Composite Measures of Brain Activation Predict Individual
Differences in Behavioral Stroop Interference
Louisa L. Smith1
, Hannah R. Snyder2, Benjamin L. Hankin3, and Marie T. Banich1
Abstract
■ The goal of the current study was to interrogate aspects of
the cascade-of-control model [Banich, M. T. Executive function:
The search for an integrated account. Current Directions in
Psychological Science, 18, 89–94, 2009; Banich, M. T. The
Stroop effect occurs at multiple points along a cascade of con-
trol: Evidence from cognitive neuroscience approaches. Frontiers
in Psychology, 10, 2164, 2019], a neurocognitive model that
posits how portions of pFC interact in a cascade-like manner
to overcome interference from task-irrelevant information, and
to test whether it could be used to predict individual differences
in cognitive control outside the scanner. Participants (n = 62)
completed two fMRI Word–Picture Stroop tasks, one containing
emotional stimuli and one containing non-emotional stimuli, as
well as a behavioral out-of-scanner Color–Word Stroop task at
each of two time points. In a departure from the traditional
approach of using a single task contrast to index neural activa-
tion across all ROIs, the current study utilized specific ROI by
contrast pairings selected based on the specific level of control
hypothesized by the cascade-of-control model to occur within
that region. In addition, data across both tasks and both time
points were combined to create composite measures of neural
activation and of behavior. Consistent with the cascade-of-
control model, individual differences in brain activation for spe-
cific contrasts within each of the three ROIs were associated
with behavioral interference on the standard Color–Word
Stroop task. Testing of alternative models revealed that these
brain–behavior relationships were specific to the theoretically
driven ROI by contrast pairings. Furthermore, such relationships
were not observed across single-task and single-time point mea-
sures, but instead emerged from the composite measures.
These findings provide evidence that brain activation observed
across multiple regions of frontal cortex, each of which likely
exerts cognitive control in a differential manner, is capable of
predicting individual differences in behavioral performance. ■
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
5
5
7
8
1
2
0
7
7
7
2
2
/
j
o
c
n
_
a
_
0
1
9
7
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
INTRODUCTION
Cognitive control allows an individual to prioritize and act
on information that is goal-relevant. Such goal-directed
behavior is implemented by neural activation in distrib-
uted regions and networks throughout the brain, which
serve to bias toward task-relevant processing and protect
against interference from competing information. The
goal of the current study is to test predictions made by
the cascade-of-control model (Banich, 2009, 2019), a neu-
rocognitive model that posits that cognitive control is
enacted via multiple regions of frontal cortex that each
resolve unique levels of interference in a cascade-like man-
ner, and to utilize this model to predict individual differ-
ences in behavioral performance outside of the magnet.
Specifically, this model asserts that different regions of
pFC play somewhat distinct roles in exerting control, and
that the degree to which a task set is effectively imple-
mented by lateral prefrontal regions determines the
amount of control that must be enacted at later stages
along the cascade. Using this model as a guiding theoretical
framework, we test the ability of multitask composite
measures of neural activation, selected on the basis of the
1University of Colorado Boulder, 2Brandeis University,
Waltham, MA, 3University of Illinois–Urbana Champaign
© 2023 Massachusetts Institute of Technology
cascade-of-control model, to predict individual differences
in out-of-scanner behavioral performance. We first present
the tenets of the cascade-of-control model and then dis-
cuss its use to examine brain–behavior relationships.
Cascade-of-control Model
Substantial research has identified regions of both lateral
and medial pFC as playing a critical role in cognitive con-
trol (Derrfuss, Brass, Neumann, & von Cramon, 2005).
One model designed to explicitly outline the neural sys-
tems required for Stroop task performance is the
cascade-of-control model (Banich, 2009, 2019). In the clas-
sic Stroop task (Stroop, 1935), participants are presented
with words printed in different ink colors and must
respond based on the ink color in which a word is written,
rather than on the meaning of the word. This is more dif-
ficult and requires more control on incongruent trials, in
which the word conflicts with the ink color, (e.g., “RED”
printed in blue ink) as compared with congruent trials,
in which no conflict exists (“RED” printed in red ink). As
discussed in Banich (2019), conflict can arise at many
levels in the Stroop task: between the more automatic,
but task-irrelevant, process of word reading as compared
with ink-color identification; between the meaning of
Journal of Cognitive Neuroscience 35:5, pp. 781–801
https://doi.org/10.1162/jocn_a_01977
“red” as compared with the meaning of “blue”; and
between responses linked to red as opposed to blue.
The cascade-of-control model posits that specific prefron-
tal regions play more prominent roles than others in
resolving conflict at these different levels. In particular,
the model proposes that abstract task set implementa-
tion occurs within inferior frontal junction, biased selec-
tion among working memory representations within
mid-dorsolateral pFC (DLPFC), and late-stage response
selection within mid-cingulate regions.
Furthermore, the cascade-of-control model aims to pro-
vide an integrative account of how these regions interact
in a cascade-like manner to overcome interference from
task-irrelevant information. Central to this model is the
idea that control is implemented at multiple levels of
interference that span stimulus inputs to motor outputs.
Furthermore, each level of interference is resolved by a
different cognitive operation that is enacted, to a degree,
by an individual brain region. Critically, ineffective control
enacted at early stages within the cascade may be com-
pensated for by downstream brain regions, effectively
sparing behavioral outcomes. Stated more concretely,
the behavior of individuals who maintain a task set poorly
may only suffer if downstream regions are unable to “pick
up the slack.” As such, it is important to consider the role
of multiple brain regions in predicting behavior, rather
than a single brain–behavior relationship in isolation.
In a departure from the traditional approach of using a
single condition contrast (e.g., incongruent vs. congruent
trials) to index neural activation across multiple ROIs
regardless of their proposed role in resolving interference,
we determine the appropriate task contrast for each indi-
vidual ROI on the basis of the cascade-of-control model.
This novel approach of using unique ROI by contrast pair-
ings is driven by the assertion that the nature of Stroop
interference differs across levels of control and allows us
to capture the activation within each ROI that is relevant
to the specific cognitive process hypothesized to occur
within that region. In the following section, we discuss
the role of each ROI in implementing control during a
standard Color–Word Stroop task. We describe the spe-
cific level of control that is engendered within each ROI
as posited by the cascade-of-control model (Banich,
2009, 2019) and provide the rationale for the chosen
ROI by contrast pairing. A discussion of the evidence that
these regions play the specific role ascribed to them is
beyond the scope of the current introduction but can be
found in detail in Banich (2019).
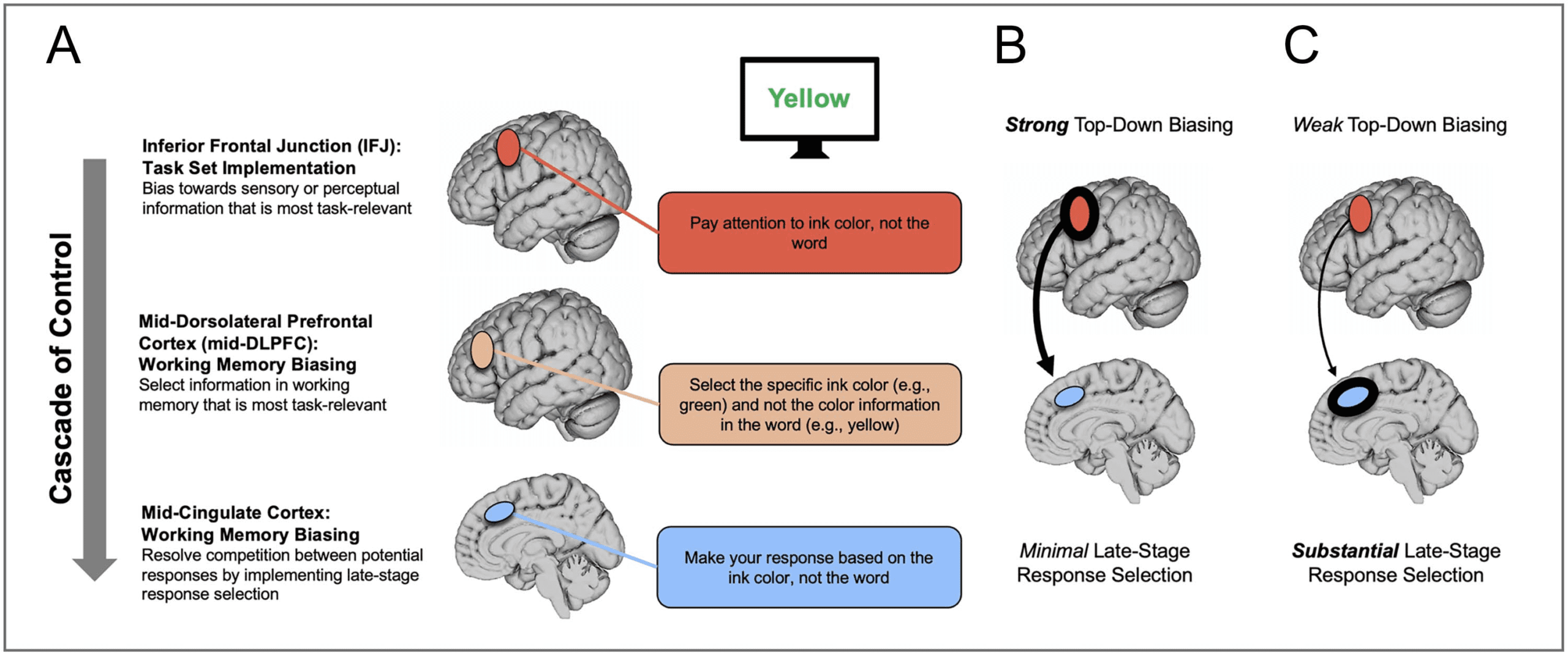
Inferior Frontal Junction: Task Set Implementation
First along the cascade is inferior frontal junction (IFJ), a
region of lateral pFC that engenders control by imple-
menting a task set that serves to bias processing toward
task-relevant and/or away from task-irrelevant processing
in sensory and perceptual regions of posterior cortex. In
the context of the Color–Word Stroop task, IFJ would
implement up-regulation of color-related processing
and/or down-regulation of word-related processing. Such
biasing occurs regardless of the current trial type and so is
present during incongruent (INC) and congruent (CON)
trials, as well as the neutral (NEU) trials also used in the
Figure 1. (A) The cascade-of-control model (Banich, 2009, 2019) outlining the brain regions that are involved in controlling interference in the Stroop
task. For an incongruent trial, such as the word “Yellow” written in green ink, control is implemented via a cascade. First, regions of IFJ enact an abstract task
set that serves to bias sensory and perceptual processing toward the task-relevant color information and/or away from the task-irrelevant word information.
Next along the cascade, mid-DLPFC regions select the task-relevant information that should be maintained in working memory (e.g., green not yellow).
At the final stage considered within the current study, caudal regions of mid-cingulate bias late-stage response selection toward the task-appropriate
response (e.g., button press for green). Importantly, the degree to which one region is active in controlling Stroop interference depends on how well
control has been implemented at prior points in the cascade. (B) Example of strong top–down biasing via IFJ. (C) Example of weak top–down biasing via IFJ.
782
Journal of Cognitive Neuroscience
Volume 35, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
5
5
7
8
1
2
0
7
7
7
2
2
/
j
o
c
n
_
a
_
0
1
9
7
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
current study, which in the context of the Color–Word
Stroop task would be trials in which the word is not
related to color. In the context of the current study, we
index relevant activation by contrasting all three trial
types (incongruent, congruent, and neutral) with fixation
(Inc + Con + Neu > Fix). This logic is consistent with
others who have argued that any regions involved in
setting a task set should be consistently observed across
task conditions (e.g., Dosenbach et al., 2006). Although
it is true that other regions may be activated by this con-
trast (e.g., perceptual processing regions), we highlight
the role of IFJ in implementing a task set based on our
prior work (e.g., Banich et al., 2000) and other findings
that this region plays a role in top–down biasing (Baldauf
& Desimone, 2014; Muhle-Karbe, Andres, & Brass, 2014;
Brass, Derrfuss, Forstmann, & von Cramon, 2005). (For
other viewpoint on other brain regions that may play a
major role in top–down biasing, see Dosenbach et al.,
2006.)
Mid-DLPFC: Working Memory Biasing
Next along the cascade, mid-DLPFC is posited to bias
selection toward the task-relevant information in working
memory, which in the case of the Color–Word Stroop task
would be the identity of the task-relevant ink color. Such
biasing is not required during neutral trials (e.g., “TABLE”
written in red ink), as the task-irrelevant word identity
does not contain task-relevant color information. In
contrast, task-relevant information is present in the
task-irrelevant dimension during both incongruent and
congruent trials, because in both cases the word names
a color. Thus, neutral trials are contrasted with both
incongruent and congruent trials to isolate relevant acti-
vation in mid-DLPFC (Inc + Con > Neu).
Mid-cingulate: Late-stage Response Selection
Finally, late-stage response-related selection is posited to
be implemented by mid-cingulate, which in the case of the
Color–Word Stroop task would bias processing toward the
motor response associated with the task-relevant word.
Relevant activation is defined by the contrast between
incongruent trials in which the task-relevant and task-
irrelevant information map to different responses, and
congruent trials in which both dimensions map to the
same response. This contrast likely also captures addi-
tional, non-response-related interference including
semantic interference. However, prior work has demon-
strated that effects within mid-cingulate are specific to
incongruent trials in which the task-irrelevant information
is response-eligible, suggesting that this interference is
primarily driven by motor mappings and not semantics
(Milham & Banich, 2005; Milham et al., 2001).
