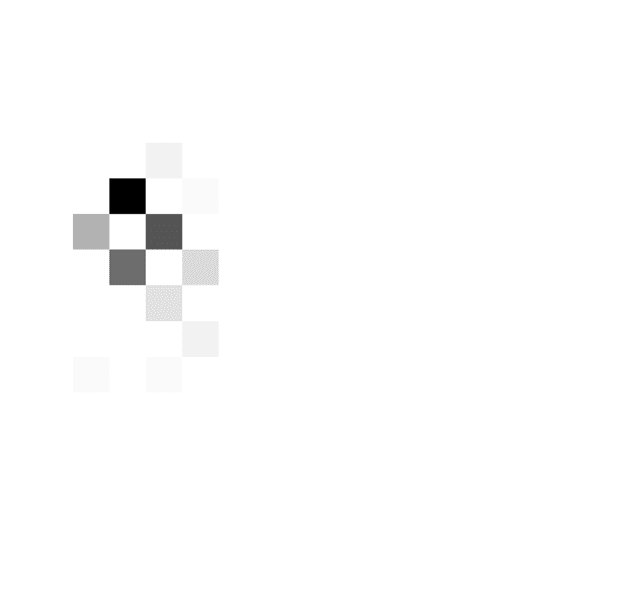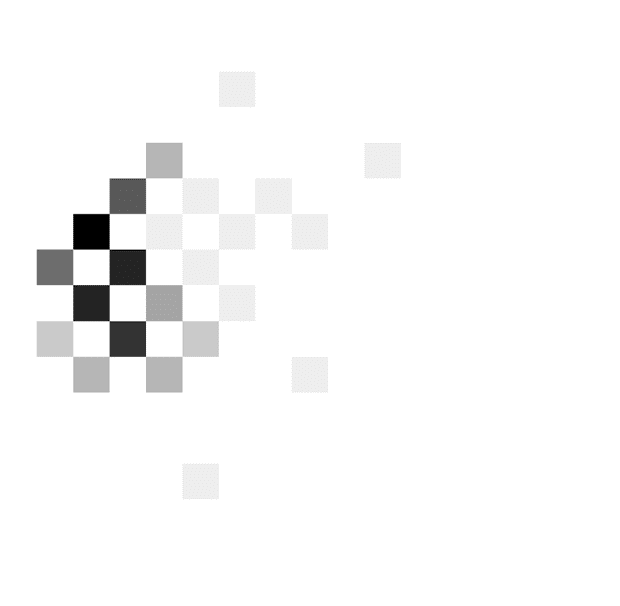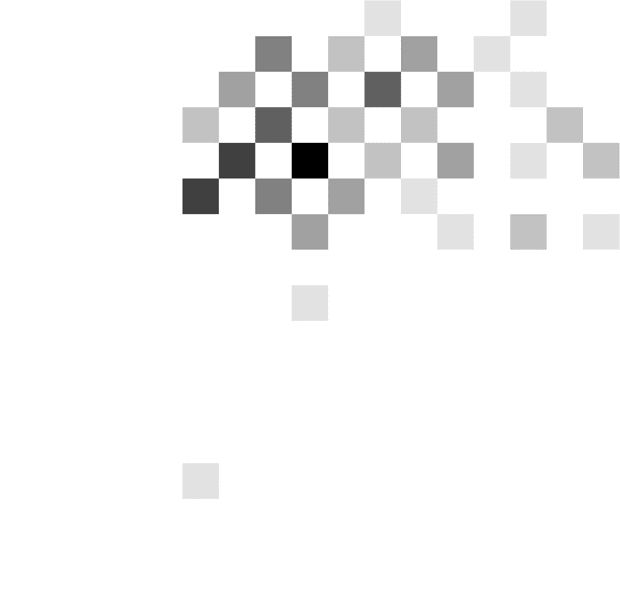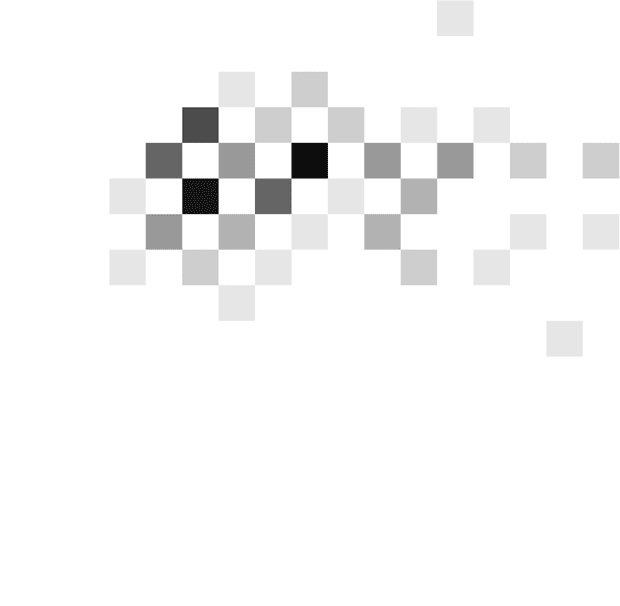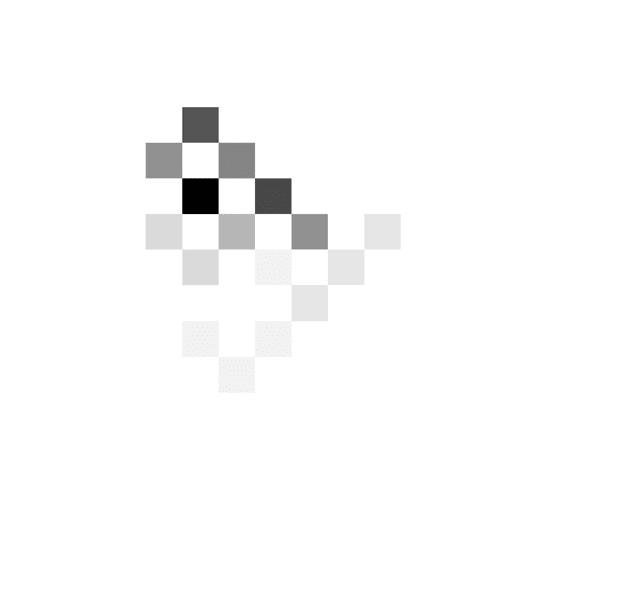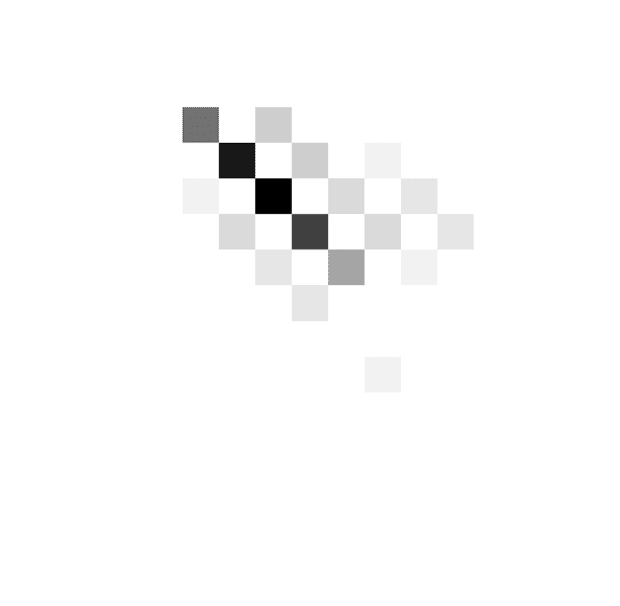Deep Brain Stimulation of the Subthalamic Nucleus Does
Not Affect the Decrease of Decision Threshold during
the Choice Process When There Is No Conflict,
Time Pressure, or Reward
Friederike Leimbach1, Dejan Georgiev1, Vladimir Litvak1, Chrystalina Antoniades2,
Patricia Limousin1, Marjan Jahanshahi1,3, and Rafal Bogacz2
Abstract
■ During a decision process, the evidence supporting alter-
native options is integrated over time, and the choice is made
when the accumulated evidence for one of the options
reaches a decision threshold. Humans and animals have an
ability to control the decision threshold, that is, the amount
of evidence that needs to be gathered to commit to a choice,
and it has been proposed that the subthalamic nucleus (STN)
is important for this control. Recent behavioral and neuro-
physiological data suggest that, in some circumstances, the
decision threshold decreases with time during choice trials,
allowing overcoming of indecision during difficult choices.
Here we asked whether this within-trial decrease of the decision
threshold is mediated by the STN and if it is affected by disrupt-
ing information processing in the STN through deep brain stim-
ulation (DBS). We assessed 13 patients with Parkinson disease
receiving bilateral STN DBS six or more months after the sur-
gery, 11 age-matched controls, and 12 young healthy controls.
All participants completed a series of decision trials, in which
the evidence was presented in discrete time points, which al-
lowed more direct estimation of the decision threshold. The
participants differed widely in the slope of their decision
threshold, ranging from constant threshold within a trial to
steeply decreasing. However, the slope of the decision thresh-
old did not depend on whether STN DBS was switched on
or off and did not differ between the patients and controls.
Furthermore, there was no difference in accuracy and RT be-
tween the patients in the on and off stimulation conditions
and healthy controls. Previous studies that have reported
modulation of the decision threshold by STN DBS or unilateral
subthalamotomy in Parkinson disease have involved either fast
decision-making under conflict or time pressure or in
anticipation of high reward. Our findings suggest that, in the
absence of reward, decision conflict, or time pressure for
decision-making, the STN does not play a critical role in mod-
ulating the within-trial decrease of decision thresholds during
the choice process. ■
INTRODUCTION
Converging behavioral and neurophysiological data sug-
