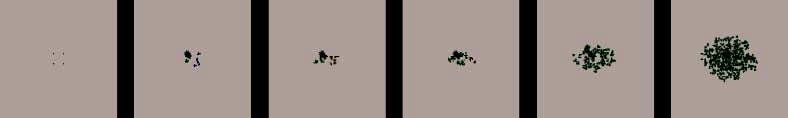An Effective Hierarchical Model
for the Biomolecular Covalent
Bond: An Approach Integrating
Artificial Chemistry and an
Actual Terrestrial Life System
Abstract Under the AChem paradigm and the programmed
self-decomposition ( PSD) model, we propose a hierarchical
model for the biomolecular covalent bond (HBCB model).
This model assumes that terrestrial organisms arrange their
biomolecules in a hierarchical structure according to the energy
strength of their covalent bonds. It also assumes that they have
evolutionarily selected the PSD mechanism of turning biological
polymers (BPs) into biological monomers (BMs) as an efficient
biomolecular recycling strategy. We have examined the validity
and effectiveness of the HBCB model by coordinating two
complementary approaches: biological experiments using existent
terrestrial life, and simulation experiments using an AChem
system. Biological experiments have shown that terrestrial life
possesses a PSD mechanism as an endergonic, genetically regulated
process and that hydrolysis, which decomposes a BP into BMs,
is one of the main processes of such a mechanism. In simulation
experiments, we compared different virtual self-decomposition
processes. The virtual species in which the self-decomposition
process mainly involved covalent bond cleavage from a BP to
BMs showed evolutionary superiority over other species in which
the self-decomposition process involved cleavage from BP to
classes lower than BM. These converging findings strongly support
the existence of PSD and the validity and effectiveness of the
HBCB model.
Tsutomu Oohashi**
Foundation for Advancement of
International Science
Osamu Uenoy
National Center of Neurology and
Psychiatry
Japan Science and Technology
Agency
Tadao Maekawaz
Yokkaichi University
Norie Kawai**
Foundation for Advancement of
International Science
Emi Nishina§
The Graduate University for
Advanced Studies
National Institute of Multimedia
Education
Manabu Honda*,y
National Center of Neurology and
Psychiatry
Japan Science and Technology
Agency
Keywords
Collective reutilizability, bond energy,
programmed self-decomposition model,
unicellular organism, hydrolysis, individual
death
* Contact author.
** Department of Research and Development, Foundation for Advancement of International Science, 1-53-11 Higashinakano, Nakano-ku,
Tokyo 164-0003, Japan. E-mail: oohashi@fais.or.jp (T.O.); nkawai@fais.or.jp (N.K.)
y Department of Cortical Function Disorders, National Center of Neurology and Psychiatry, and Japan Science and Technology Agency,
4-1-1 Ogawahigashi, Kodaira, Tokyo 187-8502, Japan. E-mail: ueno-o@ncnp.go.jp (O.U.); honda@ncnp.go.jp (M.H.)
z Faculty of Environmental and Information Sciences, Yokkaichi University, 1200 Kayou-cho, Yokkaichi, Mie 512-8512, Japan. E-mail:
maekawa@yokkaichi-u.ac.jp
§ School of Cultural and Social Studies, The Graduate University for Advanced Studies, and Research and Development Department,
National Institute of Multimedia Education, 2-12 Wakaba, Mihama-ku, Chiba 261-0014 Japan. E-mail: nishina@nime.ac.jp
n 2008 Massachusetts Institute of Technology
Artificial Life 15: 29 – 58 (2009)
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
5
1
2
9
1
6
6
2
5
6
6
a
r
t
l
.
/
.
.
2
0
0
9
1
5
1
1
5
1
0
3
p
d
.
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
T. Oohashi et al.
An Effective Hierarchical Model for the Biomolecular Covalent Bond
1 Introduction
One of the remarkable features of artificial chemistry (AChem) [5], in which analogue chemical
reactions are simulated on computers, is that it postulates that all the elementary steps of existing
terrestrial life consist of chemical phenomena, with no exceptions [3]. Moreover, AChem takes on
the construction of an even closer relationship with existent terrestrial life as its lofty goal [1, 2, 5,
26]. AChem research is thus expected to greatly advance conventional artificial life (ALife) research
by providing powerful approaches to studying actual intracellular life activities and their evolution [5,
17, 23, 24, 27]. Among such approaches, network artificial chemistry (NAC), proposed by Hideaki
Suzuki [24, 25], can be regarded as a useful paradigm that embraces various possibilities, widely
ranging from basic study to application development.
A principal feature of NAC is that it defines interelement relationships by topological networking.
Its conceptual framework and approach methodology enable NAC to express a dynamically and
continuously changing relationship, which cannot be fully defined by arrays of symbols [4] and lattice
structures [29], such as those used in conventional AChem research. NAC also focuses on the
strength of interelement linkages introduced as a hierarchy of covalent bonds, hydrogen bonds, and
van der Waals force. Then it constructed a model in which the difference in strength of the inter-
element linkage and the interaction between molecules or intramolecular units introduce molecular
structures and molecular reactivity. These two features, based on actual chemical phenomena, have
thrown light on the mechanism of cluster generation from molecular folding [25] and suggest that
this model attests to the temporal and spatial structure of an actual molecule.
Among the promising features of NAC, we are particularly interested in the hierarchical structure
of the strength of interelement linkage (bond energy). By introducing the hierarchical structure of
bond energy into chemical systems that constitute the metabolic system of terrestrial life, we can
hope to uncover the latent functionality and rationale of the chemical system.
Toward this end, we have used molecular cell biology to build a model of the hierarchical structure
of bond energy in the fundamental unit of terrestrial life, that is to say, the cell, a self-organized
chemical system consisting of molecules and their interaction and transformation. In concrete
terms, we have hierarchically categorized biomolecules in a cell into four classes on the basis of the
complexity of the interatomic network. We have also categorized biomolecular covalent bonds, which
comprise one class in Suzuki’s bond-energy hierarchy,
into three classes, based on bond energy.
We have found that these two hierarchical structures are fully compatible with one another. Accord-
ing to these classifications, we constructed a hierarchical model for the biomolecular covalent bond—
the hierarchical biomolecular covalent bond (HBCB) model—and examined its validity and effectiveness
as a way to coordinate two independent approaches: study of an existing terrestrial life system, and
AChem.
Coordination between building the model, comparing it with an existing terrestrial life system,
and examining it by means of AChem throws light on the usefulness of this study, which focuses on
AChem. Under the AChem paradigm the model itself can be described as an automaton in the form
of a chemical linkage that significantly corresponds to an existing terrestrial life system. If the model
succeeds in describing terrestrial life activities, we should be able to examine whether a phenotype of
the model actually exists in terrestrial life, by the use of cell biological methods.
However, we cannot examine the evolutionary superiority of the proposed model by comparing it
with alternative hypothetical models within the scope of biological studies of actual terrestrial life.
One reason is that an organism species as a phenotype of an alternative model may not exist.
Moreover, the time scale required for examining evolutionary superiority significantly exceeds the
operability of experiments on existing life.
AChem may therefore open the way for ultimate verification. Since AChem presupposes that all
the elementary steps of actual terrestrial life consist of chemical phenomena without exception, it is
feasible to build an automaton based on chemical linkages in imitation of existing terrestrial life and
to observe its behaviors in evolutionary simulations.
30
Artificial Life Volume 15, Number 1
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
5
1
2
9
1
6
6
2
5
6
6
a
r
t
l
.
/
.
.
2
0
0
9
1
5
1
1
5
1
0
3
p
d
.
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
T. Oohashi et al.
An Effective Hierarchical Model for the Biomolecular Covalent Bond
In this article, as a first step toward employing such perspectives, we built our HBCB model
under an NAC-inspired AChem paradigm in accord with our previously proposed programmed
self-decomposition ( PSD) model. This model assumes that terrestrial organisms arrange their bio-
molecules in a hierarchical structure according to the energy strength of their covalent bonds and
effectively utilize the hierarchical structure. Next, we examined the existence of a mechanism il-
lustrating this model by means of cell biological and biochemical methods, using existing terrestrial
life as experimental material. In parallel with this approach, we developed an experimental AChem
system by installing the HBCB model
into the SIVA series [19 – 21], our previously developed
artificial ecosystem, and examined whether life activities utilizing the HBCB model had evolutionary
superiority.
2 Construction of the Hierarchical Model for the Biomolecular Covalent Bond
2.1 Feasibility of Evolutionary Development of a Hierarchical Structure in the
Biomolecular Covalent Bond
The biological polymer (BP) responsible for essential and specific activities that actualize survival
and self-reproduction of living organisms has extremely large molecular size. At the same time, BPs
have a highly specific structure and function. Therefore, it is extremely difficult to reutilize them, not
only in another individual, but even in other tissues or organs in a single individual.
In order for living organisms to reutilize a BP, they must decompose it into smaller elements by
cleaving a considerable portion of the chemical bonds that form the BP, and build the required BP
with these elements. As the specificity of the structure and function of such elements decreases, their
general versatility increases. So, the smaller the elements of the decomposed BP are, the greater the
possibility that the decomposed product can be reused. Thus each decomposition product has a
different level of versatility as a biological resource. We call such versatility collective reutilizability.
However, reconnecting the elements into a large-scale biomolecule requires an appropriate amount
of bond energy. So the smaller the elements, the larger the amount of energy required for rebuilding.
That is to say, a kind of tradeoff takes place.
In biomolecular recycling, this relationship would exert an evolutionary pressure on terrestrial life to
keep energy loss as low as possible and make the collective reutilizability of decomposed elements as
great as possible in either living individuals or the ecosystem itself. At the same time, there would be
more evolutionary pressure to make the amount of released energy as small as possible in the
decomposing process and to keep the amount of bond energy possessed by the decomposed elements
as large as possible, because the energy to exercise the activities of terrestrial life is fundamentally
supplied in the form of chemical bond energy. Such evolutionary pressures might have induced
evolutionary selection of a reasonable hierarchical structure of covalent bonds and a biomolecular
recycling mechanism effectively utilizing such hierarchical structure. If our hypothesis is borne out, we
should be able to identify proofs that reflect the existence of such a mechanism in existent terrestrial
life. Therefore, we first attempted to hierarchically classify biomolecules constituting terrestrial life from
the viewpoint of system factors of an automaton in the form of chemical linkage.
2.2 Classification of Biomolecules into a Hierarchical Structure
A complex hierarchical structure is observed in the anatomy of terrestrial life. It can be categorized
into five classes, from highest to lowest: individual, organ, tissue, cell, organelle. We can hierarchically
classify the biomaterials that are the components of the above structures according to the complexity
of the interatomic network (Table 1). The five classes are biological polymer (BP), biological
monomer (BM), organic biomaterial (BO), inorganic biomaterial (BI), and basic bioelement (BE).
BE, the lowest fundamental class (class V), is composed of only five kinds of elements. The
second lowest class (class IV), BI, is composed of substances formed by combining the members of
the BE class. BO, the middle class (class III), is composed of substances formed by combining the
Artificial Life Volume 15, Number 1
31
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
5
1
2
9
1
6
6
2
5
6
6
a
r
t
l
.
/
.
.
2
0
0
9
1
5
1
1
5
1
0
3
p
d
.
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
T. Oohashi et al.
An Effective Hierarchical Model for the Biomolecular Covalent Bond
Table 1. Hierarchization of biomaterials of terrestrial life based on the complexity of interatomic networks.
Class name
Number of types
Substance examples
I. Biological polymer (BP)
102 – 109
Lipid, polysaccharide, protein, nucleic acid
II. Biological monomer (BM)
Several tens
Glycerol, fatty acid, hexose, amino acid, nucleotide, etc.
III. Organic biomaterial (BO)
More than ten
Pyruvic acid, acetyl-CoA, ribose, etc.
IV. Inorganic biomaterial (BI)
V. Basic bioelement (BE)
Four
Five
H2O, CO2, PO4
+
3(cid:3), NH4
C, H, O, N, P
members of the BI class. The members of the BI class and the BO class can exist independently and
stably. They have specific chemical properties that clearly differentiate them. The BM class (class II)
is composed of substances formed by combining the members of the BO class. The chemical
substance of the several tens of types in the BM class is further classified into four groups according
to their structure and function. It becomes possible to generate a remarkable diversity through
permutation and combination of the members of the BM class. This is how the highest class (class I),
BP, is composed. This class plays a fundamental role in self-reproductive activities and contains an
extremely large number of members.