The cascading nature of this system dictates that the
successful enactment of an appropriate task set by IFJ will
determine the degree of control required during late-stage
selection in mid-cingulate. Consistent with this model, in a
combined fMRI and ERP Stroop task, Silton et al. (2010)
found that the relationship between early IFJ activity
(300–400-msec post-stimulus presentation) and behav-
ioral performance was mediated by late ACC activity
(520–680 msec). Furthermore, a significant interaction
between IFJ and ACC activity suggested that ACC mainly
influences behavioral performance only when IFJ is not
optimally engaged. Alternative models, such as those in
which early ACC activity was assumed to influence later
IFJ activity, could not predict behavioral performance. In
the next section, we discuss the promise and pitfalls of
examining brain–behavior relationships within the context
of functional neuroimaging studies.
Individual Differences and Brain Activation
fMRI offers a potentially powerful tool to link brain activa-
tion to behavioral outcomes. fMRI has been used exten-
sively to identify brain regions and networks that underlie
a wide range of cognitive processes including those that
support cognitive control. Brain regions are assumed to
support a particular mental process when, across a group
of participants, activation is higher during engagement in
a task condition that requires that process as compared
with a control condition that does not (e.g., conflict vs.
no conflict). For a region to be identified through this
approach, activation differences must be consistently
observed across participants, such that the average differ-
ence is reliably distinguishable from zero. There are two
limitations, however, with these group averaged data.
First, they do not identify which regions may contribute
most directly to the accuracy or speed of a particular
mental process that is being examined. Nor, because they
are group averages, do they provide insights into how
activation of such regions may influence the ultimate
behavioral performance on the task being performed,
nor how individual differences in behavioral characteris-
tics (e.g., level of cognitive control ability) may relate to
variations in brain activation.
These issues are important because research indicates
that there are stable individual differences across a wide
range of cognitive functions including executive functions
such as inhibition, working memory, and task shifting
(e.g., Friedman et al., 2016), as well as long-term memory
(e.g., Unsworth, 2019), language abilities (e.g., Kidd,
Donnelly, & Christiansen, 2018), and attentional pro-
cessing (e.g., Chechlacz, Gillebert, Vangkilde, Petersen,
& Humphreys, 2015). For example, Friedman et al.
(2008, 2016) used structural equation modeling to iden-
tify a Common Executive Function (Common EF) latent
factor that is hypothesized to reflect the ability to main-
tain and manage task goals as well as bias processing in
accordance with those goals. In an analysis of EF stability
in 840 individuals across a 6-year period, Friedman et al.
(2016) found a cross-wave correlation of .86 for the latent
Common EF factor and a cross-wave correlation of .45
Smith et al.
783
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
5
5
7
8
1
2
0
7
7
7
2
2
/
j
o
c
n
_
a
_
0
1
9
7
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
more specifically for Stroop interference, suggesting that
individual differences in Common EF are stable across
time. Suggesting that this factor is linked to real-world
performance, Common EF has been linked to self-
reported EF, including the ability to manage and retrieve
goals (Gustavson, Miyake, Hewitt, & Friedman, 2015).
Such findings have prompted a search for the neural
underpinnings of observed individual differences because,
in part, understanding the neural underpinnings of
individual differences in cognitive control is a crucial
component of better predicting, diagnosing, treating,
and ultimately preventing brain disorders (Elliott, Knodt,
& Hariri, 2021). The typical approach used to investigate
such relationships is to examine the covariation between
the magnitude of differences in brain activation between a
critical contrast of interest and the magnitude of a behav-
ioral outcome of interest. In the case of the Stroop task,
behavioral performance is typically measured by a RT
interference score that is calculated as the difference in
RT on the control-demanding incongruent trials com-
pared with the less demanding congruent trials. Brain
regions that underlie individual differences in Stroop
interference would thus be identified as those in which
activation differences are significantly associated with the
size of the RT interference.
In this approach, the individual differences variable
(e.g., RT interference score, level of Common EF ability)
is typically included as a covariate in the general linear
model (GLM) analysis. However, such an approach has
been criticized as likely to lead to false positive findings
( Vul, Harris, Winkielman, & Pashler, 2009) and may
require very large sample sizes (> 1000; Marek et al.,
2022). However, others have argued that with reliable sin-
gle time point measures of brain and behavior, samples
sizes in the range of about 80 individuals are adequate
for detecting brain–behavior relationships (Grady, Rieck,
Nichol, Rodrigue, & Kennedy, 2021). Moreover, whereas
the focus with regard to statistical power had generally
focused on the number of participants in a study (n ),
which influences between-subjects variance, recent dis-
cussions of brain–behavior relationships in neuroimaging
studies have noted that statistical power is equally influ-
enced by the robustness of the measures obtained on each
individual, which influences within-subject variance (Chen
et al., 2022; Baker et al., 2021) and that equivalent power
can be obtained by different combinations or pairings of
between- and within-subject variance. One example of
an attempt to reduce within-subject variance are studies
employing the Midnight Scan Club data set (Gordon
et al., 2017), which involve 5 hr of rest data and 6 hr of
task data on a single individual to obtain stable individual
differences measures of both brain and behavior.
In the current study, we have considered these two
paths to achieving greater reliability in brain–behavior
correlations (Gratton, Nelson, & Gordon, 2022)—a larger
N or deeper phenotyping to obtain a more stable within-
subject measure—and take an intermediate approach.
This intermediate approach involved obtaining measures
both of brain and behavior at two distinct time points
assessed 2 years apart and averaging across them, while
also obtaining measures of brain activation across more
than one task, as discussed in more detail below.
There are several advantages to using composite mea-
sures composed of data collected across multiple tasks
and multiple time points. By varying task-related attributes
such as stimulus category and valence across the two fMRI
paradigms, we reduce the degree to which the composite
measures may reflect stimulus-specific processing. Simi-
larly, by combining measures that were collected at two
different time points, we reduce the likelihood that the
composite measures reflect state-specific factors such as
alertness, which is known to impact cognitive processes
(e.g., Killgore, 2010). In addition, composite measures
of brain activation have been shown to have higher reli-
ability than single task measures (e.g., Sheu, Jennings, &
Gianaros, 2012).
Another issue with regard to assessing brain–behavior
relationships is that it is not always obvious what particular
contrast or comparison of conditions should be used to
assess brain activation. Typically, a particular contrast is
employed for all brain areas under investigation, such as
a contrast in activation between incongruent and congru-
ent trials in the Stroop task, and then a whole-brain GLM
for this contrast with the covariate of an individual differ-
ences variable (e.g., level of executive function) is per-
formed. Yet, if one has reason to believe that different
brain regions may be more involved in particular subpro-
cesses of a given task, then such an approach is not ideal.
Rather, it may be more fruitful to utilize different contrasts
for different brain regions to examine brain–behavior rela-
tionships. To do so requires a theoretical model of the
predominant role of each major brain region involved in
a task. In the current study, we take such an approach to
guide our investigation using the cascade-of-control
framework.
Current Study
The current study investigates whether individual differ-
ences in behavioral Stroop interference on a standard
Color–Word Stroop task given outside the magnet are
predicted by composite measures of task-related activa-
tion in three regions of frontal cortex: IFJ, mid-DLPFC,
and mid-cingulate during performance of two Word–
Image Stroop tasks. We evaluate the brain–behavior rela-
tionships predicted by the cascade-of-control model,
focusing specifically on the theoretically derived ROI by
contrast pairings that reflect the cognitive processes
hypothesized to occur within each region. To do so, we
test the following theoretically motivated hypotheses:
(1) Significant associations will be observed between
behavioral interference and activation within each ROI.
(2) Because of the nature of the control that is differen-
tially engaged for each ROI, associations between ROI
784
Journal of Cognitive Neuroscience
Volume 35, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
5
5
7
8
1
2
0
7
7
7
2
2
/
j
o
c
n
_
a
_
0
1
9
7
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
activation and behavioral interference will be contrast-
specific. (3) Consistent with the findings of Silton et al.
(2010), we expect to observe a significant interaction
between activation in IFJ and mid-cingulate, such that
the relationship between mid-cingulate activation and
behavioral performance is strongest when IFJ activation
is low. We additionally hypothesize that this interaction
will also be contrast-specific.
We tested these hypotheses in a sample of 62 adult
women who participated in both fMRI and behavioral test-
ing sessions as part of a larger longitudinal study. Although
smaller than the number generally recommended to reach
ideal levels of effect stability for a single time-point obser-
vation (e.g., Grady et al., 2021; Yarkoni, 2009), this sample
size is substantially larger than is typical of fMRI studies
(Poldrack et al., 2017). At each of two time points sepa-
rated by approximately 2 years, participants completed
two fMRI Word–Picture Stroop tasks—one consisting of
affectively neutral stimuli, and the other emotional
stimuli—as well as a behavioral Color–Word Stroop task.
We then use composite measures comprising all available
data for a given individual to index brain activation and
behavior (see Methods section).
To the degree that meaningful individual differences
exist within functional brain activation during the resolu-
tion of interference, these differences should predict
behavioral performance outside of those tasks during
which the activation was measured. This approach is use-
ful because measuring activation and performance during
the same task may produce relationships between brain
activation and performance that are driven by time on task
(Grinband et al., 2011) or the same type of stimulus- or
state-specific processing as previously mentioned. To this
end, we chose to index behavioral interference using a
separate Stroop task that was administered on a different
day. We sought to minimize the likelihood that observed
brain–behavior relationships might be driven by task-
related attributes by selecting a behavioral task that
indexes cognitive control, the standard Color–Word
Stroop task, and then deriving brain activation from dis-
tinct but related word-image Stroop tasks. One important
distinction between the tasks is that word reading serves
as the task-relevant process/dimension in the fMRI tasks
but as the task-irrelevant process/dimension in the behav-
ioral task. As such, any observed relationships cannot be
driven by commonalities between the nature of the task-
relevant information or conversely the task-irrelevant
information. The tasks also differed substantially in that
word reading was paired with task-irrelevant images in
the fMRI tasks and with task-relevant ink color in the
behavioral task. Finally, the tasks differed with respect to
response type, as the fMRI tasks required finger move-
ment whereas the behavioral task required a verbal
response. As such, the metrics used to examine cognitive
control behaviorally and in the magnet are distinct, mak-
ing it unlikely that any association observed results from
stimulus or response similarity.
In summary, the current study expands upon prior work
in several important ways. First, although the cascade-of-
control model describes the role of multiple brain regions,
the current study is the first to simultaneously consider
three ROIs and their association with behavior. Second,
this is one of very few studies that utilize a theoretical neu-
rocognitive model to identify and test contrast-specific
brain–behavior relationships, as compared with the tradi-
tional approach of using a single contrast across ROIs (e.g.,
Tang, Etzel, Kizhner, & Braver, 2021). Finally, we use a
number of approaches to increase the reliability and stabil-
ity of our indices of individual differences related to brain
and behavior.
METHODS
Participants
Data from unrelated adult women were collected as part of
the Colorado Cognitive Neuroimaging Family Emotion
Research (CoNiFER) study, a two-time-point longitudinal
assessment of adolescent children and their parents. Only
female parents were included in the present analyses as an
insufficient number of male parents (n = 5) chose to par-
ticipate. All participants had previously taken part in the
Genes, Environment and Mood study (R01MH077195,
Hankin, P.I.) and an associated follow-up study
(R21MH102210, Hankin, P.I.). Families were drawn from
an unselected community sample, recruited from the
Denver metro area via public schools and using direct
mail to target zip codes to maximize demographic and
socioeconomic diversity. For details of the two samples
and studies, see Hankin et al. (2015) and Snyder, Friedman,
and Hankin (2019).
Participants were free of history of neurological insult,
were fluent English speakers, and did not report having
dyslexia or difficulty reading. Informed consent was
obtained from all participants, and all procedures were
approved by the University of Colorado institutional
review board. All inclusion/exclusion criteria were
established before data analysis. fMRI data were subject
to the following exclusion criteria: Low accuracy (accuracy
below 60% on incongruent, congruent, or neutral trials),
excessive signal dropout in frontal or temporal regions,
excessive motion (greater than 3 mm), or failure to meet
multiple criteria.
Time Point 1 (TP1): 70 women participated at TP1. Six
participants were excluded from both fMRI tasks because
of low accuracy (n = 2), excessive signal dropout (n =
2), or failure to meet multiple criteria (n = 2). No partic-
ipants were excluded due to excessive motion. An addi-
tional two participants did not complete the Color–Word
Stroop task and were excluded from analysis. Data from
six participants were included for the emotional task but
excluded from the non-emotional task because low accu-
racy (n = 4), or excessive movement (n = 2). Data from
three participants were included for the non-emotional
Smith et al.
785
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
5
5
7
8
1
2
0
7
7
7
2
2
/
j
o
c
n
_
a
_
0
1
9
7
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
task but excluded from the emotional task because of
excessive movement greater than 3 mm (n = 3). No data
were excluded due to low accuracy, excessive signal
dropout, or failure to meet multiple criteria. These
exclusions resulted in usable data from 56 participants
in the non-emotional task, 59 participants in the
emotional task, and usable data across both tasks for
53 participants.
Time Point 2 (TP2): 55 women participated at TP2. Five
participants were excluded from both fMRI tasks because
of low accuracy (n = 1), excessive signal dropout (n = 1),
excessive movement, or failure to meet multiple criteria
(n = 1). Data from 10 participants were included for the
emotional task but excluded from the non-emotional task
because of low accuracy (n = 5), excessive movement
(n = 4), or failure to complete the task (n = 1), No data
were excluded due to excessive signal dropout or failure to
meet multiple criteria. Data from two participants were
included for the non-emotional task but excluded from
the emotional task because of low accuracy (n = 1), or
excessive movement (n = 1). No participants were
excluded due to excessive signal dropout or failure to
meet multiple criteria. These exclusions resulted in
usable data from 40 participants in the non-emotional
task and 48 participants in the emotional task, and usable
data across both tasks for 38 participants.
