Cannabinoid Modulations of Resting State EEG Theta
Power and Working Memory Are Correlated in Humans
Koen B. E. Böcker1, Claudine C. Hunault2, Jeroen Gerritsen1,
Maaike Kruidenier2, Tjeert T. Mensinga2, and J. Leon Kenemans1
Abstract
■ Object representations in working memory depend on
neural firing that is phase-locked to oscillations in the theta
band (4–8 Hz). Cannabis intake disrupts synchronicity of
theta oscillations and interferes with memory performance.
Sixteen participants smoked cigarettes containing 0.0, 29.3,
49.1, or 69.4 mg Δ9-tetrahydrocannabinol (THC) in a random-
ized crossover design and performed working memory and
general attention tasks. Dose-dependent effects of THC were
observed for resting state EEG theta and beta power, working
memory (per-item search time), and attentional performance
(percent errors and RT). The THC effects on EEG theta power
and memory performance were correlated, whereas other EEG
and behavioral effects were not. These findings confirm and
extend previous results in rodents and humans, and corrobo-
rate a neurocomputational model that postulates that tem-
poral aspects of information processing in working memory
depend causally on nested oscillations in the theta and gamma
(>30 Hz) bands. ■
INTRODUCTION
Phase-locking of neuronal firing to oscillations of local
field potentials is instrumental in neural representations
of behaviorally relevant information (Singer, 1993). The
short-term maintenance of behaviorally relevant informa-
tion is called working memory. Information in working
memory is coded by object-selective neural signals in, for
example, sensory cortex (Super, Spekreijse, & Lamme,
2001). These signals are phase-locked to local oscillations
in the 4–8 Hz theta frequency band (Lee, Simpson,
Logothetis, & Rainer, 2005). Accordingly, theta power in
working memory tasks has been shown to increase during
this task, both at the scalp (Ilan, Gevins, Coleman, ElSohly,
& de Wit, 2005; Ilan, Smith, & Gevins, 2004; Jensen &
Tesche, 2002; Klimesch, 1999; Gevins et al., 1998) and in-
tracranially (Raghavachari et al., 2001).
According to a current neurocomputational model, the
(repetitive) search through multiple items in memory
would be phase-locked to theta oscillations. Individual
items in memory would be separated by phase-locking
to gamma oscillations nested within the theta rhythm
( Jensen & Lisman, 1998). The model describes the reac-
tion time (RT) distribution in a Sternberg working memory
task, in which subjects decide whether a probe stimulus
was part of a memorized set or not (Sternberg, 1975).
Cannabinoids such as Δ9-tetrahydrocannabinol ( THC;
the main psychoactive ingredient of cannabis) have long
1Utrecht Institute for Pharmaceutical Studies and Rudolf Magnus
Institute of Neuroscience, The Netherlands, 2National Institute for
Public Health and Environment (RIVM), Bilthoven, The Netherlands
been known to disrupt working memory (in rodents, e.g.,
Hampson & Deadwyler, 2000; and in man, reviewed in
Ranganathan & DʼSouza, 2006). Activation of CB1 cannabi-
noid receptors inhibits synaptic transmission at hippocam-
pal pyramidal neurons. Cannabinoids consequently disrupt
the synchronization of hippocampal theta oscillations in
local field potentials in rodents, both in rest and during
the performance of a delayed alternation (working) mem-
ory task. Recently, it has been shown that the cannabi-
noid modulations of theta synchronization and working
memory performance in rodents are correlated (Robbe
et al., 2006).
In humans, local decreases in synchronicity of local field
potentials, that is, postsynaptic potentials, become manifest
as a decrease of power in scalp-recorded EEG (Bastiaansen &
Hagoort, 2006; Klimesch, 1999). In accordance with results
in rodents, acute cannabis intake in humans induced a de-
crease in EEG theta power both in rest and during a working
memory task, and led to a deterioration of working mem-
ory performance (Ilan et al., 2004, 2005). Here we tested
whether a decrease in resting state theta power is correlated
with deterioration of memory performance in humans.
The neurocomputational model holds that theta syn-
chronization is a necessary condition for working memory
performance, namely, memory search and comparison, in
the Sternberg task. This causal proposition cannot be tested
by demonstrating simultaneous pharmacological modula-
tions of theta synchronization and memory performance be-
cause these would not exclude the alternative proposition
that theta synchronization is an epiphenomenon of work-
ing memory operations. Yet, if it would be demonstrated
© 2009 Massachusetts Institute of Technology
Journal of Cognitive Neuroscience 22:9, pp. 1906–1916
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
–
p
r
d
t
i
2
c
2
l
9
e
–
1
p
9
d
0
f
6
/
1
2
9
2
3
/
9
9
8
/
9
1
1
9
o
0
c
6
n
/
1
2
0
7
0
7
9
0
5
2
7
1
3
3
5
/
5
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
3
5
0
5
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
f
.
/
.
t
o
n
1
8
M
a
y
2
0
2
1
four cannabis cigarettes (a mixture of 700 mg tobacco and
300 mg of cannabis), referred to as joints, on each testing
day, separated by a washout period of at least 7 days. The
four joints differed in THC levels, with the low dose corre-
sponding to 29.3 mg THC, the medium dose to 49.1 mg,
and the highest dose to 69.4 mg. A cannabis batch contain-
ing less than 9 μg THC was used to fabricate a placebo ciga-
rette. Subjects were instructed to smoke the cigarettes
according to a computer-paced procedure (i.e., 3 sec for
getting ready, 2 sec for inhalation, 3 sec for breath holding,
and 32 sec for normal breathing and relaxation). The whole
cigarette was smoked in about 22 min.
Behavioral Tasks
Participants completed several tasks at various times after
cannabis intake (Figure 1). Here we report on the memory-
search task and on a general attention task that was re-
corded closely in time with the EEG. Working memory
performance was assessed using the Sternbergʼs memory
scanning test, performed 60 min after the onset of cannabis
intake. Participants had to memorize four sets of digits,
increasing in length (i.e., 2 to 5 digits), generating four in-
creasing levels of memory load. Presentation of the mem-
ory set was terminated by the subject. Subsequently, on
memorization, a series of probe digits was presented.
Probe presentation was terminated by the response, or
after 2000 msec, and followed by 500 msec presentation
of a small square that was presented on the side of the cor-
rect response and served as feedback. The interstimulus in-
terval between feedback and the next probe was 1000 msec.
If a test item belonged to the memorized set and was a
target, participants had to press the right button; if it was
a lure and did not belong to the memorized set, they had
to press the left button. In total, 56 targets and 56 lures were
presented. Performance (in)accuracy was expressed as
that THC-induced theta desynchronization in a baseline
condition (i.e., during resting state) correlates with the
deterioration in working memory performance, then this
would corroborate that theta desynchronization is causally
related to memory performance. If theta power synchroni-
zation is only an epiphenomenon of memory performance,
then THC effects on baseline theta synchronization would
not correlate with working memory performance.