gest that during decision-making between two options,
the evidence favoring one alternative over another is
integrated over time (Ratcliff & McKoon, 2008; Gold &
Shadlen, 2007; Schall, 2001). These data also suggest that
the choice is made when the amount of integrated evi-
dence reaches a particular value, referred to as a decision
threshold. Humans and animals have an ability to adjust
the decision threshold according to instructions to be fast
or accurate or according to task demands ( Wickelgren,
1977). It has been suggested that the adjustments of
decision threshold rely on the BG circuit, and several
possible mechanisms have been proposed (Bogacz,
Wagenmakers, Forstmann, & Nieuwenhuis, 2010; Forstmann
1University College London Institute of Neurology, 2University
of Oxford, 3University of Electronic Science and Technology of
China
et al., 2008; Furman & Wang, 2008; Frank, Scheres, &
Sherman, 2007). One of these theories suggests that the
threshold is controlled by the activity level in the sub-
thalamic nucleus (STN), which through its widespread
connections can control the excitability in the output of
the BG and thus gate decisions (Frank, Scheres, et al.,
2007). This theory has been supported by reports that
deep brain stimulation (DBS) of the STN affected the
extent to which patients with Parkinson disease (PD)
were able to vary their decision threshold according to
speed versus accuracy instructions on a motion discrimi-
nation task (Pote et al., 2016; Green et al., 2013), and fol-
lowing unilateral subthalamotomy, patients with PD failed
to show context-dependent modulation of decision
thresholds on a conditional stop signal task (Obeso et al.,
2014). Moreover, the patterns of neural activity in the STN
differed depending on whether the patients with PD were
asked to be fast or accurate on a motion discrimination task
(Herz et al., 2017). Furthermore, in a task-switching para-
digm, a high threshold after a cue predicting a switch was
© 2018 Massachusetts Institute of Technology. Published under a
Creative Commons Attribution 4.0 Unported (CC BY 4.0) license.
Journal of Cognitive Neuroscience 30:6, pp. 876–884
doi:10.1162/jocn_a_01252
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
0
6
8
7
6
1
7
8
7
4
4
9
/
j
o
c
n
_
a
_
0
1
2
5
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
associated with a higher activity in the STN (Mansfield,
Karayanidis, Jamadar, Heathcote, & Forstmann, 2011).
However, the theory of threshold control by the STN is
not aligned with the observation that the ability to vary
decision threshold was not predicted by the amount of
structural connectivity between the cortex and the STN
but rather between the cortex and the striatum (Forstmann
et al., 2010).
