Attention and Temporal Expectations Modulate Power,
Not Phase, of Ongoing Alpha Oscillations
Rosanne M. van Diepen1, Michael X Cohen2, Damiaan Denys1,3, and Ali Mazaheri4
Abstract
■ The perception of near-threshold visual stimuli has been
shown to depend in part on the phase (i.e., time in the cycle)
of ongoing alpha (8–13 Hz) oscillations in the visual cortex
relative to the onset of that stimulus. However, it is currently
unknown whether the phase of the ongoing alpha activity can
be manipulated by top–down factors such as attention or
expectancy. Using three variants of a cross-modal attention par-
adigm with constant predictable stimulus onsets, we examined
if cues signaling to attend to either the visual or the auditory
domain influenced the phase of alpha oscillations in the associ-
ated sensory cortices. Importantly, intermixed in all three exper-
iments, we included trials without a target to estimate the phase
at target presentation without contamination from the early
evoked responses. For these blank trials, at the time of expected
target and distractor onset, we examined (1) the degree of the
uniformity in phase angles across trials, (2) differences in phase
angle uniformity compared with a pretarget baseline, and (3)
phase angle differences between visual and auditory target con-
ditions. Across all three experiments, we found that, although
the cues induced a modulation in alpha power in occipital
electrodes, neither the visual condition nor the auditory cue con-
dition induced any significant phase-locking across trials during
expected target or distractor presentation. These results suggest
that, although alpha power can be modulated by top–down
factors such as attention and expectation, the phase of the
ongoing alpha oscillation is not under such control. ■
INTRODUCTION
The modulation of the power of oscillatory activity in the
alpha range (8–13 Hz) has been proposed to play a pivotal
mechanistic role in attention by gating information flow to
relevant sensory regions through the inhibition of irrele-
vant regions (Jensen & Mazaheri, 2010; Klimesch, Sauseng,
& Hanslmayr, 2007; Fu et al., 2001). Supporting this
hypothesis are a number of studies reporting that the
power of alpha activity is suppressed in task-relevant
regions but increased in regions processing unattended
information (Mazaheri, van Schouwenburg, et al., 2014;
Bauer, Kennett, & Driver, 2012; Bauer, Kluge, et al., 2012;
Haegens, Luther, & Jensen, 2012; Haegens, Handel, &
Jensen, 2011; Kerlin, Shahin, & Miller, 2011; Hanslmayr
et al., 2007; Jokisch & Jensen, 2007; Medendorp et al.,
2007; Rihs, Michel, & Thut, 2007; Sauseng et al., 2005; Thut
et al., 2003). Although the direct mechanism by which alpha
activity exerts functional inhibition is currently unknown,
some recent studies have suggested (Mazaheri & Jensen,
2010) and shown that a power increase has a strong inhib-
itory influence on both spike timing and firing rate of neural
activity (Haegens, Nacher, Luna, Romo, & Jensen, 2011).
In addition to alpha power (i.e., the amplitude of the
oscillation), alpha phase (i.e., the timing of the oscillation)
at stimulus onset also influences perception (Mathewson
1Academic Medical Center Amsterdam, 2University of Amsterdam,
3Netherlands Institute for Neuroscience, 4University of Birmingham
© 2015 Massachusetts Institute of Technology
et al., 2011, 2012; Mathewson, Fabiani, Gratton, Beck,
& Lleras, 2010; Busch, Dubois, & VanRullen, 2009;
Mathewson, Gratton, Fabiani, Beck, & Ro, 2009) as well
as the evoked response (Scheeringa, Mazaheri, Bojak,
Norris, & Kleinschmidt, 2011), suggesting that the alpha
cycle reflects rhythmically fluctuating states of low and high
excitability (Thut, Miniussi, & Gross, 2012; Mathewson
et al., 2011; Klimesch et al., 2007). In other words, whereas
the amplitude of an alpha oscillation reflects the degree
of excitation or inhibition, the phase angle of the alpha
oscillation indexes the excitability or inhibitory state of
the underlying neural generators within a cycle of the alpha
oscillation. This has led to the intriguing possibility that
a top–down attentional modulation of the phase of alpha
activity could therefore also serve as a mechanism for
selection of relevant information on a fast time scale
(Bonnefond & Jensen, 2012; Jensen, Bonnefond, &
VanRullen, 2012; Mathewson et al., 2011).
However, to date, there has not been a clear demon-
stration that the phase of the alpha oscillations can be
modulated by top–down task demands while accounting
for the disturbances occurring because of stimulus
evoked responses. The objective of this study was to
investigate whether the anticipation of targets and dis-
tractors modulated the phase of alpha oscillations via
top–down control, without the contamination of sensory
evoked responses that would add a waveform with a spe-
cific phase to the ongoing alpha activity and subsequently
Journal of Cognitive Neuroscience 27:8, pp. 1573–1586
doi:10.1162/jocn_a_00803
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
7
/
8
2
7
1
/
5
8
7
/
3
1
1
5
9
7
4
3
9
/
8
1
7
7
1
8
o
3
c
4
n
5
_
9
a
/
_
j
0
o
0
c
8
n
0
3
_
a
p
_
d
0
0
b
8
y
0
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
f
t
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
confounding its phase estimation. Therefore, the phase
of the ongoing alpha oscillation was calculated on less
frequent, unexpected trials containing no targets (“blank”
trials) to measure anticipatory processes not contaminated
by sensory evoked responses. These trials were incorpo-
rated into three versions of a cross-modal attention task
in which top–down modulation of the phase of alpha
oscillation would be beneficial for task performance. Brief
targets and distractors were presented at regular times,
such that their timing was predictable. In the first two
versions of the paradigm, a target and distractor in the
opposing modality (visual and auditory) were presented
together (bimodal experiments). A preceding cue served
to signal which modality was the target and which the
distractor. In the first experiment, the visual stimulus was
always presented 37 msec before the auditory stimulus,
whereas in the second task, the target and distractor were
presented simultaneously. Our rationale here was that the
arrival of a distracting stimulus half cycle before a target
might induce a different degree of phase adjustment than
when the arrival of distractor and target were simultaneous.
In the third “unimodal” version, only targets were pre-
sented, either in the visual or auditory domain. Again,
cues were presented to help prepare the relevant sensory
area for the upcoming to-be-discriminated target. Because
no distractors were presented, only anticipatory processes
related to target processing would be examined in this
version, without the contamination of signals related to
inhibition of the distractors.
On the basis of the studies described above, we hypoth-
esized that the modulation of phase would be more precise
on a temporal scale and thus more effective than modula-
tion of power to obtain the optimal state of excitability
or inhibition. To anticipate our results, examining the
degree of intertrial phase-locking as well as phase angle
differences between the visual and auditory conditions,
we did not find evidence for a cue-induced phase per-
turbation of alpha activity at the time of expected target/
distractor onset across all three experiments.
METHODS
Participants
In total, 30 participants were recruited at the University of
Amsterdam and Amsterdam University of Applied Sciences
to participate in the experiments. Eighteen (16 women,
age range = 18–28 years) participated in Experiments 1
and 3, and 12 (8 women, age range = 19–37 years,
one left-handed) participated in Experiment 2. All par-
ticipants signed informed consent documents before the
start of the experiment. Participants reported normal or
corrected-to-normal vision, no hearing disabilities, and no
history of psychiatric or neurological disorders. Participants
were compensated with A10 per hour. This study was
approved by the University of Amsterdam’s Department
of Psychology Ethics Committee.
Apparatus and Procedure
We conducted three variants of a cross-modal attention
paradigm previously used in Mazaheri, van Schouwenburg,
et al. (2014). All three experiments were run in a dimly lit
room. The participants were seated 60 cm from the mon-
itor. Before the start of the experiment, a practice session
took place. During the practice session, the percentage of
correct responses was shown after every 10 trials so that
participants and experimenters were aware of performance
levels. The practice session ended after 90 trials or when
participants performed above 70% correct. The stimuli
were presented using Presentation software (Neurobehav-
ioral Systems, Inc., Albany, CA) on a 24-in. LCD monitor
(BenQ XL2420T, Taipei, Taiwan) with a refresh rate of
120 Hz. All three experiments contained 420 trials with a
mandatory rest period for every 42 trials.
The visual stimuli, presented centrally for 25 msec
(three screen refreshes), consisted of Gabor patches
having a visual angle of ∼3.8°, a contrast of 50%, and three
possible types of orientation: −5°, 0°, and 5° clockwise. The
auditory stimuli, presented for 25 msec via speakers, were
pure tones with three possible frequencies: 500, 1000,
and 2000 Hz (including 5-msec rise and 5-msec fall
shaped by a Blackman window).