The elements of class V do not exist independently as stable biomolecules. In the following
section, we focus on the top four classes (classes I – IV ) as the hierarchical structure of biomolecules
based on complexity (Figure 1a).
We next focus on conversions of the biomolecules to other classes. These conversions are
nothing more than the generation and cleavage of covalent bonds, which are essentially attributed
to energy absorption caused by bond generation and energy release caused by bond cleavage.
Regarding the amount of input and output of bond energy due to covalent bond generation and
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
5
1
2
9
1
6
6
2
5
6
6
a
r
t
l
.
/
.
.
2
0
0
9
1
5
1
1
5
1
0
3
p
d
.
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 1. Hierarchization of biomolecules and their collective reutilizability. (a) Hierarchical structure of biomolecules of
terrestrial life. (b) The collective reutilizability of biomolecules. Both intra- and interindividual collective reutilizability of
BPs is extremely limited. However, almost all BMs, which are classified just below the BPs, are reutilized in most
terrestrial organisms. (c) Free energy released in decomposition of biomolecules. Each bar indicates the amount of
released energy associated with each step of covalent bond cleavage in the decomposition of a hexose polymer.
32
Artificial Life Volume 15, Number 1
T. Oohashi et al.
An Effective Hierarchical Model for the Biomolecular Covalent Bond
cleavage, we note the tendency for the amount to decrease when a conversion occurs between
higher classes, and for it to increase when a conversion occurs between lower classes. Thus we can
hierarchically categorize covalent bonds between biomolecules into three classes on the basis of
bond energy.
Take the example of saccharides (Figure 1c). When a polysaccharide, a hexose polymer, in the BP
class is converted to monosaccharides in the BM class, there is a release of energy of approximately
13 kJ/mol accompanied by the cleavage of the glycosidic bond. Similarly, converting a monosac-
charide in the BM class to the components of the tricarboxylic acid (TCA) cycle in the BO class
generates a release of energy of approximately 200 kJ/mol (when decomposing one glucose mole-
cule into two molecules of lactic acid). Moreover, when BO is converted to BI such as carbon dioxide
and water, a release of energy of approximately 2700 kJ/mol (when decomposing two molecules of
lactic acid) associated with the cleavage of covalent bonds is generated [11]. Thus, in the molecules
that compose terrestrial life, the hierarchy inside the covalent bond can be roughly categorized into
three classes based on bond energy. This tendency is observed not just for saccharides, but for all
BPs, including proteins, nucleic acids, and lipids. It is noteworthy that the energy release associated
with a conversion from a BP to BMs is particularly small. Bonds connecting BMs to build BPs,
such as the glycosidic bond and peptide bond, all become broken by hydrolysis, which is a cleavage
of bonds by the intervention of water (H2O) and is characterized by a small energy gap accom-
panying the reaction.
We find that a four-class hierarchical structure of biomolecules based on complexity and a three-
class hierarchical structure of covalent bonds based on bond energy are fully compatible with one
another. We previously reported on a prototype version of this hierarchical classification of bio-
molecules [18, 21].
2.3 Programmed Self-decomposition Model
In exploring the characteristics of the hierarchical structure of biomolecular covalent bonds, we
regard biomolecules as resources for composing living organisms. In terms of the collective reuti-
lizability of resources, the circulation of material in a terrestrial ecosystem, namely, its biomolecular
recycling mechanism, provides significant insights. That mechanism, examined in this article,
is
discussed below.
The terrestrial ecosystem forms a nearly closed system in that both its space and substance are
limited. Accordingly, to maintain the stability of terrestrial life activities, the space and substance
removed from the environment by life activities have to be returned to the environment. That is to
say, the ecosystem must be returned to its original state. The mechanism for restoring the terrestrial
ecosystem has conventionally been explained by the principle of biological circulation called the food
chain [16], which is a biomolecular recycling mechanism for terrestrial life. We have set up a new
hypothesis that is complementary to that of the food chain. In our view of the terrestrial ecosystem,
besides the restoration of the environment by the food chain, another hidden mechanism is
fundamentally built into every life individual, by which it autonomously decomposes itself so as to
contribute to the restoration of the environment [18, 21]. We consider the phenomenon of decom-
position by the life individual’s own effort, called self-decomposition, to be a controlled biochemical process
of returning substance and space that the individual possesses to the environment for the purpose
of restoring the environment to its original state. We call this programmed self-decomposition (PSD). We
have developed a self-reproductive, self-decomposable (SRSD) automaton using von Neumann’s self-
reproductive automaton model [30, 31] as a prototype (Figure 2).
Von Neumann’s self-reproductive automaton model can be summarized as follows: automaton A
produces an automaton according to instruction tape I (information registered on a tape). Autom-
aton B replicates tape I. Automaton C combines with A and B, and controls them. Automaton D
is composed of A+B+C. Instruction tape I carries instructions to describe the automaton, and
ID carries the instructions of D. Automaton E, composed of D+ ID, can reproduce itself. Instruc-
tion tape ID+F carries instructions describing both D and F, which can be any given automata.
Artificial Life Volume 15, Number 1
33
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
5
1
2
9
1
6
6
2
5
6
6
a
r
t
l
.
/
.
.
2
0
0
9
1
5
1
1
5
1
0
3
p
d
.
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
T. Oohashi et al.
An Effective Hierarchical Model for the Biomolecular Covalent Bond
Figure 2. Von Neumann’s self-reproductive automaton and Oohashi’s self-reproductive, self-decomposable automaton.
(a) Von Neumann’s self-reproductive automaton model. This model embodies the sequence of self-reproduction of life as
a physical machine and its evolution in that process, without the sequence becoming a vicious circle. This is an immortal-
type model without an autonomous mechanism for the restoration of the environment to its original state. (b) Oohashi’s
self-reproductive, self-decomposable (SRSD) automaton model. This model uses von Neumann’s self-reproductive
automaton model as its prototype. It has a programmed mechanism contributing to the restoration of the environment
to its original state through autonomous individual death with self-decomposition, which is an essential feature of
terrestrial life.
Automaton EF, whose instruction tape ID is replaced by ID+F, can reproduce EF and produce
another automaton F. This model expresses the sequence of self-reproduction of life as a physical
machine and its evolution in that process, without falling into a vicious circle. It is also interesting in
that automaton E can be compared to a cell, and instruction tape I to its gene. However, this is an
immortal-type model without an autonomous mechanism for the restoration of the environment to
its original state.
Oohashi’s SRSD automaton [18, 21] was developed using von Neumann’s self-reproductive au-
tomaton EF as its prototype. Automaton FZ is a modular subsystem comparable to von Neumann’s
automaton F. It functions to decompose into components the whole automaton in which it is
incorporated. Instruction tape ID+FZ carries an instruction describing automaton D and an instruc-
tion describing the newly defined automaton FZ for decomposition. Automaton EFZ is comparable
to von Neumann’s automaton EF, whose tape ID+F is replaced by tape ID+FZ. Automaton G is
composed of EFZ and FZ, that is D+FZ+ ID+FZ. This automaton G can reproduce G itself, and
produce FZ as a module within G. FZ is usually latent, but it decomposes G when activated by a
certain trigger. Two activation modes are defined for the self-decomposition automaton FZ. The
first one is activated by a signal input from outside, indicating unconformity between the life and its
habitation environment. The second mode constitutes the end of the life span. If, after a certain
length of biological time has passed or a certain set of events has occurred, there is still no signal
input from outside to trigger an action, this situation itself becomes an internal trigger that activates
34
Artificial Life Volume 15, Number 1
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
5
1
2
9
1
6
6
2
5
6
6
a
r
t
l
.
/
.
.
2
0
0
9
1
5
1
1
5
1
0
3
p
d
.
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
T. Oohashi et al.
An Effective Hierarchical Model for the Biomolecular Covalent Bond
FZ. Note that the concepts of reproduction and decomposition partially include chemical reactions
known as anabolism and catabolism.
When we investigated actual life in relation to this SRSD automaton, one significant issue was the
cost-benefit comparison between the acquisition of collective reutilizability and the loss of bond
energy induced by covalent bond cleavage associated with self-decomposition by automaton FZ. It is
possible that the requirement for improvement of efficiency acts as an evolutionary pressure and that
the genetic programs of existent terrestrial life involved in a self-decomposition mechanism have
evolved toward more efficient biomolecular reutilization.
2.4 Hierarchical Model for the Biomolecular Covalent Bond
When we look on biomolecules as the resource for the material that composes terrestrial life, a highly
significant hierarchy is observed in terms of the collective reutilizability of biomolecules (see Fig-
ure 1b). Generally, BPs in the highest class have molecular structures that differ across individuals.
They differ even across different sites in the same individual. That means that collective reutilizability
of BPs is not guaranteed, not only between different individuals, but even between different tis-
sues or organs in a single individual. In contrast, it is noteworthy that the collective reutilizability
of biomolecules markedly increases as they are converted to the BM class. They then become
reutilizable in every organ and tissue in the same individual, as well as in terrestrial life in general.
Of course, this collective reutilizability is increased by a conversion to a class lower than BM (i.e.,
the BO or BI class) in some cases, but the increase is less than that caused by a conversion from BP
to BM.
When we look at the cost-benefit comparison between the acquisition of collective reutilizability
and the energy loss induced by cleavage of biomolecular covalent bonds, these two phenomena are
seen to be in the above-mentioned tradeoff relationship. Keeping collective reutilizability as high as
possible and energy release as low as possible at the point of bond cleavage would strengthen
survival capacity. It is possible that this requirement acts as an evolutionary pressure and leaves
behind certain universal mechanisms for existent terrestrial life as the fruits of evolutionary selection.
It may also be concluded that there exists a rational hierarchy of biomolecules, from the viewpoint
of bond energy, with an optimum point at which there is an advantageous decomposition from one
particular class to another, as compared to decomposition to a higher or lower class.
We suppose that this significant point corresponds to the process by which BPs are cleaved to
BMs. Members in the BM class can be universally reutilized among almost all species, and at the
same time, a conversion between this class and the higher BP class is associated with only a very
small amount of energy input and output. It can be surmised that an evolutionary pressure toward
this significant point has worked effectively during the evolutionary process.
We have made a working hypothesis that existing terrestrial
life evolutionarily selects a bio-
molecular recycling mechanism that optimizes cost-benefit performance by routing through the BM
class. In other words, we observe a trade-off between diversity, collective reutilizability, and energy
efficiency associated with an interclass conversion in the biomolecular hierarchical structure of ter-
restrial life. The BM class is a significant point at which collective reutilizability is almost assured for
all terrestrial life and, at the same time, the amount of released energy is minimized when converting
from the higher class. Therefore, we assume that a biomolecular recycling mechanism that optimizes
cost-benefit performance by making the BM class a relay point should be the outcome of evo-
lutionary selection.
We call this hypothesis the hierarchical model for the biomolecular covalent bond (HBCB model). If the
biomolecular recycling mechanism assumed in the model actually exists and manifests itself, the
evolutionary superiority, validity, and effectiveness of the HBCB model as well as the existence of a
PSD mechanism in the actual terrestrial life system will be clearly shown. In this study we examine
the validity and effectiveness of our HBCB model in simulations of AChem in conjunction with ob-
taining corroborative evidence for the existence of a PSD mechanism in experiments using terrestrial
life system.
Artificial Life Volume 15, Number 1
35
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
5
1
2
9
1
6
6
2
5
6
6
a
r
t
l
.
/
.
.
2
0
0
9
1
5
1
1
5
1
0
3
p
d
.
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
T. Oohashi et al.
An Effective Hierarchical Model for the Biomolecular Covalent Bond
3 Investigation of the PSD Model and the HBCB Model Utilizing a Living
Terrestrial Life System
3.1 Materials and Methods
3.1.1 Investigation Design
In an experiment utilizing living cells of actual terrestrial
life, we looked for the biomolecular
recycling mechanism proposed by our HBCB model. We asked if a self-decomposition mechanism
existed in a state consistent with the PSD model and if the mainstream of chemical reactions came
about as a result of the generation of BMs by hydrolytic cleavage of a BP based on the HBCB model.