Recently, it has been hotly debated whether the deci-
sion threshold remains constant or decreases within the
choice process. On difficult trials on which both options
receive similar amounts of evidence, such decrease would
prevent excessively long deliberation (Cisek, Puskas, &
El-Murr, 2009). Neural signatures of decreasing thresholds
have been observed in firing rates of neurons in areas
involved in the choice process. Namely, it was shown that,
on average, the activity of neural populations selective
for all options increases with time, suggesting that they
receive increasing “urgency” inputs, which with time drive
all neural populations closer to the response threshold
(Churchland, Kiani, & Shadlen, 2008). However, it has
proven more challenging to establish if the decrease in
decision threshold can be inferred from behavioral data.
On the one hand, models with decreasing thresholds
described better the distribution of RTs produced by
monkeys in the motion discrimination tasks (Ditterich,
2006) and by humans when responses had to be made
before a deadline (Murphy, Boonstra, & Nieuwenhuis,
2016). On the other hand, comprehensive studies showed
that multiple data sets from classic decision-making para-
digms from humans are better explained by models with
constant rather than decreasing thresholds ( Voskuilen,
Ratcliff, & Smith, 2016; Hawkins, Forstmann, Wagenmakers,
Ratcliff, & Brown, 2015). These studies had to infer the
slope of the boundary indirectly by fitting computational
models to behavioral data, as in classic decision-making
paradigms information is presented continuously so it is
difficult to estimate what evidence the participant could
have integrated at the moment of choice. To infer the
decision boundary more directly, a recent study employed
an expanded judgment paradigm in which information
is presented in discrete steps, so that it is known what
total evidence was presented on each trial before the
participant made the choice (Malhotra, Leslie, Ludwig, &
Bogacz, 2017). This study demonstrated that humans in-
deed employ decreasing decision thresholds in certain
conditions.
Previous studies suggested that DBS disrupts the func-
tions in which the STN is thought to be involved in the
healthy brain, such as preventing prepotent responses
when they are inappropriate (Georgiev, Dirnberger,
Wilkinson, Limousin, & Jahanshahi, 2016; Wylie et al.,
2010; Thobois et al., 2007; Hershey et al., 2004; Jahanshahi
et al., 2000; for a review, see (Jahanshahi, Obeso, Baunez,
Alegre, & Krack, 2015). Specifically, Frank and colleagues
(2007) proposed that the STN is involved in changing the
height of the threshold within a trial. They studied the
effects of STN stimulation on decision-making between
alternatives that were previously associated with different
reward probabilities. Results suggested that STN stimula-
tion impaired decision-making in high-conflict situations
in which both of the presented options had been asso-
ciated with high reward probability. This was reflected in
the failure of patients with PD to slow down in high-
conflict trials when STN DBS was on, unlike when their
DBS was off or the unoperated PD and healthy controls
(Frank, Samanta, Moustafa, & Sherman, 2007). Similar
findings were reported with the same task by Cavanagh
et al. (2011), who additionally recorded scalp EEG and
showed that STN stimulation reversed the normal increase
in theta-band activity over the medial pFC, which is usually
associated with raising the decision threshold for high-
conflict trials, thus suggesting that STN DBS interferes
with the normal ability of the STN to react to decision con-
flict by modulating the decision threshold (Cavanagh et al.,
2011). In addition, STN DBS has been reported to disrupt
slowing of RTs on conflicting or difficult trials in perceptual
decision-making tasks (Green et al., 2013; Coulthard et al.,
2012) and to result in fast but errorful responses under
speed pressure (Pote et al., 2016).
In addition to the role of the STN in increasing the
threshold within a trial, it has also been suggested that
the decrease in threshold during a choice trial reflects
reduced inhibition from the STN (Ratcliff & Frank,
2012). Here, we asked if the STN is causally involved in
reducing the decision threshold with time within a trial.
To address this question, we studied the decision-making
of patients with PD with DBS applied to the STN. We
asked patients with PD to perform an expanded judg-
ment task with STN DBS switched on or off and investi-
gated whether STN DBS reduces the slope of decrease of
decision threshold.
METHODS
Participants
Thirteen patients (11 men) with a clinical diagnosis of PD
based on the U.K. Brain Bank criteria were assessed
(Hughes, Daniel, Kilford, & Lees, 1992). All patients
had bilateral STN DBS for 6 months or longer. The mean
age was 61.6 (SD = 10.04, range 42–73). To control for
possible practice and age effects, 23 neurologically healthy
participants with no history of psychiatric illness, head
injury, or alcohol or drug abuse were recruited, of whom
11 were matched in age with the patient group (6 men)
and 12 were younger (8 men). The mean age of the age-
matched group was 66.0 (SD = 12.28, range 45–82), and
the mean age of the young group was 29.3 (SD = 4.65,
range 24–35). Patients and older controls were matched
in age ( p > .05). The demographic and clinical informa-
tion for the samples are presented in Table 1. The stimu-
lation parameters for each patient are presented in Table 2.
All patients were assessed on medication.
Leimbach et al.
877
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
0
6
8
7
6
1
7
8
7
4
4
9
/
j
o
c
n
_
a
_
0
1
2
5
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Table 1. Demographic and Clinical Information for the
Three Groups
Group
PD
Age Matched
Young
Age, years
61.6 (10.04)
66.0 (12.28)
29.3 (4.65)
Sex
Male
Female
11
2
6
5
8
4
Years of education
13.8 (2.65)
16.5 (2.07)
19.4 (2.72)
MMSE
28.7 (2.02)
29.3 (0.90)
29.7 (0.89)
Digit Span
18.2 (3.74)
18.2 (3.16)
20.4 (3.75)
BDI
SAS
8.8 (3.63)
3.7 (3.07)
2.3 (2.18)
15.0 (5.45)
10.5 (5.28)
5.9 (3.18)
UPDRS motor part
On
Off
18.1 (6.85)
37.0 (9.87)
Mean and standard deviation values (in brackets) for age, years of
education, Mini Mental State Examination (MMSE), digit span, Beck
Depression Inventory (BDI), Starkstein Apathy Scale (SAS), and Unified
Parkinson Disease Rating Scale-Part III (UPDRS).