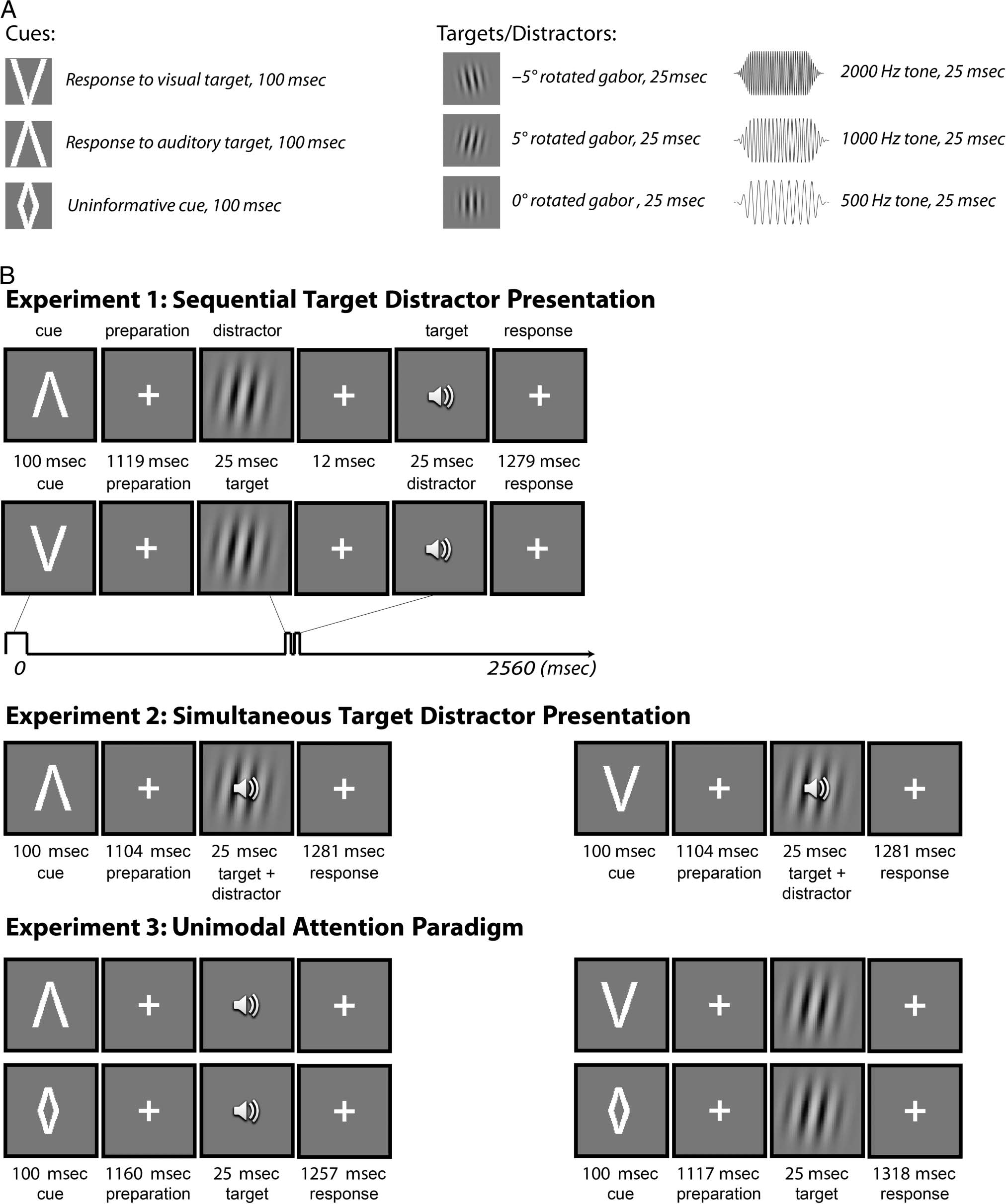
Experiment 1: Sequential Target
Distractor Presentation
In Experiment 1, the visual and auditory stimuli were pre-
sented 37 msec apart. A visual cue instructed the partici-
pants to what modality they had to perform the target
discrimination. A “V” cue (100 msec) instructed the partic-
ipants to perform spatial judgment on the upcoming visual
stimulus while ignoring the auditory stimulus. In contrast,
an “inverted V” cue (representing an “A,” 100 msec) in-
structed the participants to make a pitch judgment on
the upcoming auditory stimulus, while ignoring the visual
stimulus. The participants responded with their right hand
using a three-button mouse. A sample trial sequence is
conceptualized in Figure 1B. Using a light sensor and
sensor custom made by the EEG manufacturer (ANT,
Enschede, The Netherlands), we were able to evaluate
the synchronization of the visual and auditory stimuli as
well the jitter (represented as σ) in times of onset. The
standard deviation in onset times of the stimuli was esti-
mated by looking at the distribution time of 100 stimulus
onsets. The visual stimuli were presented 1219 msec after
cue onset (σ = 5.12 msec), and the auditory stimuli were
presented 1256 msec after cue onset (σ = 3.32 msec). Trial
duration was 2560 msec. The number of visual and audi-
tory cues was equal. All target orientations and tone fre-
quencies were presented an equal number of times.
Importantly, 84 of the 420 trials (20%) did not contain a
target and distractor (blank trials). These trials were pseu-
dorandomly intermixed with all the other trials such that
no blank trials ever occurred in succession.
1574
Journal of Cognitive Neuroscience
Volume 27, Number 8
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
7
/
8
2
7
1
/
5
8
7
/
3
1
1
5
9
7
4
3
9
/
8
1
7
7
1
8
o
3
c
4
n
5
_
9
a
/
_
j
0
o
0
c
8
n
0
3
_
a
p
_
d
0
0
b
8
y
0
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
/
t
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
7
/
8
2
7
1
/
5
8
7
/
3
1
1
5
9
7
4
3
9
/
8
1
7
7
1
8
o
3
c
4
n
5
_
9
a
/
_
j
0
o
0
c
8
n
0
3
_
a
p
_
d
0
0
b
8
y
0
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
t
f
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 1. Task figures and example sequences of Experiments 1–3 with stimulus presentation times. A trial started with the presentation of a
symbolic cue indicating whether a target in the visual or auditory domain needed to be identified. A “V” cue indicated to identify the rotation
of a grating. An “inverted V” cue (representing an “A”) indicated to identify the pitch of a tone. The symbolic cue was followed by a constant
cue–target interval in which participants were able to prepare for the upcoming target. A target was always presented in the modality indicated
by the cue. In Experiments 1 and 3, a distractor in the opposing modality to the target was presented. Participants used their right hand to
press a button to choose between three different tones and Gabor gratings.
van Diepen et al.
1575
Experiment 2: Simultaneous Target
Distractor Presentation
Experiment 2 was identical to Experiment 1 except that
visual and auditory targets were presented simultaneously
at 1204 msec after cue onset (visual target: σ = 0.88 msec,
auditory target: σ = 3.76 msec). The trial duration of this
experiment was 2510 msec.
1.25, and 1.28 components per participant for Experi-
ments 1, 2, and 3, respectively). A PCA was used to reduce
dimensionality of the data before performing the indepen-
dent component analysis. Trials with an RT < 100 msec
were considered incorrect and excluded from all data
analyses. Participants with exceptional high alpha power
values (>3 SDs from mean) were removed from analyses
(one participant in Experiment 2).
Experiment 3: Unimodal Attention Paradigm
The purpose of Experiment 3 was twofold. First, to rule
out the possibility that electrical fields from the visual
and auditory cortices canceled out at the scalp, no distrac-
tors were presented together with the targets. Second, to
manipulate attention without the need to ignore a dis-
tractor, in 60 trials (no blank trials), the informative cue
was replaced with an uninformative “diamond” cue (⋄).
This attention manipulation was possible only in Ex-
periment 3 because, although the cue was not necessary
to perform the task correctly, we reasoned that having
a cue would facilitate top–down expectation and thus
could affect the ongoing phases. The visual cue was
presented 1217 msec (σ = 4.18 msec) after cue onset,
and the auditory target was presented 1260 msec
(σ = 3.07 msec) after cue onset. Trial duration was
2560 msec.
EEG Acquisition and Processing
EEG data were acquired using a WaveGuard 10-5 cap
system developed by ANT, with 64 Ag–AgCl electrodes,
spanning from frontal, temporal, central, and occipital
scalp sites. The EEG was sampled at 1024 Hz with an
online average reference and then subsequently imported
into MATLAB (Natick, MA) for all further offline analyses.
The EOG was recorded between supraorbital and infraor-
bital sites around the right eye for vertical movement
(blinks) and outer canthi of the left and right eyes for pos-
sible horizontal eye movements. Impedance was main-
tained below 100 kΩ. Offline processing and analyses
were performed using MATLAB, functions from EEGLAB
version 13.1.1b (Delorme & Makeig, 2004), the Fieldtrip
software package (Oostenveld, Fries, Maris, & Schoffelen,
2011), and the Circular Statistics Toolbox (Berens, 2009).
The EEG data were high-pass filtered at 0.5 Hz and
epoched from −1 to +2.5 sec (−1 to +2.45 for Experi-
ment 3), time-locked to cue onset. Trials with substantial
muscle artifacts were identified and removed through
visual inspection (12.7%, 12.0%, and 15.0% for Experi-
ments 1, 2, and 3, respectively). Bad electrodes were
identified and interpolated using the spherical spline inter-
polation routing in EEGLAB (two participants with two
electrodes, two participants with one channel). Ocular
artifacts were removed using independent component
analysis (infomax algorithm) incorporated as the default
“runica” function in EEGLAB 13.1.1b (on average, 1.28,
Time–Frequency Analysis
Time–frequency representations of power and phase were
estimated per trial, using sliding Hanning tapers having an
adaptive time window of three cycles for each frequency
of interest (ΔT = 3/f ). Similar approaches were used by
Mazaheri, Nieuwenhuis, van Dijk, and Jensen (2009),
Jokisch and Jensen (2007), and Osipova et al. (2006).
Unless stated otherwise, analyses were performed on
every participant at the peak frequency in the alpha band
(average across participants: Experiment 1, 11.1 Hz, σ =
1.8; Experiment 2, 10.7 Hz, σ = 2.1; Experiment 3, 10.9,
σ = 1.8), which was determined using the 500-msec pre-
target interval from all trials.
Phase-locking Factor
To assess whether a preferred phase angle was present
for target detection or distractor inhibition, we examined
the variability (i.e., clustering or locking) of the phase
angle across trials at the time of the expected target arrival.
This was done using a measure called the phase-locking
factor (PLF), which provides a quantification of intertrial
phase-locking (Tallon-Baudry, Bertrand, Delpuech, &
Pernier, 1996). The PLF was derived by first unit normaliz-
ing the magnitude of the phase angles (obtained in the
time–frequency analysis outlined earlier), then averaging
(in the complex domain) across the trials, and getting
the absolute value of the average. A PLF value close to 0
reflects high variability of phase angles across trials,
whereas a PLF value of 1 reflects all trials having the same
phase angle. PLFs were calculated for blank trials at the
time of visual target/distractor onset. The magnitude of
the PLF is dependent on the number of phase observa-
tions; thus, an equal number of trials was randomly select-
ed from the condition containing a greater number of
trials such that the number was equal for the visual and
auditory target conditions.
Intuitively, the PLF might appear a less appropriate mea-
sure for our research question than, for example, a differ-
ence in phase between the visual and auditory conditions.
