
8
/
1
7
9
2
5
8
2
/
5
1
2
7
6
8
o
6
c
1
n
5
_
9
a
/
_
j
0
o
1
c
0
n
7
3
_
a
p
_
d
0
1
b
0
y
7
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
/
t
f
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 1. Potential effects of dopamine on WM. (A) Many studies that
have examined the effect of dopamine on WM have participants use a
binary match-to-sample paradigm. (B) Dopamine might modulate the
representation of stored information in ways that cannot readily be
detected using such methodology. For example, the resolution of the
memoranda could be of varying quality or fidelity (fuzzier representations)
but still be sufficient to provide a correct response, that is, a “yes” response
in A could correspond to very different underlying representations.
Binary report measures might fail to detect gradual changes in the fidelity
of stored information with alterations in dopaminergic stimulation.
Dopamine does not have to impact on WM in an all-or-nothing manner.
(C) An alternative modulatory effect of dopamine might be on interference
between the different memoranda, rather than on the quality with
which their features are retained. In this scenario, the fidelity of a mental
representation may be unaffected, but the features that make up the items
may become confused (swapped) during the transition from perception
to memory. For example, although the star was perceived as being
orange, it is remembered as having the color of the other item (blue).
Fallon et al.
729
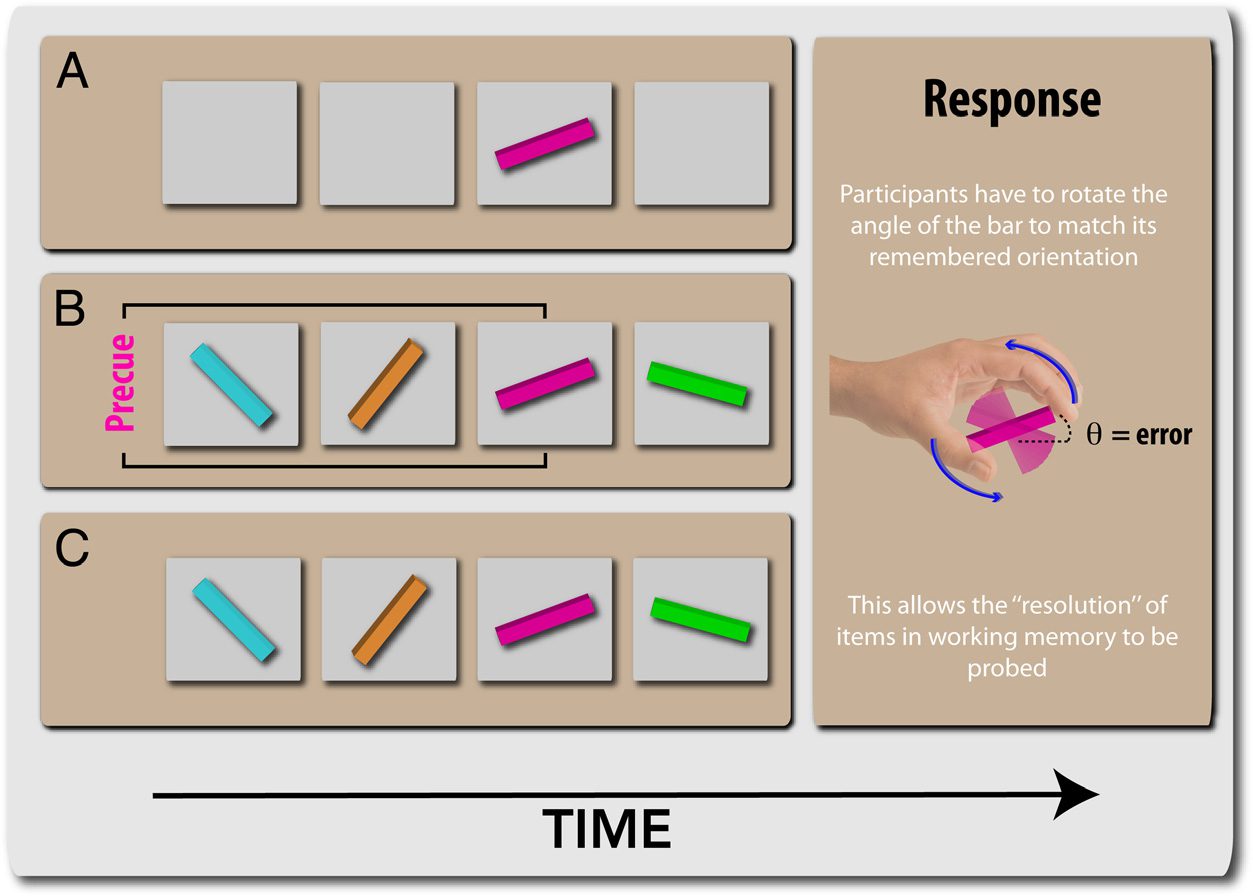
Figure 2. WM tasks.
(A) One-item task: Participants
had to maintain the orientation
of one item for a variable
duration of time before being
probed to reproduce its
orientation using a response
dial, thereby providing a
continuous measure of report
on an analog scale. (B) Cued
task: Participants were asked to
retain only the orientation of
the precued item (in this case,
pink), which was the same
color throughout a block.
(C) Uncued task: Participants
were asked to keep in mind
all four oriented bars and were
asked about one of these at
response phase.
computer error). Demographics are displayed in Table 1.
Exclusion criteria were as follows: any current major ill-
ness, current or historical incident of psychiatric illness,
and recreational drug use on more than one occasion
in the past 6 months. Participants gave written informed
consent, and the study was approved by the local ethics
committee.
Design
The study was a within-participant, double-blind placebo-
controlled design. There were three sessions: baseline,
first testing session, and second testing session. At base-
line, participants were screened for drug contraindications,
gave informed consent, and were familiarized with the
paradigm. On the first and second testing sessions, they
were administered 20 mg of domperidone (an antiemetic),
followed 20 min later by either 1.5 mg of cabergoline or
placebo (drug and placebo tablets were indistinguishable).
A 1.5-mg dose was chosen so as to be greater than that
given in a previous study where inconsistent effects on
cognitive control were observed (1.25 mg; Frank &
Table 1. Participant Demographics
Metric
Age, Years
Years of education
Mean SD Min–Max
26.6
10.2
5.8
2.1
2.8
2.1
18–36
5–14
1–11
8–14
Raven’s advanced progressive matrices
7
Digit Span Forward
11.8
Barratt Impulsivity Scale
67.2 10.7
47–84
Years of education refer to the number of years since leaving U.K.
primary school (∼11 years).
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
9
/
4
2
9
7
/
2
4
8
/
1
7
9
2
5
8
2
/
5
1
2
7
6
8
o
6
c
1
n
5
_
9
a
/
_
j
0
o
1
c
0
n
7
3
_
a
p
_
d
0
1
b
0
y
7
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
f
/
.
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
O’Reilly, 2006), with the addition of domperidone to
mitigate potential physical side effects.
Cabergoline is a long-lasting D2 agonist (Andreotti
et al., 1995). The tasks reported in this study were
administered >2 hr after ingestion. For each test session,
participants completed visual analog scales to measure
mood, affect, physical side effects, and knowledge of
the drug/placebo manipulation. Placebo/drug order was
counterbalanced across participants. There was a mini-
mum washout period of 2 weeks between the two test
sessions.
Tasks
The paradigm and related variants have been used else-
where (Zokaei, McNeill, et al., 2014; Gorgoraptis et al.,
2011). There were three conditions that differed accord-
ing to whether distractor items were present and whether
the targets/distractors were known or unknown (Fig-
ure 2). In the one-item task (Figure 2A), one colored
bar was presented at screen center. Participants’ task
was to remember the orientation of this colored bar. At
the end of the trial, they were shown a probe colored
bar, at a random orientation, with a circle around it to
signal that this was the probe. Participants rotated the
probe stimulus using a dial to match the orientation of
the probed item to their memory. In this condition, there
were four possible delays until the probe appeared.