The Mini-Mental State Examination, the Beck Depression
Inventory, and the Starkstein Apathy Scale were adminis-
tered to screen for dementia, clinical depression, and
apathy. None of the patients had cognitive impairment,
clinical depression, or apathy.
Task
Participants were asked to perform a computerized ex-
panded judgment task developed in MATLAB. During
each trial, participants were instructed to predict if a
mouse would run left or right. Each trial included multi-
ple presentations of stimuli of a mouse facing either to
the left or to the right (Figure 1A). The participants were
told that “The mouse is more likely to look in the direc-
tion it will run, but sometimes it looks in the other direc-
tion.” The stimuli were selected stochastically such that
the probability of mouse looking in the “correct” direc-
tion was .7. The same randomly pregenerated sequences
of stimuli were used for all participants. On each trial, the
stimuli were presented until a response was indicated by
the participant by pressing the appropriate right or left
buttons of a response box with their right or left index
finger, respectively. After each response, participants
were given visual feedback (correct or incorrect).
The patients performed the task in three conditions
differing in the rate of presentation of the stimuli, namely
the stimuli were presented every 200, 500, or 1000 msec.
The fastest rate (200 msec) was the same as in a previous
study of the slope of the decision threshold (Malhotra
et al., 2017). In the medium and slow rate conditions,
stimuli were presented in the center of the screen
(Figure 1A), whereas in the fast rate condition, the mice
looking left were presented on the left side of the screen,
and vice versa, to make the direction easier to identify
within a short period (Figure 1B).
Table 2. Stimulation Parameters for the Left and Right Electrodes Implanted in the STN for the 13 Patients with PD
Left STN
Right STN
Patient
1
2
3
4
5
6
7
8
9
10
11
12
13
Active
Contact
−2, −3
−0
−2
−1
−1
−1, −2
−1
−1
−0
−2
−1
−2, −3
−1
Voltage
( V)
Frequency
(Hz)
4.0
2.3
2.5
3.4
1.6
1.1
2.5
1.65
2.8
1.9
4.2
3.9
4.1
60
60
160
80
130
160
130
80
80
125
80
60
125
Pulse
Width
(μsec)
100
Active
Contact
−11
80
60
60
60
60
60
60
60
60
60
80
60
−8
−9
−5
−9
−8
−9
−8
−10
−9
−5
−5
−8
Voltage
( V)
Frequency
(Hz)
3.1
3.0
2.3
3.3
1.8
1.1
2.5
2.2
2.8
1.4
3.2
3.5
3.9
60
60
160
80
130
160
130
80
80
125
80
60
125
Pulse
Width
(μsec)
100
80
60
60
60
60
60
60
60
60
60
80
60
Mean
2.77
102.31
66.15
2.62
102.31
66.15
878
Journal of Cognitive Neuroscience
Volume 30, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
0
6
8
7
6
1
7
8
7
4
4
9
/
j
o
c
n
_
a
_
0
1
2
5
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 1. Timeline of a single
trial. (A) Slow and medium
rate conditions. In the slow
condition, the stimulus was
presented for 200 msec
followed by 800 msec of blank
screen, whereas in the medium
condition, the stimulus was
presented for 150 msec and
blank screen for 350 msec.
(B) In the fast condition, the
stimulus was presented for
66 msec followed by a blank
screen for 134 msec.
Each condition started with practice trials. During pilot-
ing, we found that some participants had a tendency to
respond after seeing only one stimulus. To illustrate the
benefit of integrating evidence, in the initial 10 practice
trials the participants were asked to wait for a “Go” cue
on the screen before pressing a button; the different
numbers of stimuli were presented on each trial. In the
next 20 practice trials (and in the main experiment), the
stimuli were presented sequentially until the participant
pressed a response button. Subsequently, participants
completed two experimental blocks of 50 trials, separated
by a break. At the end of each block, they were provided
with the percentage of correct responses.
Design
Patients completed the task with three different rates
(200, 500, and 1000 msec) on three separate days. On
each day, they were assessed with their stimulators being
switched on and with stimulators switched off. The order
of the stimulation condition was counterbalanced across
patients. The healthy control participants completed the
task only with the medium speed rate (500 msec), as we
did not expect the rate of stimulus presentation to affect
patients and controls in different ways. Control partici-
pants were also tested twice to ensure they performed
the same number of trials in the medium speed condition
as the patients, to control for practice effects.
Exclusion Criteria
As the task was relatively easy, participants typically ob-
tained high accuracy (Figure 2A), but a few patients, on
some of the study days, performed at chance level. We
excluded data from conditions in which the subjective
accuracy (defined as the fraction of trials on which re-
sponse agreed with the majority of stimuli presented
on a trial) did not significantly differ from that expected
by chance (i.e., when it was below 59.8% corresponding
to a nonsignificant Z test). This resulted in excluding
some of the data from three patients: Patient 1, both
DBS conditions with 200-msec rate and both DBS con-
ditions with 1000-msec rate; Patient 4, both DBS condi-
tions with 200-msec rate and both DBS conditions with
1000-msec rate; and Patient 11, both DBS conditions with
200 msec.
Data Analysis
For each participant and condition, we estimated the
slope of decrease of decision threshold using the method
developed and tested in a previous study (Malhotra et al.,
2017). This method assumes that a state of a decision-
maker after seeing ith stimulus during the choice process
is described by the integrated evidence for the chosen
option xi and time step ti. For example, consider a trial
in which a participant observes sequence: left, right, left,
left, and then chooses the left response. The integrated
evidence increases by 1 each time a stimulus correspond-
ing to the chosen option is presented and decreases by
1 otherwise, resulting in the following sequence: (t0 = 0,
x0 = 0), (t0 = 1, x0 = 1), (t0 = 2, x0 = 0), (t0 = 3, x0 = 1),
(t0 = 4, x0 = 2). We looked for a line in the (t, x)
space, such that the states were typically below it before
making the decision and above it after the last stimulus.