However, the phase angle of an EEG signal is (at least for
tangential dipoles) arbitrary in sign and can vary in its
physiological meaning (excitation/inhibition) between
participants. PLF is a suitable measure because it expresses
whether a phase angle is more prevalent and thus favor-
able, without retaining information about the actual phase,
1576
Journal of Cognitive Neuroscience
Volume 27, Number 8
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
7
/
8
2
7
1
/
5
8
7
/
3
1
1
5
9
7
4
3
9
/
8
1
7
7
1
8
o
3
c
4
n
5
_
9
a
/
_
j
0
o
0
c
8
n
0
3
_
a
p
_
d
0
0
b
8
y
0
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
.
t
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
and can therefore be averaged over participants with
differently positioned dipoles.
be close to 0. When only one condition exhibits intertrial
phase-locking, the PBI will take a negative value.
Statistical Significance of Intertrial Phase-locking
at the Time of Target Presentation
The PLF was estimated exclusively for blank trials as to
eliminate contamination by the early sensory responses
evoked by the target/distractor onset. Given that the PLF
is dependent on the number of observations and the
amount of the blank trials surviving artifact rejection varied
across participants, we determined the PLF and its signifi-
cance for each participant separately. We did this using
a bootstrapping procedure similar to Jensen, Hari, and
Kaila (2002). Accordingly, we generated a random set of
n phases, with n being equal to the participant’s number
of blank trials per condition, repeating this procedure
1,000 times and calculating the PLF value for each set. A
significant PLF in the alpha band (at each participant’s peak
frequency) was defined as being at or above the 95th
percentile rank of the randomly generated PLF distribu-
tion. The average number of blank trials across participants
was 35.6 (σ = 4.2), 34.3 (σ = 4.1), and 30.6 (σ = 3.8; same
for the auditory and visual target conditions), resulting in
average cutoff values of 0.291 (σ = 0.019), 0.296 (σ =
0.023), and 0.312 (σ = 0.025) for Experiments 1, 2, and
3, respectively.
Differences in the Degree of Intertrial
Phase-locking between Time of Target
Presentation and Baseline
Across participants, we statistically assessed the difference
between the PLFs (in each channel) at baseline and at the
time of expected target onset using a two-tailed t test. The
baseline was derived from the combined auditory and
visual conditions 200 msec before cue onset.
Phase Angle Differences between Visual
and Auditory Target Conditions
Previous work suggests having different phase distribu-
tions (i.e., preferred phase angle) between conditions
without differences in PLF values. Thus, we also exam-
ined the (average) phase bifurcation index (PBI; Busch
et al., 2009), which quantifies intertrial phase angle dif-
ferences between conditions. The following formula was
used:
(cid:1)
(cid:3)
(cid:1)
PLFVisual − PLFVisualþauditory
(cid:2) PLFauditory − PLFvisualþauditory
(cid:3)
When phases are phase-locked toward a different angle for
the visual and auditory target conditions, the bifurcation
index will take a positive value. When a random phase
distribution is present for both conditions, the index will
The significance level of PBI values was obtained using
a bootstrapping procedure consisting of the following
two steps: First, for each participant, trials from the visual
and auditory cue conditions were pooled. Two random
samples were drawn from the pool, and a “pseudo” PBI
was calculated. This procedure was repeated 500 times
per participant to create a random distribution of values
that fall under the null hypothesis. Second, for each
participant, a pseudo PBI value was randomly selected
from this distribution, after which the average over par-
ticipants was calculated. This was repeated 100,000 times.
For each channel, the level of significance was deter-
mined as the proportion of average pseudo PBI values
exceeding the average observed PBI.
Oscillatory Power Analysis
In addition to our phase analyses, we also examined the
effect of the attentional cues on the power of the ongoing
alpha activity. To ensure our results in the current study
were consistent with those of previous literature show-
ing attention-modulated suppression of alpha power
(Mazaheri, Fassbender, et al., 2014; Mazaheri, van
Schouwenburg, et al., 2014; Haegens et al., 2012; Haegens,
Handel, et al., 2011; Kerlin et al., 2011; Hanslmayr et al.,
2007; Jokisch & Jensen, 2007; Medendorp et al., 2007; Rihs
et al., 2007; Sauseng et al., 2005; Thut et al., 2003), here, we
did a correction for multiple comparisons. Cue-induced
power differences for the alpha peak frequency were
examined for every time point between cue and target
presentation using a nonparametric randomization test
(as described in Maris & Oostenveld, 2007) using alpha
power from every participant’s peak frequency. Monte
Carlo p values were calculated on 1,000 random partitions
in which the visual and auditory labels of power estimates
were shuffled. Type 1 errors because of multiple compari-
sons are reduced by clustering neighboring electrodes that
show a similar effect. The cluster-level test statistics ( p =
.05, two sided) reported in the Results section are calcu-
lated by averaging t values for electrodes in a given cluster.
For this analysis, the following number of trials were avail-
able for the auditory condition and visual condition, respec-
tively: 122.0556 (σ = 23.2416), 104.9091 (σ = 21.5845), and
115.5000 (σ = 14.4924) and 138.1111 (σ = 17.1323),
130.6364 (σ = 20.9775), and 124.7222 (σ = 14.4010).
RESULTS
Behavioral Data
In Experiment 1, participants were better at discrimination
of visual targets than auditory targets (96% vs. 88%, t(17) =
7.20, p = .000001). This was also reflected in RTs, which
were faster for visual discrimination than auditory discrim-
ination (630 vs. 728 msec, t(17) = −4.26, p = .0004). In
van Diepen et al.
1577
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
7
/
8
2
7
1
/
5
8
7
/
3
1
1
5
9
7
4
3
9
/
8
1
7
7
1
8
o
3
c
4
n
5
_
9
a
/
_
j
0
o
0
c
8
n
0
3
_
a
p
_
d
0
0
b
8
y
0
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
.
/
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Experiment 2, similar to the first experiment, the percent-
age of correct response was higher for visual than auditory
targets (96% vs. 81%, t(10) = 89.36, p = .000003). In line
with that RTs were faster for trials with a visual target than
for trials with an auditory target (639 vs. 748 msec, t(10) =
9.36, p = .000002). Also in Experiment 3, the unimodal
experiment, visual targets were identified faster (606 vs.
697 msec, t(17) = −7.12, p = .000002) and more often
(98% vs. 95%, t(17) = 3.24, p = .005) compared with
auditory cues.
Because of the absence of distractors in Experiment 3,
the attentional cues were not essential to perform the
task correctly. However, participants nevertheless demon-
strated significant benefits from the cuing: RTs for auditory
targets decreased from 748 msec for uninformative cues to
697 msec for informative cues (t(17) = −5.10, p = .00008).
For visual targets, the average RT decreased from 642 msec
for uninformative cues to 606 msec for informative cues
(t(17) = −5.91, p = .00002). These behavioral results
demonstrate that the attentional cues in the experiment
were indeed used by the participants, suggesting that
top–down processes facilitated target processing.
To test whether the distraction in the Experiment 1
influenced RTs (i.e., there was a distraction cost), we
compared RTs from Experiment 1 (bimodal) with Exper-
iment 3 (unimodal). For auditory targets, presentation of
a distractor slowed RTs by an average of 31 msec (t(17) =
−3.77, p = .001). For visual targets, presentation of a
distractor slowed RTs by 24 msec (t(17) = −3.29, p =
.004). These results suggest that the presence of the
distractor was detrimental to performing the task. A
mechanism for active inhibition of the distractor could
be beneficial during this task.
The attentional cues did not have any influence on
the participant’s accuracy in the task, which we specu-
lated to be because of a ceiling effect in performance.
Auditory targets were identified in 93% of the trials with
an uninformative cue and in 95% with an informative cue
(t(17) = −1.48, p = .16). Visual targets were identified in
98% of the trials with both informative and uninformative
cues (t(17) = −0.15, p = .917).
