Deep Predictive Learning in Neocortex and Pulvinar
Randall C. O’Reilly , Jacob L. Russin, Maryam Zolfaghar, and John Rohrlich
Abstract
■ How do humans learn from raw sensory experience?
Throughout life, but most obviously in infancy, we learn without
explicit instruction. We propose a detailed biological mechanism
for the widely embraced idea that learning is driven by the differ-
ences between predictions and actual outcomes (i.e., predictive
error-driven learning). Specifically, numerous weak projections
into the pulvinar nucleus of the thalamus generate top–down pre-
dictions, and sparse driver inputs from lower areas supply the
actual outcome, originating in Layer 5 intrinsic bursting neurons.
Thus, the outcome representation is only briefly activated, roughly
every 100 msec (i.e., 10 Hz, alpha), resulting in a temporal differ-
ence error signal, which drives local synaptic changes throughout
the neocortex. This results in a biologically plausible form of error
backpropagation learning. We implemented these mechanisms
in a large-scale model of the visual system and found that the simu-
lated inferotemporal pathway learns to systematically categorize 3-D
objects according to invariant shape properties, based solely on
predictive learning from raw visual inputs. These categories match
human judgments on the same stimuli and are consistent with
neural representations in inferotemporal cortex in primates. ■
INTRODUCTION
The fundamental epistemological conundrum of how
knowledge emerges from raw experience has challenged
philosophers and scientists for centuries. Although there
have been significant advances in cognitive and computa-
tional models of learning (LeCun, Bengio, & Hinton, 2015;
Watanabe & Sasaki, 2015; Ashby & Maddox, 2011) and in our
understanding of the detailed biochemical basis of synaptic
plasticity (Cooper & Bear, 2012; Lüscher & Malenka, 2012;
Urakubo, Honda, Froemke, & Kuroda, 2008; Shouval, Bear,
& Cooper, 2002), there is still no widely accepted answer to
this puzzle that is clearly supported by known biological
mechanisms and also produces effective learning at the
computational and cognitive levels. The idea that we
learn via an active predictive process was advanced by
Helmholtz in his “recognition by synthesis” proposal (von
Helmholtz, 1867/2013) and has been widely embraced in
a range of different frameworks (de Lange, Heilbron, &
Kok, 2018; Summerfield & de Lange, 2014; Clark, 2013;
George & Hawkins, 2009; Friston, 2005; Hawkins &
Blakeslee, 2004; Rao & Ballard, 1999; Elman et al., 1996;
Dayan, Hinton, Neal, & Zemel, 1995; Kawato, Hayakawa,
& Inui, 1993; Mumford, 1992; Elman, 1990).
Here, we propose a detailed biological mechanism for a
specific form of “predictive error-driven learning” based on
distinctive patterns of connectivity between the neocortex
and the higher-order nuclei of the thalamus (i.e., the pulvinar;
Usrey & Sherman, 2018; Sherman & Guillery, 2006). We hy-
pothesize that learning is driven by the difference between
top–down predictions, generated by numerous weak
University of California Davis
© 2021 Massachusetts Institute of Technology
projections into the thalamic relay cells (TRCs) in the pulvi-
nar, and the actual outcomes supplied by sparse, strong
driver inputs from lower areas. Because these driver inputs
originate in Layer 5 intrinsic bursting (5IB) neurons, the
outcome is only briefly activated, roughly every 100 msec
(i.e., 10 Hz, alpha). Thus, the prediction error is a temporal
difference in activation states over the pulvinar, from an
earlier prediction to a subsequent burst of outcome. This
temporal difference can drive local synaptic changes
throughout the neocortex, supporting a biologically plausi-
ble form of error backpropagation (Bp) that improves the
predictions over time (Lillicrap, Santoro, Marris, Akerman,
& Hinton, 2020; Whittington & Bogacz, 2019; Bengio,
Mesnard, Fischer, Zhang, & Wu, 2017; O’Reilly, 1996;
Hinton & McClelland, 1988; Ackley, Hinton, & Sejnowski,
1985). The temporal difference form of error-driven learn-
ing contrasts with prevalent alternative hypotheses that
require a separate population of neurons to compute a pre-
diction error explicitly and transmit it directly through neu-
ral firing (Lotter, Kreiman, & Cox, 2016; Ouden, Kok, &
Lange, 2012; Friston, 2005, 2010; Rao & Ballard, 1999;
Kawato et al., 1993).
In the following, our primary objective is to describe the
hypothesized biologically based mechanism for predictive
error-driven learning, contrast it with other existing pro-
posals regarding the functions of this thalamocortical cir-
cuitry and other ways that the brain might support
predictive learning, and evaluate it relative to a wide range
of existing anatomical and electrophysiological data. We
provide a number of specific empirical predictions that
follow from this functional view of the thalamocortical
circuit, which could potentially be tested by current neu-
roscientific methods. Thus, this work proposes a clear
Journal of Cognitive Neuroscience 33:6, pp. 1158–1196
https://doi.org/10.1162/jocn_a_01708
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
1
5
8
1
9
1
3
5
7
7
/
/
j
o
c
n
_
a
_
0
1
7
0
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
functional interpretation of this distinctive thalamocortical
circuitry that contrasts with existing ideas in testable ways.
A second major objective is to implement this predictive
error-driven learning mechanism in a large-scale computa-
tional model that faithfully captures its essential biological
features, to test whether the proposed learning mecha-
nism can drive the formation of cognitively useful repre-
sentations. In particular, we ask a critical question for
any predictive learning model: Can it develop high-level,
abstract representations while learning from nothing but
predicting low-level visual inputs? Most visual object rec-
ognition models that provide a reasonable fit to neuro-
physiological data rely on large human-labeled data sets
to explicitly train abstract category information via error
B (Rajalingham et al., 2018; Cadieu et al., 2014; Khaligh-
Razavi & Kriegeskorte, 2014). Thus, it is perhaps not too
surprising that the higher layers of these models, which
are closer to these category output labels, exhibited a
greater degree of categorical organization.
Through large-scale simulations based on the known
structure of the visual system, we found that our biologically
based predictive learning mechanism developed high-level,
abstract representations that significantly diverge from
the similarity structure present in the lower layers of the
network and systematically categorize 3-D objects accord-
ing to invariant shape properties. Furthermore, we found
in an experiment using the same stimuli that these cate-
gories match human similarity judgments and that they
are also qualitatively consistent with neural representa-
tions in inferotemporal (IT) cortex in primates (Cadieu
et al., 2014). In addition, we show that comparison predic-
tive Bp models lacking these biological features (Lotter
et al., 2016) did not learn object categories that go beyond
the visual input structure. Thus, there may be some im-
portant features of the biologically based model that en-
able this ability to learn higher-level structure beyond that
of the raw inputs.
It is important to emphasize that our objectives for these
simulations are not to produce a better machine-learning
algorithm per se but rather to test whether our biologically
based model can capture some of the known high-level,
cognitive phenomena that the mammalian brain learns.
Thus, we explicitly dissuade readers from the inevitable
desire to evaluate the importance of our model based on
differences in narrow, performance-based machine learning
metrics. As discussed later, there are various engineering-
level issues regarding the biologically based model’s com-
putational cost and performance, which currently limit its
ability to compete with simpler, much larger-scale Bp
models, but we do not think these are relevant to the
evaluation of the scientific questions of relevance here. In
short, this model is an instantiation of a scientific theory,
and it should be evaluated on its ability to explain a wide
range of data across multiple levels of analysis, just as every
other scientific theory is evaluated.
The remainder of the paper is organized as follows. First,
we provide a concise overview of the biologically based pre-
dictive error-driven learning framework, including the most
relevant neural data. Then, we present a small-scale imple-
mentation of the model that learns a probabilistic grammar,
to illustrate the basic computational mechanisms of the
theory. This is followed by the large-scale model of the
visual system, which learns by predicting over brief movies
of 3-D objects rotating and translating in space. We evaluate
this model and compare it to two other predictive learning
models that directly use error Bp, based on current deep
convolutional neural network (DCNN) mechanisms. Then,
we circle back to discuss the relevant biological data in
greater detail, along with testable predictions that can
differentiate this account from other existing ideas.
Finally, we conclude with a discussion of related models
and outstanding issues.
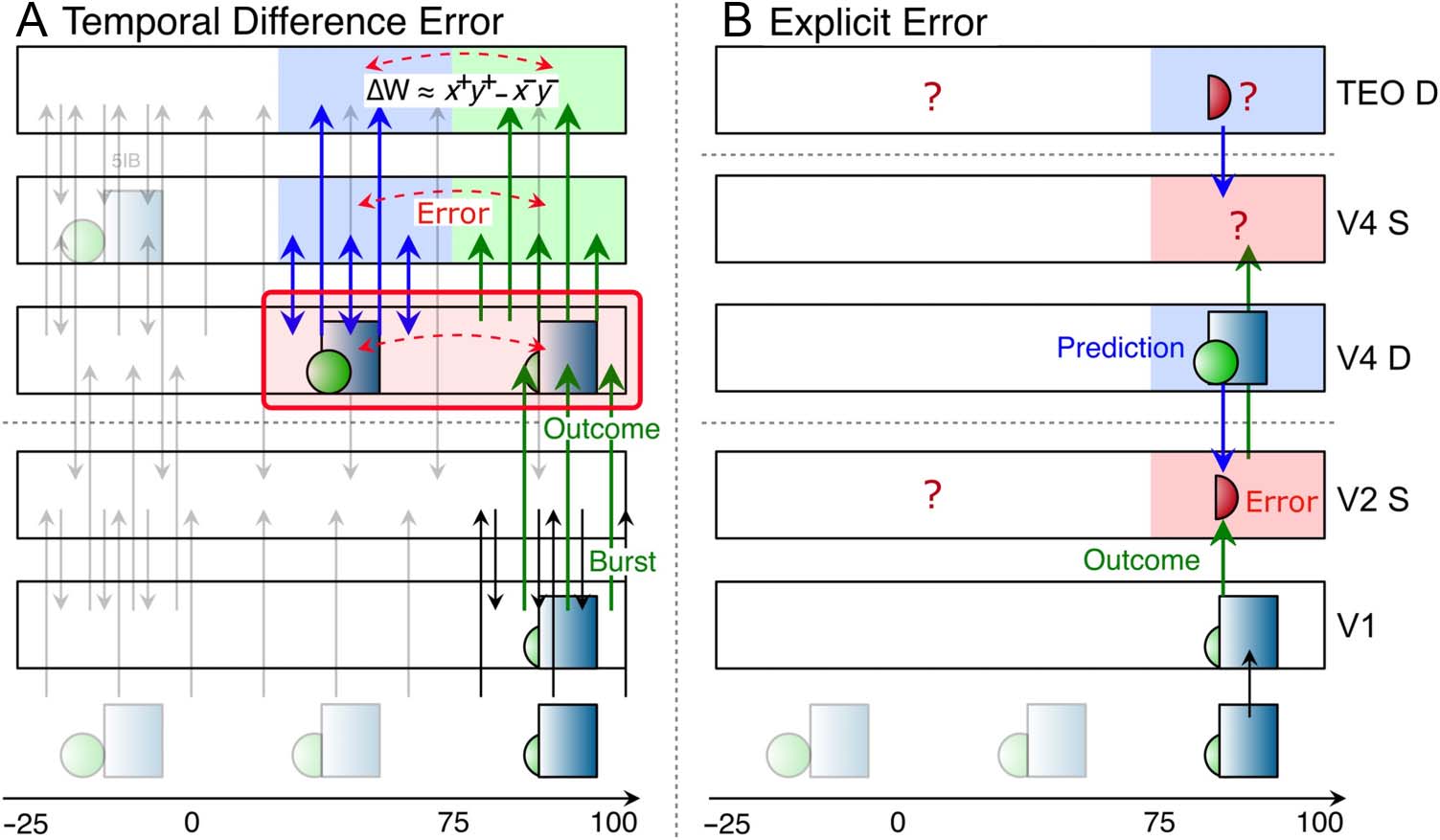
PREDICTIVE ERROR-DRIVEN LEARNING IN
THE NEOCORTEX AND PULVINAR
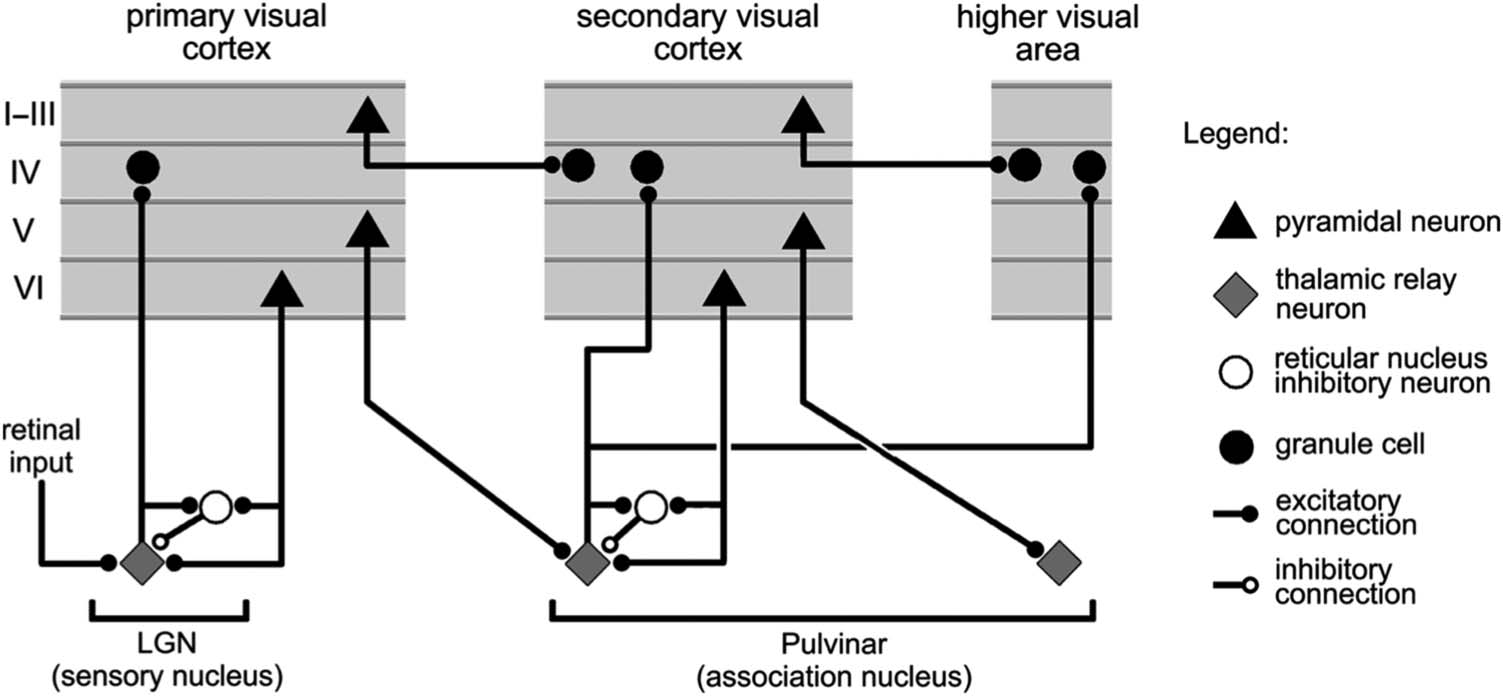
Figure 1 shows the thalamocortical circuits characterized by
Sherman and Guillery (2006; see also Usrey & Sherman,
2018; Sherman & Guillery, 2013), which have two distinct
projections converging on the principal TRCs of the pulvinar,
Figure 1. Summary figure from
Sherman and Guillery (2006)
showing the strong feedforward
driver projection emanating
from layer 5IB cells in lower
layers (e.g., V1) and the much
more numerous feedback
“modulatory” projection from
layer 6CT (corticothalamic)
cells. We interpret these
same connections as providing
a prediction (6CT) versus
outcome (5IB) activity pattern
over the pulvinar.
O’Reilly et al.
1159
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
1
5
8
1
9
1
3
5
7
7
/
/
j
o
c
n
_
a
_
0
1
7
0
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
1
5
8
1
9
1
3
5
7
7
/
/
j
o
c
n
_
a
_
0
1
7
0
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
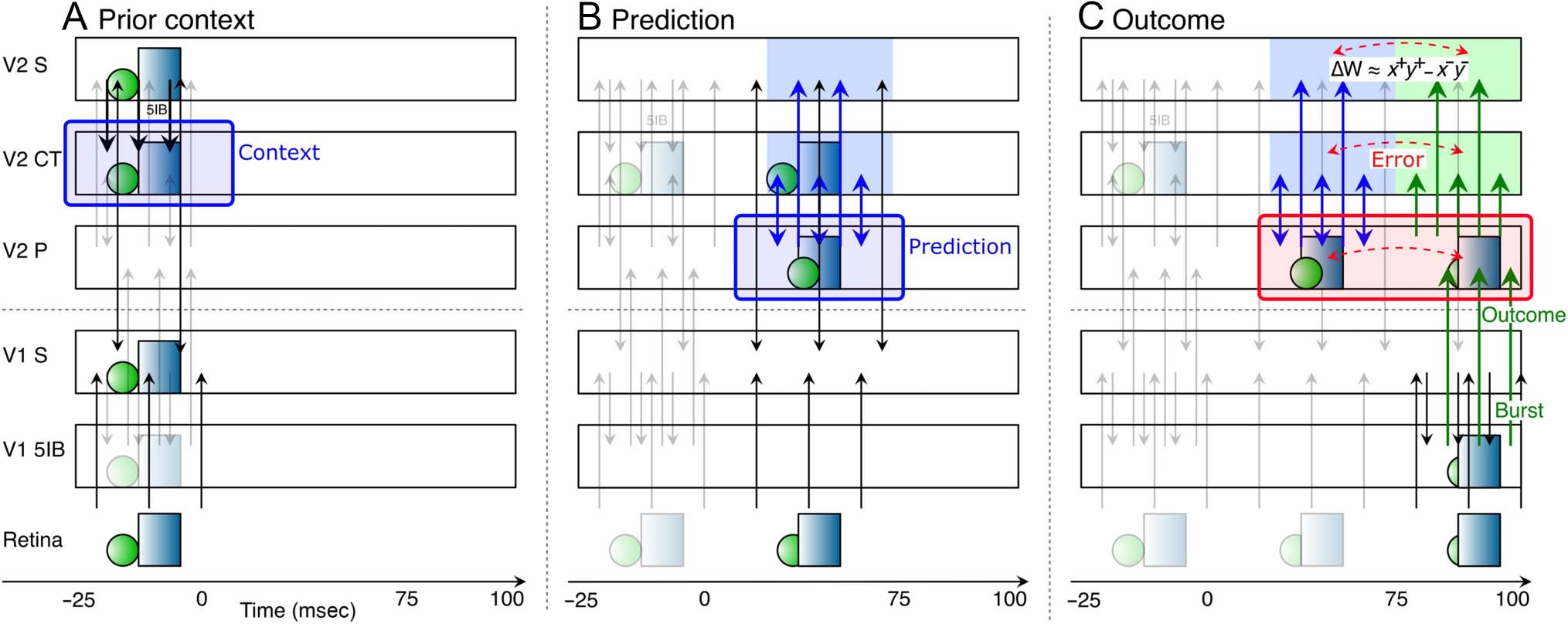
Figure 2. Corticothalamic (CT) information flow under our predictive learning hypothesis, shown as a sequence of movie frames (“Retina”), illustrating
the three key steps taking place within a single 125-msec time window, broken out separately across the three panels: (A) Prior context is updated in
the V2 CT layer, (B) which is then used to generate a prediction over the pulvinar ( V2 P), (C) against which the outcome, driven by bottom–up 5IB,
represents the prediction error as a temporal difference between the prediction and outcome states over the pulvinar. Changes in synaptic weights
(learning) in all superficial (S) and CT layers are driven from the local temporal difference experienced by each neuron, using a form of the CHL term
as shown, where the “+”: superscripts indicate outcome activations and “−” superscripts indicate prediction. CHL approximates the backpropagated
prediction error gradient experienced by each neuron (O’Reilly, 1996), reflecting both direct pulvinar error signals and indirect corticocortical error
signals as well. In specific, (A) CT context updating occurs via 5IB (not shown) in higher layer ( V2) during prior alpha (100-msec) cycle—this context
is maintained in the CT layer and used to generate predictions. (B) The prediction over pulvinar is generated via numerous top–down CT projections.
This prediction state also projects up to S and CT layers, and from S to all other S layers via extensive bidirectional connectivity, so their activation
state reflects this prediction as well. (C) The subsequent outcome drives pulvinar activity bottom–up via V1 5IB and is likewise projected to S and CT
layers, ensuring that the relevant temporal difference error signal is available locally in the cortex. The difference in activation values across these two
time points, in S and CT layers throughout the network, drives learning to reduce prediction errors. Note that the single most important property of
the 5IB is that these driver cells are not active during the prediction phase—the bursting itself may also be useful in the driving property, but that is a
secondary consideration to the critical feature of having a time when the prediction alone can be projected onto the pulvinar.
the primary thalamic nucleus that is interconnected with
higher-level posterior cortical visual areas (Halassa & Kastner,
2017; Arcaro, Pinsk, & Kastner, 2015; Shipp, 2003). One
projection consists of numerous, weaker connections
originating in deep layer VI of the neocortex (the 6CT
corticothalamic projecting cells), which we hypothesize
generate a top–down prediction on the pulvinar. The other
is a sparse (Rockland, 1996, 1998) and strong driver path-
way that originates from lower-level layer 5IB cells, which
we hypothesize provide the outcome. These 5IB neurons
fire discrete bursts with intrinsic dynamics having a period
of roughly 100 msec between bursts (Saalmann, Pinsk,
Wang, Li, & Kastner, 2012; Larkum, Zhu, & Sakmann,
1999; Franceschetti et al., 1995; Silva, Amitai, & Connors,
1991; Connors, Gutnick, & Prince, 1982), which is thought
to drive the widely studied alpha frequency of ∼10 Hz that
originates in cortical deep layers and has important effects
on a wide range of perceptual and attentional tasks (Clayton,
Yeung, & Kadosh, 2018; Jensen, Bonnefond, & VanRullen,
2012; Buffalo, Fries, Landman, Buschman, & Desimone,
2011; Mathewson, Gratton, Fabiani, Beck, & Ro, 2009;
VanRullen & Koch, 2003). Critically, unlike many other such
bursting phenomena, this 5IB occurs in awake animals
(Luczak, Bartho, and Harris, 2009, 2013; Sakata & Harris,
2009, 2012), consistent with the presence of alpha in
awake, behaving states.
The existing literature generally characterizes the 6CT pro-
jection as modulatory (Usrey & Sherman, 2018; Sherman &
Guillery, 2013), but a number of electrophysiological record-
ings from awake, behaving animals clearly show sustained,
continuous patterns of neural firing in pulvinar TRC neurons,
which is not consistent with the idea that they are only
being driven by their phasic bursting 5IB inputs (Zhou,
Schafer, & Desimone, 2016; Komura, Nikkuni, Hirashima,
Uetake, & Miyamoto, 2013; Saalmann et al., 2012; Bender
& Youakim, 2001; Robinson, 1993; Petersen, Robinson, &
Keys, 1985; Bender, 1982). Indeed, these recordings show
that pulvinar neural firing generally resembles that of the
visual areas with which they interconnect, in terms of neural
receptive field properties, tuning curves, and so forth. This
is important because our predictive learning framework
requires that these 6CT top–down projections be capable
of directly driving TRC activity. Specifically, in contrast to
the standard view, the core idea behind our theory is that
the top–down 6CT projections drive a predicted activity
pattern across the extent of the pulvinar, which precedes
the subsequent outcome activation state driven by the
strong 5IB inputs.
1160
Journal of Cognitive Neuroscience
Volume 33, Number 6
Figure 2 illustrates the temporal evolution of activity
states according to our predictive learning theory, which
is somewhat challenging to convey because the critical
signals driving learning unfold over time (O’Reilly, Wyatte,
& Rohrlich, 2014, 2017; Kachergis, Wyatte, O’Reilly, de
Kleijn, & Hommel, 2014). We hypothesize that synaptic
plasticity throughout the cortex is sensitive to the resulting
temporal differences that emerge initially in the pulvinar.
Thus, unlike other models (as we discuss in depth later),
the prediction error here is not captured directly in the
firing of a special population of error-coding neurons but
rather remains as a temporal difference error signal.
Figure 2 shows a single 125-msec time window of a
100-msec alpha cycle for the purposes of illustration
(the actual timing is likely to be more dynamic as dis-
cussed next). The activity state in pulvinar TRC neurons,
representing a prediction, as driven by the top–down 6CT
projections, should develop during the first ∼75 msec,
when the 5IB neurons are paused between bursting.
Then, the final ∼25 msec largely reflects the strong 5IB
bottom–up ground-truth driver inputs when they burst.
Thus, the prediction error signal is reflected in the tempo-
ral difference of these activation states as they develop
over time. In other words, our hypothesis is that the pul-
vinar is directly representing either the top–down predic-
tion or the bottom–up outcome at any given time, and
the temporal difference between these states implicitly
encodes a prediction error. Whereas the deep 6CT layer
is involved in generating a top–down prediction over the
pulvinar, the superficial layer neurons continuously rep-
resent the current state, simultaneously incorporating
bottom–up and top–down constraints via their own con-
nections with other areas. To ensure that the prediction
is not directly influenced by this current state represen-
tation (i.e., “peeking at the right answer”), it is important
that the 6CT neurons encode temporally delayed informa-
tion, consistent with available data (Harris & Shepherd,
2015; Thomson, 2010; Sakata & Harris, 2009).
The actual biological system is likely to be much more
dynamic than the simplistic cartoon with rigid 100-msec
timing, as shown in Figure 2, based on a set of neural mech-
anisms that can work together to enable it to more flexibly
entrain the predictive learning cycle to the environment.
These mechanisms would also tend to increase activity
and learning associated with unexpected outcomes relative
to expected ones, consistent with the observed expectation
suppression phenomena (Bastos et al., 2012; Meyer &
Olson, 2011; Todorovic, van Ede, Maris, & de Lange, 2011;
Summerfield, Trittschuh, Monti, Mesulam, & Egner, 2008).
Specifically, various underlying mechanisms result in
neural adaptation, which is generally thought to increase
neural activity and learning associated with novel inputs rel-
ative to recently familiar ones (Hennig, 2013; Grill-Spector,
Henson, & Martin, 2006; Brette & Gerstner, 2005; Müller,
Metha, Krauskopf, & Lennie, 1999; Abbott, Varela, Sen, &
Nelson, 1997). In the case where outcomes are consistent
with prior predictions (i.e., the predictions are accurate),
the same population of neurons across pulvinar and cor-
tex should be active over time, whereas unpredicted out-
comes will generally activate new subsets of neurons in
superficial cortical layers representing the current state.
Thus, because of adaptation, there should be a phasic in-
crease in activity in these superficial neurons at the onset
of unpredicted stimuli relative to predicted ones. Furthermore,
the 5IB neurons downstream of these superficial neurons
may be particularly responsive to these phasic activity in-
creases, causing their bursting to coincide preferentially
with unexpected outcomes, thereby driving the phase re-
setting of the alpha cycle to such events. Thus, during a
sequence of predicted states, the pulvinar may experience
relatively weaker or even absent 5IB driving inputs, until
an unpredicted stimulus arises. At this point, error-driven
learning would be more strongly engaged as a function of
the phasic release from adaptation and 5IB burst activa-
tion. We discuss these dynamics more later in the context
of the comparison with explicit error (EE) coding models.
We also hypothesize that 5IB preferentially drives the
synaptic plasticity processes to take place at that time,
because of the strong driving nature of the outputs from
these neurons. In computational terms originating with
the Boltzmann machine (Hinton & Salakhutdinov, 2006;
Ackley et al., 1985), this anchors the target or plus phase
to be at this point of 5IB. Furthermore, this means that
the predictive nature of the prior minus phase naturally
emerges just by virtue of it being the state before 5IB:
The learning rule automatically causes that prior state to
better anticipate the subsequent state. Thus, even if no
prediction was initially generated, learning over multiple
iterations will work to create one, to the extent that a reli-
able prediction can be generated based on internal states
and environmental inputs. Likewise, assuming relevant
activity traces naturally persist over timescales longer than
the alpha cycle, this predictive learning process can take
advantage of any such remaining traces to learn across
these longer timescales, although it is operating at the
faster alpha scale.
In short, learning always happens whenever something
unexpected occurs, at any point, and drives the develop-
ment of predictions immediately prior, to the extent such
predictions are possible to generate. In the typical labo-
ratory experiment where phasic stimuli are presented
without any predictable temporal sequence (which is
uncharacteristic of the natural world), there may often
be no significant prediction before stimulus onset, and
we would expect such stimuli to reliably drive 5IB, which
is consistent with available electrophysiological data
(Zhou et al., 2016; Komura et al., 2013; Luczak et al., 2009,
2013; Bender & Youakim, 2001; Robinson, 1993; Petersen
et al., 1985; Bender, 1982). Thus, unlike Figure 2, such
situations would start with a 5IB-triggered plus phase,
without a significant minus phase before that.
As may be evident by this point, we are mainly focused on
prediction in the sense of the humorous quote: “Prediction
is very difficult, especially about the future” (attributable
O’Reilly et al.
