Can the Brain Strategically Go on Automatic Pilot?
The Effect of If–Then Planning on
Behavioral Flexibility
Tim van Timmeren1,2
, John P. O’Doherty3, Nadza Dzinalija4, and Sanne de Wit1
Abstract
■ People often have good intentions but fail to adhere to
them. Implementation intentions, a form of strategic planning,
can help people to close this intention–behavior gap. Their
effectiveness has been proposed to depend on the mental
formation of a stimulus–response association between a trig-
ger and target behavior, thereby creating an “instant habit.”
If implementation intentions do indeed lead to reliance on
habitual control, then this may come at the cost of reduced
behavioral flexibility. Furthermore, we would expect a shift
from recruitment of corticostriatal brain regions implicated
in goal-directed control toward habit regions. To test these
ideas, we performed a fMRI study in which participants
received instrumental training supported by either implemen-
tation or goal intentions, followed by an outcome revaluation
to test reliance on habitual versus goal-directed control. We
found that implementation intentions led to increased effi-
ciency early in training, as reflected by higher accuracy, faster
RTs, and decreased anterior caudate engagement. However,
implementation intentions did not reduce behavioral flexibility
when goals changed during the test phase, nor did it affect
the underlying corticostriatal pathways. In addition, this study
showed that “slips of action” toward devalued outcomes are
associated with reduced activity in brain regions implicated
in goal-directed control (ventromedial prefrontal cortex and
lateral orbitofrontal cortex) and increased activity of the
fronto-parietal salience network (including the insula, dorsal
anterior cingulate cortex, and SMA). In conclusion, our behav-
ioral and neuroimaging findings suggest that strategic if–then
planning does not lead to a shift from goal-directed toward
habitual control. ■
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
5
6
9
5
7
2
0
8
2
8
8
1
/
j
o
c
n
_
a
_
0
1
9
9
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
INTRODUCTION
At the start of the new year, many people reflect on their
future plans and form resolutions. However, they often
fail to put their good intentions into practice (Sheeran &
Webb, 2016). Strategic “if–then” plans, also known as
implementation intentions, are an effective way to support
the translation of intentions to actions. For example,
instead of formulating an abstract plan such as “I want to
lose weight,” an implementation intention links the
intended action to a specific cue or situation, for example,
“If I get home, I will eat an apple,” thereby enhancing the
probability of success. Indeed, many studies have shown
that implementation intentions support behavior change
better than goal intentions that merely specify the
intended action or outcome (Gollwitzer & Sheeran,
2006). In addition to increasing attention to the relevant
cue, the effectiveness of if–then planning has been
proposed to rely on creating a strong associative link
between the stimulus (S) in the if-part (“home”) and the
response (R) in the then-part (eat an apple), in a manner
1University of Amsterdam, The Netherlands, 2Utrecht Univer-
sity, The Netherlands, 3California Institute of Technology, Pasa-
dena, 4Amsterdam UMC, Location VUmc, The Netherlands
akin to habits acquired through behavioral repetition
(Dickinson, 1985; Thorndike, 1911). These mentally
formed S–R associations may allow for automatic action
initiation (Gollwitzer, 2014)—a process often referred to
as strategic automaticity or “instant habits” (Gollwitzer,
1993, 1999, 2014).
The notion that merely using a verbal action-plan could
be sufficient to form a habit is fascinating, because a
central assumption in theories of habit formation is that
this process critically depends on behavioral repetition.
Support for the idea that implementation intentions
accelerate habit formation comes from research showing
that they increase (self-reported) automaticity (Orbell &
Verplanken, 2010; Parks-Stamm, Gollwitzer, & Oettingen,
2007; Brandstätter, Lengfelder, & Gollwitzer, 2001).
Therefore, implementation intentions lead to benefits
in terms of efficient goal attainment (Gollwitzer, 2014;
Gollwitzer & Sheeran, 2006). However, habits developed
through behavioral repetition also come at a cost,
namely, decreased behavioral flexibility (Dickinson,
1985). The question arises, therefore, if the use of imple-
mentation intentions also leads decreased flexibility when
goals change. This can be investigated using the
outcome-devaluation test, an experimental paradigm
originally used in rats (Adams & Dickinson, 1981) and
© 2023 Massachusetts Institute of Technology. Published under a
Creative Commons Attribution 4.0 International (CC BY 4.0) license.
Journal of Cognitive Neuroscience 35:6, pp. 957–975
https://doi.org/10.1162/jocn_a_01990
later translated to humans (de Wit, Corlett, Aitken,
Dickinson, & Fletcher, 2009; de Wit, Niry, Wariyar, Aitken,
& Dickinson, 2007; Valentin, Dickinson, & O’Doherty,
2007). In this task, participants first learn to make a
response to obtain a reward. Subsequently, the value of
the outcome associated with that response is devalued,
and the ability to flexibly adapt responding to this change
in outcome value is measured during an extinction test.
Sensitivity to outcome devaluation suggests that behavior
is based on knowledge and evaluation of their conse-
quences, and therefore under goal-directed control. If
implementation intentions lead to “instant habits,” then
we would predict reduced sensitivity to outcome devalu-
ation, reflecting a shift from goal-directed toward more
rigid, habitual control (de Wit et al., 2018; Balleine &
O’Doherty, 2010).
We have previously tested this hypothesis (van Timmeren
& de Wit, 2022), using a computerized symmetrical
outcome-revaluation task (SORT; Watson, Gladwin,
Verhoeven, & de Wit, 2022). Participants learn to make a
response (go) to certain ice cream vans to collect valuable
ice creams (and points) or to withhold a response (no-go)
to other ice cream vans delivering nonvaluable ice creams
(and a reduction of points). To investigate the effect of if–
then planning, we instructed them to use verbal imple-
mentation intentions for half of the stimuli and use goal
intentions for the other half. In the subsequent test phase,
some outcome values changed (i.e., outcome revalua-
tion). Whereas participants should continue to respond
according to the learned S–R mappings on value-
congruent trials (i.e., still-valuable and still-not-valuable),
they should flexibly adjust their behavior on value-
incongruent trials (i.e., devalued and upvalued). The
results of this previous study suggest that the use of imple-
mentation (compared with goal) intentions facilitates
instrumental learning, but also impairs performance when
some of the signaled outcome values change during the
test phase (van Timmeren & de Wit, 2022). This detrimen-
tal effect of if–then planning was observed across value-
congruent and incongruent trials, suggesting that it was
not mediated by strengthened S–R associations (as this
would have impacted the value-incongruent trials specifi-
cally). Instead, this result may have been driven by
reduced goal-directed control. Investigating the neural
processes underlying implementation intentions may
offer us a window on the underlying (goal-directed vs.
habitual) processes.
To this end, in the present study, we used fMRI to inves-
tigate the neural correlates of if–then planning of instru-
mental responses on the SORT. We capitalized on current
insights regarding the neural basis of goal-directed and
habitual control to investigate the notion that if–then
planning gives rise to “instant habits.” Decades of animal
research have provided detailed insights into the neurobi-
ology of goal-directed and habitual actions, demonstrating
that they are causally supported by anatomically distinct
but interacting corticostriatal systems (Balleine, 2019;
Balleine & O’Doherty, 2010; Yin, Knowlton, & Balleine,
2004). These findings are mirrored by (correlational)
neuroimaging evidence in humans, albeit less consis-
tently. Specifically, previous fMRI studies have found that
goal-directed control is supported by the ventromedial
prefrontal cortex (vMPFC) and caudate whereas
outcome-insensitive habitual actions depend on the pre-
motor cortex and posterior putamen/dorsal striatum
( Watson, van Wingen, & de Wit, 2018; Delorme et al.,
2016; Morris, Quail, Griffiths, Green, & Balleine, 2015;
de Wit et al., 2012; Tricomi, Balleine, & O’Doherty, 2009;
Valentin et al., 2007).
The present study is the first fMRI investigation with the
SORT, and we will therefore start with specifying our pre-
dictions regarding the general pattern of neural activity
independent of intentions. First, we expected that over
the course of training (i.e., habit acquisition) activity
would increase in regions associated with habitual control
whereas the involvement of regions implicated in goal-
directed control would decrease (Zwosta, Ruge, Goschke,
& Wolfensteller, 2018; Liljeholm, Dunne, & O’Doherty,
2015; Tricomi et al., 2009). Second, we expected neural
activity during training in these regions to be predictive
of revaluation insensitivity in the test phase ( Watson
et al., 2018; Zwosta et al., 2018; Liljeholm et al., 2015; de
Wit et al., 2009). Third, in line with previous work ( Watson
et al., 2018; Valentin et al., 2007), we hypothesized that, in
the test phase, we would find higher activity in areas impli-
cated in goal-directed action, cognitive control, and
response conflict when participants flexibly updated their
responses and equal (if anything reduced) activity in habit-
related regions. Finally, we expected that “slips of action”
would be associated with higher activity in habit regions
and reduced activity in goal-directed regions ( Watson
et al., 2018).
Our central aim was to investigate the neural basis of
implementation intentions and their effect on behavioral
flexibility. To this end, we measured neural activity related
to the effect of implementation intentions on acquisition
and flexible adjustment of instrumental actions on the
SORT. We hypothesized that the use of implementation
intentions (compared with goal intentions) during train-
ing would lead to increased habit acquisition as reflected
by higher accuracy, increased automaticity (measured
with the Self-Reported Behavioral Automaticity Index;
Gardner, Abraham, Lally, & de Bruijn, 2012), and increased
brain activity in habit regions and equal—or if anything
reduced—activity in goal-directed regions. Moreover, we
expected if–then planning to lead to increased reliance on
previously formed S–R associations in the subsequent test
phase as indicated by inflexible, habitual responding on
value-incongruent compared with value-congruent trials,
and higher activity of habit regions during the test phase.
Finally, we expected that overcoming mentally rehearsed
S–R associations (as part of an if–then plan) would require
more goal-directed control and correspondingly engage
related neural regions.
958
Journal of Cognitive Neuroscience
Volume 35, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
5
6
9
5
7
2
0
8
2
8
8
1
/
j
o
c
n
_
a
_
0
1
9
9
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
METHODS
All operationalizations, exclusion criteria, and main hypoth-
eses and analyses were preregistered on Open Science
Framework (https://osf.io/yrpxa).
Participants
Participants were recruited through the participant
portal of the University of Amsterdam Web site, flyers,
and word of mouth. We used the following inclusion
criteria: age 16–35 years, not having previously partici-
pated in a previous study using this same task, and any
contraindications for MRI. Data collection took place
between July and November 2020. Note that this is during
the first year of the COVID-19 outbreak; however, no strict
lockdowns were implemented during this period in The
Netherlands. The study was approved by the Psychology
ethics committee of the University of Amsterdam and
performed in accordance with those guidelines. All
participants gave informed consent and received either
course credit or financial compensation (15 A/hr) for
their time (total ∼2 hr). An additional A20 voucher was
given to the participant with the highest score to moti-
vate participants to perform well on the task.
Forty-seven participants were enrolled, conforming to
our preregistered sampling plan. Our sample size was
based on a previous pilot study, which found a significant
effect of implementation intentions in 35 participants using
the same task and manipulation. Moreover, a power analy-
sis with G*Power (Version 3.1.9.3) showed that our target
sample size of n = 40 should be sufficient to detect a small
behavioral effect ( f = 0.12) with an α level of .05 and
power of .8. Six participants were excluded from all analy-
ses. One participant quit half-way through participation,
and five participants were excluded based on performance
exclusion criteria (see Results for details). The remaining
41 participants (22 women, 19 men) had a mean age of
23.2 (SD = 4.1) years. All participants had normal or
corrected-to-normal vision, and all were right-handed
except one who was ambidextrous. All participants were
free of neurological or psychiatric disorders and completed
or were enrolled in higher professional education at the
time of participation, the vast majority being university stu-
dents. Two participants were native Germans who spoke
Dutch fluently; all others were native Dutch speakers.
Stimuli and Materials
Procedure
Participants performed a computerized instrumental learn-
ing task called the SORT (Figure 1; Watson, Gladwin, et al.,
2022), programmed in Presentation (Version 18.1). Partic-
ipants played a hungry skateboarder with the objective to
collect ice creams to earn points and satisfy their hunger
by pressing a response button. They were informed that
the best performing participant at the end of the study
would receive a A20 voucher. Four pictures of ice creams
were used: a Cornetto, a Magnum, a Rocket ice lolly, and a
soft serve ice cream. The task consisted of three phases.
First, participants conducted an instrumental training
phase without strategic planning outside the scanner, after
which they were moved to the MRI scanner and performed
an instrumental training phase with strategic planning
followed by a test phase (see Figure 1). The symmetrical
nature of the task stems from the inclusion of both valuable
and nonvaluable outcomes, which allows comparisons in
the test phase (when outcome values change) between
the value-congruent and value-incongruent conditions
to be made with the same response type (see Watson,
Gladwin, et al., 2022, for a more elaborate discussion on
the advantages of this task). The total experiment took
∼2 hr, of which 1 hr was spent in the scanner.
The task used here is almost identical to a previous
study in which we tested the same hypothesis behaviorally
(van Timmeren & de Wit, 2022), apart from the following
changes. To minimize head movements, we used a static
version of the task here instead of having ice cream trucks
moving across the screen. We added one block of practice
with strategic planning before being moved to the scanner,
in order for participants to once read the intentions out
loud and be able to ask questions. Moreover, we adapted
the task to promote stimulus–outcome (S-O) learning
across intention conditions, to rule out that any effect of
implementation intentions on behavioral flexibility would
be mediated by reduced contingency knowledge, as was
the case in the original behavioral study (van Timmeren
& de Wit, 2022). To this end, we changed the way in which
the blocks were composed in the first part of training
(i.e., without intentions): Instead of alternating between
two sets of four ice cream vans, each block now contains
four (out of eight) pseudorandomly selected stimuli (see
Instrumental Training section for details). More than with
the block-sets, participants are now forced to pay atten-
tion to all outcomes in the value-screen and evaluate for
which stimulus they should (not) make a response.