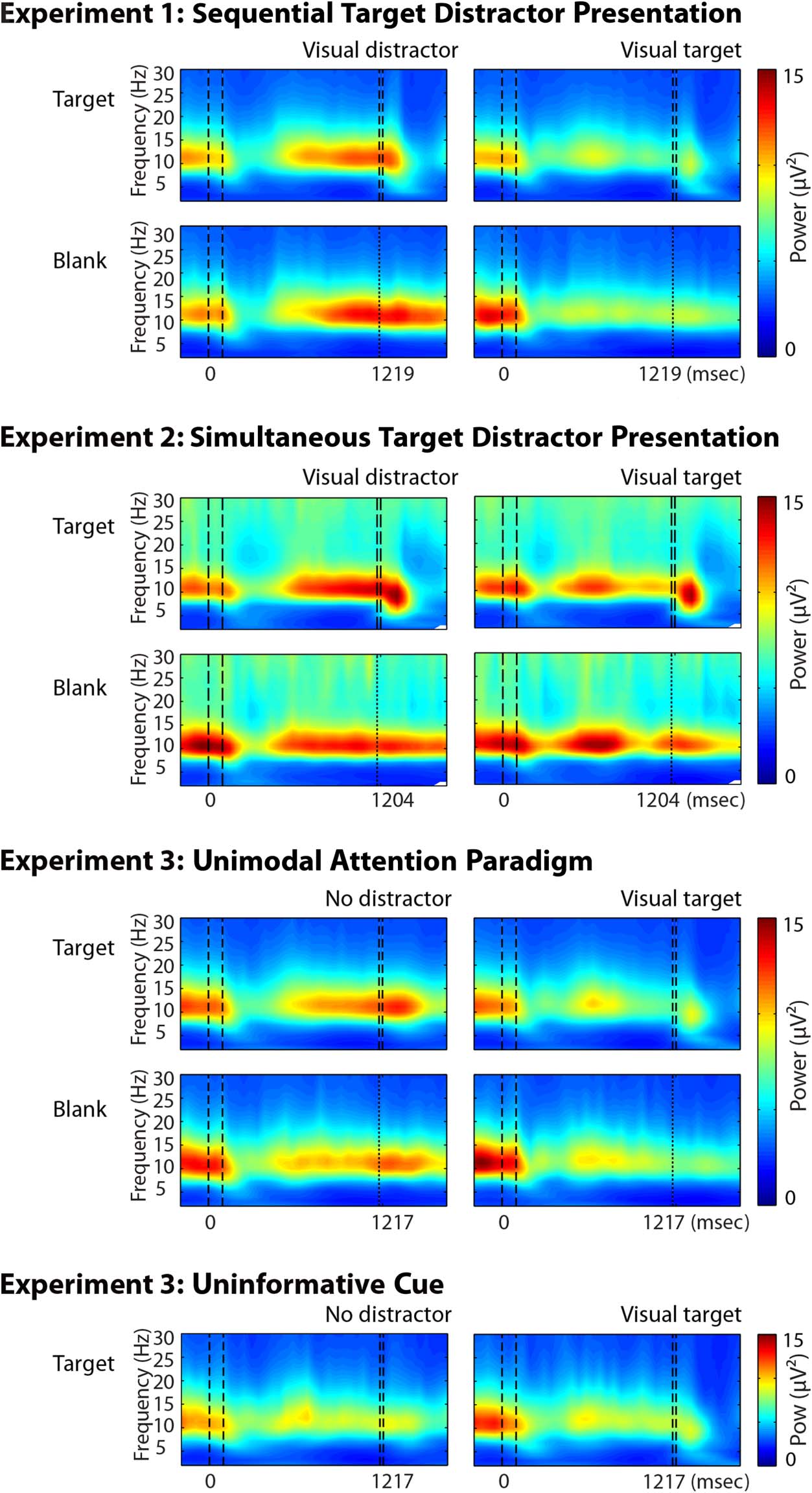
EEG Results
The onset of visual stimuli (i.e., cues and targets) evoked a
transient increase in power (Figure 2) in the theta band
and an increase in PLF in the theta and alpha bands (Fig-
ure 3) in electrodes located over the occipital cortex,
consistent with results shown in previous literature (e.g.,
Mazaheri & Jensen, 2006; Makeig et al., 2002). Moreover,
before target presentation, alpha power was highest in
occipital electrodes. Given that alpha activity from the
auditory cortex has been found difficult to measure using
scalp EEG, because of the anatomical location of the audi-
tory cortex (Mazaheri, van Schouwenburg, et al., 2014;
Bastiaansen & Brunia, 2001), we focused in part of our
subsequent analyses on the average values of electrodes
located over the occipital cortex (Oz, O1, and O2).
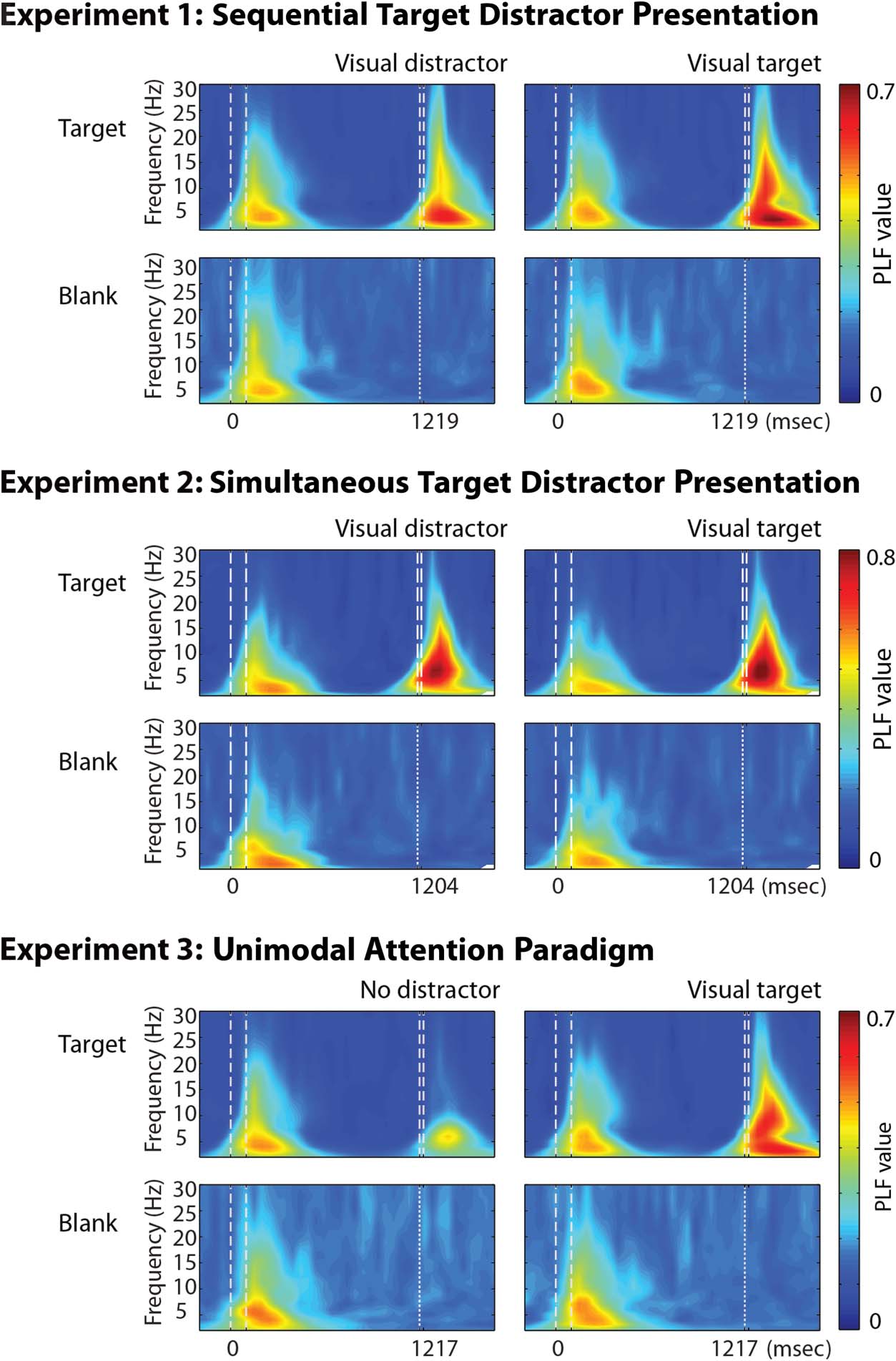
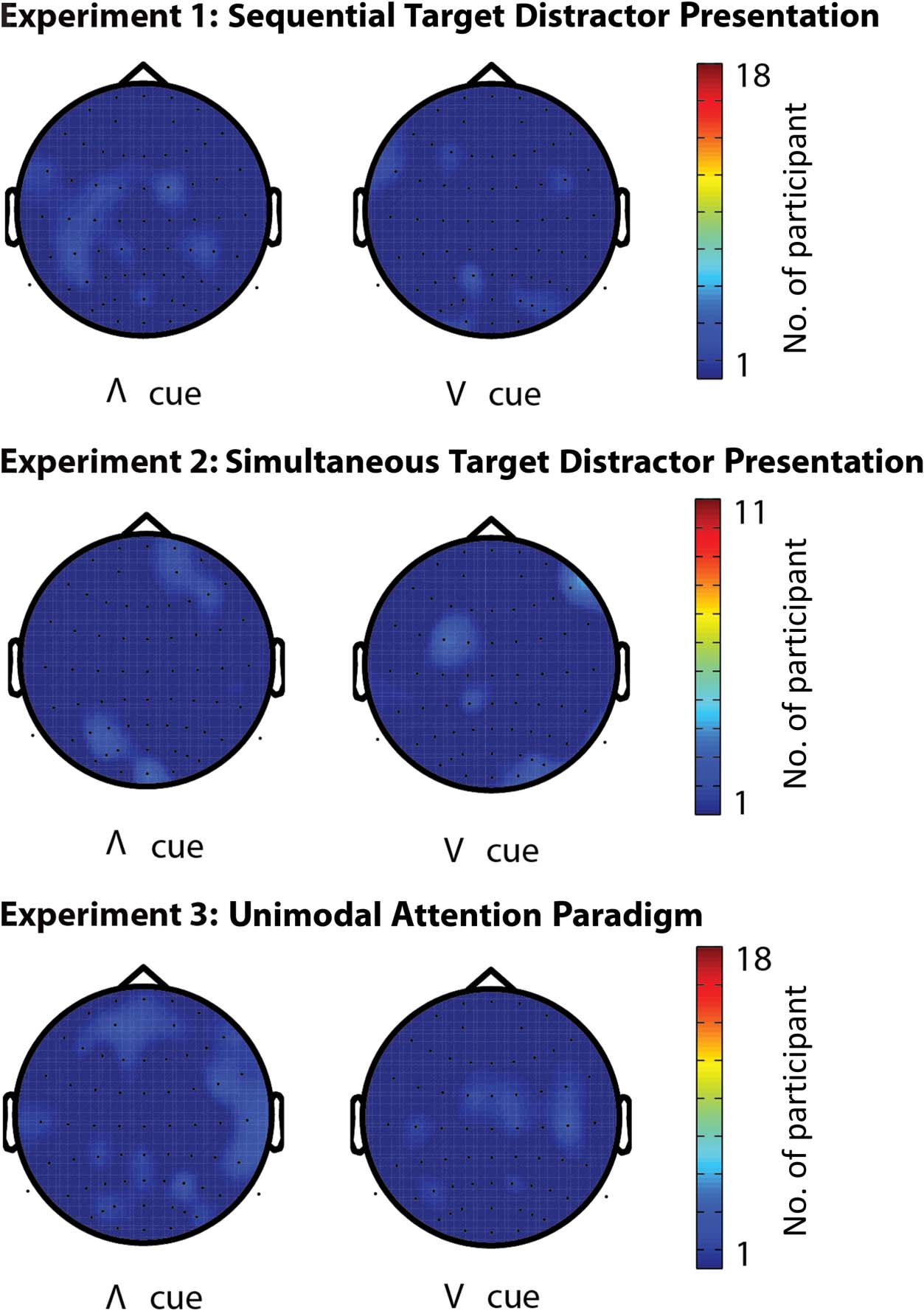
Absence of Significant PLF at Expected Arrival of Targets
We found an absence of robust PLF at the time of expected
target presentation in most participants across all three
experiments. Participants showed an average of 2.1 (σ =
4.3), 3.4 (σ = 4.9), and 2.8 (σ = 5.6) electrodes with a sig-
nificant PLF during the visual condition and 2.8333 (σ =
3.0147), 3.9167 (σ = 6.5569), and 3.7222 (σ = 5.1542)
electrodes during the auditory condition. However, these
electrodes were not neighboring one another, suggesting
that the phase-locking was spurious. The absence of a
consistent pattern (Figure 4), even with the liberal statis-
tical threshold employed, suggests that the attentional
cues did not induce a significant degree of phase-locking
at the time of expected target onset.