Furthermore, the neurocomputational model is specific
for working memory. Therefore, the relationship between
resting state theta power and working memory perfor-
mance should also be specific and should not be related
to general effects of THC intake, such as sedation. This spe-
cificity was addressed at various levels. First, to the extent
that both effects are dose-related, they should not only
be correlated over different subjects, but also within sub-
jects over doses. To optimize the chances of observing
dose-dependent relationships, the maximum dose tested
(69 mg THC) was much higher than in previous studies.
On the other hand, this dose is still ecologically valid as it
approximates the maximum THC content of cannabis ciga-
rettes in Great Britain (Potter, Clark, & Brown, 2008), The
Netherlands (Niesink, Rigter, & Hoek, 2004), and North
America (El Sohly, 2004). Second, we extended previous
findings by employing a memory task that enables the esti-
mation of per-item search time, which specifically depends
on the working memory scanning stage, and not on other
psychomotor stages (Sternberg, 1975). Third, the specificity
of possible correlations was assessed exploring correlations
with power changes in other EEG bands and with perfor-
mance in a general attention task as well.
METHODS
Participants
Sixteen male volunteers (age = 18–45 years) participated
in this study. They were recruited through advertisements
in (local) newspapers. They were selected on the basis of
their average cannabis use (between 2 and 9 cannabis cig-
arettes per month) to exclude both novice users and
users with considerable tolerance to effects of cannabis
(Perez-Reyes et al., 1991). Participants reported no history
of psychiatric diseases, and no respiratory diseases, liver
conditions, or cardiovascular problems. They neither use
medications (chronically) nor take excessive amounts of
alcohol. The study protocol was approved by the medical–
ethical committee of the Utrecht University Medical Centre.
Each participant was informed about the possible risks and
signed an informed consent form. The study was conducted
following the guidelines for Good Clinical Practice.
Design and Procedures
Cannabis Intake
The hypothesis was tested in a double-blind, placebo-
controlled, crossover design. Participants smoked one of
Figure 1. Time schedule of one testing day. Bold boxes indicate when
data for the present analyses have been recorded.
Böcker et al.
1907
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
–
p
r
d
t
i
2
c
2
l
9
e
–
1
p
9
d
0
f
6
/
1
2
9
2
3
/
9
9
8
/
9
1
1
9
o
0
c
6
n
/
1
2
0
7
0
7
9
0
5
2
7
1
3
3
5
/
5
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
3
5
0
5
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
t
.
f
.
/
o
n
1
8
M
a
y
2
0
2
1
percentage errors (including misses). The average RTs
were further broken down into a per-item search time
and a constant. Per-item search time characterizes the time
needed to compare a probe to the items in memory. For
each individual and each condition, a regression function
was fitted to the RT values for the four different memory
loads (memory set sizes 2 to 5), consisting of y = ax + b,
where a = per-item search time, x = memory load, and
b = the constant (or intercept; Jensen & Lisman, 1998;
Sternberg, 1975).
The general attention task was performed 150 min
after the onset of cannabis intake. Participants responded
to one of four different visual stimuli by pressing a button,
while ignoring the other three (Kenemans, Kok, & Smulders,
1993). The visual stimuli consisted of square-wave grat-
ings, varying in fundamental spatial frequency (high or
low, 0.6 and 4.8 cycles per degree of visual angle, respec-
tively) and orientation (horizontal or vertical). The gratings
subtended 6.67° of visual angle and were presented in the
center of a computer screen, against a gray background.
Each grating was presented for 50 msec. Eight consecutive
blocks of 128 trials (stimulus onset asynchrony = 750 to
950 msec) differed with respect to target stimulus and
responding hand. Behavioral-dependent measures were
RT, its standard deviation (sdRT; reflecting possible lapses
of attention), and the number of errors (omissions and
commissions).
On the evening prior to the first exposure, subjects were
individually trained to familiarize themselves with the tasks.
Full data on all tasks are reported elsewhere (Böcker et al.,
submitted; Hunault et al., 2009).
EEG Measurement and Analysis
EEG electrodes were applied and the EEG was recorded
just prior to the general attention task, during a 1-min inter-
val in which the participants sat quietly with eyes closed.
EEG was recorded from six midline (AFz, Fz, Cz, Pz, POz,
Oz) electrodes using an electrocap. The right mastoid was
used as a reference and a ground electrode was attached
to the forehead. Electrodes placed above and below the
right eye and at the outer canthi of both eyes were used
to measure the vertical and horizontal EOG, respectively.
Impedances were kept below 5 kΩ. Signals were amplified
using Ampligraph (EEG Technology International BV,
Leverey, the Netherlands) amplifiers with an on-line 100 Hz
low-pass filter and were sampled at a rate of 250 Hz using
Neuroscan Acquire software (Neuroscan, El Paso, TX).
The raw EEG signals were analyzed using BrainVision
Analyzer software (Brain Products, Munich, Germany).
Off-line filtering was applied (30 Hz low-pass, 12 dB/oct
and high-pass, time constant 1 sec at 24 dB/oct) and the data
were epoched for 50% overlapping 2-sec intervals, after
which artifacts were removed and the data were corrected
specifically for ocular artifacts (Gratton, Coles, & Donchin,
1983). Next, root-mean-square (RMS) power was calculated
for each interval using fast Fourier transform, after applying
a 10% Hanning window, and these values were averaged
over all intervals. The resulting resolution was 0.5 Hz. Aver-
age power in discrete EEG bands was submitted to statis-
tical analysis. To assess the validity of a possible change in
band-average theta power as reflecting a real theta power
change, time–frequency plots were evaluated. Alternative
possibilities are among others that the band-average power
decreased due to a change in peak theta frequency, with
some of the power at the sides of the peak extending partly
beyond the predefined band. Alternatively, a change in
band-average power might be secondary to a power change
and/or peak narrowing or widening in a neighboring
band. The time–frequency plots were derived by applica-
tion of Morlet wavelets that resolved the power between
0.5 and 25 Hz into 10 slightly overlapping frequency bands,
including 5–8.5 Hz.
Statistical Analyses
Both behavioral and EEG-power data were submitted to
a repeated measures MANOVA to asses the effect of
THC dose. The relationship between behavioral and
EEG power measures was assessed in two ways. First,
correlations between significant changes (69 mg placebo
condition) in EEG power and behavioral measures were
calculated. For significant power–behavior correlations,
the individual consistency of this relationship was as-
sessed, in turn, by calculating the correlation between
EEG power and the behavioral measure concerned at each
of the four doses for each subject separately. Subsequently,
it was tested whether the group-average correlation dif-
fered from zero.