Instrumental Training
At the start of the task, participants were instructed that
their goal was to collect valuable ice creams (which earn
points and alleviate hunger) and avoid collecting nonvalu-
able ice creams (which lose points and cause stomach
pain) by (not) responding to ice cream vans. There were
four different ice creams, and before each block of instru-
mental training, participants were shown which two ice
creams were valuable (in green) and which two ice creams
were not valuable (in red; Figure 1A). The position of the
valuable and nonvaluable ice creams (left/right) was coun-
terbalanced across participants. Each ice cream was associ-
ated with two out of eight vans (Figure 1B): one van always
predicting this ice cream as being valuable and the other as
being nonvaluable. Each block contained only half of the
vans: two associated with a valuable ice cream and two with
a nonvaluable ice cream. Participants were told to find out
van Timmeren et al.
959
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
5
6
9
5
7
2
0
8
2
8
8
1
/
j
o
c
n
_
a
_
0
1
9
9
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
5
6
9
5
7
2
0
8
2
8
8
1
/
j
o
c
n
_
a
_
0
1
9
9
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
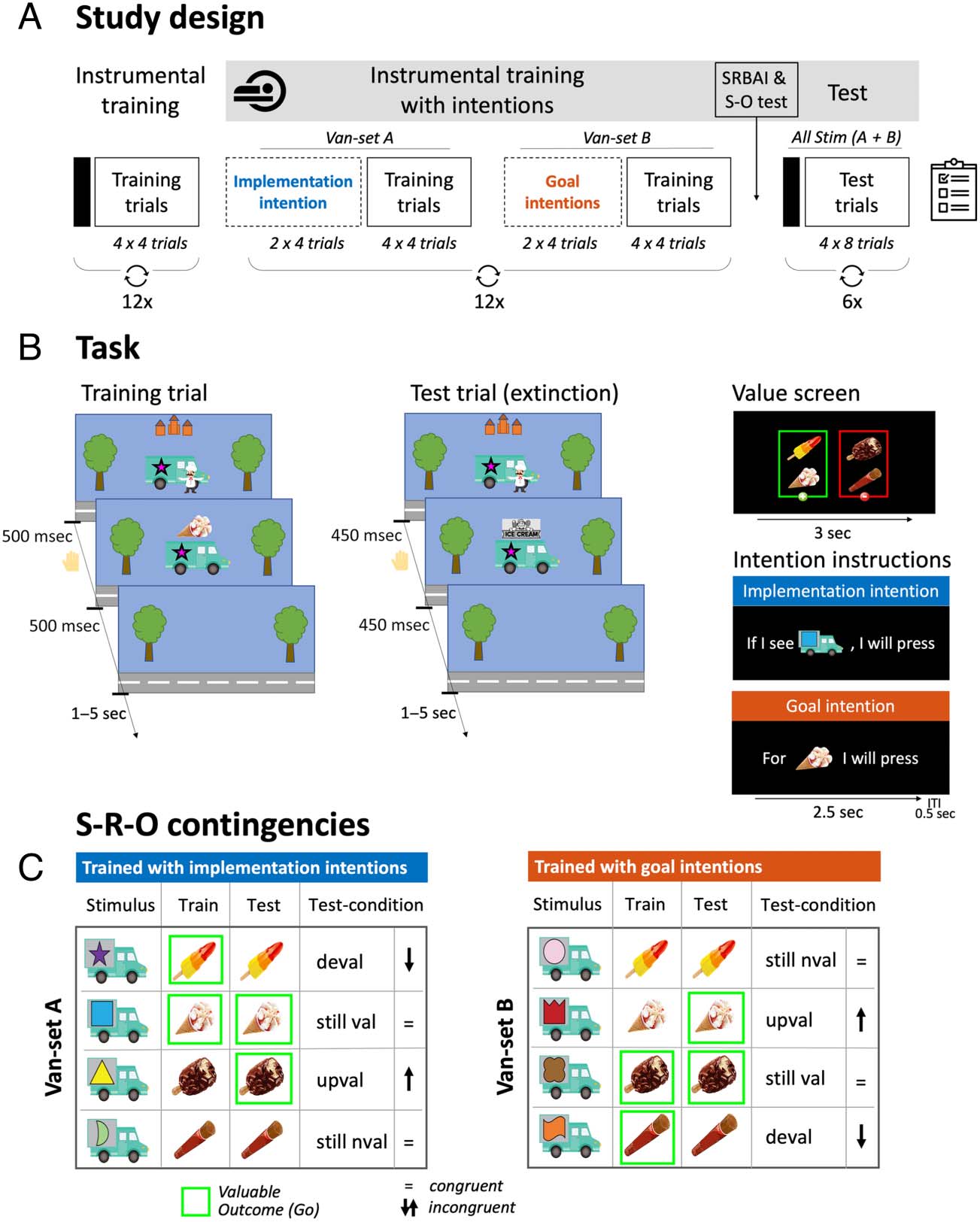
Figure 1. Overview of the study and experimental design. Participants were told they were playing a hungry skateboarder and their goal was to
collect some ice creams and not others to earn points. (A) Participants first received instrumental training. Each block started with a value-screen
(represented by the black rectangle), followed by a block of 16 training trials (see B). Each block contained four vans (pseudorandomly selected).
Training then continued with participants additionally using implementation intentions (trained with Van-Set A) or goal intentions (trained with
Van-Set B; see C), with intention instructions (see B) being presented before each instrumental learning block. Finally, participants completed six test
blocks in which all eight vans ( Van-Sets A and B) would appear intermixed and consequently the associated outcome-values of some vans changed
compared with training (see C, comparing the “Train” vs. “Test” columns). (B) Train trial: When a van was presented, participants had to decide
whether to make a response within 500 msec, after which the ice cream appeared (irrespective of a response) on top of the van for 500 msec. Test
trial: identical to train blocks, but now (i) a banner appeared on top of the van instead of the ice cream to prevent feedback about the outcome
(i.e., nominal extinction) and (ii) response time was reduced to 450 msec. Value screen: The outcome-value screen indicates which ice creams should
(in green) and should not (in red) be collected. Intention instructions: Vans were trained with either implementation intentions, indicating for which
ice cream van they should or should not make a response, or goal intentions, indicating for which ice cream they should (not) make a response.
(C) An overview of stimulus-outcome contingencies (example set) and associated values across different phases of the task. The contingencies
between each ice cream and van remained consistent throughout the whole task, but the value of each ice cream (and hence the associated
response) was stable only during training. During the critical test phase, the associated outcome values changed (were incongruent) relative to
the training value for half of the stimuli (indicated by arrows). This results in four conditions: still-valuable trials (valuable, congruent), upvalued trials
(valuable, incongruent), still-not-valuable trials (nonvaluable, congruent) and devalued trials (nonvaluable, incongruent). For example, the first
van always delivered a Rocket, which was valuable throughout training but no longer valuable during test (i.e., devalued). Shown here is an example
of the contingencies in one of six test blocks; across the test phase, the correct response for each stimulus was equally often congruent and
incongruent. Deval = Devalued; still val = still-valuable; upval = upvalued; still not = still-not-valuable trials.
by trial and error which ice cream truck delivered which ice
cream, and that the S-O contingencies would remain the
same throughout the whole task. Participants first prac-
ticed with different discriminative stimuli (scooters) and
outcomes (pizzas) for two blocks to familiarize them with
this procedure. As mentioned previously, the composition
of the blocks (i.e., which four out of eight vans were pre-
sented during this block) was now pseudorandomized.
The conditions described above allow for six unique com-
binations of four vans, which were presented twice each
960
Journal of Cognitive Neuroscience
Volume 35, Number 6
(order randomized) during this first part of training for 12
blocks. The contingencies between ice creams and vans
and which of the ice creams was valuable/nonvaluable
were randomized across participants.
Each stimulus was shown 4 times per block, constituting
16 trials. Trial order was randomized per eight trials, with
each van being presented twice in the first and twice in
the second half of a block. Each trial started with a jittered
1- to 5-sec intertrial interval. Participants were instructed
that they should respond as quickly as possible and before
the deliverer disappeared (after 500 msec). Irrespective of
the response, the associated outcome was then presented
for 500 msec. Thus, participants did not receive direct
feedback about the accuracy of their response to balance
the feedback provided for valuable and nonvaluable out-
comes and to promote goal-directed (R-O) learning and
S-O knowledge. Each block ended with a 3-sec feedback
screen that displayed accuracy and late responses in that
block and total number of points collected (Figure 1D).
Instrumental Training with Intentions
The next phase of training took part in the MRI scanner.
Participants were told that instead of seeing which ice
creams were valuable or nonvaluable, each block would
now start with sentences that would help them perform
forms
well. These sentences came in two different
(Figure 1D). Goal intentions indicated for each ice
cream whether they should make a response (R-O), for-
mulized as “If I see [picture of an ice cream], then I WILL
press.” Implementation intentions indicated for each ice
cream van if they should make a response or not (S–R),
formulized as “If I see [picture of an ice cream van] then I
WILL (NOT) press.” Each intention was presented for
2500 msec and twice per intention block (randomized
order). Half of the stimuli were trained using goal and
the other using implementation intentions. Each block
of verbal intentions was directly followed by a block of
instrumental training (identical to the previous phase)
with the corresponding stimuli. Blocks now alternated
between two sets of vans, one van-set being trained with
implementation intentions (S1–S4, “Van-Set A”) and one
with goal intentions (S5–S8, “Van-Set B”). Whether the
training started with an implementation or goal intention
block was counterbalanced across participants. At the
end of regular instrumental training and before being
moved to the scanner, participants practiced each verbal
intention without instrumental training for one block,
followed by two blocks (one for each intention type) with
instrumental training. During these first few practice
blocks outside the scanner, participants were asked to
read the intentions out loud. During the subsequent 24
blocks of training with intentions in the scanner, partici-
pants were instructed to subvocalize the intentions
instead of reading them out loud to minimize head
motion. Participants entered the scanner in a head-first
supine position and were able to view the screen using
a mirror attached to the head coil on which the task stim-
uli were presented. A button box allowed them to collect
ice creams by responding using their right index finger.
At the end of training with intentions, participants
completed a questionnaire on subjective automaticity
(Self-Report Behavioral Automaticity Index [SRBAI]) and
were tested on their S-O knowledge (details below;
Figure 1E). We had planned to additionally obtain a (pre-
intention) baseline measure of these questionnaires, but
because of a programming error, they were presented
after the practice blocks with intentions, making them
unusable as a baseline measure.
Test Phase
Participants completed six test blocks. The test phase
was similar to the first training phase (without intentions),
but with some important differences. First, as intention
blocks were no longer presented, value-screens were again
shown at the start of each block, for the duration of 4 sec.
Second, participants were told that the ice cream deliverers
placed a banner on top of their van, blocking the view of
the ice cream they delivered (i.e., nominal extinction).
Because each van still kept on delivering the same ice
cream as during training, they should base their choice
on what they learned before. Third, the feedback screens
presented at the end of each block no longer included
information on the accuracy of their responses, but only
the percentage of responses, nonresponses, and late
responses. We did this to prevent outcome-based learning
during the test phase. We explicitly instructed participants
that each block contained an equal amount of valuable and
nonvaluable outcomes so they knew they should aim for a
50%/50% distribution. Fourth, we shortened the response
window to 450 msec to force rapid responding, which has
been shown to boost the expression of habitual slips
(Hardwick, Forrence, Krakauer, & Haith, 2019). However,
because a lot of participants responded just after the
450-msec time limit, we decided to include responses up
to 600 msec for both the behavioral and fMRI analysis to
increase the number of included trials in the fMRI analyses.
This change did not significantly impact the pattern of
behavioral results, which was unsurprising as the test phase
was conducted in extinction, meaning that no performance
feedback was provided during this period. Finally and cru-
cially, participants were informed that the final phase
would be more challenging because all eight ice cream
vans would appear intermixed during each block. The cru-
cial consequence of each block containing all eight stimuli
is that half of the vans would now deliver an ice cream with
a value incongruent with the value during training. Some
ice cream vans for which they had been trained to always
make a go response during training, now delivered a (deva-
lued) ice cream that should not be collected. Vice versa,
other ice creams vans had carried nonvaluable outcomes
during training, but their signaled outcome was upvalued
and therefore required a go response. On other (value-
van Timmeren et al.
961
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
5
6
9
5
7
2
0
8
2
8
8
1
/
j
o
c
n
_
a
_
0
1
9
9
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
congruent) test trials, the signaled outcome remained the
same (i.e., still-valuable and still-not-valuable trials).
Consider for example the Rocket ice cream in Figure 1C.
In this example, during training, this ice cream is always deliv-
ered by the van with a purple star and the van with the pink
circle. During training blocks with the van with the purple
star, the Rocket is valuable and therefore requires a go
response. In contrast, during training blocks with the van
with the pink circle, the Rocket is not valuable, and partici-
pants should refrain from pressing the space bar (i.e., no-go
response). Subsequently, during the test block all (ice cream
van), stimuli are presented, and in the example illustrated in
Figure 1C, the Rocket is instructed to be currently not valu-
able. This means that the van with the purple star signals a
devalued outcome (i.e., this is value-incongruent with train-
ing and requires a different response), and the van with the
pink circle signals a still-valuable outcome (i.e., value congru-
ent; the learned response remains correct).