The uncued and cued conditions were identical except
that, in the cued condition, a 100% valid precue was pre-
sented, which indicated which of the four targets would
be probed. For example, in Figure 2B, the precued item
was pink. The same color was used as the precue
throughout a block. By contrast, in the uncued condition,
participants were not informed about which item they
were going to be tested on and had to retain all four
730
Journal of Cognitive Neuroscience
Volume 29, Number 4
items. In both conditions, they were presented with four
colored bars, displayed sequentially at screen center. The
colors of the bars in each trial were selected randomly,
with the condition that no color was repeated within a
trial, from a set of five, easy-to-discriminate samples
(red, yellow, green, blue, and purple). The orientations
of the four bars were chosen from a flat distribution. In
both conditions, items at each serial position were
probed with equal frequency.
In the one-item condition, participants were shown
only one colored bar, which appeared for 500 msec. To
enable comparison with the other distractor-present con-
ditions, the retention intervals were varied so as to match
the four possible target probe durations that occurred
when four items were presented. Thus, the delay periods
between target offset and probe were 500, 1500, 2500,
and 3500 msec. Note that, in the cued condition, distrac-
tors and targets would be distinguishable to the partici-
pant during encoding and only one item needed to be
retained, similar to the one-item condition.
In each session (drug or placebo), there were 300 trials,
split equally between the three conditions (one-item, cued,
and uncued). Within in each condition, trials were split
between each of the four serial positions (in the case of the
cued and uncued conditions) or delay periods (one-item).
Analysis
The metric of performance was angular error calculated
as the absolute angular difference between target orien-
tation and response orientation. The data were analyzed
using a three-way repeated-measures ANOVA with SPSS
22.0 (IBM, Armonk, NY). Factors included Drug (placebo,
cabergoline), Task (one-item, cued, and uncued), and Se-
rial position (first, second, third, and fourth). Note that,
in the one-item condition, as only a solitary item was
present, Delay duration was used instead. Absolute angu-
lar error was log-transformed to render it suitable to lin-
ear analysis. WM capacity (digit span) and impulsivity as
measured by the Barratt Impulsivity Scale (BIS; Patton,
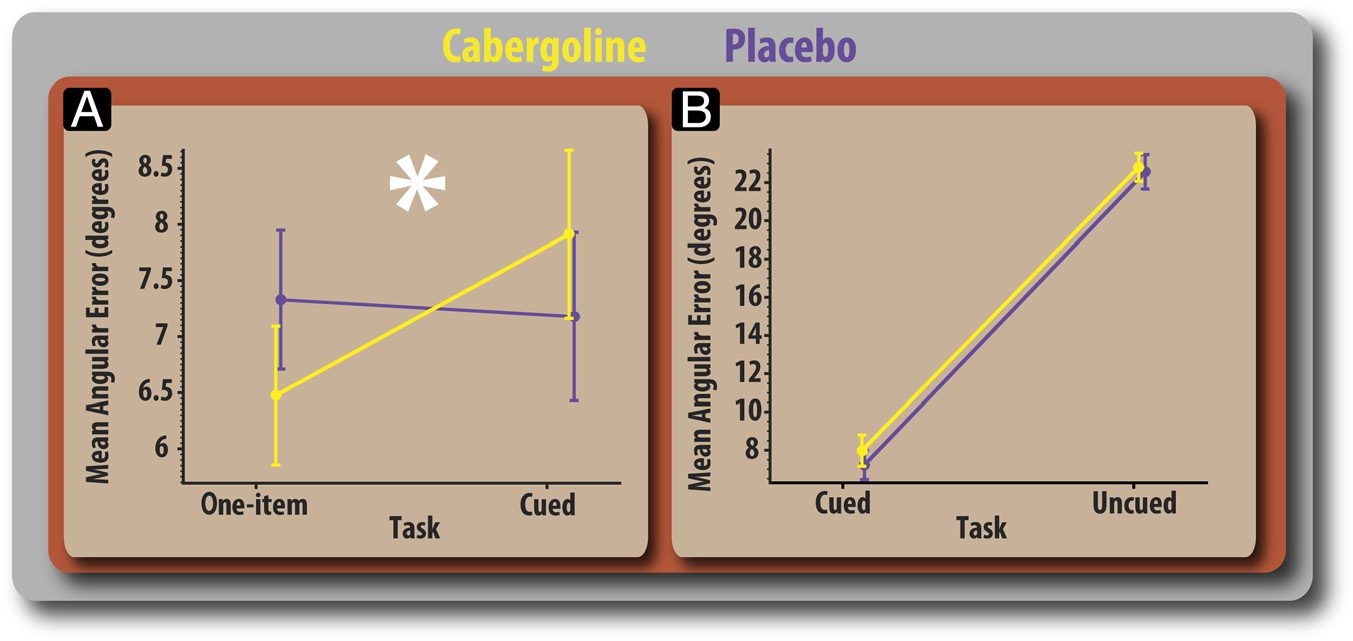
Figure 3. Absolute angular
error. (A) Comparison of
absolute mean angular error
for the one-item and cued
conditions split according to
drug sessions. (B) Comparison
of absolute mean angular
error for cued and uncued
conditions. Error bars reflect
within-participant error
(standard error of the difference
between placebo and drug
sessions).
Stanford, & Barratt, 1995) have previously been used as
proxies for striatal dopamine levels and have been found
to predict the response to dopaminergic drugs (van der
Schaaf, Fallon, ter Huurne, Buitelaar, & Cools, 2013; van
Holstein et al., 2011; Cools, Gibbs, Miyakawa, Jagust, &
D’Esposito, 2008; Cools, Sheridan, Jacobs, & D’Esposito,
2007; Kimberg & D’Esposito, 2003). Accordingly, one of
the supplemental questions we addressed was whether
digit span or impulsivity scores modulated the response
to cabergoline.
Modeling
Angular error gives us a measure of the fidelity of WM
recall. However, it may be equally informative to examine
how the pattern of errors changes with drug administra-
tion and condition. Thus, we fit a mixture model that dis-
sociates different sources of error in memory (introduced
by Bays et al., 2009; see also www.sobell.ion.ucl.ac.uk/
pbays/code/JV10/). The mixture model is described by
the following equation:
(cid:3)
(cid:1)
^θ − θ
X
(cid:3)
m
(cid:1)
^θ − φ
i
ϕκ
(cid:1) (cid:3)
p ^θ
¼ αϕκ
þ β 1
m
i
þ γ 1
2π
This model partitions errors into four different compo-
nents (Figure 3A):
1. Variability in precision (referred to as kappa [κ])
2. Probability of responding to the target orientation
3. Probability of responding to nontargets
4. Probability of guessing
Maximum-likelihood-derived parameters of κ, α, β, and
γ were obtained using expectation maximization (Myung,
2003) produced for each participant (see Bays et al.,
2009, for more details). It should be noted that the three
parameters (α, β, and γ) are not independent as they
must sum to 1.
We extracted the model parameters from each condi-
tion and drug session separately. Thus, there were 100 trials
for each drug session and condition. Because of the
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
9
/
4
2
9
7
/
2
4
8
/
1
7
9
2
5
8
2
/
5
1
2
7
6
8
o
6
c
1
n
5
_
9
a
/
_
j
0
o
1
c
0
n
7
3
_
a
p
_
d
0
1
b
0
y
7
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
.
f
t
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Fallon et al.