1161
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
1
5
8
1
9
1
3
5
7
7
/
/
j
o
c
n
_
a
_
0
1
7
0
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
to Danish author Robert Storm Petersen), whereas this
term is potentially confusingly used in a much broader
sense in most Bayesian-inspired predictive coding frame-
works (de Lange et al., 2018; Friston, 2005; Rao & Ballard,
1999). These frameworks use “prediction” to encompass
everything from genetic biases to the results of learning in
the feedforward synaptic pathways to top–down filling-in
or biasing of the current stimulus properties and fairly
rarely use it in the “about the future” sense. We think these
different phenomena are each associated with different
neural mechanisms at different timescales (O’Reilly, Hazy,
& Herd, 2016; O’Reilly, Wyatte, Herd, Mingus, & Jilk, 2013;
O’Reilly, Munakata, Frank, Hazy, & Contributors, 2012) and
thus prefer to treat them separately, while also recognizing
that they can clearly interact as well.
Thus, our use of the term “prediction” here refers specif-
ically to “anticipatory” neural firing that predicts subsequent
stimuli. We use the term “postdiction” to refer to the oper-
ation of this predictive mechanism after a stimulus has been
initially processed (to consolidate and more deeply encode,
as in an autoencoder model) and distinguish both from
top–down excitatory biasing, which directly influences the
online superficial layer neural representations of the cur-
rent stimulus (O’Reilly et al., 2013; Miller & Cohen, 2001;
Reynolds, Chelazzi, & Desimone, 1999; Desimone &
Duncan, 1995). Finally, many discussions of prediction
error in the literature include late, frontally associated pro-
cesses such as those associated with the P300 ERP compo-
nent (Holroyd & Coles, 2002). We specifically exclude these
from the scope of the mechanisms described here, which
are anticipatory, fast, and low level, as is appropriate for
the posterior cortical sensory processing areas that inter-
connect with the pulvinar.
Computational Properties of Predictive Learning in
the Thalamocortical Circuits
We next elaborate the connections between the computa-
tional properties required for predictive learning and the
properties of the circuits interconnecting the cortex and
the pulvinar, which appear to be notably well suited for
their hypothesized role in predictive learning. We begin
with a relatively established interpretation of superficial
layer processing, to contextualize subsequent points
about the special functions required of the deep layers
and the thalamus.
(cid:129) The superficial cortical layers continuously represent
the current state: The superficial layer pyramidal neu-
rons are densely and bidirectionally interconnected
with other cortical areas and update quickly to new
stimulus inputs, with continuous, relatively rapid firing
(i.e., up to about 100 Hz for preferred stimuli). These
neurons integrate higher-level top–down information
with bottom–up sensory information to resolve ambi-
guities, focus attention, fill in missing information, and
generally enhance the consistency and quality of the
online representations (O’Reilly, Hazy, & Herd, 2016;
O’Reilly, Wyatte, Herd, Mingus, & Jilk, 2013; O’Reilly
et al., 2012; Miller & Cohen, 2001; Reynolds et al.,
1999; Desimone & Duncan, 1995; Hopfield, 1984;
Rumelhart & McClelland, 1982). As noted above, we
distinguish this form of top–down processing, which
is often most evident during the period after stimulus
onset (Lee & Mumford, 2003), from the specifically
predictive, anticipatory sort.
(cid:129) Predictions must be insulated against receiving current
state information (it is not prediction if you already
know what happens): Given that the superficial layers
are continuously updating and representing the cur-
rent state, some kind of separate neural system insu-
lated from this current state information must be used
to generate predictions; otherwise, the prediction sys-
tem can just “cheat” and directly report the current
state. It may seem counterintuitive, but making the pre-
diction task harder is actually beneficial, because that
pushes the learning to capture deeper, more systematic
regularities about how the environment evolves over
time. In other words, like any kind of cheating, the
cheater itself is cheated because of the reduced pres-
sure to learn, and learning is the real goal.
(cid:129) Predictions take time and space to generate: Nontrivial
predictions likely require the integration of multiple
converging inputs from a range of higher-level cortical
areas, each encoding different dimensions of relevance
(e.g., location, motion, color, texture, shape). Thus, suf-
ficient time and space (i.e., neural substrates with rele-
vant connectivity) must be available to integrate these
signals into a coherent predicted state, and per the
above point, these substrates must be separated from
the influence of current state information. This fits with
the properties of the layer 6CT neurons and their deep
layer inputs, which we hypothesize are insulated from
superficial-layer firing by virtue of being driven locally
by the 5IB within their own cortical microcolumn, such
that the interbursting pause period provides a time
window when these deep layers can integrate and gen-
erate the prediction.
(cid:129) Biologically, this is consistent with the delayed responses
of 6CT neurons (Harris & Shepherd, 2015; Thomson,
2010; Sakata & Harris, 2009). Computationally, these neu-
rons function much like the simple recurrent network
(SRN) context layer updating (Elman, 1990; Jordan,
1989), which reflects the prior trial’s state, as discussed
in detail in the Appendix. The overall duration of the
alpha cycle may represent a reasonable compromise
between the prediction integration time and the need
to keep up with predictions tracking changes in the world.
Notably, films are typically shown at only over two times
the alpha frequency (24 Hz), suggesting a Nyquist
sampling relative to the underlying alpha processing.
(cid:129) The predicted state must be directly aligned with the
outcome state it predicts: A prediction error is a differ-
ence between two states, so these prediction and
1162
Journal of Cognitive Neuroscience
Volume 33, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
1
5
8
1
9
1
3
5
7
7
/
/
j
o
c
n
_
a
_
0
1
7
0
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
outcome states must be directly comparable such that
their difference meaningfully represents the actual pre-
diction error and not some other kind of irrelevant
encoding differences. In other words, the prediction
and the outcome must be represented in the same
“language,” so that the “words” from the prediction can
be directly compared against those of the outcome—if
the prediction was in Japanese and the outcome was in
English, it would be hard to tell whether the prediction
was correct or not. Thus, a common neural substrate
with two different input pathways is required, one
reflecting the prediction and the other reflecting the
outcome, so that both converge onto the same repre-
sentational system within this common neural substrate.
This fits well with the two pathways converging into the
pulvinar: the 6CT top–down prediction-generation
pathway and the lower-level 5IB driving inputs.
(cid:129) The outcome signal should be as veridical as possible
(i.e., directly reflecting the bottom–up outcome) and
should arise from lower areas in the hierarchy relative
to the corresponding predictive 6CT inputs: Given that
the outcome is the driver of learning, if it were to be
corrupted or inaccurate, then everything that is
learned would then be suspect. To the extent that de-
lusional thinking is present in all people (some more so
than others perhaps), this principle must be violated at
some level, but for the lowest levels of the perceptual
system at least, it is important that strongly grounded,
accurate training signals drive learning. The bottom–
up, sparse, strongly driving nature of the 5IB projec-
tions to the pulvinar can directly convey such veridical
outcome signals and ensure that they dominate the ac-
tivation of their TRC targets. On the basis of indirect
available data, it is likely that each pulvinar TRC neuron
receives only roughly one to six driver inputs (Sherman
& Guillery, 2006, 2011), such that these sparse inputs
directly convey the signal from lower layers, without
much further mixing or integration (which could dis-
tort the nature of the signal). Furthermore, these in-
puts are likely not plastic (Usrey & Sherman, 2018),
again consistent with a need for unaltered, veridical
signals. Finally, the TRC neurons are distinctive in
having no significant lateral interconnectivity (Sherman
& Guillery, 2006), enabling them to faithfully represent
their inputs. These properties led Mumford (1991) to
characterize the pulvinar as a blackboard, and we
further suggest the metaphor of a projection screen
upon which the predictions are projected.
(cid:129) The prediction error must drive learning to reduce sub-
sequent prediction errors: Obviously, this is the goal of
prediction error learning in the first place, and given
that the cortex is what generates predictions, it must
be capable of learning based on prediction error signals
represented over the pulvinar. Computationally, the
critical problem here is “credit assignment”: How do
the error signals direct learning in the proper direction
for each individual neuron, to reduce the overall
prediction error? The error Bp procedure solves this
problem (Rumelhart, Hinton, & Williams, 1986) but re-
quires biologically implausible retrograde signaling
across the entire network of neural communication
(Crick, 1989), to propagate the error proportionally
back along the same channels that drive forward activa-
tion. Bidirectional connections, which are ubiquitous
in the cortex (Markov, Ercsey-Ravasz, et al., 2014;
Felleman & Van Essen, 1991) and computationally ben-
eficial for other reasons as noted earlier, can eliminate
that problem by “implicitly” propagating error signals
via standard neural communication mechanisms along
both directions of connectivity (O’Reilly, 1996).
(cid:129) This solution to the credit assignment problem relies on
a temporal difference error signal, as originally devel-
oped for the Boltzmann machine (Ackley et al., 1985).
The bidirectional neural communication at one point
in time is encoding and sharing the prediction among
the entire network of neurons. Then, this same network
of connections is reused at another point in time to en-
code and communicate the outcome. Mathematically,
the difference in activation state across these two points
in time, locally at each individual neuron, provides an
accurate estimate of the error Bp gradient (O’Reilly,
1996). In effect, this temporal difference tells each neu-
ron which direction it needs to change its activation
state to reduce the overall error. The reuse of the very
same network of connections across both points in time
ensures the overall alignment of the two activation
states, as noted above, such that this temporal differ-
ence precisely represents the error signal. Although
various other schemes for error-driven learning in bio-
logically plausible networks have been proposed (e.g.,
Lillicrap et al., 2020; Whittington & Bogacz, 2019;
Bengio et al., 2017), the temporal difference framework
with bidirectional connectivity provides a particularly
good fit with the natural temporal ordering of predictive
learning (prediction and then outcome) and the exten-
sive bidirectional connectivity of the thalamocortical
circuits (Shipp, 2003).
(cid:129) Temporal differences in activation state across the alpha
cycle, between prediction and outcome states, must
drive synaptic plasticity: The final step needed to con-
nect all of the elements above is that neurons actually
modify their synaptic strengths in proportion to the
temporal difference error signal. We have recently pro-
vided a fully explicit mechanism for this form of learning
(O’Reilly et al., 2012), based on a biologically detailed
model of spike-timing-dependent plasticity (Urakubo
et al., 2008). We showed that, when activated by realistic
Poisson spike trains, this spike-timing-dependent plas-
ticity model produces a nonmonotonic learning curve
similar to that of the Bienenstock, Cooper, and Munro
(BCM) model (Bienenstock, Cooper, & Munro, 1982),
which results from competing calcium-driven postsyn-
aptic plasticity pathways (Cooper & Bear, 2012; Shouval
et al., 2002). As in the BCM framework, we hypothesized
O’Reilly et al.
1163
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
1
5
8
1
9
1
3
5
7
7
/
/
j
o
c
n
_
a
_
0
1
7
0
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
that the threshold crossover point in this nonmono-
tonic curve moves dynamically—if this happens on the
alpha timescale (Lim et al., 2015), then it can reflect
the prediction phase of activity, producing a net error-
driven learning rule based on a subsequent calcium
signal reflecting the outcome state. The resulting
learning mechanism naturally supports a combination
of both BCM-style Hebbian learning and error-driven
learning, where the BCM component acts as a kind of
regularizer or bias, similar to weight decay (O’Reilly
et al., 2012; O’Reilly & Munakata, 2000).
Thus, remarkably, the pulvinar and associated thalamo-
cortical circuitry appear to provide precisely the necessary
ingredients to support predictive error-driven learning,
according to the above analysis. Interestingly, although
Sherman and Guillery (2006) did not propose a predictive
learning mechanism as just described, they did speculate
about a potential role for this circuit in motor forward-
model learning and the predictive remapping phenome-
non (Usrey & Sherman, 2018; Sherman & Guillery, 2011).
In addition, Pennartz, Dora, Muckli, and Lorteije (2019)
also suggested that the pulvinar may be involved in pre-
dictive learning, but within the EE coding framework and
not involving the detailed aspects of the above-described
circuitry.
It bears emphasizing the synergy between the various
considerations above for the benefits of the pause in 5IB
firing between bursts. First, this pause is critical for creat-
ing the time window when the predictive network is repre-
senting and communicating the prediction state, without
influence from the outcome state. Furthermore, it creates
the temporal difference in activation state in the pulvinar
between prediction and outcome, which is needed for
driving error-driven learning. Thus, for both the 6CT and
pulvinar layers, the periodic pausing of 5IB neurons is
essential for creating the predictive learning dynamic.
Interestingly, by these principles, the lack of such burst/pause
dynamics in the driver inputs to first-order sensory thalamus
areas such as the lateral geniculate nucleus and medial genic-
ulate nucleus (Sherman & Guillery, 2006) means that these
areas should not be directly capable of error-driven predic-
tive learning. This is consistent with a number of models
and theoretical proposals suggesting that primary sensory
areas may learn predominantly through Hebbian-style self-
organizing mechanisms (Bednar, 2012; Miller, 1994).
Nevertheless, primary sensory areas do receive “collateral”
error signals from the pulvinar (Shipp, 2003), which could
provide some useful indirect error-driven learning signals.
Note that this form of temporal difference learning signal
is distinct from the widely used temporal-difference model
in reinforcement learning (Sutton & Barto, 1998), which is
scalar and applies to reward expectations, not sensory pre-
dictions (although see Gardner, Schoenbaum, & Gershman,
2018, and Dayan, 1993, for potential connections between
these two forms of prediction error). Finally, as we discuss
later, this proposed predictive role for the pulvinar is
compatible with the more widely discussed role it may play
in attention (Fiebelkorn & Kastner, 2019; Zhou et al., 2016;
Saalmann & Kastner, 2011; Snow, Allen, Rafal, & Humphreys,
2009; Bender & Youakim, 2001; LaBerge & Buchsbaum,
1990). Indeed, we think these two functions are synergistic
(i.e., you predict what you attend, and vice versa; Richter
& de Lange, 2019) and have initial computational results
consistent with this idea.
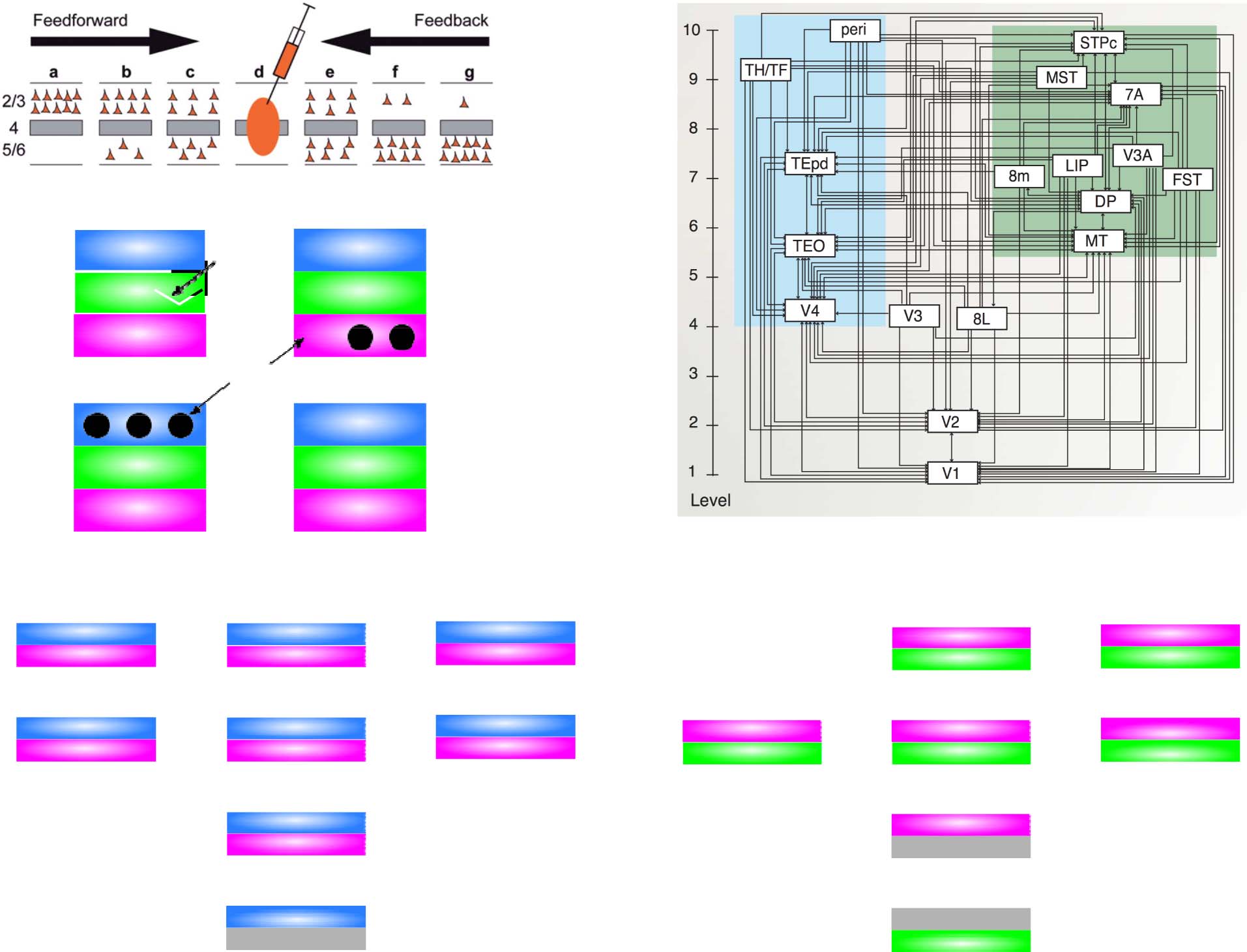
PREDICTIVE LEARNING OF TEMPORAL
STRUCTURE IN A PROBABILISTIC GRAMMAR
To illustrate and test the predictive learning abilities of this
biologically based model, we first ran a classical test of
sequence learning (Cleeremans & McClelland, 1991;
Reber, 1967) that has been explored using SRNs (Elman,
1990; Jordan, 1989). The biologically based model was
implemented using the Leabra algorithm, which is a com-
prehensive framework that uses conductance-based point
neuron equations, inhibitory competition, bidirectional
connectivity, and the biologically plausible temporal dif-
ference learning mechanism described above (O’Reilly
et al., 2012, 2016; O’Reilly & Munakata, 2000; O’Reilly,
1996, 1998). Leabra serves as a model of the bidirectionally
connected processing in the cortical superficial layers and
has been used to simulate a large number of different cog-
nitive neuroscience phenomena. It is described in the
Appendix, which also provides a detailed mapping be-
tween the SRN and our biological model.
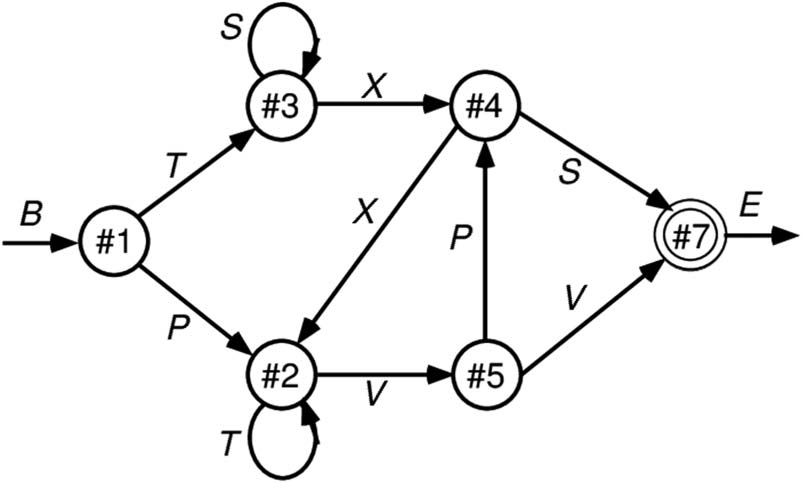
As shown in Figure 3, sequences were generated accord-
ing to a finite state automaton (FSA) grammar, as used in
implicit sequence learning experiments by Reber (1967).
Each node has a 50% random branching to two different
other nodes, and the labels generated by node transitions
are locally ambiguous (except for the B = begin and E =
end states). Thus, integration over time and across many
iterations is required to infer the systematic underlying
grammar. It is a reasonably challenging task for SRNs
and people to learn and provides an important validation
of the power of these predictive learning mechanisms.
Given the random branching, accurately predicting the
specific path taken is impossible, but we can score the
model’s output as correct if it activates either or both of
the possible branches for each state.
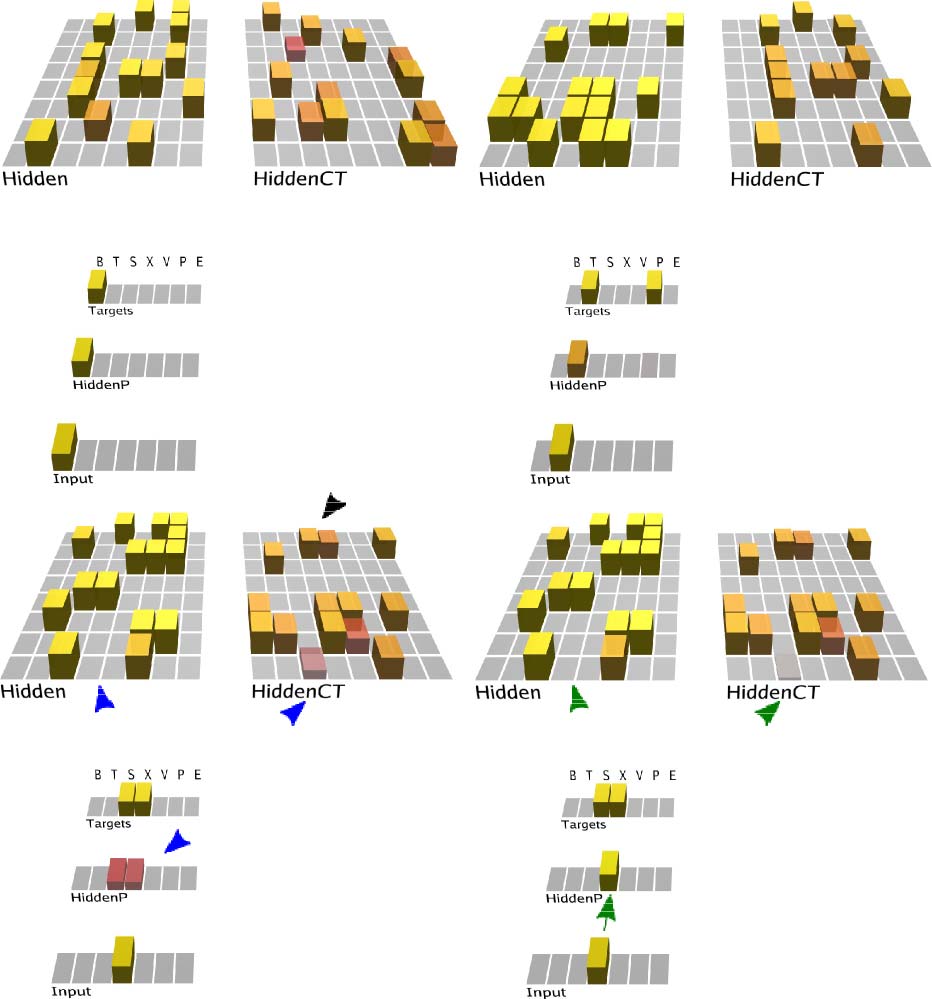
The model (Figure 4) required around 20 epochs of 25
sequences through the grammar to learn it to the point of
making no prediction errors for five epochs in a row
(which guarantees that it had completely learned the
task). This model is available in the standard emergent
distribution at github.com/emer/ leabra/tree/master
/examples/deep_fsa. A few steps through a sequence are
shown in Figure 4, illustrating how the corticothalamic
(CT) context layer, which drives the P pulvinar layer pre-
diction, represents the information present on the previ-
ous alpha cycle time step. Thus, the network is attempting
to predict the current input state, which then drives the
pulvinar plus phase at the end of each alpha cycle, as
1164
Journal of Cognitive Neuroscience
Volume 33, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
1
5
8
1
9
1
3
5
7
7
/
/
j
o
c
n
_
a
_
0
1
7
0
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
(Neupane, Guitton, & Pack, 2017; Cavanagh, Hunt, Afraz,
& Rolfs, 2010; Duhamel, Colby, & Goldberg, 1992). The only
learning signal available to the model was the prediction er-
ror generated by the temporal difference between what it
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
1
5
8
1
9
1
3
5
7
7
/
/
j
o
c
n
_
a
_
0
1
7
0
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. Predictive learning model applied to the FSA grammar shown in
the previous figure. The first three panels (A–C) show the prediction state
(end of the minus phase, e.g., the first 75 msec of an alpha cycle) of the
trained model on the first three steps of the sequence “BTX” (plus phases
also occurred but are not shown). The last panel (D) shows the plus phase
after the third step. The “Input” layer provides the 5IB drivers for the
corresponding HiddenP pulvinar layer, so the plus phase is always based
on the specific randomly selected path taken. The “Targets” layer is purely
for display, showing the two valid possible labels that could have been
predicted. To track learning, the model’s prediction is scored as accurate if
either or both targets are activated. Computationally, the model is similar
to an SRN, where the CT layer that drives the prediction over the pulvinar
encodes the activation state from the previous time step (alpha cycle),
because of the phasic bursting of the 5IB neurons that drive CT updating.
Note how the CT layer in B reflects the “Hidden” activation state in A and
likewise for C reflecting B. This is evident because we are using one-to-one
connectivity between Hidden and HiddenCT layers (which works well in
general, along with full lateral connectivity within the CT layer). Thus,
although the correct answer is always present on the Input layer for each
step, the CT layer is nevertheless attempting to predict this input based on
the information from the prior time step. (A) In the first step, the “B” label
is unambiguous and easily predicted (based on prior “E” context). (B) In
the second step, the network correctly guesses that the “T” label will come
next, but there is a faint activation of the other “P” alternative, which is
also activated sometimes based on prior learning history and associated
minor weight tweaks. (C) In the third step, both “S” and “X” are equally
predicted. (D) In the plus phase, only the Input pattern (“X” on this trial)
drives HiddenP activations, and the projections from the pulvinar back to
the cortex convey both the minus-phase prediction and the plus-phase
actual input. You can see one HiddenCT neuron, just above the arrow,
visibly changes its activation as a result (and all neurons experience
smaller changes), and learning in all these cortical (Hidden) layer neurons
is a function of their local temporal difference between the minus and
plus phases.
O’Reilly et al.
1165
Figure 3. FSA grammar used in implicit sequential learning experiments
(Reber, 1967) and in early SRNs (Cleeremans & McClelland, 1991).
It generates a sequence of letters according to the link transitioned
between state nodes, where each outgoing link to another node has
a 50% probability of being selected. Each letter (except for the B =
begin and E = end) appears at two different points in the grammar,
making them locally ambiguous. This combination of randomness and
ambiguity makes it challenging for a learning system to infer the true
underlying structure of the grammar.
shown in the last panel. On each trial, the difference be-
tween plus and minus phases locally over each cortical
neuron drives its synaptic weight changes, which accumu-
late over trials to allow accurate prediction of the sequences,
to the extent possible given their probabilistic nature.
PREDICTIVE LEARNING OF OBJECT
CATEGORIES IN IT CORTEX
Now, we describe a large-scale, systems-neuroscience im-
plementation of the proposed thalamocortical predictive
error-driven learning framework, in a model of visual pre-
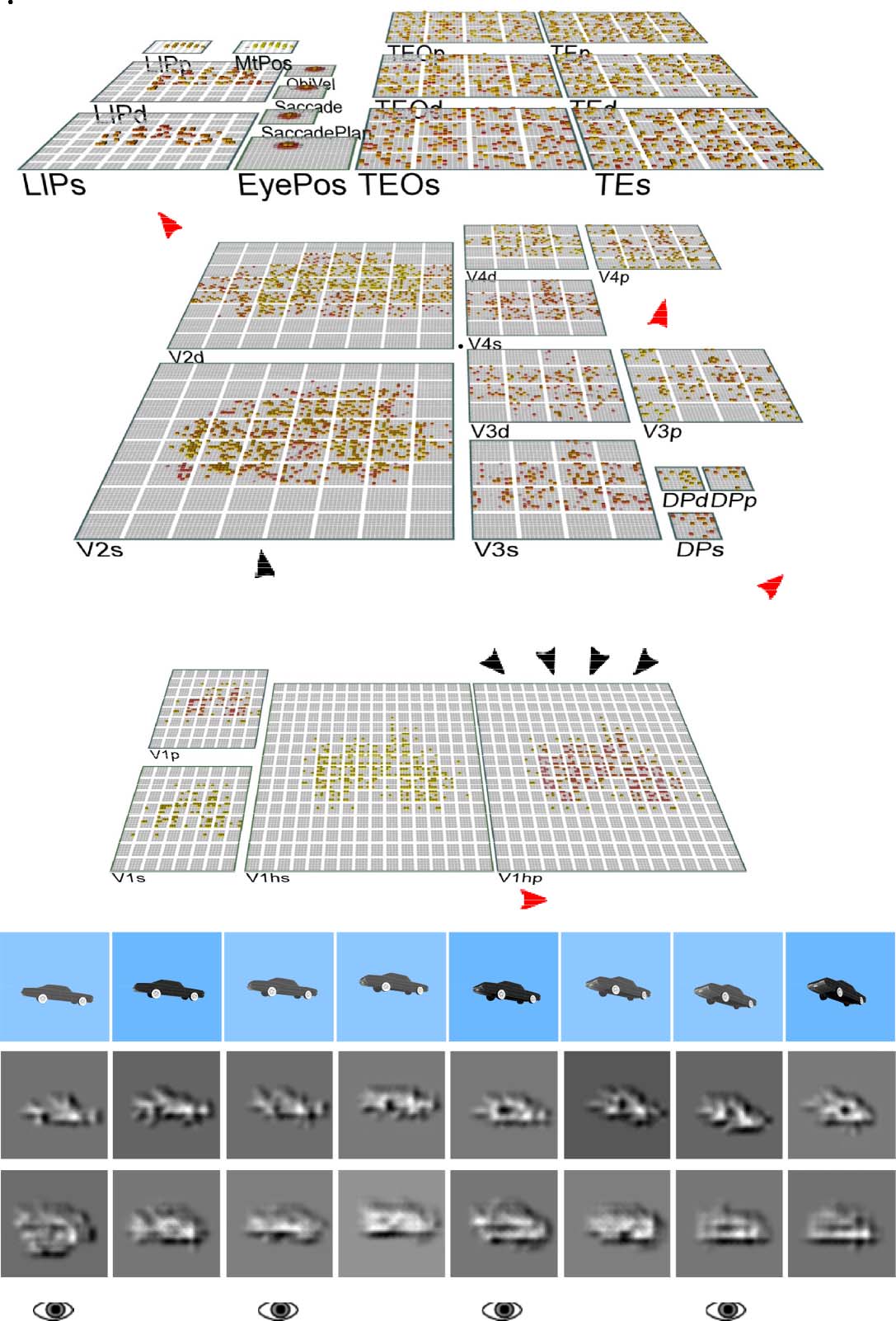
dictive learning (Figure 5). Our second major objective,
and a critical question for predictive learning, is determin-
ing whether the model can develop high-level, abstract
ways of representing the raw sensory inputs, while learning
from nothing but predicting these low-level visual inputs.