SRBAI
The SRBAI (Gardner et al., 2012) is a 4-item scale that cap-
tures self-reported habitual behavior patterns that we
adapted for to assess automaticity for (not) responding
to the ice cream vans. Participants were presented with
each ice cream van and asked to indicate the associated
response (press or not press) and the degree to which
(not) making a response was something they did: “auto-
matically,” “without having to consciously remember,”
“without thinking,” and “before I realize I am doing it.”
Each item was scored on a scale ranging from 1 (strongly
disagree) to 100 (strongly agree). The SRBAI scale was
previously shown to have good reliability and validity
(Gardner et al., 2012). Before the four SRBAI items
appeared, participants were asked to indicate which
response was associated with that stimulus (“making a
response” / “not making a response”) to test S–R knowl-
edge. Cronbach’s alpha was calculated separately for each
of the four conditions (2 intentions × 2 values), using the
eight test items (four SRBAI questions for the two stimuli
per condition). The results indicate high internal reliabil-
ity, with alpha ranging from .91 to .95. The final score was
calculated separately for each intention by taking the
mean across the four items (range: 1–100), with higher
scores reflecting more automatic behavior.
Test of Stimulus–Outcome Knowledge
Participants were asked about their knowledge of the S-O
contingencies by asking them for each ice cream vans
which ice cream it delivered. After selecting one of the four
ice creams, participants were asked to indicate how confi-
dent they were about their decision (0–100). Composite
scores, reflecting S-O knowledge, were calculated for each
intention and separately for go- and no-go-trained stimuli
by multiplying percentage of correct S-O contingencies
(0%/50%/100%) with percentage mean confidence.
Preregistered Behavioral Data Analysis
Behavioral data analyses were performed using IBM SPSS
Statistics 25 for Mac for frequentist statistics and JASP
Version 0.16.3 ( JASP Team, 2018) for Bayesian statistics.
For data analysis purposes, the training data were col-
lapsed across blocks of three, referred to as block-sets.
Accuracy is reflected by the percentage of trials on which
a correct response was made, calculated by the number of
correct responses divided by the total number of trials. In
line with the fMRI analyses, trials on which a late response
was made were not included in the analyses (of both accu-
racy and RTs). To assess that learning took place over the
first part of the training without intentions, accuracy was
analyzed using a 2 × 4 repeated-measures ANOVA with
within-subject factors Value (valuable or nonvaluable)
and Block-set (1–4). The second part of training was ana-
lyzed using a 2 × 2 × 4 repeated-measures ANOVA, with
Intention Type (implementation or goal intention) as an
additional factor. RTs for correct responses (and thus only
for valuable go trials) were analyzed with similar ANOVAs.
For the test phase, data were analyzed using a 2 × 2 × 2
repeated-measures ANOVA with three factors: Intention
Type (implementation or goal intention), Test Value (valu-
able or nonvaluable during test), and Congruency (congru-
ent or incongruent with value during training). Thus, for
each intention type there are four conditions: still-valuable
trials (valuable, congruent), upvalued trials (valuable, incon-
gruent), still-not-valuable trials (nonvaluable, congruent),
and devalued trials (nonvaluable, incongruent). Again,
RTs (including all responses up to 600 msec) were analyzed
using similar ANOVAs but now also analyzing responses on
no-go trials (i.e., responses on still-not-valuable and deva-
lued trials). Note that eight participants were excluded from
the no-go analyses because they performed perfectly on
still-not-valuable trials and thus did not make any response.
Subjective automaticity (SRBAI scores) for responding
to stimuli trained with implementation and goal intentions
at the end of training was compared using a paired t test.
Finally, the relationship between automaticity and the
“revaluation insensitivity” index was tested for both
intention types separately using correlational analyses. A
revaluation insensitivity index was calculated for each
intention type by taking the difference between accuracy
for congruent and incongruent test trials separately for go
(still-valuable minus devalued) and no-go-trained stimuli
(still-not-valuable minus upvalued), with higher revalua-
tion insensitivity scores indicating more habitual perfor-
mance. Kendall’s tau was used as the four revaluation
indices, and SRBAI scores were not normally distributed.
In the case of violations of sphericity, we report Green-
house–Geisser corrected degrees of freedom and
p values. In addition to 95% confidence intervals, partial
2) for the ANOVAs and Cohen’s d for paired
eta squared (ηp
t tests are reported as estimates of effect sizes.
We additionally conducted corresponding Bayesian
analyses. For null results ( p > .05), as preregistered, we
962
Journal of Cognitive Neuroscience
Volume 35, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
5
6
9
5
7
2
0
8
2
8
8
1
/
j
o
c
n
_
a
_
0
1
9
9
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
report the Bayes Factor01 (BF01), which quantifies the rel-
ative evidence in favor of the null hypothesis (H0) over the
alternative hypothesis (H1). For ANOVAs, we report the
BFexcl, which quantifies the extent to which the data sup-
port inclusion of the factor of interest in the model (i.e.,
the change from before posterior inclusion odds, across
matched models). Finally, although we interpret signifi-
cant findings on the basis of p < .05, we also report BFs
for comprehensiveness and transparency (i.e., BF10 or
BFincl for ANOVAs, which quantify evidence in favor of
the alternative hypothesis over H0 and is identical to
1/BF01/excl). BFs were interpreted according to Table 1 in
Wetzels and colleagues (2011), with BFs between one and
three reflecting anecdotal support, BFs larger than three
reflecting substantial support, and BFs larger than 10
reflecting strong support. In all Bayesian analyses, JASP’s
default priors (cauchy = 0.707 for t tests and r = 0.5 for
fixed and r = 1 for random effects for ANOVAs) were used.
MRI Data Acquisition
All MRIs were performed on a 3-Tesla, full-body Achieva
dStream MRI-scanner (Philips Medical Systems) equipped
with a 32-channel head coil. After entering the scanner, a
low-resolution survey scan was made to determine the
location of the field of view.
fMRI scans were acquired at a ∼30° angle from the
anterior–posterior commissure line to maximize signal
sensitivity in orbital regions (Deichmann, Gottfried,
Hutton, & Turner, 2003) using a T2*-weighted single-shot
gradient echo imaging sequence with the following
parameters: repetition time = 2000 msec; echo time =
28 msec; flip angle = 76.1°; voxel size = 3 mm3 with
0.3-mm slice gap; matrix size = 80 × 78; number of
slices = 36; field of view = 240 × 118.5 × 240 mm. The
Table 1. Imaging Results of the Training Phase (Exploratory)
training with intentions was split in two runs of 598 scans
each, whereas 415 scans were acquired for the test phase.
The first six volumes of each run were discarded to allow
T1 saturation to reach equilibrium.
A high-resolution T1-weighted structural image was
acquired before the final run (while participants com-
pleted the post-training SRBAI and SO-test) using an
MPRAGE sequence with the following parameters: voxel
size = 1 mm3; field of view = 240 × 220 × 188 mm; rep-
etition time = 8.2 msec; echo time = 3.7 msec, 220 slices,
flip angle = 8°.
fMRI Data Analysis
Image Preprocessing
MRI data were first converted to Brain Imaging Data Struc-
ture format using in-house scripts. An initial check of data
quality was done by visually inspecting the image-quality
metrics derived from MRIQC v0.15.0 (Esteban et al.,
2017). Data were preprocessed using fMRIPrep v20.1.1
(Esteban et al., 2019; RRID:SCR_016216), which is based
on Nipype 1.5.0 (Gorgolewski et al., 2011; RRID:
SCR_002502), with the default processing steps. These
included brain extraction, segmentation, and surface recon-
struction of the structural T1 image; spatial normalization
of both the structural and functional data to MNI space;
and head motion estimation, coregistration, susceptibility
distortion correction, and resampling to 2 mm3 of the
functional data. No slice-timing correction was performed.
A comprehensive description of the preprocessing pipe-
line is available here: https://osf.io/72bsh.
fMRI Statistical Analyses
The preprocessed functional data were further analyzed
using Statistical Parametric Mapping software (SPM12,
Contrast
Region
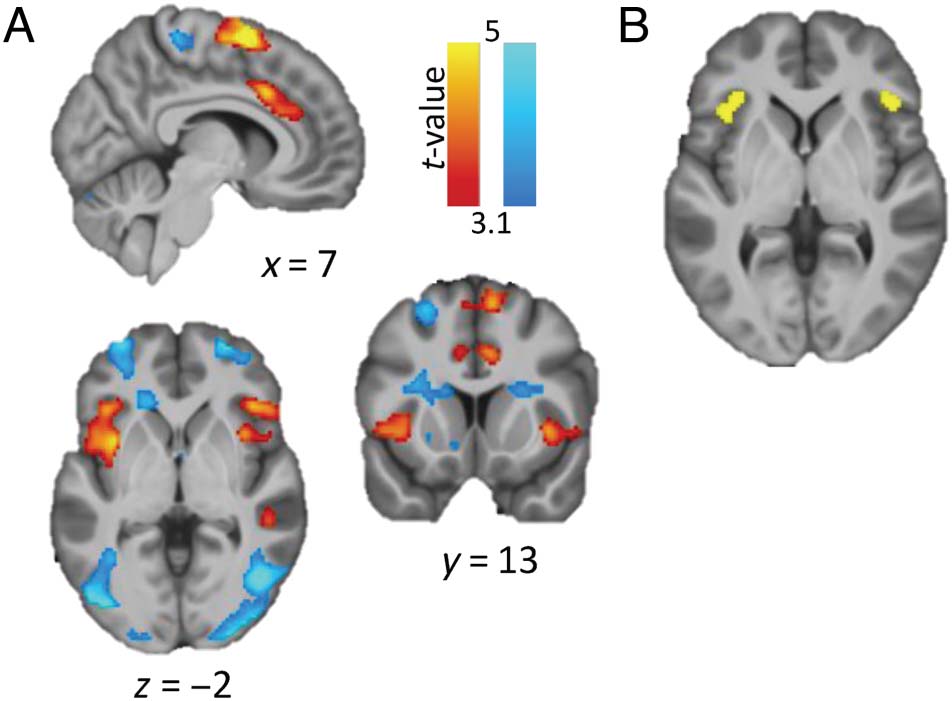
Increase over training
Caudate nucleus head
blocks (go)
Amygdalo-hippocampal
junction
Angular gyrus
Posterior putamen
Decrease over training
Anterior caudate L
blocks (go)
Anterior caudate R
Primary motor/SMA
Hippocampus/putamen
Temporal cortex L
Goal > implementation
Anterior caudate
intentions block-set 1 (Go)
SVC = small volume correction; L = left; R = right.
MNI Coordinates
(x, y, z)
Cluster Size
( Voxels)
z Score at
Peak Level
Correction
22
−10
20
26
−24
24
8
43
−46
13
6
−4
−52
−20
10
10
−24
14
−46
18
30
−14
38
4
2
−4
60
−8
−4
−4
443
348
214
34
912
537
860
657
591
40
4.37
5.17
4.92
3.96
6.53
6.39
5.44
4.83
5.69
3.69
Cluster
Peak
Peak
SVC Tricomi
Cluster
Cluster
Cluster
Cluster
Cluster
SVC striatum
van Timmeren et al.
963
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
5
6
9
5
7
2
0
8
2
8
8
1
/
j
o
c
n
_
a
_
0
1
9
9
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Wellcome Trust Centre for Neuroimaging). The data were
spatially smoothed using a Gaussian kernel with a FWHM
of 8 mm and all functional data was high pass filtered
(with a 128-sec cutoff ) to remove slow signal drifts.
First-Level Analysis
For the first-level analysis of the fMRI data, a general linear
model was constructed for each participant, concatenated
over all three runs from the training and test phase. For the
data on training with intentions, trial onsets of valuable
stimuli and nonvaluable stimuli for implementation and
goal intentions were modeled using stick functions, mak-
ing four conditions. To look at the effect of time on train-
ing, these were modeled as separate regressors per three
blocks, making four training block-sets. Only correct trials
(i.e., where an accurate (non)response was made) were
included. Blocks of verbal rehearsal of implementation
and goal intentions were additionally modeled as blocks
of 28 sec (total duration of eight 3.5-sec trials). For the test
phase, stick functions modeled the trial onsets of still-
valuable and still-not-valuable (“value-congruent”; the
outcome value is congruent with training phase) and deva-
lued and upvalued (“value-incongruent”; the outcome
value is not congruent with training phase) stimuli that
were trained with implementation or goal intentions
separately, making eight regressors. To investigate BOLD
activity during habitual (c)omission errors (habitual “slips”
in case of incongruent trials), separate regressors were
included for incorrect trials for all conditions. The follow-
ing regressors of no interest were included separately for
each run: one regressor for errors (only for training, as
test-errors/“slips” were modeled as regressors of interest)
and late trials, keypresses, feedback-displays, value-
screens (only for test phase), and six realignment parame-
ters capturing rotation and translation to correct for
residual participant motion. Three session constants were
included in the model. All onsets were then convolved
with the canonical hemodynamic response function, and
an autoregressive AR(1) model was used to correct for
serial correlations. The general linear model was regressed
against the fMRI data to generate parameter estimates for
each participant.
Regressor-specific first-level contrast images were cre-
ated for the training- and test-regressors modeling the dif-
ferent conditions of interest to construct the planned
second-level full factorial models. These contrasts of
parameter estimates were then entered into between-
subjects ANOVAs to generate group-level random-effects
statistics. To test for a difference in learning between
intention types, contrasts of parameter estimates of the
instrumental training phase were entered into a 2 × 4 ×
2 (Value × Block-set × Intention) factorial ANOVA. Fol-
lowing estimation of the second-level model, t tests were
specified by adding linear weights to each instrumental
training block-set, modeling increases over training as
[−1.5 −0.5 0.5 1.5] and decreases as [1.5 0.5 −0.5 −1.5].