731
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
9
/
4
2
9
7
/
2
4
8
/
1
7
9
2
5
8
2
/
5
1
2
7
6
8
o
6
c
1
n
5
_
9
a
/
_
j
0
o
1
c
0
n
7
3
_
a
p
_
d
0
1
b
0
y
7
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
.
f
/
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
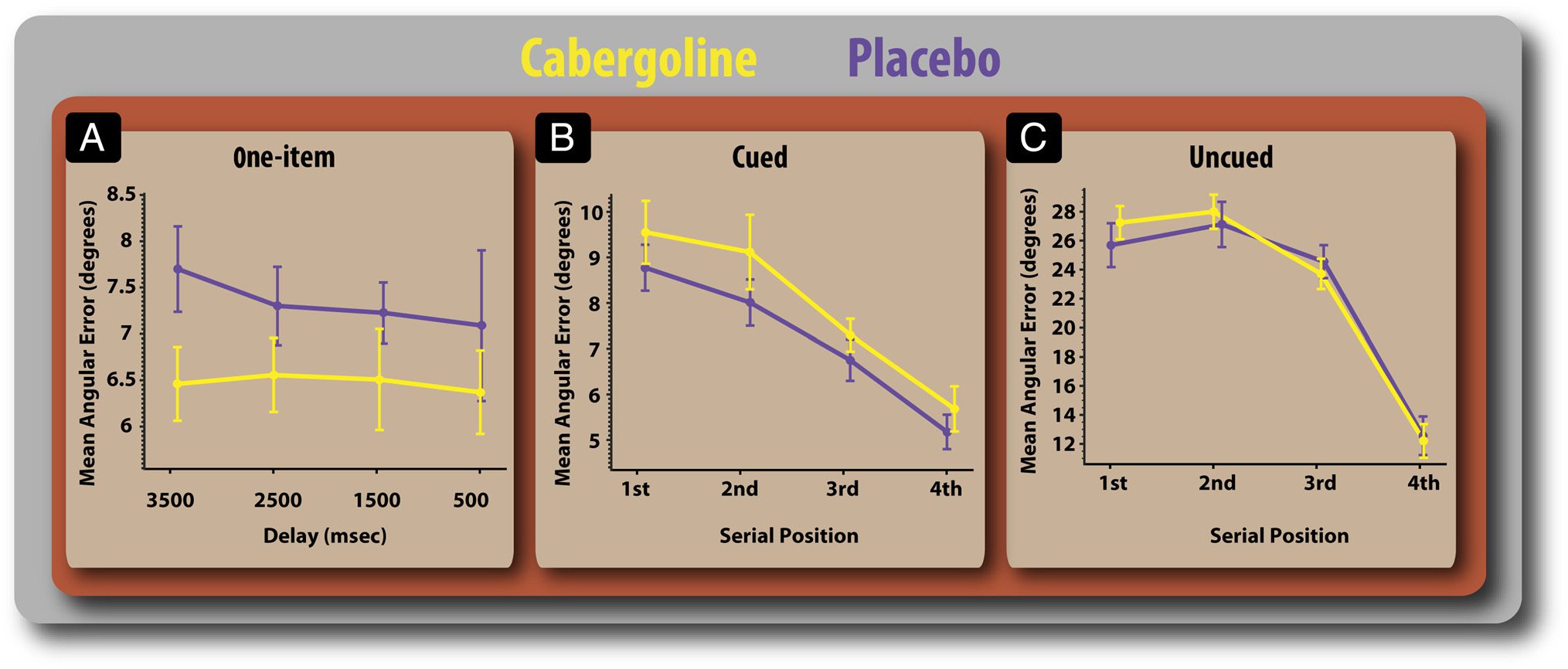
Figure 4. Details of performance in the three tasks on and off cabergoline. (A) Mean error according to retention delay in the one-item task.
Performance according to serial position of probed item in the precued (B) and uncued (C) tasks. There was no interaction between drug and delay
duration or serial position. Error bars reflect SEM.
number of trials, it was not possible to look at the effects of
delay period.
Model Evaluation
We evaluated whether the Bays et al. (2009) model was a
good fit to the data in two ways. First, we evaluated whether
the inclusion of the misbinding parameter (β) for the condi-
tions with more than one possible target (cued and uncued)
was redundant. We did this by comparing the model fit
(Akaike Information Criterion [AIC]) for the cued and uncued
conditions with and without the misbinding parameter. The
model fit (AIC) for the full model (with misbinding) was
11702, and the AIC without the misbinding parameter was
11809. This led to a delta AIC of 107.19 in favor of the model
that contained the misbinding parameter, suggesting that
behavior was best explained in terms of misbinding.
As an additional indication of the goodness of fit, we
compared the AIC for the model with and without the
guessing parameter. The AIC for the full precision model
(Bays et al., 2009) across all three conditions was 13135,
whereas the AIC for the model without the guessing
parameter was 13731, corresponding to an improvement
of 33.1 log units per participant. Thus, the full model pro-
vided a better fit of the data compared with a model that
did not contain the guessing parameter.
RESULTS
Cabergoline-modulated Error according to
Top–Down Attentional Demands
Cabergoline differentially affected recall error as a func-
tion of task, demonstrated by a significant Drug × Task
interaction (F(2, 34) = 4.19, p = .024). The potentially
multifaceted nature of this interaction necessitates that
it is broken down into three separate analyses. The first
considers how drug influences the effect that distractors
have on performance (one-item vs. four-item cued condi-
tions; Figure 3A). The second examines how cabergoline
affects recall when multiple items were presented in the
cued versus uncued conditions, each of which had four
items in the sequence (Figure 3B). Finally, the third con-
siders whether drug modulates the effect of load on re-
call (one-item vs. four-item uncued conditions).
Comparison of recall error (in degrees) for the one-
item versus four-item cued conditions revealed that caber-
goline significantly modulated the effect that distractors
had on report (F(1, 17) = 12.69, p = .002). On placebo,
participants showed no significant difference in perfor-
mance between the two tasks (t(17) = 0.67, p = .521).
In line with previous data (Gorgoraptis et al., 2011), this
demonstrates that people are able to filter distractors very
effectively. After cabergoline, however, recall error was
significantly lower (1.45° on average) for the one-item com-
pared with the four-item cued condition (t(17) = 2.61, p =
.018). Thus, cabergoline improved performance when one
item had to be retained but worsened it in the four-item
precued condition (Figure 3A). Enhanced performance
on cabergoline in the one-item task was observed at all
delays (Figure 4A).
Next, we examined whether cabergoline affected recall
error when multiple (four) items were presented in the
cued versus uncued conditions. Although precueing clearly
improved performance overall, there were no significant
effect of drug in the comparison between cued and uncued
tasks (F(1, 17) = 1.139, p = .30; Figure 3B) and no
evidence that cabergoline affected recall according to
732
Journal of Cognitive Neuroscience
Volume 29, Number 4
load (one-item vs. four-item uncued; F(1, 17) = 0.310,
p = .585).
Thus, the only effect of cabergoline was observed in
the condition where one item presented alone had to
be remembered (performance enhanced) versus the task
in which one item was precued with 100% validity in a
sequence of four items (performance deteriorated).
Recall performance improved on the drug when there
were no distractors but worsened when distractors had
to be gated out in the cued condition. This was not affected
by memory retention interval (Figure 4), indicating that
cabergoline did not alter decay of items stored in WM.
Aside from the Drug × Task interaction, there were no
significant main effect of Drug on error and no significant
interaction between Drug and Delay (Fs < 1). Other non-
drug-related effects were comparable with those observed
on this task previously (Gorgoraptis et al., 2011): Error
was greatly influenced by Task (F(2, 34) = 215.59, p <
.0001), and Serial position had a significant effect on error
(Figure 4A–C; F(3, 51) = 61.36, p < .0001), such that later
items were recalled more accurately—the classical recency
effect. Serial position had greater effects on error in the
uncued condition (Figure 4C; F(6, 102) = 24.73, p <
.0001). However, drug did not significantly affect this
relationship (F < 1).