It is advantageous for us to focus on the self-decomposition process in the experiment. Since var-
ious processes,
life
in general, it is not easy to examine the effectiveness of the HBCB model. On the other hand,
the process that leads to death and decomposition consists mostly of reactions leading to self-
decomposition and, limitedly, of other, contaminating simultaneous processes. Therefore, we consid-
ered the self-decomposition process the most appropriate choice for investigating the biomolecular
covalent bond cleavage process in living organisms in as coherent a state as possible.
including synthesis and decomposition, progress simultaneously in terrestrial
Accordingly, we first set the following four steps for verification:
1. Our most basic problem was to find an appropriate species from among existing terrestrial
life. Using such a species, we then looked for the autonomous degradation of cells that
could be interpreted as self-decomposition consistent with the PSD model.
2. We looked for evidence of the expression of self-decomposition corresponding to the
genetically programmed process.
3. We looked for evidence that self-decomposition in actual life was not a natural random
degradation of biological resources with energy release (exergonic reaction), but rather a
controlled active metabolic process with energy consumption (endergonic reaction).
4. We looked for evidence that hydrolytic reactions from a BP to BMs are the main
pathway of the autonomous degradation of cells, which can be interpreted as a
self-decomposition process.
We set up this four-step verification process as follows.
3.1.2 Test Organism
We selected the protozoan Tetrahymena pyriformis strain W [6, 15] for the experimental material for the
following reasons:
1. Tetrahymena is a unicellular organism, so cell death equals individual death. Experimenting
on Tetrahymena, therefore, excludes the problem of mixed occurrence of partial death
(including apoptosis) and individual death that is inevitable when using a multicellular
organism.
2. Tetrahymena is a eukaryotic cell and contains various kinds of independent organelles
(intracellular functional particles), so Tetrahymena can easily be compared with an automaton
model that defines the organelle as a functional modular subsystem.
3. Tetrahymena has unlimited proliferability without limitation of cell division caused by telomere
shortening. Although some species have both a macronucleus and a micronucleus and
perform sexual reproduction by conjugation, other species, such as the pyriformis used in this
study, have only a macronucleus and perform unlimited proliferation by asexual reproduction.
In this regard Tetrahymena pyriformis is analogous to the automaton model.
36
Artificial Life Volume 15, Number 1
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
5
1
2
9
1
6
6
2
5
6
6
a
r
t
l
.
/
.
.
2
0
0
9
1
5
1
1
5
1
0
3
p
d
.
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
T. Oohashi et al.
An Effective Hierarchical Model for the Biomolecular Covalent Bond
4. Tetrahymena has well-developed lysosomes, organelles in which various kinds of hydrolytic
enzymes densely accumulate. These enzymes degrade a BP into BMs by hydrolytic reaction.
This feature of the lysosome corresponds closely to the automaton FZ defined as the
functional modular subsystem for decomposition in our SRSD automaton model [18, 21].
5. The authentic type culture and the method of cultivating a pure culture are well established,
and there is no symbiosis of other gene systems in the cell. Consequently, this pure culture
contributes to a simplification of the experimental condition.
These five characteristics of Tetrahymena pyriformis make it an appropriate choice for the experi-
mental material we required for our investigation of the HBCB model and the PSD model.
3.1.3 Induction of Self-decomposition
To achieve clear-cut results in cell biological experiments, it is necessary to construct experimental
conditions in which the physiological activity of each cell is controlled in a coherent state so that self-
decomposition is released simultaneously in a large population of cultivated protozoa (approximately
105 individual cells in 1 ml of culture medium).
Two different types of triggers of self-decomposition are defined in the PSD model [18], namely,
an external factor of high unconformity with the environment (the first mode) and an internal factor
of natural life span (the second mode).
Self-decomposition as natural death at the end of the life span is a phenomenon readily observed
in nature. Since natural death is an autonomous internal phenomenon, it is difficult to avoid having a
large majority of normal living cells (individuals) and a small number of self-decomposing cells
randomly coexisting in the culture medium. Therefore, it is difficult to conduct a clear-cut experi-
ment that requires precisely controlled conditions without a time lag with respect to the biological
activity of each individual cell. Consequently, we could not adopt such a process in this experiment.
On the other hand, induction of self-decomposition by an encounter with an environment highly
incompatible with life activity can be manipulated by external factors including factitious operations.
Well-controlled experimental conditions can induce expression and progression of self-decomposition
in a way that satisfies the experimental objectives. If we wish to establish an experimental method
that induces self-decomposition in all cultured cells simultaneously, we must discover an external
message that triggers the self-decomposition program inside the gene. In other words, we must locate
environmental information signaling inadaptability, and develop a technique for simultaneously dis-
tributing such a signal to every cell in a culture. However, since such a signal might have an extremely
oppressive effect on all life activities, it is likely that this signal will cause serious damage to the
controlled physiological processes that are responsible for self-decomposition. Thus the signal might
interfere with the expression or progression of self-decomposition. We determined that the following
experimental protocol might overcome such an antinomy. First, an external signal triggering the
emergence of self-decomposition is given in a short period of time to activate the genetic program.
Next, the culture condition is immediately returned to one appropriate for life, so that the phys-
iological processes responsible for self-decomposition can progress with little damage. Based on this
principle, we developed the following method, which we call the impulse shock method.
First, to obtain the homogeneous physiological condition of every cultured cell, we subjected the
protozoan Tetrahymena [14] cells to a procedure that synchronized the cell cycles according to
established methods [32]. Next, we applied two treatments to this culture environment:
1. Impulse heat shock treatment: The temperature of the culture medium was rapidly increased to a
value unfit for survival, sustained for a short period of time, then returned rapidly to the original
optimum condition. In the present experiment, we increased the temperature of the entire culture
medium to 39jC, kept it there for 21 min, then returned it to the original temperature of 26jC [28].
2. Impulse pH shock treatment: Similarly, the hydrogen ion concentration (pH) of the Tetrahymena
culture medium was rapidly changed to a value unfit for survival, sustained for a short period
Artificial Life Volume 15, Number 1
37
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
5
1
2
9
1
6
6
2
5
6
6
a
r
t
l
.
/
.
.
2
0
0
9
1
5
1
1
5
1
0
3
p
d
.
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
T. Oohashi et al.
An Effective Hierarchical Model for the Biomolecular Covalent Bond
of time, then rapidly returned to the original optimum condition. In the present experiment,
we made the culture medium pH 4 by adding hydrochloric acid (acidification). After keeping
it there for 450 s, we quickly neutralized the culture medium to its original pH 7 by adding a
tris base, which becomes a weak basic buffer. Note that the above-mentioned examples of
concrete parameters might slightly change according to minor differences in experimental
conditions. The onset of the impulse shock treatment was designated as the 0 hour for
various observation and measurement purposes.
3.1.4 Observation of the Phenomenon of Self-decomposition
We observed the timeline of the morphological changes in the cells to confirm that the phenomenon
induced in each cell by the impulse shock method was a self-decomposition process. We used the
impulse heat shock treatment and supravital staining with acridine orange to stain the intracellular
lysosomes [13], which correspond to the automaton FZ in the PSD model. Acridine orange
specifically stains lysosomes and their contents in an acid condition. We then observed their behavior
under a fluorescence microscope.
3.1.5 Inhibition of the Self-decomposition Process
In order to investigate what kind of biochemical process self-decomposition is, the following pro-
cedures were administered immediately after returning the cultured cells to the appropriate culture
condition following the impulse shock treatment, and the influence of subsequent decomposition of
the cells was investigated in terms of population, individual condition (morphology and mobility),
and hydrolytic enzyme activity.
1. To investigate whether self-decomposition is a process programmed inside the gene, we
added actinomycin D, an inhibitor of the transcription from deoxyribonucleic acid (DNA)
to messenger ribonucleic acid (mRNA), to the culture medium.
2. To confirm if self-decomposition is an endergonic reaction, that is, an active process
requiring energy, the supply of oxygen to the medium was restricted for inhibition of
energy-requiring metabolic processes.
3. To investigate if the main reaction of the self-decomposition process is hydrolysis of a BP
into BMs, we added the reagent chloroquine, a specific inhibitor of the whole hydrolytic
enzyme group generated in the lysosome [8]. The lysosome is a strong candidate for a
decomposition module FZ in our SRSD automaton model.
3.2 Results
3.2.1 Morphological Changes in Tetrahymena Cells and Lysosomes in the
Self-decomposition Process Induced by Impulse Heat Shock Treatment
Impulse heat shock treatment of Tetrahymena cells brought about morphological changes of the cells
and changes in the behavior of the lysosomes that corresponded to the automaton FZ with its self-
decomposition function. We observed these changes by means of acridine orange supravital staining,
which causes areas in a neutral condition, such as normal cytoplasm and each of the organelles, to
appear green, and areas in an acid condition, namely the lysosome and its contents, to appear orange.
The following series of self-decomposition processes was observed synchronously in almost all of
the cells (Figure 3).
In normal living cells at the 0 hour, the cytoplasm and organelles were stained a similar green,
indicating a neutral condition, which is suitable for normal metabolic activities. There were a few
38
Artificial Life Volume 15, Number 1
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
5
1
2
9
1
6
6
2
5
6
6
a
r
t
l
.
/
.
.
2
0
0
9
1
5
1
1
5
1
0
3
p
d
.
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
T. Oohashi et al.
An Effective Hierarchical Model for the Biomolecular Covalent Bond
Figure 3. Morphological changes in Tetrahymena cells and lysosomes in the self-decomposition process induced by impulse
heat shock treatment. To observe the changes in appearance of cells and lysosomes induced by impulse heat shock
treatment, the distribution of intracellular pH was visualized by acridine orange supravital staining and photographed under
a fluorescence microscope. The green in the photographs indicates the areas or modules in a neutral condition, such as
normal cytoplasm. Orange indicates the areas or modules, including lysosomal granules, in an acid condition and the areas
throughout which the lysosomal contents were diffused. 0 hour: Normal living cells. There were a few lysosomal granules.
The entire cytoplasm is stained green, indicating a neutral condition. 1 hour: The number of lysosomal granules increased,
indicating an increase in lysosome biosynthesis. The cells lost mobility and swelled slightly. 2 hours: The number of lysosomal
granules further increased. The cells became spherical and decreased in volume, resulting in a further increase in the relative
density of lysosomes in a cell. 4 hours: Lysosomal membranes ruptured, and their contents diffused throughout the whole
cell, making the intracellular environment acidic. This condition activated the lysosomal acid hydrolytic enzymes. This finding
suggests that hydrolysis turning BPs into BMs was concentrated in this phase. Most of the cell membranes remained, and
each cell still included both intracellular components to be decomposed and hydrolytic enzymes under acidic conditions.
Such a situation would be reasonable for intensive decomposition of the intracellular contents by lysosomal enzymes.
6 hours: Cell membranes lysed, and cells decomposed into a homogenate.
particles stained orange, which were considered to be lysosomes. The cells had a pyriform shape and
were mobile.
One hour after the impulse heat shock treatment, almost all of the cells had lost mobility and had
slightly swelled. Although the cytoplasm and nucleus were green, indicating that they had maintained
ordinary life activity under neutral conditions, the number of acidic orange particles (i.e., lysosomes)
increased. These observations indicate the biosynthesis of lysosome and its contents, namely
hydrolytic enzymes. Lysosomal hydrolytic enzymes were all isolated within lysosomal granules. They
are activated specifically in a low-pH (acid) condition but not in a neutral or alkaline condition. We
concluded there had been very little hydrolysis of BPs in the cytoplasm during this phase. This
finding suggests that this phase is a preparatory stage for execution of self-decomposition.
Two hours after treatment, the number of lysosomal granules had further increased, although the
cytoplasm of the cells remained in a neutral condition. In addition, the cells had become spherical
and decreased in volume. The synergic effect of these factors resulted in a significant increase in the
relative density of the lysosomes in the cell. The increase of acidic lysosomal granules during this
period indicated an increase in lysosome biosynthesis, suggesting that an active biochemical process
in preparation for hydrolysis of self-decomposition was in progress.
Four hours after the impulse heat shock treatment, the lysosomal membranes had ruptured.