RESULTS
Performance
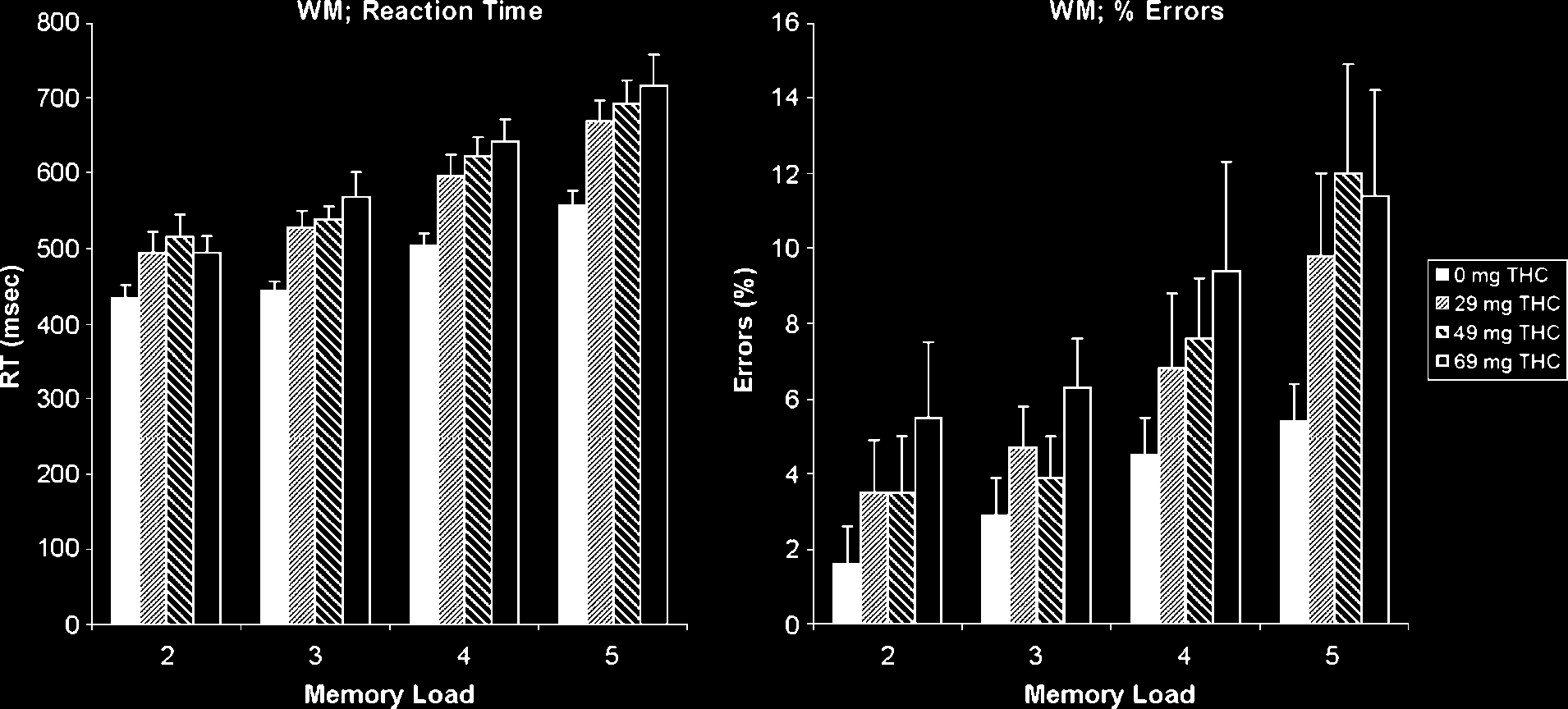
The RTs and percentage of errors in the Sternberg working
memory task are shown in Figure 2. The MANOVAs of
both RT and percentage of errors data showed main effects
of dose [F(3, 13) = 10.24, p < .005 and F(3, 13) = 4.34, p <
.05, respectively] and memory load [F(3, 13) = 38.87, p <
.0005 and F(3, 13) = 9.50, p < .005], but no interaction
[F(9, 7) = 1.17, ns and F(9, 7) < 1, ns]. The linear effects
of dose and memory load, indicating increasing RT and
increasing numbers of errors with increasing doses of
THC and increasing loads, were all significant [all F(1, 15)
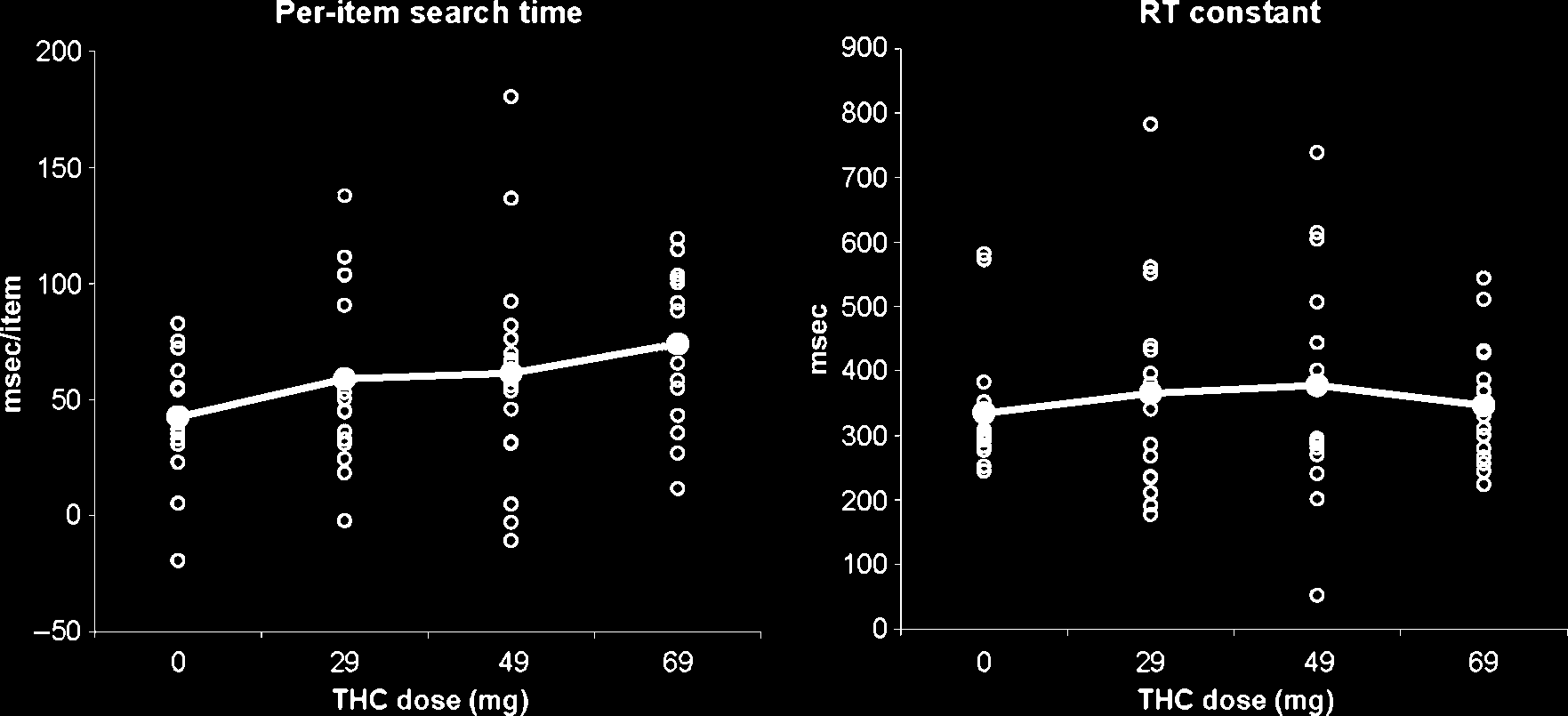
≥ 12.7, all p < .005]. After breaking down the RTs into per-
item search time and a constant (see Figure 3), only the
former revealed a cannabis effect [dose main effect: F(3,
13) = 4.01, p < .05 and F(3, 13) < 1, ns, respectively].