Denoting making a choice by 1 and waiting by 0, we can
define a sequence of actions ai, which for our sample
trial would be a0 = 0, a1 = 0, a2 = 0, a3 = 0, a4 = 1.
Leimbach et al.
879
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
0
6
8
7
6
1
7
8
7
4
4
9
/
j
o
c
n
_
a
_
0
1
2
5
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
The probability of an action can be related to the predictor
variables using the following logistic regression model,
log
P ai ¼ 1
ð
Þ
Þ
ð
P ai ¼ 0
¼ β
þ β
tti þ β
x xi
0
In the above equation, βt and βx are the regression coef-
ficients for time and evidence, respectively, and β0 is the
intercept. Given the triplet (xi, ti, ai) for each stimulus in
each trial, we estimated for each participant and condition
the βt, βx, and β0 that maximized the likelihood of the
observed triplets. Next, the slope of the decrease of deci-
sion threshold was taken as arctan(βt/βx). This slope is
equal to 0 if the threshold is constant within a trial and
takes positive values if the threshold decreases.
We assessed how the accuracy, RT, and slope of the
threshold depended on experimental group and condition.
For each of these measures, for the patients we tested
how it depended on presentation rate and STN stimulation
condition and performed repeated-measures ANOVA with
two factors: Rate (200, 500, 1000 msec) and DBS (on vs off ).
In addition, to compare performance in the medium rate
condition between the groups, an ANOVA was performed
with one factor: Group (patients with DBS on, patients with
DBS off, age-matched controls, young controls).
RESULTS
Behavior
Patients had significantly reduced motor symptoms,
as measured by the Unified Parkinson Disease Rating
Scale-Part III, when the DBS was on in comparison to
the condition when it was off (Table 1; p < .001, t = 7.96,
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
6
8
7
6
1
7
8
7
4
4
9
/
j
o
c
n
_
a
_
0
1
2
5
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 2. Comparison of behavior across experimental groups and conditions. Different colors correspond to different rate conditions (see key).
(A) Objective accuracy defined as the fraction of trials on which “Correct” feedback was given. (B) Mean RT. (C) Slope of decision threshold. In A–C, the
error bars show the SEM. (D) Distribution of slopes across patients. Each circle corresponds to data from a particular patient in a particular rate condition,
and its coordinates are the estimated slopes off and on STN DBS. The dashed line shows the identity line, so that the points below it have a higher
slope off STN DBS, whereas points above it have a higher slope on STN DBS. Matched = age-matched healthy controls; Young = young healthy controls.
880
Journal of Cognitive Neuroscience
Volume 30, Number 6
df = 12). Some patients received DBS with frequencies
above 100 Hz whereas others received DBS with lower
frequencies (Table 2), but these two groups did not
significantly differ in any behavioral measure reported
below, and thus, the analyses of pooled data from both
groups are presented.
Figure 2 (A and B) shows the accuracy and RTs of
different groups in different conditions of the experi-
mental task. In the medium rate condition, the patients
with DBS on or off, elderly and young healthy control
groups did not differ significantly in the accuracy ( p =
.70, F = 0.48, df = 3) or RT ( p = .34, F = 1.15, df =
3). The accuracy of the patients did not significantly
differ with rate ( p = .20, F = 1.66, df = 2) or with the
stimulation being on or off ( p = .74, F = 0.11, df = 1),
and there was no interaction ( p = .88, F = 0.12, df = 2).
The RT of the patients depended on the rate of the pre-
sentation of the stimuli ( p < .0001, F = 39.68, df = 2).
Thus, the patients responded faster if the stimuli were
presented at a faster rate. Nevertheless, the patients did
not maintain across rates the average number of stimuli
they were viewing before making a choice. For example,
the interval between the onset of stimuli is twice as long
in the slow (1000 msec) than in the medium condition
(500 msec), but the RT in the slow condition is far from
being twice as long as in the medium condition. Thus,
the patients on average were viewing fewer stimuli before
making a choice in the slower condition. However, the
RT did not change with the stimulation ( p = .80, F =
0.06, df = 1), and there was no interaction ( p = .87, F =
0.13, df = 2).
Slopes of the Decision Threshold
Patients showed a great diversity of slopes of decision
threshold as illustrated in Figure 3, which shows at what
combinations of time and evidence the choices were
made. The patient in Figure 3A tended to respond when
the cumulated evidence was in a particular range (around
three to seven in the DBS off condition and around two
to five in the DBS on condition) irrespective of the time
from trial onset. The behavior of this patient is best de-
scribed by a relatively flat decision threshold (slopes 7°
and 2° in the DBS off and on conditions, respectively).
By contrast, the patient shown in Figure 3B responded
at lower evidence levels as time progressed, and the be-
havior is best described by decreasing boundaries (slopes
39° and 45° in the off and on conditions). On the other
hand, the patient shown in Figure 3C tended to respond
after seeing a particular number of stimuli irrespective of
the integrated evidence (around two to three stimuli in
the DBS off condition and three to four stimuli in the
DBS on condition), and this behavior is described by
steeply decreasing thresholds (slopes 85° and 61° in the
off and on conditions). Although the moment of the re-
sponse is mostly determined by the number of stimuli
seen by this patient, the actual response (left or right) is
influenced by the stimuli seen, which allows the patient
to achieve an accuracy above chance.
Figure 2D shows the distribution of the slopes of deci-
sion threshold across patients. The behavior illustrated in
Figure 3A and C corresponds to the lowest and one of
the highest slopes, and it can be seen in Figure 2D that
most patients had estimated slopes between these ex-
tremes. Similar slopes were produced by healthy control
participants, and Figure 2C compares the mean slopes
across the groups and conditions. In the medium rate
condition, the groups did not differ significantly in the
slope ( p = .57, F = 0.68, df = 3). The slope of the
patients’ thresholds differed between the rate conditions
( p = .015, F = 4.58, df = 2), and it can be noticed in
Figure 2D that the points corresponding to the fast rate
condition (green circles) tend to have lower slopes of the
threshold. There is no systematic difference in the slopes
between 500- and 1000-msec rate conditions (Figure 2C)
despite the difference in the number of stimuli viewed
(Figure 2B), but note that the number of stimuli viewed
is not only determined by the slope of the threshold
but also by its intercept.