We showed the model brief movies of 156 3-D object exem-
plars drawn from 20 different basic-level categories (e.g.,
car, stapler, table lamp, traffic cone) selected for their over-
all shape diversity from the CU3D-100 data set (O’Reilly
et al., 2013). The objects moved and rotated in 3-D space
over eight movie frames, where each frame was sampled
at the alpha frequency (Figure 5B). Because the motion
and rotation parameters were generated at random on each
sequence, this data set consists of 512,000 unique images,
and there is no low-dimensional object category training
signal, so the usual concerns about overfitting and training
versus testing sets are not applicable: Our main question
is what kind of representations self-organize as a result of
this purely visual experience.
There were also saccadic eye movements every other
frame, introducing an additional, realistic, predictive learning
challenge. An efferent copy signal enabled full prediction of
the effects of the eye movement and allows the model to
capture the signature predictive remapping phenomenon
Figure 5. (A) The WWI deep
predictive learning model. The
dorsal “Where” pathway learns
first, using easily abstracted
spatial blobs, to predict object
location based on prior motion,
visual motion, and saccade
efferent copy signals. This
drives strong top–down inputs
to lower areas with accurate
spatial predictions, leaving the
residual error concentrated on
“What” and “What × Where”
integration. The V3 and DP
(dorsal prelunate) constitute
the What × Where integration
pathway, binding features and
locations. V4, TEO, and TE are
the What pathway, learning
abstracted object category
representations, which also
drive strong top–down inputs
to lower areas. Suffixes: s =
superficial; d = deep; and p =
pulvinar. (C) Example sequence
of eight alpha cycles that the
model learned to predict, with
the reconstruction of each
image based on the V1 Gabor
filters (“V1h recon”) and
model-generated prediction
(correlation r prediction error
shown). The low resolution
(Res) and reconstruction
distortion impair visual
assessment, but r values are well
above the rs for each V1 state
compared to the previous time
step (mean = .38, minimum of
.16 on Frame 4; see Appendix
for more analyses). Eye icons
indicate when a saccade
occurred.
predicted to see in the V1 input in the next frame and
what was actually seen.
As described in detail in the Appendix, our model was
constructed to capture critical features of the visual system,
including the major division between a dorsal “where”
pathway and a ventral “what” pathway (Ungerleider &
Mishkin, 1982), and the overall hierarchical organization
of these pathways derived from detailed connectivity anal-
yses (Markov, Ercsey-Ravasz, et al., 2014; Markov, Vezoli,
et al., 2014; Felleman & Van Essen, 1991; Rockland &
Pandya, 1979). In addition to these biological constraints,
we conducted extensive exploration of the connectivity
and architecture space and found a remarkable conver-
gence between what worked functionally and the known
properties of these pathways (O’Reilly et al., 2017). For ex-
ample, the feedforward pathway has projections from
lower-level superficial layers to superficial layers of higher
levels, whereas feedback originated in both the superficial
and deep layers and projected back to both (Felleman &
Van Essen, 1991; Rockland & Pandya, 1979). In addition,
consistent with the core features of the pulvinar pathways
discussed above, deep layer predictive (6CT) inputs origi-
nated in higher levels, whereas driver (5IB) inputs originated
in lower levels. For simplicity, we organized the model
layers in terms of these driver inputs, whereas the topo-
graphic organization of pulvinar in the brain is organized
more according to the 6CT projection loops (Shipp, 2003).
Another important set of parameters are the strength of
deep-layer recurrent projections, which influence the time-
scale of temporal integration, producing a simple biologically
based version of slow feature analysis (Wiskott & Sejnowski,
2002; Foldiak, 1991). We followed the biological data sug-
gesting that recurrence increases progressively up the visual
hierarchy (Chaudhuri, Knoblauch, Gariel, Kennedy, & Wang,
2015). It was essential that the “where” pathway learn first,
consistent with extant data (Kiorpes, Price, Hall-Haro, &
Movshon, 2012; Bourne & Rosa, 2006), including early path-
ways interconnecting lateral inferior parietal (LIP) and pulvi-
nar (Bridge, Leopold, & Bourne, 2016), and a rare asymmetric
pathway, from V1 to LIP (Markov, Ercsey-Ravasz, et al.,
1166
Journal of Cognitive Neuroscience
Volume 33, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
1
5
8
1
9
1
3
5
7
7
/
/
j
o
c
n
_
a
_
0
1
7
0
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
2014), providing a direct shortcut for high-level spatial
representations in LIP. Results from various informative
model architecture and parameter manipulations are dis-
cussed below after the primary results from the standard
intact model.
Learning curves and other model details are shown in the
Appendix. We have also implemented a full de-novo repli-
cation of the model in a new modeling framework, which
also replicated the results shown here. Furthermore, much
of the model was originally developed in the context of a set
of object-like patterns generated systematically from a set of
simple line features (O’Reilly et al., 2017), and the parame-
ters that work best in terms of combinatorial generalization
on those patterns also worked well for these 3-D objects.
Thus, we are confident that the model’s learning behavior
is not idiosyncratic to the particular set of objects used here
and represents a general capacity of the system to develop
abstract representations through predictive learning. Other
ongoing work to be reported in an upcoming publication is
applying the model to prediction of auditory speech inputs,
which has a natural temporal structure, and finding similar
results in terms of learning higher-level abstract encoding
of these auditory signals.
To directly address the question of whether the hierar-
chical structure of the network supports the development
of abstract, higher-level representations that go beyond
the information present in the visual inputs, we applied
a second-order similarity measure across the object-level
similarity matrices computed at each layer in the network
(Figure 6). This shows the extent to which the similarity
matrix across objects in one layer is itself similar to the ob-
ject similarity matrix in another layer, in terms of a corre-
lation measure across these similarity matrices. Critically,
this measure does not depend on any kind of subjective
Figure 6. Emergence of abstract category structure over the hierarchy
of layers, comparing similarity structure in each layer versus that
present in V1 (black line) or in TE (red line). Both cases, which are
roughly symmetric, clearly show that IT layers (TEO, TE) progressively
differentiate from raw input similarity structure present in V1 and,
critically, that the model has learned structure beyond that present
in the input. This is the simplest, most objective summary statistic
showing this progressive emergence of structure, whereas subsequent
figures provide a more concrete sense of what kinds of representations
actually developed. correl = correlation.
interpretation of the learned representations—it only tells
us whether whatever similarity structure was learned dif-
fers across the layers. Starting from either V1 compared
to all higher layers, or the highest TE layer compared to
all lower layers, we found a consistent pattern of progres-
sive emergence of the object categorization structure in
the upper IT pathway (TEO, TE).
This analysis confirms that indeed the IT category struc-
ture is significantly different from that present at the level of
the V1 primary visual input. Thus, the model, despite being
trained only to generate accurate visual input-level predic-
tions, has learned to represent these objects in an abstract
way that goes beyond the raw input-level information. We
further verified that, at the highest IT levels in the model, a
consistent, spatially invariant representation is present
across different views of the same object (e.g., the average
correlation across frames within an object was .901).
To better understand the nature of these learned repre-
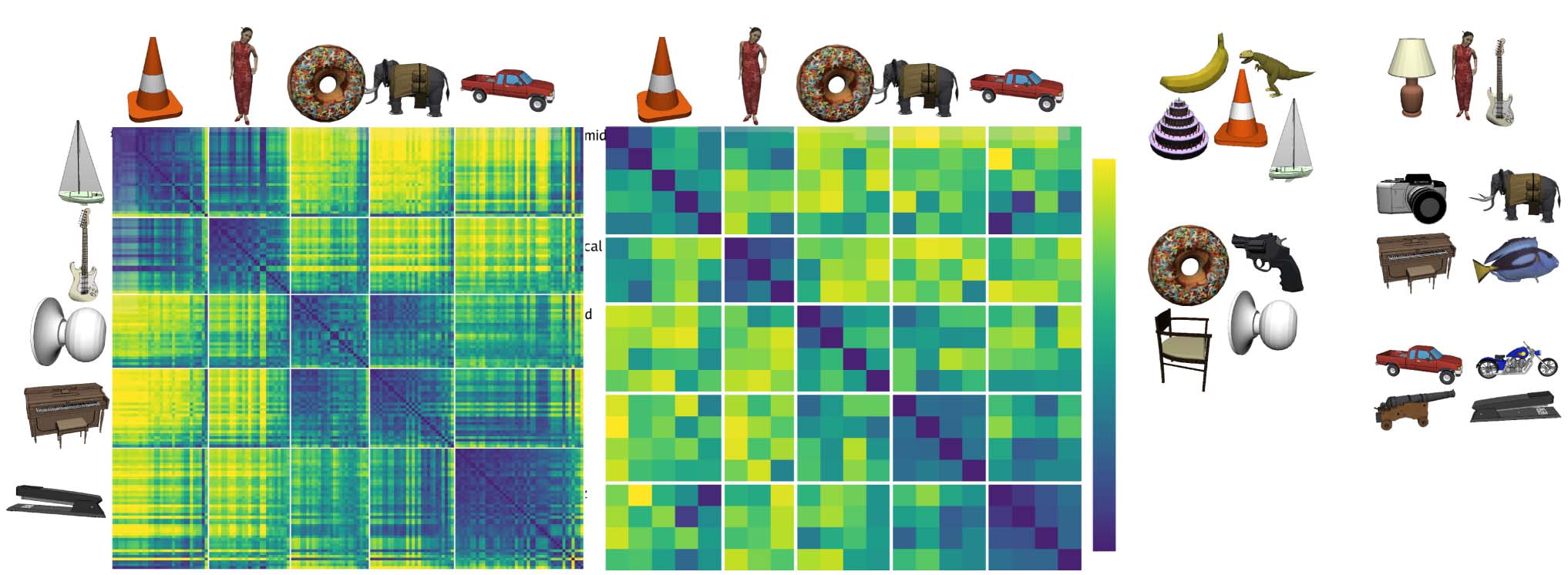
sentations, Figure 7 shows a representational similarity
analysis (RSA) on the activity patterns at the highest IT layer
(TE), which reveals the explicit categorical structure of the
learned representations (Cadieu et al., 2014; Kriegeskorte,
Mur, & Bandettini, 2008). Specifically, we found that the
highest IT layer (TE) produced a systematic organization
of the 156 3-D objects into five categories. In our admittedly
subjective judgment, these categories seemed to corre-
spond to the overall shape of the objects, as shown by the
object exemplars in Figure 7 (pyramid shaped, vertically
elongated, round, boxy/square, and horizontally elongated).
Furthermore, the basic-level categories were subsumed
within these broader shape-level categories, so the model
appears to be sensitive to the coherence of these basic-level
categories as well, but apparently, their shapes were not suf-
ficiently distinct between categories to drive differentiated
TE-level representations for each such basic-level category.
Given that the model only learns from a passive visual ex-
perience of the objects, it has no access to any of the richer
interactive multimodal information that people and animals
would have. Furthermore, as evident in Figure 5B, the
relatively low resolution of the V1 layers (required to make
the model tractable computationally) means that complex
visual details are not reliably encoded (and, even so, are not
generally reliable across object exemplars), such that the
overall object shape is the most salient and sensible basis
for categorization for this model.
Although these object shape categories appeared sensi-
ble to us, we ran a simple experiment to test whether a
sample of 30 human participants would use the same
category structure in evaluating the pairwise similarity of
these objects. Figure 7B shows the results, confirming that
indeed this same organization of the objects emerged in
their similarity judgments. These judgments were based
on the V1 reconstruction as shown in Figure 5B to capture
the model’s coarse-grained perception (see Appendix for
methods and further analysis).
The progressive emergence of increasingly abstract cat-
egory structure across visual areas, evident in Figure 6, has
O’Reilly et al.
1167
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
1
5
8
1
9
1
3
5
7
7
/
/
j
o
c
n
_
a
_
0
1
7
0
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
1
5
8
1
9
1
3
5
7
7
/
/
j
o
c
n
_
a
_
0
1
7
0
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
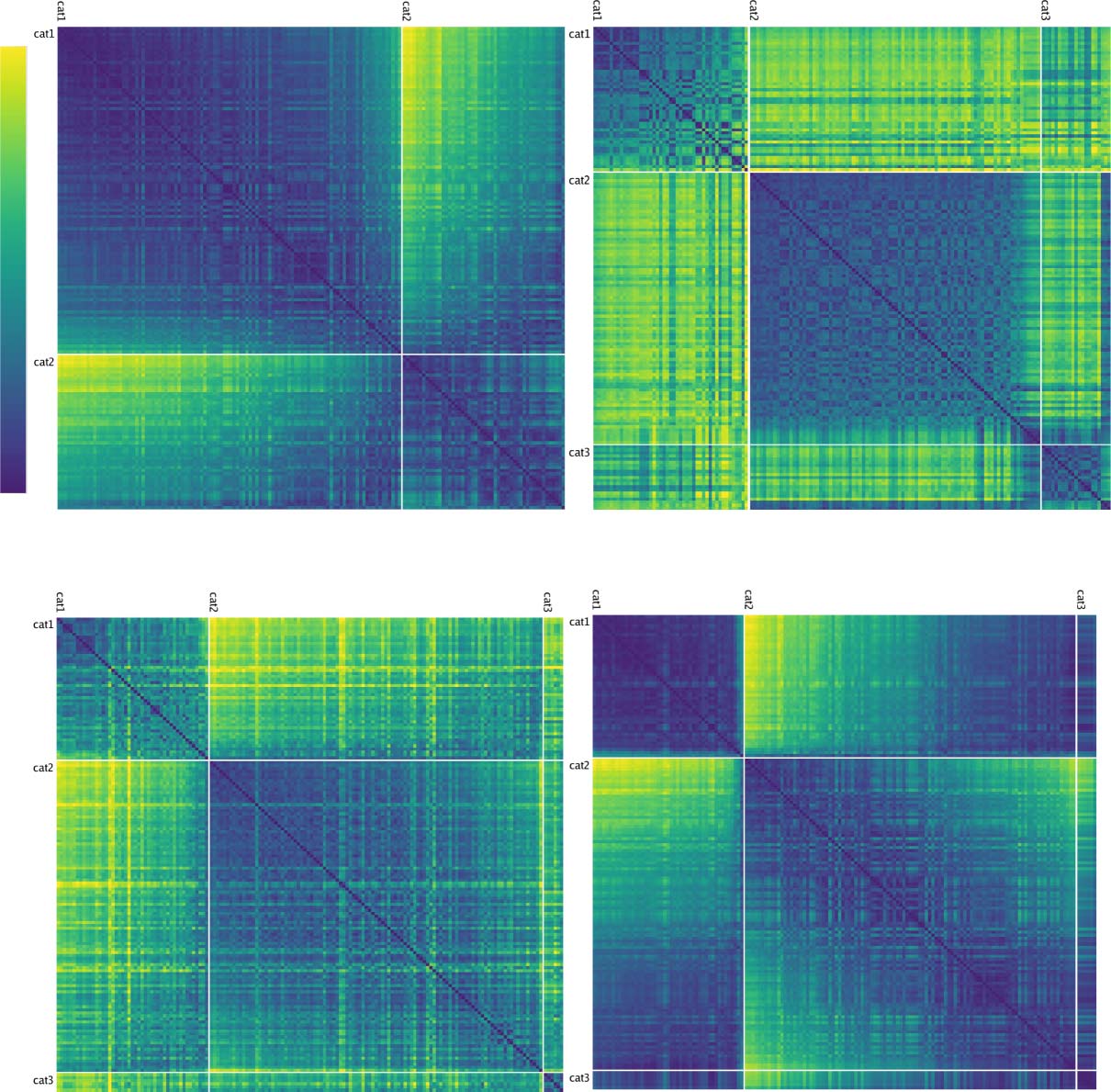
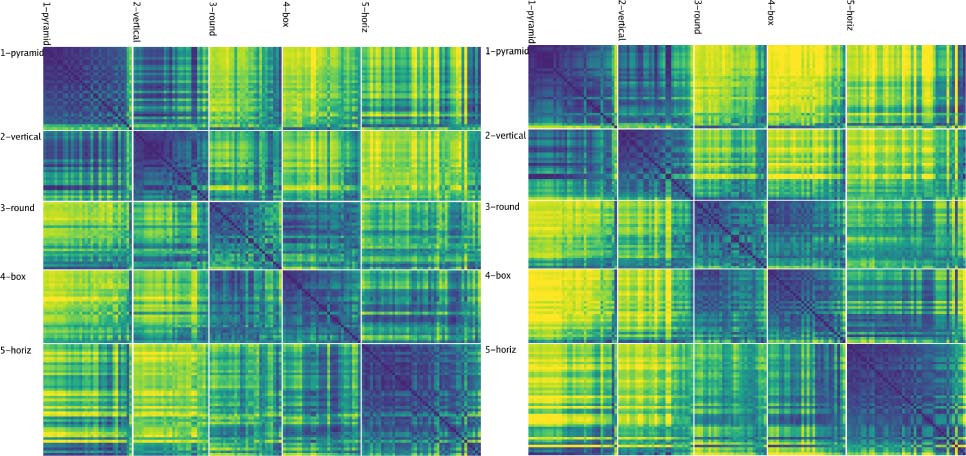
Figure 7. (A) Category similarity structure that emerged in the highest layer, TE, of the biologically based predictive learning model, showing
dissimilarity (1-correlation) of the TE representation for each 3-D object against every other 3-D object (156 total objects). Blue cells have high
similarity. Model has learned block-diagonal clusters or categories of high-similarity groupings, contrasted against dissimilar off-diagonal other
categories. Clustering maximized average within-between dissimilarity (see Appendix) and clearly corresponded to the shape-based categories,
with exemplars from each category shown. In addition, all items from the same basic-level object categories (n = 20) are reliably subsumed within
learned categories. (B) Human similarity ratings for the same 3-D objects, presented with the V1 reconstruction (see Figure 1B) to capture coarse
perception in the model, aggregated by 20 basic-level categories (a 156 × 156 matrix was too large to sample densely experimentally). Each cell is
1 − proportion of time given that object pair was rated more similar than another pair (see Appendix). The human matrix shares the same centroid
categorical structure as the model (confirmed by permutation testing and agglomerative cluster analysis; see Appendix), indicating that human
raters used the same shape-based category structure. (C) One object from each of the 20 basic-level categories, organized into the shape-based
categories. The Vertical, Box, and Horizontal categories are fairly self-evident, and the model was most consistent in distinguishing those, along with
subsets of the Pyramid (layered cake, traffic cone, sailboat) and Round (donut, doorknob) categories, whereas banana, trex, chair, and handgun were
more variable.
been investigated in recent comparisons between monkey
electrophysiological recordings and DCNNs, which provide
a reasonably good fit of the overall progressive pattern of
increasingly categorical organization (Cadieu et al., 2014).
However, these DCNNs were trained on large data sets of
human-labeled object categories, and it is perhaps not too
surprising that the higher layers closer to these category
output labels exhibited a greater degree of categorical
organization. In contrast, because the only source of learn-
ing in our model comes from prediction errors over the V1
input layers, the graded emergence of an object hierarchy
here reflects a truly self-organizing learning process.
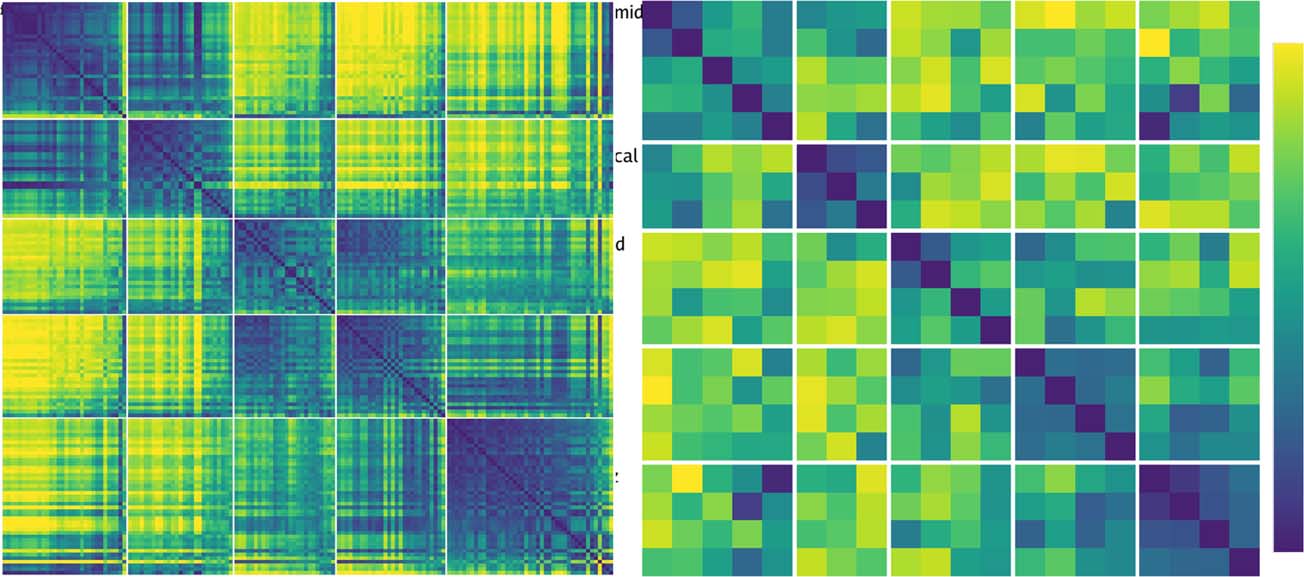
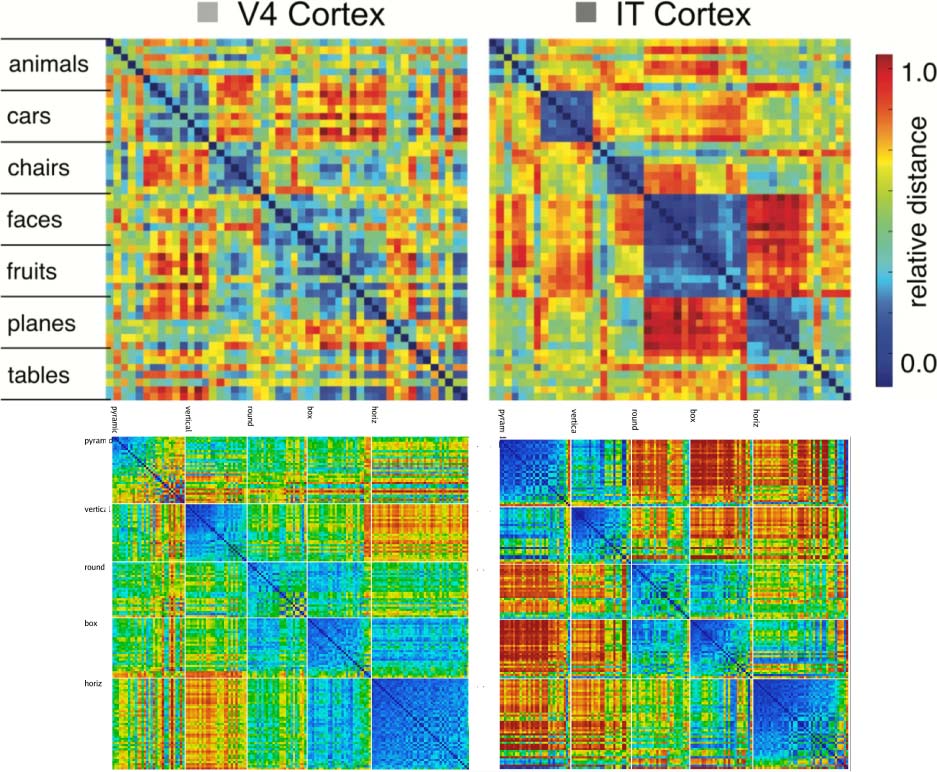
Figure 8 compares the similarity structures in Layers V4
and IT in macaque monkeys (Cadieu et al., 2014) with
those in corresponding layers in our model. In both the
monkeys and our model, the higher IT layer builds upon
and clarifies the noisier structure that is emerging in the
earlier V4 layer, showing that our model replicates the
essential qualitative hierarchical progression in the brain.
As noted, we would not expect our model to exactly repli-
cate the detailed object-specific similarity structure found
in macaques, because of the impoverished nature of our
model’s experience, so this comparison remains qualita-
tive in terms of the respective differences between V4
and IT in each model, rather than a direct comparison of
the similarity structure between corresponding layers in
the model and the macaque. In the future, when we can
scale up our model and tune the attentional processing
dynamics necessary to deal with cluttered visual scenes,
we will be able to train our model on the same images pre-
sented to the macaques and can provide this more direct
comparison.
Finally, we did not use analyses based on decoding tech-
niques, because with high-dimensional distributed neural
representations, it is generally possible to decode many
different features that are not otherwise compactly and
directly represented (Fusi, Miller, & Rigotti, 2016). In pre-
liminary work using decoding in the context of the simpler
feature-based input patterns, we indeed found that de-
coding was not a very sensitive measure of the differen-
tiation of representations across layers, which is so clearly
evident in Figure 6. Thus, as advocates of the RSA approach
have argued, measuring similarity structure evident in the
activity patterns over a given layer generally provides a
clearer picture of what that layer is explicitly encoding
(Kriegeskorte et al., 2008).
In summary, the model learned an abstract category
organization that reflects the overall visual shapes of the
objects as judged by human participants, in a way that is
invariant to the differences in motion, rotation, and scaling
that are present in the V1 visual inputs. We are not aware of
any other model that has accomplished this signature
computation of the ventral “what” pathway in a purely
self-organizing manner operating on realistic 3-D visual
1168
Journal of Cognitive Neuroscience
Volume 33, Number 6
Backpropagation Comparison Models
To help discern some of the factors that contribute to the
categorical learning in our model and provide a comparison
with more widely used error Bp models, we tested a
Bp-based version of the same “what vs. where” architecture
as our biologically based predictive error model, and we
also tested a standard PredNet model (Lotter et al., 2016) with
extensive hyperparameter optimization (see Appendix).
Because of the constraints of Bp, we had to eliminate
any bidirectional connectivity loops in the Bp version,
but we were able to retain a form of predictive learning
by configuring the V1p pulvinar layer as the final target
output layer, with the target being the next visual input
relative to the current V1 inputs.