The lysosomal contents diffused at high density throughout the whole of each cell and mixed with
the BPs. Most of the cell membranes still remained. The intracellular environment in this phase
became acidic and the lysosomal enzymes became active, suggesting that hydrolysis was rapidly
proceeding. These processes seem reasonable candidates for intensive decomposition of the intra-
cellular contents by lysosomal enzymes. Since the systems to decompose BMs, such as glycolysis and
the TCA cycle, are optimized under neutral conditions, it is unlikely for such pathways to have
contributed to decomposition in this phase. It could be assumed that the cleavage of covalent bonds
associated with hydrolysis using lysosomal enzymes to turn a BP into BMs had played a major role in
the observed self-decomposition process. Measurement of the activity of the lysosomal hydrolytic
enzymes described below supports this assumption.
Six hours after treatment, the cell membranes had lysed and the cells had decomposed into a
homogenate. This finding suggests the completion of the self-decomposition process; the BP of the
Artificial Life Volume 15, Number 1
39
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
5
1
2
9
1
6
6
2
5
6
6
a
r
t
l
.
/
.
.
2
0
0
9
1
5
1
1
5
1
0
3
p
d
.
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
T. Oohashi et al.
An Effective Hierarchical Model for the Biomolecular Covalent Bond
Figure 4. Self-decomposition is an expression of a genetic program. (a) Self-decomposition of cells and its inhibition. Top:
Self-decomposition of cells induced by impulse pH shock treatment. Cells rapidly decomposed and became homogenate
in approximately 1.5 to 2 h. Bottom: When the expression of genetic programs was inhibited by adding the transcription-
inhibiting reagent actinomycin D immediately after induction of self-decomposition by impulse pH shock treatment, some
cells remained without self-decomposition, and some of these cells started moving again and began reproducing them-
selves. (b) Timeline for the decrease in cell number. The number of cells at each time point is shown as the percentage of
the number at 0 hour. Cells rapidly decreased in number after the impulse pH shock treatment. Inhibited expression of
genetic programs suppressed the decrease in cell number. These findings suggest that the self-decomposition mechanism
is a controlled biochemical process that is programmed in the genes and induced by its expression.
cells had decomposed into BMs, which, having a high degree of collective reutilizability, were re-
turned to the environment.
3.2.2 Examination of Whether Self-decomposition Is an Expression of a
Genetic Program
To inhibit the expression of a genetic program, actinomycin D, an inhibitor of transcription from
DNA to mRNA, was added into the culture medium after the impulse shock treatment; therefore,
the large cell population often suppressed the self-decomposition process even though there was
insufficient reproduction. Furthermore, a considerable part of the population of the cell was ob-
served to have recovered from this brief arrest of cell movement and reproduction (Figure 4). These
experimental results indicate that the decomposition of cells induced by impulse shock treatment
requires gene transcription, so that it must be a genetically controlled process according to the central
dogma. This finding suggests that gene programs predicated on the PSD model do exist and func-
tion as described above.
3.2.3 Examination of Whether Self-decomposition Is an Energy-Requiring,
Active Process
When general biological metabolic processes that require an energy supply were inhibited by
restricting the supply of oxygen to the culture medium, decomposition of cells was clearly inhibited
(Figure 5). This is an important finding because, if decomposition of the cells in this system were
merely uncontrolled random degradation with release of energy, decomposition of the cells would
progress without an energy supply. On the contrary, the decomposition process did not progress
without such an energy supply. This indicates that the observed decomposition is an endergonic,
actively controlled process with reduction of entropy. These results, indicating that self-degeneration
is a genetically controlled process that requires an energy supply, support our PSD model.
40
Artificial Life Volume 15, Number 1
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
5
1
2
9
1
6
6
2
5
6
6
a
r
t
l
.
/
.
.
2
0
0
9
1
5
1
1
5
1
0
3
p
d
.
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
T. Oohashi et al.
An Effective Hierarchical Model for the Biomolecular Covalent Bond
3.2.4 Examination of Whether the Main Reaction of the Self-decomposition
Process Lies in Hydrolysis Decomposing a Biological Polymer into a
Biological Monomer
The lysosomal hydrolytic enzyme group specifically acts to cleave the covalent bonds of a BP into
BMs. The addition of chloroquine, which is a comprehensive and exhaustive inhibitor of these lyso-
somal hydrolytic enzyme activities, into the culture medium clearly inhibited the self-decomposition
of the cells (Figure 5). This means that the hydrolytic activities of the lysosomal enzymes were an
essential factor in the observed decomposition. This finding further supports our HBCB model.
3.2.5 Confirmation that Self-decomposition Is an Energy-Requiring, Genetically
Regulated Process Mediated by Lysosomal Hydrolytic Reactions
To further confirm that the self-decomposition is an energy-requiring, genetically regulated process
and is mediated by lysosomal hydrolytic reactions, we directly measured the activity of the lysosomal
enzymes during the self-decomposition process and examined whether the activity change was
affected by the inhibition of the expression of the genetic program or an energy-requiring, active
metabolic process. Two of the most common marker enzymes, representative of all lysosomal acid
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
5
1
2
9
1
6
6
2
5
6
6
a
r
t
l
.
/
.
.
2
0
0
9
1
5
1
1
5
1
0
3
p
d
.
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 5. Self-decomposition is an energy-requiring metabolic process involving hydrolysis through lysosomal acid
hydrolytic enzymes as one of the main processes. (a) Self-decomposition of cells and its inhibition. Top: Self-
decomposition of cells induced by the impulse heat shock treatment. Cells rapidly decomposed and became homogenate
in approximately 4 to 6 h. Middle: Inhibition of energy-requiring metabolic processes by restriction of the oxygen supply
immediately after impulse heat shock treatment. Self-decomposition of cells was significantly suppressed. Bottom:
Inhibition of all lysosomal hydrolytic enzyme activities by the addition of the acidotropic reagent chloroquine immediately
after impulse heat shock treatment. Self-decomposition of cells was significantly suppressed. (b) Timeline for the
decrease in cell number. The number of cells at each time point is shown as the percentage of the number at 0 hour.
Cells rapidly decreased in number after the impulse heat shock treatment. Inhibition of the energy-requiring metabolic
processes and inhibition of the lysosomal hydrolytic enzyme activities significantly suppressed the decrease in cell
number. These findings suggest that self-decomposition is an endergonic (i.e., energy-requiring) active metabolic process
and that hydrolysis, which uses lysosomal enzymes to turn BPs into BMs, plays a major role in the self-decomposition
process.
Artificial Life Volume 15, Number 1
41
T. Oohashi et al.
An Effective Hierarchical Model for the Biomolecular Covalent Bond
Figure 6. Behavior of the lysosomal acid hydrolytic enzymes in the self-decomposition process shows that self-
decomposition is an energy-requiring, genetically regulated process. During the self-decomposition process induced by
impulse pH shock treatment, we measured the intracellular activities of the most common marker enzymes of the
lysosomal acid hydrolytic enzymes: (a) h-N-acetylhexosaminidase and (b) acid phosphatase. The enzymes’ activity
increased up to approximately 10 times that of the controls 2 h after the induction of self-decomposition. Inhibition of
the transcription of genetic programs by actinomycin D (blue line) and inhibition of energy-requiring metabolic processes
by restriction of the oxygen supply significantly (yellow line) suppressed the increase in activity of the hydrolytic
enzymes. These findings further confirm that self-decomposition is an endergonic (i.e., energy-requiring) genetically
regulated process mediated by lysosomal hydrolytic enzymes that decompose BPs into BMs.
hydrolytic enzyme activity, were investigated. The activity of both marker enzymes increased up to
approximately ten times that of the control as self-decomposition progressed (Figure 6). This
finding, together with changes in intracellular pH distribution (Figure 3), confirmed that self-
decomposition proceeded as an accumulation of various hydrolytic reactions from BP to BM. In
addition, such increases in enzymatic activity were remarkably suppressed by the inhibition of the
expression of the genetic program or an energy-requiring, active metabolic process. This indicates
that a causal relationship exists between lysosomal activities and expression of the genetic program
or energy consumption. Such results confirm that self-decomposition is an energy-requiring, geneti-
cally regulated process mediated by lysosomal hydrolytic reactions.
Such experimental results, which were not contradictory but complementary among different
indexes, support the notion that existent terrestrial life utilizes a hierarchical structure as shown by
our HBCB model and that existent terrestrial
life realizes effective reuse of materials by self-
decomposition. As a survival strategy in evolution and selection, terrestrial life might acquire and
preserve the mechanism to return BPs to the environment as BMs with a high degree of collective
reutilizability through hydrolysis in the self-decomposition process.
As mentioned above, it is impossible in an experiment that uses actual terrestrial life to examine
this hypothesis by comparing it with alternative hypotheses in terms of evolutionary superiority. It is
necessary to examine the validity of the HBCB model in the context of evolution through computer
simulation, making full use of AChem activity.
4 Artificial Chemistry Simulation of Evolutionary Superiority of a Hierarchical
Structure of Biomolecular Covalent Bonds
The existence of a PSD mechanism that utilizes the hierarchical structure of biomolecular covalent
bonds has been confirmed by our biological experiments using an existent terrestrial life system. This
suggests the possibility that decomposition from the BP class to the BM class has been evolutionarily
selected. We used our SIVA-T05 (Simulator for Individuals of Virtual Automata— Terra 2005), an
experimental AChem system with which we had performed simulation experiments,1 to examine
how evolutionarily advantageous this decomposition process is.
1 Anyone who is interested in obtaining SIVA-T05 can contact the corresponding author by e-mail.
42
Artificial Life Volume 15, Number 1
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
5
1
2
9
1
6
6
2
5
6
6
a
r
t
l
.
/
.
.
2
0
0
9
1
5
1
1
5
1
0
3
p
d
.
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
T. Oohashi et al.
An Effective Hierarchical Model for the Biomolecular Covalent Bond
4.1 Architecture of SIVA-T05
4.1.1 The Design Concept of SIVA-T05
We developed a virtual ecosystem series SIVA [19 – 21] configured with Oohashi’s SRSD automaton
installed in a finite, heterogeneous environment consisting of virtual biomolecules with chemical
reactivity. Since constructing SIVA-III, a pioneering prototype for an AChem system, in 1996 [19],
we have continued to develop SIVA as a virtual ecosystem based on AChem. To promote the major
purpose of AChem, namely, the achievement of a closer relationship with existent terrestrial life,
SIVA-T05, a new version of SIVA, has been developed based on the following design concepts.
1. In accordance with the actual terrestrial environment, the virtual environment that virtual life
individuals (VLIs) inhabit in SIVA-T05 has limited amounts of space, materials, and energy.
The materials, energy, and temperature are heterogeneously distributed throughout the whole
environment.
2. The virtual biomolecules making up a VLI are hierarchically organized according to the
HBCB model. In terrestrial organisms, proteins are constructed by polymerization of amino
acids, and nucleic acids are constructed by polymerization of nucleotides. Likewise, an element
in a certain class in the hierarchical structure of virtual biomolecules consists of elements
belonging to the next lower class. In addition, interclass conversion, such as synthesis and
decomposition, is associated with release and absorption of energy corresponding to each
class.
3. As in terrestrial life, virtual biological polymers and monomers are categorized into two
groups: the constitutive information group, which serves as the gene-preserving information
about structure and function of the VLI, and the functional module group, which serves as
proteins or enzymes expressing life activities in the VLI. The constitutive information group
and the functional module group correspond to the genotype and the phenotype in terrestrial
life. Virtual biomolecules belonging to the functional module group are synthesized according
to the information described by virtual biomolecules belonging to the constitutive information
group.
4. A functional automaton for self-reproduction and self-decomposition acts as the phenotype
of a VLI, and a virtual genome acts as the genotype. The functional automaton consists of
virtual biomolecules belonging to the functional module group, whereas the virtual genome
consists of ones belonging to the constitutive information group.
5. A VLI reproduces itself by using materials and energy existing in the virtual environment.
Activities of a VLI depend on the amount of materials and energy as well as the temperature
in its habitation point.
6. For each VLI, the optimum environmental conditions are defined a priori. A VLI cannot
express its life activities when actual environmental conditions at its habitation point
markedly deviate from its optimum ones.
7. A VLI can be set to decompose itself when the environmental conditions at its habitation
point deviate from the optimum for a VLI, or when it has lived out its life span. Materials
and energy released in association with the decomposition of a VLI are restored to the
environment.