The slope of the threshold did not depend significantly
on the DBS condition ( p = .58, F = 0.32, df = 1), and
there was no interaction ( p = .27, F = 1.35, df = 2).
Figure 2D does not show any obvious tendency for the
points to lie on either side of the identity line. To quan-
tify this further, we compared the coordinates of points
in Figure 2D, with a paired Bayesian t test, using statistical
software JASP 0.8.5.1. The Bayes factor, that is, the ratio
of the probability of the null hypothesis (no difference
between slopes) over the alternative hypothesis was
4.26. The patients produced similar slopes in the two
DBS conditions, and there was a significant correlation
between the slopes produced off and on DBS across
patients and conditions (r = .78, p < .0001).
DISCUSSION
In this study, the estimated slopes of the decision thresh-
old differed significantly as a function of the rate of pre-
sentation of the stimuli, with faster rates associated with
lower slopes of decision threshold, suggesting that pa-
tients adjusted the slope of decision threshold within a
trial according to task demands, but the slope was not
affected by STN DBS. Decreasing decision thresholds
were recently also observed in a task employing a similar
expanded judgment paradigm (Malhotra et al., 2017). A
steeper decrease of the threshold in a slower rate con-
dition may be connected with a higher mental cost of
maintaining the information between the stimuli, which
has been proposed to affect the slope of decision bound-
aries (Drugowitsch, Moreno-Bote, Churchland, Shadlen,
& Pouget, 2012).
The lack of the effect of STN DBS on changing the
slope of decision threshold within a trial might be related
to the differences in the tasks. First, this study did not
Leimbach et al.
881
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
6
8
7
6
1
7
8
7
4
4
9
/
j
o
c
n
_
a
_
0
1
2
5
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. Examples of
distributions of combinations of
time and evidence at which
choices were made. Different
panels correspond to different
patients. Data in A come from
the fast rate condition, whereas
data in B and C come from the
slow rate condition. Displays in
the left column show data from
an STN DBS off condition,
whereas displays in the right
column correspond to STN DBS
on condition. In each display,
the darkness of a square
indicates the number of trials at
which the choice was made
after seeing a particular number
of stimuli (x axis) and particular
accumulated evidence ( y axis).
The color scale is normalized
such that black corresponds to
the maximum of a histogram.
The coordinates of red circles
indicate the mean number of
stimuli and evidence, whereas
the whiskers indicate standard
deviations.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
6
8
7
6
1
7
8
7
4
4
9
/
j
o
c
n
_
a
_
0
1
2
5
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
explicitly introduce any conflict between responses. Un-
like the study of Frank and colleagues (2007), where
some trials involved the choice between two options
previously associated with a high probability of reward,
in our study each stimulus provided evidence for one
option or another but never for both options. Second,
the presented experiment employed an expanded judg-
ment paradigm in which responses were much slower
than in previous studies. It is possible that in such slow
tasks the behavior is more affected by a process of forc-
ing decisions to move on to the next trial (Drugowitsch
et al., 2012) than by a process of slowing down responses
in high-conflict situations (Frank, Samanta, et al., 2007).
This dominant effect of this forcing process is manifested
by extremely steep decision thresholds employed by
some participants (Figure 3C). It is possible that the
882
Journal of Cognitive Neuroscience
Volume 30, Number 6
processes of slowing down and speeding up the decision
may have different neural bases, as patterns of activity in
distinct frequencies have been observed in the STN when
patients were asked to be accurate and fast (Herz et al.,
2017). Furthermore, STN DBS induced impulsive action
reflected as fast and errorful responding and lowering of
the decision threshold on the moving dots task was only
observed under speed instructions when patients with
PD were acting under time pressure (Pote et al., 2016).
This is consistent with the proposal that the BG in-
vigorate the decision-making process by providing an ur-
gency signal (Thura & Cisek, 2017). Third, it is possible
that, in our expanded judgment task, some participants
used explicit strategies, which contributed to the diver-
sity of slopes of decision threshold. This diversity might
have masked the effects of DBS. Finally, in the study by
Frank and colleagues (2007), decisions were based on
previously learned stimulus–action–reward associations,
and STN stimulation only produced impulsive behaviors
when decision-making in win–win situations. Our study
did not involve reward-based decisions, and the function
of the STN may be more related to decisions that lead
to positive outcomes.
The lack of the effect of DBS may also be connected
with the differences in methodology. In the study by
Frank and colleagues (2007), the patients were tested
on relatively low doses of medication that might have
magnified the effects of DBS, whereas in our study, the
patients had normal doses of their medications. Further-
more, in previous research patients with STN DBS were
tested in two different sessions on different days with
their stimulation on or off. The patients were less likely
to become fatigued during the sessions. In this investiga-
tion, patients were tested on and off stimulation during
one session with potential fatigue or “carryover” effects,
despite the counterbalancing of the order. Given many
differences in tasks and methodology between our and
previous studies, a very interesting direction of future
work would be to systematically perform a series of
experiments, including a repetition of a previous study
that showed an effect of DBS, and several conditions
each differing in just a single aspect from the repetition.
It would allow to identify which features are critical for
the effect of DBS on decision-making to be present.
Previous research looking at the acute effects of STN
stimulation on decision-making reached contradicting
conclusions. Although some authors found deficits with
STN stimulation on (Pote et al., 2016; Florin et al.,
2013; Green et al., 2013; Coulthard et al., 2012; Cavanagh
et al., 2011; Oyama et al., 2011; Rogers et al., 2011; Frank,
Samanta, et al., 2007), others reported that decision-
making remained stable or even improved with STN stim-
ulation on relative to when the DBS was off (Brandt et al.,
2015; Fumagalli et al., 2015; Boller et al., 2014; Torta
et al., 2012; van Wouwe et al., 2011). To assess risk-taking
and delay aversion, Torta and colleagues (2012) adminis-
tered the Cambridge Gambling Task and the Quick Delay
Questionnaire to assess how STN stimulation affected
willingness of patients with PD to respond later to receive
bigger rewards. The results suggested that the patients’
response delays did not change with STN stimulation
(Torta et al., 2012). These findings match our results,
considering that we did not find any changes in RT
slowing between patients in the on and off stimulation
conditions.