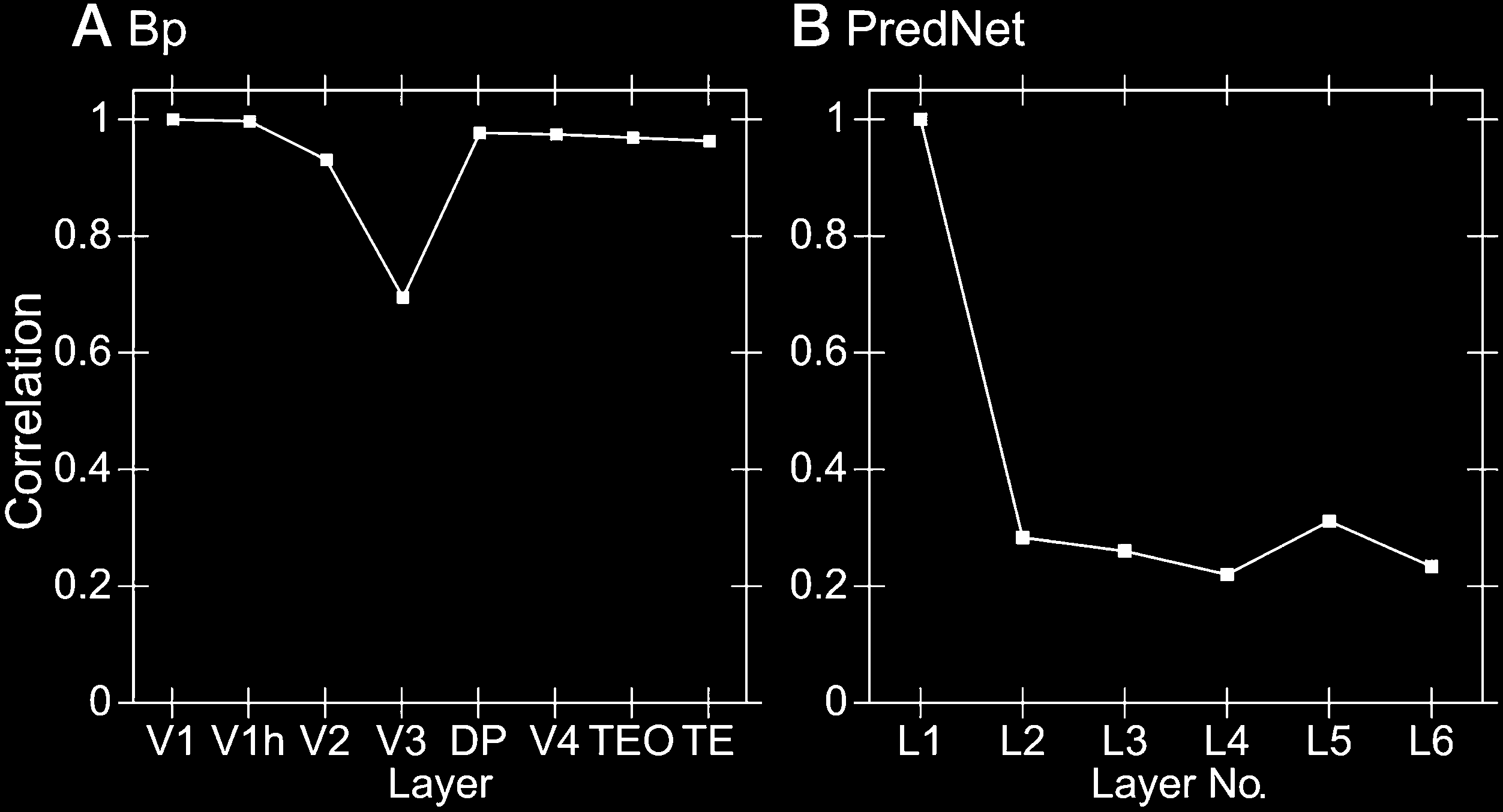
Figure 9 shows the same second-order similarity analy-
sis as Figure 6, to determine the extent to which these
comparison networks also developed more abstract repre-
sentations in the higher layers that diverge from the simi-
larity structure present in the lowest layers. According to
this simple objective analysis, they did not—the higher
layers showed no significant, progressive divergence in
their similarity structure. The PredNet model did show a
larger difference between the first layer and the rest of
the layers, because of the subsequent layers encoding er-
rors while the first layer has a positive representation of
the image, but there was no progressive difference beyond
that up into the higher layers.
Next, we examined the RSA matrices for the highest
(TE) layer in the comparison models, also in comparison
with the same for the V1 layer (Figure 10). This shows that
the TE layer in the Bp model formed a simple binary cate-
gory structure overall, which is similar to the RSA for the V1
input layer. It is also important to emphasize that the
scales on these figures are different (as shown in their
headers), such that these comparison models had much
Figure 8. Comparison of progression from V4 to IT in macaque monkey
visual cortex (top row, from Cadieu et al., 2014) versus same progression
in model (replotted using a comparable color scale). Although the
underlying categories are different, and the monkeys have a much richer
multimodal experience of the world to reinforce categories such as foods
and faces, the model nevertheless shows a similar qualitative progression
of stronger categorical structure in IT, where the block-diagonal highly
similar representations are more consistent across categories and the
off-diagonal differences are stronger and more consistent as well (i.e.,
categories are also more clearly differentiated). Note that the critical
difference in our model versus those compared in Cadieu et al. (2014)
and related papers is that they explicitly trained their models on category
labels, whereas our model is entirely self-organizing and has no external
categorical training signal. horiz = horizontal.
objects, without any explicit supervised category labels.
Furthermore, our model does this using a learning algo-
rithm directly based on detailed properties of the under-
lying biological circuits in this pathway, providing a
coherent overall account.
Figure 9. Similarity of similarity
structure across layers for the
comparison Bp models,
comparing each layer to the first
layer. (A) Bp model with the
same “what/where” structure as
the biological model. Unlike
the biologically based model
(Figure 6), the higher IT layers
(TE, TEO) do not diverge
significantly from the similarity
structure present in V1,
indicating that the model has
not developed abstractions
beyond the structure present in
the visual input. Layer V3 is most
directly influenced by spatial
prediction errors, so it differs
from both in strongly encoding
position information. (B) PredNet model, which has six layers. Layers 2–6 diverge from Layer 1, but there is no progressive change in the higher layers
as we see in our model moving from V4 to TEO. The divergence in correlation starting at Layer 2 is likely because of the fact that higher layers only
encode errors, not stimulus-driven positive representations of the input. Aside from this large distinction (which is inconsistent with the similarity in
neural coding seen in actual V1 and V2 recordings), there is no evidence of a cumulative development of abstraction in higher layers.
O’Reilly et al.
1169
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
1
5
8
1
9
1
3
5
7
7
/
/
j
o
c
n
_
a
_
0
1
7
0
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 10. (A) Best-fitting
category similarity for TE layer
of the Bp model with the same
“what/where” structure as the
biological model. Only two broad
categories are evident, and the
lower maximum (max) distance
(0.3 vs. 1.5 in the biological
model) means that the patterns
are much less differentiated
overall. (B) Best-fitting similarity
structure for the PredNet model,
in the highest of its layers (Layer
6), which is more differentiated
than Bp (max = 0.75) but also
less cleanly similar within
categories (i.e., less solidly blue
along the block diagonal) and
overall follows a broad category
structure similar to V1. (C) The
best-fitting V1 structure, which
has two broad categories and
banana, is in a third category by
itself. The lack of dark blue on the
block diagonal indicates that
these categories are relatively
weak, and every item is fairly
dissimilar from every other. (D)
The Bp TE similarity values from
A shown in the same ordering
as V1 from C, demonstrating
how the similarity structure
has not diverged very much,
consistent with the results shown
in Figure 9—the within-between
contrast differences are 0.0838
for A and 0.0513 for D (see
Appendix for details).
less differentiated representations overall. Similar results
were found in the PredNet model. Because existing work
with these models has typically relied on additional super-
vised learning and decoder-based analyses (which are es-
sentially equivalent to an additional layer of supervised
learning), these RSA-based analyses provide an important,
more sensitive way of determining what they learn purely
through predictive learning.
These results show that the additional biologically de-
rived properties in our model are playing a critical role in
the development of abstract categorical representations
that go beyond the raw visual inputs. These properties in-
clude excitatory bidirectional connections, inhibitory com-
petition, and an additional Hebbian form of learning that
serves as a regularizer (similar to weight decay) on top of
predictive error-driven learning (O’Reilly & Munakata,
2000; O’Reilly, 1998). Each of these properties could pro-
mote the formation of categorical representations.
Bidirectional connections enable top–down signals to con-
sistently shape lower-level representations, creating signifi-
cant attractor dynamics that cause the entire network to
settle into discrete categorical attractor states. Another indi-
cation of the importance of bidirectional connections is that
a greedy layer-wise pretraining scheme, consistent with a
putative developmental cascade of learning from the sensory
periphery on up ( Valpola, 2015; Bengio, Yao, Alain, &
Vincent, 2013; Hinton & Salakhutdinov, 2006; Shrager &
Johnson, 1996), did not work in our model. Instead, we
found it essential that higher layers, with their ability to form
more abstract, invariant representations, interact and
shape learning in lower layers right from the beginning.
Furthermore, the recurrent connections within the TEO
and TE layers likely play an important role by biasing the
temporal dynamics toward longer persistence (Chaudhuri
et al., 2015). By contrast, Bp networks typically lack these
kinds of attractor dynamics, and this could contribute signif-
icantly to their relative lack of categorical learning. Hebbian
learning drives the formation of representations that encode
the principal components of activity correlations over time,
which can help more categorical representations coalesce
(and results below already indicate its importance).
Inhibition, especially in combination with Hebbian learning,
drives representations to specialize on more specific sub-
sets of the space.
Ongoing work is attempting to determine which of these
is essential in this case (perhaps all of them) by systematically
introducing some of these properties into the Bp model,
although this is difficult because full bidirectional recurrent
1170
Journal of Cognitive Neuroscience
Volume 33, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
1
5
8
1
9
1
3
5
7
7
/
/
j
o
c
n
_
a
_
0
1
7
0
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
activity propagation, which is essential for conveying error
signals top–down in the biological network, is incompatible
with the standard efficient form of error Bp, and requires
significantly more computationally intensive and unstable
forms of fully recurrent Bp ( Williams & Zipser, 1992;
Pineda, 1987). Furthermore, Hebbian learning requires
dynamic inhibitory competition, which is difficult to incorpo-
rate within the Bp framework.
Architecture and Parameter Manipulations
Figure 11 shows only a few of the large number of parameter
manipulations that have been conducted to develop and test
the final architecture. For example, we hypothesized that
separating the overall prediction problem between a spatial
“where” versus nonspatial “what” pathway (Goodale &
Milner, 1992; Ungerleider & Mishkin, 1982) would strongly
benefit the formation of more abstract, categorical object
representations in the “what” pathway. Specifically, the
“where” pathway can learn relatively quickly to predict
the overall spatial trajectory of the object (and anticipate
the effects of saccades) and thus effectively regress out that
component of the overall prediction error, leaving the
residual error concentrated in object feature information,
which can train the ventral “what” pathway to develop
abstract visual categories.
Figure 11A shows that, indeed, when the “where” path-
way is lesioned, the formation of abstract categorical rep-
resentations in the intact “what” pathway is significantly
impaired. We also hypothesized that full predictive learning
(about the future), as compared to just encoding and
decoding the current state (i.e., an autoencoder, which is
much easier computationally), is also critical for the for-
mation of abstract categorical representations—prediction
is a “desirable difficulty” (Bjork, 1994). Figure 11B
shows that this was the case. Finally, consistent with our
hypothesis that Hebbian learning provides an important
bias on learning, Figure 11C shows the impairment asso-
ciated with reducing this learning bias. The significant
reduction in differentiation across all of these manipula-
tions shows that this differentiation property is not a
simple consequence of the neural architecture but rather
depends critically on the learning process, unfolding over
time with appropriate parameter values and other archi-
tectural components. Furthermore, the Bp comparison
model shares the same architecture and does not show
the differentiation across layers.
Predictive Behavior
A signature example of predictive behavior at the neural
level in the brain is the predictive remapping of visual space
in anticipation of a saccadic eye movements (Marino &
Mazer, 2016; Nakamura & Colby, 2002; Gottlieb, Kusunoki,
& Goldberg, 1998; Colby, Duhamel, & Goldberg, 1997;
Duhamel et al., 1992; Figure 12A). Here, parietal neurons
start to fire at the future receptive field location where a
currently visible stimulus will appear after a planned
saccade is actually executed. Remapping has also been
shown for border ownership neurons in V2 (O’Herron &
von der Heydt, 2013) and in Area V4 (Neupane, Guitton,
and Pack, 2016, 2020). These are examples, we believe, of
a predictive process operating throughout the neocortex
to predict what will be experienced next. A major conse-
quence of this predictive process is the perception of a
stable, coherent visual world despite constant saccades
and other sources of visual change.
Figure 12B shows that our model exhibits this predictive
remapping phenomenon. Specifically, LIP, which is most
directly interconnected with the saccade efferent copy sig-
nals, is the first to predict the new location, and it then
drives top–down activation of lower layers. This top–down
Figure 11. Effects of various
manipulations on the extent
to which TE representations
differentiate from V1. For all plots,
“Intact” is the same result shown
in Figure 6 from the intact model
for ease of comparison (A is
missing V3 and DP dorsal pathway
layers). All of the following
manipulations significantly impair
the development of abstract TE
categorical representations (i.e.,
TE is more similar to V1 and the
other layers). (A) Dorsal “where”
pathway lesions, including lateral
inferior parietal sulcus (LIP), V3,
and dorsal prelunate (DP). This
pathway is essential for regressing out location-based prediction errors, so that the residual errors concentrate feature-encoding errors that train the
“what” pathway. (B) Allowing the deep layers full access to current-time information, thus effectively eliminating the prediction demand and turning the
network into an autoencoder, which significantly impairs representation development and supports the importance of the challenge of predictive
learning for developing deeper, more abstract representations. (C) Reducing the strength of Hebbian learning by 20% (from 2.5 to 2), demonstrating
the essential role played by this form of learning on shaping categorical representations. Eliminating Hebbian learning entirely (not shown) prevented
the model from learning anything at all, as it also plays a critical regularization and shaping role on learning. Hebb = Hebbian.
O’Reilly et al.
1171
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
1
5
8
1
9
1
3
5
7
7
/
/
j
o
c
n
_
a
_
0
1
7
0
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 12. Predictive
remapping. (Top) Original
remapping data in LIP from
Duhamel et al. (1992). A shows
stimulus (star) response within
the receptive field (dashed
circle) relative to the fixation
dot (top right of fixation). (B)
Just before monkey making a
saccade to new fixation (moving
left), stimulus is turned on in
receptive field location that
will be upper right of the new
fixation point, and the LIP
neuron responds to that
stimulus in advance of the
saccade completing. The
neuron does not respond to the
stimulus in that location if it is
not about to make a saccade
that puts it within its receptive
field (not shown). This is
predictive remapping. (C)
Response to the old stimulus
location goes away as saccade is
initiated. (Bottom) Data from
our model, from individual units
in LIPd, V2d, and V2s, showing
that the LIP deep neurons
respond to the saccade first,
activating in the new location
and deactivating in the old,
and this LIP activation goes
top–down to V3 and V2 to drive
updating there, generally at
a longer latency and with less
activation especially in the
superficial layers. When the new stimulus appears at the point of fixation (after a 50-msec saccade here), the primed V2s units get fully activated by
the incoming stimulus. However, the deep neurons are insulated from this superficial input until the plus phase, when the cascade of 5IB firing drives
activation of the actual stimulus location into the pulvinar, which then reflects up into all the other layers.
dynamic is consistent with the account of predictive
remapping given by Wurtz (2008) and Cavanagh et al.
(2010), who argue that the key remapping takes place at
the high levels of the dorsal stream, which then drive top–
down activation of the predicted location in lower areas,
instead of the alternative where lower levels remap them-
selves based on saccade-related signals. The lower-level
visual layers are simply too large and distributed to be able
to remap across the relevant degrees of visual angle—the
extensive lateral connectivity needed to communicate
across these areas would be prohibitive.
NEURAL DATA AND PREDICTIONS
Having tested the computational and functional learning
properties of this biologically based predictive learning
mechanism, we now return to consider some of the most
important neural data of relevance to our hypotheses,
beyond that summarized in the introduction, including
contrasts with a widely discussed alternative framework
for predictive coding, and some of the extensive data on
alpha frequency effects, followed by a discussion of predic-
tions that would clearly test the validity of this framework.
Additional Neuroscience Data
We begin with data relevant to the basic neural-level prop-
erties of the framework. First, a central element of the pro-
posed model is the alpha cycle bursting, and subsequent
interburst pauses, in the 5IB neurons. Direct electrophysi-
ological recording of deep layer neurons shows periodic
alpha-scale bursting for continuous tones in awake animals
(Luczak et al., 2009, 2013; Sakata & Harris, 2009, 2012). In
vitro, a variety of potential mechanisms behind the genera-
tion and synchronization of the 5IB bursts driving this alpha
cycle have been identified (Franceschetti et al., 1995; Silva
et al., 1991; Connors et al., 1982). Furthermore, the pulvinar
has been shown to drive alpha-frequency synchronization
of cortical activity across areas in the alpha band in awake,
behaving animals (Saalmann et al., 2012). We review the
larger alpha frequency literature in more detail below, but
it is critical to emphasize that this alpha bursting dynamic is
1172
Journal of Cognitive Neuroscience
Volume 33, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
1
5
8
1
9
1
3
5
7
7
/
/
j
o
c
n
_
a
_
0
1
7
0
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
actually found in awake, behaving animals, because so
many other bursting and up/down state phenomena have
recently been shown to only occur in anesthetized brains,
including bursting in the thalamic TRC neurons.
In contrast to the 5IB bursting, the 6CT neurons exhibit
regular spiking behavior (Thomson, 2010; Thomson &
Lamy, 2007), providing consistent activation to the pulvi-
nar. In addition, they do not have axonal branches that
project to other cortical areas—the subpopulation that
projects to the pulvinar only project there and not to other
cortical areas (Petrof, Viaene, & Sherman, 2012), whereas
there are other Layer 6 neurons that do project to other
cortical areas. This distinct connectivity is consistent with
a specific role of this neuron type in generating predictions
in the pulvinar. The 6CT synaptic inputs on pulvinar TRCs
have metabotropic glutamate receptors that have longer
timescale temporal dynamics consistent with the alpha
period (100 msec) and even longer (Sherman, 2014), and
the 6CT neurons themselves also have temporally delayed
responding (Harris & Shepherd, 2015; Thomson, 2010;
Sakata & Harris, 2009). Furthermore, they have significantly
more plasticity-inducing N-methyl-D-aspartate receptors
compared to the 5IB projections (Usrey & Sherman,
2018). These properties are consistent with the 6CT in-
puts driving a longer-integrated prediction signal that is
subject to learning, whereas the 5IB are likely nonplastic,
and their effects are tightly localized in time.
The 5IB inputs often have distinctive glomeruli structures
at their synapses onto pulvinar neurons, which contain a
complete feedforward inhibition circuit involving a local
inhibitory interneuron, in addition to the direct strong ex-
citatory driver input (Wilson, Bose, Sherman, & Guillery,
1984). Computationally, this can provide a balanced level
of excitatory and inhibitory drive so as to not overly ex-
cite the receiving neuron, while still dominating its firing
behavior.
Although there are well-documented and widely dis-
cussed burst versus tonic firing modes in pulvinar neurons
(Sherman & Guillery, 2006), there is not much evidence of
these playing a clear role in the awake, behaving state, and
as noted earlier, the growing electrophysiological evi-
dence shows a remarkable correspondence between cor-
tical and pulvinar response properties across multiple
different pulvinar areas in this awake state. Nevertheless,
there may be important dynamics arising from these firing
modes that are more subtle or emerge in particular types
of state transitions that may have yet to be identified.
Contrast with Explicit Error Frameworks
To further clarify the nature of the present theory and intro-
duce a body of relevant data, we contrast it with the widely
discussed EE framework for predictive coding (Lotter et al.,
2016; Bastos et al., 2012; Ouden et al., 2012; Friston, 2005,
2010; Rao & Ballard, 1999; Kawato et al., 1993; Figure 13).
The hypothesized locus for computing errors in this frame-
work is in the superficial layers of the neocortex, which
are suggested to directly compute the difference between
Figure 13. Comparison
between (A) the proposed
thalamocortical temporal-
difference predictive learning
model (from Figure 2) versus
(B) the Bayesian-style EE coding
model (Bastos et al., 2012;
Friston, 2010; Rao & Ballard,
1999). The EE model holds
that superficial (S, lamina 2/3)
error-coding neurons receive
the prediction via a net inhibitory
top–down projection from
higher-level deep layer
(D) neurons and an excitatory
bottom–up projection
representing the outcome, such
that their activation represents
the difference. To encode both
signs of the error (omissions, false
alarms) with positive-only spike
rates, two separate populations
of EE neurons would be required,
or a more complicated deviation
from tonic firing level scheme. Unambiguous evidence of such EE coding neurons has not been found ( Walsh et al., 2020). In contrast, error signals in our
proposed framework remain as a temporal difference between the two states of prediction versus outcome, which enables all connectivity between cortical
areas to be excitatory and always represent a positive encoding of either the prediction or outcome. In contrast, under EE, after one error subtraction at the
lowest level, only error signals are hypothesized to flow forward to higher layers, meaning that the representations at higher layers are about increasingly
higher-order errors, not positive encodings of the environmental state at increasing levels of abstraction. These are indicated by ? because they are difficult
to picture intuitively, and they are inconsistent with extensive available data showing similar positive representations of the external world at all levels in
the visual hierarchy. Although some frameworks make claims about temporal dynamics, these are not strongly constrained by the basic computational
framework, so that also remains a question.
O’Reilly et al.
1173
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
1
5
8
1
9
1
3
5
7
7
/
/
j
o
c
n
_
a
_
0
1
7
0
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
bottom–up inputs from lower layers and top–down inputs
from higher areas. Despite many attempts to identify such
EE coding neurons in the cortex, no substantial body of
unambiguous evidence has been discovered ( Walsh,
McGovern, Clark, & O’Connell, 2020; Kok & de Lange,
2015; Kok, Jehee, & de Lange, 2012; Summerfield &
Egner, 2009; Lee & Mumford, 2003). Furthermore, because
of the positive-only firing rate nature of neural coding, two
separate populations would be required to convey both
signs of prediction error signals, or it would have to be
encoded as a variation from tonic firing levels, which are
generally low in the neocortex.
By contrast, the use of temporal difference error signals
enables all connections between cortical layers to be excit-
atory, and each layer can represent the positive encoding
of either the prediction or outcome state, at different
levels of abstraction. These properties are overwhelmingly
supported by extensive electrophysiological data about
the hierarchical organization of representations, for ex-
ample, in the visual object recognition pathway (Cadieu
et al., 2014; VanRullen & Thorpe, 2002; Kobatake &
Tanaka, 1994), and are consistent with the widely sup-
ported biased competition model for excitatory top–
down attentional effects (O’Reilly et al., 2013; Miller &
Cohen, 2001; Reynolds et al., 1999; Desimone & Duncan,
1995).
The EE approach requires net inhibitory top–down pre-
dictions, and it sends error signals forward, not positive
representations of the actual state at a given level of ab-
straction. Thus, a literal interpretation (and at least one ex-
isting implementation; Lotter et al., 2016) has only error
signals represented at all levels above the lowest level,
which is inconsistent with the positive encoding of stimuli
at various levels of abstraction across the visual hierarchy.
For example, although Issa, Cadieu, and DiCarlo (2018)
observed an error-signal-like increase in activation for
atypical faces in some posterior IT neurons, these neurons
overall had a positive stimulus encoding, with only a rela-
tively small, later, error-like modulation.
Furthermore, as discussed below, anticipatory predic-
tions typically closely resemble the subsequent stimulus-
driven activity, suggesting a positive, not inhibitory, effect
( Walsh et al., 2020; Cavanagh et al., 2010; Lee & Mumford,
2003; Duhamel et al., 1992). However, there are various
different ways of reformulating the neural implementation
of EE that can avoid some of these issues (Bastos et al.,
2012; Spratling, 2008), but perhaps, this flexibility renders
the framework difficult to falsify (Kogo & Trengove, 2015).
In any case, an extensive treatment of the issues with EE
is beyond the scope of this paper and has already been
aptly covered by Walsh et al. (2020)—our goal here is to
highlight some of the core differences as a way to clar-
ify the framework by way of contrast and in relation to
available data.
First, there are many examples of anticipatory predictive
neural firing in the brain. Of perhaps greatest relevance,
Barczak et al. (2018) recently showed that the auditory
pulvinar in monkeys exhibits predictive firing using a care-
fully controlled auditory sequence that had no first-order
acoustic differences from a background noise signal. The
pulvinar predictive activation preceded that of A1, suggest-
ing a strong predictive role for pulvinar. Unfortunately, the
deep layers of higher auditory areas that should contribute
to the formation of the pulvinar prediction were not re-
corded in this study, so their role in generating the predic-
tion could not be determined.
Nevertheless, there is extensive additional evidence for
top–down anticipatory activation of predicted stimuli,
with activity patterns closely resembling the subsequent
stimulus-driven ones ( Walsh et al., 2020). For example,
the widely replicated predictive remapping effect, simulated
in our model (Figure 12), is of this nature (Cavanagh et al.,
2010; Wurtz, 2008; Duhamel et al., 1992). The fact that
these anticipatory activations are of a positive nature,
consistent with the stimulus-driven activations, is incon-
sistent with the expected behavior of EE neurons, which
should be inhibited by the top–down prediction, while
not receiving any bottom–up stimulus.
However, the neural response to the actual predicted
stimulus itself is typically suppressed relative to unexpected
stimuli, that is, expectation suppression (Bastos et al., 2012;
Meyer & Olson, 2011; Todorovic et al., 2011; Summerfield
et al., 2008). This phenomenon is widely cited as evidence
in favor of the EE predictive coding framework, consistent
with an inhibitory effect of the expectation. Nevertheless,
despite various conflicting results and many complications
of interpretation, multiple comprehensive reviews con-
clude that it is difficult to distinguish expectation suppres-
sion from the neural adaptation effects that underlie the
well-documented repetition suppression effect ( Walsh
et al., 2020; Kok & de Lange, 2015; Vinken & Vogels,
2017; Kok et al., 2012; Summerfield & Egner, 2009; Lee
& Mumford, 2003). Furthermore, detailed single-neuron-
level recordings are the least likely to show these effects—
instead, they are most evident in aggregate signals such
as the BOLD response in fMRI, suggesting that they may
more strongly reflect population-level differences in activity,
rather than individual EE coding neurons.
As noted earlier, accurately predicted outcomes in our
framework would result in a continued adaptation of the
neural response carrying over from the prediction to the
outcome state, whereas unexpected outcomes would be
associated with two distinct patterns of activity over a
given area: first the prediction and then the outcome.
Thus, the unexpected outcome state would not be sub-
ject to the prior neural adaptation effects, and further-
more, the time-integrated aggregate activity over these
two patterns would be greater compared to the single ac-
tivity state associated with an accurately predicted outcome.
Thus, our model explains expectation suppression with-
out invoking EE neurons, meaning that considerably more
detailed and replicable experimental paradigms using
single-neuron resolution techniques are needed to distin-
guish EE from our framework.
1174
Journal of Cognitive Neuroscience
Volume 33, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
1
5
8
1
9
1
3
5
7
7
/
/
j
o
c
n
_
a
_
0
1
7
0
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Alpha Frequency Effects
The alpha frequency bursting of 5IB neurons acting as
drivers into the pulvinar naturally entrains the predictive
learning process in our model to this fundamental rhythm,
which has long been recognized as an important signature
of posterior cortical function ( VanRullen & Koch, 2003;
Varela, Toro, John, & Schwartz, 1981; Nunn & Osselton,
1974; Walter, 1953; Berger, 1929). A number of different
functional associations with alpha have been established,
and this literature is large and growing rapidly. Thus, we
refer the reader to recent reviews (Foster & Awh, 2019;
Clayton et al., 2018; VanRullen, 2016; Jensen, Bonnefond,
Marshall, & Tiesinga, 2015) while highlighting the data
most relevant to our specific framework here, organized
according to a set of key points.
(cid:129) Alpha is specifically associated with deep neocortical
layers and the pulvinar as well as with feedback path-
ways in the cortex. This has been established using di-
rect laminar-specific electrophysiological single-neuron
and local field potential recordings (Luczak et al., 2013;
Spaak, Bonnefond, Maier, Leopold, & Jensen, 2012;
Xing, Yeh, Burns, & Shapley, 2012; Buffalo et al., 2011;
Maier, Aura, & Leopold, 2011; Maier, Adams, Aura, &
Leopold, 2010) and feedforward versus feedback
manipulations (Michalareas et al., 2016; Bastos et al.,
2015; Jensen et al., 2015; van Kerkoerle et al., 2014;
von Stein, Chiang, & König, 2000). These data are con-
sistent with the 5IB alpha bursting and the major role of
cortical deep layers in driving top–down corticocortical
projections (in addition to the 6CT pathway that is spe-
cific to the pulvinar). By contrast, these same studies
show that superficial cortical layers are associated with
gamma frequency (40-Hz) dynamics. However, the next
point raises some important interpretational difficulties.
(cid:129) Increases in cortical activity levels, for example, because
of attention, produce a corresponding decrease in
alpha power, whereas decreased activity increases alpha
power (Foster & Awh, 2019; Jensen & Mazaheri, 2010;
Fries, Womelsdorf, Oostenveld, & Desimone, 2008;
Klimesch, Sauseng, & Hanslmayr, 2007; Kelly, Lalor,
Reilly, & Foxe, 2006; Worden, Foxe, Wang, & Simpson,
2000). This pattern is not exactly what you might expect if
alpha was a signature of predictive learning. Furthermore,
given that these same pulvinar and thalamocortical
pathways are also widely regarded as important for
attention (Fiebelkorn & Kastner, 2019; Zhou et al., 2016;
Saalmann & Kastner, 2011; Snow et al., 2009; Bender
& Youakim, 2001; LaBerge & Buchsbaum, 1990), this
pattern presents a challenge for many theorists. However,
it is possible to explain this pattern as arising directly
from the desynchronizing effects of cortical activity on
alpha power. Specifically, neural spiking is associated
with broadband noise, because of the highly random,
Poisson nature of spike firing, which can desynchronize
the entrainment of lower-frequency oscillations includ-
ing alpha (Solomon et al., 2017; Privman, Malach, &
Yeshurun, 2013; Waldert, Lemon, & Kraskov, 2013;
Ray & Maunsell, 2011). In other words, because cortical
activity is inherently noisy, it tends to interfere with the
coherent activity across populations of neurons needed
to produce a strong alpha frequency power signal. This
explanation is directly supported by studies manipulat-
ing and measuring cortical activity (Zhou et al., 2016;
Fries et al., 2008) and is consistent with alpha power
changes being a result of attentional modulation, but
not their cause (Antonov, Chakravarthi, & Andersen,
2020). Thus, although attention and predictive learn-
ing can both affect overall activity levels in the cortex
and thus drive changes in alpha power, alpha power it-
self is not a transparent measure of the underlying
mechanisms supporting these functions, which may
help to explain some contradictory patterns of results
(Gundlach, Moratti, Forschack, & Müller, 2020; Foster &
Awh, 2019; Keitel et al., 2019).