8. Mutation can occur in the virtual genome and may change the optimum environmental conditions
of a VLI. This may enable a VLI to live in an environment in which it originally could not. That
is to say, evolutionary adaptation can occur.
In the following sections, we describe how these concepts are implemented in SIVA-T05.
Artificial Life Volume 15, Number 1
43
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
5
1
2
9
1
6
6
2
5
6
6
a
r
t
l
.
/
.
.
2
0
0
9
1
5
1
1
5
1
0
3
p
d
.
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
T. Oohashi et al.
An Effective Hierarchical Model for the Biomolecular Covalent Bond
4.1.2 Environmental Design of SIVA-T05
To simulate the characteristics of a terrestrial environment with limited amounts of materials and
energy distributed in a finite space, the virtual space of SIVA-T05 is designed to be a two-
dimensional lattice consisting of 16 (cid:4) 16 (= 256) spatial blocks. A single spatial block is defined as
8 (cid:4) 8 (= 64) pixels for habitation points. One habitation point is occupied by one VLI and vice
versa (Figure 7a). Environmental conditions can be independently defined for each spatial block, and
those of the 64 habitation points in the same spatial block are configured to always be homogeneous.
Since all VLIs in one spatial block share the same environmental conditions, the population of VLIs
in that block significantly affects the local condition. Consequently the divergence of local
environmental conditions across the whole ecosystem is gradually emphasized along with the
proliferation of VLIs, as would happen in a terrestrial ecosystem.
The temperature gradient and the initial distribution of virtual energy and four kinds of virtual
inorganic biomaterials (see Section 4.1.3) consisting of VLIs are heterogeneous across the whole
ecosystem (Figure 7b). No substances other than virtual inorganic biomaterials exist in the initial
environment. To simulate the effects of solar energy and its diffusion and radiation in the terrestrial
ecosystem, a predefined amount of energy per time unit is refilled, and the total amount of energy in
each spatial block must not exceed a predetermined threshold. The amount of refilled energy and the
upper limit of total energy are set at appropriate levels so that a simulation does not become
meaningless, that is, not so small that no VLI can live stably, and not so large that all VLIs can always
live without any failures. In order to compare the reproduction processes among the four different
species of virtual life under identical conditions, as below, all the conditions of the four spatial blocks
at the center of the virtual ecosystem are set to be identical at the outset.
4.1.3 Composition of Virtual Biomolecules
In SIVA-T05, we have designed a new type of virtual life based on the HBCB model. Table 2 shows
the design of the hierarchical structure of virtual biomolecules based on the complexity of the
interatomic network of actual biomolecules making up terrestrial life.
Virtual biological polymers (VPs) and virtual biological monomers (VMs) are categorized into
two groups: the functional module group and the constitutive information group, which correspond
to the phenotype and the genotype, respectively, in terrestrial life.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
5
1
2
9
1
6
6
2
5
6
6
a
r
t
l
.
/
.
.
2
0
0
9
1
5
1
1
5
1
0
3
p
d
.
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 7. Environmental conditions of the virtual ecosystem SIVA-T05 are designed to be finite and heterogeneous.
(a) Spatial design. The virtual space of SIVA-T05 is a two-dimensional lattice consisting of 16 (cid:4) 16 (=256) spatial blocks,
each of which consisted of 8 (cid:4) 8 (=64) habitation points. One habitation point is occupied by one VLI and vice versa.
Environmental conditions of each spatial block can be independently defined, and those of the 64 habitation points in the
same spatial block are configured to be always homogeneous. (b) Spatial distribution of environmental conditions. Upper
left: Distribution of environmental temperature. Lower left: Initial distribution of energy stocked in each spatial block. A
predefined amount of energy is refilled from outside the virtual ecosystem to simulate the effect of solar energy. Right:
Initial distribution of four kinds of virtual inorganic biomaterials (VI). Each substance flows between neighboring spatial
blocks to restore the environment to the initial condition when the amount of a substance goes above or below that of
the predetermined level. All the environmental conditions of the four spatial blocks at the center of the virtual ecosystem
are set to be the same at the beginning in order to compare the reproduction processes of the four different species of
virtual life that are initially seeded under identical conditions.
44
Artificial Life Volume 15, Number 1
T. Oohashi et al.
An Effective Hierarchical Model for the Biomolecular Covalent Bond
Table 2 . Hierarchization of virtual biomolecules composing virtual
network.
life based on the complexity of the interatomic
Class name
Virtual biological
polymer (VP)
Functional unit
Functional module group
Constitutive information group
Polymerized functional units
Functional word
(command)
Temporary information word
Virtual codon
(variable, relational operator, etc.)
Virtual biological
O P Q R
I J K L
monomer (VM)
(4 kinds)
0 1 2 3 4 5 6 7 8 9
(14 kinds)
W X Y Z
(4 kinds)
Virtual organic
biomaterial (VO)
Virtual inorganic
biomaterial (VI)
A B C D (4 kinds, uppercase letters)
a b c d (4 kinds, lowercase letters)
Basically each substance in a certain class consists of several elements belonging to the next lower
class. For example, a virtual organic biomaterial (VO) consists of several virtual
inorganic
biomaterials (VIs), and a VM consists of several VOs. Several VMs constitute a functional unit,
which is a subclass of the VP class, and several functional units constitute a larger VP. In the present
simulation experiments, we designed five VMs as a single functional unit. A functional unit serves as
one word in the SIVA language in the functional module group, while it serves as a virtual codon
(Vcodon) in the constitutive information group (see Section 4.1.4).
The amount of released energy associated with decomposition of a virtual biomolecule into ele-
ments in a lower class, and the absorbed energy associated with synthesis of a biomolecule in a
higher class from elements, are defined as shown in Table 3. Note that the hierarchical structure of
virtual biomolecules based on complexity corresponds closely to that based on bond energy. In
principle, released energy and absorbed energy do not have the same value in actual terrestrial life.
To simplify the experimental conditions, however, the same value was used for both in this
experiment. The relationship between the amount of bond energy associated with different interclass
conversions was also simplified. The bond energy associated with conversion between the VP class
Table 3. Amount of energy released or absorbed by the conversion between virtual biomolecular classes.
Conversion between classes
Amount of energy
(units per molecule in the
lower class)
Virtual biomolecular polymer (VP) and virtual
0.1
biomolecular monomer (VM)
Virtual biomolecular monomer (VM) and
virtual organic biomaterial (VO)
Virtual organic biomaterial (VO) and virtual
inorganic biomaterial (VI)
1
10
Artificial Life Volume 15, Number 1
45
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
5
1
2
9
1
6
6
2
5
6
6
a
r
t
l
.
/
.
.
2
0
0
9
1
5
1
1
5
1
0
3
p
d
.
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
T. Oohashi et al.
An Effective Hierarchical Model for the Biomolecular Covalent Bond
and its subclasses (the functional unit classes) is the same as the one associated with conversion
between VP and VM.
4.1.4 Function of Virtual Life and Features of Virtual Life Activities
Oohashi’s SRSD automaton is installed as an artificial life form in SIVA-T05 (Figure 8). The VLI
consists of a virtual genome and functional automata. The virtual genome is a VP of the constitutive
information group and corresponds to instruction tape I in Figure 8, whereas the functional autom-
ata are VPs belonging to the functional module group and correspond to automata A, B, C, and FZ
in Figure 8. The virtual genome covers the functions of preservation, replication, and transcription
of structural and functional information about a VLI, while the functional automaton covers various
life activities of the VLI, such as synthesis, decomposition, and reproduction.
The virtual genome consists of the sequence of four kinds of VM (W, X, Y, Z in Table 2)
corresponding to the nucleotide in terrestrial life (Figures 8 and 9a). In the virtual genome, five
VMs constitute a functional unit, which serves as a Vcodon. Namely, each Vcodon is defined to
correspond to one of 18 kinds of VM (I, J, K, L; O, P, Q, R; 0 – 9 in Table 2) of the functional
module group (i.e., virtual amino acid: VAA). The sequence of Vcodons defines the sequence of the
VAAs in a functional automaton. The sequence information regarding all the automata is described
in the virtual genome. For the reproduction of a VLI, automaton B replicates the whole virtual
genome and automaton A synthesizes a functional automaton. Mimicking the transcription from
DNA to mRNA in actual terrestrial life, automaton A first copies the corresponding part of the
Vcodon sequence and then converts the transcript into a VAA sequence. Mutation can occur in either
of these copying processes (see Section 4.1.6).
SIVA-T05 executes the functions of the automata described by the SIVA language as an
interpreter, and thus the life activities of VLIs are expressed. First, a functional unit consisting of a
sequence of five VAAs serves as a hwordi in the SIVA language. A hwordi can be categorized as a
functional word, which serves as an executable hcommandi, or as a temporary information word
(Table 2). A hcommandi as a functional word covers a substantial part of the life activities of a VLI,
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
5
1
2
9
1
6
6
2
5
6
6
a
r
t
l
.
/
.
.
2
0
0
9
1
5
1
1
5
1
0
3
p
d
.
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 8. Relationship between life activities of virtual life individuals (VLIs) and the environment in SIVA-T05. Oohashi’s
self-reproductive, self-decomposable automaton is implemented in the VLI in SIVA-T05. Each VLI consists of functional
automata for self-reproduction [ D (=A+B+C)], those for self-decomposition [FZ], and an instruction tape [ID+FZ] (i.e.,
a virtual genome) that is a blueprint of all the automata. Automaton A produces all the functional automata described in
the virtual genome. Automaton B replicates the virtual genome. Automaton C constitutes a daughter VLI, combining the
automata newly synthesized by automaton A and the virtual genome replicated by automaton B, and divides it from the
parental VLI. Automaton FZ decomposes a VLI when it encounters environmental conditions unsuitable for survival or
when it lives out its life span. A VLI can reproduce itself by uptake of substances and energy that exist in the spatial block
to which its habitation point belongs. During self-decomposition, the substances and the energy generated by the de-
composition of virtual biomolecules constituting the VLI are restored to the spatial block. The occupied space (i.e., the
habitation point) is also released for utilization by another VLI.
46
Artificial Life Volume 15, Number 1
T. Oohashi et al.
An Effective Hierarchical Model for the Biomolecular Covalent Bond
Figure 9. Examples of SIVA language statements that describe Oohashi’s self-reproductive, self-decomposable (SRSD)
automaton. (a) Synthesis of a functional automaton based on the virtual genome. The functional automaton and the
virtual genome in Oohashi’s SRSD automaton model correspond to the phenotype and the genotype of terrestrial life.
Functional automata are synthesized from the information contained in the virtual genome. In the virtual genome, a
sequence of the four different kinds of the five VMs in the constitutive information group forms a virtual codon (Vcodon).
Each Vcodon corresponds to one of the VMs in the functional module group [viz., a virtual amino acid (VAA)], and the
sequence of VAAs in the functional automaton is determined by the sequence of the Vcodons. A sequence of five VAAs
corresponds to one word in the SIVA language. One or more words constitute a sentence, and one or more sentences
constitute a paragraph. One paragraph corresponds to one functional automaton. (b) An example of a functional
automaton described in the SIVA language used in the present simulation. There is an hIDi at the beginning of each
paragraph. ID1, ID2, ID3, and ID4 correspond to automata A, B, C, and FZ in Figure 8, respectively. The meanings of
the hcommandis and hvariableis used in this description are as follows: hcommandi syntha: reading information in the
virtual genome and synthesizing a functional automaton; movef: moving an automaton to the next tag on the virtual
genome; copyi: duplicating the virtual genome; divid: dividing a VLI; decsfm: decomposing itself (i.e., automaton FZ)
into VMs; decam: decomposing a functional automaton into VMs; decim: decomposing the virtual genome into VMs.
hvariablei length_AP: the total number of functional automata existing in a VLI; max_length_AP: the maximum
number of automata that a VLI can possess (usually corresponding to twice the total number of automata described in
the virtual genome); length_IM: the total
length of the virtual genome existing in a VLI; max_length_IM: the
maximum length of the virtual genome that a VLI can possess (usually corresponding to twice the total length of the
original virtual genome); unconformity: index representing the degree of unfitness; age: index representing the age
of a VLI in TC, the time unit of the simulation.
for example, syntha for synthesizing a functional automaton based on the information described
in the virtual genome, copyi for replicating the virtual genome, divid for dividing a VLI, and
decam for decomposing functional automata. A temporary information word can be a hvariablei
(e.g., length_AP representing the total number of functional automata in a VLI), hrelational
operatori (e.g., > = meaning the left side is greater than or equal to the right side), hnumberi
(ranging from 00000 to 99999), hperiodi (. , representing the end of a sentence), or hIDi (from
ID0 to ID255, representing the beginning of a paragraph).