Intertrial Phase-locking at Target Onset Was Not
Significantly Different from Baseline
We assessed if the PLF at the time of target onset was sig-
nificantly greater than baseline values across participants.
We found that PLF values at the time of expected target
arrival were not increased in any channel in Experiments 1
(sequential target distractor presentation) and 3 (unimodal
stimulus presentation). In Experiment 2 (simultaneous tar-
get distractor presentation), one channel showed an in-
crease in PLF in the parietal cortex when a visual target
was expected, and two electrodes showed an increase in
the right temporal cortex (not adjacent) when an auditory
target was expected. However, as can be seen from the
topographic maps of the t values, there was no clear pattern
of increases and decreases of PLF at the time of expected
target onset compared with the baseline period (Figure 5).
No Phase Angle Differences between Visual and
Auditory Target Conditions
We investigated if there were any phase angle differences
between visual target and auditory target (visual distractor)
conditions using the PBI. Figure 6 shows a topographic
representation of average PBI values over participants, in-
cluding marked electrodes that showed a significant PBI
from 0 calculated using a resampling procedure. We were
unable to find any clusters of electrodes showing a significant
clear phase difference present between visual and auditory
target trials at the time of expected target presentation.
The Attentional Cue-induced Modality-specific
Modulation of Alpha Power in Anticipation of
Target Presentation
In Experiment 1, alpha power over occipital/parietal areas
was greater before the presentation of the auditory target
1578
Journal of Cognitive Neuroscience
Volume 27, Number 8
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
7
/
8
2
7
1
/
5
8
7
/
3
1
1
5
9
7
4
3
9
/
8
1
7
7
1
8
o
3
c
4
n
5
_
9
a
/
_
j
0
o
0
c
8
n
0
3
_
a
p
_
d
0
0
b
8
y
0
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
f
/
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 2. Grand mean time–
frequency representations of
power per condition for trials
with target (top rows) and blank
(bottom rows) trials. Values
are averaged across electrodes
located over the occipital cortex
(electrodes Oz, O1, and O2)
and not baseline corrected. The
dark blue color indicates power
of 0 μV2. The first two dotted
vertical lines represent the time
interval of cue presentation.
The second set of vertical
dotted lines represents the
presentation interval of targets.
The expected time of target
onset for blank trials is indicated
with a single dotted line. (Left)
Trials containing an auditory
target (i.e., visual distractor in
Experiments 1 and 3), showing
an increase in alpha power
before and during presentation
of a visual distractor. (Right)
Trials with a visual target,
showing a decrease in alpha
power during preparation
and target presentation.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
7
/
8
2
7
1
/
5
8
7
/
3
1
1
5
9
7
4
3
9
/
8
1
7
7
1
8
o
3
c
4
n
5
_
9
a
/
_
j
0
o
0
c
8
n
0
3
_
a
p
_
d
0
0
b
8
y
0
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
t
.
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
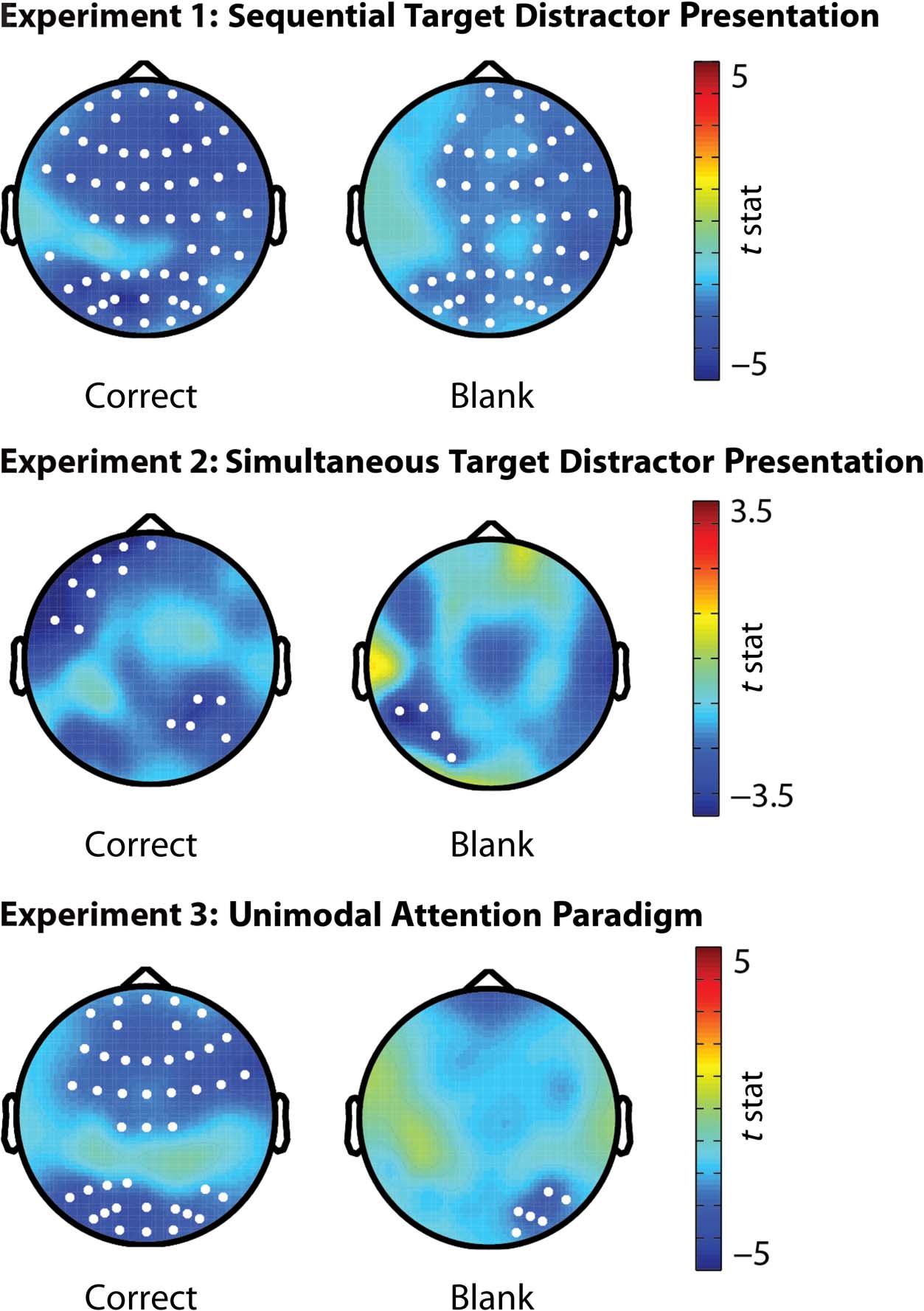
compared with the presentation of a visual target. This
difference started in electrodes located over the parietal
cortex and spread toward occipital and frontal electrodes.
For the trials containing a target, the difference was pres-
ent 550 msec after cue onset until target presentation
(t = −182.42, p = .0001, Monte Carlo p value, corrected
for multiple comparisons). For blank trials, the difference
was present from 600 msec after cue onset until target
presentation (t = −123.74, p = .003, Monte Carlo p value,
corrected for multiple comparisons; Figure 7).
The same pattern of results was observed in Experi-
ment 2 where, at 700 msec postcue until target presenta-
tion, electrodes over the occipital cortex had greater
alpha power before an expected auditory target than
expected visual target (t = −11.59, p = .049, Monte Carlo
p value, corrected for multiple comparisons).
van Diepen et al.
1579
These results are consistent with previous studies indicat-
ing that cues signaling the preparation for visual discrimina-
tion (relative to pitch discrimination) resulted in a decrease
of alpha power (9–11 Hz) in the early visual cortex (Mazaheri,
van Schouwenburg, et al., 2014; Mazaheri et al., 2010).
Finally, the informative cues in Experiment 3 also mod-
ulated alpha activity even when a target was presented
without a distractor. Alpha power was higher when an
auditory target had to be identified compared with when
a visual target had to be identified. For correct trials, the
difference started in frontal electrodes at 650 msec after
cue onset (t = −68.86, p = .006, Monte Carlo p value,
corrected for multiple comparisons) and 750 msec after
cue onset also in electrodes located over the occipital
cortex (t = −56.04, p = .008, Monte Carlo p value, cor-
rected for multiple comparisons). In blank trials, the dif-
ferences originated 950 msec after cue presentation
in left occipital electrodes and spread toward frontal
electrodes (t = −16.75, p = .028, Monte Carlo p value,
corrected for multiple comparisons).
Taken together, all three experiments showed modu-
lated pretarget alpha power depending on informative
cue presentation, such that power was higher before
identification of a visual target compared with an auditory
target. This difference was most pronounced in electrodes
located over parietal cortex.
Modality-specific Alpha Modulation Restricted to
Only Attentional Cues
We found that, in trials with an uninformative cue in
Experiment 3, no difference in alpha power was present
Figure 3. Grand mean time–
frequency representations of
PLF per condition for trials with
a target (top rows) and blank
(bottom rows) trials. Values are
averaged across electrodes
located over the occipital cortex
(electrodes Oz, O1, and O2).