Sixty-seven participants had usable data for at least one
of four fMRI observations (two time points × two tasks)
and data from at least one Color–Word Stroop task ses-
sion. Of these participants, 32 had usable data at all four
fMRI observations, 10 had usable data at three observa-
tions, and 20 had usable data at two observations. Five
had usable data for only one observation. Ideally, each par-
ticipant would have usable data at all four observation
points (two tasks × two time points). However, to main-
tain a sufficient sample size, all participants with usable
data for at least two fMRI tasks and at least one behavioral
task were included in the analysis. Thus, the final sample
consisted of 62 participants. Of these participants, 45
(73%) had usable data from at least one fMRI task at both
TP1 and TP2, and 61 (98%) had usable data from at least
one time point for both the emotion and non-emotional
tasks. The average age across both time points ranged
from 36.5 years to 65.6 years (M = 49.5 years, SD = 6.2
years).
A power analysis employing G*Power 3.1.9.4 (Faul,
Erdfelder, Lang, & Buchner, 2007) estimated an achieved
power of 1 – β = .85 (α = .05) to detect a small to
medium effect size ( f 2 = 0.15) for each regression
parameter in the primary study analysis. Although larger
sample sizes have been recommended to reach ideal
levels of effect stability in brain–behavior relationships
(e.g., Grady et al., 2021; Yarkoni, 2009), the multitask
composite measure approach utilized in the current
study required a compromise between the number of
participants and the quantity of data collected from each
participant.
Procedure
At each of two time points separated by approximately 2
years, participants completed a neuroimaging session as
well as a behavioral session. The neuroimaging and behav-
ioral sessions typically occurred within 2 months of one
another, but, in rare cases, the sessions were separated
by up to 6 months. The neuroimaging sessions consisted
of three fMRI tasks, including both an emotional and a
non-emotional Stroop task, as well as a verb generation
task that is not of interest for the current report. The
non-emotional Stroop task always preceded the emotional
task so as to not confer emotional meaning to neutral
face stimuli used in the non-emotional task. Resting state
and spectroscopy data were also collected but are not
included in the present analyses. Behavioral testing ses-
sions included a battery of questionnaires and cognitive
tasks. These included a Color–Word Stroop task, which is
the behavioral outcome of interest in the current study,
as well as a choice RT task, which is used as a control
variable.
Measures
Neuroimaging Tasks
Emotional word–emotional face Stroop task. The goal
of this task was to identify those brain regions that are
activated when cognitive control must be exerted in the
context of semantic and response conflict between two
categories of emotional items. Individuals made a seman-
tic decision about whether words were positively valenced
(i.e., “cheerful,” “joy,” “happy,” “delighted”) or negatively
valenced (i.e., “furious,” “mad,” “angry,” “rage”), and indi-
cated their response with either a right or left button
press. The words were superimposed on three types of
pictures: faces with a happy expression, faces with an
angry expression, and cars. Stimuli used in the present
study were designed to be identical to those used with
the adolescent offspring of the participants (which is the
focus of another report). As part of the goal in the adoles-
cent sample was to examine the development of prefrontal-
amygdala connectivity as assessed by fMRI, angry and
happy faces were used as they show good reliability in this
regard (Haller et al., 2018). Cars, rather than neutral faces,
were used as neutral stimuli because our prior work sug-
gests that youth in the age range we tested show highly
similar responses to faces with neutral and emotional
expressions (Banich et al., 2019).
Faces were drawn from the NIMSTIM database (Tottenham
et al., 2009), using a set of 24 posers, with two angry
expression and two happy expression images drawn from
each poser. Each poser was seen four times, twice with an
angry and twice with a happy expression. Thirty-two car
images were drawn from Herzmann and Curran (2011).
For half of the trials, the word was placed in an upper
position with regard to the object, and on half the trials,
it was placed in a lower position with regard to the object.
786
Journal of Cognitive Neuroscience
Volume 35, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
5
5
7
8
1
2
0
7
7
7
2
2
/
j
o
c
n
_
a
_
0
1
9
7
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Varying the position of the word made it less likely that an
individual could narrow their focus of attention to just the
central portion of the display where the word would
appear so as to filter out the background image.
Three types of trials were constructed. On congruent
trials, task-relevant words were drawn from the same
semantic category (e.g., a positively valenced word super-
imposed on a happy face). On incongruent trials, the
task-relevant word was drawn from the opposite semantic
category than the task-irrelevant face (e.g., a negatively
valenced word superimposed on a picture of a happy
face; a positively valenced word superimposed on an angry
face). On neutral trials, the task-irrelevant object was a car,
which has no semantic or response relationship to either
positively valenced or negatively valenced words. Each
stimulus was displayed for a fixed period of 1380 msec
(3 repetition times [TRs]), followed by a fixed intertrial
interval of 480 msec (1 TR). Sample stimuli are shown in
Figure 2.
Similar to designs used previously in our laboratory
(Andrews-Hanna et al., 2011; Banich et al., 2009), this
study was a mixed blocked/event-related design. Blocks
consisted of trials that were, on average, two thirds specific
to that block (e.g., incongruent) and one third a set of neu-
tral trials that were common across blocks, referred to as
neutral frequent trials. In the neutral block, two thirds of
trials were a set of neutral infrequent trials, and one third
the same neutral trials that were common across blocks
(i.e., neutral frequent). Six triads of blocks were created
(e.g., INC, CIN), and these were flanked in a run by fixation
blocks (Fix). Fixation blocks lasted for 23.04 sec (48 TRs)
each and served as the baseline for the contrasts discussed
below. Each run consisted of three triads, so, for example,
a run might consist of FINCFNICFCINF. Two orders of
blocks were employed, with button responses counterba-
lanced across participants.
The exact number of trials per condition differed
slightly. For half of the participants, one of the congruent
blocks contained two-third frequent neutral trials and one-
third congruent trials, whereas the other half of partici-
pants were presented with one of the incongruent blocks
consisting of two-third frequent neutral trials and one-
third incongruent trials. As a result, all participants
received 216 trials, with half of the participants seeing 44
incongruent and 48 congruent trials, and the other half
seeing 48 incongruent and 44 congruent trials, with all
participants seeing 48 neutral infrequent and 76 neutral
frequent trials.
Neuroimaging neutral word–neutral image Stroop task.
The goal of this task was to identify those brain regions
that are activated when cognitive control must be exerted
in the face of semantic and response conflict between two
categories of non-emotional items. This task was identical
to the emotional task (described above), except that the
stimuli were non-emotional. Individuals made a semantic
decision about whether words were face-related (i.e.,
“hair,” “nose,” “chin,” “eyebrow”) or house-related (i.e.,
“wall,” “roof,” “door,” “window”). The words were superim-
posed on three types of objects: faces with a neutral expres-
sion (same set of posers as used in the emotional task),
houses, and cars. Sample stimuli are shown in Figure 2.
Behavioral Color–Word Stroop Task
The goal of this task (described in more detail in Friedman
et al., 2016) was to index conflict between the prepotent
tendency to read color words and the instruction to name
Figure 2. Examples of stimuli used in the non-emotional and emotional tasks. On each trial individuals viewed an item with a word superimposed on top
of a picture and were required to make a category judgement (non-emotional: face vs. house, emotional: positive vs. negative) on the basis of the word.
Smith et al.
787
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
5
5
7
8
1
2
0
7
7
7
2
2
/
j
o
c
n
_
a
_
0
1
9
7
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
the color in which the word was printed. On each trial, a
colored stimulus (red, green, or blue) was presented on
a black background and remained on the screen until
the participant reported the color of the stimulus using
the voice-activated E-prime microphone, followed by a
1000-msec intertrial interval. Participants first completed
a block of 44 neutral trials (asterisks printed in colored
ink) after 10 practice trials. Then, to engage the prepo-
tency of word reading, participants completed a block of
44 congruent (word and color the same, e.g., “blue” in
blue ink) trials, again preceded by 10 practice trials. Partic-
ipants then completed a block of 62 incongruent trials
(word and color do not match, e.g., “blue” in red ink) with
no practice trials to minimize practice effects. Participants
then completed three mixed blocks, which are the focus of
the current analysis. These blocks consisted of all three
trial types intermixed, with 56 trials per block, for a total
of 168 trials (56 per condition).
For each subject and each trial type (e.g., mixed block
incongruent trials), outlying RTs were identified by
the Wilcox–Keselman trimming procedure ( Wilcox &
Keselman, 2003) and were excluded before calculating
means. Stroop interference was calculated as the mean
RT difference between the mixed block incongruent trials
and the mixed block neutral trials.
Control Variables
Because of the broad age range among participants, age was
included as a control variable. For participants with viable
imaging data at both TP1 and TP2, age was averaged across
the two time points. Age at a single time point was used for
participants with only one time point of imaging data.
To minimize the possibility that any observed relation-
ships between brain activation and Stroop interference
could be driven by a participant’s underlying processing
speed, all models additionally included a measure of
choice RT. Because the Stroop interference score is
indexed by an RT difference score that captures the RT
increase associated with more control-demanding trials,
baseline processing speed should generally be removed
from the score. However, in some cases, difference scores
may still be impacted by their component RTs (Chapman,
Chapman, Curran, & Miller, 1994). The choice RT task
(Snyder & Hankin, 2016) required participants to press
buttons with their right and left hands as fast as possible
when presented with left or right pointing triangles for
60 trials. The dependent variable is mean RT.
Neuroimaging
Image Acquisition
Data were acquired on a Siemens MAGNETOM PRISMA
3.0 Tesla scanner with a 32-channel head coil at the Inter-
mountain Neuroimaging Consortium on the campus of the
University of Colorado Boulder for all participants except
for 12 at the first time point for whom data were acquired
on the pre-upgrade version of the same magnet (TIM
TRIO). To reduce head motion during MRI data acquisi-
tion, foam padding was placed around participants’ heads.
Structural scans were acquired via a T1-weighted mag-
netization prepared rapid gradient echo sequence in
224 sagittal slices, with a TR = 2400 msec, echo time =
2.07 msec, flip angle = 8°, field of view = 256 mm, and
voxel size of .8 mm3.
Functional scans were acquired via multiband acceler-
ated T2*-weighted EPI (TR = 460 msec, echo time =
27.20 msec, flip angle = 44°, 56 interleaved axial slices
aligned along anterior–posterior commissure line, slice
thickness = 3 mm, field of view = 248 mm, multiband
accel. Factor = 8, gap = 1 mm, voxel size = 3.0 mm3).
Two runs were acquired for each task at each time point,
with each run consisting of 624 echo planar images, for
1248 images per task.
Preprocessing
fMRI preprocessing and GLM analyses were carried out
using the FSL suite (Smith et al., 2004). For the Stroop
tasks, the first 10 EPI volumes of each run were discarded
to allow the MRI scanner to reach steady-state stability.
Preprocessing included motion correction via ICA-based
Automatic Removal Of Motion Artifacts (ICA-AROMA)
(Pruim et al., 2015), an independent component analysis
method for removing motion, high-pass filtering (100 sec),
and brain extraction via Brain Extraction Tool (BET)
(Smith, 2002). Registration of EPI images into subject-
and standard-spaces was executed using FMRIB’s Linear
Image Registration Tool (FLIRT) ( Jenkinson, Bannister,
Brady, & Smith, 2002; Jenkinson & Smith, 2001). Individ-
ual subject EPI images were registered to that subject’s
magnetization prepared rapid gradient echo structural
image via linear Boundary-Based Registration (Greve &
Fischl, 2009) and then registered to the MNI-152 tem-
plate via 12 degrees of freedom linear transformation.
The resulting EPI images were smoothed using an 8-
mm FWHM Gaussian smoothing kernel.
GLM Analyses
As blocked analyses can have up to 35% more power than
event-related analyses (Bandettini & Cox, 2000), blocked
contrasts were used in the GLM analyses. We have found
in our prior studies that blocked contrasts in variants of
the Stroop task enable individual differences to be detected
(e.g., Andrews-Hanna et al., 2011; Banich et al., 2009).
Because both the emotional and non-emotional tasks are
identical in design, the same models were applied to both
tasks.
FMRI Expert Analysis Tool (FEAT) within FSL was used
to model the data. First-level models were run separately
for each run of each task, at each time point, for each par-
ticipant. These models consisted of three explanatory
788
Journal of Cognitive Neuroscience
Volume 35, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
5
5
7
8
1
2
0
7
7
7
2
2
/
j
o
c
n
_
a
_
0
1
9
7
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
variables, one for each block type: incongruent (Inc),
congruent (Con), and neutral (Neu). Explanatory variables
modeled the time of the entire block (i.e., also including
neutral frequent trials). Fixation blocks (Fix) served as the
nonmodeled baseline. The following contrasts of interest
were created: Inc + Con + Neu > Fix, Inc + Con > Neu,
Inc > Con, Inc > Neu. In cases where multiple trial types
were combined within a contrast, average activation was
computed across the trial types. Fixed effects analysis was
used to estimate means across Run 1 and Run 2 of each task.
Finally, four higher-level analyses were conducted to exam-
ine group main effects for both tasks at both time points.
Selection of ROIs
Bilateral brain ROIs were defined by parcels from within
the Schaefer 400-area network parcellation (Schaefer
et al., 2018), which reflects neurobiologically meaningful
areas determined by resting-state fMRI across nearly
1500 participants. The 400-area parcellation was chosen
as it provides an intermediate level of resolution that
allows the areas of interest (e.g., IFJ) to be relatively iso-
lated to a given parcel.