One or more words constitute a hsentencei, which has to include zero or more hcommandis and
one hperiodi at the end. Before a hcommandi, a hsentencei can include one or more conditional phrases,
each of which consists of a combination of a hvariablei or hnumberi and a hrelational operatori. When
there is no conditional phrase in the hsentencei, hcommandis are directly executed in the order described
in the hsentencei. If a hsentencei includes any conditional phrases, a hcommandi is executed only when
all the conditional phrases are true but not when any of the conditional phrases is false.
A collection of hsentenceis constitutes a hparagraphi. Namely, one hparagraphi consists of zero
or more hsentenceis with one hIDi at the beginning. The hsentenceis in a hparagraphi are executed
in the order described in the hparagraphi. Since the results of the conditional phrase judgments are
reset when the hsentencei ends, they do not affect the execution of the next hsentencei. The
structure of hsentencei and hparagraphi can be described as follows in Backus-Naur form:
hsentencei ::¼
ð½hvariablei hnumberi(cid:10) hrelational operatori ½hvariablei
hnumberi(cid:10)Þ(cid:12) hcommandi(cid:12) hperiodi
Artificial Life Volume 15, Number 1
47
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
5
1
2
9
1
6
6
2
5
6
6
a
r
t
l
.
/
.
.
2
0
0
9
1
5
1
1
5
1
0
3
p
d
.
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
T. Oohashi et al.
An Effective Hierarchical Model for the Biomolecular Covalent Bond
hparagraphi ::¼
hIDi hperiodi
hsentencei(cid:12)
One hparagraphi described in SIVA language corresponds to a functional automaton in the life
activities of a VLI. Figure 9b gives an example of the descriptions of a functional automaton used in
the present simulation experiment. Each functional automaton in Figure 8 corresponds to a
hparagraphi as follows: Automaton A corresponds to ID1 and synthesizes functional automata
based on the information in the virtual genome by executing syntha. Automaton B corresponds
to ID2 and replicates the virtual genome by copyi. Automaton C, corresponding to ID3,
constitutes a daughter by combining the newly synthesized automata and the replicated virtual
genome, and divides it from a parent by executing divid when all the functional automata have
been synthesized and all the virtual genomes are replicated, that is, when both the conditional
phrases length_AP >= max_length_AP and length_IM >= max_length_IM become
true (see the legend of Figure 9 for details of each variable). Automaton FZ, corresponding to
ID4, decomposes the virtual genome and all the functional automata except for itself by executing
decim and decam when the environmental conditions become unsuitable for the VLI’s survival
(namely, when the conditional phrase unconformity > 2 becomes true) or when the VLI lives
out its life span (namely, when the conditional phrase age > 20 becomes true). Furthermore,
automaton FZ decomposes itself by executing decsfm when the whole virtual genome and all the
functional automata other than itself have been decomposed (namely, when both the conditional
phrases length_IM = 0 and length_AP = 1 become true). Apart from these hparagraphis
describing functional automata, there is another hparagraphi (viz., ID0), which defines various
parameters specific to each VLI, including the optimum temperature.
Each VLI expresses its life activities by executing all hparagraphis during one time count (TC), the
unit of virtual time in SIVA-T05. The order in which a VLI in the virtual ecosystem expresses its life
activities within one TC is randomly determined at every TC.
4.1.5 Relationship between Environmental Conditions and Activities of
Functional Automaton
All activities of the functional automata are designed to be affected by the environmental
temperature and the amounts of substances and energy according to the properties of chemical
reactions in actual terrestrial life (Figure 10). First, activities of the functional automata are set to be
Figure 10. Design of the relationship between environmental conditions and the activity of a functional automaton. The
activity of each functional automaton constituting a VLI is designed to be maximal when a VLI exists in the optimum
environment specific to it and to decrease when its environmental conditions deviate from the optimum state.
(a) Relationship between the activity of a functional automaton and environmental temperature. The activity is designed
to decrease according to a pseudo-Gaussian curve as the environmental temperature deviates from the optimum
temperature for the VLI. (b) Relationship between the activity of a functional automaton and the concentration of virtual
inorganic biomaterials (VI). As the concentration of VI necessary for self-reproduction increases, the self-reproduction
activity of the functional automaton asymptotically increases according to a pseudo-Michaelis-Menten equation.
48
Artificial Life Volume 15, Number 1
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
5
1
2
9
1
6
6
2
5
6
6
a
r
t
l
.
/
.
.
2
0
0
9
1
5
1
1
5
1
0
3
p
d
.
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
T. Oohashi et al.
An Effective Hierarchical Model for the Biomolecular Covalent Bond
Figure 11. Concept of mutation and evolutionary adaptation employed in SIVA-T05. When VLIs replicate the virtual
genome for self-reproduction in SIVA-T05, at a certain probability a VM in the replicated virtual genome is randomly
replaced by another type of VM belonging to the constitutive information group (i.e., W, X, Y, or Z). Most new
phenotypes produced as a result of mutation provide negative change or no positive change in life activities, including
environmental adaptability. To a certain extent, however, a positive change in life activities can be expected to occur and
a mutant emerge whose optimal environmental conditions differ from those for its parent. Such a mutant can live in a
spatial block with environmental conditions under which its parental individual cannot. In this article, we refer to this
phenomenon as evolutionary adaptation.
maximal when the temperature of the habitation point is optimum for the VLI and to decrease
according to a pseudo-Gaussian curve when the temperature deviates from the optimum [12].
Next, activities of the functional automata related to self-reproduction are designed to asymptoti-
cally increase as the concentration of VI increases in accordance with the Michaelis-Menten equation
[12], which describes the relationship between substrate concentration and chemical reaction rate. An
actual chemical reaction in terrestrial life often shows some maximal points rather than monotonic
increase, because, as typically seen for hydrogen-ion concentration (pH), an increase in concentration of
a certain substance can affect a state of the other substances, such as the ionization state. In this
experiment, however, we adopted the simpler relationship for the first step in our investigation.
Table 3 lists the different amounts of energy required for the synthesis of the functional autom-
aton or the replication of the virtual genome according to the class to which the virtual biomolecular
materials belong. As the amount of energy necessary for reproduction increases relative to the
amount of available energy in the local environment, the activities of the functional automaton
decrease. In actual terrestrial life, energy necessary for life activities can be stored in life individuals
as the bond energy of biomolecules such as adenosine triphosphate (ATP). In this simulation
experiment, however, VLI can utilize only the energy existing in the environment, for simplification.
To implement changes in the functional automaton activities in SIVA-T05, the probability of
successful execution of a hcommandi is designed to decrease as the difference between the environ-
mental temperature and the optimum temperature for the VLI becomes greater, or as the amounts
of substances and energy necessary for reproduction increase relative to the amounts in the
environment. Under these conditions,
if the expression of life activities (i.e., the execution of
hcommandis) fails, then unconformity, which is a variable representing unfitness with respect
to the environment, increases by 1. Similarly, when there is VLI division, if the adjacent pixels of
it fails to divide itself and
a parent VLI’s habitation point are already occupied by other VLIs,
Artificial Life Volume 15, Number 1
49
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
5
1
2
9
1
6
6
2
5
6
6
a
r
t
l
.
/
.
.
2
0
0
9
1
5
1
1
5
1
0
3
p
d
.
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
T. Oohashi et al.
An Effective Hierarchical Model for the Biomolecular Covalent Bond
increases unconformity by 1. When unconformity becomes greater than the predetermined
threshold (namely, when the conditional phrase unconformity > 2 of ID4 in Figure 9b
becomes true), the self-decomposition process starts. Self-decomposition also starts when the
variable age representing the age of a VLI reaches the predetermined life span (namely, when the
conditional phrase age > 20 in ID4 becomes true). The substances and energy released by the self-
decomposition are returned to the spatial block where the VLI exists.
4.1.6 Mutation and Evolutionary Adaptation
SIVA-T05 can simulate various types of mutation occurring in the genes of actual terrestrial life.
Figure 11 illustrates the concept of mutation and evolutionary adaptation in SIVA-T05. In the
present simulation we used only the VM substitution that occurs when copying the virtual genome.
There are two situations in the life activities of VLIs in which substitution can occur: when
automaton B replicates the virtual genome for reproduction (i.e., replication of DNA in terrestrial
life), and when automaton A copies the part of the virtual genome necessary for the synthesis of a
functional automaton (i.e., transcription from DNA to mRNA in terrestrial life). When automaton
B replicates the virtual genome, the VM in the replicated virtual genome is replaced by another type
of VM belonging to the constitutive information group at a predefined probability mis_copyi_
rate. When automaton A makes a transcript of one part of the virtual genome, the VM is similarly
replaced at a predefined probability mis_syntha_rate. Thus the former case induces changes in
phenotype based on changes in genotype, while the latter one induces changes in phenotype without
changes in genotype. In both cases, the composition of the VMs, VOs, or VIs making up the
functional automata of the VLIs may consequently change. The optimal temperature defined for
each VLI may also change. A new phenotype produced by the results of mutation may often have no
effect or even a negative effect on the life activities of the VLIs. In rare cases, however, a new (i.e.,
mutant) VLI emerges that can live in an environment in which no antecedent (i.e., parent) VLI had
ever before been able to exist. Only when such a characteristic is caused by changes in the genotype
can it be passed on to offspring. In the framework of the simulation on SIVA-T05, therefore, we
define evolutionary adaptation as the phenomenon whereby offspring can live in a place with
environmental conditions under which its parental
individual cannot. If mutation produces a
sequence of VMs that cannot be interpreted in SIVA language, SIVA-T05 neglects it.
4.2 Methods of Experimental Simulation Using SIVA-T05
We conducted two experimental simulations using the AChem system SIVA-T05 to examine the
efficiency of the HBCB model. We evaluated reutilization efficiency of resources produced in the
PSD process in relation to the destination class of the decomposed VP.
For this experiment, we set three mortal species M-I, M-O, and M-M as initial virtual lives, each
of which had a different destination of decomposed VP by its automaton FZ. Here M-I is a mortal
species whose automaton FZ decomposes VP into VI. Similarly, M-O and M-M are mortal species
decomposing VP into VO and VM, respectively. Across these species, automata A and B were also
set to be able to utilize virtual biomolecules belonging to different classes for synthesis or replication.
That is, only VI was available for M-I; VI and VO for M-O; and all of VI, VO, and VM for M-M.
When multiple kinds of virtual biomolecules belonging to different classes are available, the species
utilized them in ascending order of energy amount required for the synthesis, namely, in the order
VM, VO, VI.
We seeded each VLI of the three species separately at the center habitation point in three
ecosystems with identical environmental conditions and conducted simulations of reproduction and
evolution to test the vitality of such species. As a control, we also performed another simulation with
identical conditions using immortal species I-N whose automaton FZ for self-decomposition was
inactivated. I-N is an immortal species with no decomposition. Automata A and B of the species I-N
could utilize only VI as material. Life activities of these primitive VLI were designed to fit the initial
environmental conditions of the center habitation points where they were seeded.
50
Artificial Life Volume 15, Number 1
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
5
1
2
9
1
6
6
2
5
6
6
a
r
t
l
.
/
.
.
2
0
0
9
1
5
1
1
5
1
0
3
p
d
.
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
T. Oohashi et al.
An Effective Hierarchical Model for the Biomolecular Covalent Bond
It is possible that, in the evolutionary process, these four species of VLI would coexist in an
identical ecosystem at the same time, sharing both space and substance. Therefore, we seeded each
VLI of the four species in four spatial blocks with identical environmental conditions located at the
center of one ecosystem and simultaneously started their simulations. Accordingly, these four VLIs
did not have contact with one another at the starting point. We also performed another simulation in
which the four species were seeded in the same spatial block. In this simulation, the VLIs of four
different species had contacted one another when the number of VLIs of each species was only one
or two. Since the success or failure of proliferation at this very early stage decided the whole sim-
ulation process, the results of this simulation —governed, as it was, by chance—became unstable.