The per-item search time increased linearly with THC dose
[F(1, 15) = 12.6, p < .005].
The mean RT, its standard deviation (sdRT), and the per-
centage of errors in the general attention task are shown in
Figure 4. The MANOVA showed significant effects of THC
1908
Journal of Cognitive Neuroscience
Volume 22, Number 9
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
2
l
9
e
-
1
p
9
d
0
f
6
/
1
2
9
2
3
/
9
9
8
/
9
1
1
9
o
0
c
6
n
/
1
2
0
7
0
7
9
0
5
2
7
1
3
3
5
/
5
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
3
5
0
5
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
f
.
/
t
.
.
o
n
1
8
M
a
y
2
0
2
1
Figure 2. Working memory ( WM) performance deteriorated after cannabis intake, both with respect to RT (left) and proportion of errors (right).
Both measures increase with the length of the memory set.
dose on RT [F(3, 13) = 4.42, p < .05] and percentage errors
[F(3, 13) = 4.01, p < .05], and was marginally significant
for sdRT [F(3, 13) = 3.20, p = .06].
EEG Power
Figure 5 shows the RMS power in the different frequency
bands. Dose-related effects were found in theta [F(3, 13) =
9.18, p < .05] and beta bands [12–30 Hz, F(3, 13) = 3.51,
p < .05], but not in the delta [0.05–4 Hz, F(3, 13) = 1.58,
ns] and alpha bands [8–12 Hz, F(3, 13) < 1, ns]. Theta
band power was smaller for each active dose compared
to placebo (all p < .005). Furthermore, theta power was
smaller after 69 mg than after smoking 29 mg THC intake
( p < .05). The linear, quadratic, and cubic relationships
between THC dose and theta power were all significant
[all F(1, 15) ≥ 4.99, all p < .05]. THC beta band power
showed a quadratic relationship to THC dose [F(1, 15) =
8.60, p < .05]. Power in this band was larger after 49 mg
than after smoking 69 mg THC intake ( p < .01). Both
theta and beta power were maximal at electrode Fz [at
the midline between both hemispheres, over the frontal
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
2
l
9
e
-
1
p
9
d
0
f
6
/
1
2
9
2
3
/
9
9
8
/
9
1
1
9
o
0
c
6
n
/
1
2
0
7
0
7
9
0
5
2
7
1
3
3
5
/
5
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
3
5
0
5
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
f
.
.
/
t
o
n
1
8
M
a
y
2
0
2
1
Figure 3. Individual and grand-average (bold) working memory performance after different doses of THC. RT for comparing probes to the
memory set in the Sternberg task was broken down into per-item search time (left) and a constant (right). One hour after cannabis intake, only
the former showed a statistically significant increase ( p < .05).
Böcker et al.
1909
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
2
l
9
e
-
1
p
9
d
0
f
6
/
1
2
9
2
3
/
9
9
8
/
9
1
1
9
o
0
c
6
n
/
1
2
0
7
0
7
9
0
5
2
7
1
3
3
5
/
5
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
3
5
0
5
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
t
/
.
.
.
f
o
n
1
8
M
a
y
2
0
2
1
Figure 4. Individual and grand-average (bold) general attention performance after different doses of THC. Attention tended to deteriorate
after cannabis intake with respect to RT (upper left panel) and proportion of errors (lower panel), and tended to do so for the standard
deviation of the RT (SDRT; upper right panel).
lobes, main effect of electrode: F(5, 11) = 56.98, p < .0005
and F(5, 11) = 98.14, p < .0005, for theta and beta power,
respectively]. Figure 6 shows the power spectrum of the
EEG at this electrode.
To check the validity of a change in band-average theta
power as a measure of a change in power of theta oscilla-
tions, individual time–frequency plots were inspected.
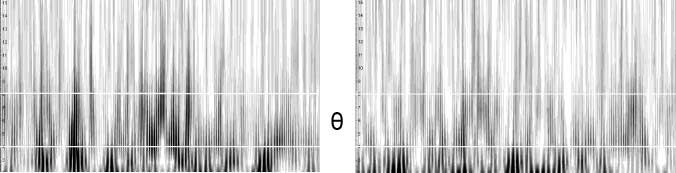
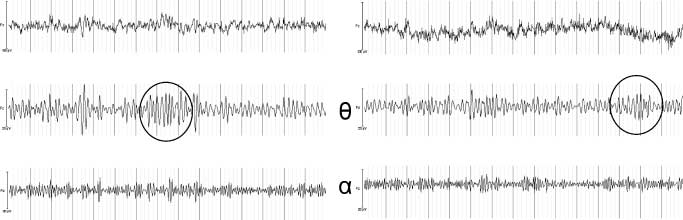
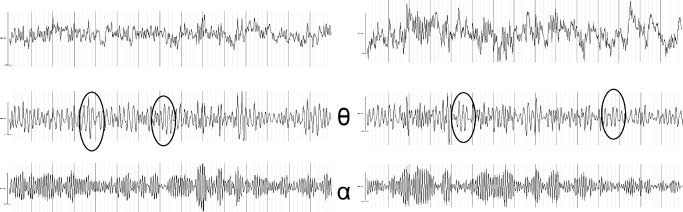
Twelve of the 16 subjects showed theta oscillations at Fz,
which were independent of oscillations in neighboring
bands for at least part of the time (Figure 7A and B show
data of two of those subjects). In an attempt to quantify
this subjective classification, 10 of these 12 subjects could
be identified quantitatively by relatively high average
theta power in the theta band under placebo conditions
(>0.80 μV) and a relatively low multiple R for the regression
of delta and alpha power over time on theta power over
1910
Journal of Cognitive Neuroscience
Volume 22, Number 9
time (R < .25). In the remaining participants, the theta
power and its modulations seemed to reflect these in
neighboring bands to a larger extent.