(cid:129) Alpha phase effects provide a more direct measure of
thalamocortical function than alpha power and have
been more consistently related to perception, attention,
and prediction (Solís-Vivanco, Jensen, & Bonnefond,
2018; Neupane et al., 2017; Jaegle & Ro, 2013; Palva &
Palva, 2011; Mathewson, Fabiani, Gratton, Beck, & Lleras,
2010; Busch, Dubois, & VanRullen, 2009; VanRullen &
Koch, 2003; Varela et al., 1981; Nunn & Osselton, 1974).
For example, weak, near-threshold stimuli are more
reliably detected and processed when presented in
the trough of the individual’s ongoing alpha cycle. Of
greatest relevance to this paper are studies showing
effects of prediction on alpha phase (Mayer, Schwiedrzik,
Wibral, Singer, & Melloni, 2016; Sherman, Kanai, Seth,
& VanRullen, 2016; Samaha, Bauer, Cimaroli, & Postle,
2015). For example, Mayer et al. (2016) showed that
prestimulus alpha phase directly correlated with the
predictability of the upcoming stimulus, and the pat-
tern of this prestimulus activation was indistinguish-
able from the subsequent stimulus activation pattern.
This is consistent with our model, and less consistent
with the EE framework, as discussed previously.
Neupane et al. (2017) found strong alpha coherence
effects in local field potential recordings distributed
across V4, associated with the predictive remapping
of receptive fields (Duhamel et al., 1992).
(cid:129) Discrete, salient, or oscillatory stimuli entrain the alpha
cycle in the brain (Spaak, de Lange, & Jensen, 2014;
Mathewson et al., 2012). Furthermore, the massive lit-
erature on ERPs may represent a significant contribu-
tion from alpha-level entrainment (Klimesch, 2011;
Gruber, Klimesch, Sauseng, & Doppelmayr, 2005;
Makeig et al., 2002). These entrainment effects are con-
sistent with the 5IB entrainment mechanisms in our
framework, as described earlier, and entrainment is
functionally important for aligning predictive learning
with relevant salient or unexpected outcomes.
(cid:129) The pulvinar contributes to synchronizing alpha phase
relationships across different brain areas (Fiebelkorn,
O’Reilly et al.
1175
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
1
5
8
1
9
1
3
5
7
7
/
/
j
o
c
n
_
a
_
0
1
7
0
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Pinsk, & Kastner, 2018; Saalmann et al., 2012). This is
consistent with the broad, convergent pattern of pro-
jections into the pulvinar from many different cortical
areas, and the corresponding broad projections back
out to these same areas (Arcaro et al., 2015; Shipp,
2003). Functionally, this convergence and synchroniza-
tion are important for integrating the contributions
from these different areas at the same time, to generate
predictions over the pulvinar.
(cid:129) The theta cycle, composed of a pair of alpha cycles,
organizes saccades as well as attentional, motor, and
mnemonic processes (Fiebelkorn & Kastner, 2019).
The theta rhythm is dominant in the medial temporal
lobe and hippocampus and has been extensively stud-
ied there (Buzsáki, 2005; Kahana, Seelig, & Madsen,
2001). Furthermore, there is a sharp peak of saccade
fixation durations at 200 msec, which suggests that
two alpha cycles are typically required for complete
processing of a given fixation. On the first cycle, the
predictions from before the eye moved may be fairly
vague depending on factors such as the size of the sac-
cade and familiarity with the environment. However,
after the first alpha cycle of a fixation, a subsequent
postdiction phase can provide an important additional
learning opportunity, to consolidate and more deeply
encode the current fixation (computationally equiva-
lent to an autoencoder). In addition, a mix of smaller
saccades (including microsaccades) and larger sac-
cades enables a range of more and less predictable out-
comes on the first alpha cycle after the saccade and
matches human behavior (Martinez-Conde, Otero-
Millan, & Macknik, 2013; Martinez-Conde, Macknik,
& Hubel, 2004).
Putting all of these points together, a particularly effec-
tive way of testing the predictions of our framework would
be measuring alpha phase changes emerging in the pre-
stimulus period as a function of predictive learning in pre-
dictable sequential stimulus streams. In addition, it would
also be important to examine theta- and alpha-cycle dy-
namics in relation to predictive learning in the context of
attention, motor control, and memory processes, to better
understand the larger systems-level temporal organization
of learning and processing in the brain (Fiebelkorn &
Kastner, 2019).
Predictions for Predictive Learning
In this section, we enumerate a set of direct, testable pre-
dictions from our framework. Before doing so, there are
several important considerations for any experimental test
of the theory. First, the nature of what is to be learned
must be matched to the pulvinar area in question. For ex-
ample, learning a new variation of basic physics in movies
at the alpha time scale (e.g., altering properties such as
gravity, inertia, or elasticity) would be appropriate for
the lower-level visual pathways. At higher visual levels
(e.g., IT cortex), it might be possible to use simple se-
quences of different objects, although it is not clear to
what extent the hippocampus or PFC might also contrib-
ute in this case (Fiser et al., 2016; Gavornik & Bear, 2014).
To distinguish pulvinar learning effects from pervasive
motor learning supported by other brain areas, it would
be most effective to directly measure activity in the pulvi-
nar and/or associated perceptual neocortical areas, instead
of involving overt behavioral performance.
Much of the learning in posterior sensory cortex should
take place early in development, requiring very early de-
velopmental interventions or genetic knockouts that are
expressed from the start (which can also have other inter-
pretational issues if not highly selective). In our models,
the bulk of the basic sensory predictive learning happens
very quickly, because the basic first-level regularities are
quite strong and relatively easily learned. Although there
are longer-term changes in the higher-level pathways in
our models, more fine-grained measurements would
likely be required to see these changes. Once this learning
has taken place, the remaining contributions of the thala-
mocortical circuit are likely more strongly weighted
toward its role in attention, as we discuss below. Finally,
directly lesioning or inactivating the pulvinar is not likely
to be very informative, because existing work has shown
dramatic effects on cortical activity (Zhou et al., 2016;
Purushothaman, Marion, Li, & Casagrande, 2012), and
furthermore, any effects could be attributed to the atten-
tional contributions of the pulvinar.
With these considerations in mind, here are a set of
strong predictions from our model that should be testable
using existing techniques. Failure to obtain the predicted
result, while adhering to all the relevant constraints, would
constitute a falsification of our model.
(cid:129) Blocking 5IB bursting mechanisms early in develop-
mental learning should disrupt learning. It should be
possible to selectively knock out or modify the channels
that cause this specific population of neurons to burst
fire, and doing so should have a significant effect on
learning in associated neocortical and pulvinar areas,
given the critical role that this burst firing plays on the
predictive learning process, as elaborated above.
(cid:129) Blocking synaptic plasticity in the pulvinar (specifically
the 6CT inputs) very early in developmental learning
should impair learning. Although most of the learning
overall should occur in the neocortex as a result of the
temporal difference error signal broadcast by the pulvi-
nar (which should remain generally intact), learning in
the 6CT projections is important, especially right at the
start, to map the emerging neocortical representations
into the space defined by the 5IB projections.
(cid:129) Temporal differences on an alpha cycle timescale actu-
ally drive synaptic plasticity in an error-driven learning
manner, in neocortical pyramidal neurons and in 6CT
inputs to the pulvinar. That is, if a pre/post pair of neu-
rons across a synapse is more active in the prediction
1176
Journal of Cognitive Neuroscience
Volume 33, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
1
5
8
1
9
1
3
5
7
7
/
/
j
o
c
n
_
a
_
0
1
7
0
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
than the subsequent outcome, the synapse should ex-
perience long-term depression, and vice versa if the
activity pattern is reversed (long-term potentiation, for
more activity in outcome than prediction). Furthermore,
if activity is essentially stable across both prediction
and outcome phases, then weights should not change
(modulo, a small level of Hebbian learning; O’Reilly
et al., 2012; O’Reilly & Munakata, 2000). This should
be directly testable using current experimental methods
and is perhaps the single most important empirical test
of this entire framework, and it also underlies many
other current approaches to error-driven learning in
the brain (Lillicrap et al., 2020; Whittington & Bogacz,
2019; Bengio et al., 2017). One general consideration is
the extent to which an awake in vivo preparation would
be required to capture all the neuromodulatory and
other factors present when this learning normally takes
place. Some suggestive evidence in such a preparation
is generally consistent with a sensitivity to relatively
short-term temporal dynamics (Lim et al., 2015), al-
though these results lacked the direct measurement
of individual neural activity across a synapse.
DISCUSSION
We have hypothesized a novel computational function for
the distinctive features of thalamocortical circuits (Usrey &
Sherman, 2018; Sherman & Guillery, 2006), as supporting
a specific form of prediction-error driven learning, where
predictions arise from the numerous top–down layer 6CT
projections into the pulvinar, and the strong, sparse driv-
ing 5IB inputs supply the bottom–up sensory-driven out-
come. The phasic bursting nature of the 5IB inputs results
in a natural temporal-difference error signal of prediction
followed by outcome, consistent with extensive neural re-
cording data. This temporal dynamic is also essential for
enabling predictions to be generated without contamina-
tion from current sensory inputs and predicts a character-
istic alpha-frequency prediction cycle based on the 10-Hz
bursting cycle of the 5IB inputs, consistent with the perva-
sive influence of alpha on perception and neural dynamics
(Foster & Awh, 2019; Clayton et al., 2018; VanRullen, 2016;
Jensen et al., 2015). In short, the hypothesized predictive
learning function fits remarkably well with a number of
well-established properties of these thalamocortical cir-
cuits, and we also provided a set of additional predictions
that could be tested to further evaluate this theory, espe-
cially in contrast to the widely discussed alternative of EE
coding neurons, which have not been unambiguously
supported across a range of empirical studies (Walsh et al.,
2020).
Furthermore, we implemented this theory in a large-
scale model of the visual system and demonstrated that
learning based strictly on predicting what will be seen next
is, in conjunction with a number of critical biologically
motivated network properties and mechanisms, capable
of generating abstract, invariant categorical representa-
tions of the overall shapes of objects. The nature of these
shape representations closely matches human shape sim-
ilarity judgments on the same objects. Thus, predictive
learning has the potential to go beyond the surface struc-
ture of its inputs and develop systematic, abstract encodings
of the environment. We found that comparison models
based on standard error Bp learning did not learn a cate-
gorical structure that went beyond the surface similarity
present in the visual input layers, and future work is focused
on narrowing down the specific mechanisms required to
drive this learning.
In addition to the predictive learning functions of the
deep/thalamic layers, these same circuits are also likely
critical for supporting powerful top–down attentional
mechanisms that have a net multiplicative effect on
superficial-layer activations (Bortone, Olsen, & Scanziani,
2014; Olsen, Bortone, Adesnik, & Scanziani, 2012). The
importance of the pulvinar for attentional processing
has been widely documented (e.g., Saalmann et al.,
2012; Bender & Youakim, 2001; LaBerge & Buchsbaum,
1990), and there is likely an additional important role of
the thalamic reticular nucleus, which can contribute a
surround-inhibition contrast-enhancing effect on top of
the incoming attentional signal from the cortex (Jaramillo,
Mejias, & Wang, 2019; Wimmer et al., 2015; Pinault, 2004;
Crick, 1984). In other work in progress, we have shown
that the deep/thalamic circuits in our model produce at-
tentional effects consistent with the abstract Reynolds
and Heeger (2009) model, whereas the contributions
of the deep layer networks to this function are broadly
consistent with the folded-feedback model (Grossberg,
1999). These attentional modulation signals cause the bi-
directional constraint satisfaction process in the superficial
network to focus on task-relevant information while
down-regulating responses to irrelevant information—in
the real world, there are typically too many objects to
track at any given time, so predictive learning must be di-
rected toward the most important objects (Richter & de
Lange, 2019; Cavanagh et al., 2010; Pylyshyn, 1989).
There are also data suggesting that the pulvinar is im-
portant for supporting confidence judgments, driven by
relative ambiguity in a random dot motion categorization
task (Komura et al., 2013). Critically for the present frame-
work, this confidence modulation only emerged in the pe-
riod after the first 100 msec of processing and manifested
as a positive correlation with confidence (i.e., more unam-
biguous stimuli resulted in higher firing rates). We can in-
terpret this as reflecting an ongoing generative postdiction
of the stimulus signal, with stronger firing associated with
more unambiguous top–down activation based on the
current internal representation. Note that this directional-
ity is the opposite of EE coding neurons, which would pre-
sumably increase with increasing error/ambiguity in the
prediction. Interestingly, inactivation of these pulvinar
neurons resulted in a substantial (200%) increase in opt-
out choices on the most ambiguous stimuli, suggesting a
O’Reilly et al.
1177
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
1
5
8
1
9
1
3
5
7
7
/
/
j
o
c
n
_
a
_
0
1
7
0
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
level of metacognitive awareness of the pulvinar signal (or
at least a direct effect of pulvinar on relevant metacognitive
processes). Predictive accuracy would be an ideal source
of metacognitive confidence signals across a wide range
of domains, suggesting another important contribution of
pulvinar even after initial learning. Jaramillo et al. (2019)
present a comprehensive model of attentional, decision-
making, and working memory contributions of the pulvi-
nar, including these confidence data, which is generally
compatible with our framework, although it does not ad-
dress any learning phenomena.
There are a number of important limitations of the cur-
rent What–Where Integration ( WWI) model, in terms of
its scale and ability to process real-world cluttered visual
scenes with multiple objects present, such as those used
in the widely studied ImageNet data set. The model is much
smaller than standard DCNN vision models, because its
computational demands are significantly higher, in a way
that also does not fit well with current graphics processing
unit (GPU)-based parallel computation hardware, because
of the relative complexity of the algorithms and the sparse-
ness of the activations. For each image, 100 cycles (of 1 msec
each) of activation updating are required to enable the bidi-
rectional activation and inhibition to integrate in a graded
manner over the alpha cycle, compared to only one such it-
eration for most feedforward DCNN models. Furthermore,
the bidirectional connectivity, extensive shortcut connec-
tions, and use of multiple cortical lamina per cortical area
result in significant increases in the number of synaptic con-
nections, which dominate the computational cost, and scale
roughly as n2 in the number of neurons n per layer across
one projection. Thus, there are 207 million connections for
the full WWI model, requiring 10 GB of RAM, and it takes
over a day to run using 32 high-performance CPU processors
with fast network interconnects, using the fastest combina-
tion of threading and parallel batch training. Doubling the
network size causes it to no longer fit in available RAM,
and yet, its high-resolution V1 layer is only 16 × 16, com-
pared to 55 × 55 for basic DCNN models such as AlexNet
and 224 × 224 for VGG16. The result is that the model
has a relatively low-resolution view of the world, as reflected
in the reconstructed images shown in Figure 5.
In addition to having a higher-resolution input to be
able to process more complex real-world cluttered images,
the model would require functional attentional dynamics
to focus processing on a small number of objects at a time,
as is well documented for humans processing complex
images. Thus, once the attentional dynamics are well inte-
grated with the predictive learning mechanisms, we can
begin to explore performance on more complex images,
subject to improved computational hardware supporting
larger network sizes.
Considerable further work remains to be done to more
precisely characterize the essential properties of our bio-
logically motivated model necessary to produce this
abstract form of learning and to further explore the full
scope of predictive learning across different domains.
We strongly suspect that extensive cross-modal predictive
learning in real-world environments, including between
sensory and motor systems, is a significant factor in infant
development and could greatly multiply the opportunities
for the formation of higher-order abstract representations
that more compactly and systematically capture the struc-
ture of the world ( Yu & Smith, 2012). Future versions of
these models could thus potentially provide novel insights
into the fundamental question of how deep an under-
standing a preverbal human, or a nonverbal primate, can
develop (Elman et al., 1996; Spelke, Breinlinger, Macomber,
& Jacobson, 1992), based on predictive learning mecha-
nisms. This would then represent the foundation upon
which language and cultural learning builds, to shape the
full extent of human intelligence.
APPENDIX
All of the materials described here, including the experi-
mental study, the computational models, and the code
to perform the representational similarity analysis, are all
available on our github account at github.com/ccnlab
/deep-obj-cat, and the new version of the emergent simu-
lation environment is at github.com/emer/leabra, which
contains extensive documentation and examples that
can be run in Python or the Go language. The best place
to start in understanding computationally how the predic-
tive learning model works is with the FSA model described
in the main text, which is available at github.com/emer
/leabra/tree/master/examples/deep_fsa. For the large and
complex WWI model, the most complete understanding
can only be had by directly examining the code, as there
are a number of details that are not efficiently captured in
this Appendix text.
REPRESENTATIONAL SIMILARITY
ANALYSIS METHODS
The different representations being compared here are
the following:
Leabra: The DeepLeabra (biological model) TE layer
representations (specifically TEs = superficial—results are
very similar for deep as well).
Bp: The TEs layer representations from the Bp version of
biological model, including “what,” “where,” and “What ×
Where” integration layers, trained with the V1p and V1hp
(low- and high-resolution pulvinar) layers as the final output
layers, using the time t target pattern from the t – 1 input
(i.e., as a predictive network).
V1: The Gabor-filtered representation of the visual input
to both of the above models, which was identical across
them.
PredNet: The highest layer (sixth layer) of the PredNet
architecture.
Expt: Similarity matrix constructed from human pair-
wise similarity judgments (see Behavioral Experiment
Methods).
1178
Journal of Cognitive Neuroscience
Volume 33, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
1
5
8
1
9
1
3
5
7
7
/
/
j
o
c
n
_
a
_
0
1
7
0
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
An optimal category cluster can be defined as one that
has high within-cluster similarity and low between-cluster
similarity. This can be operationalized by the contrast dis-
tance metric, based on a 1-correlation (dissimilarity) mea-
sure, as the difference between the average within-cluster
similarity and the average between-cluster similarity:
ð
CD ¼ 1− rin
Þ− 1− rout
ð
Þ
(1)
With distance-like 1-correlation values, this contrast distance
should be minimized (it is typically negative), or equiva-
lently, the contrast on raw correlation values can be max-
imized (it is typically a positive number—just the sign flip
of distance value). We refer to the positive numbers and
maximization here as that is more intuitive.
Starting with an initial set of clusters, a permutation-based
hill-climbing strategy was used to determine a local mini-
mum in this measure: Each item was tested in each of the
other possible categories, and if that configuration reduced
the overall average contrast distance (ACD) metric across all
items, then it was adopted and the process iterated until no
such permutation improved the metric. This algorithm can
only decrease the number of clusters (by moving all items
out of a given cluster), so different numbers of initial clus-
ters can be used to search the overall space.
Figure 14 shows the resulting categories. The Bp model
converged on the same cluster state from all starting con-
figurations tested, varying from five to two initial catego-
ries. This is the cluster set shown in Figure 10 of the
main paper and has an ACD of 0.0838 (this is relatively
low because the patterns were overall quite similar).
Likewise, the V1 patterns (which were the same across
Leabra and Bp models) reliably converged on the same
pattern (shown in Figure 10), with ACD = 0.2448.
For the PredNet Layer 6 representations, starting from
the V1 categories gave the best results of any other set
(ACD = 0.1967), and a few permutations resulted in a reli-
able solution that was arrived at from all other three cate-
gory starting points tested, shown in Figure 14 (ACD =
0.2820). This indicates that PredNet did not go much be-
yond the structure present in the input, although it did not
use the V1 Gabor filtering used in the Leabra and Bp
models (i.e., this V1-level encoding well captures the struc-
ture of the visual inputs in general). The PredNet pixel and
Layer 1 representations both converged on essentially a
single monolithic category with very low ACDs (0.0018
and 0.0013, respectively).
For the Leabra TE representations, we found a set of
centroid-shape categories that are near-best when considering
both the Leabra model and the results from the human be-
havioral experiment. Starting from these categories, the
permutation analysis converged on reducing the size of
the vertical and round categories to one item each, over a
sequence of five steps. This is consistent with the obser-
vation from Figure 7 that there are three broader catego-
ries within which the five finer-grained categories are
embedded (i.e., vertical and pyramid are overall similar
to each other, as are round and box). Nevertheless, our
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
1
5
8
1
9
1
3
5
7
7
/
Figure 14. Shape categories
used for similarity matrix plots
in the main paper. “Centroid”
shape categories are near-best
for both the Leabra model and
the Expt results and fit our
visual intuitions about overall
shape. “Bp” is reliably optimal
for the Bp model from all
starting points. “V1” reliably
optimal for V1 inputs and also
was close to the best for the
Bp and PredNet Layer 6
representations. “PredNet” is
the best stable solution for
PredNet Layer 6.
/
j
o
c
n
_
a
_
0
1
7
0
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
O’Reilly et al.
1179
initial visual intuition about the broad shape categories,
along with a bias against having single-item categories,
reinforced the use of the finer-grained centroid selection.
The average contrast difference of our centroid selection
is 0.5071, whereas the maximal result from the permuta-
tion was 0.5526, which is a relatively small proportional
difference.
Furthermore, once we had collected the human exper-
imental data (Expt), it was clear that it strongly coincided
with our original shape intuitions and with the finer-
grained five-category centroid structure. Starting from
the centroid categories, the maximal permutation made
only three changes, moving trex (T-rex) and handgun into
the horizontal category, and chair into the pyramid, going
from a distance score of 0.3083 to 0.3225, which is a rela-
tively small improvement. However, using the maximal
Expt clusters directly on the Leabra model gives a lower
ACD measure of 0.3745 (compared to 0.5071 for centroid),
so the centroid categories represent a good middle ground
between Expt and the model, and this strong shared simi-
larity structure with near-optimal cluster structures
confirms that the model and people are encoding largely
the same information.
In contrast, if we organize the Expt similarity matrix using
the Bp categories, it produces a very poor ACD measure of
0.0643 (compared to 0.3083 for the centroid categories),
strongly suggesting that people’s shape representations
are not compatible with that simple structure.
Another approach to determining clusters from similarity
matrices, “agglomerative clustering,” starts with all items as
singletons and iteratively combines the closest two into a
new cluster. The results for the Leabra and Expt similarity
matrices are shown in Figure 15, which has also color-coded
the items in terms of their category status according to the
centroid structure. Because of a strong history dependency
in the clustering process and the indeterminacy of reducing
a high-dimensional similarity structure down to two dimen-
sions, structure beyond the leaf level is not very reliable (ties
are also broken by a random number generator), but nev-
ertheless, you can clearly see that, in both cases, items from
the same cluster are almost always together as leaves in the
plots. This then provides additional converging support for
the idea that the model is learning the same kind of shape
categories as people have.
For the network layer RSA computations, activation vec-
tors were accumulated separately for each 3-D object item
and, within that, separately for each frame index of the
movie. To be able to monitor similarity metrics as the model
trained, we used a running-average integration of neural
activity across trials to accumulate the patterns. Specifically,
the current activation pattern across each layer was recorded
and averaged unit-by-unit with a time constant of τ = 10.
Critically, by integrating separately for each frame, this
running-average computation did not introduce any bias
for temporally adjacent frames to be more similar.
Nevertheless, when we computed the frame-to-frame
Figure 15. Agglomerative clustering on the Leabra and Expt representations, with the centroid categories color coded. The most reliable information
from this is the leaf-level groupings, as the rest of the structure is indeterminate and history dependent in reducing higher-dimensional structure
down to a 2-D plot. Both cluster plots show a strong tendency to group leaf items together in the same centroid categories, with a few exceptions in
each case. In addition, the Leabra plot nicely captures the broader three-category structure evident in the similarity matrix plots, within which the five
finer-grained centroid categories are organized. Overall, this provides further confirmation that the model and the human participants are organizing
the shapes in largely the same way.
1180
Journal of Cognitive Neuroscience
Volume 33, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
1
5
8
1
9
1
3
5
7
7
/
/
j
o
c
n
_
a
_
0
1
7
0
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
similarities for TE, they were quite high (.901 correlation
on average across all objects).
All the activation and general learning parameters in the
model are at their standard Leabra defaults.
BEHAVIORAL EXPERIMENT METHODS
The behavioral experiment was conducted on Amazon.com’s
MTurk Web platform under University of Colorado
institutional review board approval (19-0176), using 30
participants each categorizing up to 800 image pairs as
shown in Figure 16, using the standard simple image cat-
egorization framework with a lightly customized script.
Objects were drawn from the 156 3-D object set, but data
were aggregated in terms of the 20 basic-level categories
(car, stapler, etc.) because we could not sample all 156 ×
156 object pairs. Thus, the resulting data were aggregated
for each category pair in terms of the proportion of times
when that pair was selected when presented.
The individual images were produced by reconstructing
from the V1 transform that the computational model used
in its high-resolution V1 input layer, to give human partic-
ipants as similar of an experience as possible to how the
model “saw” the objects, and to reduce the influence of
existing semantic knowledge, which was entirely missing
in our model (Figure 16).
BIOLOGICAL MODEL METHODS
This section provides more information about the DeepLeabra
WWI model. The purpose of this information is to give
more detailed insight into the model’s function beyond
the level provided in the main text, but with a model of
this complexity, the only way to really understand it is to
explore the model itself. It is available for download at
github.com/ccnlab/deep-obj-cat/tree/master/sims/cemer.
We now have a full replication of this model in our new,
much more transparent simulation framework, available
at github.com/ccnlab/deep-obj-cat/tree/master/sims
/wwi3d—this is more readable and recommended.
Furthermore, the best way to understand this model is
to understand the framework in which it is implemented,
which is explained in great detail, with many running sim-
ulations explaining specific elements of functionality, at
CompCogNeuro.org.
Layer Sizes and Structure
Figure 5 in the main text shows the general configuration
of the model, and Table 1 shows the specific sizes of each
of the layers and where they receive inputs from.
Projections
The general principles and patterns of connectivity are
shown in Figure 17 (and Figures 1 and 2 in the main text).
As noted in the main text, the connectivity and overall
structure obeys the established principles identified in
neocortical anatomy (Markov, Ercsey-Ravasz, et al., 2014;
Markov, Vezoli, et al., 2014; Felleman & Van Essen, 1991;
Rockland & Pandya, 1979).
Detailing each of the specific parameters associated
with the different projections shown in Table 1 would take
too much space—those interested in this level of detail
should download the model from the link shown above.
There are topographic projections between many of the
lower-level retinotopically mapped layers, consistent with
our earlier vision models (O’Reilly et al., 2013). For exam-
ple the 8 × 8 unit groups in V2 are reduced down to the
4 × 4 groups in V3 via a 4 × 4 unit-group topographic pro-
jection, where neighboring units have half-overlapping
receptive fields (i.e., the field moves over two unit groups
in V2 for every one unit group in V3), and the full space is
uniformly tiled by using a wraparound effect at the edges.
Similar patterns of connectivity are used in standard
DCNNs. However, we do not share weights across units
as in a true convolutional network.
The projections from ObjVel (object velocity) and
SaccadePlan layers to LIPs, LIPd were initialized with a
topographic sigmoidal pattern that moved as a function
of the position of the unit group, by a factor of .5, whereas
the projections from EyePos were initialized with a
Gaussian pattern. These patterns multiplied uniformly
distributed random weights in the .25–.75 range, with
the lowest values in the topographic pattern having a mul-
tiplier of .6, whereas the highest had a multiplier of 1 (i.e.,
a fairly subtle effect). This produced faster convergence of
the LIP layer when doing “where” pathway pretraining
compared to purely random initial weights, consistent
with Pouget and Sejnowski (1997) and related work on
parietal gain field basis function representations.
In addition to exploring different patterns of overall con-
nectivity, we also explored differences in the relative
strengths of receiving projections, which can be set with
a wt_scale.rel parameter in the simulator. All feedforward
pathways have a default strength of 1. For the feedback
Figure 16. Example stimulus
from the behavioral experiment,
using the V1 reconstruction of the
actual input images presented to
the model, to better capture the
coarse-grained perception of
the model. Participants were
requested to choose which of the
two pairs, left or right, was most
similar in terms of overall shape.
O’Reilly et al.