In conclusion, our data suggest that stimulation of the
STN does not impair the patients’ ability to reduce their
decision threshold with time within a trial in a relatively
slow expanded judgment paradigm. This is in contrast to
previous studies suggesting a significant effect of STN
DBS on the ability to adjust the decision threshold
between trials and on adjusting threshold within a trial
in more rapid tasks performed under speed pressure or
associated with reward or conflict. Thus, our data raise
the possibility that the proposed role of the STN in
adjusting thresholds within a trial is specific to rapid
decision-making where it is important to slow down the
process when conflicting information is present or high
reward is at stake.
Acknowledgments
This research was supported by MRC grant MC_UU_12024/5.
The Wellcome Centre for Human Neuroimaging was supported
by core funding from the Wellcome Trust (203147/Z/16/Z). The
authors thank Ewa Bogacz for drawing the stimuli.
Reprint requests should be sent to Rafal Bogacz, MRC Brain
Networks Dynamics Unit, Nuffield Department of Clinical Neuro-
science, University of Oxford, John Radcliffe Hospital, Oxford,
OX3 9DU, United Kingdom, or via e-mail: rafal.bogacz@ndcn.
ox.ac.uk.
REFERENCES
Bogacz, R., Wagenmakers, E. J., Forstmann, B. U., & Nieuwenhuis,
S. (2010). The neural basis of the speed-accuracy tradeoff.
Trends in Neurosciences, 33, 10–16.
Boller, J. K., Barbe, M. T., Pauls, K. A. M., Reck, C., Brand,
M., Maier, F., et al. (2014). Decision-making under risk is
improved by both dopaminergic medication and subthalamic
stimulation in Parkinson’s disease. Experimental Neurology,
254, 70–77.
Brandt, J., Rogerson, M., Al-Joudi, H., Reckess, G., Shpritz, B.,
Umeh, C. C., et al. (2015). Betting on DBS: Effects of
subthalamic nucleus deep brain stimulation on risk taking
and decision making in patients with Parkinson’s disease.
Neuropsychology, 29, 622–631.
Cavanagh, J. F., Wiecki, T. V., Cohen, M. X., Figueroa, C. M.,
Samanta, J., Sherman, S. J., et al. (2011). Subthalamic nucleus
stimulation reverses mediofrontal influence over decision
threshold. Nature Neuroscience, 14, 1462–1467.
Churchland, A. K., Kiani, R., & Shadlen, M. N. (2008). Decision-
making with multiple alternatives. Nature Neuroscience, 11,
693–702.
Cisek, P., Puskas, G. A., & El-Murr, S. (2009). Decisions in
changing conditions: The urgency-gating model. Journal of
Neuroscience, 29, 11560–11571.
Coulthard, E. J., Bogacz, R., Javed, S., Mooney, L. K., Murphy,
G., Keeley, S., et al. (2012). Distinct roles of dopamine and
Leimbach et al.
883
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
6
8
7
6
1
7
8
7
4
4
9
/
j
o
c
n
_
a
_
0
1
2
5
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
subthalamic nucleus in learning and probabilistic decision
making. Brain, 135, 3721–3734.
Ditterich, J. (2006). Stochastic models of decisions about
motion direction: Behavior and physiology. Neural
Networks, 19, 981–1012.
Drugowitsch, J., Moreno-Bote, R., Churchland, A. K., Shadlen,
M. N., & Pouget, A. (2012). The cost of accumulating evidence
in perceptual decision making. Journal of Neuroscience, 32,
3612–3628.
Florin, E., Müller, D., Pfeifer, J., Barbe, M. T., Fink, G. R., &
Timmermann, L. (2013). Subthalamic stimulation modulates
self-estimation of patients with Parkinson’s disease and
induces risk-seeking behaviour. Brain, 136, 3271–3281.
Forstmann, B. U., Anwander, A., Schäfer, A., Neumann, J.,
Brown, S., Wagenmakers, E.-J., et al. (2010). Cortico-striatal
connections predict control over speed and accuracy in
perceptual decision making. Proceedings of the National
Academy of Sciences, U.S.A., 107, 15916–15920.
Forstmann, B. U., Dutilh, G., Brown, S., Neumann, J., von Cramon,
D. Y., Ridderinkhof, K. R., et al. (2008). Striatum and pre-SMA
facilitate decision-making under time pressure. Proceedings of
the National Academy of Sciences, U.S.A., 105, 17538–17542.
Jahanshahi, M., Obeso, I., Baunez, C., Alegre, M., & Krack, P.
(2015). Parkinson’s disease, the subthalamic nucleus,
inhibition, and impulsivity. Movement Disorders, 30, 128–140.
Malhotra, G., Leslie, D. S., Ludwig, C. J. H., & Bogacz, R. (2017).
Overcoming indecision by changing the decision boundary.
Journal of Experimental Psychology: General, 146, 776–805.
Mansfield, E. L., Karayanidis, F., Jamadar, S., Heathcote, A., &
Forstmann, B. U. (2011). Adjustments of response threshold
during task switching: A model-based functional magnetic
resonance imaging study. Journal of Neuroscience, 31,
14688–14692.
Murphy, P. R., Boonstra, E., & Nieuwenhuis, S. (2016). Global gain
modulation generates time-dependent urgency during perceptual
choice in humans. Nature Communications, 7, 13526.
Obeso, I., Wilkinson, L., Casabona, E., Speekenbrink, M.,
Bringas, M. L., Álvarez, M., et al. (2014). The subthalamic
nucleus and inhibitory control: Impact of subthalamotomy
in Parkinson’s disease. Brain, 137, 1470–1480.
Oyama, G., Shimo, Y., Natori, S., Nakajima, M., Ishii, H., Arai, H.,
et al. (2011). Acute effects of bilateral subthalamic stimulation
on decision-making in Parkinson’s disease. Parkinsonism
and Related Disorders, 17, 189–193.
Frank, M. J., Samanta, J., Moustafa, A. A., & Sherman, S. J.