Cabergoline Modulates the Fidelity of Recall When
Distractors Were Present
Given that the differences in angular error according to
drug and task reported previously might be due to several
factors (reduced precision, reduced responding to target,
responding to nontarget orientations, or increased guess-
ing), we applied a probabilistic model of response selec-
tion (Bays et al., 2009) to tease these components apart
(Figure 5A).
First, we examined kappa, which is the modeled concen-
tration of the response around the target item—a measure
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
4
2
9
7
/
2
4
8
/
1
7
9
2
5
8
2
/
5
1
2
7
6
8
o
6
c
1
n
5
_
9
a
/
_
j
0
o
1
c
0
n
7
3
_
a
p
_
d
0
1
b
0
y
7
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
/
f
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
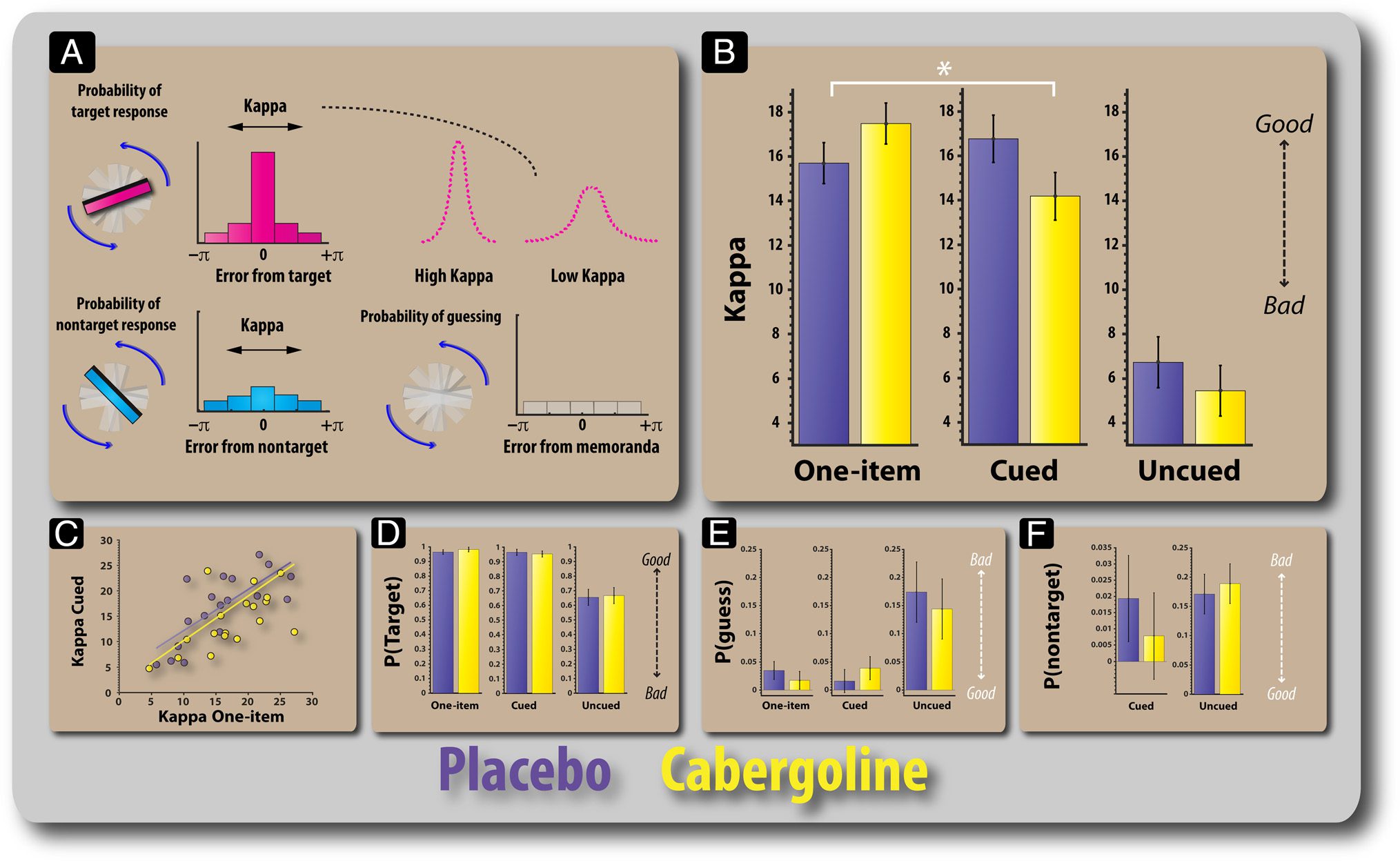
Figure 5. Modeling results for the placebo and cabergoline sessions. (A) Error can arise in recall because of increased variability in remembering the
orientations of the probed item, which is captured in the model in terms of the parameter (kappa). A high kappa corresponds to lower variability in
the precision of retained items (for targets or nontargets). Error is also expected to arise through random guessing/responses. Performance on the
task not only requires an ability to accurately remember the orientations of bars but also the ability to bind, or associate, an orientation to a specific
color. Thus, errors could arise through misbinding remembered orientations with remembered colors. For example, if the pink bar appeared at an
orientation of 40° and the cyan bar appeared at 135°, a misbinding error would be said to have occurred if they rotate the probed pink bar to the
remembered orientation of the cyan bar. (B) Kappa values according to task and drug. There was a significant difference in kappa on the cued
condition and a trend for the one-item condition. (C) Plot showing the relationship between the kappa values for the one-item and precued
conditions. (D) Probability of responding to the target (probed item) according to task and drug session. (E) Probability of random guessing
according to drug and task. (F) Probability of responding to the distractor (cued task) or nontarget (uncued task) orientation. Error bars reflect the
standard error of the difference between placebo and drug sessions.
Fallon et al.
733
of the fidelity of the item representation in WM. A high
kappa indicates a high concentration around the target
item, whereas a low kappa corresponds to a wider distribu-
tion or greater variability. Kappa values were significantly
modulated by drug and task (F(1.5, 25.2) = 4.66, p =
.028; Figure 5B and C). As for raw angular error, this was
driven by a differential effect of cabergoline in the one-item
versus cued conditions (F(1, 17) = 17.54, p = .001), with a
significant difference on the two tasks appearing only on
drug (t(17) = 2.72, p = .014). Moreover, there was a trend
toward higher kappa under cabergoline compared with
placebo in the one-item task (t(17) = 1.933, p = .07). In
the cued condition, kappa was significantly lower on caber-
goline compared with placebo (t(17) = 2.42, p = .027).
For misbinding (cued and uncued), there was no
significant interaction between drug and task (F < 1;
Figure 5E). Thus, there was no evidence that cabergoline
led to increased confusion of the feature dimensions of
memoranda or interference between stored memoranda.
There were no significant differences between drug ses-
sions in the probability of responding to the target (probed
item; Figure 5D) or in chance responding (guessing;
Figure 5E). Thus, the difference in error rate according to
drug in the cued condition appears to be due to reduced
precision of the items stored in WM. In accordance with
the lack of an effect of cabergoline on performance on
the uncued condition, there was no significant effect of
drug on kappa, probability of responding to the target
and nontargets, or guess rate.
Supplemental Analysis
A supplementary question concerns the role of individual
differences in modulating the abovementioned effects of
cabergoline on precision (kappa). To this end, we exam-
ined the role of digit span and impulsivity (BIS score) in
influencing the effects of drug and task by entering these
values as z scored covariates separately into our repeated-
measures ANOVA. Digit span was not found to modulate
the effect of cabergoline on overall precision (F < 1) or
influence, in a three-way manner, the interaction between
Drug and Task (F < 1). Similarly, total impulsivity (BIS
score) did not influence the effect of drug or the inter-
action between Drug and Task (Fs < 1).