Correlations between Performance and EEG Power
Because both theta and beta power were maximal at elec-
trode Fz, all correlational analyses were performed for
data from this electrode. The correlation between THC
modulations of theta power and per-item search time was
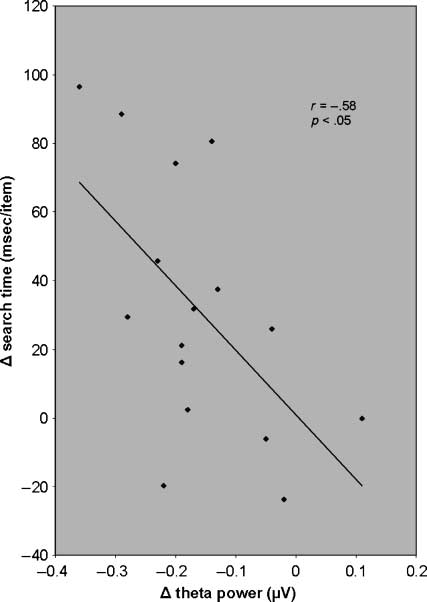
significant at the group level (r = −.58, p < .05; Figure 8).
Furthermore, it was found to be consistent over cannabis
doses at the individual level as well [mean correlation be-
tween individual theta power and per-item search time
0 trans-
over doses, r = −.30 ± .14 (M ± SEM, after Fischer Z
formation t(15) = −2.35, p < .05]. In contrast, the correla-
tion between THC modulations of theta power and the
number of errors on the working memory task was not sig-
nificant (r = −.34).
To assess the specificity of the correlation between the
increase in per-item search time and the decrease in theta
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
2
l
9
e
-
1
p
9
d
0
f
6
/
1
2
9
2
3
/
9
9
8
/
9
1
1
9
o
0
c
6
n
/
1
2
0
7
0
7
9
0
5
2
7
1
3
3
5
/
5
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
3
5
0
5
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
/
t
f
.
.
.
o
n
1
8
M
a
y
2
0
2
1
Figure 5. Individual and grand-average RMS power in each of the different frequency bands at electrode Fz.
Böcker et al.
1911
creased both theta power and working memory perfor-
mance speed. With the extended dose range that was
tested, both effects showed linear dose–response relation-
ships. Moreover, these effects were correlated, replicating
and extending the finding in rodents (Robbe et al., 2006),
whereas other cannabinoid modulations were not corre-
lated. A final indication of the specificity of the present find-
ings was the observation that, among the performance
measures derived from the Sternberg working memory
task, only the per-item search time was affected by THC,
not the constant psychomotor time.
The main aim of the study, a translational replication of
the correlation between theta synchronicity and working
memory performance, has been attained. The translational
replication of the present brain–behavior correlation was
not self-evident given the differences in theta generation
in rodents and man. These differences also lead to different
inferences regarding the physiological underpinning of
the observed correlations. In rodents there is strong cou-
pling between hippocampal and cortical theta during mem-
ory operations ( Jones & Wilson, 2005; Siapas, Lubenov, &
Wilson, 2005) that can also be observed in man (Gallinat
et al., 2006). This model would explain the presently ob-
served decrease in theta power as being a consequence of
decreased firing synchronicity in the hippocampus, such as
the desynchronization that followed application of cannabi-
noids in rats (Robbe et al., 2006). However, in man, cortical
theta has been shown to have a larger contribution from
local generators during sleep (Cantero et al., 2003) and
working memory operations (Raghavachari et al., 2006).
Therefore, the presently observed theta power decrease
could also represent the effects of THC on synchronicity
of local cortical generators of theta oscillations. As a conse-
quence of THC-induced desynchronization of either dis-
tributed or local theta generators, object-specific firing
patterns at gamma frequencies (above 30 Hz; Jensen,
2006; Jensen & Lisman, 1998) that are phase-locked to cor-
tical theta oscillations might be expected to become desyn-
chronized as well (Canolty et al., 2006; Lee et al., 2005). This
would have compromised the quality of the memory repre-
sentations. Therefore, the chances decrease that classifica-
tion of a probe stimulus and the related response decision
have completed at the end of a given search through the
items in memory (taking one full theta cycle; Jensen, 2006;
Jensen & Lisman, 1998). This would necessitate waiting for
another memory search (i.e., theta cycle) before being able
to decide and respond on some of the trials. In this way, per-
formance would slow down and become more error-prone
on average, as has been observed in the present study.
Results of intracranial EEG recordings also have signifi-
cance for mechanistic interpretations of the present results.
These recordings have shown that working memory-related
theta modulations reflect phase-resetting of spontaneous
theta (Rizzuto et al., 2003), in accordance with the neuro-
computational model ( Jensen, 2006; Jensen & Lisman,
1998). It could be assumed that THC-induced desynchroni-
zation of ongoing theta interferes with phase-resetting,
Figure 6. RMS EEG power in the theta band (4–8 Hz, average of
60 consecutive 2-sec intervals, eyes closed) decreased 2:20 hr after
administration of a cannabis cigarette containing 29, 49, or 69 mg
of THC compared to placebo ( p < 005). Electrode Fz (at the midline
between both hemispheres, over the frontal lobes) is shown.
Theta power was maximal at this location.
power, correlations with dose effects in the beta band, as well
as for performance on the general attention tasks, were cal-
culated. All dose effects were expressed as contrasts between
the highest dose and placebo. The correlation between the
drug effects on beta power (at electrode Fz) and memory
search time was −.035 and not significant. The correlations
between theta power and performance measures from the
general attention task were −.25 for RT, −.19 for sdRT, and
−.13 for error percentage, all nonsignificant.
Finally, the group-level correlation between THC modu-
lations of theta power and per-item search time was com-
puted for the subgroup of 10 subjects that clearly showed
independent theta oscillations according to the quantita-
tive criteria outlined above. This amounted to r = −.80
( p < .01).
DISCUSSION
The present translational study set out to replicate and ex-
tend the finding that cannabinoid modulation of working
memory and of synchronicity in theta oscillations were
correlated. It was found that, in humans, THC intake de-
1912
Journal of Cognitive Neuroscience
Volume 22, Number 9
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
2
l
9
e
-
1
p
9
d
0
f
6
/
1
2
9
2
3
/
9
9
8
/
9
1
1
9
o
0
c
6
n
/
1
2
0
7
0
7
9
0
5
2
7
1
3
3
5
/
5
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
3
5
0
5
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
f
.
t
.