1181
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
1
5
8
1
9
1
3
5
7
7
/
/
j
o
c
n
_
a
_
0
1
7
0
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Table 1. Layer Sizes, Showing Numbers of Units in One Pool (or Entire Layer if Pool is Missing), and the Number of Pools of Such
Units, along x, y Axes
Unites
Pools
Area
V1
V1h
Name
V1s
V1p
V1hs
V1hp
Eyes
EyePos
SaccadePlan
Saccade
Obj
V2
LIP
V3
DP
V4
ObjVel
V2s
V2d
MtPos
LIPs
LIPd
LIPp
V3s
V3d
V3p
DPs
DPd
DPp
V4s
V4d
V4p
x
4
4
4
4
21
11
11
11
10
10
1
4
4
1
10
10
10
10
10
10
10
10
10
y
5
5
5
5
21
11
11
11
10
10
1
4
4
1
10
10
10
10
10
10
10
10
10
x
8
8
16
16
8
8
8
8
8
8
4
4
4
4
4
4
y
8
8
16
16
8
8
8
8
8
8
4
4
4
4
4
4
Receiving Projections
V1s, V2d, V3d, V4d, TEOd
V1s, V2d, V3d, V4d, TEOd
V1s, LIPs, V3s, V4s, TEOd,
V1p, V1hp
V2s, V1p, V1hp, LIPd, LIPp,
V3d, V4d, V3s, TEOs
V1s
MtPos, ObjVel, SaccadePlan,
EyePos, LIPp
LIPs, LIPp, ObjVel, Saccade,
EyePos
MtPos, V1s, LIPd
V2s, V4s, TEOs, DPs, LIPs,
V1p, V1hp, DPp, TEOd
V3s, V1p, V1hp, DPp, LIPd,
DPd, V4d, V4s, DPs, TEOs
V3s, V2d, DPd, TEOd
V2s, V3s, TEOs, V1p ,V1hp,
V3p, TEOp
DPs, V1p, V1hp, DPp, TEOd
DPs, V2d, V3d, DPd, TEOd
V2s, TEOs, V1p, V1hp
V4s, V1p, V1hp, V4p,
TEOd, TEOs
V4s, V2d, V3d, V4d, TEOd
1182
Journal of Cognitive Neuroscience
Volume 33, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
1
5
8
1
9
1
3
5
7
7
/
/
j
o
c
n
_
a
_
0
1
7
0
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Table 1. (continued )
Unites
Pools
Area
TEO
TE
Name
TEOs
TEOd
TEOp
TEs
TEd
TEp
x
10
10
10
10
10
10
y
10
10
10
10
10
10
x
4
4
4
4
4
4
y
4
4
4
4
4
4
Receiving Projections
V4s, V1p, V1hp, TEs
TEOs, TEOd, V1p, V1hp,
V4p, TEOp, TEp, TEd
TEOs, V3d, V4d, TEOd, TEd
TEOs, V1p, V1hp
TEs, TEd, V1p, V1hp, V4p,
TEOp, TEp, TEOd
TEs, V3d, V4d, TEOd
Each area has three associated layers: s = superficial layer; d = deep layer (context updated by 51B neurons in the same area, shown in bold);
and p = pulvinar layer (driven by 5IB neurons from the associated area, shown in bold).
projections, which are typically weaker (consistent with
the biology), we explored a discrete range of strengths,
typically .5, .2, .1, and .05. The strongest top–down projec-
tions were into V2s from LIP and V3, whereas most others
were .2 or .1. Likewise, projections from the pulvinar were
weaker, typically .1. These differences in strength some-
times had large effects on performance during the initial
bootstrapping of the overall model structure, but in the
final model, they are typically not very consequential for
any individual projection.
Training Parameters
Training typically consisted of 512 alpha trials per epoch
(51.2 sec of real-time equivalent), for 1000 such epochs.
Each trial was generated from a virtual reality environment
in the emergent simulator, which rendered first-person
views with moving eye position onto the object tumbling
through space with fixed motion and rotation parameters
over the sequence of eight frames (see Figure 5 in the
main text for a representative example). Each frame was
rendered at a 256 × 256 resolution and processed through
our standard V1 Gabor filters, which are described in detail
in O’Reilly et al. (2013).
Because the start of each sequence of eight frames is un-
predictable, we turned off learning for that trial, which im-
proves learning overall. We have recently developed an
automatic such mechanism based on the running average
(and running variance) of the prediction error, where we
turn off learning whenever the current prediction error
z-normalized by these running average values is below
1.5 SDs, which works well and will be incorporated into fu-
ture models. Biologically, this could correspond to a con-
nection between pulvinar and neuromodulatory areas
that could regulate the effective learning rate in this way.
Figure 18A shows the learning trajectory of the model,
indicating that it learns quite rapidly. This rapid initial
learning is likely facilitated by the extensive use of shortcut
connections converging from all over the simulated visual
system onto the V1 pulvinar layers and direct projections
back from these pulvinar layers. Thus, error signals are di-
rectly communicated and can drive learning quickly and
efficiently. However, there are also extensive indirect, bi-
directional connections among the superficial layers,
which can drive indirect error Bp learning as well.
Model Algorithms
The biologically based model was implemented using the
Leabra framework, which is described in detail in previous
publications (O’Reilly et al., 2012, 2016; O’Reilly &
Munakata, 2000; O’Reilly, 1996, 1998), and summarized
here. The online textbook at CompCogNeuro.org
provides the most comprehensive description of the
framework, and github.com/emer/leabra has a summary
of all the equations (and the code itself ). There are two
main implementations of Leabra, one in the C++ emer-
gent software and a new one using Go and Python language
at the prior link. These same equations and standard pa-
rameters have been used to simulate over 40 different
models in O’Reilly and Munakata (2000), O’Reilly et al.
(2012), and a number of other research models. Thus,
the model can be viewed as an instantiation of a systematic
modeling framework using standardized mechanisms, in-
stead of constructing new mechanisms for each model
(O’Reilly et al., 2016).
The neurons use a rate code version of the adaptive ex-
ponential conductance-based point neuron model (Brette
& Gerstner, 2005), with the standard resistor-capacitor
circuit equations:
ΔVm tð Þ ¼ τ
X
c
gc tð Þ(cid:1)gc Ec − Vm tð Þ
ð
Þ;
(2)
where c represents excitatory, inhibitory, and leak channels.
Inhibition is driven by simulated interneurons in proportion
O’Reilly et al.
1183
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
1
5
8
1
9
1
3
5
7
7
/
/
j
o
c
n
_
a
_
0
1
7
0
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
1
5
8
1
9
1
3
5
7
7
/
Figure 17. Principles of connectivity in DeepLeabra. (A) Markov et al. (2014) data showing density of retrograde labeling from a given injection in a
middle-level area (“d”): Most feedforward projections originate from superficial layers of lower areas (“a–c”), and deep layers predominantly
contribute to feedback (and more strongly for longer-range feedback). (B) Summary diagram showing most feedforward connections originating in
superficial layers of lower area and terminating in Layer 4 of higher areas, whereas feedback connections can originate in either superficial or deep
layers and, in both cases, terminate in both superficial and deep layers of the lower area (adapted from Felleman & Van Essen, 1991). (C) Anatomical
hierarchy as determined by percentage of superficial layer source labeling by Markov et al. (2014)—the hierarchical levels are well matched for our
model, but we functionally divide the dorsal pathway (shown in green background) into the two separable components of a “where” pathway and a
What × Where integration pathway. (D) Superficial and deep-layer connectivity in the model. Note the repeating motif between hierarchically
adjacent areas, with bidirectional connectivity between superficial layers, and feedback into deep layers from both higher-level superficial and deep
layers, according to the canonical pattern shown in A and B. Special patterns of connectivity from TEO to V3 and V2, involving crossed super-to-deep
and deep-to-super pathways, provide top–down support for predictions based on high-level object representations. (E) Connectivity for deep layers
and pulvinar in the model, which generally mirror the corticocortical pathways (in D). Each pulvinar layer (p) receives 5IB driving inputs from the
labeled layer (e.g., V1p receives 5IB drivers from V1). In reality, these neurons are more distributed throughout the pulvinar, but it is computationally
convenient to organize them together as shown. Deep layers (“d”) provide predictive input into the pulvinar, and pulvinar projections send error
signals (via temporal differences between predictions and actual state) to both deep and superficial layers of given areas (only “d” shown). Most areas
send deep-layer prediction inputs into the main V1p prediction layer and receive reciprocal error signals therefrom. The strongest constraint we
found was that pulvinar outputs (colored green) must generally project only to higher areas, not to lower areas, with the exceptions of DPp → V3 and
LIPp → V2. V2p was omitted because it is largely redundant with V1p in this simple model.
/
j
o
c
n
_
a
_
0
1
7
0
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
to feedforward and feedback dynamics, producing sparse
distributed representations and controlling the effects of
bidirectional excitatory connections between layers.
Each neuron learns using a more biologically based ver-
sion of the contrastive Hebbian learning (CHL) algorithm,
as shown in Figure 2:
ΔCHL ¼ xþyþ− x−y−
(3)
where x is the sending activation, y is the receiving activa-
tion, and the + and − superscripts indicate activations in
the plus and minus phases, respectively. The actual learn-
ing equations, detailed at github.com/emer/leabra and in
the online textbook at CompCogNeuro.org, produce a
combination of error-driven and self-organizing factors,
which emerge out of a single learning rule that was derived
from a biologically detailed model of synaptic plasticity by
1184
Journal of Cognitive Neuroscience
Volume 33, Number 6
Figure 18. (A) Predictive
learning curve for DeepLeabra,
showing the correlation
between prediction and actual
over the two different V1 layers.
Initial learning is quite rapid,
followed by a slower but
progressive learning process
that reflects development of the
IT representations (e.g.,
manipulations that interfere
with those areas selectively
impair this part of the learning
curve). Overall prediction
accuracy remains far from
perfect, as shown in Figure 5 in
the main text, and significantly worse than the Bp-based models. This is a typical finding from Leabra models, which are significantly more
constrained as a result of bidirectional attractor dynamics, Hebbian learning, and inhibitory competition—that is, the very things that are likely
important for forming abstract categorical representations. (B) Similarity matrix over TEs layer at 200 epochs, which has less contrast and definition
(particularly evident in the off-block-diagonal differences) compared to the 1000-epoch result (C; also shown in Figure 7 in the main text).
(Urakubo et al., 2008) and is closely related to the BCM
algorithm (Bienenstock et al., 1982).
Deep Context
This section describes in detail the equations that are spe-
cific to the “deep” version of Leabra that implements the
specific predictive learning additions to the general algo-
rithm. Like the SRN (Elman, 1990; Jordan, 1989), which
the deep predictive learning model functionally resem-
bles, the primary computational specialization required
is the maintenance of prior temporal context in the CT
layer. In addition, the pulvinar layers have to be driven
by the bottom–up inputs in the plus phase, after being
driven by the CT inputs in the minus phase.
Computationally, the CT layer is specialized for main-
taining context from the previous alpha cycle, to generate
the prediction over the pulvinar layer. At the end of every
plus phase, a new CT context excitatory input is computed
from the normalized dot product of the context weights
times the sending activations, just as in the standard net
input used in Leabra:
(cid:2)
¼ xiwij
xiwij
X
(4)
(cid:3)
η
j
¼ 1
n
i
where xi are the sending activations and wij are the weights.
This net input is then added in with the standard net input at
each cycle of processing during the subsequent alpha cycle.
The relative strength of these context layer inputs was
set progressively larger for higher layers in the network,
with a maximum of four in V4, TEO, and TE. In addition,
TEO and TE received “self” context projections, which pro-
vide an extended window of temporal context into the
prior 200-msec interval, consistent with multiple sources
of neural data (Chaudhuri et al., 2015). These self projec-
tions were connected only within the narrower pool level
of units, enabling these neurons to develop mutually ex-
citatory loops to sustain activations over the multiple
trials when the same object was present. We hypothesize
that these modifications correspond to biological adapta-
tions in IT cortex that likewise support greater sustained
activation of object-level representations.
Learning of the context weights occurs as normal, but
using the sending activation states from the prior time
step’s activation.
Computational and Biological Details of
SRN-like Functionality
Predictive autoencoder learning has been explored in
various frameworks, but the most relevant to our model
comes from the application of the SRN to a range of pre-
dictive learning domains (Elman et al., 1996; Elman, 1990).
One of the most powerful features of the SRN is that it en-
ables error-driven learning, instead of arbitrary parameter
settings, to determine how prior information is integrated
with new information. Thus, SRNs can learn to hold onto
some important information for a relatively long interval,
while rapidly updating other information that is only rele-
vant for a shorter duration. This same flexibility is present
in our DeepLeabra model. Furthermore, because this tem-
poral context information is hypothesized to be present in
the deep layers throughout the entire neocortex (in every
microcolumn of tissue), the DeepLeabra model provides a
more pervasive and interconnected form of temporal inte-
gration compared to the SRN, which typically only has a
single temporal context layer associated with the internal
“hidden” layer of processing units.
An extensive computational analysis of what makes the
SRN work as well as it does, and explorations of a range
of possible alternative frameworks, has led us to an impor-
tant general principle: Subsequent outcomes determine
what is relevant from the past. At some level, this may seem
obvious, but it has significant implications for predictive
learning mechanisms based on temporal context. It means
that the information encoded in a temporal context
O’Reilly et al.
1185
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
1
5
8
1
9
1
3
5
7
7
/
/
j
o
c
n
_
a
_
0
1
7
0
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
long period in neural terms (e.g., a rapidly firing cortical neu-
ron fires at around 100 Hz, meaning that it will fire 10 times
within that context frame)? However, there is an important
transformation of the SRN context computation, which is
more biologically plausible and compatible with the struc-
ture of the deep network (Figure 19). Specifically, instead
of copying an entire set of activation states, the context
activations (generated by the phasic 5IB burst) are imme-
diately sent through the adaptive synaptic weights that
integrate this information, which we think occurs in the
6CC (corticortical) and other lateral integrative connec-
tions from 5IB neurons into the rest of the deep network.
The result is a precomputed net input from the context
onto a given hidden unit (in the original SRN terminology),
not the raw context information itself. Computationally,
and metabolically, this is a much more efficient mecha-
nism, because the context is, by definition, unchanging
over the 100-msec alpha cycle, and thus, it makes more
sense to precompute the synaptic integration, rather than
repeatedly recomputing this same synaptic integration
over and over again (in the original feedforward Bp-based
SRN model, this issue did not arise because a single step of
activation updating took place for each context update—
whereas in our bidirectional model, many activation
update steps must take place per context update).
There are a couple of remaining challenges for this trans-
formation of the SRN. First, the precomputed net input from
the context must somehow persist over the subsequent
100-msec period of the alpha cycle. We hypothesize that this
can occur via N-methyl-D-aspartate and metabotropic gluta-
mate receptor channels that can easily produce sustained
excitatory currents over this time frame. Furthermore, the
reciprocal excitatory connectivity from 6CT to TRC and back
to 6CT could help to sustain the initial temporal context
signal. Second, these contextual integration synapses re-
quire a different form of learning algorithm that uses the
sending activation from the prior 100 msec, which is well
within the time constants in the relevant calcium and
second messenger pathways involved in synaptic plasticity.
BACKPROPAGATION MODEL METHODS
The Bp version of the WWI model has the same layer sizes
and feedforward patterns of connectivity as the DeepLeabra
version. Topographically, the V1p and V1hp pulvinar layers
serve as output layers at the highest level of the network,
receiving all the various connections from deep layers as
shown in Table 1. Likewise, the LIPp served as a target out-
put layer for the “where” pathway. To achieve predictive
learning, the V1 pulvinar targets were from the scene at
time t, whereas the V1s inputs were from the scene at time
t − 1. We also ran a comparison autoencoder model that
had inputs and target outputs from the same time step,
and it showed even less systematic organization of its
higher-level representations, further supporting the notion
that predictive learning is important, across all frameworks.
Figure 19. How the DeepLeabra temporal context computation
compares to the SRN mathematically. (A) In a standard SRN, the context
(deep layer biologically) is a copy of the hidden activations from the
prior time step, and these are held constant while the hidden layer
(superficial) units integrate the context through learned synaptic
weights. (B) In DeepLeabra, the deep layer performs the weighted
integration of the soon-to-be context information from the superficial
layer and then holds this integrated value and feeds it back as an
additive net-input-like signal to the superficial layer. The context net
input is precomputed, instead of having to compute this same value
over and over again. This is more efficient and more compatible with
the diffuse interconnections among the deep layer neurons. Layer 6
projections to the thalamus and back recirculate this precomputed net
input value into the superficial layers (via Layer 4) and back into itself to
support maintenance of the held value.
representation cannot be learned at the time when that
information is presently active. Instead, the relevant
contextual information is learned on the basis of what
happens next.
This explains the peculiar power of the otherwise strange
property of the SRN: The temporal context information is
preserved as a “direct copy” of the state of the hidden layer
units on the previous time step (Figure 19), and then
learned synaptic weights integrate that copied context
information into the next hidden state (which is then
copied to the context again, and so on). This enables the
error-driven learning taking place in the current time step
to determine how context information from the previous
time step is integrated. Furthermore, the simple direct
copy operation eschews any attempt to shape this tempo-
ral context itself, instead relying on the learning pressure
that shapes the hidden layer representations to also shape
the context representations. In other words, this copy
operation is essential, because there is no other viable
source of learning signals to shape the nature of the con-
text representation itself (because these learning signals
require future outcomes, which are by definition only
available later).
The direct copy operation of the SRN is however seem-
ingly problematic from a biological perspective: How could
neurons copy activations from another set of neurons at
some discrete point in time and then hold onto those cop-
ied values for a duration of 100 msec, which is a reasonably
1186
Journal of Cognitive Neuroscience
Volume 33, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
1
5
8
1
9
1
3
5
7
7
/
/
j
o
c
n
_
a
_
0
1
7
0
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
(5)
(6)
(7)
(8)
prediction module (Âl) consists of another standard con-
volutional layer and rectified linear activation that is used
to generate predictions from the output of Rl. These pre-
dictions are then compared against the output of the input
convolutional module (Al). The errors generated in this
comparison are represented explicitly in El, which applies
a rectified linear activation to a concatenation of the pos-
itive (Al − Âl) and negative (Âl − Al) prediction errors.
These errors then become the inputs to the next layer.
¼
At
l
(cid:4)
xt
(cid:4)
(cid:4)
MaxPool ReLU Conv E t
l − 1
(cid:5)
(cid:5)
(cid:5)
;
if
if
l ¼ 0
l > 0
(cid:5)
(cid:4)
(cid:4) (cid:5)
− ReLU Conv Rt
l
(cid:5)
(cid:4)
(cid:6)
¼ ReLu At
l
− A^t
l
(cid:4)
A^t
l
E t
l
Rt
l
(cid:4)
; ReLU A^t
l
(cid:5)
(cid:7)
− At
l
¼ ConvLSTM E t
l
−1; Rt
l
−1; UpSample Rt
l þ 1
(cid:4)
(cid:5)
(cid:5)
At each time step in the video sequence, PredNet gener-
ates a prediction of the next frame. This is done as follows:
First, the Rl is computed for each layer starting from the
t depends on input
top of the hierarchy (because each Rl
t are computed in
from Rt
l , and El
a feedforward fashion (because each At
l depends on input
from the layer below, E t
lþ1), and then the At
l , A^t
l−1).
All analyses in the RSA were conducted using the repre-
sentations from the Rl layers.
Implementation Details
All experiments with the PredNet architecture were per-
formed using PyTorch. An informal hyperparameter
search was conducted to find the settings that maximized
representational similarity to the human judgments. This
was done by conducting RSA on each layer for each hyper-
parameter setting and computing, according to the cen-
troid categories derived from the human data, the
difference between the average within-category similarity
and the average between-category similarity. Our final
Figure 21. Learning curves for the PredNet model. This model
achieves the best overall prediction performance but also has the least
well-differentiated, categorical representations.
O’Reilly et al.
1187
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
1
5
8
1
9
1
3
5
7
7
/
/
j
o
c
n
_
a
_
0
1
7
0
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 20. Learning curves for the Bp version of the WWI model.
Although it achieves better predictive accuracy than the DeepLeabra
version, it fails to acquire abstract object category structure, indicating a
potential tradeoff between simplifying and categorizing inputs, versus
predicting precisely where the low-level visual features will move.
The learning curve for the predictive version is shown in
Figure 20, which shows better overall prediction accuracy
compared to the DeepLeabra model. However, as the
RSA showed, this Bp model failed to learn object categories
that go beyond the input similarity structure, indicating that
perhaps it was paying too much “attention” in learning to
this low-level structure, and lacked the necessary mecha-
nisms to enable it to impose a simplifying higher-level
structure on top of these inputs.
PREDNET MODEL METHODS
The PredNet architecture was designed to incorporate
principles from predictive coding theory into a neural
network model for predicting the next frame in a video
sequence. Details of the model can be found in the origi-
nal paper (Lotter et al., 2016), but here, we provide a brief
overview of the architecture.
Architecture
PredNet is a DCNN that is composed of layers containing
discrete modules. The lowest layer generates a prediction
of incoming inputs (i.e., the pixels in the next frame),
whereas each of the higher layers attempts to predict
the errors made by the previous layer. Each layer contains
an input convolutional module (Al), a recurrent represen-
tational module (Rl), a prediction module (Âl), and a rep-
resentation of its own errors (El). The input convolutional
module (Al) transforms its input with a set of standard con-
volutional filters, a rectified linear activation function, and
a max-pooling operation. The recurrent representation
module (Rl) is a convolutional LSTM, which is a recurrent
convolutional network that replaces the matrix multiplica-
tions in the standard long short-term memory (LSTM)
equations with convolutions, allowing it to maintain a spa-
tially organized representation of its inputs over time. The
architecture had six layers with 3, 16, 32, 64, 128, and 256
filters in the Al and Rl modules and 3 × 3 kernels through-
out the whole network. We also found that using sigmoid
and tanh activation functions in fully connected convolu-
tional LSTMs slightly improved performance, so these
were used for all experiments.
The weights in the PredNet model are trained using error
Bp. Predictions are generated, and errors are computed at
all levels of the hierarchy, but the model performs better
when only the lowest layer’s errors are backpropagated
(Lotter et al., 2016). We confirmed these results with exper-
iments that backpropagated the errors in higher layers, in
which performance (in terms of mean squared error) was
marginally reduced but the RSA results were similar. For this
reason, all reported experiments used a PredNet that was
trained by only backpropagating the lowest level error.
The model was trained using a batch size of 8 and an
Adam optimizer with a learning rate of 0.0001, with no
scheduler, for 150,000 batches. A training curve is shown
in Figure 21, showing that it achieves the best overall pre-
diction accuracy of any model we tested and yet does not
have representations that are as differentiated or categor-
ical as our biologically based model, as shown in the main
paper.
Regularization Experiments
As discussed in the main paper, our biologically based
model includes a number of important biologically moti-
vated properties that may be contributing to the develop-
ment of its categorical representations. These properties,
including excitatory bidirectional connections, inhibitory
competition, and an additional form of Hebbian learning,
may be acting as regularizers that encourage categorical
learning. We therefore tested whether standard regulari-
zation methods used in deep learning would have similar
effects on the representations developed in the PredNet
architecture. We tested (1) batch normalization, (2) drop-
out (0.1, 0.3, and 0.5), and (3) weight decay (0.01, 0.001,
0.0001, 0.00001). All experiments with batch normaliza-
tion and weight decay showed reduced performance (in
terms of both prediction error on the test set and within-
category correlation). As shown in Figure 22, dropout
marginally improved the within-category correlation
while also slightly improving prediction accuracy, so a
dropout rate of 0.1 was used for the comparison to
our biologically based model in the main paper.
Acknowledgments
We thank Dean Wyatte, Tom Hazy, Seth Herd, Kai Krueger, Tim
Curran, David Sheinberg, Lew Harvey, Jessica Mollick, Will
Chapman, Helene Devillez, and the rest of the CCN Lab for many
helpful comments and suggestions. This work was supported by
ONR grants ONR N00014-19-1-2684/N00014-18-1-2116, N00014-
14-1-0670/N00014-16-1-2128, N00014-18-C-2067, N00014-13-1-
0067, and D00014-12-C-0638.
This work utilized the Janus supercomputer, which is supported
by the National Science Foundation (award number CNS-
0821794) and the University of Colorado Boulder. The Janus su-
percomputer is a joint effort of the University of Colorado
Boulder, the University of Colorado Denver, and the National
Center for Atmospheric Research. All data and materials will be
available at github.com/ccnlab/deep-obj-cat upon publication.
Reprint requests should be sent to Randall C. O’Reilly, Department
of Psychology, Computer Science, and Center for Neuroscience,
University of California Davis, 1544 Newton Ct, Davis, CA 95618,
or via e-mail: oreilly@ucdavis.edu.
Author Contributions
Randall C. O’Reilly: Conceptualization; Formal analysis;
Funding acquisition; Investigation; Methodology; Project
administration; Software; Supervision; Validation;
Visualization; Writing – Original draft. Jacob L. Russin:
Formal analysis; Investigation; Methodology; Validation;
Writing – Review & editing. Maryam Zolfaghar:
Investigation; Methodology; Validation; Writing – Review
& editing. John Rohrlich: Conceptualization; Data curation;
Investigation; Methodology; Software; Validation; Writing –
Review & editing.
Funding Information
Randall C. O’Reilly: Office of Naval Research (http://dx.doi
.org/10.13039/100000006), grants D00014-12-C-0638,
N00014-13-1-0067, N00014-14-1-0670, N00014-18-C-2067,
and N00014-19-1-2684.
Diversity in Citation Practices
Figure 22. Effect of dropout in PredNet on RSA, as measured by the
difference between the average within-category correlation and the
average between-category correlation (using the centroid categories
derived from human data). Dropout marginally improves the category
structure learned in PredNet.
A retrospective analysis of the citations in every article pub-
lished in this journal from 2010 to 2020 has revealed a
persistent pattern of gender imbalance: Although the pro-
portions of authorship teams (categorized by estimated
1188
Journal of Cognitive Neuroscience
Volume 33, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
1
5
8
1
9
1
3
5
7
7
/
/
j
o
c
n
_
a
_
0
1
7
0
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
gender identification of first author/last author) publishing
in the Journal of Cognitive Neuroscience ( JoCN) during
this period were M(an)/M = .408, W(oman)/M = .335,
M/W = .108, and W/W = .149, the comparable proportions
for the articles that these authorship teams cited were
M/M = .579, W/M = .243, M/W = .102, and W/W = .076
(Fulvio et al., JoCN, 33:1, pp. 3–7). Consequently, JoCN
encourages all authors to consider gender balance explicitly
when selecting which articles to cite and gives them the
opportunity to report their article’s gender citation balance.
REFERENCES
Abbott, L. F., Varela, J. A., Sen, K., & Nelson, S. B. (1997).
Synaptic depression and cortical gain control. Science, 275,
220. DOI: https://doi.org/10.1126/science.275.5297.221,
PMID: 8985017
Ackley, D. H., Hinton, G. E., & Sejnowski, T. J. (1985). A learning
algorithm for Boltzmann machines. Cognitive Science, 9,
147–169. DOI: https://doi.org/10.1207/s15516709cog0901_7
Antonov, P. A., Chakravarthi, R., & Andersen, S. K. (2020). Too
little, too late, and in the wrong place: Alpha band activity
does not reflect an active mechanism of selective attention.
Neuroimage, 219, 117006. DOI: https://doi.org/10.1016
/j.neuroimage.2020.117006, PMID: 32485307
Arcaro, M. J., Pinsk, M. A., & Kastner, S. (2015). The anatomical
and functional organization of the human visual pulvinar.
Journal of Neuroscience, 35, 9848–9871. DOI: https://doi
.org/10.1523/JNEUROSCI.1575-14.2015
Ashby, F. G., & Maddox, W. T. (2011). Human Category
Learning 2.0. Annals of the New York Academy of Sciences,
1224, 147–161. DOI: https://doi.org/10.1111/j.1749-6632
.2010.05874.x, PMID: 21182535, PMCID: PMC3076539
Barczak, A., O’Connell, M. N., McGinnis, T., Ross, D., Mowery, T.,
Falchier, A., et al. (2018). Top–down, contextual entrainment of
neuronal oscillations in the auditory thalamocortical circuit.
Proceedings of the National Academy of Sciences, U.S.A., 115,
E7605–E7614. DOI: https://doi.org/10.1073/pnas.1714684115,
PMID: 30037997, PMCID: PMC6094129
Bastos, A. M., Usrey, W. M., Adams, R. A., Mangun, G. R., Fries, P., &
Friston, K. J. (2012). Canonical microcircuits for predictive
coding. Neuron, 76, 695–711. DOI: https://doi.org/10.1016
/j.neuron.2012.10.038, PMID: 23177956, PMCID: PMC3777738
Bastos, A. M., Vezoli, J., Bosman, C. A., Schoffelen, J.-M.,
Oostenveld, R., Dowdall, J. R., et al. (2015). Visual areas exert
feedforward and feedback influences through distinct
frequency channels. Neuron, 85, 390–401. DOI: https://doi
.org/10.1016/j.neuron.2014.12.018, PMID: 25556836
Bednar, J. A. (2012). Building a mechanistic model of the
development and function of the primary visual cortex.
Journal of Physiology, Paris, 106, 194–211. DOI: https://doi
.org/10.1016/j.jphysparis.2011.12.001, PMID: 22343520
Bender, D. B. (1982). Receptive-field properties of neurons in
the macaque inferior pulvinar. Journal of Neurophysiology,
48, 1–17. DOI: https://doi.org/10.1152/jn.1982.48.1.1, PMID:
7119838
Bender, D. B., & Youakim, M. (2001). Effect of attentive fixation
in macaque thalamus and cortex. Journal of Neurophysiology,
85, 219–234. DOI: https://doi.org/10.1152/jn.2001.85.1.219,
PMID: 11152722
Bengio, Y., Mesnard, T., Fischer, A., Zhang, S., & Wu, Y. (2017).