DISCUSSION
Gating entry of information into WM is an essential
function that enables humans to perform complex tasks
(D’Esposito & Postle, 2015; Gazzaley & Nobre, 2012;
Braver & Cohen, 2000). Effective gating is important,
given the limited capacity of WM (Fallon, Zokaei, & Husain,
2016), and it has been proposed that dopamine plays a
crucial modulatory role in this process (Hazy et al., 2007;
Frank & O’Reilly, 2006). The present results establish the
importance of the dopaminergic system in facilitating
gating under varying levels of top–down control. In this
study, a D2 agonist, cabergoline, was able to modulate
the resolution with which information held in WM is re-
ported. Cabergoline affected recall performance for a
single item differentially, depending on the presence of
distractors during encoding. Whereas cabergoline im-
proved recall in the absence of distractors, it worsened re-
call when distractors had to be gated out in the cued
condition (Figure 3A and B). This was not affected by the
memory retention interval (Figure 4A–C), indicating that
cabergoline did not alter the decay of items in WM.
The differential effects of drug on error in the one-item
and cued conditions appeared to result from changes in
the quality or fidelity with which relevant items were rep-
resented in memory (Figure 5A). There was no evidence
that cabergoline altered the extent to which participants
became confused between targets and nontargets, that is,
there was no increase in incorrectly conjoining a color
and an orientation—interference (Figure 5F). Taken
together, the results show that cabergoline does not ap-
pear to have a generic enhancing or deleterious effect on
WM. Rather, its effects can be predicted on the basis of
the need to exert top–down attentional control about
what information to encode into memory.
Dopamine has a long association with WM, either through
acting at the level of the frontal cortex (Smith, Swift-Scanlan,
& Boettiger, 2014; Vijayraghavan et al., 2007; Abi-Dargham
et al., 2002; Brozoski, Brown, Rosvold, & Goldman, 1979)
or the striatum (Bäckman et al., 2011; Braskie et al.,
2011; Clatworthy et al., 2009; Cools et al., 2008; Brozoski
et al., 1979). It has also been implicated in aspects of at-
tentional control (Soltani, Noudoost, & Moore, 2013;
Noudoost & Moore, 2011; Chudasama & Robbins, 2004;
Crofts et al., 2001; Servan-Schreiber, Carter, Bruno, &
Cohen, 1998). These twin and often intermingled effects
on selective attention and WM have been reconciled within
computational models and supported by empirical find-
ings that have posited an antagonistic relationship between
stability and flexibility (Fallon & Cools, 2014; Fallon, van der
Schaaf, ter Huurne, & Cools, 2017; Cools & D’Esposito, 2011;
Colzato, Waszak, Nieuwenhuis, Posthuma, & Hommel,
2010; Durstewitz & Seamans, 2008; Hazy et al., 2007;
Müller et al., 2007; Nolan, Bilder, Lachman, & Volavka,
2004). These reports argue that improving stability may
also manifest itself as impaired flexibility.
From the findings of this study, it could be argued that
cabergoline—a D2 agonist—acts to impair selective atten-
tion to task-relevant stimuli but improve WM mainte-
nance. Specifically, in the absence of distractors (pure
maintenance in the one-item condition), recall precision
was relatively enhanced under conditions of heightened
D2 stimulation. In contrast, in the presence of “known dis-
tractors” (cued condition), cabergoline actually impaired
WM performance, by worsening precision. This finding is
particularly congruent with influential theories, which
claim that WM gating occurs through modulation of the
balance between activity of the direct (go) and indirect
(no-go) pathways that link the striatum to cortex (Hazy
734
Journal of Cognitive Neuroscience
Volume 29, Number 4
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
4
2
9
7
/
2
4
8
/
1
7
9
2
5
8
2
/
5
1
2
7
6
8
o
6
c
1
n
5
_
9
a
/
_
j
0
o
1
c
0
n
7
3
_
a
p
_
d
0
1
b
0
y
7
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
/
.
t
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
et al., 2007; Frank & O’Reilly, 2006). These models have
sought to explain how and why certain features are allowed
to enter WM—a function often ascribed to the “central
executive” (Baddeley, 2012). In the context of WM, this is
achieved through a division of labor with activation of the
go pathway permitting current perceptual stimuli to enter
prefrontal-mediated mnemonic networks and activation of
the no-go pathway preventing current perceptual informa-
tion from entering these networks.
On the basis of the Frank and O’Reilly (2006) model, it
might be argued that the effects reported in our study
occur because of the postsynaptic effects of D2 stimula-
tion. Postsynaptic D2 receptors exert a generally inhibi-
tory action on neurons, through actions on adenylyl
cyclase (Nicola, Surmeier, & Malenka, 2000). Within the
Frank and O’Reilly (2006) model, cabergoline, when
acting postsynaptically, is viewed as inhibiting no-go path-
way activity, namely, in removing the net inhibitory output
of the external pallidum and subthalamic nucleus. This
inhibition of the no-go pathway may lead to a preponder-
ance of activity in the go pathway. For example, under
cabergoline, the go, that is, encode, signals from the BG
that accompany the appearance of items on the screen
may be enhanced irrespective of top–down goals and
expectations.
Enhanced, indiscriminate generation of go signals
could explain the present results. In the one-item condi-
tion, augmentation of go signals may enhance the quality
of the representation of the item to be retained, whereas
the same neurophysiological response may have delete-
rious consequences in the cued condition, where go
response to distractors needs to be suppressed. Signals
emanating from the BG have been shown to be endowed
with this capacity. It has already been shown in the
domain of attention that signals in the BG can enhance
activity in task-relevant areas of the cortex and diminish
activity in task-irrelevant areas. Neurophysiologically,
these changes may correspond to changes in neuronal
oscillations at the alpha (10 Hz) frequency, which are
thought to index functional inhibition (Jensen & Mazaheri,
2010) and have previously been found to relate to the
precision with which items are stored in WM (Myers,
Stokes, Walther, & Nobre, 2014) and distractor resistance
(Bonnefond & Jensen, 2012). Thus, stimulation of post-
synaptic D2 receptors with cabergoline, which inhibit
no-go activity, could lead to reduced inhibition of task-
irrelevant processing areas, that is, a failure in functional
inhibition. This could correspond to increased distract-
ability, which is potentially congruent from findings with
Parkinson’s disease where the administration of dopa-
minergic agents can increase distractability (Georgiev
et al., 2015; Cools, Miyakawa, Sheridan, & D’Esposito,
2010).
One finding that may appear puzzling is cabergoline’s
lack of effect on performance in the uncued task (Fig-
ures 3B and 4C). However, this may have occurred not
because cabergoline has no effect on recall in this con-
dition but because the end result of D2 stimulation is to
produce the same effects as is normally the case, that is,
placebo. There is a wealth of evidence to suggest that
items that are presented more recently are recalled with
a higher fidelity (Gorgoraptis et al., 2011) and that the
most recent (last) item presented may have a special—
privileged—state in WM (Cowan, 2011; Oberauer, 2002),
leading it to be recalled with greater accuracy. In Fig-
ure 4C, we see that there are strong serial position effects
in both placebo and cabergoline conditions for the un-
cued task. However, the mechanisms underlying this serial
position effect may be different. Under placebo, the most
recently presented item may be recalled with greater acuity
because of the normative reasons through which recent
items are recalled better (Gorgoraptis et al., 2011). In con-
trast, under cabergoline, the effect of D2 stimulation—
indiscriminate activation of go signals in the BG—may
serve to unwittingly produce the same effect. For example,
during the uncued condition in which the stimuli are
consecutively appearing, the most recently presented item
may keep getting assigned the lion’s share of mnemonic
resources, because of the D2 effects, ultimately producing
a scenario where the last (most recent) items are remem-
bered with greater fidelity. However, because this is also
the pattern that occurs in the normative situation (on pla-
cebo), no drug effects are observed. As such, the uncued
task is unable to discriminate between cabergoline and
placebo. It should be noted, though, that this explanation
is speculative. However, the issue could be resolved em-
pirically by using a retrocue design, whereby participants
are informed, after encoding, which item they are going
to be tested on. These designs have proven effective in
uncovering the mnemonic and neural basis of resource
allocation shifts during WM trials (Myers, Walther, Wallis,
Stokes, & Nobre, 2015; Zokaei, Manohar, et al., 2014).