/
.
o
n
1
8
M
a
y
2
0
2
1
which again compromises the memory representations and
slows down average performance. Alternatively, THC-in-
duced theta desynchronization might also directly interfere
with phase-locking by slowing down this process. To relate
this slowing of phase-locking to the observed increase in
working memory speed, it remains to be explained how this
slowing would depend on memory load.
The present study also replicated previous studies into
the effect of cannabinoids on working memory and EEG in
humans. These also found a decrease in working memory
speed and accuracy, as well as a decrease in EEG theta
power, both in rest and concurrent with performance of
the task (Ilan et al., 2004, 2005). In comparison with pre-
vious studies, the dose range was extended from 0–27 mg
(Ilan et al., 2005) to 0–69 mg THC in the present study.
Using the latter dose range, significant linear trends for
performance speed and EEG theta power were observed.
Given these dose-dependent effects, it is more probable
that the effects are receptor-specific rather than of a gen-
eral nature. The dose dependency of both effects was also
observed at the individual-subject level. On average, the
correlations between theta power and working memory
speed over doses were significantly positive.
Specificity was observed at several other levels. First,
the correlations were specific with respect to frequency
band. Per-item search time did not correlate with the ob-
served beta power modulations. Second, specificity with re-
spect to task was observed in that THC modulations in the
general attention task did not correlate with the decrease
in theta power. Third, within the task, only the per-item
search time, and neither the constant part nor the number
of errors, did correlate significantly with theta power. Like-
wise, theta phase-resetting in intracranial EEG was more
pronounced after presentation of a probe stimulus than
after presentation of a to-be-memorized item (Rizzuto
et al., 2003). This phase-reset was interpreted as a reflection
Figure 7. (A) Time–frequency
plot showing the power
distribution from 2 to 15 Hz
at electrode Fz over time.
Shown are 15 sec of data
under placebo and the highest
THC dose. The white horizontal
lines are at 4 and 8 HZ,
respectively (i.e., at the borders
of the theta band). Darker
compared to lighter areas
indicate time intervals with
larger power at a certain
frequency. Raw and filtered
EEG in the theta and alpha
bands is shown as well. Ellipses
indicate the clearest instances
of theta oscillations, which
are clearly independent of
either alpha or delta waves.
This subject shows a marked
reduction in theta power in
the THC condition. (B)
Corresponding data of a second
subject, with marked amplitude
fluctuations in the alpha band
and relatively constant power
in the theta band.
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
2
l
9
e
-
1
p
9
d
0
f
6
/
1
2
9
2
3
/
9
9
8
/
9
1
1
9
o
0
c
6
n
/
1
2
0
7
0
7
9
0
5
2
7
1
3
3
5
/
5
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
3
5
0
5
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
f
t
/
.
.
o
n
1
8
M
a
y
2
0
2
1
Böcker et al.
1913
splenial, lateral temporal, and medial prefrontal cortices
(Buckner, Andrews-Hanna, & Schacter, 2008). The latter
area probably generates more scalp-recorded EEG theta
when the default mode network is inactive (Scheeringa
et al., 2008). With respect to the present results, an inter-
pretation of cannabis modulations of theta power in terms
of an increase in default mode network is strengthened
by the finding that not only did theta power decrease,
but at least with 29 and 49 mg THC, beta power increased.
Similarly, default mode network activity has been shown to
correlate not only with theta power decreases but also with
beta power increases (Laufs et al., 2003). From this per-
spective, the observed correlation between theta power
decrease and per-item search time would stem from a
cannabis-induced increase in default mode network activ-
ity. Some of the effects of cannabis, such as disorganized
thought, together with cognitive decline (DʼSouza et al.,
2004), could be explained by a cannabis-induced increase
of default mode network activation that has been shown
to be related to mind wandering, or stimulus-independent
thought (Mason et al., 2007). Again, this is difficult to rec-
oncile with the specificity of the observed correlation that
was only significant for per-item search time and not for
changes in attention as assessed by the general attention
task. The present authors agree with Scheeringa et al.
(2008) that more experimental work is needed to be able
to conclude whether scalp EEG theta oscillations are speci-
fic pacemakers for scanning working memory or more gen-
erally related to decreases in default mode network activity.
The default network account of the present data, and of
theta power in general, also defines the functional sig-
nificance of resting state theta oscillations. The function
of theta oscillations is generally defined in terms of active
reentrant or looping information processing (as reviewed
in Mitchell, McNaughton, Flanagan, & Kirk, 2008). The re-
petitive serial search through multiple items in working
memory is an example of such reentrant processing. Ac-
cording to a default mode network account of theta oscilla-
tions in baseline EEG, these oscillations could be a free
running rhythmicity, with theta power serving as a state
parameter of the involved brain systems. High theta power
would indicate increased preparedness for reentrant or
looping processing. Recently, there has been much interest
in resting state brain activity. fMRI recordings of resting
state have been shown to consistently reflect fluctuations
in the activity of 10 networks (Damoiseaux et al., 2006),
including the default mode network. Their relationships
with fluctuations of resting EEG oscillations are only start-
ing to be explored (Laufs, 2008).
A potential limitation of the current study is that the re-
cordings of theta power and memory performance were
more than one hour apart. However, this did not obscure
the hypothesized relationship, and provided a first test of
causality, which was reinforced by the specificity of the
present results at various levels. A causal role of theta syn-
chronicity in memory function has also been deduced from
recent findings in the rat. These showed that not only does
Figure 8. Scatterplot showing the correlation between the theta
power decrease (69 mg–0 mg conditions) in Figures 5 and 6 and
the increase in per-item search time (idem) in Figure 3 (left) after
cannabis intake. The more negative was the theta power difference,
that is, the larger the power decrease, the larger was the elongation
of per-item search time.
of memory comparison operation, such as the per-item
search time (Sternberg, 1975). The alternative possibility
put forward by Rizzuto et al. (2003), that theta phase-
resetting was related to motor preparation, could be ruled
out in the present study by the absence of a correlation
between theta and the RT constant. The latter would have
reflected modulations in the duration of general sensory
and motor processing stages. The latter two levels of speci-
ficity are also difficult to reconcile with the conclusion that
the THC-induced decrease in theta power would reflect
increased arousal, and that the performance decrease in
working memory would result from cannabinoid modula-
tions of attention as suggested by Ilan et al. (2004, 2005).
However, a firm conclusion would necessitate a study that
combines the strong aspects of both studies (e.g., extend
the present set-up with physiological manifestations of
arousal and attention), and would take the relationship
between attention and working memory into account.
An opposite account of the present data states that
theta power increase is related to decreased activity in
a so-called default mode network (Scheeringa et al.,
2008), and vice versa. The default mode network consists
of areas that are less active during goal-directed behavior.