(Top) Trials in which a target
was shown. (Bottom) Trials in
which a target was expected
but not shown. An increase in
PLF occurs after presentation
of a stimulus (cue or target);
however, there was no
(significant) PLF observed
during the preparation for an
upcoming target or at the time
of expected target presentation.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
7
/
8
2
7
1
/
5
8
7
/
3
1
1
5
9
7
4
3
9
/
8
1
7
7
1
8
o
3
c
4
n
5
_
9
a
/
_
j
0
o
0
c
8
n
0
3
_
a
p
_
d
0
0
b
8
y
0
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
/
.
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
1580
Journal of Cognitive Neuroscience
Volume 27, Number 8
Figure 4. Topographic maps
of PLF at the time of expected
target presentation. Color
scaling specifies the number
of participants that show
significant PLF, per channel.
Left images refer to PLF over
trials with an “inverted V” cue,
indicating to respond to an
auditory target. Right images
refer to significant PLF over
trials with a “V” cue, indicating
to respond to a visual target.
The maximum number of
participants showing significant
PLF values in the same channel
was 3.
before presentation of the targets (when a separate anal-
ysis was done on every time point between cue and tar-
get presentation as well as when activity was averaged
over 450-msec pretarget interval [interval based on time
interval in which the difference was found for informative
cues]). These results suggest that the modulation of the
power of the alpha activity was because of top–down at-
tentional factors, rather than simply the physical onset of
the cues.
DISCUSSION
In the current study, we investigated whether the phase
of an alpha oscillation could be modulated by top–down
control. We utilized a series of attentional cuing para-
digms with temporally predictable targets and distractors
that also contained “blank” trials with no targets/distrac-
tors. These blank trials afforded us the opportunity to in-
vestigate the phase property of the alpha oscillations at
the time of expected target or distractor arrival without
contamination from sensory evoked responses. We
found that the distribution of the phase angles across tri-
als at the time of expected target and distractor arrival
was uniform and did not significantly differ from a precue
baseline period. These results argue against top–down
modulation of phase as a (general) mechanism for selec-
tion of relevant information.
Our results appear inconsistent with recent sugges-
tions that the prioritization of information in a stimulus
stream occurs through the top–down modulation of al-
pha phase (Bonnefond & Jensen, 2012; Jensen et al.,
2012). Although we find this idea quite intriguing, we ar-
gue that the estimates used to support evidence for
phase modulation were not fully disentangled from the
sensory evoked responses of the targets and distractors.
Another explanation for the discrepancy could be that
van Diepen et al.
1581
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
7
/
8
2
7
1
/
5
8
7
/
3
1
1
5
9
7
4
3
9
/
8
1
7
7
1
8
o
3
c
4
n
5
_
9
a
/
_
j
0
o
0
c
8
n
0
3
_
a
p
_
d
0
0
b
8
y
0
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
/
f
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
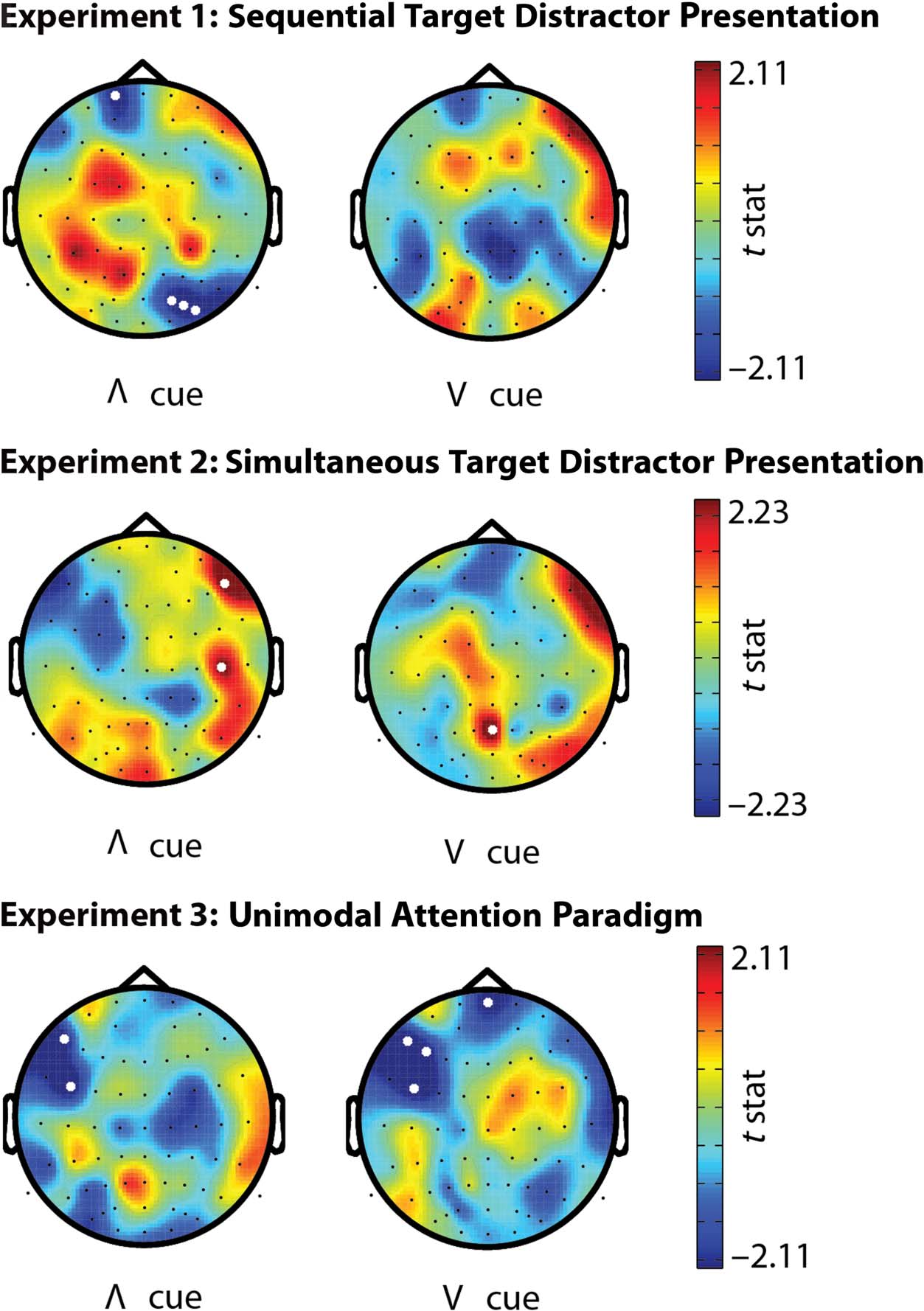
Figure 5. Scalp topographic
maps of PLF difference (t value)
of t test between baseline
and time of expected target/
distractor presentation. The
t values are positive (red
color) when PLF values are
higher at the expected target
presentation than baseline.
The range of the color bar
was chosen such that the
upper values correspond to
the critical significant values
of the t distribution. Left images
refer to PLF over trials with an
“inverted V” cue, indicating to
respond to an auditory target.
Right images refer to significant
PLF over trials with a “V” cue,
indicating to respond to a visual
target. White dots indicate
electrodes with a t value
that exceeded the statistical
threshold of p < .05 (no
correction for multiple
comparisons). No clusters
(number of neighboring
electrodes > 1) were found
in which PLF significantly
increased compared with
baseline.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
7
/
8
2
7
1
/
5
8
7
/
3
1
1
5
9
7
4
3
9
/
8
1
7
7
1
8
o
3
c
4
n
5
_
9
a
/
_
j
0
o
0
c
8
n
0
3
_
a
p
_
d
0
0
b
8
y
0
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
f
/
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
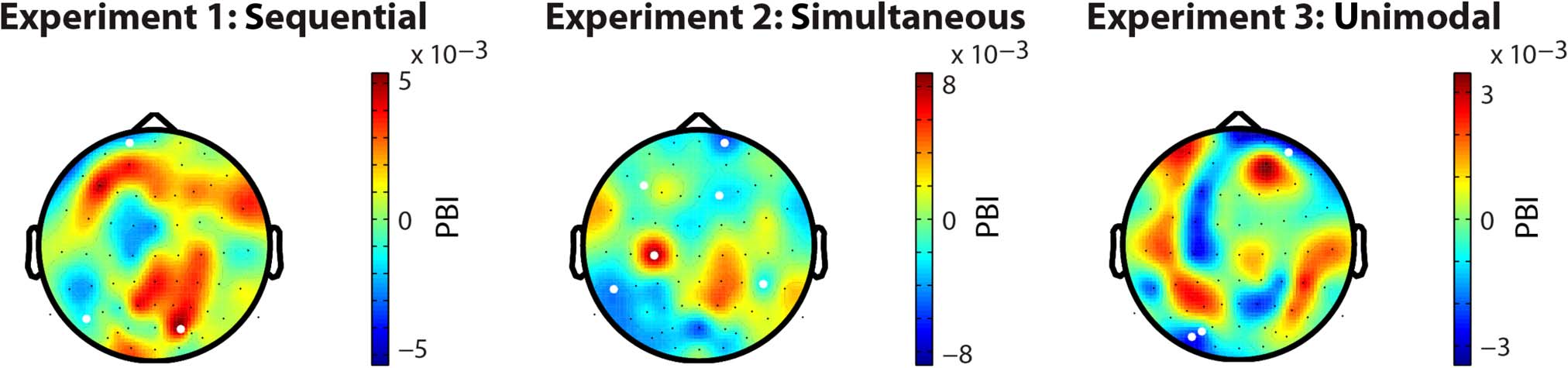
Figure 6. Topographic representations of average PBI over participants. White dots mark electrodes with a significant PBI assessed using a
bootstrapping procedure. A significant positive PBI is observed when the visual and auditory target conditions show a difference in preffered phase
angle. A negative PBI is found when a difference in PLF is present between conditions. Because the overall PLF is based on a larger amount of
trials than the auditory and visual conditions, the PBI is biased toward positive values, and an absence of any effect (in PLF or phase angle) is
represented as a value just above zero. None of the three experiments revealed a cluster of significant PBI values, indicating that no significant
difference in phase angle or PLF is present in preparation for a visual or an auditory target.