In addition, first-level contrast images were created. To
assess the effect of planning during training, contrasts were
created comparing training with implementation versus
goal intentions (across all blocks, separately for go and
no-go trials). To examine markers of goal-directed control
during test, we compared correct congruent trials with cor-
rect incongruent trials (i.e., [still-valuable go > upvalued
go] and [still-not-valuable no-go > devalued no-go]). We
also investigated situations where participants fail to adapt
to the new outcome value and continue to respond accord-
ing to the learned S–R association by comparing incorrect
incongruent trials (i.e., “slips of action”) with correct
incongruent trials. Again, separate contrasts were created
for test-go- and test-no-go trials (i.e., [devalued go > upva-
lued go] and [upvalued no-go > devalued no-go]). Finally,
we also created a similar contrast comparing incorrect
incongruent trials (slips) with correct congruent trials
(i.e., [devalued go > still-valuable go] and [upvalued no-
go > still-not-valuable no-go]). More information about
the rationale behind these contrasts is provided in the
Results section. To assess the effect of planning strategy
on test performance, the same test-phase contrasts were
constructed but looking for an interaction with intention
type (e.g., [still-valuable go > upvalued go × implementa-
tion > goal intention]). Parameter estimates generated
from these first-level analyses were entered into a
random-effects group analysis, and linear contrasts were
used to identify significant effects at the group level.
Higher level whole-brain statistical maps were corrected
for FWE at the cluster-level ( pFWE-cluster < .05) with a voxel
cluster-defining threshold of p = .001 uncorrected. When
activations did not reach statistical significance at the cluster
level, we also checked the peak-voxel level with a threshold
of p < .05 corrected ( pFWE-peak < .05). In such cases, we
clearly indicate this in the text, and we report the peak-voxel
level results so as to be as comprehensive as possible in our
reporting. Finally, in an exploratory analysis, we further
aimed to test for effects in specific regions of the striatum
given prior published findings on the role of these struc-
tures in goal-directed and habitual responding ( Watson
et al., 2018; de Wit et al., 2012; Tricomi et al., 2009; Tanaka,
Balleine, & O’Doherty, 2008; Valentin et al., 2007). In partic-
ular, we defined an anatomical ROI to examine effects in the
caudate nucleus, a region previously implicated in goal-
directed processes, as well defining a functional ROI based
on the results from Tricomi et al. (2009) that implicated the
posterior putamen in habit-related processing.
In addition, we identified several ROIs in our preregistra-
tion: for habitual control, goal-directed control, response
conflict, and implementation intentions. Three separate
masks were created based on these ROIs to apply small vol-
ume correction (SVC). Apart from a striatal ROI (encom-
passing the bilateral caudate, putamen, and NAcc from
the AAL atlas (Tzourio-Mazoyer et al., 2002); however,
applying SVC with the three preregistered ROIs did not
alter the pattern of results. This may be because of the large
number of voxels included in the ROIs (especially the goal-
964
Journal of Cognitive Neuroscience
Volume 35, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
6
9
5
7
2
0
8
2
8
8
1
/
j
o
c
n
_
a
_
0
1
9
9
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
directed mask) thereby reducing the sensitivity of the SVC.
Therefore, we have opted to stick to reporting the whole-
brain results for the confirmatory analyses. Whole-brain
t-maps (without thresholding) of the main fMRI contrasts
are available at https://neurovault.org/collections/13191/.
There was no significant difference in learning to make go
versus no-go responses (main effect of Value: F(1, 40) =
2.00, p = .17, ηp
2 = .05, BFexcl = 1.60; Block × Value inter-
action: F(1.70, 68.16) = .25, p =. 57, ηp
2 = .01, BFexcl =
22.15).
RESULTS
All analyses reported in this section were preregistered at
the start of this study, unless indicated otherwise in the
text. We generally followed the preregistered analysis plan,
but in some cases, the results prompted us to further
explore the data. We should also point out that we prere-
gistered these hypotheses before finishing data analysis of
our related behavioral study (van Timmeren & de Wit,
2022). Hence, we preregistered the same behavioral
hypotheses for this study, although the original behavioral
study only partially supported our initial predictions—a
point we will come back to in the discussion. We therefore
incidentally deviate from the preregistration to keep our
analyses in line with analyses and findings from the behav-
ioral study, which is always clearly indicated.
The total final sample used for the analyses consisted of
41 participants, after excluding the following participants.
On the basis of the preregistered exclusion criteria, no par-
ticipants were excluded on the training criterion (< 80%
accuracy in the last block-set of training), whereas three
were excluded because they made < 25% responses on
upvalued trials trained with goal intentions in the test
phase. The goal of this criterion was to ensure that partic-
ipants understood the test-phase instructions and updated
their performance accordingly, while not excluding partic-
ipants based on the manipulation of interest (i.e. imple-
mentation intentions). We additionally excluded two
participants (post hoc) based on a very low overall
response rate during the test phase. Although these partic-
ipants made (just) > 25% upvalued responses, we deviated
from the preregistration because they were outliers on the
overall response rate and responded on less than one out
of three trials during the test, despite receiving explicit
instruction to aim for a response rate of ∼50% and receiv-
ing feedback about that at the end of each block. Hence,
they did not follow the test-phase instructions and their
performance is not reliable. Note that this criterion is
independent of actual task performance (accuracy) and
that the in-/exclusion of these two participants does not
change the general pattern of behavioral nor fMRI results.
Behavioral Results
Training Phase without Intentions
As expected, participants learned to make correct responses
over the first part of training (Figure 2A), as revealed by a
significant main effect of Block-set on accuracy, F(2.46,
98.20) = 16.74, p < .001 ηp
2 = .30, BFincl = 2.81 × 105,
and a marginally significant effect of Block-set on RT,
F(2.45, 98.07) = 2.75, p = .058, ηp
2 = .06, BFincl = 0.81.
Instrumental Training with Goal versus
Implementation Intentions
Following the first 12 blocks of instrumental training
without planning, intentions were introduced during a
practice block (still outside the scanner). Although we
did not preregister to analyze those data, for completeness
and in line with our previous behavioral study with this par-
adigm investigating the same question (van Timmeren &
de Wit, 2022), we conducted a paired t test comparing
the final block of training without intentions to the practice
block. This analysis revealed that participants benefitted
from if–then planning on the valuable go trials, as reflected
by higher accuracy (M = 96.1, SD = 12.4) relative to the
preceding (pre)training block-set (baseline: M = 91.8,
SD = 9.1, Z(40) = 2.57, p = .01, d = 0.59, 95% CI [.81,
.22], BF10 = 1.34), whereas RTs were not affected, t(40) =
−.01, p = .99, d = −0.001. In contrast, the use of goal
intentions negatively impacted both accuracy (M = 87.6,
SD = 14.7, Z(40) = −1.86, p = .065, d = −0.40, 95% CI
[−.01, −.69], BF10 = 1.36) and RTs, t(40) = −2.03, p =
.049, d = −0.32, BF10 = 1.08, of go trials compared with
(pre)training. For no-go trials, no significant effects of imple-
mentation, Z(40) = 1.03, p = .31, BF01 = 5.12, or goal inten-
tion, Z(40) = .10, p = .93, BF01 = 5.68, were seen.
Subsequently, when instrumental training was resumed
during the scanning session, the 2 × 2 × 2 repeated-
measures ANOVA indicated that the advantage of if–then
planning was initially still apparent on valuable go trials
(Figure 2A). In addition to a strong main effect of Value,
driven by participants performing better overall on valu-
able compared with nonvaluable trials, F(1, 84.47) =
10.93, p = .002, ηp
2 = .22, BFincl = 18.08, we found the
expected preregistered three-way interaction between
Intention, Value, and Block-set, F(3, 103.14) = 6.45,
p < .001, ηp
2 = .14, BFincl = 857.7. Separate analyses of
valuable and nonvaluable trials revealed a significant
Intention × Block interaction for valuable, F(3, 81.78) =
6.21, p = .003, ηp
2 = .13, BFincl = 74.01, but not for non-
valuable trials, F(3, 120) = 1.88, p = .14, ηp
2 = .05, BFexcl =
2.63. The significant effect on the valuable go trials was
driven by higher accuracy with implementation compared
with goal intentions during the first block-set, Z(40) =
3.34, p < .001, d = 0.85, 95% CI [.64, .94], BF10 =
22.76. At the end of training (Block-Set 4), there was no
longer a significant effect of Intention Type on accuracy,
Z(1, 40) = −.34, p = .80, ηp
2 = −1.43, BF01 = 5.87.
The analysis of RTs (Figure 2A) revealed a main effect of
Intention Type, F(1, 40) = 12.08, p = .001, ηp
2 = .23, BFincl =
11.12, with faster responses during blocks trained with
implementation intentions (median = 365 msec, SD =
van Timmeren et al.
965
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
6
9
5
7
2
0
8
2
8
8
1
/
j
o
c
n
_
a
_
0
1
9
9
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
6
9
5
7
2
0
8
2
8
8
1
/
j
o
c
n
_
a
_
0
1
9
9
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
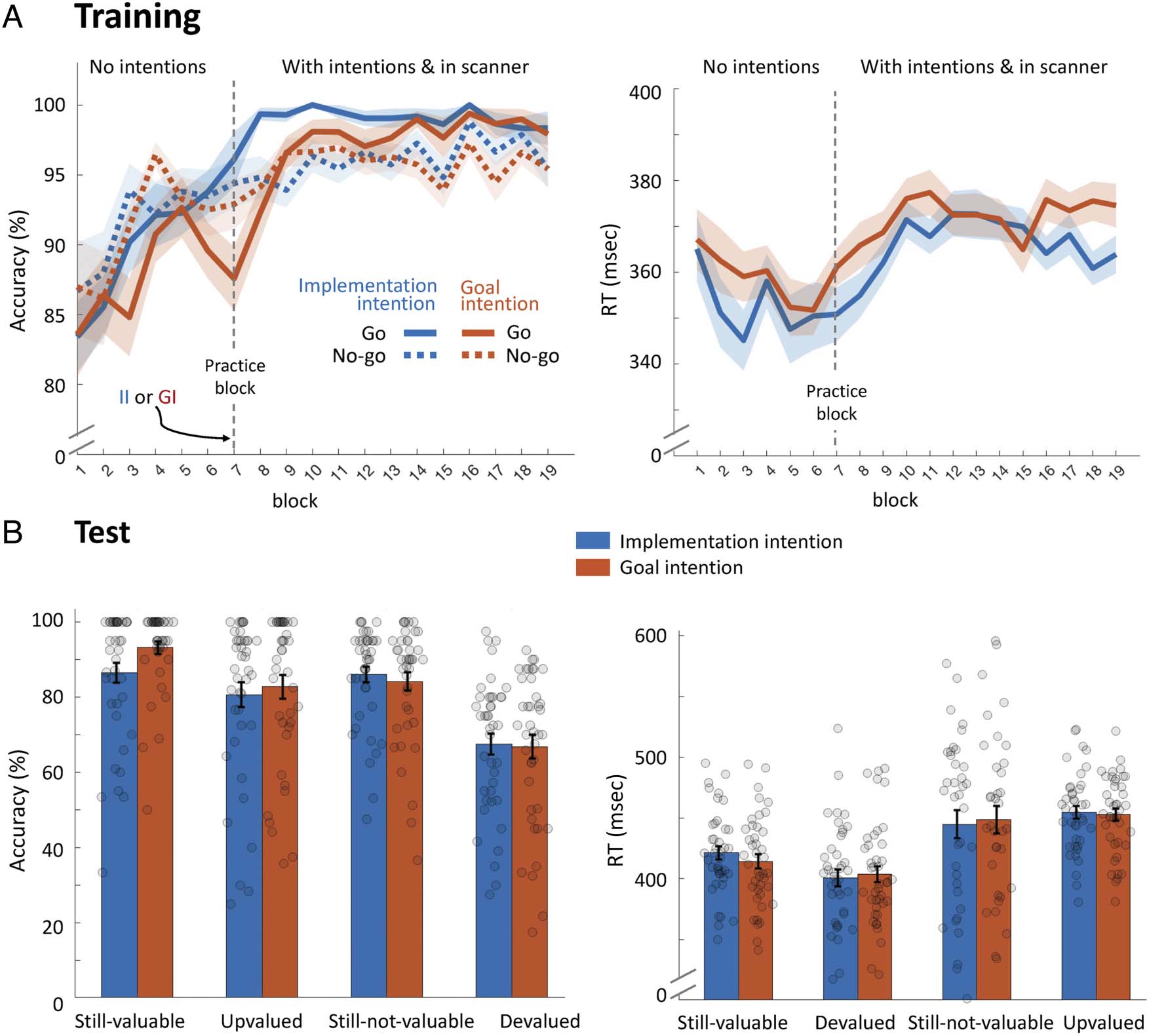
Figure 2. Behavioral results. (A) Over the course of training, participants learned to successfully respond for stimuli associated with valuable
outcomes (Go) and to withhold making a response for stimuli associated with nonvaluable outcomes (no-go), as reflected by increasing accuracy
rates. After six blocks of regular training, some stimuli continued to be trained using implementation intentions (blue) whereas others were trained
with goal intentions (blue). Following one block of practice (black dotted line), participants were moved to the scanner and resumed training
with intentions. Accuracy was significantly higher initially when using implementation intentions, but toward the end of training performance was
almost perfect for both implementation and goal intentions. Across training with intentions, participants were faster during blocks trained with
implementation versus goal intentions. (B) During the test phase, for some stimuli, the associated outcome changed in value (and thus response)
compared with training (upvalued and devalued; see Figure 1C) and participants had to flexibly update their responses accordingly. For other stimuli,
the associated value and response remained congruent with training (still-valuable and still-bot-valuable). Participants responded less accurately for
incongruent compared with congruent trials, reflecting inflexibility as a consequence of learned S–R contingencies during training. However, training
with implementation intentions did not lead to reduced flexibility. Similarly, there was no significant effect of training with implementation intentions
on RT. (Shaded) error bars represent standard error of the mean. II = implementation intentions; GI = goal intentions.