Therefore, four VLIs of different species were seeded in different spatial blocks so that they had
contact with one another only after some stable proliferation.
To evaluate the results of the simulations, we compared the life activities of the species from the
viewpoint of changes in the size of the habitation area, the number of individuals, the frequency of
reproduction, the accumulated number of reproductions, and the frequency of mutation. See our
previous report [21] for details of other conditions for the simulations.
4.3 Results of Experimental Simulations
4.3.1 Validity of the PSD Mechanism
Figure 12 shows successive changes of the VLI distribution when each species proliferated in an
independent but identically conditioned ecosystem.
At the beginning of the simulation, species I-N with no self-decomposition mechanism proliferated
prosperously. However, as the habitation area expanded, the gap between the environmental conditions
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
5
1
2
9
1
6
6
2
5
6
6
a
r
t
l
.
/
.
.
2
0
0
9
1
5
1
1
5
1
0
3
p
d
.
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 12. Mortal species in which virtual biological polymers were decomposed into virtual biological monomers in the
self-decomposition process expanded their habitation area most prominently (results of simulations in which four
species proliferated in separate ecosystems with identical environmental conditions). Immortal species I-N (first row in
gray) with no self-decomposition mechanism stopped self-reproduction and expansion of habitation area after time count
(TC) 400. VLIs of mortal species M-I (second row in yellow), M-O (third row in blue), and M-M (bottom row in red),
with a self-decomposition mechanism, repeatedly reproduced and decomposed themselves until the end of the
simulation (TC 2000). Species M-M, with a self-decomposition process oriented to the virtual monomer (VM) class,
conspicuously expanded its habitation area. Increase in population and expansion of the habitation area were more prom-
inent for species M-M than for species M-I and M-O, in which decomposition was oriented to classes lower than VM.
Artificial Life Volume 15, Number 1
51
T. Oohashi et al.
An Effective Hierarchical Model for the Biomolecular Covalent Bond
of the habitation area and their optimal conditions increased, and thus the requirements for adaptation
increased. Indicative of this tendency, the immortal species I-N with no self-decomposition mechanism
reached the limit of its adaptive area at TC 400 and stopped reproduction and expansion of its hab-
itation area.
At the same time, the VLIs of mortal species M-I, M-O, and M-M, having a self-decomposition
mechanism, repeatedly reproduced and decomposed themselves until the end of the simulation
(TC 2000). These cells formed dynamically stable populations, receiving the full benefit of their
restoring substance and space to the ecosystem by the self-decomposing mechanism. A large number
of mutations occurred, reflecting the repeated replication of the virtual genome associated with
reproduction. Thus they continuously produced offspring with a new virtual genome and a corre-
sponding phenotype that enabled them to advance into areas where the initial VLIs could not
survive. As a result, they succeeded in increasing population and expanding their habitation area.
The results of this experimental simulation were completely consistent with our previous findings
[20, 21] that show the validity of the self-decomposing mechanism in a finite, heterogeneous
habitation environment, which is a major characteristic of a terrestrial ecosystem.
4.3.2 Effect of Hierarchization of Biomolecular Covalent Bonds
Species M-I, M-O, and M-M showed significant difference in reproduction and evolution (Figures 12
and 13). Note that species M-I, M-O, and M-M each had a different type of self-decomposition
mechanism with a different destination for the decomposed VP. At the end of the simulation
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
5
1
2
9
1
6
6
2
5
6
6
a
r
t
l
.
/
.
.
2
0
0
9
1
5
1
1
5
1
0
3
p
d
.
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 13. Mortal species in which biological polymers were decomposed into virtual biological monomers in the self-
decomposition process showed a prominent advantage in reproduction and evolution (results of simulations in which
four species proliferated in separate ecosystems with identical environmental conditions). (a) Number of individuals.
(b) Frequency of reproduction. (c) Accumulated number of reproductions. (d) Frequency of mutation. Immortal species
I-N (gray), with no self-decomposition mechanism, stopped reproduction, and mutation did not occur after TC 480.
Mortal species M-I (yellow), M-O (blue), and M-M (red), all with a self-decomposition mechanism, repeatedly
reproduced and decomposed themselves until the end of the simulation (TC 2000). Mutations also repeatedly occurred.
Species M-M, with a self-decomposition process oriented to the VM class, showed greater advantage in all the indexes
than did species M-I and M-O, both with decomposition oriented to classes lower than VM.
52
Artificial Life Volume 15, Number 1
T. Oohashi et al.
An Effective Hierarchical Model for the Biomolecular Covalent Bond
Figure 14. Mortal species in which biological polymers were decomposed into virtual biological monomers in the self-
decomposition process survived and expanded their habitation area most prominently (the result of a simulation in which
four species in an identical ecosystem coexisted simultaneously and interacted). When all the species of VLIs coexisted in
an identical ecosystem, mortal species M-M (red), in which biological polymers were decomposed into VMs, showed
prominent advantages over the other three species. Immortal species I-N (gray), with no self-decomposition mechanism,
stopped reproduction after TC 200. Its habitation area was smaller than when it was seeded in a separate ecosystem.
Mortal species M-I (yellow) and M-O (blue), in which the self-decomposition process was oriented to classes lower than
the VM class, were exterminated by the middle of the simulation. Mortal species M-M survived and expanded its
habitation area up to nearly the same size it had when seeded in a separate ecosystem, and thus showed the greatest
advantage in reproduction among the four species.
(TC 2000), three species showed clear difference in both population and habitation area. By that
measure, they showed greater advantage of reproduction in the following order: [species M-M
oriented to VM class] > [species M-O oriented to VO class] > [species M-I oriented to VI class].
Species M-O and M-M showed themselves to be inferior to species M-I until about TC 400. Two
factors can be mentioned in this regard. First, species M-O and M-M used VIs as a resource for
reproduction but did not return them to the environment. This one-way consumption made the
environmental conditions recede from the optimum, making reproduction disadvantageous at the
beginning of the simulation. Second, because the VLIs of each species inhabited the optimum
habitation area at the beginning of proliferation, differences in reutilizability of resources did not
greatly affect the results.
To understand differences among the four species, we compared changes in the number of in-
dividuals, the frequency of reproduction, the accumulated number of reproductions, and the fre-
quency of mutation (Figure 13). The number of VLIs of species I-N with no self-decomposition
function reached a ceiling at 320 individuals by TC 480. On the other hand, the VLIs of species M-I,
M-O, and M-M continued reproduction and decomposition until the end of the simulation (TC 2000),
when they showed marked differences in proliferation. Reproduction of species M-M showed
overwhelming superiority in all of the indexes.
Generation of VMs by self-decomposition induced a large-scale reduction of the reproduction
cost of not only their own direct offspring but also of all VLIs inhabiting the same environment,
compared with generation of VOs or VIs by self-decomposition. This is because VOs and VIs have
a high degree of reutilizability compared to what VMs have, but involve a larger amount of energy
release and absorption accompanied by synthesis and decomposition of VPs. Such generation
enables all the VLIs in the whole ecosystem to efficiently reproduce themselves. Frequent repro-
duction should induce the occurrence of more mutation and accelerate evolutionary adaptation.
These effects would enhance the activity of species M-M more than that of other species and allow
species M-M to advance into new areas where its initial VLIs could not survive.
As shown above, the effect of the self-decomposition mechanism on reproduction and evolution
was most prominent when the destination of the self-decomposition was oriented to VM. On the
contrary, when the destination was oriented to VO or VI, some effects of self-decomposition on
reproduction and evolutionary adaptation were still preserved, but they were much less prominent.
We obtained even clearer results when the four species proliferated simultaneously in one
ecosystem (Figures 14 and 15). The reproduction of immortal species I-N was extremely constrained
from the beginning of the simulation and reached a ceiling at TC 250. Species M-I, M-O, and M-M
showed almost the same proliferation at the beginning, although species M-I showed a slight ad-
vantage over the others. After approximately TC 300, however, species M-I decreased its habitation
area due to pressure from the other species. It was exterminated at TC 367.
Artificial Life Volume 15, Number 1
53
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
5
1
2
9
1
6
6
2
5
6
6
a
r
t
l
.
/
.
.
2
0
0
9
1
5
1
1
5
1
0
3
p
d
.
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
T. Oohashi et al.
An Effective Hierarchical Model for the Biomolecular Covalent Bond
Figure 15. Mortal species in which biological polymers were decomposed into virtual biological monomers in the self-
decomposition process showed a prominent advantage in reproduction and evolution (result of a simulation in which
four species in an identical ecosystem coexisted simultaneously and interacted). (a) Number of individuals. (b) Frequency
of reproduction. (c) Accumulated number of reproductions. (d) Frequency of mutation. Immortal species I-N (gray),
with no self-decomposition mechanism, stopped reproduction and mutation after TC 250. The two mortal species M-I
(yellow) and M-O (blue), in which biological polymers were decomposed into a class lower than VM, were exterminated
in the middle of the simulation (species M-I: TC 367; species M-O: TC 1167). Mortal species M-M (red), in which self-
decomposition was oriented to the VM class, showed the greatest advantage in all the indexes among the four virtual
species.
Species M-O strove for survival evenly with species M-M until approximately TC 600, but was
exterminated at TC 1167 due to pressure from species M-M. Species M-M increased its speed of
reproduction from approximately TC 800. It showed less prominent proliferation at the beginning,
however, but became prominent gradually. It became the most successful population by TC 800 and
continued to expand its habitation area until the end of the simulation (TC 2000). Such results were
consistent with our previous study in which a virtual life form with a self-decomposition mechanism
surpassed one without it. Moreover, the results suggested that among the three self-decomposable
species, the lower the energy loss for the acquisition of collective reutilizability, the more capable the
species becomes for reproduction and evolution. The results of this simulation correspond well to
the results of the biological experiment in which the PSD mechanism observed in actual terrestrial
life involved hydrolysis as the main process that decomposes a BP into BMs.
The simulations using the AChem system SIVA-T05 suggest that VLIs with a PSD mechanism
attained superiority in reproduction and evolutionary adaptation over VLIs without the PSD mecha-
nism because they had an advantage in the recycling of habitation space and biomaterials and thus
accelerated evolutionary adaptation. This result supports our previous findings as well as the possi-
bility that the self-decomposition mechanism observed in biological experiments utilizing terrestrial
life (protozoa) may exist as a consequence of evolution.
In this experimental simulation based on the HBCB model, we also showed that the effects of
self-decomposition on the acceleration of evolutionary adaptation significantly differed according to
destination classes of the decomposition of VPs. The results showed that decomposition from the
VP class to the VM class had the greatest effect and provided the most prosperity. This suggests that
54
Artificial Life Volume 15, Number 1
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
5
1
2
9
1
6
6
2
5
6
6
a
r
t
l
.
/
.
.
2
0
0
9
1
5
1
1
5
1
0
3
p
d
.
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
T. Oohashi et al.
An Effective Hierarchical Model for the Biomolecular Covalent Bond
the species attaining a high collective reutilizability of resources with minimum energy loss by de-
composing BPs into BMs, as actual terrestrial life does, may be evolutionarily selected from among
other species that decompose BPs into various other materials.
The HBCB model, developed as a way to introduce a hierarchical structure for biomolecular
bond energy, which in turn is central to Suzuki’s NAC, provided valuable findings for understanding
actual terrestrial life. The simulation also confirmed the efficiency of this model.
5 Discussion
The concept of the AChem system proposed by Suzuki [24], namely, a hierarchy based on the bond
energies between chemical elements, has greatly inspired us. Applying his concept and our
knowledge of cell biology, we categorized biomolecules into four classes based on the complexity
of the interatomic network. We also categorized covalent bonds between biomolecules into three
classes based on bond energy. Covalent bonds are the major bond energy of actual biomolecules and
make up one of the classes in Suzuki’s hierarchy [24, 25]. We found that these two classifications fully
corresponded with one another. Incorporating characteristics of an AChem system that is tightly
coupled with actual biomolecules, we constructed the HBCB model to complement our previously
proposed PSD model. In the present study, we carried out two complementary experiments to
examine the validity and effectiveness of both our PSD and our HBCB models. We examined
models in a biological experiment on cells, utilizing actual terrestrial life, and in an AChem simulation
with controls over the evolutionary time scale.