Future studies should use such designs to unmask the
effect of dopamine on WM resource allocation.
The current study has provided support for the idea
that dopamine can affect the precision with which mental
representations are formed and acted upon in the mind.
This demonstrates that dopamine can have a graded ef-
fect in influencing the fidelity of recall, rather than the
binary effect that is often tacitly assumed. It should be
noted however that, based solely on the present work,
we are unable to specify where—in the processes be-
tween perception and storage—this graded effect origi-
nates. This is important as, although dopamine’s effect
on WM representations may be graded, this may have
been produced by the existence of a binary effect at
some other cognitive subsystem. For example, the effect
of cabergoline on precision in the cued task may have
been caused by a binary (all-or-none) effect on the proba-
bility of updating an item into WM (irrespective of whether
they are targets or distractors). In this scenario, there could
be a lower threshold for updating items into WM, meaning
that there are more items fighting for mnemonic re-
sources, which would produce the reduced precision that
Fallon et al.
735
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
4
2
9
7
/
2
4
8
/
1
7
9
2
5
8
2
/
5
1
2
7
6
8
o
6
c
1
n
5
_
9
a
/
_
j
0
o
1
c
0
n
7
3
_
a
p
_
d
0
1
b
0
y
7
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
.
/
t
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
was observed here. We cannot conclusively rule out that
such mechanisms were responsible for producing the pres-
ent results. However, such an explanation seems incapable
of accommodating all of the results as, even in the one-item
condition where there is no competition for resources, ca-
bergoline was still seen to affect recall. Thus, dopaminergic
stimulation does appear to affect the fidelity of mental
representations.
It is also possible that whether dopamine has a binary or
graded effect on mental representations may vary accord-
ing to what neural loci dopamine is affecting. One compu-
tational rationale for including the BG in WM processes is
that it allows mental representations to be selectively
updated or removed in concert with temporally precise
reward signals in the striatum (Fallon & Cools, 2014;
Chatham & Badre, 2013; O’Reilly & Frank, 2006). In con-
trast, dopaminergic stimulation of the pFC could have an
all-or-none effect on mental representations by totally
destabilizing—collapsing—the prefrontal circuits responsi-
ble for maintaining whole items, similar to what occurs
under stress (Arnsten, 2007). Thus, the precise interplay
between the frontal cortex and BG may determine how
binary or graded dopamine’s effects may be.
Despite being congruent with prior research suggest-
ing that D2 stimulation can modulate distractor resis-
tance, a novel finding from this experiment is that
changes in recall performance occur in the presence of
distractors, crucially without any corresponding change
in the probability of misbinding or interference between
items. Misbinding events occur when different stimuli
features are inappropriately combined together. In the
present context, this would correspond to pairing an ori-
entation with the wrong color. Cabergoline did not signif-
icantly affect misbinding rates on any of the tasks used
here. Thus, D2 agonism did not cause any confusion of
relevant and irrelevant items—a possibility that previous
studies have not been able to authoritatively rule out
(Figure 1C). This suggests that D2 receptors do not mod-
ulate interference between items in WM and, conversely,
that augmenting dopamine levels may have little impact
on treating cognitive impairment where such confusion
between items is prominent. Increased misbinding has,
for example, been associated with damage to the hippo-
campus (Liang et al., 2016; Pertzov, Miller, et al., 2013),
indicating that the medial-temporal lobe may be impor-
tant in correctly combining feature dimensions. It could
be argued, however, that our study is not in a place to
make this conclusion because the task, particularly the
cued condition, was too easy as no misbinding occurred
(Figure 5F). Arguing against this, however, is the fact that
drug effects on misbinding were not found in the uncued
condition, although this task is considerably harder and
does induce substantial misbinding (Figure 5F). More-
over, if cabergoline were to have had severe effects on
selective attention, then performance on the cued condi-
tion would start to resemble the uncued condition, in
that the two conditions were identical except for the fact
that participants were told in the precue condition what
item they are subsequently going to be probed on. Thus,
there was ample room for participant’s performance to
degrade if cabergoline did indeed induce misbinding.
The fact that cabergoline did not increase misbinding
in this condition, however, suggests that the memory-
enhancing effect of the precue is not influenced or per-
turbed by cabergoline. In other words, were cabergoline
to have increased misbinding (confusing targets with
nontargets), then performance would not have been at
ceiling.
The sample size in this study is relatively small. One
concern is that the results could be a false positive. How-
ever, this is unlikely to be the case. The results of this
study are highly congruent with findings from several
previous investigations that have articulated a role for
dopamine, specifically D2 stimulation, in modifying dis-
tractor resistance (Bloemendaal et al., 2015; Cools
et al., 2007; Mehta et al., 2004). They also generally con-
form to the predictions generated by computational
models (Frank & O’Reilly, 2006). The findings of this
study build on previous work by specifying the effect that
distractor suppression has on the underlying mental
representation of relevant information.
In summary, this study has revealed that dopaminergic
D2 agonists can affect the resolution of items in WM,
even when only one item needs to be maintained. How-
ever, the effect of cabergoline on mnemonic representa-
tions appeared to depend greatly on the need to gate the
entry of information into WM. There was no effect of
cabergoline on WM representations when multiple items
needed to be remembered and no gating was required.
In contrast, cabergoline impaired mnemonic representa-
tions of items encoded selectively.
Acknowledgments
This work was funded by a Wellcome Trust Principal Research
Fellowship to M. H. and a fellowship from the British Academy
to N. Z.
Reprint requests should be sent to Sean James Fallon, Depart-
ment of Experimental Psychology, University of Oxford, Oxford
OX13UD, United Kingdom, or via e-mail: sean.fallon@psy.ox.
ac.uk.
REFERENCES
Abi-Dargham, A., Mawlawi, O., Lombardo, I., Gil, R., Martinez,
D., Huang, Y., et al. (2002). Prefrontal dopamine D1
receptors and working memory in schizophrenia. Journal of
Neuroscience, 22, 3708–3719.
Alvarez, G. A., & Cavanagh, P. (2004). The capacity of visual
short-term memory is set both by visual information load and
by number of objects. Psychological Science, 15, 106–111.
Andreotti, A. C., Pianezzola, E., Persiani, S., Pacciarini, M. A.,
Strolin Benedetti, M., & Pontiroli, A. E. (1995).
Pharmacokinetics, pharmacodynamics, and tolerability of
cabergoline, a prolactin-lowering drug, after administration of
increasing oral doses (0.5, 1.0, and 1.5 milligrams) in healthy
736
Journal of Cognitive Neuroscience
Volume 29, Number 4
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
4
2
9
7
/
2
4
8
/
1
7
9
2
5
8
2
/
5
1
2
7
6
8
o
6
c
1
n
5
_
9
a
/
_
j
0
o
1
c
0
n
7
3
_
a
p
_
d
0
1
b
0
y
7
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
.
/
t
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
male volunteers. Journal of Clinical Endocrinology and
Metabolism, 80, 841–845.