Pote, I., Torkamani, M., Kefalopoulou, Z. M., Zrinzo, L.,
(2007). Hold your horses: Impulsivity, deep brain stimulation,
and medication in parkinsonism. Science, 318, 1309–1312.
Frank, M. J., Scheres, A., & Sherman, S. J. (2007). Understanding
decision making deficits in neurological conditions: Insights
from models of natural action selection. Philosophical
Transactions of the Royal Society of London, Series B:
Biological Sciences, 362, 1641–1654.
Fumagalli, M., Marceglia, S., Cogiamanian, F., Ardolino, G.,
Picascia, M., Barbieri, S., et al. (2015). Ethical safety of deep
brain stimulation: A study on moral decision-making in
Parkinson’s disease. Parkinsonism and Related Disorders,
21, 709–716.
Furman, M., & Wang, X. J. (2008). Similarity effect and optimal
control of multiple-choice decision making. Neuron, 60,
1153–1168.
Georgiev, D., Dirnberger, G., Wilkinson, L., Limousin, P., &
Jahanshahi, M. (2016). In Parkinson’s disease on a probabilistic
go/no-go task deep brain stimulation of the subthalamic
nucleus only interferes with withholding of the most prepotent
responses. Experimental Brain Research, 234, 1133–1143.
Gold, J. I., & Shadlen, M. N. (2007). The neural basis of decision
making. Annual Review of Neuroscience, 30, 535–574.
Green, N., Bogacz, R., Huebl, J., Beyer, A.-K., Kühn, A. A., &
Heekeren, H. R. (2013). Reduction of influence of task
difficulty on perceptual decision making by STN deep brain
stimulation. Current Biology, 23, 1681–1684.
Hawkins, G. E., Forstmann, B. U., Wagenmakers, E.-J., Ratcliff,
R., & Brown, S. D. (2015). Revisiting the evidence for
collapsing boundaries and urgency signals in perceptual
decision-making. Journal of Neuroscience, 35, 2476–2484.
Hershey, T., Revilla, F. J., Wernle, A., Gibson, P. S., Dowling,
J. L., & Perlmutter, J. S. (2004). Stimulation of STN impairs
aspects of cognitive control in PD. Neurology, 62, 1110–1114.
Herz, D. M., Tan, H., Brittain, J.-S., Fischer, P., Cheeran, B.,
Green, A. L., et al. (2017). Distinct mechanisms mediate
speed-accuracy adjustments in cortico-subthalamic networks.
eLife, 6, e21481.
Hughes, A. J., Daniel, S. E., Kilford, L., & Lees, A. J. (1992).
Accuracy of clinical diagnosis of idiopathic Parkinson’s
disease: A clinico-pathological study of 100 cases. Journal of
Neurology, Neurosurgery, and Psychiatry, 55, 181–184.
Jahanshahi, M., Ardouin, C. M. A., Brown, R. G., Rothwell, J. C.,
Obeso, J., Albanese, A., et al. (2000). The impact of deep
brain stimulation on executive function in Parkinson’s disease.
Brain, 123, 1142–1154.
Limousin-Dowsey, P., Foltynie, T., et al. (2016). Subthalamic
nucleus deep brain stimulation induces impulsive action
when patients with Parkinson’s disease act under speed
pressure. Experimental Brain Research, 234, 1837–1848.
Ratcliff, R., & Frank, M. J. (2012). Reinforcement-based
decision making in corticostriatal circuits: Mutual constraints
by neurocomputational and diffusion models. Neural
Computation, 24, 1186–1229.
Ratcliff, R., & McKoon, G. (2008). The diffusion decision model:
Theory and data for two-choice decision tasks. Neural
Computation, 20, 873–922.
Rogers, R. D., Wielenberg, B., Wojtecki, L., Elben, S., Campbell-
Meiklejohn, D., & Schnitzler, A. (2011). Deep brain stimulation
of the subthalamic nucleus transiently enhances loss-chasing
behaviour in patients with Parkinson’s disease. Experimental
Neurology, 231, 181–189.
Schall, J. D. (2001). Neural basis of deciding, choosing and
acting. Nature Reviews Neuroscience, 2, 33–42.
Thobois, S., Hotton, G. R., Pinto, S., Wilkinson, L., Limousin-
Dowsey, P., Brooks, D. J., et al. (2007). STN stimulation alters
pallidal-frontal coupling during response selection under
competition. Journal of Cerebral Blood Flow and Metabolism,
27, 1173–1184.
Thura, D., & Cisek, P. (2017). The basal ganglia do not select
reach targets but control the urgency of commitment.
Neuron, 95, 1160–1170.
Torta, D. M. E., Vizzari, V., Castelli, L., Zibetti, M., Lanotte, M.,
Lopiano, L., et al. (2012). Impulsivities and Parkinson’s
disease: Delay aversion is not worsened by deep brain
stimulation of the subthalamic nucleus. PLoS One, 7, e43261.
van Wouwe, N. C., Ridderinkhof, K. R., van den Wildenberg,
W. P. M., Band, G. P. H., Abisogun, A., Elias, W. J., et al.
(2011). Deep brain stimulation of the subthalamic nucleus
improves reward-based decision-learning in Parkinson’s
disease. Frontiers in Human Neuroscience, 5, 30.
Voskuilen, C., Ratcliff, R., & Smith, P. L. (2016). Comparing fixed
and collapsing boundary versions of the diffusion model.
Journal of Mathematical Psychology, 73, 59–79.
Wickelgren, W. A. (1977). Speed-accuracy tradeoff and
information processing dynamics. Acta Psychologica, 41, 67–85.
Wylie, S. A., Ridderinkhof, K. R., Elias, W. J., Frysinger, R. C.,
Bashore, T. R., Downs, K. E., et al. (2010). Subthalamic
nucleus stimulation influences expression and suppression of
impulsive behaviour in Parkinson’s disease. Brain, 133,
3611–3624.
884
Journal of Cognitive Neuroscience
Volume 30, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
6
8
7
6
1
7
8
7
4
4
9
/
j
o
c
n
_
a
_
0
1
2
5
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3