An essential property of AChem is the close correspondence with specific characteristics of
chemical substances and reactions constituting terrestrial life [5]. Consequently, the AChem system
can effectively compare and couple artificial life with an actual terrestrial one [25, 27]. A successfully
designed AChem system is expected to provide a nearly equivalent counterpart to certain forms of
existent terrestrial life. This characteristic of AChem made this approach possible.
Our biological experiment revealed that existent terrestrial life actually possesses programmed
individual death with self-decomposition. The results indicate that self-decomposition is an energy-
requiring, genetically regulated process and mainly consists of hydrolytic decomposition of a BP into
BMs. Moreover, it suggests that terrestrial life is equipped with a biomolecular recycling mechanism
that utilizes the hierarchy of covalent bonds. These results support the existence of a PSD
mechanism and the validity of the HBCB model.
In simulation experiments in which we used SIVA-T05, mortal virtual species with a self-
decomposition mechanism showed clear advantages in reproduction and evolution over immortal
virtual species without a self-decomposition mechanism. This finding validates the effect of a PSD
mechanism in a finite, heterogeneous habitation environment, which is a major characteristic of a
terrestrial ecosystem. In addition, the simulation experiments showed that the virtual species, in
which the self-decomposition process mainly involved covalent bond cleavage from the VP class to
the VM class, showed evolutionary superiority over other species in which the self-decomposition
process involved covalent bond cleavage from the VP class to a lower class than VM.
Thus the results of cell biological experiments using terrestrial life and the results of the sim-
ulation experiment of the HBCB model using SIVA-T05 almost completely supported and com-
plemented one another. The overall results strongly support the hypotheses proposed in the HBCB
model. That is, existent terrestrial life has developed a hierarchical structure based on bond energy
strength, of which the biomolecular recycling system makes use. Moreover, the balance between
energy efficiency and collective reutilizability may have been optimized as a result of the evolution of
the SRSD automaton.
The fruitful outcome of this study could not have been acquired by the single use of either an
artificial ecosystem employing computers, or biology employing living organisms. A discernible
benefit of computer simulation studies using ALife is that, by modeling and simulating various
hypotheses that are theoretically possible although not testable in real terrestrial environments, we
Artificial Life Volume 15, Number 1
55
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
5
1
2
9
1
6
6
2
5
6
6
a
r
t
l
.
/
.
.
2
0
0
9
1
5
1
1
5
1
0
3
p
d
.
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
T. Oohashi et al.
An Effective Hierarchical Model for the Biomolecular Covalent Bond
can evaluate the effectiveness and validity of such hypotheses without resorting to the real world [7,
10, 22]. This advantage allows for the study of problems otherwise out of reach, such as those
related to the ecosystem or evolution. Conventional ALife studies may not fully take into account the
fact that actual terrestrial life activities without exception consist of chemical phenomena [7, 10, 22].
Indeed, the majority of ALife studies during a certain period may have regarded that fact as a kind of
constraint, and tried to dissociate themselves from actual existing life [9]. As a result, conventional
approaches solely employing ALife are not likely to meet expectations of contributing to profound
understanding of actual life.
By organically integrating the two different approaches from artificial and actual life under the
paradigm of AChem, which takes cognizance of the fact that the basic processes of terrestrial life
without exception consist of chemical phenomena, it is expected that further insight can be acquired
that will exceed any obtained through a single approach. We believe that such positive integration will
contribute to the field of AChem.
6 Conclusion
In focusing on the hierarchical structure of biomolecular covalent bonds based on bond energy
in terrestrial life, we have constructed the HBCB model to complement our previously proposed
PSD model. We examined the validity and effectiveness of the models by organically coupling
biological and AChem experiments.
The biological experiments revealed that terrestrial life is equipped with biomolecular recycling
systems that utilize the hierarchy of covalent bonds. The results support the existence of programmed
individual death with self-decomposition. In addition it is suggested that self-decomposition is an
energy-requiring, genetically regulated process and mainly consists of hydrolytic decomposition of a
BP into BMs.
The AChem experiment revealed that, in a finite and heterogeneous environment such as that on
Earth, life that has a biomolecular recycling mechanism with PSD predominates over life without
such a mechanism. Such superiority is most conspicuous when the cleavage of covalent bonds
embodying the self-decomposition process is oriented to the level of the process in which a VP is
decomposed into VMs, where the decomposed products acquire almost complete collective
reutilizability and energy loss is minimized: This is the most plausible process based on the HBCB
model. These results support the validity and effectiveness of the HBCB model.
This approach, based on an integration of AChem and a terrestrial
deepen our understanding of terrestrial life from an evolutionary perspective.
life system, promises to
Acknowledgments
We thank Dr. K. Shimohara of Doshisha University and Dr. H. Suzuki of National Institute of
Information and Communications Technology for their valuable comments on our study; Dr. H.
Sayama of Binghamton University for his contribution to the development of the earlier version of
the SIVA series; and the members of Yamashiro Institute of Science and Culture for their technical
support. This study has been partly supported by the Japan Ministry of Education, Culture, Sports,
Science, and Technology through a Grant-in-Aid for Exploratory Research to M.H. (19659179).
References
1. Bagley, R. J., & Farmer, J. D. (1992). Spontaneous emergence of a metabolism. In C. G. Langton,
C. Taylor, J. D. Farmer, & S. Rasmussen (Eds.), Artificial life II (pp. 93 – 140). Redwood City, CA:
Addison-Wesley.
2. Bagley, R. J., Farmer, J. D., & Fontana, W. (1992). Evolution of a metabolism. In C. G. Langton,
C. Taylor, J. D. Farmer, & S. Rasmussen (Eds.), Artificial life II (pp. 141 – 158). Redwood City, CA:
Addison-Wesley.
3. Berg, J. M., Tymoczko, J. L., & Stryer, L. (2002). Biochemistry (5th ed.). New York: W.H. Freeman.
56
Artificial Life Volume 15, Number 1
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
5
1
2
9
1
6
6
2
5
6
6
a
r
t
l
.
/
.
.
2
0
0
9
1
5
1
1
5
1
0
3
p
d
.
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
T. Oohashi et al.
An Effective Hierarchical Model for the Biomolecular Covalent Bond
4. Berry, G., & Boudol, G. (1992). The chemical abstract machine. Theoretical Computer Science, 96(1), 217 – 248.
5. Dittrich, P., Ziegler, J., & Banzhaf, W. (2001). Artificial chemistries — A review. Artificial Life, 7(3),
225 – 275.
6. Hill, D. L. (1972). The biochemistry and physiology of Tetrahymena. New York: Academic Press.
7. Holland, J. H. (1975). Adaptation in natural and artificial systems. Ann Arbor: The University of
Michigan Press.
8. Kovacs, P., & Csaba, G. (1988). Effect of inhibition of endocytosis, recycling and lysosomal activity
on the insulin binding capacity and imprintability of Tetrahymena. Acta Physiologica Hungarica, 71(2),
315 – 322.
9. Langton, C. G. (1984). Self-reproduction in cellular automata. Physica, 10D, 135 – 144.
10. Langton, C. G. (1989). Artificial life. In C. G. Langton (Ed.), Artificial life (pp. 1 – 47). Reading, MA:
Addison-Wesley.
11. Lehninger, A. L. (1971). Bioenergetics: The molecular basis of biological energy transformations (2nd ed.). New York:
W. A. Benjamin.
12. McKee, T., & McKee, J. R. (2002). Biochemistry: The molecular basis of life (3rd ed.). Boston, MA:
WCB/McGraw-Hill.
13. Mpoke, S. S., & Wolfe, J. (1997). Differential staining of apoptotic nuclei in living cells: Application
to macronuclear elimination in Tetrahymena. Journal of Histochemistry and Cytochemistry, 45(5), 675 – 683.
14. Nozawa, Y., & Thompson, G. A. J. (1971). Studies of membrane formation in Tetrahymena pyriformis. II.
Isolation and lipid analysis of cell fractions. Journal of Cell Biology, 49(3), 712 – 721.
15. Nozawa, Y. (1975). Isolation of subcellular membrane components from Tetrahymena. Methods in Cell Biology,
10, 105 – 133.
16. Odum, E. P. (1971). Fundamentals of ecology (3rd ed.). Philadelphia: W.B. Saunders.
17. Ono, N., & Ikegami, T. (2000). Self-maintenance and self-reproduction in an abstract cell model. Journal
of Theoretical Biology, 206(2), 243 – 253.
18. Oohashi, T., Nakata, D., Kikuta, T., & Murakami, K. (1987). Programmed self-decomposition model
(in Japanese). Kagakukisoron, 18(2), 21 – 29.
19. Oohashi, T., Sayama, H., Ueno, O., & Maekawa, T. (1996). Artificial life based on programmed self-decomposition
model. (ATR technical report TR-H-198.) Kyoto: Advanced Telecommunications Research Institute
International.
20. Oohashi, T., Maekawa, T., Ueno, O., Nishina, E., & Kawai, N. (1999). Requirements for immortal ALife to
exterminate mortal ALife in one finite, heterogeneous ecosystem. Proceedings of the 5th European Conference on
Advances in Artificial Life (pp. 49 – 53). London: Springer-Verlag.
21. Oohashi, T., Maekawa, T., Ueno, O., Kawai, N., Nishina, E., & Shimohara, K. (2001). Artificial life
based on the programmed self-decomposition model: SIVA. Journal of Artificial Life and Robotics, 5(2),
77 – 87.
22. Ray, T. S. (1992). An approach to the synthesis of life. In C. G. Langton, C. Taylor, J. D. Farmer, &
S. Rasmussen (Eds.), Artificial life II (pp. 371 – 408). Redwood City, CA: Addison-Wesley.
23. Suzuki, H. (2001). String rewriting grammar optimized using an evolvability measure. Proceedings of the 6th
European Conference on Advances in Artificial Life (pp. 458 – 468). London: Springer-Verlag.
24. Suzuki, H. (2004). Network artificial chemistry — Molecular interaction represented by a graph. In
M. Bedau, P. Husbands, T. Hutton, S. Kumar, & H. Suzuki (Eds.), Proceedings of Ninth International
Conference on the Simulation and Synthesis of Living Systems (ALIFE9) workshop and tutorial (pp. 63 – 70).
Boston.
25. Suzuki, H. (2005). Mathematical folding of node chains in network artificial chemistry. Proceedings of Sixth
International Workshop on Information Processing in Cells and Tissues (pp. 52 – 68). York.
26. Tomita, M., Hashimoto, K., Takahashi, K., Shimizu, T. S., Matsuzaki, Y., Miyoshi, F., Saito, K., Tanida, S.,
Yugi, K., Venter, J. C., & Hutchison, C. A. (1999). E-CELL: Software environment for whole cell
simulation. Bioinformatics, 15(1), 72 – 84.
27. Tomita, M. (2001). Whole cell simulation: A grand challenge of the 21st century. Trends in Biotechnology, 19(6),
205 – 210.
Artificial Life Volume 15, Number 1
57
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
5
1
2
9
1
6
6
2
5
6
6
a
r
t
l
.
/
.
.
2
0
0
9
1
5
1
1
5
1
0
3
p
d
.
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
T. Oohashi et al.
An Effective Hierarchical Model for the Biomolecular Covalent Bond
28. Ueno, O., Nozawa, Y., & Oohashi, T. (2002). The pulse heat shock method induces cellular
decomposition process in cell death of Tetrahymena. Acta Scholae Medicinalis Universitatis in Gifu, 50(1),
10 – 19.
29. Varela, F. G., Maturana, H. R., & Uribe, R. (1974). Autopoiesis: The organization of living systems.
BioSystems, 5(4), 187 – 196.
30. von Neumann, J. (1951). The general and logical theory of automata. In L. A. Jeffress (Ed.), Cerebral
mechanisms in behavior — The Hixon symposium (pp. 1 – 41). New York: John Wiley & Sons.
31. von Neumann, J. (1966). Theory of self-reproducing automata. Urbana: The University of Illinois Press.
32. Watanabe, Y. (1963). Some factors necessary to produce division conditions in Tetrahymena pyriformis.
Japanese Journal of Medical Science & Biology, 16(6), 107 – 124.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
1
5
1
2
9
1
6
6
2
5
6
6
a
r
t
l
.
/
.
.
2
0
0
9
1
5
1
1
5
1
0
3
p
d
.
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
58
Artificial Life Volume 15, Number 1