STDP-compatible approximation of backpropagation in an
energy-based model. Neural Computation, 29, 555–577.
DOI: https://doi.org/10.1162/NECO_a_00934, PMID: 28095200
Bengio, Y., Yao, L., Alain, G., & Vincent, P. (2013). Generalized
denoising auto-encoders as generative models. In C. J. C.
Burges, L. Bottou, M. Welling, Z. Ghahramani, & K. Q.
Weinberger (Eds.), Advances in neural information
processing systems 26 (pp. 899–907). Curran Associates, Inc.
http://papers.nips.cc/paper/5023-generalized-denoising-auto
-encoders-as-generative-models.pdf
Berger, H. (1929). Über das Elektrenkephalogramm des
Menschen. Archiv für Psychiatrie und Nervenkrankheiten,
87, 527–570. DOI: https://doi.org/10.1007/BF01797193
Bienenstock, E. L., Cooper, L. N., & Munro, P. W. (1982). Theory
for the development of neuron selectivity: Orientation
specificity and binocular interaction in visual cortex. Journal
of Neuroscience, 2, 32–48. DOI: https://doi.org/10.1523
/JNEUROSCI.02-01-00032.1982, PMID: 7054394, PMCID:
PMC6564292
Bjork, R. A. (1994). Memory and metamemory considerations
in the training of human beings. In J. Metcalfe & A. P.
Shimamura (Eds.), Metacognition: Knowing about knowing
(pp. 185–205). Cambridge, MA: MIT Press.
Bortone, D. S., Olsen, S. R., & Scanziani, M. (2014). Translaminar
inhibitory cells recruited by layer 6 corticothalamic neurons
suppress visual cortex. Neuron, 82, 474–485. DOI: https://doi
.org/10.1016/j.neuron.2014.02.021, PMID: 24656931, PMCID:
PMC4068343
Bourne, J. A., & Rosa, M. G. P. (2006). Hierarchical development
of the primate visual cortex, as revealed by neurofilament
immunoreactivity: Early maturation of the middle temporal
area (MT). Cerebral Cortex, 16, 405–414. DOI: https://doi
.org/10.1093/cercor/bhi119, PMID: 15944371
Brette, R., & Gerstner, W. (2005). Adaptive exponential
integrate-and-fire model as an effective description of
neuronal activity. Journal of Neurophysiology, 94,
3637–3642. DOI: https://doi.org/10.1152/jn.00686.2005,
PMID: 16014787
Bridge, H., Leopold, D. A., & Bourne, J. A. (2016). Adaptive
pulvinar circuitry supports visual cognition. Trends in
Cognitive Sciences, 20, 146–157. DOI: https://doi.org/10
.1016/j.tics.2015.10.003, PMID: 26553222, PMCID:
PMC4724498
Buffalo, E. A., Fries, P., Landman, R., Buschman, T. J., &
Desimone, R. (2011). Laminar differences in gamma and
alpha coherence in the ventral stream. Proceedings of the
National Academy of Sciences, U.S.A., 108, 11262–11267.
DOI: https://doi.org/10.1073/pnas.1011284108, PMID:
21690410, PMCID: PMC3131344
Busch, N. A., Dubois, J., & VanRullen, R. (2009). The phase of
ongoing EEG oscillations predicts visual perception. Journal
of Neuroscience, 29, 7869–7876. DOI: https://doi.org/10
.1523/JNEUROSCI.0113-09.2009, PMID: 19535598, PMCID:
PMC6665641
Buzsáki, G. (2005). Theta rhythm of navigation: Link between
path integration and landmark navigation, episodic and
semantic memory. Hippocampus, 15, 827–840. DOI: https://
doi.org/10.1002/hipo.20113, PMID: 16149082
Cadieu, C. F., Hong, H., Yamins, D. L. K., Pinto, N., Ardila, D.,
Solomon, E. A., et al. (2014). Deep neural networks rival the
representation of primate IT cortex for core visual object
recognition. PLoS Computational Biology, 10, e1003963.
DOI: https://doi.org/10.1371/journal.pcbi.1003963, PMID:
25521294, PMCID: PMC4270441
Cavanagh, P., Hunt, A. R., Afraz, A., & Rolfs, M. (2010). Visual
stability based on remapping of attention pointers. Trends
in Cognitive Sciences, 14, 147–153. DOI: https://doi.org
/10.1016/j.tics.2010.01.007, PMID: 20189870, PMCID:
PMC2847621
Chaudhuri, R., Knoblauch, K., Gariel, M.-A., Kennedy, H., &
Wang, X.-J. (2015). A large-scale circuit mechanism for
O’Reilly et al.
1189
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
1
5
8
1
9
1
3
5
7
7
/
/
j
o
c
n
_
a
_
0
1
7
0
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
hierarchical dynamical processing in the primate cortex.
Neuron, 88, 419–431. DOI: https://doi.org/10.1016/j.neuron
.2015.09.008, PMID: 26439530, PMCID: PMC4630024
Clark, A. (2013). Whatever next? Predictive brains, situated
agents, and the future of cognitive science. Behavioral and
Brain Sciences, 36, 181–204. DOI: https://doi.org/10.1017
/S0140525X12000477, PMID: 23663408
Clayton, M. S., Yeung, N., & Kadosh, R. C. (2018). The many
characters of visual alpha oscillations. European Journal of
Neuroscience, 48, 2498–2508. DOI: https://doi.org/10.1111
/ejn.13747, PMID: 29044823
Cleeremans, A., & McClelland, J. L. (1991). Learning the
structure of event sequences. Journal of Experimental
Psychology: General, 120, 235–253. DOI: https://doi.org/10
.1037/0096-3445.120.3.235, PMID: 1836490
Colby, C. L., Duhamel, J. R., & Goldberg, M. E. (1997). Visual,
presaccadic, and cognitive activation of single neurons in
monkey lateral intraparietal area. Journal of Neurophysiology,
76, 2841–2852. DOI: https://doi.org/10.1152/jn.1996.76.5.2841
.2841, PMID: 8930237
Connors, B. W., Gutnick, M. J., & Prince, D. A. (1982).
Electrophysiological properties of neocortical neurons in
vitro. Journal of Neurophysiology, 48, 1302–1320. DOI:
https://doi.org/10.1152/jn.1982.48.6.1302, PMID: 6296328
Cooper, L. N., & Bear, M. F. (2012). The BCM theory of synapse
modification at 30: Interaction of theory with experiment.
Nature Reviews Neuroscience, 13, 798–810. DOI: https://doi
.org/10.1038/nrn3353, PMID: 23080416
Crick, F. (1984). Function of the thalamic reticular complex:
The searchlight hypothesis. Proceedings of the National
Academy of Sciences, US.A., 81, 4586–4590. DOI: https://doi
.org/10.1073/pnas.81.14.4586, PMID: 6589612, PMCID:
PMC345636
Crick, F. (1989). The recent excitement about neural networks.
Nature, 337, 129–132. DOI: https://doi.org/10.1038/337129a0,
PMID: 2911347
Dayan, P. (1993). Improving generalization for temporal
difference learning: The successor representation. Neural
Computation, 5, 613–624. DOI: https://doi.org/10.1162/neco
.1993.5.4.613
Dayan, P., Hinton, G. E., Neal, R. N., & Zemel, R. S. (1995).
The Helmholtz machine. Neural Computation, 7, 889–904.
DOI: https://doi.org/10.1162/neco.1995.7.5.889, PMID:
7584891
de Lange, F. P., Heilbron, M., & Kok, P. (2018). How do
expectations shape perception? Trends in Cognitive
Sciences, 22, 764–779. DOI: https://doi.org/10.1016/j.tics
.2018.06.002, PMID: 30122170
Desimone, R., & Duncan, J. (1995). Neural mechanisms of
selective visual attention. Annual Review of Neuroscience,
18, 193–222. DOI: https://doi.org/10.1146/annurev.ne.18
.030195.001205, PMID: 7605061
Duhamel, J. R., Colby, C. L., & Goldberg, M. E. (1992). The
updating of the representation of visual space in parietal
cortex by intended eye movements. Science, 255, 90–92.
DOI: https://doi.org/10.1126/science.1553535, PMID:
1553535
Elman, J., Bates, E., Karmiloff-Smith, A., Johnson, M., Parisi, D., &
Plunkett, K. (1996). Rethinking innateness: A connectionist
perspective on development. Cambridge, MA: MIT Press.
DOI: https://doi.org/10.7551/mitpress/5929.001.0001
Elman, J. L. (1990). Finding structure in time. Cognitive Science,
14, 179–211. DOI: https://doi.org/10.1016/0364-0213(90)
90002-E
Felleman, D. J., & Van Essen, D. C. (1991). Distributed hierarchical
processing in the primate cerebral cortex. Cerebral Cortex,
1, 1–47. DOI: https://doi.org/10.1093/cercor/1.1.1, PMID:
1822724
Fiebelkorn, I. C., & Kastner, S. (2019). A rhythmic theory of
attention. Trends in Cognitive Sciences, 23, 87–101. DOI:
https://doi.org/10.1016/j.tics.2018.11.009, PMID: 30591373,
PMCID: PMC6343831
Fiebelkorn, I. C., Pinsk, M. A., & Kastner, S. (2018). A dynamic
interplay within the frontoparietal network underlies
rhythmic spatial attention. Neuron, 99, 842–853. DOI:
https://doi.org/10.1016/j.neuron.2018.07.038, PMID: 30138590,
PMCID: PMC6474777
Fiser, A., Mahringer, D., Oyibo, H. K., Petersen, A. V., Leinweber,
M., & Keller, G. B. (2016). Experience-dependent spatial
expectations in mouse visual cortex. Nature Neuroscience,
19, 1658–1664. DOI: https://doi.org/10.1038/nn.4385, PMID:
27618309
Foldiak, P. (1991). Learning invariance from transformation
sequences. Neural Computation, 3, 194–200. DOI: https://
doi.org/10.1162/neco.1991.3.2.194, PMID: 31167302
Foster, J. J., & Awh, E. (2019). The role of alpha oscillations in
spatial attention: Limited evidence for a suppression account.
Current Opinion in Psychology, 29, 34–40. DOI: https://doi
.org/10.1016/j.copsyc.2018.11.001, PMID: 30472541, PMCID:
PMC6506396
Franceschetti, S., Guatteo, E., Panzica, F., Sancini, G., Wanke, E.,
& Avanzini, G. (1995). Ionic mechanisms underlying burst
firing in pyramidal neurons: Intracellular study in rat
sensorimotor cortex. Brain Research, 696, 127–139. DOI:
https://doi.org/10.1016/0006-8993(95)00807-3, PMID:
8574660
Fries, P., Womelsdorf, T., Oostenveld, R., & Desimone, R.
(2008). The effects of visual stimulation and selective visual
attention on rhythmic neuronal synchronization in macaque
area V4. Journal of Neuroscience, 28, 4823–4835. DOI:
https://doi.org/10.1523/JNEUROSCI.4499-07.2008, PMID:
18448659, PMCID: PMC3844818
Friston, K. (2005). A theory of cortical responses. Philosophical
Transactions of the Royal Society of London, Series B,
Biological Sciences, 360, 815–836. DOI: https://doi.org/10
.1098/rstb.2005.1622, PMID: 15937014, PMCID: PMC1569488
Friston, K. (2010). The free-energy principle: A unified brain
theory? Nature Reviews Neuroscience, 11, 127–138. DOI:
https://doi.org/10.1038/nrn2787, PMID: 20068583
Fusi, S., Miller, E. K., & Rigotti, M. (2016). Why neurons mix:
High dimensionality for higher cognition. Current Opinion
in Neurobiology, 37, 66–74. DOI: https://doi.org/10.1016
/j.conb.2016.01.010, PMID: 26851755
Gardner, M. P. H., Schoenbaum, G., & Gershman, S. J. (2018).
Rethinking dopamine as generalized prediction error.
Proceedings of the Royal Society of London, Series B,
Biological Sciences, 285, 20181645. DOI: https://doi.org/10
.1098/rspb.2018.1645, PMID: 30464063, PMCID: PMC6253385
Gavornik, J. P., & Bear, M. F. (2014). Learned spatiotemporal
sequence recognition and prediction in primary visual cortex.
Nature Neuroscience, 17, 732–737. DOI: https://doi.org/10
.1038/nn.3683, PMID: 24657967, PMCID: PMC4167369
George, D., & Hawkins, J. (2009). Towards a mathematical
theory of cortical micro-circuits. PLoS Computational
Biology, 5, e1000532. DOI: https://doi.org/10.1371/journal
.pcbi.1000532, PMID: 19816557, PMCID: PMC2749218
Goodale, M. A., & Milner, A. D. (1992). Separate visual pathways
for perception and action. Trends in Neurosciences, 15,
20–25. DOI: https://doi.org/10.1016/0166-2236(92)90344-8,
PMID: 1374953
Gottlieb, J. P., Kusunoki, M., & Goldberg, M. E. (1998). The
representation of visual salience in monkey parietal cortex.
Nature, 391, 481–484. DOI: https://doi.org/10.1038/35135,
PMID: 9461214
Grill-Spector, K., Henson, R., & Martin, A. (2006). Repetition
and the brain: Neural models of stimulus-specific effects.
1190
Journal of Cognitive Neuroscience
Volume 33, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
1
5
8
1
9
1
3
5
7
7
/
/
j
o
c
n
_
a
_
0
1
7
0
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Trends in Cognitive Sciences, 10, 14–23. DOI: https://doi.org
/10.1016/j.tics.2005.11.006, PMID: 16321563
Grossberg, S. (1999). How does the cerebral cortex work?
Learning, attention, and grouping by the laminar circuits of
visual cortex. Spatial Vision, 12, 163–185. DOI: https://doi
.org/10.1163/156856899X00102, PMID: 10221426
Gruber, W. R., Klimesch, W., Sauseng, P., & Doppelmayr, M.
(2005). Alpha phase synchronization predicts P1 and N1
latency and amplitude size. Cerebral Cortex, 15, 371–377.
DOI: https://doi.org/10.1093/cercor/bhh139, PMID:
15749980
Gundlach, C., Moratti, S., Forschack, N., & Müller, M. M. (2020).
Spatial attentional selection modulates early visual stimulus
processing independently of visual alpha modulations.
Cerebral Cortex, 30, 3686–3703. DOI: https://doi.org/10
.1093/cercor/bhz335, PMID: 31907512
Halassa, M. M., & Kastner, S. (2017). Thalamic functions in
distributed cognitive control. Nature Neuroscience, 20,
1669. DOI: https://doi.org/10.1038/s41593-017-0020-1,
PMID: 29184210
Harris, K. D., & Shepherd, G. M. G. (2015). The neocortical
circuit: Themes and variations. Nature Neuroscience, 18,
170–181. DOI: https://doi.org/10.1038/nn.3917, PMID:
25622573, PMCID: PMC4889215
Hawkins, J., & Blakeslee, S. (2004). On intelligence. New York:
Times Books.
Hennig, M. H. (2013). Theoretical models of synaptic short term
plasticity. Frontiers in Computational Neuroscience, 7,
45. DOI: https://doi.org/10.3389/fncom.2013.00154, PMID:
24198783, PMCID: PMC3812535
Hinton, G. E., & McClelland, J. L. (1988). Learning representations
by recirculation. In D. Z. Anderson (Ed.), Neural information
processing systems (NIPS 1987) (pp. 358–366). New York:
American Institute of Physics. http://papers.nips.cc/paper/78
-learning-representations-by-recirculation.pdf
Hinton, G. E., & Salakhutdinov, R. R. (2006). Reducing the
dimensionality of data with neural networks. Science, 313,
504–507. DOI: https://doi.org/10.1126/science.1127647,
PMID: 16873662
Holroyd, C. B., & Coles, M. G. H. (2002). The neural basis of
human error processing: Reinforcement learning, dopamine,
and the error-related negativity. Psychological Review, 109,
679–709. DOI: https://doi.org/10.1037/0033-295X.109.4.679,
PMID: 12374324
Hopfield, J. J. (1984). Neurons with graded response have
collective computational properties like those of two-state
neurons. Proceedings of the National Academy of Sciences,
U.S.A., 81, 3088–3092. DOI: https://doi.org/10.1073/pnas.81
.10.3088, PMID: 6587342, PMCID: PMC345226
Issa, E. B., Cadieu, C. F., & DiCarlo, J. J. (2018). Neural dynamics
at successive stages of the ventral visual stream are consistent
with hierarchical error signals. eLife, 7, e42870. DOI:
https://doi.org/10.7554/eLife.42870, PMID: 30484773,
PMCID: PMC6296785
Jaegle, A., & Ro, T. (2013). Direct control of visual perception
with phase-specific modulation of posterior parietal cortex.
Journal of Cognitive Neuroscience, 26, 422–432. DOI:
https://doi.org/10.1162/jocn_a_00494, PMID: 24116843
Jaramillo, J., Mejias, J. F., & Wang, X.-J. (2019). Engagement
of pulvino-cortical feedforward and feedback pathways
in cognitive computations. Neuron, 101, 321–336.
DOI: https://doi.org/10.1016/j.neuron.2018.11.023, PMID:
30553546, PMCID: PMC6650151
Jensen, O., Bonnefond, M., Marshall, T. R., & Tiesinga, P.
(2015). Oscillatory mechanisms of feedforward and feedback
visual processing. Trends in Neurosciences, 38, 192–194.
DOI: https://doi.org/10.1016/j.tins.2015.02.006, PMID:
25765320
Jensen, O., Bonnefond, M., & VanRullen, R. (2012). An
oscillatory mechanism for prioritizing salient unattended
stimuli. Trends in Cognitive Sciences, 16, 200–206. DOI:
https://doi.org/10.1016/j.tics.2012.03.002, PMID: 22436764
Jensen, O., & Mazaheri, A. (2010). Shaping functional
architecture by oscillatory alpha activity: Gating by inhibition.
Frontiers in Human Neuroscience, 4, 186. DOI: https://
doi.org/10.3389/fnhum.2010.00186, PMID: 21119777,
PMCID: PMC2990626
Jordan, M. I. (1989). Serial order: A parallel, distributed
processing approach. In J. L. Elman & D. E. Rumelhart (Eds.),
Advances in connectionist theory: Speech. Hillsdale, NJ:
Lawrence Erlbaum Associates.
Kachergis, G., Wyatte, D., O’Reilly, R. C., de Kleijn, R., &
Hommel, B. (2014). A continuous-time neural model for
sequential action. Philosophical Transactions of the Royal
Society of London, Series B, Biological Sciences, 369,
20130623. DOI: https://doi.org/10.1098/rstb.2013.0623,
PMID: 25267830, PMCID: PMC4186241
Kahana, M. J., Seelig, D., & Madsen, J. R. (2001). Theta returns.
Current Opinion in Neurobiology, 11, 739–744. DOI: https://
doi.org/10.1016/S0959-4388(01)00278-1, PMID: 11741027
Kawato, M., Hayakawa, H., & Inui, T. (1993). A forward-inverse
optics model of reciprocal connections between visual
cortical areas. Network: Computation in Neural Systems, 4,
415–422. DOI: https://doi.org/10.1088/0954-898X_4_4_001
Keitel, C., Keitel, A., Benwell, C. S. Y., Daube, C., Thut, G., &
Gross, J. (2019). Stimulus-driven brain rhythms within the
alpha band: The attentional-modulation conundrum. Journal
of Neuroscience, 39, 3119–3129. DOI: https://doi.org/10
.1523/JNEUROSCI.1633-18.2019, PMID: 30770401, PMCID:
PMC6468105
Kelly, S. P., Lalor, E. C., Reilly, R. B., & Foxe, J. J. (2006).
Increases in alpha oscillatory power reflect an active
retinotopic mechanism for distracter suppression during
sustained visuospatial attention. Journal of Neurophysiology,
95, 3844–3851. DOI: https://doi.org/10.1152/jn.01234.2005,
PMID: 16571739
Khaligh-Razavi, S.-M., & Kriegeskorte, N. (2014). Deep supervised,
but not unsupervised, models may explain IT cortical
representation. PLOS Computational Biology, 10, e1003915.
DOI: https://doi.org/10.1371/journal.pcbi.1003915, PMID:
25375136, PMCID: PMC4222664
Kiorpes, L., Price, T., Hall-Haro, C., & Movshon, J. A. (2012).
Development of sensitivity to global form and motion in
macaque monkeys (Macaca nemestrina). Vision Research,
63, 34–42. DOI: https://doi.org/10.1016/j.visres.2012.04.018,
PMID: 22580018, PMCID: PMC3374036
Klimesch, W. (2011). Evoked alpha and early access to the
knowledge system: The P1 inhibition timing hypothesis.
Brain Research, 1408, 52–71. DOI: https://doi.org/10.1016
/j.brainres.2011.06.003, PMID: 21774917, PMCID:
PMC3158852
Klimesch, W., Sauseng, P., & Hanslmayr, S. (2007). EEG alpha
oscillations: The inhibition-timing hypothesis. Brain Research
Reviews, 53, 63–88. DOI: https://doi.org/10.1016/j.brainresrev
.2006.06.003, PMID: 16887192
Kobatake, E., & Tanaka, K. (1994). Neuronal selectivities to
complex object features in the ventral visual pathway. Journal
of Neurophysiology, 71, 856–867. DOI: https://doi.org/10
.1152/jn.1994.71.3.856, PMID: 8201425
Kogo, N., & Trengove, C. (2015). Is predictive coding theory
articulated enough to be testable? Frontiers in Computational
Neuroscience, 9, 111. DOI: https://doi.org/10.3389/fncom
.2015.00111, PMID: 26441621, PMCID: PMC4561670
Kok, P., & de Lange, F. P. (2015). Predictive coding in sensory
cortex. In B. U. Forstmann & E.-J. Wagenmakers (Eds.),
An introduction to model-based cognitive neuroscience
O’Reilly et al.
1191
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
1
5
8
1
9
1
3
5
7
7
/
/
j
o
c
n
_
a
_
0
1
7
0
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
(pp. 221–244). New York: Springer. DOI: https://doi.org/10
.1007/978-1-4939-2236-9
Kok, P., Jehee, J. F. M., & de Lange, F. P. (2012). Less is more:
Expectation sharpens representations in the primary visual
cortex. Neuron, 75, 265–270. DOI: https://doi.org/10.1016
/j.neuron.2012.04.034, PMID: 22841311
Komura, Y., Nikkuni, A., Hirashima, N., Uetake, T., & Miyamoto, A.
(2013). Responses of pulvinar neurons reflect a subject’s
confidence in visual categorization. Nature Neuroscience,
16, 749–755. DOI: https://doi.org/10.1038/nn.3393, PMID:
23666179
Kriegeskorte, N., Mur, M., & Bandettini, P. (2008). Representational
similarity analysis—Connecting the branches of systems
neuroscience. Frontiers in Systems Neuroscience, 2, 4.
DOI: https://doi.org/10.3389/neuro.06.004.2008, PMID:
19104670, PMCID: PMC2605405
LaBerge, D., & Buchsbaum, M. S. (1990). Positron emission
tomographic measurements of pulvinar activity during an
attention task. Journal of Neuroscience, 10, 613–619. DOI:
https://doi.org/10.1523/JNEUROSCI.10-02-00613.1990,
PMID: 2303863, PMCID: PMC6570168
Larkum, M. E., Zhu, J. J., & Sakmann, B. (1999). A new cellular
mechanism for coupling inputs arriving at different cortical
layers. Nature, 398, 338–341. DOI: https://doi.org/10.1038
/18686, PMID: 10192334
LeCun, Y., Bengio, Y., & Hinton, G. (2015). Deep learning. Nature,
521, 436–444. DOI: https://doi.org/10.1038/nature14539,
PMID: 26017442
Lee, T. S., & Mumford, D. (2003). Hierarchical Bayesian
inference in the visual cortex. Journal of the Optical Society
of America, 20, 1434–1448. DOI: https://doi.org/10.1364
/JOSAA.20.001434, PMID: 12868647
Lillicrap, T. P., Santoro, A., Marris, L., Akerman, C. J., & Hinton,
G. (2020). Backpropagation and the brain. Nature Reviews
Neuroscience, 21, 335–346. DOI: https://doi.org/10.1038
/s41583-020-0277-3, PMID: 32303713
Lim, S., McKee, J. L., Woloszyn, L., Amit, Y., Freedman, D. J.,
Sheinberg, D. L., et al. (2015). Inferring learning rules from
distributions of firing rates in cortical neurons. Nature
Neuroscience, 18, 1804–1810. DOI: https://doi.org/10.1038
/nn.4158, PMID: 26523643, PMCID: PMC4666720
Lotter, W., Kreiman, G., & Cox, D. (2016). Deep predictive coding
networks for video prediction and unsupervised learning.
arXiv:1605.08104 [cs, q-bio]. http://arxiv.org/abs/1605.08104
Luczak, A., Bartho, P., & Harris, K. D. (2009). Spontaneous
events outline the realm of possible sensory responses in
neocortical populations. Neuron, 62, 413–425. DOI: https://
doi.org/10.1016/j.neuron.2009.03.014, PMID: 19447096,
PMCID: PMC2696272
Luczak, A., Bartho, P., & Harris, K. D. (2013). Gating of sensory
input by spontaneous cortical activity. Journal of Neuroscience,
33, 1684–1695. DOI: https://doi.org/10.1523/JNEUROSCI.2928
-12.2013, PMID: 23345241, PMCID: PMC3672963
Lüscher, C., & Malenka, R. C. (2012). NMDA receptor-dependent
long-term potentiation and long-term depression (LTP/LTD).
Cold Spring Harbor Perspectives in Biology, 4, a005710.
DOI: https://doi.org/10.1101/cshperspect.a005710, PMID:
22510460, PMCID: PMC3367554
Maier, A., Adams, G. K., Aura, C., & Leopold, D. A. (2010).
Distinct superficial and deep laminar domains of activity in
the visual cortex during rest and stimulation. Frontiers in
Systems Neuroscience, 4, 31. DOI: https://doi.org/10.3389
/fnsys.2010.00031, PMID: 20802856, PMCID: PMC2928665
Maier, A., Aura, C. J., & Leopold, D. A. (2011). Infragranular
sources of sustained local field potential responses in
macaque primary visual cortex. Journal of Neuroscience, 31,
1971–1980. DOI: https://doi.org/10.1523/JNEUROSCI.5300
-09.2011, PMID: 21307235, PMCID: PMC3075009
Makeig, S., Westerfield, M., Jung, T. P., Enghoff, S., Townsend,
J., Courchesne, E., et al. (2002). Dynamic brain sources of
visual evoked responses. Science, 295, 690–693. DOI: https://
doi.org/10.1126/science.1066168, PMID: 11809976
Marino, A. C., & Mazer, J. A. (2016). Perisaccadic updating of
visual representations and attentional states: Linking behavior
and neurophysiology. Frontiers in Systems Neuroscience,
10, 3. DOI: https://doi.org/10.3389/fnsys.2016.00003, PMID:
26903820, PMCID: PMC4743436
Markov, N. T., Ercsey-Ravasz, M. M., Ribeiro Gomes, A. R., Lamy, C.,
Magrou, L., Vezoli, J., et al. (2014). A weighted and directed
interareal connectivity matrix for macaque cerebral cortex.
Cerebral Cortex, 24, 17–36. DOI: https://doi.org/10.1093/cercor
/bhs270, PMID: 23010748, PMCID: PMC3862262
Markov, N. T., Vezoli, J., Chameau, P., Falchier, A., Quilodran,
R., Huissoud, C., et al. (2014). Anatomy of hierarchy:
Feedforward and feedback pathways in macaque visual
cortex: Cortical counterstreams. Journal of Comparative
Neurology, 522, 225–259. DOI: https://doi.org/10.1002/cne
.23458, PMID: 23983048, PMCID: PMC4255240
Martinez-Conde, S., Macknik, S. L., & Hubel, D. H. (2004). The
role of fixational eye movements in visual perception. Nature
Reviews Neuroscience, 5, 229–240. DOI: https://doi.org/10
.1038/nrn1348, PMID: 14976522
Martinez-Conde, S., Otero-Millan, J., & Macknik, S. L. (2013).
The impact of microsaccades on vision: Towards a unified
theory of saccadic function. Nature Reviews Neuroscience,
14, 83–96. DOI: https://doi.org/10.1038/nrn3405, PMID:
23329159
Mathewson, K., Gratton, G., Fabiani, M., Beck, D., & Ro, T.
(2009). To see or not to see: Prestimulus alpha phase
predicts visual awareness. Journal of Neuroscience, 29,
2725–2732. DOI: https://doi.org/10.1523/JNEUROSCI.3963
-08.2009, PMID: 19261866, PMCID: PMC2724892
Mathewson, K. E., Fabiani, M., Gratton, G., Beck, D. M., &
Lleras, A. (2010). Rescuing stimuli from invisibility: Inducing
a momentary release from visual masking with pre-target
entrainment. Cognition, 115, 186–191. DOI: https://doi.org
/10.1016/j.cognition.2009.11.010, PMID: 20035933
Mathewson, K. E., Prudhomme, C., Fabiani, M., Beck, D. M.,
Lleras, A., & Gratton, G. (2012). Making waves in the stream
of consciousness: Entraining oscillations in EEG alpha and
fluctuations in visual awareness with rhythmic visual
stimulation. Journal of Cognitive Neuroscience, 24,
2321–2333. DOI: https://doi.org/10.1162/jocn_a_00288,
PMID: 22905825
Mayer, A., Schwiedrzik, C. M., Wibral, M., Singer, W., & Melloni,
L. (2016). Expecting to see a letter: Alpha oscillations as
carriers of top–down sensory predictions. Cerebral Cortex,
26, 3146–3160. DOI: https://doi.org/10.1093/cercor/bhv146,
PMID: 26142463
Meyer, T., & Olson, C. R. (2011). Statistical learning of visual
transitions in monkey inferotemporal cortex. Proceedings of
the National Academy of Sciences, U.S.A., 108, 19401–19406.