The core network includes the posterior cingulate, retro-
1914
Journal of Cognitive Neuroscience
Volume 22, Number 9
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
2
l
9
e
-
1
p
9
d
0
f
6
/
1
2
9
2
3
/
9
9
8
/
9
1
1
9
o
0
c
6
n
/
1
2
0
7
0
7
9
0
5
2
7
1
3
3
5
/
5
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
3
5
0
5
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
/
.
t
f
.
o
n
1
8
M
a
y
2
0
2
1
a disruption of theta synchronicity disrupt spatial memory,
but moreover, that restoring the synchronicity (at theta
frequency) by external electrical stimulation reinstates
memory function (McNaughton, Ruan, & Woodnorth,
2006). This does not exclude, for instance, that in the pre-
sent study the theta power decrease during baseline is an
epiphenomenon of a possible THC-induced decrease in
baseline working memory activity. However, the hypoth-
esis of a causal theta–working memory relationship, that
would clarify the mechanism of action of THC on working
memory at least, seems a fruitful hypothesis for future test-
ing. Given the limitations of behavioral tasks in rodents,
and of pharmacological and physiological manipulations
and recordings in man, such studies would probably re-
quire future translational studies of phase-resetting in mon-
keys (cf. Lee et al., 2005), or more advanced analysis in man
(Onton, Delorme, & Makeig, 2005). Such an advanced anal-
ysis would also shed light on the dynamics of EEG theta
oscillations during rest (as illustrated in Figure 7), and with-
in and over trials during task performance, which is missing
from average fast Fourier transform power values. Addi-
tional analysis of the present data confirmed that the main
result, the correlation between THC-induced modula-
tions in per-item search time and theta power, was indeed
driven by subjects that showed robust theta oscillations,
independent of those in the alpha and delta bands.
A second limitation of the present study was the use of a
small number of exclusively midline electrodes. This pre-
cluded the use of EEG mapping, ICA or dipole modeling,
or a combination of these techniques (e.g., Scheeringa
et al., 2008) to assess the intracranial origin of the present
findings. When such analysis techniques were employed,
a fronto-central theta rhythm was indeed observed (Onton
et al., 2005; Gevins, Smith, McEvoy, & Yu, 1997). The Fz
maximum in the present data at least suggested that can-
nabis affected the frontal midline theta rhythm (Mitchell
et al., 2008), as it did in a previous study (Ilan et al., 2004)
which employed 40 electrodes.
In conclusion, THC modulations of EEG theta synchroni-
city and working memory speed are correlated humans
as well as in rodents. These findings connect results from
invasive neurophysiological experiments in animals with
neurocomputational models of working memory based
on analyses of working memory speed in man.
Acknowledgments
Funding for this study was provided by the Dutch Ministry of
Health, Welfare and Sport, Project V/267002, which was repre-
sented in an ad hoc advisory board that made suggestions with
respect to the study design and data analysis, but had no further
role in the writing of the report or in the decision to submit the
paper for publication.
Reprint requests should be sent to Koen B. E. Böcker, Depart-
ment of Psychopharmacology, Faculty of Science, Utrecht Uni-
versity, Sorbonnelaan 16, NL-3584 CA Utrecht, The Netherlands,
or via e-mail: K.B.E.Bocker@uu.nl.
REFERENCES
Bastiaansen, M. C. M., & Hagoort, P. (2006). Oscillatory brain
dynamics during language comprehension. In W. Klimesch
& C. Neuper (Eds.), Event-related dynamics of brain
oscillations. Amsterdam: Elsevier.
Böcker, K. B. E., Gerritsen, J., Kruidenier, M., Hunault, C. C.,
Mensinga, T. T., & Kenemans, J. L. (submitted). Acute
effects of cannabis with high Δ9-THC contents on visual
selective attention; an event-related potential study.
Buckner, R. L., Andrews-Hanna, J. R., & Schacter, D. L.
(2008). The brainʼs default network: Anatomy, function,
and relevance to disease. Year in Cognitive Neuroscience
2008, Annals of the New York Academy of Sciences,
1124, 1–38.
Canolty, R. T., Edwards, E., Dalal, S. S., Soltani, M., Nagarajan,
S. S., Kirsch, H. E., et al. (2006). High gamma power is
phase-locked to theta oscillations in human neocortex.
Science, 313, 1626–1628.
Cantero, J. L., Atienza, M., Stickgold, R., Kahana, M. J.,
Madsen, J. R., & Kocsis, B. (2003). Sleep-dependent theta
oscillations in the human hippocampus and neocortex.
Journal of Neuroscience, 23, 10897–10903.
Damoiseaux, J. S., Rombouts, S. A. R. B., Barkhof, F.,
Scheltens, P., Stam, C. J., Smith, S. M., et al. (2006).
Consistent resting-state networks across healthy subjects.
Proceedings of the National Academy of Sciences,
U.S.A., 103, 13848–13853.
DʼSouza, D. C., Perry, E., MacDougall, L., Ammerman, Y.,
Cooper, T., Wu, Y. T., et al. (2004). The psychotomimetic
effects of intravenous delta-9-tetrahydrocannabinol in
healthy individuals: Implications for psychosis.
Neuropsychopharmacology, 29, 1558–1572.
El Sohly, M. A. (2004). Quarterly report potency monitoring
project # 85. National Center for Natural Products
Research, University of Mississippi.
Gallinat, J., Kunz, D., Senkowski, D., Kienast, T., Seifert, F.,
Schubert, F., et al. (2006). Hippocampal glutamate
concentration predicts cerebral theta oscillations during
cognitive processing. Psychopharmacology, 187, 103–111.
Gevins, A., Smith, M. E., Leong, H., McEvoy, L., Whitfield, S.,
Du, R., et al. (1998). Monitoring working memory load
during computer-based tasks with EEG pattern recognition
methods. Human Factors, 40, 79–91.
Gevins, A., Smith, M. E., McEvoy, L., & Yu, D. (1997).
High-resolution EEG mapping of cortical activation related
to working memory: Effects of task difficulty, type of
processing, and practice. Cerebral Cortex, 7, 374–385.
Gratton, G., Coles, M. G., & Donchin, E. (1983). A new
method for off-line removal of ocular artifact.
Electroencephalography and Clinical Neurophysiology,
55, 468–484.
Hampson, R. E., & Deadwyler, S. A. (2000). Cannabinoids reveal
the necessity of hippocampal neural encoding for short-term
memory in rats. Journal of Neuroscience, 20, 8932–8942.
Hunault, C. C., Mensinga, T. T., Böcker, K. B. E., Schipper, C. M. A.,
Kruidenier, M., Leenders, M. E. C., et al. (2009). Cognitive
and psychomotor effects in males after smoking a combination
of tobacco and cannabis containing up to 69 mg delta-9-
tetrahydrocannabinol (THC). Psychopharmacology, 204,
85–94.
Ilan, A. B., Gevins, A., Coleman, M., ElSohly, M. A., & de Wit, H.
(2005). Neurophysiological and subjective profile of
marijuana with varying concentrations of cannabinoids.