For each of the three ROIs, the right and left hemi-
sphere parcels were determined according to the follow-
ing steps. First, a peak voxel for each ROI was identified in
the right hemisphere by the meta-analysis feature in Neu-
rosynth ( Yarkoni, Poldrack, Nichols, Van Essen, & Wager,
2011). The term “stroop” was used to identify regions of
IFJ that we posit maintain abstract task set representations
and mid-cingulate regions that we posit carry out response
selection in the face of conflicting information. To capture
the mid-DLPFC regions that we posit maintain the working
memory representation of specific items, the term “work-
ing memory” was used. In all cases, the Association Test
maps, which identify those regions that are preferentially
related to the search term, were used. One peak voxel for
each ROI was determined using FSL’s cluster function, and
the Schaefer parcel containing the peak voxel was chosen
to represent that ROI. The following right hemisphere
peak voxels (reported in Montreal Neurological Institute
coordinates) and parcels were identified: IFJ: 43, 12, 35,
parcel 350; mid-cingulate: 7, 21, 25, parcel 311; mid-
DLPFC: 42, 34, 30, parcel 351. The following three corre-
sponding left hemisphere parcels were then visually
identified: IFJ: parcel 141; mid-cingulate: parcel 107;
mid-DLPFC: parcel 142.
Calculation of Beta Weights
Beta-weights for percent signal change were used to
index brain activation. Beta-weights were calculated per
contrast and participant within each of the six parcels
using FSL’s means function. Bilateral indices of activation
were then calculated by taking the mean of the scaled
right and left hemisphere parcel beta-weights for each
of the three ROIs.
Approach and Analyses
Reliability of Brain Activation and Color–Word Stroop
Behavioral Interference
We first investigated the reliability of whole-brain activa-
tion across all 400 parcels to better understand patterns
of reliability across time points, tasks, and contrasts. Of
particular interest are the following four measures: TP1
reliability across tasks, TP2 reliability across tasks, non-
emotional task reliability across time points, and emo-
tional task reliability across time points. Reliability was
indexed by averaging across Fisher-transformed within-
subject correlations. For example, TP1 reliability was
computed by first calculating within-subject Pearson cor-
relations between activation in the 400 parcels during
the non-emotional task at TP1 with activation during the
emotional task at TP1, Fisher-transforming each correla-
tion value, averaging across subject-level correlations,
and, finally, reverse transforming the average correlation
value. The resulting correlation value can be interpreted
as a standard correlation coefficient indexing the strength
of the linear association between the two variables. Each
reliability measure was calculated separately for Inc +
Con + Neu > Fix, Inc + Con > Neu, and Inc > Con, result-
ing in 12 correlations (two time points × two tasks × three
contrasts).
The same reliability patterns were assessed within the
three ROIs. For this set of analyses, only a single Pearson
correlation was computed for each reliability measure
because each participant had only one activation value
per parcel per contrast. Rather than computing reliability
across all contrasts for all ROIs, we constrained these
analyses to our a priori ROI by contrast pairings.
Because the behavioral Stroop task was performed only
once at each time point, reliability was assessed by a single
correlation between TP1 and TP2.
Conceptual Models
To evaluate the predicted relationships between individ-
ual differences in composite measures of brain activation
and behavioral interference, we ran a single regression
model containing each composite measure of brain activa-
tion as well as the two control variables (age and choice
RT) as predictors. We additionally explored whether the
same brain–behavior relationships were present when
considering only a single observation, that is, one task at
one time point. To do so, we ran four additional models in
which the composite measures were replaced with
measures calculated within each of the individual task and
time points. All regressions were performed in R Version
4.2.2 (R Core Team, 2022).
Composite Measures of Brain Activation
To obtain more stable estimates of individual differences
in brain activation, signal change in each parcel was
Smith et al.
789
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
5
5
7
8
1
2
0
7
7
7
2
2
/
j
o
c
n
_
a
_
0
1
9
7
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
averaged across the emotional and non-emotional Stroop
tasks at both TP1 and TP2. Because participants differed in
their number of valid observation points, averages were
computed from between one and four observation points.
Participants with only one valid observation point (n = 5)
were excluded from analyses. The same method was
applied to the behavioral Stroop interference measure as
well as control variables, with averages computed across
TP1 and TP2. Participants with valid data at either TP1 or
TP2 were included in analyses.
Analysis Model
The regression equation used to test the cascade-of-
control model is presented in Equation (1).
Y ¼ β
Þ
þ β
ð
1 IFJInc þ Con þ Neu > Fix
0
2 Mid − DLPFCInc þ Con þ Neu
ð
þ β
(cid:3)
(cid:1)
þ β3 Mid − CingulateInc > Con
(cid:1)
þ β
4 IFJInc þ Con þ Neu > Fix
ð
ð
þ β
6 Age
5 ChoiceRT
Þ þ β
Þ
(cid:2) Mid − CingulateInc > Con
Þ þ ε
regard to the ROIs but assigned each of the ROIs one of
the following contrasts: Inc + Con + Neu > Fix, Inc +
Con > Neu, Inc > Neu, Inc > Con. This process resulted
in 64 possible contrast pairings (4 IFJ pairings by 4 mid-
DLPFC pairings by 4 mid-cingulate pairings).
We compared the goodness of fit of the cascade-of-
control model against each alternative model by comput-
ing the difference in Bayesian information criterion
(ΔBIC). ΔBIC is a commonly used criterion for selecting
among models and is the preferred metric when explana-
tory goodness of fit is desired over predictive accuracy
(Sober, 2002). ΔBIC was calculated by subtracting the
cascade-of-control model BIC from the alternate model
BIC, such that positive numbers indicate better fit for
the cascade model. Based on guidelines set by Raftery
(1995), we consider a difference greater than 10 to be
strong evidence of a meaningful difference.
(cid:3)
RESULTS
Behavioral Results
(1)
All variables were standardized such that the mean is
0 and the standard deviation was 1. Y represents the
outcome measure, Stroop interference. β1 captures the
expected increase in Stroop interference associated with
a 1 SD increase in activation of IFJ for an individual with
average mid-cingulate activation. β2 captures the interfer-
ence increase associated with 1 SD increases of mid-
DLPFC activation. β3 captures the expected interference
increase associated with a 1 SD increase in activation of
mid-cingulate for an individual with average IFJ activation.
β4 captures the interaction between IFJ and mid-cingulate
activation, which is a prominent feature of the cascade-of-
control model, which posits that cingulate activation is
influenced by the degree of top–down biasing by IFJ
(e.g., Silton et al., 2010). β5 and β6 capture the two control
variables, age and choice RT, respectively.
Bootstrapped Confidence Intervals
To obtain a quantitative measurement of the uncertainty
around each beta estimated by the model above, we con-
structed bootstrapped confidence intervals. Specifically,
we ran the model across 10,000 iterations in which we ran-
domly sampled, with replacement, 62 data points from our
data set (n = 62). We then used the set of beta estimates
extracted across the 10,000 iterations to generate a sam-
pling distribution and confidence interval for each effect.
To ensure that each of the tasks engendered cognitive
control as intended, we assessed patterns of behavioral
performance within each task at each time point. RT differ-
ence scores and accuracy data for the fMRI tasks and the
Color–Word Stroop task are presented in Table 1. Consis-
tent with expected task performance, one-sample t tests
indicated significant RT interference for each task at each
time point ( ps < .01).
Reliability
Whole Brain
To better understand the reliability of brain activation
across time points, tasks, and contrasts within our data,
we first investigated whole-brain reliability. The general
pattern of results is described here, and full results are pre-
sented in Table 2. Reliability differed substantially across
contrasts, with minimal differences observed in relation
to time point and task. Reliability was highest (.84–.89)
for the Inc + Con + Neu > Fix contrast, followed by the
Inc + Con > Neu contrast (.27–.32) and the Inc > Neu
contrast (.24–.33). Reliability was poor for the Inc > Con
contrast for both the time point and task measures (−.01
to .15); however, within-task reliability for both the non-
emotional task (.11) and emotional task (.15) was numer-
ically higher than the within-time point reliability at both
TP1 (.04) and TP2 (−.01).
Alternative ROI by Contrast Pairings
To test the specificity of the hypothesized ROI by contrast
pairings, we compared the cascade-of-control model to
models representing all alternative contrast pairings. To
test each pairing, we retained the model structure with
Within-ROI reliability largely followed the same contrast-
dependent pattern seen in the whole-brain results (see
Table 2). Reliability was highest within the IFJ ROI for
the Inc + Con + Neu > Fix contrast with significant cor-
relations observed across all task and time point reliability
Within-ROI
790
Journal of Cognitive Neuroscience
Volume 35, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
5
5
7
8
1
2
0
7
7
7
2
2
/
j
o
c
n
_
a
_
0
1
9
7
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Table 1. Behavioral Results
Time Point
TP1
Task
Non-emotional (Inc-Con)
Emotional (Inc-Con)
Color–Word (Inc-Neu)
TP2
Non-emotional (Inc-Con)
Emotion (Inc-Con)
Color–Word (Inc-Neu)
RT Interference
Inc Accuracy
M
55.56
39.83
158.46
48.30
44.89
147.68
SD
32.09
31.77
91.39
25.91
29.93
87.85
M
93.89
94.91
97.37
94.95
93.57
97.94
SD
5.09
4.24
2.92
3.62
5.15
2.58
RT is reported in milliseconds, and accuracy is reported as a percentage.
measures (.50–.80, ps < .001). For this contrast, within-
time point reliability appeared to be somewhat higher
(.65 and .80) compared with within-task reliability (.50
and .70). Reliability within the mid-DLPFC ROI for the
Inc + Con > Neu contrast varied considerably (−.18 to
.47), but there was no clear pattern with regard to time
point or task. Mid-cingulate reliability for the Inc > Con
contrast was poor (−.13–.16).
Color–Word
The Color–Word Stroop I-N RT interference score showed
moderate reliability from TP1 to TP2, r(48) = .56, p < .001.
Composite Measures Model
We first examined how well an individual’s behavioral
Stroop interference could be predicted by a model that
included composite activation for the contrast of interest
for each of the three ROIs as well as an interaction term
that examined whether activation in IFJ moderated the
relationship between mid-cingulate activation and inter-
ference. Model results are presented in Table 3. The
full model explained a significant amount of variance in
interference scores, F(6, 55) = 3.15 p = .010, R2 = .26,
R2
Adjusted = .17.