DOI: https://doi.org/10.1073/pnas.1112895108, PMID:
22084090, PMCID: PMC3228439
Michalareas, G., Vezoli, J., van Pelt, S., Schoffelen, J.-M.,
Kennedy, H., & Fries, P. (2016). Alpha–beta and gamma
rhythms subserve feedback and feedforward influences
among human visual cortical areas. Neuron, 89, 384–397.
DOI: https://doi.org/10.1016/j.neuron.2015.12.018, PMID:
26777277, PMCID: PMC4871751
Miller, E. K., & Cohen, J. D. (2001). An integrative theory of
prefrontal cortex function. Annual Review of Neuroscience,
24, 167–202. DOI: https://doi.org/10.1146/annurev.neuro.24
.1.167, PMID: 11283309
Miller, K. D. (1994). A model for the development of simple cell
receptive fields and the ordered arrangement of orientation
1192
Journal of Cognitive Neuroscience
Volume 33, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
1
5
8
1
9
1
3
5
7
7
/
/
j
o
c
n
_
a
_
0
1
7
0
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
columns through activity-dependent competition between
ON- and OFF-center inputs. Journal of Neuroscience, 14,
409–441. DOI: https://doi.org/10.1523/JNEUROSCI.14-01
-00409.1994, PMID: 8283248, PMCID: PMC6576834
Müller, J. R., Metha, A. B., Krauskopf, J., & Lennie, P. (1999).
Rapid adaptation in visual cortex to the structure of images.
Science, 285, 1405–1408. DOI: https://doi.org/10.1126
/science.285.5432.1405, PMID: 10464100
Mumford, D. (1991). On the computational architecture of the
neocortex. Biological Cybernetics, 65, 135–145. DOI: https://
doi.org/10.1007/BF00202389, PMID: 1912004
Mumford, D. (1992). On the computational architecture of the
neocortex. II. The role of cortico-cortical loops. Biological
Cybernetics, 66, 241–251. DOI: https://doi.org/10.1007
/BF00198477, PMID: 1540675
Nakamura, K., & Colby, C. L. (2002). Updating of the visual
representation in monkey striate and extrastriate cortex
during saccades. Proceedings of the National Academy
of Sciences, U.S.A., 99, 4026–4031. DOI: https://doi.org
/10.1073/pnas.052379899, PMID: 11904446, PMCID:
PMC122642
Neupane, S., Guitton, D., & Pack, C. C. (2016). Two distinct
types of remapping in primate cortical area V4. Nature
Communications, 7, 10402. DOI: https://doi.org/10.1038
/ncomms10402, PMID: 26832423, PMCID: PMC4740356
Neupane, S., Guitton, D., & Pack, C. C. (2017). Coherent alpha
oscillations link current and future receptive fields during
saccades. Proceedings of the National Academy of Sciences,
U.S.A., 114, E5979–E5985. DOI: https://doi.org/10.1073/pnas
.1701672114, PMID: 28673993, PMCID: PMC5530666
Neupane, S., Guitton, D., & Pack, C. C. (2020). Perisaccadic
remapping: What? how? why? Reviews in the Neurosciences,
31, 505–520. DOI: https://doi.org/10.1515/revneuro-2019
-0097, PMID: 32242834
Nunn, C. M. H., & Osselton, J. W. (1974). The influence of
the EEG alpha rhythm on the perception of visual stimuli.
Psychophysiology, 11, 294–303. DOI: https://doi.org/10.1111
/j.1469-8986.1974.tb00547.x, PMID: 4421317
O’Herron, P., & von der Heydt, R. (2013). Remapping of border
ownership in the visual cortex. Journal of Neuroscience, 33,
1964–1974. DOI: https://doi.org/10.1523/JNEUROSCI.2797
-12.2013, PMID: 23365235, PMCID: PMC4086328
Olsen, S., Bortone, D., Adesnik, H., & Scanziani, M. (2012). Gain
control by layer six in cortical circuits of vision. Nature, 483,
47–52. DOI: https://doi.org/10.1038/nature10835, PMID:
22367547, PMCID: PMC3636977
O’Reilly, R. C. (1996). Biologically plausible error-driven
learning using local activation differences: The generalized
recirculation algorithm. Neural Computation, 8, 895–938.
DOI: https://doi.org/10.1162/neco.1996.8.5.895
O’Reilly, R. C. (1998). Six principles for biologically-based
computational models of cortical cognition. Trends in
Cognitive Sciences, 2, 455–462. DOI: https://doi.org/10.1016
/S1364-6613(98)01241-8
O’Reilly, R. C., Hazy, T. E., & Herd, S. A. (2016). The Leabra
cognitive architecture: How to play 20 principles with nature
and win! In S. Chipman (Ed.), Oxford handbook of cognitive
science. Oxford, UK: Oxford University Press. http://www.
oxfordhandbooks.com/view/10.1093/oxfordhb/9780199842193
.001.0001/oxfordhb-9780199842193-e-8. DOI: https://doi.org
/10.1093/oxfordhb/9780199842193.013.8
O’Reilly, R. C., & Munakata, Y. (2000). Computational
explorations in cognitive neuroscience: Understanding the
mind by simulating the brain. Cambridge, MA: MIT Press.
DOI: https://doi.org/10.7551/mitpress/2014.001.0001
O’Reilly, R. C., Munakata, Y., Frank, M. J., Hazy, T. E., &
Contributors. (2012). Computational cognitive neuroscience
(1st ed.). Wiki Book. http://ccnbook.colorado.edu
O’Reilly, R. C., Wyatte, D., Herd, S., Mingus, B., & Jilk, D. J.
(2013). Recurrent processing during object recognition.
Frontiers in Psychology, 4, 124. DOI: https://doi.org/10.3389
/fpsyg.2013.00124, PMID: 23554596, PMCID: PMC3612699
O’Reilly, R. C., Wyatte, D., & Rohrlich, J. (2014). Learning
through time in the thalamocortical loops. arXiv:1407.3432
[q-bio]. http://arxiv.org/abs/1407.3432
O’Reilly, R. C., Wyatte, D. R., & Rohrlich, J. (2017). Deep predictive
learning: A comprehensive model of three visual streams.
arXiv:1709.04654 [q-bio]. http://arxiv.org/abs/1709.04654
Ouden, H. E. M., Kok, P., & Lange, F. P. (2012). How prediction
errors shape perception, attention, and motivation. Frontiers
in Psychology, 3, 548. DOI: https://doi.org/10.3389/fpsyg
.2012.00548, PMID: 23248610, PMCID: PMC3518876
Palva, S., & Palva, J. M. (2011). Functional roles of alpha-band
phase synchronization in local and large-scale cortical
networks. Frontiers in Psychology, 2, 204. DOI: https://doi
.org/10.3389/fpsyg.2011.00204, PMID: 21922012, PMCID:
PMC3166799
Pennartz, C. M., Dora, S., Muckli, L., & Lorteije, J. A. (2019).
Towards a unified view on pathways and functions of neural
recurrent processing. Trends in Neurosciences, 42, 589–603.
DOI: https://doi.org/10.1016/j.tins.2019.07.005, PMID:
31399289
Petersen, S. E., Robinson, D. L., & Keys, W. (1985). Pulvinar nuclei
of the behaving rhesus monkey: Visual responses and their
modulation. Journal of Neurophysiology, 54, 867–886. DOI:
https://doi.org/10.1152/jn.1985.54.4.867, PMID: 4067625
Petrof, I., Viaene, A. N., & Sherman, S. M. (2012). Two
populations of corticothalamic and interareal corticocortical
cells in the subgranular layers of the mouse primary sensory
cortices. Journal of Comparative Neurology, 520, 1678–1686.
DOI: https://doi.org/10.1002/cne.23006, PMID: 22120996,
PMCID: PMC3561675
Pinault, D. (2004). The thalamic reticular nucleus: Structure,
function and concept. Brain Research, 46, 1–31. DOI: https://
doi.org/10.1016/j.brainresrev.2004.04.008, PMID: 15297152
Pineda, F. J. (1987). Generalization of backpropagation to
recurrent neural networks. Physical Review Letters, 18,
2229–2232. DOI: https://doi.org/10.1103/PhysRevLett.59
.2229, PMID: 10035458
Pouget, A., & Sejnowski, T. J. (1997). Spatial transformations in
the parietal cortex using basis functions. Journal of Cognitive
Neuroscience, 9, 222–237. DOI: https://doi.org/10.1162/jocn
.1997.9.2.222, PMID: 23962013
Privman, E., Malach, R., & Yeshurun, Y. (2013). Modeling
the electrical field created by mass neural activity. Neural
Networks, 40, 44–51. DOI: https://doi.org/10.1016/j.neunet
.2013.01.004, PMID: 23391515
Purushothaman, G., Marion, R., Li, K., & Casagrande, V. A.
(2012). Gating and control of primary visual cortex by
pulvinar. Nature Neuroscience, 15, 905–912. DOI: https://
doi.org/10.1038/nn.3106, PMID: 22561455, PMCID:
PMC3430824
Pylyshyn, Z. (1989). The role of location indexes in spatial
perception: A sketch of the FINST spatial-index model.
Cognition, 32, 65–97. DOI: https://doi.org/10.1016/0010
-0277(89)90014-0
Rajalingham, R., Issa, E. B., Bashivan, P., Kar, K., Schmidt, K., &
DiCarlo, J. J. (2018). Large-scale, high-resolution comparison
of the core visual object recognition behavior of humans,
monkeys, and state-of-the-art deep artificial neural networks.
Journal of Neuroscience, 38, 7255–7269. DOI: https://doi
.org/10.1101/240614, PMID: 30006365, PMCID: PMC6096043
Rao, R. P., & Ballard, D. H. (1999). Predictive coding in the
visual cortex: A functional interpretation of some extra-
classical receptive-field effects. Nature Neuroscience, 2,
79–87. DOI: https://doi.org/10.1038/4580, PMID: 10195184
O’Reilly et al.
1193
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
1
5
8
1
9
1
3
5
7
7
/
/
j
o
c
n
_
a
_
0
1
7
0
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Ray, S., & Maunsell, J. H. R. (2011). Different origins of gamma
rhythm and high-gamma activity in macaque visual cortex.
PLoS Biology, 9, e1000610. DOI: https://doi.org/10.1371
/journal.pbio.1000610, PMID: 21532743, PMCID: PMC3075230
Reber, A. S. (1967). Implicit learning of artificial grammars.
Journal of Verbal Learning and Verbal Behavior, 6, 855–863.
DOI: https://doi.org/10.1016/S0022-5371(67)80149-X
Reynolds, J. H., Chelazzi, L., & Desimone, R. (1999).
Competitive mechanisms subserve attention in macaque
areas V2 and V4. Journal of Neuroscience, 19, 1736–1753.
DOI: https://doi.org/10.1523/JNEUROSCI.19-05-01736.1999,
PMID: 10024360, PMCID: PMC6782185
Reynolds, J. H., & Heeger, D. J. (2009). The normalization
model of attention. Neuron, 61, 168–185. DOI: https://
doi.org/10.1016/j.neuron.2009.01.002, PMID: 19186161,
PMCID: PMC2752446
Richter, D., & de Lange, F. P. (2019). Statistical learning attenuates
visual activity only for attended stimuli. eLife, 8, e47869. DOI:
https://doi.org/10.7554/eLife.47869, PMID: 31442202, PMCID:
PMC6731093
Robinson, D. L. (1993). Functional contributions of the primate
pulvinar. Progress in Brain Research, 95, 371–380. DOI:
https://doi.org/10.1016/s0079-6123(08)60382-9, PMID:
8493346
Rockland, K. S. (1996). Two types of corticopulvinar terminations:
Round (type 2) and elongate (type 1). Journal of Comparative
Neurology, 368, 57–87. DOI: https://doi.org/10.1002/(sici)1096
-9861(19960422)368:1%3C57::aid-cne5%3E3.0.co;2-j, PMID:
8725294
Rockland, K. S. (1998). Convergence and branching patterns of
round, type 2 corticopulvinar axons. Journal of Comparative
Neurology, 390, 515–536. DOI: https://doi.org/10.1002/(sici)
1096-9861(19980126)390:4%3C515::aid-cne5%3E3.0.co;2-3,
PMID: 9450533
Rockland, K. S., & Pandya, D. N. (1979). Laminar origins and
terminations of cortical connections of the occipital lobe in
the rhesus monkey. Brain Research, 179, 3–20. DOI: https://
doi.org/10.1016/0006-8993(79)90485-2, PMID: 116716
Rumelhart, D. E., Hinton, G. E., & Williams, R. J. (1986).
Learning representations by back-propagating errors. Nature,
323, 533–536. DOI: https://doi.org/10.1038/323533a0
Rumelhart, D. E., & McClelland, J. L. (1982). An interactive
activation model of context effects in letter perception:
Part 2. The contextual enhancement effect and some tests
and extensions of the model. Psychological Review, 89,
60–94. DOI: https://doi.org/10.1037/0033-295X.89.1.60,
PMID: 7058229
Saalmann, Y. B., & Kastner, S. (2011). Cognitive and perceptual
functions of the visual thalamus. Neuron, 71, 209–223.
DOI: https://doi.org/10.1016/j.neuron.2011.06.027, PMID:
21791281, PMCID: PMC3148184
Saalmann, Y. B., Pinsk, M. A., Wang, L., Li, X., & Kastner, S.
(2012). The pulvinar regulates information transmission
between cortical areas based on attention demands. Science,
337, 753–756. DOI: https://doi.org/10.1126/science.1223082,
PMID: 22879517, PMCID: PMC3714098
Sakata, S., & Harris, K. D. (2009). Laminar structure of
spontaneous and sensory-evoked population activity in
auditory cortex. Neuron, 64, 404–418. DOI: https://doi.org
/10.1016/j.neuron.2009.09.020, PMID: 19914188, PMCID:
PMC2778614
Sakata, S., & Harris, K. D. (2012). Laminar-dependent effects of
cortical state on auditory cortical spontaneous activity. Frontiers
in Neural Circuits, 6, 109. DOI: https://doi.org/10.3389/fncir
.2012.00109, PMID: 23267317, PMCID: PMC3527822
Samaha, J., Bauer, P., Cimaroli, S., & Postle, B. R. (2015). Top–
down control of the phase of alpha-band oscillations as a
mechanism for temporal prediction. Proceedings of the
National Academy of Sciences, U.S.A., 112, 8439–8444. DOI:
https://doi.org/10.1073/pnas.1503686112, PMID: 26100913,
PMCID: PMC4500260
Sherman, M. T., Kanai, R., Seth, A. K., & VanRullen, R. (2016).
Rhythmic influence of top–down perceptual priors in the
phase of prestimulus occipital alpha oscillations. Journal of
Cognitive Neuroscience, 28, 1318–1330. DOI: https://doi.org
/10.1162/jocn_a_00973, PMID: 27082046
Sherman, S. M. (2014). The function of metabotropic glutamate
receptors in thalamus and cortex. Neuroscientist, 20, 146–149.
DOI: https://doi.org/10.1177/1073858413478490, PMID:
23459618, PMCID: PMC4747429
Sherman, S. M., & Guillery, R. W. (2006). Exploring the thalamus
and its role in cortical function. Cambridge, MA: MIT Press.
http://www.scholarpedia.org/article/Thalamus
Sherman, S. M., & Guillery, R. W. (2011). Distinct functions for
direct and transthalamic corticocortical connections. Journal
of Neurophysiology, 106, 1068–1077. DOI: https://doi.org/10
.1152/jn.00429.2011, PMID: 21676936
Sherman, S. M., & Guillery, R. W. (2013). Functional connections
of cortical areas: A new view from the thalamus. Cambridge,
MA: MIT Press. DOI: https://doi.org/10.7551/mitpress
/9780262019309.001.0001
Shipp, S. (2003). The functional logic of cortico-pulvinar
connections. Philosophical Transactions of the Royal Society
of London, Series B, Biological Sciences, 358, 1605–1624.
DOI: https://doi.org/10.1098/rstb.2002.1213, PMID: 14561322,
PMCID: PMC1693262
Shouval, H. Z. S., Bear, M. F., & Cooper, L. N. (2002). A unified
model of NMDA receptor-dependent bidirectional synaptic
plasticity. Proceedings of the National Academy of Sciences,
U.S.A., 99, 10831–10836. DOI: https://doi.org/10.1073/pnas
.152343099, PMID: 12136127, PMCID: PMC125058
Shrager, J., & Johnson, M. H. (1996). Dynamic plasticity
influences the emergence of function in a simple cortical
array. Neural Networks, 9, 1119–1129. DOI: https://doi.org
/10.1016/0893-6080(96)00033-0, PMID: 12662587
Silva, L. R., Amitai, Y., & Connors, B. W. (1991). Intrinsic
oscillations of neocortex generated by layer 5 pyramidal
neurons. Science, 251, 432–435. DOI: https://doi.org/10.1126
/science.1824881, PMID: 1824881
Snow, J. C., Allen, H. A., Rafal, R. D., & Humphreys, G. W.
(2009). Impaired attentional selection following lesions to
human pulvinar: Evidence for homology between human and
monkey. Proceedings of the National Academy of Sciences,
U.S.A., 106, 4054–4059. DOI: https://doi.org/10.1073/pnas
.0810086106, PMID: 19237580, PMCID: PMC2656203
Solís-Vivanco, R., Jensen, O., & Bonnefond, M. (2018). Top–
down control of alpha phase adjustment in anticipation of
temporally predictable visual stimuli. Journal of Cognitive
Neuroscience, 30, 1157–1169. DOI: https://doi.org/10.1162
/jocn_a_01280, PMID: 29762100
Solomon, E. A., Kragel, J. E., Sperling, M. R., Sharan, A., Worrell,
G., Kucewicz, M., et al. (2017). Widespread theta synchrony
and high-frequency desynchronization underlies enhanced
cognition. Nature Communications, 8, 1704. DOI: https://
doi.org/10.1038/s41467-017-01763-2, PMID: 29167419,
PMCID: PMC5700170
Spaak, E., Bonnefond, M., Maier, A., Leopold, D. A., & Jensen,
O. (2012). Layer-specific entrainment of gamma-band neural
activity by the alpha rhythm in monkey visual cortex. Current
Biology, 22, 2313–2318. DOI: https://doi.org/10.1016/j.cub
.2012.10.020, PMID: 23159599, PMCID: PMC3528834
Spaak, E., de Lange, F. P., & Jensen, O. (2014). Local
entrainment of alpha oscillations by visual stimuli causes
cyclic modulation of perception. Journal of Neuroscience,
34, 3536–3544. DOI: https://doi.org/10.1523/JNEUROSCI
.4385-13.2014, PMID: 24599454, PMCID: PMC6608988
1194
Journal of Cognitive Neuroscience
Volume 33, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
1
5
8
1
9
1
3
5
7
7
/
/
j
o
c
n
_
a
_
0
1
7
0
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Spelke, E., Breinlinger, K., Macomber, J., & Jacobson, K. (1992).
Origins of knowledge. Psychological Review, 99, 605–632. DOI:
https://doi.org/10.1037/0033-295X.99.4.605, PMID: 1454901
Spratling, M. W. (2008). Reconciling predictive coding and
biased competition models of cortical function. Frontiers in
Computational Neuroscience, 2, 1–8. DOI: https://doi.org
/10.3389/neuro.10.004.2008, PMID: 18978957, PMCID:
PMC2576514
Summerfield, C., & de Lange, F. P. (2014). Expectation in
perceptual decision making: Neural and computational
mechanisms. Nature Reviews Neuroscience, 15, 745–756.
DOI: https://doi.org/10.1038/nrn3838, PMID: 25315388
Summerfield, C., & Egner, T. (2009). Expectation (and attention)
in visual cognition. Trends in Cognitive Sciences, 13, 403–409.
DOI: https://doi.org/10.1016/j.tics.2009.06.003, PMID:
19716752
Summerfield, C., Trittschuh, E. H., Monti, J. M., Mesulam, M. M.,
& Egner, T. (2008). Neural repetition suppression reflects
fulfilled perceptual expectations. Nature Neuroscience, 11,
1004–1006. DOI: https://doi.org/10.1038/nn.2163, PMID:
19160497, PMCID: PMC2747248
Sutton, R. S., & Barto, A. G. (1998). Reinforcement learning:
An introduction. Cambridge, MA: MIT Press. http://www.cs
.ualberta.ca/sutton/book/ebook/the-book.html, DOI: https://
doi.org/10.1109/TNN.1998.712192
Thomson, A. M. (2010). Neocortical layer 6: A review. Frontiers
in Neuroanatomy, 4, 13. DOI: https://doi.org/10.3389/fnana
.2010.00013, PMID: 20556241, PMCID: PMC2885865
Thomson, A. M., & Lamy, C. (2007). Functional maps of
neocortical local circuitry. Frontiers in Neuroscience, 1,
19–42. DOI: https://doi.org/10.3389/neuro.01.1.1.002.2007,
PMID: 18982117, PMCID: PMC2518047
Todorovic, A., van Ede, F., Maris, E., & de Lange, F. P. (2011).
Prior expectation mediates neural adaptation to repeated
sounds in the auditory cortex: An MEG study. Journal of
Neuroscience, 31, 9118–9123. DOI: https://doi.org/10.1523
/JNEUROSCI.1425-11.2011, PMID: 21697363, PMCID:
PMC6623501
Ungerleider, L. G., & Mishkin, M. (1982). Two cortical visual
systems. In D. J. Ingle, M. A. Goodale, & R. J. W. Mansfield
(Eds.), The analysis of visual behavior (pp. 549–586).
Cambridge, MA: MIT Press.
Urakubo, H., Honda, M., Froemke, R. C., & Kuroda, S. (2008).
Requirement of an allosteric kinetics of NMDA receptors for
spike timing-dependent plasticity. Journal of Neuroscience,
28, 3310–3323. DOI: https://doi.org/10.1523/JNEUROSCI
.0303-08.2008, PMID: 18367598, PMCID: PMC6670607
Usrey, W. M., & Sherman, S. M. (2018). Corticofugal circuits:
Communication lines from the cortex to the rest of the brain.
Journal of Comparative Neurology, 527, 640–650. DOI:
https://doi.org/10.1002/cne.24423, PMID: 29524229, PMCID:
PMC6131091
Valpola, H. (2015). From neural PCA to deep unsupervised
learning. In Advances in independent component analysis
and learning machines. (pp. 143–171). Cambridge, MA:
Academic Press.
van Kerkoerle, T., Self, M. W., Dagnino, B., Gariel-Mathis, M.-A.,
Poort, J., van der Togt, C., et al. (2014). Alpha and gamma
oscillations characterize feedback and feedforward processing
in monkey visual cortex. Proceedings of the National
Academy of Sciences, U.S.A., 111, 14332–14341. DOI: https://
doi.org/10.1073/pnas.1402773111, PMID: 25205811, PMCID:
PMC4210002
VanRullen, R. (2016). Perceptual cycles. Trends in Cognitive
Sciences, 20, 723–735. DOI: https://doi.org/10.1016/j.tics
.2016.07.006, PMID: 27567317
VanRullen, R., & Koch, C. (2003). Is perception discrete or
continuous? Trends in Cognitive Sciences, 7, 207–213. DOI:
https://doi.org/10.1016/S0042-6989(02)00298-5, PMID:
12757822
VanRullen, R., & Thorpe, S. J. (2002). Surfing a spike wave down
the ventral stream. Vision Research, 42, 2593–2615. DOI:
https://doi.org/10.1016/s0042-6989(02)00298-5, PMID:
12446033
Varela, F. J., Toro, A., John, E. R., & Schwartz, E. L. (1981).
Perceptual framing and cortical alpha rhythm. Neuropsychologia,
19, 675–686. DOI: https://doi.org/10.1016/0028-3932(81)90005-1,
PMID: 7312152
Vinken, K., & Vogels, R. (2017). Adaptation can explain
evidence for encoding of probabilistic information in
macaque inferior temporal cortex. Current Biology, 27,
R1210–R1212. DOI: https://doi.org/10.1016/j.cub.2017.09
.018, PMID: 29161556
von Stein, A., Chiang, C., & König, P. (2000). Top–down processing
mediated by interareal synchronization. Proceedings of the
National Academy of Sciences, U.S.A., 97, 14748–14753. DOI:
https://doi.org/10.1073/pnas.97.26.14748, PMID: 11121074,
PMCID: PMC18990
von Helmholtz, H. (1867/2013). Treatise on physiological
optics (Vol. 3). North Chelmsford, MA: Courier Corporation.
Waldert, S., Lemon, R. N., & Kraskov, A. (2013). Influence of
spiking activity on cortical local field potentials. Journal
of Physiology, 591, 5291–5303. DOI: https://doi.org/10.1113
/jphysiol.2013.258228, PMID: 23981719, PMCID: PMC3936368
Walsh, K. S., McGovern, D. P., Clark, A., & O’Connell, R. G.
(2020). Evaluating the neurophysiological evidence for
predictive processing as a model of perception. Annals of the
New York Academy of Sciences, 1464, 242–268. DOI: https://
doi.org/10.1111/nyas.14321, PMID: 32147856, PMCID:
PMC7187369
Walter, W. G. (1953). The living brain. Oxford, UK: W. W.
Norton.
Watanabe, T., & Sasaki, Y. (2015). Perceptual learning: Toward
a comprehensive theory. Annual Review of Psychology, 66,
197–221. DOI: https://doi.org/10.1146/annurev-psych-010814
-015214, PMID: 25251494, PMCID: PMC4286445
Whittington, J. C. R., & Bogacz, R. (2019). Theories of error
back-propagation in the brain. Trends in Cognitive Sciences,
23, 235–250. DOI: https://doi.org/10.1016/j.tics.2018.12.005,
PMID: 30704969, PMCID: PMC6382460
Williams, R. J., & Zipser, D. (1992). Gradient-based learning
algorithms for recurrent networks and their computational
complexity. In Y. Chauvin & D. E. Rumelhart (Eds.),
Backpropagation: Theory, architectures and applications.
Hillsdale, NJ: Erlbaum.
Wilson, J. R., Bose, N., Sherman, S. M., & Guillery, R. W. (1984).
Fine structural morphology of identified X- and Y-cells in
the cat’s lateral geniculate nucleus. Proceedings of the
Royal Society of London, Series B, Biological Sciences, 221,
411–436. DOI: https://doi.org/10.1098/rspb.1984.0042,
PMID: 6146984
Wimmer, R. D., Schmitt, L. I., Davidson, T. J., Nakajima, M.,
Deisseroth, K., & Halassa, M. M. (2015). Thalamic control of
sensory selection in divided attention. Nature, 526, 705–709.
DOI: https://doi.org/10.1038/nature15398, PMID: 26503050,
PMCID: PMC4626291
Wiskott, L., & Sejnowski, T. J. (2002). Slow feature analysis:
Unsupervised learning of invariances. Neural Computation, 14,
715–770. DOI: https://doi.org/10.1162/089976602317318938,
PMID: 11936959
Worden, M. S., Foxe, J. J., Wang, N., & Simpson, G. V. (2000).
Anticipatory biasing of visuospatial attention indexed by
retinotopically specific alpha-band electroencephalography
increases over occipital cortex. Journal of Neuroscience, 20,
RC63. DOI: https://doi.org/10.1523/JNEUROSCI.20-06-j0002
.2000, PMID: 10704517, PMCID: PMC6772495
O’Reilly et al.
1195
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
1
5
8
1
9
1
3
5
7
7
/
/
j
o
c
n
_
a
_
0
1
7
0
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Wurtz, R. H. (2008). Neuronal mechanisms of visual stability.
Vision Research, 48, 2070–2089. DOI: https://doi.org/10.1016
/j.visres.2008.03.021, PMID: 18513781, PMCID: PMC2556215
Xing, D., Yeh, C.-I., Burns, S., & Shapley, R. M. (2012). Laminar
analysis of visually evoked activity in the primary visual cortex.
Proceedings of the National Academy of Sciences, U.S.A., 109,
13871–13876. DOI: https://doi.org/10.1073/pnas.1201478109,
PMID: 22872866, PMCID: PMC3427063
Yu, C., & Smith, L. B. (2012). Embodied attention and word
learning by toddlers. Cognition, 125, 244–262. DOI: https://
doi.org/10.1016/j.cognition.2012.06.016, PMID: 22878116,
PMCID: PMC3829203
Zhou, H., Schafer, R. J., & Desimone, R. (2016). Pulvinar-cortex
interactions in vision and attention. Neuron, 89, 209–220.
DOI: https://doi.org/10.1016/j.neuron.2015.11.034, PMID:
26748092, PMCID: PMC4723640
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
1
5
8
1
9
1
3
5
7
7
/
/
j
o
c
n
_
a
_
0
1
7
0
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
1196
Journal of Cognitive Neuroscience
Volume 33, Number 6