1582
Journal of Cognitive Neuroscience
Volume 27, Number 8
Figure 7. Topographic
representation of alpha power
in relation to the cues across
the three experiments. The
electrodes showing a significant
decrease in alpha activity in
preparation for a target
compared with an auditory
target are marked with
white dots. Across all three
experiments, we found that
cues signalling an upcoming
visual target induced a pretarget
suppression in alpha power
over occipital electrodes.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
7
/
8
2
7
1
/
5
8
7
/
3
1
1
5
9
7
4
3
9
/
8
1
7
7
1
8
o
3
c
4
n
5
_
9
a
/
_
j
0
o
0
c
8
n
0
3
_
a
p
_
d
0
0
b
8
y
0
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
/
f
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
the study finding the strongest support for top–down
control of alpha-phase modulation utilized a working
memory paradigm rather than a cross-modal attention
task (Bonnefond & Jensen, 2012). It could be that the
suppression of distractors during maintenance in work-
ing memory is mediated through different mechanisms
than suppression of a distracting modality stream. In
addition, the cadence of stimulus presentation of their
working memory task might have been different, possibly
helping the intrinsic time estimation and thus the antici-
patory phase estimate. Finally, measurements for current
experiment are done with EEG, whereas Bonnefond and
Jensen (2012) used MEG for the working memory task.
Because EEG is more prone to blurring of signals from
several sources, phase estimates in the current paradigm
could have been distorted to a larger extent.
In contrast to the PLF results, we did find cue-related
differences in alpha power in occipital electrodes. The
power of alpha oscillations was larger after presentation
of an auditory cue compared with a visual cue, consistent
with previous visual-audio cross-modal attention studies
(Mazaheri, van Schouwenburg et al., 2014). In the exper-
iments performed in this study, the presentation of a visual
cue was meant to facilitate the processing of upcoming
stimulus, whereas presentation of the auditory cue indi-
cates that the upcoming visual stimulus needs to be ig-
nored. The absence of a pretarget difference in alpha
power after presentation of uninformative cues, together
with the slower responses in this condition, indicates that
participants used top–down modulation of alpha power as
a mechanism to prepare for an upcoming target. A number
of previous studies have found that a top–down increase in
the power alpha oscillations in task-irrelevant regions
serves as a mechanism to actively ignore distracting infor-
mation (Bauer, Kennett, et al., 2012; Bauer, Kluge, et al.,
2012; Haegens et al., 2012; Haegens, Handel, et al., 2011;
van Diepen et al.
1583
Jokisch & Jensen, 2007; Medendorp et al., 2007; Rihs et al.,
2007; Sauseng et al., 2005; Thut et al., 2003) through its
suppressive influence on spike timing and firing rate of
neural activity (Haegens, Nacher, et al., 2011; Mazaheri &
Jensen, 2010).
The absence of phase adjustment during processing
of the visual target is in line with previous research show-
ing no influence of phase on detection rates during low
alpha power (Mathewson et al., 2009, 2011). A decrease
in alpha power can induce constant vigilance for detec-
tion of (unpredictable) events. The relative decrease in
alpha in the occipital cortex after presentation of a visual
cue is sufficient to reach a state of high excitability. On
the other hand, given that inhibitory influence of high
power alpha activity is found to be pulsed instead of con-
tinuous (Mathewson et al., 2010, 2011, 2012), a phase
modulation would allow for a temporally more precise
inhibition than only a power modulation. Interestingly,
Mathewson et al. (2011) have proposed that the rhyth-
micity between excitation and inhibition phases of alpha
activity could be beneficial because it would allow for
unattended (yet valuable) information from the external
environment to filter through analogous to an “…anti-
lock brake (ABS) system of a car, in which some level of
contacts with the road surface (in our case, the external
environment) is maintained by applying pulses of braking
rather than by braking continuously.”
A handful of studies have found evidence for phase
modulation in tasks using rhythmic stimulation, includ-
ing behavioral effects. For instance, phase entrainment
of low-frequency oscillations to temporally regular pre-
sented stimulation can enhance contrast sensitivity (Cravo,
Rohenkohl, Wyart, & Nobre, 2013). Rhythmic visual stim-
ulation in the alpha range entrains activity and, at the same
time, entrains visual awareness (Mathewson et al., 2010).
Entrainment is also found when temporally expected
events need to be inhibited. Besle and colleagues (2011)
found electrodes for which low-frequency oscillations
were entrained to stimulus presentation but were phase-
shifted for an unattended stream of stimuli compared
with an attended stream. Because no trial-by-trial control
or shifts of attention are required during these tasks, it
is possible that this phenomenon is not realized by top–
down attentional control but rather bottom–up processes.
In the current experiments, we explicitly searched for
phase adjustment in the absence of entrainment. This
was realized by the use of tasks with predictable targets
in which attentional shifting was needed between trials
instead of blocks. The absence of a phase modulation
during these tasks argues (with more confidence) against
top–down phase modulation as mechanism for inhibition
or attention.
We believe that top–down modulation of the phase of
alpha oscillations is not necessary. It could be beneficial
for (single) temporally predictable events that need to be
inhibited. Apart from rhythmic stimulation, such tem-
porally predictable events are uncommon in the natural en-
vironment. Common predictable events, such as speech, can
be processed or ignored more efficiently using entrain-
ment (Calderone, Lakatos, Butler, & Castellanos, 2014;
Henry & Obleser, 2012). We suggest that top–down
modulation of phase of alpha oscillations is an unnecessary
mechanism for selection of information when only a
temporally brief stimulus is presented.
Caveats
Several task parameters could explain the absence of
alpha phase modulation in our series of experiments,
leaving open the possibility of phase modulation in other
situations. First, a phase modulation is especially effec-
tive for near-threshold stimuli (Mathewson et al., 2011;
Womelsdorf, Fries, Mitra, & Desimone, 2006). Targets pre-
sented in our design were detectable far above threshold,
such that the percentage of correct responses was fairly
high. The advantage of high task performance is that a
higher percentage of blank trials contain correct anti-
cipatory processes instead of mistakes. However, we can-
not completely discount the possibility that easiness of
the task made that phase adjustment was not essential to
perform the task correctly. On the other hand, we believe
that it is rather surprising if there were two mechanisms
for distraction suppression, one for when target detection
is easy and one for when it is difficult.
Another limitation of our task could be the length of
the ISI between cue and target. A certain amount of time
may be needed to adjust a phase, which would argue
for a long cue–target interval. However, interval timing
becomes less precise when the interval becomes longer
(Buhusi & Meck, 2005), making the estimation of target
arrival not precise enough to adjust the phase properly.
The time interval between cue and target should be varied
to find out whether an ISI exists in which both conditions
are met.
Furthermore, we did not localize the anticipatory
increase in alpha oscillations before presentation of a
visual distractor, which means that the designation of the
occipital cortex as source of the inhibitory activity is only
speculative.
Conclusion
In summary, in our tasks, attentional cues led to both
behavioral and neural changes. RTs to targets were
faster when informative cues were presented. The neural
changes involved a modulation of the power of ongoing
oscillatory activity in the alpha band. However, no sign of
a trial-by-trial phase modulation in the alpha band was
found. A modulation of phase rather than power of an
ongoing oscillation was expected because it might be
a more instantaneous and more efficient mechanism
(Pilovsky, Rosenblum, & Kurths, 2001). We presented
several task parameters that could explain the absence of
a phase modulation. Before the start of the experiment,
1584
Journal of Cognitive Neuroscience
Volume 27, Number 8
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
7
/
8
2
7
1
/
5
8
7
/
3
1
1
5
9
7
4
3
9
/
8
1
7
7
1
8
o
3
c
4
n
5
_
9
a
/
_
j
0
o
0
c
8
n
0
3
_
a
p
_
d
0
0
b
8
y
0
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
f
.
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
we hypothesized that a predictable target presentation
would be the only requirement for a phase modulation
to occur. The increase in number of conditions that might
need to be met before a phase modulation can take place
makes it less likely that such a modulation is biologically
relevant. Our conclusion is, therefore, that the phase of
the alpha oscillation is not likely under top–down control.
Acknowledgments
Ali Mazaheri was supported by a Veni grant from The Nether-
lands Organization for Scientific Research (NWO).
Reprint requests should be sent to Rosanne Van Diepen, De-
partment of Psychiatry, Academic Medical Center Amsterdam,
Meibergdreef 5, Amsterdam, Netherlands 1105 AZ, or via e-mail:
rosannevandiepen@gmail.com or Ali Mazaheri, School of
Psychology, University of Birmingham, 3.03 Hills Building, West
Midlands, Birmingham, United Kingdom B15 2TT, or via e-mail:
ali.mazah@gmail.com.
REFERENCES
Bastiaansen, M. C., & Brunia, C. H. (2001). Anticipatory
attention: An event-related desynchronization approach.
International Journal of Psychophysiology, 43, 91–107.
Bauer, M., Kennett, S., & Driver, J. (2012). Attentional selection
of location and modality in vision and touch modulates
low-frequency activity in associated sensory cortices.
Journal of Neurophysiology, 107, 2342–2351.
Bauer, M., Kluge, C., Bach, D., Bradbury, D., Heinze, H. J.,
Dolan, R. J., et al. (2012). Cholinergic enhancement of
visual attention and neural oscillations in the human brain.
Current Biology, 22, 397–402.
Berens, P. (2009). CircStat: A Matlab toolbox for circular
statistics. Journal of Statistical Software, 31, 1–21.
Besle, J., Schevon, C. A., Mehta, A. D., Lakatos, P., Goodman,
R. R., McKhann, G. M., et al. (2011). Tuning of the human
neocortex to the temporal dynamics of attended events.
Journal of Neuroscience, 31, 3176–3185.
Bonnefond, M., & Jensen, O. (2012). Alpha oscillations serve to
protect working memory maintenance against anticipated
distractors. Current Biology, 22, 1969–1974.
Buhusi, C. V., & Meck, W. H. (2005). What makes us tick?
Functional and neural mechanisms of interval timing.