17) compared with goal intentions (median = 374 msec,
SD = 20), but no significant effect of Block-set, F(2.4,
98.6) = 2.31, p = .09, ηp
2 = .05, BFexcl = 3.41, nor an inter-
action ( p = .20, ηp
2 = .04, BFexcl = 3.67).
Symmetrical Outcome-Revaluation Test
As expected, learned S–R associations had a clear impact
on performance during the test phase (Figure 2B), as
revealed by a main effect of Congruence, F(1, 40) =
65.08, p < .001, ηp
2 = .62, BFincl = 1.39 × 107. Because
test value showed significant interactions with both
Congruence, F(1, 40) = 10.73, p = .002, ηp
2 = .21, BFincl =
8.91, and Intention Type, F(1, 40) = 5.94, p = .02, ηp
2 =
.13, BFincl = 1.27, separate follow-up comparisons were
conducted for go (associated with still-valuable and
upvalued outcomes) and no-go (associated with still-
not-valuable and devalued outcomes) trials. Main effects
of Congruence were seen for both the go, F(1, 40) =
16.82, p < .001, ηp
2 = .30,, BFincl = 76.40, and no-go,
F(1, 40) = 56.46, p < .001, ηp
2 = .59, BFincl = 2.31 ×
106, stimuli. As can be seen in Figure 2B, the congruency
effect was larger for no-go trials mainly because of partic-
ipants struggling more on devalued trials, where they had
966
Journal of Cognitive Neuroscience
Volume 35, Number 6
to suppress responding to discriminative stimuli that pre-
viously signaled a valuable outcome. Importantly, we
were interested in the effect of implementation inten-
tions on test performance. First, an analysis of the go test
trials suggested that overall performance was worse when
trained with implementation compared with goal inten-
tions, F(1, 40) = 5.48, p = .02, ηp
2 = .12, although Bayesian
statistics showed that this evidence was inconclusive
(BFincl = 1.46). Importantly, in contrast to our preregis-
tered hypothesis, there was no evidence for reduced flex-
ibility as a consequence of if–then planning: The expected
interaction of congruence with intention type failed to
reach significance, F(1, 40) = 1.52, p = .23, ηp
2 = .04,
BFexcl = 1.86. Given the direct relevance of the compari-
son between intentions for our research question, we
followed these analyses up with separate (exploratory)
paired t tests for still-valuable and upvalued trials to also
report Bayesian evidence against a difference. Findings
indicate that intentions only had a significant negative
effect on (congruent) still-valuable, Z(40) = −2.55, p =
.01, d = −0.56, BFincl = 3.68, but not on (incongruent)
upvalued trials, t(40) = −.75, p = .46, BFexcl = 4.54.
Finally, for the no-go stimuli (still-not-valuable and deva-
lued), no main, F(1, 40) = .42, p = .52, BFexcl = 4.37, nor
interaction, F(1, 40) = .06, p = .81, BFexcl = 4.25, effects
of intention type were observed.
We also analyzed RTs during the test phase. A Value ×
Congruence interaction, F(1, 32) = 49.47, p < .001, ηp
2 =
.61, BFincl = 2.91 × 105, prompted separate analyses for
trials trained with go responses (still-valuable and deva-
lued) and for trials trained with no-go responses (still-
not-valuable and upvalued). Interestingly, there was a
main effect of congruence for go-trained stimuli, suggesting
significantly faster RTs on devalued trials (M = 418 msec,
SE = 8.8) relative to still-valuable (M = 443 msec, SE =
6.8; F(1, 40) = 12.56, p = .001, ηp
2 = .24, BFincl = 23.40),
in line with the idea that habitual slips of action are trig-
gered fast and efficiently before one has the chance to
suppress them. As late responses were excluded from
this analysis (following the accuracy analysis), we ran an
additional analysis including RTs for late responses to
make sure that this effect was not driven by a higher
number of (excluded) late responses on devalued trials.
This analysis showed an even stronger main effect of con-
gruence than the original analysis without late responses,
F(1, 40) = 14.84, p < .001, ηp
2 = .27, BFincl = 36.88. No
other significant effects of RTs were found (all p > .22,
BFexcl > 1.74).
Self-reported Automaticity and S-O Knowledge
Self-reported automaticity was at a high level overall
(median = 80.4%, SD = 16.7), but did not differ between
intentions, t(40) = −.98, p = .34, BF01 = 3.80, nor did sub-
jective automaticity correlate with revaluation insensitivity
for implementation (rτ = −.09, p = .57, BF01 = 4.39) or
goal intentions (rτ = .22, p = .17, BF01 = 2.03).
Following van Timmeren and de Wit (2022), we also
explored differences in S-O knowledge between intention
types and their relationship with overall test accuracy. S-O
knowledge was high (median = 89.8%, SD = 22.1) and,
contrary to our previous study, no longer differed signifi-
cantly between intention types, F(1, 40) = 2.07, p = .16,
ηp
2 = .05, BF01 = 2.6; values, F(1, 40) = 3,42, p = .07, ηp
2 =
.08, BF01 = 2.4; or their interaction, F(1, 40) = .91, p = .35,
ηp
2 = .02, BF01 = 5.88, suggesting that the adaptation we
made to the task (i.e., using a pseudorandom selection of
stimuli instead of alternating between two block-sets in
the first part of training, see Methods section) had the
desired effect. S-O knowledge did correlate positively with
test accuracy (across all four conditions) for both imple-
mentation intentions (rτ = .30, p = .008, 95% CI [.08,
.52], BF10 = 7.91) and goal intentions (rτ = .39, p <
.001, 95% CI [.21, .57], BF10 = 99.22).
Conclusions: Behavioral Results
We provide evidence for habit learning, as indicated by the
general effect of previously learned S–R mappings on the
ability to flexibly adapt responding when the cue signals a
revalued outcome (i.e., incongruent). Importantly,
although if–then planning seemed to increase efficiency
relative to goal intentions, as reflected in superior acquisi-
tion, this was not at the expense of flexibility when out-
come values changed in the test phase.
Neuroimaging Results
Instrumental Training: Across Intentions (Exploratory)
First, we were interested to explore general learning
effects across intention types because this was the first
time the SORT was used in the MRI scanner. These
analyses showed that over the course of go training (i.e.,
on valuable trials), activity increased linearly in the head
of the caudate nucleus extending into ACC (at p < .05
FWE rate corrected; pFWE-cluster < .05). Activation in the
left amygdalo-hippocampal junction and the angular
gyrus did not reach our cluster-level correction thresh-
old, but did survive voxel-level correction at p < .05
( pFWE-voxel < .05; Table 1). In this same contrast, we
also observed a cluster in the posterior putamen, which
survived a small-volume correction for the posterior
putamen ROI (i.e., pFWE < .05 with SVC, defined as a
10-mm sphere at peak value of the cluster that showed a
significant increase over training in the study of Tricomi
et al. [2009]; x = 33, y = −24, z = 0). On the other
hand, activity decreased over training in the bilateral
anterior caudate (a more ventral part of the striatum),
primary motor cortex (extending to mid-posterior cin-
gulate), hippocampus extending into the putamen, and
the left temporal cortex (all pFWE-cluster < 0.05 cor-
rected). In contrast, on no-go trials, there were no vox-
els that showed a significant linear change over training
blocks.
van Timmeren et al.
967
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
6
9
5
7
2
0
8
2
8
8
1
/
j
o
c
n
_
a
_
0
1
9
9
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Instrumental Training: Comparing Goal and
Implementation Intentions
We then examined whether strategic planning affected
instrumental training. The contrast comparing the aver-
age BOLD signal of trials trained with implementation
intentions and goal intentions did not reveal any signif-
icant activations, neither on go nor no-go trials. We
also tested for differences in learning between inten-
tions over the course of training by adding linear
weights to block-sets to compare increased activity
over block-sets during implementation intentions with
decreased activity during goal intentions, and vice versa.
However, both tests of this interaction failed to show sig-
nificant differences.
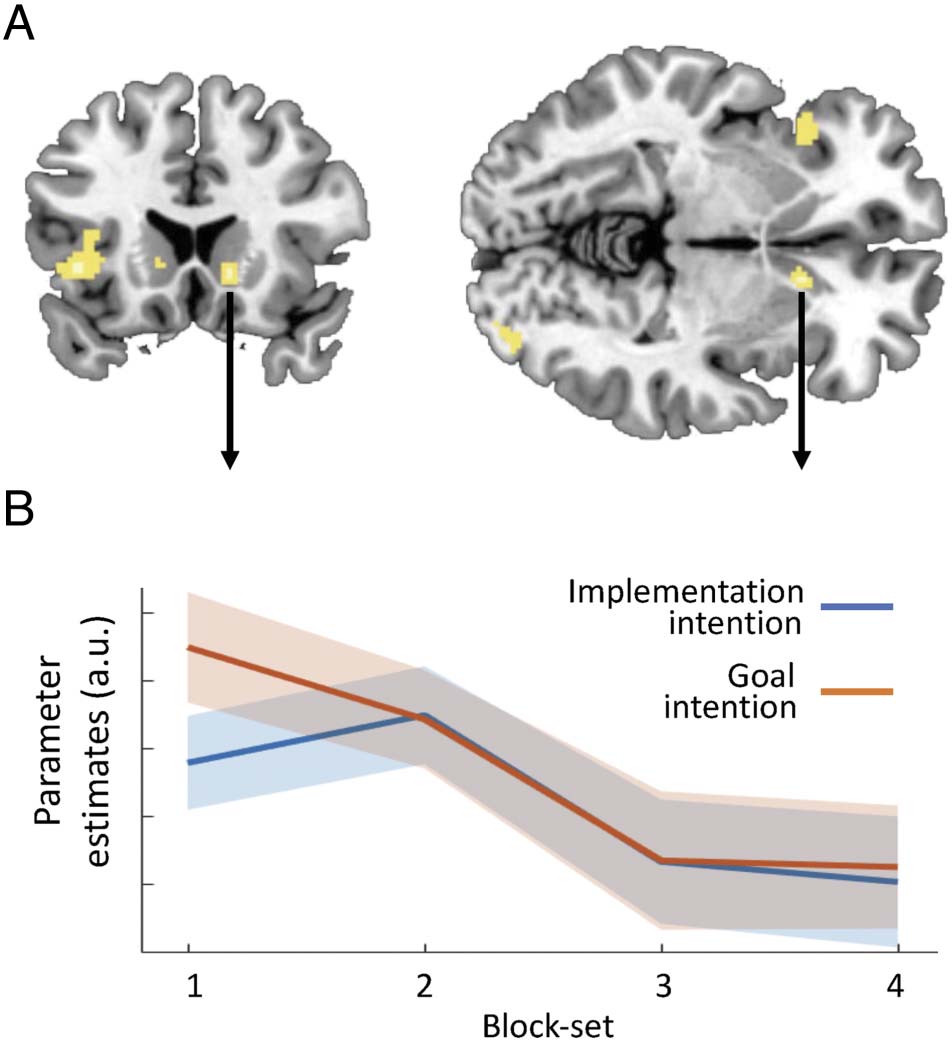
The finding that implementation intentions showed the
most pronounced effect behaviorally early in training
prompted us to conduct an exploratory analysis of only
the first training block-set. This analysis revealed signifi-
cantly decreased activation in the anterior caudate ( pFWE <
.05 with SVC, z = 3.69) on trials trained with implementa-
tion intentions compared with goal intentions (Figure 3A
and Table 1). For visual purposes, the extracted average
BOLD signal from the anterior caudate cluster is shown
separately for each block-set and intention in Figure 3B.
As can be seen here, activity was indeed lower on imple-
mentation intention trials during the first block-set only
and subsequently decreased for both intentions. A
whole-brain analysis also showed decreased activity for
implementation relative to goal intentions at an uncor-
rected threshold ( p < .001) in the right lateral orbitofron-
tal cortex (OFC; pFWE-cluster = .061, z = 4.25; x = 26, y =
50, z = 14) and the left insula ( pFWE-cluster = .28, z = 3.76;
x = −42, y = 20, z = 2). However, because these results
did not survive FWE-correction, we refrain from interpret-
ing them further. To rule out that these findings were
driven by RTs, which were significantly shorter for imple-
mentation compared with goal intentions, we performed
an additional analysis controlling for trial-by-trial RT by
including a parametric regressor (one for each of the
two training runs) with RTs for each trial. This had no sig-
nificant impact on the results, and we could qualitatively
replicate all reported findings.
Neural Predictors of Test Performance
To determine whether brain activity during instrumen-
tal training with implementation intentions was pre-
dictive of test performance, we tested whether the
average BOLD signal during training covaried with
the revaluation insensitivity score. This preregistered test
did not reveal significant neural predictors of test perfor-
mance. For completeness, we also exploratively ran this
analysis separately for goal intentions and across inten-
tions, but this similarly did not reveal any significant
results.