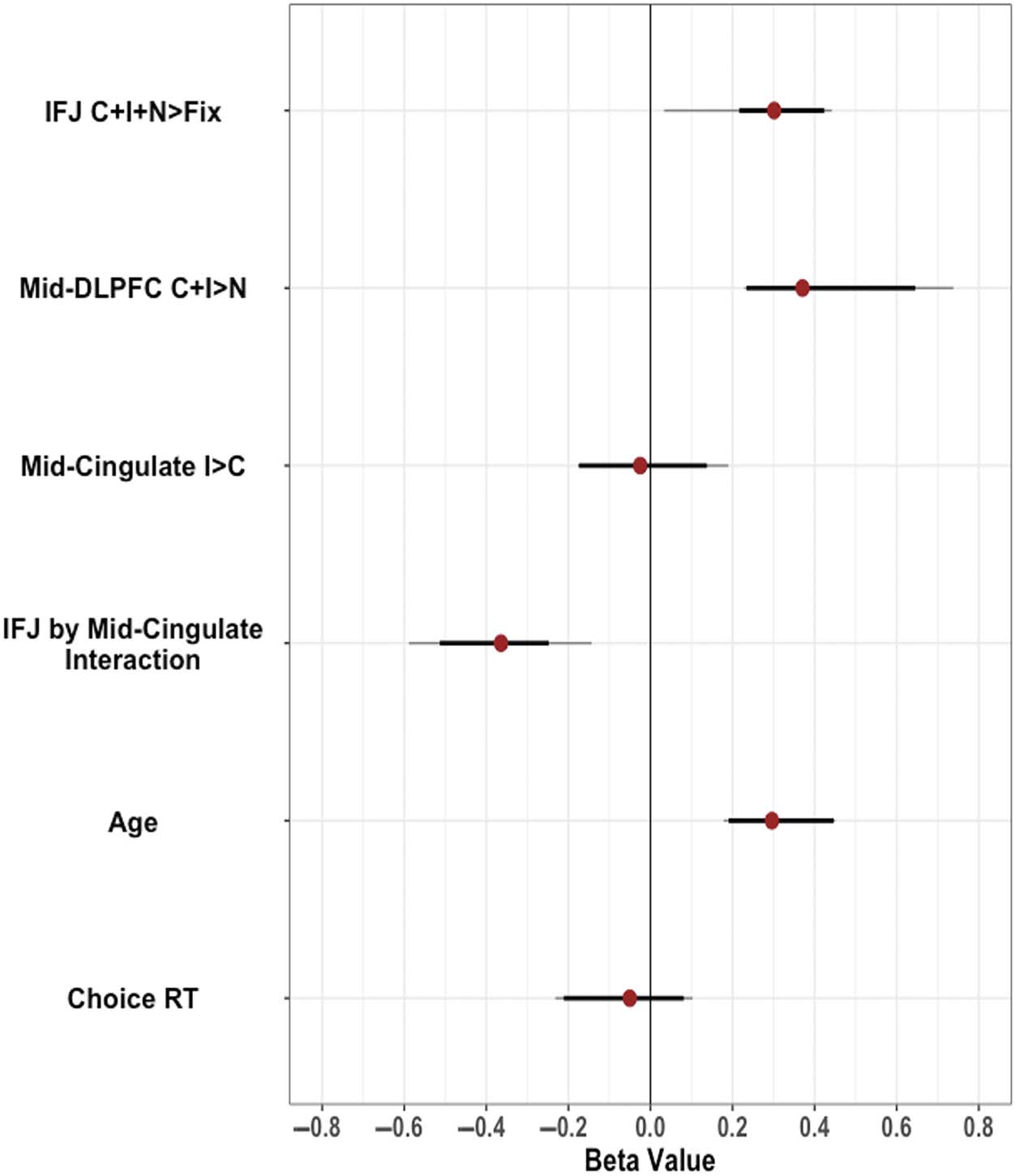
Mean activation in IFJ (Inc + Con + Neu > Fix) was
significantly positively associated with interference such
that greater activation was associated with more interfer-
ence (β = 0.31, t(55) = 2.43, p = .018). Mid-DLPFC (Inc +
Con > Neu) was also significantly positively associated
with interference (β = 0.34, t(55) = 2.73, p = .009). As
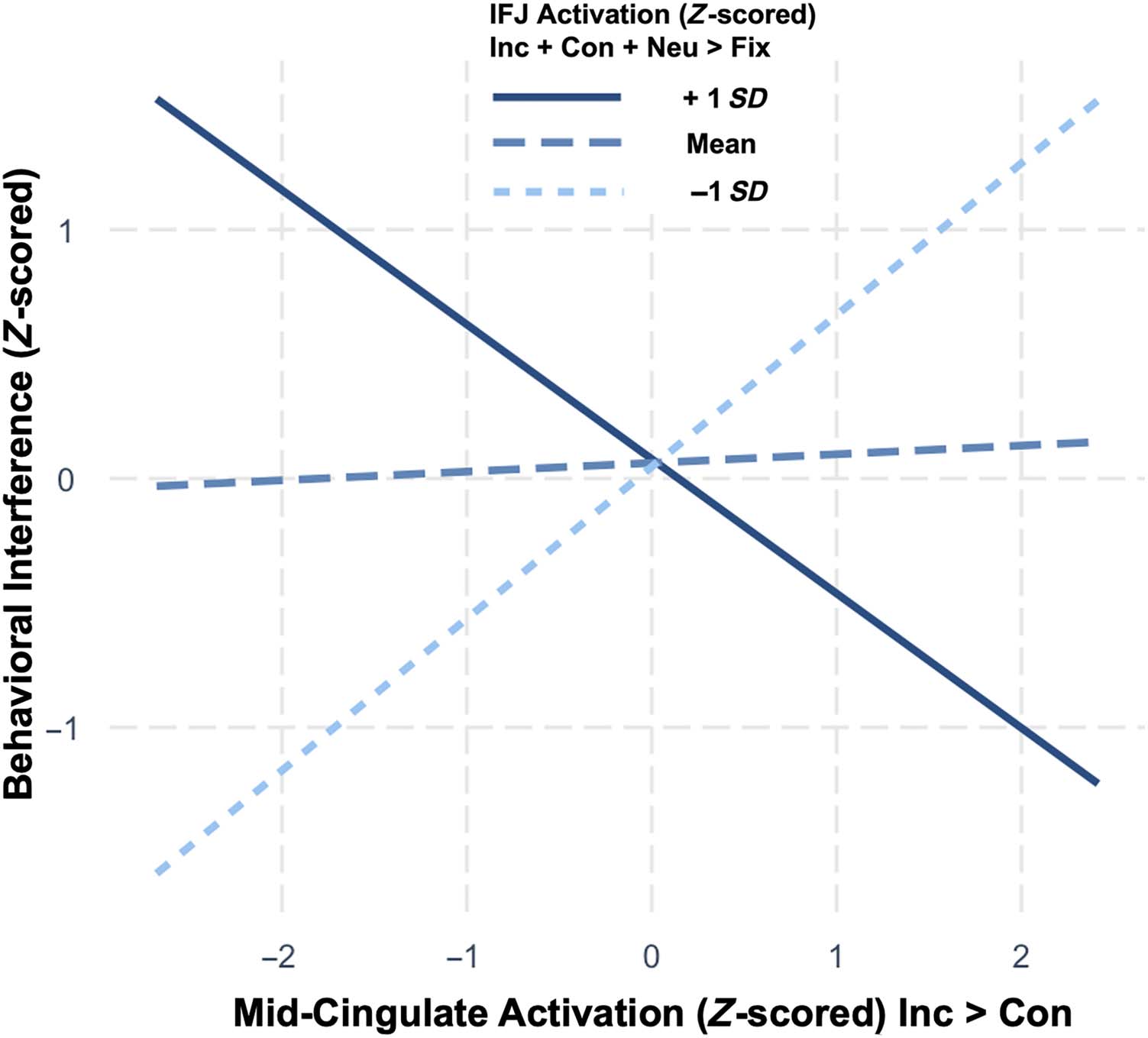
predicted, a significant interaction was observed such
that mid-cingulate activation positively predicted behav-
ioral interference when IFJ activation was low (β =
−0.36, t(55) = −2.83, p = .007). Analysis of simple slopes
Table 2. Reliability
Non-emotional Task ×
Emotional Task at TP1
Non-emotional Task ×
Emotional Task at TP2
TP1 × TP2 in
Non-emotional Task
n
53
38
34
Within-ROI Correlations
Whole-Brain Average Correlations
IFJ C + I +
N > Fix
Mid-DLPFC
C + I > N
Mid-Cingulate
I > C
C + I +
N > Fix
C + I +
N > Fix
C +
I > N
I > N
I > C
.65***
−.02
.80***
.47***
.70***
.35*
−.13
−.12
.16
.07
.89
.87
.88
.84
.27
.27
.30
.32
.28
.24
.28
.33
.04
−.01
.11
.15
TP1 × TP2 in Emotional
41
.50***
−.18
Task
Significance levels are not provided for whole-brain reliability measures as they reflect average correlations.
* p < .05. ** p < .01. *** p < .001. Smith et al. 791 l D o w n o a d e d f r o m h t t p : / / d i r e c t . m i t . e d u / j / o c n a r t i c e - p d l f / / / / 3 5 5 7 8 1 2 0 7 7 7 2 2 / j o c n _ a _ 0 1 9 7 7 p d . f b y g u e s t t o n 0 8 S e p e m b e r 2 0 2 3 Table 3. Cascade-of-Control Model Results Predictor Intercept IFJ C + I + N > Fix
Mid-DLPFC C + I > N
Mid-cingulate I > C
IFJ × Mid-cingulate interaction
Choice RT
Age
* p < .05. ** p < .01. *** p < .001. Beta 0.02 0.31 0.34 −0.02 −0.36 −0.09 0.27 SE 0.12 0.13 0.13 0.12 0.13 0.12 0.13 t-statistic 0.14 2.43 2.73 −0.17 −2.83 −0.69 2.17 95% CI −0.22, 0.25 0.06, 0.57 0.09, 0.60 −0.26, 0.22 −0.61, −0.10 −0.33, 0.16 0.02, 0.52 p .890 .018* .009** .863 .007** .495 .035* revealed a significant positive association between mid- cingulate activation and behavioral interference at low levels of IFJ activation (−1 SD; β = 0.33, t(55) = 2.03, p = .047), and a negative association at high levels of IFJ activation (+1 SD; β = −0.38, t(55) = −2.10, p = .040). Age positively predicted interference such that interference tended to be higher in older participants (β = 0.27, t(55) = 2.17, p = .035). Choice RT was not associated with interference ( p = .495). Bootstrapped Confidence Intervals Bootstrapping supported the findings reported in the pre- vious section. The 95% confidence intervals for IFJ activa- tion (0.03–0.61), mid-DLPFC activation (0.13–0.59), and the interaction between IFJ and mid-cingulate activation (−0.61 to −0.14) did not contain zero. Similarly, the 95% confidence interval for age also did not contain zero (0.03–0.54). See Figure 4. Figure 3. The relationship between mid-cingulate activation and behavioral interference as a function of IFJ activation. Plot created using interact_plot from package interactions Version 1.1.5 (Long, 2022). 792 Journal of Cognitive Neuroscience Volume 35, Number 5 l D o w n o a d e d f r o m h t t p : / / d i r e c t . m i t . e d u / j / o c n a r t i c e - p d l f / / / / 3 5 5 7 8 1 2 0 7 7 7 2 2 / j o c n _ a _ 0 1 9 7 7 p d . f b y g u e s t t o n 0 8 S e p e m b e r 2 0 2 3 Figure 4. Bootstrap distributions of beta estimates from the cascade-of-control model. The predictor variables from the linear regression are shown along the y axis. Red circles indicate the mean beta estimate. Black lines and gray lines indicate the 80% and 95% confidence intervals, respectively. l D o w n o a d e d f r o m h t t p : / / d i r e c t . m i t . e d u / j / o c n a r t i c e - p d l f / / / / 3 5 5 7 8 1 2 0 7 7 7 2 2 / j o c n _ a _ 0 1 9 7 7 p d . f b y g u e s t t o n 0 8 S e p e m b e r 2 0 2 3 Alternative ROI by Contrast Pairings We next evaluated the performance of the cascade-of- control model against 63 models with alternative contrast pairings. The models with the 10 lowest BICs are reported in Table 4. The contrast pairings hypothesized by the cascade-of-control model resulted in the lowest BIC of all 64 combinations (BIC = 189.67). The model with the Table 4. Model Comparisons ROI by Contrast Pairing IFJ C + I + N > Fix
C + I + N > Fix
C + I + N > Fix
C + I + N > Fix
C + I + N > Fix
C + I + N > Fix
C + I + N > Fix
C + I + N > Fix
C + I + N > Fix
C + I + N > Fix
Mid-DLPFC
C + I > N
I > N
I > N
C + I > N
I > C
C + I + N > Fix
C + I > N
C + I + N > Fix
I > N
C + I + N > Fix
Mid-cingulate
I > C
I > C
C + I > N
C + I > N
C + I > N
C + I > N
I > N
I > C
I > N
I > N
BIC
189.67
190.89
194.04
194.21
194.50
194.65
196.45
196.60
196.71
196.87
ΔBIC
–
1.22
4.37
4.54
4.83
4.98
6.78
6.93
7.04
7.20
ROI by contrast pairings resulting in the 10 best goodness of fit metrics. ΔBIC was calculated by subtracting the lowest BIC from that of each alter-
native model.
Smith et al.
793
Table 5. Single Observation Point Model
Time Point
Task
TP1
Non-emotional
n
53
Predictor
Intercept
IFJ C + I + N > Fix
Mid-DLPFC C + I > N
Mid-cingulate I > C
IFJ C + I + N > Fix ×
Mid-cingulate I > C
Choice RT
Age
Emotional
55
Intercept
IFJ C + I + N > Fix
Mid-DLPFC C + I > N
Mid-cingulate I > C
IFJ C + I + N > Fix ×
Mid-cingulate I > C
Choice RT
Age
TP2
Non-emotional
40
Intercept
IFJ C + I + N > Fix
Mid-DLPFC C + I > N
Mid-cingulate I > C
IFJ C + I + N > Fix ×
Mid-cingulate I > C
Choice RT
Age
Emotional
45
Intercept
IFJ C + I + N > Fix
Mid-DLPFC C + I > N
Mid-Cingulate I > C
IFJ C + I + N > Fix ×
Mid-cingulate I > C
Choice RT
Age
* p < .05.
** p < .01.
*** p < .001.
Beta
0.02
0.25
0.29
−0.03
−0.12
−0.04
0.20
0.01
0.28
0.10
−0.25
−0.19
0.06
0.25
0.00
0.17
0.03
−0.08
−0.16
−0.09
0.17
−0.03
0.18
0.15
0.13
0.22
−0.03
0.07
SE
0.13
0.14
0.14
0.14
0.16
0.14
0.14
0.13
0.14
0.14
0.13
0.13
0.14
0.14
0.17
0.19
0.19
0.19
0.16
0.18
0.19
0.15
0.16
0.16
0.17
0.19
0.16
0.17
0.11
1.72
2.13
−0.20
−0.77
−0.30
1.38
0.09
2.02
0.74
−1.92
−1.51
0.41
1.81
0.00
0.88
0.17
−0.44
−1.00
−0.47
0.94
−0.19
1.14
0.94
0.79
1.15
−0.19
0.43
t-statistic
95% CI
−0.25, 0.28
−0.04, 0.53
p
.911
.093
0.02, 0.56
.039*
−0.32, 0.26
−0.45, 0.20
−0.32, 0.24
−0.09, 0.48
.845
.448
.765
.176
−0.24, 0.27
.931
0.00, 0.57
.049*
−0.17, 0.38
−0.52, 0.01
−0.45, 0.06
−0.23, 0.35
−0.03, 0.54
−0.34, 0.34
−0.22, 0.55
−0.35, 0.41
−0.47, 0.30
−0.50, 0.17
−0.46, 0.28
−0.20, 0.55
−0.34, 0.28
−0.14, 0.51
−0.17, 0.46
−0.21, 0.48
−0.17, 0.61
−0.36, 0.30
−0.26, 0.41
.460
.060
.137
.680
.076
.998
.387
.867
.664
.326
.639
.355
.854
.262
.355
.433
.257
.853
.668
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
5
7
8
1
2
0
7
7
7
2
2
/
j
o
c
n
_
a
_
0
1
9
7
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
794
Journal of Cognitive Neuroscience
Volume 35, Number 5
second lowest BIC was identical except that activation
within mid-DLPFC was indexed by Inc > Neu, rather
than Inc + Con > Neu (BIC = 190.89). Comparing the
cascade model to the third alternative model resulted in
a ΔBIC of 4.37, demonstrating moderately better fit for
the cascade model.
Single Observation Models
Finally, we assessed whether the same relationships were
observed when data from only a single task and single time
point were used to index brain activation. No consistent
brain–behavior relationships were observed across the
four single observation models. Significant results are
reported in the text, and full results for each model are
presented in Table 5. For the non-emotional task at TP1,
mid-DLPFC activation showed a significant positive rela-
tionship with behavioral interference (β = 0.29, t(46) =
2.13, p = .039). For the emotional task within the same
session, a significant relationship was observed between
IFJ activation and behavioral interference (β = 0.28,
t(48) = 2.02, p = .049). No significant relationships were
observed for either task at TP2 ( ps > .1).