Behavioural Pharmacology, 16, 487–497.
Ilan, A. B., Smith, M. E., & Gevins, A. (2004). Effects of
marijuana on neurophysiological signals of working and
episodic memory. Psychopharmacology, 176, 214–222.
Böcker et al.
1915
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
2
l
9
e
-
1
p
9
d
0
f
6
/
1
2
9
2
3
/
9
9
8
/
9
1
1
9
o
0
c
6
n
/
1
2
0
7
0
7
9
0
5
2
7
1
3
3
5
/
5
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
3
5
0
5
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
/
f
.
t
.
o
n
1
8
M
a
y
2
0
2
1
Jensen, O. (2006). Maintenance of multiple working memory
items by temporal segmentation. Neuroscience, 139,
237–249.
Jensen, O., & Lisman, J. E. (1998). An oscillatory short-term
memory buffer model can account for data on the
Sternberg task. Journal of Neuroscience, 18,
10688–10699.
Onton, J., Delorme, A., & Makeig, S. (2005). Frontal midline
EEG dynamics during working memory. Neuroimage,
27, 341–356.
Perez-Reyes, M., White, W. R., Mcdonald, S. A., Hicks, R. E.,
Jeffcoat, A. R., & Cook, C. E. (1991). The pharmacological
effects of daily marijuana smoking in humans. Pharmacology
Biochemistry and Behavior, 40, 691–694.
Jensen, O., & Tesche, C. D. (2002). Frontal theta activity in
Potter, D. J., Clark, P., & Brown, M. B. (2008). Potency of
humans increases with memory load in a working memory
task. European Journal of Neuroscience, 15, 1395–1399.
Jones, M. W., & Wilson, M. A. (2005). Theta rhythms coordinate
hippocampal–prefrontal interactions in a spatial memory
task. PLoS Biology, 3, 2187–2199.
Kenemans, J. L., Kok, A., & Smulders, F. T. (1993). Event-related
potentials to conjunctions of spatial frequency and
orientation as a function of stimulus parameters and
response requirements. Electroencephalography
and Clinical Neurophysiology, 88, 51–63.
Klimesch, W. (1999). EEG alpha and theta oscillations
reflect cognitive and memory performance: A review
and analysis. Brain Research Reviews, 29, 169–195.
Laufs, H. (2008). Endogenous brain oscillations and related
networks detected by surface EEG-combined fMRI.
Human Brain Mapping, 29, 762–769.
Laufs, H., Krakow, K., Sterzer, P., Eger, E., Beyerle, A.,
Salek-Haddadi, A., et al. (2003). Electroencephalographic
signatures of attentional and cognitive default modes in
spontaneous brain activity fluctuations at rest. Proceedings
of the National Academy of Sciences, U.S.A., 100,
11053–11058.
Lee, H., Simpson, G. V., Logothetis, N. K., & Rainer, G. (2005).
Phase locking of single neuron activity to theta oscillations
during working memory in monkey extrastriate visual cortex.
Neuron, 45, 147–156.
Mason, M. F., Norton, M. I., Van Horn, J. D., Wegner, D. M.,
Grafton, S. T., & Macrae, C. N. (2007). Wandering minds:
The default network and stimulus-independent thought.
Science, 315, 393–395.
McNaughton, N., Ruan, M., & Woodnorth, M. A. (2006).
Restoring theta-like rhythmicity in rats restores initial
learning in the Morris water maze. Hippocampus, 16,
1102–1110.
Mitchell, D. J., McNaughton, N., Flanagan, D., & Kirk, I. J.
(2008). Frontal–midline theta from the perspective of
hippocampal “theta”. Progress in Neurobiology, 86, 156–185.
Niesink, R. J. M., Rigter, S., & Hoek, J. (2004). THC-concentraties
in wiet, nederwiet en hash in Nederlandse coffeshops,
2003–2004 [THC concentrations in cannabis, home-grown
cannabis and hashish in Dutch coffeeshops, 2003–2004].
Utrecht, The Netherlands: Trimbos Institute.
Delta(9)-THC and other cannabinoids in cannabis in England
in 2005: Implications for psychoactivity and pharmacology.
Journal of Forensic Sciences, 53, 90–94.
Raghavachari, S., Kahana, M. J., Rizzuto, D. S., Caplan, J. B.,
Kirschen, M. P., Bourgeois, B., et al. (2001). Gating of
human theta oscillations by a working memory task.
Journal of Neuroscience, 21, 3175–3183.
Raghavachari, S., Lisman, J. E., Tully, M., Madsen, J. R.,
Bromfield, E. B., & Kahana, M. J. (2006). Theta oscillations
in human cortex during a working-memory task: Evidence
for local generators. Journal of Neurophysiology, 95,
1630–1638.
Ranganathan, M., & DʼSouza, D. C. (2006). The acute effects
of cannabinoids on memory in humans: A review.
Psychopharmacology, 188, 425–444.
Rizzuto, D. S., Madsen, J. R., Bromfield, E. B., Schulze-Bonhage,
A., Seelig, D., Aschenbrenner-Scheibe, R., et al. (2003).
Reset of human neocortical oscillations during a working
memory task. Proceedings of the National Academy of
Sciences, U.S.A., 100, 7931–7936.
Robbe, D., Montgomery, S. M., Thome, A., Rueda-Orozco, P. E.,
McNaughton, B. L., & Buzsaki, G. (2006). Cannabinoids
reveal importance of spike timing coordination in
hippocampal function. Nature Neuroscience, 9,
1526–1533.
Scheeringa, R., Bastiaansen, M. C. M., Petersson, K. M.,
Oostenveld, R., Norris, D. G., & Hagoort, P. (2008).
Frontal theta EEG activity correlates negatively with the
default mode network in resting state. International
Journal of Psychophysiology, 67, 242–251.
Siapas, A. G., Lubenov, E. V., & Wilson, M. A. (2005). Prefrontal
phase locking to hippocampal theta oscillations. Neuron,
46, 141–151.
Singer, W. (1993). Synchronization of cortical activity and its
putative role in information-processing and learning.
Annual Review of Physiology, 55, 349–374.
Sternberg, S. (1975). Memory scanning: New findings and
current controversies. Quarterly Journal of Experimental
Psychology, 27, 1–32.
Super, H., Spekreijse, H., & Lamme, V. A. F. (2001). A neural
correlate of working memory in the monkey primary
visual cortex. Science, 293, 120–124.
1916
Journal of Cognitive Neuroscience
Volume 22, Number 9
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
2
l
9
e
-
1
p
9
d
0
f
6
/
1
2
9
2
3
/
9
9
8
/
9
1
1
9
o
0
c
6
n
/
1
2
0
7
0
7
9
0
5
2
7
1
3
3
5
/
5
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
3
5
0
5
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
t
.
.
.
/
f
o
n
1
8
M
a
y
2
0
2
1