Figure 3. Lower activity in the right anterior caudate early in training
for implementation compared with goal intentions. (A) Voxels that
showed significantly lower activation during the first block-set of
training with implementation compared with goal intentions on go-trials
(at pFWE < .05, small volume-corrected). The activity patterns shown
are thresholded at p < .001 uncorrected. (B) Parameter estimates
extracted from this anterior caudate cluster (peak at x = 13, y = 18,
z = −4) over block-sets. Error bars represent 95% confidence intervals.
a.u. = arbitrary units.
Symmetrical Outcome-Revaluation Test: Markers of
Goal-directed versus Habitual Performance
In the test phase, changes in outcome value create con-
flict between goal-directed control and learned S–R asso-
ciations. Specifically, to perform the correct response on
incongruent trials (i.e., upvalued go and devalued no-
go), participants have to exert goal-directed control
and override the learned S–R mapping. Conversely, on
congruent trials (still-valuable go and still-not-valuable
no-go), participants can rely on the learned S–R associa-
tions. The advantage of the symmetrical outcome-
revaluation test (compared with the original slips of
action test) is that we can compare congruent and
incongruent trials with each other unconfounded by test
outcome value (and therefore required response: i.e., go
or no-go). Therefore, to examine markers of goal-
directed control, we firstly compared upvalued go with
still-valuable go responses and found that this was asso-
ciated with increased right insula activity ( pFWE-cluster <
.05, z = 4.16; Table 2). No significant activations were
seen in the contrast between devalued no-go and still-
not-valuable no-go trials.
To identify regions where participants fail to adapt and
continue to respond according to the learned S–R
968
Journal of Cognitive Neuroscience
Volume 35, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
6
9
5
7
2
0
8
2
8
8
1
/
j
o
c
n
_
a
_
0
1
9
9
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
MNI Coordinates
(x, y, z) Max
Cluster Size
( Voxels)
z Score
(Peak)
Correction
Table 2. Imaging Results of the Test Phase
Contrast
Region
Upvalued go > still-valuable go
Insula R
Devalued slips > still-valuable go
Anterior insula L
Anterior insula R
Still-valuable go > devalued slips
vMPFC
Caudate
NAcc
Primary motor cortex
Paracentral lobule
Angular gyrus L
IPL L
38
−40
42
22
8
4
−26
−10
−30
24
26
26
42
28
20
12
−30
−52
Angular gyrus R
38
−50
IPL R
Occipital cortex
Devalued slips > upvalued go
Anterior insula L
SMA
dACC
Inferior parietal lobule L
Inferior parietal lobule R
Supramarginal gyrus
Upvalued go > devalued slips
ACC, caudate nucleus
Premotor/PMC
Lateral OFC
Superior parietal love L
Superior parietal love R
Occipital/visual cortex
−36
−36
8
8
−56
56
−36
−20
−26
−32
−28
30
−30
−74
26
8
18
−42
−4
26
22
0
62
−76
−62
−96
−2
2
−10
−4
2
4
60
66
52
58
8
−8
64
34
34
44
−8
18
42
0
36
38
16
468
611
621
388
252
336
1653
4.16
5.46
4.49
4.64
5.10
4.56
5.51
Cluster
Cluster
Cluster
Cluster
Peak
Cluster
Cluster
2510
5.33
Cluster
767
707
378
431
269
331
707
327
529
317
1482
4099
1307
5.25
4.29
5.47
4.18
4.60
4.44
4.29
4.17
4.80
4.19
4.12
5.04
6.43
Cluster
Cluster
Cluster
Cluster
Peak
Cluster
Cluster
Cluster
Cluster
Cluster
Cluster
Cluster
Cluster
L = left; R = right; NAcc = nucleus accumbens; IPL = inferior parietal lobule; (d)ACC = (dorsal) anterior cingulate cortex; PMC = primary motor
cortex; OFC = orbitofrontal cortex.
association, we contrasted incorrect incongruent trials
(devalued go and upvalued no-go) to correct incongruent
trials (upvalued go and devalued no-go, respectively), as
the latter arguably require most goal-directed control to
override the learned S–R mapping. The contrast compar-
ing devalued go responses (i.e., slips of action) with
upvalued go responses is shown in Figure 4A, and
revealed increased activity in a fronto-parietal network,
including the left anterior insula extending to the inferior
lateral prefrontal cortex, SMA, dorsal anterior cingulate
cortex, bilateral inferior parietal lobule, and supramargi-
nal gyrus (all pFWE-cluster < .05; Table 2). Conversely,
lower activity during slips of action compared with upva-
lued go responses was seen in the left anterior cingu-
late cortex extending into caudate nucleus, left lateral
lobe, and several
OFC, bilateral superior parietal
occipital/primary visual areas (all pFWE-cluster < .05,
Table 2). In addition, activation in the premotor/primary
motor cortex did not survive cluster-level correction but
did reach peak-voxel level significance ( pFWE-voxel < .05).
Although the previous contrast between devalued slips
and correct upvalued go responses maximizes the differ-
ence between habitual versus goal-directed control, the
conditions differ in terms of the original training out-
come value (as well as test value). To mitigate this, we
proceeded to compare devalued slips to still-valuable go
responses, which only differ in their test outcome value.
Thus, this contrast compares trials on which participants
correctly continued responding according to the learned
S–R association with trials on which they failed to over-
ride this association. Although we have used the same
approach previously (in the study of Watson et al.,
van Timmeren et al.
969
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
6
9
5
7
2
0
8
2
8
8
1
/
j
o
c
n
_
a
_
0
1
9
9
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
nonvaluable outcome (devalued) with stimuli signaling a
valuable outcome (upvalued or still-valuable). To control
for this, we ran some additional exploratory analyses, com-
paring activity during devalued slips with correct no-go
responses on devalued and still-not-valuable trials.
Although these contrasts are difficult to interpret by
themselves—they are themselves confounded by pressing
a button or not—looking at the overlap between all four
contrasts overcomes the value-related confounds and
hence could find a common process in the expression of
habits. To this end, we used ImCalc to create binary
images of all four contrasts thresholded at t(41) = 3.1
(equivalent to p < .001 uncorrected) and multiply them.
The result of this inclusive masking analysis, which is akin
to a conjunction analysis, shows that the bilateral anterior
insula was commonly activated across all four contrasts
(Figure 4B).
Symmetrical Outcome-Revaluation Test: Comparing
Goal and Implementation Intentions
None of the planned contrasts comparing test-phase trials
trained with implementation with goal intentions revealed
significant activation patterns.
DISCUSSION
The aim of the present study was to investigate whether
the brain can strategically go on automatic pilot. We inves-
tigated this by measuring the impact of strategic planning
(i.e., implementation intentions vs. goal intentions) on the
acquisition of instrumental actions as well as subsequent
flexible, behavioral adjustment. When strategic planning
was first introduced during the instrumental learning
phase of our paradigm, implementation intentions
improved performance relative to goal intentions. Fur-
thermore, in line with the idea that their beneficial effect
was mediated by accelerated S–R learning, an exploratory
analysis revealed that implementation intentions were
associated with reduced activity in the anterior caudate,
a brain area previously implicated in goal-directed control
( Watson et al., 2018; Liljeholm, Tricomi, O’Doherty, &
Balleine, 2011). These effects of strategic planning on per-
formance and neural activity were only apparent early in
training, with participants reaching high levels of accuracy
(and reduced activity in the anterior caudate) by the end of
the learning phase independent of intention type. Our
central question, however, was whether implementation
intentions would actually impede performance when
flexible, behavioral adjustment was required during the
subsequent outcome-revaluation test. Importantly, we
found no evidence for a detrimental effect of strategic
planning on the ability to adapt behavior to changing
outcome values, nor any effect on underlying neural activ-
ity patterns. We conclude that strategic planning of S–R
mappings may allow people to go on automatic pilot to
Figure 4. (A) Neural correlates of slips of action in the test phase,
as revealed by increased (red – yellow) and decreased (dark – light
blue) activity during devalued slips compared with upvalued responses.
Clusters that survived whole-brain FWE correction include increased
activity in a fronto-parietal network, including the left anterior insula
extending to the inferior lateral pFC, SMA, dorsal anterior cingulate
cortex, bilateral inferior parietal lobule, and supramarginal gyrus.
Conversely, lower activity was seen in the left anterior cingulate
cortex extending into caudate nucleus, premotor/primary motor
cortex, left lateral OFC, bilateral superior parietal lobe, and several
occipital/primary visual areas. Results are shown here at p < .001
(uncorrected) for visual purposes, overlaid on the mean T1 image
of all participants. (B) The bilateral anterior insula was found to be
commonly activated during devalued slips (x = ±40, y = 26, z = 2).
Shown here in yellow are the voxels that overlap between all four
contrasts comparing devalued slips relative to correct (non-)responses
during still-valuable, still-not-valuable, devalued and upvalued trials
(thresholded at p < .001 uncorrected).
2018, the “slips versus respond valuable” contrast), this
contrast was not preregistered and should thus be con-
sidered exploratory. Similar to the comparison with
upvalued go responses, this comparison of slips with
still-valuable go responses revealed increased anterior
insula activity (bilaterally) during slips, but decreased
activity in vMPFC (extending to NAcc), primary motor
cortex, paracentral lobule, a large occipital cluster, and
large parietal clusters (bilateral) including the angular
lobule (all pFWE-cluster <
gyrus and inferior parietal
0.05; Figure 4A).
As preregistered, we also compared upvalued no-go
responses (“inhibition slips”) to correct devalued (no-
go) trials, but this did not reveal any significant activation
patterns. Moreover, we were not able to conduct the
contrast between upvalued and still-valuable no-go trials,
because of the low number of omission errors on still-
valuable trials.
Our results thus identify the anterior insula as a com-
mon region associated with slips toward devalued out-
comes, as activity in this region was higher during slips
than during go responses toward upvalued and still-
valuable outcomes. However, both contrasts are con-
founded by expected value (the outcome value during
the test phase) as they both compare stimuli signaling a
970
Journal of Cognitive Neuroscience
Volume 35, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
6
9
5
7
2
0
8
2
8
8
1
/
j
o
c
n
_
a
_
0
1
9
9
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
increase behavioral efficiency, but that this does not have
to come at the expense of behavioral flexibility. Therefore,
mental rehearsal of S–R links does not appear to suffice for
the formation of a rigid habit, refuting the notion of
“instant habits” and suggesting that behavioral repetition
may be crucial for the development of rigid habits.
To shed light on the implications of these findings, we
will first discuss them in some more detail, including the
basic results (i.e., across strategies) on the relatively novel
SORT. First, during the instrumental learning phase, we
observed increasing accuracy and decreasing RTs over
the course of training, suggesting that participants
acquired the S–R mappings. This interpretation was fur-
ther supported by high levels of subjective automaticity
of responding at the end of the instrumental learning
phase and increasing involvement of two distinct parts of
the dorsal striatum: the posterior putamen, replicating
findings from Tricomi et al. (2009), and the caudate
nucleus head. Several previous fMRI studies have indi-
rectly implicated the dorsal striatum in habit learning,
either showing that with longer instrumental training this
region becomes more active ( Wunderlich, Dayan, &
Dolan, 2012; Tricomi et al., 2009) or that functional con-
nectivity with the (pre-)motor cortex increases (Zwosta
et al., 2018; Horga et al., 2015). Although increased activity
of the posterior putamen was only significant with small-
volume correction (so not very robust), activity of the
caudate nucleus as well as the hippocampus survived
whole-brain correction. Both regions have previously
been implicated in the encoding of S–R representations
(McNamee, Liljeholm, Zika, & O’Doherty, 2015). More-
over, we found decreasing activity of the primary motor
cortex (extending to mid-posterior cingulate), the hippo-
campus (extending into the putamen), bilateral temporal
cortex, and the right anterior caudate, previously implica-
ted in goal-directed control (Liljeholm et al., 2011; Balleine
& O’Doherty, 2010). In line with previous findings, these
results suggest that dissociable neural regions support
instrumental learning. Notably, these findings were
specific to learning to make go responses for valuable out-
comes. In contrast, we did not see any changes in neural
activity over the course of no-go training, despite high
accuracy and reported automaticity at the end of training.
Thus, our neuroimaging analyses do not provide evidence
for the development of “inhibition habits” ( Jahanshahi,
Obeso, Rothwell, & Obeso, 2015).
Importantly, when strategic planning was introduced
after the first 12 training blocks, implementation inten-
tions initially improved go performance (reflected by
higher accuracy during the first block-set than in the pre-
ceding (pre-)training block-set), whereas goal intentions
impaired it (reflected by lower accuracy and slower RTs).
In contrast, no-go learning (i.e., withholding a response
for nonvaluable stimuli) was not affected by planning
strategy. These findings replicate our previous results
(van Timmeren & de Wit, 2022). Furthermore, in line with
the notion of “instant habits,” this behavioral effect of
implementation intentions early in training was accompa-
nied by reduced activity in the anterior caudate relative to
goal intentions. This early effect of implementation inten-
tions quickly disappeared, however, and no differences
with goal intentions were observed on accuracy and RTs
in later training blocks, nor on subjective automaticity after
training. Therefore, in support of the notion of strategic
automaticity, it appears that instrumental acquisition ini-
tially benefitted from if–then planning, while dependency
on goal-directed control (as suggested by anterior caudate
activity) was reduced.
So far, there have been very few neuroimaging studies
that have compared the use of implementation and goal
intentions to support behavioral performance. One study
(Gilbert, Gollwitzer, Cohen, Oettingen, & Burgess, 2009)
showed that implementation intentions engaged the
medial BA10 more (and lateral BA10 less) than a control
condition, which was argued to reflect increased cue mon-
itoring (and reduced internal information processing).