DISCUSSION
Overview
The results of the present study are largely consistent with
the cascade-of-control model (Banich, 2019), providing
evidence that cognitive control is enacted via multiple
regions of frontal cortex that each resolve specific levels
of interference in a cascade-like manner. We demon-
strated that individual differences in composite measures
of brain activation within three ROIs were associated with
behavioral interference scores on a separate, out-of-
scanner task. The present study is novel in that we not only
confirmed the interactive brain–behavior relationships
hypothesized by the cascade-of-control model but also
showed that these relationships are linked to brain activa-
tion indexed at specific levels of interference (for other
evidence that distinct brain regions may code various
aspects of stimulus properties and control processes
related to performance on the Stroop task, see Freund,
Bugg, & Braver, 2021). Furthermore, this is one of rela-
tively few studies to use composite measures of brain acti-
vation collected across multiple fMRI tasks and multiple
time points. Below, we discuss the current findings as they
relate to the cascade-of-control model as well as the varia-
tion between the brain–behavior relationships observed
in the composite measures model as compared with the
single observation models, and the potential implications
of these differences.
Brain–Behavior Relationships
IFJ
Consistent with the cascade-of-control model, individuals
with less activation in IFJ showed significantly lower levels
of behavioral interference on the out-of-scanner Stroop
task. Although the directionality of activation differences
can be somewhat convoluted (e.g., see Rypma et al.,
2006), we interpret lower levels of activation in this region
as evidence of more efficient task set maintenance.
Also as predicted, the relationship between IFJ and
behavioral interference depended on mid-cingulate acti-
vation. Based on prior work demonstrating that task sets
are maintained regardless of trial type (e.g., Milham et al.,
2001), we additionally predicted and confirmed that this
relationship is specific to the Inc + Con + Neu > Fix con-
trast pairing.
Mid-DLPFC
A significant main effect was observed within mid-DLPFC
such that individuals with less activation in mid-DLFPC
showed significantly lower levels of behavioral inter-
ference. We interpret lower levels of activation in
mid-DLPFC as evidence of more efficient selection of infor-
mation in working memory. On the other hand, for those
individuals who have difficulty resolving competition,
biasing within mid-DLPFC must be ramped up to success-
fully complete the task, and thus activation is greater.
Also in line with our predictions, this mid-DLPFC rela-
tionship was specific to the Inc + Con > Neu contrast.
We had proposed that mid-DLPFC effects would be linked
to this contrast because for both incongruent and congru-
ent trials, response-relevant information is present in both
the task-relevant and task-irrelevant stimulus dimensions
(Milham & Banich, 2005). As such, both trial types should
theoretically require the selection of two pieces of infor-
mation that are being maintained in working memory.
This contrasts with neutral trials in which there is only
one source of response-relevant information that is con-
tained in the ink color. The current findings are also con-
sistent with a body of prior work demonstrating that
regions of mid-DLPFC may reduce interference from com-
peting information by buffering relevant information in
working memory (e.g., Burgess & Braver, 2010).
Mid-cingulate
As hypothesized, the relationship between mid-cingulate
activation and behavioral interference was moderated by
IFJ activation. Results further supported the hypothesized
Inc > Con contrast pairing, which captures the late-stage
response selection carried out by mid-cingulate.
Interaction between IFJ and Mid-cingulate
Central to the cascade-of-control model is the assertion
that successful competition resolution becomes depen-
dent on late-stage response selection in mid-cingulate
regions when a task set is not adequately maintained
and implemented within IFJ. Consistent with this tenet,
a significant interaction was observed between mean
Smith et al.
795
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
5
5
7
8
1
2
0
7
7
7
2
2
/
j
o
c
n
_
a
_
0
1
9
7
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
activation in IFJ (Inc + Con + Neu > Fix) and mean acti-
vation in mid-cingulate (Inc > Con). As expected, behav-
ioral interference was highest for individuals with high
levels of IFJ activation coupled with low levels of mid-
cingulate activation. As levels of mid-cingulate activation
increased, the resulting level of behavioral interference
decreased. We interpret high levels of IFJ activation as
evidence of more effortful and poorer task set implemen-
tation, which likely results in proportionally less task-
relevant information being fed downstream. Without
adequate biasing having occurred previously within the cas-
cade, the resolution of interference must occur at the final
stage of the cascade, that is, as late-stage response selection
within mid-cingulate. For those individuals who do not
successfully ramp up mid-cingulate to “pick up the slack,”
interference remains high.
The full crossover nature of the interaction was not
predicted and is somewhat intriguing. As expected, indi-
viduals who efficiently implemented a task set within IFJ
showed low behavioral interference with minimal engage-
ment of mid-cingulate. However, our results indicate that
ramping up mid-cingulate is not just unnecessary in this
case, but that doing so may actually be detrimental when
efficient biasing has already occurred within IFJ.
One potential interpretation of this pattern is that these
individuals are employing more caution at the final stage
at which control can be implemented. That is, control is
being exerted over response selection, even when the
correct response is sufficiently activated. If this were the
case, then we might expect a speed-accuracy trade-off
in which increases in mid-cingulate activation were accom-
panied by increases in accuracy. Such speed-accuracy
trade-offs are commonly observed in cognitive control
experiments and have been previously reported in work
similar to the current study (Silton et al., 2010). Unfortu-
nately, error rates in the current study were insufficient for
analysis and so this remains an open question for future
investigation.
Although the current study was not designed to inter-
rogate the temporal directionality of the relationship
between IFJ and mid-cingulate activation, the observed
interaction is consistent with prior work supporting a
directional path of information from lateral to medial
prefrontal regions. Using a combined fMRI and ERP meth-
odology, Silton et al. (2010) also observed a significant
interaction between activity within posterior regions of
dorsolateral pFC (the same area referred to as IFJ in the
current study) and cingulate regions. Critically, source-
waveform ERP mediation analysis revealed that the rela-
tionship between activity within left posterior dorsolateral
pFC and behavioral interference was mediated by activity
in right dorsal ACC. As hypothesized by the cascade-of-
control model, this pattern was specific to early activity
within posterior DLPFC (300–400 msec post-presentation)
and late activity within anterior cingulate (520–680 msec
post-presentation), with alternative models of temporal
information flow (e.g., early ACC and late posterior DLPFC)
not supported. Thus, we tentatively interpret the interac-
tion between IFJ and mid-cingulate activation in the current
study as further evidence that successful implementation
of a task set within IFJ modulated the need to resolve
response-related interference in mid-cingulate.
Alternative ROI by Contrast Pairings
When tested against all possible alternative ROI by con-
trast pairings, the cascade-of-control model demonstrated
better fit than each of the 63 alternative models. Perhaps
most interesting is the cascade model’s outperformance of
an alternative model in which activation across all ROIs
was indexed by the Inc > Neu contrast. Because behav-
ioral interference was indexed by the RT difference
between incongruent and neutral trials, it would be rea-
sonable to assume that the same difference in ROI acti-
vation, that is, the Inc > Neu contrast, would yield the
strongest associations. However, this does not appear to
be true in the current data. Similarly, an argument could
be made for using the Inc > Fix contrast to capture overall
neural engagement during the most effortful trials or,
alternatively, the Inc > Con contrast to capture activation
that occurs specifically when conflicting information is
present in both the task-relevant and task-irrelevant
dimensions. However, these models are again not sup-
ported by the data. Taken together, these results suggest
that the best index of activation for a particular region is
that which captures the cognitive operation(s) most
likely to be dependent on that region.
Composite Versus Single Observation Measures of
Brain Activation
Whereas the results of the composite measures model
were consistent with the cascade-of-control model, similar
results were not observed in the single observation
models. The results of these four models were neither
consistent with our theoretical model nor with one
another. Notably, only two significant brain–behavior rela-
tionships were present in the single observation models.
At TP1, a positive relationship was observed between mid-
DLPFC and behavioral interference for the non-emotional
task and a significant relationship between IFJ and inter-
ference was observed for the emotional task.
As discussed previously, reliability across task and time
point varied considerably. Although there was some
degree of consistency, particularly for the task versus fixa-
tion contrasts, it appears that the single time point and
single task measures of activation contain a substantial
amount of noise. There are numerous potential sources
of noise ranging from random measurement error in both
the brain and behavior measures to state-specific factors
such as alertness, which may have varied across testing
sessions. Although it is difficult to identify a specific
source, averaging across measures may have resulted in
796
Journal of Cognitive Neuroscience
Volume 35, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
5
5
7
8
1
2
0
7
7
7
2
2
/
j
o
c
n
_
a
_
0
1
9
7
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
a level of noise reduction sufficient for the observed brain–
behavior relationships to emerge.
Although not the focus of the present study, the dis-
crepancies between the composite model and the single
observation models have potential implications, most
notably for individual difference studies. Although the
growing trend has been to increase sample sizes (e.g.,
ABCD, Casey et al., 2018), there may also be considerable
value in increasing the quantity of data collected from each
individual participant. This deep phenotyping approach
has been used across a variety of cognitive neuroimaging
applications. For example, within a small sample of 10 par-
ticipants, Gordon et al. (2017) collected 6 hr of fMRI data
across three tasks in addition to multiple anatomical and
resting-state scans. The resulting Midnight Scan Club data
set has allowed for the mapping of neural organization at
the individual participant level, as well as numerous other
investigations of functional network characteristics (e.g.,
Dworetsky et al., 2021). A similar approach has been
employed by others including King, Hernandez-Castillo,
Poldrack, Ivry, and Diedrichsen (2019) who used a battery
of 26 tasks to map functional boundaries of the cerebel-
lum in 24 participants. The results of the current study
similarly provide some support for this approach as an
effective way to better identify meaningful brain–
behavior relationships.
There are, however, downsides to collecting larger
quantities of data per participant including more hours
spent in the laboratory, potentially over multiple sessions,
which can greatly increase the burden on participants.
Furthermore, issues such as missing data because of
attrition and poor data quality because of fatigue may
arise. In addition, this approach may necessitate a smaller
sample size, raising generalizability concerns. Ultimately,
the best method for testing individual differences will
depend on the experimental question and the available
resources.
Reliability
The reliability results observed within the current study
are consistent with prior work demonstrating that fMRI
task contrasts that show robust within-subject differences
often do not demonstrate high reliability (e.g., Kennedy
et al., 2022; Infantolino, Luking, Sauder, Curtin, & Hajcak,
2018). Across both the whole-brain and ROI measures,
reliability estimates differed substantially with the Inc +
Con + Neu > Fix contrast showing the highest levels of
reliability and the classical Inc > Con contrast showing
very poor levels of reliability. These results are likely driven
in part by the statistical nature of fMRI contrasts in which
one task condition is subtracted from another task condi-
tion. Although critical from a theoretical perspective, the
use of difference scores to isolate various cognitive pro-
cesses of interest negatively impacts measurement reliabil-
ity because the reliability of difference scores will always be
lower than the reliability of each component measure
(Hedge, Powell, & Sumner, 2018). As discussed by
Infantolino et al. (2018), high correlations between com-
ponent measures contribute substantially to the poor reli-
ability of any type of difference score. We see evidence of
this pattern in our results as reliability is lowest for those
task contrasts in which the cognitive and mental processes
required by each of the contrast components are most
similar, that is, incongruent and congruent trials.
Despite well-established concerns about their reliabil-
ity, the findings of the current study suggest that individual
differences in condition versus condition task contrasts
may reflect meaningful variation. Therefore, the use of
strategies to increase reliability may be of particular impor-
tance for individual differences studies in which isolating
specific cognitive processes is of particular interest. Multi-
ple such strategies including the use of composite mea-
sures were employed in the current study; however, the
suitability and value of any strategy will depend on many
study factors (see Elliott et al., 2021).
Comparisons with Other Conceptualizations
of Cognitive Control as Implemented by
Prefrontal Regions
Although the cascade-of-control model was found in the
current study to be useful in revealing brain–behavior rela-
tionships, there are other models of prefrontal regions
involved in cognitive control that might also be considered
and that differ in important ways from the model utilized
here. For example, Dosenbach et al. (2006) argue that
rather than IFJ imposing a task set, the regions responsible
for sustaining task sets are the dorsal ACC/medial superior
frontal cortex and bilateral anterior insula/frontal opercu-
lum. Determining which of these two models is more
appropriate is beyond the scope of the current paper.
In contrast to our model which posits that IFJ serves as
the critical first point of the cascade, models of cognitive
control by Badre and Nee (2018) suggest that mid-DLPFC
sits at the top of the hierarchy of control, although they do
not make specific arguments about whether that means
these regions acts before or after other regions. The model
of Koechlin and colleagues (Azuar et al., 2014; Koechlin &
Summerfield, 2007) argues for a somewhat different orga-
nization with levels of control organized in a rostral to cau-
dal manner over pFC, with more posterior regions
involved in sensory control, middle regions involved in
contextual control, and more anterior regions in episodic
control. Our model is consistent with aspects of this con-
ceptualization as it assumes that control in IFJ precedes
that of mid-DLPFC.
Our model also is distinct in important ways from the
model of Botvinick and colleagues (Botvinick, Cohen, &
Carter, 2004; Botvinick, Braver, Barch, Carter, & Cohen,
2001). In that model, the emphasis is on the anterior cin-
gulate acting as a conflict detector, which, when activated,
sends a signal to DLPFC to increase control. Our model is
distinct in that we have argued that the role of the mid-
Smith et al.
797
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
5
5
7
8
1
2
0
7
7
7
2
2
/
j
o
c
n
_
a
_
0
1
9
7
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
cingulate region is mainly to exert control at the level of
response output, rather than being a conflict detector
more generally (e.g., Milham & Banich, 2005; Milham
et al., 2001). In addition, our emphasis is on the directional
relationship in time between activity in mid-DLPFC and
ACC rather than the other way around as emphasized in
the Botvinich model (as an aside, the ACC to DLPFC feed-
back loop of Botvinick and colleagues could be easily inte-
grated into the cascade-of-control model). Future work
will be needed to adjudicate the degree to which these
various models provide contradictory or complimentary
perspectives that allow individual differences in brain
activation to be linked to individual differences in behav-
ioral performance.