Arnsten, A. F. (2007). Catecholamine and second messenger
influences on prefrontal cortical networks of “representational
knowledge”: A rational bridge between genetics and the
symptoms of mental illness. Cerebral Cortex, 17, i6–i15.
Bäckman, L., Nyberg, L., Soveri, A., Johansson, J., Andersson,
M., Dahlin, E., et al. (2011). Effects of working-memory
training on striatal dopamine release. Science, 333, 718.
Baddeley, A. (2012). Working memory: Theories, models, and
controversies. Annual Review of Psychology, 63, 1–29.
Bays, P. M., Catalao, R. F. G., & Husain, M. (2009). The precision
of visual working memory is set by allocation of a shared
resource. Journal of Vision, 9, 7.1–11.
Bays, P. M., & Husain, M. (2008). Dynamic shifts of limited working
memory resources in human vision. Science, 321, 851–854.
Bloemendaal, M., van Schouwenburg, M. R., Miyakawa, A.,
Aarts, E., D’Esposito, M., & Cools, R. (2015). Dopaminergic
modulation of distracter-resistance and prefrontal delay
period signal. Psychopharmacology, 232, 1061–1070.
Bonnefond, M., & Jensen, O. (2012). Alpha oscillations serve to
protect working memory maintenance against anticipated
distracters. Current Biology, 22, 1969–1974.
Braskie, M. N., Landau, S. M., Wilcox, C. E., Taylor, S. D., O’Neil,
J. P., Baker, S. L., et al. (2011). Correlations of striatal
dopamine synthesis with default network deactivations
during working memory in younger adults. Human Brain
Mapping, 32, 947–961.
Braver, T. S., & Cohen, J. D. (2000). On the control of control:
The role of dopamine in regulating prefrontal function and
working memory. In S. Mousell and J. Driver (Eds.), Control
of cognitive processes: Attention and performance XVIII
(pp. 713–737). Cambridge, MA: MIT Press.
Brozoski, T. J., Brown, R. M., Rosvold, H. E., & Goldman, P. S.
(1979). Cognitive deficit caused by regional depletion of
dopamine in prefrontal cortex of rhesus monkey. Science,
205, 929–932.
Chatham, C. H., & Badre, D. (2013). Working memory
management and predicted utility. Frontiers in Behavioral
Neuroscience, 7, 1–12.
Chatham, C. H., & Badre, D. (2015). Multiple gates on working
memory. Current Opinion in Behavioral Sciences, 1, 23–31.
Chudasama, Y., & Robbins, T. W. (2004). Dopaminergic modulation
of visual attention and working memory in the rodent prefrontal
cortex. Neuropsychopharmacology, 29, 1628–1636.
Chumbley, J. R., Dolan, R. J., & Friston, K. J. (2008). Attractor
models of working memory and their modulation by reward.
Biological Cybernetics, 98, 11–18.
Clatworthy, P. L., Lewis, S. J., Brichard, L., Hong, Y. T.,
Izquierdo, D., Clark, L., et al. (2009). Dopamine release in
dissociable striatal subregions predicts the different effects of
oral methylphenidate on reversal learning and spatial
working memory. Journal of Neuroscience, 29, 4690–4696.
Colzato, L. S., Waszak, F., Nieuwenhuis, S., Posthuma, D., &
Hommel, B. (2010). The flexible mind is associated with the
catechol-O-methyltransferase (COMT) Val 158 Met
polymorphism: Evidence for a role of dopamine in the
control of task-switching. Neuropsychologia, 48, 2764–2768.
Cools, R., & D’Esposito, M. (2011). Inverted-U–shaped
dopamine actions on human working memory and cognitive
control. Biological Psychiatry, 69, e113–e125.
Cools, R., Gibbs, S. E., Miyakawa, A., Jagust, W., & D’Esposito,
M. (2008). Working memory capacity predicts dopamine
synthesis capacity in the human striatum. Journal of
Neuroscience, 28, 1208–1212.
Cools, R., Miyakawa, A., Sheridan, M., & D’Esposito, M. (2010).
Enhanced frontal function in Parkinson’s disease. Brain,
133, 225–233.
Cools, R., Sheridan, M., Jacobs, E., & D’Esposito, M. (2007).
Impulsive personality predicts dopamine-dependent changes
in frontostriatal activity during component processes of
working memory. Journal of Neuroscience, 27, 5506–5514.
Cowan, N. (2011). The focus of attention as observed in visual
working memory tasks: Making sense of competing claims.
Neuropsychologia, 49, 1401–1406.
Crofts, H. S., Dalley, J. W., Collins, P., Van Denderen, J. C. M.,
Everitt, B. J., Robbins, T. W., et al. (2001). Differential effects
of 6-OHDA lesions of the frontal cortex and caudate nucleus
on the ability to acquire an attentional set. Cerebral Cortex,
11, 1015–1026.
D’Esposito, M., & Postle, B. R. (2015). The cognitive
neuroscience of working memory. Annual Review of
Psychology, 66, 115.
Durstewitz, D., & Seamans, J. K. (2008). The dual-state theory of
prefrontal cortex dopamine function with relevance to
catechol-o-methyltransferase genotypes and schizophrenia.
Biological Psychiatry, 64, 739–749.
Eckart, C., Fuentemilla, L., Bauch, E. M., & Bunzeck, N. (2014).
Dopaminergic stimulation facilitates working memory and
differentially affects prefrontal low theta oscillations.
Neuroimage, 94, 185–192.
Ester, E. F., Vogel, E. K., & Awh, E. (2012). Discrete resource
limits in attention and working memory. Cognitive
Neuroscience of Attention, 2, 99–112.
Fallon, S. J., & Cools, R. (2014). Reward acts on pFC to enhance
distractor resistance of working memory representations.
Journal of Cognitive Neuroscience, 26, 2812–2826.
Fallon, S. J., van der Schaaf, M. E., ter Huurne, N., & Cools, R.
(2017). The neurocognitive cost of enhancing cognition with
methylphenidate: Improved distractor resistance but impaired
updating. Journal of Cognitive Neuroscience, 29, 652–663.
Fallon, S. J., Zokaei, N., & Husain, M. (2016). Causes and
consequences of limitations in visual working memory.
Annals of the New York Academy of Sciences, 1369, 40–54.
Fischer, H., Nyberg, L., Karlsson, S., Karlsson, P., Brehmer, Y.,
Rieckmann, A., et al. (2010). Simulating neurocognitive aging:
Effects of a dopaminergic antagonist on brain activity during
working memory. Biological Psychiatry, 67, 575–580.
Floresco, S. B., Magyar, O., Ghods-Sharifi, S., Vexelman, C., &
Maric, T. L. (2006). Multiple dopamine receptor subtypes in
the medial prefrontal cortex of the rat regulate set-shifting.
Neuropsychopharmacology, 31, 297–309.
Frank, M. J., & O’Reilly, R. C. (2006). A mechanistic account of
striatal dopamine function in human cognition:
Psychopharmacological studies with cabergoline and
haloperidol. Behavioral Neuroscience, 120, 497–517.
Gazzaley, A., & Nobre, A. C. (2012). Top–down modulation:
Bridging selective attention and working memory. Trends in
Cognitive Sciences, 16, 129–135.
Georgiev, D., Jahanshahi, M., Dreo, J., Čuš, A., Pirtošek, Z., &
Repovš, G. (2015). Dopaminergic medication alters auditory
distractor processing in Parkinson’s disease. Acta
Psychologica, 156, 45–56.
Goldman-Rakic, P. S. (1995). Cellular basis of working memory.
Neuron, 14, 477–485.
Gorgoraptis, N., Catalao, R. F. G., Bays, P. M., & Husain, M.
(2011). Dynamic updating of working memory resources for
visual objects. Journal of Neuroscience, 31, 8502–8511.