This contrasts with our finding that implementation inten-
tions lead to reduced engagement of the anterior caudate
during the instrumental learning phase. However, their
control condition was very different to ours. Whereas they
specified the cue and the outcome that it signaled to be
available (i.e., the S-O contingency; “if the cue appears,
then I can score 5 points”), we used a goal intention con-
trol condition that specified the R-O contingency, which is
arguably more akin to a typical goal intention (e.g., “I will
exercise to lose weight”).
In the next phase of the SORT, signaled outcome values
changed, requiring flexible adaptation of responding to
the discriminative stimuli. This allowed us to determine
whether strategic planning (during training) would induce
the rigidity that is commonly regarded as a hallmark of
learned habits that are stamped in through behavioral rep-
etition. However, we failed to find convincing evidence
that if–then planning impaired the ability to flexibly adjust
responding when signaled outcome values changed. This
was despite the fact that participants struggled to adjust
learned S–R mappings overall, as reflected in a strong
main effect of congruency. Furthermore, in line with the
behavioral findings, we also found no evidence for an
impact of planning on neural activation patterns during
the extinction test phase. Therefore, this first neuroimag-
ing investigation of the effect of implementation inten-
tions on behavioral flexibility in an outcome-revaluation
paradigm failed to provide evidence for a shift from goal-
directed toward habitual control.
The evidence for intact behavioral flexibility despite if–
then planning contrasts with results from an earlier study
with this paradigm (van Timmeren & de Wit, 2022). In that
study, we found that implementation intentions impaired
test-phase performance overall, but this did not lead to
inflexibility as would be reflected by lower accuracy on
incongruent trials specifically. This general impairment
was most likely because of the fact that implementation
intentions, by focusing attention on the S–R mappings,
van Timmeren et al.
971
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
6
9
5
7
2
0
8
2
8
8
1
/
j
o
c
n
_
a
_
0
1
9
9
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
blocked learning about the S-O contingencies. To prevent
this from happening in the present study, we altered our
paradigm to promote active S-O learning at first training
phase, before intentions were introduced. As a result, par-
ticipants already acquired high levels of S-O knowledge
when they started using strategic planning. Integrating
findings from both studies, it appears that when the agent
has full knowledge of the (S-O) contingencies, implemen-
tation intentions do not impair flexibility. This finding is
encouraging, because in most applied situations in real
life, agents are perfectly aware of the three-term instru-
mental contingencies. Therefore, our results are in line
with the idea of implementation intentions being “flexibly
tenacious” (Legrand, Bieleke, Gollwitzer, & Mignon, 2017;
Gollwitzer, Parks-Stamm, Jaudas, & Sheeran, 2008):
People benefit from if–then planning when the situation
specified in their plan is encountered (here in terms of
higher accuracy and lower RTs during training), but are
goal-directed in the sense that they only act on these
planned S–R mappings when the signaled outcome is cur-
rently a goal.
Across intentions, however, we found that action slips
toward devalued outcomes were associated with
increased bilateral insula (both when compared with
still-valuable and upvalued responses), replicating findings
from the only study to date looking at neural activity dur-
ing slips of action ( Watson et al., 2018). The insula is a
functionally heterogeneous region ( Uddin, Nomi,
Hébert-Seropian, Ghaziri, & Boucher, 2017), but the ante-
rior part has been critically implicated in error and salience
processing and response inhibition (Uddin, 2015; Chang,
Yarkoni, Khaw, & Sanfey, 2013). Specifically, previous
work shows that failure to inhibit a learned response (on
the stop-signal paradigm) is associated with bilateral
insular activity (Ramautar, Slagter, Kok, & Ridderinkhof,
2006). In addition, when compared with upvalued
responses, slips were associated with increased activity
in the dorsal anterior cingulate cortex, the SMA, and pari-
etal cortex, all part of the salience network (Seeley et al.,
2007). Conversely, lower activity during slips was seen in
the vMPFC, or medial OFC, when compared with
responses for still-valuable outcomes. Previous outcome-
devaluation studies suggest that activity in this region
mediates goal-directed instrumental learning (de Wit
et al., 2012; Valentin et al., 2007). A similar contrast, com-
paring devalued action slips with responses toward upva-
lued outcomes, showed lower activity in the lateral OFC
and ACC/caudate nucleus head, regions that have also
been implicated in goal-directed control (e.g., Watson
et al., 2018; McNamee et al., 2015; de Wit et al., 2012). Over-
all, our results suggest that habitual slips of action arise as a
consequence of lapses in goal-directed control (as reflected
by decreased activity in these regions) rather than by
increased activation of S–R habit regions (i.e., the dorsal
striatum). Finally, the informal conjunction analysis of
devalued slips (Figure 4B), controlling for differences
in expected value and motor response, showed that the
anterior insula was commonly activated across all contrasts,
implicating it as a key region mediating habitual action slips.
A lack of reliable, positive markers of habits is an impor-
tant issue in human habit research ( Watson, O’Callaghan,
Perkes, Bradfield, & Turner, 2022; Kruglanski &
Szumowska, 2020; De Houwer, 2019; De Houwer,
Tanaka, Moors, & Tibboel, 2018; Watson & de Wit,
2018). In the context of the present study, it begs the
question whether habit strength independently contrib-
utes to stimulus-dependent, outcome-insensitive
responding (i.e., slips of action). A recently published
study with the SORT adds weight to this concern, as we
showed there that extensive instrumental training failed
to impair test performance ( Watson, Gladwin, et al.,
2022). The lack of reliable evidence for overtraining
effects (see also de Wit et al., 2018) could mean different
things, but our current findings may offer an interesting
explanation. Specifically, we observed that when the plan-
ning manipulation was first introduced during training,
not only did implementation intentions improve perfor-
mance, but goal intentions also significantly impaired
performance. This may indicate that participants’ sponta-
neous strategy up to that point had not been to form goal
intentions, but instead to switch as soon as they could to
the more efficient strategy of focusing on the S–R map-
pings. In other words, they may have spontaneously
formed implementation intentions (Bieleke & Keller,
2021). Therefore, rather than improving their perfor-
mance with the explicit implementation intention manip-
ulation, we impaired it in the goal intention condition.
Such an early shift to reliance on S–R associations (i.e.,
within 12 blocks of training) may explain that previous
experimental studies failed to find evidence for overtrain-
ing, as their short training conditions may already have
been sufficiently long to induce this, and beyond that
early shift additional training may not have significantly
enhanced the strength of those associations. This idea
accords well with results from a study by Pool and
colleagues (2022) who found that, following outcome
devaluation on a free-operant task, already after moderate
training (12 blocks), outcome-insensitive habitual
responding was seen in the majority of participants. Our
findings further reinforce this interpretation by showing
significant changes in neural activity over the course of
this relatively short training, with activity of the anterior
caudate (implicated in goal-directed learning) decreasing
and of the dorsal striatum (implicated in habitual control)
increasing. From our study it is unclear, however, how
activity in these regions developed in the earliest stages
of instrumental training, as that took place outside the
scanner. Future research should determine how many
behavioral repetitions it takes to permit this shift to an
S–R strategy, by assessing the effect of a goal intention
manipulation at different time points during training.
Our hypothesis is that, at the start of training, this would
not yet have a negative impact—relative to implementa-
tion intentions—but that it will after a few blocks.
972
Journal of Cognitive Neuroscience
Volume 35, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
6
9
5
7
2
0
8
2
8
8
1
/
j
o
c
n
_
a
_
0
1
9
9
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
In conclusion, we provide evidence for increased effi-
ciency but preserved flexibility following strategic if–then
planning. These behavioral findings were mirrored in our
analyses of the underlying brain activity: Implementation
intentions did not reduce the engagement of goal-directed
control when goals changed, nor increase activity in habit
regions. Therefore, our findings suggest that this strategic
planning technique supports the implementation of a new
target behavior while still allowing for flexible adjustment
when goals change.
Reprint requests should be sent to Tim van Timmeren, Depart-
ment of Social, Health and Organizational Psychology, Utrecht
University, Heidelberglaan 1, Utrecht, The Netherlands, 3508
TC, or via e-mail: t.vantimmeren@uu.nl.
Data Availability Statement
Data to recreate the main behavioral analyses (with
analysis pipeline and output) are available at OSF:
https://www.doi.org/10.17605/OSF.IO/642QU. Whole-
brain t-maps (without thresholding) of the main fMRI con-
trasts are available at https://neurovault.org/collections
/13191/.
Author Contributions
Tim van Timmeren: Conceptualization; Data curation;
Formal Analysis; Investigation; Methodology; Project
administration; Visualization; Writing—Original draft;
Writing—Review & editing. John O’Doherty: Conceptu-
alization; Writing—Review & editing. Nadza Dzinalija:
Investigation; Project administration; Writing—Review &
editing. Sanne de Wit: Conceptualization; Funding Acqui-
sition; Resources; Supervision; Writing—Original draft;
Writing—Review & editing.
Funding Information
Tim van Timmeren, VIDI grant from Dutch Research
Council (Nederlandse Organisatie voor Wetenschappelijk
Onderzoek) (https://dx.doi.org/10.13039/501100001722),
grant number: Van der Gaag Fund. Sanne de Wit, VIDI
grant from Dutch Research Council (Nederlandse Orga-
nisatie voor Wetenschappelijk Onderzoek) (https://dx
.doi.org/10.13039/501100003246), grant number:
016.145.382.
Diversity in Citation Practices
Retrospective analysis of the citations in every article pub-
lished in this journal from 2010 to 2021 reveals a persistent
pattern of gender imbalance: Although the proportions of
authorship teams (categorized by estimated gender iden-
tification of first author/last author) publishing in the Jour-
nal of Cognitive Neuroscience ( JoCN ) during this period
were M(an)/M = .407, W(oman)/M = .32, M/ W = .115,
and W/ W = .159, the comparable proportions for the arti-
cles that these authorship teams cited were M/M = .549,
W/M = .257, M/ W = .109, and W/ W = .085 (Postle and
Fulvio, JoCN, 34:1, pp. 1–3). Consequently, JoCN encour-
ages all authors to consider gender balance explicitly when
selecting which articles to cite and gives them the oppor-
tunity to report their article’s gender citation balance. The
authors of this article report its proportions of citations by
gender category to be as follows: M/M = .512, W/M = .256,
M/ W = .070, and W/ W = .163.
REFERENCES
Adams, C. D., & Dickinson, A. (1981). Instrumental responding
following reinforcer devaluation. Quarterly Journal of
Experimental Psychology, Section B, 33, 109–121. https://doi
.org/10.1080/14640748108400816
Balleine, B. W. (2019). The meaning of behavior: Discriminating
reflex and volition in the brain. Neuron, 104, 47–62. https://
doi.org/10.1016/j.neuron.2019.09.024, PubMed: 31600515
Balleine, B. W., & O’Doherty, J. P. (2010). Human and rodent
homologies in action control: Corticostriatal determinants of
goal-directed and habitual action. Neuropsychopharmacology,
35, 48–69. https://doi.org/10.1038/npp.2009.131, PubMed:
19776734
Bieleke, M., & Keller, L. (2021). Individual differences in if–then
planning: Insights from the development and application
of the if–then planning scale (ITPS). Personality and
Individual Differences, 170, 110500. https://doi.org/10.1016/j
.paid.2020.110500
Brandstätter, V., Lengfelder, A., & Gollwitzer, P. M. (2001).
Implementation intentions and efficient action initiation.
Journal of Personality and Social Psychology, 81, 946–960.
https://doi.org/10.1037/0022-3514.81.5.946, PubMed: 11708569
Chang, L. J., Yarkoni, T., Khaw, M. W., & Sanfey, A. G. (2013).
Decoding the role of the insula in human cognition:
Functional parcellation and large-scale reverse inference.
Cerebral Cortex, 23, 739–749. https://doi.org/10.1093/cercor
/bhs065, PubMed: 22437053
De Houwer, J. (2019). On how definitions of habits can
complicate habit research. Frontiers in Psychology, 10, 2642.
https://doi.org/10.3389/fpsyg.2019.02642, PubMed: 31849762
De Houwer, J., Tanaka, A., Moors, A., & Tibboel, H. (2018).
Kicking the habit: Why evidence for habits in humans might
be overestimated. Motivation Science, 4, 50–59. https://doi
.org/10.1037/mot0000065
Deichmann, R., Gottfried, J. A., Hutton, C., & Turner, R. (2003).
Optimized EPI for fMRI studies of the orbitofrontal cortex.
Neuroimage, 19, 430–441. https://doi.org/10.1016/S1053
-8119(03)00073-9, PubMed: 12814592
Delorme, C., Salvador, A., Valabrègue, R., Roze, E., Palminteri,
S., Vidailhet, M., et al. (2016). Enhanced habit formation in
Gilles de la Tourette syndrome. Brain, 139, 605–615. https://
doi.org/10.1093/brain/awv307, PubMed: 26490329
de Wit, S., Corlett, P. R., Aitken, M. R., Dickinson, A., & Fletcher,
P. C. (2009). Differential engagement of the ventromedial
prefrontal cortex by goal-directed and habitual behavior
toward food pictures in humans. Journal of Neuroscience,
29, 11330–11338. https://doi.org/10.1523/JNEUROSCI.1639
-09.2009, PubMed: 19741139
de Wit, S., Kindt, M., Knot, S. L., Verhoeven, A. A. C., Robbins,
T. W., Gasull-Camos, J., et al. (2018). Shifting the balance
between goals and habits: Five failures in experimental habit
induction. Journal of Experimental Psychology: General,
147, 1043–1065. https://doi.org/10.1037/xge0000402,
PubMed: 29975092
de Wit, S., Niry, D., Wariyar, R., Aitken, M. R. F., & Dickinson, A.