Limitations
There are several limitations to the current study. First, our
sample size was relatively small for an investigation of indi-
vidual differences. Second, the inclusion of participants
with as few as two observation points is suboptimal as
many of these participants contributed data at only one
time point or for only one task. However, because of the
trade-off with sample size, we determined this to be the
optimal cutoff. Third, our sample consisted only of women
and, as such, it is possible the observed brain–behavior
relationships may not generalize to men, yet the likelihood
of substantial sex differences in the present findings is low as
multiple studies have observed no such sex differences in
performance on Stroop tasks (see Gaillard, Fehring, &
Rossell, 2021). Fourth, the current study cannot provide
definitive evidence for the cascade-of-control model,
which argues for a directional temporal relationship
between activation in different brain regions (i.e., IFJ
activation influencing later processing in ACC). fMRI is
ill-suited for examining such temporal relationships
compared with other cognitive neuroscience methods
such as electrophysiological recordings, which can
more fruitfully address this issue (see, e.g., Silton et al.,
2010). Nonetheless, the model predicts an interaction
effect between activation in IFJ and ACC, which was
observed in the current study. Fifth, although the Stroop
task is considered a “gold standard” task of cognitive con-
trol (MacLeod, 1992), the degree to which our model is
able to predict brain–behavior relationships in other cogni-
tive control tasks remains to be seen.
provided evidence that successful engagement of a task
set within IFJ reduces the need for late-stage response
selection within mid-cingulate regions. In addition, alter-
native models could not adequately predict brain–
behavior relationships in comparison to the theoretically
derived ROI by contrast pairings that reflect the cognitive
processes hypothesized to occur within each region by
the cascade-of-control model. These findings provide evi-
dence that individual differences in engagement of mul-
tiple regions of frontal cortex, each of which contributes
differently to resolving different types of interference,
may help to explain individual differences in cognitive
control outside the neuroimaging environment. Finally,
the current study provides an example of how a “hybrid”
approach to obtaining reliable and stable measures that
considered both between- and within-subject variance
through the use of multiple measures and data collection
time points may allow for the detection of brain–behavior
relationships.
Acknowledgments
We acknowledge the help of David Caha, Kenny Carlson,
Rebecca Helmuth, Kathy Pearson, and Amy Hegarty, PhD for
their help in data collection, organization, and preliminary
aspects of analyses, as well as the staff of the Intermountain
Neuroimaging Consortium, especially Nicole Speer, Operations
Director, and Teryn Wilkes, Head MR Technologist.
Reprint requests should be sent to Louisa L. Smith, Department
of Psychology and Neuroscience, D444 Muenzinger Hall, UCB
345, University of Colorado Boulder, Boulder, Colorado
80309, or via e-mail: Louisa.Smith@colorado.edu.
Data Availability Statement
Data and analysis materials are available upon request.
Author Contributions
Louisa L. Smith: Conceptualization; Formal analysis;
Visualization; Writing—Original draft; Writing—Review &
editing. Hannah R. Snyder: Funding acquisition; Method-
ology; Writing—Review & editing. Benjamin L. Hankin:
Funding acquisition; Methodology; Writing—Review &
editing. Marie T. Banich: Conceptualization; Funding
acquisition; Methodology; Project administration;
Resources; Supervision; Writing—Review & editing.
Summary
Funding Information
The present study demonstrated that individual differ-
ences in behavioral Stroop interference measured out-
side the magnet are associated with composite measures
of brain activation in a manner consistent with the
cascade-of-control model. As hypothesized, activation
within IFJ and mid-DLPFC regions predicted perfor-
mance on an out-of-scanner Stroop task. Furthermore,
a significant interaction between IFJ and mid-cingulate
Marie T. Banich and Benjamin L. Hankin, National Institute
of Mental Health (https://dx.doi.org/10.13039/100000025),
grant number: R01MH105501.
Diversity in Citation Practices
Retrospective analysis of the citations in every article pub-
lished in this journal from 2010 to 2021 reveals a persistent
798
Journal of Cognitive Neuroscience
Volume 35, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
5
5
7
8
1
2
0
7
7
7
2
2
/
j
o
c
n
_
a
_
0
1
9
7
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
pattern of gender imbalance: Although the proportions of
authorship teams (categorized by estimated gender iden-
tification of first author/last author) publishing in the Jour-
nal of Cognitive Neuroscience ( JoCN ) during this period
were M(an)/M = .407, W(oman)/M = .32, M/ W = .115,
and W/ W = .159, the comparable proportions for the arti-
cles that these authorship teams cited were M/M = .549,
W/M = .257, M/ W = .109, and W/ W = .085 (Postle and
Fulvio, JoCN, 34:1, pp. 1–3). Consequently, JoCN encour-
ages all authors to consider gender balance explicitly when
selecting which articles to cite and gives them the oppor-
tunity to report their article’s gender citation balance.
REFERENCES
Andrews-Hanna, J. R., Mackiewicz Seghete, K. L., Claus, E. D.,
Burgess, G. C., Ruzic, L., & Banich, M. T. (2011). Cognitive
control in adolescence: Neural underpinnings and relation to
self-report behaviors. PLoS One, 6, e21598. https://doi.org/10
.1371/journal.pone.0021598, PubMed: 21738725
Azuar, C., Reyes, P., Slachevsky, A., Volle, E., Kinkingnehun, S.,
Kouneiher, F., et al. (2014). Testing the model of
caudo-rostral organization of cognitive control in the human
with frontal lesions. Neuroimage, 84, 1053–1060. https://doi
.org/10.1016/j.neuroimage.2013.09.031, PubMed: 24064070
Badre, D., & Nee, D. E. (2018). Frontal cortex and the
hierarchical control of behavior. Trends in Cognitive
Sciences, 22, 170–188. https://doi.org/10.1016/j.tics.2017.11
.005, PubMed: 29229206
Baker, D. H., Vilidaite, G., Lygo, F. A., Smith, A. K., Flack, T. R.,
Gouws, A. D., et al. (2021). Power contours: Optimising
sample size and precision in experimental psychology and
human neuroscience. Psychological Methods, 26, 295–314.
https://doi.org/10.1037/met0000337, PubMed: 32673043
Baldauf, D., & Desimone, R. (2014). Neural mechanisms of
object-based attention. Science, 344, 424–427. https://doi.org
/10.1126/science.1247003, PubMed: 24763592
Bandettini, P. A., & Cox, R. W. (2000). Event-related fMRI
contrast when using constant interstimulus interval: Theory
and experiment. Magnetic Resonance in Medicine, 43,
540–548. https://doi.org/10.1002/(SICI)1522-2594(200004)
43:4<540::AID-MRM8>3.0.CO;2-R, PubMed: 10748429
Banich, M. T. (2009). Executive function: The search for an
integrated account. Current Directions in Psychological
Science, 18, 89–94. https://doi.org/10.1111/j.1467-8721.2009
.01615.x
Banich, M. T. (2019). The Stroop effect occurs at multiple
points along a cascade of control: Evidence from cognitive
neuroscience approaches. Frontiers in Psychology, 10, 2164.
https://doi.org/10.3389/fpsyg.2019.02164, PubMed: 31681058
Banich, M. T., Mackiewicz, K. L., Depue, B. E., Whitmer, A. J.,
Miller, G. A., & Heller, W. (2009). Cognitive control
mechanisms, emotion and memory: A neural perspective
with implications for psychopathology. Neuroscience &
Biobehavioral Reviews, 33, 613–630. https://doi.org/10.1016
/j.neubiorev.2008.09.010, PubMed: 18948135
Banich, M. T., Milham, M. P., Atchley, R. A., Cohen, N. J., Webb,
A., Wszalek, T., et al. (2000). Prefrontal regions play a
predominant role in imposing an attentional ‘set’: Evidence
from fMRI. Cognitive Brain Research, 10, 1–9. https://doi.org
/10.1016/S0926-6410(00)00015-X, PubMed: 10978687
Banich, M. T., Smolker, H. R., Snyder, H. R., Lewis-Peacock, J. A.,
Godinez, D. A., Wager, T. D., et al. (2019). Turning down the
heat: Neural mechanisms of cognitive control for inhibiting
task-irrelevant emotional information during adolescence.
Neuropsychologia, 125, 93–108. https://doi.org/10.1016/j
.neuropsychologia.2018.12.006, PubMed: 30615898
Botvinick, M. M., Braver, T. S., Barch, D. M., Carter, C. S., &
Cohen, J. D. (2001). Conflict monitoring and cognitive
control. Psychological Review, 108, 624–652. https://doi.org
/10.1037/0033-295X.108.3.624, PubMed: 11488380
Botvinick, M. M., Cohen, J. D., & Carter, C. S. (2004). Conflict
monitoring and anterior cingulate cortex: An update. Trends
in Cognitive Sciences, 8, 539–546. https://doi.org/10.1016/j
.tics.2004.10.003, PubMed: 15556023
Brass, M., Derrfuss, J., Forstmann, B., & von Cramon, D. Y.
(2005). The role of the inferior frontal junction area in
cognitive control. Trends in Cognitive Sciences, 9, 314–316.
https://doi.org/10.1016/j.tics.2005.05.001, PubMed: 15927520
Burgess, G. C., & Braver, T. S. (2010). Neural mechanisms of
interference control in working memory: Effects of
interference expectancy and fluid intelligence. PloS One, 5,
e12861. https://doi.org/10.1371/journal.pone.0012861,
PubMed: 20877464
Casey, B. J., Cannonier, T., Conley, M. I., Cohen, A. O., Barch,
D. M., Heitzeg, M. M., et al. (2018). The adolescent brain
cognitive development (ABCD) study: Imaging acquisition
across 21 sites. Developmental Cognitive Neuroscience, 32,
43–54. https://doi.org/10.1016/j.dcn.2018.03.001, PubMed:
29567376
Chapman, L. J., Chapman, J. P., Curran, T. E., & Miller, M. B.
(1994). Do children and the elderly show heightened
semantic priming? How to answer the question.
Developmental Review, 14, 159–185. https://doi.org/10.1006
/drev.1994.1007
Chechlacz, M., Gillebert, C. R., Vangkilde, S. A., Petersen, A., &
Humphreys, G. W. (2015). Structural variability within
frontoparietal networks and individual differences in
attentional functions: An approach using the theory of visual
attention. Journal of Neuroscience, 35, 10647–10658.
https://doi.org/10.1523/JNEUROSCI.0210-15.2015, PubMed:
26224851
Chen, G., Pine, D. S., Brotman, M. A., Smith, A. R., Cox, R. W.,
Taylor, P. A., et al. (2022). Hyperbolic trade-off: The
importance of balancing trial and subject sample sizes in
neuroimaging. Neuroimage, 247, 118786. https://doi.org/10
.1016/j.neuroimage.2021.118786, PubMed: 34906711
Derrfuss, J., Brass, M., Neumann, J., & von Cramon, D. Y.
(2005). Involvement of the inferior frontal junction in
cognitive control: Meta-analyses of switching and Stroop
studies. Human Brain Mapping, 25, 22–34. https://doi.org
/10.1002/hbm.20127, PubMed: 15846824
Dosenbach, N. U., Visscher, K. M., Palmer, E. D., Miezin, F. M.,
Wenger, K. K., Kang, H. C., et al. (2006). A core system for the
implementation of task sets. Neuron, 50, 799–812. https://doi
.org/10.1016/j.neuron.2006.04.031, PubMed: 16731517
Dworetsky, A., Seitzman, B. A., Adeyemo, B., Neta, M., Coalson,
R. S., Petersen, S. E., et al. (2021). Probabilistic mapping of
human functional brain networks identifies regions of high
group consensus. Neuroimage, 237, 118164. https://doi.org
/10.1016/j.neuroimage.2021.118164, PubMed: 34000397
Elliott, M. L., Knodt, A. R., & Hariri, A. R. (2021). Striving toward
translation: Strategies for reliable fMRI measurement. Trends
in Cognitive Sciences, 25, 776–787. https://doi.org/10.1016/j
.tics.2021.05.008, PubMed: 34134933
Faul, F., Erdfelder, E., Lang, A.-G., & Buchner, A. (2007). G*
Power 3: A flexible statistical power analysis program for
the social, behavioral, and biomedical sciences. Behavior
Research Methods, 39, 175–191. https://doi.org/10.3758
/BF03193146, PubMed: 17695343
Freund, M. C., Bugg, J. M., & Braver, T. S. (2021). A
representational similarity analysis of cognitive control
Smith et al.
799
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
5
5
7
8
1
2
0
7
7
7
2
2
/
j
o
c
n
_
a
_
0
1
9
7
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
during Color–Word Stroop. Journal of Neuroscience, 41,
7388–7402. https://doi.org/10.1523/JNEUROSCI.2956-20
.2021, PubMed: 34162756
Freund, M. C., Etzel, J. A., & Braver, T. S. (2021). Neural coding
of cognitive control: The representational similarity analysis
approach. Trends in Cognitive Sciences, 25, 622–638. https://
doi.org/10.1016/j.tics.2021.03.011, PubMed: 33895065
Friedman, N. P., Miyake, A., Altamirano, L. J., Corley, R. P.,
Young, S. E., Rhea, S. A., et al. (2016). Stability and change in
executive function abilities from late adolescence to early
adulthood: A longitudinal twin study. Developmental
Psychology, 52, 326–340. https://doi.org/10.1037/dev0000075,
PubMed: 26619323
Friedman, N. P., Miyake, A., Young, S. E., DeFries, J. C., Corley,
R. P., & Hewitt, J. K. (2008). Individual differences in
executive functions are almost entirely genetic in origin.