Hazy, T. E., Frank, M. J., & O’Reilly, R. C. (2007). Towards an
executive without a homunculus: Computational models of
the prefrontal cortex/basal ganglia system. Philosophical
Transactions of the Royal Society of London, Series B,
Biological Sciences, 362, 1601–1613.
Jensen, O., & Mazaheri, A. (2010). Shaping functional
architecture by oscillatory alpha activity: Gating by inhibition.
Frontiers in Human Neuroscience, 4, 186.
Fallon et al.
737
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
4
2
9
7
/
2
4
8
/
1
7
9
2
5
8
2
/
5
1
2
7
6
8
o
6
c
1
n
5
_
9
a
/
_
j
0
o
1
c
0
n
7
3
_
a
p
_
d
0
1
b
0
y
7
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
.
/
t
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Kimberg, D. Y., & D’Esposito, M. (2003). Cognitive effects of
the dopamine receptor agonist pergolide. Neuropsychologia,
41, 1020–1027.
Liang, Y., Pertzov, Y., Nicholas, J. M., Henley, S. M., Crutch, S.,
Woodward, F., et al. (2016). Visual short-term memory binding
deficit in familial Alzheimer’s disease. Cortex, 78, 150–164.
Ma, W. J., Husain, M., & Bays, P. M. (2014). Changing concepts
of working memory. Nature Neuroscience, 17, 347–356.
Mehta, M. A., Manes, F. F., Magnolfi, G., Sahakian, B. J., &
Robbins, T. W. (2004). Impaired set-shifting and dissociable
effects on tests of spatial working memory following the
dopamine D2 receptor antagonist sulpiride in human
volunteers. Psychopharmacology, 176, 331–342.
Müller, J., Dreisbach, G., Goschke, T., Hensch, T., Lesch, K.-P.,
& Brocke, B. (2007). Dopamine and cognitive control: The
prospect of monetary gains influences the balance between
flexibility and stability in a set-shifting paradigm. European
Journal of Neuroscience, 26, 3661–3668.
Myers, N. E., Stokes, M. G., Walther, L., & Nobre, A. C. (2014).
Oscillatory brain state predicts variability in working memory.
Journal of Neuroscience, 34, 7735–7743.
Myers, N. E., Walther, L., Wallis, G., Stokes, M. G., & Nobre, A. C.
(2015). Temporal dynamics of attention during encoding
versus maintenance of working memory: Complementary
views from event-related potentials and alpha-band
oscillations. Journal of Cognitive Neuroscience, 27, 492–508.
Myung, I. J. (2003). Tutorial on maximum likelihood estimation.
Journal of Mathematical Psychology, 47, 90–100.
Nicola, S. M., Surmeier, D. J., & Malenka, R. C. (2000). Dopaminergic
modulation of neuronal excitability in the striatum and nucleus
accumbens. Annual Review of Neuroscience, 23, 185–215.
Nolan, K. A., Bilder, R. M., Lachman, H. M., & Volavka, J. (2004).
Catechol O-methyltransferase Val158Met polymorphism in
schizophrenia: Differential effects of Val and Met alleles on
cognitive stability and flexibility. American Journal of
Psychiatry, 161, 359–361.
Noudoost, B., & Moore, T. (2011). Control of visual cortical
signals by prefrontal dopamine. Nature, 474, 372–375.
Oberauer, K. (2002). Access to information in working memory:
Exploring the focus of attention. Journal of Experimental
Psychology: Learning, Memory, and Cognition, 28, 411.
Oberauer, K., & Hein, L. (2012). Attention to information in
working memory. Current Directions in Psychological
Science, 21, 164–169.
O’Reilly, R. C., & Frank, M. J. (2006). Making working memory
work: A computational model of learning in the prefrontal
cortex and basal ganglia. Neural Computation, 18, 283–328.
Patton, J. H., Stanford, M. S., & Barratt, E. S. (1995). Factor
structure of the Barratt impulsiveness scale. Journal of
Clinical Psychology, 51, 768–774.
Pertzov, Y., Bays, P. M., Joseph, S., & Husain, M. (2013). Rapid
forgetting prevented by retrospective attention cues. Journal
of Experimental Psychology: Human Perception and
Performance, 39, 1224.
Pertzov, Y., Miller, T. D., Gorgoraptis, N., Caine, D., Schott, J. M.,
Butler, C., et al. (2013). Binding deficits in memory following
medial temporal lobe damage in patients with voltage-gated
potassium channel complex antibody-associated limbic
encephalitis. Brain, 136, 2474–2485.
Rypma, B., Fischer, H., Rieckmann, A., Hubbard, N. A., Nyberg,
L., & Bäckman, L. (2015). Dopamine D1 binding potential
predicts fusiform BOLD activity during face-recognition
performance. Journal of Neuroscience, 35, 14702–14707.
Servan-Schreiber, D., Carter, C. S., Bruno, R. M., & Cohen, J. D.
(1998). Dopamine and the mechanisms of cognition:
Part II. D-amphetamine effects in human subjects performing
a selective attention task. Biological Psychiatry, 43,
723–729.
Smith, C. T., Swift-Scanlan, T., & Boettiger, C. A. (2014). Genetic
polymorphisms regulating dopamine signaling in the
frontal cortex interact to affect target detection under high
working memory load. Journal of Cognitive Neuroscience,
26, 395–407.
Soltani, A., Noudoost, B., & Moore, T. (2013). Dissociable
dopaminergic control of saccadic target selection and its
implications for reward modulation. Proceedings of the
National Academy of Sciences, U.S.A., 110, 3579–3584.
van der Schaaf, M. E., Fallon, S. J., ter Huurne, N., Buitelaar, J., &
Cools, R. (2013). Working memory capacity predicts effects
of methylphenidate on reversal learning.
Neuropsychopharmacology, 38, 2011–2018.
van Holstein, M., Aarts, E., van der Schaaf, M. E., Geurts, D. E.,
Verkes, R. J., Franke, B., et al. (2011). Human cognitive
flexibility depends on dopamine D2 receptor signaling.
Psychopharmacology, 218, 567–578.
van Schouwenburg, M. R., den Ouden, H. E., & Cools, R. (2013).
Selective attentional enhancement and inhibition of fronto-
posterior connectivity by the basal ganglia during attention
switching. Cerebral Cortex, 25, 1527–1534.
Vijayraghavan, S., Wang, M., Birnbaum, S. G., Williams, G. V., &
Arnsten, A. F. (2007). Inverted-U dopamine D1 receptor
actions on prefrontal neurons engaged in working memory.
Nature Neuroscience, 10, 376–384.
Wilken, P., & Ma, W. J. (2004). A detection theory account of
change detection. Journal of Vision, 4, 1120–1135.
Zhang, W., & Luck, S. J. (2011). The number and quality of
representations in working memory. Psychological Science,
22, 1434–1441.
Zokaei, N., Manohar, S., Husain, M., & Feredoes, E. (2014).
Causal evidence for a privileged working memory state in
early visual cortex. Journal of Neuroscience, 34, 158–162.
Zokaei, N., McNeill, A., Proukakis, C., Beavan, M., Jarman, P.,
Korlipara, P., et al. (2014). Visual short-term memory deficits
associated with GBA mutation and Parkinson’s disease.
Brain, 137, 2303–2311.
Zokaei, N., Ning, S., Manohar, S., Feredoes, E., & Husain, M.
(2014). Flexibility of representational states in working
memory. Frontiers in Human Neuroscience, 8, 853.
738
Journal of Cognitive Neuroscience
Volume 29, Number 4
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
4
2
9
7
/
2
4
8
/
1
7
9
2
5
8
2
/
5
1
2
7
6
8
o
6
c
1
n
5
_
9
a
/
_
j
0
o
1
c
0
n
7
3
_
a
p
_
d
0
1
b
0
y
7
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
.
t
/
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1