(2007). Stimulus-outcome interactions during instrumental
van Timmeren et al.
973
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
6
9
5
7
2
0
8
2
8
8
1
/
j
o
c
n
_
a
_
0
1
9
9
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
discrimination learning by rats and humans. Journal of
Experimental Psychology: Animal Behavior Processes, 33,
1–11. https://doi.org/10.1037/0097-7403.33.1.1, PubMed:
17227190
de Wit, S., Watson, P., Harsay, H. A., Cohen, M. X., ven de Vijver,
I., & Ridderinkhof, K. R. (2012). Corticostriatal connectivity
underlies individual differences in the balance between
habitual and goal-directed action control. Journal of
Neuroscience, 32, 12066–12075. https://doi.org/10.1523
/JNEUROSCI.1088-12.2012, PubMed: 22933790
Dickinson, A. (1985). Actions and habits: The development of
behavioural autonomy. Philosophical Transactions of the
Royal Society of London, Series B: Biological Sciences, 308,
67–78. https://doi.org/10.1098/rstb.1985.0010
Esteban, O., Birman, D., Schaer, M., Koyejo, O. O., Poldrack,
R. A., & Gorgolewski, K. J. (2017). MRIQC: Advancing the
automatic prediction of image quality in MRI from unseen
sites. PLoS One, 12, e0184661. https://doi.org/10.1371/journal
.pone.0184661, PubMed: 28945803
Esteban, O., Markiewicz, C. J., Blair, R. W., Moodie, C. A., Isik,
A. I., Erramuzpe, A., et al. (2019). fMRIPrep: A robust
preprocessing pipeline for functional MRI. Nature Methods,
16, 111–116. https://doi.org/10.1038/s41592-018-0235-4,
PubMed: 30532080
Gardner, B., Abraham, C., Lally, P., & de Bruijn, G.-J. (2012).
Towards parsimony in habit measurement: Testing the
convergent and predictive validity of an automaticity subscale
of the self-report habit index. International Journal of
Behavioral Nutrition and Physical Activity, 9, 102. https://
doi.org/10.1186/1479-5868-9-102, PubMed: 22935297
Gilbert, S. J., Gollwitzer, P. M., Cohen, A. L., Oettingen, G., &
Burgess, P. W. (2009). Separable brain systems supporting
cued versus self-initiated realization of delayed intentions.
Journal of Experimental Psychology: Learning, Memory,
and Cognition, 35, 905–915. https://doi.org/10.1037
/a0015535, PubMed: 19586260
Gollwitzer, P. M. (1993). Goal achievement: The role of
intentions. European Review of Social Psychology, 4,
141–185. https://doi.org/10.1080/14792779343000059
Gollwitzer, P. M. (1999). Implementation intentions: Strong
effects of simple plans. American Psychologist, 54, 493–503.
https://doi.org/10.1037/0003-066X.54.7.493
Gollwitzer, P. M. (2014). Weakness of the will: Is a quick fix
possible? Motivation and Emotion, 38, 305–322. https://doi
.org/10.1007/s11031-014-9416-3
Gollwitzer, P. M., Parks-Stamm, E. J., Jaudas, A., & Sheeran, P.
(2008). Flexible tenacity in goal pursuit. In J. Y. Shah &
W. L. Gardner (Eds.), Handbook of motivation science
(pp. 325–341). Guilford Press.
Gollwitzer, P. M., & Sheeran, P. (2006). Implementation
intentions and goal achievement: A meta-analysis of effects
and processes. Advances in Experimental Social Psychology,
38, 69–119. https://doi.org/10.1016/S0065-2601(06)38002-1
Gorgolewski, K., Burns, C. D., Madison, C., Clark, D., Halchenko,
Y. O., Waskom, M. L., et al. (2011). Nipype: A flexible,
lightweight and extensible neuroimaging data processing
framework in Python. Frontiers in Neuroinformatics, 5, 13.
https://doi.org/10.3389/fninf.2011.00013, PubMed: 21897815
Hardwick, R. M., Forrence, A. D., Krakauer, J. W., & Haith, A. M.
(2019). Time-dependent competition between goal-directed
and habitual response preparation. Nature Human Behaviour,
3, 1252–1262. https://doi.org/10.1038/s41562-019-0725-0,
PubMed: 31570762
Horga, G., Maia, T. V., Marsh, R., Hao, X., Xu, D., Duan, Y.,
et al. (2015). Changes in corticostriatal connectivity during
reinforcement learning in humans. Human Brain Mapping,
36, 793–803. https://doi.org/10.1002/hbm.22665, PubMed:
25393839
Jahanshahi, M., Obeso, I., Rothwell, J. C., & Obeso, J. A.
(2015). A fronto-striato-subthalamic-pallidal network for
goal-directed and habitual inhibition. Nature Reviews
Neuroscience, 16, 719–732. https://doi.org/10.1038/nrn4038,
PubMed: 26530468
JASP Team. (2018). JASP (version 0.8.6) [Computer Software].
Retrieved from https://jasp-stats.org/2018/02/28/now-jasp-0-8-6/.
Kruglanski, A. W., & Szumowska, E. (2020). Habitual behavior
is goal-driven. Perspectives on Psychological Science, 15,
1256–1271. https://doi.org/10.1177/1745691620917676,
PubMed: 32569529
Legrand, E., Bieleke, M., Gollwitzer, P. M., & Mignon, A. (2017).
Nothing will stop me? Flexibly tenacious goal striving with
implementation intentions. Motivation Science, 3, 101–118.
https://doi.org/10.1037/mot0000050
Liljeholm, M., Dunne, S., & O’Doherty, J. P. (2015).
Differentiating neural systems mediating the acquisition vs.
expression of goal-directed and habitual behavioral control.
European Journal of Neuroscience, 41, 1358–1371. https://
doi.org/10.1111/ejn.12897, PubMed: 25892332
Liljeholm, M., Tricomi, E., O’Doherty, J. P., & Balleine, B. W.
(2011). Neural correlates of instrumental contingency
learning: Differential effects of action-reward conjunction
and disjunction. Journal of Neuroscience, 31, 2474–2480.
https://doi.org/10.1523/JNEUROSCI.3354-10.2011, PubMed:
21325514
McNamee, D., Liljeholm, M., Zika, O., & O’Doherty, J. P. (2015).
Characterizing the associative content of brain structures
involved in habitual and goal-directed actions in humans:
A multivariate fMRI study. Journal of Neuroscience, 35,
3764–3771. https://doi.org/10.1523/JNEUROSCI.4677-14
.2015, PubMed: 25740507
Morris, R. W., Quail, S. L., Griffiths, K. R., Green, M. J., &
Balleine, B. W. (2015). Corticostriatal control of goal-directed
action is impaired in schizophrenia. Biological Psychiatry,
77, 187–195. https://doi.org/10.1016/j.biopsych.2014.06.005,
PubMed: 25062683
Orbell, S., & Verplanken, B. (2010). The automatic component
of habit in health behavior: Habit as cue-contingent
automaticity. Health Psychology, 29, 374–383. https://doi.org
/10.1037/a0019596, PubMed: 20658824
Parks-Stamm, E. J., Gollwitzer, P. M., & Oettingen, G. (2007).
Action control by implementation intentions: Effective
cue detection and efficient response initiation. Social
Cognition, 25, 248–266. https://doi.org/10.1521/soco.2007
.25.2.248
Pool, E. R., Gera, R., Fransen, A., Perez, O. D., Cremer, A.,
Aleksic, M., et al. (2022). Determining the effects of training
duration on the behavioral expression of habitual control
in humans: A multilaboratory investigation. Learning &
Memory, 29, 16–28. https://doi.org/10.1101/lm.053413.121,
PubMed: 34911800
Ramautar, J. R., Slagter, H. A., Kok, A., & Ridderinkhof, K. R.
(2006). Probability effects in the stop-signal paradigm:
The insula and the significance of failed inhibition. Brain
Research, 1105, 143–154. https://doi.org/10.1016/j.brainres
.2006.02.091, PubMed: 16616048
Seeley, W. W., Menon, V., Schatzberg, A. F., Keller, J., Glover, G. H.,
Kenna, H., et al. (2007). Dissociable intrinsic connectivity
networks for salience processing and executive control.
Journal of Neuroscience, 27, 2349–2356. https://doi.org/10
.1523/JNEUROSCI.5587-06.2007, PubMed: 17329432
Sheeran, P., & Webb, T. L. (2016). The intention–behavior gap.
Social and Personality Psychology Compass, 10, 503–518.
https://doi.org/10.1111/spc3.12265
Tanaka, S. C., Balleine, B. W., & O’Doherty, J. P. (2008).
Calculating consequences: Brain systems that encode the
causal effects of actions. Journal of Neuroscience, 28,
974
Journal of Cognitive Neuroscience
Volume 35, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
6
9
5
7
2
0
8
2
8
8
1
/
j
o
c
n
_
a
_
0
1
9
9
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
6750–6755. https://doi.org/10.1523/JNEUROSCI.1808-08
.2008, PubMed: 18579749
Thorndike, E. L. (1911). Animal intelligence: Experimental
studies. The Macmillan Company. https://doi.org/10.5962/bhl
.title.55072
Tricomi, E., Balleine, B. W., & O’Doherty, J. P. (2009). A specific
role for posterior dorsolateral striatum in human habit
learning. European Journal of Neuroscience, 29, 2225–2232.
https://doi.org/10.1111/j.1460-9568.2009.06796.x, PubMed:
19490086
Tzourio-Mazoyer, N., Landeau, B., Papathanassiou, D., Crivello,
F., Etard, O., Delcroix, N., et al. (2002). Automated
anatomical labeling of activations in SPM using a macroscopic
anatomical parcellation of the MNI MRI single-subject brain.
Neuroimage, 15, 273–289. https://doi.org/10.1006/nimg.2001
.0978, PubMed: 11771995
Uddin, L. Q. (2015). Salience processing and insular cortical
function and dysfunction. Nature Reviews Neuroscience, 16,
55–61. https://doi.org/10.1038/nrn3857, PubMed: 25406711
Uddin, L. Q., Nomi, J. S., Hébert-Seropian, B., Ghaziri, J., &
Boucher, O. (2017). Structure and function of the human
insula. Journal of Clinical Neurophysiology, 34, 300–306.
https://doi.org/10.1097/ WNP.0000000000000377, PubMed:
28644199
Valentin, V. V., Dickinson, A., & O’Doherty, J. P. (2007).
Determining the neural substrates of goal-directed learning
in the human brain. Journal of Neuroscience, 27, 4019–4026.
https://doi.org/10.1523/JNEUROSCI.0564-07.2007, PubMed:
17428979
van Timmeren, T., & de Wit, S. (2022). Instant habits versus
flexible tenacity: Do implementation intentions accelerate
habit formation? Quarterly Journal of Experimental
Psychology. https://doi.org/10.1177/17470218221147024,
PubMed: 36476147
Watson, P., & de Wit, S. (2018). Current limits of experimental
research into habits and future directions. Current Opinion
in Behavioral Sciences, 20, 33–39. https://doi.org/10.1016/j
.cobeha.2017.09.012
Watson, P., Gladwin, T. E., Verhoeven, A. A. C., & de Wit, S.
(2022). Investigating habits in humans with a symmetrical
outcome-revaluation task. Behavior Research Methods.
https://doi.org/10.3758/s13428-022-01922-4, PubMed:
35867208
Watson, P., O’Callaghan, C., Perkes, I., Bradfield, L., & Turner, K.
(2022). Making habits measurable beyond what they are not:
A focus on associative dual-process models. Neuroscience &
Biobehavioral Reviews, 142, 104869. https://doi.org/10.1016/j
.neubiorev.2022.104869, PubMed: 36108980
Watson, P., van Wingen, G., & de Wit, S. (2018). Conflicted
between goal-directed and habitual control, an fMRI
investigation. eNeuro, 5, ENEURO.0240-18.2018. https://doi
.org/10.1523/ENEURO.0240-18.2018, PubMed: 30310863
Wetzels, R., Matzke, D., Lee, M. D., Rouder, J. N., Iverson,
G. J., & Wagenmakers, E.-J. (2011). Statistical evidence in
experimental psychology: An empirical comparison using 855
t tests. Perspectives on Psychological Science, 6, 291–298.
https://doi.org/10.1177/1745691611406923, PubMed:
26168519
Wunderlich, K., Dayan, P., & Dolan, R. J. (2012). Mapping value
based planning and extensively trained choice in the human
brain. Nature Neuroscience, 15, 786–791. https://doi.org/10
.1038/nn.3068, PubMed: 22406551
Yin, H. H., Knowlton, B. J., & Balleine, B. W. (2004). Lesions
of dorsolateral striatum preserve outcome expectancy but
disrupt habit formation in instrumental learning. European
Journal of Neuroscience, 19, 181–189. https://doi.org/10
.1111/j.1460-9568.2004.03095.x, PubMed: 14750976
Zwosta, K., Ruge, H., Goschke, T., & Wolfensteller, U. (2018).
Habit strength is predicted by activity dynamics in
goal-directed brain systems during training. Neuroimage,
165, 125–137. https://doi.org/10.1016/j.neuroimage.2017.09
.062, PubMed: 28970144
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
6
9
5
7
2
0
8
2
8
8
1
/
j
o
c
n
_
a
_
0
1
9
9
0
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
van Timmeren et al.
975