Nature Reviews Neuroscience, 6, 755–765.
Busch, N. A., Dubois, J., & VanRullen, R. (2009). The phase
of ongoing EEG oscillations predicts visual perception.
Journal of Neuroscience, 29, 7869–7876.
Calderone, D. J., Lakatos, P., Butler, P. D., & Castellanos, F. X.
(2014). Entrainment of neural oscillations as a modifiable
substrate of attention. Trends in Cognitive Sciences, 18,
300–309.
Cravo, A. M., Rohenkohl, G., Wyart, V., & Nobre, A. C. (2013).
Temporal expectation enhances contrast sensitivity by
phase entrainment of low-frequency oscillations in visual
cortex. Journal of Neuroscience, 33, 4002–4010.
Delorme, A., & Makeig, S. (2004). EEGLAB: An open source
toolbox for analysis of single-trial EEG dynamics including
independent component analysis. Journal of Neuroscience
Methods, 134, 9–21.
Fu, K. M. G., Foxe, J. J., Murray, M. M., Higgins, B. A., Javitt,
D. C., & Schroeder, C. E. (2001). Attention-dependent
suppression of distractor visual input can be cross-modally
cued as indexed by anticipatory parieto-occipital alpha-band
oscillations. Cognitive Brain Research, 12, 145–152.
Haegens, S., Handel, B. F., & Jensen, O. (2011). Top–down
controlled alpha band activity in somatosensory areas
determines behavioral performance in a discrimination
task. Journal of Neuroscience, 31, 5197–5204.
Haegens, S., Luther, L., & Jensen, O. (2012). Somatosensory
anticipatory alpha activity increases to suppress distracting
input. Journal of Cognitive Neuroscience, 24, 677–685.
Haegens, S., Nacher, V., Luna, R., Romo, R., & Jensen, O.
(2011). Alpha-oscillations in the monkey sensorimotor
network influence discrimination performance by rhythmical
inhibition of neuronal spiking. Proceedings of the National
Academy of Sciences, U.S.A., 108, 19377–19382.
Hanslmayr, S., Aslan, A., Staudigl, T., Klimesch, W., Herrmann,
C. S., & Bauml, K. H. (2007). Prestimulus oscillations predict
visual perception performance between and within
participants. NeuroImage, 37, 1465–1473.
Henry, M. J., & Obleser, J. (2012). Frequency modulation
entrains slow neural oscillations and optimizes human
listening behavior. Proceedings of the National Academy
of Sciences, U.S.A., 109, 20095–20100.
Jensen, O., Bonnefond, M., & VanRullen, R. (2012). An
oscillatory mechanism for prioritizing salient unattended
stimuli. Trends in Cognitive Sciences, 16, 200–206.
Jensen, O., Hari, R., & Kaila, K. (2002). Visually evoked
gamma responses in the human brain are enhanced during
voluntary hyperventilation. NeuroImage, 15, 575–586.
Jensen, O., & Mazaheri, A. (2010). Shaping functional
architecture by oscillatory alpha activity: Gating by inhibition.
Frontiers in Human Neuroscience, 4, 186.
Jokisch, D., & Jensen, O. (2007). Modulation of gamma and
alpha activity during a working memory task engaging the
dorsal or ventral stream. Journal of Neuroscience, 27,
3244–3251.
Kerlin, J. R., Shahin, A. J., & Miller, L. M. (2011). Attentional
gain control of ongoing cortical speech representations in
a cocktail party. Journal of Neuroscience, 30, 620–628.
Klimesch, W., Sauseng, P., & Hanslmayr, S. (2007). EEG
alpha oscillations: The inhibition-timing hypothesis.
Brain Research Reviews, 53, 63–88.
Makeig, S., Westerfield, M., Jung, T. P., Enghoff, S., Townsend,
J., Courchesne, E., et al. (2002). Dynamic brain sources of
visual evoked responses. Science, 295, 690–694.
Maris, E., & Oostenveld, R. (2007). Nonparametric statistical
testing of EEG- and MEG-data. Journal of Neuroscience
Methods, 164, 177–190.
Mathewson, K. E., Fabiani, M., Gratton, G., Beck, D. M., &
Lleras, A. (2010). Rescuing stimuli from invisibility: Inducing
a momentary release from visual masking with pre-target
entrainment. Cognition, 115, 186–191.
Mathewson, K. E., Gratton, G., Fabiani, M., Beck, D. M., &
Ro, T. (2009). To see or not to see: Prestimulus alpha phase
predicts visual awareness. Journal of Neuroscience, 29,
2725–2732.
Mathewson, K. E., Lleras, A., Beck, D. M., Fabiani, M., Ro, T.,
& Gratton, G. (2011). Pulsed out of awareness: EEG alpha
oscillations represent a pulsed-inhibition of ongoing cortical
processing. Frontiers in Psychology, 2, 99.
Mathewson, K. E., Prudhomme, C., Fabiani, M., Beck, D. M.,
Lleras, A., & Gratton, G. (2012). Making waves in the stream
of consciousness: Entraining oscillations in EEG alpha
and fluctuations in visual awareness with rhythmic visual
stimulation. Journal of Cognitive Neuroscience, 24,
2321–2333.
Mazaheri, A., Coffery-Corina, S., Mangun, G. R., Bekker, E. M.,
Berry, A., & Corbett, B. A. (2010). Functional disconnection
of frontal cortex and visual cortex in attention deficit
hyperactivity disorder. Biological Psychiatry, 67,
617–623.
van Diepen et al.
1585
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
7
/
8
2
7
1
/
5
8
7
/
3
1
1
5
9
7
4
3
9
/
8
1
7
7
1
8
o
3
c
4
n
5
_
9
a
/
_
j
0
o
0
c
8
n
0
3
_
a
p
_
d
0
0
b
8
y
0
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
t
.
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Mazaheri, A., Fassbender, C., Coffey-Corina, S., Hartanto,
T. A., Schweitzer, J. B., & Mangun, G. R. (2014).
Differential oscillatory electroencephalogram between
attention-deficit/hyperactivity disorder subtypes and
typically developing adolescents. Biological Psychiatry,
76, 422–429.
Mazaheri, A., & Jensen, O. (2006). Posterior alpha activity is not
phase-reset by visual stimuli. Proceedings of the National
Academy of Sciences, U.S.A., 103, 2948–2952.
Mazaheri, A., & Jensen, O. (2010). Rhythmic pulsing: Linking
ongoing brain activity with evoked responses. Frontiers
in Human Neuroscience, 4, 177.
Mazaheri, A., Nieuwenhuis, I. L. C., van Dijk, H., & Jensen, O.
(2009). Prestimulus alpha and mu activity predicts failure
to inhibit motor responses. Human Brain Mapping, 30,
1791–1800.
Mazaheri, A., van Schouwenburg, M. R., Dimitrijevic, A.,
Denys, D., Cools, R., & Jensen, O. (2014). Region-specific
modulations in oscillatory alpha activity serve to facilitate
processing in the visual and auditory modalities.
NeuroImage, 87, 356–362.
Medendorp, W. P., Kramer, G. F. I., Jensen, O., Oosterveld, R.,
Schoffelen, J. M., & Fries, P. (2007). Oscillatory activity in
human parietal and occipital cortex shows hemispheric
lateralization and memory effects in a delayed double-step
saccade task. Cerebral Cortex, 17, 2364–2374.
Oostenveld, R., Fries, P., Maris, E., & Schoffelen, J. M. (2011).
FieldTrip: Open source software for advanced analysis
of MEG, EEG, and invasive electrophysiological data.
Computational Intelligence and Neuroscience, 2011,
156869.
Osipova, D., Takashima, A.,Oostenveld, R., Fernandez, G.,
Maris, E., & Jensen, O. (2006). Theta and gamma
oscillations predict encoding and retrieval of declarative
memory. Journal of Neuroscience, 26, 7523–7531.
Pilovsky, A. S., Rosenblum, M. G., & Kurths, J. (2001).
Synchronization: A universal concept in nonlinear
sciences. Cambridge: Cambridge University Press.
Rihs, T. A., Michel, C. M., & Thut, G. (2007). Mechanisms of
selective inhibition in visual spatial attention are indexed
by alpha-band EEG synchronization. European Journal
of Neuroscience, 25, 603–610.
Sauseng, P., Klimesch, W., Stadler, W., Schabus, M.,
Doppelmayr, M., Hanslmayr, S., et al. (2005). A shift of
visual spatial attention is selectively associated with human
EEG alpha activity. European Journal of Neuroscience,
22, 2917–2926.
Scheeringa, R., Mazaheri, A., Bojak, I., Norris, D. G., &
Kleinschmidt, A. (2011). Modulation of visually evoked
cortical fMRI responses by phase of ongoing occipital
alpha oscillations. Journal of Neuroscience, 31, 3813–3820.
Tallon-Baudry, C., Bertrand, O., Delpuech, C., & Pernier, J.
(1996). Stimulus specificity of phase-locked and
non-phase-locked 40 Hz visual responses in human.
Journal of Neuroscience, 16, 4240–4249.
Thut, G., Miniussi, C., & Gross, J. (2012). The functional
importance of rhythmic activity in the brain. Current
Biology, 22, R658–R663.
Thut, G., Theoret, H., Pfennig, A., Ives, J., Kampmann, F.,
Northoff, G., et al. (2003). Differential effects of
low-frequency rTMS at the occipital pole on visual-induced
alpha de synchronization and visual-evoked potentials.
NeuroImage, 18, 334–347.
Womelsdorf, T., Fries, P., Mitra, P. P., & Desimone, R. (2006).
Gamma-band synchronization in visual cortex predicts
speed of change detection. Nature, 439, 733–736.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
7
/
8
2
7
1
/
5
8
7
/
3
1
1
5
9
7
4
3
9
/
8
1
7
7
1
8
o
3
c
4
n
5
_
9
a
/
_
j
0
o
0
c
8
n
0
3
_
a
p
_
d
0
0
b
8
y
0
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
/
f
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
1586
Journal of Cognitive Neuroscience
Volume 27, Number 8