Journal of Experimental Psychology: General, 137, 201–225.
https://doi.org/10.1037/0096-3445.137.2.201, PubMed:
18473654
Gaillard, A., Fehring, D. J., & Rossell, S. L. (2021). A systematic
review and meta-analysis of behavioural sex differences in
executive control. European Journal of Neuroscience,
53, 519–542. https://doi.org/10.1111/ejn.14946, PubMed:
32844505
Gordon, E. M., Laumann, T. O., Gilmore, A. W., Newbold, D. J.,
Greene, D. J., Berg, J. J., et al. (2017). Precision functional
mapping of individual human brains. Neuron, 95, 791–807.
https://doi.org/10.1016/j.neuron.2017.07.011, PubMed:
28757305
Grady, C. L., Rieck, J. R., Nichol, D., Rodrigue, K. M., &
Kennedy, K. M. (2021). Influence of sample size and analytic
approach on stability and interpretation of brain–behavior
correlations in task-related fMRI data. Human Brain
Mapping, 42, 204–219. https://doi.org/10.1002/hbm.25217,
PubMed: 32996635
Gratton, C., Nelson, S. M., & Gordon, E. M. (2022). Brain–
behavior correlations: Two paths toward reliability. Neuron,
110, 1446–1449. https://doi.org/10.1016/j.neuron.2022.04
.018, PubMed: 35512638
individual differences. Behavior Research Methods, 50,
1166–1186. https://doi.org/10.3758/s13428-017-0935-1,
PubMed: 28726177
Herzmann, G., & Curran, T. (2011). Experts’ memory: An ERP
study of perceptual expertise effects on encoding and
recognition. Memory & Cognition, 39, 412–432. https://doi
.org/10.3758/s13421-010-0036-1, PubMed: 21264603
Infantolino, Z. P., Luking, K. R., Sauder, C. L., Curtin, J. J., &
Hajcak, G. (2018). Robust is not necessarily reliable: From
within-subjects fMRI contrasts to between-subjects
comparisons. Neuroimage, 173, 146–152. https://doi.org/10
.1016/j.neuroimage.2018.02.024, PubMed: 29458188
Jenkinson, M., Bannister, P., Brady, M., & Smith, S. (2002).
Improved optimization for the robust and accurate linear
registration and motion correction of brain images.
Neuroimage, 17, 825–841. https://doi.org/10.1006/nimg.2002
.1132, PubMed: 12377157
Jenkinson, M., & Smith, S. (2001). A global optimisation
method for robust affine registration of brain images.
Medical Image Analysis, 5, 143–156. https://doi.org/10.1016
/S1361-8415(01)00036-6, PubMed: 11516708
Kennedy, J. T., Harms, M. P., Korucuoglu, O., Astafiev, S. V.,
Barch, D. M., Thompson, W. K., et al. (2022). Reliability and
stability challenges in ABCD task fMRI data. Neuroimage,
252, 119046. https://doi.org/10.1016/j.neuroimage.2022
.119046, PubMed: 35245674
Kidd, E., Donnelly, S., & Christiansen, M. H. (2018). Individual
differences in language acquisition and processing. Trends in
Cognitive Sciences, 22, 154–169. https://doi.org/10.1016/j.tics
.2017.11.006, PubMed: 29277256
Killgore, W. D. (2010). Effects of sleep deprivation on
cognition. Progress in Brain Research, 185, 105–129.
https://doi.org/10.1016/B978-0-444-53702-7.00007-5,
PubMed: 21075236
King, M., Hernandez-Castillo, C. R., Poldrack, R. A., Ivry, R. B., &
Diedrichsen, J. (2019). Functional boundaries in the human
cerebellum revealed by a multi-domain task battery. Nature
Neuroscience, 22, 1371–1378. https://doi.org/10.1038/s41593
-019-0436-x, PubMed: 31285616
Greve, D. N., & Fischl, B. (2009). Accurate and robust brain
Koechlin, E., & Summerfield, C. (2007). An information
image alignment using boundary-based registration.
Neuroimage, 48, 63–72. https://doi.org/10.1016/j.neuroimage
.2009.06.060, PubMed: 19573611
Grinband, J., Savitskaya, J., Wager, T. D., Teichert, T., Ferrera,
V. P., & Hirsch, J. (2011). The dorsal medial frontal cortex
is sensitive to time on task, not response conflict or error
likelihood. Neuroimage, 57, 303–311. https://doi.org/10.1016
/j.neuroimage.2010.12.027, PubMed: 21168515
Gustavson, D. E., Miyake, A., Hewitt, J. K., & Friedman, N. P.
(2015). Understanding the cognitive and genetic
underpinnings of procrastination: Evidence for shared
genetic influences with goal management and executive
function abilities. Journal of Experimental Psychology:
General, 144, 1063–1079. https://doi.org/10.1037
/xge0000110, PubMed: 26389573
Haller, S. P., Kircanski, K., Stoddard, J., White, L. K., Chen, G.,
Sharif-Askary, B., et al. (2018). Reliability of neural activation
and connectivity during implicit face emotion processing in
youth. Developmental Cognitive Neuroscience, 31, 67–73.
https://doi.org/10.1016/j.dcn.2018.03.010, PubMed: 29753993
Hankin, B. L., Young, J. F., Abela, J. R., Smolen, A., Jenness, J. L.,
Gulley, L. D., et al. (2015). Depression from childhood into
late adolescence: Influence of gender, development, genetic
susceptibility, and peer stress. Journal of Abnormal
Psychology, 124, 803–816. https://doi.org/10.1037
/abn0000089, PubMed: 26595469
Hedge, C., Powell, G., & Sumner, P. (2018). The reliability
paradox: Why robust cognitive tasks do not produce reliable
theoretical approach to prefrontal executive function. Trends
in Cognitive Sciences, 11, 229–235. https://doi.org/10.1016/j
.tics.2007.04.005, PubMed: 17475536
Long, J. A. (2022). Interactions: Comprehensive, user-friendly
toolkit for probing interactions. R package version 1.1.5.
https://cran.r-project.org/package=interactions
MacLeod, C. M. (1992). The Stroop task: The “gold standard” of
attentional measures. Journal of Experimental Psychology:
General, 121, 12–14. https://doi.org/10.1037/0096-3445
.121.1.12
Marek, S., Tervo-Clemmens, B., Calabro, F. J., Montez, D. F.,
Kay, B. P., Hatoum, A. S., et al. (2022). Reproducible
brain-wide association studies require thousands of
individuals. Nature, 603, 654–660. https://doi.org/10.1038
/s41586-022-04492-9, PubMed: 35296861
Milham, M. P., & Banich, M. T. (2005). Anterior cingulate cortex:
An fMRI analysis of conflict specificity and functional
differentiation. Human Brain Mapping, 25, 328–335. https://
doi.org/10.1002/hbm.20110, PubMed: 15834861
Milham, M. P., Banich, M. T., Webb, A., Barad, V., Cohen, N. J.,
Wszalek, T., et al. (2001). The relative involvement of anterior
cingulate and prefrontal cortex in attentional control depends
on nature of conflict. Cognitive Brain Research, 12, 467–473.
https://doi.org/10.1016/S0926-6410(01)00076-3, PubMed:
11689307
Muhle-Karbe, P. S., Andres, M., & Brass, M. (2014). Transcranial
magnetic stimulation dissociates prefrontal and parietal
contributions to task preparation. Journal of Neuroscience,
800
Journal of Cognitive Neuroscience
Volume 35, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
5
5
7
8
1
2
0
7
7
7
2
2
/
j
o
c
n
_
a
_
0
1
9
7
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
34, 12481–12489. https://doi.org/10.1523/JNEUROSCI.4931
-13.2014, PubMed: 25209286
Poldrack, R. A., Baker, C. I., Durnez, J., Gorgolewski, K. J.,
Matthews, P. M., Munafò, M. R., et al. (2017). Scanning
the horizon: Towards transparent and reproducible
neuroimaging research. Nature Reviews Neuroscience, 18,
115–126. https://doi.org/10.1038/nrn.2016.167, PubMed:
28053326
Pruim, R. H., Mennes, M., van Rooij, D., Llera, A., Buitelaar, J. K.,
& Beckmann, C. F. (2015). ICA-AROMA: A robust ICA-based
strategy for removing motion artifacts from fMRI data.
Neuroimage, 112, 267–277. https://doi.org/10.1016/j
.neuroimage.2015.02.064, PubMed: 25770991
R Core Team. (2022). R: A language and environment for
statistical computing. Vienna, Austria: R Foundation for
Statistical Computing. https://www.R-project.org/
Raftery, A. E. (1995). Bayesian model selection in social
research. Sociological Methodology, 111–163. https://doi.org
/10.2307/271063
Rypma, B., Berger, J. S., Prabhakaran, V., Bly, B. M., Kimberg, D. Y.,
Biswal, B. B., et al. (2006). Neural correlates of cognitive
efficiency. Neuroimage, 33, 969–979. https://doi.org/10.1016
/j.neuroimage.2006.05.065, PubMed: 17010646
Schaefer, A., Kong, R., Gordon, E. M., Laumann, T. O., Zuo, X. N.,
Holmes, A. J., et al. (2018). Local-global parcellation of the
human cerebral cortex from intrinsic functional connectivity
MRI. Cerebral Cortex, 28, 3095–3114. https://doi.org/10.1093
/cercor/bhx179, PubMed: 28981612
Sheu, L. K., Jennings, J. R., & Gianaros, P. J. (2012). Test–retest
reliability of an fMRI paradigm for studies of cardiovascular
reactivity. Psychophysiology, 49, 873–884. https://doi.org/10
.1111/j.1469-8986.2012.01382.x, PubMed: 22594784
Silton, R. L., Heller, W., Towers, D. N., Engels, A. S., Spielberg,
J. M., Edgar, J. C., et al. (2010). The time course of activity in
dorsolateral prefrontal cortex and anterior cingulate cortex
during top–down attentional control. Neuroimage, 50,
1292–1302. https://doi.org/10.1016/j.neuroimage.2009.12
.061, PubMed: 20035885
Smith, S. M. (2002). Fast robust automated brain extraction.
Human Brain Mapping, 17, 143–155. https://doi.org/10.1002
/hbm.10062, PubMed: 12391568
Smith, S. M., Jenkinson, M., Woolrich, M. W., Beckmann, C. F.,
Behrens, T. E., Johansen-Berg, H., et al. (2004). Advances
in functional and structural MR image analysis and
implementation as FSL. Neuroimage, 23(Suppl. 1),
S208–S219. https://doi.org/10.1016/j.neuroimage.2004.07
.051, PubMed: 15501092
Snyder, H. R., & Hankin, B. L. (2016). Spiraling out of control:
Stress generation and subsequent rumination mediate the
link between poorer cognitive control and internalizing
psychopathology. Clinical Psychological Science, 4,
1047–1064. https://doi.org/10.1177/2167702616633157,
PubMed: 27840778
Snyder, H. R., Friedman, N. P., & Hankin, B. L. (2019).
Transdiagnostic mechanisms of psychopathology in youth:
Executive functions, dependent stress, and rumination.
Cognitive Therapy and Research, 43, 834–851. https://doi
.org/10.1007/s10608-019-10016-z, PubMed: 31551642
Sober, E. (2002). Instrumentalism, parsimony, and the Akaike
framework. Philosophy of Science, 69(Suppl. 3), S112–S123.
https://doi.org/10.1086/341839
Stroop, J. R. (1935). Studies of interference in serial verbal
reactions. Journal of Experimental Psychology, 18, 643–662.
https://doi.org/10.1037/h0054651
Tang, R., Etzel, J. A., Kizhner, A., & Braver, T. S. (2021).
Frontoparietal pattern similarity analyses of cognitive control
in monozygotic twins. Neuroimage, 241, 118415. https://doi
.org/10.1016/j.neuroimage.2021.118415, PubMed: 34298081
Tottenham, N., Tanaka, J. W., Leon, A. C., McCarry, T., Nurse,
M., Hare, T. A., et al. (2009). The NimStim set of facial
expressions: Judgments from untrained research participants.
Psychiatry Research, 168, 242–249. https://doi.org/10.1016/j
.psychres.2008.05.006, PubMed: 19564050
Unsworth, N. (2019). Individual differences in long-term
memory. Psychological Bulletin, 145, 79–139. https://doi.org
/10.1037/bul0000176, PubMed: 30596433
Vul, E., Harris, C., Winkielman, P., & Pashler, H. (2009).
Puzzlingly high correlations in fMRI studies of emotion,
personality, and social cognition. Perspectives on
Psychological Science, 4, 274–290. https://doi.org/10.1111/j
.1745-6924.2009.01125.x, PubMed: 26158964
Wilcox, R. R., & Keselman, H. J. (2003). Modern robust
data analysis methods: Measures of central tendency.
Psychological Methods, 8, 254–274. https://doi.org/10.1037
/1082-989X.8.3.254, PubMed: 14596490
Yarkoni, T. (2009). Big correlations in little studies: Inflated
fMRI correlations reflect low statistical power—Commentary
on Vul et al. (2009). Perspectives on Psychological Science, 4,
294–298. https://doi.org/10.1111/j.1745-6924.2009.01127.x,
PubMed: 26158966
Yarkoni, T., Poldrack, R. A., Nichols, T. E., Van Essen, D. C., &
Wager, T. D. (2011). Large-scale automated synthesis of human
functional neuroimaging data. Nature Methods, 8, 665–670.
https://doi.org/10.1038/nmeth.1635, PubMed: 21706013
Smith et al.
801
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
5
5
7
8
1
2
0
7
7
7
2
2
/
j
o
c
n
_
a
_
0
1
9
7
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3